Fossilien dokumentieren die Makroevolution
Werbung

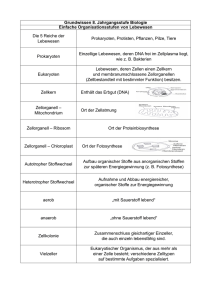
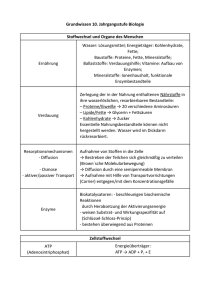
Kapitel 23: Auf den Spuren der Phylogenie: Makroevolution, Fossilbelege und Systematik (englische Version: Kap. 25) Makroevolution ist die Entstehung taxonomischer 1 Gruppen auf höherer Ebene als auf der Stufe der Arten. Aus der Makroevolution resultieren neue Gattungen, neue Familien oder sogar neue Stämme. Reptilien und Säugetiere sind beispielsweise aus der Sicht der Makroevolution miteinander verwandt, da letztere aus den Reptilien hervorgegangen sind. Makroevolution lässt sich durch Fossilbefunde belegen. Fossilien dokumentieren die Makroevolution Wie sich Fossilien bilden Sedimentgesteine enthalten am meisten Fossilien. Sedimente gelangen durch Flüsse in Meere und Moore und lagern sich Schicht für Schicht aufeinander ab. Durch den steigenden Druck entstehen im Laufe der Zeit Gesteine. Wasserlebewesen und Landtiere werden nach dem Tod auch sedimentiert, aber nur ein Bruchteil davon bleibt als Fossil erhalten. In Sedimenten eingelagerte tote Tiere werden schnell zersetzt. Nur die harten, mineralhaltigen Bestandteile bleiben erhalten. Das sind in der Regel Knochen, Zähne, Schalen und Panzer. Unter den richtigen Bedingungen wird das organische Material durch Mineralien ersetzt, indem das Grundwasser diese Stoffe in das tote Gewebe einsickern lässt. In ganz seltenen Fällen werden sogar organische Gewebe ausgegraben, die z. T. praktisch intakt geblieben sind, wie z. B. Blätter von Pflanzen. Diese Exemplare konnten kloniert und sequenziert, deren organische Zusammensetzung analysiert und die Ultrastruktur ihrer Zellen mit dem Elektronenmikroskop erforscht werden. Es können in den Gesteinen auch Abdrücke der Lebewesen entstehen, indem in den Sedimentablagerungen sich befindende Tiere zersetzt werden und eine Art Gussform hinterlassen, die von Mineralsalzen gefüllt wird, die sich kristallisieren. Es gibt auch Funde von fossilisierten Abdrücke wie Fuss- 1 Taxonomie: Der Bereich der Biologie, der sich mit der Benennung und Klassifizierung der verschiedenen Le- bensformen befasst. 1 spuren, Tierbauten etc., die als Spurenfossilien bezeichnet werden. Spurenfossilien verraten viel über das Verhalten der betreffenden Tiere. Die Erhaltung von Fossilien ist nicht nur in Sedimentgesteinen möglich, sondern auch an Orten, wo Destruenten (Bakterien, Pilze, ...) nicht vorkommen oder behindert werden. So bleibt der Kadaver samt Weichteile erhalten oder wird sehr langsam zersetzt, wie z. B. in sauren Mooren. Ein anderes Beispiel sind Insekte, die im Harz von Bäumen eingefangen bleiben oder eingefrorene Tiere. Diese Art von Fossilien sind jedoch auch sehr selten. Grenzen der Fossilzeugnisse Der Fund eines Fossils ist der Höhepunkt von einer Reihe von aufeinander abgestimmten Zufällen. Erstens muss das Tier am richtigen Ort zur richtigen Zeit sterben, damit durch Sedimentation unter günstigen Bedingungen fossilisieren kann. Zweitens muss es viele geologische Prozesse entgehen, wie z. B. Erosion, Entwicklung von viel zu hohen Druck, Gesteinsschmelze, etc. Drittens muss das Fossil an die Oberfläche – beispielsweise durch die Vertiefung eines Flussbetts, Steinbrüche, Klippenbrüche, etc. – gelangen. Abgesehen davon sind die meisten Lebewesen der Frühzeit nicht erhalten geblieben, nur die meist verbreiteten Spezies, die Schalen oder harte Skelette besassen. Der