Genetik (Vererbungslehre)
Werbung
")
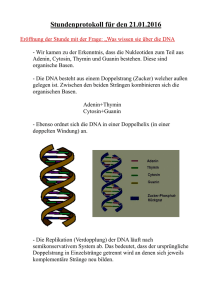
Genetik (Vererbungslehre) Ziele der Genetik sind z.B.: Herausfinden ob, warum und wie Merkmale von Eltern an direkte oder indirekte Nachkommen weiter gegeben werden. Auffinden von Genen die pos. oder neg. Auswirkungen auf Mensch, Tier und Pflanze haben. Suche nach Erklärungen für Erbkrankheiten. Versuch der Heilung von genetisch bedingten Krankheiten (Gentherapie) Im Rahmen der Gentechnik auch Herstellung von Genen, Einschleusen mit Vektoren, und Aktivierung von solchen Genen. u.v.m. Geschichte der Genetik und der Gentechnik: Ca. – 10 000 Jahre Selektion von Wildgräsern ( Urgetreideformen, vermutlich Weizen „Dinkel“) - 6 000 Jahre Bedeutung des Blütenstaubes (diözische Dattelpalmen in Ägypten) - 3 000 Jahre Haustierkreuzungen (z.B. Eselhengst und Pferdestute Maultier) Erfahrungen mit Inzucht Um 1800 Entdeckung der Spermien (bei Hunden) „Samen, die da reifen im Schoß der Mutter . . .“ ca. 20 Jahre später Entdeckung der Eizelle und der Befruchtung 1865 Gregor Mendels Erbgesetze im Buch „Versuche über Pflanzenhybriden“ 1900 Erkenntnis des Zellkerns als Träger der Erbinformation 1902 die Chromosomentheorie der Vererbung 1953 Entdeckung und Erforschung der DNA als Träger der Erbinformation 1975 Eigentlicher Beginn der Gentechnik mit der Entdeckung der Restriktionsenzyme (Chemische Stoffe zum gezielten Schneiden von DNA) 1988 Start des HUGO (=human genom organisation) oder besser des Human Genom Projects. Ziel war die vollständige Entschlüsselung des menschlichen Genoms ( Diskussion: Sequenzierung/Entschlüsselung) 2004 wurde HUGO aufgelöst, die Sequenzierung (!) war abgeschlossen. 1 I) Möglichkeiten der Zellteilung Die Mitose Eine Mutterzelle 2 identische Tochterzellen (hinsichtlich ihrer Erbinformation untereinander und mit der Mutterzelle identisch). Der Verlauf in erfolgt in 5 Phasen (Phasen sind aus dem durchgehenden Prozess herausgegriffene „Standbilder“, die mit wissenschaftlichen Namen versehen worden sind): 1. Interphase: (hier erfolgt die Verdoppelung der Erbinfo. → Replikation) Beim Menschen entsteht dabei zu jedem der 46 (23 x 2) einzelnen Stränge, welche die Erbinformation tragen eine Kopie. Diese Kopie bleibt jedoch mit dem Orginal an einem Punkt verbunden. Die so entstandenen X-förmigen Gebilde bezeichen wir künftig als Chromosomen. Die einzelnen Fäden aus denen sie bestehen sind die Chromatiden. Die Chromosomen sind zu diesem Zeitpunkt noch so dünn, dass sie im Lichtmikroskop auch bei stärkster Vergrößerung (ca. 2000fach) unsichtbar bleiben. 2. Prophase: Die Chromosomen kondensieren jetzt, d.h. sie spiralisieren sich und werden dadurch wesentlich dicker (natürlich auch kürzer!). Ab jetzt können die Chromosomen auch im Lichtmikroskop beobachtet werden. Anschließend ändert sich die Struktur der Kernmembran so, dass sie sich auflöst. Jetzt sind die Chromosomen frei im Cytoplasma. 3. Metaphase: In der Zelle entsteht jetzt unter Mitwirkung von zwei Zellorganellen, den Centriolen, ein kompliziertes Gerüst aus Proteinfäden. Diese Fäden führen von der an einem Pol liegenden Centriole weg und führen zur gegenüber liegenden. Aufgrund der Form dieses Fadengebildes wird das Ganze als Spindelapparat und die einzelnen Fäden als Spindelfäden bezeichnet. Die jetzt beginnende Bewegung der Chromosomen wird durch diese Spindelfäden bewirkt. Die Chromosomen kommen so in der Mitte der Zelle zwischen den beiden Centriolen in der Äquatorialebene - zu liegen. (Anordnung an Äquatorialebene, Aufbau des Spindelapparates), 4. Anaphase: Die Chromosomen werden jetzt unter der Zugwirkung der Spindelfäden in ihre beiden Chromatiden aufgetrennt. Je eine dieser Chromatiden kommt an den beiden Polen der Zelle - also in der Nähe der Centriolen - zu liegen. Nachdem die beiden Centriolen eine Kopie voneinander sind, befindet sich jetzt an jedem Zellpol eine vollständige Erbinformation. 5. Telophase: Rund um die beiden Chromatidengruppen wird je eine neue Kernmembran aufgebaut, die Zelle hat so kurzzeitig zwei identische Zellkerne. Durch Einstülpung der Zellmembran zwischen den beiden Zellkernen schnürt sich jetzt die Zelle langsam durch, so dass letztlich zwei neue Zellen entstanden sind. Bei Vielzellern bleiben diese Zellen aneinander haften, bei einzelligen Lebewesen trennen sie sich und bilden zwei neue, identische Lebewesen. Die neuen Zellen wachsen jetzt zur ursprünglichen Größe der Mutterzelle heran und dann beginnt im Allgemeinen der ganze Zyklus von vorne, also mit einer neuen Interphase. Die Meiose (= Reduktionsteilung = Voraussetzung für die geschlechtliche Fortpflanzung)) Eine diploide Mutterzelle (diploid = mit doppelter Erbinformation) 4 haploide Keimzellen (haploid = mit einfacher Erbinformation). Die Mutterzelle wird daher auch Keimzellenmutterzelle genannt. Die entstandenen Keimzellen sind hinsichtlich ihrer Erbinformation untereinander und im Vergleich mit der Mutterzelle auch qualitativ verschieden. 2 Auch hierbei zeigen sich typische Bewegungsabläufe (Phasen) in der Zelle. Zudem jedoch wird der ganze Prozess in zwei große Teilschritte unterteilt, die 1. und die 2. Reifeteilung. Jede dieser beiden Reifeteilungen beinhaltet die oben (Mitose!) genannten 5 Phasen: Also ... 