15 Signaltransduktionswege: eine Einführung in den
Werbung

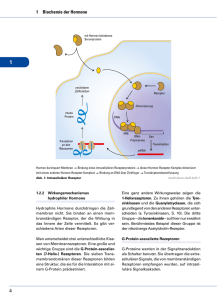
15 Signaltransduktionswege: eine Einführung in den Informationsstoffwechsel Membranrezeptoren übertragen Informationen aus der Umgebung ins Zellinnere. Die meisten Signalmoleküle sind jedoch zu gross, um die Zellmembran zu passieren. Die Signale müssen übertragen werden, ohne dass die Moleküle selbst in die Zelle gelangen. Die Informationen werden durch sekundäre Botenstoffe in das Zellinnere geleitet. Dies geschieht meist mittels Phosphorylierung von Proteinen 15.1 Rezeptoren mit sieben Transmembranhelices ändern nach der Bindung eines Liganden ihre Konformation und aktivieren G-Proteine Rezeptoren mit sieben Transmembranhelices (7TM-Rezeptoren) sorgen für die Übermittlung von Informationen. Durch die Bindung eines Liganden aus der Umgebung der Zelle führt es im 7TM-Rezeptor zu einer Konformationsänderung, die im Zellinnern zu spüren ist. Es führt zur Aktivierung eines G-Proteins in dem diese mit GTP gebunden werden. Die Aktivierten GProteine regen andere Proteine an. Diese Proteine sind die Adenylat-Cyclase-Enzyme, die dann den sekundären Botenstoff cAMP erzeugt. Durch die Zunahme der cAMPKonzentration wird die Reaktion auf das ursprüngliche Signal. Nach einer gewissen Zeit wird das GTP zu GDP und Pi versetzt. Somit fällt das G-Protein wieder in den Ausgangszustand. Ligand + Rezeptor ↓ aktivierter Rezeptor ↓ aktiviertes G-Protein ↓ aktivierte Adenylat-Cyclase ↓ höhere [cAMP] ↓ aktivierte Effektoren 15.2 Die Hydrolyse von Phosphatidylinositolbisphosphat durch die Phospholipase C lässt zwei Botenstoffe entstehen cAMP ist nicht der einzig sekundäre Botenstoff. Durch die Phosphoinositidkaskade entstehen zwei weitere Botenstoffe: Inositoltrisphosphat und Diacylglycerin. Ionisitoltrisphospat öffnet Calciumkanäle in den Membranen das endoplasmatischen Reticulums, was einen erhöten Ca2+-Spiegel im Cytosol zur Folge hat. Das Diacylglycerin aktiviert die Proteinkinase C, die Serin- und Threoninreste in Zielproteinen phosphoryliert. 15.3 Calcium ist im Cytosol ein weit verbreiteter Botenstoff Die Ca2+-Konzentration im Cytosol kann zum Zweck der der Signalübertragung sehr plötzlich gesteigert werden, idem sich in der Plasmamembran oder einer Membran im Zellinneren vorübergehend Calciumkanäle öffnen. Ca2+ eignet sich aber auch als guter Botenstoff für das Zellinnere, weil es sehr fest an Proteine binden kann. Calmodulin wird duch die Bindung von Calciumionen aktiviert. Calmodulin ist ein EF-HandProtein. D.h. es besteht aus einer Helix, einer Schleife und einer weiteren Helix und kann Ca2+ binden. Ohne Ca2+ ist Calmodulin nur im Kern hydrophob. Durch Calciumionen werden auch an der Oberfläche hydrophobe Abschnitte freigelegt, die als Anheftstellen für Zielproteine dienen. Der Komplex aus Ca2+ und Calmodulin aktiviert eine grosse Zahl von Enzymen, Pumpen und anderen Zeilproteinen. 15.4 Manche Rezeptoren bilden nach der Ligandenbindung Dimere und geben Signale durch gegenseitige Phosporylierung weiter 7TM-Rezeptoren setzen Signaltranduktionswege duch Veränderung der Tertiärstruktur in Gang, die durch die Bindung des Liganden ausgelöst werden. Bei manchen Rezeptoren führt die Bindung des Liganden zu einer Veränderung der Quartärstruktur oder anders ausgedrückt: die Rezeptoren bilden Dimere. Ein solcher Rezeptor enthält eine extrazelluläre Domäne, die den Liganden bindet. Er besitzt aber auch ene Domäne im Cytosol, die eine Proteinkinase bindet oder selbst eine solche Enzymaktivität enthält. Ein Signalübertragnugsweg dieses Typs geht vom Wachstumshormonrezeptor aus. Durch die Dimerbildung dieses Rezeptors wird die Januskinase 2 (JAK2) aktiviert. Ist die JAK2 aktiviert, kann sie andere Substrate phosphorylieren. Andere Rezeptoren enthalten in ihrer kovalenten Struktur Tyrosin-Kinase-Domänen. Beispiele sind Rezeptoren von Wachstumsfaktoren wie Insulin, Epidermiswachstumsfaktor (EGF) und Blutplättchenwachstumsfaktor. Die Phosphorylierung des EGF-Rezeptors ist über ein Adapterprotein und eine Kette von Proteinphosphorylierungreaktionen mit der Stimulation des Zellwachstums gekoppelt. Sobald der Rezeptor phosphoryliert ist, bindet die SH2-Domäne de Adapterproteins Grb-2 an die Phosphaotyrosinreste der Rezeptor-TyrosinKinase. Anschiessend zieht Grb-2 ein Protein namens Sos hinzu, dann mit Grb-2 über zwei SH3-Domänen in Wechselwirkung tritt. Sos aktiviert das Protein Ras das das Zellwachstum Reguliert. Signal (EGF) ↓ Rezeptor-Tyrosin-Kinase (EGF-Rezeptor) ↓ Adaperprotein (Grb-2) ↓ Guaninnucleotidaustauschfaktor (SoS) ↓ Effektor (Ras) ↓ Reaktion (Zellproliferation)