Fossilnachweis stellt also nur ein Bruchstück der früheren Tierwelt dar. Nichtsdestotrotz sind Fossilbelege eine bemerkenswert detaillierte Dokumentation der Makroevolution über einen immensen geologischen Zeitraum. Paläontologen verfügen über eine Vielzahl von Methoden, um Fossilien zu datieren Relative Datierung Jüngere Sedimente lagern sich auf ältere ab. Jede Schicht entsteht in einem bestimmten Zeitraum und deshalb verraten diese das Zeitalter, in dem die fossilisierten Lebewesen eingelagert werden. Die Sedimentierung ist von Umweltfaktoren abhängig, sodass durch Vergleiche Schwankungen in der Sedimentationsrate und Art der Sedimentpartikel im Laufe der Zeit feststellen kann. Zum Teil können ganze Schichten fehlen, weil der betreffende Bereich während des betreffenden Zeitraumes oberhalb des Meeresspiegels lag oder durch Erosion die Schichten abgetragen wurden. Oft ist die Bestimmung einer Gesteinsschicht durch das Vorhandensein von ähnlichen Fossilien, sog. Leitfossilien. 2 Durch Erforschung vieler unterschiedlicher Fundorte haben Wissenschafter eine geologische Skala erstellen können, die in vier verschiedenen Erdzeitalter gegliedert ist: das Präkambium, das Paläozoikum, das Mesozoikum und das Känozoikum. Der Wechsel von einem Erdzeitalter zum nächsten ist bestimmt durch Massenaussterben, explosionshafte Entstehung vieler neuer Lebensformen und extreme geologische Veränderungen. Dies lässt sich anhand der Erforschung der Gesteinsschichten und des Fossilnachweises feststellen. Jedes Zeitalter ist in Perioden und letztere wiederum in Epochen unterteilt. Die Schichtfolge von Sedimenten entspricht dem relativen Alter darin enthaltener Fossilien; sie teilt uns mit, in welcher Reihenfolge die fossilen Artengruppen evolvierten. Doch die Abfolgen von Sedimentgesteinen sagen nichts über das absolute Alter der eingebetteten Fossilien aus. Absolute Datierung Bei der absoluten Datierung wird das Alter in Jahren angegeben. Die radiometrische Datierung ist die gängigste Art der Altersbestimmung. Dabei wird die Anzahl noch vorhandener Isotopen gemessen, um das Alter zu bestimmen. Isotopen ermöglichen die Altersbestimmung, weil diese eine bestimmte Zerfallsrate haben. Die C-14-Methode eignet sich bspw. gut für die Altersbestimmung von jungen Fossilien, da C-14 eine Halbwertszeit von 5'600 Jahren aufweist. Für Fossilien, die älter als 50'000 Jahre alt sind verwendet man Isotope mit längeren Halbwertszeiten, wie z. B. Uran-238 mit einer Halbwertszeit von 4,5 Milliarden Jahren. Eine andere Methode der Altersdatierung ist die Messung der L-Aminosäuren. Bekanntlich synthetisieren Lebewesen nur L-Aminosäuren (linksdrehende Form). Nach dem Tod werden L-Aminosäuren in D-Aminosäuren umgewandelt. Da die Umwandlungsgeschwindigkeit (= Racemisierung) bekannt ist, ist eine Datierung möglich. Die Racemisierung ist aber temperaturabhängig, was bedeutet, das durch Klimaänderung die Umwandlungsgeschwindigkeit beeinflusst wird. Viele evolutionäre Neuerungen sind abgewandelte Versionen älterer Strukturen Wie evolvieren ganz neuartige Körperkonstruktionen? Ein Mechanismus ist die graduelle Verbesserung existierender Strukturen, wodurch diese neue Funktionen erfüllen können. Die Funktionen von den meisten biologischen Strukturen, wie z. B. die leichte, poröse Knochenstruktur von Vögeln, um das Fliegen zu erleichtern, war ursprünglich eine andere. Diese Funktionen wurden aber durch die Lebensumstände für