1. Erste Reifeteilung: a. Interphase1 wie bei Mitose! b. Prophase1 wie bei Mitose! c. Metaphase1 beginnt wie in der Mitose. Wenn die Chromosomen jedoch in der Äquatorialebenen liegen, legen sich die homologen Chromosomen - also z.B. die beiden Chromosomen Nr. 1 - deckungsgleich aneinander. Dieser Vorgang heißt Chromosomenpaarung (= Synapsis). Während dieser Synapsis kann es zu einem für die Evolution der Lebewesen enorm wichtigen Ereignis kommen, zur intrachromosomalen Rekombination (= Crossing Over). Dabei werden zwischen den beiden homologen Chromosomen Stücke ausgetauscht. d. Anaphase1: Anders als bei einer Mitose werden jetzt nicht die Chromosomen in ihre Chromatiden „zerrissen“, sondern es werden die homologen Chromosomen wieder getrennt (ihre Information hat sich durch das C.O. inzwischen verändert) und zu den Zellpolen gezogen. e. Telophase1: Wie bei Mitose. Nachdem in der Anaphase 1 die homologen Chromosomen getrennt worden sind, besitzt jeder Zellpol jetzt nur mehr einen einzigen Partner der homologen Chromosomen. Daher ist die Zelle jetzt bereits als haploid zu bezeichnen (mit einfachem Chromomensatz!). Kernmembranbildung und Durchschnürung können auch unterbleiben, dann gibt es einen nahtlosen Übergang zur ... 2. Zweite Reifeteilung: Eine Teilung jedes der beiden neu entstandenen Zellkerne nach dem Schema einer Mitose (ohne Interphase bzw. ohne die Kondensation in der Prophase!). Man spricht von Prophase2, Metaphase2 ... etc. Die Durchschnürung am Ende erfolgt vollständig, die Zellen trennen sich. Letztendlich entstehen so insgesamt 4 Tochterzellen, die jetzt, weil sie nur mehr die Hälfte der Erbinformation der Keimzellenmutterzelle besitzen, als haploid (= mit einfacher Erbinfo. ausgestattet) bezeichnet werden (= Keimzellen = Geschlechtszellen = Ei- oder Samenzellen). Diese Keimzellen sind mit völlig unterschiedlicher Erbinformation ausgestattet. Durch Verschmelzung von zwei solchen Keimzellen bei der Befruchtung entsteht eine neue diploide Zelle (befruchtete Eizelle). Diese kann sich dann durch Millionen von Zellteilungen nach dem Muster der Mitose (Embryonalentwicklung) zu einem Organismus entwickeln. 3 II) Klassische Genetik (Vererbungslehre) Grundbegriffe: • Gene sind Programme für die Ausbildung von Merkmalen. Das gesamte Programmpaket (=Genom) für die Ausbildung von allen Merkmalen liegt im Zellkern jeder Zelle. • Stofflich gesehen sind Gene kettenartig gebaute Moleküle mit dem Kurznamen DNA (=DNS =Desoxiribinucleinsäure) • Die Gene liegen nacheinander aufgereiht auf den Chromosomen (das ist also ein sehr langer DNA-Strang). • Die Gesamtheit der Chromosomen einer Zelle bezeichnet man als Chromosomensatz. In sogenannten Körperzellen des Menschen sind das 23 Chromosomenpaare bzw. 46 Stück. In Körperzellen gibt es für jedes Merkmal zwei Gene – eines wurde bei der Befruchtung vom Vater beigesteuert, das andere von der Mutter Körperzellen haben also eine doppelte Erbiformation (man sagt, sie sind diploid). • In den Geschlechtszellen (= Keimzellen = Ei- und Samenzellen) liegen die Gene für jedes Merkmal nur noch einfach vor (Keimzellen sind haploid). Durch die Befruchtung entsteht wieder eine doppelte Information in der befruchteten Eizelle (=Zygote). • Im Chromosomensatz des Menschen unterscheidet man die Chromosomenpaare 1 – 22, die bei beiden Geschlechtern gleich aussehen. Im Chromomenpaar Nr. 23 zeigt sich der Unterschied zwischen männlichen (ein großes und ein kleines 23er Chromosom) und weiblichen (zwei große 23er Chromosomen) Organismen XX oder XY. Die Erbgesetze von Gregor Mendel 1865 Augustinermönch Gregor Mendel Veröffentlichtes Werk „Versuche über Pflanzenhybriden“ (Hybriden sind Mischlinge)! Mendel kreuzte (=geschlechtlich fortpflanzen) reinerbige Pflanzen mit verschiedenen Merkmalen und beobachtete diese Merkmale in den Folgegenerationen. Diese Beobachtungen wurden von G. Mendel statistisch erfasst und ausgewertet. Allgemeine Schreibweise zur Darstellung von Erbgesetzmäßigkeiten → Siehe Heft!: → Körperzellen → reinerbig (=homozygot) → mischerbig (heterozygot) → Keimzellen → diploid und haploid → Reduktionsteilung → Parentalgeneration und Filialgenerationen 1 und 2. o Beobachtung eines Merkmales (= monohybrider Erbgang) und beide für ein Merkmal zuständigen Gene wirken gleich stark (= intermediärer Erbgang) Bsp. Japanische Wunderblume (Alternative: Erbsenpflanzen) rot x weiß ... Aus dem erkennbaren Ergebnis in der F1-Generation formulierte Mendel die 1. Mendel´sche Regel (=Uniformitätsregel): „Kreuzt man in der Parentalgeneration reinerbige rot blühende Pflanzen mit reinerbig weiß blühenden, so treten in der F1-Generation ausschließlich gleich gefärbte, aber 4 heterozygote Pflanzen auf. Verhalten sich die Gene intermediär (Wunderblume) so sind die Nachkommen rosa, aber ebenfalls alle gleich. → Das bedeutet aber, dass man eine homozygote, rot blühende Pflanze bei einem dominant-rezessiven Erbgang nicht von einer hetorozygoten unterscheiden kann! Die beiden haben denselben Phänotypus ( =das Erscheinungsbild) aber unterschiedlichen Genotypus ( =das Erbbild). Die Unterscheidung wird mit Hilfe einer Rückkreuzung möglich: Rückkreuzung ist die Kreuzung der zu untersuchenden Form mit der homozygot rezessiven Ausgangsform (hier also einer weiß blühenden Pflanze!) → Schema für Rückkreuzung anfertigen! Tritt in der Filialgeneration nach einer Rückkreuzung auch nur eine einzige weiß blühende Pflanze auf, heißt das, dass die getestete rot blühende Pflanze nicht reinerbig ist! Im nächsten Schritt kreuzte Mendel Pflanzen aus der F1-Generation untereinander und beobachtete die F2-Generation. → Schema für Kreuzung anfertigen! Aus dem erkennbaren Ergebnis in der F2-Generation formulierte Mendel die 2. Mendel´sche Regel (=Spaltungsregel): „Kreuzt man die Pflanzen der F1-Geration untereinander, so treten in der F2-Generation unter anderem wiederum die Merkmale der P-Generation auf. Das Verhältnis in der F2 lautet bei intermediären Merkmalen: 1(rot) : 2(rosa) : 1(weiß)“ Beobachtet man das Verhältnis bei einem dom.