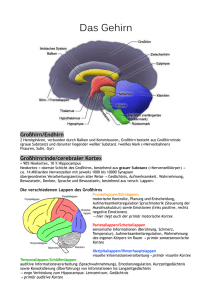
Legasthenie und reine Worttaubheit als neurologisch basale St
Werbung

/HJDVWKHQLHXQGUHLQH:RUWWDXEKHLWDOV QHXURORJLVFKEDVDOH6W|UXQJHQ Inaugural-Dissertation zur Erlangung der Doktorwürde der Philosophischen Fakultät IV (Sprach- und Literaturwissenschaften) der Universität Regensburg vorgelegt von Barbara Limmer Otlohstraße 15 93053 Regensburg 2001 Erstgutachter: Prof. Dr. Herbert E. Brekle Zweitgutachter: PD Dr. habil. Helmut Weiß Vorwort Ich möchte diesen Platz nutzen, um denjenigen zu danken, die maßgeblich daran beteiligt waren, daß dieses Dissertationsprojekt zu einem Abschluß gelangen konnte. Die Zahl der zu Bedenkenden ist groß, so daß es mir unmöglich erscheint, alle namentlich zu erwähnen. Ich möchte jedoch die wichtigsten anführen. Allen voran danke ich meinen Eltern, die mir ein Studium ermöglichten und ohne deren Unterstützung diese Arbeit nicht möglich gewesen wäre. Außerdem danke ich meinem Doktorvater, Prof. Dr. Herbert E. Brekle, der bereit war, mir ein Thema aus der Neurolinguistik zu gewähren und mir zur Bearbeitung den nötigen Freiraum ließ, für seine unermüdliche Diskussionsbereitschaft, wodurch viele seiner Gedanken Einzug in diese Arbeit gefunden haben, ohne daß sie explizit gekennzeichnet werden könnten. Seine Emeritierung im Oktober 2001 bedeutet einen unersetzbaren Verlust für die Universität Regensburg. Ebenso will ich mich bei PD Dr. habil. Helmut Weiß dafür bedanken, daß er sich als Zweitgutachter zur Verfügung gestellt hat. Weiterer Dank gilt allen Mitarbeitern des Lehrstuhls für Allgemeine Sprachwissenschaft und anderen Beschäftigten der Universität Regensburg, die wegen ihrer großen Zahl nicht genannt werden können. Schließlich danke ich einem lieben Freund für die Korrektur und Recherchearbeiten. Die zweifellos verbleibenden Fehler und Unzulänglichkeiten der Arbeit sind selbstverständlich mir anzulasten. Dieses Dissertationsprojekt wurde teilweise finanziert durch ein Stipendium der Universität Regensburg. Regensburg im August 2002 Barbara Limmer Inhaltsverzeichnis I ,QKDOWVYHU]HLFKQLV INHALTSVERZEICHNIS I ABBILDUNGSVERZEICHNIS IV TABELLENVERZEICHNIS V 1 EINLEITUNG 1 2 PROBLEMSTELLUNG 2 3 ANATOMISCHE UND PHYSIOLOGISCHE GRUNDLAGEN 4 3.1 Physiologie des Sprechapparates 3.1.1 Phonation 3.1.2 Artikulation 4 5 5 6 3.2 Kortikale Sprachzentren 3.2.1 Broca-Sprachregion 3.2.2 Wernicke-Sprachregion 3.2.3 Zusammenarbeit kortikaler Sprachzentren 3.2.4 Aphasieformen 8 8 10 10 12 3.3 Physiologie des auditiven Systems 3.3.1 Schall 3.3.2 Schallleitung zum Innenohr 3.3.3 Schalltransduktion im Innenohr 3.3.4 Signaltransformation von der Sinneszelle zum Hörnerv 3.3.5 Frequenzselektivität: Grundlage des Sprachverständnisses 3.3.6 Informationsübertragung und Verarbeitung im ZNS 14 14 16 17 19 20 21 3.4 Physiologie des visuellen Systems 3.4.1 Sehen und Blicken 3.4.2 Optisches System des Auges 3.4.3 Signalaufnahme und Signalverarbeitung in der Netzhaut 3.4.4 Signalverarbeitung im zentralen visuellen System 3.4.5 Neurophysiologische Grundlagen der Gestaltwahrnehmung 25 25 26 28 31 36 3.5 Moderne Techniken zur Lokalisation sprachrelevanter Hirnareale 3.5.1 Nuklearmedizinische Methoden 3.5.2 Intraoperativ-neurophysiologische Methoden 3.5.3 Angiographie und Wada-Test 3.5.4 Computertomographie und Magnetresonanztomagraphie 3.5.5 Grundlagen der funktionellen MR-Bildgebung (FMRI) 37 38 38 40 41 43 REINE WORTTAUBHEIT 4.1 Allgemeine Grundlagen 4.1.1 Begriffserklärungen und Definitionen 46 46 46 Inhaltsverzeichnis 4.1.2 5 6 II Historische Aspekte 48 4.2 Pathogenese 4.2.1 Reine Worttaubheit durch Störungen in den Sprachzentren des Gehirns 4.2.2 Reine Worttaubheit durch Störungen der auditiven Wahrnehmung 4.2.2.1 Untersuchungen von Dennis P. PHILLIPS 4.2.2.2 Weitere relevante Untersuchungsergebnisse 50 50 54 54 61 4.3 65 Zusammenfassung und Schlussfolgerungen LEGASTHENIE 67 5.1 Allgemeine Grundlagen 5.1.1 Begriffserklärungen und Definitionen 5.1.2 Typische Symptome 67 67 69 5.2 Pathogenese 5.2.1 Genetische Faktoren 5.2.1.1 Familien- und Segregationsstudien 5.2.1.2 Zwillingsuntersuchungen 5.2.1.3 Ergebnisse molekulargenetischer Untersuchungen 5.2.2 Auditive Verarbeitungsstörungen 5.2.2.1 Die Bedeutung der Wahrnehmung von nichtsprachlichen Reizen 5.2.2.2 Neuroanatomische Veränderungen 5.2.3 Visuelle Verarbeitungsstörungen 5.2.3.1 Magnozelluläres System 5.2.3.2 Die Bedeutung grundlegender visueller Wahrnehmungsprozesse 71 72 73 73 75 77 80 87 88 89 92 5.3 94 Zusammenfassung und Schlussfolgerungen DISKUSSION 96 6.1 Entstehungsbedingungen der reinen Worttaubheit 6.1.1 Die Postulate und Schlussfolgerungen von PHILLIPS 6.1.2 Die Pro-Position 6.1.3 Die Contra-Position 6.1.4 Schlussfolgerung 96 96 100 104 107 6.2 Entstehungsbedingungen der Legasthenie 6.2.1 Postulate und Schlussfolgerungen von TALLAL, TALCOTT und anderen 6.2.1.1 Störungen der Sehverarbeitung 6.2.1.2 Störungen der Hörverarbeitung 6.2.1.3 TALLAL-Hypothese 6.2.1.4 Neuroanatomische Veränderungen 6.2.2 Die Pro-Position 6.2.2.1 Störungen der Sehverarbeitung 6.2.2.2 Störungen der Hörverarbeitung 6.2.2.3 Legasthene Schwächen – Schwächen der raschen zeitlichen Verarbeitung 6.2.2.4 Neuroanatomische Veränderungen 6.2.3 Die Contra-Position 6.2.4 Schlussfolgerung 109 109 109 112 113 115 115 115 116 116 118 119 131 6.3 Zusammenführung der Ergebnisse aus 6.1 und 6.2 132 6.4 Zusammenfassung und Schlussfolgerung 133 Inhaltsverzeichnis 7 DIE KONSEQUENZEN FÜR DIE THERAPIE III 138 7.1 Therapie der Lese-Rechtschreib-Schwäche 7.1.1 Herkömmliche Symptomtherapie 7.1.2 Verfahren nach TALLAL und MERZENICH 7.1.3 Training des Blicklabors Freiburg 7.1.3.1 Blick-Training 7.1.3.2 Hör-Training 7.1.4 Therapie nach WARNKE 7.1.4.1 Gestörte Teilfunktionen der zentralen Hörverarbeitung 7.1.4.2 Trainingsverfahren nach WARNKE 139 139 140 143 144 146 148 149 151 7.2 153 Therapie der reinen Worttaubheit SCHLUSSBEMERKUNG 159 LITERATUR 162 Abbildungsverzeichnis IV $EELOGXQJVYHU]HLFKQLV Abbildung 1: An der Sprachkonstruktion beteiligte Hirnregionen Abbildung 2: Schematische Darstellung von äußerem, mittlerem 9 und innerem Ohr 17 Abbildung 3: Schnitt durch eine Windung der Kochlea 18 Abbildung 4: Schematische Darstellung der zentralen Hörbahn 22 Abbildung 5: Waagerechter schematischer Durchschnitt durch den linken Augapfel – von oben gesehen 27 Abbildung 6: Reduziertes Auge 27 Abbildung 7: Aufbau der Primatennetzhaut und Schema der Reaktion einzelner Neuronen auf einen Lichtreiz 28 Abbildung 8: Schema der Sehbahn im Gehirn des Menschen 32 Abbildung 9: Äußere Hirnoberfläche eines Rhesusaffen 32 Abbildung 10: Beispiel des Reizmusters „A“ 36 Abbildung 11: Schematische Zeichnung der zwei Spaltenhälften oder Chromatiden eines Chromosoms 76 Tabellenverzeichnis V 7DEHOOHQYHU]HLFKQLV Tabelle 1: Neue Ergebnisse von Zwillingsuntersuchungen 74 1 Einleitung 1 (LQOHLWXQJ Schon seit einigen Jahren sind Bildungspolitiker ebenso wie Pädagogen und Sprachwissenschaftler alarmiert. Denn immer mehr Grundschüler leiden angeblich an einer mehr oder weniger ausgeprägten Lese- und Rechtschreibschwäche (Legasthenie) und sind offenbar immer weniger in der Lage, schriftsprachliche Äußerungen zu verstehen. Es wäre sicherlich zu kurz gegriffen, die Ursachen allein in der multimedialen Reizüberflutung, dem geänderten Spiel- und Freizeitverhalten und der angeblich immer weniger stattfindenden interfamiliären Kommunikation zu suchen. Die rasanten Fortschritte in der Neurologie und der Zellforschung gaben nun Diskussionen um die Ursachen von jenen Störungen neuen Auftrieb, die sich mit der Frage beschäftigten, ob und inwieweit diese genetisch bedingt oder „krankheitsbedingt“ erworben und therapiefähig wären. In diesem Zusammenhang traten erneut Zweifel an bis dato anerkannten Paradigmen auf, die zahlreiche Autoren zu einer Wiederaufnahme von Studien über Themen wie Legasthenie und Worttaubheit veranlassten. Vorliegende Arbeit sieht sich in dieser Tradition und nimmt neueste Untersuchungen namhafter Autoren zum Anlass, die Frage nach den neurologischen Ursachen jener Phänomene genauer zu untersuchen. 2 Problemstellung 2 3UREOHPVWHOOXQJ Tests ergaben, dass Kinder, die unter den o.g. Phänomenen besonders leiden, nicht etwa einen niedrigen IQ aufweisen. Folglich müssen andere Faktoren und Ursachen existieren, um die Minderleistungen auf dem Gebiet der Hör- und Lesefähigkeit zu erklären. Diese könnten im auditiven wie im visuellen Bereich und nicht in höheren kognitiven Bereichen liegen, wofür auch neuere Studien sprechen. Dies gab im Jahr 1999 zu einer von internationalen Forschern initiierten Studie an einer Leipziger Sprachheilschule Anlass, um zu evaluieren, wie häufig diese basalen Wahrnehmungsdefizite auftreten. Die Ergebnisse bestätigen ein so genanntes „Specific Language Impairment (SLI-Syndrom)“, das, so die amerikanische Forscherin Paula TALLAL – im „akustischen Timing“ des Gehirns liegt. Nun liegt die Vermutung nahe, dass auch im visuellen Bereich diese sensorischen Wahrnehmungslücken bestehen. In der vorliegenden Arbeit wird von der Hypothese ausgegangen, dass Defizite in diesen Bereichen möglicherweise in funktional viel grundlegenderen Schichten des Gehirns zu suchen sind und eher im so genannten „vorsprachlichen Bereich“ in den basalen Hirnregionen zu lokalisieren sind. Sprache als Gesamtkomplex wäre so bedeutend weniger mächtig als bis dato angenommen, was zwangsläufig zu einem neuen Paradigma in der Legasthenie- und Worttaubheitsforschung führen würde. Gestützt werden diese Hypothesen von neuesten Forschungsergebnissen, die darauf hindeuten, dass legasthenische Defizite nicht auf Störungen beim Erkennen von Buchstaben- oder Wortbildern bzw. auf einem mangelhaften Zugriff auf Einheiten des orthographischen Gedächtnisses zurückzuführen sind. Vielmehr wurde experimentell nachgewiesen, dass die Defizite auf einer Störung beim Verarbeiten rasch wechselnder elementarer optischer Stimuli (z.B. der Richtungsänderung zweier bewegter Punkte) beruhen können. Nun existiert das Phänomen, dass dasselbe Symptom sich auch im akustischen Wahrnehmungskanal feststellen lässt: gestörte 2 Problemstellung 3 Diskriminationsfähigkeit von zwei rasch aufeinanderfolgenden Tönen bzw. deren Frequenzänderungen. Auch an Patienten, deren akustisches Spracherkennungsdefizit als Worttaubheit klassifiziert wird, zeigen neuere Untersuchungen, dass dem Syndrom offenbar eine sprach-unspezifische Störung der auditiven Wahrnehmung zugrunde liegen kann. Diese neuen Befunde sind von weitreichender Bedeutung, ergibt sich doch die Konsequenz, dass sowohl die Legasthenie als auch die Worttaubheit möglicherweise auf ein und demselben Phänomen basieren. 3 Anatomische und physiologische Grundlagen 4 $QDWRPLVFKHXQGSK\VLRORJLVFKH*UXQGODJHQ An der Sprache des Menschen sind mehrere Organsysteme beteiligt: Mit der Phonation erzeugt der Kehlkopf Schall, d.h. Stimme. Der Mund-Rachen-Raum moduliert die bei der Phonation gebildeten Töne und formt sie in verständliche Vokal- und Konsonantensequenzen um. Dieser Mechanismus wird allgemein als Artikulation bezeichnet. Phonation des Kehlkopfs und Artikulation des MundRachen-Raums werden zentral durch das motorische Sprachzentrum des Gehirns gesteuert. Sprache ist ein komplexes Phänomen und ein spezifisches Kennzeichen des Homo sapiens sapiens. Für die Entwicklung der Sprache beim Kind sowie zu ihrer ständigen Kontrolle beim Erwachsenen sind die physiologischen Hör- und Sehfunktionen erforderlich. Der Sprach-Kreis umfasst dabei u.a. die Funktionen von Ohr und Auge, der Hör- und Sehbahn, die Sprachwahrnehmung im sensorischen Sprachzentrum (Wernicke) sowie die Integration von Gefühl und Vernunft. Der Kreis setzt sich zur motorischen Steuerung der Phonation des Kehlkopfs und der Artikulation des Mund-Rachen-Raums fort. Sie beginnt in dem als motorische Sprachregion (Broca) bezeichneten Gebiet des vorderen Temporallappens im Gehirn und erreicht über neuronale Verschaltungen den Kehlkopf sowie den Mund-Rachen-Raum (Schmidt et al. 2000). Dieses System des Sprach-Kreises (Schmidt et al. 2000) soll im folgenden Kapitel im Mittelpunkt stehen. 3 Anatomische und physiologische Grundlagen 5 3K\VLRORJLHGHV6SUHFKDSSDUDWHV 3KRQDWLRQ Unter Phonation wird hier der eigentliche Sprechakt verstanden. Dieser gliedert sich in mehrere, nahezu unwillkürlich ablaufende Prozesse, die hier Schritt für Schritt nachvollzogen und erläutert werden sollen. Vor dem Sprechen wird eine Exspiration eingeleitet, bei der die Glottis geschlossen oder stark verengt wird. Als Folge entsteht im Thorax ein höherer Druck als bei normaler Exspiration, und die Stimmbänder werden auseinander gepresst. Zu diesem Zeitpunkt beginnt Luft durch die Glottis in den MundRachen-Raum zu strömen. Die Glottis bildet dabei eine Engstelle im Exspirationstrakt, als Folge davon ist die Strömungsgeschwindigkeit der Exspirationsluft dort weit höher als in der Trachea. Nach den Bernoulli-Gesetzen (Gerthsen et al. 1980) bedeutet dies, dass dort der Luftdruck so klein wird, dass sich die Glottis wieder schließt. Der Vorgang beginnt von neuem – die Stimmbänder führen permanent Bernoulli-Schwingungen aus. In ihrem Rhythmus wird der Luftstrom dabei ständig unterbrochen. So entsteht ein hörbarer Klang, die Stimme, deren Grundfrequenz, d.h. deren Tonhöhe, den Unterbrechungen des Luftstroms entspricht. Da der Luftstrom durch das Öffnen und Schließen der Stimmbänder nicht sinusförmig moduliert wird, entsteht kein reiner Ton, sondern ein Klanggemisch, das reich an Obertönen ist (Flanagan 1972). Die Häufigkeit, mit der die Glottis pro Zeiteinheit geöffnet bzw. geschlossen wird, d.h. die Grundfrequenz des erzeugten Klangs, ist in erster Linie von der Spannung der Stimmbänder und erst in zweiter Linie vom subglottischen Druck abhängig. Beide Parameter können aber durch die Kehlkopfmuskulatur und die Thoraxmuskulatur verändert werden. Je höher die Spannung der Stimmbänder bzw. je höher der subglottische Druck ist, desto höher wird die Grundfrequenz des erzeugten Klangs. 3 Anatomische und physiologische Grundlagen 6 Hierfür und zum Halten eines Tons müssen außerordentlich fein abgestimmte Kontraktionen der beteiligten Muskeln ausgeführt werden. Daran sind unter anderem Propriozeptoren in den Kehlkopfmuskeln und der Schleimhaut sowie die Kontrolle durch das Gehör beteiligt. $UWLNXODWLRQ Aus der Glottis austretende Luft gelangt zunächst in den Mund-Rachen-Raum, der als $QVDW]URKU bezeichnet wird. Dieser Raum umfasst Pharynx-, Nasen- und Mundraum. Seine Form ist sehr variabel. Das Gaumensegel (Velum) sorgt dabei für eine funktionale Abtrennung der Nasopharynx und des Rachenraums vom Mund-Rachen-Raum. Durch Zungen- und Kieferstellung kann der Mundraum in seiner Konfiguration erheblich verändert werden. Auch kann die Zunge durch Bildung eines Buckels den Mundraum in zwei Räume unterteilen. Verantwortlich für diese Veränderungen sind die Gaumenmuskulatur, die Kaumuskulatur und insbesondere die Zungenmuskulatur. Die Zunge kann dabei innerhalb des Mundraums fast jede Stellung einnehmen. Dies wird durch die Zungenbinnenmuskulatur sowie die Muskeln ermöglicht, die von verschiedenen knöchernen Ansatzpunkten in die Zunge einstrahlen, bzw. solchen, die das Zungenbein in seiner Lage verschieben können. Doch bei der Phonation entsteht nicht nur die bereits geschilderte periodische Unterbrechung des Luftstroms an der Stimmritze. Auch an anderen Engpässen innerhalb des Respirationstraktes bilden sich, sobald die Exspirationsgeschwindigkeit groß genug ist, Wirbel und Luftstromturbulenzen, die als akustisches Ereignis für die Erzeugung des Tones der Stimme mit verantwortlich sind. Es handelt sich dabei um ein relativ schwaches Rauschen, das einen breiten Frequenzbereich überstreicht. Die durch Muskelkontraktion unterschiedliche Ausformung der Räume des Ansatzrohres besitzen nun entsprechend ihrer jeweiligen Konfiguration bestimmte Eigenfrequenzen. Diese 3 Anatomische und physiologische Grundlagen 7 entstehen also, wenn die Luft in diesen Räumen zu Schwingungen anregt wird. Das an Engstellen entstehende Rauschen bzw. der obertonreiche Klang der an den Stimmbändern erzeugten Stimme enthalten auch diese Frequenzen. Durch sie wird das Ansatzrohr zur Resonanz (Gerthsen et al. 1980) angeregt, was die Amplituden dieser Frequenzen also erheblich vergrößert, sie werden schließlich überschwellig und deutlich hörbar. Das heißt, dass die Veränderung der Konfiguration des Ansatzrohres durch die unterschiedliche Ausformung der Resonanzräume auch für die jeweils verschiedene Eigenfrequenzen verantwortlich ist. Bei jeder Artikulationsstellung, d.h. jeder bestimmten Stellung von Kiefer, Zunge und Gaumensegel werden, sobald die gebildeten Hohlräume in Resonanz geraten, ganz charakteristische Frequenzen bzw. Frequenzbänder hörbar (Flanagan 1972, Gauer et al. 1972, Lenneberg 1970). Diese für die jeweilige Artikulationsstellung charakteristischen Frequenzbänder werden als )RUPDQWHQ bezeichnet. Sie hängen also praktisch nur von der Konfiguration des Ansatzrohres ab und nicht von der im Kehlkopf gebildeten Stimme. Beim normalen Sprechen werden die 9RNDOHdurch eine stimmbildende Anregung des Ansatzrohres erzeugt. Hierbei hat das Ansatzrohr eine relativ stabile Konfiguration, denn die Schallabstrahlung erfolgt direkt vom Mund (Flanagan 1972). Die dabei entstehenden Formanten sind dafür verantwortlich, dass z.B. der Vokal /a/ unabhängig von der Tonhöhe und vom Sprecher als /a/ erkannt wird oder dass trotz festgehaltener Tonhöhe die Vokale /a, e, i, o, u/ sowie die Diphthonge bzw. Umlaute /ei, ä, ö, ü, eu, au/ voneinander unterschieden werden können. Der Formant bzw. die Formanten (in den Fällen, in denen es mehrere gibt) sind also das akustische Äquivalent eines bestimmten Vokals oder mancher Konsonanten. .RQVRQDQWHQ werden dagegen in einer weniger stabilen Konfiguration des Ansatzrohres gebildet. Sie sind nicht notwendigerweise stimmhaft und werden nicht ausschließlich vom Mund abgestrahlt. Bei der Bildung von Konsonanten ist das Ansatzrohr stärker verengt als bei den Vokalen, wobei insbesondere den 3 Anatomische und physiologische Grundlagen 8 Lippen und der Zahnreihe eine große Bedeutung zukommt (Flanagan 1972, Lenneberg 1973). Die Konsonanten bilden keine einheitliche Klasse von Lauten. Es sind so genannte Reibelaute, die an einer Konstriktion entstehen, bzw. Explosionslaute, die durch das plötzliche Freigeben eines Verschlusses gebildet werden und Nasale, bei denen das Gaumensegel den Nasenraum freigibt, wodurch sich die Resonanzverhältnisse erheblich verändern. Die Konsonanten können an den drei Artikulationszonen, den Lippen (labial), den Zähnen (dental), am Gaumen (palatal) oder in der Kehle (guttural) entstehen. Schwingen gleichzeitig mit dem Erklingen eines Konsonanten auch die Stimmbänder, handelt es sich um einen stimmhaften Konsonanten. In den übrigen Fällen wird von stimmlosen Konsonanten gesprochen. Konsonanten sind akustisch durch bestimmte Frequenzanteile oder Zeitmuster charakterisiert. Bei den Reibe- und Explosionslauten wird ein breites Frequenzspektrum erzeugt, das hohe Frequenzanteile enthält, z.B. /t/, /s/, wobei die beiden Genannten sich in ihren Frequenzanteilen praktisch nicht unterscheiden, aber aufgrund ihres verschiedenen zeitlichen Verhaltens ohne weiteres unterschieden werden können (Schmidt et al. 2000). .RUWLNDOH6SUDFK]HQWUHQ %URFD6SUDFKUHJLRQ Als erster beobachtete Paul BROCA im Jahre 1861, dass Läsionen des unteren Abschnitts der dritten Stirnwindung links (Broca-Sprachregion, Brodmann Area 44 und 45, s. Abb. 1) zu einer Sprachstörung (Aphasie) führen, bei dem das Sprachverständnis noch intakt ist, die Kranken aber spontan fast nichts sprechen. 3 Anatomische und physiologische Grundlagen 9 Nach Aufforderung bringen sie zögernd mit großer Anstrengung kurze Sätze hervor, die auf die nötigsten Substantive, Abb. 1 An der Sprachkonstruktion beteiligte Hirnregionen (nach Kolb u. Whishaw 1990). Verben und Adjektive reduziert sind (Telegrammstil). Die Artikulationen erfolgen zumeist sehr mühevoll und ohne Prosodie, und die Worte sind phonetisch entstellt. Sowohl in einfachen als auch in komplexen Sätzen fehlen häufig die grammatikalischen Funktionswörter. Das Verstehen vieler Satztypen (z.B. Passivsätze) ist daher oft nicht möglich. Weiterhin treten Probleme beim Nachsprechen von Sätzen auf. Diese Form der Aphasie wird als PRWRULVFKH $SKDVLH bezeichnet und das zugehörige Hirnareal Broca-Sprachregion genannt. Das Areal liegt unmittelbar vor denjenigen Abschnitten des motorischen Kortex, die die Muskeln des Gesichts, des Kiefers, der Zunge, des Gaumens und des Rachens kontrollieren, also derjenigen Muskeln, die zur Artikulation notwendig sind. Die motorische Aphasie durch Schädigung der Broca-Region ist aber nicht auf eine Lähmung dieser Muskeln zurückzuführen. Selbst eine direkte Schädigung der Gesichtsregion des Gyrus praecentralis führt nur zu geringen kontralateralen Ausfallserscheinungen, da die Gesichtsmuskulatur beidseitig im Kortex repräsentiert ist und ein unilateraler Ausfall durch die Gegenseite kompensiert werden kann. 3 Anatomische und physiologische Grundlagen 10 :HUQLFNH6SUDFKUHJLRQ Kurz nach der Entdeckung Brocas beschrieb Carl WERNICKE 1874 einen anderen Typ der Aphasie, bei der das Sprachverständnis extrem gestört, das spontane Sprechen der Patienten aber flüssig, wenn auch wenig verständlich war. Viele Wörter sind phonematisch entstellt, sodass noch verständliche phonematische Paraphasien (z.B. „ Rille“ statt „ Rinne“ ) oder ganz unverständliche Neologismen auftreten. Oft werden Wörter durch bedeutungsverwandte ersetzt (semantische Paraphasien). Auch das Sprachverständnis ist stark reduziert, d.h. dass das Verständnis einzelner Wörter häufig nicht gelingt. Nicht zuletzt ist auch das Nachsprechen von Wörtern und Sätzen stark beeinträchtigt. Diese VHQVRULVFKH $SKDVLH korreliert auffallend häufig mit Schädigungen im Schläfenlappen, insbesondere im rückwärtigen Abschnitt der ersten Schläfenwindung (Wernicke-Region, Brodmann Area 22), also in unmittelbarer Nachbarschaft der Hörrinde. Broca- und Wernicke-Region sind in der Nähe der primären Kortizes lokalisiert, die bei der frühen Sprachentwicklung des Kindes aktiviert werden. Artikuliert das Kind einen Laut oder ein Wort, so tritt eine neuronale Aktivität sowohl im motorischen System, wo die artikulatorischen Befehle generiert werden auf, als auch im akustischen System, das durch die selbstproduzierten Lautäußerungen stimuliert wird. Konsequenterweise sind die Sprachzentren zu den sprachrelevanten primären Kortizes auch benachbart, was bei einem Ausfall zu den gezeigten Symptomen führt. =XVDPPHQDUEHLWNRUWLNDOHU6SUDFK]HQWUHQ Die Auffassung, dass die Sprachproduktion durch Strukturen des Frontalhirns (Broca) und das Sprachverständnis durch Bereiche im Temporallappen 3 Anatomische und physiologische Grundlagen 11 (Wernicke) gesteuert wird, stellt eine stark vereinfachte Sichtweise der beiden Funktionen dar. Dies postuliert das von WERNICKE (Wernicke 1874) und LICHTHEIM (Lichtheim 1885) Ende des 19. Jahrhunderts vorgeschlagene Sprachmodell, das von GESCHWIND später weiterentwickelt wurde. GESCHWIND räumt zwar der Wernicke-Region eine Funktion bei der Sprachproduktion ein, ordnet jedoch das Sprachverständnis allein den posterioren Kortexgebieten zu. Diese Sichtweise stellt zweifellos eine nicht angemessene Simplifikation dar. Denn Läsionen einer der beiden Regionen verursachen in der Mehrzahl der Fälle multimodale Störungen. Das „ motorische Sprachzentrum“ ist also keineswegs ausschließlich nur für motorische Sprachfunktionen verantwortlich, sondern auch für die Perzeption von Sprache, ebenso wie das „ sensorische Sprachzentrum“ für die Sprachproduktion notwendig ist. (Luria 1970). Neuere Positronen-Emissions-Tomographien zeigen sogar, dass bei der Perzeption von Silben und Wörtern im intakten Gehirn in der Regel Broca- und WERNICKE-Region gemeinsam aktiviert werden. Dies legt die Schlussfolgerung nahe, dass die Sprachareale sowohl bei der Sprachproduktion als auch beim Sprachverständnis zusammenarbeiten, dass also sprachverarbeitende neuronale Einheiten über den perisylvischen Kortex verteilt sind. Die Sichtweise, dass die linke Hemisphäre beim Rechtshänder sprachdominant sei, ist vor allem darauf zurückzuführen, dass bei Rechtshändern Schädigungen der linken Hemisphäre zumeist zu Aphasien fuhren. Exakter erscheint es jedoch, von einer sich gegenseitig ergänzenden Spezialisation der beiden Hemisphären zu sprechen. Denn es gibt vielerlei Hinweise darauf, dass im intakten Gehirn auch rechtshemisphärische Prozesse an der Sprachverarbeitung beteiligt sind. So sind z.B. die durch Wörter evozierten Gehirnpotenziale im EEG meist über beiden Hemisphären sichtbar, wenn manche Komponenten auch über einer Hemisphäre stärker ausgeprägt sind (Boller u. Grafmann 1988-1992). Sprachverständnis und das Erkennen von Worten (vor allem von Inhaltswörtern), die Bildung von Satzmelodie und Betonung (Prosodie) sowie die Einordnung von Sprechakten (z.B. als Frage oder als Vorwurf) sind Leistungen, zu denen die rechte Hemisphäre nicht nur beiträgt, sondern zu denen sie sogar selbständig in der Lage 3 Anatomische und physiologische Grundlagen 12 ist. Dennoch tritt beim rechtshändigen Erwachsenen nach Schädigung im perisylvischen Bereich der linken Hemisphäre in der Regel eine Aphasie auf (Heilman u. Valenstein 1993, Lichtheim 1885, Schmidt 1993). Syntaktische Regeln und Funktionswörter sind primär links in der perisylvischen Region lokalisierbar (sprachdominante Hemisphäre). Sprachverständnis, vor allem von Inhaltswörtern, findet sich aber auch rechts (Schmidt et al. 2000). $SKDVLHIRUPHQ Aphasien sind hirnorganische Sprachstörungen von Menschen, die bereits eine Sprache beherrschen. Aphasien treten in unterschiedlichen Ausprägungen auf und können verschiedene Komponenten des Sprachsystems (z.B. Phonologie, Syntax, Lexikon, Semantik) betreffen. Sprachabhängige Leistungen wie Lesen, Schreiben und Rechnen können dabei ebenfalls beeinträchtigt sein. Zusätzlich sind eine Apraxie, Agnosie oder Dysarthrie möglich. Zahlreiche Klassifikationen der Aphasien sind bekannt; die Nomenklatur ist dementsprechend chaotisch. Eine allgemeine Einteilung nach Symptomatik unterscheidet sensorische (rezeptive) und motorische (expressive) Aphasien. Spezieller unterteilt man Aphasien in Worttaubheit (Unfähigkeit, gesprochene Worte zu verstehen), Wortblindheit (Unfähigkeit, geschriebene Worte zu verstehen) und Agraphie (Unfähigkeit, Gedanken in geschriebener Form auszudrücken) und in die im engeren Sinn als motorische Aphasie bezeichnete Störung (Unfähigkeit, Gedanken sprechend auszudrücken) (Ganong 1974). Aphasien treten zum einen bei subkortikalen Läsionen in der weißen Substanz, in den Basalganglien oder im Thalamus auf. Die kortikalen aphasieverursachenden Areale betreffen primär die Areale in der Nähe der sylvischen Furche, wo Brocaund Wernicke-Region abgrenzbar sind. Obwohl bei den verschiedenen Aphasieformen unterscheidbare Läsionsorte vorliegen können, ist jedoch bei der Mehrzahl der Fälle eine genaue Zuordnung der einzelnen Sprachfunktionen zu bestimmten Kortexarealen nicht möglich. Dennoch gilt: Schädigungen der BrocaRegion (Areae 44, 45) führen meist zu (motorischen) 3 Anatomische und physiologische Grundlagen 13 Sprachproduktionsproblemen (Broca-Aphasie), Schädigungen der WernickeRegion (Area 22) und angrenzender Gebiete zu (sensorischen) Ausfällen im Sprachverständnis (Wernicke-Aphasie). In der Nachbarschaft der Wernicke-Region wurden weitere Bereiche gefunden, deren Läsion regelmäßig zu Aphasien führt: der Gyrus angularis (Brodmann Area 39), der Gyrus supramarginalis (Brodmann Area 40) sowie die mittlere Temporalwindung (Brodmann Area 37). Eine Schädigung des Gyrus angularis führt zu leichten Sprachstörungen, bei denen semantische Paraphasien auffallen und Benennstörungen im Vordergrund stehen (DPQHVWLVFKH $SKDVLH). Probleme treten vor allem mit bedeutungstragenden Inhaltswörtern auf. Das Sprachverständnisdefizit ist hierbei besonders schwach ausgeprägt. Eine Läsion des Fasciculus arcuatus, der Broca- und Wernicke-Region verbindet, einschließlich einer Läsion im oberen Temporallappen und/oder der Insula (Heilman u. Valenstein 1993, Kolb u. Whishaw 1990) hat eine /HLWXQJVDSKDVLH zur Folge. Die Fähigkeit zum Nachsprechen ist hier stark beeinträchtigt, wogegen andere sprachliche Symptome im Hintergrund stehen. Auch größere Läsionen innerhalb des perisylvischen Bereichs können zu schweren Formen der WUDQVNRUWLNDOHQ $SKDVLH führen. Das Nachsprechen gelingt zwar relativ gut, wohingegen Defizite in der Sprachproduktion (transkortikale motorische Aphasie), im Sprachverständnis (transkortikale sensorische Aphasie) oder in beiden Leistungen (gemischte transkortikale Aphasie) hervortreten. Eine Schädigung der gesamten perisylvischen Region dagegen führt zur JOREDOHQ $SKDVLH, d.h. zu einer schweren Sprachproduktionsstörung, bei der oft nur noch stereotype Silben- oder Wortfolgen geäußert werden können. Ebenso stark ausgeprägt ist das Defizit im Sprachverständnis und im Nachsprechen (Schmidt et al. 2000). 3 Anatomische und physiologische Grundlagen 14 3K\VLRORJLHGHVDXGLWLYHQ6\VWHPV KLINKE stellt zurecht fest, dass das Ohr das empfindlichste Sinnesorgan des Menschen ist (Klinke 1987). Das menschliche Gehör erlaubt es, hochkomplexe, detaillierte Informationen aus der Umwelt zu extrahieren. In erheblich höherem Grad als jeder andere Sinn ist das Gehör dabei für die menschliche Sprache und deren Entwicklung verantwortlich (Schmidt et al. 2000). 6FKDOO Schall ist der adäquate Reiz für das Ohr. Im alltäglichen Leben tritt Schall als Druckschwankungen in der Luft auf. Ein UHLQHU 7RQ ist dabei eine Sinusschwingung, die nur aus einer einzigen Frequenz besteht, was im Alltag nur sehr selten vorkommt. Subjektiv besteht ein Zusammenhang zwischen der Frequenz und der empfundenen Tonhöhe. Je höher die Schallfrequenz, desto höher wird der Ton empfunden. Außer durch seinen Frequenzgehalt wird ein Schall durch die Amplitude der entstehenden Druckschwankungen, d.h. den Schalldruck, charakterisiert. Der Schalldruckpegel (L) wird in Dezibel (dB) angegeben und ergibt einfach anzuwendende Zahlenwerte zwischen 0 und ungefähr 120 dB: Dabei gilt: L = 20 log PxP0 [dB] (Px: zu beschreibender Schalldruck, P0: Bezugsschalldruck [2x10-5 Pa]) 3 Anatomische und physiologische Grundlagen 15 Je höher der Schalldruck also ist, desto lauter erscheint der Schall. Eines der wichtigsten klinischen Untersuchungsverfahren des Gehörs ist die Tonaudiometrie, bei der dem Untersuchten reine Töne für jedes Ohr getrennt über einen Kopfhörer angeboten werden. Interessanterweise ist die subjektive Lautstärke dabei frequenzabhängig: Bei gleichem physikalischen Schalldruck werden Töne zwischen 2000 und 5000 Hz lauter empfunden als höher- oder niederfrequentere Schallsignale. Besteht die Absicht, dass der Untersuchte alle Töne isophon hört, so muss der Schalldruck frequenzabhängig ständig verändert werden. Kurven gleicher Lautstärkepegel (Isophone) werden in Phon angegeben und decken sich definitionsgemäß bei 1000 Hz mit der dB-Skala des Schalldruckpegels. Der menschliche Hörbereich erstreckt sich über Frequenzen von 20 bis 16.000 Hz und Lautstärkepegel zwischen 4 und 130 Phon. Der Hauptsprachbereich, der die Frequenzen und Lautstärken der menschlichen Sprache umfasst, liegt hierbei etwa im mittleren Bereich. Die Hörschwelle, also die Grenze, ab der ein Ton als hörbar empfunden wird, ist gleichfalls frequenzabhängig und zwischen 2000 und 5000 Hz am niedrigsten. Sie stellt eine Isophone dar (4 Phon). Die Schwellenaudiometrie misst dabei die Hörschwelle des Ohrs und bestimmt die Schalldruckpegel der niedrigsten Isophone. Die durchschnittliche Hörschwelle bei gesunden Jugendlichen wurde für alle Frequenzen bestimmt und als 0 dB V (Hörverlust) definiert. Die klinische Hörschwellenkurve wird dabei grafisch als Gerade dargestellt, sodass für den medizinischen Alltag ein übersichtliches Bild entsteht (Tonaudiogramm). 3 Anatomische und physiologische Grundlagen 16 6FKDOOOHLWXQJ]XP,QQHQRKU Das Ohr des Menschen besteht aus dem äußeren Ohr, dem Mittel- und dem Innenohr (Abb. 2). Der Schall gelangt durch die Luft des äußeren Gehörgangs bis zum Trommelfell in Form der so genannten Luftleitung. Anschließend wird er durch Schwingungen von Trommelfell und Gehörknöchelchenkette (Malleus, Incus und Stapes) bis zum ovalen Fenster des Innenohrs fortgeleitet. Gleichzeitig wird der niedrige Schallwellenwiderstand (Schallimpedanz) der Luft an die hohe Impedanz des flüssigkeitsgefüllten Innenohrs angepasst. Ohne Mittelohr würden nämlich 98% des Schalls vom Ohr reflektiert und nicht aufgenommen. So ist eine Impedanzanpassung erforderlich, die durch das funktionale Zusammenspiel von Trommelfell und Gehörknöchelchen vollzogen wird. Die Reflexion wird dadurch so stark reduziert, dass 60% der Schallenergie in das Innenohr eintreten kann. Aber auch ohne Mittelohr kann das Innenohr angeregt werden. Die Knochenleitung über die Schädelkalotte wird klinisch für Stimmgabeluntersuchungen (z.B. nach Rinne oder Weber) und für Hörprüfungen genutzt. 3 Anatomische und physiologische Grundlagen 17 Abb. 2 Schematische Darstellung von äußerem, mittlerem und innerem Ohr. Punktiert eingezeichnet ist ein Extremwert eines Schwingungszustandes des TrommelfellGehörknöchelchen-Apparates (nach Schmidt u. Thews 1977). H Hammer (Malleus) A Amboss (Incus) S Steigbügel (Stapes) 6FKDOOWUDQVGXNWLRQLP,QQHQRKU Das Innenohr liegt im Knochen der Felsenbeinpyramide und besteht aus mehreren miteinander verbundenen Knochenkanälen, dem knöchernen Labyrinth. Dieses besteht aus zahlreichen mit Perilymphe gefüllten Gängen und umgibt ein geschlossenes System von Röhren, das häutige Labyrinth. Dieses wiederum setzt sich aus den Anteilen des Gleichgewichtsorgans und dem Hörorgan, d.h. der Kochlea einschließlich des Corti-Organs zusammen. Die Kochlea – Schnecke – ist ein schlauchförmiges Organ, das in Form eines Schneckenhauses in zweieinhalb Windungen quasi eingerollt ist. Drei übereinanderliegende Skalen, von denen zwei, die Scala vestibuli und die Scala 3 Anatomische und physiologische Grundlagen 18 tympani, am so genannten Helikotrema zusammenhängen, bilden die Struktur (Abb. 3). Gegen das Mittelohr sind sie durch die Steigbügelfußplatte am ovalen Fenster bzw. durch die Membran des runden Fensters abgegrenzt. Scala vestibuli und Scala tympani sind mit der aus dem Liquor stammenden Perilymphe gefüllt, einer Flüssigkeit, die sich ähnlich wie andere extrazelluläre Flüssigkeiten zusammensetzt, also viel Na+-Ionen enthält. Zwischen diesen beiden Skalen liegt die Scala media. Diese wird durch die Reissner-Membran und das Corti-Organ von Scala vestibuli bzw. Scala tympani abgegrenzt. Das Corti-Organ befindet sich auf der Basilarmembran und enthält die mit Stereozilien besetzten Hörsinneszellen (Haarzellen). Über den Reihen der Haarzellen liegt die Membrana tectoria. Abb. 3 Schnitt durch eine Windung der Kochlea (nach Schmidt u. Thews 1977). In der Scala media befindet sich die Endolymphe, eine auffällig K+-reiche Flüssigkeit, deren Zusammensetzung den intrazellulären Flüssigkeiten ähnelt. 3 Anatomische und physiologische Grundlagen 19 Ist das Ohr nun einem Schall ausgesetzt, schwingt der Stapes mit der ovalen Fenstermembran. Im Innenohr löst das Schallsignal Auf- und Abwärtsbewegungen der kochlearen Membranen aus. Das Schallsignal bildet eine Wanderwelle in der zentralen Funktionseinheit der Kochlea, der Basilarmembran. Das Amplitudenmaximum der Wanderwelle entsteht in Abhängigkeit von der jeweiligen Reizfrequenz an einem bestimmten Ort entlang der Basilarmembran. Die wanderwelleninduzierte Auslenkung der Basilarmembran – einschließlich der sich auf ihr befindenden Haarzellen – führt nun zu einer Deflexion der Stereozilien: Die Stereozilien der äußeren Haarzellen werden durch die Tektorialmembran deflektiert, die Stereozilien der inneren Haarzellen werden durch Bewegung der Endolymphströmung abgeschert. Dadurch wird ein Prozess initiiert, der das mechanische Schallsignal in elektrische und chemische Signale umwandelt (transduziert). Als dessen Folge geben innere Haarzellen einen afferenten Transmitter an die afferenten Fasern des Hörnervs ab, wobei die äußeren Wanderwellenmaximums Haarzellen und die für die Stimulation aktive der Verstärkung inneren des Haarzellen verantwortlich sind. 6LJQDOWUDQVIRUPDWLRQYRQGHU6LQQHV]HOOH]XP+|UQHUY Innere Haarzellen erregen den Hörnerv durch einen Neurotransmitter, wahrscheinlich das Glutamat. Dieses diffundiert durch den synaptischen Spalt und verbindet sich mit AMPA-Rezeptoren (AMPA: 2-amino-3-[3-hydroxy-5methylisoxazol-4-yl]propionic acid) der Nervenzellmembran. Dadurch wird ein postsynaptisches Potenzial ausgelöst, das zu Nervenaktionspotenzialen führt (Klinke 1986, Klinke 1987, Klinke 1990, Spoendlin 1972). Die elektrischen Phänomene im Innenohr führen zu klinisch messbaren Reizfolgepotenzialen: dem Mikrophonpotenzial und dem Summenaktionspotenzial. Nimmt das Ohr nun Schall, beispielsweise Sprachlaute, 3 Anatomische und physiologische Grundlagen 20 auf und gibt die registrierten Potenziale nach weiterer Verstärkung auf einen Lautsprecher, so wird das gesprochene Wort aus dem Lautsprecher „ gehört“ . Die Mikrophonpotenziale entstehen dabei an den äußeren Haarzellen. Wird das Ohr mit einem kurzen Schallimpuls gereizt, dann kann mit einer Elektrode am Promontorium (Vorwölbung an der medialen Wand der Paukenhöhle) oder am runden Fenster ein Summenaktionspotenzial des Nervus acusticus abgeleitet werden, das durch eine synchrone Erregung vieler afferenter Nervenfasern des Hörnerven entsteht. )UHTXHQ]VHOHNWLYLWlW*UXQGODJHGHV6SUDFKYHUVWlQGQLVVHV Es ist experimentell gut nachweisbar, dass das Ohr sehr gut Tonhöhen von hintereinander angebotenen Tönen unterscheiden kann: Bei 1000 Hz werden Änderungen von 0,3%, d.h. 3 Hz, wahrgenommen (Frequenzunterschiedsschwelle). Für die Ausbildung dieser )UHTXHQ]VHOHNWLYLWlW besitzt die Kochlea einen zweistufigen Mechanismus (von Békésy 1960). Erklingt ein Ton, werden Reissner-Membran, Tektorialmembran, Corti-Organ und Basilarmembran in die bereits geschilderten ständigen Auf- und Abwärtsbewegungen, d.h. in Vibrationen, versetzt. Diese Vibrationen bleiben nicht auf den Bereich in unmittelbarer Nähe von Stapes und rundem Fenster beschränkt, sondern bilden entlang der Basilarmembran eine – nicht gleichmäßige – Wanderwelle aus, die von der Basis bis zur Spitze der Kochlea wandert. Die Amplitude der Welle nimmt in einem ersten Schritt etwas zu, wird in einem zweiten Schritt bis zu tausendfach zu einer hohen Welle mit sehr scharfer Spitze und nimmt im weiteren Verlauf plötzlich wieder ab. Diese Verstärkung ist bei niedrigen und mittleren Schalldrücken besonders auffällig. Die scharfe Spitze der Wanderwelle stimuliert innere Haarzellen, die nach dem o.g. Transduktionsprozess einen Transmitter an die afferenten Hörnervenfasern weitergeben. 3 Anatomische und physiologische Grundlagen 21 Die Wanderwellenspitze entsteht nun für jede Tonfrequenz an einem anderen Ort entlang der Basilarmembran. Hohe Frequenzen erzeugen das Maximum der Wanderwelle in der Nähe der Kochleabasis, mittlere Frequenzen in der Kochleamitte, tiefe Frequenzen an der Kochleaspitze. Für jede Tonhöhe gibt es dadurch einen bestimmten Ort der Maximalauslenkung der Wanderwelle entlang der Basilarmembran (Tonotopie der Wanderwelle). Eine einzelne Frequenz wird also nur innere Haarzellen an einem bestimmten Ort reizen, unterschiedliche Frequenzen grundsätzlich Haarzellen an unterschiedlichen Orten entlang der Basilarmembran. Ein aus mehreren Tonhöhen bestehendes Schallereignis wird dadurch längs der Basilarmembran verteilt (Frequenzdispersion) (Khanna u. Leonard 1982, Klinke 1987, Sellick et al. 1982, Tonndorf 1980). Bei niedrigem Schalldruck führen die äußeren Haarzellen zu zusätzlichen mikromechanischen Schwingungen der Reizfrequenz. Äußere Haarzellen können sich bis zu 20.000mal pro Sekunde (20 kHz) verkürzen und verlängern. Dadurch wirken sie wie Servomotoren, die nach dem ersten Schritt der Wanderwelle diese im zweiten Schritt bis zu tausendfach verstärken. Die zusätzliche Schwingungsenergie entsteht nur an dem jeweils frequenzcharakteristischen, eng umschriebenen Ort der Basilarmembran. Nur dort werden jeweils einige wenige (wahrscheinlich ca. 50) äußere Haarzellen durch die Tektorialmembran gereizt, die zusätzlich erzeugte Schwingungsenergie wird scharf lokalisiert an die inneren Haarzellen abgegeben: Die Wanderwelle wird also nur in dem sehr eng umschriebenen Bereich verstärkt (Zenner 1986, Zenner et al. 1985, Zenner et al. 1987). ,QIRUPDWLRQVEHUWUDJXQJXQG9HUDUEHLWXQJLP=16 Die von der Haarzelle als Folge des Transduktionsprozesses ausgelöste Transmitterfreisetzung (s. Kapitel 3.3.4) wird in Form einer neuronalen Erregung über Hörnerv, Hirnstamm und Hörbahn bis zum auditorischen Kortex im Temporallappen weitergeleitet (Abb. 4). Hieran sind mindestens 5-6 3 Anatomische und physiologische Grundlagen 22 hintereinander geschaltete, durch Synapsen verbundene Neuronen beteiligt. Sie verfügen über Kollaterale und Interneuronen, sodass das auditorische System ausgedehnt neuronal vernetzt ist. Abb. 4 Schematische Darstellung der zentralen Hörbahn (nach Schmidt, Thews u. Lang 2000). Der Nervus cochlearis endet aufgeteilt im Nucleus cochlearis (Beginn des zweiten Neurons). Vom ventralen Teil des Nucleus cochlearis zieht der überwiegende Teil der Fasern zu den Kernen des Corpus trapezoideum der Gegenseite. Der ipsilaterale Anteil führt unter mehreren Umschaltungen wie der dorsale Teil zur primären Hörrinde. Vom Nucleus cochlearis dorsalis verlaufen die Neuriten zur Gegenseite dicht unter den Striae medullares der Rautengrube und enden in den Kernen des Trapezkörpers. Über das nächste Neuron in der lateralen Schleife (Lemniscus lateralis) entsteht die Verbindung zum Colliculus caudalis der Vierhügelplatte. Von hier wird das dritte Neuron über das Brachium colliculi caudalis zum Corpus geniculatum mediale gelenkt. Dort erreicht das letzte Neuron über die Hörstrahlung (Radiatio acustica) die primären akustischen Rindenfelder des Schläfenlappens (Areae 41 und 42). Durch die Umschaltungen in den medialen Kniehöckern entstehen Kollateralen für Reflexe auf akustische Reize. Neuronen der Formatio reticularis sind dabei mit den afferenten Abschnitten parallel geschaltet. 