Entwicklungssteuerung durch Gene
Werbung

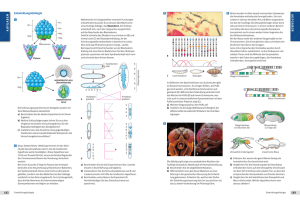
4 Entwicklungssteuerung durch Gene 176.1 Aufbau der VorderpolHinterpolPolarität in einem Embryo von Drosophila Zellkern bicoidmRNA BicoidProtein Bicoid-Protein-Gradient in Blastodermkernen 176.2 Abfolge der Expression entwicklungssteuernder Gene bei Drosophila Produkte der maternalen Polaritäts-Gene Gap-Gene PaarregelGene Segmentpolaritäts-Gene 176 Entwicklungsbiologie A B Die anschließend aktivierten Segmentpolaritäts-Gene legen die endgültigen Segmentgrenzen der Larve sowie die Ausrichtung der Segmente entlang der Körperlängsachse fest. Mutationen dieser Gene führen zu Embryonen mit veränderten Polen. Walter GEHRING entdeckte 1984, dass in den homöotischen Genen von Drosophila eine DNA-Sequenz von 180 Basenpaaren immer wiederkehrte. Er nannte diese identischen DNA-Abschnitte Homöobox. Diese wird in ein 60 Aminosäuren langes Protein translatiert, die Homöodomäne, die als Transkriptionsfaktor andere Gene kontrolliert. Bisher hat man bei allen untersuchten Organismen, Pflanzen, Hefepilzen, Seeigeln, Fröschen und Menschen, Homöobox-Gene gefunden. Diese sind scheinbar ein universelles Steuerelement, denn es gibt zum Beispiel Froschproteine, die beim Drosophila-Embryo die gleiche Wirkung bei der Differenzierung in Bauch und Rücken haben. Bei Wirbeltieren werden die Homöobox-Gene als Hox-Gene, bei Gliedertieren wie Insekten als Hom-Gene bezeichnet. Die Hox- und Hom-Gene sind gruppenweise in Clustern verteilt. Bei Drosophila liegen die Cluster auf zwei Chromosomen, bei Wirbeltieren wie Mensch und Maus sind sie auf vier Chromosomen verteilt, trotzdem können die Cluster von Maus und Insekt homologisiert werden. Die Reihenfolge der Gene eines Clusters bestimmt die Abfolge der Expression und gleichzeitig den Wirkort. Homöotische Gene sind auch bei der Gestaltbildung von Pflanzenkörpern beteiligt. Sie bestimmen bei diesen die Abfolge verschiedener Blattformen im Blütenbereich. A = Abdominalsegmente T = Thoraxsegmente 177.1 Segmentierung des DrosophilaKörpers Flügel Haltere T1 T2 T3 A5 – A8 A4 A3 A2 A1 177.2 BithoraxMutante „Doppelbrust“. Homöotische Umformung des Segments T3 zu einem zweiten Segment T2 Bei der Erforschung der pflanzlichen Entwicklung hat ein Kreuzblütler, die Ackerschmalwand Arabidopsis thaliana, eine ähnliche Stellung wie Drosophila. Sie ist besonders gut als Modell geeignet, weil ihr Entwicklungszyklus nur sechs Wochen dauert und die Pflanzen nur 10 bis 20 Zentimeter hoch wachsen. Außerdem besitzt die Pflanze ein relativ kleines Genom. Neun Gene bestimmen die Organisation des Pflanzenembryos. Über Mutanten konnte die genetische Steuerung der embryonalen Pflanzenentwicklung aufgeklärt werden. Die beobachteten homöotischen Mutationen betrafen unter anderem die Polarität der Pflanzen. Die in Magdeburg geborene Christiane NÜSSLEINVOLHARD arbeitete zusammen mit dem Amerikaner Eric WIESCHAUS über die Gestaltbildung und Entwicklungskontrollgene bei Drosophila. Sie wurde für ihre Ergebnisse als erste deutsche Wissenschaftlerin zusammen mit