6. Graduiertenforum der Fachgruppe Morphologie
Werbung

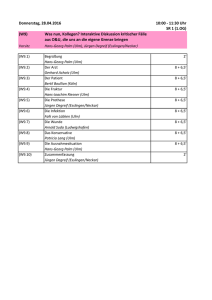
6. Graduiertenforum der Fachgruppe Morphologie der Deutschen Zoologischen Gesellschaft 10.-13. Oktober 2013 Universität Ulm 6. Graduiertenforum Morphologie 2013 Ulm Sponsoren 3 6. Graduiertenforum Morphologie 2013 Ulm 4 6. Graduiertenforum Morphologie 2013 Ulm 3 6 8 10 12 Sponsoren Willkommen Allgmeine Informationen Programmübersicht Vorträge 30 Posterbeiträge 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 31 31 32 33 34 37 41 42 43 44 Inhaltsverzeichnis Philipp Schütz, Verena Kutschera, Andreas Maas & Dieter Waloßek Das Grafikprogramm Blender und dessen Möglichkeiten zur Darstellung morphologischer Rekonstruktionen Thomas van de Kamp & Tilo Baumbach Vom Präparat zum Kunststoffmodell: 3D-Druck für Morphologen Paul Walther Novel electron microscopic methods for 3D imaging of subcellular structures Peter Rühr, Alexander Blanke & Bernhard Misof Die cephale Morpholgie von Tomocerus flavescens (Hexapoda: Collembola): Eine 3D-Rekonstruktion Thomas Schwaha Eine neue Süsswasser-Kamptozoe? Verena Kutschera, Andreas Maas & Dieter Waloßek Proximale Sklerite an Pleopoden von Malacostraca (Crustacea s. l.) Christina Nagler, Renate Matzke-Karasz & Juergen Geist A revision of the genus Tanycypris (Ostracoda, Cypricercinae) with the description of a new species and a dichoto- mous key of the genus. S. Kruppert, M. Rabus,L. Weiss M. Horstmann, C. Laforsch & R. Tollrian Geometrische Morphometrie von Wasserflöhen Marie K. Hörnig, Carolin Haug, Andy Sombke, Steffen Harzsch & Joachim T. Haug Evolution und erdgeschichtliches Auftreten von Brutpflegeverhalten innerhalb der dictyopteren Insekten Ute Eulitz Die Ootheken der Gottesanbeterinnen (Mantodea) Joachim T. Haug „Palaeo-Evo-Devo von Arthropoden – Was uns Fossilien über die Evolution von Entwicklungsmustern sagen können“ Alexandra Kerbl Kleiner Wurm ganz groß – Die Entwicklung des Nervensystem des interstitiellen „Polychaeten“ Dinophilus gyrociliatus (Annelida, Dinophilidae) Leonie Hannig, Simon Schwarz & Heinz-R. Köhler Die Streptoneurie bei Paradiesschnecken setzt keine ontogenetische Torsion voraus Mayuran Sanmugaraja & Rico Schellhorn Habitatbestimmung bei Katzen anhand der Unterarmknochen Marlene Nowak & Dr. Dorota Konietzko-Meier Eine neue Rekonstruktion des Schultergürtels und der Vorderextremität von Metoposaurus diagnosticus krasiejo- wensis Vivien Jaquier & Torsten Scheyer Die Halswirbelsäule des Giraffenhalssauriers: eine knochenhistologische Untersuchung Torsten Scheyer Integration morphologischer, knochenhistologischer und embryologischer Datensätze - drei paläobiologische Fallbei spiele (Schildkröten, Krokodile und marine Reptilien aus der Trias-Zeit) Stephan Handschuh, Hans L Nemeschkal, Brian D Metscher, Philipp Mitteröcker & Gerd B Müller Die Rollen von post-embryonalem Wachstum und sexueller Selektion in der Evolution sexual-dimorpher Extremetitä- ten in Amphipoden Verena Kutschera, Joachim T. Haug, Wafaa S. Sallam, Andreas Maas, Dieter Waloszek & Carolin Haug Tagmatisierung bei Fangschreckenkrebsen (Stomatopoda, Malacostraca, Crustacea) Tobias Reich Die Ventralseite des Peraeons bodenlebender Zehnfußkrebse (Decapoda: Reptantia) und seine Signifikanz für phylo- genetische Deutungen Peter Rühr, Alexander Blanke & Bernhard Misof Die cephale Morpholgie von Tomocerus flavescens (Hexapoda: Collembola): Eine 3D-Rekonstruktion Teilnehmerliste Übersichtsplan Universität Übersichtsplan Stadt Wichtige Adressen Empfohlene Busverbindungen 5 6. Graduiertenforum Morphologie 2013 Ulm Willkommen Liebe Teilnehmer, wir möchten Euch ganz herzlich zum 6. Graduiertenforum der Fachgruppe Morphologie der Deutschen Zoologischen Gesellschaft in Ulm willkommen heißen. Jedes Kind lernt im Laufe seines Lebens den Spruch „In Ulm und um Ulm und um Ulm herum“, doch dass Ulm mehr als diesen Vers und sein Münster zu bieten hat wollen wir mit dieser Tagung unter Beweis stellen. Wir freuen uns dieses Jahr wieder ein breites Teilnehmerfeld aus dem gesamten deutschsprachigen Raum (und darüber hinaus) willkommen heißen zu dürfen. Auch wenn der Zuspruch nicht ganz so gut wie letztes Jahr ist hoffen wir dass die familiäre Atmosphäre um so mehr zum diskutieren und ins Gespräch kommen einlädt. Die thematische Bandbreite führt einmal mehr durch das gesamte „Tierreich“ und beweist die breite Anwendbarkeit morphologischer Methoden quer durch alle Taxa. Unsere Idee war es dieses Jahr ein Fachgruppentreffen zum Thema „Lücken schließen – Morphologie fossil & rezent” durchzuführen. Methodisch gibt es viele Ansätze die Morphologie von Tieren darzustellen – mikroskopische, dreidimensionale und viele mehr. Oftmals werden die Ergebnisse morphologischer Untersuchungen phylogenetisch interpretiert. Bedauerlicherweise werden für das Erstellen von Verwandtschaftshypothesen jedoch oft nur rezente Vertreter betrachtet und fossile Repräsentanten nicht mit bedacht. Wir wollen hier einen Schwerpunkt setzen und die Wichtigkeit von Fossilien für morphologische Untersuchungen in Kombination mit phylogenetischen Interpretationen beleuchten. 6 6. Graduiertenforum Morphologie 2013 Ulm Dazu haben wir zwei Hauptredner geladen, die sich dieser Illumination annehmen wollen: Dr. Joachim T. Haug aus München wird einen Blick aus der Sicht eines Arthropodenforschers werfen. Herrn Dr. Torsten Scheyer aus Zürich konnten wir gewinnen uns den Sachverhalt aus Sicht eines Wirbeltierspezialisten zu zeigen. Selbstverständlich werden auch die Methoden bei diesem Fachgruppentreffen nicht zu kurz kommen. Prof. Dr. Paul Walther aus Ulm wird einen Hauptvortrag zum Thema dreidimensionale Rekonstruktion aus FIB-SEM (Focused Ion Beam Scanning Electron Microscopy) Daten halten. Wir freuen uns auf schöne Tage mit interessanten Vorträgen und viel Zeit für Gespräche und Diskussionen! Das Organisationsteam Verena Kutschera & Hans Peter Katzmann 7 6. Graduiertenforum Morphologie 2013 Ulm Allgmeine Informationen Tagungsort Die Tagung findet statt auf dem Campus der Universität Ulm auf dem Eselsberg in Ulm. Tagungsort ist der Hörsaal 16, Niveau 1, Gebäude N24 an der Universität Ost. Dort finden die Vorträge sowie die Postersession statt. Da sämtliche Parkmöglichkeiten an der Universität mittlerweile kostenpflichtig sind, raten wir allen Teilnehmern mit dem von uns kostenlos zur Verfügung gestellten Konferenzticket die öffentlichen Verkehrsmittel zu nutzen. Am schnellsten erreicht man die Universität mit der Linie 3 Richtung Wissenschaftsstadt/ Science Park II, die Linien 5, 6, 13, 15 und 45/48/49 können je nach Situation jedoch ebenso genutzt werden. Registrierung Die Regsitrierung findet vor und während des Icebreakers am Donnerstag, dem 10.10.2013 am Tagungsort statt. Teilnehmer die später anreisen können sich jederzeit während der Tagung bei den mit „Organisation“-Anhänger kenntlich gemachten Personen registrieren. Da der Teilnehmerbeitrag bereits vorab beglichen wurde, ist bei der Registrierung lediglich der Tagungsbeitrag in digitaler Form (Vorträge) oder gedruckter Form (Poster) mitzubringen. In unmittelbarer Nähe zum Tagungsort besteht die Möglichkeit Schließfächer für die Unterbringung von Jacken, Taschen o. Ä. zu reservieren, hierfür ist lediglich ein Vorhängeschloss nötig. Internetzugang Wir empfehlen allen Teilnehmern sich vorab über ihre Universität einen Zugang zum eduroam Netzwerk geben zu lassen, über diesen ist freier Internetzugang auf dem gesamten Universitätsgelände möglich. Sollten hierzu Fragen auftauchen stehen die Veranstalter gerne mit Rat und Tat zur Verfügung. Für Teilnehmer die keine Möglichkeit haben über einen eduroam-Zugang ins Internet zu gelangen können auf Wunsch Zugangsdaten zum Universitäts-WLAN zur Verfügung gestellt werden. Darüber hinaus ist es möglich einen zur Verfügung gestellten PC mit Internetzugang zu nutzen. 