3 Anatomische und physiologische Grundlagen 23 Die durch einen Schallreiz im Verlauf dieser Neuronen hintereinander HYR]LHUWHQ $NWLRQVSRWHQ]LDOH werden oft zur Diagnostik verwendet (evoked response audiometry, ERA). Hierbei werden dem Untersuchten – sehr geringe – Schallreize angeboten, die im Elektroenzephalogramm (EEG) zu einer Veränderung der Hirnaktivität führen. Da die Abweichungen aber sehr klein sind, wird die einzelne Reizantwort im EEG vom Rauschen völlig überdeckt. Mit Hilfe des Computers wird durch rechnerische Mittelung zahlreicher evozierter Einzelpotenziale (z.B. von 2000 Potenzialen) die spezifische akustische Reizantwort von Hörnerv und Hörbahn aus der unspezifischen Hirnaktivität im EEG erkennbar. Der Nervus cochlearis verlässt das Ohr durch den inneren Gehörgang zum Kleinhirnbrückenwinkel. Wie oben beschrieben, teilen sich seine afferenten Fasern und ziehen im Hirnstamm zum Nucleus cochlearis ventralis bzw. zum Nucleus cochlearis dorsalis, um dort zum zweiten Neuron umgeschaltet zu werden. Der Hörnerv besteht aus einer großen Zahl afferenter sowie teilweise auch efferenter (d.h. aus dem Gehirn kommender) Nervenfasern. 90% der afferenten Nervenfasern haben nur eine Synapse mit einer einzigen, nämlich einer inneren Haarzelle. Das bedeutet, dass an das Gehirn im Wesentlichen Informationen von den inneren Haarzellen weitergeleitet werden. Nach dem Ortsprinzip ist jeder Haarzelle eine ganz bestimmte Tonfrequenz zugeordnet, und die mit einer bestimmten Haarzelle synaptisch verbundene Hörnervenfaser wird bei Beschallung des Ohrs mit dieser Frequenz optimal erregt. Daher wird diese Frequenz auch FKDUDNWHULVWLVFKH )UHTXHQ] (Bestfrequenz) der Einzelfaser genannt. Die Zeitdauer eines Schallreizes wird durch die Zeitdauer der Aktivierung der Nervenfasern verschlüsselt, die Höhe des Schalldruckpegels durch die Entladungsrate. Hierbei kann eine einzelne Nervenfaser eine bestimmte Entladungsrate nicht überschreiten, sondern erreicht ab einem bestimmten Schalldruck einen Sättigungsbereich. Trotzdem kann die Information nach höherer Lautstärke weitergegeben werden, da dann eine zunehmende Zahl benachbarter Fasern rekrutiert wird. 3 Anatomische und physiologische Grundlagen 24 Der Nervus cochlearis zergliedert sich in den Nuclei cochlearis dorsalis und ventralis, wo die zweiten Neuronen beginnen. Wie oben beschrieben, zieht ein Teil der vom ventralen Nucleus cochlearis ausgehenden Neuronen zur oberen Olive der gleichen Seite, ein Teil kreuzt zur oberen Olive der anderen Seite. Auch die afferenten Nervenfasern vom dorsalen Kern kreuzen zum Nucleus lemnisci lateralis der Gegenseite. Das bedeutet, dass im zweiten Neuron ein Teil der Fasern ipsilateral verläuft, ein wesentlicher Teil der zentralen Hörbahn jedoch auf die kontralaterale Seite kreuzt. Daher ist jedes Innenohr mit der rechten und der linken Hörrinde verbunden. Außerdem können in den Nervenzellen des Olivenkomplexes erstmals im Verlauf der Hörbahn binaurale akustische Signale miteinander verglichen werden. Die höheren Neuronen verlaufen von der oberen Olive zum Teil auf der gleichen Seite, zum Teil auf der Gegenseite nach jeweils neuer Umschaltung zum Colliculus inferior und anschließend zum Corpus geniculatum mediale. Schließlich ziehen die Afferenzen als Radiatio acustica zur primären Hörrinde (Heschl-Querwindung) des Temporallappens. Die höheren Neuronen sind stark spezialisiert, was bedeutet, dass sie nur auf jeweils spezifische Schallmuster reagieren: Die einfache Kodierung des ersten und von Teilen des zweiten Neurons wandelt sich grundlegend ab dem dorsalen Nucleus cochlearis und weiter zunehmend mit jedem höheren Neuron. Zwar wird das Ortsprinzip bis zum auditorischen Kortex beibehalten, sodass bestimmte Schallfrequenzen an bestimmten Orten der Hörrinde oder der auditorischen Kerne repräsentiert sind. Zusätzlich besitzen jedoch einige vom dorsalen Nucleus cochlearis ausgehende Neuronen kollaterale Verschaltungen, die teils exzitatorisch, teils inhibitorisch wirksam sind (On-Off-Neuronen). In Folge werden einzelne Neuronen des dorsalen Kochleariskerns bei Schallreiz stets gehemmt. Höhere Neuronen der Hörbahn reagieren nicht auf reine Sinustöne, sondern auf bestimmte Eigenschaften eines Schallmusters (z.B. Sprachmuster). Es gibt dabei Fasern, die bei einer bestimmten Schallfrequenz aktiviert, durch höhere oder 3 Anatomische und physiologische Grundlagen 25 tiefere Töne jedoch gehemmt werden. Einige Neuronen reagieren auf eine Frequenzmodulation, wobei zusätzlich der Grad der Modulation von Bedeutung sein kann. Andere Nervenzellen sprechen hingegen nur auf eine Amplitudenänderung eines Tons an. Diese Spezialisierung von Neuronen auf bestimmte Eigenschaften eines Schallmusters ist im auditorischen Kortex noch ausgeprägter. Neuronen können hochspezialisiert auf den Beginn oder das Ende, auf eine Mindestzeitdauer oder eine mehrfache Wiederholung und auf bestimmte Frequenz- oder Amplitudenmodulationen eines Schallreizes sein. Es liegt daher die Schlussfolgerung nahe, dass diese bis zur Hörrinde zunehmende Spezialisierung der Neuronen auf bestimmte Eigenschaften des Schallreizes es erlaubt, Muster innerhalb des Schallreizes herauszuarbeiten und für die kortikale Beurteilung vorzubereiten (Informationsverarbeitung). Nun besteht das gesprochene Wort ausschließlich aus derartigen Mustern, die trotz eines Störschalls (z.B. Umgebungsgeräusche) erkannt werden können (Pickles 1988, Zenner 1985, Evans 1974, Manley u. Müller-Preuss 1978). 3K\VLRORJLHGHVYLVXHOOHQ6\VWHPV Licht besteht aus elektromagnetischer Strahlung mit Wellen- und Teilcheneigenschaften. Weißes Sonnenlicht verfügt dabei über die gesamte Spektralpalette aller Farbfrequenzen, wobei nur der Bereich von 400 bis etwa 750 nm Wellenlänge für das menschliche Auge sichtbar ist und als Licht wahrgenommen wird. Der Leuchtdichtebereich, innerhalb dessen gesehen werden kann, beträgt etwa 1:1011. 6HKHQXQG%OLFNHQ Die visuelle Wahrnehmung ergibt sich aus der Wechselwirkung sensorischer und motorischer Leistungen von Auge und Zentralnervensystem: Durch Augen-, Kopf- und Körperbewegungen verschieben sich die Bilder der visuellen Umwelt 3 Anatomische und physiologische Grundlagen 26 alle 0,2-0,6 s auf der Netzhaut. Das Gehirn erzeugt aus den diskontinuierlichen und unterschiedlichen Netzhautbildern eine einheitliche und kontinuierliche Wahrnehmung der visuellen Objekte und des umgebenden extrapersonalen Raums. Trotz der retinalen Bildverschiebungen werden die Raumrichtungen richtig und die Gegenstände unbewegt wahrgenommen, weil die afferenten visuellen Signale mit der Efferenzkopie der motorischen Kommandos und mit vestibulären Signalen im Gehirn verrechnet werden. Eine besonders regelhafte Form der Augenbewegungen tritt beim /HVHQauf. Der Fixationspunkt verschiebt sich beim Lesen in raschen Sakkaden (in den Kulturkreisen mit rechtsläufiger Schrift von links nach rechts) über die Zeile. Zwischen den Sakkaden liegen Fixationsperioden von 0,2-0,6 s Dauer. Ist der Fixationspunkt beim Lesen am Zeilenende angelangt, so bewegen sich die Augen meist mit einer Sakkade wieder nach links zum nächsten Zeilenanfang. Die Amplitude und die Frequenz der Lesesakkaden sind von der formalen Struktur des Textes (Größe, Gliederung, Groß- und Kleinschreibung) abhängig. Sie werden jedoch auch vom Textverständnis bestimmt. Ist ein Text unklar geschrieben oder gedanklich schwierig, treten gehäuft Regressionssakkaden auf. Dies sind Sakkaden entgegengesetzt zur normalen Leserichtung. Zahlreiche Regressionssakkaden kennzeichnen auch die Augenbewegungen eines gerade das Lesen lernenden Kindes. Kinder mit Legasthenie zeigen ebenfalls häufig Regressionssakkaden (Stein 1991). 2SWLVFKHV6\VWHPGHV$XJHV Das optische System des Auges ist ein nicht exakt zentriertes, zusammengesetztes Linsensystem, das auf der Netzhaut ein umgekehrtes und verkleinertes Bild der Umwelt entwirft: Der dioptrische Apparat besteht aus der durchsichtigen Kornea, den mit Kammerwasser gefüllten vorderen und hinteren Augenkammern, der die Pupille bildenden Iris, der Linse, die von einer durchsichtigen Linsenkapsel 3 Anatomische und physiologische Grundlagen 27 umgeben ist, und dem Glaskörper, der den größten Raum des Augapfels ausfüllt (Abb. 5). Die hintere innere Oberfläche des Auges wird von der Retina ausgekleidet. Der Raum zwischen Retina und der den Bulbus oculi bildenden festen Sklera wird durch das Gefäßnetz der Choroidea ausgefüllt. Am hinteren Pol des Auges hat die menschliche Retina eine kleine Grube, die Fovea centralis. Sie ist für das Sehen bei Tageslicht die Stelle des schärfsten Sehens und normalerweise der Schnittpunkt der optischen Achse des Auges mit der Netzhaut. Abb. 5 Waagrechter schematischer Durchschnitt durch den linken Augapfel – von oben gesehen (nach Leydhecker 1976). Im menschlichen Auge werden verschieden weit entfernte Gegenstände auf der Netzhaut dadurch scharf abgebildet, dass sich der Krümmungsradius der vorderen Linsenfläche ändert (Akkommodation, Abb. 6). Abb. 6 Reduziertes Auge. 1 Winkelgrad entspricht etwa 0,29 mm auf der Netzhaut (nach Schmidt, Thews u. Lang 2000). 3 Anatomische und physiologische Grundlagen 28 6LJQDODXIQDKPHXQG6LJQDOYHUDUEHLWXQJLQGHU1HW]KDXW Im Wirbeltierauge liegt die Schicht der Rezeptoren von der Glaskörperseite abgewandt in engem Kontakt mit den Pigmentepithelzellen, die an das Gefäßsystem der Choroidea angrenzen und den Stoffwechsel der Rezeptorzellen beeinflussen (Abb. 7). Im menschlichen Auge besteht die Rezeptorschicht aus ca. 120 Millionen Stäbchen und sechs Millionen Zapfen. Das skotopische Sehen mit den Stäbchen ist an die Beleuchtungsbedingungen einer sternenklaren Nacht angepasst, das photopische Sehen mit den Zapfen dagegen an das Tageslicht. Es gibt eine Klasse von Stäbchen und drei Klassen von Zapfen mit unterschiedlichen spektralen Absorptionskurven. Abb. 7 Aufbau der Primatennetzhaut und Schema der Reaktion einzelner Neuronen auf einen Lichtreiz. Nervus opticus (nach Schmidt, Thews u. Lang 2000). 3 Anatomische und physiologische Grundlagen 29 Die Rezeptordichte ist für die Zapfen in der Mitte der Fovea, für die Stäbchen dagegen im parafovealen Bereich am höchsten. In der Fovea centralis gibt es keine Stäbchen. Zwischen den Rezeptoren und dem Glaskörper liegen die Schichten retinaler Nervenzellen – der Horizontalzellen, Bipolarzellen, Amakrinen und Ganglienzellen, deren Axone den N. opticus bilden. Der Transduktionsprozess des Sehens wird durch die Lichtquantenabsorption in den Sehfarbstoffen der Photorezeptoren eingeleitet. Jeder der drei Zapfentypen besitzt einen spezifischen Sehfarbstoff (Zapfenopsin). Der Sehfarbstoff der Stäbchen ist das Rhodopsin. Durch einen mehrstufigen Zerfall entsteht Metarhodopsin II, das in eine intrazelluläre Enzymkaskade eingreift und über ein G-Protein (Guanyl-Nukleotid bindendes Protein) der Photorezeptormembran die cGMP-Konzentration (cGMP: zyklisches Guanosin5'-phosphat) reduziert, was eine Abnahme des Natriumleitwertes und eine Hyperpolorisation des Membranpotenzials der Photorezeptoren bei Belichtung zur Folge hat. Die Amplitude dieses hyperpolarisierenden Rezeptorpotenzials nimmt nichtlinear mit dem Photoneneinfall pro Zeiteinheit zu. Die Signale der Rezeptoren werden durch synaptische Kontakte auf die Bipolarzellen und die Horizontalzellen übertragen und in diesen Zellen durch langsame Membranpotenzialänderungen verrechnet. Die Signale der Bipolarzellen werden direkt über die Amakrinen auf die Dendritenmembran der Ganglienzellen übertragen. Eine Ganglienzelle ist in der Regel mit mehreren Bipolarzellen verbunden. Das Ausmaß dieser Signalkonvergenz ist um so größer, je weiter außen in der Netzhautperipherie eine Ganglienzelle situiert ist. Da ein Photorezeptor mit mehreren Bipolarzellen verbunden ist, besteht im retinalen Neuronennetz auch eine erhebliche Signaldivergenz. Die Signalkonvergenz von Rezeptoren und Ganglienzellen steht im Verhältnis 125:1. Konvergenz und Divergenz der Verknüpfungen in der Retina bilden die Grundlage der rezeptiven Felder. Das rezeptive Feld (RF) eines visuellen Neurons 3 Anatomische und physiologische Grundlagen 30 ist jener Bereich der Netzhaut, dessen adäquate Stimulation zu einer Aktivitätsänderung des Neurons führt. Es sind verschiedene Klassen retinaler Ganglienzellen unterscheidbar, die jeweils antagonistisch organisierte RF haben: Führt die Belichtung des RF-Zentrums zu einer Erregung, bewirkt Belichtung der RF-Peripherie eine Hemmung und umgekehrt (On- bzw. Off-Neurone). Die On-Off-Ganglienzellen reagieren auf stationäre Lichtreize meist mit einer kurzen On-Aktivierung, auf Verdunkelung mit einer kurzen Off-Aktivierung. Die Ganglienzellen des On- und des OffSystems übertragen Signale über retinale Hell-Dunkel- oder Farbwerte, während die On-Off-Ganglienzellen meist zum System der bewegungsempfindlichen Neuronen gehören. Die Impulsrate der Neuronen des On-Systems verläuft linear mit der subjektiven Helligkeit, die des Off-Systems mit der subjektiven Dunkelheit eines Gesichtsfeldbereichs korrelierend. Mit Hilfe dieser linearen Korrelationsregeln lassen sich einige elementare Phänomene des Hell-Dunkel-Sehens einfach deuten: Die Veränderung der Sehschärfe und des Simultankontrastes mit Abnahme der mittleren retinalen Beleuchtungsstärke korreliert mit einer Ausdehnung der RFZentren retinaler Neuronen. Die retinale „ Lokaladaptation“ bewirkt eine Empfindlichkeitsänderung retinaler On-Zentrums- und Off-Zentrums-Neuronen, die nach längerer gleichförmiger retinaler Belichtung zu anhaltenden Nachbildern führt. Nach den Leitungsgeschwindigkeiten der von den Ganglienzellen gebildeten Optikusaxone lassen sich die retinalen Ganglienzellen in drei Klassen einteilen. Die On-Zentrum- und Off-Zentrum-Neuronen des PDJQR]HOOXOlUHQ 6\VWHPV 0 6\VWHPV) bilden schnell leitende, dicke Axone und gehören zum achromatischen System. Die Ganglienzellen des parvozellulären Systems bilden dünnere markhaltige Axone und übertragen z.T. chromatische Information. Viele Ganglienzellen der Netzhaut haben markarme Optikusaxone. Dieses koniozelluläre System besteht überwiegend aus On-Off-Neuronen, die z.T. 3 Anatomische und physiologische Grundlagen 31 bewegungsempfindlich sind, z.T. der Steuerung der Pupillomotorik und anderer vegetativer Reaktionen dienen. Insgesamt wird durch den Transduktions- und Verarbeitungsprozess in der Netzhaut das optische Bild auf der Retina in das Erregungsmuster von mindestens 10 verschiedenen Neuronenklassen umgesetzt, was dem Prinzip einer parallelen Signalübertragung im Zentralnervensystem entspricht. 6LJQDOYHUDUEHLWXQJLP]HQWUDOHQYLVXHOOHQ6\VWHP Die visuelle Information wird durch die den Sehnerv bildenden Axone der retinalen Ganglienzellen in das Gehirn übertragen (Abb. 8). Die Sehnerven beider Augen vereinigen sich an der Schädelbasis zum &KLDVPDRSWLFXP, wo die aus der nasalen Retinahälfte stammenden Sehnervenfasern zur Gegenseite kreuzen. Die Sehnervenfasern aus der temporalen Retinahälfte ziehen ipsilateral mit den gekreuzten Axonen des anderen Sehnervs im Tractus opticus zu den ersten zentralen Schaltstationen der Sehbahn, dem Corpus geniculatum laterale (CGL), den Colliculi superiores und der prätektalen Region des Hirnstamms. Die Axone der Geniculatumzellen verlaufen durch die Sehstrahlung 5DGLDWLRRSWLFD zu den Nervenzellen der SULPlUHQ 6HKULQGH $UHD VWULDWD oder $UHD 9 der okzipitalen Großhirnrinde). Von dort gehen weitere Verbindungen zu den ÄH[WUDVWULlUHQ³ visuellen Hirnrindenfeldern (V2, V3, V4) sowie zu den visuellen Integrationsregionen in der parietalen und temporalen Großhirnrinde (vgl. hierzu auch S. 29). 3 Anatomische und physiologische Grundlagen 32 Abb. 8 Schema der Sehbahn im Gehirn des Menschen (nach Schmidt u. Thews 1976). Fast alle afferente visuelle Information erreicht die Hirnrinde über die Verbindung des CGL mit der Area V1 (Abb. 9). Aus dieser werden die Signale zu den Areae V2, V3, V4 und zu den parietalen visuellen Hirnrindenfeldern übertragen. Diese Hirnareale haben ihrerseits rückläufige Verbindungen zur Area V1. Durch die Verknüpfungen der Area V1 werden die neuronalen Signale, die unterschiedliche Struktureigentümlichkeiten unterschiedliche der visuellen Hirnrindenareale verteilt Reizmuster (Prinzip repräsentieren, der parallelen auf und rückgekoppelten Informationsverarbeitung in der Großhirnrinde). Abb. 9 Äußere Hirnoberfläche eines Rhesusaffen. Areae V1, V2, V3 und V4 sind visuelle Hirnrindenfelder im Okzipitallappen (nach Schmidt, Thews u. Lang 2000). 3 Anatomische und physiologische Grundlagen 33 Mit Hilfe YLVXHOO HYR]LHUWHU 3RWHQ]LDOH 9(3 können Störungen der visuellen Signalverarbeitung objektiv erfasst werden. Die Messung der visuell evozierten Potenziale ermöglicht die objektive Beurteilung der Funktion des afferenten visuellen Systems und der Area V1. Dabei registriert das Elektroenzephalogramm im Okzipitalbereich einen Impuls. Die EEG-Reaktionen auf Lichtreize werden mit Hilfe eines Digitalrechners gemittelt. Die Nervenzellen der &ROOLFXOL VXSHULRUHV reagieren bevorzugt auf bewegte visuelle Reizmuster und sind zum Teil auch richtungsempfindlich: Ein visuelles Reizmuster löst nur dann eine Aktivierung der Nervenzellen aus, wenn es in bestimmter Richtung durch das RF bewegt wird. Die Neuronen der Colliculi sind in „ Säulen“ senkrecht zur Oberfläche der Colliculi angeordnet, die innerhalb einer Säule haben ihr RF jeweils in der gleichen Region des Gesichtfeldes. In den tieferen Schichten der Colliculi gibt es Nervenzellen, die kurz vor Augenbewegungen aktiviert sind und die eine blickmotorische Kontrollfunktion haben (Berthoz et al. 1992, Cohen 1981, Creutzfeldt OD 1993, Polyak 1957, Grüsser u. Grehn 1991, Grüsser u. Grüsser-Cornehls 1973, Grüsser u. Henn 1991). Im &RUSXV JHQLFXODWXP ODWHUDOH &*/ enden die Optikusaxone in drei dem ipsilateralen und drei dem kontralateralen Auge zugeordneten Zellschichten. Das CGL besteht aus zwei magnozellulären und vier parvozellulären Schichten. Die Nervenzellen des CGL haben wie die Ganglienzellen der Retina konzentrisch organisierte rezeptive Felder. Mit einfarbigen Hell-Dunkel-Mustern werden im CGL zwei verschiedene Neuronenklassen unterschieden: „ Kontrastneuronen" und „ Hell-Dunkel-Neuronen". In beiden Neuronenklassen gibt es etwa gleich häufig On-Zentrums- und Off-Zentrums-Neuronen. Die Kontrastneuronen reagieren wegen verstärkter lateraler Inhibitionsprozesse nicht oder nur schwach auf diffuse Lichtreize, jedoch sehr stark auf Hell-Dunkel-Konturen im RF. Die Aktivierung der Hell-Dunkel-Neuronen hängt dagegen von der mittleren Leuchtdichte des visuellen Reizmusters im ganzen rezeptiven Feld ab (Grüsser u. Klinke 1971, Jung 1973). 3 Anatomische und physiologische Grundlagen 34 Durch die Leistung der Neuronensysteme der Netzhaut und des CGL werden die visuellen Signale nach ihren chromatischen Eigenschaften, der räumlichen Kontrastverteilung und der mittleren Leuchtdichte an den verschiedenen Gesichtsfeldstellen bewertet. Eine weitere Strukturierung des visuellen Signalflusses erfolgt durch die Neuronensysteme des primären visuellen Kortex, d.h. der $UHD 9. Ein großer Teil dieser Nervenzellen antwortet nicht mehr auf einfache Hell-Dunkel-Reize, sondern nur noch auf Konturen bestimmter Orientierungen, Konturunterbrechungen usw. Im Reaktionsmuster kortikaler visueller Neuronen ist also eine weitere „ Spezialisierung“ der visuellen Signalverarbeitung zu erkennen. Der Grad der Spezialisierung wird durch die rezeptiven Feldeigenschaften gekennzeichnet: einfache, komplexe und hyperkomplexe rezeptive Felder werden hier unterschieden (Grüsser u. Landis 1991, Livingstone u. Hubel 1988). Die Nervenzellen der Area V1 sind in sechs bis acht Schichten parallel und in Säulen senkrecht zur Hirnoberfläche angeordnet. Die rezeptiven Felder aller Nervenzellen in einer etwa 200-300 µm dicken Säule befinden sich im gleichen Bereich des Gesichtsfeldes, können jedoch eine unterschiedliche Ausdehnung haben. Gemeinsam für die Nervenzellen einer Säule ist die ähnliche Achsenorientierung der rezeptiven Felder. • (LQIDFKH UH]HSWLYH )HOGHU Ein Teil der Nervenzellen der Area V1 hat einfache rezeptive Felder mit konzentrisch oder parallel zueinander angeordneten On- oder Off-Zonen. Diffuse Belichtung des ganzen rezeptiven Feldes löst keine oder nur eine schwache Aktivierung aus. Als Achsenorientierung wird die Richtung der Grenze zwischen den On- und OffZonen bezeichnet. Eine Hell-Dunkel-Kontur löst die stärkste Aktivierung aus, wenn sie an und parallel zu der Grenze zwischen On- und Off-Zone liegt. • .RPSOH[H UH]HSWLYH )HOGHU Um Nervenzellen mit komplexen rezeptiven Feldern zu aktivieren, müssen differenzierte Reizmuster in das RF projiziert werden, z.B. Hell-Dunkel-Konturen bestimmter Orientierung oder Konturunterbrechungen bestimmter Ausdehnung. Komplexe RF sind in der 3 Anatomische und physiologische Grundlagen 35 Regel in ein exzitatorisches rezeptives Feld (ERF) und ein inhibitorisches rezeptives Feld (IRF) unterteilt. Eine gleichzeitige Reizung von ERF und IRF löst eine geringe oder keine neuronale Aktivierung aus. • %HZHJXQJVHPSILQGOLFKNHLW NRUWLNDOHU 1HXURQHQ Neuronen mit komplexem oder hyperkomplexem RF reagieren auf bewegte Reizmuster stärker als auf unbewegte. Diese Bewegungsempfindlichkeit kann als Anpassung an die beim normalen Sehen ständig vorhandenen Augenbewegungen gedeutet werden. Das zerebrale Bild der stationären Welt wird aus bewegten retinalen Reizmustern ermittelt. Zum Teil steuern die bewegungsempfindlichen kortikalen Neuronen über Verbindungen zu den Colliculi superiores die Abtastbewegungen der Augen beim Sehen und über Verbindungen mit der prätektalen Region die Akkommodation und die Pupillenweite. • %LQRNXOlUH $NWLYLHUXQJ Die Neuronen des visuellen Kortex haben in der Regel in jeder Retina ein rezeptives Feld. Ihr Entladungsmuster ist das Resultat einer binokulären Verrechnung und Grundlage des binokuläre Tiefensehens. Die H[WUDVWULlUHQ YLVXHOOHQ $UHDOH sind über die ganze okzipitale Hirnrinde ausgedehnt. Die visuellen Hirnrindenareale des Hinterhauptlappens (Areae V2, V3, V4) übernehmen die afferenten visuellen Signale aus den verschiedenen Neuronenklassen der Area V1. Hierbei erfolgt eine Aufteilung nach funktionellen Gesichtspunkten. Die Area V2 hat überwiegend die Aufgabe der visuellen Gestalterkennung stationärer Reizmuster, die Area V3 Gestalterkennung kohärent bewegter Objekte oder Objektteile und die Area V4 Objekterkennung aufgrund charakteristischer nachvollziehbar, Oberflächenfarben dass zur und ungestörten Farbkontraste. Wahrnehmung Es ist eines leicht visuellen Gegenstandes die koordinierte Aktivität aller drei Areale erforderlich ist. Die Signale aus den retinotop organisierten visuellen Hirnrindenarealen (Areae V2, V3, V4) werden in die visuellen Assoziations- und Integrationsregionen des Parietal- und Temporallappens übertragen (z.B. Gyrus angularis). Mit jedem 3 Anatomische und physiologische Grundlagen 36 neuronalen Verarbeitungsschritt nimmt die retinotope Organisation ab, bis schließlich die einzelnen rezeptiven Felder der Nervenzellen in einigen der visuellen Assoziations- und Integrationsregionen einen großen Teil des fovealen und des peripheren Gesichtsfeldes einnehmen (Maunsell u. Newsome 1987, van Essen 1979, van Essen et al. 1992). 1HXURSK\VLRORJLVFKH*UXQGODJHQGHU*HVWDOWZDKUQHKPXQJ Konturen bestimmter Orientierung und Länge, Konturunterbrechungen und Winkel (wie z.B. Schrift) sind optimale Reizmuster für die Aktivierung der verschiedenen Neuronenklassen in der Sehrinde. Im räumlichen Erregungsmuster des Neuronennetzes jeder Klasse werden jeweils andere Eigenschaften („ Gestaltmerkmale“ ) des Reizmusters „ abgebildet“ . In Abb. 10 ist am Beispiel des Reizmusters „ A“ schematisch das räumliche Erregungsmuster angegeben, das dieses Reizmuster in der Netzhaut, in den Kontrastneuronen des CGL und in den verschiedenen kortikalen Neuronenklassen auslöst. Abb. 10 Ein Leuchtbuchstabe A wird auf die Netzhaut projiziert und löst in der Rezeptorschicht der Netzhaut ein entsprechendes räumliches Erregungsmuster aus (a), das zu einem räumlichen Erregungsmuster in der retinalen Ganglienzellschicht und im Corpus geniculatum laterale (a, b) führt. Die Neuronen des visuellen Kortex reagieren entweder auf die Konturen (b), Konturen bestimmter Richtung, Konturunterbrechungen, Winkel und Ecken des A (c bis i). Die aktivierenden Gestaltmerkmale des A sind jeweils grau gezeichnet. In k ist ein Beispiel für eine visuelle Gestaltergänzung dargestellt: Drei schwarze Reizmuster sind so angeordnet, dass der Eindruck eines räumlich ausgedehnten Buchstabens A entsteht (nach Schmidt u. Thews 1977). 3 Anatomische und physiologische Grundlagen 37 Lesen ist eine höhere YLVXHOOHHirnleistung, die durch umschriebene Hirnläsionen gestört werden kann (Alexie). Durch Messung der regionalen Hirndurchblutung konnte nachgewiesen werden, dass beim Lesen eine besonders starke Aktivierung im Bereich des *\UXV DQJXODULV und des *\UXV FLQJXOL der linken Großhirnhemisphäre auftritt. Die Bedeutung dieser Großhirnrindenregionen für das Lesen ist vor allem an Patienten erkennbar, die nach einer Läsion im inneren Bereich des Gyrus angularis der linken Hirnhälfte entweder Worte nicht mehr lesen können (verbale Alexie) oder sogar Buchstaben nicht mehr erkennen, bzw. identifizieren können (litterale Alexie). Bei einer reinen Alexie kann der Patient noch schreiben, das von ihm selbst Geschriebene jedoch nicht mehr lesen (Alexie ohne Agraphie). Dehnt sich die Hirnläsion vom Gyrus angularis zum Gyrus cinguli aus, so ist die Alexie in der Regel von einer Unfähigkeit zum Schreiben (Agraphie) begleitet. Eine Läsion im Bereich der prämotorischen Hirnrinde des Frontallappens der linken Seite kann selektiv eine Agraphie ohne Alexie zur Folge haben. Die Fähigkeit, ideographische Schrift zu lesen (z.B. chinesische Schriftzeichen oder Kanji im Koreanischen und Japanischen) ist bei einer umschriebenen Läsion des linken Gyrus angularis nur wenig beeinträchtigt. Eine Alexie für diese Schriftzeichen tritt jedoch bei einer Läsion des rechten Gyrus angularis auf. 0RGHUQH7HFKQLNHQ]XU/RNDOLVDWLRQ VSUDFKUHOHYDQWHU+LUQDUHDOH Bis gegen die Mitte des 20. Jahrhunderts war man zur Lokalisation von sprachrelevanten Hirnregionen hauptsächlich auf die bei der Autopsie von verstorbenen aphasischen Patienten erhobenen pathologisch-anatomischen 3 Anatomische und physiologische Grundlagen 38 Hirnbefunde angewiesen. Mit dem Aufkommen neuer Techniken wurde es möglich, auch am Lebenden und Gesunden Regionen, die für die Sprache von Bedeutung sind, zu lokalisieren. 1XNOHDUPHGL]LQLVFKH0HWKRGHQ Das nach Applikation von Radionukliden in die Blutbahn mit Hilfe von Detektoren durchgeführte Hirnscanning bestätigt die schon früher gemachte Beobachtung, dass Patienten mit nicht flüssiger Aphasie eine Schädigung in der vorderen perisylvischen Region aufweisen und Patienten mit flüssiger Aphasie in oder um die hintere perisylvische Region geschädigt sind (Benson u. Patten 1967). Die Emissionstomographie, insbesondere die Positronen-Emissions-Tomographie (PET), ist eine wichtige Informationsquelle bei neuropsychologischen Studien der höheren Hirnfunktionen (Frackowiak 1994, Herholz et al. 1996, Raichle 1987). Mit der PET kann in verschiedenen sprachrelevanten Hirnarealen eine Aktivitätssteigerung (über vermehrten Blutfluss als Ausdruck einer gesteigerten Stoffwechselaktivität bei erhöhter Funktion der Nervenzellen) beobachtet werden, je nachdem, welche Sprachtests (Benennung von Objekten, Wortfindung u.a.m.) vorgelegt werden. ,QWUDRSHUDWLYQHXURSK\VLRORJLVFKH0HWKRGHQ Eine weitere Methode ist die von PENFIELD und BOLDREY 1937 eingeführte, von PENFIELD und ROBERTS (1959) auf die Sprachregionen ausgedehnte und 3 Anatomische und physiologische Grundlagen 39 von Ojeman et al. (1989) weiter entwickelte elektrische Stimulation kleiner Areale der operativ freigelegten Hirnoberfläche (Penfield u. Boldrey 1937, Penfield u. Roberts 1959, Ojeman et al. 1989). Die Methode kommt allerdings nur bei Patienten infrage, die sich wegen einer fokalen Epilepsie oder einem Hirntumor im Bereich des Sprachkortex einer Operation in Lokalanästhesie unterziehen müssen. Es geht darum, beim Eingriff sprachrelevante Areale möglichst zu schonen (Herholz et al. 1997, Reulen et al. 1997). Dabei wird den Patienten je nach Testwahl z.B. ein Gegenstand gezeigt, den sie benennen müssen (z.B. „ Dies ist eine Schere“ ). Bei diesen Stimulationen mit kurzer Applikation eines elektrischen Stroms von wenigen mA in kleinen, voneinander getrennten Arealen (brain mapping) zeigt sich Folgendes: Wird ein Kortexareal gereizt, das zur Benennung des Objekts nicht notwendig ist, kann das vorgezeigte Objekt einschließlich Trägersatz fehlerfrei und sprachlich korrekt identifiziert werden. Ist das gereizte Kortexareal zur Benennung des Objekts unverzichtbar, so wird die Antwort fehlerhaft (z.B. „ Säge“ statt „ Schere“ ), es liegt somit eine Sprachstörung infolge des elektrischen Reizes vor. Ist die Antwort während des Reizes unverständlich oder kommt es zu überhaupt keiner Sprachproduktion, so besteht eine Sprachhemmung bzw. eine Sprechhemmung. Die für die Sprachfunktion wichtigen kleinen Areale – etwa 1 cm2 –, im erwähnten Testfall für Benennung, variieren sowohl hinsichtlich Anzahl als auch Lokalisation innerhalb der klassischen Sprachregion von Patient zu Patient. Diese kleinen mosaikartig verstreuten Areale sind voneinander durch stumme Abschnitte getrennt und können sogar außerhalb der klassischen Sprachregion liegen. Obschon es Areale gibt, von welchen aus die Sprache gestört werden kann, so gibt es doch nur wenige Areale, die für die Sprache (im erwähnten Testfall für Objektbenennung) von essenzieller Wichtigkeit sind, was auf die große Bedeutung der Namensgebung in der Sprachfunktion hinweist. Das will aber nicht heißen, dass der Name an diesem Punkt gespeichert ist, sondern es bedeutet lediglich, dass dieses Areal als „ check-point“ wirkt und die zahlreichen, ihm zuströmenden Impulse aus dem zerebralen Netzwerk für Namensgebung zur korrekten sprachlichen Äußerung verarbeitet. Im Hinblick auf die wichtigen 3 Anatomische und physiologische Grundlagen 40 Areale außerhalb der klassischen Sprachregion stellt sich zudem die Frage nach der Plastizität des Gehirns: Andere Regionen können bei einem Prozess in sprachrelevanten Bezirken die essenzielle Funktion des „ check-point“ im Netzwerk übernehmen. Dieses Problem ist noch nicht geklärt (Huber et al. 2000). $QJLRJUDSKLHXQG:DGD7HVW Eine andere, weniger invasive Methode, um selektiv, vorübergehend und reversibel umschriebene Teile des Gehirns auszuschalten, ist im Rahmen der Angiographie auf dem Blutweg möglich (Huber et al. 2000). Bereits 1949 hatte Wada ein kurz wirksames Schlafmittel (Amobarbytal) in die A. carotis interna einer Seite injiziert und dabei herausgefunden, dass dadurch eine Großhirnhälfte für einige Minuten außer Funktion gesetzt werden kann (Wada 1949). Dieser sogenannte „ Wada-Test“ oder auch „ Amytal-Test“ wird heute als globaler Karotis-Test oder als selektiver Test (Wieser et al. 1994) eingesetzt. Die Durchführung des Wada-Tests erfolgt in der Regel im Rahmen einer diagnostischen Angiographie. Dazu wird nach örtlicher Betäubung von der Leistenarterie aus ein Katheter zunächst in der einen, dann in der kontralateralen A. carotis interna platziert, und es werden in der Regel 100-150 mg Phenobarbytal injiziert. Gleichzeitig wird im Angiographie-Raum ein EEG abgeleitet, um zu sehen, in welchem Ausmaß und in welcher Ausbreitung das Gehirn „ gelähmt“ wird. Während der Wirkung des Schlafmittels auf das Gehirn werden dann neuropsychologische Tests durchgeführt. Damit kann selektiv geprüft werden, welche Aufgaben die nicht ausgeschaltete kontralaterale Hirnhälfte zu bearbeiten und zu lösen in der Lage ist. Sozusagen als Umkehrbeweis erfolgt dann, wenn sich die primär lahmgelegte Hirnhälfte wieder erholt hat, der globale Karotis- 3 Anatomische und physiologische Grundlagen 41 Amytal-Test auf der Gegenseite. Mit diesem globalen Wada-Test lässt sich vor einem geplanten operativen Eingriff festlegen, ob bei dem Patienten – bezogen auf die Sprache – eine links- oder rechtshemisphärische oder eine gemischte Dominanz (bilaterale Sprachpräsentation) vorliegt. Im Rahmen des selektiven und superselektiven Amytal-Tests werden einzelne Hirngefäße mit einem Mikrokatheter sondiert, der durch den transfemoral eingeführten Führungskatheter in Teleskop-Technik in den Hirnkreislauf vorgeschoben werden kann. Dieser Eingriff ist schmerzfrei, erfolgt bildgesteuert und ist zwischenzeitlich ein Routineeingriff in interventionellen neuroradiologischen Zentren geworden. Die Angiographie dieses Hirngefäßes durch den Mikrokatheter zeigt, welches Hirnareal von diesem Gefäß selektiv versorgt wird und welcher Bereich des Gehirns ausgeschaltet wird, wenn 40-60 mg Amytal durch den Mikrokatheter in dieses Hirngefäß injiziert werden. Die reversiblen, etwa 10-15 Minuten dauernden neurologischen Ausfälle werden dann durch standardisierte Tests im Angiographie-Raum durch die anwesenden Neurologen und Neuropsychologen gemessen und dokumentiert. Vergleichbare Ergebnisse lassen sich erzielen, wenn während der Bearbeitung sprachlicher Aufgaben Doppler-sonographisch der Fluss in den mittleren Hirnarterien gemessen wird. Auf der sprachdominanten Seite kommt es zu einem signifikanten Flussanstieg (Rihs et al. 1999). &RPSXWHUWRPRJUDSKLHXQG0DJQHWUHVRQDQ]WRPDJUDSKLH 1967 wurde die Computertomographie (CT) eingeführt (Ambrose 1973, Haunsfield 1973), 10 Jahre später folgte die Kernspintomographie (Magnetic Resonance Imaging, MRI) (Bloch 1946, Bydder 1982, Lauterbur 1973, Mansfield 3 Anatomische und physiologische Grundlagen 42 1976, Purcell 1946). Mit diesen beiden bildgebenden Methoden wurde es möglich, das Hirnparenchym mit großer Präzision und guter Strukturauflösung von grauer und weißer Substanz (Rinde und Mark, zentrale Kerngebiete) am Lebenden darzustellen. Krankhafte Veränderungen, wie z.B. ischämische Hirninfarkte, werden mit einer bis in die kleinsten Details gehenden Genauigkeit abgebildet. Dies ist für die Aphasieforschung von großem Vorteil, da das Ausmaß der zerebralen Läsion bereits im Initialstadium (und zu jedem beliebigen späteren Zeitpunkt) erfasst und mit dem jeweiligen sprachlichen Defizit in Beziehung gebracht werden kann (Damasio 1989, Damasio 1992). So hat sich gezeigt, dass die echte sog. Broca-Aphasie mit der nicht flüssigen Sprache, dem Agrammatismus usw. nicht nur durch Läsionen des unteren Gyrus frontalis (Brodmann Area 44 und 45), sondern auch durch eine Schädigung der umliegenden frontalen Bezirke (Brodmann Areale 46, 47 und 9) sowie der darunter liegenden weißen Substanz und der Basalganglien (Kopf des Nucleus caudatus und des Putamen) verursacht sein kann. Läsionen des ursprünglichen Broca-Zentrums (hinterer Abschnitt der 3. Frontalwindung) verursachen nicht die typische Broca-Aphasie, sondern nur eine milde und vorübergehende Form ohne Halbseitenlähmung. Die Wernicke-Aphasie ihrerseits wird durch eine Schädigung verursacht, die nicht nur die obere Temporalwindung (Brodmann Area 22) einschließt, sondern auch die parietalen Regionen des Gyrus supramarginalis (Brodmann Area 40) und des Gyrus angularis (Brodmann Area 39) sowie die mittlere Temporalwindung (Brodmann Area 37). Ebenso kann bei anderen Formen der Aphasie die Läsion genau lokalisiert werden. Es zeigt sich auch, dass eine Schädigung der Basalganglien der sprachdominanten Hirnhemisphäre oder begleitende Läsionen kortikaler Strukturen zu Aphasien führen können. In gewissen Fällen lassen sich sogar Einzelheiten von aphasischen Störungen bestimmten Regionen, z.B. der vorderen Temporalregion, zuordnen. Eine Schädigung der Brodmann Area 38 (Spitze des Temporallappens) führt z.B. zu Benennungsschwierigkeiten von Personen, nicht aber zu solchen von allgemein gebräuchlichen Gegenständen. 3 Anatomische und physiologische Grundlagen 43 Eine Läsion der mittleren und unteren Temporalwindung (Brodmann Areale 20 und 21) löst eine Störung von Einzel- oder Allgemeinbezeichnungen aus, nicht aber von Verben, Adjektiven und grammatikalischen Wörtern. Das heißt aber nicht, dass diese Wörter in diesen Regionen wie in Schubladen gespeichert sind, sondern nur, dass diese Strukturen im gesamten Netzwerk intakt sein müssen, damit die entsprechenden Worte gefunden werden können. Seit ihrer Einführung in den frühen 90er-Jahren (Ogawa 1992) hat die funktionelle MR-Bildgebung (FMRI functional magnetic resonance imaging) eine zunehmende Verbreitung in der neurophysiologischen und neuropsychologischen Grundlagenforschung sowie inzwischen auch in der Klinik gefunden (Huber et al. 2000). *UXQGODJHQGHUIXQNWLRQHOOHQ05%LOGJHEXQJ)05, FMRI wird üblicherweise mit der als BOLD – ein Akronym für Blood Oxygenation Level dependent – bekannten Technik durchgeführt (Bandettini 1992, Bandettini 1994, LeBihan 1995, Moseley 1995). Diese basiert darauf, dass die zerebrale vasomotorische Autoregulation bei neuronaler Aktivierung zu einer lokal vermehrten Durchblutung führt; dies wiederum bedeutet, dass aktivierte Hirnareale mehr Sauerstoff erhalten. Diese Zunahme der Sauerstoffversorgung erfolgt jedoch überschießend, sodass der zugeführte Sauerstoff vom aktivierten Hirngewebe nicht völlig verbraucht wird. Infolgedessen enthält das Blut in den Kapillaren und den aus einem aktivierten Hirnareal drainierenden Venen noch einen hohen Sauerstoffanteil in Form von Oxyhämoglobin. Dieser physiologische Effekt kann benutzt werden, um ein aktiviertes Hirnareal zu identifizieren, da Oxyhämoglobin (HbO2) andere magnetische Eigenschaften hat als DeoxyHämoglobin (Deoxy-Hb). Diamagnetisches HbO2 hat nur einen geringen Einfluss auf das lokale Magnetfeld, während paramagnetisches Deoxy-Hb lokale Feldinhomogenitäten verursacht. Eine höhere lokale HbO2-Konzentration 3 Anatomische und physiologische Grundlagen 44 verursacht damit entsprechend weniger Signalverlust als das Deoxy-Hb in der Umgebung. Benutzt man eine Messsequenz, die für diese Signalverluste (den sog. T2-Effekt) besonders empfindlich ist, wird diese Region daher mit einem relativ höheren Signal zur Darstellung kommen. Eine Bildgebungstechnik, welche die Anforderungen der FMRI – Messen eines großen Volumens in sehr kurzer Zeit – ideal erfüllt, ist die echoplanare Bildgebung (echo planar imaging [EPI]). Obwohl Mansfield (Mansfield 1977) die zugrunde liegende Theorie schon vor mehr als 20 Jahren beschrieben hat, sind erst in den letzten Jahren MR-Scanner verfügbar geworden, bei denen die nötigen sehr leistungsfähigen Gradientensysteme mit hohen Anstiegsraten in klinischen Systemen implementiert werden konnten. EPI erlaubt eine extrem schnelle Bildakquisition (unter 100 Millisekunden für die einzelne Schicht) und ist daher geeignet, relativ schnelle physiologische Prozesse zu untersuchen. Eine komplette FMRI-Studie unter Anwendung der EPI-BOLD-Technik mit Untersuchung des gesamten Gehirns kann unter Anwendung der unten beschriebenen Aktivierungstechniken in 5-7 Minuten durchgeführt werden; sie lässt sich damit problemlos in die klinisch-diagnostische Untersuchung integrieren. Grundlage für den Nachweis einer Aktivierung ist der Vergleich zwischen zwei Zuständen, der durch den wiederholten Wechsel zwischen Ausführen einer Aufgabe (Paradigma) und einem Kontrollparadigma erreicht wird. Die zeitliche Limitierung beider Phasen ist wünschenswert, um Gewöhnungseffekte zu vermeiden. Das Kontrollparadigma kann entweder aus „ Ruhe“ (Nicht-Ausführen der Aufgabe oder irgend einer anderen ähnlichen Tätigkeit) bestehen oder ein anderes Paradigma sein, von dem aus neuropsychologischer Sicht zuverlässig erwartet werden kann, dass es zu einem anderen Hirnaktivierungsmuster fährt. Beispielsweise wechseln HUBER et al. (Huber et al. 2000) alle 24 Sekunden zwischen Aufgabe und Ruhe/Kontrollaufgabe. Mit der verwendeten FPI-Sequenz werden in 6 Sekunden 30 überlappende Schichten durch das gesamte Gehirn akquiriert, sodass in jeder Phase vier Bildserien entstehen. Insgesamt werden 3 Anatomische und physiologische Grundlagen standardmäßig 68 Serien 45 akquiriert; entsprechend werden in einer Gesamtmesszeit von etwa 6,5 Minuten 2040 Bilder gewonnen. Der relative Signalanstieg in einem aktivierten Kortexareal beträgt nur etwa 2-5% und ist in einem einzelnen Bild nicht nachweisbar. Die Aktivierungsinformation muss daher unter Heranziehung aller während einer funktionellen Aufgabe akquirierten Bilder errechnet werden. Grundsätzlich wird dazu ein statistischer Vergleich zwischen den während der Aufgabe und den während der Kontrolle (Ruhe, andere Aufgabe) akquirierten Bildern durchgeführt. Dabei werden pixelweise die Mittelwerte und Standardabweichungen aller „ Aktivierungs“ - und „ Ruhe“ -Pixel berechnet und mittels parametrischer (z-score, t-Test) oder nichtparametrischer (Wilcoxon-Test) Tests auf signifikante Unterschiede überprüft; entsprechende Korrekturfaktoren (z.B. Bonferroni-Korrektur) sind wegen des großen Stichprobenumfangs zu berücksichtigen. Die Untersuchung höherer kognitiver und motorischer Funktionen erfordert in der Regel den Einsatz spezieller Hilfsmittel (Gutbrod et al. 2000). Visuelle Information kann dem Patienten mittels eines Projektionssystems und eines Spiegels an der Kopfspule zugänglich gemacht werden, eine andere Möglichkeit sind spezielle Brillen mit kleinen integrierten LCD (liquid crystal display)Anzeigen. 