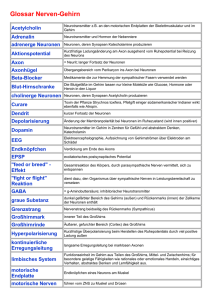
Neurobiologie
Werbung

Inoffizielles Skriptum zu Neurobiologie Univ.­Prof. Dr. Werner Sieghart (Department für Pharmazeutische Technologie und Biopharmazie) Vorlesung, 1 Semesterwochenstunde SS 2009, 300322 Di. 17:15­18:10 im gr. Seminarraum 1.Stock, Zentrum für Hirnforschung, Spitalgasse 4, 1090 Wien Geschrieben von Robert Aufreiter 1 / 77 Neurobiologie Inhaltsverzeichnis Literatur......................................................................................................................... .......................4 Dieses Skriptum............................................................................................................... ....................4 Einführung........................................................................................................................................... ......5 Aufbau des Nervensystems................................................................................................................. ......5 Zentrales Nervensystem (ZNS, Gehirn)....................................................................................... .......5 Peripheres Nervensystem (PNS).............................................................................. ...........................7 Somatisches Nervensystem...................................................................................................... .......7 Sensorisches Nervensystem..................................................................................... ..................7 Motorisches Nervensystem........................................................................................ ................8 Vegetatives (autonomes) Nervensystem................................................................................... ......8 Bildgebende Verfahren zur Untersuchung der Struktur und Funktion des Gehirns..............................8 Röntgenuntersuchungen............................................................................................................... ........9 Computertomographie (CT)............................................................................................... .................9 Neurochirurgie............................................................................................................................... .....10 Positron Emission Tomographie (PET)............................................................... .............................10 Single Photon Emission Computertomographie (SPECT)..............................................................11 Kernspinresonanztomographie (NMR­Imaging, MRI)....................................................................12 Funktiononelle magnetische Resonanztomographie (fMRI)...........................................................14 Magnetische Resonanz­Spektroskopie (NMR)............................................................................. ....14 Elektroenzephalographie (EEG).................................................................................................. ......15 Magnetenzephalographie (MEG).................................................................................. ....................16 Zellen des Nervensystems.................................................................................................................. .....17 Glia Zellen.................................................................................................................... ......................17 Oligodendrocyten und Schwann­Zellen......................................................................... ..............18 Astrocyten......................................................................................................................... .............18 Microglia.................................................................................................................................. ......18 Neuronen (Nerven Zellen).......................................................................................... .......................18 Axon....................................................................................................................................... ........20 Entwicklung des Gehirns......................................................................................................... ..........22 Elektrische Eigenschaften, Ionenkanäle....................................................................... .........................23 Ruhepotential........................................................................................................ .............................23 Aktionspotential................................................................................................................ .................27 Synapsen........................................................................................................................... ..................31 Synaptische Transmission........................................................................................... ...........................32 Chemische Transmission................................................................................................ ...................32 Präsynaptisch....................................................................................................................... ..........32 Postsynaptisch....................................................................................................... ........................34 Transmitter­aktivierte Ionenkanäle.................................................................................... ......34 G­Proteine......................................................................................................................... ........35 Elektrische Transmission........................................................................................................ ...........36 Transmittersysteme............................................................................................................ .....................36 Acetylcholin­System......................................................................................................... .................40 Noradrenalin­System............................................................................................ .............................43 Dopamin­System....................................................................................................................... .........45 Serotonin­System............................................................................................................. ..................45 GABA­System........................................................................................................................... .........46 Glutamat­System........................................................................................................................ ........49 2 / 77 Neurobiologie Peptid­Systeme......................................................................................................................... ..........50 Funktion der Sinne.................................................................................................................. ................51 Mechanorezeptoren.............................................................................................................. ..............52 Thermorezeptoren............................................................................................................ ..................53 Chemorezeptoren....................................................................................................... ........................53 Geschmacksrezeptoren............................................................................................................... ...53 Geruchsrezeptoren..................................................................................................................... ....55 Lichtrezeptoren................................................................................................................................ ...56 Reizweiterleitung und ­verarbeitung................................................................................ .................56 Funktion des motorischen Systems............................................................................................ ............57 Arbeitsmuskulatur.................................................................................................................... ..........58 Sensoren des Muskels.................................................................................................... ....................61 Muskelspindel.......................................................................................................................... ......61 Golgi Sehnenorgan.............................................................................................................. ..........62 Rezeptoren in den Gelenken........................................................................................... ..............62 Reziproke Hemmung.................................................................................................. .......................62 Integrative Hirnfunktionen.................................................................................................. ...................63 Bewegung......................................................................................................................... ..................64 Sprache.................................................................................................................... ...........................66 Spezialisierung von linkem und rechtem parietalen Assoziationscortex........................................67 Synaptische Plastizität und Lernen................................................................................... .....................68 Lernen..................................................................................................................................... ............70 LTP und LTD........................................................................................................................ .........71 Gedächtnis, Emotionen................................................................................................................ ...........73 Gedächtnis.................................................................................................................... ......................73 Emotionen..................................................................................................................................... ......74 Furcht und Angst......................................................................................................... ..................75 Motivation.................................................................................................................................... ..76 3 / 77 Neurobiologie Information Literatur Mark F. Bear, Barry W. Connors, Michael A. Paradiso "Neuroscience, Exploring the Brain", 3rd edition, Lippincott Williams & Wilkins Folien zur Vorlesung: ftp://NeuroBio:[email protected]/ Passwort wird in der Vorlesung bekannt gegeben Dieses Skriptum Dieses Skriptum beruht auf den Folien von Prof. Sieghart, meiner Mitschrift, Mitschriften anderer Studenten SS2009 (ich war nicht in jeder Vorlesung) und einigen Büchern (Bear, Connors, Paradiso "Neuroscience, Exploring the Brain"; ...). Ich habe diese Reinschrift erstellt, um den Stoff der Vorlesung zu wiederholen. Ich habe mich natürlich bemüht, alles richtig und vollständig zusammenzuschreiben. Es kann aber durchaus sein, daß etwas fehlt, mehr ist als zum Test kommt oder Fehler enthalten sind. Jeder der dieses Skriptum verwendet, tut dies auf eigene Gefahr. Version 2009­10­08 4 / 77 Neurobiologie Einführung Das Gehirn (mit Genen und Erfahrung) formt unsere Persönlichkeit. Es steuert lebensnotwendige Funktionen (z.B.: Atmung, Hunger und Durst, Wachen und Schlafen, Ruhe und Bewegung), verarbeitet Sinneseindrücke, erzeugt Gedanken / Emotionen / Verhalten und ermöglicht Lernen / Gedächtnis. Erkrankungen des Gehirns (werden in dieser Vorlesung nicht behandelt): • psychiatrische Erkrankungen (z.B.: Depression, Schizophrenie, Angst) • neurologische Erkrankungen (z.B.: Schlaganfall, Multiple Sklerose, Alzheimer, Parkinson, Schmerzerkrankungen) Die Ziele der Hirnforschung sind: • die Funktion des Gehirns zu verstehen • die Ursachen von Erkrankungen zu erkennen • neue Therapien zu entwickeln Wissenschaftsdisziplinen, die dazu beitragen das Gehirn zu verstehen: • Neuroanatomie (welche Teile des Gehirns gibt es, wie interagieren sie, ...) • Neurohistologie • Neurophysiologie • Biochemie (die Zellen des Gehirns unterscheiden sich in ihrer Biochemie von anderen Zellen des Körpers) • Molekularbiologie • Zellbiologie • Pharmakologie • Verhaltensforschung (am Tier und am Menschen) • Psychologie • Psychiatrie (Fehlverhalten, Erkrankungen) Aufbau des Nervensystems Das Nervensystem wird in Zentrales Nervensystem (ZNS) (E: central nervous system (CNS)) und Peripheres Nervensystem (PNS) (peripheral nervous system (PNS)) unterteilt. Zentrales Nervensystem (ZNS, Gehirn) Das zentrale Nervensystem besteht aus Gehirn (Encephalon) (E: brain) und Rückenmark (Medulla spinalis) (E: spinal cord). Das Gehirn besteht aus 3 Teilen: • Cerebrum (Großhirn, Telencephalon) (E: cerebrum): der größte Teil des Gehirns; liegt anterior1 und superior (d.h. vorne und oben); ist in 2 Hemisphären geteilt • Cerebellum (Kleinhirn) (E: cerebellum): liegt posterior (hinter dem Cerebrum); hat auch 2 Hemisphären • Hirnstamm (Truncus encephali) (E: brain stem): der übrige Teil des Gehirns (alles außer Cerebrum und Cerebellum); ist die Verbindung zwischen Cerebrum/Cerebellum und Rückenmark Das Cerebrum wird äußerlich in 6 Lappen (Lobi cerebri) unterteilt: • Lobus frontalis (Frontallappen, Stirnlappen) (E: frontal lobe): anteriorer Teil • Lobus parietalis (Parietallappen, Scheitellappen) (E: parietal lobe): zwischen Lobus frontalis und Lobus occipitalis 1 anterior = vorne, posterior = hinten, superior = oben, inferior = unten 5 / 77 Neurobiologie • Lobus occipitalis (Okzipitallappen) (E: occipital lobe): posteriorer Teil Lobus temporalis (Temporallappen, Schläfenlappen) (E: temporal lobe): inferior, unter/zwischen Lobus frontalis, Lobus parietalis und Lobus occipitalis • • Lobus insularis (Insel) Lobus limbicus • Die Oberfläche des Cerebrums weist viele Falten auf. Die Furchen werden als Sulci2 (E: sulci; wenn besonders tief: fissures) und die Erhebungen (Windungen) als Gyri3 (E: gyri) bezeichnet. Der Sulcus centralis (E: central sulcus) trennt Lobus frontalis und Lobus parietalis. Gyrus precentralis (E: precentral gyrus) ist der Gyrus direkt anterior und Gyrus postcentralis (E: postcentral gyrus) der Gyrus direkt posterior vom Sylcus centralis. Der Sulcus lateralis (E: lateral (Sylvian) fissure) trennt den Lobus temporalis von Lobus frontalis und Lobus parietalis. Anterior an der unteren Seite des Cerebrums ist der Bulbus olfactorius (E: olfactory bulb) (es gibt 2: an jeder Hemisphäre einen). Nerven aus der Nase (Geruchsinn) enden im Bulbus olfactorius. Der Hirnstamm besteht aus: • • • • (Diencephalon (Zwischenhirn): besteht hauptsächlich aus Thalamus und weiters aus Hypothalamus, ...) Mesencephalon (Mittelhirn) (E: midbrain): verbindet Pons und Cerebellum mit dem Cerebrum Pons (Brücke) (E: pons): zwischen Mesencephalon und Medulla oblongata Medulla oblongata (Myelencephalon) (E: medulla): zwischen Pons und Rückenmark Die Furche, die die beiden Hemisphären des Cerebrums trennt (Fissura longitudinalis cerebri) geht tief hinein bis zum Corpus callosum ((Hirn­)Balken). Das Corpus callosum ist die Hauptverbindung (fast die einzige Verbindung) zwischen den beiden Gehirnhälften. Wenn es durchtrennt wird, weiß die eine Hälfte nicht mehr was die andere tut. Nach außen (nach superior, aber immer noch in der Fissura longitudinalis cerebri) ist dem Corpus callosum der Gyrus cinguli (E: cingulate gyrus) benachbart. Corpus callosum und Gyrus cinguli sind Teile des Cerebrums. Der Fornix (E: fornix) verbindet in beiden Gehirnhälften den Hippocampus4 mit dem Hypothalamus und stellt auch eine (geringe) Verbindung zwischen den Gehirnhälften dar. Auf der dorsalen Seite des Mesencephalons sind 4 Hügel (auf jeder Seite ein oberer und ein unterer). Der obere Hügel heißt Colliculus superior (E: superior colliculus) und empfängt Informationen von von den Augen. Der untere heißt Colliculus inferior (E: inferior colliculus) und gehört zum Gehör­System. Das Chiasma opticum (E: optic chiasm) ist die Kreuzung der Sehnerven. Am Beginn des Hirnstamms befindet sich das Diencephalon (Zwischenhirn), das aus Thalamus (E: thalamus) und Hypothalamus (E: hypothalamus) besteht. Am Hypothalamus befindet sich die Hypophyse (E: pituitary). Die Nuclei mammillaris (E: mamillary bodies) des Hypothalamus empfangen Information vom Fornix und tragen zur Regulation des Gedächtnisses bei. Der Uncus5 (E: uncus) ist das vordere Ende des Gyrus parahippocampalis, befindet sich an der inferioren Seite des Gehirns und gehört zum Cerebrum. 2 3 4 5 Einzahl: Sulcus Einzahl: Gyrus Der Hippocampus gehört zum Lobus temporalis. Eigentlich: Uncus gyri parahippocampalis; (uncus (lat.) = Haken) 6 / 77 Neurobiologie Die älteste Methode festzustellen, wofür eine Gehirnregion zuständig ist, ist bei jemandem mit einem Fehlverhalten nach seinem Tod durch eine Obduktion herauszufinden, welcher Teil des Gehirn beschädigt ist. Ein Beispiel dafür ist das Gehirn auf Folie 1.12. Diese Person konnte nicht sprechen, weil das brocasche Areal geschädigt war. (Das Sprachverständnis war nicht beeinträchtigt.) Das motorische Zentrum (Bewegungszentrum) befindet sich am Gyrus precentralis. Das Sehzentrum befindet sich im Lobus occipitalis. Das Sprachzentrum I ist für das Verstehen von Sprache zuständig. Das Sprachzentrum II ist für das Sprechen zuständig. Eine weitere Methode um die Funktion von Gehirnregionen festzustellen sind neurochirurgische Experimente. Läsionen konnten oft nicht genau lokalisiert werden. Es wurden daher unter Lokalanästhesie Elektroden in der Nähe der Läsion angebracht um zu sehen wo das Gehirn noch funktioniert und wo nicht. Z.B.: • Bei Stimulation im Lobus occipitalis sieht der Patient Licht. • Stimulation im Gyrus precentralis bewirkt Zukungen im zugehörigen Körperteil. Der Gyrus postcentralis ist der sensorische Cortex6. Die somatotopische Hirnrindenkarte (E: somatotopic map), die den Körperregionen Regionen auf dem Gyrus postcentralis zuordnet, wird auch als sensorischer "Homunculus" bezeichnet (siehe Folie 1.14). Wenn eine Stelle am sensorischen Cortex stimuliert wird, hat der Patient das Gefühl als würde die entsprechende Region am Körper berührt. Regionen mit viel Empfindungen haben auch große Bereiche im sensorischen Cortex. Peripheres Nervensystem (PNS) Das periphere Nervensystem wird in das somatische Nervensystem und das vegetative (autonome) Nervensystem unterteilt. Somatisches Nervensystem Das somatische Nervensystem ist der Teil des Nervensystems, der Haut, Gelenke und Skelettmuskeln innerviert. (Anteil des Nervensystems, der die willkürlichen Funktionen regelt.) Das somatische Nervensystem wird in das sensorische Nervensystem und das motorische Nervensystem unterteilt. Sensorisches Nervensystem Primär gibt es 6 Sinne: Tastsinn, Geruchssinn, Geschmackssinn, Gleichgewichtssinn, Sehsinn, Gehörsinn. Es gibt aber noch viele weitere Sinne (z.B.: Sinne, die wahrnehmen, wie die Gliedmaßen relativ zueinander positioniert sind, ...). Bsp.: Eine Druckempfindung am Finger geht von den Sensoren über die primäre afferente7 Faser (primäres sensorisches Neuron) ins Rückenmark. Der Nerv tritt über das Hinterhorn8 ins 6 7 8 Cortex ist eine Ansammlung von Neuronen, die eine dünne Schicht bilden (üblicherweise auf der Oberfläche des Gehirns); cortex (lat.) = Rinde, Schale afferente Nerven sind Nerven, die von der Peripherie zum ZNS führen (sensorisch) (lat. afferens = zuführend) (Gegenteil: efferente Neuronen (führen vom ZNS in die Peripherie (motorisch) (lat. efferens= herausführend))) dorsaler Teil des Rückenmarks 7 / 77 Neurobiologie Rückenmark ein (verläuft dann im Hinterstrang (Funiculus dorsalis)) und endet erst im Hirnstamm. Von dort wird das Signal von einem zweiten Nerv (der im Lemniscus medialis verläuft) bis in den Thalamus weitergeleitet und von dort über einen dritten Nerv in den Gyrus postcentralis. (Bis zum Gyrus postcentralis erfolgen nur 3 Übertragungen: Sensor­>1.Nerv (im Finger), 1.Nerv­>2.Nerv (im Hirnstamm), 2.Nerv­>3.Nerv (im Thalamus).) Motorisches Nervensystem Das motorische Nervensystem (Exekutivsystem) ist absolut notwendig um mit der Umwelt Kontakt aufzunehmen. Kniesehnenreflex (E: knee­jerk reflex) (Bsp für ein monosynaptisches Reflexsystem): Wenn unterhalb der Kniescheibe (Patella) mit einem Gummihammer auf die Kniesehne (Sehne des Musculus quadriceps femoris) geschlagen wird, wird der Musculus quadriceps femoris kurz gedehnt. Afferente Neuronen übertragen diese Information ins Rückenmark (und weiter ins Gehirn). Im Rückenmark sind diese Neuronen mit Motorneuronen verbunden und bewirken über diese eine Kontraktion des Musculus quadriceps femoris. Gleichzeitig hemmen sie die Motorneuronen des antagonistischen Muskels. D.h. die Kontraktion des Muskels wird im Rückenmark ausgelöst (noch bevor die Information über die Dehnung des Muskels im Gehirn ist). (siehe Folie 1.19) Motorische Neuronen gehen über das Vorderhorn (ventral) aus dem Rückenmark. Sensorische Neuronen gehen über das Hinterhorn (dorsal) ins Rückenmark. Die Reaktion im Gehirn hängt von der Situation ab. Es ist nahezu das ganze Gehirn beteiligt, bevor ein Signal vom Gyrus precentralis (motorischer Cortex) zu den Muskeln geschickt wird. Funktionen des Gehirns: • Registrierung der Information aus der Umgebung (über die Sinne) • Vergleich dieser Information mit bisherigen Erfahrungen • Beurteilung (rational und emotional) • Initiieren von adequatem Verhalten über das motorische System • Lernen aus der Situation (Erfahrung, emotionales Gedächtnis): hilft um in Zukunft besser zu unterscheiden, was gut ist und was nicht Vegetatives (autonomes) Nervensystem Das vegetative Nervensystem innerviert die inneren Organe, das kardiovaskuläre9 System und Drüsen. (Anteil des Nervensystems, der die primär nicht dem Willen und dem Bewußtsein untergeordneten Funktionen regelt.) Sympathicus ist der Teil des vegetativen Nervensystems, der für Kampf und Flucht (Aktivität, Stress) bereit macht (steigert Blutdruck, weitet die Lungen, sorgt für ausreichend Glucose im Blut, ...). Parasympathicus ist der Teil des vegetativen Nervensystems, der für Ruhe, Ausrasten, Relaxation bereit macht. (aktiviert Ernährung, Verdauung, Metabolismus, Entspannung) (siehe Folie 1.22) Bildgebende Verfahren zur Untersuchung der Struktur und Funktion des Gehirns Die Bildgebenden Verfahren sind: • Röntgenuntersuchungen • Computertomographie (CT) 9 kardiovaskulär = Herz und Gefäße betreffend 8 / 77 Neurobiologie • • • • • • • Kernspinresonanztomographie (NMR10­Imaging, MRI11) Positron Emission Tomographie (PET) Single Photon Emission Computertomographie (SPECT) Funktiononelle magnetische Resonanztomographie (fMRI12) Magnetische Resonanz­Spektroskopie (NMR) Elektroenzephalographie (EEG) Magnetenzephalographie (MEG) Die älteste Methode um Funktionen des Gehirns zu untersuchen waren Untersuchungen von Personen (mit Verletzungen oder Veränderungen des Gehirns) nach deren Tod (post mortem Untersuchungen). Ein wesentliches Problem dabei ist, daß es viele Kompensationsfunktionen im Gehirn gibt. D.h. die Symptome des Patienten entsprachen nicht unbedingt der Schädigung des Gehirns, sondern waren (zumindest nach einiger Zeit) durch Kompensationsfunktionen überdeckt (z.B.: abgeschwächt). Mit Röntgenuntersuchungen oder Computertomographie können diese Verletzungen auch am lebenden Patienten untersucht werden. Röntgenuntersuchungen Röntgenstrahung ist elektromagnetische Strahlung mit einer Wellenlänge zwischen ca 10nm und 0.01nm. Röntgenstrahlen werden erzeugt, indem Elektronen beschleunigt werden (durch eine hohe Spannung (ca 20 000 ­ 150 000 V) zwischen einer Kathode und einer Anode) und dann auf ein Metall auftreffen. Dabei schlagen sie aus den Atomen des Metalls Elektronen (aus niedrigen Orbitalen) heraus. Elektronen, die dann in diese Orbitale zurückfallen, geben dabei Röntgenstrahung ab. Röntgenstrahlen werden umso stärker abgeschwächt je höher die Ordnungszahl des Atom, auf das sie Treffen, ist. (Z.B. schwächt Ca (Ordungszahl 20) in Knochen die Röntgenstrahlen viel stärker ab, als H (1), C (6), O (8) im übrigen Gewebe, weshalb Knochen einen deutlich stärkeren Schatten am Röntgenbild erzeugen.) Die Röntgenstrahlen können auf einer photografischen Platte oder einem Schirm (oder mit Sensoren, die das Bild an einen Computer weitergeben) aufgezeichnet (bzw. sichtbar gemacht) werden. (Die Röntgenröhre, die die Röntgenstrahlung erzeugt, befindet sich dabei auf einer Seite des Patienten und der Schrim (die photographische Platte, ...) befindet sich auf der anderen Seite und macht die Röntgenstrahen sichtbar, die durch den Patienten durchgegangen sind.) Computertomographie (CT) Bei der Computertomographie13 befindet sich eine Röngenstrahlenquelle auf einer Seite der zu untersuchenden Person und ein Röntgenstrahlendetektor auf der anderen Seite. Beide rotieren um den Körper, wobei sie immer gegenüber bleiben, sodaß der Detektor die Röngtenstrahlen registriert, die durch den Körper durchgegangen sind. Auf diese Art werden Röntgenbilder aus jeder Position auf einem Kreis um den Körper gemacht. (Jedes einzelne dieser Bilder enthält, wie bei einem normalen Röntgenbild, nur den Schatten aller Substanzen, die sich zwischen Röntgenquelle und Detektor befinden (d.h. die Menge an Röntgenstrahlen, die durch den Körper durchgegangen ist).) Aus diesen Bildern wird dann von einem Computer ein Bild eines Schnittes durch den Körper (in der Ebene des Kreises) berechnet. Dies wird für viele Kreise entlang des Körpers wiederholt, sodaß (wenn die Kreise in ausreichend geringem Abstand sind) praktisch ein 3­dimensionales Bild 10 11 12 13 NMR = nuclear magnetic resonance MRI = magnetic resonance imaging fMRI = functional MRI tómos (gr.) = Schnitt; graphía (gr.) = schreiben 9 / 77 Neurobiologie entsteht. (Bei anderen Tomographie­Methoden (z.B.: Kernspinresonanztomographie) ist die Berechnung des Schnittbildes aus den Meßdaten ähnlich (für jede Tomographie wird ein leistungsstarker Computer benötigt), aber die Art der Messung unterschiedlich, sodaß die Bezeichnung Röntgen­Computertomographie besser wäre.) Vorteile: • in der Nähe von Knochen eventuell besser als moderne Verfahren Nachteile: • komplexe Mathematik • Strahlenbelastung Neurochirurgie Unter Lokalanästhesie werden mit Elektroden leichte Stimuli im Gehirn erzeugt. Der Patient hat dann den Eindruck, daß z.B. ein Finger berührt wurde. Auf diese Weise wurde der Homunculus am sensorischen und motorischen Cortex gefunden. Positron Emission Tomographie (PET) Bei der PET wird dem Patienten eine Lösung mit einem Molekül, das ein Radionuklid14, das bei seinem Zerfall ein Positron15 freisetzt, enthält, injiziert. Die freigesetzten Positronen treffen nach kurzer Wegstrecke auf ein Elektron und werden mit diesem in 2 γ­Quanten umgewandelt, die im Winkel von 180° (d.h. genau in entgegengesetzter Richtung zueinander) abgestrahlt werden. Diese γ­Quanten werden detektiert. Aus den Positionen der beiden Detektoren, die 2 zusammengehörende γ­Quanten registriert haben, ergibt sich die Gerade, auf der sich Elektron und Positron befunden haben. Aus den Daten vieler detektierter γ­Quanten kann dann wie bei der Computertomographie ein Schnittbild berechnet werden. (Im Gegensatz zur Computertomographie wird hier nicht der Schatten einer externen Strahlung, sondern eine im Körper erzeugte Strahlung gemessen.) Es kann auch aus vielen Schnittbildern (oder durch gleichzeitiges Messen in vielen Ebenen) ein 3­dimensionales Bild erstellt werden. Weil die verwendeten Radionuklide eine geringe Halbwertszeit (HWZ) haben (z.B.: 15O (2 Minuten HWZ), 13N (10 Minuten HWZ), 11C (20 Minuten HWZ), 18F (110 Minuten HWZ)), müssen sie kurz vor der Verwendung hergestellt und in das zu verwendende Molekül eingbaut werden. D.h. das Zyklotron zur Herstellung des Radionuklids und das Labor zum Einbau des Radionuklids in ein Molekül müssen in der Nähe des PET­Geräts sein (z.B.: im selben Gebäude) (und somit kommen zu den Kosten für das PET­Gerät auch noch die Kosten für das Zyklotron, ...). Da 18F eine relativ lange Halbwertszeit hat, kann es auch ein Stück trasportiert werden (kann also von einer anderen Firma gekauft werden) und wird daher häufig für PET verwendet. Aufgrund der geringen Halbwertszeit kann die Intensität der Strahlung auch während des Messens merklich abnehmen. Weiters können die γ­Quanten vom Gewebe absorbiert werden, sodaß bei gleicher Konzentration des Radionuklids von einem tiefliegenden Organ weniger Strahlung detektiert wird, als von einem oberflächlich liegenden. Die Detektoren registrieren auch kosmische Strahlung. Diese kann aber herausgefiltert werden, weil sie nur einen Detektor auf einmal trifft, während die 2 γ­Quanten aus dem Körper immer 2 gegenüberliegende Detektoren gleichzeitg aktivieren. 