8 6. Graduiertenforum Morphologie 2013 Ulm Vorträge und Poster Die Vortragszeit beträgt 30 Minuten inklusive Diskussion. Bitte achten Sie darauf Ihren Vortrag nicht länger als 25 Minuten zu planen um eine angemessene Diskussion zu ermöglichen. Im Hörsaal steht für die Präsentationen ein PC mit Windows 7, Office 2013 und Acrobat XI zur Verfügung sowie ein Apple Macintosh mit OS X 10.8.5, Microsoft Office 2011 und Acrobat 9. Bitte geben Sie Ihre Vorträge rechtzeitig, im Idealfall bereits bei der Registrierung, spätestens jedoch in der Pause vor Ihrem Vortragsblock beim Organisationskomitee ab. Poster sollten das Format DIN A0 nicht überschreiten, für die Präsentation am Freitag Abend stehen Posterwände und Befestigungsmaterial zur Verfügung. Essen Wir freuen uns außerordentlich bei dieser Tagung wieder das Essen für alle Teilnehmer kostenlos anbieten zu können. Lediglich die Getränke beim Konferenzdinner sowie ein eventuelles Mittagessen am Sonntag müssen selbst übernommen werden, wir denken dies ist jedoch für alle Teilnehmer auch die gerechteste Lösung. Exkursion Aufgrund der geringen Anzahl an Vortragsbeiträgen fällt das Vormittagsprogramm am Sonntag leider aus, stattdessen wird eine kostenlose Exkursion in Ulm stattfinden. Geplant sind ein Besuch des Münsters mit Besteigung des höchsten Kirchturms der Welt. Im Anschluss wird uns ein kleiner Spaziergang die Donau entlang führen bevor wir im Naherholungsgebiet Friedrichsau das Donauaquarium mit Tiergarten besuchen werden. Auch dieser Programmpunkt wird für alle Teilnehmer kostenlos angeboten. 9 6. Graduiertenforum Morphologie 2013 Ulm Programmübersicht Donnerstag 10.12.2013 18:00 Uhr Icebreaker N24/H16 (siehe Plan) Freitag 11.10.2013 09:15 Uhr Begrüßung 09:30 Uhr Schütz „Das Grafikprogramm Blender und dessen Möglichkeiten zur Darstellung morphologischer Rekonstruktionen” 10:00 Uhr van de Kamp „Vom Präparat zum Kunststoffmodell: 3D-Druck für Morphologen“ 10:30 Uhr Kaffeepause 11:00 Uhr Walther “Novel electron microscopic methods for 3D imaging of subcellular structures” 12:00 Uhr Mittagspause, Essen in der Mensa (wird bezahlt) 13:30 Uhr Rühr „Die cephale Morpholgie von Tomocerus flavescens (Hexapoda: Collembola): Eine 3D-Rekonstruktion” 14:00 Uhr Schwaha „Eine neue Süßwasser-Kamptozoe?“ 14:30 Uhr Kaffeepause 15:00 Uhr Kutschera „Proximale Sklerite an Pleopoden von Malacostraca (Crustacea s. l.)” 15:30 Uhr Nagler „A revision of the genus Tanycypris (Ostracoda, Cypricercinae) with the description of a new species and a dichotomous key of the genus.” 16:00 Uhr Kruppert „Geometrische Morphometrie von Wasserflöhen” 17:00 Uhr Postersession Bier & Brezeln 19:00 Uhr Stadtführung, Treffpunkt Stadthaus 10 6. Graduiertenforum Morphologie 2013 Ulm Samstag 12.10.2013 9:30 Uhr Hörnig „Evolution und erdgeschichtliches Auftreten von Brutpflegeverhalten innerhalb der dictyopteren Insekten” 10:00 Uhr Eulitz „Die Ootheken der Gottesanbeterinnen (Mantodea)” 10:30 Uhr Kaffeepause 11:00 Uhr Haug „Palaeo-Evo-Devo von Arthropoden – Was uns Fossilien über die Evolution von Entwicklungsmustern sagen können” 12:00 Uhr Mittagspause (Pizzabestellung, wird bezahlt) + Gruppenfoto 13:30 Uhr Kerbl „Kleiner Wurm ganz groß – Die Entwicklung des Nervensystem des interstitiellen „Polychaeten“ Dinophilus gyrociliatus (Annelida, Dinophilidae)“ 14:00 Uhr Hannig „Die Streptoneurie bei Paradiesschnecken setzt keine ontogenetische Torsion voraus” 14:30 Uhr Sanmugaraja „Habitatbestimmung bei Katzen anhand der Unterarmknochen“ 15:00 Uhr Kaffeepause 16:00 Uhr Nowak „Eine neue Rekonstruktion des Schultergürtels und der Vorderextremität von Metoposaurus diagnosticus krasiejowensis” 16:30 Uhr Jaquier „Die Halswirbelsäule des Giraffenhalssauriers: eine knochenhistologische Untersuchung” 17:00–18:00 Uhr Scheyer „Integration morphologischer, knochenhistologischer und embryologischer Datensätze - drei paläobiologische Fallbeispiele (Schildkröten, Krokodile und marine Reptilien aus der Trias-Zeit” 19:00 Uhr Abschlussessen im Barfüßer (Essen nach Wahl, Getränke auf eigene Rechnung) Sonntag 13.10.2013 11:00 Uhr Münsterbegehung und -besteigung 12:30 Uhr Auf Wunsch kleine Führung durch das Münster, ansonsten vorzeitiger Aufbruch zum Spaziergang 14:00 Uhr Spaziergang zum Tiergarten Ulm 11 6. Graduiertenforum Morphologie 2013 Ulm Vorträge 12 6. Graduiertenforum Morphologie 2013 Ulm Das Grafikprogramm Blender und dessen Möglichkeiten zur Darstellung morphologischer Rekonstruktionen Philipp Schütz*, Verena Kutschera**, Andreas Maas** & Dieter Waloszek** * Institute of Experimental Ecology, Universität Ulm, Albert-Einstein-Allee 11, 89081 Ulm ** Biosystematische Dokumentation, Universität Ulm, Helmholtzstr. 20, 89081 Ulm Blender ist eine bereits sehr mächtige kostenlose Open-Source-Software, mit deren Hilfe sich 3D-Grafiken und - Animationen erstellen lassen. Das Programm wird oft wegen seiner benutzerfreundlichen Oberfläche gegenüber anderen Grafikprogrammen bevorzugt. Morphologische Daten, wie z.B. Schnittserien können in Blender importiert und entweder als Volumenmodelle dargestellt werden oder als Vorlage zur Erstellung von Meshes dienen. Blender ist dabei weniger eine Ein-Klick-Lösung zur Verarbeitung von Rohdaten, da es hauptsächlich für das Modellieren und Animieren konzipiert ist, jedoch eignet sich das Programm sehr gut zur Bearbeitung von Objekten. Es ist möglich, vorgefertigte 3D-Modelle verschiedener Formate zu importieren und diese im 3D-Fenster nachzubearbeiten. Hierbei bietet Blender eine breite Palette zur Erstellung von Shadern, über die den Objekten Farben, Texturen, Transparenz, Bumpmaps usw. zugewiesen werden können. Fertige Szenen mit benutzerdefinierter Belichtung, Hintergrund und Kameraposition können als Bilder oder als Filmsequenz gerendert werden. Da Blender von seinen Programmierern ständig überarbeitet und verbessert wird und auch immer mehr Addons zur Verfügung stehen, lässt die Software auch in Zukunft auf interessante Möglichkeiten in der 3D-Rekonstruktion hoffen. 13 6. Graduiertenforum Morphologie 2013 Ulm Vom Präparat zum Kunststoffmodell: 3D-Druck für Morphologen Thomas van de Kamp* & Tilo Baumbach* * ANKA / Institut für Photonenforschung und Synchrotronstrahlung, Karlsruher Institut für Technologie (KIT) 3D-Bildgebungsverfahren und die digitale Bearbeitung von Volumendaten sind inzwischen weitverbreitete Werkzeuge für morphologische Untersuchungen. 3D-Drucker können diese Daten greifbar machen, indem sie aus einem digitalen Datensatz ein Kunststoffmodell erzeugen. Auch wenn die Geräte häufig zur Herstellung von Prototypen für Architektur und Maschinenbau verwendet werden, kann ihr Einsatz für Biologen sehr interessant sein. So können reale 3D-Modelle zum wissenschaftlichen Erkenntnisgewinn beitragen. Die Bewegung winziger Gelenke lässt sich etwa an einem funktionellen Kunststoffmodell oft besser nachvollziehen als am virtuellen Datensatz. Auch die Herstellung von Schaumodellen für Präsentationen kann sehr nützlich sein. Für den 3D-Druck kommen je nach Druckertyp verschiedene Techniken zum Einsatz. Häufig verwendete Verfahren sind etwa das Fused Deposition Modeling, das einen schmelzfähigen Kunststoff verwendet und das Multi-Jet Modeling, bei dem ein Modell mit Hilfe eines Druckkopfes schichtweise aufgebaut wird. Beim ColorJet Printing wird ein Gipspulver durch ein Bindemittel gehärtet. Die Verfahren unterscheiden sich mitunter stark in Geschwindigkeit, Detailauflösung, Farbfähigkeit und Betriebskosten. Im Vortrag werden verschiedene 3D-Fertigungsverfahren präsentiert. Außerdem wird der 3D-Drucker der ANKA (ProJet HD 3500Plus) vorgestellt und der Weg vom biologischen Präparat zum fertigen Kunststoffmodell erläutert. Anwendungsbeispiele beinhalten funktionelle Modelle biologischer Gelenke, Schaumodelle von Bernsteininklusen und die Herstellung angepasster Probenhalter für die Mikrotomographie. 