Nervenzelle und AP
Werbung

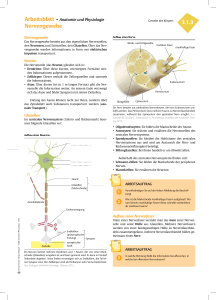
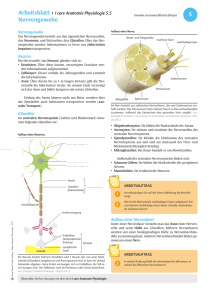
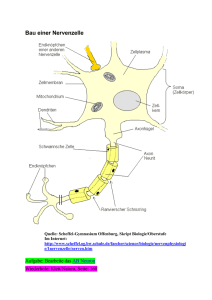
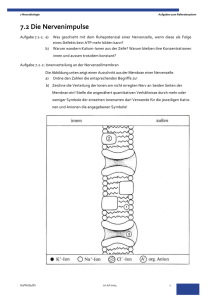
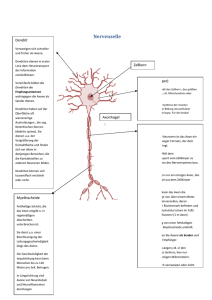
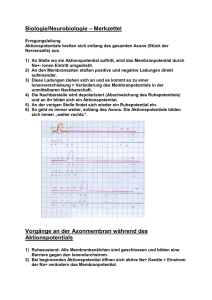
1. Bau der Nervenzelle Die Nervenzelle, auch als Neuron bezeichnet, setzt sich zusammen aus einem etwa 0,25 mm großen Zellkörper (Soma) und mehreren Fortsätzen, den Dendriten. Im Zellkörper befindet sich der Zellkern, der die Steuerung übernimmt und die Erbanlagen enthält. Die aus dem Zellkörper herausragenden Dendriten sind kurz und stark verästelt. Einer dieser Fortsätze kann sehr lang werden und wird als Nervenfaser oder Axon bezeichnet. Dieses Axon kann beim Menschen bis zu 1 m lang werden. Axone werden von Schwanschen Zellen umgeben. Da diese jedoch kürzer sind als die Axone, sind mehrere aneinandergereiht. Die Markscheiden sind an das Axon gereiht, wie Perlen an einer Schnur. Sie dienen als Isolierschicht. Die vorhandenen Zwischenräume werden als Ranviersche Schnürringe bezeichnet. Zwei dieser hintereinanderliegenden Zwischenräume haben einen Abstand von 1-2 mm. Es wird bei den Axonen unterschieden zwischen denen mit Markscheide, genannt markhaltige Nervenfasern, und denen ohne Markscheide, bezeichnet als marklose Nervenfasern. Weiterhin gibt es Verbindungsstellen zwischen mehreren Nervenzellen, sowie zwischen Nervenzellen und Muskelfasern oder Drüsenzellen. Diese Berührungsstellen heißen Synapsen. Die von Hüllen umgebenen Neuriten bezeichnet man als Nervenfasern. Ein Bündel solcher Nervenfasern bildet den Nerv, welcher die Erregung von den Sinneszellen über das Zentralnervensystem zu den Erfolgsorganen leitet. 2. Funktion der Nervenzelle 2.1. Grundlegende Aufgabe der Nervenzelle Das Neuron ist eine funktionelle Einheit des Nervensystems. Der Reiz, den das Neuron erfährt, ist eine physikalische oder chemische Einwirkung der Zelle aus der Umwelt oder dem Körperinneren. Dieser Reiz wird von Sinneszellen aufgenommen und an die Nervenzelle weitergegeben. Wenn es erregt wurde, hat das Neuron die Aufgabe den Reiz weiterzuleiten und zu verarbeiten. Das Nervensystem ist mit den Organen bzw. Organsystemen verbunden. Es steuert ihre Tätigkeit und sorgt für ihr Zusammenspiel. Es gibt verschiedene Nervenzelltypen für verschiedene Funktionen. Die Informtionen über die Sinnesorgane laufen über sensorische Nervenbahnen. Die Informationen zur Ausführung von Reaktionen laufen über motorische Bahnen. Diese beiden Typen bezeichnet man als das periphere Nervensystem. Das vegetative Nervensystem steuert die Arbeit der inneren Organe. 2.2. Erregungsleitung im Axon ohne Markscheide Die Erklärung des Erregungsverlaufes im Axon verläuft hier am Beispiel des Tintenfisches. Das Axon besitzt im Ruhezustand ein Membranpotential von ca. -90 mV. Im Labor ist es möglich durch Anlegung einer schwachen Spannung das Membranpotential auf -50 mV zu erhöhen. Diese Steigerung wird als Depolarisation bezeichnet und die Senkung des Mempranpotentials wird Hyperpolarisation genannt. Die Membran des Axons besitzt ebenfalls ähnliche Eigenschaften wie die eines Kondensators. Sie besitzt eine geringe Leitfähigkeit und trennt dabei die beiden intrazellulären und extrazellulären Flüssigkeiten, die auch als elektrische Leiter dienen. Je geringer der Abstand der beiden Flüssigkeiten und je größer die Fläche mit der sie an den Nichtleiter stoßen, um so größer die Kapazität des ´Kondensators´, das heißt das Fassungsvermögen für elektrische Ladungen. Dies bedeutet dicke Axone besitzen aufgrund ihrer großen Membranoberfläche eine große Kapazität. Je größer die oben beschriebene künstlich angelegte Spannung, je stärker ändert sich das Membranpotential über einen bestimmten Wert. Dieser Wert wird als Schwellenwertbezeichnet. Wird dieser Schwellenwert überschritten, kehrt sich die Ladungsverteilung an der Membran kurzzeitig um. Diese Spannungsänderung dauert etwa 1 ms an und wird als Aktionspotential, spike oder impuls bezeichnet. Das hier geltende ´Alles oder nichts´Gesetz besagt, daß es nur die Möglichkeit des Membranpotentials über dem Schwellenwert oder unter diesem gibt, eine 3.Variante ist nicht möglich. Die Höhe des Aktionpotentials ist unabhängig von der höhe und Dauer der angelegten Spannung. Das entstandene Spike wandert über das Axon entlang zum Axonende. 