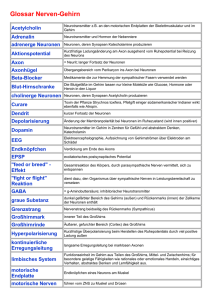
Neuronale Botenstoffe in einfachen Nervensystemen und ihr
Werbung

Aus dem Institut für Tierökologie und Zellbiologie Stiftung Tierärztliche Hochschule Hannover Neuronale Botenstoffe in einfachen Nervensystemen und ihr Einfluss auf funktionelle und strukturelle Plastizität Habilitationsschrift zur Erlangung der Venia legendi an der Stiftung Tierärztliche Hochschule Hannover vorgelegt von Dr. rer. nat. Michael Stern Hannover 2009 Tag der nicht öffentlichen wissenschaftlichen Aussprache: 07.06.2010 Für Svenja, Lisa und Britta Inhaltsverzeichnis Abkürzungsverzeichnis 1 1. Verzeichnis der Veröffentlichungen, die Bestandteil der Habilitationsschrift sind 2 2 Einleitung 2.1 Einfache Nervensysteme 4 4 2.1.1 Das Nervensystem von Insekten 4 2.1.2 Zellkultursysteme 6 2.2 Neuronale Botenstoffe 8 2.2.1 Neurotransmission 8 2.2.2 Synaptische Plastizität 10 2.2.3 Neuronale Botenstoffe in der Entwicklung 11 3 Fragestellungen 15 4 Ergebnisse 16 4.1 Zentrifugale Neuronen in der Sehbahn der Heuschrecke und ihre Modulation durch Octopamin 16 4.2 Serotonin bei der Entwicklung des enterischen Nervensystems der Heuschrecke 19 4.3 Stickstoffmonoxid im Zentralnervensystem der Heuschrecke und sein Einfluss auf axonale Regeneration 23 4.4 Neurotransmitter und synaptische Modulation durch Stickstoffmonoxid in menschlichen Modellneuronen 4.5 Neurotransmitter an der neuromuskulären Synapse von Onychophoren 5 Studienübergreifende Diskussion 5.1 Neurotransmitter bei neuronaler Plastizität und Entwicklung 30 36 39 39 5.1.1 Octopamin 39 5.1.2 Serotonin im enterischen Nervensystem 40 5.1.3 Stickstoffmonoxid im Heuschrecken-ZNS 42 5.1.4 Stickstoffmonoxid im Vertebraten-ZNS 45 5.2 Ausblick 47 6 Literatur 48 7 Zusammenfassung 58 8 Summary 59 9 Danksagung 60 10 Darstellung des eigenen Anteils an den wissenschaftlichen Veröffentlichungen 62 1 Abkürzungsverzeichnis cGMP Zyklisches Guanosinmonophosphat GABA Gamma-Amino-Buttersäure GAD Glutamatdecarboxylase ENS Enterisches Nervensystem NO Stickstoffmonoxid NOS Stickstoffmonoxid-Synthase SERT Serotonin-(Wiederaufnahme-)Transporter sGC NO-Rezptor (lösliche Guanylylzyklase) PM1 Protocerebrum-Medulla-Neuron 1 PM4 Protocerebrum-Medulla-Neuron 4 PNS Peripheres Nervensystem ZNS Zentralnervensystem 1 Verzeichnis von Veröffentlichungen die Bestandteil der Habilitationsschrift sind 1 Verzeichnis der Veröffentlichungen, die Bestandteil der Habilitationsschrift sind (chronologisch) Veröffentlichung 1: Pätschke A, Bicker G, Stern M (2004) Regeneration of proctolinergic neurons in the ventral nerve cord of the locust. Dev. Brain Res. 150:73-76. Veröffentlichung 2: Stern M, Knipp S, Bicker G (2007) Embryonic differentiation of serotonin-containing neurons in the enteric nervous system of the locust (Locusta migratoria). J. Comp. Neurol. 501:38-51. Veröffentlichung 3: Stern M, Bicker G (2008a) Nitric oxide regulates axonal regeneration in an insect embryonic CNS. Dev. Neurobiol. 68:295-308. Veröffentlichung 4: Stern M, Bicker G (2008b) Mixed cholinergic/glutamatergic neuromuscular innervation of Onychophora: A combined histochemical/electrophysiological study. Cell Tissue Res. 333:333-338. Veröffentlichung 5: Stern M (2009) The PM1 neurons, movement sensitive centrifugal visual brain neurons in the locust: anatomy, physiology, and modulation by identified octopaminergic neurons. J. Comp. Physiol. A 195:123-137. Veröffentlichung 6: Bicker G, Stern M (2009) NO als Regulator neuronaler Motilität und Regeneration in einfachen Nervensystemen. Neuroforum 1/09:4-12. 2 1 Verzeichnis von Veröffentlichungen die Bestandteil der Habilitationsschrift sind Veröffentlichung 7: Podrygajlo G, Tegenge MA, Gierse A, Paquet-Durand F, Tan S, Bicker G, Stern M (2009) Cellular phenotypes of human model neurons (NT2) after differentiation in aggregate culture. Cell Tissue Res. 336:439-452. Veröffentlichung 8: Tegenge MA, Stern M, Bicker G (2009) Nitric oxide and cyclic nucleotide signal transduction modulates synaptic vesicle turnover in human model neurons. J. Neurochem. 111:1434-1446. Veröffentlichung 9: Stern M, Böger N, Eickhoff R, Kerßen U, Lorbeer C, Ziegler M, Martinelli GP, Holstein GR, Bicker G (2010) Development of nitrergic neurons in the nervous system of the locust embryo. J. Comp. Neurol. 518:1157–1175. Veröffentlichung 10: Stern M (submitted) Serotonin controls development of serotonergic neurons in the enteric nervous system of the locust (Locusta migratoria). Neurosci. Lett., under review. 3 2 Einleitung 4 2 Einleitung 2.1. Einfache Nervensysteme 2.1.1. Das Nervensystem von Insekten Das menschliche Gehirn ist eine der komplexesten Strukturen im Universum. Will man sich der zellulären Basis seiner Entwicklung und Funktion nähern, ist es oft einfacher, grundsätzliche Vorgänge an weniger komplexen Nervensystemen oder gar an Zellkultursystemen zu untersuchen. Dabei muss natürlich die Komplexität des ausgewählten Modellsystems der Komplexität der Fragestellung entsprechen. Das Insektennervensystem ist bei aller Einfachheit hinreichend weit entwickelt, um vielfältige Sinnesleistungen, räumliche und zeitliche Orientierung und ein reichhaltiges Verhaltensrepertoire zu ermöglichen, das Lernen, Gedächtnis und Kommunikation einschließt (v. Frisch, 1965). Das Nervensystem von Insekten und vielen anderen Wirbellosen besteht aus dem Zentralnervensystem (ZNS) mit dem Gehirn und einer durch paarige Nervenstränge (Konnektive) verbundenen Kette von Ganglien (Abb. 1), dem enterischen Nervensystem (ENS) mit enterischen Ganglien und Nervenplexus (Abb. 2), und dem peripheren Nervensystem (PNS). Alle Ganglien und auch das Gehirn sind aus einer Rinde aus an der Informationsverarbeitung nicht beteiligten Zellkörpern und einer zentralen Neuropil-Region aufgebaut, in welcher sich Axone, Dendriten und synaptische Kontakte befinden. Über periphere Nerven steht das ZNS mit dem PNS in Verbindung. Im Gehirn befinden sich neben zentralen Steuereinheiten für das Verhalten auch die Verarbeitungssysteme für Informationen aus den wichtigsten Sinnesorganen, nämlich den Antennen und den Komplexaugen. Die Verarbeitung olfaktorischer Information von den Antennen erfolgt zunächst in den Antennalloben, und weiter in den Pilzkörpern. Visuelle Information von den Photorezeptoren wird in den optischen Loben des Gehirns in mehreren Verarbeitungsstufen zunehmender Komplexität parallel für jeden einzelnen Bildpunkt verarbeitet. Auf unterschiedlichen Ebenen wird Information von sogenannten Tangentialneuronen abgegriffen und zur Weiterverarbeitung ins zentrale Gehirn geleitet. In geringerem Maße erfolgt auch ein Informationsfluß in umgekehrter Richtung, vom zentralen Gehirn in die primären Verarbeitungszentren wie z.B. die optischen Loben, um dort die Informationsverarbeitung dem Verhaltenskontext entsprechend zu modulieren. Um diese Signalwege zu verstehen, ist es notwendig, die beteiligten Zellen (zentrifugale Neuronen) 2 Einleitung 5 anatomisch, physiologisch und auf die von ihnen verwendeten Neurotransmitter zu charakterisieren. Abb. 1: Die Heuschrecke hat ein einfaches Strickleiter-Nervensystem. Rechts ein Ausschnitt aus dem Gehirn mit einem intrazellulär markierten PM4Interneuron. (aus: Bicker und Stern, 2002) Als ENS bezeichnet man sowohl bei Wirbeltieren als auch bei Wirbellosen das System von Nervenfasern und Ganglien, die den Verdauungskanal innervieren. Beim Insekt ist es mit dem Gehirn und dem endokrinen System verbunden und reguliert die Nahrungsaufnahme, – verdauung und –weiterleitung im Darmtrakt, sowie andere Verhaltensmuster, an denen der Darm beteiligt ist, z.B. die Häutung, bei der das Insekt Luft schluckt, um durch die dadurch bedingte Volumenzunahme die alte Haut zu sprengen. Die vom ENS generierten, oft rhythmischen, neuronalen Entladungsmuster und ihre verhaltensabhängige Modulation werden seit Jahrzehnten an Crustaceen (review: Harris-Warrick und Marder, 1991) und in der Gegenwart auch bei Insekten (review: Ayali, 2004) untersucht. Außerdem hat sich das ENS in den letzten Jahren als ein interessantes Modellsystem für entwicklungsneurobiologische Fragestellungen herausgestellt. Im Insektennervensystem stellt das ENS eine Besonderheit dar. Im Gegensatz zu der relativ stereotyp ablaufenden Entstehung des ZNS, bei dem sich fest lokalisierte Neuroblasten im Stammzellmodus teilen, und festgelegte neuronale Nachkommenschaft produzieren (review: Goodman und Bate, 1981), sind bei der Bildung des ENS äquale Teilungen und umfangreiche Migrationsvorgänge beteiligt (review: Hartenstein, 1997). Damit ähnelt die ENS-Entwicklung beim Insekt eher der Gehirnentwicklung beim Wirbeltier, wo in der ventrikularen Zone geborene Neuronen an ihren Bestimmungsort im Cortex migrieren (review: Hatten, 1999). Das Insekten-ENS besteht aus Nervengeflechten, die Vorder- und Mitteldarm umspannen, sowie einigen damit assoziierten Ganglien und Nerven (Abb. 2). Es entwickelt sich aus einer kleinen Gruppe ektodermaler neurogener Placoden 2 Einleitung 6 Abb.2: Links: Übersicht über den Heuschreckendarm (aus Ganfornina et al., 1996) Rechts: Schematische Darstellung des ENS bei 40% (A) und 50% (B) der Embryonalentwicklung (aus Stern et al., 2007). c: caecum, ca: corpus allatum, en: Oesophagealnerv, f: Vorderdarm, fc, fgc: Frontalkonnective, fg: Frontalganglion, fp: Vorderdarmplexus h: Enddarm, hg: Hypocerebralganglion, hn: Enddarmnerven, ig: Ingluvialganglion, m, mg: Mitteldarm, mp: Mitteldarmplexus, mt: Migrationszunge, rn; recurrenter Nerv, nz1, nz2, nz3: neurogene Placoden. auf dem Vorderdarm, von denen aus Neuronen anterior und lateral migrieren, um die Ganglien des Vorderdarms zu bilden (Abb. 2). Später erfolgt eine zweite Migrationswelle, ausgehend von zungenförmigen Gruppen von Neuronen, die sich an der Grenze zwischen Vorder- und Mitteldarm sammeln, um nach Schließen des Mitteldarms in 4 Zugstraßen posterior zu migrieren und den Mitteldarmplexus zu bilden (Ganfornina et al., 1996; Haase und Bicker, 2003). 2.1.2 Zellkultursysteme Eine sinnvolle Alternative zur Untersuchung von mehr oder weniger intakten Nervensystemen kann die Arbeit mit geeigneten Zellkultursystemen sein. Insbesondere ist es aus verschiedenen Gründen erstrebenswert, ein Zellkultur-basiertes Modellsystem für die menschliche Gehirnentwicklung zur Verfügung zu haben (review: Paquet-Durand und Bicker, 2007). Dazu gehört die Erforschung der physiologischen Grundlagen und möglicher Therapieperspektiven bei einer Reihe von Erkrankungen des Zentralnervensystems, wie z.B. Schlaganfall (Casper et al., 2003), Parkinson (Biskup und Moore, 2006), Alzheimer (Roberson und Mucke, 2006), amyotrophe Lateralsklerose (Boilee et al., 2006), etc. Weiterhin besteht großer Bedarf an Testsystemen für neutoxikologische und aktuell entwicklungsneurotoxikologische Fragestellungen (Radio und Mundy, 2008; Hill et al., 2008; Stern et al., 2009b). Allerdings ist selbst für die neurowissenschaftliche Grundlagenforschung humanes embryonales Hirngewebe nur schwer und unter starken ethischen Bedenken zu erhalten. Für pharmakologische oder toxikologische Screening-Verfahren in Großmaßstab ist 2 Einleitung 7 dieser Weg erst recht nicht gangbar. Aufgrund von Spezies-Unterschieden können menschliche Neuronen nicht in jedem Fall durch leichter zugängliches Hirngewebe etwa von Nagetieren ersetzt werden. Im schlimmsten Fall können sich Pharmaka die in einer Spezies neuroprotektiv wirken, schädliche Effekte bei einer anderen Art bewirken (Munir et al., 1995; Hoyte et al., 2004). Als Alternative bieten sich menschliche Zelllinien an, wenn sich zeigen lässt, dass sich diese im Hinblick auf die zu bearbeitenden Fragestellungen natürlichem menschlichen Hirngewebe entsprechen. Ein Beispiel ist die Zelllinie Ntera-2 (NT2), eine aus einem menschlichen Hodentumor hervorgegangene neuronale Vorläufer-Zelllinie. Unter nicht-differenzierenden Bedingungen teilen die Zellen sich ca. alle 24 Stunden und exprimieren für neuronale Progenitorzellen typische Proteine wie z. B. die Intermediärfilamente Nestin und Vimentin, zeigen aber keine Marker für reife neuronale Zellen (Podrygajlo et al., 2009). Durch Behandlung mit dem Morphogen Retinsäure können NT2-Zellen zu postmitotischen Nervenzellen differenziert werden (Andrews, 1984; Pleasure et al., 1992). Ein Nachteil bei der Generierung reiner postmitotischer Neurone aus NT2-Zellen war bisher die lange dafür benötigte Zeit (ca. 2 Monate). Unter Verwendung einer Kulturmethode als sphärische Zellaggregate unter nichtadhärenten Bedingungen konnte die benötigte Zeit zur Gewinnung reiner Neurone in unserem Labor auf unter einen Monat verringert werden (Paquet-Durand et al., 2003, Abb. 3). Dazu werden NT2-Zellen in Gegenwart von 10 µM Retinsäure zunächst für eine Woche in nichtadhärenten Petrischalen kultiviert, so dass sich sphärische Aggregate bilden, in denen sich Neuronen entwickeln (Abb. 3b). Diese Zellaggregate werden weiterhin unter Retinsäure in adhärenter Kultur inkubiert, wobei die Zellen aus den Aggregaten auswandern (Abb. 3c). Danach folgt eine Behandlung mit Mitoseinhibitoren, so dass die nicht differenzierten Zellen absterben (Abb. 3d). Die übrig bleibenden postmitotischen Neuronen können dann gewonnen und auf geeignetem Untergrund in Kultur genommen werden (Abb. 3e) und bieten so eine regelmäßig verfügbare Quelle menschlicher Neuronen. 2 Einleitung 8 Abb. 3: Differenzierung von NT2-Zellen zu Neuronen (aus Podrygajlo et al., 2009) 2.2 Neuronale Botenstoffe 2.2.1 Neurotransmission In mehrzelligen Organismen kommunizieren Zellen miteinander hauptsächlich auf chemischem Wege. Im Nervensystem gibt es sowohl Kommunikation von Nervenzellen untereinander als auch mit Sinneszellen und effektorischen Zellen, beispielsweise in Muskeln und Drüsen. Diese Kommunikation erfolgt normalerweise an spezialisierten Strukturen, den Synapsen. An diesen Kontaktstellen bilden prä- und postsynaptische Zellen Spezialisationen ihrer Zytoplasmamembranen aus, die der Signalübertragung dienen. Auf der präsynaptischen Seite sind dies sogenannte aktive Zonen, in denen sich mit Neurotransmittern gefüllte Vesikel und die für die Fusion dieser Vesikel mit der präsynaptischen Membran verantwortliche Proteine befinden. Auf der postsynaptischen Seite sind dies vor allem Ansammlungen von Neurotransmitter-Rezeptoren. Ein durch eine Depolarisation der präsynaptischen Zelle ausgelöster Kalzium-Einstrom löst die Fusion der synaptischen Vesikel mit der Zytoplasmamembran aus, was zur Ausschüttung des Neurotransmitters führt. Dieser diffundiert über den synaptischen Spalt und bindet an der postsynaptischen Membran an für ihn spezifische Rezeptoren, die dann eine Reaktion der postsynaptischen Zelle bewirken. Die zwei wichtigsten Klassen von Neurotransmitter-Rezeptoren sind Rezeptor-Ionenkanäle und G-Protein gekoppelte Rezeptoren (Zimmermann, 1993). Rezeptor-Ionenkanäle sind meist aus 4 oder 5 Untereinheiten aufgebaut, die zusammen einen die Zytoplasmamembran durchquerenden Kanal bilden. Dieser öffnet sich bei Bindung des spezifischen Liganden und ermöglicht einen Ionenfluss über die Membran und bewirkt damit eine Membranpotential- 2 Einleitung 9 Wirkmechanismus Ionenkanal (nicotinisch) G-Protein gekoppelt (muscarinisch) in Vertebraten nicotinisch v.a. motorisch, muscarinisch im ZNS in Evertebraten vor allem sensorisch, häufiger Transmitter im ZNS, bei einigen Gruppen motorisch Ionenkanal G-Protein gekoppelt häufigster excitatoricher Transmitter im ZNS motorisch (bei einigen Gruppen) Glycin Ionenkanal, auch als Kofaktor bei Glutamat-Rezeptoren inhibitorischer Transmitter im ZNS nicht nachgewiesen GABA Ionenkanal G-Protein gekoppelt häufigster inhibitorischer Transmitter inhibitorischer Transmitter, auch motorisch G-Protein gekoppelt selten Ionenkanal modulatorisch, dämpfend, im Gehirn, excitatorisch im ENS modulatorisch, dämpfend, im ZNS, excitatorisch im ENS Dopamin G-Protein gekoppelt modulatorisch modulatorisch, Lernen Noradrenalin G-Protein gekoppelt modulatorisch, aktivierend nicht nachgewiesen Octopamin G-Protein gekoppelt nicht nachgewiesen modulatorisch, aktivierend, Lernen Tyramin G-Protein gekoppelt nicht nachgewiesen modulatorisch Histamin Ionenkanal G-Protein gekoppelt modulatorisch, Gewebshormon modulatorisch, in einigen Gruppen sensorisch (Auge) G-Protein gekoppelt modulatorisch modulatorisch, motorisch im ENS intrazelluläre Rezeptoren modulatorisch, retrograder messenger, Lernen modulatorisch, Lernen modulatorisch, retrograder messenger Name Acetylcholin Aminosäuren Glutamat Biogene Amine Serotonin Neuropeptide, z.B. Proctolin FMRFamide Gasförmige Transmitter Stickstoffmonoxid NO Kohlenstoffmonoxid CO intrazelluläre Rezeptoren Tab.1: einige häufige Neurotransmitter ENS-Entwicklung, sonst unklar 2 Einleitung 10 änderung. Dabei bestimmt die Auskleidung des Kanals mit unterschiedlich geladenen Aminosäureresten die eventuelle Spezifität des Rezeptors für bestimmte Ionen. G-Protein gekoppelte Rezeptoren bestehen aus einer einzigen, mehrmals die Membran durchspannenden Polypeptidkette, die bei Bindung des spezifischen Liganden intrazellulär mit einem trimeren G-Protein interagiert. Durch das so aktivierte G-Protein kann eine biochemische Signalkaskade angestoßen werden, vermittelt durch intrazelluläre second messenger wie z.B. die zyklischen Nucleotide cAMP und cGMP oder durch Kalzium. Als Neurotransmitter kommen sowohl niedermolekulare Substanzen als auch Peptide vor (Tab.1). Zu den niedermolekularen Substanzen gehören die klassischen Neurotransmitter wie Acetylcholin, die Aminosäuren Glutamat, Glycin und Gamma-Amino-Buttersäure (GABA), sowie die biogenen Amine Serotonin, Dopamin und Noradrenalin (bei Wirbellosen ersetzt durch Octopamin). Außer diesen vesikulär ausgeschütteten Neurotransmittern und Peptiden gibt es noch atypische neuronale Botenstoffe wie die Gase Stickstoffmonoxid (NO) und Kohlenstoffmonoxid (CO), welche nicht-vesikulär ausgeschüttet werden. Stattdessen, diffundieren diese kleinen apolaren Moleküle sofort nach ihrer Synthese in der "präsynaptischen" Zelle durch die Zellmembranen und binden im Inneren der EmpfängerZelle an zytoplasmatische Rezeptoren wie z.B. die lösliche Guanylyl-Cyclase (sGC), die daraufhin den second messenger cGMP produziert. 