14 6. Graduiertenforum Morphologie 2013 Ulm Novel electron microscopic methods for 3D imaging of subcellular structures Paul Walther* * Zentrale Einrichtung Elektronenmikroskopie, Universität Ulm Electron microscopy is experiencing a remarkable comeback in life science. This is mainly because new methods have been developed. The main challenges of visualizing subcellular structures in 3D is sample preparation preventing artifact formation. We believe that cryo-fixation protocols are the method of choice to arrest cells at defined physiological conditions to preserve a life-like state. Afterwards these samples are dehydrated at cold temperature (freeze substituted) and embedded in a resin. Different protocols are now available to obtain a 3D dataset. With scanning transmission electron microscopic (STEM) tomography, semi-thin sections up to a thickness of about 1 µm can be recorded in 3D. The resolution is very good in X and Y (in the range of a few nanometers, depending from the structural preservation during specimen preparation), and reasonably well in Z (limited by the missing wedge problem). Eukaryotic cells, however, have a thickness of up to 10 µm or more, therefore, even with STEM tomography only a small portion of the cell can be recorded. Alternative methods are serial sectioning tomography and focused ion beam/ scanning electron microscopic (FIB/SEM) tomography. Serial sectioning only requires an ultra microtome and an electron microscope as equipment. It is easy to search for an area of interest and the sections can be stored and reused e. g. to further magnify an area of interest. The resolution in Z is, however, restricted to the section thickness (about 100 nm). With FIB/SEM tomography the resolution in X and Y can be as good as in TEM images of the same plastic embedded material. When very thin layers of the embedding medium are removed with the focused ion beam in between two SEM images of the block face, even isotropic resolution can be obtained. 15 6. Graduiertenforum Morphologie 2013 Ulm Die cephale Morpholgie von Tomocerus flavescens (Hexapoda: Collembola): Eine 3D-Rekonstruktion Peter Rühr*, Alexander Blanke* & Bernhard Misof* * Zoologisches Forschungsmuseum Alexander Koenig, Zentrum für Molekulare Biodiversitätsforschung, Adenauerallee 160, 53113 Bonn, Die Springschwänze (Collembola), eine primär flügellose Gruppe der Hexapoden, werden zur Gruppe der Entognatha (Collembola + Protura + Diplura) gezählt. Die drei Taxa sind unter anderem dadurch charakterisiert, dass die Mandibeln und Maxillen durch laterale Mundfalten in der Kopfkapsel eingeschlossen sind (Entognathie). Neuere Publikationen stellen die Monophylie der Entognatha in Frage. Daher werden von der detaillierten Erforschung der inneren und äußeren Kopfmorphologie, besonders der Mundwerkzeuge, des Endoskeletts und der Muskulatur, wichtige Hinweise auf die Evolution der Entognathie sowie die Verwandtschaftsverhältnisse der primär flügellosen Hexapodengruppen untereinander und zu den Ectognatha erwartet. Der erste Schritt, der nötig ist, um die Kopfmuskulatur und andere cephale Strukturen der Collembolen mit denen der Ectognatha phylogenetisch zu untersuchen, ist die Homologisierung dieser Strukturen. Die Kopfmuskulatur der drei entognathen Taxa konnte bisher nicht erfolgreich mit der der Ectognatha homologisiert werden. Synchrotron-micro-Computertomographie (SR-µCT) der cephalen Strukturen kann hierbei helfen die komplizierten Verhältnisse innerhalb des Collembolenkopfes sichtbar zu machen und Homologieannahmen über die klassischen Kriterien der Lage und spezifischen Qualität zu ermöglichen. Die Daten, die der dreidimensionalen Rekonstruktion des Kopfes von Tomocerus flavescens zu Grunde liegen, wurden mittels SR-µCT an der Swiss Light Source des Paul-Scherrer-Instituts erstellt. Anschließend erfolgte die Rekonstruktion mittels den frei erhältlichen Programmen Reconstruct und Blender. Die nicht destruktive und schnelle Technik der SR-µCT liefert extrem hochauflösende Scanbilder ohne Schnittartefakte. Der Vergleich der 3D-Rekonstruktion des Kopfes von T. flavescens mit umfassenden, früheren Arbeiten an ectognathen Insektenköpfen führt zu ersten Homologiehypothesen. Muskeln innerhalb des Collembolenkopfes könnten ein Homologon zu Muskeln ectognather Insekten darstellen. Dadurch könnte es jetzt möglich sein, die Muskulatur der Collembolen in bereits bestehende Forschungsarbeiten zu integrieren, die sich mit der Evolution der Kopfmuskulatur der Ectognatha beschäftigt. 16 6. Graduiertenforum Morphologie 2013 Ulm Eine neue Süsswasser-Kamptozoe? Thomas Schwaha* * Universität Wien, Department für integrative Zoologie, Althanstraße 14, 1090 Wien Kamptozoen oder Entoprocten sind eine sehr kleine Gruppe von etwa 180 Arten vorwiegend mariner, sessile Tiere. Die meisten dieser Arten leben solitär und filtrieren mit einer cilienbesetzten Tentakelkrone Nahrungspartikel herbei. Ein paar Arten bilden auch zusammenhängende Kolonien. Derzeit sind nur zwei Arten beschrieben die im Süsswasser auftreten: Urnatella gracilis und Loxosomatoides sirindhornae. Morphologische Untersuchungen an diesen beiden Arten zeigten, dass die Besiedelung des Süsswassers offensichtlich starke Modifikationen des Nephridialsystems zur Folge hatte. Konkret setzt sich dieses System aus zahlreichen untereinander verbundenen Terminalorganen zusammen, die im Wesentlich der Osmoregulation und wahrscheinlich kaum der Exkretion dienen. Eine kürzlich gefundene tropische Süsswasserkamptozoe scheint eine neue Art darzustellen. In diesem Beitrag werde ich basierend auf morphologische Merkmalen darstellen, warum diese Art eine noch nicht beschriebene darstellt und weshalb sogar vielleicht eine neue Gattung notwendig ist. 17 6. Graduiertenforum Morphologie 2013 Ulm Proximale Sklerite an Pleopoden von Malacostraca (Crustacea s. l.) Verena Kutschera*, Andreas Maas* & Dieter Waloßek* * Biosystematische Dokumentation, Universität Ulm, Helmholtzstr. 20, 89081 Ulm Krebse sind u. a. durch das Vorhandensein eines Proximalen Enditen an allen Beinen hinter der ersten Antenne charakterisiert. Dieser Proximale Endit war anfänglich vielleicht nicht mehr als eine skleritische Cuticularplatte median in der Gelenkmembran des Basisgelenks mit wenigen Borsten darauf. In der weiteren Evolution (durch Taxa belegt), entstanden andere Formen des Proximalen Enditen, z. B. eine deutliche Auswölbung, stärkere Beborstung und neue Funktionalitäten wie freie Beweglichkeit durch Muskeln. Ferner wurde der Proximale Endit zu einer ringförmigen Struktur, Coxa genannt, umgewandelt. Sie umschließt das Bein unterhalb des Basipoditen, der durch Membrananteile von diesem getrennt ist (weitere Membrananteile bilden dabei das neue Beingelenk). Somit haben alle nachantennularen Beine der Krebse entweder einen solchen Proximalen Enditen oder eine Coxa als sein Derivat, aber niemals beide Strukturen. Eine Ausnahme bilden die so genannten Pleopoden, die Beine des hinteren Körperabschnittes (Pleon) der Malacostraca, zu denen prominente Vertreter wie Hummer, Krabben und Kellerasseln gehören. Wir haben die Pleopoden verschiedener Malacostraca mit Alizarinfärbung und Fluoreszenzmikroskopie untersucht. Beide Methoden erlauben das Unterscheiden von verkalkten und membranösen Teilen der Kutikula, was zum Nachweis kleiner Kalkplättchen, Sklerite an der Basis der Pleopoden führte. Diesen morphologischen Befund diskutieren wir dahingehend, ob es sich bei den Skleriten um eine Neubildung der Malacostraca, um einen Proximalen Enditen oder gar eine Coxa handelt? Auch die Evolution derartiger membrangebundener Sklerite innerhalb der Malacostraca soll mit diesem Vortrag beleuchtet werden. 