14 Radionuklid = Atom, dessen Kern radioaktiv zerfällt 15 Positron (=Antielektron) ist das Antiteilchen des Elektrons. Es hat praktisch gleiche Eigenschaften (z.B.: Masse) wie das Elektron, aber genau entgegengesetzte Ladung (d.h. es ist positiv geladen). Wenn 1 Elektron mit einem Positron zusammenkommt, werden beide vollständig in Energie umgewandelt, die in Form von elektromagnetischer Stahlung (γ­Quanten (=Photonen mit Wellenlänge im Bereich der y­Stahlung)) abgegeben wird. 10 / 77 Neurobiologie Es kann jedes Molekül verwendet werden, in das ein Radionuklid eingebaut werden kann (und das der Körper verträgt). Häufig wird FDG (18F­Fluordesoxyglucose) verwendet, das wie Glucose aufgenommen wird. Es können auch Moleküle verwendet werden, die an bestimmte Rezeptoren binden, ... . PET kann auch verwendet werden um Blutgefäße darzustellen und um Blutfluß, Rezeptorverteilung und Neuropharmakologie zu untersuchen. FDG wird wie Glucose phosphoryliert. Das resultierende FDG­6­Phosphat wird aber nicht weiter umgesetzt und reichert sich daher in der Zelle an. Dem Patienten wird eine Lösung mit FDG gegeben, dann wird einige Zeit gewartet, sodaß es sich in den stark Glucose verbrauchenden Organen anreichert, und dann wird die PET­Messung durchgeführt. Auf diese Weise können Gehirnareale gefunden werden, die aufgrund starker Aktivität viel Glucose verbrauchen. (Dazu bekommt der Patient eine bestimmte Aufgabe (z.: ein Rechenbeispiel) und dann wird festgestellt welche Gehirnareale aktiv waren.) Mit 11C­ oder 18F­Desoxyglucose können Metabolismus, Tumore und Degenerationen untersucht werden. Vorteile: • Es kann jedes beliebige radioaktiv markierte Molekül verwendet werden (mit Spezifität für bestimmte Organe, ...). • (relativ geringe Strahlung) • Störungen durch kosmische Strahlung können erkannt und somit entfernt werden. • sehr vielseitig Nachteile: • kurze Halbwertszeit der Radionuklide • teuer • komplexe Mathematik • Strahlenbelastung (wenn auch relativ gering) • sehr aufwendig Bsp. (Bild siehe Folie 2.18): Beim Sehen von Worten ist der occipitale Cortex aktiv, beim Lesen sind motorischer und sensoricher Cortex aktiv und beim Erzeugen von Worten der frontale Cortex. Bsp. (Bild siehe Folie 2.19): Musik ist eher im rechten Temporallappen und Sprache eher im linken. Bsp. (Bild siehe Folie 2.20): Das obere und untere Rasterbild werden in leicht unterschiedlichen Hirnarealen verarbeitet. Single Photon Emission Computertomographie (SPECT) Bei der SPECT wird dem Patienten eine Lösung mit einem Molekül, das ein Radionuklid, das bei seinem Zerfall γ­Strahlung freisetzt, enthält, injiziert. Diese γ­Strahlen werden mit einer Kamera aufgenommen. Die Kamera rotiert langsam um den Patienten, sodaß, wie bei der Computertomographie, Bilder aus vielen verschiedenen Winkeln aufgenommen werden. Aus diesen Bildern wird dann ein Schnittbild berechnet. Viele Schnittbilder zusammen ergeben wieder ein 3­ dimensionales Bild. SPECT ist der PET sehr ähnlich. Es gibt 2 wesentliche Unterschiede: • Die γ­Quanten (γ­Strahlen) stammen direkt von einem Radionuklid (statt von einem Positron, das von einem Radionuklid stammt). Es können Radionuklide mit deutlich längeren Halbwertszeiten als bei PET verwendet werden. Es ist also kein Zyklotron in der 11 / 77 Neurobiologie • Nähe nötig. Es werden (weil sie nicht von ein Positron stammen) nicht 2 γ­Quanten im Winkel von 180° abgestrahlt, sondern nur 1 γ­Quant (1 Photon) pro Zerfall. Dadurch können Störungen aufgrund kosmischer γ­Strahlung nicht erkannt und somit nicht herausgefiltert werden. SPECT wird sehr häufig verwendet (z.B.: zum Diagnostizieren von Schlaganfällen oder zur Darstellung von Blutgefäßen). Vorteile: • Es kann jedes beliebige radioaktiv markierte Molekül verwendet werden (mit Spezifität für bestimmte Organe, ...). • Radionuklide mit längeren Halbwertszeiten als bei PET • sehr vielseitig • billiger und weniger aufwenig als PET Nachteile: • Strahlenbelastung (wenn auch relativ gering) • Störungen durch kosmische Strahlung können nicht erkannt werden • geringere Auflösung als PET Bsp. (Bild siehe Folie 2.22): p­J­Amphetamin reichert sich im Gewebe in Abhängigkeit von der Durchblutung an. Technetium geht nicht durch die Gefäßwand und wird daher zur Darstellung von Blutgefäßen verwendet. Kernspinresonanztomographie (NMR­Imaging, MRI) Atome mit ungerader Anzahl an Protonen oder Neutronen (z.B.: 1H, 13C, 31P)16 haben einen Spin17 und somit ein magnetisches Moment18. Solche Atome präzessieren19 in einem (starken) Magnetfeld um die Richtung des Magnetfeldes. Dieser Präzessionsbewegung entspricht eine genau festgelegte Frequenz (Resonanzfrequenz), die von der Art des Atoms und der Stärke des Magnetfeldes abhängt (z.B.: 42,58MHz für Wasserstoff in einem Magnetfeld der Stärke 1 Tesla). Wenn Radiowellen genau dieser Frequenz eingestrahlt werden (d.h. ein (zusätzliches) Magnetfeld, das sich mit dieser Frequenz ändert, angelegt wird), kann deren Energie die Atome auf ein höheres Energieniveau heben. Nach Abschalten der Radiowellen fallen die Atome auf ihr ursprüngliches Energieniveau zurück und geben dabei Energie in Form von Radiowellen ab. (Durch das starke Magnetfeld werden die Atom in die Richung des Feldes ausgerichtet. Durch Anlegen des Wechselfeldes (Magnetfeld, das sich mit vorgegebener Frequenz ändert) werden die Atome ausgelenkt und präzessieren dann (das ist das höhere Energieniveau). Durch diese Präzession wird in einer Meß­Spule ein Strom induziert (von den Atomen abgestrahlte Radiowellen), der von einem Computer aufgezeichnet wird. Nach Abschalten des Wechselfeldes richten sie sich mit der Zeit (Millisekunden bis Sekunden) wieder ganz im starken Magnetfeld aus. Entsprechend gehen die abgestrahlten Radiowellen zurück und in der Meß­Spule wird immer weniger Strom induziert.) Die Stärke des Signals hängt von der Menge der angeregten Kerne ab. Die Signaldauer (d.h. bis wieder keine Radiowellen mehr abgestrahlt werden) hängt von der Wechselwirkung der angeregten Atome (meist 16 Z.B.: Protonen haben Spin +1/2 oder ­1/2. D.h. der Spin zweier Protonen mit entgegengesetztem Spin ist 0 (+½ + ­½ = 0). Bei einem Atom mit ungerader Anzahl an Protonen kann sich der Spin nicht vollständig aufheben. (analoges gilt für Neutronen) 17 Den Spin kann man sich ca als eine Art Eigenrotation vorstellen, auch wenn er nicht wirklich eine Rotation (oder Drehimpuls) im Sinn der klassischen Physik ist. 18 Stark vereinfacht kann man sich diese Atome als magnetische Kreisel vorstellen. 19 Präzessieren bedeutet, daß sich die Achse, um die der Kreisel rotiert langsam bewegt (das Ende der Ache beschreibt einen Kreis.) D.h. der Kreisel rotiert schnell um seine Drehachse und die Drehachse (mitsamt rotierendem Kreisel) dreht sich langsam um eine Achse senkrecht auf des Kraftfeld (z.B. das Gravitationsfeld der Erde bei einem herkömmlichen Kreisel oder das Magnetfeld bei einem Atom). 12 / 77 Neurobiologie H) mit anderen Atomen ab (d.h. vom Molekül in dem sie sich befinden und damit auch vom Gewebe (z.B.: Gewebe mit viel/wenig H2O oder anderen H­hältigen Molekülen)). Weil die Resonanzfrequenz der Präzession von der Stärke des Magnetfeldes abhängt, kann bei fixer Frequenz der Radiowellen durch unterschiedliche Feldstärken festgelegt werden, ob die Atome durch die Radiowellen angeregt werden. Es wird ein Gradient angelegt und dadurch eine Ebene festgelegt in der die Atome angeregt werden. (Das Magnetfeld ist auf einer Seite etwas stärker als auf der anderen. Dazwischen gibt es irgendwo eine Ebene in der die Feldstärke genau so ist, daß die Resonanzfrequenz mit den Radiowellen übereinstimmt.) (Um dann eine andere Ebene auszuwählen kann entweder der Gradient geändert werden (erlaubt eine beliebige Änderung der Ebene) oder die Frequenz der Radiowellen geändert werden (erlaubt eine Parallelverschiebung der Ebene).) (Durch weitere Magnetfeld­Gradienten können Phase (Gradient direkt nach Einstrahlen der Radiowellen angelegt) und Frequenz (Gradient während des Messens angelegt) einzelner Punkte in dieser Ebene festgelegt werden, sodaß die Informationen im gemessenen Strom in Informationen der Einzelnen Punkte der Ebene zerlegt werden können.) (Bei MRI wird mit einer Spule ein Strom gemessen, der Informationen über alle Atome einer Ebene enthält. D.h. die direkten Meßwerte sind im Gegensatz zur Computertomographie (wo sie Röntgebilder sind), keine für Menschen brauchbare Information und müssen auf jeden Fall von einem Computer verarbeitet werden, bevor sie irgendwie verwendet werden können.) D.h. das starke homogene Magnetfeld wird von einem Magnetfeld­Gradienten überlagert, der die Ebene, aus der Informationen gesammelt werden sollen, festlegt. Da der Gradient beliebig hineingelegt werden kann, kann auch diese Ebene beliebig festgelegt werden (d.h. im Gegensatz zur Computertomographie braucht sie nicht parallel zu irgendeiner Ebene des Geräts sein). Es wird also isotrope20 Information geliefert. Mit MRI können nicht nur Schichtbilder, sondern wirklich 3­ dimensionale Bilder gemacht werden. (Durch Änderungen der Frequenz der Radiowellen können der Reihe nach viele Ebenen (ohne Änderung des Gradienen) gemessen werden. Dann kann durch Änderung (z.B.: Drehung) des Gradienten die Richtung der Ebenen geändert werden und wieder mit einer Abfolge von Frequenzen viele Schnittbilder gemacht werden. ...) (Für MRI gibt es auch Kontrastmittel, die verwendet werden können um einige Strukturen besser sichtbar zu machen. Im Gegensatz zur PET können die Aufnahmen auch ohne Injektion irgendeiner Lösung gemacht werden. Die Verwendung von Kontrastmitteln ist optional und wird nur für bestimmte Untersuchungen angewandt.) Vorteile: • keine Strahlenbelastung (Es wird keinerlei ionisierende Strahlung verwendet (keine Röntgenstrahlung, keine Radioaktivität, ...), sondern nur Magnetfelder (allerdings ein sehr starkes (zur Zeit bis maximal ca 7 Tesla)).) • nicht invasiv (keine Injektion einer Lösung nötig) • Es kann isotrope Information geliefert werden (wirklich 3­dimensionale Bilder). • Bei 2­dimensionalen Bildern kann die Ebene der Schnittbilder beliebig festgelegt werden. (Die einzelnen Schnittbilder können auch entlang eines beliebigen (gekrümmten) Pfades aufgenommen werden.) • sehr gute Gewebsauflösung und Gewebekontrast (Organe und Gewebe können gut unterschieden werden. (Der Kontrast zwischen verschiedenen Weichteilgeweben (die mit Röntgenstrahlung kaum unterschieden werden können) ist sehr gut, weil die Messung auf Wasserstoffatomen beruht und auch deren Bindung (in wechem Molekül sie sind, ...) miteinbezieht.) (Z.B. können weiße und graue Substanz21) im Gehirn gut unterschieden 20 isotrop = richtungsunabhängig 21 weiße Substanz (des Telencephalons) = Substantia alba graue Substanz (des Telencephalons) = Substantia grisea 13 / 77 Neurobiologie werden.)) • gute Auflösung (Punkte mit ca 1mm Durchmesser (oder noch weniger)) Nachteile: • Aufgrund des starken Magnetfeldes dürfen keine (ferromagnetischen) Metalle in die Nähe des Geräts kommen, weil sie sonst hineingezogen werden. Weiters können elektrische Geräte (z.B.: Herzschrittmacher) geschädigt werden. • hohe Kosten • relativ lautes Klopfgeräusch Beispiele (siehe Folien 2.26 bis 2.33): guter Kontrast zwischen grauer und weißer Substanz im Gehirn Knochen (Wirbeln), Bandscheiben, Rückenmark sind gut erkennbar Das Blut bewegt sich in der Zeit zwischen der Anregung durch die Radiowellen und der Abstrahlung, die dann gemessen wird. Dadurch kann man Bilder machen, auf denen zu sehen ist, was in eine Schicht hineinfließt. Blut und Gefäße sind auch ohne Kontrastmittel gut sichtbar. Es können Adern im Gehirn gefunden werden, die Platzen könnten (man kann dann vorbeugend operieren). Weiters können Verschlüsse von Arterien gefunden werden. Tumore sind gut zu erkennen. Aufgrund der genauen 3­dimensionalen Darstellung kann die Operation gut geplant werden. Funktiononelle magnetische Resonanztomographie (fMRI) MRI liefert Informationen über die Anatomie. fMRI ist MRI, die Informationen über die Funktion liefert. Oxigeniertes Hämoglobin ist im Gegensatz zu nicht oxigeniertem Hämoglobin paramagentisch und beeinflußt dadurch das Magnetfeld. Daher kann oxigeniertes Hämoglobin mit MRI sichtbar gemacht werden. fMRI liefert Bilder, die die Menge an oxigeniertem Hämoglobin in jedem Punkt anzeigen. Je aktiver ein Teil des Gehirn ist, desto mehr Blut wird hingeleitet und desto mehr oxigeniertes Hämoglobin ist in diesem Teil. fMRI zeigt also anhand der Stärke der Durchblutung, wie aktive eine Gehirnregion ist. Es können Gehirnareale identifiziert werden, die bei bestimmten Tätigkeiten (z.B.: Emotionen) aktiviert werden. Vorteile: • wie bei MRI • nicht invasiv • kann schnell durchgeführt werden • gut Auflösung (ca 3 mm3) Nachteile: • wie bei MRI Bsp. (siehe Folie 2.35): Aktivitäten im visuellen System (Wahrnehmung), der Amygdala22 (Emotionen) und im orbitofrontalen Kortex beim Betrachten von Gesichtern mit starkem emotionalen Ausdruck Magnetische Resonanz­Spektroskopie (NMR) NMR bildet die Basis von MRI. Bei der NMR wird ein starkes Magnetfeld erzeugt, in dem sich die 22 Die Amygdala gehört zu den Nuclei basales (im weiteren Sinne), die im Cerebrum liegen. 14 / 77 Neurobiologie Spins der Atomkerne (die nicht Spin 0 haben) ausrichten. Dann wird ein kurzer Radiofrequenz­Puls (der viele verschiedene Schwingungen enthält) erzeugt, der eine Auslenkung (höheren Energiezustand) bewirkt. Nach dem Puls gehen die Atomkerne wieder in den ausgerichteten Zustand zurück. Die dabei abgegebene Strahlung induziert dabei in einer Meßspule einen Strom, der gemessen wird. Das Magnetfeld, das auf einen Atomkern wirkt, hängt zu einem geringen Teil von der chemischen Umgebung (d.h. in welchem Molekül es ist, an welche Atome es gebunden ist, ...) des Atoms ab. (Die durch die chemische Umgebung bewirkte Änderung des Magnetfeldes (und damit der Frequenz der Strahlung) im Vergleich zum Atom ohne diese chemische Umgbung heißt chemische Verschiebung. Sie wird auf die Frequenz der Strahlung ohne die chemische Umgebung (das unveränderte starke Magnetfeld) bezogen und in ppm angegeben.) D.h. die abgegebene Strahlung ist leicht unterschiedlich, je nach chemischer Umgebung des Atoms. Der gemessene Strom enthält die verschiedenen Strahlungen von Atomen in vielen unterschiedlichen chemischen Umgebungen. Daraus werden die Daten nach Atomen in gleicher Umgebung aufgetrennt (mittels Fourier­Transformation berechnet), sodaß man die Information erhält welche verschiedenen chemischen Umgebungen des untersuchten Atoms (z.B.: H) in der Probe (bzw. dem untersuchten Teil des Patienten) vorhanden sind. Aus der Information über die chemsichen Umgebungen der H­Atome, kann auf das Molekül, in dem sie sich befinden, zurückgeschlossen werden. Die NMR kann daher zur Identifikation von chemischen Substanzen verwendet werden. Da sich im Gewebe immer viele verschiedene Moleküle befinden, ergeben sich Mischspektren, die Auskunft über die Zusammensetzung des Gewebes liefern. Die Peakhöhe ist proportional zur Konzentration der Substanzen. Meist werden 1H­Atome (oder 13C, 31P) für die Spektren verwendet. Die chemische Verschiebung hängt von der Stärke des starken Magnetfeldes ab. D.h. je stärker das Magnetfeld, desto besser und genauer die Messungen. Es ist möglich, NMR Messungen von einzelnen cm3 im Gehirn zu machen. Man kann dann eine einzelne Substanz auswählen und für das ganze Gehirn die Verteilung dieser Substanz anzeigen lassen. Damit können z.B. kaputte Enzyme gefunden werden, ohne daß eine Probe genommen werden müßte. (Weiters kann aus der Konzentration von H+ der pH bestimmt werden.) Vorteile: • ähnlich wie bei MRI • nicht invasiv • Substanzen an beliebigen Stellen im Gehirn können bestimmt werden Nachteile: • ähnlich wie bei MRI Elektroenzephalographie (EEG) Bei der Elektroenzephalographie wird die Spannung an mehreren Stellen auf dem Kopf gemessen (die Eletroden berühren die Kopfhaut). Die Spannungsschwankungen auf der Kopfhaut werden durch die elektrische Aktivität der Neuronen im Gehirn verursacht (besonders der Hirnoberfläche, weil die meßbare Intensität mit zunehmender Entfernung schnell abnimmt). Aufgezeichnet werden die Spannungen relativ zu einer Vergleichselektrode. Die Meßwerte werden entweder auf Papier gezeichnet oder in einem Computer gespeichert. (Eine rechenintensive Weiterverarbeitung mit einem Computer ist nicht nötig.) EEG mißt die durch viele Neuronen gemeinsam verursachten Spannungsschwankungen direkt und nicht nur erhöhte Durchblutung/O2­Gehalt wie bei fMRI oder Metabolismus (z.B.: Umsetzung von 15 / 77 Neurobiologie Glucose) wie bei PET oder SPECT. EEG hat eine sehr gut Zeitauflösung (Millisekunden), aber eine schlechte Ortsauflösung (einige cm). D.h. die von jeder einzelnen Elektrode gemessenen Spannungeschwankungen werden von jeweils einem einige cm2 großen Gehirnareal verursacht (und sind somit die Summe der Aktivitäten aller Neuronen dieses Areals). (fMRI (und auch PET, SPECT) haben eine bessere Ortsauflösung, aber eine schlechtere Zeitauflösung (Sekunden bis zu Minuten).) EEG Anwendungen: • Epilepsien23: EEG wird verwendet um Epilepsien zu erkennen. • Pharmako­EEG: Bei einem Pharmako­EEG wird gemessen, ob eine dem Patienten verabreichte Substanz z.B. stimulierende oder sedative Wirkung hat. Alle psychoaktiven Substanzen werden mit dieser Methode an Freiwilligen getestet, bevor sie freigegeben werden. • EEG­Mapping • Kohärenzanalysen: Mit EEG ist erkennbar, ob bestimmte Gehirnareale im gleichen Rhythmus schwingen und somit gerade zusammenarbeiten. Vorteile: • billig • gute Zeitauflösung • Äktivität der Neuronen wird direkt gemessen • nicht invasiv Nachteile: • schlechte Ortsauflösung Bsp. (siehe Folie 2.40): (seizure = (plötzlicher) Anfall; convolution = (epileptischer) Anfall; petit mal = kleiner epileptischer Anfall) (LT = linker Temporallappen, RT = rechter Temporallappen, LF = linker Frontallappen; LO = linker Okzipitallappen, ...) Das obere EEG24 zeigt einen starken Anfall, bei dem das ganze Gehirn betroffen ist. EEG wird in der Medizin hauptsächlich für das Identifizieren/Orten/Klassifizieren von epileptischen Anfällen verwendet. Magnetenzephalographie (MEG) Bei Magnetenzephalographie werden Magnetfelder gemessen, die von den elektrischen Strömen im Gehirn erzeugt werden. (Die Ströme verursachen sowohl die Spannungsschwankungen auf der Kopfhaut (mit EEG gemeßbar), als auch Magnetfelder (mit MEG gemeßbar).) Die Sensoren (supraleitende Spulen) brauchen den Kopf nicht berühren, sondern nur in unmittelbarer Nähe des Kopfes sein (meist einige 100 Spulen, die in einer Art Helm um den Kopf angeordnet sind). Aus den Meßwerten kann (bisher nur relativ ungenau und nicht eindeutig) berechnet werden, wo der Strom, der das Magnetfeld erzeugt hat, gefloßen ist. D.h. welche Gruppen von Neuronen im Gehirn aktiv waren. Wie bei EEG muß auch bei MEG in vielen (tausenden) Neuronen der Strom in die gleiche Richtung fließen, damit ein meßbares Signal entsteht. Weil die vom Gehirn erzeugten Magnetfelder sehr schwach sind (einige fT (femto Tesla; 10­15T)) muß der Raum, in dem die Messung stattfindet, sehr gut gegen Magnetfelder (Magnetfelder von Autos, Aufzügen, Erdmagnetfeld, ...) von außen abgeschirmt sein. Weiters werden durch spezielle Sensoren und bei der Berechnung Störungen herausgefiltert. 23 "Epilepsie" ist eine Sammelbezeichnung für chronische Erkrankungen des Gehirns bei denen immer wieder durch exzessive Entladungen von Neuronen verursachte Krämpfe auftreten 24 EEG steht nicht nur für Elektroenzephalographie, sondern auch für Elektroenzephalogramm 16 / 77 Neurobiologie Wie beim EEG ist die zeitliche Auflösung sehr gut (Millisekunden). Die mit MEG gemessenen Magnetfelder werden hauptsächlich von intrazellulärem Strom erzeugt, während die Spannungen beim EEG hauptsächlich durch extrazellulären Strom zustandekommen. Wie EEG wird auch MEG für Epilepsie­Untersuchungen verwendet. Weiters können funktionelle Untersuchungen gemacht werden und rhythmische Aktivitäten untersucht werden. MEG wird z.B. für Sprachuntersuchungen verwendet oder der Patient bekommt eine Aufgabe und es wird gemessen, welche Gehirnareale bei der Bearbeitung der Aufgabe aktiv sind. Nur EEG und MEG erlauben Untersuchungen in Echtzeit. Alle anderen Methoden liefern erst im nachhinein und nur gesamt über einen längeren Zeitraum (Sekunden bis Minuten) welche Gehirnareale aktiv waren (z.B.: die Durchblutung nimmt nicht in der selben Millisekunde zu, in der ein Gehirnareal stärker aktiv wird, sondern erst etwas verzögert). Vorteile: • gute Zeitauflösung • Äktivität der Neuronen wird direkt gemessen • nicht invasiv Nachteile: • teurer als EEG (abgeschirmter Raum, flüssiges Helium zum Kühlen der supraleitenden Spulen, ...) Zellen des Nervensystems Das Gehirn besteht aus ca 1010­1012 Neuronen (Nerven Zellen) und weiters aus Glia Zellen und Ependym Zellen. Es gibt ca 10 mal mehr Glia Zellen im Gehirn als Neuronen, aber das Volumen aller Glia Zellen im Gehirn zusammen ist ca gleich groß wie das Volumen aller Neuronen im Gehirn zusammen. Die Ependym Zellen bilden die Oberfläche der Hirnhaut und der Gefäße (sie werden in dieser Vorlesung nicht weiter behandelt). Es ist schwer die Zahl an Neuronen im Gehirn genau zu bestimmen, weil die Dichte der Neuronen in jedem Gehirnareal anders ist. Es gibt viele verschiedenen Arten von Nerven Zellen und von Glia Zellen. Glia Zellen Glia Zellen ist ein Sammelbegriff für praktisch alle Zellen im Gehirn, die keine Nervenzellen sind (auch Ependym Zellen werden gelegentlich zu den Glia Zellen gerechnet). D.h. die Zusammenfassung zu Glia Zellen bedeutet nicht, daß alle diese Zelltypen nahe miteinander verwandt sind (z.B.: Microglia sind spezialisierte Makrophagen). (Astrocyten und Oligodendrocyten werden als Macroglia zusammengefaßt.) Die wichtigsten Glia Zellen sind Oligodendrocyten, Schwann­Zellen, Astrocyten und Microglia. Glia Zellen kontrollieren die synaptische Aktivität durch Regulation des Glukose­Transports und des Blutflusses. (Glia Zellen nehmen Glucose aus dem Blut auf und wenn sie viel Glukose verbrauchen bewirken sie eine Erhöhung des Blutflusses um mehr Glukose zu bekommen (d.h. Bildgebende Verfahren, mit denen die Gehirnaktivität aufgrund der Durchblutung angezeigt wird (z.B.: fMRI), messen eigentlich die Aktivität der Glia Zellen).) Weiters kontrollieren sie die Hirnentwicklung und Synaptogenese durch Wachstumsfaktoren (GDNF25, TNFα26, ...) und spezifische Signale an Neuronen. 25 GDNF = glia derived neurotrophic factor 26 TNFα = tumor necrosis factor α 17 / 77 Neurobiologie Es findet viel Austausch an Information zwischen Glia Zellen und Neuronen statt. Die Glia Zellen beeinflussen Erregbarkeit, ... von Nervenzellen. Es gibt auch viele Rezeptoren auf Glia Zellen (nicht nur auf Neuronen). Glia Zellen sind bisher aber deutlich weniger erforscht als Neuronen. Oligodendrocyten und Schwann­Zellen Sowohl Oligodendrocyten als auch Schwann­Zellen bilden die Myelinscheide (E: myelin sheath) um Axone von Neuronen. Die deutlichsten Unterschiede sind, daß Oligodendrocyten nur im ZNS (Gehirn und Rückenmark) vorkommen und ein Oligodendrocyt gleichzeitig die Myelinscheiden mehrerer Axone bilden kann (d.h. ein Oligodendrocyt umwickelt getrennt mehrere Axone), während Schwann­Zellen nur im PNS (gesamtes Nervensystem ausser ZNS) vorkommen und eine Schwann­ Zelle immer nur die Myelinscheide eines Axons bildet. Die Myelinscheide bildet eine elektrische Isolierung des Axons. Bis auf einige freie Stellen (Einschnürungen), die als Ranvier­Schnürringe bezeichnet werden (und für die Erregungsweiterleitung wichtig sind), ist das ganze Axon umwickelt. Die Myelinscheide (und damit auch der Oligodendrocyt) ist weiß. Teile des Gehirns, die viel Oligodendrocyten enthalten sind daher heller (weiße Substanz) als Teile mit weniger Oligodendrocyten (graue Substanz). Astrocyten Astrocyten haben mit ihren Endfüßchen direkten Kontakt einerseits zu Blutgefäßen und andererseits zu Neuronen. Astrocyten bilden die Blut­Hirn­Schranke. Alle Stoffe, die zwischen Blut und Neuronen ausgetauscht werden, müssen durch Astrocyten hindurch. Sie müssen durch aktiven Transport von den Astrocyten aufgenommen werden, wobei allerdings lipophile Substanzen leichter passieren können. (Unter anderem muß auch Glukose durch die Astrocyten hindurch. Glucose wird in den Astrocyten verarbeitet und erst das Produkt dieser Verarbeitung wird an die Neuronen zur Energiegewinnung weitergegeben.) Weiters füllen die Astrocyten den Raum zwischen den Neuronen (und halten dadurch die Neuronen an ihrem Platz) und regulieren auch die chemische Zusammensetzung der Flüssigkeit außerhalb der Zellen (z.B. entfernen sie (gebrauchte) Neurotransmitter). Microglia Micorglia sind der Hauptbestandteil der Immunabwehr im Gehirn. Als Phagocyten beseitigen sie auch abgestorbene Zellen und schädliche Stoffe. Neuronen (Nerven Zellen) Es gibt wahrscheinlich ca 1000 verschiedene Typen von Neuronen mit unterschiedlicher Gestalt und Größe. Ein Neuron besteht aus Soma, Dendriten und Axon. Ein Neuron hat meist nur ein Axon (E: axon). Das Axon beginnt beim Soma mit dem Axon Hügel (E: axon hillock). Das Axon ist (meist) nicht verzweigt. Das Ende (oder die Enden) des Axons wird als Bulbus terminalis (präsynaptische Terminale) (E: axon terminal, terminal bouton) bezeichnet. Ein vom Neuron ausgehendes Aktionspotential beginnt im Axon Hügel und wird über das Axon zu anderen Neuronen geleitet. Axone sind über Synapsen mit Dendriten anderer Neuronen verbunden. Die Synapse (E: synapse) ist der Kontakt­Bereich zwischen zwei Neuronen über den Information von einem Neuron in ein anderes transferiert wird. D.h. Synapsen bestehen aus Bulbus terminalis des Axons, dem synaptischen Spalt und einem Stück der Oberfläche eines Dendriten (bzw. Spines auf einem Dendriten). Ein Neuron hat (meist) viele Dendriten (E: dendrites). Dendriten sind verzweigt. Bei einigen Neuronen befinden sich auf den Dendriten viele kleine Vorsprünge/Auswüchse, die als Spines 18 / 77 Neurobiologie (Dornfortsätze) bezeichnet werden. Spines vergrößern die Oberfläche der Dendriten und dienen dem Kontakt mit Axonen. Ein Neuron hat sehr viele (10000ende) Synapsen an den Dendriten. Dendriten funktionieren als Antennen für Signale von Axonen anderer Neuronen: Die Signale anderer Neuronen kommen von deren Axonen über die Synapsen (in die Spines und weiter) in die Dendriten des Neurons. Das Soma (E: soma) ist der Zellkörper. Das Soma enthält (wie alle tierischen Zellen) Organellen: Mitochondrien, Zellkern, glattes und raues ER (Endoplasmatisches Reticulum), Ribosomen und Golgi Apparat. (Neuronen enthalten besonders viel raues ER.) Neuronen können mit verschiedenen Farbstoffen spezifisch angefärbt werden, sodaß nur bestimmte Neuronen sichtbar werden. Z.B. Golgi­Färbung färbt die Pyramidenzellen (die Farbe in den Zellen ist Silberchromat). Der freie Raum zwischen den Neuronen ist mit Liquor cerebrospinalis (Gehirnflüssigkeit) gefüllt. Der Liquor cerebrospinalis dient der Dämpfung von Erschütterungen. Weiters können Stoffe von den Zellen in Liquor cerebrospinalis hinausdiffundieren. Das Cerebellum steuert (unter anderem) die Feinmotorik. Die Kleinhirnrinde enthält 5 Typen von Nervenzellen (Körnerzellen, Purkinjezellen, Golgi­Zellen, Korbzellen, Sternzellen), die in 3 Schichten angeordnet sind. Die oberste (äusserste) Schicht heißt Molekularschicht (Stratum moleculare) (E: molecular layer) und enthält Korbzellen (E: basket cells), Sternzellen (E: stellate cells), Parallelfasern (das sind die Axone der Körnerzellen) und die Dendriten der Purkinjezellen. Die mittlere Schicht heißt Purkinjezellschicht (Stratum purkinjense) (E: purkinje cell layer) und enthält die Purkinjezellen (E: Purkinje cells). Die unterste (innerste) Schicht heißt Körnerzellschicht (Stratum granulosum) (E: granular layer) und enthält Körnerzellen (E: granule cells) und Golgi­Zellen (E: Golgi cells). Das Cerebellum enthält 1011 Körnerzellen. Es gibt im Cerebellum mehr Körnerzellen als es im Cortex cerebri27 (E: cerebral cortex) Nervenzellen gibt. Die Purkinjezellen sind die einzigen Neuronen des Kleinhirns, die eine Ausgabe aus dem Kleinhirn produzieren (sie liefern Informationen an andere Teile des ZNS). Die Purkinjezellen haben einen stark verzweigten Dendritenbaum, wobei die Dendriten einer Zelle (fast) vollständig in einer Ebene liegen. Die Parallelfasern liegen senkrecht zu den Ebenen der Purkinjezellen­Dendriten. Purkinjezellen haben Synapsen (d.h. erhalten Informationen) von den Parallelfasern und den Kletterfasern (und von Korbzellen und Sternzellen). Purkinjezellen sind hemmende Zellen (d.h. Signale von Purkinjezellen inhibieren die Zielneuronen). Eingaben erhält das Kleinhirn über die Moosfasern (E: mossy fibers) und die Kletterfasern (E: climbing fibers). Die Moosfasern kommen vom Hirnstamm und vom Rückenmark und innervieren28 Körnerzellen. Die Kletterfasern kommen von der Medulla oblongata und innervieren Purkinjezellen. Die Körnerzellen sind erregende Zellen (d.h. Signale von Körnerzellen erhöhen die Wahrscheinlichkeit der Erzeugung eines Signals in der Zielzelle). Die Somata der Körnerzellen liegen in der Körnerzellschicht, aber die Axone gehen in die Molekularschicht, verlaufen dort parallel zur Oberfläche und werden als Parallelfasern bezeichnet. Da die Axone der Körnerzellen senkrecht durch die Ebenen der Dendriten der Purkinjezellen gehen, geht jedes Axon durch die Dendritenbäume vieler Purkinjezellen und kann somit Synapsen zu vielen Purkinjezellen haben. Das Cerebellum erhält Informationen aus den Cerebrum und dem Körper und kann durch Abstimmung dieser Informationen die Details von Bewegungen steuern (d.h. die Feinmotorik). 