einen neuartigen Gebrauch umfunktioniert, was durch natürliche Selektion über 3 eine Reihe von Zwischenstadien erfolgte. Bei den zweibeinigen Reptilienvorfahren der Vögel war z. B. dieser poröser Knochenbau ideal für eine flinke und schnelle Gangart. Möglicherweise dienten die flügelartigen Vordergliedmassen, bevor sie zum Fliegen eingesetzt wurden, zum Fangen von Insekten und anderen kleinen Beutetieren. Diese Umfunktionierung von biologischen Strukturen wird als Präadaptation bezeichnet. Gene, welche die Entwicklung steuern, spielen eine wichtige Rolle bei evolutionären Neuerungen In manchen Fällen verursachen geringfügige genetische Abweichungen grosse Unterschiede zwischen Lebewesen, wie z. B. zwischen Mensch und Schimpanse. Allometrisches Wachstum ist ein Beispiel dafür. Es führt zu unterschiedlicher Wachstumsrate verschiedener Körperteile und trägt auf diese Weise dazu bei, ein Lebewesen zu formen. Verändern sich die Wachstumsraten nur geringfügig, führt das zu grossen Unterschieden. Genetische Veränderungen können ausserdem die Reihenfolge, in der verschiedenen Körperteile ihre Entwicklung beginnen und beenden ändern. Neotenie ist eine wichtige Konsequenz davon. Dabei handelt es sich um Lebewesen, die als erwachsene, geschlechtsreife Organismen Merkmale erhalten, die bei ihren Vorfahren Juvenilmerkmale waren. Abgesehen von Neotonie und allometrische Unterschiede ist die Gehirngrösse ein weiterer, bedeutender Faktor, der Menschen von Schimpansen unterscheidet. Das viel grössere Gehirn bei Menschen ist auf ein verlängertes Wachstum dieses Organs zurückzuführen. Solche evolutionäre Veränderungen in der zeitlichen Abstimmung oder Geschwindigkeit der Entwicklung werden als Heterochronie bezeichnet. Räumliche Veränderungen in der Anordnung von Körperteilen werden als Homöose bezeichnet. In Fossilien Reihen erkennbare Trends bedeuten nicht, dass die Makroevolution zielgerichtet verläuft Das heutige Pferd stammt von einer viel kleineren Art, die aus dem Eozän stammt. Da diese Art (Hyracotherium) sich von Buschblättern anstatt von Gräsern ernährte, war ihr Gebiss anders gebaut. Ausserdem besass diese vier Zehen statt nur einen, wie es bei den modernen Pferden der Fall ist. Aus dem Hyracotherium evolvierten durch adaptive Radiation zahlreiche andere Arten, die jedoch 4 nicht alle aneinandergereiht werden können, da es im Widerspruch zur Theorie der zusammenhängenden, fortschreitenden Evolution stehen würde. Es handelt sich also in diesem Fall um einen weit verzweigten Stammbaum nur eine bestimmte Linie davon führt zum heutigen Pferd. Alle andere sind ausgestorben. Wenn bei einer Art der evolutionäre Trend nicht ersichtlich ist, weil bspw. die neu entstandenen Arten genauso oft kleiner wie grösser als ihre Stammart sind, dann entscheiden die Merkmale derjenigen Art mit der längsten Existenzdauer und der grössten adaptiven Radiation. Hier ist also von Artenselektion die Rede, da nur diejenige Art die Trendrichtung bestimmt, die sich am besten durchsetzt, indem es durch gute Anpassung am meisten „Nachkommen“ erzeugt. Die Makroevolution hat eine biogeographische Triebfeder in der Kontinentalverschiebung Die kontinentale Plattenverschiebung ist die eigentliche Ursache für die Verteilung des Lebens und für den Verlauf der Makroevolution, die durch Massenaussterben und explosionsartige Entstehung von vielen neuen Lebensformen. Das Driften der Kontinente verursachte ständige geographische