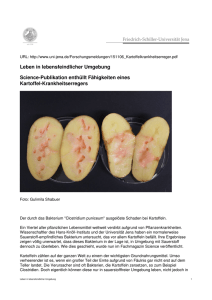
Charakterisierung der Wechselwirkung von Selen mit dem
Werbung

Thema: CHARAKTERISIERUNG DER WECHSELWIRKUNG VON SELEN MIT DEM BAKTERIUM AZOSPIRILLUM BRASILENSE MASTERARBEIT Zur Erlangung des akademischen Grades MASTER OF SCIENCE Studiengang Biotechnologie und Angewandte Ökologie vorgelegt von: Anika Maffert geboren am 01.03.1989 in Herzberg/Elster eingereicht am: 30.09.2013 Erstgutachter: Prof. Dr. rer. nat. habil. Thomas Wiegert Fakultät Mathematik/Naturwissenschaften Hochschule Zittau/Görlitz Zweitgutachter: Dr. rer. nat. Manja Vogel Institut für Ressourcenökologie Helmholtz-Zentrum Dresden-Rossendorf e.V. Zusammenfassung I. S e i t e | II ZUSAMMENFASSUNG Das Bakterium Azospirillum brasilense gehört zur Gruppe der Plant growth-promoting rhizobacteria (PGPR) (Kloepper et al. 1991), da es die Fähigkeit besitzt das Wachstum zu fördern und infolge dessen den Ertrag bestimmter Pflanzen zu erhöhen. Ein natürliches Habitat der Bakterien ist der Boden. Insbesondere lagert es sich an der Wurzeloberfläche von Nutzpflanzen, wie beispielsweise Mais, Weizen und Reis, an. Dabei geht das Bakterium A. brasilense mit den Pflanzen eine symbiotische Bindung ein. Das Spurenelement Selen (Se), welches in vielen Böden, Gesteinen und Gewässern vorkommt, ist essentiell für viele Organsimen. Zu beachten ist, dass die Aufnahme von Selen in höheren Konzentrationen toxisch und im Gegensatz dazu die Aufnahme von nur geringen Mengen zu einem Selendefizit führen kann. Aufgrund der Toxizität sind die wasserlöslichen Oxyanionen Selenit (SeO32-) und Selenat (SeO42-) von besonderem ökologischen Interesse. Aus der Literatur ist bekannt, dass einige Bakterienstämme die Fähigkeit besitzen Selenit und Selenat zu weniger toxischem elementaren Selen (Se(0)) zu reduzieren. Dieses elementare Selen bildet extra- oder intrazelluläre Nanopartikel aus. In der vorliegenden Arbeit wurde die Wechselwirkung des Bakteriums Azospirillum brasilense mit den wasserlöslichen Oxidationsstufen des Selens (Se(IV) und Se(VI)) unter aeroben Bedingungen untersucht. Dabei wurde die mikrobielle Reduktion von Selenit durch das Bakterium beobachtet, die Auswirkung der Variation der Selenitkonzentrationen untersucht und entstandene Se(0)-Nanopartikel charakterisiert. Die in der Arbeit untersuchten Aspekte, gaben Aufschlüsse über die Toleranzen des Bakteriums A. brasilense gegenüber Selenat und Selenit. Es wurde gezeigt, dass nur das Oxyanion Selenit toxisch auf das Bakterium wirkt. Das Selenat wurde während des Kultivierungszeitraumes nicht in den Metabolismus des Bakteriums aufgenommen. Durch die Variation der Selenitkonzentrationen von 1 mM bis 5 mM im Medium wurde herausgefunden, dass A. brasilense Konzentrationen bis 2,5 mM toleriert. Bei einer Selenitkonzentration von 5 mM konnte nicht eindeutig bestimmt werden, in wie weit das Bakterium diese Konzentration verträgt. Zusammenfassung S e i t e | III Die Ergebnisse der Hydrid-Generation Atomabsorptionsspektrometrie (HG-AAS) und der Massenspektrometrie mit induktiv gekoppeltem Hochfrequenzplasma (ICP-MS) verdeutlichten die zeitliche Abnahme der Selenitkonzentration im Medium und der damit verbundenen Umsetzung von Selenit zu Se(0)-Nanopartikeln. Die vollständige Umsetzung des Selenits aus dem Medium erfolgte ca. sieben Tage nach Beginn der Kultivierung bei einer Selenitkonzentration von 1 mM. Bei der Kultivierung des Bakteriums in Gegenwart von Selenit konnte festgestellt werden, dass die Bakterien Nanopartikel produzierten. Diese wurden mittels Lichtmikroskopie und Rasterelektronenmikroskopie (REM) visualisiert und konnten mittels energiedispersiver Analyse der Röntgenstrahlung (EDX) als Selen-Schwefel-Gemisch identifiziert werden. Dabei konnte gezeigt werden, dass die Nanopartikel außerhalb der Bakterienzelle lokalisiert sind. Mittels Dynamischer Lichtstreuung (DLS) wurde die Partikelgrößenverteilung und der damit verbundene durchschnittliche hydrodynamische Partikeldurchmesser von etwa 400 nm bestimmt. Durch die Zetapotentialmessung wurde ermittelt, dass die Partikel eine stark negativ-geladene Oberfläche besitzen und somit die Tendenz aufweisen keine Agglomerate zu bilden, da die Partikel sich aufgrund der Ladung abstoßen. Die Struktur dieser Selennanopartikel wurde mit Hilfe der Raman-Spektroskopie untersucht. Dabei wurden Selenketten, Se8-Ringstrukturen und Selen-Schwefel- Valenzschwingungen ermittelt. Durch Inkubieren der Nanopartikelsuspension bei unterschiedlichen Temperaturen im Bereich von 35 °C bis 250 °C wurde die Thermostabilität analysiert. Bis 35 °C konnte kein signifikanter Unterschied zur unbehandelten Probe mittels UV/Vis-Spektroskopie und DLS beobachtet