-rez. Erbgang so zeigt sich ein Verhältnis 3(rot) : 1(weiß). Mendel hat bereits erkannt, dass hier dennoch auch das Verhältnis 1 : 2 : 1 : vorliegt. Allerdings gilt dieses Verhältnis nur für für den Genotypus! 5 o Beobachtung zweier Merkmale (= dihybrider Erbgang) dominant-rezessive Vererbung Bsp. 2 Kaninchenrassen: Rasse A: weiß und langhaarig Rasse B: schwarz und kurzhaarig Annahme: Die Merkmale „langhaarig“ und „schwarz“ dominieren! → Kreuzungsschema bis zur F2 durchführen (Kombinationsquadrat anfertigen)! Bei der Erstellung der Keimzellen, die in der F1 gebildet werden, ging G. Mendel davon aus, dass tatsächlich jede Kombination der untersuchten Gene möglich ist (Voraussetzung: es muss jedes Merkmal – aber nur durch nur ein Gen - vertreten sein!) Aus dem erkennbaren Ergebnis in der F2-Generation (Kombinationsquadrat) formulierte Mendel die 3. Mendel´sche Regel (=Kombinationsregel): „Die Merkmale aller Organismen vererben sich voneinander unabhängig und sind beliebig kombinierbar“ Hier irrte sich G. Mendel! Er hatte bei seinen Kreuzungsversuchen Merkmale beobachtet - das konnte er zu diesem Zeitpunkt noch nicht wissen - deren Gene sich auf verschiedenen Chromosomen befanden. Für solche ist eine beliebige Kombinierbarkeit tatsächlich gegeben. Hätte Mendel Merkmale beobachtet, deren Gene sich auf dem gleichen Chromosom befinden (gekoppelte Gene), hätte er die beliebige Kombinierbarkeit nicht feststellen können. Diese Entdeckung machte ein anderer Wissenschafter, Thomas Morgan um ca. 1910 bei seinen Kreuzungsversuchen mit der Fruchtfliege Drosophila melanogaster. Er fand heraus, dass bestimmte Gene am gleichen Chromosom liegen und daher als „gekoppelt“ bezeichnet werden können. Durch bestimmte Vorgänge bei der Meiose (Keimzellenbildung) kann diese Koppelung jedoch durchbrochen werden. Durch Beobachtung der Häufigkeit des Auftretens solcher Koppelungsbrüche fand Morgan ein Werkzeug zur Erstellung der ersten Chromosomenkarten. Damit konnte er eine Aussage machen wo am jeweiligen Chromosom sich bestimmte Gene befinden müssen. 6 Die Koppelungsversuche von Thomas Hunt Morgan (1866 – 1945): (Die unterstrichenen Merkmale verhalten sich gegenüber den nicht unterstrichenen dominant!) Versuchsmaterial: Drosophila melanogaster (Gemeine Obst- oder Essigfliege) Normalform (=Wildtyp): grau, normale Flügel, normale Augen (braun) (gekoppelte Gene!) Mutante 1: schwarz, stummelflügelig(gekoppelt) Mutante 2: schwarz, rote Augen (gekoppelt) Die von T. Morgan durchgeführten Kreuzungen: 1. Normalform x Mutante1 es entstehen ausschließlich Hybriden (grau, normale Flügel) 2. Hybridenmännchen der F1 rückgekreuzt mit homozyg. rez. Weibchen es entstehen phänotypische Wildformen (heterozygot!) und Mutanten 1 im Verhältnis 1:1. → Kreuzungsschema erstellen! (Aufgrund der Koppelung können die Kombinanten Grau mit Stummelflügeln und Schwarz mit normalen Flügeln nicht entstehen!) 3. Hybridenweibchen rückgekreuzt mit homoz. rez. Männchen hier entstehen zusätzlich mit einer Häufigkeit von 18% die eigentlich „verbotenen“ Kombinationen (Grau, Stummelflügel / Schwarz Normalflügel) Schlussfolgerung: 18% der Nachkommen sind von einer intrachromosomalen Rekombination (Entkoppelung) betroffen der Abstand der betroffenen Gene am Chromosom beträgt daher nach T. Morgan daher 18 Austauscheinheiten (= Morganeinheiten!) 4. Normalform mit Mutante 2 → Hybriden (Grau mit braunen Augen) 5. Hybridenweibchen mit homozygot rez. Männchen der Mutante 2 es entstehen phänotypische Wildformen (heterozygot!), die Mutanten 2 und zusätzlich mit Häufigkeit von 6% die eigentlich „verbotenen“ Kombinanten Grau mit roten Augen und Schwarz mit braunen Augen → Kreuzungsschema erstellen! Schlussfolgerung: Der Austauschwert zwischen Körperfarbe und Augenfarbe (rot) wurde also mit 6 ermittelt, derjenige zwischen Augenfarbe und Flügellänge konnte mit 12 vorhergesagt und schließlich auch experimentell bestätigt werden! SKIZZE ! Körperfarbe -----6------Augenfarbe -----12------ Flügellänge ------------------------------------ 18 ------------------------------------ Durch diese Überlegungen ergab sich erstmals die Möglichkeit der Konstruktion von Chromosomenkarten, weil durch die Bestimmung der Austauschwerte bei diesen Kreuzungsexperimenten auf den Abstand der Gene am Chromosom geschlossen werden konnte! (Hier also: Das Gen für die Flügellänge ist vom Gen für die Körperfarbe weiter entfernt (18 Einheiten) als das Gen für die Augenfarbe (nur 6 Einheiten)) Schlussfolgerung: Nicht gekoppelte Gene folgen der freien Kombinierbarkeit, bei gekoppelten Genen ist der Austauschwert proportional zum Abstand der Gene am Chromosom (also keine beliebige Kombinierbarkeit! 