Imagebroschüre MPG-ASMB - Max-Planck
Werbung

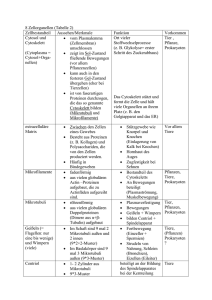
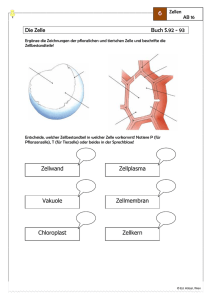
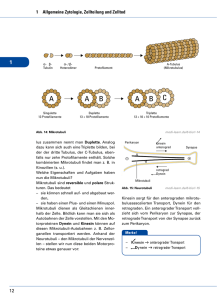
Max-Planck-Arbeitsgruppen für strukturelle Molekularbiologie Forschungsthemen im Überblick Die Max-Planck-Arbeitsgruppen wurden 1985 gegründet, um die am Deutschen Elektronensynchrotron (DESY) vorhandene Synchrotronstrahlung für die Untersuchung der Struktur und Dynamik von Biomolekülen einzusetzen. Die Kenntnis der Struktur von Biomolekülen ist wesentliche Voraussetzung für das Verständnis von biologischen Prozessen auf molekularer und zellulärer Ebene, sowie für Anwendungen in Biotechnologie, Medizin oder Pharmazie. Die bei DESY vorhandene Synchrotronstrahlung erlaubt es wegen ihrer hohen Intensität und variablen Wellenlänge, Röntgenstrukturanalysen von Biomolekülen schnell, schonend und mit hoher Auflösung durchzuführen. Nur so gelingt es, den steigenden Bedarf an Strukturanalysen zu befriedigen und die Methoden einem internationalen Kreis von Nutzern zur Verfügung zu stellen. Dazu werden Techniken der Strahlführung, Probenbehandlung, Datenerfassung und -verarbeitung entwickelt. Durch die Aufklärung des Genoms des Menschen und anderer Organismen, sowie durch den Bedarf an rekombinanten Proteinen in Biotechnologie und Medizin wird die Nachfrage nach Strukturanalysen stark zunehmen. Diese wird nur durch die verstärkte Nutzung der Synchrotronstrahlung befriedigt werden können. Dies ist besonders wichtig vor dem Hintergrund des bei DESY geplanten freien Elektronenlasers (TESLA-FEL) und des Ausbaus des Speicherrings PETRA-III für Synchrotronexperimente. Diese Anlagen werden in einigen Jahren die weltweit stärksten Quellen von Röntgenlicht darstellen. Hier fehlt noch das Hintergrundbild! Die drei Arbeitsgruppen bearbeiten unterschiedliche biologische Forschungsprojekte: Die Gruppe Ribosomenstruktur beschäftigt sich mit der Aufklärung der Struktur und Funktion des Ribosoms, eines Aggregats aus vielen Proteinen und Nukleinsäuren, das in der lebenden Zelle für die Herstellung neuer Proteine (Proteinbiosynthese) und damit für die Übersetzung der Erbinformation verantwortlich ist. Wegen seiner Komplexität ist die Analyse von Ribosomen ein Testmodell für die Leistungsfähigkeit der Kristallographie mit Synchrotronstrahlung. Bei der Gruppe Proteindynamik geht es um Reaktionsmechanismen von Enzymen und um schnelle Konformationsänderungen bei Proteinen, die die biologischen Funktionen bestimmen. Ein Schwerpunkt ist die „zeitaufgelöste Proteinkristallographie“, bei der die Intensität der Synchrotronstrahlung eingesetzt wird, um Strukturveränderungen im Mikro- bis Nanosekundenbereich zu erfassen. Die Gruppe Zytoskelett befasst sich mit den Proteinfasern der Zelle, speziell mit Mikrotubuli und Motorproteinen, die für Zellbewegung, Zellteilung und Differenzierung wichtig sind. Von besonderer Bedeutung ist hier das Tau-Protein, das bei der Alzheimer-Krankheit in den Nervenzellen aggregiert und ein Merkmal für ihre Degeneration darstellt. Hier fehlt noch das Hintergrundbild! Ribosomen: Proteinfabriken der Zelle Die Erzeugung von Proteinen ist für jeden Organismus existenziell. In der Zelle erfolgt die Produktion der Eiweiße im Rahmen der