18 6. Graduiertenforum Morphologie 2013 Ulm A revision of the genus Tanycypris (Ostracoda, Cypricercinae) with the description of a new species and a dichotomous key of the genus. Christina Nagler*, Renate Matzke-Karasz** & Juergen Geist* * Lehrstuhl für Aquatische Systembiologie, Wissenschaftszentrum Weihenstephan für Ernährung Landnutzung und Umwelt, Technische Universität München, Mühlenweg 22, 85354 Freising ** Department für Geo- und Umweltwissenschaften, Paläontologie und Geobiologie, LudwigsMaximilians Universität München, Richard-Wagner-Straße 10, 80333 München Die Taxonomie und Systematik der Cypricercinae, einer Unterfamilie der Ostrakoden, ist noch nicht vollständig geklärt. Sie vereinen alle Arten, die den Triebel loop – ein Merkmal des Attachment der kaudalen Rami – aufweisen. Das Ziel dieser Studie ist es den systematischen Status der Gattung Tanycypris zu überprüfen und einen Bestimmungsschlüssel für diese Gattung zu erstellen. Im Rahmen einer faunistischen Analyse der Ostrakoden im Stadtgebiet Münchens wurden im Jahr 2011 Proben aus Wasserpflanzen-Schaubecken in einem Gewächshaus des Botanischen Gartens genommen. Neben der ubiqitären Art Cypridopsis vidua (O.F. Müller, 1776), wurden vier nicht heimische Arten der Unterfamilie Cypricercinae gefunden. Diese konnten als Chlamydotheca arcuata (Sars, 1901), Strandesia bicuspis (Claus, 1892), Tanycypris centa (Chang, Lee & Smith 2012) und als eine noch nicht beschriebene Art der Gattung Tanycypris bestimmt werden. Während des Prozesses der Beschreibung der neuen Art wurden eine Vielzahl an taxonomischen Unsicherheiten in der Gattung Tanycypris aufgedeckt, die zu einer Revision der neun Arten, die momentan dieser Gattung angehören, führten. Die Analysen basieren hauptsächlich auf archiviertem Typenmaterial, das aus verschiedenen Sammlungen angefordert wurde. Verschiedene Mikroskopie-Techniken wurden angewandt, um taxonomisch relevante Merkmale zu identifizieren und zu zeichnen. Außerhalb der Gattung Tanycypris wurden verschiedene andere Arten der Cypricercinae untersucht, um ihren taxonomischen Status zu überprüfen; diese waren: Cypricercus inermis, Diaphanocypris meridana, Dolerocypris sagitta, Herpetocypris bonettoi und Nealecypris obtusa. Dies war nötig, da einige von diesen Arten zeitweise als Tanycypris behandelt wurden. Basierend auf den Merkmalen des Carapax, der Anwesenheit des Wouters Organ, der dorsalen Seta auf dem ersten Segment der ersten Antenne und der Konfiguration der b-, c- und d-Seta des ersten Thorakopoden, wurde ein Bestimmungsschlüssel für die Gattung Tanycypris entwickelt. Der systematische Status von Dolerocypris marina und Herpetocypris telavivensis bleibt unsicher, weil das Material der Typen trotz intensiver Bemühungen nicht gefunden werden konnte. Genauso sind genetische Analysen wünschenswert, um deren taxonomische Einteilung zu verfestigen. 19 6. Graduiertenforum Morphologie 2013 Ulm Geometrische Morphometrie von Wasserflöhen S. Kruppert*, M. Rabus**,L. Weiss* M. Horstmann*, C. Laforsch** & R. Tollrian* *Department of Animal Ecology, Evolution & Biodiversity; Ruhr- University Bochum, Germany; **University of Bayreuth, Animal Ecology I, Bayreuth) Die Morphologie eines Organismus’ ist in der Regel an sein natürliches Umfeld angepasst, was sich geradezu beispielhaft in den gut untersuchten Räuber-induzierten morphologischen Verteidigungen von Daphnia widerspiegelt. Um die Funktion morphologischer Strukturen zu untersuchen, bietet sich die geometrische Morphometrie an. Die Genauigkeit solcher Untersuchungen steht und fällt mit der Beschreibbarkeit und Verlässlichkeit von Messpunkten, sogenannten Landmarks. Strukturen, die sich durch Kurven und homogene Oberflächen (oder sich wiederholende Muster) auszeichnen, wie beispielsweise der Körper von Daphnia, sind bei der Determinierung von Landmarks und ihrer morphometrischen Beschreibung problematisch. Wir bieten eine Möglichkeit eben solche Strukturen akkurat zu beschreiben, und präsentieren diese am Modelorganismus Daphnia pulex. Wir haben dazu 8 Individuen von Daphnia pulex mit Congo Rot gefärbt und an einem confocalen Laser-Scanning Mikroskop als Bilderserie (Image Stack) aufgenommen. Anhand dieser Stacks konnten wir 184 gleichmäßig verteilte Landmarks mit Hilfe eines Rasters beschreiben. Anschließend haben wir diese Landmarks in ein dreidimensionales kartesisches Koordinatensystem übertragen. Basierend auf diesen Koordinaten kreierten wir ein generalisiertes Model der Oberfläche von Daphnia’s Morphologie. Dieses Model ermöglicht uns nun den Vergleich von unterschiedlichen Morphotypen, die aufgrund unterschiedlicher Umweltbedingungen ausgeprägt werden können. Ein solcher Vergleich bietet die Chance bisher unerkannte Eigenschaften eines Räuberinduzierten Morphotyps zu beschreiben. 20 6. Graduiertenforum Morphologie 2013 Ulm Evolution und erdgeschichtliches Auftreten von Brutpflegeverhalten innerhalb der dictyopteren Insekten Marie K. Hörnig*, Carolin Haug**, Andy Sombke*, Steffen Harzsch* & Joachim T. Haug** * Ernst-Moritz-Arndt-Universität Greifswald, Zoologisches Institut und Museum, Cytologie und Evolutionsbiologie, Soldmannstraße 23, 17487 Greifswald ** Ludwig-Maximilians-Universität (LMU) München, Biozentrum, Biologie II, Großhaderner Straße 2, 82152 Planegg-Martinsried Dictyoptera ist eine Gruppe der polyneopteren Insekten und umfasst heute Schaben, Mantodeen und Termiten. Dictyopteren waren mit den sogenannten Schabenartigen bereits vor ca. 300 Millionen Jahren in den karbonischen Wäldern vertreten. Aber auch aus jüngeren Zeitepochen gibt es zahlreiche Funde fossiler Dictyopteren; hierbei machen Schaben und Schabenartige den größten Anteil aus, Mantodeen und Termiten sind sehr viel seltener. Moderne Dictyopteren zeigen zahlreiche Abstufungen von Sozialund Brutpflegeverhalten. Dazu gehören u.a. die Ausbildung einer Oothek (zum Schutz der Eier), das Tragen der Oothek am Körper bis hin zur Ovoviviparie, das Versorgen der Nymphen durch die Eltern und schließlich Eusozialität (bei Termiten). Fossilien liefern wichtige Daten für die Rekonstruktion der Evolution morphologischer Strukturen, können aber eher selten zum Verständnis der evolutionären Entwicklung von Verhalten beitragen. Wir stellen Beispiele vor, bei denen Fossilien direkte und/oder indirekte Hinweise auf Aspekte des Brutpflegeverhaltens innerhalb der Dictyopteren bieten können und damit auf das zeitliche Auftreten dieser Verhaltensweisen. Frühe bekannte Dictyopteren aus dem Karbon besitzen einen langen Ovipositor, was darauf hindeutet, dass Dictyopteren vor 300 Millionen Jahren noch keine Ootheken bildeten. Schaben mit einem reduzierten Ovipositor, Mantiden (ebenfalls mit kurzem Ovipositor) und fossile Ootheken wurden erst von kreidezeitlichen Fundstätten wie der Crato-Formation in Brasilien (110 Millionen Jahre alt) beschrieben. Dies zeigt, dass diese einfache Form der Brutpflege in der Kreide entwickelt war, aber keine älteren Hinweise darauf existieren. Den ältesten Hinweis auf Sozial- und weiterreichendes Brutpflegeverhalten innerhalb der Dictyopteren bietet ein 50 Millionen Jahre alter baltischer Bernstein. Dieser enthält eine Gruppe von Schabennymphen. Ihre Morphologie weist daraufhin, dass sie nicht selbständig, sondern auf die Fürsorge von Elterntieren angewiesen waren. Fossilien wie die hier vorgestellten, stellen einen wertvollen Beitrag für die Rekonstruktion der Evolution des Brutpflegeverhaltens dar und ermöglichen vor allem auch die zeitliche Einordnung des Auftretens der verschiedenen Ausprägungen von Brutpflege innerhalb der Dictyoptera. 