7 III) Molekulargenetik III.1) Das Versuchsmaterial der molekulargenetischen Grundlagenforschung Als Versuchsmaterial dienen die einfachsten „Lebewesen“ mit genetischer Information: IV.1.1) Bakterien Bestuntersuchtes Bakterium Escherichia Coli (bildet einen Großteil der menschlichen Darmflora) In freier Natur überall dort, wo Exkremente abgebaut werden (Coli-Titer-Bestimung / Trinkwasser < 1 Keim/100 ml Wasser) Bakterienmerkmale: • Geringe Masse ( ca. 1/1000 eines tierischen Einzellers ) • Kernäquivalent ( = Nucleoid ), keine Chromosomenkondensation möglich, keine Kernhülle • Zellmembran bildet Mesosomen (Orte intensiver Zellatmung – Mitochondrien fehlen!) Skizze! • Bei photosynthet. Aktiven Bakterien Thylakoide (ebenfalls Membraneinstülpungen) als Orte der Photosynthese keine Chloroplasten! • Wichtiges Strukturelement ist die Zellwand regelmäßiges Geflecht aus KH und Proteinen Murein-Hülle Wirkungsweise von Antibiotika! • Die Mureinhülle kann Wandstrukturen ausbilden, welche eine äußerst resistente Kapsel ausbilden Antibiotikaresistenz! • Vermehrung meist durch einfache Abschnürung Konjugationsvorgänge finden statt, dies führt zu diploiden Zygoten. Die Entwicklung dieser Zygoten zu den haploiden Normalformen ist noch unerforscht! Bakterien sind haploid! • Viele Bakterien können widerstandsfähige Überdauerungsformen ausbilden Sporenbildung Bedeutung der Sterilisation in der Medizin IV.1.2) Viren Virusmerkmale: • • • Können schwer als Lebewesen bezeichnet werden, weil ihnen typische Lebenskennzeichen fehlen. Kompliziert gebaute Molekülverbände aus Proteinen und NS Nucleoproteide Fortpflanzungsmechanismus von Viren 8 III.2) Die Nucleinsäuren als Träger der Erbinformation Experimente zum Beweis der NS als Träger d. Erbinfo.: - Exp. Von Avery (1944) mit Pneumokokken und Mäusen (Nachweis der Transformation) Linder 3 Isolierte DNA von virulenten Pneumokokken vermengt mit harmlosen Pneumokokken. Es treten einige virulente Formen auf, weil sie durch Transformation die DNA der virulenten Stämme übernommen haben. - Die mutagene Wirkung von UV-Strahlen setzt bei jenen Wellenlängen ein, die im Absorptionsspektrum der DNS fehlen Erklärung! III.3) Aufbau und Vorkommen der Nucleinsäuren III.3.1) Die DNS ( = DNA ) - Übersetzung Kommt vor im Zellkern, in Chloroplasten und Mitochondrien vor Aufbau aus Zucker (Desoxiribose), Phosphatrest, organische N-hältigen Basen (A, C, G, T) Zucker + Base = Nucleosid Nucleosid + Phosphat = Nucleotid Desoxiribosestruktur: Ribosestruktur: - Gesetz der spezifischen Basenpaarung - Doppelhelixmodell der DNA von Watson und Crick (1953) Auf eine volle Windung der Helix fallen 10 Nucleotide bei einer Länge von 3,4 nm Die Holme der Leiter bilden Zucker und Phosphatbausteine, die Sprossen entstehen durch die Basenpaare 2 H-Brückenbindungen zw. Adenin und Thymin – 3 H-Brücken zw. Cytosin und Guanin. 9 III.3.2) Die RNS ( = RNA ) Ribose als Zucker - Uracil anstelle von Thymin - einsträngig - wesentlich kürzer III.4) Die Replikation der DNA Bedeutung: Verdoppelung der Erbinformation im Verlauf der Zellteilungen. III.4.1) Theoretische Überlegungen zum Verlauf • Modell der konservativen Replikation: Man stellt sich eine komplette Neusynthese der Doppelhelix vor • Modell der dispersiven Replikation: Zerbrechen des Elternstranges und Ergänzen der Bruchstücke durch Neusynthese • Modell der semikonservativen Replikation: Die beiden Helixstränge werden getrennt, anschließend erfolgt schrittweise eine Anlagerung der komplemetären Nucleotide an die beiden Stränge! III.4.2) Beweis für den semikonservativen Charakter der Repl. 10 Das MESELSON - STAHL Experiment: Zellkultur von Escherichia Coli in einem Nährmedium mit dem Stickstoffisotop N15. Nach einiger Zeit ist in alle Basenbausteine der DNS N15 eingebaut! Anschließend die Bakterien in eine Kultur mit ausschließlich normalem N14 überführen. Alle neu gebildete DNA muß ab diesem Zeitpunkt ausschließlich normalen Stickstoff beinhalten. Nach Verdoppelung der Population findet sich ausschließlich halbschwere DNA in den Bakterien, nach der nächsten Verdoppelung „halbschwere“ und „normale“ DNA im Verhältnis 1: 1! III.4.3) Der Verlauf der Repl. Helicasenwirkung Trennung der Komplementärstränge durch Lösen der H-Brücken Reißverschlussprinzip! Anlagerung von Nucleosidtriphosphaten unter Diphosphatabspaltung an komplementäre Nucleotide des Elternstranges. Verknüpfung der Nucleotide durch spezielle Enzyme DNA-Polymerasen SKIZZE! III.5) Der genetische Code Die genetische Information liegt in der Nucleotidsequenz (Basensequenz !) verschlüsselt vor! III.5.1) Die Realisierung der genetischen Information Sie erfolgt durch Enzyme (Proteine = Polypeptide), welche dann ganz bestimmte merkmalsbildende Stoffwechselprozesse des Körpers auslösen. Daher muss in der DNA ein Programm für den Aufbau verschiedenster Enzyme verschlüsselt vorliegen! Wiederholung Aufbau von Proteinen aus 20 verschiedenen Aminosäuren! Im Endeffekt entspricht also die Nucleotidsequenz der DNA der Aminosäurensequenz von benötigten Proteinen. Dabei müssen allerdings 4 verschiedene Nucleotide 20 Aminosäuren codieren! 11 III.5.2) Die Merkmale des genetischen Code • Die Basenabfolge dreier Nucleotide (Basen!) der DNA codiert eine Aminosäure! CODOGEN! Begründung: 1 Base....., 2 Basen ......, 3 Basen .... 