Taschenatlas Ernährung - Biesalski / Grimm - Beck-Shop
Werbung

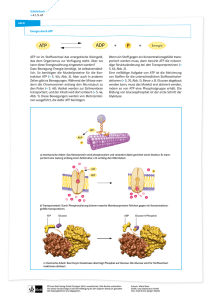
Taschenatlas Ernährung von Hans Konrad Biesalski, Peter Grimm Neuausgabe Taschenatlas Ernährung – Biesalski / Grimm schnell und portofrei erhältlich bei beck-shop.de DIE FACHBUCHHANDLUNG Thematische Gliederung: Ernährungsmedizin, Diätetik Thieme 2007 Verlag C.H. Beck im Internet: www.beck.de ISBN 978 3 13 115354 8 Inhaltsverzeichnis: Taschenatlas Ernährung – Biesalski / Grimm 30 Energiehaushalt Energie in Geweben Das Gehirn ist fast ausschließlich auf Glucose als Energielieferant angewiesen. Da es oxidierbare Substanzen nicht speichern kann, muss es stndig mit Glucose versorgt werden. Dies kann nur durch einen Mindest-Glucosegehalt im Blut gewhrleistet werden (A). Das Gehirn verbraucht ca. 120 g Glucose pro Tag; im Hungerzustand kann es in begrenztem Umfang auch Ketonkrper verarbeiten. Im Gegensatz zum Gehirn besitzt die Muskulatur enorme Glykogenspeicher. Beim Abbau entsteht daraus innerhalb der Muskelzellen Glucose-6-Phosphat, das jedoch nicht mehr hydrolisiert werden kann. Es geht daher ausschließlich in die Glykolyse ein und kann nicht in Form von Glucose an das Blut abgegeben werden. Insbesondere bei pltzlicher Aktivittssteigerung ist Glucose der wichtigste Brennstoff fr die Muskelzelle. Das unter anaeroben Bedingungen gebildete Lactat wird ins Blut abgegeben. Die im Fettgewebe gespeicherten Triglyceride stellen den wesentlichen Brennstoffvorrat des Menschen dar. Zur Veresterung von Fettsuren bentigt das Fettgewebe aktiviertes Glycerin. Da jedoch das Enzym hierzu, die Glycero-Kinase, im Fettgewebe fehlt, kann bei der kontinuierlich ablaufenden Hydrolyse von Triglyceriden das anfallende Glycerin nicht verwendet werden. Das aktivierte Glycerin muss daher der Glykolyse entnommen werden. Aus diesem Grund ist die Zelle fr die Fettsynthese auf eine ausreichende Glucosezufuhr angewiesen – ein Umstand, den sich viele zweifelhafte „Diten“ zunutze machen. Die Leber ist das zentrale Organ fr die Regulation des Stoffwechsels. Sie ist in der Lage, große Mengen an Glucose aufzunehmen, in Form von Glykogen zu speichern und bei Bedarf zur Stabilisierung des Blut-Glucose-Spiegels bereit zu stellen. Solange ausreichend Brennstoffe vorhanden sind, synthetisiert die Leber auch Fettsuren, verestert sie und transportiert sie dann ber in der Leber gebildete Lipoproteine zu den peripheren Geweben. Im Hungerzustand wandelt die Leber jedoch Fettsuren verstrkt zu Ketonkrpern um (B). Ihre Synthese in der Leber beginnt, wenn das Angebot an Acetyl-CoA dessen Abbaugeschwindigkeit im Citrat-Zyklus bersteigt. Die Ketonkrper werden von allen Geweben außer der Leber selbst zur Energiegewinnung herangezogen. Gleichzeitig werden aus dem Proteinabbau entstehende Aminosuren zur Gluconeogenese genutzt, um den Mindestbedarf an Glucose zu gewhrleisten. Rein rechnerisch wrden die Fettreserven des Menschen fr mehr als 2 Monate ausreichen. Allerdings