Protein-Biosynthese. Dieser komplexe Prozess besteht aus einer Vielzahl individueller Schritte, unter Beteilung vieler unterschiedlicher Helfer oder Co-Faktoren, die wiederum zum größten Teil nichts anderes als eben solche Proteine sind. Zentrales Element der ProteinBiosynthese ist jedoch das Ribosom. Das Ribosom ist – an biologischen Maßstäben gemessen – ein gewaltiges Molekül, mit einem Molekulargewicht von mehr als 2 Millionen Dalton. Bis zu 50.000 dieser Zellorganellen, die mehr als 80% der Zellenergie konsumieren, kann eine Zelle beherbergen. Das Ribosom setzt sich immer aus zwei verschieden großen ribosomalen Untereinheiten zusammen, die im Falle des prokaryontischen Ribosoms auch als 30S und 50S Untereinheiten bezeichnet werden. Beide Untereinheiten, die jeweils für spezifische Aufgaben in der Proteinsynthese verantwortlich sind, bestehen zu etwa 2/3 aus ribosomaler RNA und einer verschiedenen Anzahl ribosomaler Proteine. Der modulare, flexible Aufbau und die räumliche Entkopplung der verschiedenen molekularen Mechanismen machen das Ribosom zu einer außerordentlich effizienten Proteinfabrik. Der erste Schritt der Proteinsynthese ist die Initiation, die im Wesentlichen die mRNA für das Ribosom verfügbar macht, also die fehlerfreie Übersetzung und Verifizierung des genetischen Codes durch die kleine (30S) Untereinheit einleitet. Das Start-Codon der mRNA bindet die komplementäre Initiator-tRNA, die die erste Aminosäure des zu erzeugenden Proteins bereit stellt. In der Folge werden die verschiedenen Aminosäuren von den jeweiligen tRNA-Molekülen geliefert und entsprechend dem genetischen Bauplan miteinander verknüpft (Elongation). Diese fundamentale Reaktion wird durch die 50S Untereinheit, oder genauer durch deren 23S rRNA, im Peptidyl-Transferase-Zentrum (PTC) katalysiert. Das Ende der Proteinsynthese wird dem Ribosom schließlich durch ein Stop-Codon auf der mRNA signalisiert, der Prozess mithilfe verschiedener Release- und Termination-Faktoren beendet (Termination) und die ribosomalen Untereinheiten für weitere Aufgaben wiederverwertet. Die Proteinsynthese ist gleichzeitig ein bevorzugtes Ziel einer großen Zahl verschiedener Antibiotika, die diesen Prozess zum Erliegen bringen, und so eine Zelle, vorzugsweise eines Erregers, eliminieren. Um die zugrunde liegenden Mechanismen der Proteinsynthese - aber auch deren Hemmung durch Antibiotika - zu verstehen, muss man den Aufbau des Ribosoms und dessen Wechselwirkungen mit verschiedenen Substraten möglichst präzise bestimmen. Die Struktur der 30S Untereinheit von Thermus thermophilus (Bild links) gibt Aufschluss über die Prozessierung der mRNA, also der Übersetzung des genetischen Codes. Pharmakologisch nutzvoll sind Untersuchungen an Komplexen mit Antibiotika, wie zum Beispiel Edeine und Tetracycline (rechts). Edeine ist eines der Antibiotika, die eine spezifische Konformation des Ribosoms einfrieren, und so die notwendige Mobilität unterbinden. Edeine bindet an einer phylogenetisch konservierten Region, also einem Bereich des Ribosoms, der in allen Organismen praktisch identisch ist. Als Folge stört Edeine die Proteinsynthese nicht nur in bakteriellen Erregern, sondern in jeder Zelle, was dieses Antibiotikum als Medikament weitgehend unbrauchbar