4 Reine Worttaubheit 46 5HLQH:RUWWDXEKHLW $OOJHPHLQH*UXQGODJHQ %HJULIIVHUNOlUXQJHQXQG'HILQLWLRQHQ Der Begriff der UHLQHQ:RUWWDXEKHLW wurde Anfang des 20. Jahrhunderts geprägt, um das neuropsychologische Syndrom einer Störung des auditiven Objektund/oder Worterkennens zu beschreiben. Die reine Worttaubheit ist ein Syndrom, das sich aus einem vollständigen initialen Hörverlust (Rindentaubheit) heraus entwickeln kann. Sie ist gekennzeichnet durch eine Beeinträchtigung des auditiven Sprachverständnisses, des Nachsprechens und des Diktatschreibens, bei besser erhaltener Spontan- und Schriftsprache (Engelien 2001). Es ist eine generelle, oft sehr große Unaufmerksamkeit auf Gehörseindrücke aller Art vorhanden; die Worttaubheit ist in den einwandfreien Fällen eine bleibende. Mit der maximalen Worttaubheit kontrastiert die nicht oder nur spurweise gestörte, auffallend gute innere Sprache. Die Wortfindung ist tadellos, Paraphasien fehlen ganz oder sind selten. Einzelne der Kranken lesen und schreiben fast fehlerfrei; bei anderen bestehen Schreibstörungen, die aber nicht zum Wesentlichen des Bildes zu gehören scheinen. Entwickelt hat sich diese Aphasieform bisher immer aus einem Syndrom, in dem anfangs über kurze Zeit eine Wernicke-Aphasie bestanden hatte. Die Störung des Sprachverständnisses blieb bestehen, während die innere Sprache sich in kürzerer oder längerer Zeit weitgehend erholte (Pötzl 1919). Charakteristisch für die reine Worttaubheit ist der Unterschied zwischen dem Verstehen von phonologischer und dem Nichtverstehen von semantischer Information: Zwar kann der Patient ein gesprochenes Wort nachsprechen, ihm ist es jedoch nicht möglich, ein gesprochenes Wort zu verstehen. Der Patient kann 4 Reine Worttaubheit 47 verstehen, was er liest. Dagegen muss die Fähigkeit zu schreiben nicht unbedingt intakt bleiben. Besteht eine Worttaubheit, bei der der Zugriff auf die richtigen phonologischen Repräsentationen des Gehirns unversehrt ist, jedoch die Zuordnung zu den entsprechenden semantischen Repräsentationen misslingt, so bleibt die Fähigkeit zu schreiben erhalten. Ist die phonologische Analyse der Sprache zwar korrekt, jedoch kann der erzeugte phonologische Code nicht der phonologischen Repräsentation zugeordnet werden, so ist auch das Schreiben nicht mehr möglich.(Kohn u. Friedman 1986). Auch nach MAYER (Mayer 2001) ist die Diskrepanz von erhaltener auditiver Wahrnehmungsfähigkeit und Diskriminationsfähigkeit gegenüber einer Unfähigkeit des Erkennens des auditiven Objektes (d.h. der semantischen Bedeutung des komplexen akustischen Reizes) charakteristisch. Oft erleben die Patienten die verbalen Mitteilungen anderer Menschen ,,als Geräusch". Vokalische und suprasegmentale Eigenschaften sprachlicher Äußerungen (Intonation) scheinen weniger betroffen zu sein als Konsonanten und ermöglichen in gewissen Grenzen residuale sprachlich-perzeptive Leistungen. Ein Stück weit lässt sich auch durch Lippenlesen oder Verlangsamung des Sprechtempos das auditive Defizit kompensieren. Soweit untersucht, ist zusätzlich immer eine Störung der Diskrimination nonverbaler Geräusche, der Wahrnehmung von Musik, der Zeitauflösung akustischer Signale (Ordnungs- und Fusionsschwellen) und/oder der Lokalisation von Schallquellen im Raum zu beobachten (Mayer 2001). Die reine Worttaubheit ist selten. Oft tritt sie in Zusammenhang mit anderen Aphasieformen auf (vgl. 3.2.4). 4 Reine Worttaubheit 48 +LVWRULVFKH$VSHNWH Im Jahre 1919 beschrieb der Arzt Otto PÖTZL einen Fall reiner Worttaubheit, d.h. eines Patienten, „ dem das Wortlautverständnis fehlte, während er Geräusche, Klänge, die kontinuierliche Stimmgabelreihe bis auf eine geringe Verkürzung der Perzeptionsdauer vollkommen hörte“ (Pötzl 1919). Nach einem Unfall litt der Patient unter einer linksseitigen Hörstörung. Neun Jahre später begannen Schwindelanfälle und häufige Kopfschmerzen. Es entwickelte sich eine hochgradige Aufmerksamkeitsstörung für alle Gehörseindrücke. Im Verlauf der Erkrankung wandelte sich die reine Worttaubheit des Patienten in eine gewöhnliche mit Störungen der inneren Sprache um, sodann zeigte sich wieder eine Umwandlung der gewöhnlichen Worttaubheit in eine reine Worttaubheit. Der Patient starb am 5.4.1916. Die Autopsie des verstorbenen Patienten stellte einen großen Herd im Schläfenlappen und Scheitellappen der rechten Hirnhälfte fest; der linksseitige Herd zeigte sich am Hirnrelief durch eine kleine, auffallend atrophische Stelle im Temporallappen. Auch andere Wissenschaftler beschrieben zu Anfang des 20. Jahrhunderts Fälle reiner Worttaubheit (Bonvicini 1918, Liepmann u. Pappenheim 1914), wobei sie großen Wert auf die genaue Darstellung des autoptischen Befunds legten. Wie die Pioniere der Aphasiologie (Broca 1861, Wernicke 1874) nahmen die Wissenschaftler lokalisierte Zentren für die sprachlichen Äußerungen und das Sprachverständnis an, die relativ unabhängig und nur durch unidirektionale Bahnen miteinander verbunden seien (sog. Diagrammatiker). Diese vereinfachte Auffassung wurde jedoch auch kritisiert. So griff unter anderem Pierre MARIE 1906 die Konzeption von BROCA und WERNICKE, d.h. den Unterschied zwischen motorischer und sensorischer Aphasie, an, wobei er sich auf eine genaue 4 Reine Worttaubheit 49 Untersuchung des von BROCA beschriebenen Gehirns des Patienten Leborgne stützte (Marie 1906). Dabei stellte er fest, dass durch die Destruktion nicht nur der hintere Abschnitt der 3. Frontalwindung betroffen war, sondern auch die obere Temporalwindung. (Die computertomographische Untersuchung dieses Gehirns 1980 durch CASTAIGNE et al. [Castaigne et al. 1980] bestätigte Pierre MARIEs Beobachtung.) Pierre MARIE (Marie 1906) vertrat die Ansicht, dass jeder aphasische Patient einen gewissen Defekt im Wortverständnis aufweise und dass seine intellektuellen Fähigkeiten vermindert seien. Eine Läsion, die auf den hinteren Abschnitt der 3. Frontalwindung beschränkt sei, führe nur zu einer Anarthrie, also einer Sprachstörung rein motorischer Art, ohne Beeinträchtigung der Intelligenz, inneren Sprache und des Sprachverständnisses. Henry HEAD (Head 1926) entwickelte am Anfang des 20. Jahrhunderts im Anschluss an Jackson auf Grund von Tests eine dynamische Konzeption. Er hielt den Versuch, anatomische Veränderungen in verschiedenen Hirnbezirken als Zentren für Sprache, Lesen, Schreiben und Wortgedächtnis anzusehen, für verfehlt. Für ihn spielten mentale Funktionsstörungen bei Sprachstörungen eine wichtige Rolle. Er klassifizierte die Sprachstörungen deshalb in vier Gruppen: verbale, syntaktische, nominale und semantische. Auf die große Zahl von Publikationen über Sprachstörungen und deren Einteilung in Syndrome in den folgenden Jahrzehnten kann im Rahmen dieser Übersicht nicht eingegangen werden, weshalb auf die Arbeiten von Sir R. BRAIN vom Jahre 1961 (Brain 1961) und von BENSON und GESCHWIND (Benson u. Geschwind 1985) verwiesen sei. 4 Reine Worttaubheit 50 3DWKRJHQHVH Mehrere Untersuchungsergebnisse deuten jedoch daraufhin, dass der reinen Worttaubheit eine sprachunspezifische Störung zugrunde liegt. So vertreten zahlreiche Forscher die These, dass am Anfang der Sprachstörungen Probleme bei der elementaren akustischen Diskrimination insbesondere nur kurz dargebotener Laute bestehen. Andere Wissenschaftler sind jedoch der Ansicht, dass die reine Worttaubheit durch Störungen in den Sprachzentren des Gehirns entsteht. 5HLQH:RUWWDXEKHLWGXUFK6W|UXQJHQLQGHQ6SUDFK]HQWUHQ GHV*HKLUQV Die neuen Techniken (PET, kortikale Stimulation, CT, MRI, FMRI, s. 3.5) erlauben eine präzise Lokalisation am Lebenden und eine Korrelation mit den vorhandenen Sprachstörungen und lassen die Entwicklung einer Sprachstörung im Verlauf der Zeit verfolgen. Dank dieser Techniken sind neue Konzepte über die neuronalen Systeme und ihre funktionelle Bedeutung für Sprache und Gedächtnis entstanden. Es zeigte sich, dass nicht umschriebene Zentren für bestimmte sprachliche Leistungen zuständig sind, sondern dass eine synchronisierte Aktivität in ausgedehnten neuronalen Netzwerken von Regionen in der Hirnrinde, in den subkortikalen Kerngebieten und den Verbindungen dieser Bezirke in reziproker Weise stattfindet (Huber 2000). Dies ist möglicherweise der Grund, warum in der Literatur keine Einigkeit darüber besteht, wo die Schädigungen, die zu einer reinen Worttaubheit führen, genau in den Sprachzentren des Gehirns lokalisiert sind. Es ist sogar umstritten, ob eine bestimmte Aphasieform mit einer Schädigung eines bestimmten Ortes verbunden ist. SCHMIDT et al. sind der Überzeugung, dass die verschiedenen Aphasieformen auf abgrenzbaren Unterschieden der Läsionsorte im Gehirn beruhen. Die transkortikale sensorische Aphasie, bei der die Fähigkeit 4 Reine Worttaubheit 51 nachzusprechen zwar gut erhalten ist, wohingegen Defizite im Sprachverständnis ausgeprägt sind, beruhe auf Läsionen in der Nähe des linken perisylvischen Bereichs (Schmidt et al. 2001). HUBER et al. glauben dagegen, dass die alte Theorie, dass Aphasien mit lokalisierten Läsionen verbunden seien (verschiedene Aphasiesyndrome mit spezifisch lokalisierten Läsionen), nur teilweise ihre Gültigkeit behalten habe. Die Auffassung, dass die Zuordnung einer verlorenen Funktion auf einen einzigen umschriebenen geschädigten Bezirk möglich sei, müsse korrigiert werden (Huber et al. 2000). Auch OTSUKI et al. stellen beim Vergleich verschiedener wissenschaftlicher Beiträge fest, dass keine Beziehung zwischen dem Aphasietyp und dem pathologischen Befund herzustellen war (Otsuki et al. 1998). Die Worttaubheit kann einerseits mit bilateralen Läsionen der Temporalregionen verbunden sein (Auerbach et al. 1982), zum anderen mit Läsionen, die auf den linken Temporallappen beschränkt sind (Luria 1976). ELLIS u. YOUNG geben an, dass Läsionen im Temporallappen der linken Hemisphäre eine reine Worttaubheit verursachen können. Der linke Temporallappen kontrolliere bei den meisten rechtshändigen Menschen zahlreiche Sprachfunktionen (Ellis u. Young 1988). PRAAMSTRA et al. beschreiben einen Patienten, der nach einem linksseitigen temporoparietalen Infarkt worttaub wurde und nach einem rechtseitigen Infarkt auch das semantische Sprachverständnis verlor (Praamsta et al. 1991). PRAAMSTA et al. sind der Überzeugung, dass die Ursache der reinen Worttaubheit auf kombinierten Defekten beruht. Auch im Falle VON STOCKERTs war es zu einer bilateralen Läsion der Temporalregionen gekommen (von Stockert 1982). Nach einem Selbstmordversuch litt der 23-jährige Patient unter einer reinen Worttaubheit. Der Mann hatte sich eine Kugel durch den Kopf geschossen, und die Kugel hatte die Schädeldecke in unmittelbarer Nähe der rechten Temporalregion zwei cm oberhalb des Ohres durchbohrt, das Gehirn in ventrokaudaler Richtung 4 Reine Worttaubheit 52 durchdrungen und war in der linken temporoparietalen Region in der Nähe der Schädeldecke stecken geblieben (von Stockert 1982). Das rechte Temporalhorn des Seitenventrikels wurde verletzt und füllte sich mit Blut. Am nächsten Tag wurde eine Operation aufgrund eines progressiven temporalen Hämatoms durchgeführt. Während der folgenden Tage erlangte der Patient das Bewusstsein wieder, aber sein Zustand war weiterhin verwirrt. Der Patient war unkooperativ und reagierte inadäquat, wenn er angesprochen wurde. Langsam besserte sich der Zustand über mehrere Wochen. Anschließend blieben eine leichte linksseitige Ataxie, ein bilateraler oberer Gesichtsfeldausfall und eine Verständigungsschwierigkeit bestehen, die als primäre Worttaubheit diagnostiziert wurde (von Stockert 1982). Auch acht Monate später hatte sich der neurologische Zustand nicht gebessert. Noch immer bestanden die leichte Ataxie, der Gesichtsfeldausfall und eine schwere Störung des auditiven Wahrnehmungsvermögens, die dem Patienten eine Unterhaltung unmöglich machte (von Stockert 1982). Die Sprache des Patienten war schnell, aber nicht dysarthrisch. Bis zu einem gewissen Grade konnte er Sätzen folgen. Er hatte mehr Schwierigkeiten, einzelne Phoneme und Silben zu verstehen als Worte bzw. Sätze. Dies machte sich besonders dann bemerkbar, wenn er zwischen zwei Wörtern unterscheiden musste, die nur in einer einzigen Silbe verschieden waren (von Stockert 1982). Es stellte sich heraus, dass der Patient unter einer Agnosie für Bilder litt, aber nicht für wirkliche Objekte. Der Patient hatte auch keine Probleme, geschriebene Wörter zu verstehen. Beim Schreiben machte er einige Fehler, die als Aphasie interpretiert werden konnten. Rechnen war absolut normal. Das Zeichnen war langsam und ungeschickt, und der Patient verirrte sich in den kleineren Details. Er hatte Probleme bei der räumlichen Orientierung. Auch gelang es ihm kaum, Geräusche zu lokalisieren und deren Bedeutung zu verstehen (von Stockert 1982). 4 Reine Worttaubheit 53 Einsilbige Wörter waren für den Patienten leichter verstehbar als zweisilbige. Ein zweiteiliger Hörtest mit 25 ein- und zweisilbigen Objektnamen wurde mit dem Patienten durchgeführt. Von den 25 Paaren identifizierte er 9 rechtsseitig und 10 linksseitig korrekt. Dieses Resultat zeigte, dass die Verletzung offenbar keine Seite eindeutig stärker betraf. Die Verständnisschwierigkeit nahm bei längeren Wörtern zu, trat jedoch weniger stark bei Sätzen in Erscheinung (von Stockert 1982). Um die Rolle, die Phonologie und Semantik bei der reinen Worttaubheit spielen, festzustellen, führte VON STOCKERT zwei Untersuchungen durch. Zuerst wurden drei Listen von Silben, die aus der Struktur Konsonant-Vokal-Konsonant aufgebaut waren, dem Patienten über Kassettenrecorder mit Kopfhörer angeboten. Der Patient wurde aufgefordert, das Gehörte sofort zu wiederholen. Es stellte sich heraus, dass bei Variation des ersten Konsonanten sowie bei Variation des Vokals die Fehlerrate relativ gering war, jedoch bei Veränderung des zweiten Konsonanten hoch. VON STOCKERT schloss daraus, dass der Patient Probleme hatte, Gehörtes in eine richtige Reihenfolge zu bringen. Bei der zweiten Untersuchung sollte der Patienten ein-, zwei- und dreisilbige Wörter sowie Silbenkombinationen ohne Sinn nachsprechen. Auch in diesem Fall schaffte der Patient das Nachsprechen kürzerer Wörter wesentlich besser. Falls der Patient ein Wort nicht verstand, konnte er nicht sagen, ob das Gehörte ein Wort oder eine sinnlose Silbenkombination war. Das Nachsprechen der sinnlosen Silbenkombination gelang wesentlich schlechter als das der Wörter mit Sinn. Oft tauschte der Patient eine sinnlose Silbenkombination gegen ein ähnlich klingendes Wort mit Sinn aus. Offensichtlich erreichte den Patienten ein gestörtes phonemisches Muster und er versuchte, das wirkliche Wort in einem inneren Wörterbuch zu finden. Im Gegensatz zum Gesunden nahm seine Verständnisschwierigkeit mit der Anzahl der Silben zu (von Stockert 1982). Als der Patient an drei aufeinander folgenden Tagen mit identischen Silbenlisten geprüft wurde, nahm seine Störung nur unwesentlich ab. Daraus schloss VON STOCKERT, dass Kompensationsmechanismen auf einem höher geordneten 4 Reine Worttaubheit 54 Mechanismus auf semantischer Ebene beruhen und daher nicht bei einzelnen Wörtern oder Silben zu beobachten sind (von Stockert 1982). Insgesamt kam VON STOCKERT zu dem Schluss, die reine Worttaubheit würde durch Störungen höherer kortikaler Funktionen verursacht. Aus den Ergebnissen der auditiven Analyse1 folgerte VON STOCKERT, dass der zugrunde liegende Mechanismus der Erkrankung darin besteht, dass es nicht gelingt, akustische Stimuli in eine richtige Reihenfolge zu bringen (von Stockert 1982). 5HLQH :RUWWDXEKHLW GXUFK 6W|UXQJHQ GHU DXGLWLYHQ :DKUQHKPXQJ Verschiedene Untersuchungsergebnisse deuten daraufhin, dass der reinen Worttaubheit eine sprachunspezifische Störung im primären auditorischen Kortex zugrunde liegt. Bestimmte Frequenzspektren akustischer Stimuli können nicht erfasst werden. 8QWHUVXFKXQJHQYRQ'HQQLV33+,//,36 PHILLIPS (Phillips 1999) ist davon überzeugt, dass genaue Angaben über die Eigenschaften des Hörprozesses einen Einblick in pathologische Prozesse, insbesondere der reinen Worttaubheit, ermöglichen. Hierbei spielen die ]HLWDEKlQJLJHQ &KDUDNWHULVWLND GHV +|USUR]HVVHV, die die sich zeitlich verändernden Eigenschaften der gesprochenen Sprache widerspiegeln, eine wichtige Rolle (Phillips 1990). Die zeitabhängige Struktur der gesprochenen 1 Bei der auditiven Analyse geht es nicht allein um die genaue Unterscheidung ähnlicher Sprachlaute, sondern auch darum, aus dem Redestrom alle Laute in der richtigen Reihenfolge herauszuhören. 4 Reine Worttaubheit 55 Sprache kann auf drei Ebenen beschrieben werden, die sich durch ihre mit der Zeit verändernden Muster voneinander unterscheiden. Diese sind die sich zeitlich verändernden Wellenformen des Schalldrucks, die zeitabhängige Abstimmung der phonetisch bedeutsamen Elemente und die zeitabhängigen Variationen der Tonhöhe. Das zeitliche Muster dieses Ereignisses hängt von der Frequenz, die im Laut enthalten ist, ab (Phillips u. Farmer 1990). Die Wellenformen des Schalldrucks stellen die Feinstruktur sich zeitlich verändernder Variationen dar. Das zeitabhängige Muster hängt von der Frequenz, die im Laut enthalten ist, ab. So beinhaltet die Wellenform geflüsterter Vokale Periodizitäten hoher Amplituden, die die Pulsierungen der Glottis abbilden. Da ihre Frequenz bei 100 Hz liegt, ist die Schwingungsdauer fast 10 Millisekunden lang. Auf diesen Schwingungen überlagern sich die feineren Wellen, die die Formanten darstellen, sodass ein Vokal identifiziert werden kann. Die Formanten werden mit Hilfe des Vokaltrakts geformt. Diese Frequenzen liegen im kHzBereich, sodass die Schwingungsdauer im Bereich von Zehntelmillisekunden liegt. Die Konsonanten, insbesondere die Stopp-Konsonanten sind geräuschvoll: Sie bestehen aus gefilterten Breit-Band-Signalen mit unregelmäßigen Wellenformen (Phillips u. Farmer 1990). Die akustischen Ereignisse, die die Phoneme kennzeichnen, zeichnen sich durch einen langsameren Charakter aus (Phillips u. Farmer 1990). Die Frequenzveränderungen, die beispielsweise für die Diphthonge oder die Formantenübergänge typisch sind, haben ein zeitabhängiges Muster, das im Bereich von Millisekunden bis zu zehn Millisekunden liegt, zum Teil auch länger. Daraus folgt, dass die Stopp-Konsonanten besondere Beachtung verdienen (Phillips u. Farmer 1990). Die Stopp-Konsonanten beginnen mit einem kurzen „ Ausbruch“ , der ein nahezu kontinuierliches Spektrum zeigt, dessen höchste Schalldruckwerte abhängig sind von den Resonanzeigenschaften des oberen Vokaltrakts zur Zeit der Artikulation. In der folgenden Ausatmungsphase verändern sich die Schalldruckwerte, wenn die Mundhöhle ihre Form ändert. Was zwei Konsonanten, deren Artikulation am gleichen Ort stattfindet, voneinander 4 Reine Worttaubheit 56 unterscheidet, ist die Zeitdauer vom „ Ausbruch“ bis zum Beginn der stimmhaften Aussprache (voice onset time, VOT). Die wichtigen akustischen Eigenschaften von Stopp-Konsonanten sind ihre flüchtige Natur und ihre zeitabhängige Struktur (Phillips u. Farmer 1990). Der „ Ausbruch“ dauert nur ein paar Millisekunden, und die Formantenübergänge erstrecken sich über einige zehn Millisekunden. Interessanterweise können Stopp-Konsonanten auf der Grundlage eines Signals, dessen Dauer im Bereich von einigen zehn Millisekunden liegt, identifiziert werden. Die zeitabhängigen Variationen der Tonhöhe folgen den Pulsierungen der Glottis (Phillips u. Farmer 1990). PHILLIPS u. FARMER (1990) schließen aus dieser Darstellung, dass die sich zeitabhängig verändernden Wellenformen der Sprachlaute vollständig deren phonetische Identität bestimmen. Zum anderen stellen PHILLIPS u. Farmer (1990) fest, dass noch nicht genau bekannt sei, welche Ebene des Nervensystems für die zeitabhängige Darstellung benutzt wird, um die Sprachsignale zu verarbeiten. Studien über die Fähigkeit des Kochlearnervs, Sprachsignale zu codieren, haben sich hauptsächlich mit der Fähigkeit befasst, mit der Neuronen oder Neuronenpopulationen die sich zeitabhängig verändernden Wellenformen der Sprachlaute repräsentieren können. Psychologische Studien dagegen haben die sich zeitabhängig verändernden Wellenformen als weniger wichtig gesehen, sondern die eher groben spektralen Variationen in dem phonetisch relevanten zeitlichen Bereich (Phillips u. Farmer 1990). Die %HVWLPPXQJ GHU )UHTXHQ]OFNH ist ein Mittel, um den zeitabhängigen Verlauf des Hörvermögens zu bestimmen (Phillips 1999). Typischerweise werden dem Hörer zwei relativ lange (d.h. Hunderte von Millisekunden dauernde) Töne angeboten. Einer dieser Töne ist in seiner Mitte für kurze Zeit unterbrochen. Aufgabe des Hörers ist es zu bestimmen, welcher der beiden Töne dies ist. Bei der Untersuchung wird die kürzeste erkennbare Unterbrechung bestimmt. Hierbei können der der Frequenzlücke vorangehende Tonabschnitt und der ihr folgende gleiche (within-channel) oder unterschiedliche Frequenzen (between-channel) haben. Bei gleicher Frequenz ist eine Unterbrechung der Nervenimpulse der 4 Reine Worttaubheit 57 entsprechenden Hörnervenfasern zum Zeitpunkt der Frequenzlücke erkennbar (Phillips 1999). Ist die Bandbreite der Trägerwellen hoch und ihre Amplitude signifikant über dem Schwellenwert, so werden Frequenzlücken von 2 bis 3 msec erkannt. In verschiedenen Experimenten untersuchte PHILLIPS, welche Wirkungen Geräusche, die eine Frequenzlücke abgrenzen, auf das Erkennungsvermögen der Lücke haben, und zwar sowohl in der Between-Channel- als auch in der WithinChannel-Alternative (Phillips 1997). In einem Versuch wurden die Personen Paaren engbandiger Geräuschfolgen ausgesetzt. Der der Frequenzlücke vorangehende Ton hatte eine Frequenz von 2 kHz und die Frequenz des folgenden wurde variiert. Die Erkennung der Frequenzlücke gelang sehr schlecht, wenn die spektrale Unterscheidung der beiden Geräusche, die die Frequenzlücke begrenzten, hoch war. Dies legt die Vermutung nahe, dass die Erkennung der Frequenzlücke sich verschlechtert, wenn hierzu ein Vergleich der Aktivitäten in verschiedenen Wahrnehmungskanälen erforderlich ist. In einem anderen Experiment zeigte sich, dass eine Verlängerung des der Frequenzlücke vorangehenden Tones ihre Erkennung in der Between-Channel-Alternative verschlechterte. Insgesamt zeigten die Versuchsergebnisse, dass der Erkennungsvorgang, der eine zeitabhängige Korrelation der Aktivitäten in verschiedenen Wahrnehmungskanälen erfordert (Between-Channel-Alternative), sich deutlich von der Within-Channel-Alternative unterscheidet. Die Bestimmung der Frequenzlücke erfordert es, eine Unterbrechung zu erkennen und ihren Zeitpunkt zu bestimmen. Die Between-Channel-Alternative ist aus zwei Gründen von besonderem Interesse. Zum einen kann sie neue Einblicke gewähren in die Mechanismen der Unterscheidung beim Hören von Sprache. Der Fluss der Sprache wird von Zeitabschnitten unterbrochen, in denen die Klänge von sehr niedriger Amplitude sind oder sogar von vollständiger Stille. Die wichtigsten „ Frequenzlücken“ der Sprache sind die phonetisch bedeutsamen - sie lassen Stopp-Konsonanten erkennbar werden. Eine über 30 msec dauernde Pause wird als lang erkannt. Zum anderen können die beim Hören wichtigen räumlichen 4 Reine Worttaubheit 58 Strukturen erforscht werden. So gibt es Hinweise darauf, dass wenigstens zwei räumliche Kanäle beim Hören von Bedeutung sind (linke und rechte Hemisphäre). Hörgestörten Kindern fällt es schwer, eine Frequenzlücke zu erkennen und sie einzuordnen. Die Ergebnisse des mit Hilfe nonverbaler Stimuli durchgeführten Tests korrelieren mit dem Ausmaß der Hörstörung. Wird das Hörvermögen trainiert, so führt dies zu einer Reduktion der Sprachstörung. Es ist daher wahrscheinlich, dass bestimmte Hörstörungen mit Störungen der Sprachentwicklung bei Kindern verbunden sind. Anhand der Ergebnisse eines weiteren Versuchs stellte PHILLIPS fest, dass der SULPlUH DXGLWRULVFKH .RUWH[ wahrscheinlich eine wichtige Rolle bei der Unterscheidung verschiedener Sprachelemente spiele. Bei 94 Neuronen des primären auditorischen Kortex der Katze bestimmte PHILLIPS die Genauigkeit, mit der die Nervenzellen auf Töne oder Geräusche in Form von elektrischen Spikes2 reagierten (Phillips 1990). Die Genauigkeit der Spike-Reaktion wurde mit Hilfe der Standardabweichung der Latenzzeit der ersten Spike-Antwort bestimmt. Die Stimuli wurden systematisch in Frequenz, Amplitude und/oder Wiederholungsrate variiert. Innerhalb eines Neurons war die Variabilität der ersten Spike-Antwort zumeist proportional zu ihrer durchschnittlichen Latenzzeit. In den Fällen, in denen eine systematische Beziehung zwischen der Genauigkeit des Antwortverhaltens und der durchschnittlichen Latenzzeit bestand, ergab sich eine lineare Korrelation bei bis zu 90% der Varianz. Bei allen 94 Neuronen lagen die minimalen Latenzzeiten im Bereich von 0,15-1,5 msec. Diese Daten deuten darauf hin, dass das Antwortverhalten auf Reize in Form von Spikes im auditorischen Kortex eine Genauigkeit zeigt, die nur wenig schlechter ist als die in den Fasern des Hörnervs (Phillips 1990). 2 Spike: Spitze, Kurvenzacke. Spikes können während der Reaktion auf einen Stimulus von der Oberfläche einer Nervenstruktur abgeleitet werden. Es sind spitze Potenziale mit steil ansteigender und abfallender erhöhter Amplitude. 4 Reine Worttaubheit 59 Aus dem zeitlichen Abstand von neuronalen Antworten auf zwei in einem kurzen Abstand folgende Reize folgt die wahrzunehmende Unterscheidbarkeit dieser Laute (Phillips 1990). PHILLIPS stellte fest, dass die Höhe des hierzu erforderlichen zeitlichen Abstands nicht mit Sicherheit bekannt sei (Philipps 1990). Aus den Untersuchungen an den 94 Neuronen des primären auditorischen Kortex der Katze schätzt PHILLIPS, dass ein Abstand zwischen 0,45 und 4,5 Millisekunden für eine wahrzunehmende Unterscheidbarkeit erforderlich sei. Die Neuronen mit den präzisesten Spike-Reaktionen erzeugten gewöhnlich Werte zwischen 0,15 und 1,5 Millisekunden (Philipps 1990). PHILLIPS stellte fest, dass bei menschlichen Hörern die Auflösung wahrscheinlich auch von der Konfiguration des Stimulus und der gestellten Aufgabe abhängig sei (Phillips 1990). Es bestünde jedoch Übereinstimmung darin, dass im einfachsten Fall der Unterscheidung von zwei zeitlich getrennten Lauten die Auflösung etwa bei 1 bis 2 Millisekunden liegen würde (Albert u. Bear 1974, Auerbach et al. 1982), ein Bereich der nahe an dem in PHILLIPS Versuch ermittelten liegt. Die Genauigkeit des Antwortverhaltens der kortikalen Neuronen reicht aus, um den zeitabhängigen Verlauf einzelner Sprachkomponenten darzustellen. Dies lässt darauf schließen, dass der primäre auditorische Kortex bei der Unterscheidung verschiedener Sprachelemente eine wichtige Rolle spielt. Auf jeden Fall zeichnet sich die beste neuronale Population durch ihre schnelle und genaue Reaktionsfähigkeit aus, welche die Unterscheidung vermittelt, und es ist anzunehmen, dass der Prozess, welcher die wahrnehmbare Unterscheidung aufzeichnet, Zugriff auf diese Neuronenpopulation hat (Phillipps 1990). Unter einer UHLQHQ:RUWWDXEKHLW leiden diejenigen Patienten, deren sprachliche Unterscheidungsfähigkeit trotz einer erhaltenen, audiometrisch messbaren Wahrnehmungsfähigkeit stark vermindert ist (Phillips u. Farmer 1990). Die reine Worttaubheit ist eine seltene Störung der auditorischen Wahrnehmungsfähigkeit, 4 Reine Worttaubheit 60 die offenbar auf sprachliche Laute beschränkt ist (Phillips u. Farmer 1990). Lesen, schreiben, sprechen und die Unterscheidung nichtsprachlicher Laute sind typischerweise erhalten, aber der Patient ist nicht in der Lage, gesprochene Sprache zu unterscheiden. Bei reiner Worttaubheit sind somit die Wahrnehmung und die Unterscheidungsfähigkeit nichtsprachlicher Laute intakt. Weitet sich die Störung auf nichtsprachliche Laute aus, wird die Krankheit als auditorische Agnosie bezeichnet. Fälle von auditorischer Agnosie haben fast immer eine Beeinträchtigung der Wahrnehmung von Sprachlauten, obwohl die Erkennung der Sprache fast intakt bleiben kann (Phillips u. Farmer 1990). Diesen beiden Gruppen kann die kortikale Taubheit gegenüber gestellt werden, die einen vollständigen Wahrnehmungsverlust beinhaltet (Phillips u. Farmer 1990). Die reine Worttaubheit ist beschränkt auf eine Läsion des auditorischen Kortex (Phillips u. Farmer 1990). Dies macht die Erkrankung aus zwei Gründen interessant. Zum einen bedeutet es, dass eine spezifisch linguistische Funktion einer klassischerweise sensorischen Hirnregion zugeschrieben wird. Zum anderen können die Patienten der Forschung wichtige Daten liefern, mit deren Hilfe die Funktionen des auditorischen Kortex bestimmt werden könnten (Phillips u. Farmer 1990). Patienten mit Läsionen des primären auditorischen Kortex erreichen bei ClickFusions-Tests3 Werte im Bereich von 15 bis 30 Millisekunden (Auerbach et al. 1982). Aus diesen Messdaten scheint unmittelbar zu folgen, dass der primäre auditorische Kortex die erreichbare zeitabhängige Auflösung vermittelt (Phillips 1990) und der Verlust des primären auditorischen Kortex die zeitabhängige Auflösung um eine Größenordnung verschlechtert. 3 Hörtest zur Frequenzauflösung. Es wird festgestellt, ab welchem Zeitabstand zwei in unterschiedlichen Intervallen dargebotene Töne getrennt wahrnehmbar sind. 4 Reine Worttaubheit 61 Zwei Studien haben rein worttaube Patienten bezüglich ihrer Fähigkeit untersucht, VOT-Zeiten zu unterscheiden (Phillips u. Farmer 1990). Ein an reiner Worttaubheit erkrankter Patient zeigte eine gestörte Unterscheidungsfähigkeit von aus Konsonant und Vokal bestehenden Silben, die sich in einer VOT-Zeit von 20 msec unterschieden, auch wenn die Silben VOT-Zeiten hatten, die nahe den Werten lagen, die die phonetische Grenze zwischen den wichtigen stimmhaften und stimmlosen Stopp-Konsonanten bestimmten. Vergleichbare Werte zeigte ein anderer Patient mit einer eher generalisierten kortikalen Hörstörung. Auch diese Ergebnisse deuten also daraufhin, dass der reinen Worttaubheit eine zeitabhängige Verarbeitungsstörung zugrunde liegt (Phillips u. Farmer 1990). Die reine Worttaubheit, die bisher als eine Störung höherer Ordnung angesehen wurde, ist somit offenbar eine grundlegende Störung der Wahrnehmung (Phillips 1995). Während die reine Worttaubheit früher als Agnosie bezeichnet wurde (Teuber 1968), muss sie mittlerweile wahrscheinlich als eine notwendige Folge einer Wahrnehmungsstörung nahe beieinander liegender akustischer Ereignisse gelten. (Phillips 1995). Dennoch kann die Erkennung von Sprache als eine Hierarchie von Prozessen angesehen werden, die mit der Erkennung des Klangs beginnt, mit der Unterscheidung des Klangs fortschreitet, ihn geeigneten phonetischen Markern zuordnet, den Klang als ein Wort erkennt und schließlich seine Bedeutung aus dem semantischen Gedächtnis zuordnet. Das Ausmaß, in welchem alle diese Prozesse voneinander unterscheidbar sind, ist eine empirische Frage (Phillips 1995). :HLWHUHUHOHYDQWH8QWHUVXFKXQJVHUJHEQLVVH NOTOYA et al. stellten einen Fall vor, bei dem die reine Worttaubheit offenbar auf einer Wahrnehmungsstörung der Lautheit und einer gestörten zeitabhängigen Auflösung des Gesprochenen beruhte (Noyota et al. 1991). Die japanische Patientin litt unter einer reinen Worttaubheit in Verbindung mit einem Landau- 4 Reine Worttaubheit 62 Kleffner-Syndrom4. Erste Störungen machten sich im Alter von vier Jahren bemerkbar. Im Alter von 20 Jahren hatte sich die Hörstörung nicht signifikant gebessert. Die Patientin konnte weder Vokale noch aus Vokalen und Konsonanten bestehende Silben erkennen. Bei den Konsonanten konnte sie zwischen stimmlosen und stimmhaften unterscheiden, aber sie hatte große Probleme, die Konsonanten innerhalb jeder Gruppe zu identifizieren (Notoya et al. 1991). Die Patientin war zunächst ein gesundes Kind mit normaler motorischer Entwicklung. Ihre ersten Worte sprach sie mit 1 Jahr und 6 Monaten. Nach Erklärung ihrer Eltern verfügte sie über ein normales Sprachverständnis, Vokabular und eine regelrechte Satzbildung. Ihre Sprachentwicklung war schneller als die ihrer jüngeren Geschwister. Zu Beginn des 4. Lebensjahres begannen ihre Eltern, Zweifel an ihrem Hörvermögen zu entwickeln, weil sie ihren Kopf nicht zur Seite wandte, wenn sie beim Namen gerufen wurde. Sie versuchte oft, die Worte zu verstehen, indem sie von den Lippen ablas (Notoya et al. 1991). Im Alter von 4 Jahren und 3 Monaten war sie wegen eines Verkehrsunfalls stationär behandelt worden. Zu dieser Zeit war sie bewusstseinsklar, aber ihr EEG zeigte Abnormitäten. Es wurde jedoch berichtet, dass ihr EEG bereits Veränderungen vor dem Unfall gezeigt hatte. Im Alter von 5 Jahren hatte sie mehrere generalisierte Krampfanfälle. Das EEG zeigte zunächst Spike-and-Wave-Wellen. Als die Patientin zwölf Jahre alt war, wechselte es in den Normalzustand. CT und MRI waren immer normal gewesen (Notoya et al. 1991). Als die Patientin im Alter vom zwanzig Jahren NOTOYA et al. vorgestellt wurde, schienen ihr sprachlicher Ausdruck und ihre Artikulation 4 Das Landau-Kleffner-Syndrom ist eine Störung, bei der ein Kind mit zuvor normaler Sprachentwicklung sowohl rezeptive als auch expressive Sprachfertigkeiten verliert, wobei jedoch die allgemeine Intelligenz erhalten bleibt; der Beginn der Störung ist begleitet von paroxysmalen Auffälligkeiten im EEG (fast immer im Temporallappenbereich, gewöhnlich bilateral, jedoch oft mit ausgedehnteren Veränderungen) und in der Mehrzahl der Fälle auch von epileptischen Anfällen. 4 Reine Worttaubheit 63 intakt zu sein, jedoch war ihr Sprachverständnis stark beeinträchtigt. Vor längerer Zeit hatte die Patientin in Verbindung mit dem Verlust des Sprachverständnisses ihre Sprechweise verändert. Sie sprach in kurzen Sätzen und zeigte Artikulationsprobleme. Zunächst war die Sprache der Patientin flüssig gewesen mit normaler Prosodie und Artikulation sowie korrekter Grammatik. Die Länge der Phrasen und der Wortschatz schienen normal und ohne verbale phonematische Paraphasien zu sein. Die Bennennung gesehener Objekte war vollständig und korrekt. Das Verständnis auch einfacher Wörter war dagegen sehr gering (5%). Die Wiederholung einzelner Wörter und Phrasen war ebenfalls stark beeinträchtigt (Notoya et al. 1991). Es waren mehrere Hörtests durchgeführt worden. Im Alter von 20 Jahren wurden jedem Ohr reine Töne präsentiert. Die Hörschwelle des linken Ohrs lag bei 15 dB und die des rechten Ohrs bei 14 dB. Im Alter von 7 Jahren waren die evozierbaren Hirnstammpotenziale getestet worden. Sie hatten sich normal gezeigt. Audiometrische Tests waren im Alter von 7, 12, 16 und 20 Jahren durchgeführt. Hierbei war die Patientin kaum in der Lage, die Stimuli zu unterscheiden. Ihre Sprachunterscheidungsfähigkeit lag zunächst unter 30%, verbesserte sich jedoch im Alter von 20 Jahren auf 45%. Die Fähigkeit der Patientin, nichtsprachliche Laute zu erkennen, wurde im Alter von 6 und 20 Jahren geprüft. Die Patientin musste Bildern die passenden Klänge zuordnen, z.B. einer Violine den Klang einer Geige. Der Patientin gelang dies in allen Fällen. Im Alter von 18 Jahren wurde ein phonemischer Unterscheidungstest nach Auerbach et al. (1982) durchgeführt. Die Patientin musste zunächst die Vokale /a, e, i, o, u/ identifizieren. Dies gelang ihr vollständig. Bei der Unterscheidung von StoppKonsonanten jedoch hatte sie große Schwierigkeiten (Notoya et al. 1991). Dieses Resultat stimmt mit den Untersuchungsergebnissen, die PHILLIPS (1990) von Patienten mit reiner Worttaubheit erhielt, überein. 4 Reine Worttaubheit 64 NOTOYA et al. (1991) sind überzeugt, dass ihre Patientin unter einer reinen Worttaubheit leidet. Sie stellen fest, dass in der japanischen Sprache die Vokale etwa 100 bis 200 Millisekunden andauern. Das Schwirren der stimmhaften StoppKonsonanten (/b/, /d/, /g/) reicht von 94 bis zu 113 Millisekunden, der Ausbruch der stimmlosen Stopp-Konsonanten (/p/, /t/, /k/) liegt bei 10 bis 42 Millisekunden. NOTOYA et al. sind überzeugt, dass ihre Patientin die kurzzeitigen Differenzen zwischen Stopp-Konsonanten nicht unterscheiden kann – zum einen aufgrund einer Insensibilität bezüglich der Lautheit, zum anderen aufgrund eines Defektes in der zeitabhängigen Auflösung (Notoya et al. 1991). Auch SAFFRAN et al. (1976) stellten einen Patienten vor, der unter reiner Worttaubheit litt. Wie bei NOTOYA et al. hatte der Patient insbesondere Probleme in der Diskriminierung von verbalen Verschlusslauten (StoppKonsonanten). Dies weist nach SAFFRAN et al. daraufhin, dass die Störung auf einem vorsprachlichen Niveau beruht (Saffran et al. 1976). Bei einem von OTSUKI et al. beschriebenen Fall reiner Worttaubheit war eine kortikale Atrophie feststellbar, die hauptsächlich in der linken oberen Temporalregion lokalisiert war, d.h. dem Ort der primären Hörrinde (Otsuki et al. 1998). Es handelte sich um einen 67-jährigen rechtshändigen Patienten, der innerhalb eines Zeitraums von neun Jahren eine reine Worttaubheit entwickelte, ohne dass andere Minderungen geistiger Fähigkeiten auftraten. Das MRI zeigte eine kortikale Atrophie insbesondere in der linken oberen Temporalregion. Die Prüfung des Hörvermögens ergab eine schwere Störung im Unterscheidungsvermögen von Silben oder Wörtern der japanischen Sprache trotz einer ausreichenden Hörschärfe. Der Patient zeigte ebenfalls eine Beeinträchtigung in der zeitabhängigen Hörunterscheidung, insbesondere im Click-Fusions-Test und im Click-Zähltest. 4 Reine Worttaubheit 65 BADDELEY u. WILSON (1991) beschrieben einen Fall von Worttaubheit, bei dem die Fähigkeit der Patientin, Wörter zu wiederholen eingeschränkt war, obwohl die audiometrisch und tympanometrisch gemessenen Werte im Normbereich lagen. Auch hatte die Patientin Schwierigkeiten, Laute aus der Umgebung zu erkennen. Ihr phonologisches visuelles Gedächtnis und ihr auditorisches Gedächtnis waren nur geringfügig beeinträchtigt. Baddeley und Wilson vermuteten daher, dass bei der Patientin grundlegende auditive Wahrnehmungsprozesse gestört waren: Es werde angenommen, dass die Patientin eine Hörstörung habe, die vor dem phonologischen Erkennen liege (Baddeley u. Wilson 1991). Die Ergebnisse all dieser Untersuchungen stimmen offenbar mit der Aussage von Phillips überein, dass die reine Worttaubheit auf einer grundlegenden zeitabhängigen Störung im primären auditorischen Kortex beruhen würde. =XVDPPHQIDVVXQJXQG6FKOXVVIROJHUXQJHQ Die reine Worttaubheit ist ein Syndrom, das sich aus einem vollständigen initialen Hörverlust (Rindentaubheit) heraus entwickeln kann. Charakteristisch für die reine Worttaubheit sind eine Beeinträchtigung des auditiven Sprachverständnisses, des Nachsprechens und des Diktatschreibens, wobei die Spontan- und Schriftsprache besser erhalten bleiben (Engelien 2001). Verschiedene Untersuchungsergebnisse deuten daraufhin, dass der reinen Worttaubheit eine sprachunspezifische Störung im primären auditorischen Kortex zugrunde liegt. So zeigten die Untersuchungen von PHILLIPS, dass die Genauigkeit des Antwortverhaltens der Neuronen des primären auditorischen Kortex ausreicht, um den zeitabhängigen Verlauf einzelner Sprachkomponenten darzustellen (Phillips 1990). 4 Reine Worttaubheit 66 Zudem weisen mehrere Befunde von PHILLIPS daraufhin, dass die reine Worttaubheit eine Manifestation einer basalen zeitlichen Verarbeitungsstörung auf vorsprachlichem Niveau ist, d.h. dass der reinen Worttaubheit eine sprachunspezifische Störung zugrunde liegt. So ist die Wahrnehmung der Frequenzlücke eine wichtige vorsprachliche Grundlage zum Verstehen von Sprache, und insbesondere Stopp-Konsonanten sind aufgrund ihrer Frequenzlücke erkennbar. PHILLIPS stellte fest, dass das Wahrnehmungsvermögen langer Vokale bilaterale Läsionen des primären auditorischen Kortex übersteht, jedoch das von Stopp-Konsonanten nicht (Phillips 1990). Da Stopp-Konsonanten aufgrund ihrer Frequenzlücken erkennbar werden, folgt, dass der primäre auditorische Kortex beim Erkennen der Frequenzlücke (und damit der Wahrnehmung von Sprache) eine entscheidende Rolle spielt. Dies weist zudem daraufhin, dass die Störung auf einem vorsprachlichen Niveau beruht: Bestimmte Frequenzspektren akustischer Stimuli können nicht erfasst werden. Die Ergebnisse von PHILLIPS wurden durch andere Untersuchungsbefunde bestätigt, u.a. die von NOTOYA et al., SAFFRAN et al., OTSUKI et al. und BADDELEY u. WILSON (Notoya et al. 1991, Saffran et al. 1976, Otsuki et al. 1998, Baddeley u. Wilson 1991). Ist das primäre Hörfeld geschädigt, so verfügt der zerebrale Kortex offenbar nicht mehr über eine akustische Darstellung von Signalen mit der entsprechenden zeitlichen Struktur, die für eine normale Spracherkennung erforderlich ist. Neuronale Antworten auf akustische Signale sind zeitlich weniger differenziert. Dies führt dazu, dass das Erkennungsvermögen der Patienten für die entsprechenden Laute abnimmt. 5 Legasthenie 67 /HJDVWKHQLH $OOJHPHLQH*UXQGODJHQ %HJULIIVHUNOlUXQJHQXQG'HILQLWLRQHQ Der Begriff der Legasthenie wurde 1916 von dem Neurologen und Psychiater P. RANSCHBURG geprägt. Er wird gleichbedeutend mit der XPVFKULHEHQHQ/HVH 5HFKWVFKUHLE6FKZlFKH/56 benutzt (Warnke 1996). Legasthenie ist die Bezeichnung für Schwächen beim Erlernen von Lesen, Schreiben und Rechtschreiben, die weder auf eine allgemeine Beeinträchtigung der geistigen Entwicklung, noch auf unzulänglichen Unterricht zurückgeführt werden können5. Probleme beim Erlernen von Lesen und Rechtschreiben werden derzeit bei mindestens 15% aller Schulkinder beobachtet. Dies kann aber nicht als Zeiterscheinung gesehen werden: Bereits im Jahr 1896 wurde von dem englischen Arzt MORGAN ein Bericht über einen normalbegabten Jugendlichen veröffentlicht, der das Lesen nicht erlernen konnte. MORGAN prägte den Begriff der „ angeborenen Wortblindheit“ , um diese Lernstörung von anderen bereits bekannten Hirndefekten zu unterscheiden. Berühmte Beispiele für die umschriebene Lese-Rechtschreib-Schwäche sind Winston CHURCHILL, Albert EINSTEIN und Hans Christian ANDERSEN. Das ICD-106 unterscheidet die isolierte Rechtschreibstörung (lediglich die Rechtschreibung, aber nicht das Lesen ist beeinträchtigt) und die Lese- 5 Entsprechend ist die Legasthenie (Dyslexie, umschriebene Lese-Rechtschreibschwäche) unter Ziffer F81.0 in der Internationalen Klassifikation der Krankheiten von 2000 (ICD 10) definiert. 