27 Cortex cerebri = Großhirnrinde (graue Substanz an der Oberfläche des Cerebrums) 28 Neuron A innerviert Neuron B, bedeutet, daß Neuron A Synapsen zu Neuron B hat, über die Informationsfluß von Neuron A zu Neuron B erfolgt 19 / 77 Neurobiologie Die Neuronen des Kleinhirns sind multipolare Neuronen (d.h. sie haben mehrere Dendriten/Axone). Der Hippocampus ist für Lernen und Gedächtnis wichtig. Die weit verzweigten Dendriten sind dazu da, daß viele Axone Kontakt mit ihnen haben können (Bild siehe Folie 3.11). Neuronen empfangen/transportieren eingehende Informationen (sensorisches System) und verarbeiten Informationen. Sie vermitteln Lernen, Gedächtnis und Emotionen. Sie führen Entscheidungen des Gehirns aus (motorisches System). Axon Die Nerven von Tintenfischen sind nicht myelinisiert (sie haben keine Myelinscheide29). Die Geschwindigkeit der Erregungsleitung in einem Axon hängt vom Durchmesser des Axons ab: je dicker, desto schneller ist die Leitung entlang des Axons. In Tintenfischen sind daher Neuronen, die Signale schnell weiterleiten müssen, sehr dick. Das Axon eines Neurons, daß für die Flucht wichtig ist, hat einen Durchmesser von bis zu 1mm (Riesenaxon des Tintenfischs). Dieses Axon wird wegen seiner Größe (und damit einfacheren Handhabbarkeit) häufig für Forschungen verwendet. (Z.B. ist es für das Messen des Potentials viel einfacher Elektroden in ein 1mm dickes Axon einzubringen als in ein 0.01 mm dickes (oder noch dünneres).) Das Plasma eines Axons (Axoplasma) kann aus dem Axon entfernt werden, ohne die Membran des Axons zu zerstören: Das Axon wird auf eine Gummiunterlage gelegt. Dann wird mit einem Gummiroller mehrmals leicht über das Axon gerollt und so das Axoplasma am Ende herausgedrückt. Es kann eine Kanüle angeschlossen werden und damit Flüssigkeit in und durch die verbleibende Membran des Axons geleitet werden. Auf diese Weise können axonale Membran und Axoplasma isoliert und getrennt untersucht werden. Wird in die isolierte Membran eine Flüssigkeit geleitet, die eine hohe K+ und eine niedrige Na+ Konzentration hat, und das Axon in eine Flüssigkeit gelegt, die eine niedrige K+ und hohe Na+ Konzentration hat (d.h. innen viel K+ und wenig Na+ und außen umgekehrt), so stellt sich das Ruhepotentioal von ­60mV im Inneren im Vergleich zu außen ein und ein Aktionspotential (ca +50mV) kann ausgelöst werden und wird durch das Axon geleitet. D.h. das Axon verhält sich genauso wie ein Axon, das noch sein urspüngliches Plasma enthält. Ein Axon ist von Axolemma umgeben und enthält Microtubuli, Neurofilamente, Actin Microfilamente, Mitochondrien, axoplasmatisches Reticulum, Lysosomen und Vesikel. Axolemma: Zellmembran des Axons Microtubuli: Polymer aus dem Protein Tubulin; gerade Röhre mit ca 20nm Durchmesser; Teil des Cytoskeletts; Neurofilament: eine bestimmte Art von Intermediärfilament (es gibt viele verschiedene Proteine, die Intermediärfilamente bilden können); ca 10nm Durchmesser; Teil des Cytoskeletts Microfilament: Polymer aus dem Protein Actin; Strang mit ca 5nm Durchmesser; Teil des Cytoskeletts axoplasmatisches Reticulum: Es ist dem endoplasmatischen Reticulum ähnlich, hat aber eine etwas andere Zusammensetzung. Es ist am Anfang des Axons mit dem rauen Endoplasmatischen Reticulum in Soma verbunden. Lysosomen: sind membranumgrenzte Organellen und dienen der Verdauung (z.B.: Abbau von zelleigenen Stoffen) (microtrabeculares System: Netzwerk aus Filamenten mit ca 2nm Durchmesser; vermutlich existiert es gar nicht, sondern war nur eine Folge der Präparationmethoden bei der Untersuchung von Cytoplasma in den Jahren zwischen 1970 und 1980) Ein wesentlicher Unterschied des Axons im Vergleich zum Soma ist, daß das Axon keine Ribosomen und kein raues endoplasmatisches Reticulum enthält. Jedes Neuron, das Sinnesreize von der Zehe in Richtung ZNS leitet, geht von der Zehe bis ins Rückenmark und erst im Rückenmark wird das Signal auf ein anderes Neuron übertragen. D.h. 29 Die Myelinscheide ermöglicht besonders schnelle Erregungsleitung, ohne daß das Axon dafür dicker sein muß. 20 / 77 Neurobiologie diese Neuronen sind ca 1 m lang. Proteine müssen aktiv transportiert werden, damit sie bis ans Ende das Axons kommen können. Es gibt daher ein axoplasmatisches Transportsystem. Es funktioniert indem Vesikel den Microtubuli entlang wandern. Unterschiede zwischen Axon und Dendriten: Axon Dendriten ein Neuron hat meist nur 1 Axon ein Neuron hat viele Dendriten ein Axon ist wenig verzweigt Dendriten sind stark verzweigt im Axon gibt es keine Proteinsynthese (weil es keine Ribosomen gibt) in Dendriten gibt es Proteinsynthese (in Dendriten gibt es Ribosomen) die Mirotubuli im Axon sind gerichtet die Mirotubuli in Dendriten sind ungerichtet im Axon gibt es kein MAP II in Dendriten gibt es MAP II im Axon gibt es tau (τ) Proteine in Dendriten gibt es keine τ Proteine im Axon ist ein Aktionspotential möglich in Dendriten gibt es kein Aktionspotential (Es ist aber eine Rückpropagierung des Aktionspotentials möglich, die eine Depolarisierung bewirkt.) (Axone haben auf ihrer ganzen Länge ca gleichen Durchmesser.) (Dendriten werden meist immer schmäler) Im Axon sind die Microtubuli einheitlich mit ihrem Plus­Ende in Richtung Terminale30 und mit ihrem Minus­Ende in Richtung Soma orientiert. Transport im Axon erfolgt sowohl in Richtung Terminale als auch in Richtung Soma. Der Transport in Richtung Terminale heißt anterograder Transport. Es gibt 2 Arten von anterogradem Transport: schneller anterograder Transport und langsamer anterograder Transport. Mit dem langsamen anterograden Transport werden hauptsächlich Bestandteile des Cytoskeletts (Tubulin, Neurofilamente, Actin) und lösliche Proteine transportiert. Es ist unabhängig von den Microtubuli. Seine Geschwindigkeit ist ca 1­10mm pro Tag. Mit dem schnellen anterograden Transport werden Vesikel und Mitochondrien trasnportiert. Er erfolgt entlang den Micotubuli. Kinesin (bzw. Proteine der Kinesin­Familie), das an die Vesikel gebunden ist, bindet auch an die Microtubuli und bewegt die Vesikel in Richung Plus­Ende des Microtubulis. Die Geschwindigkeit des schnellen anterograden Transports ist ca 10­40 cm pro Tag (100­400 mm pro Tag) (d.h. ca 2­3 Tage von der Zehe bis zum Rückenmark), also ca 100 mal so schnell wie der langsame anterograde Transport. Der Transport in Richtung Soma heißt retrograder Tansport. Mit dem retograden Transport werden Endosomen31, Mitochondrien, neurophile Signale (z.B.: Wachstumsfaktoren) und Viren transportiert. Der retrograde Transport erfolgt in der selben Weise wie der schnelle anterograde Transport, nur in die entgegengesetzte Richtung und mit einem anderen Motorprotein: Dynein. Dynein bewegt die Vesikel entlang den Microtubuli in Richtung Minus­Ende. (Z.B. Herpesviren dringen an Axon­Enden ein und nutzen den retrograden Transport um sich zum Soma bringen zu lassen.) Die an das Vesikel gebundenen Motorproteine entscheiden, wohin das Vesikel transportiert wird. 30 Terminale = das Ende des Axons (üblicherweise Teil einer Synapse) 31 Das Endosom ist eine membranbegrenzte Organelle. Es bringt Material, das durch Endocytose neu in die Zelle aufgenommen wurde, zu den Lysosomen. 21 / 77 Neurobiologie Ein Vesikel, das Kinesin gebunden hat, bewegt sich in Richtung Terminale, ein Vesikel, das Dynein gebunden hat, bewegt sich in Richtung Soma. Der Transport kann untersucht werden, indem radioaktiv markierte Proteine (oder radioaktiv markierte Aminosäuren, die dann in Proteine eingebaut werden) in die Zelle gegeben werden. Die Proteine werden dann im Axon transportiert und man kann durch Messung der Radioaktivität feststellen, wie schnell und wohin sie transportiert werden. (Z.B. kann auf diese Weise festgestellt werden, wohin die Axone einer Gruppe von Neuronen im Gehirn gehen bzw. wo sie Enden.) Beispiele für Motorproteine sind: Myosin (ist im Muskel an der Muskelkontraktion beteiligt), Kinesin und Dynein. (Bilder siehe Folie 3.17) Es gibt viele verschiedene Proteine in den Proteinfamilien der Kinesine und Dyneine (z.B.: Kinesin­I, Kinesin­II, KIF17, KIF13A, ... (Bilder siehe Folie 3.18)). Einige davon kommen nur im ZNS vor, andere im ganzen Körper. Kinesine und Dyneine benötigen ATP für den Transport. Es ist nur von wenigen dieser Proteine bekannt, wofür sie genau benötigt werden. Allerdings scheinen sie eine wichtige Funktion zu haben, weil es so viele verschiedene gibt. Die Stoffe/Proteine/... werden in Vesikeln dorthin transportiert, wo sie in der Zelle gebaucht werden. Einige Vesikel müssen in Dendriten gebracht werden, andere in Synapsen oder in Spines oder ... . Vermutlich gibt es so viele verschiedene Kinesine, weil jedes Kinesin sein Vesikel zu einem anderen Ziel bringt. Eine Art von Kinesin bringt das Vesikel zu einer Verzeigung, dort wird das Vesikel von einer anderen Art von Kinesin übernommen und weitertransportiert. Bei der nächsten Verzweigung übernimmt es wieder eine andere Art von Kinesin, bis das Vesikel an seinem Ziel ist. (Dies ist bisher nur eine Vermutung und noch nicht bewiesen.) Entwicklung des Gehirns Das ZNS entwickelt sich aus dem Neuralrohr. Die Neuronen (bzw. deren Vorläuferzellen) müssen in die richtige Schicht (z.B.: des Isocortex, der aus 6 Schichten besteht, oder des Cerebellums, das 3 Schichten hat) kommen. Geleitet werden sie dabei durch lösliche Signale (können Gradienten aufbauen) und stationäre Signale (z.B.: an Zellmembranen gebunden). Glia Zellen bauen sich als erstes auf. Die Neuronen wandern dann entlang den Glia Zellen. Die Zellen wandern in die Richtung in der die Konzentration der Wachstumsfaktoren am größten ist. Nach Verankerung der Zelle (durch Adhesionsproteine) in ihrem Zielgebiet, differenziert sie sich in ihre endgültige Form und Fortsätze (Axon und Dendriten) werden ausgebildet. Der längste Fortsatz wird das Axon, die anderen werden Dendriten. (Wenn das Axon während der Entwicklung abgeschitten wird, entwickelt sich der längste Dendrit zum Axon. Damit die Information, daß das Axon abgeschnitten wurde, zum Soma kommt, sodaß die Entwicklung des längsten Dendriten geändert werden kann, ist retrograder Tansport nötig.) Wohin das Axon wächst, wird durch verschiedene Signal­Moleküle gesteuert (durch Wachstumsfaktoren und stationäre Signale). Das Axon kann aufgrund der Konzentration (z.B. in Gradienten des Signal­Moleküls) von bestimmten Molekülen die Richtung ändern, die Seite des Gehirns wechseln, Haken schlagen, in Richtung höherer/niedrigerer Konzentration weiterwachsen, ... . Einige Moleküle bewirken ein weiteres Wachstum des Neurons. Genauso gibt es Moleküle die verhindern, daß das Axon weiterwächst, sodaß es, wenn in einem Gebiet dieses Molekül vorhanden ist, nicht in dieses Gebiet, sondern in eine andere Richtung weiter wächst ("No­Go"). Wenn das Axon in die Nähe seiner Zielzelle (der zukünfigen postsynaptischen Zelle) kommt, werden über Adhesionssignale (Adhesionsproteine) Kontakte hergestellt und schließlich Synapsen ausgebildet. 22 / 77 Neurobiologie Im Axon werden nicht einzelne Proteine, sondern Vesikel mit Proteine transportiert (also ganze vorgefertige Packete für den Aufbau von Synapsen). Wenn sich nach der Bindung an die Zielzelle die Synapse bildet, ist schon nach 30 Minuten eine Struktur erkennbar und nach 90 Minuten ist es schon eine echte Synapse. (Bilder siehe Folie 3.20) Die Spines der Dendriten sind nicht statisch, sondern bewegen und verändern sich ständig (werden auch neu gebildet und wieder abgebaut). Sie sind flexibel und plastisch. Die Bewegungen erfolgen über das Cytoskelett. In Dendriten werden nicht nur Vesikel mit Proteinen transportiert, sondern auch mRNA. Mit dieser werden dann an ihrem Ziel im Dendriten (z.B.: bei einem Spine / einer Synapse) von Ribosomen Proteine hergestellt. Das Protein ZBP1 bindet im Soma an die mRNA. Solange ZBP1 an die mRNA gebunden ist wird sie nicht translatiert. Erst wenn ZBP1 phosphoryliert wird und die mRNA frei gibt, erfolgt die Proteinsynthese. Das erlaubt eine rasche Umsetztung. Z.B.: beim Lernen, daß eine heiße Herdplatte Schmerz verursacht (der Anblick der heißen Herdplatte wird mit dem Schmerz verbunden), müssen einige Synapsen gezielt verstärkt werden. Elektrische Eigenschaften, Ionenkanäle Ein Neuron besteht aus Perikaryon (Soma), Axon und Dendriten. Es enthält einen Zellkern (Nukleus), der einen Nukleolus32 enthält, ER33, Mitochondrien, ... . Das Axon (Neurit) beginnt mit dem Ursprungskegel des Axons (Axon Hügel). Das Axon kann auch etwas verzweigt sein. Die Enden des Axons heißen Nervenendigungen (Terminale). Das Axon ist (zumindest in Mammalia) meist von einer Myelinscheide umgeben, die durch Ranvier'sche Schnürringe unterbrochen ist. (Bild siehe Folie 4.3) Eine Zelle kann zwischen 10000 und 100000 Synapsen haben. Nerven, die z.B. von den Zehen kommen, haben ihre erste Übertragung von einer Nervenzellen auf die nächste erst im Rückenmark. D.h. das Axon des ersten Neurons ist ca 1 m lang. Es ist eine schnelle Übertranung des Signals entlang des Axons nötig. Ruhepotential Alle Zellen (nicht nur Neuronen) haben ein Membranpotential. Das Membranpotential ist die elektrische Spannung über die Membran. Es ist also die elektrische Spannung im Inneren der Zelle im Vergleich zur elektrischen Spannung außerhalb der Zelle. D.h.: Membranpotention = Spannung innen ­ Spannung außen Das Ruhe­Membranpotential eines Neurons ist ca ­65 mV. D.h. das Innere des Neurons ist (um 65mV) negativer als die Umgebung des Neurons. Das Membranpotential kann gemessen werden indem eine Elektrode (Mikroelektrode) in die Zelle gestochen wird und mit einem Voltmeter die Spannung im Vergleich zu einer Elektrode, die nur in die Umgebung um das Neuron geht, gemessen wird. Die gemessende Spannung kann verstärkt und mit einem Oszilloskop angezeigt, oder der zeitliche Verlauf aufgezeichnet werden. Das Membranpotential eines Neurons in Ruhe ist gleichbleibend auf ca ­65mV. (Bild siehe Folie 4.4 A) Das Membranpotential kommt durch eine ungleiche Ladungsverteilung an der Membran zustande. Im Inneren der Zelle sind mehr negative Ladungen als positive und außen sind mehr positive als negative. Das Ruhepotential wird hauptsächlich durch K+­Ionen verursacht. 32 Der Nukleolus ist eine Struktur im Zellkern, in der ribosomale RNA transcribiert wird und ribosomale Untereinheiten zusammengesetzt werden. 33 ER = endoplasmatisches Retikulum 23 / 77 Neurobiologie Außerhalb der Zelle ist die Konzentration an K+ niedrig, an organischen Anionen34 auch niedrig und an Na+, Ca++ und Cl­ hoch. Im Inneren der Zelle ist die Konzentration an K+ hoch, an organischen Anionen auch hoch und an Na+, Ca++ und Cl­ niedrig. (jeweils als Vergleich der Konzentrationen des selben Ions innerhalb und außerhalb der Zelle (z.B.: die Konzentration an Ca++ (2mM) außerhalb der Zelle ist niedriger als die an K+ (5mM) außerhalb der Zelle aber viel höher als die Ca++­ Konzentration (0.0002mM) im Inneren der Zelle)) Für das Zustandekommen des Membranpotentials ist die Semipermeabilität der Membran wesentlich: im Ruhezustand ist die Membran hauptsächlich für K+ durchlässig (aufgrund von K+­ Kanälen), aber nicht für die anderen Ionen. Aufgrund des K+­Gradienten (die Konzentration an K+ in der Zelle ist ca 20 mal so hoch wie außerhalb) fließen K+­Ionen aus der Zelle hinaus. Ohne diesen K+­Ausstrom würden die negativen und positiven Ladungen einander aufheben (sowohl innerhalb als auch außerhalb der Zelle). Weil aber nur K+ die Membran passieren kann und die anderen Ionen (insbesondere auch die Anionen (organische und Cl­)) nicht, wandern nur positive Ladungen, und zwar aus der Zelle hinaus. D.h. im Inneren sind dann mehr negative Ladungen als positive, sodaß eine negative Gesamtladung resultiert, und außerhalb sind dann mehr positive Ladungen als negative, sodaß eine positive Gesamtladung resultiert. Nachdem positive und negative Ladungen einander anziehen, sammeln sich die überschüssigen negtiven Ionen (innen) und die überschüssigen positiven Ionen (außen) direkt an der Membran (anstatt sich auf die ganze Zelle bzw. äußere Umgebung zu verteilen). Weiters wird mit zunehmendem Ladungsunterschied zwischen außen und innen das Bestreben der K+­Ionen entlang des Ladungsgradienten wieder in die Zelle hineinzuwandern immer größer (auch das Bestreben der anderen Ionen, aber diese können nicht durch die Membran). Bei einem bestimmten Potential (Gleichgewichtspotential) wandern pro Zeiteinheit gleich viele K+­Ionen aus der Zelle hinaus (aufgrund des Konzentrationsunterschieds) wie hinein (aufgrund des Ladungsunterschieds) und somit ist kein Gesamtfluß an K+ mehr vorhanden. (Die K+­Kanäle lassen K+ in beide Richtungen durch. Die Richtung des Gesamtflusses an K+ wird durch den Konzentrationsgradienten und das Potential festgelegt.) (Das Gleichgewichtspotential hängt natürlich vom Konzentrationsgradienten ab, weil es gerade so stark ist, daß es die Ionen­ Wanderung aufgrund des Konzentrationsgradienten genau aufhebt.) Schon sehr geringe Unterschiede in der Ladungsverteilung sind ausreichend um ein Potential zu erzeugen, das den Konzentrationsgradienten kompensiert. D.h. die Konzentration an K+ innerhalb und außerhalb der Zelle ändert sich durch die Wanderung der K+ nur sehr wenig. (Z.B.: Bei einer Zelle mit 50µm Durchmesser, 100mM K+ innen und 5mM K+ außen ist die nötige Konzentrationsänderung um das Gleichgwichtspotential zu erreichen ca 0.00001mM. D.h. im Inneren ändert sich die K+­Konzentration von 100mM auf 99.99999mM.) Der Konzentrationsunterschied von K+ und Na+ zwischen innen und außen wird von der Zelle mittels Natrium­Kalium­Pumpen aktiv aufrecht erhalten. Eine Natrium­Kalium­Pumpe (Na+/K+­ ATPase) befindet sich in der Membran der Zelle und transportiert pro Schritt 2 K+ in die Zelle hinein und 3 Na+ aus der Zelle hinaus. Da die Pumpe für beide Ionen gegen den Konzentrationsgradienten arbeitet, benötigt sie dafür Energie in Form von ATP. Nachdem die Na+/K+­ATPase pro Schritt 3 positive Ladungen aus der Zelle hinaus und nur 2 positive Ladungen in die Zelle hinein transportiert, trägt sie auch (etwas) zum Ladungsunterschied zwischen innen und außen bei. Die Membran ist im Ruhezustand zwar hauptsächlich für K+ durchlässig, aber auch etwas für Na+, das aufgrund seiner höheren Konzentration außen von außen (150mM) nach innen (15mM) fließt. Wäre sie nur für K+ durchlässig, dann wäre das Gleichgewichtspotential ca ­80mV. Aufgrund der zusätzlichen geringen Durchlässigkeit für Na+ liegt das Gleichgewichtspotential insgesamt bei ca ­65mV. Die Na+/K+­Pumpe baut nur den grundsätzlichen Konzentrationsgradienten auf und spielt keine 34 Anionen sind negativ geladen 24 / 77 Neurobiologie Rolle bei schnellen Ladungs­/Spannungsschwankungen. Bei schnellen Spannungsschwankungen fließen nur wenige Ionen durch die Membran. Schnelle Spannungsschwankungen werden durch das Öffnen von Ionenkanälen bewirkt, genauso, wie das Ruhepotential durch die offenen K+­Kanäle zustande kommt. Es gilt ganz allgemein (nicht nur für K+), daß, wenn über eine Membran ein Konzentrationsunterschied eines Ions besteht und die Membran permeabel für das Ion ist, sich auf der Seite der Membran, auf der die Konzentration des Ions höher ist, Ladungen des umgekehrten Vorzeichens der Ladung des Ions sammeln. (D.h. falls das Ion positiv geladen ist, wird die Seite mit der hoheren Konzentration negativ geladen; falls das Ion negativ geladen ist, wird die Seite mit der höheren Konzentration positiv geladen.) (Natürlich unter der Voraussetzung, daß die Ladungen des Ions durch andere Ionen, die die Membran nicht passieren können, ausgeglichen waren (bevor die Ionen begannen durch die Membran zu wandern).) Nachdem das Gleichgewichtspotential (hauptsächlich) durch das Verhältnis der Ionenkonzentrationen auf der Außen­ und der Innenseite bestimmt ist, läßt es sich auch daraus berechnen. Die Formel dafür heißt Nernst Gleichung und lautet: Eion: Gleichgewichtspotential des Ions; [ion]o bzw. [ion]i: Konzentration des Ions außen (outside) bzw. innen (inside) [ion ]o RT E ion =2.303 log [mol]; R: Gaskonstante (8.316 J K­1 mol­1); T: absolute zF [ion ]i Temperatur [K]; z: Ladungszahl des Ions (z.B.: Cl­: ­1, Ca++: +2); F: Faraday Konstante (96522 A s mol­1) Die Konstante 2.303 ergibt sich dadurch, daß der dekadische Logarithmus (log10) statt dem natürlichen Logarithmus (ln, loge) verwendet wird: ln(x) ≈ 2.303 * log10(x). Eion (das Gleichgewichtspotential) gibt die Spannung an, die nötig ist damit die Ionen nicht aufgrund des Konzentrationsgradienten auf die andere Seite der Membran wandern. R und F sind Konstanten und für ein gegebenes Ion ist z konstant. Wenn auch die Temperatur fix RT (z.B.: 20°C = 293.15K) vorgegeben ist, ist 2.303 =: c konstant und damit ist dann zF [ion]o E ion =c∗log . [ion ]i 8.316∗ 273.1537 RT Bei Körpertemperatur (37°C) ist c = 2.303 = 2.303 ≈ 0.06154 V / z = zF z∗96522 61.54 mV / z. Beispiele: [ K + ]o 61.54 mV ∗log + K : E K = 1 [ K ]i Bei einem Verhältnis außen : innen = 1 : 20 ergibt sich dann: 1 E K =61.54 mV ∗log ≈ ­80.07 mV 20 D.h. wenn die Membran nur für K+ permeabel ist, ist das Gleichgewichtspotential ca ­80mV. + + + [ Na+ ]o 61.54 mV ∗log Na : E Na = 1 [ Na+ ]i Bei einem Verhältnis außen : innen = 10 : 1 ergibt sich dann: 10 E Na =61.54 mV ∗log ≈ +61.54 mV 1 + + + [Cl - ]o [Cl - ]o 61.54 mV ∗log = E Cl =−61.54 mV ∗log Cl : E Cl = −1 [Cl - ]i [Cl - ]i ­ - - 25 / 77 Neurobiologie Ca : E Ca ++ ++ [Ca ++ ]o [Ca++ ]o 61.54 mV = ∗log = E Ca =30.77 mV∗log 2 [Ca++ ]i [Ca ++ ]i ++ Nach der Nernst Gleichung hängt das Gleichgewichtspotential (abgesehen von Konstanten) nur vom Verhältnis der Ionen­Konzentrationen, der Landungszahl des Ions und der Temperatur ab, aber nicht davon wie viele Ionenkanäle es gibt (bzw. wie viele geöffnet sind) und wie gut/schnell die Ionen durch diese Kanäle passieren können. (Es ist nur notwendig, daß die Ionen die Membran irgendwie passieren können.) Wenn die Membran nur für eine einzige Art von Ion (z.B.: nur für K+) permeabel ist, bestimmt dieses eine Ion das Membranpotential und dann kann dieses Membranpotential über die Nernst­ Gleichung berechnet werden. Beispiele (Ionen­Konzentrationen in und um Neuronen und das resultierende Membranpotential, wenn die Membran nur für das jeweilige Ion durchlässig ist): Ion Konz. außen Konz. innen Verhältnis Eion (bei 37°C) K+ Na+ Ca++ 5 mM 100 mM 1 : 20 ­80 mV 150 mM 15 mM 10 : 1 62 mV 2 mM 0.0002 mM 10000 : 1 123 mV Cl­ 150 mM 13 mM 11.5 : 1 ­65 mV Ca++ ist ein wichtiger second Messenger in der Zelle. Daher muß seine Konzentration sehr niedrig gehalten werden. Wenn Kanäle für verschiedene Ionen gleichzeitig offen sind, ist das resultierende Membranpotential von allen diesen Ionen abhängig. In diesem Fall ist es auch wichtig, wie gut/schnell die verschiedenen Ionen die Membran passieren können. D.h. das Membranpotential hängt dann auch von der Permeabilität der Membran für diese Ionen (bzw. von Verhältnis der Permeabilitäten) ab. Dies wird in der Goldman Gleichung berücksichtigt: Em: Membranpotential; [X+]o bzw. [X+]i: + + [ K ]o b∗[ Na ]o Konzentration des Ions X+ (K+ oder Na+) außen E m=58 mV ∗log bzw. innen; b: Verhältnis der Permeabilitäten der [ K + ]i b∗[ Na+ ]i Membran für Na+ und K+ (PNa+ : PK+) 8.316∗ 273.1520 RT (58 mV ist das Ergebnis von 2.303 bei 20°C: 2.303 ≈ 58 mV) zF 1∗96522 Die Goldman Gleichung kann für beliebige einfach geladene Ionen verwendet werden (also z.B. nicht für Ca++), wobei für negativ geladenen Ionen (z.B.: Cl­) die Konzentration innen ([X­]i) oberhalb des Bruchstrichs steht und die Konzentration außen ([X­]o) unter dem Bruchstrich. In dieser allgemeine Form lautet die Goldman Gleichung: + P X ∗[ X ]o ∑ P Y ∗[Y ]i ∑ RT E m =2.303 ∗log wobei PX die Permeabilität der Membran für das Ion X ist. X+ F ∑ P ∗[ X + ]i ∑ P ∗[Y - ]o + X+ - Y- und Y­ stehen jeweils für mehrere verschiedene Ionen innerhalb der Summen (die Indizes wurden weggelassen), d.h. [X+] + [X+] kann z.B. für [K+] + [Na+] stehen. (Wenn man nur die beiden Ionen K+ und Na+ einsetzt, über und unter dem Bruchstrich jeweils durch PK+ dividiert und 2.303*RT/F für 20°C ausrechnet erhält man die obige Goldman Gleichung.) Wenn man nur ein einziges Ion in die Goldman Gleichung einsetzt ergibt sich die Nernst Gleichung. (Natürlich nur für z = ±1, weil diese Goldman Gleichung für mehrfach geladene Ionen nicht geeignet ist (weshalb z auch nicht vorkommt).) Bsp. (bei 37°C, Konzentrationen aus der obigen Tabelle): 26 / 77 Neurobiologie Wenn die Membran für K+ und Na+ gleich durchlässig ist, dann ist b = 1/1 = 1. [ K + ]ob∗[ Na + ]o [ K + ]o1∗[ Na + ]o = 61.54 mV ∗log + = Em = 61.54 mV ∗log + [ K ]ib∗[ Na + ]i [ K ]i1∗[ Na + ]i 51∗150 155 61.54 mV ∗log = 61.54 mV ∗log ≈ 61.54 mV * 0.1296 ≈ 7.98 mV 1001∗15 115 D.h. es ist nicht einfach der Mittelwert der Membranpotentiale von Membranen, die nur für eines der beiden Ionen durchlässig sind (der Mittelwert wäre (­80 + 62) / 2 = ­9 (≠ 7.98)). Bsp. (bei 37°C, Konzentrationen aus der obigen Tabelle): Wenn die Membran für K+ 40 mal so permeabel ist wie für Na+, dann ist b = 1/40 = 0.025: [ K + ] b∗[ Na + ]o [ K + ]o0.025∗[ Na + ]o 61.54 mV ∗log = Em = 61.54 mV ∗log + o = [ K ]ib∗[ Na + ]i [ K + ]i0.025∗[ Na + ]i 50.025∗150 8.75 61.54 mV ∗log = 61.54 mV ∗log ≈ 61.54 mV * ­1.06 ≈ 65 mV 1000.025∗15 100.375 Aktionspotential Wenn die Spannung im Inneren des Axons weniger negativ wird (z.B.: von ­65mV auf ­40mV), wird ab einem bestimmten Schwellwert ein Aktionspotential ausgelöst. (Ist die Änderung zu gering (z.B.: von ­65mV auf ­60mV) dann hat sie keine Wirkung (d.h. es wird kein Aktionspotential ausgelöst).) Die Änderung des Potentials auf ein weniger negatives Potential wird als Depolarisation bezeichnet. D.h. wenn positive Ladungen in die Zelle gebracht werden, bewirkt das eine Depolarisation in der Zelle. Diese Änderung der Spannung kann z.B. durch eine Elektrode oder durch das Öffnen von Ionenkanälen (z.B.: aufgrund eines Reizes aus der Umgebung (z.B.: aus einem Sinnesorgan)) verursacht werden. Bei einem Aktionspotential wird für kuze Zeit die Ladung im Inneren des Axons positiv. (Das Aktionspotential wird wegen dieser Umkehr der Ladung auch als Umkehrpotential bezeichnet.) Das Potential ändert sich von ­65 mV auf +40 mV, fällt dann auf einen Wert, der sogar noch etwas niedriger als die ­65mV (z.B.: ca ­80mV) ist, und geht dann wieder in das Ruhepotential von ­65mV über. (Ein Aktionspotential dauert ca 2 ms.) Direkt nach einem Aktionspotential kann kein neues Aktionspotential ausgelöst werden (nicht beeinflußbare Periode), sondern erst kurze Zeit später. (Bild siehe Folie 4.10 B) (Z.B.: es wird (zusätzlich zu der Meßelektrode) eine Elektrode in das Axon eingeführt und an dieser eine positive Spannung angelegt, die das Potential im Axon von ­65mV auf ­35mV anhebt. Durch diese Änderung steigt die Spannung im Axon weiter auf +40mV, fällt dann auf ­80mV und steigt dann wieder auf ­65mV. Sobald das Aktionspotential beginnt (d.h. die Spannung höher steigt als von der Elektrode bewirkt (hier: > ­35mV)), ist der weitere Verlauf unabhängig davon, ob die Elektrode eingeschaltet bleibt oder nicht. Falls die Spannung an der Elektrode auch nach dem Aktionspotential noch eingeschaltet bleibt, wird kurze Zeit später wieder ein Aktionspotential ausgelöst, ... .) Die Änderung des Potentials auf ein noch negativeres Potential wird als Hyperpolarisation bezeichnet. (z.B.: von ­65mV auf ­80mV) Ein Aktionspotential wird entweder ausgelöst oder nicht. Wenn es ausgelöst wird, hat es immer den gleichen Verlauf und die gleiche Höhe. Es kann kein stärkeres oder schwächeres Aktionspotential geben, sondern nur ein Aktionspotential oder kein Aktionspotential. Ein Aktionspotential bleibt auch immer gleich und wird nicht schwächer, wenn es einem Axon entlang läuft. Aktionspotentiale können nur in Axonen ausgelöst werden. In anderen Teilen des Neurons und in 27 / 77 Neurobiologie anderen Zellen löst eine Depolarisation kein Aktionspotential aus. Ein Aktionspotential kommt dadurch zustande, daß sich ab einer bestimmten Spannung, dem Schwellwert, spannungsabhängige Na+­Ionenkanäle öffnen. Durch die Na+­Kanäle strömt Na+ in das Axon, sodaß die Spannung im Axon erhöht wird. Durch den Na+ Einstrom könnte das Potential maximal auf ca +62mV (vgl. Bsp. zur Nernst­Gleichung) steigen, aber die spannungsabhängigen Na+­Kanäle schließen sich nach kurzer Zeit, schon vor erreichen des Maximalwerts, wieder (entweder Zeitabhängig oder aufgrund der höheren Spannung). Wenn das Membranpotential in den positiven Bereich kommt, beginnen sich spannungsabhängige K+­Ionenkanäle zu öffnen. Durch das Schließen der spannungsabhängigen Na+­Kanäle und das ca gleichzeitige Öffnen der spannungsabhängigen K+­Kanäle wird das Membranpotential schnell wieder negativ und fällt sogar unter den Wert des Ruhepotentials (mit den spannungsabhängigen K+­Kanälen sind mehr K+­Kanäle offen als nur beim Ruhepotential). Dann schließen sich die spannungsabhängigen K+­Kanäle wieder und die spannungsabhängigen Na+­Kanäle gehen wieder in ihren Ausgangszustand über, sodaß ein neues Aktionspotential ausgelöst werden kann. Zusammenfassung: Ereignis Wirkung Potential Ruhepotential ­65mV Depolarisation bis Schwellwert (z.B.: durch Elektrode) spannungsabhängige Na+­Kanäle öffnen sich ­40mV spannungsabhängige Na+­Kanäle offen Membranpotential wird positiv +40mV Membranpotential positiv spannungsabhängige K ­Kanäle öffnen sich +40mV Zeit oder Spannung spannungsabhängige Na+­Kanäle schließen +40mV sich (inaktiviert) spannungsabhängige Na+­Kanäle geschlossen und spannungsabhängige K+­ Kanäle offen Membranpotential sinkt auf einen Wert unter dem Ruhepotential (Hyperpolarisation) + spannungsabhängige K+­Kanäle schließen Ruhepotential; es kann wieder ein sich und spannungsabhängige Na+­Kanäle Aktionspotential ausgelöst werden gehen in ihren Ausgangszustand über ­80mV ­65mV Spannungsabhängige Na+­Kanäle haben 3 Zustände: • Ausgangszustand: geschlossen; öffnet sich wenn Potential > Schwellwert • offen: offen; geht nach kurzer Zeit (< 1 ms) oder bei bestimmter Spannung in Zustand inaktiviert über • inaktiviert: geschlossen; kann sich nicht öffnen; geht bei Ruhepotential (oder Hyperpolarisation) in den Ausgangszustand über Die Änderung des Potentials (die Depolarisation) breitet sich von der Stelle am Axon, an der sich die Na+­Kanäle geöffnet haben, in beide Richtungen entlang des Axons aus (allerdings nur eine kurze Strecke). Dadurch wird die Spannung in benachbarten Teilen des Axons über den Schwellwert erhöht und somit auch dort ein Aktionspotential ausgelöst, wenn sich die Na+­Kanäle im Ausgangszustand befinden. Wenn ein Aktionspotential einem Axon entlang wandert, kann nur in der Richtung vor dem Aktionspotential ein weiteres Aktionspotential ausgelöst werden, weil sich hinter den Aktionspotential gerade das vorhergehende Aktionspotential befunden hat und die Na+­ Kanäle somit noch nicht wieder im Ausgangszustand sind. (Meist wird ein Aktionspotential am 28 / 77 Neurobiologie Axon Hügel ausgelöst und wandert dann in Richtung Terminale entlang des Axons. Wenn in der Mitte eines Axons ein Aktionspotential ausgelöst wird, wandert es von dort aus in beide Richtungen (zum Soma und zur Terminale).) (Das Axon kann mit einer Zündschnur verglichen werden: wird die Zündschnur an einem Ende angezündet, läuft das Feuer nur in eine Richtung, wird sie in der Mitte angezündet, läuft das Feuer in beide Richtungen.) Bsp.: Zeit Axon Hügel Axon Anfang Axon (etwas weiter) Axon (noch weiter) 0 ­65mV ­65mV ­65mV ­65mV 1 +40mV ­65mV ­65mV ­65mV 2 +40mV ­35mV ­65mV ­65mV 3 ­65mV +40mV ­65mV ­65mV 4 ­65mV +40mV ­35mV ­65mV 5 ­65mV +40mV (Na+­Ka. inaktiv.) +40mV ­65mV 6 ­65mV ­35mV (Na+­Ka. inaktiv.) +40mV ­35mV 7 ­65mV ­80mV +40mV (Na+­Ka. inaktiv.) +40mV 8 ­65mV ­65mV ­35mV (Na+­Ka. inaktiv.) +40mV 9 ­65mV ­65mV ­80mV +40mV (Na+­Ka. inaktiv.) Eine Spannungsänderung am Axon Hügel (Zeitpunkt 1) löst ein Aktionspotential im Axon aus (Zeitpunkt 3). Das Aktionspotential am Anfang des Axons bewirkt eine Schwellwertüberschreitung etwas weiter im Axon (Zeitpunkt 4), die dort ein Aktionspotential auslöst (Zeitpunkt 5). Das Aktionspotential an der Stelle "etwas weiter" bewirkt eine Schwellwertüberschreitung (Zeitpunkt 6) am Anfang des Axons und an der Stelle "noch weiter" im Axon. Am Anfang des Axons kann jetzt aber kein Aktionspotential ausgelöst werden, weil die Na+­Kanäle noch nicht wieder im Ausgangszustand sind. An der Stelle "noch weiter" war bis jetzt das Ruhepotential und somit kann dort ein Aktionspotential ausgelöst werden. Wenn die Na+­Kanäle am Anfang des Axons wieder im Ausgangszustand sind (Zeitpunkt 8), ist an Stelle "etwas weiter" schon kein Aktionspotential mehr und das Aktionspotential an Stelle "noch weiter" ist zu weit weg um eine Wirkung auf den Anfang des Axons zu haben. Somit ist der Schwellwert am Anfang des Axons nicht mehr überschritten, sodaß dort natürlich das Ruhepotential bleibt (und kein Aktionspotential ausgelöst wird). Das selbe gilt für jede andere Stelle im Axon (wenn gerade ein Aktionspotential an der Stelle war, kann kein neues ausgelöst werden). Dadurch kann das Aktionspotential nur in eine Richtung wandern. Aufgrund der geringen elektrischen Isolationswirkung der Membran eines (unmyelinisierten) Axons wird das sich ausbreitende Potential schnell schwächer und ist schon nach kurzer Strecke (ca maximal einige 10 µm) nicht mehr stark genug um ein Aktionspotential auszulösen. Dadurch kann ein Aktionspotential nur in direkt benachbarten Teilen des Axons ein neues Aktionspotential auslösen. Durch Vergrößerung des Durchmessers des Axons wird der Querschnitt des Axon stärker vergrößert (propotional zu r2), als der Umfang (proportional zu r). Dadurch spielt die schlechte Isolation der Membran eine geringere Rolle (durch eine relativ geringere Fläche können weniger Ionen passieren und somit ist der Widerstand (die Isolation) relativ höher) und die positiven Ladungen (und damit das Potential) können sich weiter im Axon ausbreiten. Die Myelinscheide erhöht die Isolation der Membran um ein vielfaches und bewirkt so eine