Myoarthropathien des Kausystems: VI - Die
Werbung

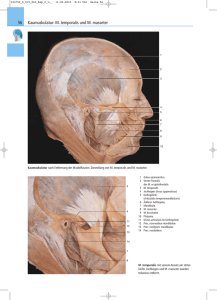
Praxis Myoarthropathien des Kausystems: VI - Die Kaumuskulatur. Neue wissenschaftliche Erkenntnisse Hans J. Schindler, Jens C. Türp Abb. 1: Verlauf der Kaumuskeln zwischen Schädel und Unterkiefer Indizes: Motorische Einheiten, Muskelfasertypen, heterogene Aktivierbarkeit, Biomechanik, Okklusionsschienen. Die Kaumuskeln unterscheiden sich in Mikromorphologie, Histochemie und Elektrophysiologie deutlich von der Muskulatur der Extremitäten und des Rumpfes. Auch die motorische Kontrolle des Kausystems zeigt Charakteristika, die bei der übrigen Skelettmuskulatur nicht vorhanden sind. Die Kenntnis dieser Besonderheiten hilft, die pathophysiologischen Hintergründe der Entstehung und der Therapie von Kaumuskelschmerzen besser zu verstehen. Die Kaumuskulatur Die Bewegungen des Unterkiefers werden im Wesentlichen durch die Kaumuskulatur 1 bewirkt. Diese setzt mit sieben paarigen 426 Einzelmuskeln am Unterkiefer an. Die Funktion der betreffenden Muskeln ergibt sich aus der Kenntnis ihrer Wirkungslinien, d.h. ihres dreidimensionalen Verlaufs von Ursprung zu Ansatz (Tab. 1). Der individuelle Muskel ist aus einer Vielzahl von Muskelzellen aufgebaut, die aufgrund ihrer kontraktilen Proteine (Aktin, Myosin) aktiv Kraft entwickeln können. Die elastischen Eigenschaften vor allem der extrazellulären Bindegewebsmatrix sorgen darüber hinaus bei passiver Dehnung für eine Spannungsentwicklung und Rückstellkraft in der Muskulatur. Die außergewöhnliche Verbindung der Mandibula mit dem Schläfenbein über die beiden Kiefergelenke erlaubt dem Unterkiefer mit Hilfe der neuromuskulären Kon1 Die suprahyoidale Muskulatur wird als „akzessorische Kaumuskulatur“ nicht getrennt betrachtet, sondern ebenfalls zu den Kaumuskeln gerechnet. ZAHN PRAX 8, 9, 426-430 (2005) Praxis Tab. 1: Wesentliche Funktionen der Kaumuskulatur Sonderform der Muskulatur: Die menschliche Kaumuskulatur trolle ein Bewegungsrepertoire, das im Vergleich zu den anderen großen Gelenken des menschlichen Körpers einzigartig ist. Diese Sonderstellung des Kauapparates hat im Laufe der Evolution dazu beigetragen, daß sich die Kaumuskeln in ihrer Mikromorphologie, Histochemie und Elektrophysiologie deutlich von der Muskulatur der Extremitäten und des Rumpfes unterscheiden. Aber auch die motorische Kontrolle des Kausystems zeigt Besonderheiten, die bei der übrigen Skelettmuskulatur nicht zu finden sind [4]. Makroskopische Morphologie der Kaumuskulatur Betrachtet man den Frontalschnitt des Kopfes in Abbildung 1, so sind die ausgeprägten Abwinkelungen der drei Schließmuskeln Temporalis, Pterygoideus medialis und Masseter zum Schädel und auch zueinander zu beachten. Diese Angulationen gestatten es den Muskeln, zusammen mit der nahezu horizontalen Ausrichtung des Pterygoideus lateralis eine Vielfalt von Kraftvektoren im Bereich der Okklusion zu generieren. Eine solche Konfiguration individueller Muskeln in Bezug zu einem Knochenelement (in diesem Fall dem Unterkiefer) ist im muskuloskelettalen System des menschlichen Körpers einmalig. Bei näherer Betrachtung des Frontalschnitts durch die wichtigsten Kaumuskeln ist eine weitere Besonderheit zu erkennen, nämlich ZAHN PRAX 8, 9, 426-430 (2005) die durch Sehnenspiegel vielfach unterteilte (kompartimentierte) multiple Fiederung von Masseter, Temporalis und Pterygoideus medialis [9]. Diese Fiederung wiederholt gewissermaßen, so könnte man es interpretieren, innerhalb des einzelnen Muskels die Angulationen der individuellen Muskeln zueinander. Eine derart strukturierte Muskulatur bezeichnet man als „komplexe“ Muskulatur. Sie zeichnet sich durch ein hohes Kraftpotential