6 Ab 1. Januar 2000 ist nach dem Sozialgesetzbuch V (durch das Gesundheitsstrukturgesetz geänderter ³295 SGB V) die Pflicht zur Diagnosenverschlüsselung nach ICD 10 (International 5 Legasthenie 68 Rechtschreib-Störung (das Lesen und fakultativ auch die Rechtschreibung sind beeinträchtigt). Abzugrenzen ist die „ erworbene Wortblindheit“ (vgl. 3.4.5). Diese wird diagnostiziert, wenn Lesen und Schreiben bereits erlernt sind, aber durch Erkrankungen bzw. Verletzungen bestimmter Hirnregionen verloren gehen. Diese Störungen werden als Alexie (Verlust des Lesens) oder Agraphie/Dysgraphie (Verlust/ Störung des Schreibens) bezeichnet (Warnke1990). Ungenau ist es, wenn allgemein von Lese-Rechtschreib-Schwäche die Rede ist. Hiermit kann entweder die Legasthenie gemeint sein oder eine ÄDOOJHPHLQH/HVH 5HFKWVFKUHLE6FKZlFKH³ Diese allgemeine Lese-Rechtschreib-Schwäche bezeichnet als Oberbegriff alle Formen von Problemen bei Lesen, Schreiben und Rechtschreiben, auch wenn sie zum Beispiel durch Minderbegabung, organische Behinderung oder psychische Erkrankung entstehen. Für die pädagogische Förderung ist eine eng gefasste Definition nicht notwendig, da alle Kinder ihren Fähigkeiten entsprechend unterstützt werden sollten, aber im Sinne einer einheitlichen Verwendung von Begriffen ist die Unterscheidung wichtig. Grundsätzlich lässt sich die Legasthenie als ein Syndrom auffassen, d.h. ein gemeinsames Auftreten Erscheinungsbild führen. von Symptomen, Kennzeichnend die für zu die einem bestimmten umschriebene Lese- Rechtschreib-Schwäche ist die Problematik beim Erlernen von Lesen, Schreiben und Rechtschreiben, die weder auf eine allgemeine Beeinträchtigung der geistigen Entwicklung, noch auf unzulänglichen Unterricht zurückgeführt werden kann, bei durchschnittlicher oder oft überdurchschnittlicher Intelligenz. Das Missverhältnis zwischen den Leistungen beim Lesen und Schreiben sowie den übrigen Lernleistungen ist offensichtlich. Diese Schwierigkeiten sind weder erziehungs- noch milieubedingt. Sie sind weder auf eine allgemeine Beeinträchtigung der geistigen Entwicklung noch auf einen unzulänglichen Classification of Diseases – Internationale Klassifikation der Krankheiten, Verletzungen und Todesursachen) wirksam geworden. 5 Legasthenie 69 Unterricht zurückzuführen. Eine Legasthenie ist häufig in einer Familie mehrfach zu beobachten und das ganze Leben lang anhaltend. Auch nach der Verbesserung der Lese- und Recht-Schreib-Leistung wirkt die partielle Lernschwäche fort (Bundesverband Legasthenie e.V. http://www.legasthenie.net 18.6.2001). Zusammenfassend lässt sich somit feststellen, dass Legasthenie eine Schwäche beim Erlernen von Lesen, Schreiben und Rechtschreiben ist. Oftmals besteht eine Unsicherheit im Umgang mit der geschriebene Sprache ein Leben lang (Forschungsgruppe Lese-Rechtschreibstörung http://www.kjp.uni-marburg.de/kjp. Legast/index.htm 18.6.2001). Weil sowohl in der Schule als auch in unserer Gesellschaft das Lesen und Rechtschreiben einen sehr hohen Stellenwert besitzen, entstehen aufgrund dieser Schwäche oft sehr große Probleme. Vor allem von Seiten der Eltern und der Lehrerinnen und Lehrer ist eine Förderung der Stärken dieser Kinder und das Verständnis für ihre so oft vergeblichen Bemühungen enorm wichtig. 7\SLVFKH6\PSWRPH Eine umschriebene Lese-Rechtschreib-Schwäche zeigt sich in der Regel deutlich zwischen dem zweiten und vierten Schuljahr. Die folgende Übersicht soll nur als Aufzählung von möglichen Symptomen verstanden werden. Die beschriebenen Fehler können einzeln oder in Kombination auftreten. Typisch für die LRS sind alle gravierenden Probleme beim Lesen und/oder Rechtschreiben im Vergleich zu Leistungen gleichaltriger Kinder. Das Lesen kann stockend, buchstabierend oder flüchtig und sehr fehlerhaft sein. Buchstaben und Worte werden ausgelassen, falsch serialisiert (z.B. Metathese) 5 Legasthenie 70 oder durch willkürliche Hinzufügungen verändert. Es können folgende häufig auftretende Fehler unterschieden werden (Warnke 1992): • Verwechslungen von Buchstaben im Wort wie b-d, p-q, u-n, • Umstellungen von Buchstaben innerhalb eines Wortes, • Auslassungen, • Einfügungen, • Regelfehler. Häufig kommen Verwechslungen detailähnlicher Buchstaben wie t und f vor. Es treten auch oft Richtungsverwechslungen wie d/b/p/q auf. (Altmann 2001): Bei b/d und p/q handelt es sich um vertikalaxiale Spiegelungen. Die Buchstaben b und d sind deshalb besonders schwierig auseinander zu halten, weil sie sowohl visuell ähnlich sind als auch phonologisch sehr ähnliche Laute bilden. Wichtig ist, dass es meist nicht eine mangelnde visuelle Differenzierung ist, die die Buchstabenverwechslungen verursacht. So werden etwa die Buchstaben n und u nur selten miteinander verwechselt, obwohl sie visuell sehr ähnlich sind. Da diese Buchstaben aber Laute abbilden, die wenig Ähnlichkeit mit einander haben, können sie auch besser auseinander gehalten werden (Tursky 2001). Umgestellt werden bevorzugt visuell ähnliche Buchstaben (z.B. Trum statt Turm) (Tursky 2001). Ausgelassen werden insbesondere Vokale (Altmann 2001). Eingefügt werden häufig nur gering visuell bzw. auditorisch wahrnehmbare Konsonanten (z.B. Koft statt Kopf) (Müller 2001). Ursache der Regelfehler ist das Nichtbeachten spezifischer Rechtschreibregelungen (Schpiegel statt Spiegel, Bletter statt Blätter, Ferkäuferin statt Verkäuferin), die in der deutschen Orthographie häufig vorkommen (May 1995). Die ordnungsgemäße Steuerung des Blicks entlang der Zeile von Wort zu Wort ist eine wichtige Voraussetzung für das Erlernen der Schriftsprache. Obwohl neuromotorische Störungen bislang nicht nachgewiesen werden konnten, zeigte eine Untersuchung von über 1.000 Kindern, dass bis zu 50% der Legastheniker 5 Legasthenie 71 Störungen in der Kontrolle der Blickmotorik haben (Beratung Blicklabor http://www.brain.uni-freiburg.de/bbl 18.6.2001) (vgl. 5.2.3). In Fällen, in denen ein Zusammenhang mit den Augenbewegungen besteht, berichten die Kinder auf Anfrage oft, dass sie beim Lesen die Buchstaben auf dem Papier nicht stillstehend, sondern sich bewegend sehen (Beratung Blicklabor http://www.brain.uni-freiburg.de/bbl 18.6.2001). Außerdem verlieren die Kinder oft die Textstelle, überspringen häufig Zeilen und übersehen einzelne Worte oder Wort-Teile. Charakteristisch für eine Legasthenie sind Häufigkeit und hohe Stabilität von Fehlern. Selbst nach wiederholtem Üben kann das Kind meist nicht selbst erkennen, ob das Wort richtig oder falsch gelesen bzw. geschrieben ist. Es gibt aber auch Kinder mit Legasthenie, die nicht diese typischen Fehler machen. Bei Legasthenie finden sich häufig auch allgemeine visuelle und/oder akustische Wahrnehmungsstörungen, visuelle Aufmerksamkeitsprobleme, Rhythmusstörungen und Probleme der Grob- und Feinmotorik. Sie können eventuell bereits im Vorschulalter beobachtet werden, dürfen aber nicht mit Aufmerksamkeitsstörungen und Hyperaktivität verwechselt werden. Wird die Legasthenie nicht rechtzeitig erkannt und das Kind nicht zusätzlich gefördert, geht den Kindern der Spaß an der Schule schnell verloren. Es treten Verhaltensstörungen auf, und die Persönlichkeitsentwicklung der Kinder kann behindert werden. Diese sekundären Erscheinungen werden oft mit den Ursachen der Legasthenie verwechselt. 3DWKRJHQHVH Bei jedem Kind ist eine individuelle Kombination von verschiedenen Faktoren in unterschiedlicher Gewichtung für die LRS verantwortlich (Schroth 1996). 5 Legasthenie 72 Erbfaktoren scheinen in vielen Fällen eine Rolle zu spielen. Verbindungen mit Chromosom 6 und 15 wurden nachgewiesen, andere werden zur Zeit erforscht. Derzeit gibt es keinen genetischen Diagnose-Test. Es konnten Hinweise für eine Reihe von Fehlleistungen gefunden werden, die mit Lese-Rechtschreib-Schwächen verbunden sind. Darunter finden sich phonologische Schwächen, Schwierigkeiten bei der zeitlichen Verarbeitung visueller und auditorischer Reize sowie deren Abhängigkeit von Aufmerksamkeitsprozessen und Schwächen der motorischen Steuerung. So stehen Unregelmäßigkeiten der Blicksteuerung und binokuläre Instabilität mit LeseRechtschreib-Schwächen in Verbindung. Sowohl Seh- als auch Hörschwächen können bei der Entstehung von LeseRechtschreib-Schwächen ein Rolle spielen. Einige neuroanatomische Befunde lassen sich zur Stützung der Hypothese einer visuellen bzw. auditiven Pathogenese der LRS anführen. *HQHWLVFKH)DNWRUHQ Seit den ersten Fallbeschreibungen wurde ein familiär gehäuftes Auftreten der Störung beschrieben. Mehrere Segregationsanalysen (Segregration: Aufspaltung der Erbfaktoren während der Reifeteilung der Geschlechtszellen) und Zwillingsstudien unterstützen die Bedeutsamkeit von genetischen Faktoren für diese Störung. Segregationsanalysen sprechen für das Vorliegen eines geschlechtsgebundenen, additiven oder autosomal dominanten Erbganges. Zwillingsstudien belegen eine hohe Heritabilität für die Bereitschaft, eine Rechtschreibstörung zu entwickeln, und eine mittelhohe für die Bereitschaft, eine Lesestörung zu entwickeln. Anhand von Kopplungsanalysen konnte gezeigt 5 Legasthenie 73 werden, dass relevante Loci für die Störung auf dem Chromosom 15 und 6 liegen (Schulte-Körne et al. 2001). )DPLOLHQXQG6HJUHJDWLRQVVWXGLHQ Eine familiäre Häufung der Lese-Rechtschreib-Schwäche wurde in mehreren unabhängigen Stichproben beschrieben. Das Wiederholungsrisiko für Geschwister liegt zwischen 38-62%. Es fehlen jedoch große systematische Studien, die einen Aufschluss über die familiäre Ähnlichkeit bzgl. der basalen visuellen und/oder auditiven Störungen geben könnten. Es wurde zwar immer wieder in einzelnen Familien über Auffälligkeiten in der Sprachentwicklung und bezüglich phonologischer Fähigkeiten (Hoien et al. 1989) berichtet, die auf eine mögliche familiäre Subgruppe hinweisen könnten, weitergehende Daten fehlen aber (s. hierzu auch Schulte-Körne et al. 1993). In der bisher umfangreichsten Segregationsanalyse (Pennington et al. 1990) von 204 Familien mit insgesamt 1.698 Individuen aus vier unabhängigen Stichproben konnte ausgehend von einer Prävalenz von 7,5% und einem Geschlechterverhältnis von 1,8:1 (männlich:weiblich) in drei Stichproben ein Hauptgeneffekt mit reduzierter Penetranz bei weiblichen Familienmitgliedern nachgewiesen werden. In der vierten Stichprobe wurde eine polygene Vererbung der LRS favorisiert. =ZLOOLQJVXQWHUVXFKXQJHQ Neuere Zwillingsuntersuchungen zeigen, dass bei der Lesefähigkeit der genetische Anteil an der beobachteten Gesamtvarianz zwischen 3% und 60% liegt, für die Rechtschreibung, die insgesamt seltener untersucht wurde, liegt der 5 Legasthenie 74 Anteil zwischen 60% und 70% (Tab. 1). Die großen Streubreiten schwächen allerdings die Vermutung, dass die Erblichkeit eine so große Rolle spielt. Tabelle 1: Neuere Ergebnisse von Zwillingsuntersuchungen (Schulte-Körner et al. 2001) Stevenson (1991) ! "# $ 51/62 48/61 47/47 28/50 Olson et al. (1994) 183/126 151/105 132/92 93/68 # Schätzung für verschiedene Schweregrade +nicht signifikant *p (probability) < 0.05 % &')( *+' &! ! & -,).)/ Wort-Lesen Rechtschreibung phonologisches Dekodieren orthographische Fähigkeiten (Lesen von sog. irregulären Wörtern) h2= 03-41#+ h2= 66-69* h2= 36-82* Wort-Lesen phonologisches Dekodieren (Lesen von Nichtwörtern) orthographische Fähigkeiten (PseudohomophoneErkennen) Phonologische Bewusstheit (s. 5.2.2) h2= 12-68+ h2=47* h2=59* h2=56* h2=60* Von STEVENSON (1991) wurde der Einfluss des IQ und des Schweregrades auf die Heritabilitätsschätzung von Lesefähigkeit, Rechtschreibung, phonologischen und orthographischen Fähigkeiten untersucht. Es zeigte sich, dass bei Berücksichtigung des IQ höhere Schätzungen der Heritabilität resultieren. Ein systematischer Effekt des Schweregrades zeigte sich hingegen nicht, wobei die Stichproben zur Überprüfung des Effekts z.T. sehr klein waren. Die Heritabilität für die phonologische Bewusstheit (s. 5.2.2) und für die phonologische Dekodierfähigkeit ist mittel- bis sehr hoch. Die Heritabilität der orthographischen Fähigkeiten schwankt je nach Untersuchungsmethode beträchtlich (Tab. 1). Vor dem Hintergrund der signifikanten Korrelationen, die zwischen den einzelnen Fertigkeiten beobachtet werden, untersuchten OLSON et al. (1994), in welchem Ausmaß der Zusammenhang zwischen Wort-Lesen und phonologischer 5 Legasthenie 75 Bewusstheit sowie Wort-Lesen und orthographischen Fähigkeiten durch gemeinsame genetische Varianzanteile erklärt werden kann. Der genetische Anteil an dem Zusammenhang zwischen Wort-Lesen und phonologischer Bewusstheit beträgt demnach ca. 75% sowie ca. 69% an dem Zusammenhang zwischen WortLesen und orthographischen Fähigkeiten. (UJHEQLVVHPROHNXODUJHQHWLVFKHU8QWHUVXFKXQJHQ Bislang liegen für die Lese-Rechtschreib-Störung noch keine Ergebnisse aus systematischen Genom-Scans vor. Die bislang durchgeführten Untersuchungen postulieren Genloci auf den Chromosomen 6 und 15. Unter der Annahme eines autosomal dominanten Erbganges wurde erstmals 1983 eine Kopplung auf Chromosom 15 beschrieben (Smith et al. 1983). Dieser Befund konnte von einer dänischen Arbeitsgruppe nicht bestätigt werden (Bisgaard et al. 1987). Allerdings wurde diese Studie hinsichtlich des methodischen Vorgehens kritisiert, da die Phänotypbestimmung ausschließlich auf anamnestischen Angaben beruhte. In Erweiterung ihrer Stichprobe von 1983 fanden SMITH et al. (1991) erste Hinweise für genetische Heterogenität. Neben der Kopplung auf Chromosom 15, die jetzt etwas weiter distal (15q15) stehend als zuvor (15cen) beobachtet wurde, fand sich eine Kopplung auf Chromosom 6 (6p21.3) (Erklärung: 15q15 bedeutet: in Position 15 (rechts stehende Zahl) auf dem langen Arm (q) des Chromosoms 15 (links stehende Zahl); 15cen bedeutet: auf dem Centromer des Chromosoms 15, 6p21.3 bedeutet: in Position 21.3 auf dem kurzen Arm (p) des Chromosoms 6). Der Genlocus auf Chromosom 6 wurde durch die Arbeit von CARDON et al. (1994) bestätigt. In 2 unabhängigen Stichproben fanden die Autoren Kopplung, wobei in dieser Studie der Phänotyp der Leseschwäche als quantitatives Merkmal aufgefasst wurde. 5 Legasthenie 76 Abb. 11 Schematische Zeichnung der zwei Spalthälften oder Chromatiden eines Chromosoms (nach Singer u. Berg 1992). GRIGORENKO et al. (1997) untersuchten an ausgedehnten Familien Kopplungen zu fünf verschiedenen Phänotypen, die ein breites Spektrum der Lesefertigkeiten abdecken. Auf Chromosom 6p21.3 (Erklärung: 6p21.3 bedeutet: in Position 21.3 (rechts stehende Zahl) auf dem kurzen Arm (p) des Chromosoms 6 (links stehende Zahl)) zeigte sich eine signifikante Kopplung mit phonologischer Bewusstheit, auf Chromosom 15q21 (Erklärung: 15q21 bedeutet: in Position 21 (rechts stehende Zahl) auf dem langen Arm (q) des Chromosoms 15 (links stehende Zahl)) mit dem Wort-Lesen. In einer Untersuchung deutscher Familien konnte für die Bereitschaft, eine Legasthenie zu entwickeln, ebenfalls eine Kopplung auf Chromosom 15q21 (Erklärung: 15q21 bedeutet: in Position 21 (rechts stehende Zahl) auf dem langen Arm (q) des Chromosoms 15 (links stehende Zahl)) nachgewiesen werden (Schulte-Körne et al. 1998c). Auf Chromosom 6 hingegen fand sich mit diesem Phänotyp kein Hinweis auf Kopplung. 5 Legasthenie 77 Es wird angenommen, dass der Locus auf Chromosom 6 relevant für phonologische Prozesse ist, die wiederum eine hohe Korrelation mit der Lesefähigkeit aufweisen. Der Locus auf Chromosom 15 ist hingegen möglicherweise für die nichtphonologischen Anteile an der Lesefähigkeit relevant, z.B. für visuelle Prozesse (orthographische Fähigkeiten). Derzeit sind von mehreren internationalen Gruppen (World Health Organization, International Association of Genetic Technologists) systematische Kopplungsuntersuchungen des gesamten Genoms in Arbeit bzw. in Planung. Diese Untersuchungen werden mit großer Wahrscheinlichkeit Hinweise auf weitere Genloci bei der LRS ergeben. Die Identifizierung der verantwortlichen Gene wird ein Verstehen der zugrundeliegenden neurobiologischen Zusammenhänge und ihre Beziehung zu basalen Prozessen der phonologischen und visuellen Informationsverarbeitung ermöglichen. $XGLWLYH9HUDUEHLWXQJVVW|UXQJHQ Es wird angenommen, dass die auditive Verarbeitung von Sprache auf verschiedenen Ebenen geschieht (Gerd Schulte-Körne et al. http://www.kjp.unimarburg.de/kjp.legast/index.htm 18.6.2001). Das unterste Verarbeitungsniveau ist gekennzeichnet durch die Wahrnehmung von nichtsprachlichen Reizen. Die hierauf folgende Ebene ist bestimmt durch die Fähigkeit zur Sprachwahrnehmung. Das nächsthöhere phonologischen und komplexere Bewusstheit. Verarbeitungsniveau Phonologische Bewusstheit ist das der bezeichnet die Fähigkeit, die lautlichen Strukturen der geschriebenen Sprache zu erkennen und mit den lautlichen Strukturen zu operieren. 5 Legasthenie 78 Auf die Bedeutung der phonologischen Bewusstheit für den Schriftspracherwerb bei Vorschul- und Grundschulkindern weisen mehrere Untersuchungen hin. Zu den Verfahren zur Überprüfung von phonologischer Bewusstheit zählen z.B. Laut-zu-Wort-Zuordnung (Ist ein /s/ in Sonne?), Wort-zu-Wort-Zuordnung (Beginnen „ Maus“ und „ Mond“ gleich?) oder Reime erkennen (Reimen sich „ Haus“ und „ Maus“ ?) (Yopp 1988). Für den Lese- und Leselernprozess stellt die phonologische Bewusstheit offenbar eine entscheidende Voraussetzung dar. FRITH (1985) bezeichnet in ihrem Stufenmodell zur Entwicklung von Lesefähigkeit eine Stufe als alphabetische oder phonologische Entwicklungsstufe, die durch die Aneignung von BuchstabenLaut-Kenntnissen gekennzeichnet ist. Auf dieser Entwicklungsstufe gelingt es dem Leser erstmals, nicht nur anhand des visuellen Erkennens von Buchstaben (von FRITH als logographische Stufe bezeichnet) und Buchstabenkombinationen Wörter zu lesen, sondern über die lautliche Zuordnung die Aussprache von Wörtern zu erkennen und damit die Wörter selbst wiederzuerkennen. Die Bedeutung der phonologischen Bewusstheit für den Schriftspracherwerb wurde in einer Reihe von Längsschnittsstudien belegt. Die phonologische Bewusstheit, überprüft in der Kindergartenzeit, war ein bedeutsamer Prädiktor für die Lese- und Rechtschreibentwicklung während der Grundschulzeit. Dieses Ergebnis wurde in mehreren Studien repliziert und in mehreren Sprachen nachgewiesen (Englisch: Bryant et al. 1989, Wagner et al. 1994, Französisch: Algeria et al. 1982, Italienisch: Cossu et al. 1988, Schwedisch: Lundberg et al. 1980, Dänisch: Lundberg et al. 1988). Für die deutsche Schriftsprache zeigten KLICPERA und GASTEIGER-KLICPERA (1994), LANDERL und WIMMER (1994), NÄSLUND (1990) und MARX et al. (1993) anhand von Längsschnittstudien, dass Laut-Erkennen, Laut-Ersetzen und Reim-Erkennen, gemessen zur Zeit des Kindergartenbesuchs, prädiktiv für die Lese- und Rechtschreibleistung von Grundschulkindern in der zweiten und dritten Klasse war (Landerl und Wimmer 1994, Marx et al. 1993), und dass leseschwache Dritt-, 5 Legasthenie 79 Viert- und Achtklässler im Vergleich zu durchschnittlich lesenden Kindern der gleichen Klassenstufe bedeutsame Schwächen im Bereich der phonologischen Bewusstheit aufwiesen (Klicpera und Gasteiger-Klicpera, 1994). Im vorangehenden Abschnitt wurde die Bedeutung von phonologischer Bewusstheit für die Lese-Rechtschreib-Schwäche dargestellt. Unter der Annahme, dass die Wahrnehmung von Einzelsegmenten der Sprache ebenfalls eine Voraussetzung für den erfolgreichen Schriftspracherwerb ist, ist zu fragen, ob Lese-Rechtschreib-Schwache auch Schwächen in der Lautwahrnehmung und unterscheidung aufweisen, d.h. ob Schwächen in der Verarbeitung von Schallfolgen, die durch einen raschen Wechsel unterschiedlicher akustischer Spektren gekennzeichnet sind, bestünden. Anhand von einfachen Lautunterscheidungstests konnte in einer Reihe von Untersuchungen diese Annahme bestätigt werden (Godfrey et al., 1981, Werker und Tees, 1987, Manis et al., 1997). Lese-Rechtschreib-Gestörte hatten Schwächen in der Identifizierung und Unterscheidung von Lauten, die sich im Wesentlichen durch die „ voiceonset-time“ (z.B. /ba/ und /pa/) unterschieden. Die Formanten eines Lauts sind diejenigen Frequenzbereiche des Anregungssignals (hervorgerufen durch die Schwingung der Stimmlippen oder durch eine enge Konstriktion im Mundraum), die abhängig von der Stellung der Artikulatoren vom Ansatzrohr verstärkt werden. Spektrogramme mit hervorgehobenen Formantübergängen für die Silben /ba/, /da/ und /ga/ zeigen Unterschiede bei den F2- und F3-Transitionen, aber eine einheitliche F1Transition. Lese-Rechtschreib-Schwache hatten Schwierigkeiten in der Diskrimination dieser Laute, die sich durch die Transition des zweiten und dritten Formanten voneinander unterschieden. Diese Schwäche zeigte sich nicht nur bei Schulkindern, sondern auch bei Erwachsenen (Steffens et al. 1992). 5 Legasthenie 80 'LH%HGHXWXQJGHU:DKUQHKPXQJYRQQLFKWVSUDFKOLFKHQ5HL]HQ Die bereits dargestellten Befunde zeigen, dass Lese-Rechtschreib-Schwache Störungen im Bereich der Sprachwahrnehmung und -verarbeitung aufweisen. In verschiedenen Untersuchungen konnte zudem nachgewiesen werden, dass ein auditives Informationsverarbeitungsdefizit häufig bereits auf nichtsprachlicher Ebene vorliegt. Da die Wahrnehmung nichtsprachlicher Reize Voraussetzung für die Sprachverarbeitung in höheren Ebenen ist, kommt dieser Störung mit hoher Wahrscheinlichkeit eine große Bedeutung bei der Entstehung der Legasthenie zu. Eine Untersuchung von SCHULTE-KÖRNE et al. (1998a) zeigte, dass LeseRechtschreib-Schwache keine Schwächen in der Wahrnehmung von Sinustönen hatten, wenn die Töne mit einem Interstimulusintervall (ISI) von 590 ms präsentiert wurden. Untersuchungen von TALLAL et al. (1993) unterstützen jedoch die Hypothese, dass Leseschwache dann Schwächen in der Wahrnehmung von nichtsprachlichen Reizen aufweisen, wenn diese in einem sehr kurzen zeitlichen Abstand nacheinander präsentiert werden. Von Paula TALLAL wurde der Begriff des 6SHFLILF/DQJXDJH,PSDLUPHQW (SLI) geprägt, um eine nicht altersgemäße Entwicklung der Sprechfähigkeit bei ansonsten normalen kognitiven Fähigkeiten zu beschreiben (Tallal u. Piercy 1973, Tallal 1993). Der Ausdruck SLI („ spezifische Beeinträchtigung der Sprachentwicklung“ ) betont, dass die Kinder zwar normal intelligent, jedoch sprachlich zurückgeblieben sind. Die Sprachverzögerung beträgt mindestens ein Jahr, der Handlungs-IQ liegt im Durchschnittsbereich. Ausgeschlossen werden insbesondere alle Kinder, die unter einem sensorischen Hörverlust, genereller mentaler Retardierung, Lähmungen oder Sensibilitätsverlusten der Mundmuskulatur oder anderen neurologischen oder psychiatrischen Störungen leiden. 5 Legasthenie 81 TALLALs Studien begannen 1970. Ihr Ziel war es, die schweren Ausfälle der Wahrnehmung und Produktion von Sprache zu verstehen, die kennzeichnend für die meisten SLI-Kinder sind. TALLAL et al. (1993) bemerkten, dass es vor dem Studium des Sprechvermögens per se vernünftig sein würde, die Unversehrtheit der einzelnen akustischen Prozesse zu beurteilen, die das komplexe akustische Spektrum von Sprache bestimmen. So ist es sicherlich wichtig festzustellen, dass ein Kind über ein normales Hörvermögen verfügt, bevor Defizite in seiner Fähigkeit, Sprache zu verarbeiten oder zu produzieren, interpretiert werden können. Mit diesen Voraussetzungen begannen TALLAL et al. (1993), eine Testreihe zu entwickeln, um die Wahrnehmung, Unterscheidungsfähigkeit, Erinnerungsvermögen zeitliche Reihenfolge, für die Integration, Assoziation, Geschwindigkeit Reihenfolge und das von akustischen Ereignissen festzustellen. TALLAL verglich leseschwache Kinder mit nicht-leseschwachen Kindern anhand von Aufgaben zum Erkennen der Reihenfolge von zwei Tönen bzw. zwei komplexen Tonkombinationen (Tallal 1980). Die Reihenfolge der Reize war entweder 1-1, 1-2, 2-1 oder 2-2. Die Kinder mussten zur erfolgreichen Bewältigung der Aufgaben anhand einer ausgedehnten Übungsphase die Zuordnung der vier Lösungsmöglichkeiten zu den entsprechend farbig markierten Tasten erlernen. Ein Gruppenunterschied zeigte sich erst, wenn das Interstimulusintervall zwischen zwei Reizen geringer als 305 msec war. Dieses Ergebnis legt die Vermutung nahe, dass sich bei sehr kurzem Interstimulusintervall bereits auf nichtsprachlicher Ebene eine Schwäche zeigt, die möglicherweise im direkten Zusammenhang mit der Sprachwahrnehmung steht (Tallal et al. 1993). 5 Legasthenie 82 Weitere Untersuchungen von TALLAL et al. (1993) bestätigten die Annahme, dass grundlegende sprachgestörten zentralnervöse Kindern auf deren Verarbeitungsstörungen Unfähigkeit beruhen, auch bei sensorische Informationen zu integrieren, die in schneller Folge im Zentralnervensystem ankommen. Offenbar besteht ein enger Zusammenhang zwischen Dysphasie und Legasthenie (Tallal et al. 1993). So konnten Längsschnittstudien zeigen, dass Kinder, deren Sprachentwicklung verzögert ist, ein hohes Risiko haben, eine Legasthenie zu entwickeln. Zudem sind sich die neuropsychologischen Profile von Kindern mit einer verzögerten Sprachentwicklung und von Kindern mit einer Leseschwäche sehr ähnlich. Zeitabhängige Mechanismen im Nervensystem spielen eine zentrale Rolle in den grundlegenden Prozessen der Informationsverarbeitung und -produktion und sind von großer Bedeutung für die normale Entwicklung und Erhaltung des phonologischen Systems. Dass zeitabhängige Mechanismen eine wichtige Rolle bei der Entwicklung des sprachverarbeitenden Systems spielen, folgt auch aus den Untersuchungsergebnissen von Phillips. Er stellte fest, dass es hörgestörten Kindern schwer fällt, eine Frequenzlücke zu erkennen und sie einzuordnen (s. 4.2.2.1). Die Störung kann zur primären Worttaubheit führen: Der Fluss der Sprache wird von Zeitabschnitten unterbrochen, in denen die Klänge von sehr niedriger Amplitude sind oder sogar von vollständiger Stille. Die wichtigsten „ Frequenzlücken“ der Sprache sind die phonetisch bedeutsamen - sie lassen Stoppkonsonanten erkennbar werden. Wie bei TALLAL ergaben auch die Forschungsergebnisse von PHILLIPS, dass sprachunspezifische Störungen eine wichtige Rolle spielen. So zeigten seine Untersuchungen, dass der primären Worttaubheit eine Funktionsbeeinträchtigung im primären auditorischen Kortex zugrunde liegt (s. 4.2.2.1). TALLAL et al. (1993) nehmen an, dass die Verarbeitungsstörung eine Kaskade von Folgen auslöst. Das Defizit betrifft somit mehrere sensorische Modalitäten und auch das motorische Nervensystem. Im weiteren Verlauf ist die Entwicklung 5 Legasthenie 83 eines normalen phonologischen Systems unmöglich, insbesondere normal zu sprechen und zu lesen. Die Störung im akustischen Sinneskanal führt dazu, dass zu wenige für die Sprachentwicklung wichtige Lautreize aus der Umwelt das Gehirn erreichen. So verbinden sich die dortigen Nervenzellen nur unzureichend zu Einheiten, die bestimmten Lauten zugeordnet sind. Die verringerte Fähigkeit, die raschen Schwankungen der Tonhöhe zu identifizieren, geht mit Schwierigkeiten beim Aussprechen mancher Wörter einher. Das bedeutet, dass sowohl Sprach- als auch Leseprobleme ihren Ursprung in einer ungenügend entwickelten Fähigkeit der Verarbeitung und Decodierung schneller Impulse haben. TALLAL et al. (1993) stellten fest, dass die Versuchsergebnisse der sprach- und lesegestörten Kinder in bemerkenswerter Weise übereinstimmten. Längsschnitterhebungen zeigten, dass die Mehrheit der Kinder, die bereits im Kindergartenalter eine verzögerte Sprachentwicklung zeigten, überdurchschnittliche Schwierigkeiten beim Lesenlernen entwickelte (Tallal et al. 1988). Die neurologische Ursache der unzureichenden Verarbeitung und Decodierung schneller Impulse ist bislang unbekannt. MERZENICH et al. (1993) nehmen an, dass ein Mangel an Erfahrung in der frühen Entwicklung dem Nervensystem so schaden könne, dass es ihm unmöglich sei, schnelle Informationseingaben zu verarbeiten. Da das Gehirn gewährleisten müsse, dass es sich in Einklang mit der äußeren Welt befinde, passe sich die Verarbeitung aller Sinnesmodalitäten der langsamsten an. Aus ihren Untersuchungen schließen MERZENICH et al. (1993), dass Defizite in der zeitlichen Verarbeitung von Sprache erlernt sein können. Merkmale von Augenbewegungen beim Lesen sowie Studien, die eine Verbesserung beim Lesen 5 Legasthenie 84 ergaben, wenn das periphere Gesichtsfeld verdeckt wurde, stimmen mit der Hypothese überein, dass leseschwache Kinder einem größeren Bereich ihre Aufmerksamkeit schenken als dies normalerweise der Fall ist. Zudem sind MERZENICH et al. (1993) davon überzeugt, dass sich neuronale Veränderungen während eines fehlerhaften Lernvorgangs bilden. Hierbei spielt ein Mangel an Erfahrungen, bedingt durch unzureichende Stimulierung, eine wichtige Rolle. Diese unzureichende Reizexposition beginnt bereits im Mutterleib und ist dort von besonderer Bedeutung, da der den Lernvorgängen zu Verfügung stehende Axonenüberschuss bereits nach der Geburt abnimmt. Die Entstehung kohärenter Stimulusrepräsentationen während des Lernens und die vermutliche Plastizität grundlegender „ Uhrzeiten“ in der Hirnrinde während der kindlichen Entwicklung und des weiteren Lebens sind abhängig von grundlegenden Mechanismen der Synapsenverstärkung. Die Liste molekularer Elemente, die von der Modifikation synaptischer Wirksamkeit während des Lernvorgangs abhängt, ist lang. Ein Defekt in irgend einem von wenigstens mehreren hundert Proteinen kann zu einer Verzögerung des Lernvorgangs führen. Es konnte gezeigt werden, dass Kinder mit Legasthenie von normaler Intelligenz sind, wenn sie in vom Lesen unabhängigen Tests untersucht werden. Das lässt vermuten, dass Lesen auf der Fähigkeit eines nicht oder kaum assoziativen Systems beruht. Offenbar beruht die neuronale Veränderung, die zur Legasthenie führt, selektiv auf plastischen Veränderungen, die Eigenschaften der zeitlichen Verarbeitung von Eingaben kontrollieren (Tallal et al. 1993). LLINAS (1993) stellt eine andere Hypothese vor. Er nimmt an, dass eine besondere innere „ Uhr“ , die die Rate der neuronalen Entladungen kontrolliert, bei Legasthenikern gestört sei. LLINAS erklärt, dass die Frequenz der neuronalen Entladungen im Kortex bei normalen Menschen im 40-Hz-Bereich liege. Es werde angenommen, dass diese neuronalen Oszillationen wichtige Komponenten seien, um sensorische Informationen in kortiko-thalamo-kortikale Netzwerke hineinzulassen. 5 Legasthenie 85 Aufgrund einer zellulären Dysfunktion noch unbekannter Genese sind die normalen Eigenschaften neuronaler Schaltkreise, die verantwortlich sind für die zeitlichen Aspekte des Denkens, bei Legasthenie in einem bestimmten Maße verändert. LLINAS (1993) nimmt an, dass diese Veränderung dazu führt, dass das Nervensystem nur innerhalb bestimmter zeitlicher Rahmen normal funktioniert. Er glaubt, dass die Regelwidrigkeiten zwei wichtige Folgen haben: Zum einen ist es den Neuronen nicht mehr möglich, eine ausreichende Anzahl von Oszillationen mit höheren Frequenzen (35-45 Hz) zu erzeugen, zum anderen solche Rhythmen im Anschluss an sensorische Stimulationen mit kurzen zeitlichen Abständen neu einzustellen. LLINAS (1993) stellt fest, dass seine Hypothese sehr gut erklären könnte, warum Legasthenikern eine geeignete Triggerung oder Neueinstellung von Frequenzen im 40-Hz- bzw. 10-Hz-Bereich fehlt. Dies wird von anderen Wissenschaftlern bestätigt. Wie PÖPPEL (1997) feststellte, bildet das Gehirn innere Zustände im 40-Hz-Bereich, die für die Wahrnehmung von zentraler Bedeutung sind. Dabei spielen zwei grundlegende Zeitgrenzen eine wichtige Rolle: eine nach unten im Bereich von 30 bis 40 Millisekunden und eine nach oben von 3 Millisekunden. Die 30 Millisekunden stellen dabei die untere Ordnungsschwelle dar, bis zu der zwei verschiedene Reize sicher als nicht gleichzeitig und demnach als verschieden identifiziert werden können. Dieses untere Zeitfenster beruht auf der Synchronisation der Neuronen, die durch die im 40-Hz-Bereich oszillierende Kreisschleife Thalamus-Kortex vorgegeben wird. Hierdurch schafft das Gehirn Systemzustände, die quasi zeittote Zonen darstellen. Innerhalb dieser 30 Millisekunden existiert für die menschliche Wahrnehmung kein Fluss der Zeit im Newtonschen Sinn, kein Vorher und kein Nachher. Damit gibt sich das Gehirn neutrale präsemantische Arbeitsplattformen vor, in die sich dann alle Hirnaktivitäten einordnen. Die Wahrnehmung unterliegt somit durch die 30-Millisekunden-Zeitfenster einer präsemantischen automatischen Integration, die die einlangenden Impulse bündelt und strukturiert. Dieser Integrationsmechanismus sorgt dafür, dass Ereignisse zu überschaubaren Erlebnissen zusammengefasst werden, die als Jetzt erlebt werden. Die obere 5 Legasthenie 86 Schwelle von 3 Millisekunden ergibt sich aus der Beobachtung, dass zwei sensorische Reize nur innerhalb dieses Zeitraums exakt miteinander verglichen werden können, dass bezüglich der Zeitwahrnehmung nur Intervalle von 2 Millisekunden genau reproduziert werden können, während darüber die Schätzung einsetzt, dass auch eine sensomotorische Aktion nur für 2 Millisekunden präzise antizipiert werden kann. Auch erstreckt sich die Konzentration jeweils nur über ein Intervall von 2 bis 3 Millisekunden, nach dem das Gehirn anfragt, ob es etwas Neues gibt bzw. ob sich das Verweilen bei dem bisherigen Gegenstand noch lohnt (Pöppel 1997). Zwar gelang bisher noch kein Nachweis, jedoch ist anzunehmen, dass eine entwicklungsbedingte Veränderung des ZNS die inneren Oszillationsraten verlangsamen kann. Eine der wichtigsten Folgen wäre, dass es unmöglich sei, sensorische oder motorische Informationen zu verarbeiten, die in schneller Abfolge, d.h. im Zehn-Millisekunden-Bereich, ablaufen würden. Dies ist genau das Defizit, das sprach- oder lesegestörte Kinder betrifft (Llinas 1993). Die Annahme gestörter kortiko-thalamo-kortikaler Netzwerke wird bestärkt durch magnetresonanztomographische Untersuchungen. Jernigan et al. (1991) berichten über eine signifikante Reduktion der grauen Substanz in subkortikalen Strukturen (einschließlich des Corpus striatum und des Thalamus) sowie in kortikalen Strukturen, die bekannt sind, der Sprachverarbeitung zu dienen. TALLAL et al. (1993) weisen darauf hin, dass sich die Frage stellt, warum Kinder mit solch schweren Störungen der zeitlichen Verarbeitung von Sinnesreizen relativ normal leistungsfähig sind. Der zeitliche Rahmen der Störung liegt im Zehn-Millisekunden-Bereich, und es muss angenommen werden, dass nur Verarbeitungsprozesse, die in diesem zeitlichen Rahmen ablaufen, beeinträchtigt sind (Phillips 1993). 5 Legasthenie 87 Obgleich es wahr ist, dass Buchstaben auf einer Seite für den Leser unbeweglich wirken, und zwar für jeden möglichen Zeitraum, ist die visuelle Identifizierung von Graphemen nur dann möglich, wenn die visuelle Darstellung mit der begrenzt dauernden neuronalen Darstellung des passenden Phonems assoziiert werden kann. TALLAL et al. (1993) nehmen an, dass sprach- und lesegestörte Kinder – entsprechend ihrer grundlegenden Störung der auditiven Verarbeitung – keine stabilen und unbeweglichen neuronalen Darstellungen von Phonemen bilden können. Wie LIBERMANN (1985) annimmt, gelingt es diesen Kindern niemals, eine phonologische Bewusstheit zu erreichen. Zahlreiche Studien unterstützen diese Annahme (Goswani 1993). Diese Daten bestärken die Vermutung, dass Störungen der zeitlichen Verarbeitung im Hörsystem ausreichen, um eine Entwicklung des Lesenlernens zu unterbrechen. Obwohl die zeitlichen Verarbeitungsstörungen alle Sinnesmodalitäten betreffen, besteht somit die Möglichkeit, dass Störungen in anderen Modalitäten bei manchen Kindern das Lesen nicht direkt beeinflussen könnten (Tallal et al. 1993). 1HXURDQDWRPLVFKH9HUlQGHUXQJHQ Ergebnisse von MRI- und PET-Untersuchungen sowie Verhaltensstudien bei gesunden Erwachsenen und Erwachsenen mit erworbenen Läsionen weisen daraufhin, dass die Sprachverarbeitung in der linken Hemisphäre lokalisiert ist (Tallal et al. 1993). Diese Spezialisierung für die Verarbeitung sensorischer und motorischer Ereignisse, die in schneller Folge auftreten, könnte sich während der Evolution ausgebildet haben. TALLAL et al. (1991) beobachteten unterschiedliche Muster der zerebralen Lateralisation in Gehirnen von SLI-Kindern – verglichen mit denen von gesunden, und zwar sowohl im Präfrontalkortex als auch in der parietalen Region. Es fand sich eine hochsignifikante Korrelation zwischen dem Ausmaß der hemisphärischen Asymmetrie dieser kortikalen Bereiche und dem Ausmaß des 5 Legasthenie 88 Defizits bezüglich der zeitlichen Verarbeitung akustischer Ereignisse, das in den Tests von TALLAL et al. (1991) festgestellt wurde. In mehreren Untersuchungen fand eine Arbeitsgruppe um GALABURDA bei Patienten mit Lese-Rechtschreib-Störungen Veränderungen der linksseitigen temporalen Hirnrinde (Galaburda et al. 1994). Primäres und sekundäres Hörzentrum sowie das rezeptive Sprachzentrum sind in dieser Hirnregion lokalisiert (vgl. 3.2 u. 3.3.6). Bei LRS-Kindern ist die physiologische Asymmetrie zugunsten der linken Hirnhälfte aufgehoben, und wiederholt wurden Abnormitäten der dort gelegenen Hirnwindungen belegt. In neuerer Zeit wurden die Befunde durch magnetresonanztomographische Untersuchungen bestätigt. ROSEN (1992) und FITSCH (1994) erzeugten im Tierversuch Hirnabnormitäten, die denen von LRS-Patienten entsprechen. Die Tiere zeigten im Erwachsenenalter Defizite bei der zeitlichen Auflösung auditiver Reize. 1997 berichteten die Autoren über unterschiedliche Ausfälle bei männlichen und weiblichen Tieren. Nur bei männlichen Tieren wurden auditive Diskriminationsschwächen und eine sekundäre Verminderung der Zahl von Riesenzellen in funktionell abhängigen basalen Kernen (Corpus geniculatum mediale) gefunden. ROSEN und FITSCH sind überzeugt, dass sich aus derartigen Befunden Schlussfolgerungen für Störungen der Laut- und Schriftsprache beim Menschen ziehen lassen können. 9LVXHOOH9HUDUEHLWXQJVVW|UXQJHQ Störungen der Verarbeitung visueller Informationen bei LRS wurden im Bereich der Okulomotorik und bei der Verarbeitung von Reizen, die im Wesentlichen vom magnozellulären System verarbeitet werden, beschrieben. 5 Legasthenie 89 Zusammenhänge von Lese-Rechtschreib-Schwäche mit der Blickmotorik waren schon mehrfach vermutet worden, Studien ergaben jedoch zunächst keine deutlichen Auffälligkeiten. Erst die Untersuchung der Anti-Sakkaden-Aufgabe offenbarte ein häufig auftretendes Defizit: die willentliche Komponente, die in den früheren einfacheren Tests nicht gemessen worden war. Kinder mit einer Lese- und Rechtschreibschwäche zeigen häufig Regressionssakkaden, d.h. Sakkaden entgegengesetzt zur normalen Leserichtung (Fischer u. Weber 1990, Kotchen u. Guthrie 1980, Stark et al. 1991). Störungen in der Kontrolle der Blickmotorik zeigten sich außerdem in einem Rückstand der • Entwicklung der Fixationsfähigkeit, d.h. mehr reflexartige Blicksprünge (Eden et al. 1994), • willentlichen Kontrolle über die Durchführung der Blicksprünge (Biscaldi et al. 1998), • zeitlichen Genauigkeit der Reaktionszeiten (Stark et al. 1991), • Treffsicherheit (Stark et al. 1991). Da eine Abhängigkeit der Entwicklung der Okulomotorik beim Lesen von der Entwicklung der Lesefähigkeiten besteht (Olson et al. 1991), ist allerdings zur Zeit noch unklar, inwieweit die beschriebenen Auffälligkeiten kausal mit der Lesestörung in Zusammenhang stehen oder nur ein Epiphänomen darstellen. 0DJQR]HOOXOlUHV6\VWHP Die Legasthenie steht offenbar in Verbindung mit Normabweichungen neuroanatomischer und hirnfunktioneller Merkmale. Besondere Bedeutung kommt möglicherweise einer Nervenbahn zu, die von der Netzhaut stammende 5 Legasthenie 90 Informationen zum Thalamus und weiter zum visuellen Kortex leitet. Es gibt Hinweise dafür, dass diese von Magnozellen gebildete Bahn bei Legasthenikern weniger leistungsfähig ist (Tallal 2000). Die Magnozellen sind für die Vorverarbeitung schnell hintereinander eintreffender Informationen verantwortlich. Bereits in den Achtzigerjahren hatte TALLAL festgestellt, dass viele Legastheniker ungefähr 0,3 Sekunden brauchen um Laute wie /da/ und /ba/ voneinander zu unterscheiden – deutlich länger als die 0,008 Sekunden, die Sechs- bis Achtjährige normalerweise für diese Aufgabe benötigen (TALLAL 1980). Kann man die Klänge einer Sprache nicht unterscheiden, führt dies auch zu Schwierigkeiten beim Erlernen der Grammatik, des Lesens und des Schreibens (Tallal 2000). Im Vordergrund der aktuellen Forschung zu den Ursachen der LRS im Bereich der visuellen Verarbeitung stehen die Funktionen des magno- und des parvozellulären Systems. Das magno- (M) und das parvozelluläre (P) System haben ihren Ursprung in retinalen Ganglionzellen (vgl. 3.4.3). Im Bereich der Fovea finden sich überwiegend Zellen des P-Systems, hingegen sind die Zellen des M-Systems überwiegend peripher zu finden. Das M- und das P-System projizieren in getrennte Schichten des Corpus geniculatum laterale und weiter in den primären visuellen Kortex. Der magnozelluläre Anteil des primären visuellen Kortex (V1) projiziiert über die Area V5 überwiegend in den parietalen Kortex, hingegen erreichen den temporalen Kortex sowohl parvo- als auch magnozelluläre Bahnen (Merigan und Maunsell 1993). Eine Arbeitsgruppe um GALABURDA fand in histologischen Untersuchungen des Corpus geniculatum laterale (vgl. 3.4.4) bei fünf LRS-Patienten eine Verminderung der Zahl der Nervenzellen in der Magnozellschicht (Galaburda u. Livingston 1993). Dies könnte in Zusammenhang mit einer visuellen Wahrnehmungsschwäche stehen: Im Corpus geniculatum laterale wird die visuelle Information umgeschaltet und zur optischen Hirnrinde weitergeleitet. 