64 Kombinationsmöglichkeiten Experimenteller Beweis: Sie gelang durch Experimente von TMV mit deren Wirtszellen (Pflanzenzellen – Tabakpflanzen!) Veränderung der Erbinfo. der Viren durch ACRIDINE (ähnliche Struktur wie Nucleotide, heften sich immer an den Anfang der DNA) • • Veränderung der Information durch einzelne Acridine Aufbau von völlig unterschiedlichem Viruseiweiß in der Wirtszelle. • Veränderung durch Acridin-Zweiergruppen gleiches Ergebnis • Veränderung durch Dreiergruppen Produktion von fast identischem Viruseiweiß ERKLÄRUNG !!! Der genetische Code ist degeneriert: Man kann von der Basensequenz auf die Aminosäuresequenz schließen, aber nicht umgekehrt! Einige Tripletts codieren bestimmte Funktionen: UAA, UAG, UGA Stop Funktion AUG Start der Verknüpfung mit Meth. • Es gibt keine überlappende Codierung! Beispiel! • Die Codogens sind nicht gegeneinander abgegrenzt, sie grenzen direkt aneinander! • Der genetische Code ist universell gültig! Ein wichtiges Argument für die Evolutionstheorie!!! III.5.3) Die Wirkungsweise der Enzyme Energiediagramm !!! Enzyme sind Wirkungs- und substratspezifisch Erklärung! 12 III.5.4) Die Biosynthese der Proteine (der Enzyme!) • • Übersichtsskizze Die Transkription Als Ergebnis liegt die Erbinfo. für ein Gen als mRNA vor (Codon) • Die Translation tRNA Moleküle ( Aufbau! ) sind die eigentlichen Übersetzer des genet. Code Anticodon • Ergänzung: Spleißen (splicing) der DNA: Introns, Exons, Reifung der DNA . . . Bei der Reifung unterscheidet man drei Hauptschritte: • • • Capping → das 5´ Ende wird mit mehreren 7-Methylguaninmolekülen besetzt (wichtig für das spätere Anlagern an das Ribosom) Polyadenylation → an das 3´ er Ende werden bis zu 200 A-Nucleotide angehängt (Funktion unbekannt!) Eigentliches splicing unter Einwirkung mehrerer Enzyme III.5.5) Genwirkungsketten • Begriffsdefinition • Beispiel: Phenylalaninstoffwechsel des Menschen Eiweiß Enz.A Phenalalanin Enz.B Tyrosin Enz.C Homogentisinsäure . . . . . Kohlendioxid, Wasser • Fehlt Enzym D Albinismus Fehlt Enzym B Stau an Phenylalanin Umbau zu Phenylketon Phenylketonurie (mit Fehlentwicklungen des Nervensystems) IV.) Geschlechtsdifferenzierung – Geschlechtsbestimmung IV.1) DieDifferenzierung des Geschlechtes BEGRIFF: Bisexuelle Potenz! IV.1.1) Phänotypische Geschlechtsdifferenzierung Davon spricht man, wenn die bisexuelle Potenz durch Außeneinflüsse in eine der beiden möglichen Richtungen realisiert wird! Beispiele: Putzerfische: Leben in Haremsverbänden (bis zu 200 Weibchen auf 1 Männchen. Stirbt das Männchen, übernimmt ein Weibchen die Funktion des Männchens. In diesem Fall bewirken psychische Faktoren das Heranreifen von Hodengewebe aus embryonalen Blastemen, die in der Gonadenanlage erhalten geblieben sind! 13 Bonellia viridis (mariner Wurm!): geschlechtlich undifferenzierte Larven entwickeln sich zu weiblichen Tieren, setzen sich die Larven jedoch am Rüsssel eines geschlechtsreifen Weibchens fest, übernehmen sie von diesen Hormone, welche innerhalb von sechs Tagen eine Differenzierung zum Männchen bewirken. Werden die Larven nach drei Tagen vom Weibchen getrennt, entwickeln sich Intersexe. Reptilienfortpflanzung: Temperaturabhängigkeit IV.1.2) Genotypische Geschlechtsdifferenzierung Dabei wird die bisexuelle Potenz durch Gene in eine Richtung entschieden Geschlechtsrealisatoren! Typisch für die meisten Tiere (Mensch!) und alle zweihäusigen Pflanzen M-Faktoren induzieren die Ausbildung des weiblichen Geschlechtes, M-Faktoren das Gegenteil. Faktoren können sich auf Heterosomen und/oder auf Autosomen befinden (Drosophila: WFaktoren auf X / M-Faktoren auf Autosomen) Mensch: W-Faktoren auf X, M-Faktoren auf Y, Autosomen geschlechtsneutral. M-Faktoren mit deutlicher Dominanz (1:3 bis 1:4!) Männer sind heterogametisch, Frauen homogametisch! Y induziert in der frühembryonalen Anlage der Geschlechtsorgane die Hodenbildung. Nach erfolgter Differenzierung bestimmen die bereits gebildeten Hormone die weitere sexuelle Ausbildung des Organismus! IV.2) Die Bestimmung des Geschlechtes Durch hormonelle Störungen kann ein Mensch äußerlich undifferenziert erscheinen, obwohl sein Geschlecht genetisch eindeutig fixiert ist phänotypischer Zwitter. Sein genotypisches Geschlecht kann durch verschiedene Methoden bestimmt werden: IV.2.1.) Karyogramm IV.2.2.) Barr-Körperchen IV.2.3.) Drumsticks 14 IV.3) Geschlechtschromosomenanomalien Norm. Mann Norm. Frau Turner Syndrom ----Triplo-X Syndrom Tetra-X Syndrom Klinefelter Synd. ----Triplo-XY Synd. Heterosomen XY XX XO YO YY XXX XXXX XXY (XXXY) XYY XXYY XXXY Barr-K. _ 1 0 0 0 2 3 1 (2) 0 1 2 Phänotypus M W W letal letal W W M W M M M Fruchtbarkeit + + - +/leicht eingeschr. +/+/- IV.4) Geschlechtschromosomengebundene Vererbung X-Chromosom W-Faktoren + andere Gene / Y Chromosomen enthält nur M-Faktoren. Daraus ergibt sich eine geschlechterspezifische Vererbbarkeit bestimmter Krankheiten: X-chromosomal-rezessive Vererbung: Z.B. Bluterkrankheit: Erbgang (Bluter, Bluterin, Konduktorin) 1:10 000 bei Männern, 1: 100 Mio. bei Frauen Rot-Grün-Sehschwäche: 8% der Männer, 0,5% der Frauen V) Mutation – Modifikation • Veränderungen in der Erbsubstanz • Bei Tieren vererbbar, wenn die Keimbahn betroffen ist, bei Pflanzen ist Vererbbarkeit immer möglich (jede Zelle kann Keimbahnzelle werden! Erklärung! 