und eine ausgeprägte feinmotorische Kapazität aus. Im Gegensatz dazu sind Pterygoideus lateralis und Digastricus durch eine einfache Struktur gekennzeichnet. Diese beiden Muskeln müssen während der funktionellen Kieferbewegungen keine hohen Kräfte entwickeln und sie haben bei ihrer Kontraktion in der Regel wenig Widerstand zu überwinden. Mikroskopische Morphologie der Kaumuskulatur Blickt man noch tiefer in diese Muskulatur hinein, so sind weiter Besonderheiten zu erkennen, nämlich eine sog. Gruppierung der Muskelfasern (-zellen) einzelner motorischer Einheiten zu kleinen Faserbündeln (Faszikeln) [10] (Abb. 2a).2 Eine solche Anordnung der Muskelfasern auf engstem Raum ist eine physiologische Eigenheit der Kaumuskulatur, die beim Extremitätenmuskel als pathologisches Zeichen gewer2 Eine motorische Einheit besteht aus einem Motoneuron und allen von ihm gesteuerten Muskelzellen / Muskelfasern. 427 Praxis 2A 2B Abb. 2: Schematische Darstellung motorischer Einheiten im Muskelquerschnitt. A: Gruppierung der motorischen Einheiten in der Kaumuskulatur; B: Mosaikmusterartige Anordnung der Muskelzellen unterschiedlicher motorischer Einheiten in den Extremitäten am Beispiel des M. biceps brachii tet würde, nämlich als Folge einer fehlgeleiteten Re-Innervation nach Muskelläsionen. Betrachtet man zum Vergleich den Bizeps brachii als Vertreter des einfachen Extremitätenmuskels, so erkennt man sofort, daß hier die motorischen Einheiten ganz anders aussehen: sie sind im Stile eines Mosaikmusters eng miteinander verflochten (Abb. 2b) und erstrecken sich darüber hinaus über weite Bereiche des Muskelquerschnittes. Auch zeichnet sich die Muskulatur der Extremitäten durch deutlich größere Faserdurchmesser aus. Muskelfasereigenschaften Zu den strukturellen Besonderheiten der Kaumuskulatur gesellen sich weitere Eigenheiten. Im Wesentlichen lassen sich in der quergestreiften Muskulatur drei Fasertypen differenzieren. Sie werden entsprechend ihrem Gehalt an myofibrillärer ATPDie Kaumusase (verantwortliches Enzym für die Konkulatur traktionsgeschwindigkeit) und mitochonzeichnet sich drialem Enzym (verantwortlich für die Ausdurch besondauer) als sog. S-, FR- und FF-Fasern [2] dere Möglichgekennzeichnet (Tab. 2). In der Extremität keiten der sind die drei Fasertypen relativ gleichmäßig Kontraktion über den Muskelquerschnitt verteilt, und aus die motorischen Einheiten setzen sich aus gleichen Fasertypen zusammen. Betrachten wir die Kaumuskeln, so lassen sich im Vergleich zur Extremitätenmuskulatur fundamentale Unterschiede erkennen. Die Verteilung der Fasertypen ist heterogen; es dominieren die S-Fasern. Auch die diversen Muskelregionen (z.B. tiefe, oberflächliche resp. anteriore, intermediäre und posteriore Bereiche in Masseter und Temporalis) lassen unterschiedliche Verteilungsmuster 428 erkennen, d.h. die relativen Anteile der Fasern variieren zwischen den Regionen [3]. Darüber hinaus sind die S-Fasern in der Kaumuskulatur elektrophysiologisch nicht, wie zu erwarten, langsame Fasertypen, sondern schnell zuckende Fasern [7]. Doch nicht genug: Gehen wir bis in die Ebene der kontraktilen Elemente und betrachten uns das Myosin, so sind ebenfalls Besonderheiten im Vergleich zur Extremität zu entdecken. Es werden hier nämlich Myosine gebildet (Myosin-Isoforme), die in den Extremitäten überhaupt nicht vorkommen. Zudem findet man motorische Einheiten und einzelne Muskelfasern, die aus sehr unterschiedlichen Myosin-Isoformen bestehen [6], d.h. aus Myosinen, die sich in ihrem Kontraktionsverhalten voneinander unterscheiden. Dieser in den Extremitätenmuskeln unbekannte Sachverhalt in Verbindung mit der heterogenen Faserverteilung verleiht der Kaumuskulatur