macht. Tetracycline, eines der ersten Breitband-Antibiotika, bindet im Gegensatz zu Edeine an vielen verschiedenen Stellen innerhalb des Ribosoms. Primär wichtig aber ist ein Tetracycline-Molekül, das die Positionierung der tRNA an die Dekodierungs-Stelle und somit den Aufbau des Proteins verhindert. Die ersten Versuche, Ribosomen zu kristallisieren, wurden bereits in den frühen 1980ern unternommen, aber erst in den letzten Jahren durch die Bestimmung der Röntgenstrukturen von Erfolg gekrönt. Alle Ribosomen, die bislang kristallisierbar waren, entsprangen sehr robusten Organismen. Exemplarisch ist Deinococcus radiodurans (unten links), auch bekannt als Conan the Bacterium, das selbst extreme Strahlung überstehen kann. Ursprünglich aus verdorbenem Dosenfleisch isoliert, dient es heute zur Dekontaminierung hochgiftiger Stoffe - und soll nach Plänen der NASA demnächst den Mars besiedeln. Die 50S Untereinheiten dieses ansonsten harmlosen Bakteriums haben so gute und stabile Kristalle geliefert, dass die Interpretation der Elektronendichtekarte (mitte links) ein zu 99% vollständiges Bild der Struktur (rechts) mit einer Auflösung von 3Å ergab. Weitere Komplexe mit verschiedenen Substraten und Co-Faktoren haben es erlaubt, die Geheimnisse der Proteinsynthese, der katalytischen Aktivität der Ribosomen weitgehend zu lüften. Das katalytische Zentrum des Ribosoms ist zudem Angriffspunkt wichtiger Klassen von Antibiotika, so dass diese Ergebnisse damit auch den Weg zur Entwicklung dringend benötigter, neuer Antibiotika ein wenig geebnet haben. Mit der Verknüpfung der einzelnen Aminosäuren zu einer linearen Polypeptidkette ist allerdings noch kein funktionelles Protein gewonnen. Dies erfordert die Faltung der linearen Kette in ihre komplexe dreidimensionale Struktur. Damit das ungefaltete Protein nicht vorzeitig beschädigt wird, wird es durch den ribosomalen Exit Tunnel der 50S Untereinheit gefädelt (Bild links). Am Eingang des Tunnels wirken die Makrolide-Antibiotika, Erythromycin und dessen semi-synthetische Abkömmlinge. Die Makrolide blockieren den Eingang zum Tunnel (rechts oben), und verhindern so die Passage der Polypeptidkette. Wer den Hamburger Elbtunnel durchqueren muss, erfährt bisweilen, wie wirkungsvoll dieser Mechanismus sein kann. Das Ribosom kann also auch in der Anwesenheit der Makrolide noch eine kurze Peptidkette synthetisieren. Manche Zellen nutzen diese Eigenschaft zur Resistenz mittels der Produktion von Penta-Peptiden, die die Makrolide bildlich aus dem Tunnel heraus bürsten, und die Aktivität des Ribosoms wieder herstellen. Eine der neuesten Makrolide-Entwicklungen ist Azithromycin, das sich durch besondere pharmakokinetische Eigenschaften auszeichnet. Es ist auch in anderer Hinsicht speziell, da es bislang das einzige Antibiotikum ist, das zwei eng miteinander verbundene Bindungsstellen aufweist, und so die eigene Wirkung potenziert (unten rechts). Die Struktur der 50S Untereinheit mit einem anderem Makrolid, Troleandomycin, wies eine unerwartete Änderung der Struktur des Tunnels auf. Normalerweise läuft Protein L22 an der Wand des Tunnels entlang (Bild oben), durch die Bindung von Troleandomycin wird jedoch eine Konformationsänderung von L22 induziert, dessen Loop nunmehr quer durch den Tunnel