sind nur ca. 3 kg Protein mobilisierbar, was den normalen Glucosebedarf des Nervengewebes fr ca. 15 Tage decken knnte. Eine lngere Fastenperiode kann nur dank der Anpassungsfhigkeit der Nervenzellen berlebt werden (C). Die Glucoseverwertung wird drastisch eingeschrnkt und das entstehende Energiedefizit durch Ketonkrper gedeckt. Der verminderte Bedarf an Aminosuren fr die Gluconeogenese schont die Speicher an verfgbarem Muskelprotein, so dass auch mehrwchige Hungerperioden berlebt werden knnen. H. K. Biesalski, P. Grimm, Taschenatlas der Ernhrung (9783131153548) c2007 Georg Thieme Verlag KG Energie in Geweben 31 A. Brennstoffreserven eines 70kg schweren Mannes [kcal] Blut Leber Gehirn Glucose oder Glykogen 60kcal 390 kcal 8kcal Muskel 1200 kcal Fettgewebe 80 kcal Triglyceride 45kcal 450 kcal 0kcal 450 kcal 135 000 kcal mobilisierbare Proteine 0 kcal 390 kcal 0kcal 24 000 kcal 37 kcal B. Brennstoffumsatz in der Leber bei längerem Hungern von Muskulatur Protein 1 Harnstoff wird zu den Nieren transportiert und mit dem Urin ausgeschieden Glucose gelangt mit dem Blut zum Gehirn 2 4 5 8 Glucose Fettsäuren Ketonkörper Aminosäuren Proteinabbau liefert glucogene Aminosäuren. aus dem Fettgewebe kommende Fettsäuren werden als Brennstoffe zu Acetyl-CoA oxidiert P Anhäufung von Acetyl-CoA begünstigt die Synthese von Ketonkörpern. Glucose-6phosphat Harnstoff Acetyl-CoA Phosphoenolpyruvat NHe 6 Zwischenprodukte des Citrat-Zyklus werden der Gluconeogenese zugeführt Leberzelle Ketonkörper gelangen mit dem Blut zum Gehirn, wo sie als Brennstoff dienen Acetoacetyl-CoA 7 Wegen Mangels an Oxalacetat gelangt Acetyl-CoA nicht in den Citrat-Zyklus; Acetyl-CoA sammelt sich an 3 Oxalacetat CitratZyklus Citrat C. Brennstoffmetabolismus im Hungerzustand Glucoseproduktion durch Gluconeogenese 100 100 80 75 40 Ketonkörper 5 150 Plasmaspiegel [mM] Brennstoffnutzung im Gehirn 4 Glucose 3 2 Fettsäuren 50 1 20 Glucose Ketonkörper Abbau von GlucoseproMuskelprotein duktion Leber 0 1 2 3 4 Fastentage 5 6 7 3.Tag 40.Tag gebildete oder verbrauchte Menge [g] in 24 Stunden H. K. Biesalski, P. Grimm, Taschenatlas der Ernhrung (9783131153548) c2007 Georg Thieme Verlag KG 32 Energiehaushalt Kontrolle des Energiehaushalts Die Produktion des energiereichen ATP ist grundstzlich an dessen Spaltung zu ADP gekoppelt (A). Die Notwendigkeit dieser Anpassung ergibt sich aus der Tatsache, dass nur wenige Gramm ATP, ADP bzw. AMP im menschlichen Organismus zur Verfgung stehen. Um den tglichen Energiebedarf von ca. 2000 kcal zu decken, bedarf es aber ca. 80 kg an freien Adenin-Nucleotiden. Um dies zu gewhrleisten, muss jedes ADP-Molekl tglich mehrere tausend Mal – unter strenger Kontrolle – phosphoryliert und wieder dephosphoryliert werden. Die Regulation dieser Prozesse geschieht ber eine Vielzahl ineinandergreifender Mechanismen. Als Stellgrße kann die Energieladung einer Zelle betrachtet werden: [ATP] + 12 [ADP] Energieladung = [ATP]+ [ADP]+ [AMP] Sie