werden. Ab 50 °C agglomerierten die Partikel, sodass eine instabile Suspension entstand. Je höher die Temperaturen stiegen, desto weniger blieb von den Nanopartikeln erhalten. Bei einer Inkubation bei 250 °C konnten keine Partikel mehr nachgewiesen werden. Diese Arbeit liefert einen wesentlichen Beitrag für die Aufklärung der Wechselwirkungen zwischen Mikroorganismen und giftigen Substanzen. Die Umwandlung toxischer Stoffe, wie beispielsweise Selenit, durch A. brasilense zu weniger toxischen elementaren Selen, kann für die Wasseraufbereitung oder für die Regenerierung von selenbelasteten Böden von Nutzen sein. Die Nanopartikel könnten in Industrie (Photovoltaik) und in der Medizin (Pharmakotherapie) Anwendung finden. Inhaltsverzeichnis II. S e i t e | IV INHALTSVERZEICHNIS I. ZUSAMMENFASSUNG................................................................................................... II II. INHALTSVERZEICHNIS ............................................................................................... IV III. ABKÜRZUNGSVERZEICHNIS ....................................................................................... VI IV. ABBILDUNGSVERZEICHNIS .....................................................................................VIII V. TABELLENVERZEICHNIS ............................................................................................ XI 1 EINLEITUNG ................................................................................................................. 1 2 THEORETISCHE GRUNDLAGEN ................................................................................... 3 2.1 Das Bakterium Azospirillum brasilense ............................................................... 3 2.2 Selen ........................................................................................................................ 4 2.2.1 Allgemeines ................................................................................................... 4 2.2.2 Vorkommen ................................................................................................... 5 2.2.3 Verwendung ................................................................................................... 6 3 MATERIAL UND METHODEN ....................................................................................... 7 3.1 Chemikalien ........................................................................................................... 7 3.2 Bakterienstamm..................................................................................................... 8 3.3 Kulturmedien und Lösungen ............................................................................... 8 3.3.1 Azo-Medium zur Kultivierung von Azospirillum brasilense......................... 8 3.3.1.1 Stammlösung für das Azo-Medium ................................................... 9 3.3.1.2 Spurenelementlösung (10-fach) ......................................................... 9 3.3.2 Reagenzien für die Proteinbestimmung ......................................................... 9 3.3.2.1 Lösung A .......................................................................................... 10 3.3.2.2 Lösung B .......................................................................................... 10 3.3.3 Salzlösung zur Isolierung der Nanopartikel ................................................. 10 3.4 Zellaufschlusspuffer/Lagerungspuffer .............................................................. 11 3.4.1 Kaliumphosphatpuffer (0,1 M) .................................................................... 11 3.4.2 PBS-Puffer ................................................................................................... 11 3.4.3 Lysispuffer ................................................................................................... 11 3.5 Methoden .............................................................................................................. 12 3.5.1 Erhaltung der Stammkultur .......................................................................... 12 3.5.2 Kultivierung von A. brasilense .................................................................... 12 3.5.3 Chemische Reduktion von Selenit und Selenat ........................................... 13 3.5.4 Mikrobielle Reduktion von Selenit und Selenat .......................................... 13 3.5.5 Lichtmikroskopie ......................................................................................... 14 3.5.6 Zellaufschluss mittels basischer Verseifung ................................................ 14 3.5.7 Proteinbestimmung ...................................................................................... 