1. dia
Werbung

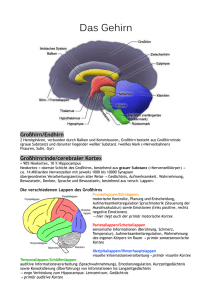
Aufbau der Großhirnrinde Der Kortex lässt sich in motorischen, sensorischen und Assoziationskortex einteilen. Sensorischer und motorischer Kortex nehmen im Vergleich zum Assoziationskortex nur einen kleinen Teil der Kortexoberfläche ein. Wir unterscheiden: die primär sensorischen Kortexareale, die ausschließ lich auf eine Sinnesmodalität reagieren; die primär motorischen Kortexareale, die direkt die Willkürmotorik steuern; die sekundären (oder auch unimodalen) sensorischen bzw. motorischen Kortexareale, meist in der Umgebung der primären Kortexareale liegen die (polymodalen) Assoziationskortizes, die mit höheren kognitiven, motorischen und emotionalen Funktionen befasst sind Der phylogenetische und ontogenetische Zuwachs an Hirnrinde beim Menschen ist primär auf die enorme Ausdehnung der polymodalen Assoziationsfelder zurückzuführen. Kortexschichten Der Kortex ist in sechs Schichten aufgebaut. Neuronal unterscheidet man zwei Hauptzelltypen: Pyramiden-und Sternzellen Neurone des Kortex: 2 Haupttypen-die erregende (exzitatorische) Pyramiden (80%)- und die überwiegend hemmende (inhibitorische) Sternzellen Die Kortex Schichten: -Die spezifischen Eingänge aus den Sinnessystemen gelangen über die thalamischen Fasern in die Schichten III, IV und V, in denen die Zellkörper der Pyramidenzellen liegen. Assoziationsfasern, Kommissurenfasern und unspezifische thalamische Fasern führen an die Dendriten von den Schichten I und II. Die Schichten I-V empfangen primäre Afferenzen -Die Schichten V und VI sind dagegen Ausgangsschichten (Efferenzen) Überträgersubstanzen: Die Pyramidenzellen benutzen als Überträgersubstanz (Transmitter) meist eine erregende Aminosäure, vor allem Glutamat. Obwohl die meisten Sternzellen hemmende Transmitter, gammaAminobuttersäure (GABA) ausschütten, enthalten einige der erregenden Sternzellen Neuropeptide (CCK, VIP). Viele der afferenten Fasern benutzen die Monoamine Noradrenalin und Dopamin, andere Acetylcholin, Serotonin und Histamin. NO (Stickoxid) spielt eine Rolle bei der anhaltenden Aktivierung von Zellensembles Die biologischen Grundlagen I. Die Neuropsychologie Analysiert die Interaktion von Gehirn und Verhalten Descartes -Leib-Seele-Problem Läsionsstudien Die bildgebenden Verfahren-Positronen-Emissions-Tomographie (PET) funktionale Magnetresonanztomographie (fMRT), Elektroenzephalogramm (EEG), Der Vorteil von PET und fMRI liegt in der guten räumlichen Auflösung. Gleichzeitig ist die zeitliche Auflösung schlecht. Das EEG liefert gute zeitliche Auflösung, aber kaum räumliche Informationen Repräsentationen psychischer Funktionen im Gehirn Hirnstamm Zerebellum Basalganglien, limbisches System, Hypothalamus Die emotionale Bewertung von Information findet in der Amygdala statt. Der Hippocampus spielt dagegen eine zentrale Rolle bei der Einspeicherung neuer Gedächtnisinhalte Die biologischen Grundlagen II. Neokortex Der Frontallappen (Stirnlappen),-Broca-Areal Die zentrale Rolle des Frontallappens liegt in der Planung und Steuerung von Verhalten. Der Temporallappen (Schläfenlappen) -(WernickeAphasie) Der Temporallappen ist für alle Prozesse relevant, die mit der Wahrnehmung auditorischer Information und dem Langzeitgedächtnis zusammenhängen. Der Parietallappen (Scheitellappen) Im Parietallappen liegen die Areale der sensorischen Repräsentation des Körpers und steuert gnostische Funktionen Der Okzipitallappen Aufgabe der okzipitalen Kortexareale ist die visuelle Informationsverarbeitung. Physiologie des Kortex I. Die Synapsen der Pyramidenzellen sind aktivitätsabhängig modifizierbar (plastisch). Dies führt zur Bedeutung des Kortex als einem großen assoziativen Speicher, d. h. dort ist das Wissen niedergelegt, das im Laufe eines Lebens erworben und beim Denken genutzt wird. Die kollektive elektrische Aktivität der Kortexneurone kann mithilfe von Elektroden auf der Kopfhaut registriert werden,die als Elektroenzephalogramm, EEG, bezeichnet werden.Ihre Frequenzen liegen zwischen 0-80 Hz und ihre Amplituden in der Größenordnung von 1-100 μV Mit der Magnetoenzephalographie, MEG, können magnetische Felder erfasst werden. Diese entstehen durch die elektrische Hirnaktivität. Das EEG entsteht überwiegend durch extrazelluläre Ströme der Pyramidenzellen in der Hirnrinde, das MEG resultiert aus intrazellulären Strömen -der a-Rhythmus wird durch rhythmisch entladende Schrittmacherzellen des Thalamus in den Kortex übertragen -&-Wellen, die nicht wie üblich im Schlaf, sondern bei gespannter