15 • Mutationen sind zufällig und richtungslos ??? • Unterscheide: Genmutationen - Chromosomenbaumut. - Chromosomenzahlmut. (=Genommut.) V.1) Genmutationen o Die häufigste Mutationsform (Mutationsrate zw. 10 –5 und 10 –9 Erklärung!) o Beispiele: Hämophilie, Albinismus, Phenylketonurie, Sichelzellenanämie (rezessive Verhaltensweise) o Sichelzellenanämie Beschreibung! Resistenz gegen Malaria Heterozygote Träger sind phänotypisch gesund, aber ebenfalls resistent Extrem hohe Häufigkeit von heterozygoten Trägern in Malariaregionen (gegen 60%) Malariazyklus V.2) Chromosomenbaumutationen Die Zahl der Chromosomen bleibt erhalten, die Form und/oder Struktur ändert sich. Chromosomen brechen auseinander, die Bruchstücke können ... • ... sich auflösen Deletion (Cri du chat Syndrom = Deletion am Chromosom Nr. 5) • ... an der Schwesterchromatide angehängt werden Duplikation • ... sich umgekehrt wieder einfügen Inversion • ... an ein nicht homologes Chromosom angehängt werden Translokation (z.B. Philadelphiachromosom = t(9/22) → Auslöser für Leukämien. 16 V.3) Chromosomenzahlmutationen V.3.1) Die Aneuploidie Einzelne Chromosomen fehlen, oder liegen mehrfach vor. Ursachen sind Fehlverteilungen bei der Meiose Monosomien, Trisomien, Tetrasomien . . . Die lebensfähigen Aneuploidien des Menschen Trisomie 21, 13, 18 ! (Andere Trisomien sind offensichtlich letal! Die Trisomie 21 (Mongolismus, Down-Syndrom) • Ursache • Krankheitsbild: Schräge Augachsen, abgeflachtes Gesicht, breite Nase, Kurzfingrigkeit, Schwachsinn, große Zunge, typischer Verlauf der Handfurchen • Zusammenhang der Häufigkeit mit dem Alter der Mutter: 0,o5% bei 20-jährigen - 1% bei 40 jährigen - 8% bei 50 jährigen Frauen Grafik: V.3.2) Die Euploidie • Gesamter Chromosomensatz liegt vermindert oder vermehrt vor: n (Haploidie!), 3n, 4n, . . • Polyploidie bei Pflanzen weit verbreitet, bei Tieren selten, beim Menschen unbekannt (10% aller vorzeitigen, natürlichen Schwangerschaftsabgänge (=Abortus) sind polypoid! V.3.2.1) Die Autopolyploidie • Der arteigene Chromosomensatz liegt vervielfacht vor. Künstliche Auslösung durch Colchizinbehandlung Verhindert den Aufbau eines funktionellen Spindelapparates diploide Keimzellen Befruchtung Tetraploider Organismus! • Vorteil: Polyploide Pflanzen haben größere Zellen und daher einen üppigeren Wuchs. • Nachteil: Polyploide Pflanzen sind häufig steril, weil bei der Meisose häufig Störungen in der Chromosomenverteilung auftreten. 17 V.3.2.2) Die Allopolyploidie • Ausgangsmaterial sind Artbastarde, die normalerweise steril sind, weil es zu keiner Synapsis kommen kann Verschiedene Chromosomen!!! Wird der Chromosomensatz eines Bastards durch Colchizin verdoppelt, so liegt Allopolyploidie vor (2 doppelte, aber verschiedene Chromosomensätze ) Solche allopolyploide Pflanzen sind in der Regel fertil, weil jedes Chromosom wieder einen Synapsispartner findet! • Verwendung: Zucht neuer Kulturpflanzen, welche die Vorteile anderer Pflanzen vereinigen und dennoch fruchtbar sind Die meisten heutigen Kulturpflanzen sind allopolyploid! • Allopolyploidie tritt vereinzelt auch natürlich auf Hauptsächlich in Pflanzen auf Extremstandorten Sie besitzen ein größeres Reservoir an verschiedenen Allelen und damit ein höheres Potential zur Anpassung gegenüber der Umwelt! V.4) Allele Man versteht darunter Gene, die an einander entsprechenden Stellen zweier Chromosomen liegen. 2 gleiche Allele Homozygotie / 2 verschiedene Allele Heterozygotie. Multiple Allele: Alle Allele, die durch Mutationen eines Gens oder bereits mehrerer Allele entstanden sind. Z. B.Die menschlichen Blutgruppen 3 Allele IA, IB, i (IA zu IB intermediär, beide gegenüber i dominant) daraus ergeben sich folgende Geno-/Phänotypen: Genotyp I AI A I Ai I BI B I Bi I AI B ii Phänotyp A homozyg. A heterozyg. B homozyg. B heterozyg. AB (immer heterozyg.) 0 (immer homozyg.) Forensischer Vaterschaftsausschluss!!! (Wiederholung!) Z.B. Kind mit 0 kann unmöglich einen Vater der Gruppe AB haben Kind mit AB kann keinen Vater der Gruppe 0 haben Kind mit Kind mit AB kann keinen Vater mit A haben, wenn die Mutter A hat. etc. 18 V.5) Ursachen für Mutationen (Mutagene Faktoren) = Mutagene Die größte Bedeutung kommt folgenden Mutagenen zu: V.5.1) Strahlung • UV- Strahlung mit Wirkungsmaximum bei 260 nm • Röntgenstrahlung ( entspricht radioakt. Gamma-Strahlung ) • Kosmische Strahlung (eine Mischstrahlung ) • Radioaktive Gesteine (hauptsächlich Gamma ) • Größten Anteil am terrestrischen Strahlungsaufkommen hat das gasförmige RADON (Rn 222) als Ergebnis der Zerfallsreihe von Uran und Thorium. Mittelwert in Bq/m3 (Kernzerfall pro Sekunde und m3) Freiluft Raumluft Trinkwasser Boden (1m Tiefe) 15 50 (extreme Schwankungen!) 5000 5000 – 500 000 Effektive Strahlendosis pro Person (mSv/Jahr) Radon Terrestrische Strahlung Kosmische Srahlung Strahlung aus Nahrungsmitteln 1,1 mSv/Jahr 0,4 0,3 0,3 Sievert = absorbierte Energiemenge in Joul pro Kilogramm Körpermasse x Wirkungsfaktor der Strahlung. V.5.2) Chemikalien • Strukturanaloge von DNA-Bausteinen (Z.B. 5-Brom-Uracil) Verbindet sich mit Guanin anstelle von Adenin . . . . . . • Nitrit (NO2) bewirkt Umsetzung von Cytosin zu Uracil • Acridine Siehe hinten! 