Veränderungen, aber zwei Episoden der Kontinentalverschiebung waren evolutionär gesehen entscheidend für den Verlauf der Makroevolution. Ein markantes Ereignis war sicher die Vereinigung aller Kontinente zu einer riesigen Landmasse namens Pangea (= alles Land). Da durch diese Vereinigung viele Küstenregionen verschwanden, entstand ein trockenes unausgeglichenes Klima, das für die Landmasse kennzeichnend war. Die Meeresbecken vertieften sich, sodass die flachen Meeresregionen, wo die meisten Arten vorkamen, austrockneten, was zu einem Massenaussterben führte. Durch die neuen klimatischen und biologischen Bedingungen kam es auch am Land zu einem Massenaussterben, was zur Neugestaltung des Lebens führte. Auch entscheidend für das Massenaussterben waren die massiven Vulkanausbrüche zu dieser Zeit. Der atmosphärische Staub von den Ausbrüchen hielt genügend Sonnenlicht zurück, um die Temperatur prägnant zu senken. Das zweite einschneidende Ereignis im Laufe der Makroevolution war das Auseinanderdriften der Kontinente, was zu einer Diversifikation des Lebens in den so neu entstandenen Kontinenten führte, da diese nun evolutionär voneinander unabhängig waren. Diese Theorie der Kontinentaltrennung erklärt die verblüffende Ähnlichkeit von Fossilien, die in unterschiedlichen Orten ausgegraben wurden und die heutige Verbreitung der Lebewesen. 5 Die Geschichte des Lebens ist geprägt durch Massenaussterben gefolgt von adaptiver Radiation der überlebenden Arten Die Fossilfunde deuten darauf hin, dass die Evolution nicht glatt verlief, sondern geprägt war von Massenaussterben gefolgt von explosionsartigen adaptiven Radiationen. Die Phasen dazwischen waren ziemlich ruhig. → Punktualismus Viele taxonomische Gruppen haben früh in ihrer Geschichte eine reiche Vielfalt hervorgebracht und zwar nach der Evolution einiger neuartiger Merkmale, die ihnen eine neue Adaptationszone zugänglich machte. Es ist aber meistens nicht möglich, dass neuentwickelte Tierarten in bereits besetzten Adaptationszonen ihren Nutzen daraus ziehen können. Die adaptive Radiation und die Besetzung von neuen Adaptationszonen bei den Säugetieren sind möglicherweise erst nach dem Aussterben der Dinosaurier zu Stande gekommen. Eine Art kann aussterben, weil ihr Lebensraum zerstört wurde oder sich in eine für sie ungünstige Richtung veränderte. Nicht nur physikalische Faktoren, wie z. B. Temperaturveränderungen, sondern auch biologische können für das Aussterben einer Art verantwortlich sein. Bspw. das Mutieren einer Art kann sich negativ auf eine andere auswirken. Der deutliche Floren- und Faunenschnitt am Ende der Kreidezeit, in dem mehr als die Hälfte aller marinen Arten und zahlreiche Familien landlebender Pflanzen und Tiere zum Verhängnis wurde, wird von vielen Wissenschafter auf die Einschlaghypothese gestützt. Diese Hypothese besagt, dass ein Asteroid von mehreren Kilometern Durchmesser eine Staubwolke erzeugte, welche die Sonne weltweit verdunkelte. Das führte zu sauren Niederschlägen, Temperaturabstiege und zu einer massiven Verminderung der Photosynthesebetreibung der Pflanzen, was der Kollaps vieler Ökosysteme verursachte. Beweise für diese Hypothese liefert der entdeckte Riesenkrater Chicxulub, eine 65 Mio. Jahre alte Narbe unterhalb der Küstensedimente der mexikanischen Halbinsel Yucatán. Angeblich soll der Krater einen Durchmesser von etwa 300 km haben. Ein weiterer Beweis dafür ist die zwischen mäsozoischen