2.2.2 Synaptische Plastizität Unter synaptischer Plastizität versteht man die Veränderbarkeit der Stärke der synaptischen Übertragung. Die chemische Signalübertragung bietet vielfältige Möglichkeiten, die Informationsverarbeitung an der Synapse zu modulieren (review: Hawkins et al., 2006). Neuromodulation kann sowohl auf der präsynaptischen als auch auf der postsynaptischen Seite erfolgen. Bei präsynaptischer Modulation wird die bei einer Erregung ausgeschüttete Neurotransmittermenge verändert, bei postsynaptischer Modulation ändert sich die Antwortstärke auf eine gegebene Menge Neurotransmitter. Synaptische Plastizität spielt sich auf unterschiedlichen Zeitskalen ab. Sehr kurzfristige Effekte können durch temporär veränderte Ionenkonzentrationen (vor allem Kalzium) ausgelöst werden. Mittelfristige Veränderungen beruhen meist auf durch biochemische Signalkaskaden angestoßene kovalente Modifikationen (z.B. Phosphorylierung/Dephosphorylierung) von Proteinen. Dies können z.B. Rezeptoren oder Ionenkanäle sein, die die Erregbarkeit der Zelle verändern, oder auch Proteine, die die vesikuläre Transmitterfreisetzung beeinflussen. Langfristige Veränderungen an Synapsen beruhen meist auf differentieller Genaktivität, und beinhalten geänderte 2 Einleitung 11 Expressionsmuster von prä- oder postsynaptischen Proteinen, wie z.B. Rezeptoren und Transportern, aber auch strukturelle Veränderungen, d.h. Wachstum von Neuriten und Neubildung von synaptischen Strukturen. Insgesamt deckt der Begriff "Neuronale Plastizität" sowohl erfahrungsabhängige Prozesse (Habituation, assoziatives Lernen) als auch erfahrungsunabhängige Entwicklungsprozesse ab. 2.2.3 Neuronale Botenstoffe in der Entwicklung In den letzten Jahrzehnten hat sich mehr und mehr herausgestellt, dass der Einfluss von Neurotransmittern auf strukturelle synaptische Plastizität eigentlich nur ein Spezialfall ist. Tatsächlich haben Neurotransmitter einen erheblichen Einfluss auf alle Aspekte der Entwicklung im Nervensystem, und vielerorts auch in nicht-neuronalen Geweben. Dies betrifft so unterschiedliche Vorgänge wie die Proliferation von neuronalen Vorläuferzellen, neuronale Differenzierung, Migration und Auswachsen von Neuronen, Synaptogenese und synaptische Reifung (reviews: Lauder, 1988; Nguyen et al., 2001). Acetylcholin zum Beispiel, bei Wirbeltieren ein klassischer Neurotransmitter sowohl in der Peripherie wie im zentralen Nervensystem, hat in der Hirnentwicklung Auswirkungen auf Zellproliferation, Migration, Überleben von Nervenzellen und Auswachsen von Neuriten bei diversen Wirbeltieren und Wirbellosen (Lauder und Schambra, 1999). Störungen des cholinergen Systems während der Entwicklung, z.B. durch Pestizide, können deshalb zu Hirnschäden führen. Auch GABA, der wichtigste inhibitorische Transmitter im ZNS, hat während der Hirnentwicklung einen wichtigen Einfluss auf Proliferation, Migration und Synaptogenese (review: Wang und Kriegstein, 2009). Serotonin hat außer seiner seit langem gut bekannten Rolle als Neurotransmitter und Neuromodulator auch morphogene Eigenschaften in der Entwicklung des Nervensystems und anderer Organe sowohl bei Wirbeltieren als auch bei Evertebraten. Im Wirbeltier ist es zum Beispiel an der Regulierung der Synaptogenese beteiligt (Mazer et al., 1997) oder bei der Neuralleistenzellwanderung in der Maus (Moiseiwitsch und Lauder, 1995). Bei Evertebraten ist die entwicklungsbiologische Rolle von Serotonin vor allem bei Mollusken untersucht worden. In der Schnecke Tritonia reguliert 5-HT Zellteilungen schon vor der Ausbildung eines Nervensystems (Buznikov et al., 2003). Später ist Serotonin im Schneckennervensystem an der Steuerung von Neuritenwachstum und –verzweigung beteiligt (Goldberg und Kater, 1989; Diefenbach et al., 1995). Auch bei Krebsen hat Serotonin Einfluss auf Auswachsen und Verzeigen von Neuronen (Sullivan at al., 2000). Im Insektenembryo ist vor allem die Entwicklung einer kleinen Gruppe von serotonergen Neuronen in ZNS sowohl in 2 Einleitung 12 Heuschrecken (Taghert und Goodman, 1984; Condron, 1999; Stern und Bicker, 2008; siehe Kapitel 4.3) als auch bei Drosophila sehr gut untersucht (Valles und White, 1988; Lundell und Hirsh, 1998; Sykes und Condron, 2005). Weniger gut untersucht ist die Entwicklung des serotonergen Systems im enterischen Nervensystem (ENS), mit der sich die vorliegende Arbeit beschäftigt (Stern et al., 2007). Dass Stickstoffmonoxid (NO) mit seinem second messenger cGMP eine neuromodulatorische Wirkung im Wirbeltiergehirn haben kann, ist seit mehreren Jahren bekannt (zusammengefasst in Garthwaite, 2008). Auch im Nervensystem von Insekten kommt das NO/cGMP-System vor. Sowohl die Freisetzung von NO durch eine Ca2+/Calmodulin stimulierte NOS aus kultivierten Neuronen als auch die NO-abhängige Synthese von cGMP in den Zielzellen konnten im Insektengehirn dargestellt werden (Müller und Bicker, 1994; Bicker et al., 1997). Einige funktionelle Erkenntnisse über die Modulation neuronaler Plastizität in Insektennervensystemen sind in den letzten Jahren gewonnen worden. So wirkt NO zum Beispiel als retrograder Botenstoff bei der Lichtadaptation im visuellen System der Heuschrecke (Schmachtenberg und Bicker, 1999) oder es stimuliert die Freisetzung von Neurotransmittervesikeln an der neuromuskulären Synapse von Drosophila-Larven (Wildemann und Bicker, 1999). Ausschüttung von NO kann im Suboesophagealganglion der Heuschrecke motorische Muster auslösen (Rast, 2001) und im Zentralkomplex des Hirns das Stridulationsverhalten von Grashüpfern unterdrücken (Wenzel et al., 2005; Weinrich et al., 2008). Insgesamt sind die Erkenntnisse über NO-vermittelte Neuromodulation allerdings noch sehr lückenhaft, und es sind in den nächsten Jahren sicher noch weitere Ergebnisse zu erwarten. Es mehren sich auch die Hinweise of eine wichtige Bedeutung des NO/cGMP-Signalweges für die Entwicklung von Nervensystemen, im Wirbeltier z.B. bei der Neurogenese, bei der Migration und beim Neuritenwachstum (Hess et al., 1993; Packer et al., 2003; Moreno-Lopez et al., 2004). Auch im Insektennervensystem gibt es seit einigen Jahren Hinweise auf eine Wirkung von NO auf Entwicklungsprozesse (reviews: Bicker, 2007; Bicker und Stern, 2009). Null-Mutationen im NOS-Gen führen bei Drosophila zu embryonaler oder frühlarvaler Letalität (Regulski et al., 2004), woraus abzuleiten ist, dass NO als essentieller Entwicklungsregulator fungiert. Schon recht gut untersucht ist die Wirkung von NO auf die Migration enterischer Neurone auf dem Mitteldarm der Heuschrecke (Haase und Bicker, 2003; Knipp und Bicker, 2009a). Die in einer neurogenen Zone des ektodermalen Vorderdarms geborenen Neurone des Mitteldarmplexus durchlaufen eine Phase schneller 2 Einleitung 13 Zellwanderung, während der sie sich als „Kettenmigration“ auf vier geradlinig verlaufenden Zugstraßen über die Oberfläche des Mitteldarms bewegen, um sich danach auf dem Darm zu verteilen. Während der Migrationsphase bilden die enterischen Neuronen NO-abhängig cGMP. Es gibt verschiedenen Möglichkeiten, pharmakologisch in die NO/cGMPSignalkaskade einzugreifen (Abb. 4) Durch pharmakologische Manipulationen im Embryo-Kultursystem konnte gezeigt werden, dass das NO/cGMP-System die Geschwindigkeit dieser Migration reguliert (Haase und Bicker, 2003; Knipp und Bicker, 2009a). Blockierung des Signalweges führt zu einer Verlangsamung der Migration. Abb. 4: NO-cGMP-Signalweg und pharmakologische Eingriffsmöglichkeiten. Aktivierende Pharmaka sind grün unterlegt, blockierende rot. Außerdem kann man NO in Form von spontan zerfallenden und dabei NO freisetzenden Substanzen applizieren (blau). (aus Bicker und Stern, 2009) Auch an anderen Stellen des peripheren Nervensystems ist eine Beteiligung des NO/cGMPSignalweges an Entwicklungsvorgängen bekannt. In den Extremitätenanlagen von Insektenembryonen legen sogenannte Pionierneurone die erste Bahn von der Peripherie ins 2 Einleitung 14 ZNS, der später sensorischen und motorischen Axone folgen. Diese Pionierneurone sind cGMP-positiv und ihr Auswachsen ist z.B. in der Heuschreckenantenne abhängig von NO (Seidel und Bicker, 2000). Auch in den Beinanlagen lässt sich dies zeigen (Pätschke und Bicker, 2007). Dies passt zu Zellkultur-Untersuchungen an Wachstumskegeln von Neuronen der Schnecke Helisoma, deren Wachstumsverhalten konzentrationsabhängig von NO reguliert wird (Trimm und Rehder, 2004). Ob und wo NO auch bei der Entwicklung des Zentralnervensystems von Insekten eine Rolle spielt, ist bisher erst ansatzweise untersucht. In der Puppe von Drosophila ist das NO-cGMP-system bei der Umgestaltung des visuellen Systems, insbesondere beim Aufbau der retinalen Projektionen in den optischen Lobus des Gehirns beteiligt (Gibbs und Truman, 1998; Gibbs et al., 2001). In der Puppe des Tabakschwärmers (Manduca sexta) beeinflusst NO im optischen Lobus die Steroidhormonkontrollierte Zell-Proliferation (Champlin und Truman, 2000). Ebenfalls in TabakschwärmerPuppen konnte gezeigt werden, dass bestimmte Neuronen im Gehirn NO-abhängig cGMP synthetisieren (Schachtner et al., 1998). Mit Ausnahme einer Studie von Ball und Truman (1998), die eine NO-abhängige Guanylyl-Cyclaseaktivität in bestimmten Neuronen zeigte, die zeitlich mit der Phase der Synaptogenese korrelierte gab es aus dem Zentralnervensystem des Heuschreckenembryos bisher keine Informationen über das Vorhandensein oder gar eine mögliche entwicklungsbiologische Beteiligung des NO-cGMP-Systems während der Embryonalentwicklung. Gerade über diese Spezies existieren aber besonders viele entwicklungsbiologische Arbeiten über die Entstehung des Zentralnervensystems. Vor allem die Pionierarbeiten am Bauchmark aus der Gruppe um C. S. Goodman aus den 1980er Jahren (review: Goodman und Bate, 1981; Thomas et al., 1984; Sanchez et al., 1995) und die Arbeiten über die Hirnentwicklung von G. S. Boyan, J. L. Williams und Mitarbeitern aus den letzten 15 Jahren (z.B. Boyan und Williams, 2000; Boyan et al., 2008) sind hier zu nennen. Darum besteht hier dringender Forschungsbedarf. 3 Fragestellungen 15 3 Fragestellungen Aus dem Themenkomplex der Bedeutung neuroaktiver Botenstoffe für die neuronale Plastizität wurden folgende Fragestellungen bearbeitet und werden im Folgenden vorgestellt: 1. Wie ist octopaminerge Modulation synaptischer Übertragung im visuellen System im Heuschreckenhirn organisiert, und welche Rolle spielen dabei zentrifugale Neuronen ? 2. Wie entwickeln sich serotonerge Neuronen im Heuschrecken-ENS und gibt es Evidenz für eine funktionelle Bedeutung von Serotonin bei dessen Entwicklung? 3. Ist Stickstoffmonoxid an Entwicklungsvorgängen im Zentralnervensystem der Heuschrecke beteiligt? 4. Können Entwicklungsvorgänge unter dem Einfluß neuronaler Botenstoffe in einem menschlichen neuronalen Zellkultursystem untersucht werden? 5. Sind Neurotransmitter auch geeignet, neuronale Plastizität unter einem phylogenetischen Aspekt zu betrachten? 4 Ergebnisse 16 4 Ergebnisse 4.1 Zentrifugale Neuronen in der Sehbahn der Heuschrecke und ihre Modulation durch Octopamin (Veröffentlichung 5) Im visuellen System der Heuschrecke tritt Neuromodulation zum Beispiel bei Habituationsund Dishabituationsvorgängen im Bewegungsdetektor-System auf. Eine Anzahl von bewegungssensitiven visuellen Neuronen hat Aufgaben bei der Kollisionsvermeidung im Flug sowie bei der Detektion von Feinden und der Einleitung von entsprechenden Ausweich- oder Fluchtmanövern. Dieses Bewegungsmelder-System zeigt Habituation bei ständiger Wiederholung desselben Bewegungs-Reizes, der daraufhin fast überhaupt nicht mehr beantwortet wird. Dies ist biologisch sinnvoll, da es inadäquat wäre wenn die Heuschrecke z.B. auf ein regelmäßig im Wind flatterndes Blatt mit Fluchtsprüngen reagieren würde. Wenn jedoch in der Umgebung des Tieres Veränderungen auftreten, die auf eine mögliche FeindAnnäherung hinweisen könnten, muss das System wieder “aufmerksam” werden und dishabituieren. Dies ist für das bewegungssensitive DCMD-Neuron schon seit langem bekannt (Rowell, 1971). Eine ganze Reihe von Reizen (Licht, Geräusche, Wind, Berührung) lösen dort Dishabituation aus. Neuronen, die diese Dishabituation vermitteln können, müssen drei Kriterien erfüllen: 1. sie müssen aus dem zentralen Gehirn in den optischen Lobus projizieren, 2. sie müssen auf die Dishabituation auslösenden Reize antworten, und 3. sie müssen einen Neuromodulator ausschütten, der Dishabituation bewirken kann. Es gibt im Heuschreckenhirn einige für diese Aufgabe in Frage kommende Neuronen, die experimentell gut zugänglich sind, da ihre bis zu 60 µm großen Zellkörper unter dem Präparationsmikroskop freigelegt und identifiziert werden können (Stern et al., 1995). Eines von diesen, das Neuron PM1, wurde im Laufe der vorliegenden Arbeit charakterisiert (Stern, 2009). Es existieren auf jeder Seite des Gehirns zwei Exemplare dieses Typs, die anatomisch und physiologisch nicht zu unterscheiden sind. Sie verzweigen sich im Proto- und Deutocerebrum, wo auch der Zellkörper lokalisiert ist, und senden ihr Axon in den optischen Lobus, wo sie sich in eine ausgedehnte Arborisation in der Medulla verästeln (Abb. 5A). PM1 ist multimodal und antwortet auf visuelle Reize (Licht, Objektbewegung), sowie auf 4 Ergebnisse 17 Abb. 5: Das Neuron PM1 und seine Modulation durch Octopamin. Morphologie gezeichnet nach A: intra- zellulärer Kobaltfärbung. B-C: Doppelfärbung GAD (rot) und PM1 intrazellulär (grün) im optischen Lobus (B) und im zentralen Gehirn (C). D: Zeitverlauf der Antworten von PM1 auf wiederholte Bewegungsreize vor und nach BadApplikation von 5x10-4 M Octopamin. E Schematische Darstellung eines Versuchs zum Einfluss des octopaminergen PM4 auf PM1. F: Simultanableitungen von PM4 und PM1, PM4-Aktivität verstärkt die Antworten von PM1. al: Antennallobus, lob: Medulla, Protocerebrum, pc: ventrolaterales Lobula, med: vlp: Protocerebrum. (verändert aus: Stern, 2009) mechanische Stimulation der Antennen und Haarsensillen am Kopf. Das Antwortverhalten auf visuelle Bewegungsreize entspricht dem einer ganzen Reihe von visuellen Interneuronen der Heuschrecke (Rind, 1987; Gewecke und Hou, 1993; Stern und Gewecke, 1993), deren Aufgabe wahrscheinlich im Kontext von Fluchtauslösung, Kollisionsvermeidung und ganz allgemein für Objektdetektion liegt. Es antwortet auf Bewegungen vor allem kleiner Objekte, hat keine Präferenz für eine bestimmte Bewegungsrichtung, und zeigt schnelle Habituation bei Wiederholung des selben Reizes (Stern, 2009). Der Neurotransmitter dieses Neurons ist GABA, was mit Hilfe von Doppelmarkierungen mit einem in den Zellkörper injizierten Farbstoff und Antikörpern gegen GABA oder das GABA synthetisierende Enzym GlutamatDecarboxylase (GAD) nachgewiesen werden konnte (Abb. 4B, C). Die funktionelle Polarität (Eingangsregion im zentralen Hirn, Ausgang im optischen Lobus) wird durch das Vorhandensein von GAD in den Verzweigungen im optischen Lobus, aber dessen Fehlen in den Verzweigungen im zentralen Gehirn angezeigt. Damit konnte GAD hier zum ersten Mal im Insektenhirn als funktioneller Polaritätsmarker verwendet werden. Bestätigt wird dieser Befund durch doppelte intrazelluläre Ableitungen vom Zellkörper und aus dem Axon im optischen Lobus, die eine Leitungsrichtung vom Zentralhirn in die Peripherie anzeigen. Die funktionelle Bedeutung des PM1-Neurons ist noch nicht abschließend geklärt, kann aber im 4 Ergebnisse 18 Zusammenhang damit stehen, dass die Antworten des bewegungssensitiven visuellen Systems auf Antennenbewegungen durch Ausschüttung von GABA unterdrückt oder abgemindert werden. So kann verhindert werden, dass das Tier von Bewegungen der eigenen Fühler erschreckt und inadäquat zur Flucht veranlasst wird. In der vorliegenden Arbeit wurden die Eigenschaften von PM1 als bewegungssensitives visuelles Interneuron genutzt, um die neuromodulatorische Beeinflussung von Habituationsvorgängen zu untersuchen. Es ist bereits in vorangegangen Arbeiten gezeigt worden, dass Bad-appliziertes Octopamin (das Wirbellosen-Analogon zu unserem Noradrenalin) Dishabituation in einem visuellen Neuron bewirken kann, genauso wie durch elektrische Stimulation bewirkte Octopaminausschüttung aus einem weiteren zentrifugalen Neuron, dem in Abb. 1 abgebildeten PM4 (Stern et al., 1995; Bacon et al., 1995; Stern, 1999). Bei den Untersuchungen am DCMD wurde mit Hilfe extrazellulärer Ableitungen vom Halskonnektiv gearbeitet. Das in dieser Studie untersuchte PM1-Neuron bietet den Vorteil der leichten Zugänglichkeit für intrazelluläre Untersuchungen, die es ermöglichen, einen direkten Effekt des Neuromodulators auf das untersuchte Neuron auszuschließen. Vielmehr muss angenommen werden, dass die Neuromodulation an den Eingangssynapsen der der bewegungssensitiven Neuronen im optischen Lobus erfolgt. In der vorliegenden Arbeit konnte gezeigt werden, daß die Habituation der Antwort von PM1 auf wiederholte visuelle Bewegungsreize durch Bad-Applikation von Octopamin aufgehoben werden kann (Abb. 5D) und dass die Habituation statt dessen auch durch gleichzeitige intrazelluläre Stimulation des octopaminergen Neurons PM4 aufgehoben wird (Abb. 5E, F). Diese Modulation des PM1-Antwortverhaltens kann pharmakologisch durch Antagonisten des Octopamin-Rezeptors inhibiert werden. Somit wissen wir jetzt, dass die Modulation der Habituation durch PM4 nachgewiesenermaßen octopaminerg ist, und dass octopaminerge Modulation visueller Habituation nicht auf das DCMD-Neuron beschränkt ist, sondern wahrscheinlich weit im visuellen System verbreitet ist. Mit dem hier charakterisierten PM1-Neuron gibt es jetzt ein für intrazelluläre Untersuchungen leicht zugängliches Testsystem für die Funktionsanalyse des visuellen Systems und seiner Beeinflussung durch neuronale Botenstoffe, wie hier am Beispiel des biogenen Amins Octopamin gezeigt wurde. 