Aufbau und Funktionen der Zellmembranen
Werbung

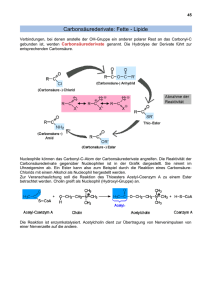
Lipide (Wiederholung) Wiederholung Lipide (Wiederholung) Unter dem Begriff Lipide werden verschiedene fettartige organische Verbindungen zusammengefasst, die in chemischer Hinsicht zum Teil recht unterschiedlich aufgebaut sind. Ihr gemeinsames Merkmal ist die gute Löslichkeit in apolaren Lösungsmitteln wie Hexan, Diethylether und Chloroform; in Wasser sind Lipide unlöslich. Manche sind allerdings infolge ihrer Grenzflächenaktivität auch in wässriger Umgebung existenzfähig. Eine übliche Klassifizierung der Lipide unterscheidet: freie Fettsäuren Neutralfette Glycerinphosphatide Sphingolipide Steroide Terpene 2015 1 Einige häufige Fettsäuren cis 2010V1 Fettsäuren Eine Fettsäure (hier Palmitinsäure) besteht aus einer hydrophoben Kohlenwasserstoffkette und einer hydrophilen Carbonsäure-Gruppe. Hydrophiler Carbonsäure-“Kopf“ “amphiphil“ Hydrophober Kohlenwasserstoff-“Schwanz“ 2006V1 2 Ungesättigte Fettsäuren Um diese Doppelbindung ist eine freie Rotation nicht möglich – die Kette ist geknickt. Meistens „cis“ ungesättigte Fettsäure (Ölsäure) gesättigte Fettsäure (Stearinsäure) 2006V1 Trigylceride (Neutralfette) Neutralfette sind Fettsäure-Ester des Glycerins. Glycerol (Glycerin) Triglycerid Die Konsistenz des Fettes ist abhängig vom Gehalt an ungesättigten Fettsäuren. Je höher der Anteil an ungesättigten Fettsäuren, desto flüssiger wird das Fett (Öl!), d.h. es hat einen niedrigeren Schmelzpunkt. 2006V1 3 Raumbedarf von gesättigten und ungesättigten Fettsäuren 02_19_fatty side chains.jpg Cis versus trans-Fettsäuren! 2016 2006V1 Glycerinphosphatide Glycerinphosphatide (“Phospholipide“) sind die Hauptbestandteile der Zellmembranen. In den Glycerinphosphatiden sind zwei der OH-Gruppen des Glycerins mit Fettsäuren verestert, die dritte OH-Gruppe hängt an einem Phosphat, das seinerseits mit einem weiteren Alkohol verbunden ist. Je nach der Alkoholverbindung unterscheidet man: • Phosphatidylcholin (Lecithin) • Phosphatidylethanolamin • Phosphatidylserin • Phosphatidylinosit(ol) Phospholipide 2010V1 4 Glycerinphosphatide Das Bild k ann zurzeit nicht angezeigt werden. hydrophile Gruppe Cholin hydrophobe Fettsäureschwänze Phospholipide 2010V1 Glycerinphosphatide (Lecithin) Phospholipide 2008V1 5 Sphingolipide Sphingolipide bestehen ebenfalls aus einem hydrophoben Bereich mit zwei langen Kohlenwasserstoffketten und einem polaren Bereich, der sich aus ein oder mehreren Zuckerresten zusammensetzt. Das Basismolekül ist das Sphingosin, ein Aminoalkohol. Eine Fettsäure ist über eine Amidbindung an die Aminogruppe gebunden. 2010V1 Sphingolipide Einfaches Glycolipid Ganglioside: verzweigte Zuckerketten, oft mit Sialinsäure 2010V1 6 Weitere Lipide Zwei weitere häufige Lipid-Typen sind die Steroide und die Poly-Isoprenoide. Beide Verbindungsklassen basieren auf dem Isopren. Isopren 2006V1 Poly-Isoprenoide Dolichol ist ein Polymer des Isoprens und wirkt als Überträger von aktivierten Zuckermolekülen bei der Biosynthese von Glykoproteinen. Dolichol-Phosphat Als weiteres Beispiel für Poly-Isoprenoide das Retinol (Vitamin A) : 2010V1 7 Steroide Die Basis-Struktur