1.2 Die Bestäubung
Werbung

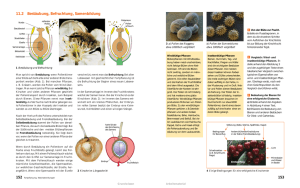
3 BFW-Berichte 144 Die Ernährung der Bäume spielt offenbar für den generativen Prozess eine besondere Rolle. Vor beinahe hundert Jahren wurde die Bedeutung eines weiten „C/N – Verhältnisses“ innerhalb des Vegetationskegels auf die Blühreife der Pflanzen erkannt (KLEBS, 1913). Daher ist es auch nicht verwunderlich, dass zahlreiche positive Effekte einer Mineraldüngung auf den Fruchtund Samenertrag von Bäumen beschrieben wurden (MATSCHKE, 1982). Andererseits werden bei fehlender oder nicht ausreichender Nährstoffversorgung trotz reichlicher Blütenbildung nur wenige Zapfen oder Früchte gebildet, was nach Erfahrung des Autors manchmal bei insektenblütigen Baumarten (Prunus avium, Tilia sp.) zutrifft. Schon HARTIG (1889) wies darauf hin, dass eine Vollblüte bzw. Vollmast für die betreffenden Bäume einen starken „Aderlass“ darstellt, da dadurch große Mengen an Reservestoffen verbraucht werden; das nur periodisch reichliche Fruktifizieren und die geringere Blütenbildung in den Zwischenjahren der meisten Waldbaumarten ist dadurch erklärbar. Noch in den Sechzigerjahren des vorigen Jahrhunderts wurden anhand von Untersuchungen Zuwachsverluste zwischen 15 – 20 % in Mastjahren von Eiche und Buche festgestellt (NEMEC, 1956 und FIELDING, 1960), die aber seit 1990 nicht mehr so ausgeprägt ersichtlich sind. Auswirkungen des Klimawandels – bessere Nährstoffversorgung (Lösung durch höhere Niederschlagsmengen) sowie mehr Energie (Strahlung) könnten dazu beitragen. Nach LYR et al. (1967) entscheiden die Mengenverhältnisse der Phytohormone in Verbindung mit gleichzeitiger Genexpression in den Meristemen sowie die Ausbildung und der Transport von Assimilaten (sog. “source-sink“ Verhältnisse) über eine mögliche vegetative oder generative Entwicklung der Bäume. GRUBER (2003) testete die Fruktifikation der Buche unter dem Einfluss verschiedener Witterungsfaktoren in den Jahren 1986 bis 2001 um die Frage der Vorhersage zu klären und die Auswirkung von Mastjahren auf den Zuwachs zu bestimmen. 1.2 Die Bestäubung Unter Bestäubung versteht man die Ablagerung von Pollen auf Narben oder Samenanlagen. Die Windbestäubung (Anemophilie) überwiegt bei den Waldbaumarten – im Gegensatz zur übrigen Flora. Alle heimischen Gymnospermen und viele Angiospermen (Alnus, Betula, Carpinus, Castanea, Corylus, Fagus, Fraxinus, Juglans, Platanus, Populus, Quercus und Ulmus) sind Windbestäuber. Anemogame Blüten haben weder Duftstoffe noch einen Schauapparat und blühen zumeist vor dem Blattaustrieb. Von sekundärer Anemogamie spricht man, wenn die Bestäubung in der ersten Blühphase durch Insekten und später nach Eintrocknung des Pollenkitts über die Luftbewegung erfolgt (Acer, Salix, Tilia). Die Bestäubung hängt bei den windblütigen Baumarten nicht so sehr wie lange Zeit angenommen von den Witterungsbedingungen ab, da männliche Blüten (Antheren) ihre Pollen nur bei günstigen Bestäubungsbedingungen (Wärme, bis zu einer bestimmten Luftfeuchtigkeit sowie nur geringere Luftbewegung) entlassen und auch nur dann die weiblichen Blüten geöffnet sind. Oft wird je nach Reifegrad die gesamte Pollenmenge eines Baumes innerhalb weniger Blühtage (ca. 10) abgegeben (ROBITSCHECK et al., 2008). Trotz der nachweislich großen Flugweiten (Alnus, Betula, Corylus...) die kleinere Baumpollen zurücklegen können, ist die