14 3.5.8 Massenspektrometrie mit induktiv gekoppeltem Plasma (ICP-MS) ........... 16 3.5.9 Hydrid-Generation (Hydridtechnik) Atomabsorptionsspektrometrie (HG-AAS) .................................................................................................... 16 3.5.10 Isolierung der Nanopartikel ......................................................................... 17 3.5.11 UV/Vis-Spektroskopie ................................................................................. 18 3.5.12 Rasterelektronenmikroskopie in Kombination mit der energiedispersiven Röntgenspektroskopie (REM-EDX) ............................................................ 18 3.5.13 Transmissionselektronenmikroskop in Kombination mit der energiedispersiven Röntgenspektroskopie (TEM-EDX) ............................. 20 3.5.14 Bestimmung des Zetapotentials ................................................................... 20 3.5.15 Dynamische Lichtstreuung (DLS) ............................................................... 22 Inhaltsverzeichnis 4 5 6 7 8 9 Seite |V 3.5.16 Raman-Spektroskopie der Nanopartikel ...................................................... 23 3.5.17 Thermostabilitätstest .................................................................................... 23 3.5.18 Bestimmung des Totalen organischen Kohlenstoffgehalts (TOC) .............. 24 ERGEBNISSE ............................................................................................................... 26 4.1 Wachstumskurve von A. brasilense ................................................................... 26 4.2 Reduktion von Selenat und Selenit durch das Bakterium A. brasilense ........ 28 4.2.1 Wachstumsverlauf in Gegenwart von Selen ................................................ 30 4.2.2 Messung der Selenit- und Selenatkonzentration .......................................... 33 4.2.3 Einfluss der Konzentration auf die mikrobielle Reduktion von Selenit und das Wachstum von A. brasilense .......................................................... 36 4.2.4 Wachstumsverlauf des Bakteriums A. brasilense bei unterschiedlichen Selenitkonzentrationen ................................................................................. 37 4.2.5 Messung der unterschiedlichen Selenitkonzentrationen .............................. 40 4.3 Charakterisierung der Nanopartikel ................................................................. 41 4.3.1 Farbgebung Nanopartikelsuspension nach der Isolierung ........................... 41 4.3.2 Elektronenmikroskopie in Kombination mit der energiedispersiven Röntgenspektroskopie (EDX) ...................................................................... 44 4.3.3 Bestimmung der Oberflächenladung durch Zetapotentialmessungen ......... 48 4.3.4 Dynamische Lichtstreuung .......................................................................... 49 4.3.5 Raman-Spektroskopie .................................................................................. 54 4.3.6 Thermostabilitätstest .................................................................................... 55 4.3.6.1 UV/Vis-Spektroskopie ..................................................................... 56 4.3.6.2 Dynamische Lichtstreuung............................................................... 58 4.3.6.3 Bestimmung des TOC-Gehaltes ....................................................... 60 DISKUSSION ................................................................................................................ 62 5.1 Reduktion von Selenit/Selenat ............................................................................ 62 5.2 Charakterisierung der Nanopartikel ................................................................. 65 AUSBLICK................................................................................................................... 70 LITERATURVERZEICHNIS .......................................................................................... 72 EIDESSTATTLICHE ERKLÄRUNG ............................................................................... 76 DANKSAGUNG ............................................................................................................ 