der Steroide ist ein kondensiertes Ringsystem bestehend aus drei Sechserringen und einem Fünferring: 2010V1 Cholesterol 2010V1 8 Vom Cholesterol abgeleitete Hormone 2010V1 3-Andostrenol Humanes Pheromon 2013 GW 9 Lipid-Aggregate Fettsäuren haben einen hydrophilen Kopf und einen hydrophoben Schwanz. In Wasser können sie einen Oberflächenfilm oder kleine Micellen bilden: Triglyceride (Fette) bilden große sphärische Fett-Tröpfchen in der Zelle: Phospholipide und Glykolipide bilden selbst-schließende Lipid-Doppelschichten, die das Bauprinzip aller zellulären Membranen sind: 2006V1 Die biologische Membran Zellmembran und Membranen der Organellen 2016 10 Die verschiedenen Kompartimente einer eukaryontischen Zelle, die von einer Membran umschlossen sind: endoplasmatisches Reticulum Zellkern Peroxisom Lysosom Golgi Apparat Vesikel Mitochondrium Plasmamembran 2010V1 Plasmamembran und interne Membranen als Barriere 11_01_Cell.membranes.jpg 2010V1 11 Funktionen der Plasmamembran 11_02_plasma.membra.jpg 2010V1 Transportprozesse durch zelluläre Membranen 12_05_carrier_proteins.jpg 2010V1 12 Modell der Zellmembran LipidDoppelschicht (5 nm) Elektronenmikroskopische Aufnahme der ErythrocytenMembran Protein Lipid “Flüssig-Mosaik-Modell”: Beweglichkeit der Membranlipide (und eingelagerter Membranproteine) 2010V1 Basalmembran / Basallamina von Endothelzellen der Kapillaren Blut-Hirn Schranke https://www.google.at/url?sa=i&rct=j&q=&esrc=s&source=images&cd=&cad=rja&uact=8&ved=0ahUKEwj3o62IrZrKAhWJuRQKHTqlB7oQj B0IBg&url=http%3A%2F%2Fwww.nature.com%2Fnrc%2Fjournal%2Fv3%2Fn5%2Ffig_tab%2Fnrc1073_F5.html&psig=AFQjCNFMwpicZ4 nKWCVZHf510v5FOpVMjA&ust=1452347198219707 https://www.google.at/search?q=vaskul%C3%A4res+Endothel&source=lnms&tbm=isch&sa=X&ved=0ahUKEwjS2JeZrJrKAhWB2BQKHcLMCtkQ_AUIBygB&biw=1702&bih=902#imgrc=I9v81vTDq42BQM%3A 2016 13 Aufbau von Glycerinphosphatiden (Phospholipiden) 11_06_Phosphatidylch.jpg 2010V1 Die Lipid-Bestandteile der Zell-Membran hydrophiler Abschnitt 11_07_amphipathic.jpg hydrophober Abschnitt 2010V1 14 Ausrichtung der Phospholipide in der Membran Amphipathische (amphiphile) Moleküle hydrophiler Kopf Wasser PhospholipidDoppelschicht zwei hydrophobe FettsäureSchwänze Phospholipid-Molekül 2010V1 Triglyceride Phospholipide hydrophob 11_10_Fat_phospholip.jpg amphipathisch 2010V1 15 Einbau von Cholesterol in die Zellmembran Phospholipid Cholesterin polare Kopfgruppe starrer ebener Steroidring unpolarer Kohlenwasserstoff-Schwanz Cholesterol Cis versus trans-Fettsäuren! Membran wird rigieder 2016 Die Lipid-Doppelschicht im Querschnitt 11_11_bilayer.in.H20.jpg “Flüssig-Mosaik-Modell”: Beweglichkeit der Membranlipide (und eingelagerter Membranproteine) 2010V1 16 Leichte Reparatur bei Verletzungen 11_12_sealed.compar.jpg 2010V1 Liposomen 11_13_liposomes.jpg 2010V1 17 Beweglichkeit der Phospholipide im Membranverband 11_15_Phospho,move.jpg 2010V1 Membran- Fluidität hängt ab von Temperatur – höhere Temperatur höhere Fluidität Zusammensetzung – kürzere Fettsäuren können weniger gut interagieren höhere Fluidität – ungesättigte cis-Fettsäuren können sich weniger gut zusammenpacken höhere Fluidität – mehr Cholesterol führt