21 6. Graduiertenforum Morphologie 2013 Ulm Die Ootheken der Gottesanbeterinnen (Mantodea) Ute Eulitz* * Museum für Tierkunde, Senckenberg Naturhistorische Sammlungen Dresden, Königsbrücker Landstrasse 159, 01129 Dresden, Die Ordnung der Gottesanbeterinnen (Mantodea) umfasst rund 2.400 gültige Arten in 436 Gattungen und 15 Familien. Es sind thermophile, räuberische Insekten, die in den Tropen und Subtropen weit verbreitet sind. Die innere Stammesgeschichte ist weitgehend ungeklärt, innerhalb der Mantodea werden wenige Autapomorphien für die Taxa genannt. Molekulare Analysen zeigen eine künstliche Unterteilung bei vielen der traditionellen Gruppen. Weibliche Gottesanbeterinnen legen ihre Eier zum Schutz in großen Eipaketen, sog. Ootheken, ab. Die Oothek besteht aus einer schaumigen Masse, die durch akzessorische Drüsen abgesondert werden. Ootheken werden an Wänden, auf Steinen, an Blättern, Zweigen oder Stängel abgelegt, und in der Erde vergraben. Die Zahl der schlüpfenden Nymphen variiert je nach Art zwischen 10 bis 300. Im Gegensatz zu den Schaben (Blattodea) weisen die Eipakete der Mantiden eine enorme strukturelle Vielfalt auf, die dennoch charakteristisch und taxonspezifisch ist. Bisher gibt es keine Arbeiten, in welchen Ootheken zur Rekonstruktion der Mantiden-Phylogenie genutzt wurden. Dieses Projekt wird die erste integrative Studie sein und diese Lücke füllen. 22 6. Graduiertenforum Morphologie 2013 Ulm “Palaeo-Evo-Devo von Arthropoden – Was uns Fossilien über die Evolution von Entwicklungsmustern sagen können” Joachim T. Haug* * Ludwig-Maximilians-Universität (LMU) München, Biozentrum, Biologie II, Großhaderner Straße 2, 82152 Planegg-Martinsried Fossilien erlauben uns den Blick zurück in die Vergangenheit. Sie vermitteln uns einen Eindruck, wie einstmals gelebt-habende Organismen ausgesehen haben. Auch wenn es keinesfalls so ist, dass ausgestorbene Organismen automatisch „primitiver“ sind als ihre modernen Verwandten, so besitzen doch viele Vertreter fossiler Arten Eigenschaften, die sich nicht mehr in der modernen Fauna finden. Unter diesen kann sich dann eine solche Eigenschaftskombination befinden, die eine fossile Art als Schwesterart eines modernen Monophylums ausweist und somit potentiell die Merkmalsevolution zu diesem hin genauer auflöst. Dies gilt nicht nur für morphologische Merkmale adulter Organismen, sondern auch für solche früherer Ontogenesestadien inclusive Larvenstadien. Damit lassen sich auch ganze ontogenetische Sequenzen ausgestorbener Arten rekonstruieren und dadurch dann die evolutiven Veränderungen der Ontogenese morphologischer Strukturen herleiten. Es werden Beispiele aus verschiedenen Gruppen der Gliederfüßer gezeigt. Besonders interessant sind hierbei Monophyla, in denen besonders auffällige ontogenetische Muster entwickelt sind. Hierzu gehören solche Gruppen, deren Vertreter sich durch eine Metamorphose entwickeln. Metamorphose beschreibt einen kurzen Zeitraum der post-embryonalen Entwicklung, bei dem sich die Morphologie des Organismus auf drastische Weise verändert. Unter den Gruppen, die eine ausgeprägte Metamorphose zeigen, sind die holometabolen Insekten, aber auch verschiedene Innengruppen der Malacostraca, der höheren Krebse. Relativ frühe postembryonale Entwicklungsstadien sowohl verschiedener Krebse als auch verschiedener Insekten finden sich in der Tat im Fossilbericht. So kommen beispielsweise die äußerst bizarr wirkenden Larvenstadien von Bärenkrebsen und Langusten zu Tausenden in verschiedenen Lagerstätten des Erdmittelalters vor. Während die meisten dieser fossilen Larven die prinzipielle Morphologie ihrer modernen Gegenstücke aufweisen, gibt es dennoch einige Formen, die deutlich hiervon abweichen. Diese Fossilien erlauben somit den Einblick in die frühe Spezialisierung und Diversifikationsphase der frühen Bärenkrebse und Langusten. Neben weiteren Beispielen sollen die Möglichkeiten und Herausforderungen eines Palaeo-Evo-Devo-Ansatzes und die potentiellen Fragestellungen, die er beantworten kann, beleuchtet werden. 23 6. Graduiertenforum Morphologie 2013 Ulm Kleiner Wurm ganz groß – Die Entwicklung des Nervensystem des interstitiellen „Polychaeten“ Dinophilus gyrociliatus (Annelida, Dinophilidae) Alexandra Kerbl* * Sektion für Meeresbiologie, Department für Biologie, Universität Kopenhagen, Strandpromenaden 5, 3000 Helsingør, Dänemark. Das Nervensystem ist eines der bestuntersuchten Organsysteme in verschiedenen Tiergruppen und gilt als eines der konserviertesten Systeme. Dennoch gibt es nur sehr wenige Studien über die Entwicklung des Nervensystems, was besonders auf die Vertreter der Spiralia zutrifft. Anneliden nehmen hier aufgrund der reichen Palette an Bauplänen und Lebensstilen eine besondere Stellung ein. Eine der eher speziellen Familien sind die Dinophilidae. Sie leben interstitiell und weisen daher starke Reduktionen der Körpergröße, der Zellzahl, der äußeren Merkmale und der internen Organisation im Vergleich zu den makroskopischen Anneliden auf. Eine Art, Dinophilus gyrociliatus, weist außerdem noch die Ausprägung von Zwergmännchen auf. Da diese Tiere eventuell progenetischen Ursprungs sind, ist die Untersuchung der Entwicklung vor allem des Nervensystems mit verschiedenen Methoden (Immunozytochemie und CLSM, Serienschnitte, Licht- und Elektronenmikroskopie mit 3D-Rekonstruktion sowie in der Folge auch die Verfolgung molekularer Ansätze) das Ziel dieser Studie. In erwachsenen Weibchen besteht das Nervensystem aus dem Gehirn, dem ventralen Nervenstrang und dem peripheren sowie dem stomatogastrischen Nervensystem. Das Gehirn wird von 500 bis 800 Zellen gebildet, von denen 6 bis 8 mit antiSerotonin-Antikörpern markiert werden können. 7 longitudinale Nervenbündel und 18 Kommissuren sowie ein engmaschiges Netzwerk aus feinen Nervenfasern zeichnen den ventralen Nervenstrang aus. Die frühen Entwicklungsstadien sind noch nicht genau analysiert, doch die ersten Anzeichen des serotonergen Nervensystems können bereits vor dem Schlüpfen am 6. Tag nach Ablage der Kokons festgestellt werden. Kurz nach dem Schlüpfen sind Nervenfasern im Gehirn und im ventralen Nervenstrang schon klar erkennbar, ebenso wie einzelne Perikarya nahe bei den Kommissuren. Eine detaillierte Rekonstruktion des Nervensystems während der Entwicklung wird in Zukunft als Plattform für Untersuchungen dienen, die zum einen Vergleiche zu anderen Arten (wie zum Beispiel Platynereis dumerilii) ermöglichen, zum anderen aber auch auf die Kartierung von genetischer Information im Nervensystem abzielen, die wiederum über mehrere Zweige der Spiralia hinweg verglichen werden kann. 24 6. Graduiertenforum Morphologie 2013 Ulm Die Streptoneurie bei Paradiesschnecken setzt keine ontogenetische Torsion voraus Leonie Hannig*, Simon Schwarz* & Heinz-R. Köhler* *Physiologische Ökologie der Tiere, Institut für Evolution und Ökologie, Universität Tübingen, Konrad-Adenauer-Straße 20, D - 72072 Tübingen. Organismen in der Umwelt sind gegenüber vielen Substanzen ausgesetzt, u.a. auch gegenüber Schwermetallen, die zum Beispiel über Autoabgase emittiert werden. Wir haben die Toxizität des Schwermetalls Platin, das in Autokatalysatoren verwendet wird, untersucht und zu diesem Zwecke Embryonen der Paradiesschnecke Marisa cornuarietis während ihrer Entwicklung gegenüber Platinchlorid exponiert. Wir konnten 2+ beobachten, dass hohe Konzentrationen von Platin die Entwicklung einer externen Schale verhindern, indem das differentielle Wachstum von Mantelrand und Schalendrüse gestoppt wird. Da das schalenbildende Gewebe somit nicht nach außen wächst, kommt 2+ es zu einer Internalisierung der Schale. Gegenüber Platin exponierte Individuen besitzen somit einen nackten Eingeweidesack, auf dem caudal die Kieme liegt. Ohne differentielles Wachstum von Mantelrand und Schalendrüse findet allerdings auch keine ontogenetische Torsion statt. Diese Torsion, eine Rotation des Visceropalliums relativ zum Cephalopodium, ist nach Lehrbuchmeinung die Ursache für die Streptoneurie der Schnecken, die Überkreuzung der Pleurovisceralkonnektive, und bei „prosobranchen“ Schnecken die Positionierung von Kiemen und Anus vor dem Herzen. Um die Struktur 2+ des Nervensystems bei gegenüber