WIESCHAUS und dem Amerikaner Edward LEWIS 1995 mit dem Nobelpreis geehrt. Arabidopsis P O RT R ÄT 176.3 Expression von Segmentierungsgenen im DrosophilaEmbryo. A Fluoreszenzmarkierung der Produkte der Segmentierungsgene; B Strukturierung der Larve in 14 Segmente homöotische Gene Von rund 20 000 Drosophila-Genen sind etwa 100 an der Steuerung von Entwicklungsprozessen beteiligt. Durch Ausschalten einzelner Gene erzeugten Christiane NÜSSLEIN-VOLHARD und Eric WIESCHAUS Mutanten. Anhand des veränderten Phänotyps konnten sie auf die eventuell gestörte Funktion eines Entwicklungskontrollgens schließen. In einer hierarchischen Kaskade kontrollieren diese Gene die Entwicklung eines Drosophila-Embryos in drei Stufen: • Im noch unbefruchteten Ei werden Vorderund Hinterpol bestimmt; • dann wird die Gliederung des Embryos in Segmente festgelegt und • schließlich werden die unterschiedliche Morphologie und die Funktionsmöglichkeiten der Segmente bestimmt. Die Entwicklungskontrollgene lassen sich in maternale und zygotische Gene einteilen. Letztere stehen unter der direkten Kontrolle der maternalen Gene. Zum Beispiel wird das maternale Polaritätsgen bicoid in Zellen des Eierstocks transkribiert. Die bicoid-mRNA wird in den Vorderpol des Eies eingelagert und nach der Eiablage in das Protein Bicoid translatiert. Dessen Konzentration nimmt vom Vorder- zum Hinterpol ab. Das Protein Bicoid kontrolliert seinerseits nachgeschaltete zygotische Entwicklungskontrollgene. Ist das bicoid-Gen (lat., zwei Hinterleibern ähnlich) mutiert, entsteht ein kopf- und brustloser Embryo. Durch zygotische Gap-Gene (engl., Lücke) wird der noch ungegliederte Embryo entlang der Körperlängsachse grob in breite Zonen unterteilt. Mutieren diese Gene, fallen in der Larve breite Bereiche aus, beispielsweise bei einer Mutante Kopf- und Brust(Thorax)-Segmente sowie zwei weitere Segmente. Die Produkte der Gap-Gene aktivieren Paarregel-Gene: Durch sie wird der Insektenembryo zellulär gegliedert und die Zellen werden bestimmten Körperabschnitten zugeordnet. Mutieren diese Gene, fällt jedes zweite Segment aus, daher rührt der Name Paarregel-Gen. Nachdem die Aktivität der SegmentierungsGene den Drosophila-Embryo in regelmäßige Segmente unterteilte, werden diese von homöotischen Genen (gr. homoio, gleichartig) in Kopf-, Thorax- oder Hinterleibssegmente differenziert. Mutieren solche Gene, entstehen Körperabschnitte, die bereits vorhandenen ähneln, weil Teile wie Antennen, Flügel, Beine oder Augen gebildet werden, die nicht zu den Segmenten passen. Die meisten homöotischen Gene gehören entweder zum Antennapedia-Komplex, der die Bildung der Körperteile am Kopf und den vorderen Brustsegmenten steuert, oder zum Bithorax-Komplex. Dieser bestimmt die Körperanhängsel des hinteren Brustsegmentes und der Hinterleibs(Abdominal)-Segmente. Edward LEWIS forscht seit 1978 intensiv zum Bithorax-Komplex. Die zugehörige BithoraxMutante „Doppelbrust“ wurde schon 1923 von BRIDGES beschrieben. Aber erst LEWIS konnte ihr Zustandekommen erklären. Christiane Nüsslein-Volhard (*1942) Entwicklungsbiologie 177