deutlich weitere Ausbreitung des Potentials. (Die Ausbreitung des Potentials entlang des Axons geht schnell, aber das Potential wird mit zunehmender Entfernung schwächer. Der Einstrom von Na+­Ionen an einer Stelle dauert eine Zeit, baut aber ein Aktionspotential fixer Größe auf. Je besser ein Axon isoliert ist, desto weniger wird das Potential entlang des Axons geschwächt, desto weiter können die Aktionspotentiale auseinander liegen und desto schneller wird das Signal weitergeleitet.) Ein Aktionspotential ist immer gleich stark, weil es an jeder Stelle des Axons (durch Fließen von 29 / 77 Neurobiologie Na+­Ionen durch die Membran) neu aufgebaut wird (aufgrund einer Schwellwertüberschreitung an dieser Stelle). Die Schwellwertüberschreitung wird durch eine Ausbreitung des Potentials innerhalb des Axons bewirkt, wobei diese Wirkung des Aktionspotentials auf benachbarte Gebiete mit zunehmender Entfernung schwächer ist. D.h. das Aktionspotential ist immer gleich stark, aber die Wirkung eines Aktionspotentials hat nur beschränkte Reichweite. Durch Aufbau eines neuen Aktionspotentials innerhalb dieser Reichweite kann ein Signal entlang des Axons wandern. (Streng genommen wandert nicht ein Aktionspotential entlang des Axons, sondern es entstehen entlang des Axons immer neu Aktionspotentiale, jeweils durch die Auswirkungen das Aktionspotentials direkt davor.) Ein Test für Multiple Sklerose (bei Multipler Sklerose wird die Myelinscheide zerstört) ist die Auslösung eine Aktionspotentials in einem langen Axon (z.B.: im Bein) und das Messen wann es nach einer bestimmten Strecke ankommt. Ist die Myelinisierung geschädigt (Demyelinisierung), so dauert es länger als bei einem gesunden Menschen. In unmyelinisierten Axonen (beim Menschen) werden Signale mit einer Geschwindigkeit von ca 1m pro Sekunde weitergeleitet. (Es gibt auch bei (gesunden) Menschen unmylinisierte Axone: z.B. bestimmte ("brennende") Schmerzen werden über unmyelinisierte Axone geleitet.) Die Myelinscheide verbessert die Isolation des Axons. Ranvier­Schnürringe35 befinden sich im Abstand von ca 2 mm. Nur bei den Ranvier­Schnürringen enthält die Membran des Axons spannungsabhängige Ionenkanäle. Somit kann auch nur bei den Ranvier­Schnürringen ein Aktionspotential auftreten. Durch die bessere Isolation kann sich ein Aktionspotential innerhalb des Axons von einem Ranvier­Schnürring bis zum nächsten auswirken und somit dort wieder ein Aktionspotential auslösen. Das Aktionspotential "springt" praktisch von einem Ranvier­Knoten zum nächsten. Diese Art der Erregungsleitung wird als saltatorische Reizleitung bezeichnet. Durch die Myelinscheide wird die Geschwindigkeit der Signalleitung von 1m/sec (unmyelinisiert, langsame Reizleitung) auf 120m/sec (saltatorischen Reizleitung) erhöht. Die Ionenkanäle (z.B.: der spannungsabhängige Na+­Kanal) sind wirklich Kanäle und keine Transporter. Durch einen Kanal können 108 Na+­Ionen pro Sekunde passieren, ein Transporter würde nur maximal 104 Ionen pro Sekunde schaffen. Die Ionenkanäle können durch Toxine selektiv blockiert werden (z.B. Tetrodotoxin blockiert nur Na+­Kanäle (aber nicht die K+­Kanäle)). Weiters können Toxine genutzt werden um einen gewünschten Ionenkanal zu reinigen. Z.B.: elektrische Fische haben eine hohe Konzentration an Ionenkanälen. Aus einer Lösung, die Bestandteile von Zellen dieser Fische enthält, können durch Bindung an ein spezifisches Toxin und Auftrennung über eine Säule genau die Ionenkanäle, an die das Toxin bindet, isoliert werden. TTX (Tetrodotoxin) und STX (Saxitoxin) sind Toxine, die spezifisch an Na+­Ionenkanäle binden. Tetrodotoxin ist das Gift des Kugelfischs. (Es sterben immer wieder Menschen, die einen Kugelfisch essen, bei dem das Gift nicht vollständig entfernt wurde.) Ein spannungsabhängiger Na+­Kanal besteht aus 4 transmembran­Domänen (von denen jede aus 6 α­Helices, die durch die Membran gehen, besteht). N­ und C­Terminus befinden sich beide in der Zelle. Der spannungsabhängiger Na+­Kanal ist ein einziges Protein. (Bilder siehe Folie 4.14) Jede der 4 Domänen enthält einen Finger (E: pore loop) (auf Folie 4.14 rot dargestellt), der von außen in den Kanal hinein geht und die Selektivität (für Na+) des Kanals bewirkt. Damit ein Na+­Kanal funktioniert, benötigt er bestimmte Eigenschaften. Die wichtigsten 35 Ranvier­Schnürringe werden auch als Ranvier­Knoten bezeichnet 30 / 77 Neurobiologie Bereiche/Eigenschaften sind: • Spannungssensor (E: voltage sensor): ist in α­Helix S4 jeder Domäne enthalten • Gate (Öffnungsmechanismus): öffnet den Kanal bei Depolarisation (­40mV) • Ionenselektivität: durch die Finger bewirkt; es können nur Na+­Ionen durch den Kanal durch • Inaktivierungsmechanismus: verschließt den Kanal bei positivem Potential (+40mV) oder kurze Zeit nach dem Öffnen + Ein Na ­Kanal verfügt über mehrere verschiedene Schließmechanismen (eine für das Verschließen bei Ruhepotential (Gate) und einer für das Inaktivieren (Verschließen) nach dem Öffnen bei positiver Spannung (Inaktivierungsmechanismus)). K+­Kanäle sind ähnlich aufgebaut, wie Na+­Kanäle. Sie gehören zur selben Protein­Familie. Es gibt viele verschiedene K+­Kanäle. Im Gegensatz zum Na+­Kanal besteht der K+­Kanal aus 4 einzelnen Proteinen, von denen jedes einer Domäne des Na+­Kanal­Proteins entspricht. Am N­Terminus (in der Zelle) ist ein positiver Bereich, der den Kanal verschießen kann. Beim Durchgang durch den Kanal (z.B. Na+­Kanal) verlieren die Ionen (z.B. Na+) ihre Hydrathülle. Die Selektivität des Kanals entsteht durch Carboxylgruppen(­COOH) und Hydroxygruppen (­OH), die den Na+­Ionen erlauben praktisch ohne Energieunterschied die Hydrathülle aufzugeben und den Kanal zu passieren. Synapsen Im Axon wird das Signal mittels elektrischer Reizleitung transportiert. Der elektrische Reiz (das Aktionspotential) geht aber nicht einfach über die Synapse in das nächste Neuron, sondern wird in der Synapse durch Transmittersubstanzen (Trägersubstanzen) übertragen. In der präsynaptischen Membran (d.h. der Membran des Axon­Endes) sind spannungsabhängige Ca ++ ­Kanäle (E: voltage­gated calcium channels). Wenn ein Aktionspotential im Axon­Ende (Terminale) ankommt, bewirkt die Depolarisation der Membran, daß sich diese Ca++­Kanäle öffnen. Nachdem die Ca++­Konzentration in der Zelle sehr niedrig (0.0002 mM) und der Konzentrationsunterschied von Ca++ zwischen außen und innen sehr groß (10000:1) ist, strömt Ca++ in die Zelle solange die Kanäle geöffnet sind und die Ca++­Konzentration in der Zelle erhöht sich deutlich (auf bis zu 0.1 mM in der Nähe der Ca++­Kanäle). Die erhöhte Ca++­Konzentration bewirkt die Exocytose der Neurotransmitter aus Vesikeln, die in der Zelle in der Nähe der Membran gelagert sind. Bei der Exocytose verschmilzt die Membran des Vesikels mit der Membran der Zelle und der Inhalt des Vesikels (hier: die Neurotransmitter) wird nach außen (hier: in den synaptischen Spalt) freigesetzt. Die Fusion der Vesikel mit der Membran läuft sehr schnell ab, sodaß es kaum eine Verzögerung (nur ca 0.2ms) zwischen dem Einströmen von Ca++ und der Freisetzung der Neurotransmitter gibt. (Damit es so schnell gehen kann ist es auch nötig, daß die Vesikel schon nahe der Membran gelagert sind.) Die spannungsabhängigen Ca++­Kanäle sind ähnlich aufgebaut wie die Na+­Kanäle. Sie gehören zur selben Protein­Familie. Ca++­Kanäle bestehen aus mehreren Untereinheiten, wobei die α1­Untereinheit die eigentliche Pore bildet (sie enthält alle 4 Domänen in einem Protein (wie der Na+­Kanal)). Weiters haben spannungsabhängige Ca++­Kanäle mehrere akzessorischen Untereinheiten (z.B.: β­Untereinheit, α2δ­Untereinheit, γ­Untereinheit), die ihre Eigenschaften beeinflussen (z.B. Modifikation der Spannungsabhängigkeit). 31 / 77 Neurobiologie Synaptische Transmission Der Einstrom von Ca++ in das Ende des präsynaptischen Axons bewirkt die Fusion von Neurotransmitter­gefüllten Vesikeln mit der Membran. Dadurch werden die Neurotransmitter aus den Vesikeln in den synaptischen Spalt (E: synaptic cleft) freigesetzt. Pro Aktionspotential fusionieren nur ein paar (ca 1­10) Vesikel mit der Membran. Die Vesikel befinden sich nur an bestimmten Stellen der Membran. Die Stellen, an denen die Vesikel an die Membran gebunden sind und an denen sie bei Ca++­Einstrom mit der Membran verschmelzen, heißen aktive Zonen (E: active zones) (oder docking stations). Die freigesetzten Neurotransmitter reagieren mit Rezeptoren in der Membran der postsynaptischen Zelle. (D.h. meist mit Rezeptoren in der Membran eines Spines eines Dendriten der postsynaptischen Zelle.) Es war lange umstritten (bis ca 1921), ob die Signalweitergabe über die Synapse chemisch oder elektrisch erfolgt. Otto Loewi (Professor in Graz) wies 1921 mit folgendem Experiment nach, daß es sich um eine chemische Übertragung handelt (und bekam dafür den Nobelpreis). (Die parasympathische Steuerung des Herzens erfolgt durch den Vagus­Nerv (Nervus vagus). Dieser Nerv verlangsamt den Herzschlag.) Loewi stimulierte den Nervus vagus eines (herausoperierten) Froschherzens. (Das Herz schlug daraufhin langsamer.) Dann gab er die Flüssigkeit, die das Herz umgab, zu einem anderen Froschherz, woraufhin auch dieses begann langsamer zu schlagen. D.h. das, was das langsamere Schlagen bewirkte, mußte in der Flüssigkeit und damit ein chemischer Stoff (und kein elekrisches Signal) sein. (Später stellte sich heraus, daß es sich bei diesem Stoff um Acetylcholin handelt.) Die Synapsen zwischen Nervus vagus und Herz sind also chemische Synapsen. (Inzwischen weiß man, daß es sowohl chemische als auch elektrische Synapsen gibt. Aber elektrische Synapsen kommen seltener vor.) Chemische Transmission Präsynaptisch Die Fusion eines Vesikels mit der Membran verläuft über den SNAP­SNARE­Mechanismus (SNAP36­SNARE37 mediierte Vesikelfusion) (E: SNARE­mediated lipid fusion). In der Membran der Zelle und der Membran des Vesikels sind SNAREs. SNAREs sind Proteine (der SNARE­Familie) und haben ein lipophiles Ende, das in der Membran verankert ist und ein (längeres) lipophobes Ende, das in das Cytosol hinausragt. In der Vesikelmembran sind v­SNAREs und in der Zellmembran sind t­SNAREs. Die lipophoben Enden von v­SNAREs und t­SNAREs38 können aneinander binden. Indem die SNAREs in Vesikelmembran und Zellmembran aneinander binden ziehen sie das Vesikel näher zur Zellmembran bis die beiden Membranen fusionieren. Der Mechanismus der Fusion der beiden Lipiddoppelschichten ist der selbe wie beim Verschmelzen von 2 Seifenblasen. (Bilder siehe Folie 5.6) Die Vesikel befinden sich schon an der docking Stelle der Membran. Wenn das Signal zum Verschmelzen mit der Membran (erhöhte Ca++­Konzentration) kommt, können sie sofort mit der Membran fusionieren. VAMP39 ist das v­SNARE in der Membran des Vesikels, SNAP­2540 und Syntaxin sind die t­ 36 SNAP steht für soluble NSF attachment protein (oder für synaptosome­associated protein); NSF = N­ethyl­maleimide­ sensitive fusion protein 37 SNARE steht für SNAP receptor 38 v­SNARE, t­SNARE: Das v steht für vesicle und das t für target. 39 VAMP steht für vesicle­associated membrane protein. VAMP ist eine anderer Name für Synaptobrevin. VAMP ist ein v­SNARE. 40 Das 25 in SNAP­25 steht für die molekulare Masse des Proteins: 25kDa. SNAP­25 ist ein t­SNARE. 32 / 77 Neurobiologie SNAREs in der Membran der Zelle. n­Sec141 ist an Syntaxin gebunden und VAMP kann erst an SNAP­25 und Syntaxin binden, wenn n­Sec1 entfernt wurde. Durch Ca++ wird die Bindung verstärkt, sodaß es zur Fusion kommt. (Zwei SNAP­25, ein VAMP und ein Syntaxin bilden ein Bündel aus 4 Helices, das die Fusion der Membranen bewirkt.) α­SNAP und NSF bewirken die Rückfaltung (Endocytose) des Vesikels in die Zelle nachdem die Neurotransmitter freigesetzt wurden. (Bilder siehe Folie 5.7) Der Lebenszyklus eines Vesikels mit Neurotransmittern besteht aus • Docking: binden an die Membran (in einer aktiven Zone) • Priming/Prefusion: erfordert ATP • Fusion mit der Zellmembran und Exocytose: erfolgt erst bei sehr hoher Ca++­Konzentration; der Neurotransmitter wird in den synaptischen Spalt freigesetzt • Endocytose: Zurückholen der Vesikelmembran aus der Membran der Zelle • Translokation von der Zellmembran weg: Transport zu einem Endosom • Fusion mit einem frühen Endosom • Abspaltung vom Endosom • Aufnahme der Neurotransmitter • Translokation zur Zellmembran: Transport zur Zellmembran Nach der Translokation kann das Vesikel entweder sofort wieder Neurotransmitter aufnehmen, oder zuerst mit einem Endosom verschmelzen. Wenn viele Aktionspotentiale ankommen, müssen Vesikel mit Neurotransmittern schnell nachgeliefert werden. (Neurotransmitter können im Soma hergestellt und zu den Synapsen transportiert werden oder direkt am Ende des Axons hergestellt werden oder aus der Synapse wieder aufgenommen werden.) Es gibt 3 verschiedene Möglichkeiten der Fusion und Neurotransmitterfreisetzung: • collapse: Das Vesikel verschmitzt vollständig mit der Zellmembran und entleert seinen gesamten Inhalt in den synaptischen Spalt. (Es ist heute aber bekannt, daß das Vesikel nicht vollständig mit der Membran verschmilzt, sondern (die Vesikel­Membran) nach dem Entleeren mittels Endocytose wieder rückgewonnen wird.) • kiss and run: Die Fusion mit der Zellmembran ist nur kurz. Es wird dabei nur ein kleiner Teil der Neurotransmitter aus dem Vesikel freigesetzt. Danach verläßt das Vesikel die docking­Stelle wieder (obwohl es noch nicht leer ist). • kiss and stay: Die Fusion mit der Zellmembran ist nur kurz. Es wird dabei nur ein kleiner Teil der Neurotransmitter aus dem Vesikel freigesetzt. Das Vesikel bleibt aber an der docking­Stelle (bis es leer ist) und kann beim nächsten Aktionspotential wieder Neurotransmitter freisetzen. Zusammenfassung der synaptischen Transmission: Im präsynaptischen Neuron werden die Neurotransmitter synthetisiert und kommen in Vesikel. Wenn ein Aktionspotential am Ende des Axons des präsynaptischen Neurons ankommt, werden die Transmitter aus dem Vesikel in den synaptischen Spalt freigesetzt. Die Transmitter reagieren mit den Rezeptoren in der Membran des postsynaptischen Neurons, wodurch sich das Potential in der postsynaptischen Zelle ändert. Die Neurotransmitter werden aus dem synaptischen Spalt entfernt. Z.B. können sie wieder in die präsynaptische Zelle aufgenommen werden und können dort wieder in Vesikel kommen. Einige Neurotransmitter (z.B.: GABA, Glutamat) werden von Transportern wieder in die Zelle und in Vesikel aufgenommen. Diese Transporter haben meist 12 (8­12) Transmembrandomänen und sind 41 n­Sec1 ist ein anderer Name für Munc18­1 33 / 77 Neurobiologie in der selben Familie wie ABC­Transporter42. Die Transporter sind spezifisch für bestimmte Neurotransmitter. Sie Energie für die Aufkonzentration des Neurotransmitters (d.h. für den Transport gegen den Konzentrationsgradienten des Neurotransmitters) stammt aus dem Cotransport von Ionen entlang des Konzentrationsgradienten der Ionen. Die Transporter in der Zellmembran und die in der Membran der Vesikel sind verschieden. Die Transporter in der Membran transportieren Na+ zusammen mit dem Neurotransmitter in die Zelle und die Transporter in der Vesikelmembran transportieren H+ aus dem Vesikel und gleichzeitig den Neurotransmitter in das Vesikel (Vesikel haben ATP getriebene H+­Pumpen, die die H+­Konzentration im Vesikel relativ hoch halten (saurer pH)) (d.h. die Transporter in der Vesikelmembran sind eigentlich Antiporter (und nicht Cotransporter wie die in der Zellmembran)). Postsynaptisch Aktionspotentiale haben immer die gleiche Gestalt und Größe. Aktionspotentiale gibt es nur in Axonen. Dendriten haben keine spannungsabhängigen Ionenkanäle und somit gibt es in Dendriten kein Aktionspotential. Wenn in einem Dendriten eine Depolarisation erfolgt, so kann diese kein Aktionspotential auslösen, sondern muß sich ohne weitere Verstärkung bis zum Soma (genau genommen bis zum Axon Hügel) ausbreiten, um eine Wirkung zu haben (d.h. um ein Aktionspotential im Axon auszulösen). Transmitter­aktivierte Ionenkanäle Transmitter­aktivierte Ionenkanäle werden auch als ionotrope Rezeptoren bezeichnet. In der Synapse befinden sich in der Membran der postsynaptischen Zelle Transmitter­aktivierte Ionenkanäle (meist Na+­Kanäle). Wenn ein Neurotransmitter an sie bindet öffnen sie sich, Na+ strömt in die postsynaptische Zelle und es kommt zur Depolarisation im Dendriten der postsynaptischen Zelle. Diese Depolarisation wird als (kleines) exzitatorisches postsynaptisches Potential (EPSP43) bezeichnet. Es ist im Vergleich zum Aktionspotential im präsynaptischen Axon, das die Ausschüttung der Neurotransmitter bewirkt hat, um ca 0.5 bis 1 ms verzögert und ist relativ schwach. Eine einzige solche Depolarisation reicht nicht für ein neues Aktionspotential im präsynaptischen Axon aus. Es sind mehrere Aktionspotentiale (nacheinander im selben präsynaptischen Axon oder ungefähr gleichzeitig in mehreren verschiedenen Axonen, die Synapsen mit der selben postsynaptischen Zelle haben) nötig um ein neues Aktionspotential im Axon der postsynaptischen Zelle auszulösen (d.h. um Reizweiterleitung zu bewirken). Bei vielen sensorischen Neuronen, beginnt das Axon nicht am Soma, sondern hat nur irgendwo in der Mitte eine Verbindung zum Soma. Die spike­generating zone ist nahe einem Ende des Axons und die Synapse ist am anderen Ende des Axons. Die spike­generating zone ist die Stelle an der das Aktionspotential (üblicherweise) ausgelöst wird. Bei Neuronen, bei denen das Axon am Soma beginnt, ist der Axon Hügel die spike­generating zone. Information über die Intensität eines Reizes wird durch die Frequenz der Aktionspotentiale übertragen. Je stärker der Reiz, desto höher die Frequenz. Information über die Dauer eines Reizes wird durch die Dauer des Schickens von Aktionspotentialen übertragen. D.h. je stärker der Reiz desto mehr Aktionspotentiale werden pro Zeiteinheit geschickt und es werden so lange Aktionspotentiale geschickt, so lange der Reiz vorhanden ist. Bsp.1: Ein geringer Reiz, der kurze Zeit andauert, kann z.B. 4 Aktionspotentiale bewirken, die zur 4­maligen Ausschüttung von Neurotransmittern in der Synapse führen. Bsp.2: Wenn der Reiz stärker ist, ist die Depolarisation stärker (z.B.: ­35mV (statt ­45mV wie in Bsp.1)) und dadurch werden leichter und somit öfter Aktionspotentiale ausglöst. D.h. es werden in 42 ABC­Transporter = ATP­binding cassette transporter 43 EPSP = excitatory postsynaptic potential 34 / 77 Neurobiologie der selben Zeit mehr Aktionspotentiale generiert. (Wenn der Reiz z.B. genauso lang dauert wie in Bsp.1, dann werden dennoch mehr Aktionspotentiale geschickt aber die Zeit zwischen dem ersten und letzten Aktionspotential des Reizes ist gleich geblieben wie bei Bsp.1.) (Bild siehe Folie 5.12) Ausser diesen erregenden Synapsen gibt es auch noch hemmende Synapsen. Wenn die postsynaptischen Transmitter­abhängigen Ionenkanäle Kanäle für Cl­ (statt für Na+) sind, dann wird durch das Öffnen der Kanäle das Zellinnere negativer (bis zu ­65mV). (Cl­ ist außen höher konzentriert als innen und somit fließen negative Ionen (Cl­) in die Zelle.) D.h. wenn im präsynaptischen Axon ein Aktionspotential in der Synapse ankommt, bewirkt dies (über die Neurotransmitter und die Cl­­Kanäle) letztlich eine Hyperpolarisation im Dendrit der postsynaptischen Zelle (falls das Potential vorher > ­65mV war). Diese vorübergehende Hyperpolarisation der postsynaptischen Membran wird als hemmendes postsynaptisches Potential (IPSP44) bezeichnet. Eine Hyperpolarisation löst keine weiteren Hyperpolarisationen aus (wie eine Depolarisation Aktionspotentiale auslöst) und die Hyperpolarisation durch Cl­ geht nur bis ­65mV und ist somit nicht wesentlich niedriger als das Ruhepotential. Ein IPSP kann also nur dann eine Auswirkung haben, wenn ca gleichzeitig eine Depolarisation erfolgt, die durch die Hyperpolarisation ausgeglichen oder zumindes abgeschwächt werden kann. D.h. ein IPSP hat nur zusammen mit einem ca gleichzeitigen EPSP (von einer anderen Synapse) eine Auswirkung (und zwar die, daß das EPSP geschwächt wird). (Z.B.: Wenn ein Muskel kontrahiert wird, muß der Gegenmuskel entspannt werden.) Alle Impulse, die ein Neuron ca gleichzeitig erreichen, werden in diesem aufsummiert und wenn die Gesamtdepolarisation am Axon Hügel ausreicht, wird ein Aktionspotential generiert. Es erfolgt sowohl räumliche Summierung (E: spatial summation) als auch zeitliche Summierung (E: temporal summation). Bei der räumlichen Summierung werden EPSP und IPSP, die über verschiedene Synapsen die selbe postsynaptische Zelle erreichen in dieser aufsummiert. Nachdem EPSP und IPSP eine kurze Zeit lang anhalten (einige Millisekunden), ist auch eine zeitliche Summierung möglich. Ein EPSP oder IPSP, das die Zelle erreicht solange noch etwas von einem vorhergehenden EPSP oder IPSP vorhanden ist, wird mit diesem (d.h. mit dem noch vorhandenen Rest) summiert. Die Informationsverarbeitung im Gehirn findet in jedem Neuron statt. Ob ein Aktionspotential im Axon des Neurons generiert wird, hängt von EPSPs, IPSPs und der Erregbarkeit des Neurons ab. Die EPSPs und IPSPs werden summiert und nur wenn die Summe höher als der durch die Erregbarkeit festgelegte Schwellwert ist, wird ein neues Aktionspotential erzeugt und somit der Reiz weitergeleitet. G­Proteine G­Protein gekoppelte Rezeptoren werden auch als metabotrope Rezeptoren bezeichnet. (Sie können auf den ganzen Metabolismus der Zelle wirken.) Die Neurotransmitter können in der postsynaptischen Membran nicht nur an Transmitter­aktivierte Ionenkanäle binden, sondern auch an G­Protein­gekoppelte Rezeptoren. Wenn ein Neurotransmitter an einen G­Protein­gekoppelten Rezeptor bindet, wird das zugehörige G­Protein aktiviert (indem GDP gegen GTP ausgetauscht wird) und das G­Protein dissoziiert in α­ Untereinheit und βγ­Untereinheit. Sowohl α­ als auch βγ­Untereinheit können Effektor­Proteine aktivieren. Diese Effektor­Proteine können G­Protein­abhängige Ionenkanäle oder Enzyme, die Second Messenger herstellen, sein. Die Wirkung von G­Proteinen ist langsamer, hält dafür aber 44 IPSP = inhibitory postsynaptic potential 35 / 77 Neurobiologie länger an, als die Wirkung von Transmitter­aktivierten Ionenkanälen. Durch G­Protein­abhängige Ionenkanäle kommen langsame EPSPs und langsame IPSPs zustande. (Im Gegensatz zu den raschen EPSPs und IPSPs, die durch die Transmitter­aktivierten Ionenkanäle verursacht werden.) G­Protein­aktivierte Enzyme stellen Second Messenger (z.B.: cAMP) her und diese können über weitere Proteine (PKA45, transcriptionsregulierende Enzyme, ...) viele Funktionen der Zelle beeinflussen. Sie können z.B. die Expression von Genen für Ionenkanäle erhöhen, sodaß mehr Ionenkanäle hergestellt werden und somit die Durchlässigkeit der Membran erhöht wird. Sie können auch auf existierende Ionenkanäle wirken (indem sie sie phosphorylieren). Der Kanal bleibt dann offen bis er wieder dephosphoryliert wird. (D.h. es ist ein eigenes Signal nötig um diese Ionenkanäle wieder zu schließen.) Dadurch können Neuronen reguliert werden, sodaß man sich auf etwas konzentrieren kann und unnötige Information aus der Umgebung abgeschottet wird. (Z.B. spürt man, wenn man steht, nicht die ganze Zeit sein eigenes Gewicht auf seinen Fußsohlen.) Second Messenger Systeme wirken in einer Kaskade (in jedem Schritt kann jedes Enzym viele andere des nächsten Schritts aktivieren, sodaß z.B. 1 Neurotransmitter auf viele Ionenkanäle wirken kann). Es kommt dadurch zu einer Verstärkung und Verbreiterung des Effekts. (Z.B.: ein Neurotransmitter aktiviert einen Rezeptor, dieser aktiviert mehrere G­Proteine; jedes G­Protein aktiviert mehrere Adenylyl Cyclasen; jede Adenylyl Cyclase stellt mehrere cAMP aus ATPs her; die cAMP aktivieren PKAs und jede PKA phosphoryliert mehrere Ionenkanäle) Die Erregbarkeit eines Neurons kann durch G­Protein­gekoppelte Rezeptoren (z.B.: langsame EPSPs und IPSPs) reguliert werden. Das ermöglicht die Konzentration auf bestimmte Tätigkeiten. Elektrische Transmission Elektrische Synapsen (über diese erfolgt die elektrische Transmission) bestehen aus Gap Junction Kanälen. In einer elektrischen Synapse sind die Membranen der präsynaptischen und der postsynaptsichen Zelle sehr nahe beisammen (ca 3nm). Ein Gap Junction Kanal besteht aus 2 Connexons (je 1 in jeder Zelle). Ein Connexon besteht aus 6 Connexins. (Connexins sind Proteine.) Durch Gap Junctions wird eine direkte Verbindung zwischen den Cytoplasmen der beiden Zellen hergestellt. Gap Junction Kanäle haben einen relativ großen Durchmesser und lassen viele verschiedene Ionen durch. Im Gegensatz zu chemischen Synapsen haben elektrische Synapsen keine Richtung. Egal in welcher der beiden Zellen eine Potentialänderung auftritt, sie wirkt sich direkt auf die andere Zelle aus. Durch eine elektrische Synapse fließen Ionen aus einer Zelle in die nächste. Die Übertragung ist daher sehr schnell. Elektrische Synapsen kommen auch im ZNS von Mammalia vor. Vermutlich sind sie nicht wichtig für die Weitergabe eines Signals auf nachfolgende Neuronen, sondern für die Verbindung von Neuronen, die parallel laufen und das selbe Signal übertragen. Transmittersysteme Im Gehirn kommen viele Substanzen vor, die keine Neurotransmitter sind, und es gibt Neurotransmitter, die auch (oder hauptsächlich) andere Aufgaben haben (z.B.: Glycin ist eine proteinogene Aminosäure aber auch ein Neurotransmitter). Wenn im Gehirn eine Substanz gefunden 45 PKA = protein kinase A 36 / 77 Neurobiologie wird, die ein Neurotransmitter sein könnte, muß überprüft werden, ob es sich wirklich um einen Neurotransmitter handelt. Nur wenn sie die Transmitterkriterien erfüllt, kann man sicher sein, daß sie ein Neurotransmitter ist. Transmitterkriterien: Jeder Neurotransmitter muß • im Gehirn vorkommen (muß im Neuron gespeichert werden) • im Gehirn synthetisiert werden (meist direkt im Neuron) • in Synapsen freigesetzt werden (muß bei Stimulation vom Neuron freigesetzt werden) • exzitatorische oder inhibitorische Wirkung haben • aus dem synaptischen Spalt eliminiert werden (wenn ein Neurotransmitter dauerhaft im synaptischen Spalt bliebe, würde das postsynaptische Neuron ständig stimuliert; andere Substanzen (die auf das postsynaptsiche Neuron keine Wirkung haben), können aber dauerhaft im synaptichen Spalt sein) • der endogenen Substanz in allem gleichen (wenn in einem Experiment die gefundene Substanz auf ein postsynaptisches Neuron gegeben wird, muß dieses genauso reagieren, wie wenn es durch das präsynaptsiche Neuron (durch Freisetzen des Neurotransmitters) stimuliert wird) Ein Neurotransmitter ist nur für eine bestimmte Art von Synapsen ein Neurotransmitter und für andere Synapsen (in denen das postsynaptische Neuron andere Rezptoren hat) nicht. Wichtige Transmittersubstanzen: Name und Struktur Antagonist46 Vorkommen Acetylcholin (ACh) Curare, Atropin in 5­10% der Synapsen im Gehirn (in neuromuskulären Verbindungen, in autonomen Ganglien47, in parasympathischen Nervenendigungen, im Nucleus caudatus) Noradrenalin48 (NE) Ergotamin (α), Propranolol (β) in 1­2% der Synapsen im Gehirn (in sympathischen Nervenendigungen, im unteren Hirnstamm, im Hypothalamus, in der Substantia nigra49) Holoperidol, Spiroperidol in 15% der Synapsen im Striatum50; sonst sehr wenig (in Corpus Striatum51 und Nucleus caudatus) Dopamin (DA) 46 Ein Antagonist blockiert die Wirkung des Neurotransmitters. (Ein Agonist ruft die gleiche Wirkung wie der Neurotransmitter hervor.) 47 Ein Ganglion ist ein Nervenknoten (eine Ansammlung von Neuronen im peripheren Nervensystem). Autonome Ganglien sind Ganglien des autonomen Nervensystems. 48 Noradrenalin heißt auf Englisch norepinephrine (USA) oder noradrenaline (GB). 49 Substantia nigra ist ein Teil des Mesencephalon 50 Striatum ist ein Teil des Corpus striatum 51 Das Corpus striatum ist Teil der Nuclei basales und diese sind im Inneren des Cerebrums. 37 / 77 Neurobiologie Serotonin (5­Hydroxytryptamin (5­HT)) Methergolin in 0.1­1% der Synapsen im Gehirn (im Hypothalamus und Nucleus caudatus52) Glycin (Gly) Strychnin in 25% der Synapsen im Rückenmark; sonst wenig (in Rückenmark, eventuell auch im Cerebellum) γ­Aminobuttersäure (GABA) Bicucullin in 25­30% der Synapsen im Gehirn (in Rückenmark, Cerebellum, Hirnrinde53) Glutamat (Glu) in 25% aller Synapsen im Gehirn Dopamin ist nicht nur selbst ein Neurotransmitter, sondern auch die Vorstufe von Noradrenalin (und Noradrenalin ist die Vorstufe von Adrenalin). Glycin kommt als Neurotransmitter im Rückenmark vor. Glycinerge Interneuronen im Rückenmark sind dafür verantwortlich, daß sich, wenn sich ein Muskel kontrahiert, der Gegenmuskel entspannt. Glycin und Glutamat sind nicht nur Neurotransmitter, sondern auch proteinogene Aminosäuren und kommen somit in allen Zellen vor (aber nur in wenigen als Neurotransmitter). GABA ist auch eine Aminosäure (NH2­Gruppe und COOH­Gruppe), wird aber nicht für den Aufbau von Proteinen verwendet. Auch Peptide können als Neurotransmitter wirken. Einige neuroaktive Peptide (wirken als Transmitter oder als Modulatoren) sind: • Peptide im Nervennetz des Magen­Darm­Trakts54 VIP (vasoaktives intestinales Peptid): ist vermutlich auch ein Neurotransmitter im Gehirn CCK­8 (cholecystokinin octapeptide): wurde ursprünglich in der Galle gefunden; heute weiß man, daß es mit Angst zu tun hat Substanz P: ist ein Neurotransmitter in Synapsen für die Schmerzempfindung Neurotensin Methionin Enkephalin 52 Nucleus caudatus ist ein Teil des Striatum 53 Hirnrinde = Cortex cerebri (graue Substanz an der Oberfläche des Cerebrums) 54 Der Magen­Darm­Trakt hat ein dichtes Nervennetz, das auf Englisch als "gut­brain" bezeichnet wird. 38 / 77 Neurobiologie Leucin Enkephalin Insulin Glucagon Hypothalmus Releasing­Hormone TRH (thyrotropin releasing hormone) LHRH (luteinisierendes Hormon Releasing Hormon) 55 Somatostatin Hypophysen­Peptide ACTH (adrenocorticotropes Hormon) β­Endorphin α­MSH (α­Melanocyte stimulating hormone) andere Peptide Angiotensin II Bradykinin Vasopressin Oxytocin Carnosine Bombesin • • • Auch Hormone (z.B.: Insulin) können Neurotransmitter sein. (Das ist ein Beispiel dafür, daß die Transmitterkriterien für neue Substanzen genau überprüft werden müssen. Insulin könnte im Gehirn als Hormon oder als Neurotransmitter oder als Regulator wirken.) Einige Peptid­Neurotransmitter sind Opioidpeptide (z.B.: Endorphine, Enkephaline). Opioidpeptide sind Liganden der Opioid­Rezeptoren (die auch als Opiatrezeptoren bezeichnet werden). Es gibt Synapsen, in denen mehrere Neurotransmitter vorkommen (die vom selben Axon­Ende freigesetzt werden). Es können 2 (oder mehrere) klassische Neurotransmitter sein, oder ein (oder mehrere) klassicher und ein (oder mehrere) Peptid­Neurotransmitter oder 2 (oder mehrere) Peptid­ Neurotransmitter. Es kann auch ein erregender und ein hemmender Neurotransmitter in der selben Synapse vorkommen. Einige Peptid­Neurotransmitter, die zusammen mit klassischen Neurotransmittern auftreten: klassischer Neurotransmitter Peptid­Neurotransmitter Acetylcholin VIP Noradrenalin Somatostatin Enkephalin Neurotensin Dopamin Cholecystokinin Enkephalin Adrenalin Enkephalin Serotonin Substanz P TRH (Vermutlich werden klassische Transmitter relativ rasch freigesetzt und wirken meist auf 55 Somatostatin kann mit SRIF (somatotropin release inhibiting factor) oder GHRIH (growth hormone release inhibiting hormone) abgekürzt werden. 