und känozoischen Erdschichten sich befindende Tonschicht, die reich an Iridium ist. Iridium ist ein sehr seltenes Element auf der Erde. Gegner dieser Hypothese sind der Meinung, dass für ein derartiges Massenaussterben ein Meteoriteneinschlag nicht unbedingt ausschlaggebend war. Sie behaupten, dass irdische Prozesse wie Vulkanismus, Klimaveränderungen wegen der Kontinentaldrift völlig ausreichend sind. Ein wichtiger Punkt für die Aufklärung dieses Ereignisses ist die Geschwindigkeit, mit der sich das Verschwinden der Arten ereignete. Ein schnelles Verschwinden spricht für den Meteoriteneinschlag, während einem allmählichen Verschwinden eher irdische Ursachen zugrunde liegen könnten. Wie dem auch sei, diejenigen Arten, die solche Umweltbedingungen standhalten können, sind die Basis für die Entwicklung von neuen Radiationen, die viele der freigewordenen Adaptivzonen wieder besetzen können. 6 Die Systematik verbindet die biologische Mannigfaltigkeit zu einem phylogenetischen System Die Entwicklungsgeschichte einer Art oder einer Gruppe verwandter Arten wird als Phylogenie bezeichnet. Die Phylogenie einer Gruppe wird traditionell als Stammbaum dargestellt. Taxonomie und hierarchische Klassifikation Linné entwickelte im 18. Jhd. ein System der Taxonomie, in dem jede Art einen zweiteiligen Namen erhielt (Binomen). Dieses System bezeichnet man als binäre Nomenklatur, die aus dem Namen der Gattung und dem Artennamen besteht. Eine Gattung kann aus mehreren verwandten Arten zusammengesetzt sein. Mit steigender Hierarchie wird die Klassifizierung allgemeiner: verwandte Gattungen gehören zur selben Familie, Familiengruppen wurden in Ordnungen eingeteilt, diese wiederum in Klassen, Klassen in Stämme und schliesslich Stämme in Reiche (s. Anhang 3!). Die Taxonomie verfolgt zwei Ziele: erstens nahe verwandte Lebewesen auseinanderzuhalten und sie in Arten einzuteilen sowie zweitens die Differenzierungsmerkmale zu beschreiben. Dazu gehört – gemäss dem Linnéschen Prinzip – die Benennung neu entdeckter Arten. Es gibt bei der Klassifizierung auch Zwischenkategorien wie Unter- und Überfamilien. Eine taxonomische Kategorie bezeichnet man kurz als Taxon (Plural: Taxa!). Die Beziehung zwischen Klassifikation und Phylogenie Abgesehen von der Taxonomie beinhaltet die Systematik auch die evolutionären Beziehungen zwischen den Arten. Dafür werden Artstammbäume erstellt, welche die Phylogenie von Arten darstellt. Dabei unterscheidet man zwischen monophyletischen, polyphyletischen und paraphyletischen evolutionären Beziehungen. monophyletisches Taxon: Alle Arten des Taxons stammen vom gleichen Vorfahren, der keine in einem anderen Taxon vorkommende Spezies hervorbrachte (s. Abb. 23.12, Taxon 1!). polyphyletisches Taxon: Mitglieder des Taxons stammen von zwei oder mehr Vorfahren, die nicht allen gemeinsam sind (s. Abb. 23.12, Taxon 2!). paraphyletisches Taxon: gemeinsamer Vorfahren, der aber nicht nur den in diesem Taxon enthaltenen Arten gemeinsam ist (s. Abb. 23.12, Taxon 3!) Ideal wäre ein monophyletisches Taxon, weil es die Entwicklungsgeschichte der Lebewesen genau klassifiziert. 