4 Ergebnisse 19 4.2 Serotonin bei der Entwicklung des enterischen Nervensystems der Heuschrecke (Veröffentlichungen 5 und 10) Abb. 6 Serotonin im enterischen Nervensystem der Heuschrecke. (verändert aus: Stern et al., 2007) Oben: Halbschematische Zeichnung des Darms einer L1-Heuschreckennymphe, Serotonin-Immunreaktivität ist schwarz eingezeichnet. Unten: Detailfotos des Präparates, das der Zeichnung zugrunde lag (von links: Frontalganglion, Vorderdarmplexus, Ingluvialganglion, Mitteldarmplexus), Abkürzungen: ca: Caecum, en: Oesophageal-Nerv, fc: Frontal-Konnective, fg: Frontalganglion, fp: Vorderdarmplexus, hg: Hypocerebralganglion, hig: Enddarm, ig: Ingluvialganglion, mp: Mitteldarmplexus, rn: recurrenter Nerv Serotonin findet sich im gesamten enterischen Nervensystem in den Plexus des Vorder- und Mitteldarms, sowie in den Neuropilregionen der enterischen Ganglien (Abb. 6). Bis auf wenige Ausnahmen sind diese auf Fortsätze von etwa 40 serotonin-immunreaktiven Zellkörpern im Frontalganglion zurückzuführen. Im Verlaufe der vorliegenden Arbeit wurde die embryonale Entwicklung dieser Zellen analysiert. Die ersten Serotonin-immunreaktiven Zellen traten auf kurz bevor der Embryo 50% seiner Entwicklung abgeschlossen hatte. Um die dann sehr schwache Färbung zu verstärken und auch Details sichtbar zu machen, wurden die Präparate vor der Fixierung mit 5 µM Serotonin präinkubiert. Es ist bekannt, dass serotonerge Neuronen unter diesen Bedingungen Serotonin aus dem Kulturmedium aufnehmen. Der dieser Serotonin-Aufnahme zugrundeliegende spezifische Serotonin- 4 Ergebnisse 20 Transporter (SERT) ist sowohl in Wirbeltieren als auch bei Drosophila gut charakterisiert (Barker et al., 1998) und kommt auch in der Heuschrecke vor (Condron, 1999). Schon bei erst zu 40% entwickelten Embryonen ließ sich SERT-Aktivität in einigen Neuronen des sich gerade bildenden Frontalganglions zeigen. Ohne Präinkubation mit Serotonin zeigen die Zellen zu diesem Zeitpunkt noch keine Serotonin-Immunreaktivität. Die SERT-Aktivität ist spezifisch, denn sie lässt sich mit einem spezifischem SerotoninaufnahmeBlocker, dem Antidepressivum Fluoxetine (Prozac) vollständig hemmen. Erst später beginnen die Zellen mit der Serotonin-Synthese, so dass sich im Frontalganglion während der Entwicklung immer mehr Zellen finden die zur Serotonin-Aufnahme befähigt sind, als schon selbst Serotonin synthetisieren. Erst mit dem Abschluss der Entwicklung, bei ca. 80% der Embryonalentwicklung gleichen sich die Zahlen an, und alle SERT-aktiven Neuronen produzieren auch selbst Serotonin (Abb. 7). Die Aufnahme der SERT-Aktivität Abb. 7 Serotonin (schwarz) und SERT (weiß) im Frontalganglion während der Entwicklung des Heuschreckenembryos (aus Stern et al., 2007). Links: Jeder Punkt ist der Mittelwert ± S.E.M. der Anzahl immunreaktiver Zellkörper aus mindestens fünf Präparaten. (105% = frisch geschlüpfte Larven des ersten Stadiums (L1). Rechts: Beispiel, SERT bei 53%, Pfeil zeigt auf migrierendes Neuron, das schon SERT-Aktivität zeigt. Maßstab: 100 µm. beginnt schon sehr früh, oft noch während der Migration der Neuronen von der neurogenen Zone zu ihrem Zielort im Frontalganglion (Abb. 7). Nach Erreichen ihres Bestimmungsortes bilden die serotonergen Zellen einen posterior wachsenden Neuriten, der sich im Hypocerebralganglion gabelt und über die paarigen Ingluvialganglien den Mitteldarmplexus innerviert. Dabei scheinen die auswachsenden serotonergen Fortsätze den über den Mitteldarm migrierenden Neuronen der zweiten Migrationsphase dicht zu folgen. Ob dieses Phänomen eine rein zeitliche Koinzidenz darstellt, oder ob die migrierenden Neuronen den für die nachfolgenden Fasern bestimmten Weg im Sinne eines Pionierneurons (Goodman und Bate, 1981) vorgeben, wurde mit einem pharmakologischen Manipulationsexperiment 4 Ergebnisse 21 untersucht (Stern et al., 2007). Dazu wurden Heuschreckenembryonen dorsal geöffnet, um Zugang zum Darm zu gewährleisten, und für 24 h in Kultur genommen, wobei bei einer experimentellen Gruppe das NO-cGMP-Signalsystem mit dem NO-Synthase-Inhibitor 7-NI ausgeschaltet wurde (siehe Kap. 4.3). Diese Manipulation verlangsamt die Migration der NOresponsiblen Neuronen, sollte aber keinen Effekt auf die nicht NO-responsiblen auswachsenden serotonergen Neuriten haben. Trotzdem verlangsamte sich nicht nur die neuronale Migration sondern in gleichem Maße auch das Auswachsen der serotonergen Neuriten, die stets hinter dem am weitesten migrierten Mitteldarmneuron blieben. Damit kann der Pioniercharakter der migrierenden Neuronen als erwiesen gelten. Es stellte sich nun die Frage nach der möglichen Funktion der frühen Synthese von Serotonin, und der noch früheren Serotonin-Aufnahmekapazität der Neuronen des Frontalganglions. Da zu diesem frühen Zeitpunkt noch nicht einmal der Mitteldarm ausgebildet ist, und auch die an der Rhythmus-Genese für die Darmperistaltik maßgeblichen Ingluvialganglien nicht ausgebildet sind, ist eine andere Rolle als die Steuerung der Darmmotorik wahrscheinlich. Da Serotonin neben seinen Funktionen als Neurotransmitter auch für seine entwicklungssteuernden Eigenschaften bekannt ist, sollte in Gewebekulturversuchen das serotonerge System manipuliert werden, um eventuelle Auswirkungen auf das sich entwickelnde ENS festzustellen (Stern, z. Publ. eingereicht). Abb. 8 SERT-Expression in enterischen Neuronen wird durch Serotonin reguliert. A-C: ENS gefärbt gegen Horseradish-Peroxidase als generellem neuronalen Marker (grün) und Serotonin (rot), wobei die Ganglien vor der Fixierung für 20 min in 5 µM Serotonin präinkubiert worden, so dass Zellen die den Serotonin-Transporter SERT exprimieren, Serotonin aus dem Medium aufnehmen konnten. A: Start bei 47%, B: nach 48 h in Kultur in normalem Zellkulturmedium (L15), C: ohne Kultur, 53%, das Stadium das nach 48 h in Kultur erreicht wird. D: Gemittelte Anzahl (±S.E.M.) SERT-positiver Zellen pro Frontalganglion aus 10 Präparaten wie in A-C. E: Gemittelte Anzahl (±S.E.M.) SERT-positiver Zellen pro Frontalganglion aus 10 Präparaten von 40% ENS vor bzw. nach 48 h in Kultur in ovo oder in L15, L15 mit 1 µM Serotonin (5-H-T) und/oder 2,5 µM des SerotoninAntagonisten Mianserin. Statistische Signifikanzen (ANOVA mit Tukey's post hoc test): ** p<0,01, n.s. nicht signifikant (verändert aus Stern, z. Publ. eingereicht). 4 Ergebnisse 22 Es zeigte sich, dass in enterischen Nervensystemen in Gewebe-Kultur mehr serotonerge Neuronen gebildet werden als normal. Bei der Entwicklung von 47% nach 53% steigert sich normalerweise die Anzahl der SERT-exprimierenden Neuronen von 22 auf 30 (Abb. 8A, C, D). Nimmt man 47% ENS für 48 h in Kultur, was ungefähr einem Stadium von 53% entspricht, steigert sich die Zahl jedoch auf ca. 45 (Abb. 8B, D), das sind mehr Zellen als selbst im adulten Stadium Serotonin synthetisieren. Während die Hämolymphe von Insektenembryonen Serotonin in hohen nanomolaren Konzentrationen enthält (Colas et al., 1995), befindet sich in normalem L15 Zellkulturmedium kein Serotonin. Darum wurde als nächstes dem Medium, in dem ENS kultiviert wurden, Serotonin beigegeben. Tatsächlich konnte die Zugabe von 1µM Serotonin verhindern, dass die Anzahl SERT-positiver Zellen in Zellkultur unnormal anstieg (Abb. 8E). Es gab keinen Unterschied mehr zu zeitgleich in ovo (bei natürlicher Serotonin-Konzentration) kultivierten Embryonen. Da die absolute Zellzahl pro Ganglion sich zwischen den Versuchen nicht signifikant unterschied, kann ein indirekter Effekt, z.B. über Beeinflussung von Proliferation oder Migration, ausgeschlossen werden. Es ist vielmehr so, dass der Anteil der serotonergen Zellen an der Gesamtzellzahl im Frontalganglion steigt. Der inhibierende Effekt des Serotonins auf die Ausprägung des serotonergen Phänotyps war spezifisch, denn er ließ sich durch gleichzeitige Zugabe des Serotoninrezeptor-Antagonisten Mianserin aufheben (Abb. 8E). Damit ist hier zum ersten Mal gezeigt worden, dass Serotonin einen direkten Einfluß auf die Expression eines TransmitterPhänotyps haben kann. 4 Ergebnisse 23 4.3 Stickstoffmonoxid im Zentralnervensystem der Heuschrecke und sein Einfluß auf axonale Regeneration (Veröffentlichungen 1, 2, 3, 6 und 9) In diesen Arbeiten wurde untersucht, ob ähnlich wie im peripheren Nervensystem, NO auch Wachstumsvorgänge im Zentralnervensystem reguliert. Dazu musste zunächst untersucht werden, ob die NO-Synthase während der Entwicklung des ZNS dort sowohl vorhanden als auch aktiv ist. Hierzu wurden Komponenten des NO-cGMP-Signalweges im embryonalen und postembryonalen Nervensystem mit vier verschiedenen Nachweismethoden dargestellt (Stern et al., 2010). Dies sind die NADPH-Diaphorase-Technik, immunzytochemische Markierung der NOS, immunzytochemische Markierung von Citrullin und immunzytochemische Markierung von cGMP. Die NADPH-Diaphorase-Technik macht es sich zu Nutze, dass 1. die NO-Synthase NADPH umsetzt und 2. dass sie im Gegensatz zu anderen NADPH umsetzenden Enzymen beständig gegenüber (milder) Fixation ist. Diese Färbereaktion wird allgemein als Nachweis für das Vorhandensein von NOS akzeptiert, und zwar sowohl bei Vertebraten (Dawson et al., 1991; Hope et al., 1991) als auch bei Evertebraten (Müller and Bicker, 1994). Allerdings gibt es auch einige Beispiele, bei denen die NADPH-Diaphorase-Reaktion sowohl falsch positive als auch falsch negative Ergebnisse erzielte (z.B. Gibson and Nighorn, 2000). Darum wurde als Ergänzung ein universelles Antiserum gegen NOS (uNOS) eingesetzt, das gegen eine konservierte Region sämtlicher im Wirbeltier vorkommender NOS-Proteine gerichtet ist, und das auch in Wirbellosen zur Markierung der NOS bereits erfolgreich verwendet worden ist (Kurylas et al., 2005). Dieses Antiserum wurde sowohl an Gewebeschnitten als auch im Western Blot eingesetzt. Als weitere Ergänzung diente der Nachweis von Citrullin. Diese Aminosäure entsteht als Nebenprodukt bei der Synthese von NO und wird im Nervensystem nicht schnell enzymatisch abgebaut (Martinelli et al., 2002). Somit wird Citrullin in NO-produzierenden Zellen akkumuliert und kann dort nach entsprechender Fixierung mit einem monoklonalen Antikörper nachgewiesen werden. Während mit diesen drei Methoden die Sender-Seite des NO-cGMP-Signalweges lokalisiert werden kann, dient die immunzytochemische Färbung von cGMP der Lokalisation der Empfänger-Seite. Dazu wird ein Präparat vor der Fixierung mit einem NO-Donor und einem Phosphodiesterase-Inhibitor inkubiert, so dass NO-abhängig cGMP gebildet wird, und nicht von Phosphodiesterasen wieder abgebaut werden kann. 4 Ergebnisse 24 Es ergab sich an Paraffinschnitten des adulten Nervensystems eine gute Übereinstimmung zwischen NADPH-Diaphorase-Färbung, uNOS und Citrullin, mit einer Ausnahme. Citrullin ließ sich im Pilzkörper nicht nachweisen, obwohl dort mit den anderen Färbemethoden eine deutliche und strukturierte Färbung vorhanden war. Dies mag auf den ersten Blick überraschen, findet seine Erklärung jedoch darin, dass die Neuronen der Pilzkörper nur bei koinzidentem Input von bestimmten Eingängen verschiedener Sinnesmodalitäten aktiv sind (sogenanntes 'sparse coding', Perez-Orive et al., 2002), und deshalb wahrscheinlich die meiste Zeit kein NO freisetzen. Dieses Ausbleiben der Citrullin-Färbung im Pilzkörper ist daher auch ein guter Nachweis der Aktivitätsabhängigkeit dieser Färbung. Abb 9: Schematische Darstellung der Entwickung des NO-cGMP-Signalweges im Heuschreckengehirn (aus Stern et al., 2010). Zeitpunkte (in Prozent der Embryonalentwicklung) der ersten Nachweisbarkeit von Komponenten des NO/cGMP-Signalweges im Heuschreckengehirn. al: Antennallobus, cbl: Zentralkomplex, untere Einheit, cbu: Zentralkomplex, obere Einheit, lam: Lamina, lob: Lobula, med: Medulla, ol dist: distaler optischer Lobus, ol prox: proximaler optischer Lobus, pc: Protocerebrum (Einzelzellen). 4 Ergebnisse 25 Im Folgenden wurde das Vorhandensein und die Aktivität der NOS im Verlauf der Embryonalentwicklung des Heuschrecken-ZNS untersucht. Im Western Blot von ganzen Embryonen ließ sich NOS ab 30% der Embryonalentwicklung nachweisen. Im Gehirn wurden die ersten Zellen bei 43% mit der NADPH-Diaphorase-Technik angefärbt, im Bauchmark bei 47%. Im Vergleich zeigte sich, dass die Citrullin-Färbung wesentlich später einsetzte als die NADPH-Diaphorase-Färbung. Dies lässt darauf schließen, dass das Enzym zwar schon früher vorhanden aber noch nicht konstitutiv aktiv ist. In den meisten Regionen war die von NO induzierbare cGMP-Färbung noch früher nachzuweisen als NADPH-Diaphorase und Citrullin. Dementsprechend existiert der Rezeptor schon lange vor dem Liganden. Auffallend ist auch, dass die NOS in den wichtigsten Neuropil-Regionen des Gehirns wie den Pilzkörpern, dem Zentralkomplex, den Antennalloben und den optischen Loben erst nachgewiesen werden kann, wenn diese zumindest in ihrem grundlegenden Aufbau schon etabliert sind (Abb. 9). Dementsprechend ist es unwahrscheinlich, dass der NO-cGMP-Signalweg zu ihrer Entstehung beiträgt. Möglich ist allerdings ein Einfluß von NO auf nachfolgende Prozesse wie z.B. Synaptogenese und synaptische Reifung. Abb. 10: NO-Synthase in der embryo- nalen Bauchganglienkette. A: NADPHDiaphorase-Färbung bei 90%. B: CitrullinImmunfluoreszenzfärbung Schematische Verteilung bei der 75%. C: Citrullin- immunreaktiven Zellkörper bei 75% (aus 43 analysierten Präparaten), schwarz: in mehr als 66% aller Präparate, grau: in 33%-66% aller Präparate, weiß: in weniger als 33% aller Präparate. (verändert aus Stern et al., 2010) Auch in der ventralen Ganglienkette erscheint die NO-induzierte cGMP-Immunreaktivität früher (bei 45%, Ball und Truman, 1998) als die NADPH-Diaphorase-Färbung. CitrullinImmunreaktivität setzt erst bei ca. 75% ein, und ist auch dann sehr variabel (Abb. 10). Wird 4 Ergebnisse 26 allerdings ein Nerv bei der Präparation verletzt, sieht man auch schon früher (bei 60%), und dann auch viel regelmäßiger eine Citrullin-Färbung (Stern et al., 2010). Dies lässt sich dadurch erklären, dass bei einer Axotomie an der verletzten Stelle Kalzium einströmt und die NOS zur NO-Synthese stimuliert. Es ist also zu erwarten, dass bei Schädigungen NO freigesetzt wird, welches dann auf benachbarte Neurone wirken kann. Aus diesem Grunde wurde die weitere Untersuchung eines möglichen Einflusses von NO auf axonales Wachstum im ZNS mit Hilfe eines Regenerationsparadigmas untersucht (Stern und Bicker, 2008a). Zunächst musste gezeigt werden, dass das Zentralnervensystem der Heuschrecke prinzipiell zur Regeneration befähigt ist. Bis vor kurzem hielt sich die Auffassung, dass dies nicht der Fall sei, basierend auf einem Artikel von Edwards (1969), die von späteren Autoren so übernommen wurde. Um dies zu klären, wurde bei Heuschreckennymphen eines der paarigen Halskonnektive gequetscht, was zur Axotomie der darin befindlichen Nervenbahnen führt (Pätschke et al., 2004). Das andere Halskonnektiv wurde unbeeinträchtigt gelassen und diente als interne Kontrolle. Zu verschiedenen Zeitpunkten nach der Operation wurden die Nervensysteme der Versuchstiere präpariert und mit einem Antiserum gegen das Neuropeptid Proctolin gefärbt. Dieses kommt in den Halskonnektiven nur in zwei einzelnen Axonen vor die aus den Thorakalganglien Richtung Unterschlundganglien und Gehirn ziehen (Keshishian und O'Shea, 1985). Diese proctolinergen Axone sind hervorragend geeignet, um die Reaktion auf Axotomie am Beispiel identifizierter Zellen zu untersuchen, die von Tier zu Tier wiedergefunden