weist, und an der gegenüber liegenden Tunnelwand eine neue Bindung eingeht (mitte), und somit den Tunneldurchgang stark verengt. Dieses Ergebnis weist auf einen universellen Mechanismus hin: Bei der Produktion des Secretory Monitoring Proteins (SecM) legt das Ribosom eine Pause ein, und zwar nur, wenn eine spezifische Aminosäure-Sequenz innerhalb des Tunnels im Bereich von L22 liegt. Diese vorübergehende Unterbrechung der Proteinsynthese ist ein wichtiger Regulations-Mechanismus, der ausschließlich durch die Struktur des Tunnels bestimmt wird. Modifiziert man einige Aminosäuren innerhalb des Loop, so dass L22 nicht mehr in der Lage ist, die alternative Konformation einzunehmen, so findet auch die Unterbrechung der Proteinsynthese nicht mehr statt, und SecM wird unabhängig von der Sequenz kontinuierlich erzeugt. Die Vermutung liegt nahe, dass L22 verantwortlich für die Regulation von SecM ist (unten). Proteindynamik: Enzyme in Aktion Viele biologische Prozesse erfordern zu ihrem Verständnis die Kenntnis der räumlichen Struktur der beteiligten Proteine und ihrer Änderung im Verlauf der Reaktionen. Ein Beispiel, das sowohl für Grundlagenforschung als auch für industrielle Pharmaforschung von Bedeutung ist, sind enzymatische Reaktionen. Die Bestimmung des dreidimensionalen Aufbaus der Enzyme ist eine Voraussetzung zur Untersuchung ihrer Wirkungsweise und zur gezielten Entwicklung medizinischer Wirkstoffe. Dazu müssen die Lagen der einzelnen Atome dieser Makromoleküle mit einer Genauigkeit von etwa einem Zehntel eines Atomdurchmessers bestimmt werden. Dieses Ziel kann mit Hilfe der Kristallstrukturanalyse erreicht werden. Hierzu müssen Proteinkristalle gezüchtet und in einer Serie von Röntgenbeugungsaufnahmen untersucht werden. Die MPG betreibt zu diesem Zweck eine Messstation an einem DESY-Synchrotron, das die erforderliche extrem intensive Röntgenstrahlung zur Verfügung stellt. Die kristallographische Auswertung der äußerst umfangreichen Messdaten erfolgt mit Hilfe eines leistungsfähigen Verbundes von Rechnern. Die Forschung mit Synchrotronstrahlung ist eines der dynamischsten Gebiete der molekularen Strukturbiologie und anderer experimenteller Naturwissenschaften. Durch die Weiterentwicklung der vorhandenen Synchrotronstrahlungsquellen und den Bau eines Röntgenlasers (FEL) nimmt das DESY einen Spitzenplatz im weltweiten Vergleich ein. Bestimmung der dreidimensionalen Struktur von Proteinen Aus den Beugungsaufnahmen kann durch mathematische Verfahren die Elektronendichteverteilung berechnet und die Lage der Atome bestimmt werden. Die dazu durchgeführte Fouriertransformation erfordert insbesondere die Kenntnis der Phasen von Strukturfaktoren. Dieses zentrale Problem der Kristallstrukturanalyse kann mit Hilfe der anomalen Dispersion und der Durchstimmbarkeit der Synchrotronstrahlung gelöst werden. Das Ergebnis dieses „MAD-Verfahrens“ ist ein Strukturmodell, das die Grundlage für weitere Untersuchungen wie etwa die der Bindung von Reaktionspartnern bildet. Die Methoden der Datensammlung und –auswertung werden zur Zeit weiter beschleunigt und automatisiert. Dies ermöglicht die schnelle Bestimmung von Proteinstrukturen, etwa für Strukturgenomik. Auch hochkomplexe Multiproteinstrukturen können so bis zu