19 V.5.3) Physikalische Einflüsse Temperaturschocks, Elektromagnetische Strahlung bestimmter Wellenlängen (Handy´s !?) etc. Modifikationen Rein phänotypische Veränderungen bei Lebewesen, welche also unter keinen Umständen vererbbar sind. Beispiele: Löwenzahn, Ertrag von Kulturpflanzen auf unterschiedlichen landwirtschaftlichen Böden, . . . ) V) Tier- und Pflanzenzucht unter genetischen Aspekten V.1) Individualauslese (Ziel ist Homozygotie!) Man wählt aus einer Pflanzenpopulation Pflanzen mit günstigen Eigenschaften aus und sät deren Samen auf isolierte Beete. Durch wiederholte Auslese kann der Effekt zunehmend verstärkt werden, Reinerbigkeit wird durch hohe Inzuchtgrade erreicht. V.2) Kombinationszüchtung (Ziel ist Homozygotie!) Es werden verschiedene Rassen oder Sorten derselben Art gekreuzt! Zielsetzung: • • Vereinigung der gewünschten Eigenschaften zweier Sorten in einer einzigen Herauszüchten neuer Eigenschaften, die bisher, weil rezessiv, nicht phänotypiusch auftraten V.3) Hybridzüchtung (Ziel ist Heterozygotie) Der Heterosiseffekt: Er tritt bei Hybriden homozygoter Eltern auf, wenn diese zwar verschiedenen, aber sehr ähnlichen Sorten (Rassen) angehören. Diese Hybriden weisen gesteigerte Leistungsfähigkeit auf. Müssen immer neu hergestellt werden Beispiel: Hybridmaissorten V.4) Mutationszüchtung Zuchtsorten werden mutagen bestrahlt, die entstandenen Mutationen nach dem Prinzip der Individualauslese weiterselektiert. Bsp. Opaque 5 Mais für Mittelamerika! V.5) Produktion transgener Pflanzen Künstliche Herstellung einer DNA mit der gewünschten Basensequenz (codiert das gewünschte Protein, z.B. mit hohem Anteil an der gewünschten essentiellen Aminosäure) → Einbringen dieser DNA in Pflanzenzellen (Genkanone, Elektroporation, Mikroinjektion, Vektoren) → Beobachten, ob eingeschleuste Gene auch exprimieren. 20 Genkanone: DNA wird auf Goldkügelchen aufgedampft, die Kügelchen wiederum auf einem größeren Projektil. Das Projektil wird gegen Pflanzengewebe gerichtet und abgefeuert. Beim Aufprall auf ein Schutzgitter bleibt das Projektil hängen, viele Goldkügelchen fliegen durch ihre Trägheit weiter, durchdringen die Zellwände und bleiben im Zytoplasma hängen. Viele Zellen sterben dabei ab, einige wenige überleben und exprimieren u.U. die Fremd-DNA. Elektroporation Durch gezielte Elektroimpulse wird die Zellmembran kurzzeitig perforiert, in dieser Zeitspanne kann Fremd-DNA aus der Umgebung aufgenommen werden. Mikroinjektion Mechanisches Einbringen von wenigen Nanolitern DNA in eine Wirtszelle. Vektoren → Trojanisches Pferd. o Agrobacterium tumefaciens A.t. befällt Pflanzenzellen und regt diese zum unkontrollierten Wachstum (Tumor) an, indem es Plasmide mit dem entsprechenden „Tumorgen“ in die Wirtszellen einschleust. Gentechniker haben dieses Tumorgen entfernt und können an dieser Stelle beliebige andere Gene einbauen. o Phagen → ... V.5) Klonung V.5.1) Klonung in der Pflanzenzucht Stecklingsvermehrung, Vermehrung über Zwiebeln, Knollen, Ausläufer etc. V.5.2) Klonung höherer Organismen Das Embryonensplitting: Die gezielte Klonung aus Körperzellen (Kerntransfer): V.6) Andere Fortpflanzungstechnologien Extrakorporale Befruchtung (In vitro Fertilisation → IVF) Leihmutterschaft Präimplantationsdiagnostik (→ PID) Multipler Embryonentransfer 21 VI) Gentechnik/Gentechnologie Bei zelltechnischen Verfahren werden Gene wenn überhaupt, dann nur im Gesamtverband, also als ganzes Genom verschoben oder kombiniert. Bei Gentechnologischen Verfahren wird in das Genom eingegriffen! → Unterscheide zwischen Grüner-, Roter- und Grauer Gentechnik! VI.2) Gentechnische Verfahrensweisen VI.2.1) Grundbegriffe Die Restriktionsenzyme (Chemische Scheren): Enzyme, welche imstande sind, die DNA an bestimmten Sequenzen zu zerschneiden. Erstmalig in Bakterien entdeckt, welche gegen bestimmte Viren immun geworden waren. Restriktionsenzyme werden heute in beliebigen Sequenzen in biochemischen Labors designed. Z.B. Schnittstelle des Restriktionsenzyms Mnl1 lautet 5´ CCTC(N)7 3´ → . . . . . .. Die Gelelektrophorese: Die einzelnen DNA-Fragmente liegen in Ionenform vor, bewegen sich also im elektrischen Gleichstromfeld. Die Strömungsgeschwindigkeit wird durch die Molekülgröße und die Dichte des Mediums, in welchem die Elekrophorese stattfindet, bestimmt. Man verwendet ein definiertes, erstarrtes Gel als Flussmittel und gibt die aufzutrennenden Lösungen am Startpunkt dazu elektrisches Feld kurze Abschnitte wandern schneller als lange nach einer bestimmten Zeit (ca. 1 Stunde) lassen sich im Gel bereits die aufgetrennten Flußbande erkennen. Nachweis von Mutationen durch Restriktionsfragmentlängen Übung: Bekannte, weil häufig auftretende Mutation auf einem Exon zum Insulinaufbau (Entspräche einer genetisch bedingten Diabetes mellitus): Sequenz eines Exons (Großbuchstaben) mit kurzem Abschnitt des benachbarten Introns (Kleinbuchstaben) zum Insulinaufbau in der Normalform und bei Vorliegen einer Mutation von G nach A im 6. Triplett. 