ein schier unerschöpfliches Repertoire an Kontraktionseigenschaften. Funktionelle Eigenschaften der Kaumuskulatur Die Frage, die sich an dieser Stelle zwangsläufig stellt lautet: Hat diese komplexe Architektur der Kaumuskeln eine funktionelle Entsprechung? Diese Frage ist mit einem eindeutigen „Ja“ zu beantworten. Die Entsprechung ist die sog. heterogene Aktivierbarkeit (= differenzierte Aktivierbarkeit) ZAHN PRAX 8, 9, 426-430 (2005) Praxis Tab. 2: Unterschiede zwischen dem S-, FR- und FF-Fasertyp der komplexen Kaumuskulatur [1]. Unter dieser Eigenschaft versteht man die Tatsache, daß der Verband der Motoneuronen, der einen individuellen Kaumuskel versorgt, in Subgruppen unterteilt ist, die kleiEin Vorteil ne Bereiche oder sog. funktionelle Kombei der Nahrungszer- partimente des Muskels selektiv kontrollieren können. Dies gestattet bei unterkleinerung: schiedlichen motorischen Aufgaben sowohl Die Kaueine zeitlich als auch eine im (relativen) Ermuskeln können ihre regungsniveau differenzierte Aktivierung der diversen Muskelregionen (KompartiWirkungslinie variieren mente). Prinzipiell läßt sich der individuelle Muskel so als Einheit verschiedener eigenständiger kleinerer Muskeln verstehen, die verschieden gerichtete Kraftvektoren entwickeln können. erung der Kontraktion wird dabei so bewerkstelligt, daß die Motoneuronen, die unterschiedliche Größe und damit auch unterschiedliche Reizschwellen besitzen, entsprechend der Reizstärke der einlaufenden Signale sequentiell erregt werden; also die kleinen Motoneuronen vor den großen. Da die motorischen Einheiten gleichmäßig über den Muskelquerschnitt verteilt sind, kommt es auf diese Weise zu einer homogenen Aktivitätssteigerung des Muskels [5]. Bedeutung der differenzierten Aktivierbarkeit für die Physiologie und Pathophysiologie der Kaumuskulatur Dieser Form der Aktivierung steht in der Extremität (als wesentlichem Prinzip) die sog. homogene Aktivierung gegenüber. Bei der homogenen Aktivierung erhalten alle Motoneuronen eines individuellen Muskels den gleichen synaptischen Input, d. h. identische Informationen von den Steuerorganen. Die Feinsteu- Die physiologische Bedeutung der heterogenen Aktivierbarkeit ist darin zu sehen, daß der individuelle Kaumuskel durch dieses Kontrollprinzip seine Wirkungslinie sowohl in anteroposteriorer als auch in mediolateraler Richtung variieren kann. Dieser Vorteil kommt besonders bei der Nahrungszerkleinerung zum Tragen, bei der eine präzise Orientierung des Kraftvektors in Bezug A B Abb. 3: Funktionsmusteränderungen in der Kaumuskulatur (M. masseter schematisch). A: Verteilung der aktiven Muskelregionen in Interkuspidation; B: Aktivitätsverteilung nach Lageveränderung des Unterkiefers ZAHN PRAX 8, 9, 426-430 (2005) 429 Praxis zum Nahrungsbolus notwendig ist. Darüber hinaus kann diese Eigenschaft aber auch die Ätiologie begrenzter Muskelläsionen (Mikrotraumata, lokale Erschöpfung) einleuchtend erklären, da eine differenzierte Aktivierung von diskreten Muskelregionen das Entstehen von lokalen Überlastungen eher plausibel macht als die homogene Aktivierung eines ganzen Muskels [11]. Regionaler Muskelschmerz findet auf diese Weise eine stichhaltige Erklärung (Abb. 3). Bedeutung der differenzierten Aktivierbarkeit für die Therapie von Muskelschmerzen Die regionalen Aktivierungszustände in den individuellen Kaumuskeln, wie sie z. B. bei Muskelkontraktion in maximaler Interkuspidation zu beobachten sind, werden durch eine Änderung der Kieferrelation (z. B. Entlastung: durch die Inkorporation von OkklusionsEine Ändeschienen) deutlich modifiziert [8]. Dies ist rung des als neuromuskuläre Adaptation an die neue Funktions„Startposition“ zu werten, die