39 / 77 Neurobiologie Transmitter­aktivierte Ionenkanäle und Peptid­Transmitter erst bei hoher Stimulation und wirken meist auf G­Protein gekoppelte Rezeptoren. (Das ist jedoch noch nicht endgültig bewiesen.)) Acetylcholin­System Das Acetylcholin­System heißt auch cholinerges System und Neuronen, die Acetylcholin freisetzen, heißen cholinerge Neuronen. Acetylcholin (ACh) ist ein Neurotransmitter: • der neuromuskulären Synapse: Acetylcholin ist der Transmitter zwischen Nerv und Muskel. (Acetylcholin ist die Substanz, die Otto Loewi bei seinem Versuch mit den beiden Froschherzen fand.) • der präganglionären Neuronen • des parasympathischen Systems Funktionen von Acetylcholin im ZNS: • Aufrechterhaltung des Bewußtseins • Sehbahn (colliculus superior, visueller Cortex) • Hörbahn (nucleus cochlearis) • Schmerzperzeption56 • Gedächtnis (Mangel an Acetylcholin spielt eine Rolle bei Alzheimer'scher Erkrankung) (Bild siehe Folie 6.7 (cholinerge Neuronen sind auf dem Bild gelb)) Acetylcholin­Neuronen gehen vom Rückenmark bis in die Nähe des Ziel­Gewebes und dann erfolgt eine Übertragung auf Neuronen, die direkt mit dem Ziel­Gewebe verbunden sind. Diese Neuronen, die das letzte Stück zum Ziel­Gewebe gehen, sind beim sympathischen System Noradrenalin­ Neuronen und beim parasympathischen System cholinerge Neuronen. (Beim sympathischen System liegen die Zellkörper in Ganglien. Von den Ganglien bis zum Gewebe verlaufen Noradrenalin­ Neuronen.) Acetylcholin wird (nach der Freisetzung in den synaptischen Spalt) durch das Enzym Acetylcholinesterase abgebaut. Acetylcholinesterasehemmer werden verwendet um Demenz (z.B.: bei Alzheimer­Krankheit) zu behandeln. 5­10% aller Synapsen im Gehirn setzen Acetylcholin frei. Cholinerge Neuronen (bzw. ihre Axone) gehen vom pontomesencephalotegmental complex57, von Nucleus septalis medialis58 und von Nucleus basalis Meynert59 aus in wichtige Teile des Gehirns. Vom pontomesencephalotegmental complex im Hirnstamm gehen sie in den Thalamus und das Vorderhirn60. Von Nucleus septalis medialis und Nucleus basalis Meynert im Vorderhirn gehen sie in den Neocortex61 und den Hippocampus62. Das Enzym Cholinacetyltransferase (ChAT) stellt Acetylcholin her, indem es eine Acteyl­Gruppe (CH3CO­) von einem Acetyl­CoA auf Cholin (HOCH2CH2(N+)(CH3)3) überträgt: 56 Perzeption = Reizaufnahme durch Sinneszellen 57 Der pontomesencephalotegmental complex (das Tegmentum pontis oder ein Teil davon) liegt im Pons und besteht aus Nucleus tegmentalis pedunculopontinus und Nucleus tegmentalis posterolateralis. 58 Nucleus septalis medialis (E: medial septal nucleus) liegt im basalen Telencephalon. 59 Nucleus basalis Meynert (E: basal nucleus of Meynert) liegt im basalen Telencephaon. 60 Das Vorderhirn (Prosencephalon) (E: forebrain) besteht aus Telencephalon (Cerebrum) und Diencephalon. 61 Der Neocortex (Isocortex) ist der jüngste und größte Teil (~90%) der Hirnrinde (Cortex cerebri (E: cerebral cortex)). 62 Der Hippocampus ist ein weiterer Teil des Cortex cerebri. Der Hippocampus liegt im Lobus temporalis. 40 / 77 Neurobiologie CH3CO­CoA + HOCH2CH2(N+)(CH3)3 ­> CH3CO­OCH2CH2(N+)(CH3)3 + CoA (ChAT kommt nur in Acetylcholin­Neuronen vor. Acetylcholin­Neuronen können daher daran erkannt werden, daß sie ChAT enthalten.) Cholin kann in den Neuronen nicht hergestellt werden, sondern muß aus der Umgebung der Zelle aufgenommen werden. Das Enzym Acetylchoinesterase (AChE) baut Acetylcholin ab, indem es Acetylcholin in Essigsäure (CH3COOH) und Cholin spaltet: CH3CO­OCH2CH2(N+)(CH3)3 +H2O ­> CH3COOH + HOCH2CH2(N+)(CH3)3 Die Neuronen setzen auch Acetylcholinesterase in den synaptischen Spalt frei. Dort baut es Acetylcholin ab. Das Neuron nimmt dann Cholin (und Essigsäure) wieder auf und stellt daraus neuerlich Acetylcholin her. Die Neuronen haben einen für Cholin spezifischen Transporter, der Cholin aus dem synaptischen Spalt (oder der Umgebung des Neurons) in die Zelle aufnimmt. Cholin wird aus der Umgebung des Neurons aufgenommen. Im Ende des Axons wird daraus von ChAT Acetylcholin hergestellt und durch einen Acetylcholin­Transporter in Vesikeln aufkonzentriert. Bei einem Aktionspotential wird es aus dem Vesikel in den synaptischen Spalt freigesetzt, bindet an ACh­Rezeptoren der postsynaptischen Zelle und wird schließlich von AChE in Cholin und Essigsäure gespalten. Das Cholin wird wieder in die Zelle aufgenommen. ... Acetylcholinesterasehemmer (z.B. in Medikamenten gegen Demenz) hemmen AChE, verlangsamen dadurch den Abbau von Acetylcholin und verlängern (d.h. verstärken) somit die Wirkung von Acetylcholin im synaptischen Spalt. Ähnliche Stoffe können auch als Gifte (z.B. Kampfgas) eingesetzt werden. Wenn AChE vollständig ausgeschaltet wird, wird ACh nicht mehr abgebaut und die postsynaptische Zelle wird dauerhaft stimuliert. In neuromuskulären Synapsen bewirkt das, daß der Muskel dauerhaft kontrahiert ist (die Folge ist Tod durch Atemlähmung). Eine Substanz, die die gleiche Wirkung hervorruft, wie ein Neurotransmitter, wird als Agonist des Neurotransmitters bezeichnet. Eine Substanz, die die Wirkung des Neurotransmitters verhindert, wird als Antagonist des Neurotransmitters bezeichnet. Ein Neurotransmitter kann auf mehrere verschiedene Rezeptoren wirken. Die verschiedenen Rezeptoren des selben Neurotransmitters werden als Subtypen des Neurotransmitter­Rezeptors bezeichnet. Es gibt Agonisten, die nur auf einen Rezeptor­Subtyp wirken. Die Rezeptor­Subtypen eines Neurotransmitters können mittels solcher Agonisten unterschieden werden (und werden meist nach dem Agonisten benannt). (Zu einem Neurotransmitter kann es mehrere verschiedene Rezeptoren geben. Aber ein Rezeptor gehört immer nur zu einem Neurotransmitter.) Acetylcholin wirkt auf 2 verschiedene Rezeptor­Subtypen: nikotinische Acetylcholin­Rezeptoren (nAChR) und muskarinische Acetylcholin­Rezeptoren (mAChR). Nikotin ist ein Agonist, der auf nikotinische Acetylcholin­Rezeptoren wirkt, aber nicht auf muskarinische. Curare ist ein Antagonist, der nur auf nikotinische Acetylcholin­Rezeptoren wirkt, aber nicht auf muskarinische. Muskarin (als Agonist) und Atropin (als Antagonist) wirken auf muskarinische Acetylcholin­ Rezeptoren, aber nicht auf nikotinische. (Acetylcholin wirkt natürlich als Agonist auf beide Subtypen.) 41 / 77 Neurobiologie Curare bindet wie Acetylcholin an nikotinische Rezeptoren, bewirkt aber keine Reaktion der postsynaptischen Zelle und verhindet, solange es gebunden ist, daß Acetylcholin an den Rezeptor bindet. Muskarin ist ein Gift des Fliegenpilzes (Amanita muscaria). Atropin ist das Gift der Tollkirsche (Atropa belladonna). Beide blockieren das parasympatische Nervensystem. Nikotinische Acetylcholin­Rezeptoren sind ionotrope Rezeptoren (Ionenkanäle). (Die Acetylcholin­ Rezeptoren der Skelettmuskeln sind nikotinische Rezeptoren.) Muskarinische Acetylcholin­Rezeptoren sind metabotrope Rezeptoren (G­Protein­gekoppelt). (Der Rezeptor im Herz beim Versuch von Otto Loewi ist ein muskarinischer Rezeptor.) Der nikotinische Acetylcholin­Rezeptor ist praktisch der Prototyp des Neurotransmitter­Rezeptors, weil er der erste Rezeptor war, dessen Struktur aufgeklärt wurde. Er besteht aus 5 Untereinheiten: zwei α­Untereinheiten, und je eine β­, γ­ und δ­ Untereinheit (die γ­Untereinheit ist in adulten Tieren durch die ε­Untereinheit ersetzt). (Bild siehe Folie 6.12) Wenn Acetylcholin an den nikotinischen Acetylcholin­Rezeptor bindet, vergrößert sich seine Permeabilität für Na+ und K+. Nachdem K+­Ionen auch im Ruhezustand die Membran passieren können, wirken sich vor allem die durchließenden Na+­Ionen aus, und die Folge ist ein EPSP. Nikotinischen Acetylcholin­Rezeptor sind erregende Rezeptoren. Nikotinische Acetylcholin­Rezeptoren sind nicht alle gleich, sondern unterscheiden sich je nach Art der Neuronen. Die niktotinischen Acetylcholin­Rezeptoren in der neuromuskuläre Endplatte bestehen aus 2 α1­ und je einer β1­, γ­(oder ε­) und δ­ Untereinheit. Sie haben die Agonisten Acetylcholin und Nikotin und die Antagonisten Curare und Bungarotoxin. Bungarotoxin ist das Gift der Schlange Bungarus multicinctus (eine Giftnatter). Die niktotinischen Acetylcholin­Rezeptoren in den autonomen Ganglien und im ZNS bestehen aus anderen α­und β­Untereinheiten (α2­α9, β2­β4) und enthalten keine γ­, ε­ und δ­ Untereinheiten (z.B. gibt es einen Rezeptor, der aus 2 α2­ und 3 β4­Untereinheiten besteht und einen anderen, der aus 5 α7­Untereinheiten besteht). Die niktotinischen Acetylcholin­Rezeptoren in den autonomen Ganglien haben die Agonisten 63 Acetylcholin und Nikotin und Antagonisten, die als Ganglienblocker bezeichnet werden (z.B.: Azamethonium). Auch bei den muskarinischen Aceytlcholin­Rezeptoren gibt es verschiedene: mR1, ... mR5. Muskarinische Rezeptoren kommen in Synapsen von Neuronen, die vom Parasympathikus auf das Organ wirken, vor (z.B. beim Herz). Weil muskarinische Acetylcholin­Rezeptoren über G­Proteine wirken, hält ihre Wirkung länger an als das Signal vorhanden ist. Muskarinische Aceytlcholin­Rezeptoren haben die Agonisten Acetylcholin und Muskarin, den Antagonist Atropin und indirekte Agonisten, die die Cholinesterase hemmen (z.B.: Physostigmin). (Physostigmin hat eine relativ kurze Halbwertszeit von ca 3 Stunden und sollte daher 3 (besser 4) mal am Tag genommen werden. Physostigmin wird verwendet um z.B. die Alzheimer'scher Krankheit zu behandeln.) (Viele dieser Agonisten und Antagonisten werden in Medikamenten verwendet.) Es gibt mehr als 1000 G­Protein gekoppelte Rezeptoren (GPCR). Bei vielen ist die Sequenz bekannt, aber man kennt noch keinen Agonisten ("orphan receptors"). Viele bilden Homo­ oder Heterodimere. Der Großteil der G­Protein gekoppelten Rezeptoren dürfte für das Geruchsempfinden 63 Ganglienblocker verhindern Übertragung in autonomen Ganglien des parasympathischen und symatischen Nervensystems. 42 / 77 Neurobiologie verwendet werden. G­Protein gekoppelte Rezeptoren haben 7 transmembran­Domänen. G­Protein gekoppelte Rezeptoren werden in 6 Familien (A ­ F) eingeteilt: • A (Rhodopsin­ähnliche): z.B.: Rhodopsin, Adrenoceptoren • B (Secretin Rezeptor Familie): z.B.: Neuropeptid Rezeptoren • C (metabotrope Glutamat­Rezeptoren und Pheromone): z.B.: mGluR (metabotrope Glutamat­ Rezeptoren) • D, E, F: 3 weitere Familien, die bei Pilzen oder Bakterien vorkommen Im Herzmuskel wird durch das G­Protein des muskarinischen Rezeptors ein K+­Kanal geöffnet, was bewirkt, daß das Herz dann langsamer schlägt: Wenn Acetylcholin an den muskarinischen Rezeptor bindet, wird im G­Protein GDT durch GTP ersetzt, die βγ­Untereinheit des G­Proteins wandert zu einem passenden K+­Kanal, bindet an ihn und der K+­Kanal öffnet sich. In vielen anderen Fällen aktiviert das G­Protein eines muskarinische Rezeptoren keinen Ionenkanal, sondern ein second messenger System. Noradrenalin­System Noradrenalin heißt auf Englisch norepinephrine (USA) oder noradrenaline (GB) und wird mit NE abgekürzt. Die Neurotransmitter Dopamin, Noradrenalin und Adrenalin werden aus der Aminosäure Tyrosin hergestellt und enthalten alle Brenzcatechin (E: Catechol) (1,2­Dihydroxybenzol). Diese 3 Neurotransmitter werden unter der Bezeichnung Catecholamine zusammengefaßt. Die Systeme dieser Neurotransmitter heißen auch catecholaminerges System. Noradrenalin ist ein Neurotransmitter: • des sympathischen Nervensystems • des Noradrenalin Systems im Gehirn Funktionen von Noradrenalin: • Aufrechterhaltung des Wachzustandes • Intellekt (Noradrenalin ist wichtig für Aufmerksamkeit und Konzentration) • Regulation anderer Systeme • Depressive Erkrankung (Mangel an Noradrenalin kann Depression bewirken) (Bild siehe Folie 6.17 (Noradrenlain­Neuronen sind auf dem Bild grün)) Noradrenalin­Neuronen des sympathischen Nervensytems gehen von den Ganglien bis zum Ziel­ Gewebe. Das Soma liegt dabei im Ganglion. (Die Übertrangung vom Rückenmark bis zu den Ganglien erfolgt auch beim symathischen Nervensystem mittels cholinerger Neuronen.) (Im parasympathischen Nervensystem kommen keine Noradrenalin­Neuronen vor.) Noradrenalin kommt im Gehirn vor allem im Locus caeruleus64 vor. (Der Locus caeruleus ist bläulich, weil Dopamin β­hydroxylase (das Enzym, das Noradrenalin synthetisiert) Kupfer enthält.) Von dort gehen Axone in den Großteil des Gehirns (in Lobus temporalis, Hypothalamus, Neocortex, Cerebellum und Rückenmark). Die Noradrenalin­Neuronen im Locus caeruleus haben eine erregende Wirkung auf die anderen Neuronen des Gehirns. Obwohl es nur wenige Neuronen sind, innervieren sie praktisch das ganze Gehirn (aufgrund ihrer starken Verzweigung). (Z.B.: eine Ratte hat nur ca 1500 Noradrenlin­Neuronen im Gehirn, aber diese innervieren das ganze Gehirn.) Dopamin, Noradrenalin und Adrenalin sind Catecholamine und haben die gleiche Biosynthese. Das Ausgangsmolekül ist Tyrosin. Tyrosin Hydroxylase (TH) hydroxyliert es zu L­DOPA (3,4­ 64 Der Locus caeruleus (alte Bezeichnung: Locus coeruleus) befindet sich im Pons. 43 / 77 Neurobiologie Dihydroxyphenylalanin). Dieser erste Schritt ist der geschwindigkeitsbestimmende. Hier findet auch die Regulation statt. L­DOPA wird dann vom Enzym Aromatische­L­Aminosäure­Decarboxylase (DOPA­ Decarboxylase) zu Dopamin (3,4­Dihydroxyphenylethylamin) decarboxyliert. Dopamin wird dann in Vesikel aufgenommen. Falls es weiterverarbeitet wird, erfolgt der nächste Schritt (zu Noradrenalin) im Vesikel. (D.h. auch das Enzym für diesen Schritt wird in das Vesikel aufgenommen.) In Noradrenlin­Neuronen wird Dopamin dann im Vesikel durch Dopamin β­Hydroxylase zu Noradrenalin (3,4­Dihydroxyphenyl­ethanolamin) hydroxyliert. In Adrenalin­Neuronen wird Noradrenalin dann durch Phenylethanolamin­N­methyltransferase zu Adrenalin65 ((R)­4­(1­Hydroxy­2­(methylamino)ethyl)benzen­1,2­diol) methyliert. Dopamin, Noradrenalin und Adrenalin werden wieder in die Zelle aufgenommen und in Vesikel transportiert (für die neuerliche Freisetzung) oder abgebaut. Die Inaktivierung (der Abbau) erfolgt entweder durch Methylierung der 3­Hydroxylgruppe des Catecholrings oder durch oxidative Entfernung der Aminoguppe oder beides hintereinander. Die Methylierung der 3­Hydroxylgruppe des Catecholrings wird vom Enzym COMT (catechol­O­ methyl transferase) katalysiert. Die Entfernung der Aminogruppe wird vom Enzym MAO (monoamin oxidase) katalysiert. Das Enzym Aldehyd­Dehydrogenase macht aus dem ­CHO, das von MAO statt der Aminogruppe eingesetzt wurde, ein ­COOH. Statt der Aldehyd­Dehydrogenase kann bei Noradrenalin und Adrenalin auch die Aldehyd­Reduktase, die aus dem ­CHO ein ­CH2OH macht, auf MAO folgen. COMT und MAO(inklusive Aldehyd­Dehydrogenase oder Aldehyd­Reduktase) wandeln Dopamin, Noradrenalin und Adrenalin in HVA66, VMA67 und MHPG68 um, wobei das Molekül sowohl zuerst von COMT und dann von MAO als auch in umgekehrter Reihenfolge verarbeitet werden kann. Dopamin wird in HVA abgebaut. Noradrenalin und Adrenalin werden in Vanillinmandelsäure (VMA) (wenn Aldehyd­Dehydrogenase nach MAO kommt) oder 3­Methoxy­4­ hydroxyphenylethylenglycol (MHPG) (wenn Aldehyd­Reduktase nach MAO kommt) abgebaut. (Strukturformeln und Zwischenprodukte siehe Folie 6.19) Es gibt 2 Gruppen von Noradrenalin­Rezeptoren: α­Rezeptoren und β­Rezeptoren. (Beide sind weiter in Subtypen unterteilt (z.B.: α1A, α1B, α2A, β1, β2).) Noradrenerge α­Rezeptoren vermitteln Kontraktion der glatten Muskulatur (z.B.: Blutgefäße, Uterus, ...). Agonisten sind (der Stärke nach geordnet) Adrenalin (Epi) > Noradrenalin (NE) > Phenylephrine (PE) > Isoproterenol (Iso). Antagonisten sind Phentolamin und Phenoxybenzamin. Noradrenerge β­Rezeptoren vermitteln Entspannung der glatten Muskulatur (z.B.: Bronchien). Agonisten sind (der Stärke nach geordnet) Isoproterenol (Iso) > Adrenalin (Epi) > Noradrenalin (NE) > Phenylephrine (PE). Antagonisten sind Propranolol und Dichloroisoproterenol. Herzprobleme, Blutdruck, ... sind durch Medikamente, die auf den Sympathikus wirken, beeinflußbar. Welche Wirkung Noradrenalin hat, hängt davon ab, welche Rezeptoren vorhanden sind. Wenn Noradrenalin auf verschiedene Gewebe getropft wird, kontrahieren sich einige und andere relaxieren sich. 65 66 67 68 Adrenalin heißt auf Englisch epinephrine (USA) oder adrenaline (GB). HVA = homovanillic acid VMA = vanilmandelic acid (vanillyl madelic acid) MHPG = 3­methoxy­4­hydroxy phenylglycol 44 / 77 Neurobiologie Noradrenerge Rezeptoren sind metabotrope Rezeptoren. Es gibt verschiedene G­Proteine, die verschiedene Wirkungen haben (z.B.: Aktivierung oder Inhibierung der Adenylyl Cyclase (wandelt ATP in cAMP um)). Je nachdem mit welchem G­Protein ein Rezeptor gekoppelt ist, hat die Bindung von Noradrenalin einen andere Wirkung auf die Zelle. Es sind heute 3 α1­, 3 α2­ und 3 β­Rezeptoren bekannt. Die α1­Rezeptoren bewirken eine Erhöhung von Inositol Triphosphat (IP3) und Diacylglycerol (DG) in der Zelle (diese bewirken eine Änderung der Ca++­Konzentration und diese bewirkt dann z.B. die Kontraktion). Die α2­Rezeptoren bewirken eine Verringerung der Menge an cAMP. (Bewirkt Relaxation der glatten Muskulatur.) Die β­Rezeptoren bewirken eine Erhöhung der Menge an cAMP und das bewirkt die Relaxation. (Im Herzmuskel bewirkt es Kontraktion.) Dopamin­System Dopamin wird mit DA abgekürzt. Es ist die Vorstufe des Noradrenalins bei dessen Biosynthese. Dopamin ist ein Neurotransmitter: • des Dopamin Systems im Gehirn Funktionen von Dopamin: • Es ist ein wichtiger Teil des Motivationssystems. • Schizophrenie (Schizophrenie kann mit hohen Dopamin Werten zusammenhängen) • Parkinson'sche Erkrankung (zur Behandlung von Parkinson'scher Erkrankung werden Medikamente verwendet, die Dopamin­Werte erhöhen (z.B.: Vorläufer in der Dopamin­Biosynthese)) Dopamin kommt im Gehirn vor allem in der Substantia nigra und im ventralen Teil des Tegmentums69 vor. Von dort gehen Axone in das Striatum und den Lobus frontalis. Die motorischen Störungen bei Parkinson'scher Krankheit werden durch das Absterben dopaminerger Neuronen in der Substantia nigra verursacht. Das ventrale Tegmentum ist für die Motivation wichtig. Es sind 5 Dopamin Rezeptoren bekannt: D1, D2, D3, D4 und D5. D1­ und D5­Rezeptor stimulieren die Adenylyl Cyclase, wenn Dopamin an den Rezeptor bindet. D2­, D3­ und D4­Rezeptor hemmen die Adenylyl Cyclase, wenn Dopamin an den Rezeptor bindet. Die meisten Neuroleptika (für die Behandlung von Schizophrenie) wirken auf D2, D3 und D4. Nur wenige wirken auf D1 und D5. Serotonin­System Serotonin wird mit 5­HT (5­Hydroxytryptamine) abgekürzt. Es ist nicht sehr häufig, hat aber wichtige regulatorische Funktionen. Serotonin ist ein Neurotransmitter: • des Serotonin Systems im Gehirn (0.1% bis 1% aller Synapsen verwenden Serotonin) Funktionen von Serotonin: • Biosynthese des Melatonins (Melatonin (N­acetyl­5­methoxytryptamin) wird vor allem bei Dunkelheit • • hergestellt und ist Schlaf­fördernd. Melatonin wird aus Serotonin hergestellt) Regulation vieler anderer Systeme (Stimmung, emotionales Verhalten, Schlaf, ...) Depressive Erkrankung (Mangel an Serotonin kann die Wahrscheinlichkeit für Depression erhöhen) 69 Das Tegmentum liegt im Mesencephalon (und zum Teil im Pons). 45 / 77 Neurobiologie Serotonin wird aus der Aminosäure Tryptophan synthetisiert. (Stukturformeln siehe Folie 7.4) Das Enzym Tryptophan Hydroxylase stellt aus L­Tryptophan das Zwischenprodukt 5­Hydroxy­L­ tryptophan (5­HTP) her (es wird eine ­OH­Gruppe angehängt). Aus diesem wird dann vom Enzym Aromatische L­Aminosäure Decarboxylase (E: L­aromatic amino acid decarboxylase) Serotonin hergestellt (es wird ­COOH entfernt). Serotonin wird zu 5­Hydroxyindolessigsäure (5­HIAA) abgebaut. Dabei wandelt MAO (monoamin oxidase) Serotonin in 5­Hydroxyindol­Acetaldehyd um und dieses wird von Aldehyd Dehydrogenase in 5­HIAA umgewandelt. Serotonin kann auch zu Melatonin weiter verarbeitet werden. Serotonin wird dabei vom Enzym N­ Acetylase in N­Acetylserotonin umgewandelt und dieses wird von Hydroxyindol­O­ Methyltransferase in Melatonin umgewandelt. Weiters kann Serotonin von Sulfotransferase in Serotonin­O­sulfat oder von N­Methyltransferase in Bufotenin umgewandelt werden. Tryptophan wird von den Neuronen aus der Umgebung aufgenommen (Tryptophan wird vom Darm aus der Nahrung aufgenommen und kommt mit dem Blut ins Gehirn und in die Flüssigkeit, die die Neuronen umgibt). Das Endprodukt des Serotonin­Abbaus ist 5­Hydroxyindolessigsäure (E: 5­hydroxyindoleacetic acid). Melatonin ist das Hormon der Zirbeldrüse (E: pineal gland). Die Produktion von Melatonin ist in der Nacht (im Dunkeln) viel stärker als am Tag (bei Licht). Es wird vermutet, daß Melatonin den Tag/Nacht­Rhythmus steuert. (Die Wirkung von Melatonin als Schlafmittel ist umstritten.) Serotonin kommt im Gehirn vor allem in den Nuclei raphes70 (Raphekerne) (E: raphe nuclei) vor. Von dort gehen Axone in das Rückenmark und praktisch das ganze Gehirn (Cerebellum, Lobus temporalis, Hypothalamus, Thalamus, Nuclei basales71 (E: basal ganglia), Neocortex). Jeder Nucleus raphes innerviert ein bestimmtes Gebiet (z.B.: nur das Rückenmark). Die serotonergen Rezeptoren werden in Familien (5­HT1 bis 5­HT7) unterteilt. Einige dieser Rezeptor­Familien werden weiter unterteilt (5­HT1A bis 5­HT1F, 5­HT2A bis 5­HT2C, 5­HT5A bis 5­ HT5B). Alle Serotonin­Rezeptoren außer 5­HT3 sind metabotrope Rezeptoren (also G­Protein gekoppelt). 5­HT3 ist ein Ionenkanal (Ligand­gesteuerter Na+/K+­Ionenkanal). GABA­System GABA ist die Abkürzung für γ­aminobutyric acid. GABA ist eine Aminosäure, aber keine α­ Aminosäure, sondern eine γ­Aminosäure (und somit natürlich auch keine proteinogene Aminosäure). GABA ist ein Neurotransmitter: • von 30% aller Synapsen im Gehirn • GABA ist der häufigste hemmende Neurotransmitter im ZNS Funktionen von GABA: • Kontrolle der Erregbarkeit des Gehirns 70 Die Nuclei raphes liegen in Medulla oblongata, Pons und Mesencephalon. 71 Die Nuclei basales (Basalganglien) sind Striatum (Nucleus caudatus und Putamen), Globus pallidus (und andere). Sie liegen im Cerebrum. 46 / 77 Neurobiologie • • • • Kontrolle des Wachzustands Kontrolle der Angst Kontrolle der Kognition Kontrolle von Lernen und Gedächtnis GABA wird aus Glutamat hergestellt. Das Enzym GAD (glutamic acid72 decarboxylase (Glutamat­ Decarboxylase)) wandelt Glutamat in GABA um. (Damit wird in einem einzigen Schritt aus dem wichtigsten exzitatorischen Neurotransmitter der wichtigste inhibitorische Neurotransmitter hergestellt.) Als Neurotransmitter wurde GABA entdeckt indem Aminosäuren (unter anderem Glutamat und GABA) auf Nerven gegeben wurden. Glutamat wirkte erregend und GABA hemmend. (Glutamat ist eine proteinogene Aminosäure und kommt daher in allen Zellen vor. Dies ist ein Beispiel dafür wie wichtig es ist, Kriterien zu haben mit denen beurteilt werden kann, ob es sich um einen Neurotransmitter handelt.) Muscimol kann wie GABA wirken und kann durch Bicuculline gehemmt werden. Baclofen kann wie GABA wirken und kann durch Phaclofen gehemmt werden. GABA hat mehr als einen Effekt: • Der erste Effekt kann durch Bicuculline inhibiert werden und wird als GABAA­Effekt bezeichnet. • Der zweite Effekt kann durch Phaclofen oder 2­OH­Saclofen inhibiert werden (aber nicht durch Bicuculline) und wird als GABAB­Effekt bezeichnet. Es gibt 2 Subtypen von GABA­Rezeptoren: GABAA­ und GABAB­Rezeptoren. Agonisten der GABAA­Rezeptoren sind: GABA, Muscimol, Isoguvacine Antagonisten der GABAA­Rezeptoren sind: Bicuculline Agonisten der GABAB­Rezeptoren sind: GABA, Baclofen (β­(p­chlorophenyl)­GABA) Antagonisten der GABAB­Rezeptoren sind: Phaclofen (β­(p­chlorophenyl)­3APPA), 2­OH­Saclofen (3­amino­2­(4­chlorophenyl)­2­hydroxy­propylsulphonic acid), (3APPA (3­aminopropylphosphonic acid) ist ein partieller Antagonist) (Strukturformeln siehe Folie 7.9) GABAA ist ein Cl­­Ionenkanal. GABAB ist ein metabotroper Rezeptor (und wirkt daher langsamer als GABAA). GABAB­Rezeptoren sind Heterodimere und bestehen aus den beiden Untereinheiten GBR173 und GBR274. GBR1 und GBR2 sind 7­transmembran­Proteine. GBR1 und GBR2 sind über eine Coiled­ Coil­Domäne (an der Zellinnenseite der Membran) miteinander verbunden. Die Untereinheit GBR1 ist an ein G­Protein gekoppelt. Die α­Untereinheit des G­Proteins inhibiert die Adenylyl Cyclase75. Die βγ­Untereinheit kann GIRKs76 (bestimmte K+­Kanäle) aktivieren oder Ca++­Kanäle inhibieren. Das Aktivieren (Öffnen) von K+­Kanälen bewirkt, daß K+ aus der Zelle fließt (weil die K+­ Konzentration in der Zelle höher ist als außen). Es kommt somit zu einer Hyperpolarisation. D.h. es führt zu einem IPSP. (Das Inhibieren (Schließen) von Ca++­Kanälen verhindern das Fließen von Ca++ von außen nach 72 73 74 75 76 glutamic acid = glutamate GBR1 = GABAB­Receptor 1 (auch abgekürzt mit GABAB1 oder GABBR1) GBR2 = GABAB­Receptor 2 (auch abgekürzt mit GABAB2 oder GABBR2) Adenylyl Cyclase wird mit AC abgekürzt. Sie wird auch (bzw. wurde früher) als Adenylat­Cyclase bezeichnet. GIRK = G­protein coupled inwardly rectifying potassium channels (potassium = K) 47 / 77 Neurobiologie innen (die Ca++­Konzentration ist außerhalb der Zelle höher als innen). Und trägt somit zum Erhalt des negativen Potentials im Inneren der Zelle bei. D.h. es führt zu einem IPSP.) GABAA­Rezeptoren, an die GABA gebunden ist, lassen Cl­ in die Zelle einströmen (die Cl­­ Konzentration ist außerhalb der Zelle höher als innen) und führen somit zu einem IPSP. Ob bei geöffneten Cl­­Kanälen Cl­ in die Zelle strömt und somit das Zellinnere negativer macht (d.h. inhibierend wirkt) oder aus der Zelle strömt und somit das Zellinnere positiver macht (d.h. exzitatorisch wirkt), hängt davon ab, ob die Cl­­Konzentration außerhalb der Zelle oder im Inneren höher ist. Bei der Entwicklung des Nervensystems (im Fötus) ist die Cl­­Konzentration in der Zelle höher als außen und somit wirkt GABA an GABAA­Rezeptoren während der Entwicklung des Nervensystems exzitatorisch. Da sowohl GABAA­ als auch GABAB­Rezeptoren bei Aktivierung ein IPSP hervorrufen, wirkt GABA immer (außer während der Entwicklung des Nervensystems) hemmend. GABAerge Neuronen sind somit (meist) inhibitorisch. GABAA­Rezeptoren sind ionotrope Rezeptoren. Sie gehören zur selben Super­Familie wie die nikotinischen Acetylcholin­Rezeptoren (und 5­HT3­Rezeptoren und Glycin­Rezeptoren) und bestehen aus 5 Untereinheiten. In den meisten Fällen sind es 2 α­, 2 β­ und 1 γ­Untereinheit. Es gibt 6 verschiedene α­Untereinheiten (α1 bis α6), 3 verschiedene β­Untereinheiten (β1 bis β3) und 3 verschiedene γ­Untereinheiten (γ1 bis γ3). Seltener kann eine β­ durch eine θ­Untereinheit und/oder die γ­ durch eine δ­, ε­ oder π­Untereinheit ersetzt sein. (Es gibt auch 3 verschiedene ρ­Untereinheiten (ρ1 bis ρ3). Diese bilden aber kein Heteropentamer mit den anderen Untereinheiten, sondern Pentamere, die aus 5 ρ­ Untereinheiten bestehen. Diese werden als GABAA­ρ Rezeptoren (früher GABAC­Rezeptoren) bezeichnet und sind auch Liganden­gesteuerte Cl­­Ionenkanäle.) GABAA­Rezeptoren sind die wichtigsten depolarisierenden (d.h. inhibitorischen) Neurotransmitter­ Rezeptoren. Mit Hilfe von Antikörpern gegen die Untereinheiten α1 ­ α6, β1 ­ β3, γ1 ­ γ3 und δ wurden Bilder der Verteilung dieser GABAA­Rezeptor­Untereinheiten im Rattenhirn erstellt. (Bilder siehe Folie 7.12) Z.B.: α1 kommt fast überall im Gehirn vor. α4 kommt nur in einem deutlich engeren Bereich vor. γ3 wird fast gar nicht exprimiert. Diese Untereinheiten haben eine spezifische regionale Verteilung. Es gibt viele Medikamente (und auch andere Moleküle), die an GABAA­Rezeptoren binden: • GABA • Benzodiazepine: sind relaxierende Medikamente (z.B. Valium (Diazepam) ist ein Benzodiazepin) (Benzodiazepine binden an einer anderen Stelle an GABAA als GABA und verstärken die Wirkung von GABA. D.h. durch Benzodiazepine wirkt GABA noch hemmender als normal. (Benzodiazepine sind positiv allosterische Modulatoren des GABAA­Rezeptors.)) • • • • • • • • • Barbiturate: sind Schlafmittel (und können auch zur Vollnarkose verwendet werden) Steroide: wirken ähnlich wie Benzodiazepine; (es gibt einen Zusammenhang mit Stimmungsschwankungen) Anästhetika (Narkotika) Picrotoxin Avermectin Loreclezole R05­4864 La3+ Zn++ 48 / 77 Neurobiologie Propofol Chlormethiazol • Ethanol: Alkohol wirkt als Agonist auf GABAA­Rezeptoren • γ­Butyrolactone • Furosemide Diese Substanzen sind allosterische77 Regulatoren des GABAA­Rezeptors. Sie können die Wirkung von GABA verstärken oder schwächen. • • Im präsynaptischen Neuron wird GABA von GAD aus Glutamat synthetisiert und über das Protein VGAT78 in der Vesikel­Membran in Vesikel aufgenommen. Nach der Freisetzung in den synaptischen Spalt bindet es an GABAA­ und GABAB­Rezeptoren der postsynaptischen Zelle. GABA wird über GAT79 in der Membran der präsynaptischen Zelle und von Glia­Zellen wieder aufgenommen und wieder in Vesikel gebracht oder (in Glia­Zellen) durch GABA­T80 abgebaut (zu Glutamin). Es können sich auch in der präsynaptischen Zelle (in der Terminale des Axons) GABAB­Rezeptoren befinden. Diese bewirken über das G­Protein die Inhibierung (Schließung) von Ca++­Kanälen und verringern somit die Freisetzung von Neurotransmittern, sodaß, wenn ein Aktionspotential ankommt, kein Neurotranmitter (oder zumindest weniger) in den synaptischen Spalt freigesetzt wird. Glutamat­System Glutamat ist eine proteinogene Aminosäure. Glutamat ist der bedeutendste erregende Neurotransmitter. Glutamat ist ein Neurotransmitter: • von 25% aller Synapsen im Gehirn • Glutamat ist der häufigste erregende Neurotransmitter im ZNS Funktionen von Glutamat: • Glutamat ist an vielen Funktionen beteiligt • Glutamat ist der Gegenspieler von GABA • Epilepsie (Glutamat löst Epilepsie aus und GABA reduziert Epilepsie) (Mikroinjetion von • Glutamat in ein Neuron löst spontane Depolarisation aus, wie sie bei Epilepsie vorkommt) Lernen und Gedächtnis (Glutamat spielt eine wichtige Rolle bei synaptischer Plastizität) • Neurodegeneration (Glutamat­Übererregung kann zu Neurodegeneration führen) (Hohe Glutamat­Konzentrationen in der Umgebung eines Neurons können zur Apoptose des Neurons führen. (Glutamat bewirkt das Öffnen von NMDA­Rezeptor­Kanälen, durch die Ca++ in die Zelle einströmt. Zu hohe Ca++­ Konzentration in der Zelle löst Apoptose aus.) Dieser Mechanismus ist an Schlaganfällen, Alzheimer'scher Krankheit, ALS (Amyotrophe Lateralsklerose), ... beteiligt.) Glutamat wird aus α­Ketoglutarat hergestellt. Das Enzym Glutamat Dehydrogenase wandelt α­Ketoglutarat, das aus dem Citrat­Zyklus stammt, in Glutamat um. Glutamat kann zu Glutamin weiter verarbeitet werden (vom Enzym Glutaminase). (Aus Glutamin kann auch wieder Glutamat hergestellt werden und daraus wieder α­Ketoglutarat.) Sowohl Glutamat als auch Glutamin sind im Zentrum vieler metabolischer Wege. Glutamat ist Ausgangsstoff für z.B. Glutathion, Polyamine, Harnstoff, GABA und Glutamin für z.B. Amino­Zucker, Glycoproteine, Nukleotide. 