Aufmerksamkeit in vorderen Kortexbereichen auftreten, dürften vom Hippokampus generiert werden Die Neurotransmitter und das Verhalten Die Neurotransmitter können je nach dem Ort ihrer Wirkung und dem Zusammenspiel mit anderen Neurotransmittern eine unterschiedliche Wirkung haben. Das dopaminerge System, mesolimbische,nigrostriatale,tuberoinfundibulare Das serotonerge System Schlaf-Wach-Rhythmus Stimmung Das noradrenerge System – arousal-, Stimmung Das Elektroenzephalogramm I Das Elektroenzephalogramm (EEG) Hans Berger (1929) misst mit Oberflächenelektroden an standardisierten Ableitpunkten auf der Kopfhaut die bioelektrische Aktivität bestimmter Gehirnregionen. Das Spontan-EEG zeigt die Potenzialschwankungen, die ohne einen Einfluss von außen im Wachzustand oder im Schlaf zu messen sind. Vier Typen von Frequenzbändern nach ihrer dominierenden Frequenz : Alpha-Wellen -synchronisiertes EEG. Beta-Wellen-Alpha-Blockade-EEG- Desynchronisation Theta-Wellen -und Delta-Wellen Das EEG-Muster unterscheidet sich je nach Lebensalter Das Elektroencephalogramm II Die evozierten Potenziale (ereigniskorrelierte Potenziale, EKP) sind Veränderungen der elektrischen Aktivität, die durch ein Reizereignis hervorgerufen (= evoziert) werden. nach auslösendem Reiz spricht man:von visuellen-,akustischen-und somatosensorischen evozierten Potenziale Contingent Negative Variation" - Bereitschaftspotenzial P300-Aufmerksamkeitsprozessen Im Allgemeinen ist eine Negativierung im EEG ein Indikator für eine kortikale Mobilisierung und eine Positivierung auf eine Deaktivierung hinweist Physiologie des Kortex II. PET. Eine analoge Methode, bei der aus Radioisotopen freigesetzte Positronen erfasst werden, nennt man Positron-EmissionsTomographie (PET) Funktionelle Magnetresonanztomographie (fMRT) Aktive Hirnareale werden durch die funktionelle Magnetresonanztomographie (Kernspintomographie), ein örtlich besonders gut auflösendes bildgebendes Verfahren dargestellt; in diesen Bereichen ist der Gehalt an oxygenisiertem Blut, das weniger paramagnetisch ist, besonders hoch Zerebrale Asymmetrie I. Die beiden Hemisphären des Neokortex weisen zwar unterschiedliche Arten von Informationsverarbeitung auf, für Verhalten und Denken ist aber die Zusammenarbeit der rechten und linken Hemisphäre unerlässlich Hemisphärenasymmetrien. Menschen mit einseitigen Hirnläsionen, von Patienten mit durchtrenntem Corpus callosum (Split Brain) Evolution der zerebralen Asymmetrie Die zerebrale Asymmetrie entwickelt sich möglicherweise in utero; die Lateralität von Händigkeit, Sprache und visuell-räumlicher Funktionen könnte dennoch weitgehend unabhängig voneinander auftreten Ursache der zerebralen Asymmetrien. ca. 75 % der Erdbevölkerung anzutreffende Bevorzugung der rechten Hand hat mit dem aufrechten Gang des Menschen zu tun Darwin: die zerebrale Asymmetrie ist eine evolutionäre der Sprachentwicklung Die Ausprägung unterschiedlicher Talente könnte mit der Lateralisierung für bestimmte Verhaltensweisen zusammenhängen Vorbedingung Zerebrale Asymmetrie II. Geschlechtsunterschiede der Lateralisierung Gemeinsamer anatomischer Unterschied: die variablere intrakortikale axonale Kommunikation der linken Hirnhemisphäre, bedingt durch variablere Myelinisierung der intrahemisphärischen Verbindungen auf der linken Seite Die rechte und die linke Hirnhemisphäre unterscheiden sich in ihrem makro- und mikroanatomischen Aufbau. Bestimmte Denkmuster und Bewegungsprogramme werden dabei von einer Hemisphäre bevorzugt: von der rechten Hemisphäre wird eine auf Ähnlichkeit und visuell-räumliche Gestalten ausgerichtete Informationsverarbeitung, von der linken syntaktisch-sprachliche und sequenziellkausale Verarbeitung praktiziert. Die Lateralisation und die Hemisphärendominanz Die Lateralisation von Funktionen wird einerseits von genetischen Faktoren wie Händigkeit und Geschlecht und andererseits von Faktoren der Umwelt (z. B. Anregungsgrad, spezielle Anforderungen) beeinflusst. Dominanz der linken Hemisphäre Sprache,-Analyse schneller sequenzieller Informationen Dominanz der rechten Hemisphäre räumliche Prozesse -Verarbeitung nicht-sprachlicher Informationen mit einer Spezialisierung auf visuellräumlichen Funktionen Die neuronale Plastizität Die Fähigkeit des Gehirns, sich zu reorganisieren und ausgefallene Funktionen auf verschiedene Weise zu kompensieren Die neuronale Plastizität als Grundlage des Lernens Neubildung von Neuronen,-die Anzahl der Synapsen und der synaptischen Dornen (Spines) nimmt zu anregende Umgebung -Deprivation -Hebb-Synapsen Die neuronale Plastizität und die Regeneration „Sprouting" - das Aussprossen von Axonkollateralen Plastizität des Gehirns und Lernen I Entwicklung und Lernen Frühe Erfahrungen und Interaktion mit der Umgebung steuern Wachstum und Verbindung von Nervenzellen Lernen und Reifung. Alle Lernprozesse sind Ausdruck der Plastizität des Nervensystems, aber nicht jeder plastische Prozess bedeutet Lernen. Reifung- genetisch programmierte Wachstumsprozesse zu Veränderungen des zentralen Nervensystems führen, die als unspezifische Voraussetzung für Lernen fungieren. Inaktivierung und Absterben unbenutzter Neurone. Durch simultanes Feuern wird nicht nur die Stärke der Verbindung der kooperierenden Synapsen erhöht, sondern gleichzeitig die der inaktiven benachbarten Synapsen geschwächt. Durch die simultan aktiven Synapsen wird aktivitätsabhängig der Nervenwachstumsfaktor (Nerve Growth Factor, NGF) von den benachbarten Synapsen »abgezogen«. Bei nicht Vorhandensein des Nervenwachstumsfaktors oder eines ähnlichen, auf den postsynaptischen Zellen aktivierten Wachstumsfaktors sterben die benachbarten nicht-aktiven Zellen ab (»pruning«). Plastizität des Gehirns und Lernen II Hebb-Synapsen (Donald Hebb) Die Hebb-Regel stellt die neurophysiologische Grundlage der Bildung von Assoziationen dar. Hebb-Synapsen haben die Eigenheit, bei simultaner Erregung ihre Verbindung zu verstärken Die Aktivation und das Bewusstsein Aktivierung oder „Arousal" meint eine allgemeine Funktionsanregung des Organismus mit dem Ziel der Handlungsvorbereitung. Indikatoren: erhöhte Aktivität des aufsteigenden retikulären Aktivierungssystems (ARAS), erhöhte Sympathikusaktivität Je höher die Frequenzen des EEGs, desto höher der Grad des Bewusstseins. Die Aktivierung und die Leistung -Yerkes-Dodson-Gesetz – umgekehrte UForm Die umgekehrte U-Form wird je nach Schwierigkeitsgrad der Aufgabe verzerrt Die Orientierungsreaktion und die Aktivierung Orientierungsreaktion verändert das Aktivierungsniveau des gesamten Organismus und versetzt ihn so in die Lage, Reize, die für ihn bedeutsam sein könnten, zu erfassen und auf sie reagieren zu können. Die Orientierungsreaktion richtet die Aufmerksamkeit auf Reize, die neu und unerwartet sind. Die Habituation und Defensivreaktion Habituation oder Gewöhnung. Defensivreaktion hat eine Abwendung vom schädigenden Reiz (Flucht) oder ein Abwenden der Bedrohung (Angriff) zum Ziel. Neurobiologie der Aufmerksamkeit Formen der Aufmerksamkeit: Automatisierte (nicht-bewusste) Aufmerksamkeit findet im sensorischen Gedächtnis und im Langzeitgedächtnis statt. Die Reaktion auf einen Reiz erfolgt automatisch, wenn der ankommende Reiz in ein gespeichertes Reiz-Reaktionsmuster passt Kontrollierte (bewusste) Aufmerksamkeit spielt sich im limitierten Kapazitätskontrollsystem(LCCS) ab. Diese Form der Aufmerksamkeit tritt nur nach neuen, nicht eindeutigen oder biologisch bedeutsamen Reizen und vor Willenshandlungen in Aktion und führt zu einer Begrenzung der Reizverarbeitung und Reaktionsausführung. Kortikale Mechanismen Der bewussten Aufmerksamkeit liegt die synchrone Depolarisation der apikalen Dendriten des Neokortex zugrunde. Diese tritt als Folge der Aktivierung eines ausgedehnten neuronalen Netzwerks, einschließlich präfrontaler und assoziativer Kortexareale. Die Aufzeichnung langsamer Hirnpotentiale (Negativierung bei Aufmerksamkeitsmobilisierung), ereigniskorrelierter Potentiale (zum Erfassen des Zeitablaufs) und der lokalen Hirndurchblutung erlauben die Aufzeichnung dieser Vorgänge beim Menschen. Subkortikale Aktivierungssysteme I. Retikulärformation Die Retikulärformation stellt die anatomische und physiologische Basis des Wachbewusstseins dar Abtrennung des Hirnstamms vom Zwischenhirn (Cerveau isole (isoliertes Vorderhirn)Durchtrennung der Medulla oblongata (Encephale isole, isoliertes Hirn) Subkortikale Aktivierungssysteme II. Aktivierungssysteme Ein anatomisch und neurochemisch heterogenes System des medialen Hirnstamms ist für die Steuerung tonischer (länger anhaltender) Wachheit verantwortlich. Dieses System ist die Retikulärformation des Mittelhirns. Die dienzephalen Ausläufer der Retikulärformation, vor allem der Nucleus reticularis thalami und Teile der Basalganglien sind mit selektiven Aufmerksamkeitsprozessen befasst. Präfrontaler und parietaler Kortex und Gyrus cinguli sind die obersten Entscheidungsinstanzen für die Auswahl biologisch bedeutsamer und für die Hemmung irrelevanter Information (Top-down-Aufmerksamkeit) Neurochemie des Bewusstseins