Platin exponierten Schnecken zu untersuchen, wurden 3-D-Modelle von adulten Tieren auf Grundlage von histologischen Schnitten angefertigt. Zusätzlich wurden exponierte und nicht exponierte Embryonen von M. cornuarietis immunhistochemisch untersucht und es wurden ebenfalls 3-D-Modelle angefertigt. Zur Färbung des Nervensystems wurden Antikörper gegen alpha-Tubulin, Serotonin und FMRFamid verwendet, welche mittels konfokaler Laser-Scanning2+ Mikroskopie lokalisiert wurden. Die Modelle zeigen, dass auch bei gegenüber Platin exponierten Schnecken Streptoneurie vorliegt, obwohl bei ihnen keine horizontale Rotation des Visceropalliums stattfindet. Streptoneurie und ontogenetische Torsion scheinen somit– zumindest bei der untersuchten Art –unabhängig voneinander zu sein. 25 6. Graduiertenforum Morphologie 2013 Ulm Habitatbestimmung bei Katzen anhand der Unterarmknochen Mayuran Sanmugaraja* & Rico Schellhorn* * Steinmann-Institut für Geologie, Mineralogie und Paläontologie, Rheinische Friedrich-Wilhelms-Universität Bonn Rezente Katzen bewohnen verschiedene Habitate. Man findet sie heutzutage sowohl in offenen Gebieten, wie z.B. Wüsten oder Steppen, als auch in Wäldern, aber auch im Gebirge. Die Anpassungen an das jeweilige Habitat sind an den Unterarmknochen erkennbar. Wir haben in unserer Studie Radius und Ulna von 19 rezenten Katzenarten, von denen das bevorzugte Habitat bekannt ist, vermessen. In den verschiedenen Diagrammen, die mithilfe der Daten erstellt wurden, sind die Arten, die offene Gebiete bevorzugen, von den Waldbewohnern getrennt. Katzen, die in offenen Habitaten leben und damit an eine laufintensive Lebensweise angepasst sind, haben relativ lange und dünne Unterarmknochen. Dagegen haben Waldbewohner relativ kürzere und dickere Radii und Ulnae. Da diese einfache Methode bei rezenten Katzenarten anwendbar ist, wurde sie auch für die Bestimmung von Habitaten von fossilen Arten genutzt, u.a. bei Panthera atrox und Smilodon fatalis von der berühmten pleistozänen Fundstelle Rancho La Brea in Los Angeles (USA). Die Werte dieser beiden Arten lassen eine Anpassung an das Waldhabitat vermuten. 26 6. Graduiertenforum Morphologie 2013 Ulm Eine neue Rekonstruktion des Schultergürtels und der Vorderextremität von Metoposaurus diagnosticus krasiejowensis Marlene Nowak* & Dr. Dorota Konietzko-Meier ** *Steinmann-Institut für Geologie, Mineralogie und Paläontologie, Nußallee 8, 53115 Bonn, Deutschland ** Steinmann-Institut für Geologie, Mineralogie und Paläontologie, Nußallee 8 53115 Bonn, Deutschland Temnospondylen sind frühe Amphibien, die vom frühen Karbon bis in die Kreide vorkamen. Zu ihnen zählt das Taxon Metoposaurus, das aus vielen Fundstellen in Europa beschrieben ist. In der vorliegenden Arbeit werden ein Schultergürtel und eine damit assoziierte Vorderextremität von Metoposaurus diagnosticus krasiejowensis präsentiert. Das Fossil wird in der Sammlung der Abteilung für Biosystematik der Universität von Oppeln (Polen) unter der Katalognummer UOPB 00097 verwahrt. Das Material stammt aus der karnischen oder norischen Lagerstätte Krasiejów im Südwesten von Polen. Artikulierte Funde von Temnospondylen mit Postcranium sind selten und deshalb in der Literatur kaum beschrieben. Daher bietet das Fossil UOPB 00097 die Möglichkeit in zukünftigen phylogenetischen Untersuchungen Merkmale für das Postcranium für Metoposaurus diagnosticus krasiejowensis zu vergeben. Es wird der artikulierte Fund beschrieben, vermessen und auf dessen Grundlage Verhältnisse von Schultergürtel und Extremitätenknochen erstellt. Die Verhältnisse dienen als erster Ansatz für die Abschätzung von Verhältnissen im Skelett von Metoposaurus. Außerdem wird auf die Pentadactylie der Hand eingegangen, die bisher für Metoposauriden nicht beschrieben ist. Die bisher angenommene Vierfingrigkeit bei Metoposauriden richtete sich nach artikulierten Funden von Dutuitosaurus ouazzoui (Dutuit 1976), ein naher Verwandter von Metoposaurus. Außerdem ist es anhand der Morphologie dieses Fossils möglich, die Hypothese von Sulej (2007) zu widerlegen, wonach Metoposaurus diagnosticus krasiejowensis einen Lokomotionstyp wie die Plesiosaurier nutzte, um sich in der Wassersäule zu bewegen. Allerdings kann der Lokomotionsmodus für Metoposaurus diagnosticus krasiejowensis im Rahmen dieser Arbeit nur kurz angerissen werden. Jedoch bietet das Fossil Ansätze für zukünftige Untersuchungen, auch bezüglich der Lokomotion der Temnospondylen. 27 6. Graduiertenforum Morphologie 2013 Ulm Die Halswirbelsäule des Giraffenhalssauriers: eine knochenhistologische Untersuchung Vivien Jaquier* & Torsten Scheyer* * Paläontologisches Institut und Museum, Karl Schmid-Strasse 4, Universität Zürich, CH-8006 Zürich, Schweiz Im Laufe der Evolution gibt es bei einigen Tiergruppen unabhängig die Tendenz, lange Halswirbelsäulen auszubilden. Absolut gesehen sind die sauropoden Dinosaurier die ungeschlagenen ‚Langhälse‘ mit Halswirbelsäulen von bis zu 15 Metern, doch bezogen auf die Körpergrösse gibt es auch vor allem im aquatischen Milieu mehrere Saurier welche extrem lange Hälse besassen (z.B. Plesiosaurier). Einen extremen Fall stellt auch der Giraffenhalssaurier Tanystropheus (Protorosauria, Archosauromorpha) aus der Mittel- und Obertrias (ca. 242 bis ca. 210 Millionen Jahre) dar, dessen Hals über die Hälfte der Gesamtkörperlänge ausmachte. Bei Tanystropheus besteht der Hals aus nur 13, extrem verlängerten Wirbeln. Ein nahe Verwandter hingegen, Dinocephalosaurus aus der Trias von China, hatte einen ebenso langen Hals, aber mehr als 25 Wirbel. Solch verlängerte Hälse erfordern spezielle Anpassungen, sowohl an den Knochenbau als auch an das umgebende Weichgewebe. So besitzt Tanystropheus zum Beispiel extrem verlängerte Halsrippen, die sich über mehrere Wirbelzentren erstrecken. Bis heute unklar ist allerdings, wie dieses Tier seinen Hals gehalten hat. Das Fehlen von direkten Nachfahren oder modernen Äquivalenten im Tierreich erschwert zudem funktionsmorphologische Untersuchungen, sowie Aussagen zur Lebensweise der ausgestorbenen Tiere. So wurden einige verschiedene Hypothesen hervorgebracht, welche die Halswirbelsäule von einer sehr beweglichen, gekrümmten, Fragezeichen-ähnlichen Haltung bis hin zu einer sehr gerade gestreckten, fast unbeweglichen Haltung rekonstruierten. Hier stellen wir erstmals die Knochenmikrostruktur der Wirbel und der Halsrippen von Tanystropheus vor, um im Vergleich zu anderen ausgestorbenen (z.B. Dinosaurier) und rezenten Tieren (z.B. Vögel) Aussagen über Aufbau und Eigenschaften der Halswirbelsäule und deren Haltung zu machen. 28 6. Graduiertenforum Morphologie 2013 Ulm Integration morphologischer, knochenhistologischer und embryologischer Datensätze - drei paläobiologische Fallbeispiele (Schildkröten, Krokodile und marine Reptilien aus der Trias-Zeit) Torsten Scheyer * * Paläontologisches Institut und Museum, Karl Schmid-Strasse 4, Universität Zürich, CH-8006 Zürich, Schweiz Die Entdeckung und Erforschung fossiler Reptilien reicht bis ins 17. Jahrhundert zurück, wobei sich diese frühen Untersuchungen meist rein auf die äusserliche Anatomie der Knochen beschränkten. Auch die Knochenmikrostrukturanalyse und die Anfänge der vergleichenden Embryologie haben in dieser Zeit ihre Wurzeln, zu einer Integration der drei Teilgebiete kam es aber erst verstärkt im 20 Jahrhundert. Neben der Osteologie, erlaubt speziell die Untersuchung fossiler Knochenmikrostrukturen, im Vergleich zu denen heutiger Reptilien, die Gewinnung und Interpretation von ‚Life History‘-Daten, einer Informationsquelle die normalerweise nur in der Rezent-Biologie zur Verfügung steht. Daneben können, in seltenen Fällen, unter besonderen Fossilisationsbedingungen (Typ Konservatlagerstätte) auch Entwicklungsserien von Wirbeltieren zum Vergleich mit heute lebenden Reptilien herangezogen werden. Diese