werden können. Es zeigte sich, dass innerhalb von sechs Tagen das distale, vom Zellkörper abgetrennte Segment der axotomierten Neuronen im Unterschlundganglion vollständig degeneriert war (Abb. 11). Nach neun Tagen waren im Unterschlundganglion erste von posterior wieder eingewachsene Proctolin-immunreaktive Fortsätze zu sehen und nach 21 Tagen war die typische Verzweigungsform mit vier dichten Büscheln Proctolinimmunreaktiver Fortsätze wiederhergestellt. Damit ist gezeigt, dass Neuronen des Heuschrecken-ZNS sehr wohl zur Regeneration fähig sind. 4 Ergebnisse 27 Abb. 11: proctolinerger Regeneration Neurone Heuschreckennymphen in nach Quetschen des Halskonnektivs (verändert aus Pätschke et al., 2004). Oben: Darstellung des Schematische Heuschrecken- ZNS, Pfeilspitze zeigt auf die Quetschstelle. A-D: Fotografien, EH: Zeichnungen immunzyto- chemisch gefärbter proctolinerger Fasern des Unterschlundganglions zu verschiedenen Zeitpunkten nach der Quetschung. Um jetzt an diesem Tier die Mechanismen der Regeneration untersuchen zu können, und insbesondere zu testen ob das NO/cGMP-system die Regeneration beeinflussen kann, stößt man bei den adulten oder juvenilen postembryonalen Insekten schnell an Grenzen. Es ist für solche Untersuchungen zwingend notwendig, über die Dauer des Experiments Pharmaka zu applizieren. Dies wird vor allem in Hinblick auf die lange Dauer des Regenerationsprozesses schwierig. Alternativ lässt sich die Regeneration an einem Embryo-Kultursystem untersuchen (Stern und Bicker, 2008a). Der späte Embryo (ab 65% Entwicklung) hat schon ein weitgehend vollständiges Nervensystem und enthält funktionelle Synapsen. Sein Nervensystem bietet aber, da es sich noch im Aufbau befindet, ein wachstumsfreundlicheres Milieu als das postembryonale. Außerdem sind die von regenerierenden Axonen zu bewältigenden Strecken im kleinen Embryo viel kürzer als in der erwachsenen Heuschrecke, so dass auch aus diesem Grund kürzere Regenerationszeiten zu erwarten sind. Für eine begrenzte Zeit ist es möglich, Embryonen in Kultur zu halten und mit geeigneten Maßnahmen sicherzustellen, dass Pharmaka, die ins Kulturmedium gegeben werden, an den zu untersuchenden Wirkort 4 Ergebnisse gelangen. Dieser 28 Kulturansatz sollte auch für die axonale Regeneration im Heuschreckenembryo angewandt werden. Abb. 12: Stickstoffmonoxid fördert die Regeneration serotonerger Axone im Heuschreckenembryo. A: Filet-Präparat eines zu 65% entwickelten Heuschreckenembryos in Kultur. Das Zentralnervensystem ist freigelegt. B: Eines der beiden Konnektive zwischen zwei Abdominalganglien gequetscht wird an zwei (Pfeilspitzen). Stellen Danach wird das Präparat für 48 h bei 30°C kultiviert. C: Fluoreszenzfärbung der vier in jedem Ganglion vorhandenen intersegmentalen serotonergen Interneuronen, 48 h nach dem Quetschen. D: Unter Kontrollbedingungen regenerieren nach 48 h ca. 40% der gequetschten Axone, in Gegenwart des NO-Donors NOC-18 (500 µM) mehr als 60%. E: Abfangen von intern produziertem NO mit dem Scavenger cPTIO reduziert die Regeneration, dies lässt sich im Rescue-Versuch mit membranpermeablem cGMP (8Br-cGMP) teilweise aufheben. F: Inhibition der NO-abhängigen sGC mit ODQ (200µM) reduziert Regeneration, dies wird im Rescue mit 8BrcGMP wieder aufgehoben. Daten in D-F sind Mittelwerte aus mindestens 10 Embryonen. G: NADPHDiaphorase-Färbung zeigt, dass zu diesem Zeitpunkt NO produzierende Neurone vorhanden sind. H: Konfokale Aufnahme einer Doppel-Färbung gegen Serotonin und cGMP nach Stimulation mit einem NO-Donor. Viele Neurone antworten auf NO mit cGMP-Produktion, die serotonergen sind auch darunter. (aus. Bicker und Stern, 2009) Zu diesem Zweck musste zunächst ein Embryo-Kultursystem entwickelt werden, das einen freien Zugang für Pharmaka gewährleistet, ohne Wachstums- und Regenerationsvorgänge zu sehr zu beeinträchtigen. Dazu wird der Embryo dorsal geöffnet, der Darm entnommen und das Nervensystem freigelegt, indem mit zwei feinen Pinzetten das ventrale Velum entfernt wird, welches das Nervensystem von der Körperhöhle trennt. Danach wird der Embryo auf einen Tropfen unpolymerisiertem Sylgard gegeben und mit Zellkulturmedium überschichtet. Aufgrund der in diesem Stadium schon vorhandenen hydrophoben Cuticula breitet sich der 4 Ergebnisse 29 Embryo zwischen dem hydrophoben Sylgard und dem Zellkulturmedium aus (Abb. 12A). Dieses Präparat kann über mehrere Tage in Kultur gehalten werden. Quetscht man nun in solchen Präparaten einen der paarigen Nerven, die die Ganglien des Bauchmarks verbinden, werden die darin verlaufenden Axone zerstört (Abb. 12B). Mit immunzytochemischen Methoden kann man die dadurch bedingte Degeneration und spätere Regeneration einzelner Axone verfolgen und quantifizieren. Dazu wurden exemplarisch die serotonergen Axone herausgegriffen, die aus den bei Heuschrecken (Taghert und Goodman, 1984; Condron, 1999) und Drosophila (Lundell und Hirsh, 1998) eingehend untersuchten Interneuronen S1 und S2 entspringen. Durch jedes abdominale Konnektiv laufen in diesem Entwicklungsstadium genau vier serotonerge Axone, die sich einfach mit einem Antikörper gegen Serotonin anfärben lassen (Abb. 12C). Innerhalb von zwei Tagen regenerieren ca. 40% der Axone über die Quetschstelle hinaus, und nach 4 Tagen in Kultur haben einzelne Axone das Neuropil des nächsten Ganglions erreicht. Appliziert man nun während der Kultur Substanzen, die NO freisetzen, erhöht sich dieser Prozentsatz signifikant (Abb. 12D). Nicht nur extern zugefügtes Stickstoffmonoxid, sondern auch intern produziertes kann Regeneration fördern: Fängt man nämlich intern produziertes NO mit einem NO-Scavenger ab, ist die Regeneration reduziert (Abb. 12E). Wie oben erwähnt, kommen als interne NO-Quelle neben Hämozyten vor allem NOS exprimierende Neurone in Frage, deren Axone durch den Quetschvorgang geschädigt werden, was zum Kalziumeinstrom und zur Kalziumabhängigen NO-Produktion führt. Die NO-Produktion in gequetschten Axonen konnte in der vorliegenden Arbeit mit Hilfe der CitrullinImmunomarkierung nachgewiesen werden (Stern et al., 2010). Wie im peripheren Nervensystem erfolgt auch im ZNS die Wirkung von NO zumindest teilweise über cGMP als second messenger. Wird die als NO-Rezeptor fungierende sGC inhibiert, ist die Regeneration reduziert, lässt sich aber im Rescue-Versuch durch Zugabe von 8Br-cGMP teilweise wiederherstellen (Abb. 12F). Die serotonergen Interneurone selbst, die hier als Beispiel betrachtet wurden, wie auch eine Vielzahl anderer Zellen vermag auf NOStimulation mit erhöhter cGMP-Produktion zu antworten, wie sich immunzytochemisch nachweisen lässt (Abb. 12H), so dass man von einem allgemeinen Effekt ausgehen kann. 4 Ergebnisse 30 4.4 Neurotransmitter und synaptische Modulation durch Stickstoffmonoxid in menschlichen Modellneuronen (Veröffentlichungen 7 und 8) In diesem Teil der Arbeit soll ein einfaches Modellsystem mit menschlichen Neuronen in Kultur vorgestellt werden, an dem die im Vorausgehenden an Evertebraten-Nervensystemen verwendeten Methoden und Fragestellungen auf den Menschen übertragen werden sollen. Bevor die mit der in unserem Labor entwickelten Methode (Abb. 3) quasi im Schnellverfahren generierten Zellen nun als in vitro-Modell für das menschliche Gehirn eingesetzt werden können, muss gezeigt werden, dass sie normalen menschlichen Neuronen entsprechen und sich auch nicht wesentlich von den im früher üblichen langsamen Verfahren generierten NT2-Neuronen unterscheiden. Darum wurden die nach der neuen Methode gewonnen NT2-Neuronen im Verlaufe der vorliegenden Arbeit umfassend charakterisiert (Podrygajlo et al., 2009a). Insbesondere wurde untersucht, ob alle entstehenden Zellen neuronale Charakteristika aufweisen, ob Anzeichen für synaptische Kontakte feststellbar sind und welche neuronalen Botenstoffe die Neuronen verwenden. Als neuronale Marker dienten die für Neuronen typische Mikrotubuli-Untereinheit ß-Tubulin III (ßTubIII), das vor allem in Dendriten vorkommende Mikrotubuli-assoziierte Protein 2 (Map2), das in phosphorylierter Form vor allem in Axonen vorkommende Phospho-Tau-1 (Tau) und das in präsynaptischen Endigungen lokalisierte Synapsin. Als Marker für undifferenzierte Zellen diente das Intermediärfilament Nestin (Tab. 2). Tage nach Neuronengewinnung Nestin -14 (RA) -10 (Inhib.) 0 7 14 28 ++ ++ 0 0 0 0 ßTubIII 0 + ++ ++ ++ ++ Map2 0 + ++ ++ ++ ++ Tau - - - 0 ++ ++ Synapsin - - - 0 ++ ++ Tab. 2: Zeitliche Entwicklung einiger typischer Marker-Proteine während der NT2-Differenzierung und Neuronenkultur. (++: in mehr als 90% der Zellen, +: in 10-20% der Zellen, 0: in weniger als 10% der Zellen, -: in keiner Zelle, RA: während der adhärenten Kultur in Gegenwart von Retinsäure, Inhib.: Zu Beginn der Behandlung mit Mitoseinhibitoren) 4 Ergebnisse 31 Es ergab sich, dass die ersten ßTubIII- und Map2-positiven Neuronen während der Differenzierungsphase in Retinsäure-Medium auftraten. Am Ende der Differenzierungsphase enthalten die Kulturen 10-20% ßTubIII- und Map2-positive Zellen. Die gewonnenen Neuronen enthalten nach dem erneuten Aussäen weniger als 5% Nestin-positive, undifferenzierte Zellen, die sich aber nicht mehr teilen. Nach dem Aussäen beginnen die Neuronen, Fortsätze auszubilden, in denen sich nach ca. 2 Wochen axonale und synaptische Marker nachweisen lassen (Tab. 2). Glia-Typische Marker, wie z.B. glial fibrillary acidic protein (GFAP) wurden zu keinem Zeitpunkt gefunden, so dass davon auszugehen ist, dass hier eine reine Neuronen-Kultur vorliegt. Im Folgenden wurden Neurotransmitter bzw. Neurotransmitter-spezifische Marker bei ausgewachsenen Kulturen (28 Tage in Kultur) untersucht (Abb. 13, Tab. 3). Die Neurotransmitter Serotonin, Glutamat, GABA und Dopamin lassen sich direkt mit gegen sie gerichteten Antikörpern markieren. Das ist beim Acetylcholin nicht möglich, da die Verbindung instabil ist und sich nicht fixieren lässt. Statt dessen kamen Antiseren gegen das Acetylcholin synthetisierende Enzym Cholin-Acetyl-Transferase (ChAT) und gegen den vesikulären Acetylcholin-Transporter (VAChT) zum Einsatz. Auch die Befunde bei Glutamat, GABA und Dopamin wurden abgesichert durch Immunfärbungen gegen einen vesikulären Glutamat-Transporter (VGluT1), das GABA synthetisierende Enzym Glutamat- Decarboxylase (GAD) und das Dopamin synthetisierende Enzym Tyrosin-Hydroxylase (TH). In jedem Fall ergab sich eine Bestätigung des Befundes. Abb. 13: Neurotransmitter Immunfluoreszenzfärbungen in NT2-Neuronen. gegen Neurotransmitter bzw. Neurotransmitter-spezifische Markerproteine an 2 Wochen alten NT2-Neuronenkulturen. ChAT: CholinAcetyl-Transferase, GAD: Glutamat-Decarboxylase, 5HT: Serotonin, vGluT1: vesikulärer Glutamat-Transporter 1, Maßstab 50 µm (100 µm für Glutamat und GABA). (verändert aus Podrygajlo et al., 2009a) 4 Ergebnisse 32 Der hohe Anteil von cholinergen NT2-Neuronen spricht dafür, dass diese Kultur eher Ähnlichkeiten mit spinalen als mit Hirn-Neuronen hat. Im Rückenmark kommt eine größere Anzahl cholinerger (Moto-)Neurone vor. In einem Versuch, den ohnehin schon hohen Prozentsatz cholinerger Neuronen noch weiter zu steigern, wurde pharmakologisch in das BMP/sonic hedgehoc-System eingegriffen, das maßgeblich an der Motoneuron- Differenzierung im Rückenmark beteiligt ist. Allerdings veränderten weder die Zugabe von sonic hedgehoc-Protein oder dem sonic hedgehoc-Rezeptor aktivierenden Faktor Purmorphamin, noch die Inhibition des BMP-Signalweges mit dem Faktor noggin den Prozentsatz cholinerger Neuronen. Möglicherweise reicht allein die hohe RetinsäureKonzentration, die zur Differenzierung der NT2-Neuronen eingesetzt wurde aus, um einen größtmöglichen Anteil cholinerger Neuronen zu erzeugen. Retinsäure ist dafür bekannt, als caudalisierendes Signal zu wirken (Maden, 2002), so dass es nicht verwunderlich ist, einen für Rückenmarks-Neuronenpopulationen typischen cholinergen Anteil zu erhalten. Transmitter Prozentsatz Neuronen Acetylcholin 37% Glutamat 40% GABA 15% Serotonin 2% Dopamin 0% Tab. 3: Neurotransmittersyteme in NT2-Neuronen, 28 Tage in Kultur. Dargestellt ist der Prozentsatz der Neuronen (bezogen auf DAPI-gefärbte Zellkerne) die mindestens einen der getesteten Marker für das entsprechende Transmittersystem aufweisen. Für eine caudale Identität der NT2-Neurone spricht auch der sehr geringe Anteil serotonerger, und die völlige Abwesenheit dopaminerger Neuronen, die beim Menschen nur im Gehirn vorkommen. In konventionell erzeugten NT2-Neuronen konnte ein dopaminerger Phänotyp durch Zugabe von Lithium (Zigova et al., 1999, 2000) oder die Applikation einer Mixtur von FGF1 und verschiedenen Co-Aktivatoren (Iacovitti et al., 2001) erreicht werden. Diese Versuche führten bei unseren NT2-Neuronen in keinem Fall zu dopaminergen Neuronen (Podrygajlo et al., 2009a). Zusammenfassend lässt sich sagen, dass die mit dem optimierten Verfahren generierten NT2-Neuronen bezüglich ihrer Neurotransmitter eine inhomogene Population darstellen, wie es ja auch bei den Neuronen des menschlichen ZNS der Fall ist. 4 Ergebnisse 33 Als nächstes stellt sich die Frage, ob es in der NT2-Neuronenkultur funktionelle Verbindungen gibt. Ein Hinweis darauf liefert die immunzytochemische Lokalisierung der synaptischen Proteine Synapsin und Synaptotagmin. Synapsin ist ein lösliches Protein, das Neurotransmittervesikel an das Zytoskelett bindet. Synaptotagmin ist ein in die synaptische Vesikelmembran integriertes Protein, das als Kalziumsensor bei der aktivitätsabhängigen Exozytose dient. Beide Proteine lassen sich in NT2-Neuronen nachweisen, und zwar zeitabhängig (Tegenge et al., 2009). Nach 7 Tagen in Kultur ist die Färbung noch schwach, aber nach 14 Tagen ist sie deutlich und erscheint punktiert. Das Vorhandensein von Neurotransmitter-spezifischen Markern und präsynaptischen Proteinen allein reicht nicht aus, um die Existenz funktioneller Synapsen nachzuweisen. Im Verlauf der vorliegenden Arbeit wurde diese Fragestellung mit zwei unterschiedlichen Methoden angegangen, der aktivitätsabhängigen Markierung mit Antikörpern gegen die luminale SynaptotagminDomäne (lu-Syt) und der FM 1-43-Imaging-Technik, bei der ein Fluoreszenzfarbstoff, der in geöffnete Vesikel gelangt, sich vorübergehend an die luminale Vesikelmembran anlagert und somit recycelte synaptische Vesikel markiert. (Tegenge et al., 2009). Abb. 14: Markierung synaptischer Aktivität mit Antikörpern gegen Synaptotagmin. A: Antikörper (grün) gegen die luminale Domäne des Vesikel-Proteins Synaptotagmin (rot) befinden sich in der Ringer-Lösung. B: Bei Depolarisation öffnen sich synaptische Vesikel, Antikörper können eindringen und an Synaptotagmin binden. C: Nach dem Waschen und Fixieren sind die beteiligten synaptischen Vesikel markiert. D: Fluoreszenzaufnahme wie in C, Pfeilspitzen zeigen auf synaptische Punkte, rot: lu-Syt, blau: DAPI (Zellkerne). Es zeigte sich, dass in entsprechend alten Kulturen (2 Wochen und älter) bei Depolarisation durch eine Kalium-reiche Lösung die Aufnahme der lu-Syt-Antikörper erfolgte (Abb. 14), nicht aber ohne Depolarisation oder in Abwesenheit von Kalzium. Dies ist ein Indiz für Depolarisations- und Kalzium-abhängige Exozytose und Vesikel-Recycling. Zu demselben Ergebnis führte FM 1-43 Imaging: Bei Depolarisation in Kalziumhaltiger Lösung wurde FM 1-43 aufgenommen. Bei erneuter Depolarisation in Gegenwart von Kalzium erfolgte eine Entfärbung (Abb. 15; Tegenge et al., 2009). 4 Ergebnisse 34 Abb. 15: Synapsen-Imaging mit FM 1-43. A: FM 1-43 (grün) befindet sich in der Ringer-Lösung. B: Bei Depolarisation öffnen sich synaptische Vesikel und der Farbstoff kann eindringen. C: Nach dem Waschen sind die Vesikel fluoreszenzmarkiert. D: Bei erneuter Depolarisation entweicht der Farbstoff aus den geöffneten Vesikeln. E: Die Synapse ist wieder entfärbt F: NT2-Neuriten nach der FM 1-43-Aufnahme (wie in C) G: Selber Ausschnitt wie in F, nach dem depolarisations- und Kalziumabhängigen Entfärben (wie in E). Pfeilspitzen weisen auf synaptische Punkte. Doppelfärbungs-Experimente, in denen ein fixierbares Analogon des FM 1-43 verwendet wurde, konnten belegen, dass die FM 1-43-Aufnahme-Orte mit Synaptotagmin kolokalisiert waren, woraus zu schließen ist, dass es sich hierbei tatsächlich um präsynaptische Regionen handelt. Ob diesen präsynaptischen Freisetzungsorten auch die entsprechenden postsynaptischen Strukturen gegenüberliegen, konnte im Verlauf der vorliegenden Arbeit noch nicht geklärt werden. Geeignete Marker-Proteine für postsynaptische Strukturen, z.B. PSD95, konnten in NT2-Neuronen nicht nachgewiesen werden. In früheren Arbeiten an konventionell präparierten NT2-Neuronen war elektrophysiologische und elektronenmikroskopische Evidenz für funktionelle Synapsen nur bei Ko-Kultur mit Gliazellen zu erreichen (Hartley et al., 1999). Es gibt aber inzwischen in den nach unserer verbesserten Methode hergestellten NT2-Neuronen Evidenz für funktionelle Neurotransmitter-Rezeptoren, wie z.B. Glutamat-Rezeptoren vom NMDA-Typ (PaquetDurand und Bicker, 2004) und vom AMPA-Typ sowie GABA-Rezeptoren (Podrygajlo et al., 2009b). Außerdem konnten in der letztgenannten Arbeit auch Miniatur-postsynaptische Ströme (bei denen es sich um postsynaptische Antworten auf spontane Neurotransmitterfreisetzung handelt) sowohl für GABA als auch für Glutamat nachgewiesen werden – eine direkte Evidenz für funktionelle Synapsen. Im Weiteren wurde nun untersucht ob die synaptische Exozytose durch Neuromodulatoren, insbesondere Stickstoffmonoxid, beeinflusst wird. Dies lag nahe, denn ca. die Hälfte aller NT2-Neuronen reagiert in Gegenwart eines NO-Donors mit der Produktion von zyklischem GMP. Selbst ohne Zugabe von externem NO ist immer ein kleiner Teil der Neurone cGMPimmunreaktiv, was auf die Aktivität einer in der Kultur vorkommenden NOS schließen lässt. 