77 Abkürzungsverzeichnis III. S e i t e | VI ABKÜRZUNGSVERZEICHNIS °C Grad Celsius A. brasilense Azospirillum brasilense Azo-Medium Medium für Azospirillum brasilense BCA Bicinchoninic Acid BSA Bovine Serum Albumin (Rinderserumalbumin) c Konzentration Cu Kupfer d.nm Durchmesser in nm DLS Dynamische Lichtstreuung DSMZ Deutsche Sammlung von Mikroorganismen und Zellkulturen EDX Energy Dispersive X-Ray (Energiedispersive Analyse der Röntgenstrahlung) g Fallbeschleunigung HG-AAS Hydrid-Generation Atomabsorptionsspektrometrie ICP-MS Inductive Coupled Plasma Mass Spectrometry (Massenspektrometrie mit induktiv gekoppeltem Hochfrequenzplasma) K Kontrolle konz. konzentriert(e) min. Minute(n) MW Mittelwert m/z Verhältnis Masse zu Ladung N Atomanzahl im Molekül OD Optische Dichte pH Negativ dekadischer Logarithmus der Wasserstoffionenkonzentration PBS Phosphate Buffered Saline (Phosphatgepufferte Salzlösung) pI Isoelektrischer Punkt REM Rasterelektronenmikroskopie RT Raumtemperatur S Schwefel Se Selen Abkürzungsverzeichnis S e i t e | VII Se(0) Selen mit der Oxidationsstufe 0 Se(-II) Selen mit der Oxidationsstufe -2 Se(IV) Selen mit der Oxidationsstufe 4 Se(VI) Selen mit der Oxidationsstufe 6 St.Abw. Standardabweichung t Zeit td Verdopplungszeit TEM Transmissionselektronenmikroskopie TOC Total Organic Carbon (Totaler organischer Kohlenstoff) TRIS Trishydroxymethylaminomethan UV Ultraviolett (ultravioletter Spektralbereich) µ Wachstumsrate VE-Wasser vollentsalztes Wasser Vis Visible (sichtbarer Spektralbereich) x0 OD-Wert am Anfang der exponentiellen Phase xt OD-Wert am Ende der exponentiellen Phase Abbildungsverzeichnis IV. S e i t e | VIII ABBILDUNGSVERZEICHNIS Abbildung 1: Reduktion von Selenat zu Selenit und elementaren Selen ........................ 2 Abbildung 2: Bakterium A. brasilense auf festem Nährmedium ................................... 3 Abbildung 3: Mikroskopische Aufnahme von A. brasilense .......................................... 3 Abbildung 4: Farbgebung nach 1 h Inkubation ............................................................. 15 Abbildung 5: BSA-Eichkurve zur Proteinbestimmung mit dem BCA-Reagenz .......... 15 Abbildung 6: Typische Kurve nach Auftragen des Zetapotentials gegen den pH-Wert (Malvern Instruments 2007) .................................................................... 22 Abbildung 7: Stahlautoklav mit eingesetzten Teflonbecher ......................................... 24 Abbildung 8: Bestimmungsformen des Kohlenstoffs ................................................... 25 Abbildung 9: Logarithmische Darstellung der Wachstumskurve von A. brasilense .... 26 Abbildung 10: Farbverlauf der Kontrolle ........................................................................ 28 Abbildung 11: Farbverlauf der Selenatkultur .................................................................. 28 Abbildung 12: Farbverlauf der Selenitkultur .................................................................. 28 Abbildung 13: Farbliche Entwicklung der Kontroll-, Selenat- und Selenitkultur nach sieben Tagen Kultivierung ...................................................................... 29 Abbildung 14: Mikroskopische Aufnahme von A. brasilense im Kontrollmedium (ohne Nanopartikel) ................................................................................ 30 Abbildung 15: Mikroskopische Aufnahme von A. brasilense mit Nanopartikeln im Selenatansatz (ohne Nanopartikel) .......................................................... 30 Abbildung 16: Mikroskopische Aufnahme von A. brasilense mit Nanopartikeln im Selenitansatz ............................................................................................ 30 Abbildung 17: Hellfeldaufnahme der Nanopartikel in der Selenitkultur ........................ 30 Abbildung 18: Logarithmische Darstellung der OD-Werte der Kontroll-, Selenit- und Selenatkultur ........................................................................................... 31 Abbildung 19: Darstellung der Gesamtzellproteinkonzentration der Kontroll-, Selenitund Selenatkultur .................................................................................... 32 Abbildung 20: Vergleich der Messungen der Selenitkonzentrationen der HG-AAS und der ICP-MS ............................................................................................. 34 Abbildung 21: Zeitlicher Verlauf der Selenkonzentration im Selenit- und Selenatansatz ........................................................................................... 35 Abbildung 22: Zeitlicher Verlauf der Selenat- und Selenitintensität und den daraus resultierenden Selennanopartikelanteil ................................................... 36 Abbildungsverzeichnis Abbildung 23: S e i t e | IX Farbentwicklung nach sieben Tagen bei unterschiedlichen Selenitkonzentrationen ............................................................................ 37 Abbildung 24: Wachstumskurven der Kontrollkultur und den Ansätzen mit unterschiedlichen Selenitkonzentrationen ............................................... 