5´ AAC GGT TGA ACC CCC CAG TTA GTT TGG AAT AAG AAT AAC CCC AAG A (=Mutation) AAA GAT TAA GAT TAG TAG GCA CAT CAA ATC TTT TGG ACA GAA TTC CGA AGG GTT ATA ATG GGG GAT gta agg gtt aga aaa aaa gaa tgg gcg ccc tgg 3´ Die Schnittstelle des Restriktionsenzyms lautet 5´ GTTA(N)3 3´ 22 Hinweis: Die DNA ist eine Doppelhelix !!! Fragestellung: Wie groß sind die Produkte (in bp = Basenpaare), die beim Schneiden mit dem oben genannten RE aus dem PCR – Produkt entstehen, wenn: 1. ein Wildtyp vorliegt (Normalform): 2. die Mutation homozygot vorliegt: 3. die Mutation heterozygot vorliegt: Skizziere das Ergebnis einer durchgeführten Gelelektrophorese, bei der ein DNALängenmarker verwendet wird der Kontroll-DNA mit 5, 10, 15 ... bis 100 bp enthält: DNA – fingerprinting Introns besitzen eine bestimmte Reihenfolge von Basen (entspricht einer Sequenz), die sich mehrmals wiederholen kann. Es gibt hunderte solcher DNA-Abschnitte, die sich nach Längeneinheiten in Minisatelliten und Mikrosatelliten (= STR´s) einteilen lassen. Heutzutage werden in der Kriminaltechnik nur noch STR´s verwendet, die so genannten Short Tandem Repeats. Wie der Name schon sagt, bestehen sie aus kurzen, ca. 2-4 Basen langen und sich immer wiederholenden DNA-Abschnitten. Von mehr als 100 repetetiven DNAAbschnitten hat man sich in der Kriminalistik mittlerweile auf ca. 30-40 mit 120 bis 350 bp geeignet, die in der Population sehr variabel sind. Diese Genorte, auch Loci genannt, nennt man VNTR-Loci (Variable Numbers of Tandem Repeats). Mit Hilfe der PCR werden diese VNTR-Loci vielfach kopiert und durch ein Gel aufgetrennt. Beispiel aus LINDER Teil 3: 23 Der Southern Blot (nach Edwin Southern) Anwendungsbeispiele: a) Nachweis einer bekannten Nucleotidsequenz (eines Gens) in einer Probe (Liefert Antwort auf die Frage: Ist das Gen vorhanden, oder nicht? → Auch als AIDS Test anwendbar): b) DNA-Probe mit Restriktionsenzym schneiden (mehrere Schnittstellen) DNA-Längendiagnose durch Gelektrophorese Übertragung durch Diffusion auf Membranfolien (= Blotten) Hybridisierung mit einem zum gesuchten Gen komplementären DNA-Einzelstrang der radioaktiv markiert ist Auflegen auf Röntgenfilm Ergebnis: ist der gesuchte DNA-Abschnitt vorhanden, wurde er hybridisiert und taucht am Röntgenfilm als schwarzer Balken (=Bande) auf. Nachweis der Sichelzellenanämie im Southern Blot Durch die Substitution von A T wird die Erkennungssequenz des Restriktionsenzyms Mst II (CCTGAGG) zerstört. Das durch PCR klonierte Hämoglobingen wird mit Mst II behandelt und anschließend im Southern Blot analysiert. Zur Hybridisierung verwendet man einen DNA-Einzelstrang, der zum längeren Abschnitt der geschnittenen DNA komplementär ist: Ergebnis: Bei gesunden Menschen zeigt sich eine Bande in der Größe von 1,15 kb. Mischerbige zeigen eine zusätzliche Bande von 1,35 kb, reinerbig Kranke nur diese 1,35 kb-Bande. Sticky ends: Durch bestimmte Restriktionsenzyme entsteht nicht ein glatter Schnitt durch die DNA. Es bleibt einer der beiden Stränge um beliebig viele Nucleotide länger als der andere Skizze: Anwendung: Genklonung in vivo Beschreibung! Großer Erfolg: Produktion von Insulin durch gentechnisch veränderte Bakterien. 24 Die Polymerase-Kettenreaktion PCR (Genklonung in vitro) Sie dient dazu, DNA oder DNA-Abschnitte die nur in Spuren vorliegen (z.B. aus einer einzigen Zelle!) beliebig zu vervielfachen und damit andere gentechnische Arbeitsschritte zu ermöglichen. Die Primer: Man versteht darunter relativ kurze DNA-Sequenzen, welche als Ansatzstelle der DNA Poymerase fungieren. Die Replikation der DNA geht immer von diesen Primern aus. Auch Primer können in beliebiger Sequenz designed werden. Vektoren: Vektoren sind Träger unterschiedliche Komponenten, die dazu dienen, meist künstlich hergestellte DNA-Sequenzen (Gene!) in eine Zelle einzuschleusen (Trojanische Pferde). Vektoren sollen immer marker beinhalten. Diese ermöglichen es festzustellen, ob ein Vektor erfolgreich eingesetzt wurde, oder ob nicht: • Plasmide: Siehe hinten. Als marker werden meist Resistenzgene gegen Antibiotika eingeschleust. • Viren Meist Bakteriophagen. Die Viren müssen zuerst so umgestaltet werden, daß negative Auswirkungen unmöglich werden (Humanmedizin!) 1) Extraktion und Reinigung der DNA: Die Zelle mit enthaltener DNA wird enzymatisch aufgelöst (Lipasen, Proteinasen, Carbohydrasen), eventuelle Gewebsreste durch Zentrifugation entfernt. 2) Das DNA-Extrakt wird vermengt mit: • Zwei speziellen, auf die geschnittenen DNA-Stränge designten primern (extrem konservative Bereiche!) • Hitzestabiler DNA-Polymerase (entdeckt in Bakterien aus Vulkanschloten und unterozeanischen, heißen Quellen). • Gemenge aus DNA-Nucleotiden • Wasser 3) Denaturierung der DNA durch Erhitzen auf 90°C Parallelstränge trennen sich Abkühlen Primer lagern sich an kompementäre Bereiche Polymerase beginnt angelagerte Nucleotide zu verbinden es entstehen zwei komplementäre Stränge DNA ist verdoppelt . . . . . . . . . . . . . . . . . . . . . . . . Übungsbeispiel !!! 25 Die DNA-Analyse (Sequenzierung) Ziel ist die Erforschung einer genauen Basensequenz eines durch PCR vervielfältigten DNABereiches. Man benötigt dazu zusätzlich zur normalen PCR-Ausrüstung Didesoxinucleotide, welche eine Polymerisierung der Nucleotide unterbrechen. Diese Didesoxinucleotide sind zusätzlich mit 4 unterschiedlichen Farbstoffen markiert (für die 4 Basen A, T, C, G) Während der PCR entstehen unterschiedlich lange DNA-Einzelstränge mit einem Endnucleotid T oder A oder C oder G. Diese sind unterschiedlich markiert! Auftrennung dieser Lösung mit Elektrophorese ! Erklärung! 26 27