hoher Auflösung bestimmt werden, z.B. die des Proteasoms, das in der Zelle falsch gefaltete oder sonstwie anomale Proteine beseitigt. Das 20S-Proteasom von Hefe, dessen Struktur aufgrund von Messungen an der Beamline der MPG-ASMB aufgeklärt wurde, besteht aus 14 verschiedenen Proteinen mit einem gesamten Molekulargewicht von 700.000. Es enthält etwa 45.000 Atome, wobei Wasserstoffatome nicht eingerechnet sind. Für die Messungen werden im allgemeinen Kristalle mit Kantenlängen um 0.1 mm benötigt. Zur Verringerung von Strahlenschäden werden viele Experimente bei tiefen Temperaturen (100 K) durchgeführt. Im Falle hoher kristalliner Ordnung können extrem hohe Auflösung (< 1 Å) und äußerst geringe Fehler in den Atomlagen erreicht werden. Untersuchung dynamischer Vorgänge in Proteinen Die Wirkungsweise von Proteinen auf molekularer Ebene ist in der Regel mit transienten Änderungen ihrer dreidimensionalen Struktur verknüpft. So treten etwa bei enzymatischen Reaktionen Konformationsänderungen auf, die für die Spezifizität, Richtung und Geschwindigkeit der Biokatalyse wesentlich sind. Die Packung der Proteinmoleküle im Kristall, die große mit Solvent gefüllte Zwischenräume aufweist, ermöglicht eine Untersuchung der Kristallstruktur in verschiedenen Reaktionszuständen des Proteins. Je nach Charakteristik der Reaktion kann sie durch Eindiffusion von Reaktionspartnern oder durch externe Anregung, etwa mittels Laserphotolyse, im Kristall gestartet werden. Intermediäre Konformationen können auf der Grundlage von Schockfrieren oder mit Verfahren der zeitaufgelösten Röntgenbeugung untersucht werden. Bei zeitaufgelöster Proteinkristallographie sind bisher Zeitskalen bis in den Nanosekundenbereich erreicht worden; der Röntgen-FEL wird in Zukunft die Anwendungsgrenzen hin zu wesentlichen kürzeren Zeiten (Femto- bis Pikosekunden) und komplexeren Strukturen erweitern. Durch Auswertung der diffusen Streuung von Proteinkristallen kann im Prinzip Information über korrelierte Bewegungen im Proteinmolekül bzw. in der molekularen Packung erhalten werden. Komplementär zu experimentellen Beugungs- und Streuverfahren werden theoretische Verfahren zur Simulation der Moleküldynamik eingesetzt. Ausgangspunkt ist dabei die durch die Kristallstrukturanalyse bestimmte dreidimensionale Struktur. Zytoskelett: Architektur und Bewegung der Zelle Die Arbeitsgruppe „Zytoskelett“ beschäftigt sich mit dem Innengerüst der Zelle, das für die Form von Zellen, für die Bewegung, für Materialtransport, für Zellteilung und Zelldifferenzierung verantwortlich ist. Das Zytoskelett wird aus drei Fasersystemen aufgebaut, den Aktinfilamenten, Intermediärfilamenten und Mikrotubuli. Diese Fasern können sich aus ProteinUntereinheiten selbständig auf- und wieder abbauen, d.h. sie sind dynamisch, und sie haben die Fähigkeit der „Selbstorganisation“. Hinzu kommen zahlreiche Proteine, die an die Fasern andocken können, die Fasern miteinander verbinden oder Verbindungen mit anderen Zellkomponenten herstellen können, z. B. der äußeren Zellmembran oder der Kernmembran. Eine wichtige Klasse dieser assoziierten Proteine sind die „Motorproteine“. Sie können chemische Energie (in Form von Adenosintriphosphat) in mechanische Energie umwandeln und dadurch Lasten