Monoaminerge, glutamaterge und cholinerge Systeme des Hirnstamms modulieren dieTätigkeit vieler Hirnregionen und des Rückenmarks Formen von Lernen und Gedächtnis I. Nicht-assoziatives Lernen Habituation (Gewöhnung) und Sensitivierung sind die einfachsten Formen von (nicht-assoziativem) Lernen Associatives Lernen Lernen durch Konditionierung Bei der klassischen Konditionierung wird ein neutraler Reiz mit einem vital bedeutsamen Reiz assoziiert; bei der operanten Konditionierung wird ein zu lernendes Verhalten verstärkt bzw. gehemmt Prägung ist eine spezielle Form von assoziativem Lernen. Sie beruht auf einer angeborenen Sensibilität für bestimmte Reiz-Reaktions-Verkettungen in einem bestimmten Abschnitt der Entwicklung eines Lebewesens. (Konrad Lorenz) Formen von Lernen und Gedächtnis II. Lernen durch Biofeedback Durch assoziative Lernvorgänge können autonome und zentralnervöse Vorgänge verändert werden. Operantes Lernen von physiologischen Reaktionen wurde zu einer wichtigen Behandlungsmethode Der fundamentale Mechanismus, der allem Lernen zugrunde liegt, ist die Assoziation. Beim klassischen Konditionieren wird die Assoziation über zeitlich simultan auftretende Reize erworben (Kontiguität). Beim instrumentellen Konditionieren erfolgt die Assoziation über Kontiguität und die Verursachung einer Konsequenz nach einer Verhaltensweise (Kontingenz). Deklaratives und Prozedurales Gedächtnis Zwei Gedächtnissysteme werden unterschieden: Das prozedurale (Verhaltens-) Gedächtnis und das deklarative (Wissens-) Gedächtnis oder implizites und explizites Gedächtnis Das Verhaltensgedächtnis wird auch implizites Lernen und Gedächtnis genannt. Implizit, d. h. auch ohne Beteiligung des Bewusstseins, werden Verhaltensweisen erworben und wiedergeben. Das Wissensgedächtnis bezeichnet man auch als explizites Lernen und Gedächtnis. Zum Erwerb und zur Wiedergabe von Wissen und Ereignissen braucht man meist expliziten, bewussten Zugriff zum Gedächtnismaterial. Sensorisches Gedächtnis-Die sehr kurze, nichtbewusste Speicherung aller ankommenden Information erfolgt durch das sensorische Gedächtnis Kognitives Lernen Das Wissensgedächtnis ist für die Speicherung von Episoden und Wissen zuständig; man unterteilt es grob in Kurz- und Langzeitgedächtnis Einfluss der Umgebung I. Lernen und Erfahrung sind auf Reize aus der Umgebung angewiesen und führen zu verschiedenen strukturellen Änderungen, vor allem an kortikalen Dendriten Ort und Art des Lernens: die apikalen dendritischen Synapsen und Spines können als ein wesentlicher Ort des Lernens betrachtet werden. Kortikale Karten Durch Lernprozesse kommt es zur Ausbreitung oder Reduktion kortikaler Repräsentationen und Karten Phantomempfindungen und -schmerzen Einfluss der Umgebung II. Lernprozess Für einen erfolgreichen Lernprozess sind verschiedene Parameter notwendig: Genetisch bestimmte Reifung des Nervensystems, Ausbildung spezifischer synaptischer Verbindungen unter dem Einfluss von Umwelteinflüssen, Abbau »überflüssiger« synaptischer Verbindungen (Pruning) unter dem Einfluss von Umwelteinflüssen. Da eine stimulierende Umgebung die Voraussetzung für die Modifikation der synaptischen Verbindungen darstellt, gelingt diese in anregender Umgebung besser als in verarmter. Neuronale Grundlagen Die makroskopischen und mikroskopischen Veränderungen des Gehirns durch Lernen folgen der Hebb-Regel: gleichzeitige Aktivierung einer Zelle verstärkt die mitbeteiligte Synapsen und die benachbarten, nicht aktivierten Synapsen werden gleichzeitig abgeschwächt. Proteinbiosynthese und Langzeitgedächtnis I. Konsolidierung und Langzeitgedächtnis sind mit Änderungen der Genexpression und Proteinsynthese verbunden Gedächtnisinhalte haben immer in neuronalen Netzen oder Assemblies ihre Entsprechung und sind nicht auf molekulare Kaskaden reduzierbar Proteinexpression Die Expression neuer Proteine nach simultaner Erregung hängt von der Aktivierung von cAMP-Reaktions-ElementBindungsproteinen (CREB) ab Second Messengers, welche durch die anhaltende Erregung oder Hemmung der postsynaptischen Zelle synthetisiert werden, regen über die RNA-Synthese die Expression von Proteinen an. Proteinbiosynthese und Langzeitgedächtnis II. Molekulare Lernprozesse: Kurzzeitgedächtnis Einfache Assoziationsbildungen entstehen durch eine Verstärkung der synaptischen Verbindungen zwischen denjenigen sensorischen Neuronen, die konditionalen (CS) und unkonditionalen (US) Reiz an die efferenten Neurone leiten. Die Gleichzeitigkeit der beiden ankommenden Erregungen löst eine Kaskade