5 Legasthenie 91 Die Magnozellen des in der Tiefe des Gehirns gelegenen Kerns sind für die Vorverarbeitung von Informationen, die schnell hintereinander eintreffen und nur einen geringen Kontrast aufweisen, verantwortlich. Ebenso werden dem MSystem Funktionen zur globalen Analyse von visueller Information zugeordnet (Merigan u. Maunsell 1993). In der Schicht der Parvozellen hingegen werden langsam wechselnde visuelle Reize mit hohem Kontrast verarbeitet. Das P-System mit hoher Orts- und Farbauflösung verarbeitet stationäre bzw. langsame Reize und verarbeitet aufgrund seiner hohen Ortsauflösung im Wesentlichen lokale Aspekte bzw. Detailaspekte von visuellen Reizen (Breitmeyer u. Glanz 1976). GALABURDA und LIVINGSTONE (1993) fanden bei Legasthenikern Unterschiede in der Struktur und Funktion der thalamokortikalen magnozellulären Systeme, aber intakte parvozelluläre Systeme. Es zeigten sich signifikante zelluläre Unterschiede auf der Ebene des Thalamus, und zwar nicht nur im Corpus geniculatum laterale, das visuelle Informationen weiterleitet, sondern auch im Corpus geniculatum mediale, das auditive Informationen zum Kortex überträgt. Dies sind offenbar wichtige empirische Daten, die auf eine mögliche direkte Verbindung zwischen anatomischen, physiologischen und Verhaltensdaten hinweisen. Auch STEIN und TALCOTT (1999) sind der Überzeugung, dass bei LeseRechtschreib-Schwäche hauptsächlich die Magnozellen betroffen sind. STEIN und TALCOTT stellten die Hypothese auf, dass bei Legasthenikern – möglicherweise infolge einer Immunreaktion der Mutter – während der fetalen Entwicklung junge Nervenzellen im Gehirn geschädigt würden. In Folge sei ihr Zusammenwachsen behindert und die Ausbildung der M-Bahn von der Netzhaut zum lateralen Corpus geniculatum und weiter zum visuellen Kortex gestört (Stein u. Talcott 1999). 5 Legasthenie 92 'LH%HGHXWXQJJUXQGOHJHQGHUYLVXHOOHU:DKUQHKPXQJVSUR]HVVH Eine Forschergruppe um die Physiologen TALCOTT und STEIN (1999) wies nach, dass bei Legasthenikern grundlegende visuelle Wahrnehmungsleistungen beeinträchtigt sein können. In den Untersuchungen von TALCOTT und STEIN gelang es Legasthenikern erst ab einer höheren Anzahl von sich bewegenden Punkten auf einem Computerbildschirm zu erkennen, dass diese plötzlich in gleiche Richtung bewegt werden. Je mehr Schwierigkeiten sie bei dieser Aufgabe hatten, desto stärker war auch ihre Leseschwäche ausgeprägt. Dieser Befund macht es sehr wahrscheinlich, dass die Lesefähigkeit auf der Fertigkeit beruht, rasche Veränderungen elementarer Sinneseindrücke zu registrieren. Die Legasthenie hängt somit mit einer Schwäche beim Verarbeiten optischer Eindrücke im Gehirn zusammen. Dabei besteht offenbar ein Zusammenhang mit Defiziten bei akustischen Sinnesleistungen. So hatten Kinder, die bei der Untersuchung TALCOTTs et al. ein schlechtes Testergebnis zeigten, auch Schwierigkeiten, zwischen richtig und falsch buchstabierten Wörtern zu unterscheiden. Je deutlicher dieses doppelte Defizit ausgeprägt ist, desto schwerer war die Leseschwäche (Talcott et al. 2000). TALCOTT et al. (2000) heben hervor, dass Lesenlernen darauf beruht, sowohl die gesprochenen (phonologischen) als auch die geschriebenen (orthographischen) Eigenschaften einer Sprache zu verstehen. Bereits früher durchgeführte Untersuchungen bestätigen die Ergebnisse von TALCOTT et al. (2000). So wiesen MANIS et al. (1997) nach, dass 7 von 25 Legasthenikern Schwächen bei der nichtsprachlichen Wahrnehmung von Sprache hatten, aber nur 1 Kind in der Vergleichsgruppe mit altersgerechter Leseentwicklung. MANIS et al. schließen daraus, dass manche Legastheniker ein nichtsprachliches Wahrnehmungsdefizit haben, das die Sprachverarbeitung hemmt und zur Legasthenie führt (Manis et al. 1997). 5 Legasthenie 93 Untersuchungen solcher Art lassen allerdings die wichtige Frage unbeantwortet, ob die Legasthenie tatsächlich ein eigenständiges pathologisches Phänomen ist. TALCOTT et al. (2000) haben mittlerweile deutliche Hinweise dafür gefunden, dass dies nicht zutrifft. Ihren Studien zufolge lässt sich die Legasthenie in ein Spektrum, das alle Lesefähigkeiten umfasst, einordnen (Talcott et al. 2000). Für ihre visuellen und akustischen Tests wählten TALCOTT et al. insgesamt 32 zehn Jahre alte Kinder aus, bei denen keine Anzeichen einer Leseschwäche zu beobachten waren. Die visuelle Aufgabe bestand wiederum darin, die Richtungsänderung bewegter Punkte auf einem Bildschirm zu erkennen. Bei dem akustischen Test sollten die Kinder angeben, welcher von zwei rasch hintereinander erzeugten Tönen eine Modulation aufwies. Solche raschen Änderungen der Tonhöhe müssen auch beim Dekodieren der gesprochenen Sprache registriert werden. Ähnlich wie bei Legasthenikern konnten die Forscher anhand der beiden ermittelten Sinnesleistungen auch bei den „ normalen" Lesern den Grad der Lesefertigkeit voraussagen. Die Kinder, die beim visuellen Leistungstest schlecht abschnitten, vermochten nur schwer zwischen richtig und falsch buchstabierten Wörtern zu unterscheiden. Eine verringerte Fähigkeit, die raschen Schwankungen der Tonhöhe zu identifizieren, ging dagegen mit Schwierigkeiten beim richtigen Aussprechen mancher Wörter einher. Diese Ergebnisse deuten daraufhin, dass Leseschwäche nur das ungünstige Ende einer normalen Fähigkeitsverteilung bildet (Tallal 2000). TALLAL und MERZENICH (Barinaga 1996) entwickelten ein ComputerTrainingsprogramm, das die Verarbeitung schneller akustischer und visueller Reize im Gehirn fördert. TALLAL und MERZENICH konnten zeigten, dass die Sprachübungen die Sprachentwicklung von Kindern, die mehrere Jahre hinter dem Durchschnitt lagen, gesprochene Sprache zu verstehen, deutlich fördern konnten (s. Kapitel 7). 5 Legasthenie 94 =XVDPPHQIDVVXQJXQG6FKOXVVIROJHUXQJHQ Hauptziel dieses Kapitels war es, neue Untersuchungsergebnisse zur Pathogenese der Legasthenie darzustellen. Legasthenie ist die Bezeichnung für Schwächen beim Erlernen von Lesen, Schreiben und Rechtschreiben, die weder auf eine allgemeine Beeinträchtigung der geistigen Entwicklung, noch auf unzulänglichen Unterricht zurückgeführt werden können. Typisch sind Verwechslungen von Buchstaben im Wort wie b-d, p-q, u-n, Umstellungen von Buchstaben innerhalb eines Wortes, Auslassungen und Einfügungen. Es ist möglich, dass die Bereitschaft zur Legasthenie hereditär ist. Auditive Verarbeitungsstörungen spielen bei der Entwicklung der Legasthenie eine wichtige Rolle. In verschiedenen Untersuchungen konnte nachgewiesen werden, dass ein auditives Informationsverarbeitungsdefizit bereits auf nichtsprachlicher Ebene vorliegt. Die Untersuchungen TALLALS bestätigten die Annahme, dass grundlegende zentralnervöse Verarbeitungsstörungen die Ursache einer Legasthenie sind. Es ist unmöglich, sensorische Informationen zu integrieren, die in schneller Folge im Zentralnervensystem ankommen (Tallal et al. 1993). Die Störung im akustischen Sinneskanal führt dazu, dass zu wenige für die Sprachentwicklung wichtige Lautreize aus der Umwelt das Gehirn erreichen. So verbinden sich die dortigen Nervenzellen nur unzureichend zu Einheiten, die bestimmten Lauten zugeordnet sind. Wie bei der Entstehung der primären Worttaubheit (Phillips 1999, s. Kapitel 4) spielen zeitabhängige Mechanismen offenbar auch bei der Entwicklung der Legasthenie eine wichtige Rolle. Es wird angenommen, dass eine besondere innere „ Uhr“ , die die Rate der neuronalen Entladungen kontrolliert, bei Legasthenikern gestört sei (Llinas 1993). Bei der Wahrnehmung spielen zwei 5 Legasthenie 95 grundlegende Zeitgrenzen eine wichtige Rolle: eine nach unten im Bereich von 30 bis 40 Millisekunden und eine nach oben von 3 Millisekunden. Das untere Zeitfenster beruht auf der Synchronisation der Neuronen, die durch die im 40-HzBereich oszillierende Kreisschleife Thalamus-Kortex vorgegeben wird. Die Wahrnehmung unterliegt somit durch die 30-Millisekunden-Zeitfenster einer präsemantischen automatischen Integration, die die einlangenden Impulse bündelt und strukturiert. Die obere Schwelle von 3 Millisekunden ergibt sich aus der Beobachtung, dass zwei sensorische Reize nur innerhalb dieses Zeitraums exakt miteinander verglichen werden können (Pöppel 1997). Es ist anzunehmen, dass bei sprach- und lesegestörten Kindern die inneren Oszillationsraten verlangsamt sind (Llinas 1993). Die Annahme gestörter kortiko-thalamo-kortikaler Netzwerke wird durch magnetresonanztomographische Untersuchungen bestärkt (Jernigan et al. 1991). Störungen der Verarbeitung visueller Informationen bei Legasthenie wurden im Bereich der Okulomotorik und bei der Verarbeitung von Reizen, die im Wesentlichen vom magnozellulären System verarbeitet werden, beschrieben. Die von Magnozellen gebildete Bahn leitet von der Netzhaut stammende Informationen zum Thalamus und weiter zum visuellen Kortex. Es gibt deutliche Hinweise dafür, dass diese Bahn, die der Vorverarbeitung schnell hintereinander eintreffender Informationen dient, bei Legasthenikern weniger leistungsfähig ist (Tallal 2000). Offenbar sind bei vielen Legasthenikern grundlegende visuelle Wahrnehmungsleistungen beeinträchtigt (Talcott und Stein 1999), wobei ein Zusammenhang mit Defiziten bei akustischen Sinnesleistungen besteht (Talcott 2000). Aus den Ergebnissen ergeben sich wichtige Schlussfolgerungen für die Behandlung einer Legasthenie: Es sollten Trainingsprogramme eingesetzt werden, die die Verarbeitung schneller akustischer und visueller Reize im Gehirn fördern. 6 Diskussion 96 'LVNXVVLRQ (QWVWHKXQJVEHGLQJXQJHQGHUUHLQHQ:RUWWDXEKHLW 'LH3RVWXODWHXQG6FKOXVVIROJHUXQJHQYRQ3+,//,36 Die Untersuchungen von PHILLIPS ergaben, dass die reine Worttaubheit auf einer grundlegenden Wahrnehmungsstörung beruht. Offenbar deutet schon die Organisation des auditiven Systems daraufhin, dass auf einer basalen Stufe der auditiven Wahrnehmung Sprachlaute erkannt und analysiert werden (Phillips 1995). So ist die Wahrnehmung von Sprachlauten eine Hierarchie von Prozessen, die mit der Erkennung des Lautes und seiner phonetischen Klassifizierung beginnt. Laute werden als Wort erkannt und diesen die aus dem semantischen Gedächtnis abgerufene Bedeutung zugeordnet (Phillips 1990). Es gibt mindestens vier Prinzipien der Organisation des auditiven Systems (Phillips 1995). Das erste ist die tonotope Organisation: Innerhalb jedes Kerngebiets der zentralen Hörbahn (und innerhalb der meisten auditiven Rindenbezirke) sind die Neuronen schichtweise so angeordnet, dass die stärksten Erregungen von Neuronen einer Schicht vom gleichen kochlearen Ort stammen. Da einem bestimmten kochlearen Ort eine bestimmte Frequenz zugeordnet werden kann, haben die Neuronen einer Schicht die gleiche charakteristische Frequenz, auf die sie am empfindlichsten reagieren. In der räumlichen Anordnung der Neuronen spiegeln sich somit die verschiedenen Frequenzen der Kochlea wieder. Die spektrale Zerlegung des Signals, die von der Kochlea durchgeführt wurde, wird im Gehirn aufrecht erhalten. Das zweite Prinzip betrifft die Muster der Konvergenz und der Divergenz von Nervenzellen der afferenten Hörbahn. Diese Muster zeigen sogar bei Erwachsenen ein gewisses Maß an Modellierbarkeit. Die zugrunde liegenden Mechanismen sind noch unbekannt, es gibt jedoch Hinweise darauf, dass die 6 Diskussion 97 Verformbarkeit das morphologische Substrat des auditiven Lernens oder der Erholung pathologischer Prozesse darstellt (Phillips 1995). Die sensorische Repräsentation kann mehrere Formen annehmen, z.B. als Lokalisation in einem bestimmten Hirngebiet oder als Verteilung einer zellulären Eigenschaft. Die sensorische Repräsentation ist veränderlich. Experimentell verursachte Taubheit führt zu einer signifikanten Änderung der Verknüpfungen im auditiven System (Phillips 1995). Mehrere Untersuchungen von PHILLIPS weisen daraufhin, dass die reine Worttaubheit eine Manifestation einer basalen zeitlichen Verarbeitungsstörung auf vorsprachlichem Niveau ist, d.h. dass der reinen Worttaubheit eine sprachunspezifische Störung im primären auditorischen Kortex zugrunde liegt. Eine Anzahl von Hinweisen zeigt an, dass der primäre auditorische Kortex eine Rolle bei der Wahrnehmung, aber nicht bei der linguistischen Verarbeitung von Lauten spielt. PHILLIPS maß bei 94 Neuronen des primären auditorischen Kortex die Genauigkeit, mit der die Nervenzellen auf Töne oder Geräusche reagierten (Phillips 1990). Bei den 94 Neuronen lagen die Latenzzeiten7 im Bereich von 0,15 bis 1,5 Millisekunden. Das bedeutet, dass die Genauigkeit des Antwortverhaltens der kortikalen Neuronen ausreicht, um den zeitlichen Verlauf einzelner Sprachkomponenten darzustellen. Daraus folgt, dass der primäre auditorische Kortex bei der Unterscheidung verschiedener Sprachabschnitte eine Rolle spielt. Außerdem zeigten die Ergebnisse, dass die Antworten auf Reize im Kortex eine Genauigkeit des Antwortverhaltens zeigten, die nur etwas schlechter als die der Fasern des Kochlearnervs ist. Pathologische Veränderungen in der primären Hörrinde führen somit offenbar zu einer Beeinträchtigung grundlegender auditiver Wahrnehmungsprozesse, die Basis des Sprachverständnisses sind. Eines der wichtigsten diagnostischen Kriterien der Genauigkeit, mit der Nervenzellen auf Töne oder Geräusche reagieren ist die Prüfung, inwieweit die Frequenzlücke (4.2.2.1) erkannt werden kann. Hören ist die Verarbeitung 6 Diskussion 98 akustischer Ereignisse, die über die Zeit verteilt sind, und die Bestimmung der Frequenzlücke ist ein Mittel, um den zeitlichen Verlauf des Hörvermögens zu bestimmen (Phillips 1999). Die Wahrnehmung der Frequenzlücke ist eine wichtige vorsprachliche Grundlage zum Verstehen von Sprache. Der Fluss der Sprache wird von Zeitabschnitten unterbrochen, in denen die Klänge von sehr niedriger Amplitude sind oder der Zeitabschnitt sogar von vollständiger Stille ist. Die wichtigsten „ Frequenzlücken“ der Sprache sind die phonetisch wichtigen – sie lassen Stopp-Konsonanten erkennbar werden. Eine über 30 Millisekunden dauernde Pause wird als lang erkannt. Bei den Untersuchungen von PHILLIPS spielten hauptsächlich zeitliche Variationen im Bereich von 1 Millisekunde bis hin zu 10 Millisekunden eine Rolle, die die so genannten Stopp-Konsonanten phonetisch charakterisieren. In mehreren Untersuchungen prüfte PHILLIPS, welche Einfluss Geräusche, die eine Frequenzlücke abgrenzen, auf das Erkennungsvermögen der Lücke haben, und zwar sowohl in der Between-Channel- als auch in der Within-ChannelAlternative (Phillips 1997). In einem Versuch wurden die Versuchspersonen Paaren engbandiger Geräuschfolgen ausgesetzt. Der der Frequenzlücke vorangehende Ton hatte eine Frequenz von 2 kHz, und die Frequenz des folgenden wurde variiert. Die Erkennung der Frequenzlücke gelang sehr schlecht, wenn die spektrale Unterscheidung der beiden Geräusche, die die Frequenzlücke begrenzten, hoch war. Dies legt die Vermutung nahe, dass die Erkennung der Frequenzlücke sich verschlechtert, wenn hierzu ein Vergleich der Aktivitäten in verschiedenen Wahrnehmungskanälen des auditiven Systems erforderlich ist. Dies deutet daraufhin, dass die Erkennung der Frequenzlücke ein komplexer Vorgang der Wahrnehmung ist. Anderseits ist die Erkennung der Frequenzlücke eine wichtige Grundlage zum Verstehen von Sprache. Die wichtigsten „ Frequenzlücken“ der Sprache sind die phonetisch bedeutsamen – sie lassen Stopp-Konsonanten erkennbar werden. Grundlegende auditive Wahrnehmungsprozesse wie die Erkennung der Frequenzlücke sind somit für das Sprachverständnis von entscheidender Bedeutung. 7 Latenzzeit: von der Leitungsgeschwindigkeit des Nervs abhängiger Zeitraum zwischen 6 Diskussion 99 Nicht alle Bereiche des Sprachunterscheidungsvermögens sind bei der reinen Worttaubheit gleichermaßen betroffen. Insbesondere stellte PHILLIPS fest, dass das Wahrnehmungsvermögen langer Vokale bilaterale Läsionen des primären auditorischen Kortex übersteht, jedoch das von Konsonanten, insbesondere StoppKonsonanten, nicht (Phillips 1990). Die Laute von Stopp-Konsonanten zeigen zwei Ebenen zeitlicher Organisation. Die feine Wellenform, die im Bereich unterhalb von Millisekunden liegt, spezifiziert gänzlich die langsamen spektralen Wechsel, die den Konsonanten phonetisch identifizieren. Die gestörte Unterscheidungsfähigkeit von Stopp-Konsonanten bei Worttaubheit zeigt ein zeitliches Verarbeitungsdefizit an, das Stopp-Konsonanten betrifft, aber Vokale nicht. Dies liegt im Bereich von einer bis zu zehn Millisekunden und charakterisiert die Stopp-Konsonanten phonetisch. Da Stopp-Konsonanten aufgrund ihrer Frequenzlücken erkennbar werden, folgt, dass der primäre auditorische Kortex beim Erkennen der Frequenzlücke (und damit der Wahrnehmung von Sprache) eine entscheidende Rolle spielt. Dies weist daraufhin, dass die Störung auf einem vorsprachlichen Niveau beruht: Bestimmte Frequenzspektren akustischer Stimuli können nicht erfasst werden. Die Fehler beim Erkennen von Konsonanten, die von Patienten mit reiner Worttaubheit gemacht werden, können im stimmhaften Aussprechen und/oder in der Artikulation liegen. Ist das primäre Hörfeld geschädigt, so verfügt der zerebrale Kortex nicht mehr über eine akustische Darstellung von Signalen mit der entsprechenden zeitlichen Struktur, die für eine normale Spracherkennung erforderlich ist. Neuronale Antworten auf akustische Signale sind zeitlich weniger differenziert. Dies führt dazu, dass das Erkennungsvermögen der Patienten für die entsprechenden Laute abnimmt. Es stellt sich die Frage, warum das Unterscheidungsvermögen anderer Konsonanten (z.B. nasaler) erhalten bleibt. Eine Ursache könnte darin liegen, dass Reiz und Reizantwort 6 Diskussion 100 diese Konsonanten länger sind und weniger zeitlich differenziert sind als StoppKonsonanten (Metz-Lutz u. Dahl 1984). Bei den meisten Untersuchern besteht Übereinstimmung darin, dass für die reine Worttaubheit fast immer eine Läsion eines oder beider primärer Hörfelder im linken und/oder rechten Temporallappen pathognomonisch ist (Phillips 1990). Dies ist von Bedeutung aus zwei Gründen. Einer ist, dass die Diagnose der reinen Worttaubheit eine spezifisch linguistische Funktion einer klassischerweise sensorischen Hirnregion zuschreibt. Der zweite ist, dass die Störung diejenigen Patienten identifiziert, deren psychoakustische Mängel eine wichtige Datenbasis bieten könnte, um die Hörfunktionen des auditorischen Kortex zu bestimmen (Phillips 1990). 'LH3UR3RVLWLRQ Die Untersuchungen von PHILLIPS wurden durch mehrere Studien über das Spracherkennungsvermögen bestätigt. Wie bei PHILLIPS wurden Untersuchungen durchgeführt, die die Erkennung der Frequenzlücke bestimmten. Sie bestätigten die Ergebnisse von PHILLIPS, dass bei reiner Worttaubheit die Erkennung der Frequenzlücke vermindert ist. Insbesondere wurden Studien durchgeführt, die das zeitliche Auflösungsvermögen für nichtsprachliche Laute direkt bei den Patienten maßen. Drei Studien stellten bei Patienten mit reiner Worttaubheit die minimale Zeit zwischen zwei Lautübergängen fest, die erforderlich ist, um die Laute getrennt wahrnehmen zu können (Albert u. Bear 1974, Hendler et al. 1990, Schwarz u. Tomlinson 1990). Lagen die Zeiten bei Gesunden zwischen 1 und 3 Millisekunden, lagen sie bei worttauben Patienten zwischen 15 und 30 Millisekunden, und bei keinem der Patienten lag das Intervall unterhalb von 2 msec (Albert u. Bear 1974). Insbesondere bei Patienten mit schwereren Läsionen waren die Zeiten deutlich erhöht. 6 Diskussion 101 Bei Patienten mit reiner Worttaubheit wurde zudem deren Fähigkeit getestet, Phoneme zu identifizieren. Wie bei PHILLIPS wurde festgestellt, dass die Patienten größere Schwierigkeiten hatten, Verschlusslaute zu identifizieren als Vokale (Eggermont 1991). EGGERMONT stellt fest, dass seinem Ergebnis dadurch eine besondere Bedeutung verliehen wird, dass die akustischen Signale der Verschlusslaute zeitlich differenziert und aperiodisch sind (z.B. Zischen, Einatmen) (Eggermont 1991). Im Gegensatz hierzu zeigten bei den Untersuchungen von EGGERMONT die akustisch konstanten Vokale keine zeitlichen akustischen Veränderungen, die phonetisch von Bedeutung sind. Die zeitliche Struktur der akustischen Ereignisse lag bei Verschlusslauten im Bereich von einer bis zehn Millisekunden, d.h. im Bereich der Zeiteinstellung des primären Hörfelds, aber nicht im Bereich anderer zerebraler Bereiche (Eggermont 1991). Die Ergebnisse von EGGERMONT bestätigen somit die von PHILLIPS, dass der zeitliche Verlauf eine entscheidende Rolle bei der Identifizierung von Phonemen spielt, die im primären Hörfeld stattfindet. Auch neurophysiologische Ergebnisse bekräftigen die Aussagen von PHILLIPS. Die grundlegende funktionelle Organisation des primären auditorischen Kortex ähnelt im Prinzip der von Primaten und Katzen (Brugge 1975). Er ist tonotopisch organisiert, was darauf hinweist, dass seine afferente Bahn die frequenzspezifischen Kanäle bewahrt hat, die in der auditorischen Peripherie, wie beispielsweise der Kochlea, entstanden sind. Dies führt zu der Schlussfolgerung, dass die kortikale Repräsentation des Spektrums von jedem beliebigen auditorischen Ereignis auf der räumlichen Verteilung der Aktivität beruht, die im Bereich des neuronalen Mosaiks evoziert wurde, das die Anordnung der Frequenzen der Kochlea wiedergibt (Brugge 1975). Es gibt zwei Möglichkeiten, die Fähigkeit der kortikalen oder auditorischen Neuronen zu bestimmen, die Zeit-Abstimmung von Stimulusereignissen zu codieren (Hall 1991). Eine ist das „ Zittern“ in der Synchronisation von Antworten auf einen transitorischen Stimulus. Im Kortex der Katze sind die 6 Diskussion 102 Standardabweichungen dieser Spike-Zeiten8 relativ klein, gewöhnlich unter 1,5 msec, und sind vergleichbar mit denen im Kochlearnerv (Hall 1991). Das zweite Mittel, mit dem Hörnerven die zeitliche Struktur eines Lauts anzeigen, beruht auf dessen Fähigkeit, Spike-Entladungen mit Stimulus-Frequenzen in eine Phasenübereinstimmung zu bringen. Direkte Messungen von kortikalen Zellen zeigen, dass die optimalen Frequenzen unter 100 Hz liegen und dass Frequenzen über 4000 Hz nicht in Phasenübereinstimmung gebracht werden können. Das bedeutet, dass die Laute der Sprache in Phasenübereinstimmung gebracht werden können. Antworten auf Reize im Kortex zeigten eine Genauigkeit des Antwortverhaltens, die nur etwas schlechter als die der Fasern des Kochlearnervs war (Creutzfeld et al. 1980). Dies bestätigt wieder – wie ALBERT und BEAR feststellten (Albert u. Bear 1974) –, dass die Einschränkung der Spracherkennung bei primärer Worttaubheit eine Manifestation einer eher grundlegenden zeitlichen Verarbeitungsstörung des auditiven Systems auf einer unteren Verarbeitungsebene (Hörnerv) ist, die sich auf eine höhere (den Kortex) fortpflanzt. Mehrere Untersuchungen bestätigen die Ergebnisse PHILLIPS, dass für die reine Worttaubheit fast immer eine Läsion des einen oder der beiden primären Hörfelder pathognomonisch ist (Phillips 1990). Die Worttaubheit kann einerseits mit bilateralen Läsionen der Temporalregionen verbunden sein (Auerbach et al. 1982), zum anderen mit Läsionen, die auf den linken Temporallappen beschränkt sind (Luria 1976). Bereits 1974 wurde eine neurophysiologische Studie von ALBERT und BEAR bei Tieren durchgeführt. Sie zeigte insbesondere, dass dem primären Hörfeld eine besondere Rolle zukommt in der Darstellung von Klängen mit zeitlichen Komponenten in einem Bereich von 200 bis 4000 Hz (Albert und Bear 1974), d.h. in dem Bereich, in dem die menschliche Sprache liegt. 8 Spike: Spitze, Kurvenzacke. Spikes können während der Reaktion auf einen Stimulus von der Oberfläche einer Nervenstruktur abgeleitet werden. Es sind spitze Potenziale mit steil ansteigender und abfallender erhöhter Amplitude. 6 Diskussion 103 Bei dem von OTSUKI et al. beschriebenen Fall reiner Worttaubheit war eine kortikale Atrophie feststellbar, die hauptsächlich in der linken oberen Temporalregion lokalisiert war, d.h. dem Ort der primären Hörrinde (Otsuki et al. 1998). Es handelte sich um einen 67-jährigen rechtshändigen Patienten, der innerhalb eines Zeitraums von neun Jahren eine reine Worttaubheit entwickelte, ohne dass andere Minderungen geistiger Fähigkeiten auftraten. NOTOYA et al. beschrieben einen Fall reiner Worttaubheit, der die Ergebnisse von PHILLIPS bestätigt (Notoya et al. 1991). Die Patientin litt unter einer reinen Worttaubheit in Verbindung mit einem Landau-Kleffner-Syndrom (s. 4.2.2.2). Erste Störungen machten sich im Alter von vier Jahren bemerkbar. Im Alter von 20 Jahren hatte sich die Hörstörung nicht signifikant gebessert. Die Patientin konnte zunächst weder Vokale noch aus Vokalen und Konsonanten bestehende Silben erkennen. Später hatte sie keine Schwierigkeiten mehr, Vokale zu erkennen, aber ihre Identifizierung von Konsonanten verbesserte sich kaum. Bei den Konsonanten konnte sie zwischen stimmlosen und stimmhaften unterscheiden, aber sie hatte große Probleme, die Konsonanten innerhalb jeder Gruppe zu identifizieren. Diese Ergebnisse lassen darauf schließen, dass es der Patientin unmöglich ist, kurzzeitig andauernde Konsonanten zu erkennen. Die zugrunde liegende Störung ist eine Insensibilität für Lautheit und ein Defekt in der zeitlichen Auflösung, d.h. eine Störung im vorsprachlichen Niveau. BADDELEY und WILSON (1976) untersuchten einen Fall von Worttaubheit, bei dem die Fähigkeit der Patientin, Worte zu wiederholen, eingeschränkt war, obwohl die audiometrisch und tympanometrisch gemessenen Werte im Normbereich lagen. Auch hatte die Patientin Schwierigkeiten, Laute aus der Umgebung zu erkennen. Dagegen waren ihr phonologisches visuelles Gedächtnis und ihr auditorisches Gedächtnis nur geringfügig beeinträchtigt. Aufgrund dieser Ergebnisse kamen BADDELEY und WILSON zu dem Schluss, dass bei der Patientin grundlegende auditive Wahrnehmungsprozesse gestört waren (Baddeley u. Wilson 1976). 6 Diskussion 104 SAFFRAN et al. (1976) beschrieben einen Fall einer reinen Worttaubheit. Es handelte sich um einen 37-jährigen Patienten, der seit Jahren unter epileptischen Anfällen und einer peripheren Neuropathie aufgrund eines Alkoholabusus litt. Der Patient hatte insbesondere Probleme in der Diskrimination von verbalen Verschlusslauten (Stopp-Konsonanten). Nach SAFFRAN et al. weist dies daraufhin, dass die Störung auf einem vorsprachlichen Niveau beruht. Die reine Worttaubheit sei eine Störung der zeitlichen Verarbeitung des auditorischen Stimulus. 'LH&RQWUD3RVLWLRQ Nicht alle Untersucher unterstützen die Aussage PHILLIPS, dass die reine Worttaubheit eine Manifestation einer eher grundlegenden zeitlichen Verarbeitungsstörung auf vorsprachlichem Niveau sei. Die Wissenschaftler sind sich auch nicht einig darüber, wo die Schädigungen, die eine reine Worttaubheit bedingen, genau in den Sprachzentren des Gehirns lokalisiert sind. Es besteht noch nicht einmal Klarheit darüber, ob eine bestimmte Aphasieform mit einer Schädigung eines bestimmten Ortes verbunden ist. SCHMIDT et al. sind überzeugt, dass den verschiedenen Aphasieformen abgrenzbare und verschiedene Läsionsorte im Gehirn zugrunde liegen würden. Die transkortikale sensorische Aphasie, bei der die Fähigkeit nachzusprechen zwar gut erhalten ist, wohingegen Defizite im Sprachverständnis ausgeprägt sind, werde durch Läsionen in der Nähe des linken perisylvischen Bereichs verursacht (Schmidt et al. 2001). HUBER et al. meinen dagegen, dass die Theorie, dass Aphasien mit lokalisierten Läsionen (verschiedene Aphasiesyndrome mit spezifisch lokalisierten Läsionen) verbunden seien, veraltet sei und heutzutage teilweise ihre Gültigkeit verloren 6 Diskussion 105 habe. Die Meinung, dass die Zuordnung einer verlorenen Funktion auf einen einzigen umschriebenen geschädigten Bezirk möglich sei, müsse berichtigt werden (Huber et al. 2000). HUBER et al. (2000) sind der Überzeugung, dass noch unbekannt sei, wo der Entstehungsort der reinen Worttaubheit genau liege. Nach Meinung der Forscher deutet vieles daraufhin, dass dies kein einziger klar abgrenzbarer Bereich sei, sondern mehrere gleichzeitig ablaufende Mechanismen an verschiedenen Orten eine Rolle spielten. HUBER et al. (2000) sind überzeugt davon, dass insbesondere die neuen Techniken (PET, kortikale Stimulation, CT, MRI, FMRI, siehe 3.5) eine genaue Lokalisation am Lebenden und eine Verbindung zu den vorhandenen Sprachstörungen erlauben. Zudem lassen sie die Entwicklung einer Sprachstörung im Verlauf der Zeit verfolgen. Es zeige sich, dass nicht mehr eng umschriebene Zentren für bestimmte sprachliche Leistungen zuständig sind, sondern dass eine synchronisierte Aktivität in ausgedehnten neuronalen Netzwerken von Regionen in der Hirnrinde, in den subkortikalen Kerngebieten und den Verbindungen dieser Bezirke in reziproker Weise stattfindet (Huber 2000). Verschiedene Wissenschaftler kommen nach Falluntersuchungen zu dem Schluss, dass die reine Worttaubheit auf kombinierten Defekten beruhe. Hier gilt der von VON STOCKERT beschriebene Fall erneut als beispielgebend: Nach einem Selbstmordversuch durch eine schwere Schussverletzung bei der beide Temporalregionen betroffen waren litt der 23-jährige Patient unter einer reinen Worttaubheit. Trotz intensiver Rehabilitationsbemühungen blieb eine leichte linksseitige Ataxie, ein bilateraler oberer Gesichtsfeldausfall und eine Verständigungsschwierigkeit bestehen. Intensive Untersuchungen bestätigten die Vermutung, dass es sich hier um primäre Worttaubheit handelt. Der neurologische Zustand blieb auch mehrere Monate danach noch nahezu unverändert bestehen, d. h. noch immer bestanden die leichte Ataxie, der Gesichtsfeldausfall und eine schwere Störung des auditiven Wahrnehmungsvermögens, die dem Patienten eine Unterhaltung unmöglich machte. (von Stockert 1982). 6 Diskussion 106 Seine Sprache war schnell, aber nicht dysarthrisch. Bis zu einem gewissen Grade konnte er Sätze verstehen. Er hatte mehr Schwierigkeiten, einzelne Phoneme und Silben zu verstehen als Worte bzw. Sätze. Dies machte sich besonders dann bemerkbar, wenn er zwischen zwei Worten unterscheiden musste, die nur in einer einzigen Silbe verschieden waren. Die Verständnisschwierigkeit nahm bei längeren Worten zu, trat jedoch weniger stark bei Sätzen in Erscheinung. Nach mehreren Tests, in denen der Patient gehörte Silbenfolgen (Worte und sinnlose Silbenkombinationen) nachsprechen sollte, kam VON STOCKERT zu dem Ergebnis, dass den Patienten offensichtlich ein gestörtes phonemisches Muster erreichte und er versuchte, das wirkliche Wort in einem „ inneren Wörterbuch“ zu finden. VON STOCKERT stellte fest, dass im Gegensatz zum Gesunden die Verständnisschwierigkeit des Patienten mit der Anzahl der Silben zunahm. Insgesamt kam VON STOCKERT (1982) zu dem Schluss, dass die reine Worttaubheit durch Störungen höherer kortikaler Funktionen verursacht werde. Seiner Meinung nach bestand der zugrunde liegende Hauptmechanismus der Erkrankung darin, dass es nicht gelingt, akustische Stimuli in eine richtige Reihenfolge zu bringen. Beim Vergleich verschiedener wissenschaftlicher Beiträge stellten OTSUKI et al. fest, dass keine Beziehung zwischen dem Aphasietyp und dem pathologischen Befund festzustellen sei (Otsuki et al. 1998). So kann nach OTSUKIs Meinung kein bestimmter Kortexbereich gefunden werden, dessen Läsion eine reine Worttaubheit hervorruft. PRAAMSTA et al. stellen fest, dass die Ursache der reinen Worttaubheit auf kombinierten Defekten beruhe (Praamsta et al. 1991). Die Wissenschaftler beschreiben einen Fall, der nach einem linksseitigen temporoparietalen Infarkt worttaub wurde und nach einem rechtseitigen Infarkt auch das semantische Sprachverständnis verlor. Daraus schließen PRAAMSTA et al., dass die reine Worttaubheit auf mehreren, miteinander Schädigungen beruhe (Praamsta et al. 1991). in Zusammenhang stehenden 6 Diskussion 107 Auch eine andere Aussage PHILLIPS blieb nicht unwidersprochen. So zeigten die Untersuchungen von PHILLIPS, dass die Antworten auf Reize im Kortex eine Genauigkeit des Antwortverhaltens zeigten, die nur etwas schlechter als die der Fasern des Kochlearnervs ist. Dies steht im Gegensatz zu früheren Ergebnissen zu der langsamen Antwort kortikaler Zellen, die wenigstens eine Ordnung schlechter ist als diejenige der Fasern des Hörnervs und vieler kochlearer Nukleuszellen. So stellen CARNEY und GEISLER (1986) fest, es sei bemerkenswert, dass die Fasern des Kochlearnervs nur eine Synapse entfernt von der Basilarismembran seien und sich mit jeder zusätzlichen Synapse auf der Hörbahn die Genauigkeit des Codes verringern würde – teilweise aufgrund des Zitterns, das durch chemische synaptische Vorkommnisse verursacht würde und teilweise durch neuronale Anpassung. In diesem Zusammenhang sei die Darstellungsgenauigkeit der zeitlichen Struktur insbesondere der Vokalstruktur auf der Ebene des ventralen Nucleus cochlearis bereits deutlich erniedrigt, und es sei verständlich, dass die Genauigkeit im primären auditorischen Kortex zu gering sei, um die zeitliche Verschlüsselung der Vokalstruktur zu unterstützen. 6FKOXVVIROJHUXQJ Leidet ein Patient unter einer grundlegenden auditorischen Wahrnehmungsstörung, so ist es für ihn unmöglich, Laute richtig zu erkennen, und zwar sowohl sprachliche als auch nichtsprachliche. Ist diese Fähigkeit erloschen, ist es jedoch auch nicht mehr möglich, Silben und Worte zu identifizieren oder gar dem Gehörten eine semantische Struktur zuzuordnen. Das bedeutet, es ist auch die Fähigkeit verloren gegangen, Sprache zu erkennen, also eine Fähigkeit, die den höheren Sprachzentren zugeschrieben wird. Somit pflanzt sich eine Störung auf der frühen Verarbeitungsebene auf die höheren fort. Beispielsweise ist in dem von VON STOCKERT beschriebenen Fall die Verständnisschwierigkeit bei Phonemen besonders hoch (von Stockert 1982). 6 Diskussion 108 Dies lässt erkennen, das der Patient über eine intakte innere Struktur höherer Sprachzentren verfügt und somit auf ein Lexikon zurückgreifen kann, das es ihm möglich macht, aus dem gestörten empfangenen Klangmuster ein sinnvolles Wort zu rekonstruieren. Bei Phonemen gelingt dies jedoch nicht immer. Stellen Wissenschaftler fest, dass eine Worttaubheit auf kombinierten Defekten höherer geistiger Funktionen beruhe, so beziehen sie sich zumeist auf die Beschreibung eines Falles, der in der Tat von multiplen Traumata der Rindenoberfläche betroffen ist. Es ist dann nicht auszumachen, welcher Bezirk die „ reine Worttaubheit“ und welcher Bezirk andere Dysfunktionen verursacht. So beziehen sich die Fallbeschreibungen von OTSUKI, VON STOCKERT und PRAAMSTA auf Patienten, bei denen mehrere Läsionen der Rindenoberfläche bestanden. Daher ist davon auszugehen, dass der primäre auditorische Kortex von der Erkrankung nicht allein betroffen war (Otsuki et al. 1998, von Stockert 1982, Praamsta et al. 1991). So ist verständlich, dass die Patienten unter kombinierten Defekten litten. Insbesondere hatte der von PRAAMSTA beschriebene Fall nach einer Läsion im linken Temporallappen nachfolgend einen Infarkt im rechten Temporallappen erlitten. Erst im Anschluss an den zweiten Schlaganfall kam es zu einem schweren Sprachverständnisdefizit. Auch andere Feststellungen, die im Gegensatz zu den Aussagen von PHILLIPS stehen, sind zweifelhaft. Die Aussage von CARNEY und GEISLER (1986), dass die Darstellungsgenauigkeit der zeitlichen Struktur von Lauten im Bereich des primären Hörkortex deutlich geringer sei als die im Bereich des Kochlearnervs, bezieht sich in erster Linie auf Vokale. Auch PHILLIPS stellte fest, dass das Erkennungsvermögen für Vokale bei primärer Worttaubheit erhalten bliebe, nicht jedoch das für Stopp-Konsonanten (Phillips 1995). Dies lässt darauf schließen, dass die Erkennung der zeitlichen Struktur von Vokalen nicht im primären auditorischen Kortex stattfindet, wohl aber das für Stopp-Konsonanten. 6 Diskussion 109 (QWVWHKXQJVEHGLQJXQJHQGHU/HJDVWKHQLH 3RVWXODWHXQG6FKOXVVIROJHUXQJHQYRQ7$//$/ 7$/&277XQGDQGHUHQ Auch andere Untersuchungen, die sich jedoch auf ein anderes Krankheitsbild, nämlich die Legasthenie beziehen, kamen zu dem Ergebnis, dass der Wahrnehmung nichtsprachlicher Reize eine höhere Bedeutung zukommt, als bisher angenommen wurde. So beschreiben mehrere Wissenschaftler – insbesondere Paula TALLAL sowie Joel TALCOTT – das reibungslose Funktionieren der optischen und akustischen Sinneswahrnehmungen als unumgängliche Voraussetzung für das erfolgreiche Erlernen von Schreiben und Lesen. In verschiedenen Untersuchungen konnte nachgewiesen werden, dass bei Legasthenikern häufig ein auditives bzw. visuelles Informationsverarbeitungsdefizit bereits auf nichtsprachlicher Ebene vorliegt. Da die Wahrnehmung nichtsprachlicher Reize Voraussetzung für die Sprachverarbeitung in höheren Ebenen ist, kommt dieser Störung mit hoher Wahrscheinlichkeit eine große Bedeutung bei der Entstehung der Legasthenie zu. 6W|UXQJHQGHU6HKYHUDUEHLWXQJ TALCOTT und STEIN stellten fest, dass etwa 75 Prozent der legasthenen Kinder in Tests schwächer abschneiden, die eine rasche visuelle Informationsverarbeitung verlangen. Zudem entdeckten TALCOTT et al., dass erwachsene legasthene Versuchspersonen auf langsame oder kontrastreiche visuelle Reize genauso gut reagierten wie gute Leser. Bei schnell bewegten Reizen mit niedrigem Kontrast aber schnitten die legasthenen Versuchspersonen im Vergleich mit Erwachsenen ohne Leseschwächen deutlich schlechter ab (Stein u. Talcott 1999). 6 Diskussion 110 In den Untersuchungen von TALCOTT und STEIN (1999) gelang es Legasthenikern, erst ab einer höheren Anzahl von sich bewegenden Punkten auf einem Computerbildschirm zu erkennen, dass diese plötzlich in die gleiche Richtung bewegt werden. Je mehr Schwierigkeiten sie bei dieser Aufgabe hatten, desto stärker war auch ihre LRS ausgeprägt. TALCOTT und STEIN sind überzeugt, dass dieser Befund es sehr wahrscheinlich mache, dass die Lesefähigkeit auf der Fertigkeit beruht, rasche Veränderungen elementarer Sinneseindrücke wahrzunehmen. Somit hängt die Legasthenie mit einer Schwäche beim Verarbeiten optischer Eindrücke im Gehirn zusammen. TALCOTT und STEIN untersuchten zwei Verarbeitungskanäle der menschlichen Sehbahn, die ihren Anfang in unterschiedlichen Zellen der Retina nehmen und zu zwei unterschiedlichen Zellschichten (der magnozellulären und der parvozellulären) im Corpus geniculatum laterale leiten. Von dort werden die Seheindrücke zum visuellen Kortex weitergeleitet. Der magnozelluläre Anteil des primären visuellen Kortex (V1) projiziert über die Area V5 überwiegend in den parietalen Kortex (so genannte M-Bahn), hingegen erreichen den temporalen Kortex sowohl parvo- als auch magnozelluläre Bahnen (Merigan und Maunsell 1993). Die großen Zellen im Corpus geniculatum laterale (großzelliger oder magnozellulärer Verarbeitungsweg) sind äußerst empfindlich für schwache Kontraste und verarbeiten rasch aufeinander folgende bzw. sehr kurz anhaltende Seheindrücke. Die magnozellulären Hirnzellen dienen zudem der Integration von Informationen zwischen Fixationen und Augenbewegungen (Lovegrove 1982). Signale aus diesem System werden bis in Gebiete der Hirnrinde weitergeleitet, die auch an der Steuerung von Augenbewegungen beteiligt sind. Die kleinen Zellen im Corpus geniculatum laterale (kleinzelliger oder parvozellulärer Verarbeitungsweg) verarbeiten Farben, starke Kontraste und lang anhaltende bzw. unbewegte Sehreize. Außerdem sind sie für das räumliche Sehen zuständig. 6 Diskussion 111 TALLAL stelllte fest, dass die Legasthenie offenbar in Verbindung mit Normabweichungen neuroanatomischer und hirnfunktioneller Merkmale steht, wobei der M-Bahn eine besondere Bedeutung zukommt. Es gibt Hinweise dafür, dass diese von Magnozellen gebildete Bahn bei Legasthenikern weniger leistungsfähig ist (Tallal 2000). GALABURDA und LIVINGSTONE entdeckten in histologischen Präparaten des Corpus geniculatum laterale bei LRS-Patienten eine Verminderung der Zahl der Nervenzellen in der Magnozellschicht (Galaburda u. Livingston 1993). Dies könnte in Zusammenhang mit einer visuellen Wahrnehmungsschwäche stehen: Wie bereits erwähnt, wird im Corpus geniculatum laterale die visuelle Information umgeschaltet und zur optischen Hirnrinde weitergeleitet. Auch TALCOTT et al. (1997) fanden bei Untersuchung von Hirnschnitten ehemals legasthener Personen, dass die großzelligen Schichten im seitlichen Kniehöcker von legasthenen Gehirnen weniger organisiert waren und kleinere Zellkörper enthielten im Vergleich mit Gehirnen von ehemals nichtlegasthenen Personen. Da das Lesen sehr rasche Informationsverarbeitung erfordert, könnte die unzureichende Leistungsfähigkeit des großzelligen Verarbeitungskanals Leselern- und Leseschwierigkeiten mit visuellem Schwerpunkt erklären (Talcott et al. 1997). Zwar fanden GALABURDA und LIVINGSTONE (1993) bei Legasthenikern Unterschiede in der Struktur und Funktion der magnozellulären Systeme, aber intakte parvozelluläre Systeme. Es zeigten sich signifikante zelluläre Unterschiede auf der Ebene des Thalamus, und zwar nicht nur im Corpus geniculatum laterale, das visuelle Informationen weiterleitet, sondern auch im weiteren Verlauf des visuellen Systems. Daher stellten STEIN und TALCOTT die Hypothese auf, dass bei Legasthenikern – vielleicht als Folge einer mütterlichen immunogenen Reaktion – während der fetalen Entwicklung junge Nervenzellen im Gehirn geschädigt würden. Dadurch komme es zu einer Behinderung ihres Zusammenwachsens. Ebenso sei die 6 Diskussion 112 Ausbildung der M-Bahn von der Netzhaut zum lateralen Corpus geniculatum und weiter zum visuellen Kortex gestört (Stein u. Talcott 1999). 6W|UXQJHQGHU+|UYHUDUEHLWXQJ Von Paula TALLAL wurde der Begriff des Specific Language Impairment (SLI) geprägt. Diese Störung benennt eine nicht altersgemäße Entwicklung der Sprechfähigkeit bei ansonsten normalen kognitiven Fähigkeiten (Tallal u. Piercy 1973, Tallal 1993). Die Sprachverzögerung beträgt mindestens ein Jahr, der Handlungs-IQ liegt im Durchschnittsbereich. Als TALLAL et al. (1985) sprachverzögerte Kinder mit Leselernschwierigkeiten untersuchten, kamen die Wissenschaftler zu dem Ergebnis, dass 98 Prozent der sprachbehinderten Kinder von Kontrollkindern allein mit Hilfe einer Testreihe unterschieden werden konnten, die schnelle Sprachproduktion oder rasche Lautunterscheidung bzw. eine Unterscheidung von rasch aufeinander folgenden Berührungsreizen erforderte. Dabei stellten TALLAL et al. (1985) eine zeitliche Verlangsamung der Hörverarbeitung bei Legasthenikern fest. Unter anderem machte sich diese in einer mangelhaften „ Filterfunktion“ des Gehörs bemerkbar: Einen bestimmten Sprecher aus einem „ akustischen Salat“ von Störgeräuschen und anderen Sprechern herauszuhören und zu verstehen stellt für Legastheniker eine besondere Schwierigkeit dar (Tallal et al. 1985). TALLAL et al. (1985) konnten nachweisen, dass das bei Legasthenie häufig vorkommende auditive Informationsverarbeitungsdefizit bereits auf nichtsprachlicher Ebene vorliegt. Da die Wahrnehmung nichtsprachlicher Reize Voraussetzung für die Sprachverarbeitung in höheren Ebenen ist, kommt dieser Störung mit hoher Wahrscheinlichkeit eine große Bedeutung bei der Entstehung der Legasthenie zu (Tallal et al. 1985). 