Arbeiten folgen in der Regel dem Sparsamkeitsprinzip, wonach näher verwandte fossile Reptilienarten nicht nur bestimmte gemeinsame äußerliche anatomische Merkmale aufweisen, sondern dass sie auch Charakteristiken in ihrer Knochenhistologie und Entwicklung gemein haben. Eine weitere Annahme ist, dass die aktuellen systematischen und funktionellen Studien von der Integration solch verschiedener Datensätze profitieren, da diese, ähnlich einem ‚Total Evidence‘-Ansatz, durch eine breitere und somit stabilere Datenbasis unterstützt werden. Anhand von drei Fallbeispielen verschiedener Wirbeltiergruppen wird aufgezeigt, wie die unterschiedlichen morphologischen Datensätze unterschiedlicher Betrachtungsebenen kombiniert werden können, um sowohl systematische Fragestellungen zu beantworten, als auch funktionale und ökologische Aspekte innerhalb der Evolution der Reptilien zu ergründen. Hierbei sollen speziell neuere Untersuchungen zum Ursprung der Schildkröten und der Entwicklung des Schildkrötenpanzers, der Evolution und Paläobiologie von ca. 245 Millionen Jahre alten, sekundär marinen Reptilien aus der Triaszeit, sowie die Untersuchung von jüngeren, ca. 6 Millionen Jahre alten, Krokodilfaunen aus der Neotropis vorgestellt werden. Zusammen mit einem besseren Verständnis über die Rolle der ausgestorbenen Tiere in vergangenen Ökosystemen erlauben die Untersuchungen der Fossilien somit einen Einblick in die allgemeinen Muster und Prozesse, welche der Evolution zugrunde liegen. 29 6. Graduiertenforum Morphologie 2013 Ulm Posterbeiträge 30 6. Graduiertenforum Morphologie 2013 Ulm Die Rollen von post-embryonalem Wachstum und sexueller Selektion in der Evolution sexual-dimorpher Extremetitäten in Amphipoden Stephan Handschuh* **, Hans L Nemeschkal**, Brian D Metscher**, Philipp Mitteröcker** & Gerd B Müller** *** * VetCore Facility for Research, University of Veterinary Medicine Vienna, Vienna, Austria ** Department of Theoretical Biology, University of Vienna, Vienna, Austria *** Konrad Lorenz Institute for Evolution and Cognition Research, Altenberg, Austria In dieser Studie wurden Aspekte der Evolution der Pereon-Extremitäten von Amphipoden (Crustacea, Malacostraca) mittels morphologischer Daten aus einer rezenten Population untersucht. Dafür wurden drei-dimensionale microCT Datensätze im mikroskopische Auflösungsbereich durch lineare Messstrecken quantifiziert. Das Sample bestand aus 200 microCT Scans von 100 Tieren (50 Präkopula-Paare) des invasiven Süßwassergammariden Dikerogammarus villosus (Sowinski, 1894). Es wurden sowohl die morphologische Integration von Extremtitäten im post-embryonalen Wachstum als auch die Korrelationsmuster zwischen verpaarten Individuen untersucht. Dadurch wird gezeigt, wie sexuelle Selektion zur evolutionären Modifikation der Rumpfextremitäten der Amphipoden beitragen kann. Der Fokus der Studie lag auf den beiden Gnathopodenpaaren (zweite und dritte Pereonextremtitäten). Diese sind bei D. villosus, wie bei vielen Amphipodenarten, sexual-dimorph. Die Gnathopoden erfüllen neben der Nahrungsaufnahme auch eine wichtige Rolle im Paarungsverhalten, wo sie dem Männchen während Präkopula-Amplexus und Kopula als Klammerorgane dienen. Wir zeigen, dass die Größe der männlichen Gnathopoden sexuell selegiert wird, und geben damit Hinweise zur Evolution dieser hoch dimorphen Strukturen. 31 6. Graduiertenforum Morphologie 2013 Ulm Tagmatisierung bei Fangschreckenkrebsen (Stomatopoda, Malacostraca, Crustacea) Verena Kutschera*, Joachim T. Haug**, Wafaa S. Sallam***, Andreas Maas*, Dieter Waloszek* & Carolin Haug** * Biosystematic Documentation, Ulm University, Helmholtzstr. 20, 89081Ulm, Germany ** Department Biology II, University of Munich, Großhaderner Str. 2, 82152 Planegg-Martinsried, Germany *** Department of Marine Science, Suez Canal University, Ismailia, 41522, Egypt Unter Tagmatisierung versteht man die Einteilung eines segmentierten Körpers in funktionelle Einheiten, sogenannte Tagmata. Die Identifizierung dieser Tagmata fußt auf vier Hauptkriterien, 1. der Verbindung von Segmenten, 2. ähnlicher Dorsalmorphologie, 3. ähnlicher Ventralmorphologie einschließlich der Anhänge und 4. der räumlichen Nähe der Segmente. Wir untersuchten Erugosquilla massavensis (Kossmann, 1880) und nutzten die Autofluoreszenz der Kutikula, um membranöse von sklerotisierten Bereichen zu unterscheiden. Damit wurden sechs Tagmata unterschieden: Tagma I, die sensorische Einheit; Tagma II, die vordere Nahrungsverarbeitungseinheit; Tagma III, die hintere Nahrungsverarbeitungseinheit; Tagma IV, die Laufbeineinheit; Tagma V, die Schwimm- und Atmungseinheit; und Tagma VI, der Schwanzfächer. Die segmentale Zusammensetzung der von uns erkannten Tagmata weicht allerdings stark von bisher propagiertem Wissen ab: Die sensorische Einheit umfasst neben Augen und erster Antenne auch die zweite Antenne, welche somit kein Teil der vorderen Nahrungsverarbeitungseinheit ist, wie bisher gedacht. Der erste Maxillipede gehört nicht zur vorderen Nahrungsverarbeitungseinheit sondern zur hinteren. Außerdem steht der fünfte Maxillipede den vorderen ventral räumlich sehr nahe, gehört somit zur hinteren Nahrungsverarbeitungseinheit. Dorsal gleicht dieses Segment jedoch sehr dem nachfolgenden, das zum Laufbeinabschnitt gehört. All dies wurde in dieser Form bisher noch nicht dokumentiert, trägt jedoch maßgeblich zum Verständnis der Evolution zu den heutigen Fangschreckenkrebsen (Verunipeltata) bei. 32 6. Graduiertenforum Morphologie 2013 Ulm Die Ventralseite des Peraeons bodenlebender Zehnfußkrebse (Decapoda: Reptantia) und seine Signifikanz für phylogenetische Deutungen Tobias Reich*, Verena Kutschera*, Andreas Maas* & Dieter Waloszek* * Biosystematische Dokumentation, Universität Ulm, Helmholtzstraße 20, 89081 Ulm Nach wie vor werden die Verwandtschaftverhältnisse innerhalb der reptanten Decapoda kontrovers diskutiert. Aus den letzten 20 Jahren gibt es zwei wesentliche Publikationen zur Phylogenie der Reptantia, die umfangreiche morphologische Daten verwenden. Beide, eine von Scholtz & Richter (1995), die andere von Dixon et al. (2003), weichen in einigen Punkten erheblich voneinander ab, da sie relevante Merkmale verschieden bewerten. Dies führt zu unterschiedlichen Stellungen bestimmter Taxa im System, im Wesentlichen betrifft es die Stellung der Langusten und Bärenkrebse (Achelata), der Flusskrebse (Astacida) und der Hummer (Homarida). Die maßgeblichen Merkmale beziehen sich dabei auf den Laufbeinabschnitt (Peraeon) der Tiere und vor allem auf die Beschaffenheit und den Verschmelzungsgrad der ventralen Brücken zwischen linken und rechten Laufbeinen (Sternite). Zur Unterscheidung von kalzifizierten und membranösen Bereichen der Cuticula wurde die Methode der Alizarinfärbung als besonders geeignet erachtet und auf insgesamt 20 Vertreter der Decapoda angewendet. Um die Veränderungen am Peraeon unabhängig vom Verschmelzungsgrad der Sternite beurteilen zu können, wurde zudem eine morphometrische Beurteilung der Proportionen des Peraeons durchgeführt. Die Ergebnisse unterstützen die Hypothese von Dixon et al. (2003), bei der Flusskrebse und Hummer einen gemeinsamen Ursprung haben und die Schwestergruppe aller übrigen Eureptantia bilden, während die Achelata die Schwestergruppe der Meiura (Mittelschwanzkrebse und Krabben) sind. Es ergaben sich demnach deutliche Hinweise darauf, dass die von Scholtz & Richter (1995) eingeführten Taxa Macrochelata und Fractosternalia obsolet sind. 33 6. Graduiertenforum Morphologie 2013 Ulm Die cephale Morpholgie von Tomocerus flavescens (Hexapoda: Collembola): Eine 3D-Rekonstruktion Peter Rühr*, Alexander Blanke* & Bernhard Misof* * Zoologisches Forschungsmuseum Alexander Koenig, Zentrum für Molekulare Biodiversitätsforschung, Adenauerallee 160, 53113 Bonn, Die Springschwänze (Collembola), eine primär