transportieren oder verschieben (z. B. Zellorganellen wie Mitochondrien, oder andere Komponenten des Zytoskeletts). Die Zytoskelettfasern werden durch verschiedene „assoziierte Prote- Abbildung rechts: Mikrotubuli bilden ein dynamisches Korsett der Zelle, das sich ständig den wechselnden Anforderungen der Zelle anpassen muss. Die Abbildung zeigt im Hintergrund eine Momentaufnahme von Mikrotubuli, die innerhalb einiger Millisekunden eingefroren wurden und so in ihrem Zustand festgehalten wurden (Aufnahme mit Kryo-Elektronenmikroskopie). Wachsende Mikrotubuli haben gerade Enden, während zerfallende Mikrotubuli am Ende gekrümmt und ausgefranst sind. Das Nebeneinander von Auf- und Abbau ist typisch für den Zustand von Zellen zwischen zwei Teilungen. Es ist aber auch möglich, alle Mikrotubuli in einer Lösung zu synchronisieren, so dass sie gleichzeitig zwischen Phasen des Wachstums und des Zerfalls hin- und herschwingen. Dies stellt einen biochemischen Oszillator dar, der durch den Verbrauch von energiereichen Phosphatverbindungen (Guanosintriphosphat) in Schwung gehalten wird. Rechts unten ist eine Serie von Röntgenbeugungsexperimenten zu sehen, bei denen die oszillierenden Mikrotubuli mit einer zeitlichen Auflösung von einigen Sekunden verfolgt wurden. Aus der Form der Kurven kann man die Struktur der Zwischenzustände berechnen. In lebenden Zellen werden solche Strukturumwandlungen häufig durch chemische Signalübertragungsketten reguliert. Das Bild rechts oben zeigt einen „Strauß“ von Mikrotubuli (aufgenommen mit Video-Dunkelfeldmikroskopie), die von einem gemeinsamen Zentrum auswachsen und durch ein Zusatzprotein (Tau-Protein) stabil gehalten werden. Wenn das Tau-Protein abgelöst wird (indem eine Phosphatgruppe übertragen wird), werden die Mikrotubuli instabil, der Strauß von Mikrotubuli bricht zusammen. Das Enzym, das diesen Übergang einleitet (eine sog. Protein-Kinase), besitzt in manchen pathologischen Zuständen der Zelle eine erhöhte Aktivität (z. B. in erkrankten Neuronen der Alzheimerkrankheit). ine“ stabilisiert. Ein bekanntes Beispiel für das Zusammenspiel verschiedener Zytoskelett-Proteine ist die Zellteilung; dabei bilden Mikrotubuli einen Spindelapparat aus, der die Chromosomen mit Hilfe von Motorproteinen auseinanderziehen kann. Bei den Nervenzellen stellen Mikrotubuli die „Gleise“ für den Transport in den Zellfortsätzen (Axone, Dendriten) dar, wo Lasten über weite Entfernungen zu den Nervenendigungen (Synapsen) geliefert werden müssen. Die Abbildung links stellt die Ergebnisse der Strukturuntersuchung des Motorproteins Kinesin dar. Das Strukturmodell (links unten) zeigt einen doppelköpfigen Protein-Komplex, bei dem die beiden Köpfe die Funktion der „Motoren“ übernehmen; sie sind durch eine „Deichsel“ miteinander verbunden (doppelte alpha-Helix, rot). Der obere Bildteil veranschaulicht, wie die beiden Motor-Köpfe des Kinesins auf dem Gleis eines Mikrotubulus entlangwandern, indem sie abwechselnd an die Protein-Untereinheiten andocken (blau/grün; das Gleis ist hier treppenförmig unterbrochen, um verschiedene Phasen der Bewegung zu unterscheiden). Damit wird eine Last von links nach rechts gezogen (am oberen Ende der Doppelhelix, hier nicht gezeigt). Obwohl die Moleküle nur wenige Nanometer