6 Diskussion 113 TALLAL et al. (1985) sind überzeugt, dass auch die Verarbeitung in den Hörbahnen bei legasthenen Kindern sehr häufig beeinträchtigt ist (Tallal et al. 1985). So können die Zellen der entsprechenden Schaltstelle im Corpus geniculatum mediale kurz anklingende Hörreize nicht verarbeiten. Dies ist der Grund, warum es einem hohen Prozentsatz legasthener Menschen nicht möglich ist, die nur kurz anklingenden Konsonanten (vor allem b–p, d–t, g–k) und die ähnlichen Kurzvokale (o–u, i–e) zu unterscheiden (Tallal et al. 2000). 7$//$/+\SRWKHVH TALLAL et al. (1993) nehmen einen Zusammenhang zwischen auditiven und – wie STEIN und TALCOTT (1999) – auch visuellen Problemen an. Sowohl die Unterscheidung von Sprachlauten im gesprochenen Wort als auch die von Buchstabenkombinationen des geschriebenen Wortes erfordern eine rasche zeitliche Verarbeitung. Offenbar besteht dabei ein Zusammenhang mit Defiziten bei akustischen Sinnesleistungen: Kinder, die bei der Untersuchung TALCOTT‘s et al. (1993) schlecht abschnitten, hatten auch große Probleme, zwischen richtig und falsch buchstabierten Wörtern zu unterscheiden. Je ausgeprägter sich dieses doppelte Defizit zeigte, desto schwerer war die Leseschwäche (Talcott et al. 1993). Die Untersuchungen von TALLAL et al. (1993) bekräftigen die Hypothese, dass Leseschwache dann Schwächen in der Wahrnehmung von nichtsprachlichen Reizen aufweisen, wenn diese in einem sehr kurzen zeitlichen Abstand nacheinander präsentiert werden. In dieser Untersuchungsreihe wurden legasthene mit nichtleseschwachen Kindern anhand von Aufgaben zum Erkennen der Reihenfolge von zwei Tönen bzw. zwei komplexen Tonkombinationen verglichen (Tallal 1980). 6 Diskussion 114 Ein Unterschied zwischen den beiden Gruppen zeigte sich erst dann, wenn der Zeitabstand zwischen zwei Reizen geringer als 305 msec war. Aus diesem Ergebnis kann gefolgert werden, dass sich bei sehr kurzem Interstimulusintervall bereits auf nichtsprachlicher Ebene eine Schwäche zeigt, die möglicherweise im direkten Zusammenhang mit der Sprachwahrnehmung steht (Tallal et al. 1993). Insgesamt spielen zeitabhängige Mechanismen im Nervensystem offenbar eine zentrale Rolle in den grundlegenden Prozessen der Informationsverarbeitung. Daher sind sie von großer Bedeutung für die normale Entwicklung des phonologischen Systems. Offenbar besteht ein enger Zusammenhang zwischen Dysphasie und Legasthenie (Tallal et al. 1993). So zeigten Studien, dass Kinder, deren Sprachentwicklung verzögert ist, ein hohes Risiko haben, Legasthenie zu entwickeln (Tallal 2000). Es kann davon ausgegangen werden, dass eine Störung im akustischen Sinneskanal dazu führt, dass zu wenige für die Sprachentwicklung wichtige Lautreize aus der Umwelt das Gehirn erreichen. So verbinden sich die betroffenen Nervenzellen nur ungenügend zu Einheiten, die bestimmten Lauten zugeordnet sind. Die verringerte Fähigkeit, die raschen Schwankungen der Tonhöhe zu identifizieren, geht mit Schwierigkeiten beim Aussprechen mancher Wörter einher. Das bedeutet, dass sowohl Sprach- als auch Leseprobleme ihren Ursprung in einer ungenügend entwickelten Fähigkeit der Verarbeitung und Decodierung schneller Impulse haben (Tallal 2000). Die visuelle Identifizierung von Graphemen ist nur dann möglich, wenn die visuelle Darstellung mit der begrenzt dauernden neuronalen Darstellung des passenden Phonems assoziiert werden kann. TALLAL et al. (1993) nehmen an, dass sprach- und lesegestörte Kinder- entsprechend ihrer grundlegenden Störung der auditiven Verarbeitung – keine stabilen und unbeweglichen neuronalen Darstellungen von Phonemen bilden können. Wie LIBERMANN (1985) erklärte, gelingt es diesen Kindern dann nicht, eine phonologische Bewusstheit zu erreichen. 6 Diskussion 115 1HXURDQDWRPLVFKH9HUlQGHUXQJHQ TALLAL et al. (1991) stellten, verglichen mit gesunden Kindern, verschiedene Muster der zerebralen Lateralisation in Gehirnen von SLI-Kindern fest. Insbesondere kann bei SLI-Kindern die physiologische Asymmetrie zugunsten der linken Hirnhälfte aufgehoben sein, und wiederholt wurden Abnormitäten der dort gelegenen Hirnwindungen belegt. Diese Abnormitäten zeigten sich teilweise sowohl im Präfrontalkortex als auch in der parietalen Region. Es fand sich eine hochsignifikante Korrelation zwischen dem Ausmaß der hemisphärischen Asymmetrie dieser kortikalen Bereiche und dem Ausmaß des Defizits bezüglich der zeitlichen Verarbeitung akustischer Ereignisse. 'LH3UR3RVLWLRQ 6W|UXQJHQGHU6HKYHUDUEHLWXQJ WARNKE und REMSCHMIDT (1982) wiesen nach, dass bei mindestens 5 bis 10 Prozent der legasthenen Kinder visuell-räumliche Wahrnehmungsschwierigkeiten anzutreffen sind. Diese Defizite zeigten sich in der Analyse und im Kodieren von visueller Information. In neurobiologischen Befunden konnte gezeigt werden, dass bei einem Teil der Personen mit LRS Besonderheiten in der Anatomie und der Hirnfunktion des Systems visueller Informationsverarbeitung zu erkennen sind. Es wurden Anomalien der neuronalen Migration in den magnozellulären Schichten des lateralen Nucleus geniculatus gefunden. Außerdem zeigten Kinder mit LRS linkszentral abnorme Verläufe im visuell-evozierten Potenzial (Warnke und Remschmidt 1992). Die Annahme gestörter kortiko-thalamo-kortikaler Netzwerke konnte durch magnetresonanztomographische Untersuchungen unterstützt werden. JERNIGAN et al. (1991) wiesen eine signifikante Reduktion der grauen Substanz in subkortikalen Strukturen (einschließlich des Corpus striatum und des Thalamus) 6 Diskussion 116 sowie in kortikalen Strukturen nach, die bekannt sind, der Sprachverarbeitung zu dienen. 6W|UXQJHQGHU+|UYHUDUEHLWXQJ Nach Untersuchungen an legasthenen Kindern kamen MANIS et al. (1997) zu mit denen von TALLAL (2000) übereinstimmenden Schlüssen. MANIS et al. (1997) wiesen nach, dass 7 von 25 Legasthenikern Schwächen bei der nichtsprachlichen Wahrnehmung von Sprache hatten, aber nur 1 Kind in der Vergleichsgruppe mit altersgerechter Leseentwicklung. MANIS et al. schließen daraus, dass Legastheniker ein nichtsprachliches Wahrnehmungsdefizit haben können, das die Sprachverarbeitung verzögert und zur Lese-Rechtschreib-Schwäche führt (Manis et al. 1997) – eine Aussage, die mit derjenigen von Paula TALLAL (2000) übereinstimmt. /HJDVWKHQH6FKZlFKHQ±6FKZlFKHQGHUUDVFKHQ]HLWOLFKHQ 9HUDUEHLWXQJ Die Untersuchungen von TALLAL et al. (1993) unterstützen die Hypothese, dass Leseschwache dann Schwächen in der Wahrnehmung von nichtsprachlichen Reizen aufweisen, wenn diese in einem sehr kurzen zeitlichen Abstand nacheinander präsentiert werden. Auch andere Wissenschaftler kommen zu diesem Ergebnis. So entdeckten LIVINGSTONE et al., dass erwachsene legasthene Versuchspersonen auf langsame oder kontrastreiche visuelle Reize genauso gut reagierten wie gute Leser. Bei schnell bewegten Reizen mit niedrigem Kontrast aber schnitten die legasthenen Versuchspersonen im Vergleich mit Erwachsenen ohne Leseschwächen deutlich schlechter ab (Livingstone et al. 1991). 6 Diskussion 117 Auch MERZENICH et al. (1993) und LLINAS (1993) stellen fest, dass Legastheniker schnelle Impulse unzureichend verarbeiten und decodieren. Aus ihren Untersuchungen schließen MERZENICH et al. (1993), dass Defizite in der zeitlichen Verarbeitung von Sprache erlernt sein können. Die erhaltenen Daten bestärken die Vermutung, dass Störungen der zeitlichen Verarbeitung im Hörsystem ausreichen, um eine Entwicklung des Lesenlernens zu unterbrechen. So könne ein früher Erfahrungsmangel das Nervensystem so stark schädigen, dass ihm eine schnelle Informationsverarbeitung unmöglich sei. Da das Gehirn aber gewährleisten müsse, dass es sich in Einklang mit der äußeren Welt befinde, passe sich seine Verarbeitung der Sinne dem langsamsten an. MERZENICH et al. (1993) sind der Überzeugung, dass sich neuronale Veränderungen aufgrund eines fehlerhaften Lernvorgangs bilden würden. Ein Defekt in nur einem von mehreren hundert Proteinen kann zu einer Verlangsamung des Lernprozesses führen. Da Legastheniker normal intelligent sind, ist zu vermuten, dass Lesen auf der Fähigkeit eines nicht oder wenig assoziativen Systems beruht. MERZENICH et al. (1993) glauben, dass die neuronale Veränderung, die eine Legasthenie verursacht, auf Veränderungen beruht, die die Eigenschaften der zeitlichen Verarbeitung von Eingaben kontrollieren (Merzenich et al. 1993). LLINAS (1993) stellt eine andere Hypothese vor. Nach LLINAS (1993) ist bei Legasthenikern eine besondere innere „ Uhr“ , die die Rate der neuronalen Entladungen kontrolliert, gestört. So liegt nach LLINAS (1993) die Frequenz der neuronalen Entladungen im Rindenbereich normalerweise im 40-Hz-Bereich. LLINAS (1993) nimmt an, dass diese neuronalen Schwingungsmuster wichtige Voraussetzungen seien, um sensorische Informationen in kortiko-thalamokortikale Netzwerke hineinzulassen. Werden die inneren Oszillationsraten verlangsamt, z.B. wegen einer Veränderung des ZNS während der Embryonalentwicklung, so ist es nach der Auffassung von LLINAS (1993) unmöglich, sensorische oder motorische Informationen zu verarbeiten, die im Zehn-Millisekunden-Bereich ablaufen, d.h. die Störung, von der sprach- oder 6 Diskussion 118 lesegestörte Kinder betroffen sind. LLINAS (1993) bemerkt, dass seine Hypothese sehr gut erklären könnte, warum Legasthenikern eine geeignete Triggerung oder Neueinstellung von Frequenzen im 40-Hz- bzw. 10-Hz-Bereich fehlt. Ergebnisse aus früheren Untersuchungen (McGuire 1989) weisen zudem daraufhin, dass es nicht nur in der visuellen und auditiven Verarbeitung, sondern auch in anderen Wahrnehmungssystemen die Unterteilung in rasche und langsame Kanäle gibt, z.B. für Berührungsreize. Dann wäre auch zu erwarten, dass legasthene Kinder ebenso bei der Wahrnehmung von Melodie, Rhythmus und Bewegungen langsamer sind als andere Kinder. Es stellt sich die Frage, warum Kinder mit solch schweren Störungen der zeitlichen Verarbeitung von Sinnesreizen relativ normal leistungsfähig sind. Der zeitliche Rahmen der Störung liegt im Zehn-Millisekunden-Bereich. Daher kann angenommen werden, dass nur Verarbeitungsprozesse, die in diesem zeitlichen Rahmen ablaufen, beeinträchtigt sind (Phillips 1993). 1HXURDQDWRPLVFKH9HUlQGHUXQJHQ Wie TALLAL (1991) wiesen auch andere Forschungsgruppen neuroanatomische Veränderungen bei Legasthenikern nach. In mehreren Untersuchungen fand eine Arbeitsgruppe um GALABURDA bei Patienten mit Lese-Rechtschreib-Störungen Veränderungen der linksseitigen temporalen Hirnrinde (Galaburda et al. 1994). Primäres und sekundäres Hörzentrum sowie das sensorische Sprachzentrum sind in dieser Region lokalisiert (vgl. 3.2 u. 3.3.6). Bei LRS-Kindem ist die physiologische Asymmetrie zugunsten der linken Hirnhälfte aufgehoben, und wiederholt wurden Abnormitäten der dort gelegenen Hirnwindungen belegt. Im Tierversuch konnten ROSEN (1992) und FITSCH (1994) Hirnabnormitäten erzeugen, die denen von LRS-Patienten entsprechen. Die Tiere zeigten im Erwachsenenalter Defizite bei der zeitlichen Auflösung auditiver Reize. 6 Diskussion 119 'LH&RQWUD3RVLWLRQ Sind TALLAL, TALCOTT und andere Wissenschaftler (s. 6.2.2.) davon überzeugt, dass eine nichtsprachlichen Legasthenie visuellen und durch Störungen akustischen der elementaren Diskriminationsfähigkeiten verursacht werde, vertreten alternativ hierzu andere Forscher die Meinung, dass legasthenische Defizite auf Störungen höherer Ebenen der Sprachverarbeitung beruhen würden. Zur genauen Darstellung der unterschiedlichen Positionen wird das von FRITH und PROBST verwendete ,QIRUPDWLRQVYHUDUEHLWXQJ 6FKHPD GHU gelegt (Frith zugrunde http://www.rrz.uni-hamburg.de 18.9.2001): SKRQRORJLVFKHQ 1986, PROBST Phonologische Informationsverarbeitung ist die Nutzung lautsprachlicher Informationen beim Erlernen von gesprochener und geschriebener Sprache. FRITH (1986) entwickelte ein Dreiphasenmodell für den Lese-Rechtschreib-Prozess. Im ORJRJUDSKHPLVFKHQ 6WDGLXP ist die Worterkennung an visuellen und auditiven Merkmalen orientiert. Beim Lesen besteht die Konsequenz des Wiedererkennens in der Zuordnung der phonologischen Repräsentation. Wenn detailliertere Merkmale wahrgenommen werden können, nutzen Kinder diese Strategie auch für das Schreiben. Nach TALLAL und TALCOTT ist dieses Stadium bei LRS in erster Linie betroffen. Im nachfolgenden DOSKDEHWLVFKHQ 6WDGLXP werden die Graphem- Korrespondenz-Regeln genutzt, d.h. ein Wort wird Buchstabe für Buchstabe erlesen (phonologisches Rekodieren), und im RUWKRJUDSKLVFKHQ6WDGLXP werden die Wörter innerlich repräsentiert, d.h. sie werden im Gedächtnis gespeichert und sind abrufbar. Die orthographische Strategie wird wie die logographemische zuerst für das Lesen gebraucht. Hierbei werden Wörter in orthographische Einheiten, z.B. in Morpheme oder häufig vorkommende Buchstabenfolgen, gegliedert und deshalb schneller erfasst. Nach Erreichen eines gewissen Niveaus dieser Strategie für das Lesen nutzen Kinder sie auch für das Schreiben. 6 Diskussion 120 Die Begründung für die Darstellung des Schriftspracherwerbs in Stufen liegt für FRITH in der Tatsache, dass die kognitive Entwicklung in Phasen oder Stufen verläuft und nicht kontinuierlich (Küspert 1998, Scheerer-Neumann 1998). Viele Autoren (Günther 1998, Eisenberg, 1995, Steinweger 1999) sind der Auffassung, dass eine XQ]XUHLFKHQGH %HKHUUVFKXQJ GHU 3KRQHP*UDSKHP =XRUGQXQJ eine entscheidende Rolle bei der Entwicklung einer LRS spielt. Insbesondere das Schreiben ist betroffen. Denn nach der Phonemanalyse besteht der weitere Weg der Teilhandlungen beim Schreiben in der Umsetzung der gehörten Laute in die konventionelle Schrift (Steinweger 1999). So muss beim Schreiben jedem Laut ein Graphem zugeordnet werden, und dieser Schritt kann Probleme mit sich bringen, da für einzelne Laute mehrere Realisierungsmöglichkeiten existieren. Der Laut [k] wird beispielsweise schriftlich dargestellt mit K (Kind), CK (Ecke), C (Comic), CC (Mocca), CH (Chlor), [k] und [s] in manchen Wörtern mit X (Hexe), [k] im Auslaut in deutschen Wörtern mit G (Lauf weg!) (Möckel 1997). Die Zeichenfolge im Schriftsystem entspricht der Lautfolge. Wichtig ist dabei das Schreiben der Grapheme von links nach rechts entsprechend der Phonemfolge im gesprochenen Wort. Der letzte Schritt in der Umsetzung des gesprochenen Wortes in die Schrift ist das Zu-Ende-bringen des aufzuschreibenden Wortes. Das letzte Phonem im Wort sagt, welches das letzte Graphem ist, das ganz rechts am Ende des Wortes realisiert werden muss (Möckel 1997). Das dann geschriebene Wort genügt nicht unbedingt den orthographischen Ansprüchen. Beim Schreiben wird nur darauf geachtet, dass die Lautzeichen in der richtigen Reihenfolge bis zum letzten Laut des Wortes geschrieben werden (Möckel 1997). Die Verschriftung der Lautsprache – aus didaktischer Sichtweise von SASSENROTH (1998) – erfolgt nach bestimmten Prinzipien: • phonematisches Prinzip: Versuch, die akustische Gestalt eines Wortes genau wiederzugeben 6 Diskussion • 121 morphematisches Prinzip: Regel, dass ein und dasselbe Morphem, d.h. die kleinste bedeutungstragende Einheit, trotz lautlicher Veränderung immer gleich geschrieben wird • semantisches Prinzip: Regel, dass lautgleiche Begriffe unterschiedlich verschriftet werden • grammatisches Prinzip: Regelung der Groß- und Kleinschreibung • historisches Prinzip: Verschriftung aufgrund der früheren Sprechweise • graphisch-formales Prinzip: Regel der Verschriftung, die der besseren Lesbarkeit dient Das phonematische Prinzip gilt als das wichtigste Prinzip. Es beinhaltet die lautgetreue Verschriftung (Eichler 1992). Die Schwierigkeit, das phonematische Prinzip zu realisieren, beruht darauf, dass es keine 1:1-Beziehung zwischen Phonemen und Graphemen gibt. Diese Vieldeutigkeit birgt für die Kinder, die schreiben lernen, Probleme. Nach MÖCKEL (1997) gelten bei der Phonem-Graphem-Korrespondenz folgende Basisregeln: • Jedem Phonem eines gesprochenen Wortes entspricht ein Graphem. • Die Reihenfolge der Phoneme im gesprochenen Wort entspricht der Reihenfolge der Grapheme im geschriebenen Wort. • Sämtliche Phoneme eines gesprochenen Wortes werden verschriftlicht (Möckel 1997). • Wichtig bei der Phonem-Graphem-Korrespondenz ist das Verstehen des Zuordnungscodes. Die mangelnde Erfassung der regelhaften Zuordnungssysteme stellt also eine Schwierigkeit beim Schrifterwerb dar. Somit ist das Erlernen des Lesens und des Schreibens von einer Vertiefung der Einsicht in den Aufbau der Sprache und in die Regelmäßigkeiten der Abbildung der Sprache durch die Schrift abhängig, einer Entwicklung, die in Stufen erfolgt. Dies erklärt die These, dass eine Legasthenie in der mangelnden Ausbildung 6 Diskussion 122 unterschiedlicher Teilfertigkeiten begründet ist (Klicpera u. Gasteiger-Klicpera 1995). Es stellt sich zudem die Frage, ob legasthene Kinder GHQhEHUJDQJLQGLHLQGHQ (QWZLFNOXQJVPRGHOOHQ EHVFKULHEHQH QlFKVWH 3KDVHQLFKWEHZlOWLJHQ können. Bei großen Schwierigkeiten wäre der Wechsel von der logographischen zur alphabetischen Strategie, bei geringeren der Übergang von der alphabetischen zur orthographischen Strategie betroffen. Am Anfang des Schreibenlernens ist der Übergang von der logographischen zur alphabetischen Stufe von besonderem Interesse (Probst http://www.rrz.uni-hamburg.de 18.9.2001). Sind TALLAL und TALCOTT davon überzeugt, dass eine Legasthenie auf einer grundlegenden sprachunspezifischen Wahrnehmungsstörung beruhe (Tallal 2000, Talcott 2000), gehen andere Wissenschaftler (s.u.) davon aus, dass LRS durch Verarbeitungsstörungen sprachlicher Elemente verursacht werde, d.h. durch eine Einschränkung der phonologischen Informationsverarbeitung. Zur genaueren Darstellung dieser These sei zunächst der Aufbau der phonologischen Informationsverarbeitung erklärt. Sie wird in drei Teilgebiete gegliedert (Probst http://www.rrz.uni-hamburg.de 18.9.2001): Die SKRQRORJLVFKH %HZXVVWKHLW (siehe auch 5.2.2) bezeichnet die Fähigkeit, sprachliche Einheiten (Worte, Reime) zu erkennen und mit ihnen zu operieren. Sie erfährt noch eine weitere Unterteilung nach SKOWRONEK und MARX (Blässer 1984). Bei der phonologischen Bewusstheit im weiteren Sinne geht es um das Erkennen von größeren Spracheinheiten, wie z.B. Wörtern, Silben und Reimen. Bei der phonologischen Bewusstheit im engeren Sinne geht es um das Erkennen von kleinen lautsprachlichen Einheiten wie z.B. die Phonemersetzung (wenn man /a/ durch /i/ in dem Wort „ Wand“ ersetzt) oder die Phonemanalyse ( welche Laute hört man in dem Wort „ Uhr“ ). Die phonologische Bewusstheit von Kindern mit LRS ist meist mangelhaft ausgebildet. So gelingt es ihnen z.B. oft nicht, Phoneme in einem Wort zu hören und zu identifizieren. 6 Diskussion 123 Beim SKRQRORJLVFKHQ 5HNRGLHUHQ während des Zugriffs auf das semantische Gedächtnis geht es um die Fähigkeit, schriftliche Symbole (geschriebene Wörter) zu rekodieren (in die lautsprachliche Struktur zu übertragen), um deren Bedeutung aus dem Langzeitgedächtnis abzurufen. Bei Kindern mit LRS ist die Geschwindigkeit, mit der auf das semantische Gedächtnis zurückgegriffen wird, niedrig: Sollen Wörter benannt werden, dauert dieses länger, als bei Kindern ohne LRS (Probst http://www.rrz.uni-hamburg.de 18.9.2001). Das SKRQHWLVFKH 5HNRGLHUHQ im Arbeitsgedächtnis betrifft das Kurzzeitgedächtnis. In ihm sind schriftliche Symbole lautsprachlich repräsentiert, d. h. ein Wort wird Buchstabe für Buchstabe erlesen und dann zu einem vollständigen Wort verbunden. Das Arbeitsgedächtnis ist also für die Artikulationsgeschwindigkeit und Genauigkeit zuständig. Auch in diesem Bereich zeigen legasthene Kinder deutlich schwächere Leistungen. Mehrere Wissenschaftler gehen davon aus, dass LRS auf einer Einschränkung der phonologischen Informationsverarbeitung beruht, d.h. insbesondere auf einer verringerten Möglichkeit, Buchstaben- bzw. Wortbilder zu erkennen (d.h. auf einer Störung der phonologischen Bewusstheit) bzw. auf einem mangelhaftem Zugriff auf das semantische und das Arbeitsgedächtnis (d.h. auf Störungen des phonologischen bzw. phonetischen Rekodierens) beruht. Die Wiener Längsschnittuntersuchung von KLICPERA und GASTEIGERKLICPERA (1993) ergab, dass die Komponenten der phonologischen Informationsverarbeitung reliable Prädiktoren für die Ausbildung des Lesens und Schreibens sind: Sind diese gut ausgebildet, dann ist es wahrscheinlich, dass sich keine Schwierigkeiten im Lesen und Schreiben entwickeln. Auch neuroanatomische Befunde zeigten hirnelektrische Auffälligkeiten in den linkshemisphärischen Regionen, die mit der sprachlichen Informationsverarbeitung verknüpft sind. KLICPERA und GASTEIGERKLICPERA kommen in ihrer Untersuchung zu dem Ergebnis, dass die primäre 6 Diskussion 124 Ursache der LRS in der Störung des Worterkennungsvorgangs liegt. Diese beruhe auf einer Störung von Kodierungsprozessen, die vor allem auch bei Gedächtnisaufgaben bemerkbar sind (Klicpera u. Gasteiger-Klicpera 1993). Ähnlich meint VELLUTINO, dass die Lese-Rechtschreib-Schwäche auf eine beschränkte Fähigkeit, Sprache zum Verschlüsseln nichtsprachlicher Information zu verwenden, zurückzuführen ist (Vellutino 1987). Danach wäre Legasthenie ein „ unterschwelliger Sprachfehler", der in phonetischen Kodierungsstörungen (der Unfähigkeit, die Lautgestalt eines Wortes wieder verfügbar zu repräsentieren, um es sich besser merken zu können), in fehlerhafter phonemischer Segmentierung (der Unfähigkeit Wörter in Sprachlaute zu zerlegen) und in einem unterentwickeltem Wortschatz sowie Schwierigkeiten, Wörter und Sätze grammatisch zu unterscheiden, wurzelt. Viele Wissenschaftler sind der Ansicht, dass insbesondere eine Störung der phonologischen Bewusstheit die Ursache der Lese-Rechtschreib-Störung sei. Im Gegensatz zu TALLAL und TALCOTT, die der Auffassung sind, dass eine Wahrnehmungsstörung nichtsprachlicher Reize die Legasthenie verursacht, gehen diese Forscher somit davon aus, dass ein höheres und komplexeres Verarbeitungsniveau der Sprachverarbeitung betroffen sei. Trotz der Tatsache, dass die phonologische Bewusstheit erst Anfang der 80er-Jahre in das Feld der Forschung rückte und somit noch relativ neu ist (Küspert 1998), gibt es viele Längsschnittstudien, die einen Zusammenhang zwischen metalinguistischen Fähigkeiten – speziell der phonologischen Bewusstheit – und dem späteren Schriftspracherwerb nachweisen (Küspert, 1998). Zu nennen sind die Untersuchungen von LIBERMAN (Liberman 1966) und SHAYWITZ (Shaywitz 1996), die korrelativen Längsschnittstudien von BRADLEY und BRYANT (1985), von LUNDBERG, OLOFSSON und WALL (1980) sowie von LANDERL, LINORTNER und WIMMER (1992). 6 Diskussion 125 LIBERMAN ist der Auffassung, dass Lesen auf einer bewussten Ebene erlernt werden müsse (Liberman 1966). Die Aufgabe des Lesers sei es nämlich, visuelle Wahrnehmungen in Gesprochenes umzuwandeln, d.h. Graphemen die entsprechenden Phoneme zuzuordnen. Damit dies erreicht werden könne, müsse der Leser zunächst die innere phonologische Struktur des gesprochenen Wortes bewusst wahrnehmen, d.h. er müsse über die Fähigkeit der phonologischen Bewusstheit verfügen. Als Nächstes müsse er einsehen, dass die Orthographie des Wortes dessen Phonologie repräsentiere. Dieser Vorgang finde statt, wenn ein Kind lesen lerne. Ist ein Kind legasthenisch, so beeinträchtige ein Defizit innerhalb des sprachverarbeitenden Systems auf phonologischer Ebene die Fähigkeit, das geschriebene Wort in die zugrunde liegenden phonologischen Komponenten zu zergliedern. Obwohl Worte im Prinzip verstanden werden können, kann diese Fähigkeit nicht genutzt werden, weil die Worte nicht identifiziert werden können. Auch SHAYWITZ vertritt die Ansicht, dass es einen Zusammenhang zwischen der phonologischen Bewusstheit und der Fähigkeit des Lesens und Schreibens geben würde (Shaywitz 1996). Beginnend in 445 zufällig ausgewählten Kindergärten, führte sie von 1983 bis 1996 eine Längsschnittstudie durch, die die Kinder bezüglich ihrer Lese-Rechtschreib-Fähigkeiten untersuchte. Bei 20% der Schulkinder wurde eine Legasthenie festgestellt. Diese Prozentzahl stimmt mit dem Ergebnis einer früheren von LIBERMAN durchgeführten Studie überein (Liberman 1976). Sie zeigte, dass die phonologische Struktur eines Wortes den meisten Kindern in einem Alter von vier bis sechs Jahren bewusst wird. In dem Versuch wurden Kinder befragt, wie viele Phoneme sie in einer Folge von Worten hörten. Keines der Vierjährigen konnte diese Aufgabe korrekt lösen, jedoch 17% der Fünfjährigen und 80% der Sechsjährigen. Ein Vergleich der beiden Studien unterstützt nach der Auffassung von SHAYWITZ die These, dass ein Zusammenhang zwischen phonologischer Rechtschreib-Fähigkeit bestehen würde. Bewusstheit und der Lese- 6 Diskussion 126 BRADLEY und BRYANT (Bradley u. Bryant 1980) stellten fest, dass die Schulung der phonologischen Bewusstheit deutlich die Lese-RechtschreibFähigkeit eines Kindes verbessert. In ihrer über einen Zeitraum von drei Jahren durchgeführten Studie wurde in einer Gruppe die phonologische Bewusstheit trainiert, in der anderen Gruppe Übungen durchgeführt, die nicht auf den Lautstrukturen von Worten beruhte. BRADLEY und BRYANT wiesen einen Zusammenhang zwischen der phonologischen Bewusstheit im weiteren Sinne und dem späteren Schriftspracherwerb nach. Untersucht wurden 368 Kindergartenkinder im Alter von vier bis fünf Jahren, die über keinerlei Lese- oder Schreibkompetenz verfügten, in Bezug auf ihre Fähigkeit, Laute zu kategorisieren. Sie sollten hierbei Reime (z.B. Endreim: men – red – bed – fed), Alliterationen (z.B. Wortanfangsalliteration: peg – pen – well – pet) und gleiche Phonemgruppen im Wortinneren (z.B. lot – pot – cot – hat) erkennen. Mit Hilfe standardisierter Tests wurden drei Jahre später die Lese- und Rechtschreibfähigkeiten der Kinder überprüft. Um die Gültigkeit dieser Tests zu erhärten, wurde außerdem auf einen Mathematiktest und einen Intelligenztest für die allgemeinen kognitiven Fähigkeiten zurückgegriffen. Die Lautkategorisierungsaufgabe wies die größte Korrelation hinsichtlich der beiden Lesetests auf, gefolgt von der Korrelation zum Rechtschreibtest. Der Mathematiktest korrelierte am wenigsten (Küspert 1998). Trotz der starken Bedeutung von Faktoren wie Intelligenz und Erziehung für die Vorhersage späterer Leistungen im Bereich der Schriftsprache wiesen die Ergebnisse auf die phonologische Bewusstheit als einen wichtigen spezifischen Faktor für die Prognose der späteren Lese- und Rechtschreibleistungen hin. Allerdings muss bei dieser Studie bedacht werden, dass die phonologische Bewusstheit nur durch Lautkategorisierungsaufgaben nachgewiesen wurde, denn weitere Variablen hätten die Bedeutung der Lautkategorisierung um ein Weiteres reduziert (Küspert 1998). 6 Diskussion 127 Die Untersuchungen von LUNDBERG, OLOFSSON und WALL (1980) berücksichtigten diesen Aspekt. Sie prüften durch Reimaufgaben, Silbensegmentierung und Silbensynthese die phonologische Bewusstheit im weiteren Sinn, durch Phonemsegmentierung und Phonemsynthese sowie Phonemvertauschung und Lokalisation eines Phonems in einem Wort die phonologische Bewusstheit im engeren Sinn. Sie untersuchten bei ihrer Längsschnittstudie 143 Vorschulkinder im Durchschnittsalter von etwa sieben Jahren. Am Ende des ersten und zu Beginn des zweiten Schuljahres wurden Aussagen über die Intelligenz, die Lese- (hier das stille Lesen) und die Rechtschreibleistung der Kinder getroffen. Im Hinblick auf den Zusammenhang zwischen der phonologischen Bewusstheit im Vorschulalter und der späteren Leseleistung fielen die Ergebnisse unterschiedlich aus. Die Aufgaben zur phonologischen Bewusstheit im engeren Sinn wiesen eine größere Korrelation zur späteren Leseleistung auf als die phonologische Bewusstheit im weiteren Sinn. Die Leistungen bei Aufgaben zur Phonemenvertauschung und der Reimproduktion galten als verlässliche Faktoren bei der Vorherbestimmung der späteren Leseleistung (Küspert 1998). Die Ergebnisse dieser Untersuchung veranschaulichen das herrschende Problem der phonologischen Bewusstheit. Diese nimmt eine bedeutende Rolle in Bezug auf den Schriftspracherwerb ein, der kausale Zusammenhang ist aber nicht geklärt. Die Experten scheiden sich bei der Beantwortung der Frage, ob phonologische Bewusstheit als Voraussetzung für den Schriftspracherwerb gilt oder Folge von diesem ist (Steinweger 1999). WAGNER und TORGESEN untersuchten 1987 die Daten von LUNDBERG, OLOFSSON und WALL und kamen zu dem Ergebnis, dass die im Vorschulalter vorhandenen Schriftsprachfertigkeiten eine bedeutende Position einnehmen (Wagner u. Torgesen 1987). Denn bei Konstanthaltung der anfänglichen Lesekenntnisse verfehlten die Korrelationen zwischen phonologischer Bewusstheit und Leseleistung die Signifikanzgrenze (Küspert 1998). 6 Diskussion 128 1992 untersuchten LANDERL et al. in zwei Studien die Bedeutung der phonologischen Bewusstheit für den Schriftspracherwerb im Deutschen (Landerl et al. 1992). Bei der ersten Untersuchung wurden Kindern zu Anfang des ersten Schuljahres zwei Aufgaben zur phonologischen Bewusstheit gestellt – eine Aufgabe zur Erkennung von Alliterationen und Reimen und eine Aufgabe zur Ersetzung von Lauten. Am Ende des ersten Schuljahres wurde die Lese- und Rechtschreibfähigkeit überprüft. Im Gegensatz zu der Alliterations- und Reimaufgabe, die keinen deutlichen Zusammenhang zu der späteren Lese- und Rechtschreibleistung erkennen ließ, zeigte die Lauterkennungsaufgabe einen Zusammenhang zu der späteren Fähigkeit im Lesen und Schreiben. Allerdings wurde ebenfalls festgestellt, dass auch die Kinder, die bei der Aufgabe zur Lauterkennung anfangs schlecht abgeschnitten hatten, keine wirklichen Schwierigkeiten beim Lese- und Schrifterwerb entwickelten. Die zweite Studie untersuchte am Ende des ersten Schuljahres, ob es Unterschiede in der phonologischen Bewusstheit bei durchschnittlichen und schlechten Lesern gab. Während bei der Reimerkennungsaufgabe keine Unterschiede in den Leistungen beider Gruppen vorlagen, zeigte sich, dass bei der Lauterkennungsaufgabe die normalen Leser den schlechten deutlich überlegen waren. Dieses Ergebnis lässt den Schluss zu, dass nicht die Fähigkeit, Alliterationen und Reime zu erkennen, also mit Silben zu operieren, sondern die Operation mit Einzellauten in einem engen Zusammenhang mit der Schriftsprache steht (Küspert 1998). Bei den Studien wurden die rudimentären Schriftsprachkenntnisse der Kinder nicht berücksichtigt. Das lässt die Annahme zu, dass tatsächlich die Einsicht in die phonologische Struktur der Sprache als Konsequenz der Einsicht in das alphabetische Prinzip der Schriftsprache entstanden sein könnte (Küspert 1998). Die Einseitigkeit der Längsschnittstudien ist zu kritisieren. Bis auf die Reanalyse von WAGNER und TORGESEN wurde nur die Bedeutung der phonologischen Bewusstheit für den Schrifterwerb untersucht, welchen Einfluss der Schrifterwerb 6 Diskussion 129 auf die phonologische Bewusstheit nimmt, wurde dagegen nicht berücksichtigt (Steinweger 1999). Die hier als Beispiel aufgeführten Studien für die korrelativen Längsschnittstudien verdeutlichen, dass der SKRQRORJLVFKHQ %HZXVVWKHLW offenbar eine wichtige Rolle für den Schriftspracherwerb zukommt. Störungen des phonologischen bzw. phonetischen Rekodierens werden durch '\VIXQNWLRQHQGHV*HGlFKWQLVVHV verursacht. Bei Legasthenikern wurde ein zu geringer Zeitaufwand für das Memorieren schriftsprachlicher Informationen sowie ungenügende Wiederholungsstrategien beobachtet. EGERER konnte eine Schwäche des Kurzzeitgedächtnisses bei legasthenen Kindern feststellen (Egerer 1988). Auch MILZ sieht die Störungen des Gedächtnisses und speziell des Kurzzeitgedächtnisses als mögliche Ursache der Legasthenie (Milz 1989). Somit zeigen zahlreiche Untersuchungen, dass eine korrekte phonologische Informationsverarbeitung erforderlich ist, um ein fehlerfreies Lesen und Schreiben zu erlernen. Dies widerlegt aber die These von TALCOTT und TALLAL, dass eine Lese-Rechtschreib-Störung durch Störungen der elementaren nichtsprachlichen visuellen und akustischen Diskriminationsfähigkeiten verursacht werde, nicht eindeutig. Um eine phonologische Bewusstheit bzw. phonetisches und phonologisches Rekodieren zu erlernen, muss offensichtlich eine niedrigere Ebene der Sprachverarbeitung, nämlich das Erkennen nichtsprachlicher Reize, korrekt funktionieren. Es ist evident, dass bei einer Wahrnehmungsstörung nichtsprachlicher Reize im Sinne von TALLAL und TALCOTT keine phonologische Bewusstheit erreicht werden kann. Ob allerdings immer Wahrnehmungsdefizit das einer von LRS TALLAL zugrunde postulierte liegt, wird spezifische von einigen Wissenschaftlern bezweifelt. In diesem Zusammenhang führten SABISCH und FERSTL eine Studie durch (Sabisch u. Ferstl http://www.uni.leipzig.de 3.10.2001). Ziel dieser Studie war es, Kinder einer Leipziger Sprachheilschule mit 6 Diskussion 130 einer Diskriminationsaufgabe zu testen, um zu evaluieren, wie häufig dieses basale Wahrnehmungsdefizit auftritt. Zwölf Versuchspersonen im Alter von acht bis zehn Jahren wurden aus einer Gruppe von 42 Kindern ausgewählt. Sie erfüllten das Kriterium eines Handlungs-IQ von über 85 bei schlechterem verbalen IQ. Um diese Werte zu bestimmen, verwendeten SABISCH und FERTL den HAWIK-R-Intelligenztest. Zusätzlich wurden die Sprachfähigkeiten mit Hilfe des Heidelberger Sprachentwicklungstests, die Leseleistung mit dem Züricher Lesetest, und die phonologischen Fähigkeiten mit Hilfe einer eigens entwickelten Spoonerism-Aufgabe erfasst. Zwölf weitere Kinder, die nach Alter und Handlungs-IQ den Kindern der Versuchsgruppe entsprachen, aber einen höheren verbalen IQ aufwiesen bildeten die Kontrollgruppe. Für die auditorische Wahrnehmungsaufgabe lernten die Kinder zuerst die Zuordnung von zwei komplexen Tönen (100 Hz und 305 Hz) zu zwei Maustasten. In der Diskriminationsaufgabe wurden dann zehn verschiedene Interstimulusintervalle von 30 ms bis 428 ms in je acht Durchgängen geprüft. Die Ergebnisse zeigten, dass die Kontrollgruppe die Aufgabe signifikant besser bewältigte als die Versuchsgruppe. Ein Einfluss des Interstimulusintervalls konnte jedoch nicht gefunden werden. Sogar bei dem kürzesten Intervall von 30 ms konnten alle Kinder der Versuchsgruppe mehr als 75% der Folgen richtig nachtippen. Partielle Korrelationen, in denen die Gruppe ausfaktorisiert wurde, zeigten weiterhin, dass die auditorische Diskriminationsfähigkeit mit Sprachleistungen in Beziehung stand. Lesefähigkeit, allgemeine Sprachfähigkeit und phonologische Fähigkeiten korrelierten positiv mit der Diskriminationsleistung. Nach diesen Ergebnissen weisen offenbar nicht alle Kinder mit entwicklungsbedingten Sprachstörungen das von TALLAL postulierte spezifische Wahrnehmungsdefizit auf, nämlich dass Leseschwache dann Schwächen in der Wahrnehmung von nichtsprachlichen Reizen aufweisen, wenn diese in einem sehr kurzen zeitlichen Abstand nacheinander präsentiert werden. Dennoch wurde auch 6 Diskussion 131 in der Studie von SABISCH und FERTL (2001) ein Zusammenhang zwischen Sprachleistungen und der Diskrimination von nichtverbalen auditorischen Reizen gefunden. 6FKOXVVIROJHUXQJ Phonologische Informationsverarbeitung, die Nutzung von lautsprachlichen Informationen bei dem Erlernen von gesprochener und geschriebener Sprache, ist erforderlich, um Lesen und Rechtschreibung zu erlernen. Hierzu ist es notwendig, die Phonem-Graphem-Zuordnung sicher zu beherrschen. Auch sind der Erwerb der phonologischen Bewusstheit sowie ein richtiges phonologisches und phonetisches Decodieren für das Erlernen gesprochener und geschriebener Sprache unabdingbar. Aber der Wahrnehmung sprachlicher Elemente liegt das richtige Erkennen nichtsprachlicher Laute zugrunde. Werden Stopp-Konsonanten wie /b/, /d/ oder /p/, die nur 40 Millisekunden lang schwingen, nicht erkannt, so ist das richtige Erlernen von Lesen und Rechtschreibung unmöglich. Können Kinder schnelle akustische Signale nicht auseinander halten, so überlappen sich diese Einheiten, und das Gehörte im Gehirn führt zu Sprachstörungen. Möglicherweise kann eine LRS auch noch auf anderen Wahrnehmungsstörungen nichtsprachlicher Elemente beruhen (Sabisch u. Ferstl http://www.uni.leipzig.de 3.10.2001). Zudem ist ungeklärt, ob die Legasthenie tatsächlich ein eigenständiges pathologisches Phänomen ist. So haben TALCOTT et al. (2000) mittlerweile deutliche Hinweise dafür gefunden, dass dies nicht zutrifft. Für ihre visuellen und akustischen Tests wählten TALCOTT et al. Kinder aus, bei denen keine Anzeichen einer Leseschwäche zu beobachten waren. Die visuelle Aufgabe 6 Diskussion 132 bestand wiederum darin, die Richtungsänderung bewegter Punkte auf einem Bildschirm zu erkennen. Bei dem akustischen Test sollten die Kinder angeben, welcher von zwei kurz hintereinander erzeugten Tönen eine Modulation aufwies. Solche raschen Änderungen der Tonhöhe müssen auch beim Dekodieren der gesprochenen Sprache registriert werden. Die Kinder, die beim visuellen Leistungstest schlecht abschnitten, vermochten nur schwer zwischen richtig und falsch buchstabierten Wörtern zu unterscheiden. Eine verringerte Fähigkeit, die raschen Schwankungen der Tonhöhe zu identifizieren, ging dagegen mit Schwierigkeiten beim richtigen Aussprechen mancher Wörter einher (Tallal 2000). Das deutet daraufhin, dass Legasthenie nur das ungünstige Ende einer normalen Fähigkeitsverteilung darstellt. =XVDPPHQIKUXQJGHU(UJHEQLVVHDXVXQG PHILLIPS und TALLAL kommen beide zu einem übereinstimmenden Ergebnis: Ursache eines Defizits ist eine Störung im akustischen Timing des Gehirns, die das Erkennen schnell aufeinander folgender Sinnesereignisse unmöglich macht. Insbesondere die Wahrnehmung so genannter Stopp-Konsonanten ist erschwert. Während alle Vokale für wenigstens 100 Millisekunden akustisch schwingen, sind so genannte Stopp-Konsonanten wie /b/, /d/ oder /p/ nur 40 Millisekunden hörbar. Ihr Übergang zu Vokalen wie dem /a/ erfolgt also sehr schnell – zu schnell für Menschen, die legasthen sind oder an einer reinen Worttaubheit leiden. Interessant ist somit, dass ein- und dieselbe nichtsprachliche Wahrnehmungsstörung offenbar die Grundlage zweier verschiedener Krankheiten sein kann, nämlich einer LRS und einer reinen Worttaubheit. Dies weist auf die Bedeutung der Wahrnehmungsstörung hin. Normalerweise kann ein Erwachsener eine korrekte Auskunft geben, wenn ihm über Kopfhörer z.B. ein Ton in das rechte und ein anderer in das linke Ohr 6 Diskussion 133 gegeben wird und wenn beide Töne im Abstand von mindestens 40 Millisekunden aufeinander folgen. Erwachsene mit einer reinen Worttaubheit nach einem Schlaganfall benötigen demgegenüber einen zeitlichen Abstand von mindestens 100 Millisekunden, um ein richtiges Urteil abzugeben. Eine ähnliche Verlangsamung fand man auch bei legasthenen Kindern. Zwar liegt die Ordnungsschwelle, also der nötige zeitliche Abstand zwischen zwei Reizen, bei Kindern im zweiten Schuljahr noch bei 100 bis 120 Millisekunden. Das ist entwicklungsbedingt. Leseschwache Schüler mit erhöhter Ordnungsschwelle aber brauchen etwa 200 und mehr Millisekunden, um sagen zu können, welchen Ton sie zuerst gehört haben (Tallal et al. 2000). Auch andere schnell aufeinander folgende Sinnesereignisse, z.B. optische, sind für Legastheniker oft nur als undifferenzierte Reizkulisse wahrnehmbar. Dieser Defekt, möglicherweise entstanden während der fetalen Entwicklung, (Stein u. Talcott 1999) führt dazu, dass zu wenige für die Sprachentwicklung wichtige Lautreize aus der Umwelt das Gehirn erreichen. Als Antwort auf diese Reize knüpfen die dortigen Nervenzellen phonetische Einheiten, die bestimmten Lauten zugeordnet sind. Können Kinder schnelle akustische bzw. optische Reize nicht auseinander halten, dann überlappen sich die Einheiten, und diese fehlerhafte neuronale Verknüpfung führt zu Sprachstörungen (Tallal 2000). Es ist davon auszugehen, dass auch eine reine Worttaubheit auf einer fehlerhaften bzw. unzureichenden Verknüpfung von Neuronen beruht, die zu einer Wahrnehmungsstörung schnell aufeinander folgender Reize führt (Phillips 1995). =XVDPPHQIDVVXQJXQG6FKOXVVIROJHUXQJ Die Untersuchungen von PHILLIPS weisen daraufhin, dass die reine Worttaubheit eine Manifestation einer mehr grundlegenden zeitlichen Verarbeitungsstörung auf vorsprachlichem Niveau iat, d.h. dass der reinen Worttaubheit eine sprachunspezifische Störung im primären auditorischen Kortex 6 Diskussion 134 zugrunde liegt (Phillips 1997). Eine Anzahl von Hinweisen zeigt an, dass der primäre auditorische Kortex eine Rolle bei der Wahrnehmung, aber nicht bei der linguistischen Verarbeitung von Lauten spielt. Die Genauigkeit des Antwortverhaltens der kortikalen Neuronen reichte in den Untersuchungen von PHILLIPS aus, um den zeitlichen Verlauf einzelner Sprachkomponenten darzustellen. Dies lässt darauf schließen, dass der primäre auditorische Kortex bei der Unterscheidung verschiedener Sprachabschnitte eine Rolle spielt. Die Ergebnisse zeigen zudem, dass die Antworten auf Reize im Kortex eine Genauigkeit des Antwortverhaltens zeigten, die nur etwas schlechter als die der Fasern des Kochlearnervs ist (Phillips 1990). Eines der wichtigsten diagnostischen Kriterien der Genauigkeit, mit der Nervenzellen auf Töne oder Geräusche reagieren ist die Prüfung, inwieweit die Frequenzlücke (4.2.2.1) erkannt werden kann. Die wichtigsten „ Frequenzlücken“ der Sprache sind die phonetisch bedeutsamen – sie lassen Stopp-Konsonanten erkennbar werden (Phillips 1990). Nicht alle Bereiche des Sprachunterscheidungsvermögens sind bei der reinen Worttaubheit gleichermaßen betroffen. Insbesondere wurde festgestellt, dass das Wahrnehmungsvermögen langer Vokale bilaterale Läsionen des primären auditorischen Kortex übersteht, jedoch das von Stopp-Konsonanten, nicht. Die gestörte Unterscheidungsfähigkeit von Stopp-Konsonanten bei Worttaubheit zeigt ein zeitliches Verarbeitungsdefizit an, das Stopp-Konsonanten betrifft, aber Vokale nicht. Dies liegt im Bereich von einer bis zu zehn Millisekunden und charakterisiert die Stopp-Konsonanten phonetisch (Phillips 1990). Die Untersuchungen von PHILLIPS wurden von mehreren Studien über das Spracherkennungsvermögen bestätigt. Wie bei PHILLIPS wurden Untersuchungen durchgeführt, die die Erkennung der Frequenzlücke bestimmten. Sie bestätigten die Ergebnisse von PHILLIPS, dass bei reiner Worttaubheit die Erkennung der Frequenzlücke vermindert ist (Albert u. Bear 1974, Hendler et al. 1990, Schwarz u. Tomlinson 1990). Bei Patienten mit reiner Worttaubheit wurde zudem deren Fähigkeit getestet, Phoneme zu identifizieren. Wie bei PHILLIPS 6 Diskussion wurde 135 festgestellt, dass die Patienten größere Schwierigkeiten hatten, Verschlusslaute zu identifizieren als Vokale (Eggermont 1991). Mehrere Untersuchungen bestätigen die Ergebnisse von PHILLIPS, dass grundlegende auditive Wahrnehmungsstörungen Ursache der reinen Worttaubheit sein können (Baddeley u.Wilson 1976, Saffran et al. 1976). Andere Wissenschaftler – insbesondere Paula TALLAL sowie Joel TALCOTT – beschreiben das reibungslose Funktionieren der optischen und akustischen Sinneswahrnehmungen als unumgängliche Voraussetzung für das erfolgreiche Erlernen des Schreibens und Lesens. TALCOTT und STEIN hatten festgestellt, dass etwa 75 Prozent der legasthenen Kinder in Tests schwächer abschneiden, die eine schnelle visuelle Informationsverarbeitung verlangen (Stein u. Talcott1999). Zudem entdeckten TALCOTT et al., dass erwachsene legasthene Versuchspersonen auf langsame oder kontrastreiche visuelle Reize genauso gut reagierten wie gute Leser. Offenbar steht die Legasthenie in Verbindung mit Normabweichungen neuroanatomischer und hirnfunktioneller Merkmale, wobei der M-Bahn eine besondere Bedeutung zukommt. Es gibt Hinweise dafür, dass diese von Magnozellen gebildete Bahn bei Legasthenikern weniger leistungsfähig ist (Tallal 2000). STEIN und TALCOTT stellten die Hypothese auf, dass bei Legasthenikern – möglicherweise infolge einer Immunreaktion der Mutter – während der fetalen Entwicklung junge Nervenzellen im Gehirn geschädigt würden. In Folge sei ihr Zusammenwachsen behindert und die Ausbildung der M-Bahn von der Netzhaut zum lateralen Corpus geniculatum und weiter zum visuellen Kortex gestört (Stein u. Talcott 1999). TALLAL et al. stellten eine zeitliche Verlangsamung der Hörverarbeitung bei Legasthenikern fest. Unter anderem äußerte sich diese in einer mangelhaften „ Filterfunktion“ des Gehörs: Einen bestimmten Sprecher aus einem „ akustischen Salat“ von Störgeräuschen und anderen Sprechern herauszuhören und zu verstehen stellt für sie eine besondere Schwierigkeit dar (Tallal et al. 1985). 