4 Ergebnisse 35 In der Tat konnte die neuronale NOS in NT2-Neuronen sowohl im Western Blot als auch mit Hilfe von Immunfluoreszenzfärbungen nachgewiesen werden (Tegenge et al., 2009). Um einen möglichen Einfluss von NO/cGMP auf die synaptische Vesikelfusion untersuchen zu können, wurden 28 Tage alte NT2-Neuronen in Gegenwart von Pharmaka, die in den Signalweg eingreifen (Abb. 4.) mit Antikörpern gegen die luminale Domäne des Synaptotagmins inkubiert (wie in Abb. 14). NO bewirkte eine Aufnahme der Antikörper, in ähnlicher Weise wie bei Depolarisation mit Hilfe hoher extrazellulärer Abb. 16: NO/cGMP-Signalweg bei der synaptischen Vesikelfusion (verändert aus Tegenge et al., 2009). A: Markierung mit anti-lu-Syt erfolgt unter depolarisierenden Bedingungen (high KCl) oder in Gegenwart eines NO-Donors (SNP= 1mM Natriumnitroprussid), nicht aber in Kontrolllösung (KRH = Krebs-Ringer mit HEPES) oder in Gegenwart des NO-Donors und 50 µM des cGC-Inhibitors ODQ. B: Entfärbung mit FM 1-43 markierter synaptische Punkte in Gegenwart der NO-Donoren SNP (1mM) und NOC-12 (100µM) und unter depolarisierenden Bedingungen (high KCl). C: Nach 5 min verbleibende Fluoreszenzintensität synaptischer Punkte nach Behandlung mit Kontrolllösung (KRH), Depolarisation (high KCl), 1mM SNP, und 1mM SNP + 200 µM ODQ. Alle Daten sind Mittelwerte ±S.E.M. von mindesten 15 synaptischen Punkten. Kaliumkonzentrationen. Dieser Effekt ließ sich durch Zugabe des sGC-Inhibitors ODQ blockieren (Abb. 16A). Mit FM 1-43 markierte synaptische Vesikel entfärbten sich in Gegenwart von NO-Donoren mit einem Zeitverlauf, der dem bei Depolarisation durch hohe Kaliumkonzentration sehr ähnlich ist (Abb. 16B). Auch dieser Effekt von NO ließ sich durch Inhibition der löslichen Guanylyl-Cyclase mit ODQ hemmen (Abb. 16C). In weiteren Experimenten konnte gezeigt werden, dass die synaptischen Vesikel auch durch direkte Zugabe membranpermeabler cGMP-Analoga zur Fusion gebracht werden können. 4 Ergebnisse 36 4.5 Neurotransmitter an der neuromuskulären Synapse von Onychophoren (Veröffentlichung 4) Die in diesem Kapitel dargestellten Befunde beziehen sich nicht in erster Linie auf Synapsen und ihre Botenstoffe unter dem entwicklungsneurobiologischen Gesichtspunkt, sondern unter einem phylogenetischen. Verwandtschaftsverhältnisse zwischen Tiergruppen wurden bis in die jüngste Gegenwart vor allem anhand anatomischer und funktionsmorphologischer Kriterien ermittelt (z.B. Henning, 1980). Erst in neuester Zeit stand in Form der molekularbiologischen Stammbaumanalyse ein Werkzeug zur Überprüfung der bei manchen Gruppen auch anhand klassischer Kriterien kontrovers diskutierten Verwandtschaftsverhältnisse zur Verfügung. Diese führten zu einigen bedeutenden Umgruppierungen im Stammbaum der Tiere (Aguinaldo et al., 1997; Giribet et al., 2001). Dieses neue Konzept ist noch immer nicht überall akzeptiert und es besteht dringender Bedarf an zusätzlichen unabhängigen Daten, die weder morphologischer Natur sind, noch Gensequenzen darstellen. Schon in einer früheren Arbeit konnten wir zeigen, dass die Analyse der chemischen Architektur des Nervensystems hier einen hilfreichen Beitrag leisten kann (Haase et al., 2001). In der vorliegenden Arbeit wurde die neuromuskuläre Synapse der Onychophoren untersucht, einer urtümlichen Tiergruppe, die in der südlichen Hemisphäre mit einigen Dutzend Arten verbreitet ist (Abb. 17). Nach morphologischen Kriterien stellen sie Abb. 17: Peripatoides novaezealandiae, ein Onychophor aus Neuseeland (verändert aus Stern und Bicker, 2008b) ein Bindeglied zwischen den Arthropoden und den Anneliden dar, allerdings mit überwiegend Anneliden-ähnlichen Eigenschaften. Nach DNA-Sequenzanalysen gehören sie in die neu aufgestellte Gruppe der Ecdysozoa, zu denen die Arthropoden, nicht aber die Anneliden gehören (Aguinaldo et al., 1997). Diese Einteilung wird durch unseren Befund gestützt, dass Onychophoren-Neuronen auf ihrer Oberfläche ein Zucker-Epitop exprimieren, das für die Ecdysozoa typisch ist (Haase et al., 2001). Bezüglich der neuromuskulären Synapse gleichen sich alle daraufhin untersuchten Ecdysozoa durch die Verwendung von Glutamat als Transmitter, während viele andere Tiergruppen, unter anderem Wirbeltiere, und auch die Anneliden, cholinerge neuromuskuläre Synapsen aufweisen. Über die Neuromuskuläre Synapse der Onychophoren gibt es einige wenige Arbeiten aus den 1950er und 1960er Jahren 4 Ergebnisse 37 (Ewer und van den Berg, 1954; Florey und Florey, 1965), die auf eine cholinerge Transmission schließen lassen – ein Gegensatz zu den neueren Daten. Mit den heutigen verbesserten Methoden der Immunfluoreszenzfärbung und mit elektrophysiologischen Methoden wurde diese Diskrepanz in der vorliegenden Arbeit einer kritischen Prüfung unterzogen (Stern und Bicker, 2008b). Die neuromuskulären Synapsen lassen sich gut mit Antikörpern gegen Synapsin darstellen (Abb. 18A, C). Es zeigen sich zumindest zwei in ihrer Ausdehnung und Gestalt sehr unterschiedliche Typen von Synapsen. Der eine Typ lässt sich auch mit der Acetylcholinesterase-Färbung (Karnowsky und Roots, 1964) darstellen, einem zytochemischen Marker für cholinerge Synapsen (Abb. 18B). Der andere Typ lässt sich mit einem Antikörper gegen Glutamat färben (Abb. 18D). Abb. 18: Acetylcholin und Glutamat an der neuromuskulären Synapse von P. novaezealandiae. A, C: Synapsin-Immunfluoreszenzfärbungen an zwei unterschiedlichen Typen von neuromuskulären Synapsen. B: Acetylcholinesterase-Färbung an einer Synapse wie in A. D: Glutamat-Immunfluoreszenzfärbung an einer Synapse wie in B. (verändert aus Stern und Bicker, 2008b) Es gibt also prinzipiell histochemische und immunzytologische Evidenz für beide Transmitter. Durch intrazelluläre Ableitungen von Muskelpotentialen konnte dieser Befund gestützt werden. Bei Applikation kurzer Pulse des entsprechenden Neurotransmitters auf den Muskel, verursachte in den Longitudinalmuskeln zwar nur Acetylcholin, nicht aber Glutamat eine Depolarisation (Abb. 19A), in anderen Muskeln bewirkte Glutamat allerdings sehr wohl eine deutliche Exzitation (Abb. 19B, C). Dass diese Tatsache früher übersehen wurde, ist nicht überraschend, denn gerade die großen Longitudinalmuskeln, die für die damaligen Untersuchung am einfachsten zur Verfügung standen zeigen, als Ausnahme, diese doppelte cholinerg/glutamaterge Innervierung nicht. 4 Ergebnisse 38 Abb. 19: Elektrische Antworten verschiedener Muskeln auf Druck-appliziertes Acetylcholin und Glutamat. A: Longitudinalmuskeln antworten mit Depolarisationen auf einen 50 ms Puls von 100 µM Acetylcholin (Ach), nicht aber auf einen gleichlangen Puls von 100 µM Glutamat (Glut). B: Diagonalmuskeln antworten stark auf Acetylcholin und schwach auf Glutamat. C: Ringmuskeln antworten gleichstark auf beide Neurotransmitter. Maßstäbe: vertikal 20 mV, horizontal 2 s. (verändert aus Stern und Bicker, 2008b) Hier ist jedenfalls Evidenz präsentiert worden, dass Onychophoren sowohl Glutamat als auch Acetylcholin als neuromuskulären Transmitter verwenden, und die chemische Ausstattung dieser Synapse ist nicht länger ein Gegenargument gegen die moderne phylogenetische Einteilung der Onychophoren. Möglicherweise ist der neuromuskuläre Transmitter generell nicht das am besten geeignete phylogenetische Kriterium. Cholinerge neuromuskuläre Transmission ist im Tierreich weit verbreitet, aber es ist nicht klar, ob dies einen basalen Phänotyp oder das Ergebnis konvergenter Evolution darstellt. Sowohl Deuterostomier wie Wirbeltiere oder Echinodermaten (Devlin, 2001) als auch nicht-ecdysozoische Protostomier verwenden am Muskel Acetylcholin, wie z.B. Anneliden (Wallace, 1981) und Mollusken (Keating und Lloyd, 1999), aber auch die nach Aguinaldo et al. (1997) zu den Ecdysozoa gehörenden Nematoden (Duerr et al., 2008). In Mollusken gibt es sogar, wie hier für die Onychophoren gezeigt, sowohl cholinerge als auch glutamaterge Motoneuronen (Keating und Lloyd, 1999). Neuere Untersuchungen haben gezeigt, dass sogar bei Vertebraten der neuromuskuläre Transmitter keineswegs starr festgelegt ist. Embryonale Wirbeltiermuskeln exprimieren neben Acetylcholinrezeptoren auch solche für Glutamat, GABA und Glycin, und die normalerweise ausschließlich cholinerge Transmission kann durch Manipulation der neuronalen Erregbarkeit im embryonalen Rückenmark in eine glutamaterge Richtung verschoben werden (Borodinsky und Spitzer, 2007). 5 Studienübergreifende Diskussion 39 5 Studienübergreifende Diskussion 5.1 Neurotransmitter bei neuronaler Plastizität und Entwicklung In den vorgestellten Arbeiten wurden einige niedermolekulare neuronale Botenstoffe, insbesondere Octopamin, Serotonin und Stickstoffmonoxid, unter dem Aspekt ihres Einflusses auf die neuronale Plastizität betrachtet. 5.1.1 Octopamin Beim Octopamin wurde am Beispiel der Dishabituation im visuellen System kurzfristige neuronale Plastizität untersucht. Solche kurzfristigen Effekte sind gerade für das Octopamin und verwandte biogene Amine in diversen sensorischen Systemen bei Insekten nachgewiesen worden (reviews: Roeder, 1999, 2005), im visuellen System allerdings nur am Beispiel eines einzelnen gut beschriebenen Neurons, dem DCMD der Heuschrecke (Bacon et al., 1995; Stern, 1999). Durch die vorliegende Arbeit (Stern, 2009) wurde die naheliegende Vermutung bestätigt, dass sich jedoch die octopaminerge Modulation des Bewegungsdetektorsystems nicht auf diese einzelne Zelle beschränkt, sondern dass es sich um ein generelles Phänomen handelt, und dass dasselbe octopaminerge Neuron, PM4, eine Vielzahl von visuellen Interneuronen beeinflussen kann. Ähnliche, vermutlich homologe octopaminerge Neuronen sind inzwischen auch schon in verschiedenen anderen Insekten gefunden worden (Sinakevitch et al., 2005, Sinakevitch und Strausfeld, 2006), so dass man davon ausgehen kann, dass es sich hier um einen evolutionär konservierten Mechanismus handelt. Bei Wirbeltieren wird Dishabituation und Aufmerksamkeit in ähnlicher Weise von multimodalen Neuronen aus dem locus coeruleus bewirkt, vermittelt durch das bei Wirbeltieren zum Octopamin homologen Catecholamin Noradrenalin (Aston-Jones et al., 1999). Bei dem wirbellosen Tier, das im Hinblick auf synaptische Plastizität wohl am besten untersucht ist, der Meeresschnecke Aplysia, wird sowohl Dishabituation als auch assoziatives Lernen durch ein anderes biogenes Amin, nämlich Serotonin, vermittelt (Hawkins et al., 2006). Bei Insekten wird assoziatives Lernen durch Octopamin vermittelt (Menzel et al., 1999; Schwaerzel et al., 2003), demselben Modulator, der wie in der vorliegenden Arbeit gezeigt, Dishabituation bewirkt. Auch hier ist ein einzelnes identifiziertes octopaminerges Neuron maßgeblich beteiligt, dessen Aktivität die dem Lernvorgang zugrundeliegende synaptische Modulation bewirkt (Hammer, 1993). Allerdings kann man bei Insekten zwischen 5 Studienübergreifende Diskussion 40 appetetivem und aversivem Lernen unterscheiden. Bei Drosophila wird nur appetetives Lernen über octopaminerge Modulation vermittelt, beim aversiven Lernen übernimmt Dopamin diese Rolle (Schwaerzel et al., 2003). Ob Dopamin auch eine Rolle bei der Dishabituation spielen kann, ist bisher nicht untersucht. Die neuroanatomische Grundlage dafür ist allerdings auch im visuellen System der Heuschrecke gegeben, wo extensive Dopamin-immunreaktive Verzweigungen im optischen Lobus vorhanden sind (Wendt und Homberg, 1992), die sich mit den octopaminergen Verzweigungen überlappen. Mit dem in der vorliegenden Arbeit charakterisierten PM1-Neuron gibt es jetzt ein für intrazelluläre Untersuchungen leicht zugängliches Testsystem für die Funktionsanalyse des visuellen Systems an dem diese und weitere Fragestellungen untersucht werden können. 5.1.2 Serotonin im enterischen Nervensystem Serotonin ist in mancher Hinsicht unter den biogenen Aminen der Gegenspieler zum Octopamin. Während Octopamin die Antworteigenschaften sowohl motorischer als auch sensorischer Systeme verstärkt und die Mobilisierung von Stoffwechselreserven fördert (review: Roeder, 1999), induziert Serotonin Verdauungsprozesse und reduziert oft die Stärke sensorischer Eingänge. Als Beispiel sei nur das luftströmungs-sensitive System der Schabe genannt, bei denen die von sensorischen Afferenzen ausgelösten postsynaptischen Potentiale in den giant interneurons durch Octopamin verstärkt und durch Serotonin abgeschwächt werden (Hill und Blagburn, 2001). Serotonin spielt sowohl bei Wirbeltieren als auch bei Wirbellosen eine wichtige Rolle für die Darmmotilität, weshalb Störungen im serotonergen System in beiden Tiergruppen Probleme bei der Darmperistaltik und der Verdauung verursachen (Gershon, 2004; Trumm und Dorn, 2000). Im Darm von Insekten beeinflusst Serotonin einerseits direkt die Darmmuskulatur (Molaei und Lange, 2003; Luffy und Dorn 1992), andererseits kann es in den enterischen Ganglien generierte motorische oder prämotorische Rhythmen beeinflussen (Luffy und Dorn, 1992). Dies ist auch beim stomatogastrischen System des Hummers gezeigt (Harris-Warrick und Marder, 1991). In der vorliegenden Arbeit wurde die Entwicklung des serotonergen Systems im Heuschrecken-ENS untersucht. Dabei erwies es sich als nützlich, dass die serotonergen Neuronen schon lange bevor sie beginnen Serotonin zu synthetisieren, den SerotoninWiederaufnahme-Transporter, SERT, exprimieren. Dieser befähigt die Zellen, Serotonin aus der Umgebung aufzunehmen, so dass es möglich war, die Migration und das Auswachsen der serotonergen Neuronen schon kurz nach ihrer Geburt zu verfolgen. Dieser Transporter ist bei Wirbeltieren sehr gut untersucht, denn auf ihm beruhen die Wirkung von Kokain und vieler 5 Studienübergreifende Diskussion 41 Medikamente, vor allem Antidepressiva (Butler und Meegan, 2008). Bei Insekten ist er vor allem in Drosophila gut charakterisiert (Barker et al., 1998), kommt aber auch im Heuschrecken-ZNS vor (Condron, 1999). Es stellte sich heraus, dass die serotonergen Neuronen schon bald nach ihrer Geburt, und noch während der Migration in ihr Zielgebiet im Frontalganglion SERT exprimierten, was darauf schließen lässt, dass ihr Schicksal zu diesem Zeitpunkt schon determiniert war. Die Determination einer kleinen Gruppe serotonerger Neuronen (S1, S2 und S3) wird im Bauchmark von Heuschrecken- und Drosophila-Embryonen schon seit Jahren intensiv untersucht (Taghert und Goodman, 1984; Condron 1999; Lundell und Hirsh, 1998; Dittrich et al., 1997). Dabei beruht die Determination eines serotonergen Phänotyps auf einer Kaskade von Transkriptionsfaktoren, an deren Anfang der Faktor engrailed steht. Die Neuronen des ENS entwickeln sich aber auf eine andere Art und Weise als im ZNS, und sie exprimieren auch kein engrailed (Stern, z. Publ. eingereicht). Die serotonergen Neuronen des ENS erhalten schon während ihrer Migration zum Zielort ihren Transmitter-Phänotyp. Deshalb können lokale Merkmale des Zielgewebes nicht allein die Determination bestimmen, anders als z.B. bei den Mitteldarm-Neuronen in Manduca, die ihren Transmitter FMRFamid erst nach Erreichen ihres Bestimmungsortes exprimieren (Copenhaver und Taghert, 1989). Möglicherweise stellt der serotonerge Phänotyp sogar den default state dar, der durch das Vorhandensein von Serotonin selbst negativ reguliert wird. Eine Autoregulation der SERTExpression konnte in der vorliegenden Arbeit in vitro nachgewiesen werden. Diese könnte in vivo durch im Embryo zirkulierendes Serotonin bewirkt werden. Für Drosophila-Embryonen ist eine hohe Serotonin-Konzentration gemessen worden (Colas et al., 1995). Da die serotonergen Neuronen Serotonin aufnehmen können, wären sie bei Vorhandensein der notwendigen präsynaptischen Maschinerie in der Lage, Serotonin in direkter Nähe zu den migrierenden enterischen Neuronen auszuschütten, obwohl sie es noch nicht selbst synthetisieren können. Dieses Phänomen eines 'geborgten Transmitters' ist auch aus dem embryonalen stomatogastrischen Ganglion von Crustaceen (Richards et al., 2003) und im Wirbeltierhirn (Lebrand et al., 1998) bekannt. Zu dieser Hypothese passt im ENS der Heuschrecke das frühe Auftreten des präsynaptischen Proteins Synapsin (unpublizierte Ergebnisse). Somit könnte eine Aufgabe der frühen Expression serotonerger Marker in den enterischen Neuronen die Regulation der Anzahl serotonerger Zellen selbst sein. Eine andere Aufgabe könnten diese Neuronen bei der Entwicklung der Nervengeflechte des Mitteldarms haben. Der Mitteldarmplexus entwickelt sich durch die geradlinige Migration von enterischen Neuronen auf vier Zugstaßen über die Darmoberfläche, die Verzweigungen 5 Studienübergreifende Diskussion 42 zur endgültigen Ausbildung des Plexus erfolgen erst sehr spät. Im Gegensatz dazu bildet sich der Vorderdarmplexus durch sofort vielfältig verzweigte Migration (Knipp und Bicker, 2009b). Auch die Innervation durch serotonerge Neurite unterscheidet sich. Auf dem Mitteldarm folgen die serotonergen Fortsätze direkt den vordersten migrierenden Neuronen, während serotonerge Neuriten auf dem Vorderdarm erst dann auswachsen, wenn der Plexus fast fertig ausgestaltet ist (Stern et al., 2007). Es ist nicht auszuschließen, dass Serotonin vorzeitige Verzweigungen auf dem Mitteldarm inhibiert, denn Serotonin ist auch aus anderen Systemen für das Unterdrücken von neuronalen Verzweigungen bekannt (Diefenbach et al., 1995). 