Durchmesser haben, kann man die Bewegung mit Hilfe der Videomikroskopie in Echtzeit sichtbar machen (Modell rechts unten). Dazu bringt man einen „Rasen“ von doppelköpfigen Motorproteinen auf einem Mikroskopierglas auf und lässt einen Mikrotubulus (violett) darübergleiten, oder man belädt ein Vesikel (rosa) mit einem Motor und läßt es am Mikrotubulus in Pfeilrichtung entlangwandern. Das Verständnis solcher „Nanomaschinen“ könnte in Zukunft dazu beitragen, Schaltelemente oder Motoren für nanotechnologische Anwendungen zu entwickeln. Entsprechend ihrer grundlegenden Funktion für die „Infrastruktur“ der Zelle spielen Proteine des Zytoskeletts bei verschiedenen Krankheitsprozessen eine Rolle. So besteht die KrebsKrankheit aus einer unkontrollierten Teilung von Zellen. Die Wirkung einiger Zytostatika beruht darauf, die Mikrotubuli oder Motorproteine der Teilungsspindel zu vergiften, so dass sich die Krebszellen nicht mehr teilen können. Krankhafte Veränderungen des TauProteins spielen unter anderem bei der Alzheimer-Demenz eine Rolle, weil dort das Zytoskelett zusammenbricht, so dass das Tau-Protein zu unlöslichen Fasern verklumpt. Es bilden sich die „paarigen helikalen Filamente“ der Alzheimerkrankheit, die die Nervenzellen verstopfen und sie absterben lassen. Diese Vorgänge lassen sich im Reagenzglas oder in Zellmodellen nachstellen und untersuchen. Dabei werden biochemische, molekularbiologische, zellbiologische und biophysikalische Methoden kombiniert. Der Schwerpunkt der Arbeiten liegt auf der Untersuchung der Selbstorganisation der Mikrotubuli, der Struktur Mikrotubuli-abhängiger Motorproteine, dem Mikrotubuli-assoziierten Tau-Protein und seiner Rolle in der Alzheimerkrankheit. Die Intensität der Synchrotronstrahlung erlaubt es z. B., Wachstum und Zerfall von Mikrotubuli in Echtzeit durch Beugung des Röntgenlichts zu verfolgen, Röntgenbilder von sehr kleinen Proben von Alzheimer-Fasern aufzunehmen, oder die Struktur von kristallisierten Motorproteinen mit hoher Auflösung zu bestimmen. Die Proteine können rekombinant in großen Mengen in Bakterienkulturen hergestellt werden, sie lassen sich gezielt abwandeln und somit in ihrer Funktion überprüfen. Die Proteine können an fluoreszierende „Laternen“ angekoppelt und in Nervenzellen eingeschleust werden, so dass man ihre Verteilung und Funktion direkt vor Ort in lebenden Zellen beobachten kann. Abbildung links: Fasern mit einer typischen Schraubenstruktur sind charakteristische Ablagerungen in Nervenzellen, die in der Alzheimer-Krankheit langsam aber unaufhaltbar absterben. Diese „paarigen helikalen Filamente“ bestehen weitgehend aus dem Tau-Protein, das seine normale Funktion der Stabilisierung der Mikrotubuli-Transportschienen verloren hat. Der eigentliche Auslöser der pathologischen Entartung von Tau-Protein ist noch unbekannt, aber es konnten in den letzten Jahren einige strukturelle Eigenschaften der anomalen Tau-Fasern aufgeklärt werden, und es ist inzwischen möglich, die Umwandlung von normalem zu pathologischem Tau-Protein im Reagenzglas zu vollziehen. Damit kann der molekulare Prozess systematisch untersucht werden, und man kann jetzt gezielt nach Substanzen suchen, die die Entartung von Tau-Protein aufhalten oder rückgängig machen. Das