Wahrscheinlichkeit einer Bestäubung weiblicher Blühorgane auf große Entfernungen gering. Nach ROHMEDER (1972) sind hauptsächlich folgende Gründe dafür maßgeblich: Größere Entfernungen und besonders Höhenunterschiede der Wuchsorte bedingen zumeist Unterschiede in den Zeitpunkten der Blüte. Solche Unterschiede können - da die weiblichen Blüten nur wenige Tage (baumartenspezifisch) befruchtungsbereit sind, die Bestäubung verhindern. Ebenso sinkt die Trefferwahrscheinlichkeit mit Zunahme der Entfernung durch die Verdünnung der Pollenkonzentration (Schrotschußdynamik). Die von den meisten anemogamen Baumarten produzierten Pollenmengen sind enorm. Wenn man jedoch bedenkt, dass ein großer Teil der freigesetzten Pollen verloren geht – beim Windtransport mehr als bei der Insektenbestäubung, so erscheint diese Massenproduktion notwendig. Nach POHL (1936) produzieren Baumarten eine sehr unterschiedliche Anzahl von Pollen pro Blüte (Tabelle 1). Tabelle 1: Zahl der produzierten Pollen pro Blüte (Hauptbaumarten) Pollen pro Blüte Fichte ca. 200.000 Kiefer 160.000 Eiche 40.000 Buche 12.000 Pollen: sind mehrzellige, von vielschichtiger Wand umgebene, in den Pollensäcken (Antheren) entstehende Gebilde und stellen den männlichen Gametophyten dar. Eine vegetative Zelle und die generative Antheridiumzelle sind von einer cellulosereichen Innenschicht, der Intine und einer widerstandsfähigen Außenwand, der Exine umgeben. Als wesentliche Merkmale werden dabei Anzahl, 4 22 Jahre Blüh- und Fruktifikationsuntersuchungen der Waldbaumarten im Lehrforst Ofenbach / Rosalia Abbildung 1: Pollen von verschiedenen Nadel- und Laubbäumen, Sträuchern, Gräsern und Kräutern. 1 Fichte, 2 Eibe, 3 Wacholder, 4 Lärche, 5 Eiche, 6 Rotbuche, 7 Tanne, 8 Pappel, 9 Hasel, 10 Kiefer, 11 Zirbe, 12 Linde, 13 Weide, 14 Ahorn, 15 Esche, 16 Walnuß, 17 Edelkastanie, 18 Ulme, 19 Hainbuche, 20 Birke, 21 Grünerle, 22 Weißerle, 23 Süßgras, 24 Sauergras, 25 Roggen, 26 Wegerich, 27 Beifuß, 28 Kornblume, 29 Schafgarbe, 30 Brennessel, 31 Ampfer, 32 Gänsefuß, 33 Glockenblume. 34 Bibernelle, 35 Frühlingsheide Art und Lage der Keimstellen (Aperturen) herangezogen sowie die Beschaffenheit der Pollenaußenwand (SCHÜTT et al., 1992). Die Keimstellen (Aperturen) können als Poren (porate Pollen), Falten (= Colpen, colpate Pollen) ausgebildet werden oder nur schwach angedeutet bzw. nicht vorhanden sein (inaperturate Pollen). Als Anpassung an die anemogame Verbreitung werden bei einigen Pinaceen-Gattungen (Abies, Picea und Pinus) durch Abheben der Exine Luftsäcke (Vesiculae) gebildet. Die folgende Abbildung 1 (KRAL, 1983) zeigt verschiedene wesentliche Pollen in Idealdarstel- lung, wogegen sie in Präparaten in jeder Lage auftreten können, was ihre Bestimmung manchmal erschwert. 1.3 Forschungsbereich PALYNOLOGIE Nach STRAKA (1975) kann dieser weite Forschungsbereich, entsprechend der besonderen Zielsetzungen in mehrere Teilbereiche gegliedert werden: • Grundlagenforschung (Morphologie und Systematik der Pollen) • PALEO – Palynologie (fossile Pollen und Sporen in Ablagerungen) • KRYO – Palynologie (Gletscherforschung) • AERO – Palynologie (Windverbreitung der Pollen und Sporen) • IATRO – Palynologie (Allergieforschung) • MELITO – Palynologie (Honigpollenanalyse) • PHARMA – Palynologie (Prüfung von Arzneimitteln) • PHY TOPATHOLOG. – Palynologie (Pilzsporen) • FORENSISCHE – Palynologie (Kriminaltechnik)