eine optimale musters kann Kraftübertragung im Bereich der Okklusion den Schmerz sichert. Diese „Funktionsmusteränderung“ reduzieren als Folge von Lageveränderungen des Unterkiefers liefert eine plausible Erklärung für die therapeutische Wirkung von Okklusionsschienen. Durch die Veränderung regionaler Aktivierungszustände kann es nämlich zur Entlastung überlasteter Bereiche kommen, was in Folge zu einer Erholung und damit zu Schmerzlinderung oder Schmerzbeseitigung in diesen Muskelregionen führt (Abb. 3). Klinische Phänomene wie die therapeutische Wirksamkeit sehr unterschiedlicher Schienentypen bei verschiedenen Kieferpositionen, aber auch von okklusalen Einschleifmaßnahmen, sind mit dieser Vorstellung gut zu erklären [12]. Unter dieser Sicht ist die therapeutische Wirkung jeder okklusalen Intervention als Veränderung der Kieferrelation und nicht als Wiederherstellung einer vermeintlich „idealen“ Zuordnung beider Kiefer zu interpretieren. Fazit Die Kaumuskulatur unterscheidet sich in Struktur und funktionellen Eigenschaften grundlegend von der querge- 430 streiften Muskulatur anderer Körperregionen. Die differenzierte Aktivierbarkeit der Kaumuskulatur liefert ein einleuchtendes Modell für die Entstehung regionaler Muskelschmerzen. Darüber hinaus wird die therapeutische Wirkung okklusaler Interventionen auf der Basis dieses motorischen Kontrollprinzips weitreichend erklärbar. Dr. med. dent. Hans J. Schindler Hirschstr. 105 D-76137 Karlsruhe Priv.-Doz. Dr. med. dent. Jens C. Türp Klinik für Rekonstruktive Zahnmedizin und Myoarthropathien, Universitätskliniken für Zahnmedizin Universität Basel Hebelstr. 3, CH-4056 Basel Literatur 1. Blanksma N. G., van Eijden T. M., van Ruijven L. J., Weijs W. A.: Electromyographic heterogeneity in the human temporalis and masseter muscles during dynamic tasks guided by visual feedback. J Dent Res 76, 542-551 (1997). 2. Burke R. E.: The structure and function of motor units. In Karpati G., Hilton-Jones D., Griggs R.C. (Hrsg): Disorders of Voluntary Muscles. Cambridge University Press, Cambridge 2001, 3-25. 3. Eriksson P. O., Thornell L. E.: Histochemical and morphological muscle-fibre characteristics of the human masseter, the medial pterygoid and the temporal muscles. Arch Oral Biol 28, 781-795 (1983). 4. Hannam A. G., McMillan A. S.: Internal organization in the human jaw muscles. Crit Rev Oral Biol Med 5, 55-89 (1994). 5. Henneman E., Somjen G., Carpenter D.O.: Excitability and inhibitability of motoneuurons of different sizes. J Neurophysiol 28, 599-620 (1965). 6. Korfage J.A., Brugman P., Van Eijden T.M.: Intermuscular and intramuscular differences in myosin heavy chain composition of the human masticatory muscles. J Neurol Sci 178, 95-106 (2000). 7. Nordstrom M. A., Miles T. S.: Fatigue of single motor units in human masseter. J Appl Physiol 68, 26-34 (1990). 8. Schindler H. J., Rong Q., Spieß W. E. L.: Der Einfluss von Aufbissschienen auf das Rekrutierungsmuster des Musculus temporalis. Dtsch Zahnärztl Z 55, 575-581 (2000). 9. Schumacher G. H.: Funktionelle Morphologie der Kaumuskulatur. Gustav Fischer, Jena 1961. 10. Stalberg E., Eriksson P. O., Antoni L., Thornell L. E.: Electrophysiological study of size and fibre distribution of motor units in the human masseter and temporal muscles. Arch Oral Biol 31, 521-527 (1986). 11. Tonndorf M. L., Hannam A. G.: Motor unit territory in relation to tendons in the human masseter muscle. Muscle Nerve 17, 436-443 (1994). 12. Türp J. C., Schindler H. J.: Zum Zusammenhang zwischen Okklusion und Myoarthropathien: Einführung eines integrierenden neurobiologischen Modells. Schweiz Monatsschr Zahnmed 113, 964-977 (2003). ZAHN PRAX 8, 9, 426-430 (2005)