77 78 79 80 Allosterie bedeutet, daß das regulierende Molekül an anderer Stelle gebunden wird, als das Substrat. VGAT = vesicular GABA transporter GAT = GABA Transporter GABA­T = GABA­Transaminase 49 / 77 Neurobiologie Es gibt ionotrope und metabotrope Glutamat­Rezeptoren. Ionotrope Glutmat­Rezeptoren: • AMPA81­Rezeptoren: Tetramer aus den Untereinheiten GluR1 bis GluR4; (1 AMPA­Rezeptor besteht aus 4 Untereinheiten; bei den meisten sind es 2 GluR2 und 2 andere (GluR1, GluR3, GluR4) Untereinheiten) • NMDA82­Rezeptoren: Teramer aus den Untereinheiten NR1 und NR2A bis NR2D; (1 NMDA­ Rezeptor besteht aus 2 NR1­ und 2 NR2­Untereinheiten) Kainat­Rezeptoren: Tetramer aus den Untereinheiten GluR5 bis GluR7 und KA1, KA2 Metabotrope Glutmat­Rezeptoren: • mGluR: es gibt die 8 mGlu­Rezeptoren mGluR1 bis mGluR8 (AMPA, NMDA und Kainat sind Agonisten der jeweiligen Rezeptoren.) • NMDA­Rezeptoren lassen erst dann Ionen (Ca++, Na+ und K+) durch, wenn Glutamat gebunden ist und die Membran depolaristiert ist. Durch das Binden von Glutamat öffnet sich der Kanal. Bei einem Potential von ­65mV wird der Kanal aber sofort von Mg++­Ionen verstopft. Mg++ gibt den Kanal erst bei einem deutlich weniger negativem Potential (­30mV) frei und nur dann können Ca++, Na+ und K+ den Kanal passieren. D.h. NMDA­Rezeptoren sind Glutamat­abhängige und spannungsabhängigen Ionenkanäle. Damit ein NMDA­Rezeptor Ionen durchläßt, muß einerseits ein Aktionspotential im Axon­Ende des präsynaptischen Neurons ankommen und andererseits die postsynaptische Zelle (durch das Öffnen anderer Ionenkanäle (z.B.: AMPA­Rezeptoren) in der selben und in anderen Synapsen dieser postsynaptischen Zelle) soweit depolarisiert sein, daß Mg++ den NMDA­Kanal freigibt. D.h. Ca++­ Einstrom (und Na+­Einstrom und K+­Ausstrom) durch NMDA­Rezeptoren erfolgt nur, wenn präsynaptische Zelle und postsynaptische Zelle gleichzeitig depolarisiert sind (NMDA­Rezeptoren wirken somit als Koinzidenz­Detektoren). (Der Ca++­Einstrom bewirkt dann eine dauerhafte Verstärkung der Synapse. D.h. die Verbindungen zwischen Neuronen, die gleichzeitig aktiviert sind, werden verstärkt. Das ermöglicht assoziatives Lernen. (Z.B.: Beim Angreifen einer heißen Herdplatte werden die Neuronen, die die Information "Herdplatte" übermitteln und die Neuronen, die die Information "Schmerz" übermitteln, gleichzeitig aktiviert und daher stärker verbunden, sodaß man danach bei Herdplatte auch sofort an Schmerz denkt.)) Peptid­Systeme Es gibt nicht nur klassische Neurotransmitter, sondern auch Peptide, die als Neurotransmitter wirken. Für eine Liste einiger Peptid­Neurotransmitter siehe Seite 38. Einige Peptide spielen auch im ZNS als Neurotransmitter eine Rolle. Die meisten davon haben regulatorische Funktionen. Für eine Tabelle mit einigen Beispielen, welche Neurotransmitter gemeinsam in einer Synapse vorkommen, siehe Seite 39. (Dale's Gesetz, daß jede Synapse nur einen Neurotransmiter hat, ist daher falsch.) Es werden mehrere Signale über den selben Weg (über die selbe Synapse) übertragen. Es gibt sogar Synapsen in denen sowohl Glutamin als auch GABA vorkommt. Peptid­Neurotransmitter werden im Soma aus Vorstufen synthetisiert und dann in Vesikeln zur 81 AMPA = α­amino­3­hydroxy­5­methyl­4­isoxazole propionate 82 NMDA = N­methyl­D­aspartate 50 / 77 Neurobiologie Synapse transportiert. (Die Synthese erfolgt im ER (Vorstufen) und Golgi­Apparat (Umwandlung in aktive Neurotransmitter). Da es im Axon weder ER noch Golgi­Apparat gibt, muß die Synthese im Soma erfolgen.) (Viele klassische Neurotransmitter werden direkt im Axon­Ende hergestellt und nur die Enyme, die diese Neurotransmitter herstellen, werden im Soma produziert.) Peptid­Neurotransmitter werden nach der Freisetzung in die Synapse angebaut und müssen daher immer neu synthetisiert werden. (Viele klassische Neurotransmitter werden wieder in die präsynaptische Zelle aufgenommen und können wiederverwendet werden.) Weil Peptid­Neurotransmitter immer neu synthetisiert und vom Soma zum Axon­Ende transportiert werden müssen, sind sie für das Neuron viel aufwändiger als klassische Neurotransmitter und können somit nur regulatorisch sein. Sie werden nur bei besonders starker Erregung freigesetzt. Peptid­Neurotransmitter wirken auf G­Protein gekoppelte Rezeptoren. Über die Pepetid­Systeme ist noch wenig bekannt. Bedeutung der Peptid­Systeme: • vielfältige Regulationsfunktionen • Schmerz (Substanz P) • Regulation des Schmerzempfindens (Opiate) • Motivationssystem (Opiate) • Angst (Cholecystokinin System) Funktion der Sinne Es gibt Sinne für • Exterozeption: Wahrnehmung der Außenwelt; (die klassischen 5 Sinne und der Gleichgewichtssinn) Sehen Hören Riechen Schmecken Tasten Gleichgewicht • Propriozeption: Wahrnehmungen aus dem Bewegungsapparat und von der Körperoberfläche Muskellänge und ­spannung Gelenkstellung Schmerz • Interozeption: Wahrnehmung aus inneren Organen Chemosensoren Osmosensoren Barosensoren (Blutdruck­Sensoren) ... Sensoren sind entweder Neuronen oder andere Zellen, die Neuronen erregen. Eine primäre Sinneszelle ist ein Neuron, von dem ein Teil (z.B.: ein Stück des Axons) der Sinneswahrnehmung dient. Eine sekundäre Sinneszelle ist eine Zelle, die kein Neuron ist, sondern nur der Sinneswahrnehmung dient. Die wahrgenommene Information wird über eine Synapse an ein Neuron weitergegeben. 51 / 77 Neurobiologie Die Sinneszellen sind Rezeptoren (Detektoren) für chemische und physikalische Reize: Chemorezeptoren: chemische Substanzen; (z.B.: Geruch, Geschmack, CO2­Konzentration im Blut) Mechanorezeptoren: Berührung, Schall; (z.B.: Tasten, Gehör) Photorezeptoren: Licht (z.B.: Sehen) Thermorezeptoren: Temperatur; (z.B.: Temperatursensoren in der Haut) Elektrorezeptoren: elektrisches Feld; (z.B.: bei einigen Fischen) Magnetorezeptoren: magnetisches Feld; (z.B.: bei Zugvögeln und Insekten) Mechanorezeptoren Es gibt viele Mechanorezeptoren in der Haut: • Merkel­Tastscheiben (E: Merkel's Disk) • freie Nervenenden (mit mechanosensitiven Ionenkanälen) • Meissner­Körperchen (E: Meissner's Corpuscle) • Haarfollikelrezeptoren (E: Hair follicle receptor) • Vater­Pacini­Körperchen83 (E: Pacinian corpuscle) • Ruffini­Körperchen (E: Ruffini's ending) Alle diese Mechanorezeptoren enthalten unmyelinisierte, verzweigte Axone mit mechanosensitiven Ionenkanälen. Ob sich diese Ionenkanäle öffnen, hängt von der Dehnung der umgebenden Membran ab. Die Haare sind Teil des sensorischen Systems. Haarfollikelrezeptoren sind Neuronen, die die Haare (bzw. deren Wurzel) umwickeln. Verschiedene Mechanorezeptoren haben verschiedene bevorzugte Stimulus Frequenzen und Druckstärken. Dadurch können verschiedene mechanische Empfindungen wahrgenommen werden (z.B.: Vibration, Druck, Zug an einem Haar, Verbiegen eines Haares, ...). Vater­Pacini­Körperchen nehmen vor allem Vibrationen (d.h. Veränderungen im Druck) mit Frequenzen im Bereich von 200Hz bis 300Hz wahr und Meissner­Körperchen Vibrationen von 50Hz. Merkel­Tastscheiben und Ruffini­Körperchen erzeugen so lange ein Signal, wie der Druck andauert. Vater­Pacini­Körperchen erzeugen ein Signal, wenn sich der Druck ändert, aber nicht wenn er gleich bleibt. (Bild siehe Folie 8.7) Ein Vater­Pacini­Körperchen besteht aus einem Axon und einer Hülle aus mehreren Schichten Bindegewebe. Wenn auf ein intaktes Vater­Pacini­Körperchen gedrückt wird, wird im Axon ein Aktionspotential erzeugt. Wenn ohne Änderung weiter gedrückt wird, nimmt dennoch die Depolarisation ab und es wird kein weiteres Aktionspotential erzeugt. Erst wenn wieder ausgelassen wird, erfolgt eine erneute Depolarisation und damit ein Aktiononspotential. Wenn die Hülle entfernt und der Druck direkt auf das Axon ausgeübt wird, dauert die Depolarisation so lange an wie der Druck und endet erst, wenn wieder ausgelassen wird. Für die Wahrnehmung von Druck ist das Axon ausreichend (auch ohne Hülle erfolgt eine Depolarisation) aber für die Wahrnehmung von Vibration ist die Hülle notwendig. Wenn die Hülle vorhanden ist, verteilt sich der Druck schnell gleichmäßig über die ganze Hülle und ist daher nach kurzer Zeit um das ganze Axon gleich. Wenn Druck auf das Vater­Pacini­Körperchen ausgeübt wird, besteht zu Beginn kurz ein Druckunterschied zwischen der Stelle der Druckausübung und anderen Stellen des Axons (die Membran wird verformt und die Ionenkanäle öffnen sich). Dann verteilt sich der Druck gleichmäßig, die Membran des Axons ist nicht mehr verformt und somit schließen sich die Ionenkanäle wieder. Beim Auslassen besteht wieder für kurze 83 Vater­Pacini­Körperchen sind nach Abraham Vater und Filippo Pacini benannt. 52 / 77 Neurobiologie Zeit ein Druckunterschied. Das ermöglicht die Art und Qualität der Berührung zu empfinden. Geräusche werden über mechanosensitive Schallrezeptoren in der Cochlea (Hörschnecke im Innenohr) wahrgenommen. Die Cochlea ist flüssigkeitsgefüllt. Wenn Schall auf das Ohr trifft, wird diese Flüssigkeitssäule in Schwingungen versetzt. Diese Schwingungen werden von Haarzellen mittels Stereocilia wahrgenommen. In der Endolymphe (die Flüssigkeit, die die Stereocilia umgibt) ist die Konzentration von K+ höher als in den Zellen (und damit als in den Stereocilia), sodaß bei Öffnung von K+­Kanälen K+ in die Zelle strömt (statt aus dieser heraus) und damit zur Depolarisation der Zelle führt. (In der Endolymphe ist die Konzentration an K+ 150mM und an Na+ 1mM (normalerweise ist die Konzentration von K+ außerhalb der Zellen ca 5mM).) Jeder Kanal ist mit dem vorhergehenden Cilium verbunden. Wenn die Cilia gerade stehen, ist der Kanal wenig geöffnet. Wenn sie in eine Richtung gebogen sind, ist er geschloßen, wenn sie in die andere Richtung gebogen sind, ist er weiter offen. Dadurch strömt je nach Biegung der Cilia mehr oder weniger K+ in die Zelle. Die Depolarisation der Haarzelle bewirkt, daß sich Ca++­Kanäle öffnen, die wie bei Neuronen die Freisetzung eines Neurotransmitters auslösen. Je nachdem an welcher Stelle der Cochlea sich eine Haarzelle befindet, wird sie bei einer anderen Frequenz angeregt. (Bild siehe Folie 8.8 (TRPA1: TRP = transient receptor potential (Rezeptor­Familie); A steht für Ankyrin (Rezeptor­ Subfamilie))) Thermorezeptoren Ein Temperaturrezeptor ist Neuron, das bestimmte Ionenkanäle enthält. Diese Ionenkanäle gehören zur Familie der TRP84­Kanäle. Die meisten Kanäle dieser Familie (und alle 6 bisher bekannten Temperaturrezeptor­Kanäle) haben 6 Transmembrandomänen. Die Temperaturrezeptor­Kanäle sind TRPA185 (ANKTM1), TRPM886, TRPV387, TRPV4, TRPV1 (VR1) und TRPV2 (VRL1). Diese Kanäle sind für verschiedene Temperaturbereiche sensitiv (z.B.: TRPA1 für ca 10°C bis 18°C (ein Kälterezeptor; produziert umso mehr Aktionspotentiale, je kälter es ist), TRPV4 für ca 29°C bis 45°C (ein Wärmerezeptor; produziert umso mehr Aktionspotentiale, je wärmer es ist)). Aus der Mischung der Informationen von den verschiedenen Temperatursensoren kann das Gehirn die Temperatur ermitteln. (Bild siehe Folie 8.9) (Das Temperaturempfinden des Menschen ist eher schlecht.) Die Thermorezeptoren können auch durch Chemikalien ausgelöst werden. Capsaicin (in Paprika und Chili) aktiviert TRPV1 und löst damit Wärmeempfinden aus. Menthol (in Pfefferminze) aktiviert TRPM8 und löst damit Kälteempfinden aus. Chemorezeptoren Geschmacksrezeptoren Es gibt 5 Geschmacksqualitäten: • salzig (Na+­kanäle): erkennt Na+ • sauer (H+­Kanäle): erkennt H+ (d.h. niedrigen pH) 84 85 86 87 TRP = transient receptor potential TRPA = TRP ankyrin (eine Subfamilie der TRP­Kanäle) TRPM = TRP melastatin (eine Subfamilie der TRP­Kanäle) TRPV = TRP vanilloid (eine Subfamilie der TRP­Kanäle) 53 / 77 Neurobiologie bitter (G­Protein) süß (G­Protein) • umami88 (G­Protein): erkennt Glutamat Alle Geschmacksrezeptoren sind sekundäre Sinneszellen. • • (Bilder siehe Folie 8.11) Geschmackszellen für salzig haben Na+­Kanäle, die immer offen sind (Amilorid­sensitive Na+­ Kanäle89). Wenn die Na+­Konzentration außerhalb (d.h. im Mund) hoch ist, fließen Na+­Ionen in die Zelle. Das führt zur Depolarisation. Durch diese (leichte) Depolarisation öffnen sich spannungsabhängige Na+­Kanäle und die Depolarisation wird soweit verstärkt, daß sich spannungsabhängige Ca++­Kanäle öffnen. Der Ca++­Einstrom bewirkt, wie in Neuronen, die Freisetzung eines Neurotransmitters. Geschmackszellen für sauer funktionieren ähnlich wie Geschmackszellen für salzig. H+ fließt durch Amilorid­sensitive Na+­Kanäle in die Zelle (das alleine würde es nicht erlauben salzig und sauer zu unterscheiden) und blockiert zusätzlich K+­Kanäle. Beides trägt zur Depolarisation bei. Geschmackszellen für bitter, süß und umami enthalten in der Membran G­Protein gekoppelte Rezeptorproteine. Es gibt eine Art von Rezeptorprotein für süß, eine Art für umami und ca 30 Arten für bitter. Jede Geschmackszelle enthält entweder das süß­Rezeptroprotein oder das umami­ Rezeptorprotein oder bitter­Rezeptorproteine. Der weitere Weg ist für alle 3 Arten von Geschmackszellen gleich. Wenn ein Geschmacksstoff an das Rezeptorprotein bindet, wird das G­Protein aktiviert. Dieses aktiviert die Phospholipase C, die daraufhin PIP290 in IP391 umwandelt. IP3 aktiviert spezielle Na+­Kanäle, sodaß Na+ in die Zelle strömt und eine Depolarisation bewirkt. Die Depolarisation bewirkt das Öffnen von Ca++­Kanälen. Weiters kann IP3 die Freisetzung von Ca++ aus intrazellulären Speichern ins Cytosol auslösen. Ca++ bewirkt dann wieder die Freisetzung eines Neurotransmitters, der ein Neuron aktiviert. Es gibt nur ein Rezeptorprotein (das aus den 2 Untereinheiten T1R2 und T1R3 besteht) für süß. An dieses können jedoch viele verschiedene Substanzen binden (die somit alle süß schmecken). Es gibt auch nur ein Rezeptorprotein für umami. Dieses besteht aus den 2 Untereinheiten T1R1 und T1R3. Es gibt ca 30 verschiedene Rezeptorproteine für für bitter (die alle nur aus einer Untereinheit (aus der T2R­Familie) bestehen). Auf einer bitter­Geschmackszelle befinden sich aber viele dieser Rezeptoren, sodaß eine bitter­Geschmackszelle von vielen verschiedenen Substanzen gleichartig aktiviert wird. D.h. alle diese verschiedenen Substanzen schmecken bitter, ohne weitere Unterscheidung. Viele giftige Substanzen sind bitter. Um diese verschiedenen giftigen Substanzen wahrzunehmen sind verschiedene Rezeptorproteine nötig. Die wichtige Information ist aber nur, daß es sich um eine giftige Substanz handelt und nicht um welche davon. Die Geschmacksqualitäten (salzig, sauer, bitter, süß und umami) können vom Gehirn unterschieden werden, weil jede Geschmackszelle nur eine davon wahrnimmt und von jeder Geschmackszelle 1 Neuron ausgeht. Der Gesamtgeschmack der Substanzen im Mund (z.B.: eines Nahrungsmittels) setzt sich aus diesen 5 Geschmacksqualitäten zusammen. D.h. die Mischung aus diesen Empfindungen erlaubt es dem Gehirn den Geschmack (z.B.: eines Nahrungsmittels) zu identifizieren. Die 88 89 90 91 umami kommt vom japanischen Wort für köstlich diese Na+­Kanäle werden durch Amilorid blockiert PIP2 = phosphatidylinositol­4,5­bisphosphate IP3 = inositol triphosphate (ein intrazellulärer Botenstoff) 54 / 77 Neurobiologie Geschmackswahrnehmung erflogt also mittels population coding. (Population coding ist die Repräsentation von Information durch die Kombination der Aktivität vieler Neuronen.) Geruchsrezeptoren Die Geruchsrezeptoren befinden sich ganz oben in der Nasenhöhle an der Siebplatte92 (E: cribriform plate). Auf der Siebplatte (d.h. auf der Seite des Gehirns) befinden sich die Bulbi olfactorii (E: olfactory bulbs). An der Unterseite der Siebplatte (also in der Nasenhöhle) ist das Epithelium olfactorium (Riechepithel) (E: olfactory epithelium). (Die 3 Hauptzelltypen des Epithelium olfactorium sind Riechsinneszellen (olfactorische Neuronen) (E: olfactory receptor cells), Stützzellen (E: supporting cells) und Basalzellen (E: basal cells).) (Bild siehe Folie 8.14) Auf dem Epithelium olfactorium befindet sich immer eine Schleimschicht (E: mucus layer). Geruchssubstanzen lösen sich in dieser Schleimschicht und regen die aus dem Epithelium olfactorium in den Schleim hineinragenden Cilien der Riechsinneszellen an. Die Nerven (d.h. die Axone der Riechsinneszellen) gehen durch die Siebplatte in die Bulbi olfactorii. (Bei einem starken Schlag gegen den Kopf können sie reißen, was den Verlust des Geruchssinns zur Folge hat.) (Die "Geschmacksempfindung" besteht zum Teil auch aus dem Geruch des Nahrungsmittels. (Bei einem Schnupfen scheinen Nahrungsmittel weniger Geschmak zu haben, obwohl nur der Geruchssinn und nicht der Geschmackssinn beeinträchtigt ist.)) Geruchssinneszellen sind primäre Sinneszellen (d.h. Neuronen). Wenn Geruchsstoffe (E: odorant molecules) an Geruchsrezeptorproteine (E: odorant receptor protein) binden, aktiviert das Geruchsrezeptorprotein ein Golf­Protein93. Das Golf­Protein aktiviert daraufhin die Adenylyl Cyclase, was die Produktion von cAMP zur Folge hat. Das cAMP bindet an spezielle Kationenkanäle, die sich dadurch öffnen und Na+ und Ca++ in die Zelle strömen lassen. Ca++ bewirkt das Öffnen von Ca++­abhängigen Cl­­Kanälen durch die Cl­ aus der Zelle strömt. Kationeneinstrom (Na+, Ca++) und Anionenausstrom (Cl­) bewirken eine Depolarisation. Damit wird das sensorische Signal in ein elektrisches umgesetzt. Das Rezeptorpotential, das durch Kationeneinstrom (Na+, Ca++) und Anionenausstrom (Cl­) im Dendriten, an dem sich die Cilien mit den Rezeptorproteinen befinden, entsteht, wirkt sich bis ins Soma aus und verursacht dort Aktionspotentiale, solange es besteht. Diese Aktionspotential breiten sich über das Axon aus. Es gibt über 1000 G­Protein gekopelte Rezeptorproteine. Viele davon sind Geruchsrezeptorproteine. Es gibt also sehr viele verschiedene Geruchssensoren. (Im Gegensatz zu bitter­Geschmacksrezeptorzellen enthält jede Riechsinneszelle nur wenige (oder nur 1) verschiedene Geruchsrezeptorproteine und somit gibt es viele verschiedene Riechsinneszellen.) Jede Geruchssinneszelle exprimiert nur einen Typ von Geruchsrezeptorprotein. An ein Geruchsrezeptorprotein können aber viele verschiedene Moleküle binden. Allerdings lösen verschiedene Molelüle unterschiedlich starke Reaktionen in der Zelle aus (d.h. verschiede Anzahl (bzw. Frequenz) an Aktionspotentialen). Die verschiedenen Geruchsinneszell­Typen sind zufällig verteilt. Durch Kombination der Sigale von mehreren Geruchssinneszellen können verschiedene Gerüche genau unterschieden werden. (z.B.: Die 3 Geruchssinneszellen am Bild reichen aus um die 4 Gerüche eindeutig zu unterscheiden (Bild siehe Folie 8.17). Die 3 Zellen am Bild sind nur ein Beispiel. Es gibt viel mehr verschiedene Geruchsrezeptorproteine und damit auch viel mehr 92 Die Siebplatte (Lamina cribrosa ossis ethmoidalis) ist ein Teil des Siebbeins (Os ethmoidale). Sie bildet das Dach der Nasenhöhle und damit die Grenze zwischen Gehirn und Nasenhöhle. 93 Golf­Proteine sind spezielle G­Proteine, die nur in olfactorischen Rezeptorzellen vorkommen. 55 / 77 Neurobiologie verschiedene Geruchsinneszell­Typen.) Das unterschiedliche Ansprechen verschiedener Rezeptoren ergibt insgesamt den Geruch einer Substanz (population coding). Lichtrezeptoren Das Photopigment (in Stäbchen: Rhodopsin) ist ein G­Protein gekoppelter Rezeptor. D.h. Umwandlung von Licht in ein Signal in einem Neuron funktioniert ähnlich wie die Umwandlung eines chemischen Signals in ein Signal in einem Neuron. Licht, das auf das Photopigment trifft, (oder ein chemisches Signal) bewirkt die Aktivierung des G­Proteins und dieses aktiviert Enzyme, die eine Änderung der Menge eines bestimmten second Messengers auslösen. Dadurch wird die Durchlässigkeit bestimmter Ionenkanäle geändert und somit auch das Membranpotential. Im Fall von Licht ist der second Messenger cGMP, das G­Protein ist Transducin, die Menge des second Messengers wird reduziert und Na+­Kanäle schließen sich. Photorezeptoren haben cGMP94­gesteuerte Na+­Kanäle. cGMP wird kontinuierlich hergestellt und bewirkt, daß die Kanäle offen sind. Licht führt zur Aktivierung der cGMP­Phosphodiesterase, die cGMP zerstört. Durch die Zerstörung von cGMP schließen sich die Na+­Kanäle. Im dunkeln haben Photorezeptoren aufgrund des Na+­Einstroms ein Membranpotential von ca ­30mV. Bei Licht wird das Membranpotential negativer (bis zu ­60mV), weil der Na+­Einstrom geringer wird. D.h. Photorezeptoren werden durch Licht hyperpolarisiert. (Licht führt somit zu einer Reduktion der Neurotransmitterausschüttung in die Synapse zwischen Photorezeptor und Neuron.) Photorezeptoren sind Stäbchen und Zapfen. Sie sind sekundäre Sinneszellen. Stäbchen dienen dem schwarz/weiß­Sehen und sind zahlreicher als Zapfen. Zapfen dienen dem Farbsehen (es gibt 3 verschiedenen Zapfen (für blau, grün und rot)). Rhodopsin besteht aus Opsin (ein Protein mit 7 Transmembrandomänen) und Retinal (ein Derivat von Vitamin A). Retinal ist der Teil, der durch Licht verändert wird. Wenn Retinal durch Licht in die gestreckte Konformation übergeht (von 11­cis­Retinal in all­trans­ Retinal), aktiviert es das Opsin und dieses das G­Protein. (für ein Bild der Licht­aktivierten biochemischen Kaskade in einem Photorezeptor siehe Folie 8.21) Der Mechanismus der Phototransduktion (die Kaskade vom Retinal bis zum Schließen der Na+­ Kanäle) ist in Zäpfchen der selbe wie in Stäbchen. Zäpfchen enthalten aber 3 verschiedene Photo­ Pigmente (für die Farben Blau, Grün und Rot). (Jedes Zäpfchen enthält nur eine Art von Photopigment. D.h. es gibt auch 3 Arten von Zäpfchen.) (Wellenlängenbereiche der 3 Photo­Pigmente siehe Folie 8.22) Die Kombination der Information aus den verschiedenen Zäpfchen ergibt zusammen die Farbe. Reizweiterleitung und ­verarbeitung Die Stärke eines Reizes spiegelt sich im Rezeptorpotential wider. Je stärker der Reiz ist, desto stärker ist auch das Rezeptorpotential95. Ob und wie viele Aktionspotentiale aus diesem Rezeptorpotential resultieren hängt vom Schwellwert der Zelle ab. Die Stärke des Rezeptorpotentials hängt von der Stärke des Reizes ab, aber Aktionspotentiale sind immer gleich stark. Für ein stärkeres Rezeptorpotential werden daher Aktionspotentiale in kürzerer Abfolge (d.h. mit höherer Frequenz) generiert. Die Aktiononspotentiale werden generiert, solange das 94 cGMP = cyclic guanosine monophosphate 95 Das Rezeptorpotential ist das Membranpotential des Rezeptors. D.h. die direkte Folge des Öffnens von Ionenkanälen aufgrund eines Reizes. 56 / 77 Neurobiologie Rezeptorpotential den Schwellwert überschreitet. Wenn auf die Fingerspitze ein Druck ausgeübt wird, wird dieser von Rezeptorenden in ein elektrisches Signal umgewandelt. Die Neuronen gehen in das Rückenmark, kreuzen auf die andere Seite und verlaufen im Tractus spinothalamicus96 (E: spinothalamic tract) über den Hirnstamm und das Mesencephalon bis zum Thalamus und von dort weiter in den somatosensorischen Cortex (Gyrus postcentralis). (Umschaltungen von einem Neuron auf das nächste erfolgen im Rückenmark und im Thalamus.) Im somatosensorischen Cortex besteht Somatotopie. D.h. Information aus benachbarten Rezeptoren kommt in benachbarte Zellen im Gehirn (vgl. Homunculus). Zwei Mechanismen der Reizverarbeitung sind Laterale Hemmung und Absteigende Hemmung. (Bilder siehe Folie 8.25) Laterale Inhibition bedeutet, daß jedes Neuron seine benachbarten Neuronen inhibiert. Das führt zu einer Kontrastverstärkung bei der Empfindung. Bsp.: Wenn 3 Druckrezeptoren nebeneinander sind und es erfolgt ein Druck auf die beiden äußeren (und ein schwächerer Druck auf den mittleren), dann inhibieren die beiden äußeren das mittlere Neuron und das Gehirn bekommt dann nur von den beiden äußeren Signale, aber nicht vom mittleren. (Der mittlere inhibiert dabei auch die beiden äußeren, aber da der Druck auf den mittleren schwächer ist und die äußeren nur von einem Neuron (dem mittleren) inhibiert werden (während das mittlere von 2 Neuronen inhibiert wird), kommen dennoch (die meisten) Signale der äußeren zum Gehirn.) Absteigende Hemmung bedeutet, daß Signale vom Gehirn bewirken, daß sensorische Neuronen gehemmt werden. Z.B. wird Schmerz in bestimmten Situationen (z.B. Lebensgefahr) nicht gespürt. Funktion des motorischen Systems Um Kontakte mit der Umwelt aufzunehmen, ist das motorische System notwendig. Kommunizieren ist nur über das motorische System möglich (z.B.: für das Sprechen werden Muskeln benötigt). Das motorische System ist die Grundlage des Verhaltens. Das menschliche Verhalten erfordert die koordinierte Aktivierung von nahezu 700 Muskeln (schon allein an Gesichtsmuskeln gibt es eine große Anzahl). Komplizierte Verhaltenmuster können allerdings auch erzeugt werden, ohne daß das Gehirn beteiligt ist (z.B. kommt es vor, daß ein Huhn, nachdem ihm der Kopf abgeschlagen wurde, noch mit den Flügeln flattert (das Flattern geht vom Rückenmark aus)). Regelkreise im Rückenmark kontrollieren stereotype, repetitive Verhaltensmuster. Das Gehirn löst diese Verhaltensmuster aus und kann sie auch modifizieren. (Der Eingriff des Gehirns kann auch stören: eingelernte Verhaltensmuster funktionieren meist besser, wenn das Gehirn den automatischen Ablauf nicht beeinflußt (d.h. wenn man nicht darüber nachdenkt).) Vieles ist als fertiges Programm abgelegt, das vom Gehrin nur gestartet werden muß (z.B.: mit einem Fußball auf ein Tor schießen). Das motorische System verfügt über 2 Muskeltypen: glatte Muskulatur und quergestreifte Muskulatur. Zur glatten Muskulatur gehört die Muskulatur des Verdauungstraktes (Regulation der Peristaltik97) und der Arterien (Regulation von Blutfluß und Blutdruck (Kontraktion der Arterien erhöht den Blutdruck)). 96 Tractus spinothalamicus ist eine lange aufsteigende Leitungsbahn im Rückenmark (vom Rückenmark zum Thalamus). Eine lange Leitungsbahn ist eine Gruppe gebündelter Nervenfasern (gleichen Ursprungs und Ziels) in der weißen Substanz des Rückenmarks. 97 Peristaltik = wellenförmig fortschreitende Wandbewegung von Hohlorganen 57 / 77 Neurobiologie Zur quergestreiften Muskulatur gehören der Herzmuskel und die Skelettmuskeln. Die Skelettmuskeln machen den Großteil der Muskelmasse aus. Sie bewegen Knochen um die Gelenke (Bewegung, Gestik) und die Augen im Kopf, kontrollieren die Atmung und den Gesichtsausdruck (Mimik) und produzieren Sprache. Der Herzmuskel kontrahiert sich auch ohne Innervation rhythmisch. (Sinusknoten98, AV­Knoten99). Die Innervation reguliert die Frequenz, also den Puls. Die glatte Muskulatur und der Herzmuskel werden vom autonomen Nervensystem innerviert und reguliert. Die Skelettmuskeln werden vom somatischen Nervensystem innerviert und gesteuert. Das somatische motorische System besteht aus den Skelettmuskeln und den Teilen des Nervensystems, die sie steuern. Arbeitsmuskulatur (Arbeitsmuskulatur sind die Muskelfasern des Muskels, die die Kontraktion bewirken. Im Gegensatz zu Muskelfasern in Sensoren im Muskel.) Jeder Skelettmuskel ist von Bindegewebe umschlossen. Dieses bildet an den Enden des Muskels die Sehnen. Jeder Muskel besteht aus sehr vielen Muskelfasern. Jede Muskelfaser wird durch einen einzelnen Axon­Ast der unteren Motor­Neuronen innerviert. Die Hinterwurzel (Radix posterior), die in das Hinterhorn im Rückenmark geht, enthält sensorische Nervenfasern. Die Vorderwurzel (E: ventral root) (Radix anterior), die aus dem Vorderhorn (E: ventral horn) im Rückenmark kommt, enthält motorische Nervernfasern. Außerhalb des Rückenmarks vereinigen sich Hinterwurzel und Vorderwurzel zu einem Spinalnerv100 (E: spinal nerve), der somit sensorische und motorische Nervenfasern enthält. Vorderhorn (Cornu anterius) und Hinterhorn (Cornu posterius) gehören zur grauen Substanz im Inneren des Rückenmarks. Eine Muskelfaser ist eine einzige Zelle, die aber viele Zellkerne enthält. Sie hat einen Durchmesser von ca 20µm­80µm und ist bis zu einigen cm lang (um auf die ganze Länge des Muskels zu kommen sind meist mehrere Muskelfasern hintereinander bzw. überlappend). Eine Muskelfaser enthält viele Myofibrillen, die den Großteil des Volumens der Zelle einnehmen und ca so lang sind wie die Zelle selbst. Myofibrillen enthalten Myosinfilamente und Actinfilamente, die für die Querstreifung (ca 1.5µm breite Streifen) verantwortlich sind ("quergestreifte Muskulatur"). Jede Muskelfaser wird von einem eigenen Axon­Ast innerviert (aber ein Motorneuron innerviert viele Muskelfasern, weil es viele Axon­Äste hat). Mehrere Muskelfasern werden zu einem Primärbündel zusammengefaßt (Durchmesser: ca 1mm). Mehrere Primärbündel werden zu einem Sekundärbündel zusammengefaßt (Durchmesser: einige mm). Die Sekundärbündel sind die mit freiem Auge sichtbare Faserung der Muskeln. Alpha Motor Neuronen101 aktivieren direkt die Muskelfasern. Das Soma jedes Alpha Motor Neurons liegt im Vorderhorn im Rückenmark. Es muß für jede Tätigkeit die richtige Stärke eingesetzt werden. Z.B. wenn man ein Ei hält wird es bei zu viel Kraft zerdrückt und bei zu wenig fällt es hinunter. Die meisetn Bewegungen benötigen nur eine relativ schwache Muskelkontraktion (z.B.: gehen, reden, schreiben). Starke Muskelkontraktionen sind nur selten nötig (z.B.: Heben von Gewichten, Joggen (bei jedem Schritt muß das gesamte Gewicht des Körpers abgefangen und wieder wegegstoßen werden)). 98 Der Sinusknoten ist eine Anhäufung spezifischen Herzmuskelgewebes im rechten Vorhof des Herzens. In ihm entstehen die normalen Kontraktionsreize. (physiologischer Schrittmacher des Herzens) 99 Der AV­Konten (Atrioventrikularknoten) ist das sekundäre Erregungsbildungszentrum und befindet sich in der rechten Vorhofwand des Herzens. 100 Ein Nerv ist ein Bündel Axone im peripheren Nervensystem. 101 Alpha Motor Neuronen innervieren die Muskelfasern, die die Kontraktion des Muskels bewirken (Arbeitsmuskulatur). (Im Unterschied zu den Gamma Motor Neuronen, die die Muskelfasern in Muskelspindeln innervieren. Muskelspindeln sind Sensoren innerhalb der Skelettmuskeln, die Informationen über deren Länge liefern.) 