5.1.3 Stickstoffmonoxid im Heuschrecken-ZNS Die in den letzten Jahren deutlich verbesserte Methodik der Darstellung NO produzierender Neuronen hat es ermöglicht, die embryonale Entwicklung des nitrergen Systems im Heuschreckenembryo detailliert zu untersuchen. So ist z.B. die Darstellung nitrerger Neuronen durch die Optimierung der Fixierungsmethoden durch Ott und Kollegen stark verbessert worden (Ott und Elphick, 2002, 2003). Die cGMP-Immunfärbung wurde durch die Einführung des allosterischen cGC-Aktivators YC-1 (Ott et al., 2004) um ein Vielfaches sensitiver und ermöglicht jetzt auch die Auflösung feinster Details. Mit der CitrullinImmunfärbung (Martinelli et al., 2002) hat man zum erstenmal die Möglichkeit, auch die Aktivität der NO-Synthase, und nicht nur ihre bloße Anwesenheit in hoher räumlicher Auflösung zu lokalisieren. Die zeitliche Auflösung ist mit dieser Methode natürlich gering, da die immunzytochemische Methode akkumuliertes Citrullin in Zellen nachweist, die in einem nicht definierbaren Zeitraum vor dem Zeitpunkt der Fixierung eine länger andauernde, oder möglicherweise auch eine kurze aber sehr starke NOS-Aktivität hatten. Eine höhere zeitliche Auflösung könnten eventuell Imaging-Verfahren, z.B. mit dem NO-indizierenden Farbstoff Diaminofluorescein (Kojima et al., 1998), bieten, die in Einzelfällen auch schon am sich entwickelten Insektennervensystem angewendet wurden (Champlin und Truman, 2000). Allerdings ist die Anwendung dieses Farbstoffes nicht einfach, z.B. wegen seiner Interaktion mit zellulärem Kalzium (Suzuki et al., 2001), und auch die räumliche Auflösung ist in lebendem Gewebe nicht sehr gut. Möglicherweise sind hier aber in Zukunft z.B. mit Hilfe der 2-Photonen-Laser-Scanning-Mikroskopie Verbesserungen zu erwarten. Bis dahin muss die Citrullin-Immunfärbung ausreichen, um NOS-Aktivität zellulär nachzuweisen. Mit diesen verbesserten Methoden ist es gelungen, einige bemerkenswerte Erkenntnisse über die Entwicklung des nitrergen Systems im Insekten-ZNS zu gewinnen (Stern et al., 2010). 5 Studienübergreifende Diskussion 43 Erste NADPHd-positive Neuronen lassen sich sowohl im Gehirn als auch in der Bauchganglienkette schon am Ende der ersten Hälfte der Embryonalentwicklung nachweisen. Die großen sensorischen und motorischen Verarbeitungszentren im Hirn, wie z.B. die optischen Loben, die Antennalloben und der Zentralkomplex, exprimieren NOS aber erst relativ spät während ihrer Entwicklung, wenn zumindest ihre grobe Anatomie schon festgelegt ist. Die Pilzkörper exprimieren NOS sogar erst in nennenswertem Maße einige Tage nach dem Schlupf, wenn zumindest ihre prinzipielle Verschaltung abgeschlossen ist. Weiterhin setzt selbst in den Strukturen, die schon während der Embryogenese NOS exprimieren, die Citrullin-Immunreaktivität deutlich später ein, in vielen Regionen erst kurz vor dem Schlupf. Diese Befunde sprechen gegen eine Beteiligung von NO beim Auswachsen und der Wegfindung von Neuriten zum Aufbau der komplexen Hirnstruktur. Im peripheren Nervensystem gibt es vielfältige Hinweise auf eine essentielle Beteiligung des NO-cGMPSignalweges gerade beim Aufbau der grundlegenden Strukturen, etwa bei den Pionierneuronen in der Antenne (Seidel und Bicker, 2000) und im Bein (Pätschke und Bicker, 2007), oder beim Aufbau des Mitteldarmplexus (Haase und Bicker, 2003; Stern et al., 2007; Knipp und Bicker, 2009). Dies ist möglicherweise im Heuschrecken-ZNS prinzipiell anders, denn für NO-Produktion gibt es in dem entsprechenden Zeitfenster keine Evidenz, es sei denn man postuliert eine NO-Quelle außerhalb der Neuronen des ZNS. Beim Tabakschwärmer Manduca z.B. spielt von Gliazellen produziertes NO eine wichtige Rolle beim Aufbau der Antennallobus-Struktur während der Puppenentwicklung (Gibson et al., 2001). Im Heuschreckenhirn ist NOS zwar auch in Gliazellen gefunden worden, die die Pilzkörper umschließen (Bicker und Hähnlein, 1995), aber nicht im Antennallobus, der bei der Heuschrecke weitgehend frei von Gliazellen ist. Eine ganz andere externe NO-Quelle könnten Hämozyten sein, von denen bis zu 30% NOS exprimieren, und die auch schon in der ersten Hälfte der Embryonalentwicklung aktiv sind (Stern und Bicker, 2008a; Stern et al., 2010). Mit dem sich entwickelnden peripheren Nervensystem kommen die Hämozyten in direkten Kontakt, und könnten hier tatsächlich an der NO-Regulation des Neuritenwachstums beteiligt sein. Für das schon recht früh geschlossene ZNS und insbesondere das Gehirn trifft dies allerdings nicht zu, und da Hirngewebe NO sehr effektiv und schnell inaktiviert (Garthwaite, 2008), wäre der Wirkungsradius solcher einzelner externer NO-Quellen auch sehr begrenzt. Falls tatsächlich eine frühe externe NO-Quelle vorhanden und wirksam wäre, stünde vielen Zellen schon der NO-Rezeptor sGC zur Verfügung, der im Bauchmark der Heuschrecke zwar vor allem während der zweiten Hälfte der Embryonalentwicklung stark exprimiert ist 5 Studienübergreifende Diskussion 44 (Truman et al., 1996), aber der auch schon bei ca. 40% der Entwicklung dort, sowie auch im Gehirn vorkommt (Stern et al., 2010). Dass zumindest einige identifizierte Neuronen nicht nur den funktionellen NO-Rezeptor exprimieren, sondern dass auch deren Auswachsen von NO über Vermittlung von cGMP positiv beeinflusst wird, konnte in der Regenerations-Studie gezeigt werden (Stern et al., 2008a). Da zu diesem Zeitpunkt ca. 50 weitere Neuronen pro Hemiganglion NO-abhängig cGMP produzieren, ist zu erwarten, dass auch das Wachstum anderer Axone durch verletzungsbedingte NO-Freisetzung beeinflusst wird. Selbst nicht direkt NO-responsive Zellen können indirekt einem nitrergen Einfluss unterliegen, wie z.B. für das Auswachsen der serotonergen Axone auf dem Mitteldarm gezeigt (Stern et al., 2007). Die funktionelle Signifikanz des Regenerationsvermögens embryonaler Neuronen ist nicht klar, da eine Verletzung des ZNS im in der Erde geschützten Ei kaum auftreten wird. Möglicherweise zeigt sich hier im Embryo schon das Regenerationsvermögen, das im postembryonalen Nervensystem von Bedeutung sein kann und dort auch im Verlauf der vorliegenden Arbeit nachgewiesen worden ist (Pätschke et al., 2004). Neuronale Regeneration wurde schon bei verschiedenen Gruppen der Evertebraten untersucht, z.B. bei Krebsen (Bittner, 1991), Blutegeln (Carbonetto und Muller, 1972) und Schnecken (Fredman und Nutz, 1988; Moffet, 1995). Bei Insekten gibt es relativ viele Daten über Regeneration im peripheren Nervensystem, sowohl auf der zellulären als auch auf der Verhaltens-Ebene, z.B. im auditorischen System der Heuschrecke (Lakes-Harlan und Pfahlert, 1995), oder dem mechanosensorischen Fluchtreflexsystem von Grillen (Chiba und Murphey, 1991) und Schaben (Stern et al., 1997). Über Regeneration im Zentralnervensystem weiß man bei Insekten allerdings noch recht wenig, dort vor allem von Schaben (Spira et al., 1987) und Grillen (Roederer und Cohen, 1983). Gerade die Identifizierung von Faktoren, die ZNSRegeneration fördern, ist aus medizinischer Sichtweise sehr interessant. Im Wirbeltier- und Menschen-ZNS gibt es, im Gegensatz zum PNS, eine ganze Reihe von Faktoren, die zentralnervöse Regeneration behindern und meist sogar verhindern. Ein Großteil der Probleme beruht auf Vernarbungsprozessen unter Beteiligung diverser GliazellTypen und anderer Zellen. Die Hemmung des Wiederauswachsens verletzter Neurone wird über eine Vielzahl von Molekülen vermittelt (reviews: Horner und Gage, 2000; Schwab, 2002) Dazu gehören z.B. Bestandteile der Basallamina, inhibitorische Wachstumsfaktoren oder Chondroitin-Sulfat-Proteoglykane (CSPGs). Einige dieser Faktoren sind inzwischen identifiziert und kloniert, z.B. das Protein Nogo, und es konnte gezeigt werden, dass deren inhibitorischer Einfluss auf ZNS-Regeneration mit spezifischen Antikörpern vermindert 5 Studienübergreifende Diskussion 45 werden kann (review Schwab, 2002), allerdings nur graduell und mit starken Nebenwirkungen. Die zusätzliche Anwendung regenerationsfördernder Faktoren könnte hier entscheidende Fortschritte bringen (Benowitz and Yin, 2007). Es konnte kürzlich gezeigt werden, dass zyklisches AMP die Regenerationsfähigkeit von spinalen Rattenneuronen entscheidend beeinflußt (Neumann et al., 2002, Qiu et al., 2002). Solche positiven Regulatoren lassen sich am besten dort untersuchen, wo die negativen Faktoren weitgehend fehlen - bei Wirbellosen, wie z.B. dem Heuschreckenembryo. Der Einfluss von NO auf Regeneration geschädigten Nervengewebes ist auch aus anderen Organismen bekannt, z.B. aus dem ZNS des Blutegels (Duan et al. 2005), aber dort wirkt NO indirekt über die Vermittlung von Mikroglia. Auch bei der Regeneration des peripheren Nervensystems von Wirbeltieren scheint es eine Rolle zu spielen (Keilhoff et al., 2002), hier allerdings durch Förderung der für Regeneration wichtigen vorhergehenden Wallerschen Degeneration. 5.1.4 Stickstoffmonoxid im Vertebraten-ZNS Im Wirbeltier-ZNS ist NO als Modulator synaptischer Transmission schon seit einiger Zeit nachgewiesen, z.B. bei der Langzeitpotenzierung in Hippocampus-Neuronen (Arancio et al., 1996). Auch auf der Ebene der strukturellen Plastizität hat Stickstoffmonoxid in diesem System einen entscheidenden Einfluss. Nikonenko et al. (2008) konnten zeigen, dass die NOFreisetzung aus hippocampalen Neuronen retrograd eine multiaxonale Innervation der dendritischen Spines induzieren kann. Auch auf die Migration von Neuronen und neuronalen Vorläuferzellen scheint NO eine Rolle zu spielen, und zwar sowohl als Sender, z.B. im menschlichen Rückenmark (Foster und Phelps, 2000) als auch als Empfänger z.B. bei Neuroblasten des rostralen migratorischen Stroms (Guiterrez-Mecinas et al., 2007). Dies lässt sich auch in vitro an der menschlichen neuronalen Vorläuferzelllinie NT2 nachweisen. Die Migration neuronaler Vorläufer aus sphärischen Aggregaten (siehe Abb. 3) wird durch das NO/cGMP-Signalsystem beschleunigt (Tegenge und Bicker, 2009). Dies zeigt wiederum die Parallelität von Entwicklungsvorgängen bei menschlichen Neuronen und bei der Entwicklung des ENS von Insekten, bei denen Migration ebenfalls durch NO stimuliert wird (Haase und Bicker, 2003; Knipp und Bicker, 2009a). Dass das NO/cGMP-Signalsystem bei der Exozytose synaptischer Vesikel eine Rolle spielt haben die hier dargestellten Ergebnisse an NT2-Neuriten in Kultur gezeigt (Tegenge at al., 2009). Der genaue Mechanismus dieses Einflusses auf die Vesikelfusion ist noch nicht geklärt, aber die Beteiligung der Proteinkinase G konnte durch pharmakologische 5 Studienübergreifende Diskussion 46 Manipulationen belegt werden. Es ist bekannt, dass Phosphorylierung synaptischer Proteine die Vesikelfusion maßgeblich beeinflusst (Leenders und Sheng, 2005). Auch bei Wirbellosen ist dies gezeigt worden, z.B. anhand der Phosphorylierung von Synapsin bei der Langzeitpotenzierung an Schnecken-Synapsen (Fiumara et al., 2007). In diesem Zusammenhang ist zu erwähnen, dass NO auch die Vesikelfusion an der neuromuskulären Synapse von Drosophila stimuliert (Wildemann und Bicker, 1999). Diese NO-abhängige Exozytose ist Kalzium-unabhängig, und damit auch unabhängig von neuronaler Aktivität. Kalzium-unabhängige, spontane Exozytose ist ein weit verbreitetes Phänomen, das auch an NT2-Neuronen nachgewiesen werden konnte (Podrygajlo et al., 2009b). In den letzten Jahren hat sich gezeigt, dass diese nicht aktivtätsabhängige Neurotransmitterfreisetzung wichtig für synaptische Reifung und synaptische Stabilität ist (review: Wasser und Kavalali, 2009). Hierzu könnte NO-induzierte vesikuläre Exozytose einen Beitrag leisten. Auch so prinzipelle Eigenschaften der postsynaptischen Zelle wie die Expression des passenden Rezeptors können durch den Neurotransmitter des präsynaptischen Neurons bestimmt werden. So exprimieren z.B. frühembryonale Wirbeltiermuskeln nicht nur die zu erwartenden Acetylcholin-Rezeptoren, sondern zunächst auch in geringer Menge Rezeptoren für Glutamat, GABA und Glycin, und können durch experimentelle Änderungen der Erregbarkeit spinaler Neuronen, die einen anderen Transmitter als Acetylcholin enthalten, dazu gebracht werden, z.B. funktionelle glutamaterge oder GABAerge Synapsen auszubilden (Borodinsky und Spitzer, 2007). Solches aktivitätsabhängiges "Transmitter-RezeptorMatching" kann sogar in adulten Ratten gezeigt werden. Brunelli et al. (2005) konnten zeigen dass nach Operationen bei denen der motorische Nerv durchtrennt und durch einen von glutamatergen Axonen durchzogenen Nerv ersetzt wurde, vollständige glutamaterge neuromuskuläre Synapsen ausgebildet wurden. Somit können zumindest neuromuskuläre Synapsen abhängig vom Phänotyp der innervierenden Neuronen unterschiedliche Neurotransmitter verwenden. Das kann auch dazu führen, dass dieselbe Muskelfaser von Motoneuronen unterschiedlichen Transmittertyps innerviert wird und z.B. gleichzeitig cholinerge und glutamaterge Synapsen ausbildet, so wie in der vorliegenden Arbeit an der neuromuskulären Synapse von Onychophoren gezeigt wurde (Stern und Bicker, 2008b). 5 Studienübergreifende Diskussion 47 5.2 Ausblick Neben den direkten Erkenntnissen über neuronale Plastizität hat die vorliegende Arbeit auch einen Beitrag zur Etablierung von Modellsystemen zur weiteren Erforschung dieser Phänomene geleistet. Überdies lassen sich solche Modellsysteme auch in der pharmakologischen und toxikologischen Forschung einsetzen. Insbesondere die Entwicklung eines Modells für axonale Regeneration im ZNS, das für pharmakologische Manipulationen zugänglich ist, kann bei der Suche nach regenerationsfördernden Substanzen helfen. Das menschliche neuronale NT2-Zellkultursystem, das nun in einer zeit- und kosteneffizienten Variante als gut charakterisiert gelten kann, kann nicht nur zur Aufklärung entwicklungsneurobiologischer Fragestellungen beitragen, sondern auch für toxikologische Untersuchungen eingesetzt werden. Im REACH Programm der Europäischen Union, bei dem ca. 140.000 Substanzen hinsichtlich ihres toxischen Potentials bewertet werden müssen, werden Tests zur entwicklungsbedingten Neurotoxizität empfohlen wenn sich Hinweise darauf aus anderen Studien ergeben haben (ECHA, 2009). In diesem Rahmen kann die Verwendung von Zellkultursystemen zu einer signifikanten Verringerung von Tierversuchen beitragen, mit dem zusätzlichen Vorteil, dass es sich bei NT2 um menschliche Zellen handelt. Erste entwicklungsneurotoxikologische Studien mit NT2 sind bereits durchgeführt worden, und die Möglichkeit des Einsatzes dieser Zelllinie für high-throughput screening von potentiell entwicklungsneurotoxischen Substanzen ist zur Zeit in unserem Labor in Erprobung (Stern et al., 2009). 