6 Diskussion 136 Zudem nehmen TALLAL et al. (1993) einen Zusammenhang zwischen visuellen und auditiven Problemen an. Sowohl die Unterscheidung von Sprachlauten im gesprochenen Wort als die von Buchstabenkombinationen des geschriebenen Wortes erfordern eine schnelle zeitliche Verarbeitung. Die Untersuchungen von TALLAL et al. (1993) unterstützen die Hypothese, dass Leseschwache dann Schwächen in der Wahrnehmung von nichtsprachlichen Reizen aufweisen, wenn diese in einem sehr kurzen zeitlichen Abstand nacheinander präsentiert werden. In diesen Untersuchungen wurden leseschwache Kinder mit nichtleseschwachen Kindern anhand von Aufgaben zum Erkennen der Reihenfolge von zwei Tönen bzw. zwei komplexen Tonkombinationen verglichen (Tallal 1980). Wie TALLAL und TALCOTT konnten auch andere Wissenschaftler bei Legasthenikern Störungen der Hör- (Warnke u. Remschmidt 1992) und Sehverarbeitung (Manis et al. 1997) nachweisen. Es wurden bei Legasthenie zudem Störungen der phonologischen Informationsverarbeitung, d.h. der Nutzung von lautsprachlichen Informationen beim Erlernen von gesprochener und geschriebener Sprache nachgewiesen. Um eine phonologische Informationsverarbeitung sicher zu bearbeiten, ist es notwendig, eine Phonem-Graphem-Zuordnung zuverlässig zu beherrschen. Auch sind der Erwerb einer phonologischen Bewusstheit sowie ein richtiges phonologisches und phonetisches Decodieren für das Erlernen gesprochener und geschriebener Sprache unabdingbar. Dies steht jedoch nicht im Widerspruch zu der These von TALCOTT und TALLAL, dass eine Lese-Rechtschreib-Störung durch Störungen der elementaren nichtsprachlichen visuellen und akustischen Diskriminationsfähigkeiten verursacht werde. Denn um eine phonologische Bewusstheit zu erreichen bzw. phonetisches und phonologisches Rekodieren zu erlernen, muss natürlich eine niedrigere Ebene der Sprachverarbeitung, nämlich das Erkennen nichtsprachlicher Reize, korrekt funktionieren. Es ist offensichtlich, dass bei einer 6 Diskussion 137 Wahrnehmungsstörung nichtsprachlicher Reize im Sinne von TALLAL und TALCOTT keine phonologische Bewusstheit erreicht werden kann. Die Ergebnisse von PHILLIPS und TALLAL stehen in Übereinstimmung miteinander: Ursache eines Defizits ist eine Störung im akustischen Timing des Gehirns, sodass das Erkennen schnell aufeinander folgender Sinnesereignisse unmöglich wird. Somit haben sowohl sensorische Worttaubheit als auch Legasthenie eine gemeinsame Ursache, und dies weist auf die Bedeutung der Wahrnehmungsstörung hin. Die Untersuchungen von PHILLIPS und TALLAL zeigen somit, dass die Rolle der eigentlichen Sprachzentren lange überschätzt wurde und es wahrscheinlich in Zukunft zu einem Paradigmenwechsel kommen wird. 7 Die Konsequenzen für die Therapie 138 'LH.RQVHTXHQ]HQIUGLH7KHUDSLH Aus den vorangegangenen Kapiteln folgt, dass Legasthenie und reine Worttaubheit zumeist keine Sprachverarbeitungsstörungen sind. Offenbar beruhen legasthenische Defizite nicht auf Erkennungsstörungen von Buchstaben- oder Wortbildern bzw. auf einem mangelhaftem Zugriff auf Einheiten des orthografischen Lexikons, sondern es konnte experimentell nachgewiesen werden, dass das Defizit auf einer Verarbeitungsstörung schnell wechselnder elementarer optischer Stimuli (z.B. Richtungsänderungen zweier Punkte) zurückzuführen ist. Ein ähnliches Merkmal lässt sich auch bei der akustischen Wahrnehmung feststellen: Hier besteht eine verminderte Unterscheidungsfähigkeit von zwei schnell aufeinanderfolgenden Tönen bzw. deren Frequenzänderung. Beide Defizite erwiesen sich bei derselben Versuchspopulation als hoch korreliert. Vergleichbare Ergebnisse lieferten neuere Untersuchungen an Patienten, deren akustische Spracherkennungsdefizite als reine Worttaubheit klassifiziert wurden. Offenbar liegt diesem Syndrom eine sprachunspezifische Störung im primären auditorischen Kortex zugrunde: Bestimmte Frequenzspektren akustischer Stimuli können nicht erfasst werden. Diese Untersuchungsergebnisse wecken die Hoffnung, sowohl eine LeseRechtschreib-Schwäche als auch eine reine Worttaubheit durch das Trainieren elementarer sensorischer Fähigkeiten fördern zu können. TALLAL und MERZENICH entwickelten bereits eine Art Computerspiel, das spezifisch die bei der Legasthenie beeinträchtigten Wahrnehmungs- und Unterscheidungsleistungen verbessert (Barinaga 1996). Im Folgenden wird zunächst eine Übersicht über verschiedene therapeutische Verfahren der LRS und der reinen Worttaubheit gegeben und insbesondere das von TALLAL und MERZENICH entwickelte Verfahren beschrieben, sodann auf die Therapie der reinen Worttaubheit eingegangen. Es kann davon ausgegangen werden, dass das Üben elementarer Hörfähigkeiten, beispielsweise der Unterscheidungsfähigkeit schnell aufeinander 7 Die Konsequenzen für die Therapie 139 folgender Töne bzw. deren Frequenzänderung, zu einer deutlichen Besserung der reinen Worttaubheit führen würde. 7KHUDSLHGHU/HVH5HFKWVFKUHLE6FKZlFKH +HUN|PPOLFKH6\PSWRPWKHUDSLH In vielen Therapieeinrichtungen sowie herkömmlichen Nachhilfe- und Förderinstituten, die aufgrund der wachsenden Not betroffener Familien immer zahlreicher werden, wird regelmäßig nach dem Grundsatz „ Mehr desselben, also üben, üben, üben" gearbeitet. Dieser Ansatz missachtet die Bedürfnisse der Betroffenen und ignoriert den heutigen Stand des Wissens auf dem Gebiet der LRS-Forschung. Oft bieten Nachhilfestudios nichts anderes an als ein rein oberflächlich arbeitendes Konzept, das in keinerlei Weise die neueren Forschungsergebnisse, insbesondere die von TALLAL und TALCOTT, berücksichtigt, nach deren Erkenntnissen Legasthenie auf einer Wahrnehmungsstörung nichtsprachlicher Reize beruht und keinesfalls – wie immer noch zu hören ist – von mangelndem Fleiß oder mangelnder Intelligenz geprägt ist. So lange die Betroffenen bestimmte Laute nicht oder nicht richtig hören und/oder nicht richtig sehen können, ist jedes Üben, das nicht genau hierauf in erster Linie eingeht, völlig verfehlt. Wird nicht die Wahrnehmbarkeit nichtsprachlicher Reize gefördert, ändert auch der Einsatz neuer Medien wie beispielsweise des Computers nichts. Wichtig erscheint in diesem Zusammenhang auch, dass zusätzliche Angebote für ein häusliches Training gemacht werden, sodass dort auf täglicher Basis dem 7 Die Konsequenzen für die Therapie 140 betroffenen Kind Unterstützung zuteil werden kann. Den Betroffenen muss schnell und wirksam geholfen wird – dies wird manchmal aus den Augen verloren. Bedauerlich sind in diesem Zusammenhang Aussagen von Interessenverbänden und auch von Psychologen, wonach „ die Kinder lernen müssten, mit ihrem Defizit zu leben“ . Natürlich brauchen Kinder mit einem langen Leidensweg, die schon von einer Institution zur anderen gereicht wurden, irgendwann die notwendige professionelle psychologische Betreuung. Wenn es aber gelingt, den Kindern frühzeitig wirksam zu helfen, ist dies oft genug gar nicht erforderlich. 9HUIDKUHQQDFK7$//$/XQG0(5=(1,&+ Die Untersuchungen von TALLAL ergaben, dass bei vielen Legasthenikern elementare sensorische Fähigkeiten gestört sind (Tallal 2000). So stellten auch STEIN und TALCOTT fest, dass etwa 75 Prozent der legasthenischen Kinder in Tests schwächer abschneiden, die eine rasche visuelle Informationsverarbeitung verlangen (Stein u. Talcott 1999). Zudem entdeckten TALCOTT et al., dass erwachsene legasthenische Versuchspersonen auf langsame oder kontrastreiche visuelle Reize genauso gut reagierten wie gute Leser (Talcott et al. 2000). Bei schnell bewegten Reizen mit niedrigem Kontrast aber schnitten die legasthenischen Personen im Vergleich mit Erwachsenen ohne Leseschwächen deutlich schlechter ab. Zudem stellten TALLAL et al. (1985) eine zeitliche Verlangsamung der Hörverarbeitung bei Legasthenikern fest. Unter anderem machte sich diese in einer mangelhaften „ Filterfunktion“ des Gehörs bemerkbar: Einen bestimmten Sprecher aus einem Gewirr von Störgeräuschen und anderen Sprechern herauszuhören und zu verstehen stellt für Legastheniker eine besondere Schwierigkeit dar (Tallal et al. 1985). 7 Die Konsequenzen für die Therapie 141 TALLAL et al. (1985) konnten nachweisen, dass das bei Legasthenie häufig vorkommende auditive Informationsverarbeitungsdefizit bereits auf nichtsprachlicher Ebene vorliegt. Da die Wahrnehmung nichtsprachlicher Reize Voraussetzung für die Sprachverarbeitung in höheren Ebenen ist, kommt dieser Störung mit hoher Wahrscheinlichkeit eine große Bedeutung bei der Entstehung der Legasthenie zu (Tallal et al. 1985). TALLAL et al. (1985) sind überzeugt, dass auch die Verarbeitung in den Hörbahnen bei legasthenischen Kindern sehr häufig beeinträchtigt ist. So können die Zellen der entsprechenden Schaltstelle im Corpus geniculatum mediale kurz anklingende Hörreize nicht verarbeiten. Dies ist der Grund, dass viele Legastheniker etwa 0,3 Sekunden brauchen, um kurz anklingende Konsonanten (wie z.B. b–p) zu unterscheiden – sehr viel länger als die 0,008 Sekunden, die nicht-legasthenische Sechs- bis Achtjährige für diese Aufgabe benötigen (vgl. Kap. 6.2.1.2). Kann man die Laute der Sprache nicht unterscheiden, führt dies zu Schwierigkeiten beim Erlernen der Grammatik, des Lesens und des Schreibens (Tallal 2000). Aufgrund dieser Befunde ist es sehr wahrscheinlich, dass die Lesefähigkeit durch das Trainieren grundlegender sensorischer Fähigkeiten gefördert werden kann. Dies konnte auch bereits in Versuchen bestätigt werden. Um betroffenen Kindern zu helfen, spielte TALLAL Sprachaufnahmen mit Hilfe von Computern je nach Bedarf so langsam ab, dass die Patienten die einzelnen Vokale und Konsonanten erkennen konnten. Schritt für Schritt beschleunigte TALLAL dann die Sprachwiedergabe (Tallal 2000). Inzwischen wird diese Therapiemethode in Form von Computerspielen angeboten. TALLAL und MERZENICH entwickelten ein Computer- Trainingsprogramm, das die Verarbeitung schneller akustischer und visueller Reize im Gehirn fördert. Die beiden Wissenschaftler konnten zeigten, dass die Sprachübungen die Sprachentwicklung von Kindern, die mehrere Jahre hinter 7 Die Konsequenzen für die Therapie 142 dem Durchschnitt lagen, gesprochene Sprache zu verstehen, deutlich fördern konnten (Barinaga 1996). Das von TALLAL und MERZENICH entwickelte Trainingsprogramm ist eine Art Computerspiel. Das Programm dehnt die Laute schneller Konsonanten in Texten von 40 auf 80 Millisekunden und lässt sie überdeutlich ertönen. Um die Trainings-Einheiten für die Kinder spannend zu machen, bauten die Wissenschaftler sie in vertonte Video-Lernspiele ein. Dabei können die Kinder beispielsweise Jagd auf fliegende Kühe machen, die über einem Bauernhof kreisen. Gelingt es, zwei einander ähnliche Töne zu unterscheiden und dies per Tastendruck zu signalisieren, belohnt der Computer den Erfolg, indem er die Kuh im Stall bei ihren Artgenossen landen lässt (Barinaga 1996). Die Tests verliefen überaus erfolgreich: Nach einem Monat intensiver Übung drei Stunden am Tag, fünf Tage die Woche plus Hausaufgaben holten die Fünf- bis Zehnjährigen gegenüber einer Kontrollgruppe mit Kindern, die zwar die gleichen Lernspiele, aber mit normal schneller Sprache absolvieren, einen Entwicklungsrückstand von ein bis zwei Jahren auf. In einem zweiten, knapp drei Wochen langen Test mussten die Kinder zwischen schnell folgenden, aber nicht sprachlichen akustischen Signalen unterscheiden – mit wachsendem Schwierigkeitsgrad. Auch dieser scheinbar unspezifische Crash-Kurs brachte die Kinder deutlich voran. Die verbesserten Leistungen hielten noch bis zum Ende des Trainings an. Nach Angaben von TALLAL habe das Training das Sprachverständnis der Kinder offenbar nachhaltig gefördert (Tallal http://www.pna.org 3.10.2001). Inzwischen haben 25.000 amerikanische Kinder mit dem von TALLAL und MERZENICH entwickelten Computerspiel „ Fast for Word“ trainiert – an fünf Tagen in der Woche 100 Minuten täglich. „ Fast for Word“ dehnt schnelle Konsonanten in Texten und lässt sie überdeutlich ertönen. Die Ergebnisse waren ähnlich erfolgreich wie die der Tests: Nach sechs bis acht Wochen machten 90% der Kinder im Lesen einen Entwicklungssprung von zwei Jahren (Begley 2000). 7 Die Konsequenzen für die Therapie 143 7UDLQLQJGHV%OLFNODERUV)UHLEXUJ Eine gestörte Aufnahme visueller Informationen bei LRS kann durch Fehlfunktionen im Bereich der Okulomotorik verursacht sein (vgl. 5.2.3). Auch dies kann zu einer Beeinträchtigung grundlegender visueller Wahrnehmungsleistungen führen. So offenbarte die Untersuchung der Anti-Sakkaden-Aufgabe ein häufig auftretendes Defizit der willentlichen Komponente, welches in den früheren einfacheren Tests nicht feststellbar war. Dies führt dazu, dass Kinder mit einer Lese- und Rechtschreibschwäche häufig Regressionssakkaden, d.h. Sakkaden entgegengesetzt zur normalen Leserichtung zeigen (Fischer u. Weber 1990, Kotchen u. Guthrie 1980, Stark et al. 1991). Störungen in der Kontrolle der Blickmotorik machen sich außerdem in einem Rückstand der Entwicklung der Fixationsfähigkeit bemerkbar, d.h. der willentlichen Kontrolle über die Durchführung der Blicksprünge, der zeitlichen Genauigkeit der Reaktionszeiten und der Treffsicherheit. Die ordnungsgemäße Steuerung des Blicks entlang der Zeile von Wort zu Wort ist eine wichtige Voraussetzung für das Erlernen der Schriftsprache. Eine Untersuchung von über 1.000 Kindern zeigte, dass bis zu 50% der Legastheniker Störungen in der Kontrolle der Blickmotorik haben (Beratung Blicklabor http://www.brain.uni-freiburg.de/bbl 18.6.2001) (vgl. 5.2.3). In Fällen, in denen ein Zusammenhang mit den Augenbewegungen besteht, berichten die Kinder auf Anfrage oft, dass sie beim Lesen die Buchstaben auf dem Papier nicht stillstehend, sondern „ rumtanzend“ sehen. Außerdem verlieren die Kinder oft die Textstelle, überspringen häufig Zeilen und übersehen einzelne Worte oder WortTeile. Eine Arbeitsgruppe um FISCHER, Neurophysiologische Biophysik an der Universität Freiburg, entwickelte ein spezielles Trainingsprogramm gegen LRS 7 Die Konsequenzen für die Therapie 144 (Fischer http://www.optom.de 21.11.2001). Es richtet sich gegen Störungen der Blickmotorik, d.h. greift an einem anderen Ansatzpunkt als das Verfahren von TALLAL und MERZENICH an und kann dieses evtl. ergänzen bzw. bei einem anderen Patientenkollektiv wirksam werden. Zwar stellte FISCHER fest, dass eine generelle Heilbarkeit von Legasthenie nach derzeitigem wissenschaftlichen Kenntnisstand unwahrscheinlich sei, betonte jedoch, dass es das Ziel sein solle, eine erträgliche Schulkarriere zu ermöglichen. Therapeutische Maßnahmen bei Legasthenie müssten gezielt erfolgen und verlangten daher eine ausführliche genaue Diagnose (Fischer http://www.optom.de 21.11.2001). %OLFN7UDLQLQJ Wie TALLAL sind auch FISCHER und WEBER davon überzeugt, dass grundlegende visuelle und auditive Wahrnehmungsdefizite eine bedeutende Rolle bei der Entwicklung einer LRS spielen: Nach Aussagen von FISCHER und WEBER zeigen etwa 50 Prozent aller Legastheniker Störungen elementarer zerebraler Hör- und Sehfunktionen (Fischer u. Weber 1990). FISCHER und WEBER sind der Überzeugung, dass die unzureichende Kontrolle der Blickbewegung der Augen eine entscheidende Rolle bei der Entstehung der LRS spielt. Auf diese Weise wären Legastheniker nicht imstande, ihr Augenmerk schnell genug von einzelnen Buchstaben und Wörtern zu lösen. Wie TALLAL stellte FISCHER fest, dass leserechtschreibgestörte Kinder im Vergleich zu ihren Klassenkameraden auch Laute weniger gut unterscheiden können (Fischer http://www.optom.de 21.11.2001, Tallal 2000). Ob und in welchem Ausmaß Kinder mit Lese-Rechtschreib-Schwäche unter einer solchen unzureichenden Informationsverarbeitung leiden, testete FISCHER mit speziellen Geräten. Sie projizieren einen Laserpunkt an die Wand, dem die Kinder bewusst mit den Augen folgen sollen oder von dem sie – im zweiten Test – bewusst wegblicken sollen. Zugleich werden die Augenbewegungen des Kindes aufgezeichnet und analysiert. So kann gemessen werden, wie weit das reflexbestimmte (zum Punkt hinschauen) und das willentliche (vom Punkt wegsehen) Blicken bei einem Kind 7 Die Konsequenzen für die Therapie 145 entwickelt sind. Bei LRS-Schwäche erwies sich in den Untersuchungen von FISCHER das willentliche Blicken häufig als unterentwickelt. Die Kinder gaben zu stark ihren Reflexen nach – deshalb konnten sie sich beim Lesen auch nicht auf Worte oder Buchstabengruppen konzentrieren und sprangen oft im Text hin und her. Für das willentliche Blicken ist – wie für viele andere Koordinationsaufgaben auch – der präfrontale Kortex zuständig. In diesem Bereich und vor allem in der vorgelagerten Schaltstelle zwischen Seh- beziehungsweise Hörnerv und dem Stirnhirn, dem Corpus geniculatum, wurden bei einigen Legasthenikern Veränderungen gefunden, nämlich eine zu geringe Ausprägung der Magnozellen (Fischer 21.11.2001). http://www.optom.de Dieser Befund stimmt mit Forschungsergebnissen TALLALs überein, nach denen es Hinweise dafür gibt, dass die von Magnozellen gebildete Bahn bei Legasthenikern weniger leistungsfähig ist (Tallal 2000, vgl. 5.2.3.1). Werden Störungen der Blickmotorik festgestellt, ist ein Training der Blickkontrolle angezeigt, um die Voraussetzungen für einen zuverlässigen Leseprozess zu verbessern. Spezielle Trainingsgeräte wurden vom Blicklabor in Freiburg entwickelt. Wie FISCHER betont, sollte das Training jedoch nur nach einer genauen Diagnose erfolgen, da mit unterschiedlichen Trainings-Varianten verschiedene Aspekte der Blicksteuerung bzw. der Hörens trainiert werden und durch falsches Training die Situation auch verschlimmert werden könne (Fischer http://www.optom.de 21.11.2001). Mit dem so genannten Fix-Train-Gerät sollen die Kinder drei bis sechs Wochen lang je zehn Minuten täglich üben. Beispielsweise müssen sie per Knopfdruck anzeigen, nach welcher Seite ein kleiner Pfeil zeigt, der für Sekundenbruchteile an unterschiedlichen Stellen des Bildschirms erscheint. Steigt die Trefferquote über einen vorgegebenen Schwierigkeitsstufe. Wert, so schalten die Geräte auf die nächste 7 Die Konsequenzen für die Therapie 146 Obwohl das Training nur einige Wochen dauert, hält der Effekt sehr lange, vermutlich dauerhaft, an. Das Blick-Training kann zwar nicht das Lesen-Lernen ersetzen, jedoch werden durch das Training die Voraussetzungen für den Erfolg anschließender Fördermaßnahmen des Lesens und der Rechtschreibung verbessert bzw. erst geschaffen. Etwa 80 Prozent von 110 bislang nachuntersuchten Schülern haben mit dieser Methode ihre Blicksteuerung optimiert (Fischer http://www.optom.de 21.11.2001). Ersten Untersuchungen zufolge verbesserten sich bei 33 Prozent der Kinder die Lese- und Rechtschreibleistungen allein durch die Übungen, obwohl sie keine zusätzliche pädagogische Förderung erhielten. Bei den meisten der übrigen Kinder zeigten sich zumindest kleine Verbesserungen: Die Handschrift war besser lesbar, beim Lesen sprang der Blick nicht mehr so oft im Text hin und her, und die Schrift blieb konstant auf der Zeile. FISCHER betont jedoch, dass viele Kinder auch nach dem Blicktraining noch spezielle Förderung benötigen – etwa durch Pädagogen und Logopäden (Fischer http://www.optom.de 21.11.2001). Die Blickübungen sollten lediglich die Voraussetzungen für eine erfolgreiche Teilnahme am Deutschunterricht schaffen. Wahrscheinlich kann das Blicktraining nach FISCHER somit das Verfahren von TALLAL und MERZENICH ergänzen, insbesondere bei lese-rechtschreibgestörten Kindern mit unzureichender Kontrolle der Blickmotorik. +|U7UDLQLQJ Basierend auf der Erkenntnis, dass ein Teil der legasthenischen Kinder Entwicklungsrückstände in der Hörwahrnehmung zeigt, entwickelte die Arbeitsgruppe um FISCHER auch dafür Tests, erfasste Normdaten und entwarf ein Trainings-Programm. Das hierfür entwickelte Gerät FonoFix beherrscht fünf sprachfreie Tests der Hörwahrnehmung, deren Schwierigkeit der jeweiligen 7 Die Konsequenzen für die Therapie 147 Leistung des Kindes automatisch angepasst wird (Fischer http://www.optom.de 21.11.2001). In Übereinstimmung mit TALLAL ist FISCHER davon überzeugt, dass eine wesentliche Voraussetzung für richtiges Hören und Verstehen, insbesondere Voraussetzung für das Verstehen von gesprochener Sprache, die Fähigkeit sei, Unterschiede in den akustischen Signalen zu erkennen. Diese Fähigkeiten werden erst im Laufe des Lebens erworben und können im Alter eingeschränkt sein (Fischer http://www.optom.de 21.11.2001, Tallal 2000). Auch bei gesunden Sinnesorganen können bei der Verarbeitung der Informationen im Gehirn Fehler bzw. Entwicklungsrückstände auftreten und zu Lernstörungen und deren Folgen führen. Es ist deswegen wichtig, diese Leistungen quantitativ erfassen und mit Normdaten vergleichen zu können (Fischer http://www.optom.de 21.11.2001). Die Arbeitsgruppe um FISCHER (Universität Freiburg) entwickelte in Zusammenarbeit mit der Klinik für Pädaudiologie und Phoniatrie (Medizinische Hochschule Hannover) fünf sprachfreie Hörtests und normierte diese anhand der Daten von über 400 Personen für Altersstufen zwischen 6 und 35 Jahren. FonoFix ermittelt den Schwellenwert und – nach Angabe des Alters – den Prozentrang der Testperson. Im Einzelnen werden bestimmt: • Lautstärken-Differenzierung: Unterschiedlich laute Töne sind Teil der Aussprache einzelner Wörter. Hier wird geprüft, wie gut man zwei unterschiedlich laute Töne unterscheiden kann. • Tonhöhen-Differenzierung: Ebenso sind Teile gesprochener Wörter unterschiedlich in der Tonhöhe. Hier wird geprüft, wie gut man zwei unterschiedlich hohe Töne unterscheiden kann. • Gap-Erkennung (vgl. Frequenzlücke, 4.2.2.1): Wie PHILLIPS bereits beschrieb, kommen in gesprochener Sprache kurze Pausen – Frequenzlücken – vor, die erkannt und richtig interpretiert werden müssen, z.B. bei tonlosen 7 Die Konsequenzen für die Therapie 148 Konsonanten (Phillips 1999). Die Fähigkeit, diese kurze Lautunterbrechungen zu erkennen, wird mit der Gap-Erkennung getestet. • Zeitordnungs-Schwelle: Damit die Laute in der Reihenfolge wahrgenommen werden, wie sie erzeugt wurden, wird die Fähigkeit benötigt, zwei deutlich verschiedene Töne, die kurz aufeinander folgen, nach ihrer zeitlichen Reihenfolge richtig einordnen zu können (einohrige Erkennung der zeitlichen Reihenfolge). • Seitenordnungs-Schwelle: Dieser Test prüft, inwieweit die Fähigkeit vorhanden ist zu erkennen, ob ein Ton zuerst dem rechten oder zuerst dem linken Ohr angeboten wurde (beidohrige Erkennung der zeitlichen Reihenfolge). Die Anwendung von FonoFix zeigte eine Entwicklung dieser Hörleistungen bis zum 20. Lebensjahr. Kinder mit Legasthenie bestanden diese Tests im Durchschnitt signifikant schlechter als gleichaltrige Kontrollkinder. Etwa die Hälfte der legasthenischen Kinder war betroffen (Fischer http://www.optom.de 21.11.2001). FISCHER entwickelte das Trainingsprogromm FonoTrain zum Zwecke des Hörtrainings und stellte fest, dass die gestörten Funktionen durch tägliches Üben verbessert werden konnten. Es können speziell die Funktionen trainiert werden, bei denen Schwächen bestehen. FonoTrain speichert die Trainingsdaten der verschiedenen Aufgaben während des Trainings, sodass noch nachträglich der Trainingsverlauf in einem Protokoll nachvollzogen werden kann (Fischer http://www.optom.de 21.11.2001). 7KHUDSLHQDFK:$51.( Ein weiteres Therapiekonzept, nämlich das von WARNKE, geht von einer Beeinträchtigung grundlegender auditiver Wahrnehmungs- und 7 Die Konsequenzen für die Therapie 149 Verarbeitungsprozesse im Gehirn als Ursache der Legasthenie aus (Warnke 1995). *HVW|UWH7HLOIXQNWLRQHQGHU]HQWUDOHQ+|UYHUDUEHLWXQJ Nach WARNKE sind folgende fünf Teilfunktionen bei zentraler Fehlhörigkeit, also bei zentralen Hörverarbeitungsstörungen, beeinträchtigt (Warnke 1995): • auditives Ortungsvermögen, • Tonhöhen-Unterscheidungsvermögen, • phonematische Diskrimination, • auditive Ordnungsschwelle, • auditiv-motorische Umsetzung. Unter dem auditiven Ortungsvermögen wird die Fähigkeit verstanden, Richtungsunterschiede einer Schallquelle wahrzunehmen. Es liegt im Vorderbereich bei 2 Winkelgraden, seitlich bei 10 Winkelgraden und hinten bei 5 Winkelgraden. Nur aufgrund dieser Fähigkeit ist es möglich, aus einem Störlärm eine gewünschte Schallquelle herauszuhören und zu verstehen. Dieser so genannte Partyeffekt stellt die ins Auditive übertragene Figur-Grundwahrnehmung aus dem visuellen Bereich dar. Eine Messmethode ohne Hilfsmittel besteht darin, dass das zu testende Kind die Augen schließt und der Untersucher aus verschiedenen Richtungen jeweils eine kurze Silbe vorspricht. Das Kind soll dann genau in die Richtung deuten, aus der es die Silbe gehört zu haben glaubt. Noch genauere Feststellungen erlaubt die Verwendung eines so genannten Rauschgenerators, der ein weißes Rauschen erzeugt und jeweils kurz aus verschiedenen Richtungen eingeschaltet wird. Das zu testende Kind zeigt auch hierbei mit geschlossenen Augen in die Richtung, aus der es die Tonbursts wahrgenommen hat (Warnke 1996). Bezüglich des Tonhöhen-Unterscheidungsvermögens können normal hörende Menschen mindestens 5 Tonhöhenunterschiede zwischen zwei 7 Die Konsequenzen für die Therapie 150 aufeinanderfolgenden Tönen von 1-2% wahrnehmen, d.h. angeben, ob der erste oder der letzte von zwei aufeinanderfolgenden Tönen der höhere bzw. der tiefere ist. Kinder mit einer zentralen Fehlhörigkeit können dagegen sehr selten einen Halbtonschritt (entsprechend 5,9%), häufig sogar nicht einmal einen Ganztonschritt (entsprechend 12,2%) und manchmal sogar nicht einmal eine kleine Terz (18,9%) in dieser Weise voneinander unterscheiden. Das lässt sich mit einem so genannten Viertongeber VTG-1000 in Anlehnung an eine Arbeit von TALLAL (1993) feststellen. Die phonematische Diskrimination oder Wahrnehmungs-Trennschärfe ist die Fähigkeit des Menschen, ähnlich klingende Laute voneinander zu unterscheiden. Herkömmlich wird sie häufig mit so genannten Minimalpaaren, wie beispielsweise „ Tanne-Kanne" oder „ Nagel-Nadel", überprüft. Deutlich aussagekräftiger ist ein Wahrnehmungs-Trennschärfe-Test mit sinnfreien Wörtern, wie z.B. „ ETI – EKI – EPI – EGI – ..", weil er einen Rückgriff auf den lexikalischen Wortschatz ausschließt (Warnke 1996). Die auditive Ordnungsschwelle ist diejenige Zeitspanne, die zwischen zwei Sinnesreizen mindestens verstreichen muss, damit sie getrennt wahrnehmbar sind und in eine Reihenfolge, also eine Ordnung, zu bringen sind. Die auditive Ordnungsschwelle liegt etwa ab dem neunten Lebensjahr zwischen 30-40 Millisekunden, bei Sechsjährigen etwa um 60 Millisekunden. Bei sprachauffälligen Kindern ist sie zumeist um das Zwei- bis Dreifache verlangsamt (Warnke 1996). In mehreren Versuchsreihen wies der amerikanische Wissenschaftler WOLFF (Wolff u. Drake 1984, Wolff 1993) nach, dass die auditiv-motorische Umsetzung bei sprachauffälligen Kindern zumeist gestört ist. So haben Kinder mit zentraler Fehlhörigkeit insbesondere dann ganz erhebliche Schwierigkeiten beim Umsetzen eines Metronomtaktes in synchrones Fingerklopfen, wenn sie aufgefordert werden, denselben Finger – z.B. den Zeigefinger – beider Hände abwechselnd genau im Takt zum Metronom zu benutzen. Stellt man das Metronom mittels 7 Die Konsequenzen für die Therapie 151 seiner Skalierung zunächst auf 150 Schläge pro Minute; das entspricht einem Schlagabstand von 400 Millisekunden, so sollte dies auch ein Kind mit zentralen Hörverarbeitungsproblemen noch schaffen. Ist dies der Fall, so erhöht man in mehreren Schritten bis auf 270 Schläge pro Minute, entsprechend 222 Millisekunden. Dies gelingt einem Kind mit zentraler Hörverarbeitungsstörung nicht mehr. 7UDLQLQJVYHUIDKUHQQDFK:$51.( Das schwerpunktmäßig eingesetzte Trainingsverfahren besteht – mit der Zielsetzung einer Verbesserung vor allem der Ordnungsschwelle und der Wahrnehmungs-Trennschärfe – aus zwei prinzipiell auch getrennt einsetzbaren, vorzugsweise aber zu kombinierenden Einzelabläufen (Warnke 1995): Ordnungsschwellen-Training: Da die Ordnungsschwelle eine wichtige Voraussetzung in der zentralen Hörverarbeitung für eine einwandfreie Wahrnehmungs-Trennschärfe ist, wurde ein Gerät entwickelt, das die Ordnungsschwelle trainiert (Warnke 1996). Das Trainieren der Ordnungsschwelle geht davon aus, durch einen Synergie-Effekt zwischen der zentralen Seh- und Hörverarbeitung betroffenen die Kinder zumeist zur schnellere Beschleunigung visuelle ihrer Ordnungsschwelle langsameren der auditiven Ordnungsschwelle zu benutzen. Messen lässt sich die Ordnungsschwelle mit dem handgehaltenen Gerät: Zunächst wird die auditive Ordnungsschwelle mit Hilfe von zwei aufeinanderfolgenden Klicks festgestellt, dann die visuelle mit zwei aufeinanderfolgenden Lichtblitzen. Beim eigentlichen Training wird die auditive Ordnungsschwelle mit Hilfe der visuellen verbessert: Das Gerät wird in eine neue Stellung VISUELL+AUDITIV umgeschaltet. Dabei nimmt das Kind den auditiven Sinnesreiz, also die beiden Klicks, absolut synchron mit dem visuellen Sinnesreiz wahr, also dem Aufleuchten der beiden Leuchtdioden. Bei vier zutreffenden Reaktionen verkürzt sich der Abstand zwischen den Reizen um 10 Millisekunden und verlängert sich bei einer falschen Reaktion um denselben 7 Die Konsequenzen für die Therapie 152 Betrag. Bei regelmäßigem Einsatz dieser gleichzeitigen auditiv-visuellen Übung erzielten die meisten Benutzer im Verlauf schon weniger Wochen deutlich bessere Werte sowohl ihrer auditiven als auch ihrer visuellen Ordnungsschwellen im Vergleich zu den ursprünglichen Messungen (Warnke 1996). Training der Hemisphären-Koordination: Bei diesem Teil des Trainings wird davon ausgegangen, dass die Beeinträchtigung der zentralen Hörverarbeitung schwerpunktmäßig in der unzureichenden Koordination der beiden Hirnhemisphären begründet liegt. Ansatzpunkte dafür gibt es in einigen Untersuchungen von Wolff (Wolff 1993). In mehreren aufeinander aufbauenden Stufen mit wochenweiser Steigerung des Schwierigkeitsgrades wurden verschiedene Übungen zum Mithören, Mitsingen und Mitsprechen entwickelt. Das Charakteristische an diesem Verfahren ist das stetige Hin- und Herwandern der über Kopfhörer angebotenen Lieder und Texte zwischen den beiden Ohren des Kindes. Im Einzelnen hört das Kind mittels Kopfhörer zunächst über ein spezielles Gerät von einem CD-Spieler kommende Sing- bzw. Sprechtexte, beispielsweise die eigens dafür in Kunstkopf-Stereofonie aufgezeichneten Hifino-Geschichten, die also mit Hilfe des Gerätes in einem bestimmten zeitlichen Rhythmus zwischen seinen beiden Ohren hin- und herpendeln. Sobald das Kind dazu in der Lage ist, soll es diese Texte auch in ein Mikrofon mitsingen bzw. mitsprechen oder nachsprechen, wobei seine eigene Stimme ebenfalls – und zwar gegenläufig zur Modellstimme von der CD – zwischen seinen beiden Ohren hin- und herpendelt: Wenn also die Modellstimme beispielsweise gerade von rechts kommt, hört das Kind seine eigene Stimme von links; während die Modellstimme nach links überwechselt, kommt ihm die eigene Stimme von links nach rechts entgegen. So werden ständig bestehende neuronale Verknüpfungen verstärkt und neue Verknüpfungen geschaffen. Dieser Teil des Trainings wird von den zahlreichen bisher damit bereits arbeitenden Kindern und teilweise auch Erwachsenen so bewertet, dass sie dadurch eine ganz neue Beziehung zur Sprache und mehr Sicherheit im Umgang mit ihrer eigenen Sprache gefunden haben. Eine Umfrage 7 Die Konsequenzen für die Therapie 153 bei 50 Familien nach sechsmonatigem Training ergab bereits Verbesserungen der Leseleistungen bei 94% der Trainierenden und Verbesserungen der Rechtschreibleistungen bei 86% der Trainierenden (Warnke 1996). Die beiden oben beschriebenen Trainingsverfahren sind ursprünglich mit der Zielsetzung entstanden, in der häuslichen Sphäre eine bestehende und fortzuführende Betreuung durch Logopäden, Sprachheiltherapeuten, Legasthenietherapeuten und vergleichbare Berufe flankierend zu unterstützen. Inzwischen aber sind bereits zahlreiche Angehörige dieser Berufsgruppen dazu übergegangen, auch in ihre Therapien bestimmte Elemente aus den vorstehend geschilderten Methoden einzubeziehen (Warnke 1996). Für Vorschulkinder und Erstklässler, die noch nicht lesen können, wird eine in Kunstkopfstereofonie aufgezeichnete CD mit 36 Kinderliedern im Leistungsvermögen von Fünf- bis Siebenjährigen verwendet, bei denen die trainierenden Kinder mitsingen. Übungsdiktate werden über Kopfhörer angesagt, wobei natürlich die Stimme der Therapeutin zwischen den beiden Ohren des Kindes hin- und herwandert, während gleichzeitig eine dosierbare räumliche Störgeräuscheinblendung die Figur-Grund-Wahrnehmung des Kindes schult. So entstandene Diktate werden durch Schüler und Therapeutin über zwei Mikrofone synchron „ nachgelesen", wobei die Therapeutin bei einem Fehler nur kurz innehält, um dem Schüler die Chance zum eigenständigen Erkennen des Fehlers zu geben. Lautsprachliche Arbeit wird durch „ Synchronsprechen“ von Therapeut und Kind motivierender und effizienter (Warnke 1996). 7KHUDSLHGHUUHLQHQ:RUWWDXEKHLW In Übereinstimmung mit den Ergebnissen von TALLAL, TALCOTT, FISCHER und WARNKE stellte auch PHILLIPS fest, dass die Ursache eines Defizits eine Störung im akustischen Timing des Gehirns sein kann, sodass das Erkennen 7 Die Konsequenzen für die Therapie 154 schnell aufeinander folgender Sinnesereignisse unmöglich wird. PHILLIPS untersuchte die reine Worttaubheit. Somit kann davon ausgegangen werden, dass sowohl die reine Worttaubheit als auch die Legasthenie auf einer grundlegenden Wahrnehmungsstörung basiert. PHILLIPS stellte fest, dass die reine Worttaubheit auf Laute begrenzt ist, die in einem zeitlichen Rahmen von Millisekunden oder weniger liegen. Von PHILLIPS konnten neurophysiologische Beweise präsentiert werden, dass der primäre auditorische Kortex eine spezifische Rolle in der Repräsentation in diesem kleinen Zeitrahmen spielt. Bei einer eingeschränkten Funktionsfähigkeit bzw. sogar einem Ausfall eines Kortexbereiches ist die plastische Kapazität adulter Kortizes Grundlage vieler medizinischer Rehabilitationsprozesse (Dinse et al. 1997). So können beispielsweise bei reiner Worttaubheit an anderen Orten entstehende kortikale Erregungsmuster offensichtlich vom Patienten neu gelernt werden und nach einiger Zeit den entsprechenden Signalen zugeordnet werden. Auf diese Weise kann das Sprachverständnis bis zu einem gewissen Grad wiederhergestellt werden (Godde 2001). Jedoch ist zu beachten, dass das Wiedererlangen des Sprachverständnisses nach dem Auftreten einer reinen Worttaubheit als Folge einer Schädigung des primären auditorischen Kortex ein langwieriger und für die Betroffenen und deren Angehörigen ein mühevoller Prozess ist (Mezger http://home.algaeu.org 21.11.2001). Ein Abschluss der spontanen Rückbildung wird überwiegend zwischen einem und sechs Monaten nach Erkrankungsbeginn angegeben. Danach wird das Ausmaß einer weiteren Verbesserung der sprachlichen Funktionen ohne entsprechende logopädische Therapie als nur noch geringfügig eingeschätzt. Eine Untersuchung hierzu an Patienten, deren reine Worttaubheit nicht behandelt worden war, zeigte deutliche Leistungsverbesserungen zwischen einer ersten Untersuchung 4-6 Wochen nach der Erkrankung und einer zweiten Untersuchung 3 Monate nach der ersten Untersuchung. Nur noch eine geringe weitere spontane 7 Die Konsequenzen für die Therapie 155 Besserung war drei Monate nach der zweiten Untersuchung festzustellen. Da innerhalb der Gruppe der an reiner Worttaubheit Erkrankten eine große interindividuelle Variabilität besteht, sind statistische Ergebnisse nur eingeschränkt auf Einzelfälle anwendbar. Auch kann bei unterschiedlicher Ätiologie, z.B. nach einem Schädelhirntrauma mit evtl. mehrwöchigem Koma, der Spontanverlauf nicht vergleichbar sein, und noch größere individuelle Unterschiede, je nach medizinischen Begleitumständen, sind möglich. Ein anderer Aspekt des Verlaufs ist der teilweise zu beobachtende Syndromwandel, z.B. von reiner Worttaubheit zu Global-Aphasie, von Global-Aphasie zu Broca-Aphasie, von Broca- oder Wernicke-Aphasie zu amnestischer Aphasie und in allen Gruppen zu nicht klassifizierbaren Aphasien (Schmidt et al. 2000). Nach dem oben Dargelegten muss eine objektivierbare Verbesserung der Sprachleistung jenseits von 6 Monaten nach einem Hirninfarkt auf jeden Fall als Effekt einer Therapie angesehen werden. Der Beginn therapeutischer Bemühungen soll aber keinesfalls erst nach Abschluss der zu erwartenden spontanen Remission einsetzen. Um das vorhandene Restitutionspotential möglichst völlig auszuschöpfen, wird empfohlen, möglichst früh mit einer kompetenten Behandlung oder, wenn dies nicht möglich ist, zumindest mit unspezifischer sprachlicher Aktivierung durch Laien einzusetzen. Durch Untersuchungen ist erwiesen, dass sowohl Behandlung durch nicht geschultes Personal fehlender Behandlung überlegen ist als auch, dass verschiedene Methoden, durch ausgebildete Sprachtherapeuten angewandt, noch jeweils besser als die nicht professionell durchgeführte Therapie sind, vorausgesetzt, eine ausreichend intensive Behandlung von bis zu 1 Jahr Dauer und mindestens 3 Stunden pro Woche. Allgemein anerkannt ist die Notwendigkeit rechtzeitig begonnener und genügend intensiver und langfristiger Behandlung von wenigstens 3 bis 4 Stunden pro Woche über 12 und mehr Monate. Mehrwöchige Phasen sehr intensiver, ambulanter oder stationärer Behandlung sind aller Erfahrung nach auch effektiver als die häufig noch anzutreffende einmal wöchentliche ambulante Behandlung über einen längeren Zeitraum. 