58 / 77 Neurobiologie Die Kontraktionssträrke wird im Rückenmark bei der Aktivierung der Alpha Motor Neuroen reguliert. Eine Möglichkeit die Stärke zu regulieren, ist die Feuerrate (d.h. die Frequenz der Aktionspotentiale). Eine andere ist verschieden viele Alpha Motor Neuronen zu aktivieren. Ein Alpha Motor Neuron zusammen mit allen Muskelfasern, die es innerviert, wird als motorische Einheit (E: motor unit) bezeichnet. Alle Alpha Motor Neuronen, die den selben Muskel innervieren, zusammen werden als Motor Neuron Pool bezeichnet. Wenn nur wenig Kraft nötig ist, werden nur einige Alpha Motor Neuronen aktiviert, und somit kontrahieren sich nur einige Muskelfasern des Muskels. Wenn viel Kraft nötig ist, werden alle Alpha Motor Neuronen des Motor Neuron Pools aktiviert und der ganze Muskel kontrahiert sich. Je mehr Muskelfasern aktiviert werden, desto größer ist die Muskelkraft. Die Muskelkontraktion wird auch über die Feuerrate der Alpha Motor Neuronen reguliert. Jedes Aktionspotential in einem Alpha Motor Neuron bewirkt die Freisetzung von Acetylcholin in den Synapsen des Alpha Motor Neurons zu Muskelfasern. Acetylcholin löst in einer Muskelfaser ein EPSP aus und das EPSP bewirkt eine Muskelzuckung (d.h. eine Kontraktion der Faser). Für einen konstanten Muskeltonus (d.h. für eine kontinuierliche Kontraktion) sind kontinuierliche Aktionspotentiale erforderlich. Bei niedriger Frequenz (z.B.: 5 Hz) erfolgen viele einzelne Muskelzuckungen hintereinander. Bei hoher Frequenz (z.B.: 40 Hz) gehen die einzelnen Zuckungen direkt ineinander über, sodaß eine kontinuierliche Kontraktion ohne erkennbare Einzelzuckungen entsteht. Hohe Frequenz der Aktionspotentiale bewirkt zeitliches Aufsummieren der Antwort. (Weil bei hoher Frequenz jede Zuckung schon beginnt, bevor die vorhergehende zu Ende ist, summieren sich mehrere fast gleichzeitige Zuckungen auf, sodaß die Kontraktion auch stärker ist als das Maximum einer einzelnen Zuckung.) Muskelfasern entstehen durch die Fusion von Myoblasten102. Eine Muskelfaser ist somit eine Zelle mit mehreren Zellkernen. Die Zellmembran einer Muskelfaser heißt Sarkolemma und ist erregbar, d.h. sie kann Aktionspotentiale erzeugen (wie die Membran eines Axons). Das Sarkolemma enthält spannungsabhängige Na+­Kanäle, die ermöglichen, daß ein Aktionspotential durch das Sarkolemma läuft. Das EPSP aufgrund der Acetylcholin­Freisetzung in der Synapse zwischen Alpha Motor Neuron und Muskelfaser löst in der Muskelfaser ein Aktionspotential aus, das sich über die ganze Muskelfaser ausbreitet. Durch das Aktionspotential am Sarkolemma wird die Konformation von spannungsabhängigen Ca++­Kanälen im Sarkolemma geändert, sodaß sie sich öffnen. Jeweils 4 solche Ca++­Kanäle sind zu einer Tetrade zusammengeschlossen und mit einem Ca++­Kanal im sarkoplasmatischen Retikulum103 gekoppelt. Wenn die Kanäle der Tetrade aktiviert werden, öffnet sich auch der Ca++­Kanal im sarkoplasmatischen Retikulum und durch diesen wird mehr Ca++ aus dem sarkoplasmatischen Retikulum ins Cytosol freigesetzt als durch die Tetrade aus der Umgebung. Die erhöhte Konzentration an Ca++ im Cytosol bewirkt die Kontraktion der Myofibrillen104. Die Membran enthält Einstülpungen in die Zelle, die ein Netzwerk aus Tunneln in der Zelle bilden und T­Tubuli105 heißen. Das Aktionspotential breitet sich natürlich auch in die T­Tubuli aus und erreicht somit auch sarkoplasmatisches Retikulum tief in der Muskelfaser. 102 Myoblasten sind Vorläuferzellen der Zellen des Muskelgewebes. 103 Das sarkoplasmatische Retikulum ist das endoplasmatische Retikulum der Muskelfaser. Es dient der Speicherung von Ca++­Ionen. (gr. sarkós = Fleisch) 104 Eine Myofibrille ist ein Bündel aus Actin und Myosin, das in Längsrichtung von einem Ende der Muskelzelle zum anderen geht (und am Ende jeweils mit der Membran verbunden ist). (gr. mys, myós = Muskel (eigentlich Maus)) 105 T­Tubuli (transverse tubules, transversale Tubuli) enthalten extracelluläre Flüssigkeit (weil sie Einstülpungen der Membran sind). 59 / 77 Neurobiologie Eine Myofibrille wird durch Z­Linien (auch als Z­Scheiben bezeichnet) in Untereinheiten unterteilt. Eine solche Untereinheit, bestehend aus 2 benachbarten Z­Linien und dem Teil dazwischen, heißt Sarkomer. An den Z­Scheiben sind dünne Filamente befestigt, die aber (in der entspannten Muskelfaser) nicht bis zu den dünnen Filamenten der nächsten Z­Scheibe reichen. D.h. die dünnen Filamente gehen von den beiden Enden des Sarkomers in Richung Mitte, reichen aber nicht ganz bis zur Mitte. Die dünnen Filamente bestehen aus Actin. Zwischen den dünnen Filamenten befinden sich dicke Filamente. Diese verlaufen parallel zu den dünnen Filamenten, beginnen aber nicht bei der Z­Scheibe, sondern erst ein Stück innerhalb des Sarkomers. Sie gehen über die Mitte des Sarkomers und überlappen auch mit den dünnen Filamenten der anderen Hälfte des Sarkomers. D.h. sie überlappen mit den dünnen Filamenten beider Hälften des Sarkomers, reichen aber nicht bis zu den Enden des Sarkomers. Die dicken Filamente bestehen aus Myosin. (Die Streifen der quergestreiften Muskulatur kommen durch die unterschiedliche optische Dichte von Actin (optisch weniger dicht (heller)) und Myosin (optisch dichter (dunkler)) zustande. (Die Z­ Linie ist eine dunkle Linie in der hellen I­Bande (nur Actin). Die dunkle Bande zwischen 2 I­Banden heißt A­Bande (Myosin (auch wo es mit Actin überlappt; der Überlappungsbereich ist noch etwas dunkler als der Rest der A­Bande)).)) Bei der Kontraktion des Muskels ziehen die dicken Filamente die dünnen Filamente in Richtung Mitte des Sarkomers, sodaß die beiden Z­Scheiben näher zusammen kommen und somit das Sarkomer kürzer wird. Die Länge der Filamente (dünne und dicke) ändert sich dabei nicht, sondern ihr Überlappungsbereich wird größer, weil die dünnen weiter in die Mitte gezogen werden. (Dies wird als Gleitfilament­Mechanismus (E: sliding­filament model) der Muskelkontraktion bezeichnet.) Das Protein Troponin ist an die dünnen Filamente gebunden und verhindert (bei Abwesenheit von Ca++) das Binden von Myosin an die dünnen Filamente. Wenn ausreichend Ca++ vorhanden ist, sodaß es an Troponin bindet, gibt Troponin die Bindungsstellen für Myosin frei. Molekularer Ablauf der Muskelkontraktion: • Das Myosin der dicken Filamente bindet an das Actin der dünnen Filamente (dabei wird ATP zu ADP + Pi gespalten; Ca++ verstärkt auch diese ATPase­Aktivität (Myosin ist somit eine Ca++­abhängige ATPase)). • Darauf folgt eine Konformationsänderung des Myosins, durch die der Myosin­Kopf stärker abgeknickt wird und die Filamente ein kleines Stück gegeneinander verschoben werden (dabei wird ADP abgegeben). • Durch neuerliche Bindung eines ATP an das Myosin löst sich der Myosin­Kopf wieder vom Actin (und streckt sich wieder). (Die Bindungsstelle kann dann wieder von Troponin bedeckt werden.) Dann kann er wieder (etwas weiter vorne) an Actin binden, durch Konformationsänderung die Filamente verschieben, ... . Solange ausreichend Ca++ und ATP vorhanden sind, kann sich dieser Prozess wiederholen und der Myosin­Kopf wandert am Actin­Filament entlang. Ca++ wird ständig von Pumpen in das sarkoplasmatische Retikulum transportiert. Wenn keine Aktionspotentiale von Nerven mehr ankommen, nimmt die Ca++­Konzentration ab und Troponin bedeckt wieder die Myosin­Bindungstellen des Actins. Myosin kann sich zwar noch (unter ATP­ Aufnahme) vom Actin­Filament lösen, findet aber keine neue Bindungsstelle und die Filamente gleiten wieder auseinander. Weil ATP für das Lösen der Bindung zwischen Myosin und Actin nötig ist, kann Myosin, wenn kein ATP mehr vorhanden ist (z.B.: bei Toten), zwar noch an Actin binden, bleibt aber dann im gebundenen Zustand. Dadurch kommt die Totenstarre zustande. 60 / 77 Neurobiologie Sensoren des Muskels Die Sensoren im Muskel dienen der Propriozeption (d.h. der Wahrnehmung von Eigenschaften (Bewegung, relative Positionen der Körperteile, ...) des eigenen Körpers). Muskelspindel Die Dehnung des Muskels (d.h. eine äußere Kraft, die den Muskel dehnt) wird von Muskelspindeln gemessen. (D.h. sie nehmen Änderungen der Muskellänge wahr.) Muskelspindeln befinden sich im Muskel, zwischen den Muskelfasern eingelagert. Eine Muskelspindel ist eine fibröse106 Kapsel (ca 2­10mm lang und 0.2mm dick), die modifizierte Skelettmuskelfasern enthält. (Unter anderem sind diese Muskelfasern deutlich dünner als extrafusale Fasern: die ganze Muskelspindel hat ca den gleichen Durchmesser wie eine extrafusale Faser.) (Die Muskelfasern der Arbeitsmukulatur (also die außerhalb der Muskelspindeln) heißen extrafusale Fasern, die Muskelfasern der Muskelspindeln heißen intrafusale Fasern.) Die intrafusalen Fasern werden in einen mittleren (dickeren) Bereich und die 2 Bereiche auf beiden Seiten des mittleren Bereichs (d.h. bei den Enden der Muskelspindel) unterteilt. Der mittlere Bereich enthält hauptsächlich Zellkerne und kaum Myofibrillen, sodaß er nicht kontrahierbar ist. Er ist von sensorichen Neuronen (Gruppe­Ia­Neuronen) innerviert. Die beiden Endbereiche enthalten Myofibrillen, sind von motorischen Neuronen (γ­Motor­Neuronen) innerviert und können sich kontrahieren. In der Mitte der Muskelspindel sind die Muskelfasern von sensorischen Gruppe­Ia­Nervenfasern umwickelt. Wenn der Muskel und damit auch jede enthaltene Muskelspindel gedehnt wird, löst die Dehnung der Muskelfasern der Muskelspindel über mechanosensitive Ionenkanäle der Gruppe­Ia­ Nervenfasern Aktionspotentiale in diesen Nervenfasern aus. Diese Neuronen gehen über das Hinterhorn ins Rückenmark. Gruppe­Ia­Nervenfasern sind die größten (am dicksten myelinierten) und damit schnellsten Neuronen des Körpers. Wenn ein Muskel gedehnt (gestreckt) wird, kontrahiert er sich. Das wird als Muskel­Streckreflex107 (E: stretch relfex (myotatic reflex)) bezeichnet. Die Dehnung wird von den Muskelspindeln wahrgenommen. Die Gruppe­Ia­Neuronen der Muskelspindeln werden stärker depolarisiert und die Feuerfrequenz erhöht sich. Die Gruppe­Ia­ Neuronen der Muskelspindeln haben im Rückenmark Synapsen zu den Alpha Motor Neuronen des selben Muskels. Die höhere Anzahl an Aktionspotentialen in den Gruppe­Ia­Neuronen führt daher zu einer stärkeren Depolarisation der Alpha Motor Neuronen, die dadurch schneller feuern und somit die Kontraktion des Muskels auslösen. Der Knie­Sehnen Reflex (E: knee­jerk reflex) ist ein Beispiel für einen Muskel­Streckreflex. Durch den Schlag auf die Sehne (E: tendon) des Musculus quadriceps wird der Muskel gedehnt und somit der Muskel­Streckreflex ausgelöst. Wenn ein Muskel kontrahiert ist, werden dadurch auch die Muskelspindeln verkürzt. Wenn der kontrahierte Muskel dann ein Stück gedehnt wird, könnte diese Dehnung von den Muskelspindeln nicht wahrgenommen werden, weil sie aufgrund der Kontraktion immer noch kürzer sind als in einem (gedehnten) entspannten Muskel. Damit die Muskelspindeln trotz Kontraktion des Muskels eine Dehung noch wahrnehmen können, werden die Muskelfasern der Muskelspindel auch verkürzt. Die Muskelfasern einer Muskelspindel werden von Gamma­Motorneuronen innerviert. Aktionspotentiale der Gamma­Motorneuronen bewirken eine Kontraktion der End­Bereiche der Muskelfasern der Muskelspindel (also der Aufhängung des sensorischen Teils) (der dickere Mittelteil mit den sensorischen Gruppe­Ia­Neuronen kann sich 106 fibrös = faserreich 107 Muskel­Streckreflex = Dehnungsreflex = myotatischer Reflex 61 / 77 Neurobiologie nicht kontrahieren). (Am Bild (Folie 9.13) ist die Muskelspindel vergrößert und neben (statt in) dem Muskel dargestellt.) Dadurch wird bei einer Dehnung des Muskels wieder der Mittelteil der Muskelspindel gedehnt und somit die Dehnung wahrgenommen. Golgi Sehnenorgan Golgi Sehnenorgane (E: Golgi tendon organ) befinden sind in der Sehne (meist am Übergang zwischen Muskel und Sehne) und messen die Muskelspannung. Ein Golgi Sehnenorgan besteht hauptsächlich aus kollagenen Fasern, ist von einer Kapsel umgeben und wird von sensorischen Ib­ Neuronen innerviert. (Sie sind bis zu 1.6mm lang und bis zu 0.12mm dick.) Eine der Aufgaben von Golgi Sehnenorganen ist zu verhindern, daß der Muskel reißt. Die Ib­ Neuronen haben im Rückenmark Synapsen zu inhibitorischen Interneuronen und diese haben (auch im Rückenmark) hemmende Synapsen zu den Alpha Motor Neuronen des selben Muskels. Wenn die Spannung im Muskel zu hoch wird, hemmen die Ib­Neuronen über diesen Weg die Alpha Motor Neuronen und verringern dadurch die Kontraktion des Muskels. Dies wird als umgekehrter myotatischer Reflex (E: reverse myotatic reflex) bezeichnet. Die Golgi Sehnenorgane liefern auch bei Spannungen, die noch nicht zu hoch sind, Informationen und dienen damit der Regulierung der Muskelspannung. Sie ermöglichen dadurch feine Manipulationen (z.B.: wenn man ein Ei hält darf man weder zu fest noch schwach zugreifen). Golgi Sehnenorgane messen die Muskelspannung und nicht Längenänderungen des Muskels. D.h. sie nehmen Spannungsänderungen auch wahr, wenn sich die Länge des Muskels nicht ändert. Sie liefert Informationen über die Spannung, unabhängig davon, ob diese durch eine äußere Kraft oder durch Kontraktion des Muskels selbst bewirkt wird. (Bild siehe Folie 9.15) Muskelspindel und Golgi Sehnenorgan ergänzen einander um Muskelspannung und Muskellänge zu regulieren. Rezeptoren in den Gelenken Es gibt auch in den Gelenken Propriozeptoren108. Im Bindegewebe der Gelenke (Gelenkskapsel) befindet sich eine Vielzahl an Propriozeptiven Axonen. Sie sprechen auf Änderungen des Winkels des Gelenks und der Richtung und Geschwindigkeit der Gelenkbewegung an. Gemeinsam mit Muskelspindel, Golgi Sehnenorgan und Rezeptoren in der Haut liefern sie Informationen über die Stellung des Gelenks. Reziproke Hemmung Hemmung von Muskeln (bzw. deren Alpha Motor Neuronen) ist genauso wichtig wie die Kontraktion. Wenn ein Muskel kontrahiert wird, muß der Gegenmuskel (Antagonist) relaxiert werden. Diese Hemmung des Gegenmuskels wird als reziproke Hemmung bezeichnet und wird durch hemmende Interneuronen umgesetzt. Z.B. sind die Ia­Neuronen von den Muskelspindeln nicht nur exzitatorisch mit den Alpha Motor Neuronen des selben Muskels, sondern auch über hemmende Interneuronen inhibitorisch mit den Alpha Motorneuronen des Gegenmuskels verbunden. Der Muskel­Streckreflex bewirkt also gleichzeitig mit der Kontraktion des Muskels die Entspannung des Gegenmuskels. (Ansonsten würde durch die Dehnung des Gegenmuskels auch der Muskel­Streckreflex des Gegenmuskels ausgelöst und beide Muskeln würden sich aufgrund ihrer Muskel­Streckreflexe ständig gegeneinander anspannen.) 108 Rezeptoren, die der Propriozeption dienen (Rezeptoren in Skelettmuskulatur, Sehnen und Gelenkkapseln) 62 / 77 Neurobiologie Reziproke Hemmung ist auch für absichtliche Bewegungen nötig. Wenn z.B. der Ellenbogen bewußt abgebogen wird, muß gleichzeitig der Streckmuskel relaxiert werden, sonst würde im Streckmuskel der Muskel­Streckreflex ausgelöst und der beabsichtigten Bewegung entgegenarbeiten. Alpha Motor Neuronen bekommen Input von oberen Motor Neuronen im Gehirn, von Ia­Neuronen (von Muskelspindeln) und von Interneuronen (sowohl hemmend als auch erregend) im Rückenmark. Bei Muskel­Streckreflexen (wie z.B. dem Kniesehenreflex) haben die sensorischen Neuronen Synapsen direkt mit den Alpha Motor Neuronen (ohne, daß weitere Neuronen dazwsichen geschaltet wären). Solche Reflexsysteme heißen monosynaptische Reflexsysteme. Beim Kniesehnenreflex wird der Musculus quadriceps (Extensor109 des Kniegelenks) durch einen Schlag auf seine Sehne gedehnt und somit sein Muskel­Streckreflex ausgelöst. Die afferenten Fasern (Ia­Neuronen von den Muskelspindeln) aktivieren die Alpha Motor Neuronen des Musculus quadriceps und inhibieren gleichzeitig (über Interneuronen) die Alpha Motor Neuronen des Flexors110. (Bild siehe Folie 9.19) Die Informationen aus den Sensoren werden von den Ia­Neuronen nicht nur direkt an die Alpha Motor Neuronen des selben Muskels weitergegeben, sondern auch über hemmende Interneuronen an die Alpha Motorneuronen des Gegenmuskels und an das Gehirn. Die sensorische Information geht über Medulla und Thalmus in den somatosensorischen Cortex (Gyrus postcentralis), wird dort verarbeitet und, falls eine Reaktion ausgelöst wird, geht diese vom motorischen Cortex (Gyrus precentralis) über das Rückenmark (absteigende Bahnen) zu den Muskeln. (Der Weg über das Gehirn ist natürlich viel langsamer (längere Strecke, viele Schaltungen und Verarbeitung) als der Reflex.) Integrative Hirnfunktionen Funktionelle Organisation des Nervensystems: • sensorische Funktion: Das Nervensystem erhält und transportiert Informationen von Sensoren über externe und interne Reize. • integrative Funktion: Das Nervensystem analysiert eingehende sensorische Informationen, verknüpft sie mit gespeicherter Information und löst adäquates Verhalten aus. Emotionen spielen dabei eine große Rolle. • motorische Funktion: Das Nervensystem initiiert Muskelkontraktionen (für Bewegung, Mimik, Gestik, Sprache) und Drüsensekretion (z.B.: Auslösung von Stress). Der Cortex wird in Hirnrindenfelder unterteilt: • primäre Felder: Das sind die sensorischen Zentren (z.B.: somatosensorsiches Primärfeld im Gyrus postcentralis (Brodmann­Felder 3, 1, 2; S1; primärer somatosensorischen Cortex)), in denen die betreffenden Sinnesbahnen enden, und die motorischen Zentren (z.B.: somatomotorisches Primärfeld im Gyrus precentralis (Brodmann­Feld 4; M1, primärer Motorcortex)), die einfache Bewegungen auslösen. • unimodale Felder höherer Ordnung: Sie liegen neben den primären Feldern. Hier erfolgt die erste Integration der Sinnesinformation: wahrgenommenes wird erkannt und zugeordnet (z.B.: Gesichtserkennung, Erkennen von Wörtern und Melodien). • Assoziationsfelder: Hier wird die Informationen aller Sensoren integriert und mit motorischer Leistung und Motivation verknüpft. Aktuelle Information wird mit früherer verglichen und gespeichert. 109 Extensor = Strecker 110 Flexor = Beuger 63 / 77 Neurobiologie Der Motor Cortex umfaßt die Felder 4 und 6, wobei Feld 6 weiter in PMA (prämotorisches Areal (E: premotor area)) und SMA (supplementär­motorisches Areal (E: supplementary motor area)) unterteilt wird. Der posteriore parietale Cortex (somatosensorischer Assoziationscortex) umfaßt die Felder 5 und 7. In Feld 5 werden somatosensorische und propriozeptive Informationen verarbeitet. In Feld 7 werden visuelle Informationen mit somatosensorischen/motorischen Informationen verknüpft. (Feld 7 liegt zwischen somatosensorischen Feldern (Brodmann Felder 3, 1, 2) und dem visuellen Cortex (Brodmann Felder 17, 18, 19).) Der präfrontale Cortex (frontaler Assoziationskortex) und der posteriore parietale Cortex treffen Entscheidungen und aktivieren Felder 6 (motorische Planung) und 4 (absteigender Tractus111 corticospinalis (Pyramidenbahn) (E: descending corticospinal tract)). (Bilder siehe Folie 10.5 (M1 ist der primäre motorische Cortex. S1 ist der primäre somatosensorische Cortex. Es besteht eine somatotopische Zuordung der Bereiche des primären motorischen und primären somatosensorischen Cortex zu den Körperregionen. Diese wird als Homunculus bezeichnet.)) Der präfrontale Cortex ist wichtig für bewußte Entscheidungen. Zu seinen Funktionen gehören: • zeitliche und räumliche Strukturierung von Sinneswahrnehmungen • geplantes und kontextgerechtes Handeln • Steuerung der Motivation • Arbeitsgedächtnisprozesse Läsionen des präfrontalen Cortex führen zu Schizophrenie112 und Persönlichkeitveränderung, erschweren geplante Handlungen und verringern Fähigkeiten auf den Gebieten von Moral, Regeln und Problemlösen. Bewegung Das zentrale motorische System ist in eine Hierarchie aus 3 Kontroll­/Steuerungsstufen eingeteilt: • Strategie (höchste Stufe) (was will ich erreichen): Festlegung des Ziels der Bewegung und der Bewegungsstrategie; beteiligt sind der assoziative Cortex und die Basal­Ganglien • Taktik (mittlere Stufe) (wie erreiche ich dieses Ziel): Abfolge der Muskelkontraktionen; beteiligt sind motorischer Cortex und Cerebellum (Das Cerebellum steuert die Feinmotorik.) • Durchführung (unterste Stufe): Aktivierung der Motorneuronen und der Interneuronen113 und Anpassung der Stellung des Körpers im Raum; beteiligt sind Hirnstamm und Rückenmark Strategie: Bsp. Ein Baseballspieler bereitet sich auf seinen Wurf vor: • Strategie: Entscheidung wie der Ball geworfen wird (schneller Ball, gekrümmte Flugkurve, ...) unter Berücksichtigung von Wissen und Erfahrung (Stärken und Schwächen des Batters, wie wurde der Ball letztes Mal geworfen und hat ihn der Batter erwischt, ...) • Taktik: Bewegungsablauf • Durchführung: Kontraktion einzelner Muskeln um den Bewegungsablauf durchzuführen Der Cortex sammelt alle Informationen (visuelle, akkustische, Propriozeption über die Position des Körpers im Raum). Eine Strategie für die Erreichung des Ziels wird festgelegt. Die Entscheidung 111 Tractus = Strang, Bahn (vor allem Nervenbahnen des Gehirns und Rückenmarks) 112 Schizophrenie: eine endogene Psychose; chrakterisiert durch Verlust des Kontakts zur Realität; mit vielgestaltigen Störungen der Persönlichkeit, des Denkens, des Wahrnehmens, der Stimmung/Affektivität, ... 113 Interneuronen sind alle Neuronen, die weder Sensor­ noch Motor­Neuronen sind. 64 / 77 Neurobiologie über Art und Ziel der Bewegung wird getroffen. Taktik: An der motorischen Planung sind der assoziative Cortex und die Basal­Ganglien (Nucleus caudatus, Putamen, Globus pallidus, Nucleus subthalamicus und Subtantia nigra) beteiligt. (Nucleus caudatus und Putamen werden zum Striatum zusammengefaßt.) Der Informationsfluß erfolgt vom Cortex in die Basalganglien (Striatum und weiter in Globus pallidus), von dort in den ventral­lateralen Nucleus des Thalamus (genauer: in VLo114) und dann wieder zurück in den Cortex in Feld 6 (SMA). Dies wird als motorische Schleife (E: motor loop) bezeichnet. Die Funktion der motorischen Schleife ist die Auswahl und Initiation von bewußten Bewegungen. Störungen der motorischen Schleife sind Teil von z.B. Morbus Parkinson115 und Chorea Huntington116. Das Cerebellum kontrolliert die Abfolge der Muskelkontraktionen und ist für die Koordination der Feinmotorik zuständig. Axone aus dem sensorisch­motorischen Cortex innervieren Kerne (Nuclei) im Pons und von dort das Cerebellum. Dieser Weg wird als cortico­ponto­cerebellare Projektion (E: corticopontocerebellar projection) bezeichnet. Auch hier liegt eine Schleife vor: Sie geht über VLc117 zurück in den Cortex in Feld 4. Die cortico­ponto­cerebellare Projektion enthält 20*106 Axone. Das Cerebellum informiert mittels diese Schleife den primären Motor Cortex über Bewegungsrichtung, Muskeltiming und ­kraft der geplanten Bewegung. Läsionen in diesem Weg haben Ataxie118 zur Folge. Durchführung: Bsp. Baseballspieler: • Nach der Planung wird die Bewegung im motorischen Cortex (Feld 4) ausgelöst. • Die adequat getimte Aktivierung von Motorneuronen im Rückenmark erzeugt eine koordinierte Bewegung von Schulter, Ellenbogen, Handgelenk und Fingern. • Der Hirnstamm koordiniert die notwendigen Anpassungen der Haltung des Körpers zur Erhaltung des Gleichgewichts. • Motorneuronen des Hirnstamms bewirken, daß die Augen des Werfers am Ziel fixiert bleiben, obwohl sich Kopf und Körper bewegen. • Die Flugbahn des Balls ist ballistisch (durch Anfangsrichtung und ­geschwindigkeit festgelegt). Die Armbewegung des Werfers wird ebenso als ballistisch bezeichnet, weil sie nicht mehr geändert werden kann sobald sie ausgelöst wurde. • Die Bewegung wird beeinflußt durch die Propriozeption von Gliedern und Körper vor der Bewegung und durch Antizipation (Vorwegnahme) von (vorhersehbaren) Änderungen während des Wurfs. • Sensorische Information während der Bewegung kann den Wurf nicht mehr beeinflussen (weil die Wurfbewegung dafür zu schnell abläuft). Sie ist aber für die Verbesserung 114 VLo steht für ventral lateral nucleus pars oralis. VLo ist ein Teil des Nucleus ventralis laterales. Die Nuclei ventrales laterales werden mit VL angekürzt und sind Teil des Thalamus. 115 Morbus Parkinson: eine Bewegungsstörung, verursacht durch Degeneration dopaminerger Neuronen in der Substantia nigra 116 Chorea Huntington: erbliche Krankheit, charakterisiert durch Demenz und Fehlfunktionen beim Bewegungsablauf (z.B.: unwillkürliche Muskelkontraktionen), (Zusammenhang mit Degeneration von Neuronen in Basalganglien und im Cortex) 117 VLc steht für ventral lateral nucleus pars caudalis. VLc ist ein Teil des Nucleus ventralis laterales. 118 Ataxie ist eine Störung von Bewegungsabläufen: die Bewegungen sind unkoordiniert und ungenau. 65 / 77 Neurobiologie zukünfiger ähnlicher Bewegungen wichtig. Bewegungs­Lernen erfolgt im Cerebellum: die Bewegungsabsicht wird mit dem verglichen, was wirklich passiert und zukünftige Bewegungen werden entsprechend adaptiert. Das adequate Fuktionieren der Motorkontroll­Hierarchie hängt stark von sensorischer Information ab (sensorische Information vor dem Wurf für die Durchführung und während des Wurf für zukünftige Verbesserungen). Das motorische System des Gehirns ist also eigentlich ein sensorisch­ motorisches System. Zur Entwicklung integrativer Funktionen des Gehirns gehören: • Erkennen der Umwelt durch den Säugling Summe aller sensorischen Informationen Erkennen von Zusammenhängen Kind­Mutter: Wärme, Geborgenheit Beschaffenheit von Dingen • Entwicklung von Bewußtsein Hunger, Durst, Schmerz, Emotionen • Erlernen komplexer motorischer Programme • Entwicklung von Aufmerksamkeit Motivation (Antrieb, seine Ziele zu erreichen) • Lernen und Gedächtnis • Denken und Sprache Sprache Zu den primären Feldern und Assoziationsfeldern für Sprache gehören das Broca­Zentrum (E: Broca's area) (motorische Sprachregion), Wernicke­Zentrum (E: Wernicke's area) (sensorische Sprachregion), der Gyrus supramarginalis (E: supramarginal gyrus) und Teile des primären motorischen Cortex (Muskelkontraktionen für das Sprechen), das auditorische Primärfeld (Hören). Bei Schädigungen in diesen Bereichen kommt es zu Aphasie119. Bei den meisten Menschen befinden sich die Sprachregionen in der linken Gehirnhälfte. Die Sprachregionen sind auch für Lesen und Schreiben wichtig (beim Lesen werden aus den Schriftzeichen Wörter erstellt, die in den Sprachregionen ausgewertet werden). Bei Broca'scher Aphasie ist ein Bereich im motorisch­assoziativen Cortex betroffen (das Broca­ Zentrum). Der Patient kann nicht sprechen, kann aber Sprache verstehen und Text lesen und verstehen. Das Broca­Zentrum ist in der Nähe des Gyrus precentralis (d.h. des primären motorischen Cortex). Bei Wernicke'scher Aphasie ist ein Bereich des Lobus temporalis betroffen (das Wernicke­ Zentrum). Der Patient versteht Sprache nicht. Er kann sprechen, spricht aber sinnloses Zeug. Das Wernicke­Zentrum stellt den Zusammenhang zwischen Lauten, Worten und Bedeutung her. Bei Leitungs­Aphasie (E: conduction aphasia) kann der Patient Sprache verstehen und flüssig sprechen, kann aber Worte nicht korrekt wiederholen. Bei dieser Aphasie ist die Verbindung zwischen Wernicke­Zentrum und Broca­Zentrum unterbrochen. (Das Wernicke­Geschwind Modell ist ein Modell der Sprachverarbeitung im Gehirn und inkludiert Interaktionen von Broca­Zentrum, Wernicke­Zentrum und sensorischen und motorischen Arealen.) Beim Hören von Worten ist hauptsächlich der auditorische Cortex (und das Wernicke­Zentrum) 119 Aphasie ist der teilweise oder vollständige Verlust von sprachlichen Fähigkeiten durch Schädigung des Gehirns (oft ohne Beeinträchtigung anderer Gehirnfunktionen). 66 / 77 Neurobiologie aktiv. Beim Erzeugen von Worten ist hauptsächlich das Broca­Zentrum aktiv. (Bilder siehe Folie 10.15) Beim Lesen von Worten kommt die Information vom primären visuellen Cortex in den Gyrus angularis (E: angular gyrus) und wird dort in Signale umgewandelt, die vom Wernicke­Zentrum wie gesprochen Worte verarbeitet werden können. Die weitere Verarbeitung erfolgt wie beim Hören von gesprochenen Worten. (Bilder siehe Folie 10.16) Spezialisierung von linkem und rechtem parietalen Assoziationscortex Die beiden Gehirnhälften sind nicht identisch. Sie haben unterschiedliche Aufgaben. Z.B. wird Sprache meist links und Musik meist rechts verarbeitet. Bei jeder höheren kognitiven Tätigkeit ist das ganze Gehirn beteiligt. Die Zuordnung von Funktionen zu bestimmten Teilen erfolgt durch Untersuchung was nicht mehr funktioniert, wenn ein bestimmter Teil des Gehirns geschädigt ist. (Z.B.: daß das Broca­Zentrum für Sprechen zuständig ist, bedeutet, daß ein Mensch ohne Broca­Zentrum nicht sprechen kann, bedeutet aber nicht, daß ein Broca­Zentrum alleine (ohne das restliche Gehirn) sprechen kann.) Weiters sind alle diese Zuordnungen eher diffus (die Bereiche für eine Funktion können nicht genau abgegrenzt werden, Funktionen können bei verschiedenen Menschen in unterschiedlichen Gehirnhälften liegen, Aufgaben können bei Verletzung von anderen Regionen übernommen werden, ...). Linke Hälfte: Integration von visuellem System und Sprache, rechnen, symbolisches Denken Läsionen bewirken: • Störungen beim Lesen und Schreiben • Beeinträchtigung von abstraktem Denken rechte Hälfte: Aufmerksamkeitssteuerung bei räumlicher Orientierung, räumliche Lokalisation, Musik Läsionen bewirken: • Störung der räumlichen Orientierung • Schwierigkeiten beim Werkzeuggebrauch • Ignorieren einer Körperhälfte Mit PET­Untersuchungen können die unterschiedlichen Spezialisierungen der Gehirnhälften gezeigt werden. Beim Hören von Sprache ist hauptsächlich die linke Hälfte aktiv, beim Hören vom Musik hauptsächlich die rechte Hälfte. (Bilder siehe Folie 10.18) Die beiden Gehirnhälften sind über das Corpus callosum (Hirnbalken) miteinander verbunden. Das Corpus callosum enthält 200*106 Axone. Es wurde bei Epilepsie­Patienten durchtrennt um ein Übergreifen der Epilepsie auf die andere Gehirnhälfte zu verhindern. Personen mit durchtrenntem Corpus callosum werden als Split­Brain­Personen bezeichnet. Sie zeigen normalerweise keine Auffälligkeiten, aber mit bestimmten Experimenten sind Unterschiede zu Menschen mit intaktem Corpus callosum erkennbar. Nach Durchtrennung des Corpus callosum kontrollieren praktisch zwei unabhängige Gehirne die beiden Körperhälften. Die komplementäre Hemisphären­Spezialisierung (linke Hälfte: reden, sprechen, schreiben, ...; rechte Hälfte: Musik, Sprechmelodie, Idiome (z.B.: "mit dem Zaumpfahl winken")) kann deutlicher erkennbar sein. Wenn einer Split­Brain­Person in einem Experiment das Wort "Ball" gezeigt wird, sodaß es nur in der linken Hälfte des Gesichtsfeldes ist, dann kann diese Person nicht sagen, welches Wort sie sieht, 67 / 77 Neurobiologie kann aber mit der linken Hand aus mehreren Objekten das auswählen, das dem Wort entspricht. Das Bild des Wortes wird nur von der rechten Gehirnhälfte wahrgenommen (die Nerven des linken Sehfelds gehen in die rechte Gehrinhälfte) und die rechte Gehirnhälfte kann nicht sprechen. Die linke Gehrinhälfte, die sprechen kann, kann das Wort aber nicht sehen. Die rechte Gehirnhälfte, die das Wort gesehen hat, kontrolliert die linke Seite des Körpers und kann somit mit der linken Hand das richtige Objekt auswählen. (Die Sprachregionen sind zwar in der linken Gehirnhälfte, aber die rechte kann auch einzelne Zeichen oder kurze Worte verstehen.) Synaptische Plastizität und Lernen Synaptische Transmission erfolgt an der Kontaktstelle zwischen prä­ und postsynaptischem Neuron. Neurotransmitter werden freigesetzt, binden an Rezeptoren am Zielneuron und werden dann wieder entfernt. Synapsen passen ihre Funktion durch Regulation der Biosynthese, der Freisetzung und der Rezeptoren dem Bedarf an. Regulation der Biosynthese erfolgt durch: • Feedback Hemmung: Die Tyrosinhydroxylase (wandelt Tyrosin in L­DOPA um) wird durch ihre Folgeprodukte gehemmt. • Modulation der Enzymaktivität: Die Tyrosinhydroxylase kann phosphoryliert werden. Wenn sie phosphoryliert ist, wirken die Folgeprodukte weniger hemmend. • Enzym Biosynthese: Wenn mehr Tyrosinhydroxylase vorhanden ist, wird auch mehr L­DOPA hergestellt. Die Biosynthese bestimmter Enzyme wird z.B. bei Stress erhöht. Regulation der Freisetzung erfolgt durch: • Autorezeptoren: Autorezeptoren (z.B.: α2­Noradrenlin­Rezeptor) schalten die Freisetzung ab (z.B. indem sie die spannungsabhängigen Ca++­Kanäle blockieren). Wenn genug Noradrenalin im synaptischen Spalt ist, sodaß es auch an viele Autorezeptoren bindet, wird