7 Die Unterscheidung von Homologie und Analogie Die Systematik ist grösstenteils eine vergleichende Wissenschaft. Aufgrund morphologischer Vergleiche zwischen Lebewesen schliesst sie auf den bestehenden Verwandtschaftsgrad. Ähnlichkeit infolge gemeinsamer Abstammung wird als Homologie bezeichnet. Doch nicht alle morphologisch ähnliche Lebewesen müssen einen gemeinsamen Vorfahren haben. Es kann auch sein, dass Lebewesen ähnlich evolvieren, weil sie ählichen oder gleichen Umweltbedingungen ausgesetzt sind und somit durch natürliche Selektion analoge Anpassungen herausgebildet haben. Dies wird konvergente Evolution genannt. Ähnlichkeit aufgrund von Konvergenz nennt man Analogie. Wenn man die Entwicklungsgeschichte durch Artenvergleiche rekonstruiert, muss man Stammbäume allein auf der Basis von homologen Ähnlichkeiten erstellen. Es gibt drei Kriterien, um Homologien zu erkennen: homologe Organe, d. h. gleiche Lage der Organe ähnliche Embryonalentwicklung Komplexität von Strukturen: Je komplexer eine Struktur ist, desto grösser ist die Wahrscheinlichkeit, dass diese homolog sind. Die Molekularbiologie bietet effiziente neue Hilfsmittel für die Systematik Je weniger Arten stammesgeschichtlich voneinander entfernt sind, desto grösser ist die Übereinstimmung von Aminosäuresequenzen der Proteine. Vergleich von DNA oder RNA Ein Vergleich der Gene oder Genome ist das direkteste Mass einer stammesgeschichtlichen Verwandtschaft. Dafür gibt es drei Methoden: DNA-RNA-Hybridisierung, Restriktionskartierung und DNASequenzierung. DNA-DNA-Hybridisierung: DNA-Einzelstränge zweier Organismen werden vereinigt und dabei achtet man wie gut diese sich verknüpfen können. Je besser die Verknüpfung ist, desto grösser ist der Verwandtschaftsgrad. Damit lässt sich die Ähnlichkeit zweier Genome abschätzen, die genaue Übereinstimmung der Nukleotidabfolge wird dadurch jedoch nicht ermittelt. → Restriktionskartierung Restriktionskartierung: Mit Hilfe von Restriktionsenzymen wird die DNA in Abschnitten gespalten. Diese Fragmente werden durch Elektrophorese aufgetrennt und mit denjenigen, der anderen Art verglichen, die durch dieselben Restriktionsenzyme fragmentiert wurden. Nahe verwandte Arten 8 weisen ähnliche Fragmentmuster auf. Weil man vom Kerngenom zahlreiche Fragmente erhält, eignet sich diese Methode nur für kleine DNA-Abschnitte. Man verwendet häufig die kleine mitochondriale DNA für solche Analysen. Für systematische Zwecke ist die mtDNA zusätzlich von Vorteil, weil diese etwa 10mal schneller mutiert als das Kerngenom, was selbst die Differenzierung von zwei Populationen der gleichen Art zulässt. DNA-Sequenzierung: Das ist die genaueste und effektivste Methode, um die DNA zweier Arten zu vergleichen, denn es gibt die genaue Abfolge der Nukleotide wieder. Durch Vergleich entsprechender DNA-Abschnitte zweier Arten lässt sich genau feststellen wie stark die betreffenden Arten sich im Laufe der Evolution auseinanderentwickelt haben. Da sich die ribosomale RNA (rRNA) im Vergleich zum Rest der DNA sehr langsam ändert, ist deren Analyse sehr effektiv, um die Artbildung bis hin zur Entstehung des Lebens zurückzuverfolgen. Analyse fossilisierter DNA Mit Hilfe der Polymerasekettenreaktion waren die Systematiker in der Lage, die Analyse der Nukleotidsequenz auf DNA-Spuren im fossilen Material auszudehnen. Molekulare Uhren Durch Vergleich von homologen Proteinen verschiedener Taxa mit einem gemeinsamen Vorfahren kann man herausfinden, wann die Verzweigung im Stammbaum erfolgte und wie lange sich die betreffenden Taxa auseinanderentwickelt haben. Dabei werden Unterschiede in der Aminosäurenabfolge untersucht. Allerdings muss man Proteine verwenden deren Evolutionsrate pro Zeiteinheit konstant ist, wie es bspw. bei Cytochrom