Röntgenbeugungsbild (rechts oben) zeigt, dass das anomale Tau-Protein eine neue Struktur annimmt (das sog. beta-Faltblatt), die stark zur Vernetzung mit anderen Proteinen neigt. Die Beispiele illustrieren, wie eine Kombination von Methoden aus der Strukturforschung, Biochemie und Zellbiologie bei Fragestellungen der biologischen Grundlagenforschung an lebenden Zellen eingesetzt werden kann. Sie zeigen außerdem, wie sich daraus sehr direkte Konsequenzen für biomedizinische oder biotechnologische Anwendungen ergeben können. Von der geplanten Erweiterung der Forschungseinrichtungen mit Synchrotronstrahlung bei DESY in Hamburg (Ausbau des Speicherrings PETRA-III, Bau des Röntgenlasers TESLAXFEL) werden in Zukunft noch wesentlich weitergehende Impulse für die biologische Grundlagenforschung und für die Verzahnung verschiedener Bereiche der Wissenschaft ausgehen. Abbildung links: Die Suche nach Bedingungen, die für das allmähliche Absterben von Nervenzellen in der Alzheimer-Krankheit verantwortlich sein könnten, führt unter anderem auf einen Defekt im zellulären Transportsystem hin. Es scheint, dass das Tau-Protein nicht nur Mikrotubuli (die Gleise des Transports) stabil hält, sondern sie auch „verkleben“ und damit unbrauchbar machen kann. Das Ergebnis ist, dass die vom Zellinneren nach außen gerichteten Transportvorgänge gestört werden, so dass die äußeren Bereiche der Zelle unterversorgt bleiben. Die große Abbildung zeigt Zellen, bei denen Tau-Protein mit einem grünen Fluoreszenz-Farbstoff („green fluorescent protein“, GFP) im Fluoreszenzmikroskop sichtbar gemacht wird; man sieht deutlich die Anlagerung an die faserförmigen Mikrotubuli der Zelle. Der gelbe Bereich im Zentrum besteht aus einer Ansammlung von Mitochondrien, den „Kraftwerken“ der Zelle. Normalerweise müßten diese Organellen gleichmäßig über die ganze Zelle verteilt sein, aber wegen des defekten Transportsystems bleiben sie im Inneren liegen. Besonders eindrucksvoll wird dies bei Nervenzellen (kleines Bild). Im normalen Zustand (links) sind die Mitochondrien (rote Punkte) überall im Zellkörper und in den langen Zellfortsätzen verteilt, so dass die Zelle gleichmäßig mit Energie versorgt wird. Wird das Transportsystem durch eine Fehlfunktion des Tau-Proteins defekt (rechts), dann können die Mitochondrien nicht mehr in die Zellfortsätze einwandern, sondern bleiben im Zellkörper liegen. Das führt zur Unterversorgung der Zellfortsätze, zum Absterben der Synapsen, wo Nerven miteinander Kontakt halten, und schließlich zum Tod der ganzen Nervenzelle. Quelle: DESY Max Planck-Arbeitsgruppen für strukturelle Molekularbiologie Arbeitsgruppe Ribosomenstruktur Leiterin: Prof. Dr. Ada Yonath E-mail: [email protected] Arbeitsgruppe Proteindynamik (Außenstelle des Max-Planck-Instituts für Biochemie) Leiter: Dr. Hans-Dieter Bartunik E-mail: [email protected] Arbeitsgruppe Zytoskelett (Außenstelle des Max-Planck-Instituts für medizinische Forschung) Leiter: Prof. Dr. Eckhard Mandelkow (Sprecher der Arbeitsgruppenleiter) E-mail: [email protected] Weitere Informationen geben wir Ihnen gerne unter: [email protected] MPG-ASMB Notkestraße 85 Gebäude 25b 22607 Hamburg Telefon: +49-40-8998-2801 Telefax: +49-40-8971-6810 E-mail: [email protected] www.mpasmb-hamburg.mpg.de