intrazellulärer Vorgänge aus, die zu verstärkter Ca2+-Konzentration und erhöhter Transmitterausschüttung führen. Langzeitgedächtnis: Für die Überführung der einmal gelernten Information ins Langzeitgedächtnis wird Langzeitpotenzierung im Hippokampus und Kortex verantwortlich gemacht. Die abschließende Fixierung der Information im Langzeitgedächtnis erfolgt schließlich durch Anregung oder Hemmung der vom genetischen Apparat gesteuerten Synthesen von Kanalproteinen der Zellmembran. Die Bildung von Langzeit-Gedächtnisspuren hängt von der Synthese neuer Proteine ab, welche die Erregbarkeit der postsynaptischen Zellmembran dauerhaft modifizieren. Durch die Neustrukturierung der postsynaptischen Membran wird eine dauerhafte Modifikation der Erregbarkeit dieser Zelle in einem Zellensemble erreicht und die Entladungswahrscheinlichkeit und Oszillation eines spezifischen Zellensembles verändert. Neuropsychologie von Lernen und Gedächtnis I. Lernen von Fakten und Ereignissen Das Gedächtnissystem des medialen Temporallappens ist für die Herstellung von assoziativen Verbindungen bei deklarativem (explizitem) Lernen verantwortlich Deklaratives Lernen ist von der Intaktheit des Hippokampus, des entorhinalen Kortex und der darüberliegenden perirhinalen und parahippocampalen Kortizes abhängig Kontextlernen: Das hippokampale System verbindet im Kortex isolierte Gedächtnisinhalte zu einem größeren Kontext Lernen von Fertigkeiten:Prozedurales (implizites) Lernen ist von der Funktionstüchtigkeit motorischer Systeme und der Basalganglien abhängig Neuropsychologie von Lernen und Gedächtnis II. Neurobiologische Grundlagen Deklaratives Lernen: Die bewusste Speicherung und das Abrufen von Wissen benötigt das mediale Temporalsystem und den Hippokampus. Implizietes Lernen: Klassische Konditionierung und der Erwerb von Fertigkeiten ist auf die Intaktheit der beteiligten sensomotorischen Systeme und der Basalganglien angewiesen. Neuropsychologie von Gedächtnis Anterograde Amnesien treten nach der beidseitigen Entfernung oder Zerstörung des medialen Temporallappens und der darunter liegenden Strukturen wie Hippokampus und Teilen des limbischen Systems auf. Die Patienten können keinerlei neue explizite Informationen behalten und wiedergeben, lernen aber durchaus motorische und kognitive Fertigkeiten implizit neu. Retrograde Amnesie: Die Patienten können Ereignisse, die vor einer Hirnschädigung liegen nicht erinnern. Sprachentwicklung Sprache hat sich vermutlich im Laufe der Evolution des Menschen aus dem Gebrauch von (nicht mehr ausreichender) Gestik entwickelt Koppelung von Sprachentwicklung und Werkzeuggebrauch in der Entwicklung des Kindes Sprachkortizes Aphasien und Lokalisation: präfrontale „motorische Sprachregion” Broca-Region (Brodmann Areae 44 und 45), postzentrale „sensorische” Wernicke-Region (Area 22), perisylvischer Kortex (gyrus angularis, supramarginalis) Die Broca- und Wernicke-Regionen sind nicht auf motorische bzw. sensorische Sprachfunktionen beschränkt; auch rechtshemisphärische Prozesse sind an der sprachverarbeitung beteiligt Klinisch häufige Aphasieformen I Beim Aphasiker sind in der Regel alle sprachlichen Modalitäten von der Störung betroffen (Sprachproduktion, Sprachverständnis, Nachsprechen, Schreiben, Lesen etc.). Selektive organische Sprachstörungen, die nur eine Modalität betreffen, sind selten. Alle Aphasien beinhalten also Störungen des Benennens von Objekten, der Produktion und des Verständisses von Sätzen, sowie des Lesens (Alexie) und Schreibens (Agraphie). Bei umschriebenen Läsionsorten im Gehirn können eine Reihe aphasischer Syndrome durch ihre jeweils charakteristischen Symptome voneinander abgegrenzt werden: Broca-Aphasie: Sprachproduktionsprobleme stehen im Vordergrund. Artikulationen erfolgen meist sehr mühevoll und ohne Prosodie. Wörter sind phonematisch entstellt. In komplexen Sätzen fehlen häufig die grammatikalischen Funktionswörter. Das Verständnis vieler Satztypen (z.B. Passivsätze) ist oft nicht möglich. Probleme beim Nachsprechen von Sätzen treten auf. Organische Grundlage: Schädigung der Broca-Region und angrenzender Gebiete. Klinisch häufige Aphasieformen II. Wernicke Aphasie: Sprachproduktion ist zwar »flüssig«, jedoch oft unverständlich. Viele Wörter sind phonematisch entstellt, sodass noch verständliche phonematische Paraphasien (z.B. »Spille« statt »Spinne«) oder ganz unverständliche Neologismen auftreten. Oft werden Wörter durch bedeutungsverwandte ersetzt (semantische Paraphasien). Das Sprachverständnisdefizit ist sehr