2.2.1. Ursachen des Aktionspotentials In der Membran des Axons befinden sich substratspezifische Poren, die nur für Natrium- bzw.Kaliumionen durchlässig sind. Diese Durchlässigkeit ist abhängig vom oben erklärten Membranpotential, das heißt, daß es sich hier um spannungsgesteuerte Natrium- und Kaliumporen handelt. In der Ruhe sind die Natriumporen geschlossen und die Kaliumporen zum Teil offen. Kommt es zur Depolarisation, öffnen sich einige Natriumporen. Jedoch sind immer noch mehr Kaliumporen geöffnet als Natriumporen. Steigt die Spannung über den Schwellenwert, so öffnen sich alle Natriumporen. Die Anzahl der Kaliumporen bleibt beim gesamten Prozeß gleich. Daraus folgt, daß mehr Natriumionen nach innen strömen als Kaliumionen nach außen. Das Innere des Axons ist kurzzeitig positiv geladen. Nun schließen sich die Natriumporen und alle Kaliumporen öffnen sich. Das Membranpotential sinkt nun wieder auf den Ruhewert. Der Versuch das Axon an dieser Stelle wieder künstlich zu depolarisieren schlägt fehl, da diese Stelle für einen kurzen Moment unerregbar ist. Dieser Zustand, der als refraktärbezeichnet wird, tritt auf, da der Kondensator eine gewisse Zeit braucht, um sich zu be- und entladen. Bei einer sehr starken Erregung öffnen sich die Natriumporen nach einer absoluten Refraktärzeit von 1-2 ms wieder. 2.2.2. Weiterleitung des Aktionspotentials Wenn an einer Stelle A ein Reiz ausgelöst wird, entsteht dort ein Aktionspotential und positive und negative Ladungen grenzen ohne trennende Membran aneinander. Da sich die unterschiedlichen Ladungen anziehen, kommt es zu Ionenströmen, sogenannten Ausgleichströmungen. Diese senken das Membranpotential des Nachbarn. Wird auch diese Stelle bis unter den Schwellenwert depolarisiert, kommt es auch hier zu einem ´Spike´. Diese neu gereizte Stelle senkt ihrerseits wieder die benachbarte Stelle, so daß auch Stelle A wieder die alte Polarität besitzt. Jedoch kommt es hier nicht mehr zum Aktionspotential wegen der oben beschriebenen Unerregbarkeit. Bei der Weiterleitung der Erregung ist zu beachten, das dickere Axone den Schwellenwert früher erreichen als dünnere, da hier ein geringerer elektrischer Widerstand des Innenmediums vorherscht. Das heißt dickere Axone leiten die Erregungen schneller als dünnere. 2.3. Erregungsleitung im Axon mit Markscheide An den Stellen an denen das Axon eine Markscheide besitzt, findet eine andere Leitung statt als an Stellen ohne Markscheide. Da hier keine Natriumporen vorhanden sind, können sich auch keine Impulse ausbilden. Nur an den Schnürringen zwischen zwei benachbarten Markscheiden können Aktionspotentiale entstehen. Von einer erregten Stelle A gehen wieder Ausgleichsströmchen aus, die sich zum nächsten Schnürring fortpflanzen, so daß auch dieser polarisiert wird. Die Stellen, die mit der Markscheide umhüllt sind, haben eine sehr geringe Membrankapazität, weil der Abstand zwischen Innen- und Außenmedium sehr groß ist. Deshalb kann dieser Teil des Axons schnell entladen werden. Die Erregung pflanzt sich hier sehr schnell fort (bis zu 120 m/s). An den Schnürringen ist wieder ein geringer Abstand des Außen- und Innenmediums, so daß die Erregung immer von Schnürring zu Schnürring springt. Dieser Vorgang wird als saltatorische Erregungsleitung bezeichnet. Beim Axon mit Markscheide beträgt die Übertragungsgeschwindigkeit etwa 25 m/s. Der Vorteil der Markscheide ist also eine schnellere Leitung der Erregung, eine erhebliche Materialersparnis und ein geringerer Energiverbrauch. Die restlichen Vorgänge sind gleich denen im Axon mit Markscheide.