dadurch die Freisetzung verringert. • präsynaptische Rezeptoren: GABA bewirkt (z.B.) über Cl­­Kanäle eine Hyperpolarisation der Membran, sodaß ein exzitatorisches Signal inhibiert wird. direktes postsynaptisches Feedback: Dies erfolgt durch ein direktes Signal von der postsynaptischen Zelle an die präsynaptische Zelle und kommt vermutlich relativ oft vor, ist aber nur wenig untersucht. monosynaptisches Feedback: Beim monosynaptischen Feedback (monosynaptische negative feedback loop) hat das Neuron auch (inhibierende) Synapsen zu sich selbst. D.h. wenn das Neuron ein Aktionspotential durch das Axon schickt, geht dieses durch einen Zweig des Axon auch zur Synapse zum selben Neuron und hemmt dadurch das Neuron selbst (verhindert also weitere Aktionspotentiale). polysynaptisches Feedback: Beim polysynaptischen Feedback (polysynaptische negative feedback loop) wird das Ausgangsneuron durch eine (oder mehrere) Synapse eines anderen Neurons gehemmt, wobei dieses andere Neuron direkt oder indirekt durch das Ausgangsneuron aktiviert wird. Regulation der Rezeptoren: • Rezeptor Empfindlichkeit • Rezeptor Menge • Rezeptor Expression Ein Autorezeptor wird vom eigenen freigesetzten Neurotransmitter aktiviert. Präsynaptische Rezeptoren werden von anderen Neurotransmittern aktiviert. Postsynaptische Rezeptoren können herunter­ oder hochreguliert werden. Die Synapse ist sehr veränderbar und paßt sich an den Bedarf an. 68 / 77 Neurobiologie Wenn ein Ligand an einen Rezeptor bindet, kann das zur Phosphorylierung des Rezeptors und diese zur Endocytose des Rezeptors führen. Die Endocytose erfolgt indem Clathrin­Proteine an die Membran binden und die Einstülpung der Membran bewirken. Die GTPase Dynamin trennt dann die Einstülpung ab, sodaß sie ein Vesikel wird. Nach der Endocytose kann der Rezeptor recycled oder abgebaut werden (oder weiter im signaling­pathway verwendet werden). (Bild siehe Folie 11.7) Es gibt relativ viel Austausch von Rezeptoren zwischen Zellmembran und Membranen im inneren der Zelle (Vesikel, ER, Golgi). Durch Regulation der Neusynthese und des Austauschs zwischen den Membranen kann die Menge der Rezeptoren in den Synapsen gesteuert werden. Neue Rezeptoren kommen über ER und Golgi­Apparat in Vesikel, die dann enlangt von Microtubuli zu den Synapsen gebracht werden. (Bild siehe Folie 11.8 (AP2 = adaptor protein 2 (hilft beim Bilden der Vesikel durch Clathrin; es hält die Ladung (hier die Rezeptoren) in der Einstülpung))) Verschiedene Synapsen des selben Neurons können verschiedene Rezeptoren enthalten. Damit ein Rezeptor nur an bestimmte Synapsen geliefert wird, werden für verschiedene Rezeptoren auch verschiedene Adaptoren und verschiedene Motorproteine für den Transport zur Synapse verwendet. Adaptorproteine (z.B.: PDZ, GRIP1) binden die Rezeptoren (und damit die Vesikel, in deren Membran sich die Rezeptoren befinden) an die Motorproteine (z.B.: KIF17; KIF5 A,B,C). Diese Adaptoproteine spielen nicht nur beim Transport, sondern auch noch an der Synapse eine Rolle. (Bilder siehe Folie 11.9) Wenn eine Synapse überstimuliert wird, so wird die Menge an Rezeptoren in dieser Synapse reduziert. Es ist möglich, daß benachbarte Synapsen verschieden stark stimuliert werden und somit können auch benachbarte Synapsen unterschiedlich viele Rezeptoren haben. Medikamente beeinflussen die Plastizität der Synapse: Medikament Folgen Hemmung der Biosynthese der Neurotransmitter Unterfunktion der Synapse supersensitive Rezeptoren Aktivierung der Freisetzung der Neurotransmitter Überfunktion der Synapse subsensitive Rezeptoren Blockade der Rezeptoren supersensitive Rezeptoren erhöhte Freisetzung des Neurotransmitters erhöhter Turnover120 Aktivierung der Rezeptoren subsensitive Rezeptoren veringerte Freisetung des Neurotransmitters verringerter Turnover Hemmung der Biosynthese führt zur Unterfunktion des Synapse. Um dieser Unterfunktion entgegen zu wirken wird die postsynaptische Seite der Synapse sensitiver. (Es werden auch kurze Zeit mehr Neurotransmitter aus den Vesikeln freigesetzt, aber da weniger produziert werden und somit weniger nachgeliefert werden, verringert sich die Freisetzung.) Das System versucht Veränderungen auszugleichen. Bsp.: Wenn man Nasentropfen gegen Schnupfen nimmt, so wirken sie beim ersten Mal schnell und für lange Zeit. Aber mit jedem Mal, wenn man sie nimmt, wirken sie langsamer, weniger gut und weniger lang. 120 Turnover gibt an wie viel pro Zeiteinheit umgesetzt (Freisetzung, Wiederaufnahme, Metabolisierung) wird 69 / 77 Neurobiologie Wenn ein Medikament, das die Aktivität der Synapse verstärkt, lange genommen wird, so werden die Rezeptoren in der Synapse herunterreguliert. (Das kann Entzugserscheinungen zur Folge haben, wenn das Medikament dann nicht mehr genommen wird.) Lernen Lernen hängt mit der Plastizität des Gehirn zusammen. Informationen können in Form von Veränderungen der Synapsen gespeichert werden. Drei Formen des Lernens (von nicht­deklarativem Wissen): • Gewöhnung (E: habituation): Ein schwacher Stimulus erzeugt Toleranz für diesen Stimulus. (Ein wiederholter Reiz bewirkt dauerhafte Abschwächung der Reaktion auf diesen Reiz.) (Z.B.: eine Armbanduhr drückt immer leicht auf den Arm, aber dieser Druck wird überlicherweise nicht mehr wahrgenommen.) (Dies ist eine Form von nicht­assoziativem Lernen.) • Sensitisierung (E: sensitization): Ein starker Stimulus erhöht defensive Reflexe gegen diesen Stimulus. (Ein Reiz bewirkt dauerhafte Verstärkung der Reaktion auf diesen Reiz.) (Dies ist eine Form von nicht­assoziativem Lernen.) • Klassische Konditionierung (E: classical conditioning): (Indem gleichzeitig 2 Stimuli erfolgen, von denen der erste mit einer bestimmten Reaktion verbunden ist, wird auch der zweite direkt mit dieser Reaktion verbunden, sodaß er danach auch diese Reaktion hervorruft (selbst wenn er dann ohne den ersten erfolgt).) (Dies ist eine Form von assoziativem Lernen.) Die Meeresschnecke Aplysia californica hat nur wenige Neuronen (ca 20000), von denen einige (ca 50) sehr groß (bis zu 1mm Druchmesser) sind, und wurde daher von Eric Kandel für die Erforschung des Lernens verwendet. (Z.B. können die Aktionspotentiale im prä­ und im postsynaptischen Neuron einer bestimmten Synapse gemessen und somit Verstärkung oder Abschwächung der Synapse beim Lernen direkt verfolgt werden.) (Eric Kandel bekam für diese Arbeiten den Nobelpreis). (Bild von Aplysia siehe Folie 11.13) An Aplysia können die 3 Fromen des Lernen erforscht werden: • Gewöhung: Wenn Aplysia am Siphon leicht berührt wird, zieht sie die Kieme ein. Wenn dies öfters durchgeführt wird, wird die Kieme bei einer leichten Berührung nicht mehr eingezogen. • Sensitisierung: Erfolgen die Berührungen des Siphons bei Schmerzen (z.B.: wenn ein Elektroschock (z.B. am Schwanz) erfolgt) wird die Kieme bei jeder leichten Berührung immer sofort eingezogen (auch bei späteren Berührungen ohne Elektroschock). • Assoziation (z.B.: klassische Konditionierung): Wenn kurz (<1 Sekunde) nach der Berührung des Siphons ein Elektroschock (z.B. am Schwanz) erfolgt und das einige Male wiederholt wird, bewirkt danach schon der Elektroschock alleine das Zurückziehen der Kieme. Bei der klassischen Konditionierung wird ein Stimmulus mit einem anderen verknüpft indem beide gleichzeitig erfolgen. Einer der beiden Stimuli löst von vornherein eine Reaktion aus (z.B. Sehen eines Stück Fleisches löst bei einem Hund Speichelfluß aus). Dieser wird als unkonditionierter Stimulus (E: unconditioned stimulus (US)) bezeichnet. Der andere (dieser löst von vornherein keine Reaktion aus) wird als kontitionierter Stimulus (E: conditional stimulus (CS)) bezeichnet (z.B.: Läuten einer Glocke). Wenn beide Stimuli mehrmals ca gleichzeitig erfolgen, bewirkt danach der kontitionierte Stimulus alleine auch schon die selbe Reaktion wie zuvor nur der unkonditionierte Stimulus. (Z.B.: Wenn 70 / 77 Neurobiologie einem Hund mehrmals ein Stück Fleisch gezeigt und immer gleichzeitig eine Glocke geläutet wird, bewirkt dann das Läuten der Glocke alleine schon Speichelfluß.) (Die erlernte Reaktion zum kontitionierten Stimulus wird als bedingter Reflex (E: conditioned response (CR)) bezeichnet.) Klassische Konditionierung wurde von Iwan Pawlow (E: Ivan Pavlov) erforscht. Als Beispiel wird meist der Pawlowsche Hund verwendet. LTP und LTD Langzeit­Potenzierung (E: long term potentation (LTP)) ist eine lang andauernde Steigerung der Effektivität synaptischer Übertragung, die durch bestimmte Arten von Konditionierung bewirkt wird. Der Tractus perforans (E: perforant path) geht von der Area entorhinalis (E: entorhinal cortex) zu den Körnerzellen (E: granule cells) im Gyrus dentatus des Hippocampus. (Area entorhinalis, Tractus perforans und Hippocampus sind Teil der Hippocampusformation, die sich im Lobus temporalis befindet.) (Der Hippocampus besteht aus Gyrus dentatus und Cornu ammonis. CA1, CA2, CA3, CA4 sind Teile des Cornu ammonis (CA).) (In dem Beispiel auf Folie 11.15 wurde die LTP der Synapsen zwischen Tractus perforans und Körnerzellen untersucht. (Tetanus ist eine Art von (hochfrequenter) repetitiver Stimulation. (Z.B. wenn die Frequenz der Aktionspotetniale, die bei einem Muskel ankommen, hoch ist, dann verschmelzen die einzelnen Muskelkontrationen zu einer einzigen lange Kontraktion, die als vollständiger Tetanus bezeichnet wird.))) LTP entspricht der Sensitisierung. Ein wiederholter starker Stimulus bewirkt, daß danach ein schwacher Stimulus eine stärkere Reaktion hervorruft. Bsp.: In einem Bündel präsynaptischer Axone (im Tractus perforans) können durch elektrische Stimuli Aktionspotentiale ausgelöst werden. Wenn nur ein einzelner solcher Stimulus erfolgt, bewirkt das eine leichtes EPSP im den (postsynaptischen) Körnerzellen. Wenn schnell hintereinander viele solche Stimuli erfolgen, sodaß die postsynaptischen Zellen stark depolarisiert werden, dann bewirkt danach ein einzelner Stimulus auch ein starkes EPSP in den postsynaptischen Zellen. (LTP erfolgt genau dann, wenn präsynaptische Zelle und postsynaptische Zelle gleichzeitig starkt depolarisiert sind. Das kann z.B. durch viele Aktionspotentiale in kurzer Zeit erreicht werden: die präsynaptische Zelle ist während eines Aktionspotentials ohnehin depolaristiert und in der postsynaptische Zelle summieren sich die EPSP zu einer starken Depolarisation auf.) NMDA Rezeptor Antagonisten blockieren LTP und Lernen. Es sind also NMDA­Rezeptoren für LTP verantwortlich. LTP ist ein Ca++­abhängiger Prozess (NMDA­Rezeptoren sind Ionenkanäle für Ca++ (und andere Ionen)). Es gibt jedoch viele verschiedene Mechanismen der LTP und der hier beschriebene ist nur einer davon. Der NMDA­Rezeptor ist ein ionotroper Glutamat Rezeptor (siehe Seite 50). NMDA­Rezeptoren öffenen sich wenn Glutamat gebunden ist, werden aber sofort von Mg++ verstopft. Das Mg++ gibt den Ionenkanal erst frei, wenn die Membran depolarisiert ist. Dann erst können andere Ionen (vor allem Ca++) den NMDA­Rezeptor passieren. Da NMDA­Rezeptoren von Mg++ bei ­65mV verschlossen und bei ­30 mV freigegeben werden, sind sie spannungsabhängig (zusätzlich zur ihrer Glutamatabhängigkeit). NMDA­Rezeptoren alleine haben keine Wirkung, weil die Membran zuerst durch andere Ionenkanäle depolaristiert werden muß. Meist kommen NMDA­Rezeptoren zusammen mit AMPA­ Rezeptoren vor. Besondere Eigenschaften der NMDA­Rezeptoren: • zeitliche Integration: NMDA­Rezeptoren funktionieren als Koinzidenz­Detektoren. Sie 71 / 77 Neurobiologie • • erkennen wenn präsynaptische (Glutamatfreisetzung) und postsynaptische Zelle (Depolarisation der Membran) gleichzeitig erregt sind. LTP: NMDA­Rezeptoren bewirken aktivitätsabhängige Änderungen der synaptischen Effizienz rhythmische Aktivität: NMDA­Rezeptoren spielen eine Rolle bei der Erzeugung von Rhythmen. Ein Aktionspotential in der präsynaptischen Zelle bewirkt die Freisetzung von Glutamat aus der präsynaptischen Zelle. Das Glutamat bindet an AMPA­Rezeptoren (AMPAR) und NMDA­ Rezeptoren (NMDAR) der postsynaptischen Zelle, die AMPA­Rezeptoren öffnen sich und es entsteht ein EPSP im Dendriten der postsynaptischen Zelle. Ein einziges EPSP an einer Synapse reicht üblicherweise nicht für eine Depolatisation der postsynaptsichen Zelle (und damit für ein neues Aktionspotential) aus (es ist nicht stark genug, um sich bis zum Axon­Hügel auszuwirken). Wenn viele Aktionspotentiale innerhalb kurzer Zeit in der Synapse ankommen oder an vielen Synapsen der selben postsynaptischen Zelle ca gleichzeitig Aktionspotentiale ankommen, so reicht die Summe der EPSP aus um die postsynaptische Zelle so weit zu depolarisieren, daß ein neues Aktionspotential (im Axon der postsynaptischen Zelle) entsteht (die Summe der EPSPs wirkt sich bis zum Axon­Hügel aus). Das neue Aktionspotential kann sich auch zurück in die Dendriten auswirken und die Depolarisation der Membran der Dendriten verstärken (BPAP (back­propagating action potential)). Wenn die Depolarisation der postsynaptischen Zelle (solange noch Glutamat in der Synapse ist) stark genug ist, öffnen sich auch die NMDA­Rezeptoren und Ca++ strömt in die Zelle ein. (Weiters öffnen sich auch spannungsabhängige Ca++­Kanäle (VGCC (voltage­gated calcium channel)).) Durch den Ca++­Einstrom werden Kinasen aktiviert, die lokale Änderungen der Synapse und Änderngen der Transcription im Zellkern bewirken können. Dadurch kommt es zur Verstärkung der Synapse. (Es werden dabei nicht alle Synapsen der postsynaptischen Zelle verstärkt, sondern nur die, die aktiv waren.) (Bild siehe Folie 11.19) Alles, was gleichzeitig passiert, wird gemerkt. Plastizität gibt es nur zwischen gleichzeitig aktiven Neuronen. Bei der Sensitisierung erfolgt ein starker Stimulus (d.h. viele Aktionspotentiale in kurzer Zeit), der die NMDA­Rezeptoren aktiviert. Bei der Assoziation erfolgen ein schwacher (1 Aktionspotential) und ein starker (viele Aktionspotentiale) Stimulus über verschiedene Synapsen gleichzeitig. Durch das starke Signal wird das postsynaptsiche Neuron aktiviert (somit die Membran depolarisiert) und, weil gleichzeitig das schwache Signal in seiner Synapse die Freisetzung von Glutamat bewirkt, öffnen sich auch NMDA­ Rezeptoren in der Synapse über die das schwache Signal gekommen ist. Somit wird die Synapse des schwachen Signals verstärkt und kann ab dann ein stärkeres EPSP erzeugen, auch ohne das andere (starke) Signal. Bsp.: Wenn man eine heiße Herdplatte angreift, ist das schwache Signal das Sehen der Herdplatte und das starke Signal ist der Schmerz. Ein Neuron, das beide Signale aus verschiedenen präsynaptischen Neuronen bekommt wird durch das Schmerz­Signal aktiviert und verstärkt dann auch die Synapse über die zur gleichen Zeit das Herdplatte­Signal gekommen ist. Wenn man dann später wieder eine Herdplatte sieht (d.h. ein Herdplatte­Signal kommt), reicht das aus um dieses postsynaptische Neuron zu aktivieren und man erinnert sich an den Schmerz vom letzten Mal (ohne daß man dazu die Herdplatte angreifen müßte). Bei der Sensitisierung aktiviert ein einzelner starker Reiz AMPA­ und NMDA­Rezeptoren. Die 72 / 77 Neurobiologie NMDA­Rezptoren bewirken Ca++­Einstrom und führen somit zu einer schwachen Erhöhung der synaptischen Effizienz. Durch Wiederholung verstärkt sich die synaptische Effizienz immer mehr. Lernen erfordert Wiederholungen. Assoziatives Lernen geht besser. Zum Lernen gehört auch eine aktive Korrektur. Diese ist durch LTD (long term depression) möglich. (Vergessen ist LTD.) Bsp.: Die Schaffer­Kollateralen (Schaffer collateral) sind Nervenbahnen im Hippocampus, die CA3 mit CA1 verbinden. Wenn in den Schaffer­Kollateralen Aktionspotentiale mit niedriger Frequenz (1­3Hz) für ca 5­10 Minuten erzeugt werden, bewirkt das eine LTD der Synapsen zwischen Schaffer­Kollateralen und den Pyramidenzellen im CA1. Einzelne Aktionspotentiale lösen danach viel geringere EPSP in den Pyramidenzellen aus als vorher. LTD kommt in vielen Hirnregionen vor. Auch für die LTD gibt es mehrere Mechanismen. Der hier beschriebene Mechanismus der LTD ist dem Mechanismus der LTP sehr ähnlich. Allerdings erfolgt bei der LTD nur ein geringer Ca++­ Einstrom durch die NMDA­Rezeptoren. D.h. geringer Ca++­Einstrom bewirkt LTD und starker Ca++­ Einstrom bewirkt LTP. (Bilder siehe Folie 11.21) (Ein Beispiel für LTD ist das Abgewöhnen einer Angst vor (z.B.) Spinnen: Am Anfang sieht man sich ein Bild einer Spinne an, später eine Spinne aus größere Entfernung, dann aus geringerer Entfernung, ... . Dadurch gewöhnt man sich an den Anblick der Spinne und gewöhnt sich dabei die Angst ab.) Auch bei motorischem Lernen kann mittels LTD korrigiert werden. Lernen ist Anpassung und Anpassung muß in beide Richtungen gehen (merken und vergessen). Motorisches Lernen im Cerebellum korrigiert die Muskelkoordination in beide Richtungen. Das Gedächtnis sitzt in den beteiligten Hirnregionen (motorische Programme im Rückenmark, motorisches Lernen im Cerebellum). Im Rahmen der Plastizität können sich nicht nur Synapsen verstärken, sondern es können auch neue Synapsen entstehen. Wenn aus einer Synapse zwei werden, wird dadurch die Stärke der Verbindung verdoppelt. Für das Langzeitgedächtnis ist Umbau/Neusynthese von Synapsen und damit Proteinbiosynthese nötig. Der Patient H.M. hatte Temporallappen­Epilepsie. Diese wurde geheilt indem der ganze Temporallappen chirurgisch entfernt wurde. Danach konnte sich der Patient H.M. zwar an alles vor der Operation erinnern, aber Hippocamus bedingtes Lernen war ihm nicht mehr möglich. D.h. er konnte sich an eben gelerntes nicht erinnern. (Z.B. erkannte er Leute, die er 5 Minuten zuvor gesehen hatte, nicht wieder.) Motorisches Lernen war ihm aber noch möglich. (Z.B.: Nachziehen einer gezeichneten Figur mit einem Stift konnte sich bei täglichem Üben über mehrere Tage hinweg verbessern.) Gedächtnis, Emotionen Gedächtnis Gedächtnis ist die Aufbewahrung von gelernten Informationen (Merk­ und Erinnerungsfähigkeit). Das Gedächtnis kann nach der Dauerhaftigkeit der Informationsspeicherung eingeteilt werden in: • Ultrakurzzeit Gedächtnis (E: working memory) (Sekunden): Es kommt durch kurzzeitig 73 / 77 Neurobiologie • • aktive Hirnbahnen zustande. Das Ultrakurzzeitgedächtnis ist das Arbeitsgedächtnis. Es enthält Informationen, an die man gerade denkt (z.B.: eine Telefonnummer in der Zeit zwischen dem Lesen im Telefonbuch und dem Wählen am Telefon). Kurzzeit Gedächdnis (E: short­term memory) (Minuten, Stunden): Es kommt durch postsynaptische Mechanismen (erhöhte Rezeptoraktivität (durch Phosphorylierung) und mehr Rezeptoren in der Membran) und präsynaptische Mechanismen (verstärkte Freisetzung von Glutamat) zustande. Langzeit Gedächtnis (E: long­term memory) (Tage, Wochen Jahre): Es kommt durch Protein Synthese und prä­ und postsynaptische Strukturänderungen zustande (neue Synapsen, Verstärkung von Synapsen). Das Gedächtnis kann auch nach der Art der Information eingeteilt werden: • deklaratives Gedächtnis: Dies ist das Gedächtnis für Tatsachen (Schulwissen, Ereignisse, ...). Die Speicherung erfolgt über den Hippocampus. (An diesem Gedächtnis sind der (mediale Teil des) Lobus temporalis und das Diencephalon beteiligt.) • nicht­deklaratives (unbewußtes) Gedächtnis: Dies ist das Gedächtnis für Fähigkeiten, Gewohnheiten, Programme der Skelettmuskulatur und emotionelle Reaktionen. (z.B.: Musikinstrumente spielen (man kann nicht sagen was man genau macht (z.B.: wie viele Millisekunden zwischen dem Drücken auf die Seiten sind)); Angst wenn man eine Schlange oder Spinne sieht (wenn man noch nie eine gesehen hat, weiß man nicht, daß man davor Angst hat); Werfen des Balls beim Baseball; ...) Emotionen Emotionen sind unsere ganz persönlichen Gefühle gegenüber Personen und Situationen. Sie sind sehr spezifisch für die jeweilige Person (verschiedene Personen haben ganz unterschiedliche Emotionen für die selben Personen und Situationen). Wir lernen früh, Ereignisse mit uns in Beziehung zu setzen. Wenn etwas für einen selbst gut und angenehm ist, wird man versuchen, diese Situation wieder herbeizuführen. Wenn etwas für einen selbst unangenehem oder bedrohlich ist, wird man versuchen, diese Situation zu vermeiden. (Situationen und Menschen werden bewertet: z.B.: auf wen kann man sich verlassen, wenn man Hilfe braucht, ...) Daraus entwickeln sich unsere Emotionen: • Zuneigung, Freundschaft, Liebe • Abneigung, Feindschaft, Hass • Angst, Eifersucht • Motivation, Ziele zu erreichen (z.B.: positive Gefühle für einen Menschen haben das Ziel zur Folge, mit diesem Menschen mehr zusammen zu sein) (z.B.: Eifersucht bewirkt, daß man sich besonders viel um seine Freundin kümmert) Alle Erlebnisse und Situationen werden auch emotional beurteilt. (z.B.: Wenn einem ein Arzt mit einem Gummihammer auf das Knie schlägt, ist man deshalb nicht böse auf den Arzt. Wenn einem aber jemand, der einem unsympathisch ist, auf das Knie schlägt, ist man sehr wohl böse.) Emotionale Erinnerungen sind sehr langelebig und schwer zu ändern. Emotionen prägen unser gesamtes Leben. Emotionen können nicht nur auf Personen, sondern auch auf Situationen bezogen sein. Die Emotionen entstehen in limbischen Hirnarealen. Das limbische System (und was dazu gehört) ist nicht genau definiert. Lobus limbicus war ursprünglich eine 74 / 77 Neurobiologie Zusammenfassung der Strukturen rund um das Corpus callosum. Viele dieser Strukturen haben mit Emotionen zu tun. Das limbische System sind Strukturen rund um das Corpus callosum und/oder Strukturen die (vermutlich) mit Emotionen zu tun haben. (Es gibt Strukturen, die beim Corpus callosum liegen, aber nichts mir Emotionen zu tun haben und es gibt Strukturen, die nicht beim Corpus callosum liegen, aber schon mit Emotionen zu tun haben.) Das limbische System liegt zum Teil (z.B.: Amygdala, Area septalis) im Telencephalon, zum Teil (z.B.: Corpus mammilare) im Diencephalon und zum Teil im Mesencephalon. Zum limbischen121 System gehören auf jeden Fall: • Hippocampus: Der Hippocampus hat nicht direkt mit Emotionen zu tun, sondern ermöglicht es, sich an den Kontext der Emotionen zu erinnern. • Area entorhinalis • Gyrus cinguli (enthält die Area cingularis (E: cingulate cortex)) • Area septalis (Septum): (Der Strich der Bezeichnung "Septum" im Bild (Folie 12.11) geht zum Septum • pellucidum, das nur ein Teil der Area septalis ist und (fast) keine Neuronen enthält.) Nucleus accumbens (liegt im Telencephalon) Amygdala: Die Amygdala ist sehr wichtig für Emotionen (vor allem für Furcht, Angst und Aggression). • Corpus mammillare (E: mamillary body) (liegt im Diencephalon) • Thalamus • Fornix (ist ein wichtiges Faserbündel und verbindet Hippocampos, Area entorhinalis, ...) • und einige weitere Strukturen Das limbische System befindet sich im inneren Teil des Cortex und ist phylogenetisch sehr alt. Auch Tiere haben ein limbisches System und Emotionen. • Furcht und Angst Furcht und Angst sind Gefühle der Bedrohung des Individuums. Sie sind Primäremotionen, wie Liebe, Hass, Aggression und wichtig für unser überleben. Furcht ist etwas reales, eine konkrete Situation, die jetzt da ist. (z.B.: wenn jemand mit einer Maschinenpistole zur Tür hereinkommt) Angst ist etwas (noch) nicht reales. (z.B.: es könnte jemand mit einer Maschinenpistole zur Tür hereinkommen) Lebewesen, die Angst/Furcht haben überleben länger. (Ein Tier, das sich nicht vor Raubtieren fürchtet, wird nicht lange überleben.) Angst ist sehr spezifisch (bei jeder Person anders). Auch Tiere haben Angst. (Versuch: 2 gekreuzte Stege (in Form eines "+") in für das Tier großer Höhe, wobei einer davon Wände hat und der andere nicht. Ein Tier, das Angst (vor dem hinunterfallen) hat, geht nur auf dem Steg mit den Wänden.) Tranquilizer122 sind Medikamente, die angslösend und entspannend wirken. Auch Alkohol wirkt angstlösend und hilft Angst zu überwinden. Einige Beispiele für Tranquilizer sind (Strukturformeln siehe Folie 12.13): Ethanol (Alkohol), Phenobarbital (ein Barbiturat), Methaqualon (Methylquinazolon), Meprobamat Viele Tranquilizer sind Benzodiazepine. Benzodiazepine sind die stärksten angstlösenden Medikamente. Einige Beispiele für Benzodiazepine, die als Tranquilizer verwendet werden sind (Strukturformeln siehe Folie 12.14): Diazepam (Valium®), Chlordiazepoxid (Librium®), Bromazepam (Lexotanil®), Flunitrazepam (Rohypnol®) Eine Nebenwirkung der angstlösenden Medikamente ist, daß sie alle mehr oder weniger müde machen. 121 Limbus = lat. Rand (weil das limbische System rund um das Corpus callosum liegt, also dessen Rand bildet) 122 engl. tranquillize = beruhigen 75 / 77 Neurobiologie Der GABAA­Rezeptor ist ein Cl­­Ionenkanal und somit (meist) ein inhibiorischer Rezeptor. Benzodiazepine binden an eine bestimmte Stelle (Benzodiazepin­Bindungstelle(/Rezeptor)) der GABAA­Rezeptoren und bewirken, daß die Rezeptoren stärker auf den Neurotransmitter GABA reagieren. (GABAA­Rezeptoen haben auch eine Bindungsstelle für Ethanol, das dadurch auch die Wirkung von GABA verstärkt.) Die dadurch verstärkte inhibitorische Wirkung führt zur Beruhigung. Midazolam und Bretazenil (Ro16­6028) sind Benzodiazepine, die Angst reduzieren. Flumazenil kann als Gegenmittel bei Überdosierungen von Benzodiazepin­Tranquilizern verwendet werden. Über die Benzodiazepin­Bindungstselle von GABAA­Rezeptoren kann Angst nicht nur reduziert, sondern auch ausgelöst werden. Ro15­4513 und Ro19­4603 haben die gegenteilige Wirkung von Benzodiazepin­Tranquilizern und lösen somit Angst aus. (Bei Versuchen an Freiwilligen traten nach Verabreichung solcher angstauslösender Stoffe massive Angstzustände auf.) GABAA­Rezeptoren modulieren die Angst direkt (sonst könnte über sie keine Angst ausgelöst werden). Auch andere Systeme können Angst beeinflussen: Serotonin (5­HT) (es gibt auch Tranquilizer, die auf das Serotonin­System wirken (SSRI123 verlangsamen die Wiederaufnahme von Serotonin, sodaß es länger in der Synapse bleibt und somit länger/stärker wirkt)), Noradrenalin (NE), CCK (Cholecystokinin), ... (Bilder (inklusive Strukturformeln) siehe Folie 12.15) Motivation Die Gehirnbereiche des Motivations­ und Belohnungssystems wurden durch Selbst­Stimulations Experimente identifiziert: Einer Ratte wurde eine Elektrode ins Gehirn implantiert, sodaß das Gehirn lokal stimuliert werden konnte. Dann konnte die Ratte über eine Taste in ihrem Käfig die Elektrode (und damit die Stimulation) selbst auslösen. Je nachdem in welchem Teil des Gehirns die Elektrode implantiert war, wurde die Elektrode von der Ratte öfter oder weniger oft ausgelöst. Dadurch konnten die Gehirnteile identifiziert werden, in denen die Stimulation von der Ratte am meisten belohnend empfunden wurde, und die somit zum Motivations­ und Belohnungssystem gehören. Am meisten wurden Gehirnteile von den Ratten selbst stimuliert, die die Freisetzung von Dopamin bewirken, vor allem dopaminerge Neuronen im ventralen Tegmentum (E: ventral tegmental area). Der wichtigste Teil des Motivationssystems ist das ventrale Tegmentum. Weiters spielen auch Amygdala, Nucleus accumbens, Hypothalamus und Septum eine Rolle. (Das mesolimbische Dopamin­System ist der Teil des Gehirns, der für Motivation zuständig ist. Es besteht aus dem ventralen Tegmentum, dem Nucleus accumbens, der Amygdala und weiteren Strukturen. (Die mesolimbische Bahn verbindet das ventrale Tegmentum mit dem limbischen System.)) Es wird durch Acetylcholin, Enkephalin und Substance P stimuliert. Das Motivations­ und Belohnungssystem unterstützt Verhalten, das Überleben sichert. Essen, Trinken, Sex werden als angenehm empfunden und daher wiederholt ausgeführt. Es wird von Sinnesreizen (z.B.: süßer Geschmack, Gewürze, Geruch, Aussehen der Speisen, nette Umgebung, soziale Interaktion, Sympathie) unterstützt. Das System ist erinnerungsfähig. (Wenn man weiß, daß ein Lokal gut ist, geht man immer wieder hin. Man erinnert sich an das Essen und den Kontext.) Es beeinflußt unser Verhalten. (Wenn etwas als angenehm empfunden wird, wird es oft wiederholt.) Das System ist stets offen für neue Aktivitäten: 123 SSRI = serotonin­selective reuptake inhibitors 76 / 77 Neurobiologie Neugier: z.B.: man probiert auch andere Lokale Hoffnung auf Wohlbefinden, Zufriedenheit, Glück, Macht • unabhängig von Kosten und Risiko verfolgt (z.B.: bei Sucht) • löst oft komplexe Verhaltensweisen aus (z.B.: Leute fahren einem Theaterstück nach, wenn es in einer anderen Stadt aufgeführt wird) Eigene Motivation wird durch äußere Motivation verstärkt: • Freunde, gemeinsames Erleben: z.B.: Dinge, die von Freunden gelobt werden erscheinen positiver • frühere positive Erfahrungen mit ähnlichen Aktivitäten: z.B.: man kommt wieder gerne mit Freunden zusammen Motivation ist im Entzug erhöht (z.B.: wenn man hungrig ist, schmeckt das Essen besser). • • All das trifft auch auf Suchtdrogen zu. Motivation bewirkt Planung und Durchführung, sodaß es zur Belohnung kommt und diese vertärkt die Motivation. Alle Suchtdrogen (z.B.: Kokain, Amphetamine, Heroin, Ethanol, Benzodiazepine, Cannabinoide, Nikotin) modulieren/stimulieren direkt oder indirekt das mesolimbische Dopamin­System. Sie brauchen nicht Spaß machen (z.B.: die erste Zigarette schmeckt niemandem). Sie lösen einen Belohnungsreiz aus und dieser führt zur Wiederholung. Die Suchtmitteleinnahme und ihr Kontext wird belohnt. Suchtmittel haben eine stärkere und länger andauernde Wirkung als andere Dinge, die Belohnung auslösen. Dies führt zur hinunter­Regulation des Motivations­/Belohungssystems, also zur Toleranz: Motivation und Stimmung sind reduziert und (nur) eine erneute Drogeneinnahme verhilft wieder zu Wohlbefinden. Die Toleranz führt damit zu Dosissteigerungen. Bei Abwesenheit der Droge kommt es zu Entzugssymptomatik aufgrund physischer Abhängigkeit. Nachdem die Funktion der Rezeptoren (um die Überstimulierung auszugleichen) reduziert wurde, ist ohne die Droge die Übertragung in den Synapsen nicht mehr ausreichend. (Entziehungen werden heute mit Medikamenten durchgefüht.) Beim Entzug ist ein wesentliches Problem, bei dem Medikamente nicht helfen, die psychische Abhängigkeit ("craving"124) aufgrund von Kontext­Lernen: Die Droge bewirkt Belohnung und diese Belohnung wird im Kontext gemerkt (mit wem man geraucht hat, welche Empfindungen man gehabt hat, ...). (Z.B.: Wenn jemand, der erfolgreich einen Entzug gemacht hat, und physisch nicht mehr von der Droge abhängig ist, wieder mit den Leuten zusammenkommt, mit denen er die Droge genommen hat, kann die Erinnerung an diesen Kontext des Droge­Nehmens zu einem Rückfall führen.) Die Suchtmittel­Einnahme ist oft belohnt worden und ist daher ein überragendes Ziel. Das "Drogengedächtnis" erinnert auch an die Umstände des Drogenkonsums. Sucht ist der Kontrollverlust über die Drogeneinnahme und die zwanghafte Suche und Einnahme von Drogen unabhängig von den Konsequenzen. Sucht ist nicht auf Drogen beschränkt. Beispiele für andere Dinge, auf die sich Sucht beziehen kann: Essen (z.B.: Schokolade), Arbeit, Sex, Shopping, Sammelleidenschaft (Briefmarken, Schuhe, CDs, ...), Publicity, Training und Fitness, ... (Dinge bei denen man sich sehr unwohl fühlt, wenn man sie nicht mehr hat.) 124 engl. craving = heftige Begierde 77 / 77 Neurobiologie