ausgeprägt. Das Verständnis einzelner Wörter gelingt häufig nicht. Das Nachsprechen von Wörtern und Sätzen ist beeinträchtigt. Organische Grundlage: Schädigung der WernickeRegion und angrenzender Gebiete. Globale Aphasie: Schwerste Sprachproduktionsstörung, bei der oft nur noch stereotype Silben oder Wortfolgen geäußert werden können. Ebenso stark ausgeprägtes Defizit im Sprachverständnis und im Nachsprechen. Organische Grundlage: Schädigung der gesamten perisylvischen Region. Klinisch häufige Aphasieformen III. Amnestische Aphasie: Leichte Sprachstörung, bei der semantische Paraphasien auffallen und Benennstörungen im Vordergrund stehen. Probleme treten vor allem mit bedeutungstragenden Inhaltswörtern auf. Das Sprachverständnisdefzit ist schwach ausgeprägt. Organische Grundlage: Schädigung des Gyrus angularis oder anderer Areale, die dem linken perisylvischen Kortex eng benachbart sind. Gelegentlich führt bei Rechtshändern Schädigung der rechten Hemisphäre zu amnestischer Aphasie (»gekreuzte Aphasie«). Aphasien treten auch bei subkortikalen Läsionen in der weißen Substanz, in den Basalganglien oder im Thalamus auf. Diese subkortikalen Aphasien mit einem anfänglichen Mutismus bilden sich in der Regel rasch zurück Neuronale Grundlagen von Sprache. Beim Menschen sind syntaktische Regeln und Funktionswörter primär links in der perisylvischen Region lokalisierbar (sprachdominante Hemisphäre). Sprachverständnis, vor allem von Inhaltswörtern findet sich aber auch rechts. Die Assoziationsareale des Neokortex: Höhere geistige Funktionen I Assoziationsareale sind Rindenfelder, die keine eindeutigen sensorischen, sensiblen oder motorischen Funktionen aufweisen, sondern das Zusammenwirken zwischen den einzelnen Sinnessystemen und den motorischen Arealen integrieren (»assoziieren«). Exekutive Funktionen Der präfrontale Assoziationskortex ist für die zielorientierte, exekutive Planung des Verhaltens und das Arbeitsgedächtnis wichtig Evolution des präfrontalen Kortex- »spezifisch menschlichen« Eigenschaften Das Ausüben von Selbstkontrolle ist eine beim Menschen am weitesten fortgeschrittene Funktion, die an präfrontale Hirnregionen gebunden ist Läsion der Integrationsleistung – »pseudopsychopathisches« Zustandsbild – fehlende Empathie - Schizophrenie Die Assoziationsareale des Neokortex: Höhere geistige Funktionen II Perzeptive Funktionen Der parietale Assoziationskortex ist mit der Steuerung komplexer, sensorischer Reizverarbeitung, der visuellen Aufmerksamkeit, mit Handlungsplanung und mit räumlichen Funktionen befasst Läsionen des Parietallappens Kontralateraler Neglekt Agnosien (»Seelenblindheit«) Taktile oder visuelle Agnosie Prosopagnosie bedeutet das Nichterkennen von Gesichtern. Der parietale Assoziationskortex ist mit der Steuerung sensorischer Reizverarbeitung (optisch, taktil, akustisch, vestibulär) befasst. Die parietalen Felder ermöglichen Aufmerksamkeit und Lokalisierung (»Wo«?) sensorischer Reizquellen. Der Temporalkortex ist dabei in seinem unteren Abschnitt mit Erkennen und Bedeutungsanalyse vor allem visueller Reize (bei Ausfall Agnosie), in seinem medialen Teil mit explizitem Gedächtnis und superior mit akustischen Funktionen und Sprache verbunden. Der anteriore Pol und Amygdala dienen höheren emotional-sozialen Funktionen. Emotionen als physiologische Anpassungsreaktionen Psychische Kräfte und psychische Funktionen Das Auftreten, die Intensität und die Richtung psychischer Funktionen (Denken, Wahrnehmung, Lernen) wird durch Motivation (Trieb) und Emotionen bestimmt. Verhaltensweisen werden durch positive oder negative Verstärker gefördert oder unterdrückt. Motivationen sind Antriebszustände (psychische Kräfte), die von zentralen Erregungsschwellen im Gehirn abhängen und die Wahrscheinlichkeit bestimmter Verhaltensweisen erhöhen oder senken. Sie werden auch als Triebe bezeichnet und sind im Gehirn entweder homöostatisch oder nichthomöostatisch organisiert. Emotionen sind kurzzeitige vom Gehirn organisierte Reaktionen (psychische Kräfte), die alle Verhaltensweisen mitbestimmen; sie bestehen aus subjektiv benennbaren Gefühlen und vegetativen, neuroendokrinen und somatomotorischen Reaktionen. Die sechs verschiedenen Basisemotionen (primär): Angst, Furcht, Trauer, Abscheu, Freude und Überraschung können psychophysiologisch unterschieden werden. Sie sind durch die parallel ablaufenden subjektiven Gefühle, motorischen Reaktionen und vegetativen Reaktionen charakterisiert. Die Emotionen regulieren Anpassungen des Verhaltens bei wechselnden