c der Fall ist, um aussagekräftige Resultate zu erhalten. DNA-Vergleiche sind als Uhr zum Datieren von Verzweigungspunkten in Stammbäumen sogar zuverlässiger. Molekulare Uhren werden geeicht, indem man die Anzahl der Aminosäure- oder Nukleotidunterschiede für eine Reihe von evolutionären Verzweigungspunkten, die durch Fossilfunde gut dokumentiert sind, gegen die Zeit aufträgt. Diese Art und Weise der Datierung funktioniert nur, wenn die Mutationsrate der betrachteten Gene bzw. Proteine konstant ist. Dies ist bei nahe verwandten Arten der Fall, jedoch nicht bei weiter entfernten Spezies. Freilich bedeutet eine konstante Mutationsrate, dass ein beträchtlicher Anteil der DNAVeränderungen auf neutrale Mutationen zurückzuführen ist, die das gesamte Genom schliesslich mehr verändern als natürliche Selektion. Das wird von vielen Biologen in Frage gestellt somit auch die Zuverlässigkeit der absoluten Datierung durch molekulare Uhren. Die Zuverlässigkeit einer relativen Datierung wird allerdings nicht angefochten. Punktuell starke Veränderungen von Proteinen durch Funktionswechsel können die absolute Zeitskala auch verfälschen. Deshalb müssen für eine genaue phylogenetische Rekonstruktion abgesehen von molekularen Daten alle anderen möglichen Hinweise (Morphologie, Fossilnachweise, ...) berücksichtigt werden. 9 Die Kladistik ist eine phylogenetische Systematik, die nur monophyletische Taxa zulässt Es gibt zwei Merkmale für die Erstellung eines Stammbaumes. Eines wäre der Punkt, wo die Verzweigungen auseinandergehen, d. h. der Zeitpunkt (relativ) der Entstehung eines neuen Taxons. Das zweite Merkmal wäre das Ausmass der Divergenz zwischen den Zweigen, was aufzeigt, wie stark sich zwei Taxa unterscheiden, seit sie von einem gemeinsamen Vorfahren abzweigten. Je nachdem, was für ein Merkmal stärker gewichtet wird, kann man zwischen drei verschiedenen Systematiktypen unterscheiden: Die Phänetik, die Kladistik und die klassische Evolutionssystematik. Phänetik (numerische Taxonomie) Die Einordnung der Taxa erfolgt ausschliesslich aufgrund messbarer Daten (darum numerische Taxonomie). Dabei werden so viele anatomische Eigenschaften wie möglich miteinander verglichen. Die Phänetik hat wenige Befürworter, weil man der Auffassung ist, dass Ähnlichkeit kein Indiz für Verwandtschaft ist. Kladistik In der Kladistik stellt man phylogenetische Beziehungen als Kladogramme dar. Kladen sind Evolutionszweige. In Kladogramme ordnet man die Taxa in der Reihenfolge deren Entstehung ein. Taxa mit dem selben Ursprung haben den gleichen Vorfahren, und somit weisen diese Homologien auf. Die Art und Weise der Phylogenie-Darstellung berücksichtigt jedoch nicht den evolutionären Divergenzgrad der Taxa. Ein Kladogramm ist aufgebaut aus Arten die sowohl ursprünglichen (plesiomorphen) als auch abgeleiteten (apomorphen) Merkmale aufweisen. Da jeder neue Punkt auf dem Kladogramm, von dem Abzweige ausgehen, eine neuartige Synapomorphie (abgeleitete Homologie) darstellt, die nur für die nächst kommenden Arten charakteristisch ist, müssen zuerst diejenigen Arten aufs Kladogramm eingetragen werden, die plesiomorphe Merkmale aufweisen, d. h. die allen Arten oberhalb gemeinsam haben (s. Abb. 23.15!). Klassische evolutionäre Systematik Die klassische evolutionäre Systematik versucht das Ausmass der evolutionären Divergenz und die Reihenfolge der Verzweigungen zu berücksichtigen. Dort, wo es nicht möglich ist, wird