6 Literatur 48 6. Literatur Aguinaldo AM, Turbeville JM, Linford LS, Rivera MC, Garey JR, Raff RA, Lake JA (1997) Evidence for a clade of nematodes, arthropods and other moulting animals. Nature 387:489493 Andrews PW (1984) Retinoic acid induces neuronal differentiation of a cloned human embryonal carcinoma cell line in vitro. Dev Biol 103:285-293 Arancio O, Kiebler M, Lee CJ, Lev-Ram V, Tsien RY, Kandel ER, Hawkins, RD (1996) Nitric oxide acts directly in the presynaptic neuron to produce long-term potentiation in cultured hippocampal neurons. Cell 87:1025-1035 Aston-Jones G, Rajkowski J, Cohen J (1999) Role of locus coeruleus in attention and behavioral flexibility. Biol Psychiatry 46:1309-1320 Ayali A (2004) The insect frontal ganglion and stomatogastric pattern generator networks. Neurosignals 13:20-36 Bacon JP, Thompson KSJ, Stern M (1995) Identified octopaminergic neurons provide an arousal mechanism in the locust brain. J Neurophysiol 74:2739-2743 Ball EE, Truman JW (1998) Developing grasshopper neurons show variable levels of guanylyl cyclase activity on arrival at their targets. J Comp Neurol 394:1-13. Barker EL, Perlman MA, Adkins EM, Houlihan WJ, Pristupa ZB, Niznik HB, Blakely RD (1998) High affinity recognition of serotonin transporter antagonists defined by speciesscanning mutagenesis. J Biol Chem 273:19459-19468 Benowitz LI, Yin Y (2007) Combinatorial treatments for promoting axon regeneration in the CNS: strategies for overcoming inhibitory signals and activating neurons’ intrinsic growth state. Develop Neurobiol 67:1148-1165 Bicker G (2007) Pharmacological approaches to nitric oxide signalling during neural development of locusts and other model insects. Arch Insect Biochem Physiol 64:43-58 Bicker G, Hähnlein I (1995) NADPH diaphorase expression in neurons and glial cells of the locust brain. NeuroReport 6:325-328 Bicker G, Schmachtenberg O, de Vente J (1997) Geometric considerations of nitric oxidecyclic GMP signalling in the glomerular neuropil of the locust antennal lobe. Proc R Soc Lond B 264:1177-1182 Bicker G, Stern M (2002) Neuronale Botenstoffe des Insektengehirns. TiHo Forschung fürs Leben 2002:25-30 Bicker G, Stern M (2009) NO als Regulator neuronaler Motilität und Regeneration in einfachen Nervensystemen. Neuroforum 1/09:4-12 Biskup S, Moore DJ (2006) Detrimental deletions: mitochondria, aging and Parkinson's disease. Bioessays 28:963-967 Bittner GD (1991) Long-term survival of anucleate axons and its implication for nerve regeneration. Trends Neurosci 14:188-193 Boillee S, Vande Velde C, Cleveland DW (2006) ALS: a disease of motor neurons and their nonneuronal neighbors. Neuron 52:39-59 Carbonetto S, Muller KJ (1972) A regenerating neurone in the leech can form an electrical synapse on its severed axon segment. Nature 267:450-451 6 Literatur 49 Borodinsky LN, Spitzer NC (2007) Activity-dependent neurotransmitter-receptor matching at the neuromuscular junction. Proc Natl Acad Sci USA 104:335-340 Boyan G, Williams L (2000) Building the antennal lobe: engrailed expression reveals a contribution from protocerebral neuroblasts in the grasshopper Schistocerca gregaria. Arthropod Struct Dev 29:267-274 Boyan GS, Williams JL, Herbert Z (2008) Fascicle switching generates a chiasmal neuroarchitecture in the embryonic central body of the grasshopper Schistocerca gregaria. Arthropod Struct Dev 37:539-544 Brunelli G, Spano P, Barlati S, Guarneri, B, Barbon A, Bresciani R, Pizzi M (2005) Glutamatergic reinnervation through peripheral nerve graft dictates assembly of glutamatergic synapses at rat skeletal muscle. Proc Natl Acad Sci USA 102:8752-8757 Butler SG, Meegan MJ (2008) Recent developments in the design of anti-depressive therapies: targeting the serotonin transporter. Curr Med Chem 15:1737-1761 Buznikov GA, Nikitina LA, Voronezhskaya EE, Bezuglov VV, Willows AOD, Nezlin LP (2003) Localization of serotonin and its possible role in early embryos of Tritonia diomedea (Mollusca: Nudibranchia). Cell Tissue Res 311:259-266 Casper, ML, Barnett, E, Williams, GI, Halverson, JA, Braham, VE, Greenlund, KJ (2003) Atlas of Stroke Mortality: Racial, Ethnic, and Geographic Disparities in the United States, Atlanta GA: Department of Health and Human Services, Centers for Disease Control and Prevention Champlin DT, Truman JW (2000) Ecdysteroid coordinates optic lobe neurogenesis via a nitric oxide signaling pathway. Development 127:3543-3551 Chiba A, Murphey RK (1991) Connectivity of identified central synapses in the cricket is normal following regeneration and blockade of presynaptic activity. J Neurobiol 22:130-142 Colas J-F, Launay J-M, Kellermann O, Rosay P, Maroteaux L (1995) Drosophila 5-HT2 serotonin receptor: coexpression with fushitarazu during segmentation. Proc Natl Acad Sci USA 92:5441-5445 Condron BG (1999) Serotonergic neurons transiently require a midline-derived FGF signal. Neuron 24:531-540 Copenhaver PF, Taghert PH. (1989) Development of the enteric nervous system in the moth. I. Diversity of cell types and the embryonic expression of FMRFamide-related neuropeptides. Dev Biol 131:70-84 Dawson TM, Bredt DS, Fotuhi M, Hwang PM, Snyder SH (1991) Nitric oxide synthase and neuronal NADPH diaphorase are identical in brain and peripheral tissues. Proc Natl Acad Sci USA 88:7797-7801 Devlin CL (2001) The pharmacology of g-aminobutyric acid and acetylcholine receptors at the echinoderm neuromuscular junction. J exp Biol 204:887-896 Diefenbach TJ, Sloley BD, Goldberg JI. 1995. Neurite branch development of an identified serotonergic neuron from embryonic Helisoma: evidence for autoregulation by serotonin. Dev Biol 167:282-293. Dittrich R, Bossing T, Gould AP, Technau GM, Urban J (1997) The differentiation of the serotonergic neurons in the Drosophila ventral nerve cord depends on the combined function of the zinc finger proteins Eagle and Huckebein, Development 124:2515-2525 6 Literatur 50 Duan Y, Haugabook SJ, Sahley CL, Muller KJ (2003) Methylene blue blocks cGMP production and disrupts directed migration of microglia to nerve lesions in the leech CNS. J Neurobiol 57:183-192 Duerr JS, Han H-P, Fields SD, Rand JB (2008) Identification of major classes of cholinergic neurons in the nematode Caenorhabditis elegans. J Comp Neurol 506:887-896 ECHA (2009) ECHA general report 2008, ECHA-09-A-01-EN. Edwards, JS (1969) Postembryonic development and regeneration of the insect nervous system. Adv Ins Physiol 6:97-137 Ewer DW, van den Berg R (1954) A note on the pharmacology of the dorsal musculature of Peripatopsis. J exp Biol 31:497-500 Fiumara F, Milanese C, Corradi A, Giovedi S, Leitinger G., Menegon A, Montarolo PG, Benfenati F, Ghirardi M (2007) Phosphorylation of synapsin domain A is required for posttetanic potentiation. J Cell Sci 120:3228-3237 Florey E, Florey E (1965) Cholinergic neurones in the Onychophora: A comparative study. Comp Biochem Physiol 15:125-136 Foster JA, Phelps PE (2000) NADPH-diaphorase reveals presumptive sympathetic primary afferents in the developing human spinal cord. Auton Neurosci 84:111-117 Fredman S, Nutz P (1988) Regeneration of identified neurons and their synaptic connections in the nervous system of Aplysia. Am Zool 28:1099-1108 Frisch K v. (1965) Tanzsprache und Orientierung der Bienen. Springer, Heidelberg Ganfornina MD, Sanchez D, Bastiani MJ. 1996. Embryonic development of the enteric nervous system of the grasshopper Schistocerca americana. J Comp Neurol 372:581-596 Garthwaite J (2008) Concepts of neural nitric oxide-mediated transmission. Eur J Neurosci 27:2783-2802 Gershon MD (2004) Review article: serotonin receptors and transporters - roles in normal and abnormal gastrointestinal motility. Aliment Pharmacol Ther 20(Suppl 7):3-14 Gewecke M, Hou T (1993) Visual brain neurons in Locusta migratoria. In: Wiese K, Gribakin FG, Popov AV, Renninger G (eds) Sensory systems of arthropods Birkhäuser, Basel, pp. 119144 Gibbs SM, Becker A, Hardy RW, Truman JW (2001) Soluble guanylate cyclase is required during development for visual system function in Drosophila. J Neurosci 21:7705-7714 Gibbs SM, Truman JW (1998) Nitric oxide and cyclic GMP regulate retinal patterning in the optic lobe of Drosophila. Neuron 20:83-93 Gibson NJ, Nighorn A (2000) Expression of nitric oxide synthase and soluble guanylyl cyclase in the developing olfactory system of Manduca sexta. J Comp Neurol 422:191-205 Gutièrrez-Mecinas M, Crespo C, Blasco-Ibáñez JM, Nácher J, Varea E, Martínez-Guijarro FJ (2007) Migrating neuroblasts of the rostral migratory stream are putative targets for the action of nitric oxide. Eur J Neurosci 26:392-402 Gibson NJ, Rössler W, Nighorn AJ, Oland LA, Hildebrand JG, Tolbert LP. 2001. Neuron-glia communication via nitric oxide is essential in establishing antennal-lobe structure in Manduca sexta. Dev Biol 240:326-339 6 Literatur 51 Giribet G, Edgecombe GD, Wheeler WC (2001) Arthropod phylogeny based on eight molecular loci and morphology. Nature 413:157-161 Goldberg JI, Kater SB (1989) Expression and function of the neurotransmitter serotonin during development of the Helisoma nervous system. Dev Biol 131:483-495 Goodman CS, Bate M (1981) Neuronal development in the grasshopper. Trends Neurosci 4:163-169 Haase A, Stern M, Wächtler K, Bicker G (2001) A tissue-specific maker of ecdysozoa. Dev Genes Evol 211:428-433 Haase A., Bicker G (2003) Nitric oxide and cyclic nucleotides are regulators of neuronal migration in an insect embryo. Development 130:3977-3987 Hammer M (1993) An identified neuron mediates the unconditioned stimulus in associative olfactory learning in honeybees. Nature 366: 59-63. Harris-Warrick RM, Marder E (1991) Modulation of neural networks for behaviour. Annu Rev Neurosci 14:39-57 Hartenstein V (1997) Development of the insect stomatogastric nervous system. Trends Neurosci 20:421-427 Hartley RS, Margulis M, Fishman PS, Lee VM, Tang CM (1999) Functional synapses are formed between human NTera2 (NT2N, hNT) neurons grown on astrocytes. J Comp Neurol 407:1-10 Hatten ME (1999) Central nervous system neuronal migration. Annu Rev Neurosci 22:511-539 Hawkins RD, Kandel ER, Bailey CH (2006) Molecular Mechanisms of Memory Storage in Aplysia. Biol Bull 210: 174-191 Henning W (1980) Phylogenetische Systematik. Paul Parey, Berlin Hess DT, Patterson SI, Smith DS, Skene JH (1993) Nature 366:562-556 Hill EJ, Woehrling M, Prince M, Coleman MD (2008) Differentiating human NT2/D1 neurospheres as a versatile in vitro 3D model system for developmental neurotoxicity testing. Toxicology 249:243-250 Hill ES, Blagburn JM (2001) Presynaptic effects of biogenic amines modulating synaptic transmission between identified sensory neurons and giant interneurons in the first instar cockroach. J Comp Physiol A 187:633-645 Hope BT, Michael GJ, Knigge KM, Vincent SR (1991) Neuronal NADPH diaphorase is a nitric oxide synthase. Proc Natl Acad Sci USA 88:2811-2814 Horner PJ, Gage FH (2000) Regeneration in the damaged central nervous system. Nature 407:963-970 Hoyte L, Barber PA, Buchan AM, Hill MD (2004) The rise and fall of NMDA antagonists for ischemic stroke. Curr Mol Med 4:131-136 Iacovitti L, Stull ND, Jin H (2001) Differentiation of human dopamine neurons from an embryonic carcinomal stem cell line. Brain Res 912:99-104 Karnovsky MJ, Roots L (1964) A “direct-coloring” thiocholine method for cholinesterases. J Histochem Cytochem 12:219-221 Keating C, Lloyd PE (1999) Differential modulation of motor neurons that innervate the same muscle but use different excitatory transmitters in Aplysia. J Neurophysiol 82:1759-1767 6 Literatur 52 Keilhoff G, Fansa H, Wolf G (2002) Differences in peripheral nerve degeneration/regeneration between wild-type and neuronal nitric oxide synthase knockout mice. J Neurosci Res 68:432441 Keshishian H, O’Shea M (1985) The distribution of a neuropeptide transmitter in the postembryonic grasshopper central nervous system. J Neurosci 5:992-1004 Knipp S, Bicker G (2009a) Regulation of enteric neuron migration by the gaseous messenger molecules CO and NO. Development 136:85-93 Knipp S, Bicker G (2009b) A developmental study of enteric neuron migration in the grasshopper using immunological probes. Dev Dyn 238:2837-2849 Kojima H, Nakatsubo N, Kikuchi K, Kawahara S, Kirino Y, Nagoshi H, Hirata Y, Nagano T (1998) Detection and imaging of nitric oxide with novel fluorescent indicators: diaminofluoresceins. Anal Chem 70:2446-2453 Kurylas AE, Ott SR, Schachtner J, Elphick MR, Williams L, Homberg U (2005) Localization of nitric oxide synthase in the central complex and surrounding midbrain neuropils of the locust Schistocerca gregaria. J Comp Neurol 484:206-233 Lakes-Harlan R, Pfahlert C (1995) Regeneration of axotomized tympanal nerve fibres in the adult grasshopper Chorthippus biguttulus (L.) (Orthoptera: Acrididae) correlates with regaining the localization ability. J Comp Physiol A 176:797-807 Lauder JM (1988) Neurotransmitters as morphogens. Prog Brain Res 73:365-387 Lauder JM, Schambra UB (1999) Morphogenetic roles of acetylcholine. Environ Health Perspect 107(Supp 1):65-69 Lebrand C, Cases O, Wehrle R, Blakely RD, Edwards RH, Gaspar P (1998) Transient developmental expression of monoamine transporters in the rodent forebrain. J Comp Neurol 401: 506 -524 Lebrand C, Cases O, Wehrle R, Blakely RD, Edwards RH, Gaspar P(1998) Transient developmental expression of monoamine transporters in the rodent forebrain. J Comp Neurol 401:506-524 Leenders AG, Sheng ZH (2005) Modulation of neurotransmitter release by the second messenger-activated protein kinases: implications for presynaptic plasticity. Pharmacol Ther 105:69-84 Luffy D, Dorn A (1992) Immunohistochemical demonstration in the stomatogastric nervous system and effects of putative neurotransmitters on the motility of the isolated midgut of the stick insect, Carausius morosus. J Insect Physiol 37:287-299 Lundell MJ, Hirsh J (1998) eagle is required for the specification of serotonin neurons and other neuroblast 7-3 progeny in the Drosophila CNS. Development 125:463-472 Maden M (2002) Retinoid signalling in the development of the central nervous system. Nat Rev Neurosci 3:843-853 Martinelli GP, Friedrich VL Jr, Holstein GR (2002) L-citrulline immunostaining identifies nitric oxide production sites within neurons. Neuroscience 114:111-122 Mazer C, Muneyyirci J, Taheny K, Raio N, Borella A, Whitaker-Azmitia P (1997) Serotonin depletion during synaptogenesis leads to decreased synaptic desnsity and learning deficits in the adult rat: a possible model of neurodevelopmental disorders with cognitive deficits. Brain Res 760:68-73 6 Literatur 53 Menzel R, Heyne A, Kinzel C, Gerber B, Fiala A (1999) Pharmacological dissociation between the reinforcing, sensitizing, and response-releasing functions of reward in honeybee classical conditioning. Behav Neurosci 113:744-754 Moffett SB (1995) Neural regeneration in gastropod molluscs. Progr Neurobiol 46:289-330 Moiseiwitsch JRD, Lauder JM (1995) Serotonin regulates mouse cranial neural crest migration. Proc Natl Acad Sci USA 92:7182-7186 Molaei G, Lange AB (2003) The association of serotonin with the alimentary canal of the african migratory locust, Locusta migratoria: distribution, physiology and pharmacological profile. J Insect Physiol 49:1073-1082 Moreno-López B, Romero-Grimaldi C, Noval JA, Murillo-Carretero M, Matarredona ER, Estrada C (2004) Nitric oxide is a physiological inhibitor of neurogenesis in the adult mouse subventricular zone and olfactory bulb. J Neurosci 24:85-95 Müller U, Bicker G (1994) Calcium-activated release of nitric oxide and cellular distribution of nitric oxide-synthesizing neurons in the nervous system of the locust. J Neurosci 74:75217528 Munir M, Lu L, McGonigle P (1995) Excititoxic cell death and delayed rescue in human neurons derived from NT2 cells. J Neurosci 15:7847-7860 Neumann S, Bradtke F, Tessier-Lavigne M, Basbaum AI (2002) Regeneration of sensory axons within the injured spinal cord induced by intraganglionic cAMP elevation. Neuron 34:885893 Nguyen L, Rigo JM, Rocher V, Belachew S, Malgrange B, Rogister B, Leprince P, Moonen G (2001) Neurotransmitters as early signals for central nervous system development. Cell Tissue Res 305:187-202 Nikonenko I, Boda B, Steen S, Knott G, Welker E, Muller D (2008) PSD-95 promotes synaptogenesis and multiinnervated spine formation through nitric oxide signaling. J Cell Biol 183:1115-1127 Ott SR Delago A, Elphick MR (2004) An evolutionarily conserved mechanism for sensitization of soluble guanylyl cyclase reveals extensive nitric oxide-mediated upregulation of cyclic GMP in insect brain. Eur J Neurosci 20:1231-1244 Ott SR, Elphick MR (2002) Nitric oxide synthase histochemistry in insect nervous systems: methanol/formalin fixation reveals the neuroarchitecture of formaldehyde-sensitive NADPH diaphorase in the cockroach Periplaneta americana. J Comp Neurol 448: 165-185 Ott SR, Elphick MR (2003) New techniques for whole-mount NADPH-diaphorase histochemistry demonstrated in insect ganglia. J Histochem Cytochem 51: 523-532 Packer MA, Stasiv Y, Benraiss A, Chmielnicki E, Grinberg A, Westphal H, Goldman SA, Enikolopov G (2003) Nitric oxide negatively regulates mammalian adult neurogenesis. Proc Natl Acad Sci USA 100:9566-9571 Pätschke A, Bicker G (2007) Influence of NO/cGMP on the growth of peripheral pionee neurons in the grasshpper. Göttingen Neurobiology Report 2007:T2-3B Pätschke A, Bicker G, Stern M (2004) Axonal regeneration of proctolinergic neurons in the central nervous system of the locust. Dev Brain Res 150:73-76 Paquet-Durand F, Bicker G (2004) Hypoxic/ischaemic cell damage in cultured human NT-2 neurons. Brain Res 1011:33-47 6 Literatur 54 Paquet-Durand F, Bicker, G (2007) Human model neurons in studies of brain cell damage and neural repair. Curr Mol Med 7:541-554 Paquet-Durand F, Tan S, Bicker G (2003) Turning teratocarcinoma cells into neurons: rapid differentiation of NT-2 cells in floating spheres. Brain Res Dev Brain Res 142:161-167 Perez-Orive J, Mazor O, Turner GC, Cassenaer S, Wilson RI, Laurent G (2002) Oscillations and sparsening of odor representations in the mushroom body. Science 297:359-365 Pleasure SJ, Page CP, Lee VM (1992) Pure, postmitotic, polarized human neurons derived from NTera 2 cells provide a system for expressing exogenous proteins in terminally differentiated neurons. J Neurosci 12:1802-1815 Podrygajlo G, Tegenge MA, Gierse A, Paquet-Durand F, Tan S, Bicker G, Stern M (2009) Cellular phenotypes of human model neurons (NT2) after differentiation in aggregate culture. Cell Tissue Res. 336:439-452 Podrygajlo G, Song Y, Schlesinger F, Krampfl K, Bicker G (2009b) Synaptic currents and transmitter responses in human NT2 neurons differentiated in aggregate culture. Neurosci Lett. Epub ahead of print, doi:10.1016/j.neulet.2009.10.092 Radio NM, Mundy WR (2008) Developmental neurotoxicity testing in vitro: models for assessing