Umweltkonstellationen, Annäherungs- und Vermeidungsverhalten und lenken Entscheidungen (auch im sozialen Kontext). Die zentralen Repräsentationen der Emotionen Emotionen (Gefühle, motorische, vegetative und neuroendokrine Reaktionen) sind in bestimmten Großhirnarealen (Cingulum anterior et posterior, Insula, präfrontalen Kortexarealen), Amygdala, Hypothalamus und Hirnstamm repräsentiert. Für jede Emotion ist diese Repräsentation spezifisch. Die zentralen Repräsentationen erhalten kontinuierliche afferente Rückmeldungen aus den Körpergeweben. Furchtverhalten Bestimmte Kerngebiete der Amygdala steuern über afferente Verbindungen von Thalamus und Kortexarealen und efferente Verbindungen zu Hypothalamus und oberem Hirnstamm die Emotion Furcht. Störungen der neuronalen Regulation von Emotionen führen zu psychopathologischen Veränderungen und/oder somatischen Erkrankungen. Die Emotionen Freude und Sucht Positive Verstärkung im Gehirn Belohnungssysteme im Hirnstamm und im limbischen System erzeugen Gefühle der Freude und sind für positive Verstärkung wichtig Nach J. Olds positives Verstärkungssystem genannt Mesolimbisches Dopaminsystem Dopamin des mesolimbischen Dopaminsystems wirkt als universelles positives Antriebsignal vor allem im Nucleus accumbens Dopaminagonisten wie Amphetamin und Kokain (beides süchtig machende Substanzen) fördern die positive Verstärkung. Opiate stimulieren indirekt die dopaminergen Neurone im ventralen Tegmentumdes Mittelhirns,aber auch Neurone in Nucleus accumbens, lateralem Hypothalamus, Pallidum und periaquäduktalem Grau.Auch das noradrenerge System hat bei Reizung meist positiv verstärkende Effekte. Dopaminantagonisten, wie z. B. Neuroleptika, hemmen die positive Verstärkung und führen zu Anhedonie (»Lustlosigkeit«) Negative Verstärkersysteme I. Hirnregionen, deren Reizung zu Aversion und Vermeidung führen, werden als negative Verstärkersysteme (Bestrafungssysteme) bezeichnet. Ihre neuronalen Strukturen sind weniger gut lokalisiert, da sie mit den zentralen Systemen zur endogenen Kontrolle von Schmerzen (Opioid und nicht Opioid, und den Regionen, die Sättigung und Ekel auslösen, überlappen. Sie befinden sich wahrscheinlich periventrikulär im Mesenzephalon. Eine relativ einheitliche anatomische und neurochemische zentralnervöse Struktur, wie wir sie für positive Verstärkung finden, scheint nicht zu existieren. Negative Verstärkersysteme II.. Pharmaka und negative Verstärkersysteme. Die negativen Verstärkersysteme hemmen die mesolimbischen positiven Verstärkersysteme. Transmitter in dem (dem positiven System benachbart gelegenen) negativen periventrikulären System sind Serotonin (5-HT), Cho-lezystokinin, Substanz P und andere an Sättigung und Schmerz beteiligte Neuromodulatoren. Sie haben viele Funktionen und sind nicht spezifisch für Aversion und Bestrafung. Dennoch beeinflussen viele Substanzen über das serotonerge System unser Verhalten: Antidepressiva verbessern die Stimmung durch Hemmung der Wiederaufnahme von 5-HT in die 5-HT-Neurone. Ecstasy (3,4-Methylendioxymethamphetamin) stimuliert den 5-HT2Rezeptor und verbessert Stimmung und Antrieb. Kokain hemmt sowohl die Wiederaufnahme von Dopamin wie auch Serotonin und stimuliert somit beide Systeme. Halluzinogene wie LSD (Lysergsäurediaethylamid) und Psilocybin stimulieren den 5-HT2-Rezeptor. Sie erzeugen außer Hallizunationen auch negative Gefühle (Panik, Paranoia). Herabgesetzte Verfügbarkeit von Serotonin am Rezeptor ist häufig mit gesteigerter Aggression und Autoaggression korreliert. Sucht Suchtverhalten ist eine extreme Form positiv motivierten Verhaltens; es unterscheidet sich quantitativ von der positiven Motivation durch verstärkte Aversionssymptome bei Entzug und, je nach Sucht, durch Entwicklung von Toleranz Suchtentstehung. Erfolgt die Aktivierung des Verstärkungssystems nicht mehr durch physiologische Reize, sondern werden Neurone des positiven Antrieb erzeugenden Systems direkt (chemisch) gereizt, kann, wenn die zeitlichen Abstände zwischen diesen Aktivierungen kurz sind, Sucht entstehen. Rolle der Umwelt. Sucht kann in ihren biologischen Grundlagen ohne Berücksichtigung der Umgebung, in der sie entsteht und aufrecht erhalten wird, nicht verstanden werden. Die biologischen Mechanismen, die einer Sucht zugrunde liegen, werden nur in ganz bestimmten Umgebungsbedingungen und nur bei umschriebenen Konsequenzen in dieser Umgebung (z.B. unter Stress) aktiviert. Sucht und mesolimbisches Dopaminsystem I. Die neuronale Grundlage der Sucht liegt in der Förderung der