flügellose Gruppe der Hexapoden, werden zur Gruppe der Entognatha (Collembola + Protura + Diplura) gezählt. Die drei Taxa sind unter anderem dadurch charakterisiert, dass die Mandibeln und Maxillen durch laterale Mundfalten in der Kopfkapsel eingeschlossen sind (Entognathie). Neuere Publikationen stellen die Monophylie der Entognatha in Frage. Daher werden von der detaillierten Erforschung der inneren und äußeren Kopfmorphologie, besonders der Mundwerkzeuge, des Endoskeletts und der Muskulatur, wichtige Hinweise auf die Evolution der Entognathie sowie die Verwandtschaftsverhältnisse der primär flügellosen Hexapodengruppen untereinander und zu den Ectognatha erwartet. Der erste Schritt, der nötig ist, um die Kopfmuskulatur und andere cephale Strukturen der Collembolen mit denen der Ectognatha phylogenetisch zu untersuchen, ist die Homologisierung dieser Strukturen. Die Kopfmuskulatur der drei entognathen Taxa konnte bisher nicht erfolgreich mit der der Ectognatha homologisiert werden. Synchrotron-micro-Computertomographie (SR-µCT) der cephalen Strukturen kann hierbei helfen die komplizierten Verhältnisse innerhalb des Collembolenkopfes sichtbar zu machen und Homologieannahmen über die klassischen Kriterien der Lage und spezifischen Qualität zu ermöglichen. Die Daten, die der dreidimensionalen Rekonstruktion des Kopfes von Tomocerus flavescens zu Grunde liegen, wurden mittels SR-µCT an der Swiss Light Source des Paul-Scherrer-Instituts erstellt. Anschließend erfolgte die Rekonstruktion mittels den frei erhältlichen Programmen Reconstruct und Blender. Die nicht destruktive und schnelle Technik der SR-µCT liefert extrem hochauflösende Scanbilder ohne Schnittartefakte. Der Vergleich der 3D-Rekonstruktion des Kopfes von T. flavescens mit umfassenden, früheren Arbeiten an ectognathen Insektenköpfen führt zu ersten Homologiehypothesen. Muskeln innerhalb des Collembolenkopfes könnten ein Homologon zu Muskeln ectognather Insekten darstellen. Dadurch könnte es jetzt möglich sein, die Muskulatur der Collembolen in bereits bestehende Forschungsarbeiten zu integrieren, die sich mit der Evolution der Kopfmuskulatur der Ectognatha beschäftigt. 34 6. Graduiertenforum Morphologie 2013 Ulm Notizen 35 6. Graduiertenforum Morphologie 2013 Ulm Notizen 36 6. Graduiertenforum Morphologie 2013 Ulm Teilnehmerliste Dieterich, Andreas Institut für Evolution und Ökologie Physiologische Ökologie der Tiere Eberhard Karls Universität Tübingen, Konrad-Adenauer-Straße 20, D-72072 Tübingen [email protected] Eulitz, Ute Museum of Zoology (Museum für Tierkunde) Senckenberg Natural History Collections Dresden, Königsbrücker Landstraße 159, D-01109 Dresden [email protected] Handschuh, Stephan VetCore Facility for Research Imaging Unit Veterinärmedizinische Universität Wien, Veterinärplatz 1, A-1210, Wien [email protected] Hannig, Leonie Physiologische Ökologie der Tiere, Institut für Evolution und Ökologie, Universität Tübingen, Konrad-Adenauer-Straße 20, D-72072, Tübingen [email protected] Haug, Joachim T. Department Biology II University of Munich LMU, Großhaderner Straße 2, D-82152, Planegg-Martinsried [email protected] Heethoff, Michael Evolutionary Biology of Invertebrates Institute for Evolution and Ecology University of Tübingen, Auf der Morgenstelle 28E, D-72076, Tübingen [email protected] Hörnig, Marie Ernst-Moritz-Arndt-Universität Greifswald, Zoologisches Institut und Museum, Cytologie und Evolutionsbiologie, Soldmannstraße 23, D-17487, Greifswald [email protected] Jaquier, Vivien Paläontologisches Institut und Museum, Universität Zürich, Karl Schmid-Strasse 4, CH-8006, Zürich [email protected] 37 6. Graduiertenforum Morphologie 2013 Ulm Teilnehmerliste Katzmann, Hans Peter Arbeitsgruppe Biosystematische Dokumentation, Universität Ulm, Helmholtzstraße 20, D-89081 Ulm [email protected] Kerbl, Alexandra Department of Biology, Marine Biology Section University of Copenhagen, Strandpromenaden 5, DK-3000, Helsingør, [email protected] Kruppert, Sebastian Department of Animal Ecology, Evolution & Biodiversity, Ruhr- University Bochum, Universitätsstraße 150, D- 44801, Bochum [email protected] Kutschera, Verena Arbeitsgruppe Biosystematische Dokumentation, Universität Ulm, Helmholtzstraße 20, D-89081, Ulm [email protected] Michalik, Peter Zoologisches Institut und Museum Ernst-Moritz-Arndt Universität Greifswald, J.-S.Bach-Str. 11/12, D-17489, Greifswald [email protected] Nagler, Christina Lehrstuhl für Aquatische Systembiologie, Wissenschaftszentrum Weihenstephan für Ernährung Landnutzung und Umwelt, Technische Universität München, Mühlenweg 22, D-85354, München [email protected] Nowak, Marlene Steinmann-Institut für Geologie, Mineralogie und Paläontologie, Nußallee 8, D-53115, Bonn [email protected] 38 6. Graduiertenforum Morphologie 2013 Ulm Teilnehmerliste Reich, Tobias Arbeitsgruppe Biosystematische Dokumentation, Universität Ulm, Helmholtzstraße 20, D-89081, Ulm [email protected] Rühr, Peter Zoologisches Forschungsmuseum Alexander Koenig (ZFMK) Leibniz-Institut für Biodiversität der Tiere - Zentrum für Molekulare Biodiversität (ZMB), Adenauerallee 160, D-53113 , Bonn [email protected] Sanmugaraja, Mayuran Steinmann-Institut für Geologie, Mineralogie und Paläontologie, Rheinische FriedrichWilhelms-Universität Bonn, , D-53115, Bonn [email protected] Scheyer, Torsten Paläontologisches Institut und Museum Universität Zürich, Karl Schmid-Straße 4, CH8006, Zürich [email protected] Schütz, Philipp Arbeitsgruppe Biosystematische Dokumentation, Universität Ulm, Albert-Einstein-Allee 11, D-89081, Ulm [email protected] Schwaha, Thomas Department of Integrative Zoology , Althanstraße 14, A-1090, Wien [email protected] van de Kamp, Thomas Karlsruhe Institue of Technology, Institue for Photon Science and Synchrotron Radiation, Hermann-von-Helmholtz-Platz 1, D-76344, Eggenstein-Leopoldshafen [email protected] 39 6. Graduiertenforum Morphologie 2013 Ulm Teilnehmerliste Walther, Paul Zentrale Einrichtung Elektronenmikroskopie, Universität Ulm, Albert-Einstein-Allee 11, D-89081, Ulm [email protected] 40 6. Graduiertenforum Morphologie 2013 Ulm Übersichtsplan Universität 41 6. Graduiertenforum Morphologie 2013 Ulm Übersichtsplan Stadt © OpenStreetMap.org 42 6. Graduiertenforum Morphologie 2013 Ulm Wichtige Adressen Tagungsort Hörsaal 16, Universität Ost Universität Ulm Albert-Einstein-Allee 11, 89081 Ulm Konferenzdinner Barfüßer Brauhaus Neu-Ulm Paulstraße 4, 89231 Neu-Ulm Treffpunkt Stadtführung Stadthaus Ulm Münsterplatz 50, 89073 Ulm Exkursion Münster Ulm Münsterplatz 21, 89073 Ulm Tiergarten Ulm Friedrichsau 40, 89073 Ulm 43 6. Graduiertenforum Morphologie 2013 Ulm Empfohlene Busverbindungen Die Fahrzeit vom Theater zur Universität beträgt ca. 10 Minuten, der Fußweg von der Haltestelle Universität Süd zum Tagungshörsaal bei konservativer Schätzung 5 Minuten. Busse am Theater fahren in der Regel im 10 Minutentakt, mit zeitlichem Versatz. Da am Samstag ein anderer Fahrplan als unter der Woche gilt führen wir unten die empfohlenen Busverbindungen auf. Donnestag 10.10.2013 Abfahrtszeit Ankunftszeit Linie Richtung 17:33 17:42 3 Wissenschaftsstadt 17:43 17:52 3 Wissenschaftsstadt 17:53 18:02 3 Wissenschaftsstadt 18:03 18:12 3 Wissenschaftsstadt 18:13 18:22 3 Wissenschaftsstadt Abfahrtszeit Ankunftszeit Linie Richtung 08:33 08:43 3 Wissenschaftsstadt 08:38 08:48 3 Science Park II 08:43 08:53 3 Wissenschaftsstadt 08:48 08:58 3 Science Park II 08:53 09:03 3 Wissenschaftsstadt Abfahrtszeit Ankunftszeit Linie Richtung 08:17 08:26 3 Wissenschaftsstadt 08:32 08:41 3 Wissenschaftsstadt 08:47 08:56 3 Wissenschaftsstadt 09:02 08:11 3 Wissenschaftsstadt 09:17 09:26 3 Wissenschaftsstadt Freitag 11.10.2013 Samstag 12.10.2013 44 Grafiken, Layout & Design: Hans Peter Katzmann, Verena Kutschera Umschlagbilder: Joachim T. Haug, Hans Peter Katzmann, Torsten Scheyer