zu niedriger Fluidität (rigidere Membranen) 2016 18 Asymmetrische Verteilung der Lipid-Komponenten Phosphatidylcholin Glycolipid Sphingomyelin Cholesterol Phosphatidylserin Phosphatidylethanolamin Phosphatidylinosit(ol) 2010V1 Synthese von Membranen 2010V1 19 Synthese von Membranen 11_18_Flippases.jpg 2010V1 Vesikulärer Transport z.B.: Transport vom ER Lumen zur Plasmamembran 11_19_memb.vesicles.jpg Identität der Membran-Disposition: Zytosolisch bleibt zytosolisch, Lumen der Vesikel (intrazellulär) wird extrazellulär 2010V1 2016 20 Permeabilität einer Lipid-Doppelschicht für verschiedene Molekülklassen O2 CO2 N2 Benzol Gase, kleine apolare Moleküle H2O Glycerin Ethanol kleine ungeladene polare Moleküle größere ungeladene polare Moleküle Ionen Aminosäuren Glucose Nucleotide H+ , Na+, HCO3- , K+, Ca++ , Cl-, Mg++ synthetische Lipid-Doppelschicht 2010V1 Einige Funktionen von Plasmamembran-Proteinen Transportproteine Verbindungsproteine Rezeptoren EXTRAZELLULÄRER RAUM Enzyme 11_20_memb.proteins.jpg CYTOSOL 2010V1 21 Beispiele für Proteine der Plasmamembran Funktionelle Protein Klasse Transportproteine Na+-Pumpe Pyruvat/H+ Symporter Ankerproteine Integrine Signal Recognition Particle Receptor Funktion pumpt Na+ aus der Zelle hinaus und K+ hinein verbinden intrazelluläre Aktinfilamente mit extrazellulären Matrixproteinen Rezeptoren Platelet-derived Signalübertragung growth factor (PDGF)- (extrazelluläres PDGF Rezeptor Wachstum / Zellteilung) Insulin Receptor Enzyme Adenylatcyclase Succinat Dehydrogenase (Komplex II der Atmungskette) katalysiert die Erzeugung von intrazellulärem cAMP als Antwort auf extrazelluläre Signale 2016 Möglichkeiten, Membran-Proteine mit der Lipid-Doppelschicht zu verknüpfen LipidDoppelschicht Glycosyl-phosphatidyl-Inositol Anker (GPI-Anchor) 2010V1 22 Transmembran-Abschnitt von Proteinen hat meist -Helix-Struktur hydrophobe AminosäureSeitenkette Wasserstoff-Brücke Phospholipid -Helix 2010V1 Mehrere -Helices bilden eine hydrophile Pore durch die Membran gefüllte Wasserpore Transmembrane -Helix 11_24_hydrophl.pore.jpg Lipid-Doppelschicht 2010V1 23 Dreidimensionale Struktur von Porin 11_25_Porin.proteins.jpg 2010V1 Möglichkeiten, die laterale Mobilität von Membran-Proteinen zu beschränken Bindung an den Zell-Cortex Membranskelett Bindung an extrazelluläre Matrixmoleküle ECM= extrazelluläre Matrix 11_35_lateral.mobility.jpg Bindung an Proteine an der Oberfläche einer anderen Zelle Diffusionsbarrieren 2010V1 24 Begrenzung von Membran-Proteinen auf bestimmte Membran-Domänen apikale Plasmamembran laterale Plasmamembran 11_39_tight.junction.jpg basale Plasmamembran 2010V1 Die Glycocalyx einer eukaryontischen Zelle TransmembranGlycoprotein Adsorbiertes Glycoprotein Transmembran Proteoglycan Zellhülle (Glycocalyx) 11_32_sugar.coated.jpg LipidDoppelschicht 2010V1 25 Die Erkennung von Kohlenhydraten an der Oberfläche von Neutrophilen 11_33_neutrophils.jpg Anderes Beispiel: Erkennung Spermium-Eizelle 2010V1 Das Membranskelett der Erythrocyten-Membran Cytosolische Seite der Membran Spectrin Actin VerbindungsProtein Transmembran-Proteine 11_30_blood.cells.EM.jpg 2016 26 Detergenzien lösen Membranproteine 2010V1 27