chemical effects on neurite outgrowth. Neurotoxicology 29:361-376 Regulski M, Stasiv Y, Tully T, Enikolopov G (2004) Essential function of nitric oxide synthase in Drosophila. Curr Biol 14:881-882 Richards KS, Simon DJ, Pulver SR, Beltz BE, Marder E (2003) Serotonin in the developing stomatogastric system of the lobster, Homarus americanus. J Neurobiol 54:380-392 Rind FC (1987) Non-directional movement sensitive neurons of the locust optic lobe. J Comp Physiol A 161:477-494 Roberson ED, Mucke L (2006) 100 years and counting: prospects for defeating Alzheimer’s disease. Science 314:781-784Rast, 2001 Roeder T (1999) Octopamine in invertebrates. Prog Neurobiol 59:533-561 Roeder T (2005) Tyramine and octopamine: ruling behavior and metabolism. Annu Rev Entomol 50:447-477 Roederer E, Cohen MJ (1983) Regeneration of an identified central neuron in the cricket. J Neurosci 3:1835-1847 Rowell CHF (1971) Variable responsiveness of a visual interneurone in the free-moving locust, and its relation to behaviour and arousal. J Exp Biol 55:727-747 Sanchez D, Ganfornina MD, Bastiani MJ (1995) Contributions of an orthopteran to the understanding of neuronal pathfinding. Imm Cell Biol 73:565-574 Schachtner J, Klaassen L, Truman JW (1998) Metamorphic control of cyclic guanosine monophosphate expression in the nervous system of the tobacco hornworm, Manduca sexta. J Comp Neurol 396:238-252 Schmachtenberg O, Bicker G (1999) Nitric oxide and cyclic GMP modulate photoreceptor cell responses in the visual system of the locust. J Exp Biol 202:13-20 Schwab ME. 2002. Increasing plasticity and functional recovery of the lesioned spinal cord. Prog Brain Res 137:351-359 6 Literatur 55 Schwaerzel M, Monastirioti M, Scholz H, Friggi-Grelin F, Birman S, Heisenberg M (2003) Dopamine and octopamine differentiate between aversive and appetitive olfactory memories in Drosophila. J Neurosci 23:10495-10502 Seidel C, Bicker G (2000) Nitric oxide and cGMP influence axonogenesis of antennal pioneer neurons. Development 127:4541-4549 Sinakevitch I, Niwa M, Strausfeld NJ (2005) Octopamine-like immunoreactivity in the honey bee and cockroach: comparable organization in the brain and subesophageal ganglion. J Comp Neurol 488:233-254 Sinakevitch I, Strausfeld NJ (2006) Comparison of octopamine-like immunoreactivity in the brains of the fruit fly and blow fly. J Comp Neurol 494:460-475 Spira ME, Zeldes D, Hochner B, Dormann A (1987) The effects of microenvironment on the redifferentiation of regenerating neurones: neurite architecture, acetylcholine receptors and Ca2+ channel distribution. J exp Biol 132:111-131 Stern M (1999) Octopamine in the locust brain: Cellular distribution and functional significance in an arousal mechanism. Micr Res Tech 45:135-141 Stern M (2009) The PM1 neurons, movement sensitive centrifugal visual brain neurons in the locust: anatomy, physiology, and modulation by identified octopaminergic neurons. J. Comp. Physiol. A 195:123-137 Stern M (z. Publ. eingereicht) Serotonin controls development of serotonergic neurons in the enteric nervous system of the locust (Locusta migratoria). z. Publ. Eingereicht bei Neurosci Lett Stern M, Bicker G (2008a) Nitric oxide regulates axonal regeneration in an insect embryonic CNS. Dev Neurobiol 68:295-308 Stern M, Bicker G (2008b) Mixed cholinergic/glutamatergic neuromuscular innervation of Onychophora: A combined histochemical/electrophysiological study. Cell Tissue Res 333:333-338 Stern M, Böger N, Eickhoff R, Kerßen U, Lorbeer C, Ziegler M, Martinelli GP, Bicker G (2010) Development of nitrergic neurons in the nervous system of the locust embryo. J Comp Neurol 518:1157–1175 Stern M, Ediger VL, Gibbon CR, Blagburn JM, Bacon JP (1997) Regeneration of cercal filiform hair sensory neurons in the first-instar cockroach restores escape behaviour. J Neurobiol 33:439-458 Stern M, Gewecke M (1993) Spatial sensitivity profiles of motion sensitive neurons in the locust brain. In: Wiese K, Gribakin FG, Popov AV, Renninger G (eds) Sensory systems of arthropods Birkhäuser, Basel, pp. 184-195 Stern M, Gierse A, Tan S, Bicker G (2009) Human Ntera-2 cells as a predictive in vitro test system for developmental neurotoxicity. Göttingen Neurobiology Report 2009, T1-4A Stern M, Knipp S, Bicker G (2007) Embryonic differentiation of serotonin-containing neurons in the enteric nervous system of the locust (Locusta migratoria). J Comp Neurol 501:38-51 Stern M, Thompson KSJ, Zhou P, Watson DG, Midgley JM, Gewecke M, Bacon JP (1995) Octopaminergic neurons in the locust brain: morphological, biochemical, and electrophysiological characterisation of potential modulators of the visual system. J Comp Physiol A 177:611-625 6 Literatur 56 Sullivan JM, Benton JL, Beltz BS (2000) Serotonin depletion in vivo inhibits the branching of olfactory projection neurons in the lobster deutocerebrum. J Neurosci 20:7716-7721 Suzuki N, Kojima H, Urano Y, Kikuchi K, Hirata Y, Nagano T (2001) Orthogonality of calcium concentration and ability of 4,5-diaminofluorescein to detect NO. J Biol Chem 277:47-49 Sykes PA, Condron BG (2005) Development and sensitivity to serotonin of Drosophila serotonergic varicosities in the central nervous system. Dev Biol 286:207-216 Taghert PH, Goodman CS (1984) Cell determination and differentiation of identified serotoninimmunoreactive neurons in the grasshopper embryo. J Neurosci 4:989-1000 Tegenge MA, Bicker G (2009) Nitric oxide and cGMP signal transduction positively regulates the motility of human neuronal precursor (NT2) cells. J Neurochem 110:1828-1841 Tegenge MA, Stern M, Bicker G (2009) Nitric oxide and cyclic nucleotide signal transduction modulates synaptic vesicle turnover in human model neurons. J Neurochem 111:1434-1446 Thomas JB, Bastiani MJ, Bate M, Goodman CS (1984) From grasshopper to Drosophila: a common plan for neuronal development. Nature 310:203-207 Trimm KR, Rehder V (2004) Nitric oxide acts as a slow-down and search signal in developing neurites. Eur J Neurosci 19:809-818 Truman JW, De Vente J, Ball EE (1996) Nitric oxide-sensitive guanylate cyclase activity is associated with the maturational phase of neuronal development in insects. Development 122:3949-3958 Trumm P, Dorn A (2000) Effects of azadirachtin on the regulation of midgut peristalsis by the stomatogastric nervous system in Locusta migratoria. Phytoparasitica 28:7-26 Valles AM, White K (1988) Serotonin-containing neurons in Drosophila melanogaster: development and distribution. J Comp Neurol 268:414-428 Wallace BG (1981) Neurotransmitter chemistry. In Muller KJ, Nicholls JG, Stent GS (eds) Neurobiology of the Leech. Cold Spring Harbor Publications, pp147-172 Wang DD, Kriegstein AR (2009) Defining the role of GABA in cortical development. J Physiol 587:1873-1879 Wasser CR, Kavalali ET (2009) Leaky synapses: regulation of spontaneous neurotransmission in central synapses. Neuroscience 158:177-188 Weinrich A, Kunst M, Wirmer A, Holstein GR, Heinrich R (2008) Suppression of grasshopper sound production by nitric oxide-releasing neurons of the central complex. J Comp Physiol A 194:763-776 Wendt B, Homberg U (1992) Immunocytochemistry of dopamine in the brain of the locust Schistocerca gregaria. J Comp Neurol 321:387-403. Wenzel B, Kunst M, Günther C, Ganter GK, Lakes-Harlan R, Elsner N, Heinrich R (2005) Nitric oxide/cyclic guanosine monophosphate signaling in the central complex of the grasshopper brain inhibits singing behavior. J Comp Neurol 488:129-139 Wildemann B, Bicker G (1999) Nitric oxide and cyclic GMP induce vesicle release at Drosophila neuromuscular junction. J Neurobiol 39:337-346 Zigova T, Barroso LF, Willing AE, Saporta S, McGrogan MP, Freeman TB, Sanberg PR (2000) Dopaminergic phenotype of hNT cells in vitro. Brain Res Dev Brain Res 122:87-90 6 Literatur 57 Zigova T, Willing AE, Tedesco EM, Borlongan CV, Saporta S, Snable GL, Sanberg PR (1999) Lithium chloride induces the expression of tyrosine hydroxylase in hNT neurons. Exp Neurol 157:251-258 Zimmermann H (1993) Synaptic transmission. Cellular and molecular basis. Thieme, Stuttgart, New York. 7 Zusammenfassung 58 7 Zusammenfassung In der vorliegenden Arbeit wurde der Einfluss einiger niedermolekularer neuronaler Botenstoffe auf neuronale Plastizität und Entwicklung an einfachen Nervensystemen untersucht. Gegenstand der Untersuchungen waren die Nervensysteme von Wirbellosen, vor allem Heuschrecken, und aus einer Krebszelllinie differenzierte menschliche Neuronen in Zellkultur. Das angewendete Methodenspektrum beinhaltete neuroanatomisch- immunzytologische Methoden in Kombination mit elektrophysiologischen und live-imaging Techniken, sowie in vitro-Gewebekultur und Zellkultur mit pharmakologischer Manipulation von zellulären Signaltransduktionswegen. Ein GABA-erges visuelles Interneuron aus dem Heuschreckenhirn wurde elektrophysiologisch charakterisiert und die Modulierbarkeit seiner Antwortmuster durch ein identifiziertes octopaminerges Interneuron wurde pharmakologisch nachgewiesen. Die Entwicklung der serotonergen Neuronen des enterischen Nervensystems der Heuschrecke wurde beschrieben und ihr Serotonin-Aufnahmesystem wurde nachgewiesen. Es wurde mit pharmakologischen Kulturexperimenten in vitro gezeigt, dass die Entwicklung ihres serotonergen Phänotyps unter autoregulatorischer Kontrolle von Serotonin selbst steht. Im Zentralnervensystem der Heuschrecke wurde die Entwicklung des StickstoffmonoxidcGMP-Signalweges mit einander ergänzenden histochemischen und ImmunfluoreszenzMethoden im Verlauf der Embryogenese verfolgt. Axonale Regeneration wurde im Zentralnervensystem der Heuschrecke zum ersten Mal nachgewiesen und ein Embryokultursystem entwickelt, um Regeneration unter pharmakologischer Behandlung in vitro zu studieren. Daran wurde der regenerationsfördernde Einfluss des NO-cGMP-Systems gezeigt. Aus der menschlichen Zelllinie Ntera-2 nach einer verbesserten Methode gewonnene Neuronen wurden umfassend hinsichtlich ihrer Neurotransmitter und präsynaptischer Proteine charakterisiert. Vesikuläre Exozytose wurde an diesem System erstmalig nachgewiesen und ihre Beeinflussung durch Stickstoffmonoxid und zyklische Nucleotide mit Imaging-Verfahren gezeigt. Die neuromuskulären Synapsen eines Onychophoren wurden elektrophysiologisch und immunzytochemisch charakterisiert. Die Innervierung der Muskeln durch multiple exzitatorische Neurotransmitter (Glutamat und Acetylcholin) wurde so nachgewiesen 8 Summary 59 8 Summary In the present study, the effect of low molecular weight neuroactive substances on neuronal plasticity and development were studied in simple nervous systems. Subjects of the studies were the nervous systems of invertebrates, in particular locusts, and human neurons differentiated from a carcinoma cell line. Applied methods included neuroanatomical and immunocytological techniques combined with electrophysiology and live imaging as well as cell and tissue culture under pharmacological treatment. A GABAergic visual interneuron of the locust brain was characterized anatomically and electrophysiologically and the modulation of its response patterns by an identified octopaminergic neuron was pharmacologically proven. The development of serotonergic neurons in the enteric nervous system of the locust was described and its serotonin uptake system was demonstrated. In pharmacological in vitro experiments it was shown that determination of their serotonergic phenotype is under autoregulatory control of serotonin itself. Using complementary histochemical and immunofluorescence techniques, the development of the nitric oxide/cGMP signaling pathway was studied in the locust central nervous system (CNS). Axonal regeneration was shown for the first time in locust CNS and an embryo culture system for the in vitro study of CNS regeneration under drug application was developed. Using this method, it was shown that NO/cGMP signaling enhances axonal regeneration. Neurotransmitters and synaptic proteins of neurons differentiated from the human cell line Ntera-2 using an improved method were characterized. Vesicular exocytosis was demonstrated for the first time in these cells. Using live imaging techniques, the effect of NO and cyclic nucleotides on presynaptic release was shown. The neuromuscular junctions of an onychophoran were characterized immunocytochemically and electropysiologically. It was shown that muscles receive excitatory input from multiple neurotransmitters (glutamate and acetylcholine). 9 Danksagung 60 9 Danksagung Mein besonderer Dank gilt Herrn Prof. Dr. Gerd Bicker für die langjährige Unterstützung und Ermunterung, die vielfältigen Anregungen, die gewährte wissenschaftliche Freiheit und die ständige Diskussionsbereitschaft. Ich bedanke mich bei allen Koautorinnen und Koautoren, die mit ihrem Fleiß und Engagement einen großen Teil der hier vorgestellten Arbeiten erst möglich gemacht haben. Bei allen ehemaligen und derzeitigen Mitgliedern der AG Zellbiologie und insbesondere bei Frau Saime Tan bedanke ich mich für die jederzeit angenehme Gesellschaft, Ablenkung, und vielfältige Diskussionen (nicht nur über wissenschaftliche Themen). Meiner Familie danke ich für die Unterstützung, die Geduld, die Liebe und das Verständnis, die mir den nötigen Rückhalt für die Anfertigung dieser Arbeit gegeben haben. 9 Danksagung 61 10 Darstellung des eigenen Anteils an den wissenschaftlichen Veröffentlichungen 62 10 Darstellung des eigenen Anteils an den wissenschaftlichen Veröffentlichungen Veröffentlichung 1: Pätschke A, Bicker G, Stern M (2004) Regeneration of proctolinergic neurons in the ventral nerve cord of the locust. Dev. Brain Res. 150:73-76. Idee: Bicker, Stern Durchführung der Experimente: Pätschke Auswertung der Experimente: Pätschke, Stern Diskussion, Beratung: Bicker, Stern Manuskript: Stern Veröffentlichung 2: Stern M, Knipp S, Bicker G (2007) Embryonic differentiation of serotonin-containing neurons in the enteric nervous system of the locust (Locusta migratoria). J. Comp. Neurol. 501:38-51. Idee: Stern; Knipp, Bicker Durchführung der Experimente: Stern, Knipp Auswertung der Experimente: Stern, Knipp Diskussion, Beratung: Bicker Manuskript: Stern Veröffentlichung 3: Stern M, Bicker G (2008a) Nitric oxide regulates axonal regeneration in an insect embryonic CNS. Dev. Neurobiol. 68:295-308. Idee: Stern, Bicker Durchführung der Experimente: Stern Auswertung der Experimente: Stern Diskussion, Beratung: Bicker Manuskript: Stern 10 Darstellung des eigenen Anteils an den wissenschaftlichen Veröffentlichungen 63 Veröffentlichung 4: Stern M, Bicker G (2008b) Mixed cholinergic/glutamatergic neuromuscular innervation of Onychophora: A combined histochemical/electrophysiological study. Cell Tissue Res. 333:333-338. Idee: Bicker, Stern Durchführung der Experimente: Stern Auswertung der Experimente: Stern Diskussion, Beratung: Bicker Manuskript: Stern Veröffentlichung 5: Stern M (2009) The PM1 neurons, movement sensitive centrifugal visual brain neurons in the locust: anatomy, physiology, and modulation by identified octopaminergic neurons. J. Comp. Physiol. A 195:123-137. Idee: Stern Durchführung der Experimente: Stern Auswertung der Experimente: Stern Manuskript: Stern Veröffentlichung 6: Bicker G, Stern M (2009) NO als Regulator neuronaler Motilität und Regeneration in einfachen Nervensystemen. Neuroforum 1/09:4-12. Idee: Bicker, Stern Manuskript: Bicker, Stern 10 Darstellung des eigenen Anteils an den wissenschaftlichen Veröffentlichungen 64 Veröffentlichung 7: Podrygajlo G, Tegenge MA, Gierse A, Paquet-Durand F, Tan S, Bicker G, Stern M (2009) Cellular phenotypes of human model neurons (NT2) after differentiation in aggregate culture. Cell Tissue Res. 336:439-452. Idee: Bicker, Stern Durchführung der Experimente: Stern, Podrygajlo, Tegenge, Gierse, Paquet-Durand, Tan Auswertung der Experimente: Stern, Podrygajlo, Tegenge, Gierse, Paquet-Durand Diskussion, Beratung: Bicker Manuskript: Stern, Podrygajlo Veröffentlichung 8: Tegenge MA, Stern M, Bicker G (2009) Nitric oxide and cyclic nucleotide signal transduction modulates synaptic vesicle turnover in human model neurons. J Neurochem. 111:1434-1446. Idee: Bicker, Tegenge, Stern Durchführung der Experimente: Tegenge Auswertung der Experimente: Tegenge Diskussion, Beratung: Bicker, Stern Manuskript: Tegenge, Bicker, Stern Veröffentlichung 9: Stern M, Böger N, Eickhoff R, Kerßen U, Lorbeer C, Ziegler M, Martinelli GP, Holstein GR, Bicker G (2010) Development of nitrergic neurons in the nervous system of the locust embryo. J. Comp. Neurol. 518:1157–1175 Idee: Stern, Bicker Durchführung der Experimente: Stern, Böger, Eickhoff, Kerßen, Lorbeer, Ziegler Auswertung der Experimente: Stern, Böger Diskussion, Beratung, Material: Bicker, Martinelli, Holstein Manuskript: Stern 10 Darstellung des eigenen Anteils an den wissenschaftlichen Veröffentlichungen 65 Veröffentlichung 10: Stern M (submitted) Serotonin controls development of serotonergic neurons in the enteric nervous system of the locust (Locusta migratoria). Neurosci. Lett., under review. Idee: Stern Durchführung der Experimente: Stern Auswertung der Experimente: Stern Manuskript: Stern