7 Die Konsequenzen für die Therapie 156 Im Gegensatz zur Therapie der Legasthenie wurden bei der reinen Worttaubheit bisher keine therapeutischen Verfahren entwickelt, welche insbesondere die Verarbeitung schneller akustischer Reize im Gehirn fördern. Der Grundsatz der therapeutischen Verfahren Behandlungsphasen heißt unterschieden vielmehr werden „ üben, (Mezger üben“ , wobei drei http://home.algaeu.org 21.11.2001): • Die Aktivierungsphase mit dem Ziel sprachlicher Stimulierung in den ersten 4-6 Wochen. Diese sollte möglichst bald nach Erkrankungsbeginn einsetzen, in einer Intensität wie es der Allgemeinzustand und die Belastbarkeit zulassen. Sämtliche Mitarbeiter auf der Krankenstation und die entsprechend aufgeklärten und angeleiteten Angehörigen sollten einbezogen werden. • Die störungsspezifische Übungsphase mit Behandlung vorwiegend der sprachsystematischen Defizite, anfangs überwiegend in Einzelbehandlung, später auch zusätzlich in kleinen Gruppen, insgesamt bis zu einem Jahr, in besonderen Fällen auch länger. • Die Konsolidierungsphase, in der jetzt der Patient zunehmend lernen soll, vorhandene sprachliche Fähigkeiten zur Kommunikation möglichst optimal einzusetzen, was überwiegend durch Gruppentherapie realisiert werden kann. Die in den drei Phasen jeweils definierten Behandlungsziele sind fließend ineinander übergehend und deshalb nicht künstlich streng zu trennen, sondern auch parallel nebeneinander und gleichzeitig in das Behandlungskonzept zu integrieren. Häufig wird eine vollständige Wiederherstellung der sprachlichen Fähigkeiten nicht zu erreichen sein, weshalb das Behandlungsziel dann die Erlangung bestmöglicher kommunikativer Fähigkeiten zur Bewältigung alltäglicher Situationen und individueller Bedürfnisse ist mit Verzicht auf stets grammatikalisch korrekte Äußerungen und unter Miteinbeziehung auch nonverbaler Verständigungsmöglichkeiten (Grehl et al. 2000). Zahlreiche Faktoren, die zum Teil komplex und in ihrer wechselseitigen Abhängigkeit wenig überschaubar sind, bestimmen den Verlauf einer reinen 7 Die Konsequenzen für die Therapie 157 Worttaubheit und damit den therapeutischen Erfolg. Dennoch können einige Regeln aufgestellt werden. Bezüglich der Ätiologie hat eine reine Worttaubheit bei Verschluss der A. carotis interna, vermutlich wegen der günstigen Kollateralversorgung, und bei Verschluss der A. cerebri posterior, wie auch nach gedecktem Schädelhirntrauma, einen oft günstigen Verlauf, während Verschlüsse der A. cerebri media oder einzelner ihrer Äste sehr stabile Syndrome hervorrufen, ebenso wie auch hypoxische und enzephalitische Gewebsschäden. Intrazerebrale Blutungen haben eine Prognose offensichtlich abhängig im Wesentlichen vom Ausmaß der Gewebeläsionen in der sprachrelevanten Region, was auf die Bedeutung von Lokalisation und Größe der Gewebsschädigung hinweist, auch wenn sonst die Größe der Läsion im CT erstaunlich wenig Voraussagen bezüglich der Schwere der Sprachstörung zulässt (Grehl et al. 2000). Bei den kreislaufbedingten Worttaubheiten gilt auch ein Zusammenhang zwischen Rückbildungschance und Ausmaß des initialen sprachlichen Defizits. Insbesondere muss bei noch erheblichen sprachlichen Symptomen 2 Wochen nach Erkrankungsbeginn mit länger bestehenden Beeinträchtigungen gerechnet werden (Schmidt et al. 2001). Die das höhere Lebensalter häufig begleitenden Faktoren wie schlechterer allgemeiner Gesundheitszustand, Herz-Kreislauf-Erkrankungen und ungünstigere psychosoziale Bedingungen haben mehr als das Alter selbst einen negativen Einfluss. Gleichzeitig vorliegende zusätzliche neuropsychologische Beeinträchtigungen haben eine ungünstigere Prognose zur Folge. Linkshändigkeit soll hingegen mit einer besseren Rückbildungsmöglichkeit verbunden sein (Grehl et al. 2000). Eine bleibende Läsion im Bereich des primären auditorischen Kortex bedingt in der Regel eine Einbuße sprachlicher Fähigkeiten. Eine optimale Sprachtherapie bedeutet infolgedessen das maximale Ausschöpfen des von den prognostischen 7 Die Konsequenzen für die Therapie 158 Faktoren beeinflussten vorhandenen Rückbildungspotentials. Dies aber erfordert eine ausreichend intensive, genügend lange anhaltende und kompetente Behandlung, möglichst in einer interdisziplinären Zusammenarbeit von Ärzten, Therapeuten und Psychologen, unter Miteinbeziehung der Angehörigen (Grehl et al. 2000). Schlussbemerkung 159 6FKOXVVEHPHUNXQJ Die Untersuchungsergebnisse von TALLAL, TALCOTT und PHILLIPS ergaben, dass sowohl die Legasthenie als auch die reine Worttaubheit zumeist keine Sprachverarbeitungsstörungen sind. So beruhen legasthenische Defizite in erster Linie auf einer Störung beim Verarbeiten rasch wechselnder elementarer optischer und akustischer Stimuli. TALLAL und TALCOTT stellen das reibungslose Funktionieren der optischen und akustischen Sinneswahrnehmungen als unumgängliche Voraussetzung für das erfolgreiche Erlernen von Schreiben und Lesen dar. In verschiedenen Untersuchungen konnten sie nachweisen, dass bei Legasthenikern häufig ein auditives bzw. visuelles Informationsverarbeitungsdefizit bereits auf nichtsprachlicher Ebene vorliegt. Da die Wahrnehmung nichtsprachlicher Reize Voraussetzung für die Sprachverarbeitung in höheren Ebenen ist, kommt dieser Störung mit hoher Wahrscheinlichkeit eine große Bedeutung bei der Entstehung der Legasthenie zu (Tallal 2000). Die Untersuchungen von PHILLIPS weisen daraufhin, dass auch die reine Worttaubheit eine Manifestation einer mehr grundlegenden zeitlichen Verarbeitungsstörung auf vorsprachlichem Niveau sei. PHILLIPS stellte fest, dass der reinen Worttaubheit eine sprachunspezifische Störung im primären auditorischen Kortex zugrunde liegt (Phillips 1997). Eine Anzahl von Hinweisen zeigt an, dass der primäre auditorische Kortex eine Rolle bei der Wahrnehmung, aber nicht bei der linguistischen Verarbeitung von Lauten spielt. Die Genauigkeit des Antwortverhaltens der kortikalen Neuronen reichte in den Untersuchungen von PHILLIPS aus, um den zeitlichen Verlauf einzelner Sprachkomponenten darzustellen. Dies lässt darauf schließen, dass der primäre auditorische Kortex bei der Unterscheidung verschiedener Sprachabschnitte eine Rolle spielt. Die Ergebnisse zeigen zudem, dass die Antworten auf Reize im Kortex eine Genauigkeit des Antwortverhaltens zeigten, die nur etwas schlechter als die der Fasern des Kochlearnervs ist (Phillips 1990). Schlussbemerkung 160 Eines der wichtigsten diagnostischen Kriterien der Genauigkeit, mit der Nervenzellen auf Töne oder Geräusche reagieren ist die Prüfung, inwieweit die Frequenzlücke (4.2.2.1) erkannt werden kann. Die wichtigsten „ Frequenzlücken“ der Sprache sind die phonetisch bedeutsamen – sie lassen Stopp-Konsonanten erkennbar werden (Phillips 1990). Nicht alle Bereiche des Sprachunterscheidungsvermögens sind bei der reinen Worttaubheit gleichermaßen betroffen. Insbesondere wurde festgestellt, dass das Wahrnehmungsvermögen langer Vokale bilaterale Läsionen des primären auditorischen Kortex übersteht, jedoch das von Stopp-Konsonanten nicht. Die gestörte Unterscheidungsfähigkeit von Stopp-Konsonanten bei Worttaubheit zeigt ein zeitliches Verarbeitungsdefizit an, das Stopp-Konsonanten betrifft, aber nicht Vokale. Dies liegt im Bereich von einer bis zu zehn Millisekunden und charakterisiert die Stopp-Konsonanten phonetisch (Phillips 1990). Die Forschungsergebnisse von TALLAL, TALCOTT und PHILLIPS lassen Schlussfolgerungen für die Therapie zu: Sowohl eine Lese-RechtschreibSchwäche als auch eine reine Worttaubheit müssten durch das Trainieren elementarer sensorischer Fähigkeiten gefördert werden können. Damit sollte das Konzept des starren Übens von Lesen und Schreiben abgelöst werden können durch ein wirksameres. Dies konnte bereits bezüglich der Legasthenie bestätigt werden. So entwickelten TALLAL und MERZENICH eine Art Computerspiel, das spezifisch die bei der Legasthenie beeinträchtigten Wahrnehmungs- und Unterscheidungsleistungen verbessert. TALLAL stellte fest, dass ein auf diese Weise erfolgendes Training das Sprachverständnis der Kinder nachhaltig fördert (Tallal http://www.pna.org 3.10.2001). Auch andere Wissenschaftler gehen von der Auffassung aus, dass eine LRS durch das Trainieren nichtsprachlicher Elemente, insbesondere der Ordnungsschwelle sowie der HemisphärenKoordination, nachhaltig gefördert werden könne (Warnke 1995). Ergänzt werden können diese therapeutischen Verfahren durch das von FISCHER entwickelte Blicktraining (Fischer http://www.optom.de 21.11.2001). Werden Störungen der Blickmotorik festgestellt, ist ein Training der Blickkontrolle Schlussbemerkung 161 angezeigt, um die Voraussetzungen für einen zuverlässigen Leseprozess zu verbessern. Zusätzlich entwickelte FISCHER ein Hörtraining, das wie das TALLAL-Verfahren die Fähigkeit übt, elementare Unterschiede in den akustischen Signalen zu erkennen. FISCHER geht hierbei auf die Frequenzlücke ein, deren Bestimmung ein wichtiges Mittel ist, um den zeitabhängigen Verlauf des Hörvermögens zu bestimmen (Phillips 1999). So fällt es Patienten mit reiner Worttaubheit schwer, eine Frequenzlücke zu erkennen und sie einzuordnen. Obwohl sich zwischen Legasthenie und reiner Worttaubheit offenbar zahlreiche Parallelen auftun, die immer wieder darauf hinweisen, dass auch der reinen Worttaubheit eine sprachunspezifische Störung zugrunde liegt, bei der bestimmte Frequenzspektren akustischer Stimuli nicht erfasst werden können, wurden im Gegensatz zur Therapie der Legasthenie bei der reinen Worttaubheit bisher keine therapeutischen Verfahren entwickelt, welche insbesondere die Verarbeitung schneller akustischer Reize im Gehirn fördern. Der Grundsatz der zur Zeit üblichen therapeutischen Verfahren heißt vielmehr noch immer „ üben, üben“ , und zwar das Einüben von sprachlichen Elementen – ein Grundsatz, der nach den Erfolgen der neuen LRS-Therapieverfahren überdacht werden sollte. Literatur 162 /LWHUDWXU Alexander AW, Anderson HG, Heilman PC, Voeller KKS, Torgesen JK. Phonological awareness training and remediation of analytic decoding deficits in a group of severe dyslexics. Annals of Dyslexia 41, 1991, 193-206 Algeria J, Pignot E, Morais J. Phonetic analysis of speech and memory codes in beginning readers. Memory and Cognition 10, 1982, 451-456 Altmann T. Thema: Legasthenie und Lese-Rechtschreibschwäche. http://www.theresa_LRS.htm, 14.9.2001 Ambrose J. Computerized transverse axial scanning (tomography) II. Clinical application. Br L Radiol 46, 1973, 1023-1047 Auerbach AH, Allard T, Naeser M, Alexander MP, Albert ML. Pure word deafness: Analysis of a case with bilateral lesions and a defect at the prephonemic level. Brain 105, 1982, 271-300 Baddeley A, Wilson A B. A case of word deafness with preserved spatial implications for the structure and function of short-ter memory, Brain 73, 203-223 Ball EW, Bachman BA. Phoneme segmentation training: effect on reading readiness. Annals of Dyslexia 38, 1988, 208-225 Bandettini PA, Wong EC, Hinks RS, Tikofsky RS, Hyde JS. Time course EPI of human brain function during task activation. Magn Reson Med 25, 1992, 390-397 Bandettini PA, Wong EC, Jesmanowicz A, Hinks RS, Hyde JS. Spin-echo and gradient-echo EPI of human brain activation using BOLD contrast: a comparative study at 1.5 T. NMR Biomed 7, 1994, 12-20 Barinaga M. Giving Language Skills a Boost. Science, 271, 1996, 27-28 Begley S. Rewiring Your Gray Matter. Newsweek, 2000 Benson DF, Geschwind N. Aphasia and related disorders: a clinical approach. In Baker AR, Joynt RJ (eds.). Clinical Neuoradiology 1, 1985, 193-238 Literatur 163 Benson DF, Patten DH. Meuse of radioactivc isotopes in the localization of aphasia-producing lesions. Corrtex, 3, 1967, 58-71 Benton AL, Joynt RJ. Early descriptions of aphasia. Arch Neurol 3, 1960, 205-222 Beratung Blicklabor http://www.brain.uni-freiburg.de/bbl 18.6.2001 Berthoz A, Graf W, Vidal PP (eds). The head-neck sensory motor system. Oxford University Press, Oxford 1992 Biscaldi M, Gezeck S, Stuhr V. Poor saccadic control correlates with dyslexia; Neuropsychologia 36, 11891202, 1998 Bisgaard ML, Eiberg H, Moller N, Niebuhr E, Mohr J. Dyslexia and chromosome 15 heteromorphism:negative lod score in a Danish material. Clin Genet 32, 1987, 118-119 Bloch F. Nuclear induction. Physiol Rev 70, 1946, 460–473 Boller F, Grafmann J (eds). Handbook of neuropsychology. Vol 1-4. Elsevier, Amsterdam, 1988-1992 Brain R. Speech Disorders. Aphasia, apraxia and agnosia. Washington, Butterwerth, 1961 Breitmeyer BG, Ganz L. Implications of sustained and transient channels for theories of visual pattern masking, saccadic suppression and information processing; Psychological Review 83, 1976, 1-36 Broca P. Remarques sur le siège de la faculté de language articué, suivie d'une observation d`aphémie (perte de la parole). Bull Soc Anat 36, 1861, 330-357 Bruck M. Word recognition skills of adults with childhood diagnoses of dyslexia. Developmental Psychology 26, 1990, 439-454 Bryant PE, Bradley L, MacLean M, Crossland J. Nurseery rhymes, phonological skills and reading. Journal of Child Language 16, 1989, 407-428 Bundesverband Legasthenie e.V. http://www.legasthenie.net 18.6.2001 Literatur 164 Bydder GM, Steiner RE, Young IR. Clinical NMR imaging of the brain: 140 cases. AJR Am J Roentgenol 3, 1982, 459-80 Cardon LR, Smith SD, Fulker DW, Kimberling WJ, Pennington BF, DeFries JC. Quantative trait locus for reading disability on chromosome 6. Science 266, 1994, 276-279 Castaigne P, Lhermitte F, Signoret JL, Abelante R. Description et étude scannographiqe du cerveeau de Leborgne (la découverte de Broca). Rev Neurol 136, 563-583 Cohen B (ed). Vestibular and oculomotor physiology. Ann NY Acad Sci, 1981, 374 Cohen R, Kelter S, Komeda M, Meier E. Sprache und Denken. In Von Stechow A, Schepping M (Hrsg.). Beiträge aus der Aphasieforschung. Fortschritte in der Semantik. Grammatikalische und sprachliche Prozesse. Acta Humaiora. Weinheim, 1998, 79-112 Cossu G, Shankweiler D, Liberman IY, Katz L, Tola G. Awareness of phonological segments and reading ability in Italien children. Applied Psycholinguistics 9, 1988, 1-16 Creutzfeldt OD. Cortex cerebri. Performance, structural and functional organization of the cortex. Max Planck Institut für Biophysikalische Chemie, Göttingen, 1993 Damadian R. Tumor detection by nuclear magnetic resonance. Science 1971, 1151-1153 Damasio H. Aphasia. N Engl J Med 326, 1992, 531-539 Damasio H. Neuroimaging contributions to the understanding of aphasia. Handbook of Neuropsychology 2, 1989, 3-46 Defior S, Tudela P. Effect of phonological training on reading and writing acquistion. Reading and Writing. 6, 1994, 299-320 Demb JB, Boyton GM, Heeger DJ. Brain activity in visual cortex predicts individual differences in reading performance. Proc Natl Acad Sci 94, 1997, 13363-13366 Dilling H, Mombour W, Schmidt MH. International Classification of Mental Diseases, ICD-10 (German edition). Huber, Bern, 1991 Literatur 165 Dinse HR, Godde B, Hilger T, Haupt SS, Spengler F, Zepka R. Short-Term Functional Plasticity of Cortical and Thalamic Sensory Representation and Its Implication for Information Processing, vol. 73 of Brain Plasticity. Advances in Neurology, chapter 13, 159-178 Dumont JJ, Oud J, van Mameren-Schoehuizen G, Jakobs M, van Herpen, M, van Bekerom F. Effectiveness of dyslexia treatment. In Pavlidis GT. (Hrsg.) Perspectives on dyslexia, vol 2, 293-325, Wiley, Chichester 1990 Eden GF, VanMeter JW, Rumsey JM, Maisog JM, Woods RP, Zeffiro TA. Abnormal processing of visual motion in dyslexia revealed by functional brain imaging. Nature 382, 1996, 66-70 Ellis A, Young A. Human Cognitive Neuropsychology. Lawrence Erlbaum Asso, Hove, 1988, 144-160 Engelien A. [email protected], 25.2.2001 Esser G, Schmidt M. Die langfristige Entwicklung von Kindern mit Lese-Rechtschreibschwäche. Zeitschrift für Klinische Psychologie, 22, 1993, 100-116 Evans EF. Neuronal process for the detection of acoustic patterns and for sound localization. In Schmidt FO, Worden FG (eds). Neurosciences – Third Study Program. MIT Press, New York, 131, 1974 Felton RH, Naylor CE, Wood FB. Neuropsychological profile of adult dyslexics. Brain and Language 9, 1990, 485-497 Fergusson DM, Lynskey MT. Early reading difficulties and later conduct problems. J Child Psychol Psychiatry. Nov;38(8), 1997, 899-907 Fischer B. Beratung Blicklabor. http://www.brain.uni-freiburg.de/bbl 12.11.2001 Fischer B. F. FonoFix. http://www.optom.de 21.11.2001 Fisher B, Weber H. Saccadic reaction times of dyslexic and age-matched normal subjects. Perception 19, 1990, 805-818 Flanagan JL. Speech Analysis, Synthesis and Perception. Second Edition, Springer, Berlin, Heidelberg, New York, 1972 Literatur 166 Forschungsgruppe Lese-Rechtschreibstörung http://www.kjp.uni-marburg.de/kjp.legast/index.htm 18.6.2001 Frackowiak RS, Functional mapping of verbal memory and language. Trends Neurosci 17, 1994, 109-115 Frith, U. Beneath the surface of developmental dyslexia. In Patterson KE, Marshall JC, Coltheart M (Hrsg.): Surface dyslexia. London, 1985 Galaburda A, Livingstone M. Evidence for a magnocellular defect in developmental dyslexia. In Tallal P, Galaburga AM, LLinás, von Euler C. Temporal Information Processing in the Nervous System: Special Reference to Dyslexia and Dysphagia. Annals of The New York Academy of Sciences, 62, 1993, 70-82 Galaburda AM, Menard MT, Rosen GD. Evidence for aberrant anatomy in developmental dyslexia. Proc Natl Acad Sci USA, 91, 17, 1994, 8010-8013 Ganong WF. Lehrbuch der Medizinischen Physiologie. Springer, Berlin, Heidelberg, New York, 1974 Gauer O-H, Kramer K, Jung R (Hrsg). Physiologie des Menschen, Bd. 12: Hören, Stimmen, Gleichgewicht. Urban & Schwarzenberg, München, 1972 Gerthsen C, Kneser HO, Vogel H. Physik. Springer, Berlin, Heidelberg, New York, 1980 Giard MH, Perrin F, Pernier J, Bouchet P. Brain generators implicated in the processing of auditory stimulus deviance: a topographic event-related potential study. Psychophysiology 27, 1990, 627-640 Glogauer W. Rechtschreibleistung und Intelligenz. Eine empirische Untersuchung. Psychologie in Schule und Erziehung 24, 1977, 287-292 Godde B. Auditorischer Cortex: Repräsentation und Plastizität. http://www.neuroinformatik.ruhr-unibochum.de 21.11.2001 Godfrey JJ, Syrdal-Lasky AK, Millay KK, Knox CM. Performance of dyslexic children on speech perception tests. Journal of Experimental Child Psychology 32, 1981, 401-424 Goswani U. Phonological skills and learning to read. In: Tallal P, Galaburga AM, LLinás, von Euler C. Temporal Information Processing in the Nervous System: Special Reference to Dyslexia and Dysphagia. Annals of The New York Academy of Sciences, 62, 1993, 296-311 Grehl H, Reinhardt F, Neuberger J. Neurologie. Thieme, Stuttgart, New York, 2000 Literatur 167 Grigorenko EL, Wood FB, Meyer MS, Hart LA, Speed WC, Shuster A, Pauls DL. :Susceptibility loci for distinct components of developmental dyslexia on chromosome 6 and 15. Am J Hum Genet 60, 1997, :27-39 Grüsser O-J, Grehn F. Visuelles System. In Hierholzer K, Schmidt RF (Hrsg) Pathophysiologie des Menschen. VCH, Weinheim, 1991, Kap 30 Grüsser O-J, Grüsser-Cornehls U. Neuronal mechanisms of visual movement perception and some psychophysical and behavioral correlations. In Jung R (ed). Handbook of sensory physiology, vol VII/3a. Springer, Berlin, Heidelberg, New York, 1973, 333 Grüsser O-J, Grüsser-Cornehls U. Periodische Aktivierungsphasen visueller Neurone nach kurzen Lichtreizen verschiedener Dauer. Pflugers Arch Gesamte Physiol 275, 1962, 292 Grüsser O-J, Henn V. Okulomotorik und vestibuläres System. In Hierholzer K, Schmidt RF (eds) Pathophysiologie des Menschen. VCH, Weinheim, 1991, KaP 32 Grüsser O-J, Klinke R (Hrsg). Zeichenerkennung durch biologische und technische Systeme. Springer, Berlin, Heidelberg, New York, 1971 Grüsser O-J, Landis T. Visual agnosias and other disturbances of visual perception and cognition, vol XII. In Cronly-Dillon JR (ed). Vision and visual dysfunction. MacMillan, London, 1991 Gutbrod K, Nirkko A, Ozdoba C, Heinemann D, Ptak R, Schroth G, et al. Selektive Aktivierung der Sprachgebiete mit Hilfe der funktionellen Magnetresonanztomographie (FMRI). In Voss A (Hrsg.). Neurologische Rehabilitation Gestern-Heute-Morgen. In press 2000 Haffner J, Zerahn-Hartung C, Pfuller U, Parzer P, Strehlow U, Resch F. Effekte und Bedeutung von spezifischen Rechtschreibproblemen bei jungen Erwachsenen. Zeitschrift für Kinder- und Jugendpsychiatrie und Psychotherapie, 26, 1998, 124-135 Head H Aphasia and Kindred Disorders of Speech. Zambridge University Press, London, 1926 Heilman KM, Valenstein E (eds). Clinical neuropsychology. Third edition, Oxford University Press, Oxford, 1993 Herholz K, Reulen HJ, von Stockhausen HM, Thiel A, Ilmberger J, Keuler J et al. Preoperative activation and intraoperative stimulation of language-related areas in patients with glioma. Neursurgery 41, 1997, 12531262 Literatur 168 Herholz K, Thiel A, Wienhard K, Pietrzyk U, von Stockhausen HM, Krabe H, et al. Individual functional anatomy of verb generation. Neuroimage3, 1996, 185-194 Herrholz K, Reulen HJ, von Stockhausen HM, Thiel A, Ilmberger J, Keuler J et al. Preoperative activation and intraoperative stimulation of language-related areas in patients with glioma. Neursurgery 41, 1997, 12531262 Hesse G. Tagung für Hals-Nasen-Ohren-Ärzte, Hannover, Januar 2001 Hoien T, Lundberg I, Larsen JP, Tonnessen FE. Profiles of creading related skills in dyslexic families. Read Writ Interdisc J 1, 1989, :381-392 Holligan C, Johnston R. The use of phonological information by good and poor readers in memory and reading tasks. Memory and Cognition 16, 1988, 522-532 Hounsfield G. Computerized transverse axial scanning (tomography). I. Description of system. Br L Radiol 46, 1973, 46, 1016-1022 Huber P, Gutbrod K, Ozdoba C, Nirkko A, Lövblad O, Schroth G. Zur Geschichte der Aphasiologie und Sprachlokalisation im Gehirn. Schweiz Med Wochenschr 130, 2000, 49-59 Ijzendoorn, MH, Bus AD. Meta-analytic confirmation of the nonword reading deficit in developmental dyslexia. Reading Research Quarterly 29, 1994, 267-275 Jacksozz JH. Clinical remarks on cases of defects of expression (by words, writing, songs etc.) Lancet 1, 864, 604-605 Jernigan T, Hesselink J, Tallal P. Cerebral structure on magnetic resonance imaging in language-learning impaires children. Arch Neurol 48, 1991, 539-545 Jorm A, Share D. Phonological recoding and reading acquisition. Applied Psycholinguistics 4, 1983, 103-147 Jung R (Hrsg). Central processing of visual information. A: Integrative function and comparative data. Handbook of Sensory Physiology, Bd. VII/3B, Springer, Berlin, Heidelberg, New York, 1973 Kerstholt MT, Van Bon WHJ, Schreuder R. Training in phonemic segmentation: the effects of visual support. Reading and Writing 6, 1994, 361-386 Literatur 169 Khanna SM, Leonard DBG. Basilar membrane tuning in the cat Kochlea. Science 215, 1982, 305-306 Kleist K. Gehirnpathologie. Barth, Leipzig, 1934 Klicpera C, Gasteiger-Klicpera B. Die langfristige Entwicklung der mündlichen Lesefähigkeit bei schwachen und guten Lesern. Zeitschrift für Entwicklungspsychologie und Pädagogische Psychologie 26, 1994, 278-290 Klicpera C, Schabmann A, Gasteiger-Klicpera B. Lesen- und Schreibenlernen während der Pflichtschulzeit: Eine Längsschnittuntersuchung über die Häufigkeit und Stabilität von Lese- und Rechtschreibschwierigkeiten in einem Wiener Schulbezirk. Zeitschrift für Kinder- und Jugendpsychiatrie, 21, 1993, 214-255 Klinke R. Die Verarbeitung von Schallreizen im Innenohr. HNO 35, 1987, 139 Klinke R. Gleichgewichtssinn, Hören, Sprechen. In Schmidt RF, Thews G (Hrsg). Physiologie des Menschen, 24. Auflage, Springer, Berlin, 1990 Klinke R. Neurotransmission in the inner ear. In Flock A, Wersäll J (eds). Cellular mechanisms in Hearing. Elsevier, Amsterdam, 1986, 235-244 Kochnower R, Richardson E, DiBenedetto BA. Comparison of the phonic decoding ability of normal and learning disabled children. Journal of Learning Disability 16, 1983, 348-351 Kohn SE, Friedman RB. Word-Meaning Deafness: A Phonlogical-Semantic Dissociation. Cognitive Neuropsychology 3 (3), 1986, 291-308 Kolb B, Whishaw IQ. Human neuropsychology. Third edition, Freeman, New York, 1990 Kosslyn SM, Andersen RA (eds). Frontiers in cognitive neuroscience. MIT Press, Carnbridge, Massachusetts, 1992 Kotchen TA, Guthrie GP. Renin-angiotensin-aldosterone and hypertension. Endocr Rev 5, 1, 1980, 78-99 Küspert P. Phonologische Bewußtheit und Schriftspracherwerb. Peter Lang, Frankfurt/Main, 1998 Landerl K, Wimmer H. Phonologische Bewusstheit als Prädiktor für Lese- und Schreibfertigkeiten in der Grundschule. Zeitschrift für Pädagogische Psychologie 8, 1994, 153-164 Literatur 170 Lauterburg P. Image formation by inducted local interactions: examples employing nuclear magnetic resonance. Nature 242, 1973, 190-191 LeBihan D. Functional Magnetic Resonance Imaging of the brain. Ann Intern Med 122, 1995, 296-303 Lenneberg EH. Biologische Grundlagen der Sprache, Suhrkamp, Franke, 1973 Lewis C, Hitch, GJ, Walker P. The prevalence of specific arithmetic difficulties and specific reading difficulties in 9- to 10-year-old boys and girls. Journal of Child Psychology and Psychiatry 35, 1994, 283-92 Liberman IY, Shankweiler D. Phonology and the problems of learning to read and write. Rem Spec Educ 6, 1985, 8-17 Lichtheim L. On aphasia. Brain 7, 1885, 433-484 Lichtheim L. Über Aphasie. Dtsch Arch Klin Med 36, 1885, 204-268 Livingstone MS, Hubel DH. Segregation of form, color, movement and depth: anatomy, physiology and perception. Science 240, 1988, 740 LLinas R. Is dyslexia a dischronia? In: Tallal P, Galaburga AM, LLinás, von Euler C. Temporal Information Processing in the Nervous System: Special Reference to Dyslexia and Dysphagia. Annals of The New York Academy of Sciences, 62, 1993, 70-82 Lundberg I, Frost J, Peterson O. Effects of an extensive programme for stimulating phonological awareness in pre-school children. Reading Research Quaterly 23, 1988, 163-184 Lundberg I, Olofsson A, Wall S. Reading and spelling skills in the first years predicted from phonemic awareness in kindergarten. Scandinavian Journal of Psychology 21, 1980, 159-173 Luria AR. Basic problems in neurolinguistics. The Hague, Mouton, 1976 Luria AR. Die höheren kortikalen Funktionen des Menschen und ihre Störungen bei örtlichen Hirnschädigungen. VEB Deutscher Verlag der Wissenschaften, Berlin, 1970 Manis FR, Mc-Bride-Chamg C, Seidenberg MS, Keating P, Doi, LM, Munson B, Petersen A. Are speech perception deficits associated with developmental dyslexia? Journal of Experimental Child Psychology 66 (2), 1997, 211-235 Literatur 171 Manley JA, Müller-Preuss P. Response variability of auditory cortex cells in the squirrel monkey to constant stimuli. Exp Brain Res 32, 1978, 171-180 Mansfield P. Multi-planar image formation using NMR spin echoes. J Phys C: Solid State Phys 10, 1977, L55–L58 Mansfield P. Proton spin imaging by NMR. Contemp Phys 17, 1976, 553-576 Marie P. Revision de la question de l'aphasie. La troisième circonvolution frontale gauche ne joue aucun rôle spécial dans la fonction du language. Sem Médicale 26, 1906, 241-247 Marx H, Jansen H, Mannhaupt G, Skowronek, H. Prediction of difficulties in reading and spelling on the basis of the Bielefelder screening. In Grimm H, Skowronek H (Hrsg). Language acquisition problems and reading disorders: Aspects of diagnosis and intervention. Berlin, 1993 Maunsell u. Newsome. Visual processing in monkey extrastriate cortex. Annu Rev Neurosci 10, 1987, 363 May P. Rechtschreiblernen in Ost und West. In Niemeyer W. Kommunikation und LeseRechtschreibschwäche. Winkler, Bochum, 1995 Mayer KC. http://mitglied.tripod.de/NeurologePsychiater/glossar.htm, 14.12.2001 Merigan WH, Maunsell JHR. How parallel are the primate visual pathways? Annual Review of Neuroscience, 16, 1993, 369-402 Merzenich MM, Schreiner WJ, Wang X. Neural Mechanims Underlying Temporal Integration, Segmentation, and Input Sequence Representation: Some Implications for the Origin of Learning Diabilities. In: Tallal P, Galaburga AM, LLinás, von Euler C. Temporal Information Processing in the Nervous System: Special Reference to Dyslexia and Dysphagia. Annals of The New York Academy of Sciences, 62, 1993, 1-22 Mezger GH. Die Aphasie. http://home.algaeu.org 21.11.2001 Moseley ME, Glover GH. Functional MR imaging – capabilities and limitations. Neuroimaging Clin N Am 5, 1995, 161-91 Müller E. Erste Hinweise zum Erkennen von LRS/Legasthenie für Eltern und Schüler http://www.vcg.ba/schulpsy/LRS/rsd.html, 14.9.2001 Literatur 172 Näslund JC. The interrelationships among preschool predictors of reading acquisition for German children. Reading and Writing 2, 1990, 327-360 Notoya M, Suzuki S, Furukawa M, Enokido H. A case of pure word deafness assosiated with LandauKleffner Syndrome: a long-term study of auditory disturbance. Auris Nasus Larynx (Tokyo) 18, 297-305, 1991 Ogawa S, Tank DW, Menon R, Ellerman JM, Kim SG, Merkle H et al. Intrinsic signal changes accompanying sensory stimulation: functional brain mapping with magnetic resonance imaging. Proc Natl Acad Sci USA 89, 1992, 5951-5955 Ojeman G, Ojeman J, Lettich E, Berger M. Cortical language localization in left dominant hemisphere. J Neurosurg 71, 1989, 316-326 Olson RK, Rack JP, Conners FA. Eye movements and their heritability in dyslexic and normal readers. In Schmid, R, Zambarbieri D (Eds.). Oculomotor control and cognitive processes, Amsterdam: North-Holland, 1991, 503-513 Olson RK, Forsberg H, Wise B. Genes, environment, and development of orthographic skills. In VW Berninger (Hrsg) The varieties of orthographic knowledge I: theoretical and developmental issues. Kluwer, Dordrecht, 1994, 27-71 Olson RK, Wise B, Conners F, Rack J. Organization, heritability, and remediation of component word recognition and language skills in disabled readers. In: Carr T, Levy BA. (Hrsg.). Reading and its development: component skills approaches. New York, 1990 Otsuki M, Soma Y, Sato M, Homma A, Tsuji S. Slowly Progressive Pure Word Deafness. Eur Neurol 39, 1998, 135-140 Paulesu E, Frith U, Snowling M, Gallagher A, Morton J, Frackowiak RS, Frith CD. Is developmental dyslexia a disconnection syndrome? Evidence from PET scanning. Brain 119, 1996, 143-157 Penfield W, Boldrey E. Somatic motor and sensory representation in the cerebral Kortex of man as studied by electrical stimulation. Brain 60, 1937, 381-443 Penfield W, Roberts L. Speech and brain mechanisms. University Press, Princeton London, 1959 Literatur 173 Pennington BF, Gilger J, Pauls D, Smith SSA, Smith SD, DeFries JC. Evidence for a major gene transmission of developmental dyslexia. JAMA. 18, 1991, 1527-1534 Pennington BF, Van Orden GC, Smith SD, Green PA, Haith MM. Phonological processing skills and deficits in adult dyslexics. Child Development 61, 1990, 1753-1778 Phillips DP. Response timing constraints on the cortical representation of sound time structure. J Acoust Soc Am 88 (3), 1990, 1403-1411 Phillips DP. Neural representation of stimulus times in the primary auditory cortex. In: Tallal P, Galaburga AM, LLinás, von Euler C. Temporal Information Processing in the Nervous System: Special Reference to Dyslexia and Dysphagia. Annals of The New York Academy of Sciences, 62, 1993, 104-118 Phillips PD. Central Auditory Processing: A View from Auditory Neuroscience. The American Journal of Otology, Vol 16, No. 3, 338-352, 1995 Phillips PD. Detection of silent intervals between noises activating different perceptual channels: Some properties of “ central“ auditora gap detection. J Acoust Soc Am 101, 6, 1997, 3694-3705 Phillips DP. Auditory Gap Detection, Perceptual Channels and Temporary Resolution in Speech Perception. J Am Acad Audiol 10, 1999, 343-354 Phillips DP, Farmer ME. Acquired word deafness, and the temporal grain of sound representation in the primary auditory cortex. Behavioural Brain Research, 40, 1990, 85-94 Pickles JO. An Introduction to the Physiology of Hearing. Second edition. Academic Press, 1988 Plath P. Zentrale Hörstörungen. Schriftenreihe GEERS-Stiftung Band 10, 1995 Poeppel D. A critical review of PET studies of phonological processing. Brain and Language 55, 1996, 317351 Polyak S. The vertebrate visual system. University of Chicago Press, Chicago, 1957 Pöppel E. A hierarchical model of temporal perception. Trends in Cognitive Sciences 1, 1997, 56-61 Literatur 174 Pöppel E. Grenzen des Bewußtseins. DVA Stuttgart, 1985, 9-42 Praamsta P, Hagoort P, Maassen B, Curl T. Word deafness and auditory cortical function. A case history and hypothesis. Brain 114, 1991, 1197-1225 Purcell EM, Torrey HC, Pound RF. Resonance absorption by nuclear magnetic moments in a solid. Physiol Rev 69, 1946, 37-38 Rack JP, Snowling MJ, Olson RK. The nonword reading deficit in developmental dyslexia: A review. Reading Research Quarterly 27, 1992, 29-53 Raichle M. Positron emission tomography. In Mountcastle V (ed.). Handbook of Physiology. Bethesda, 1987 Reulen HJ, Schmid UD, Ilmberger J, Eisner W, Bise K. Tumorchirurgie im Sprachkortex in Lokalanästhesie. Nervenarzt 68, 1997, 813-824 Rihs F, Sturzenegger M, Gutbrod K, Schroth G, Mattle HP. Determination of language dominance: Wada test confirms functional transcranial Doppler sonography. Neurology 52, 1999, 1591-1596 Rumsey JM, Andreason P, Zametkin AJ, Aquino T, King AC, Hamburger SD, Pikus A, Rapoport JL, Cohen RM. Failure to activate the left temporoparietal cortex in dyslexia. An oxygen 15 positron emission tomographic study. Archives of Neurology 49, 1992, 527-534 Rutter M, Yule W. The concept of specific reading retardation. Journal of Child Psychology and Psychiatry 16, 1975, 181-197 Saffran EM, Marin OSM, Yeni-Komshian GH. An Analysis of Speech Perception in Word Deafness. Brain and Language 3, 1976, 209-228 Salmelin R, Service E, Kiesila P, Uutela K, Salonen O. Impaired visual word processing in dyslexia revealed with magnetoencephalography. Annals of Neurology 40, 1996, 157-162 SAS Institute Inc. SAS Language: Reference, Version 6. SAS Institute Inc., Cary, NC, 1990 Scheerer-Neumann G. Rechtschreibtraining mit rechtschreibschwachen Hauptschülern auf kognitionspsychologischer Grundlage: Eine empirische Untersuchung. Westdeutscher Verlag, Opladen, 1988. Literatur 175 Scherg M, Vajsar J, Picton TW. A source analysis of human auditory evoked potentials. Journal of Cognitive Neuroscience 1 1989, 336-355 Schmidt RF (Hrsg). Neuro- und Sinnespsychologie. Springer, Berlin, Heidelberg, New York, 1993 Schmidt RF, Thews G, Lang, F (Hrsg). Physiologie des Menschen. 28. Auflage. Springer Verlag, Berlin, Heidelberg, New York, 2000 Schneider W, Küspert P, Roth E, Vise M, Marx H. Short- and long-term effects of training phonological awareness in kindergarten: evidence from two German studies. Journal of Experimental Child Psychology 66, 1997, 311-340 Schneider W, Vise M, Reimers P, Blaesser B. Auswirkungen eines Trainings der sprachlichen Bewusstheit auf den Schriftspracherwerb in der Schule. Zeitschrift für Pädagogische Psychologie 8, 1994, 177-188 Schulte-Körne G, Deimel W, Bartling J, Remschmidt H. Die Bedeutung der auditiven Wahrnehmung und der phonologischen Bewusstheit für die Lese-Rechtschreib-Schwäche. Sprache, Stimme und Gehör 22, 1998a, 25-30 Schulte-Körne G, Deimel W, Bartling J, Remschmidt H. Auditory processing and dyslexia: evidence for a specific speech deficit. NeuroReport 9, 1998b,:337-340 Schulte-Körne G, Deimel W, Müller K. Gutenbrunner C, Remschmidt H. Familial aggregation of spelling disability. J Child Psychol Psychiat 37, 1996, 817-822 Schulte-Körne G, Deimel W, Remschmidt H. Die Bedeutung von phonologischer Rekodierfähigkeit und orthographischem Wissen für die Rechtschreibfähigkeit Erwachsener. Zeitschrift für Klinische Psychologie 26, 1997, 210-217 Schulte-Körne G, Grimm T, Nöthen MM, Müller-Myhsok B, Cichon S, Vogt IR, Propping P, Remschmidt H. Evidence for linkage of spelling disability to chromosome 15. Am J Hum Genet 63, 1998c, 279-282 Schulte-Körne G, Remschmidt H, Hebebrand J. Zur Genetik der Lese-Rechtschreib-Schwäche. Zeitschrift für Kinder- und Jugendpsychiatrie 21, 1993, 242-252 Literatur 176 Schulte-Körne G, Remschmidt H, Warnke A. Selektive visuelle Aufmerksamkeit und Daueraufmerksamkeit bei legasthenen Kindern. Eine experimentelle Untersuchung. Zeitschrift für Kinder- und Jugendpsychiatrie 19, 1992, 99-106 Sellick PM, Patuzzi B, Johnstone BM. Measurement of basilar membrane motion in the guinea pig using Moessbauer technique. J acoust Soc Amer 72, 1982, 131-141 Shaywitz SE, Fletcher JM, Holahan JM, Shneider AE, Marchione KE, Stuebing KK, Francis DJ, Pugh KR, Shaywitz BA.Persistence of dyslexia: the Connecticut Longitudinal Study at adolescence. Pediatrics. 1999 Dec;104(6):1351-9 Shaywitz SE, Shawitz BA, Fletcher Escobar MD. Prevalence of reading disability in boys and girls: results of the Connecticut longitudinal study. JAMA 264, 1990, 998-1002 Siegel LS, Ryan EB. Development of grammatical sensitivity, phonological. and short-term memory skills in normally achieving and learning disabled children. Developmental Psychology 24, 1988, 28-37 Smith SD, Kimberling WJ, Pennington BF. Screening for multiples genes influencing dyslexia.. Read Writ Interdisc J 3, 1991, 285-298 Smith SD, Kimberling WJ, Pennington BF, Lubs HA. Specific reading disability: identification of an inherited form through linkage analysis. Science 219, 1983, 1345-1347 Snowling MJ. Phonemic deficits in developmental dyslexia. Psychological Research. 43, 1981, 219-234 Snowling MJ. The development of grapheme-phoneme correspondence in normal and dyslexic readers. Journal of Experimental Child Psychology 29, 1980, 294-305 Spoendlin H. Innervation densities of the Kochlea. Acta oto-laryngol 73, 1972, 235 Stark LW, Giveen SC, Terdiman JF. Specific dyslexia and eye movements; IN: Stein, J. F. (ed): Vision and visual dysfunction, 12, Macmillian, London, 1991, 203-232 Stark RE, Heinz JM. Perception of stop consonants in children with expressive and receptive-expressive language impairements. Journal of Speech and Hearing Research 39, 1996, 676-686 Steffens ML, Eilers RE, Gross-Glenn K, Jallad B. Speech perception in adult subjects with familiar dyslexia. Journal of Speech and Hearing Research 35, 1992, 192-200 Literatur 177 Stein JF. Vision and visual dyslexia. Vol XIII. In von Cronly-Dillon JR (eds) Vision and visual dysfunction. MacMillan, London, 1991 Stevenson J. Which aspects of processing text mediate genetic effects? Read Writ Interdisc J 3, 1991, 249269 Stockert von TR. On the Structure of Word Deafness and Mechanisms Underlying the Fluctuation of Disturbances of Higher Cortical Functions, 1982 Talcott JB, Witton C, McClean MF, Hansen PC, Rees A, Green GGR, Stein FF. Dynamic sensory sensitivity and children’s word decoding skills. Proc Natl Acad Sci, USA 97, 2000, 2592-2957 Tallal P, Galaburga AM, LLinás, von Euler C. Temporal Information Processing in the Nervous System: Special Reference to Dyslexia and Dysphagia. Annals of The New York Academy of Sciences, 62, 1993, 122 Tallal P, Miller S, Fitch RH. Neurobiological Basis of Speech: A Case for the Preeminence of Temporal Processing. In: Tallal P, Galaburga AM, LLinás, von Euler C. Temporal Information Processing in the Nervous System: Special Reference to Dyslexia and Dysphagia. Annals of The New York Academy of Sciences, 62, 1993, 1-22 Tallal P, Piercy M. Defects of non-verbal auditory perception in children with developmental aphasia. Nature 241, 1973, 468-469 Tallal P, Sainburg R, Jernigan T. Neuropathology of developmental dysphasia. Read Writ 4, 1991, 65-79 Tallal P. Temporal Processing in the Nervous System – Special Reference to Dyslexia and Dysphasia. Annals of the New York Academy of Sciences, Volume 682, 1993, 442 Tallal P. The science of literacy: From the laboratory to the classroom. Proc Natl Acad Sci, USA 97, 2000, 2402-2404 Tallal P et al. Temporal processing deficits of language-learning impaired children ameliorated by training. Science 271, 1996, 77-80 Tallal P. Auditory temporal perception, phonics, and reading disabilities in children. Brain and Language 9, 1980, 182-198 Literatur 178 Tallal P, Miller S, Fitch RH. Neurobiological basis of speech: a case for the preeminence of temporal processing. In Tallal P, Galaburda A, MLlinás RR, Euler C von (Hrsg.): Temporal information processing in the nervous system: special reference to dyslexia and dysphasia. New York, 1993 Teuber HL. Alteration of perception and memory in man. In: Weiskrantz L, ed. Analysis of behavioral change. New Yoork, Harper Row, 1968 Tonndorf J. Dimensional analysis of Kochlear models. J acoust Soc Amer 32, 1980, 493-497 Tursky I. Lese-/Rechtschreibschwäche. http://www.bildungsservice.at/schulpsychologie/ docs/BeilageLRSS99.htm, 14.9.2001 van Essen DC. Visual areas of the mammalian cerebral cortex. Annu Rev Neurosci 2, 1979, 227 Wada J. A new method for determination of the side of cerebral speech dominance. A preliminary report on the intracarotid injection of sodium amytal in man. Med Biol 14, 1949, 221-222 Wagner RK, Torgesen JK, Rashotte CA. Development of reading related phonological processing abilities: new evidence of bidirectional causality from latent variable longitudinal study. Developmental Psychology 30, 1994, 73-87 Warnke A, Remschmidt H, Hennighausen K. Verbal information processing in dyslexia--data from a followup experiment of neuro-psychological aspects and EEG. Acta Paedopsychiatrica 56, 1994, 203-208 Warnke A. Legasthenie und Hirnfunktion. Bern 1990 Warnke F. CD Dyslexie und HörLateralität. VAK-Verlag, Freiburg 1996 Warnke F. CD-Lateraltraining 1 und 2. MediTECH, Wedemark, 1998 Warnke F. Der Takt des Gehirns. VAK-Verlag Freiburg, 1995 Warnke F. Was Hänschen nicht hört. VAK-Verlag Freiburg, 1995 Watson BU, Miller TK. Auditory perception, phonological processing, and reading ability/disability. Journal of Speech and Hearing Research 36, 1993, 850-863 Literatur 179 Weinschenk C, Funke EH, Gieseler L, Pieh KH, Winhold N. Über die Häufigkeit der kongenitalen Legasthenie im zweiten Grundschuljahr (II.). Psychologische Rundschau 19, 1968, 44-51 Werker JF, Tees RC. Speech perception in severely disabled and average reading children. Canadian Journal of Psychology 41, 1987, 48-61 Wernicke C. Der aphasische Symptomenkomplex. Eine psychologische Studie auf anatomischer Basis. Kohn und Weigert, Breslau, 1874 Wieser HG, Valavanis A, Roos A, Isler P, Renella RR. Selective and superselective temporal lobe amytal Tests: neuroradiological, neuroanatomical, and electrical data. In Manelis J, Bental F, Loeber JN, Dreifuss I (eds.). Advances in epileptology, Vol 17, Ravcn Press, New York, 1994, 20-27Békésy G von. Experiments in Hearing. McGraw Hill, New York, 1960 Wolff PH, Cohen C, Drake C. Impaired motor timing control in specific reading redardation. Neuropsychologia 22 (5), 1984, 587-600 Wolff PH. Impaired Temporal Resolution in Developmental Dyslexia. Annals of the New York Academy of Sciences, Volume 682, 1993 Yopp HK. The validity and reliability of phonemic awareness tests. Reading Research Quaterly 23, 1988, 159-177 Zenner HP, Zimmermann U, Gitter AH. Fast motility of isolated mammalian auditory sensory cells. Biochem biophys Res Commun 1, 1987, 304-308 Zenner HP, Zimmermann U, Schmitt U. Reversible contraction of isolated mammalian Kochlear hair cells. Hear Res 18, 1985, 127-133 Zenner HP. Hören. Thieme, 1985 Zenner HP. Motile responses in outer hair cells. Hear Res 22, 1986, 83-90 Zwicker E, Fastl H. Psychoacoustics – Facts & Models. Springer, Berlin, 1990