kann einen Wert zwischen 0 (nur AMP) und 1 (nur ATP) annehmen. ATP erzeugende Stoffwechselwege werden durch eine hohe Energieladung gehemmt, ATP-verbrauchende dagegen werden angeregt. Die Energieladung einer Zelle ist, hnlich wie der pH-Wert, gepuffert: Sie liegt meist zwischen 0,80 und 0,95. Die Regulation der am Energiestoffwechsel beteiligten Enzyme kann durch verschiedene Mechanismen erfolgen. So kann z. B. die Enzymmenge ber die Genexpression im Zellkern gesteuert werden. Bei einigen Enzymen wird auch deren Abbaugeschwindigkeit kontrolliert. Von besonderer Bedeutung ist die Kontrolle der Enzymaktivitt durch reversible Modifikation. Z. B. wird die Glykogenphosphorylase, die den Abbau von Glykogen katalysiert, im Glucosemangel durch Phosphorylierung eines bestimmten Serinrestes aktiviert. Die – theoretische – Kopplung ATP-verbrauchender an ATP-produzierende Prozesse kann jedoch auch teilweise außer Kraft gesetzt werden (B). Verbraucht die Zelle ATP, so steht in den Mitochondrien auch kein ADP zur Verfgung. Dadurch wird die ATP-Synthese gehemmt, so dass sich NADH anhuft. Das entstehende hohe NADH/NAD+-Verhltnis hemmt schließlich den Citrat-Zyklus, so dass die Energieproduktion zum Erliegen kommt. In der Praxis kann jedoch die Oxidation von der Phosphorylierung entkoppelt werden: Der Elektronentransport in der Atmungskette luft unvermindert ab, wobei statt ATP nun Wrme produziert wird. Die Phosphofructokinase unterliegt ebenfalls einer allosterischen Hemmung durch ATP (C). Fr die katalysierte Reaktion wird ATP verbraucht. Die Fructose1,6-Biphosphatase katalysiert die Rckreaktion, ohne dafr jedoch ATP zu bentigen. Laufen beide Reaktionen ab, so wird in der Summe ATP „verschwendet“. Derartige „nutzlose“ Substratzyklen bezeichnet man als Leerzyklen, im Englischen als Futile Cycles. Die individuellen Unterschiede in der Aktivitt solcher Zyklen (Fetthydrolyse und Reveresterung u. a.) knnen pro Tag bis zu 500 kcal ausmachen. Sie bilden eine biochemische Grundlage fr die unterschiedliche Nhrstoffausnutzung und damit fr das variierende Krpergewicht bei gleichen Ernhrungsgewohnheiten und ußeren Bedingungen. H. K. Biesalski, P. Grimm, Taschenatlas der Ernhrung (9783131153548) c2007 Georg Thieme Verlag KG Kontrolle des Energiehaushalts 33 A. Energieliefernde und -verbrauchende Prozesse Energieträger (endogen, exogen) Körpersubstanz Arbeit COw NHe energieliefernde Prozesse energieverbrauchende Prozesse H Ow ADP ATP Vorstufen HwO B. Gekoppelte/entkoppelte Energieproduktion vorbereitende Reaktionen gemeinsame Endstrecke biologische Oxidation Fettsäuren entkoppelt COw Wärme ATP CitratZyklus Kohlenhydrate NADH áOw Atmungskette ATP HwO NAD + ATP ADP Aminosäuren ATP P Harnstoff gekoppelt C. Futile Cycle ATP ADP PFK Eq P P Glykogen Fructose-6phosphat Fructosebisphosphat Lactat P P Ausgangsverbindung Endverbindung P Ew H wO FBPase H. K. Biesalski, P. Grimm, Taschenatlas der Ernhrung (9783131153548) c2007 Georg Thieme Verlag KG