subjektiv entschieden, welches Kriterium wichtiger ist. 10 Ist eine neue Synthetische Theorie der Evolution notwendig? Durch die Verwendung evolutionärer Kriterien wird das Darwinsche Konzept befolgt, nämlich, dass alle Arten einen gemeinsamen Vorfahren haben, von welchem sie sich durch kontinuierliche Mutationen im Laufe der Evolution auseinanderentwickelt haben. In der modernen Wissenschaft werden mehrere Disziplinen zu einem systematischen Konzept, der Synthetischen Evolutionstheorie, vereinigt. Auch diese moderne Theorie ist eine Bekräftigung der Darwinschen Theorie. Diese Synthetische Theorie kommt zum Schluss, dass evolutionäre Veränderungen über eine riesige Zeitspanne resultieren. Aus einer Reihe von Mikroevolutionen (Änderung der Allelfrequenzen in Populationen) resultiert also die Makroevolution. Neu an der Synthetischen Theorie ist, dass neue Arten nicht nur infolge natürlicher Selektion, sondern auch infolge genetischer Drift rasch und nichtadaptiv entstehen können. Die Hauptschwerpunkte der Synthetischen Evolutionstheorie sind natürliche Selektion und Gradualismus. Wie schon erwähnt, gibt es Verfechter der Gradualismus-Theorie, weil sie der Meinung sind, dass viele in den Fossilreihen beobachtbare Übergänge nicht durch ein Hochrechnen von mikroevolutionären Ereignissen erklärbar sind. Es handelt sich dabei oft um punktuelle Änderungen. Nun kontern die Befürworter mit dem Argument, dass der Fossilnachweis unvollständig ist und dass es ausserdem eine Definitionssache ist, ob etwas als punktuell oder graduell bezeichnet wird. Einige Wissenschafter bevorzugen eine hierarchische Theorie, in der auf den verschiedenen Evolutionsstufen unterschiedliche Mechanismen zum Tragen kommen. Somit verliert natürliche Selektion an Bedeutung. Andere sind der Auffassung, dass die Evolution stark von Zufallsereignissen beeinflusst wird, wie z. B. Kontinentaldrift oder Massenaussterben. Sie sind also Befürworter des sog. historischen Zufalls. Eine weitere Theorie, die Darwinismus strikt ablehnt, ist die Frankfurter Evolutionstheorie. Diese Theorie geht davon aus, dass Individuen sich aktiv verändern im Gegensatz zur Darwinschen Theorie, wo Organismen passiv durch Umweltveränderungen evolvieren. Die Evolution wird also gemäss der Frankfurter Evolutionstheorie durch die Organismen selbst bestimmt, indem sie aus sich selbst heraus evolvieren und sich die Umwelt aktiv erschliessen. Nun drängt sich die Frage auf, ob die Frankfurter Evolutionstheorie eine Darstellung desselben Phänomens ist, das einfach von einer anderen Perspektive betrachtet wird. Während sich die Synthetische Theorie auf die Wechselwirkungen zwischen Organismus und Umwelt konzentriert, befasst sich die Frankfurter Evolutionstheorie mit den Wechselwirkungen zwischen Organismus und seinen Substrukturen. So gesehen könnte diese neue Theorie die Synthetische Evolutionstheorie ergänzen, vorausgesetzt, dass diese sich als wahr erweist. Vielleicht wird sich die Synthetische Theorie zukünftig ein paar „Schönheitsoperationen“ unterziehen müssen; dass heisst, dass man die relative Bedeutung der einzelnen Evolutionsmechanismen eventuell anders gewichten und z. T. durch neue Theorien ergänzen wird. Auf jeden Fall wird sich die Evolutionsbiologie weiterentwickeln und auf diesem Weg sind neu erscheinende Theorien wie die 11 Frankfurter Evolutionstheorie wertvolle Denkansätze, um die Entwicklung und Perfektionierung voranzutreiben. 12