synaptischen Übertragung im mesolimbischen Dopaminsystem Während das Verlangen (die Suche) nach der Droge kontinuierlich ansteigt (die Sucht im engeren Sinne!), nimmt gleichzeitig die erzeugte Euphorie (Suchtbefriedigung) ab. Beiden Verhaltensweisen liegen unterschiedliche Mechanismen zugrunde.Die Aktivität im Nucleus accumbens nimmt in der Phase der Suche stark zu, nicht jedoch in der Phase der Suchtbefriedigung (Euphorie). Sucht und mesolimbisches Dopaminsystem II. Rückfall in die Sucht. Für die gleich hohe Rückfallhäufigkeit bei allen Süchten sind weniger Toleranz und Abstinenzreduktion verantwortlich, sondern die gelernten Anreizwerte aller Situationen und Gedanken, die in der Vergangenheit mit der Substanzeinnahme assoziiert waren. Im Laufe wiederholter Einnahme süchtig-machender Substanzen wird die Sensibilität des dopaminergen Systems größer, was zum Anstieg des Verlangens bei Auftritt von Hinweisreizen für die Aufnahme der Substanz führt. Die Freude oder Lust, die durch das Suchtmittel erzeugt werden, ist davon wenig berührt. Ebenso sind Abstinenzerscheinungen für die meisten Rückfälle nicht verantwortlich, die in der Regel lange nach Abklingen des Entzugs auftreten. Um Süchte wieder zum Verschwinden zu bringen (Extinktion), müssen dieselben Situationen, die mit der Einnahme des Suchtmittels assoziiert waren, wiederholt ohne Einnahme der Substanz dargeboten werden. Vermutlich nimmt auf diese Weise die Verstärkung der synaptischen Übertragung (z.B. im mesolimbischen Dopaminsystem), die sich bei der Entstehung der Sucht gebildet hat, wieder ab. Neuroadaptation des mesolimbischen Systems I. Kurzzeit- und Langzeitwirkung von süchtig-machenden Substanzen beruhen auf unterschiedlichen molekularen Mechanismen Akute Einnahme einer süchtig-machenden Substanz. Die Bindung der zugeführten Substanz an die Dopamin-oder Opiatrezeptoren der Neurone des mesolimbischen Dopaminsystems aktiviert G-Proteine, welche die Aktivität der Adenylatzyklase hemmen Dies führt zur Abnahme der Aktivität von cAMP und cAMP-abhängigen Proteinkinasen. Durch die Reduktion der cAMP-Aktivität wird auch die Phosphorylierung von Ionenkanälen und vermutlich anderer zellulärer Effektoren reduziert. Chronische Einnahme einer süchtigmachenden Substanz. Die intrazelluläre Signalübertragung ändert sich radikal bei chronischer Einnahme: die Aktivität des Adenylatzyklase-cAMP-Systems nimmt zu und die Aktivität der cAMP- oder Ca2+-abhängigen Proteinkinasen führt zu Phosphorylierung von Transkriptionsfaktoren im Zellkern. Die Transkriptionsvorgänge haben unter anderem eine Hochregulation der Postrezeptorsignalkette für den dopaminergen D2Rezeptor und eine Herunterregulation für den D2Rezeptor zur Folge (G-Protein, Adenylatzyklase usw.,). Neuroadaptation des mesolimbischen Systems II. Neuroadaptation. Im chronischen Zustand schrumpfen die dopaminergen Neurone des mesolimbischen Systems, während die Neurone im Ncl. accumbens mit dem kompensatorischen cAMP-Anstieg und der beschleunigten Transkription überaktiv werden, wenn nicht die an die Rezeptoren bindende Substanz erneut zugeführt wird. Die Affinität der D2-Rezeptoren für Dopamin nimmt mit zunehmender Drogeneinnahme ab. Diese Veränderung verschwindet Wochen nach Entzug wieder, während die durch Transkription erzeugten intrazellulären Änderungen über längere Zeit anhalten. Die biochemischen, morphologischen und physiologischen Veränderung der Neurone (hier des mesolimbischen Systems), die bei chronischer Einwirkung von Suchtsubstanzen stattfinden, werden als Neuroadaptation bezeichnet. Sucht Dopaminagonisten wie Amphetamin und Kokain (beides süchtig machende Substanzen) fördern die positive Verstärkung. Erfolgt die Aktivierung des Verstärkungssystems nicht mehr durch physiologische Reize, sondern direkt (chemisch), kann Sucht entstehen. Eine solche direkte chemische Aktivierung kann durch viele Suchterzeugenden Substanzen geschehen (wie z. B. Heroin, Morphin, Kokain, Marijuana, Amphetamine, Barbiturate, Nikotin und Alkohol). Das mesolimbische positive Verstärkungssystem bildet die gemeinsame anatomische Endstrecke für die Entwicklung und Aufrechterhaltung von Sucht. Blockade oder Zerstörung dieses Systems nimmt allen Situationen, in denen hohe positive Erregung (»Lust«) z. B. durch Drogeneinnahme erzeugt wird, ihren Anreizwert und führt zum Erliegen der Sucht. Die Neurone des mesolimbischen Systems verändern sich biochemisch, anatomisch und physiologisch bei chronischer Einwirkung von Drogen. Dieser Zustand wird als Neuroadaptation bezeichnet.