37 Abbildung 25: Zeitlicher Verlauf der Gesamtzellproteinkonzentration der Kontrollkultur und den Ansätzen mit unterschiedlichen Selenitkonzentrationen ............................................................................ 39 Abbildung 26: Zeitlicher Verlauf der Gesamtselenkonzentration der unterschiedlichen Selenitkonzentrationen ............................................................................ 40 Abbildung 27: Suspension der isolierten Nanopartikel (1 mM-Selenitansatz (Ansatz 1)) ............................................................................................... 42 Abbildung 28: Suspension der isolierten Nanopartikel (1 mM-Selenitansatz (Ansatz 2)) ............................................................................................... 42 Abbildung 29: Suspension der isolierten Nanopartikel (2,5 mM-Selenitansatz) ............ 42 Abbildung 30: Suspension der isolierten Nanopartikel (5 mM-Selenitansatz) ............... 42 Abbildung 31: Chemischen Reduktion von Selenit (Bildung von rotem Niederschlag) 43 Abbildung 32: Chemische Reduktion von Selenat (kein Niederschlag) ......................... 43 Abbildung 33: UV/Vis Spektren des chemisch reduzierten Selenits .............................. 43 Abbildung 34: A. brasilense mit Nanopartikeln aus dem Selenitansatz (Methode ohne Fixierung) ................................................................................................ 44 Abbildung 35: Nanopartikel aus dem Selenitansatz (Methode ohne Fixierung) ............ 44 Abbildung 36: A. brasilense mit Nanopartikeln aus dem Selenitansatz (Fixierung mit Glutaraldehyd) ......................................................................................... 45 Abbildung 37: EDX-Spektrum der Nanopartikel ............................................................ 45 Abbildung 38: Bakterien ohne Nanopartikel aus dem Selenatansatz (Fixierung mit Glutaraldehyd)......................................................................................... 46 Abbildung 39: EDX-Spektrum aus dem Selenatansatz ................................................... 46 Abbildung 40: TEM-Aufnahme der Nanopartikel .......................................................... 47 Abbildung 41: EDX-Spektrum der Nanopartikel ............................................................ 47 Abbildung 42: EDX-Spektrum der von Nanopartikel umgebenden Substanz ................ 48 Abbildung 43: Zetapotentiale aller Nanopartikelsuspensionen der Selenitansätze mit Mittelwert aller Zetapotentialmessungen mit Standardabweichung ....... 48 Abbildung 44: Partikelgrößenverteilung der 1 mM-Selenitansätze ................................ 49 Abbildung 45: Autokorrelationsfunktion ........................................................................ 51 Abbildungsverzeichnis Seite |X Abbildung 46: Durchschnittliche Partikelgrößen der 1 mM-Selenitansätze ................... 52 Abbildung 47: Durchschnittliche Partikelgrößen der 2,5 mM-Selenitansätze ................ 52 Abbildung 48: Partikelgrößenverteilung der Nanopartikel bei unterschiedlichen Selenitkonzentrationen ............................................................................ 53 Abbildung 49: Darstellung der durchschnittlichen Partikelgröße aller 1 mMSelenitkulturen und Ermittlung des durchschnittlichen Durchmessers .. 54 Abbildung 50: Raman-Spektrum der Nanopartikelsuspension ....................................... 55 Abbildung 51: Aufnahme der Ausgangslösung und der bei unterschiedlich hohen Temperaturen inkubierten Nanopartikelsuspension ................................ 56 Abbildung 52: UV/Vis-Spektren der Nanopartikelsuspension bei unterschiedlichen Inkubationstemperaturen ......................................................................... 57 Abbildung 53: UV/Vis-Spektren der Nanopartikelsuspension bei einer Inkubation von 100 °C und 250 °C .................................................................................. 58 Abbildung 54: Partikelgrößenverteilung der Ausgangslösung und der Proben, die bei 35 °C, 50 °C und 100 °C inkubiert wurden............................................. 59 Abbildung 55: Grafische Darstellung des TOC-Gehaltes der Nanopartikelsuspension vor und nach der Inkubation bei unterschiedlichen Temperaturen ............... 60 Tabellenverzeichnis V. S e i t e | XI TABELLENVERZEICHNIS Tabelle 1: OD-Werte und die jeweiligen Zeiten am Anfang und Ende der exponentiellen Phase, Wachstumsrate und Verdopplungszeit ................ 27 Tabelle 2: Zeitpunkt und OD-Wert des Maximums der Kontroll-, Selenat- und Selenitkultur ............................................................................................ 31 Tabelle 3: Zeitpunkt und OD-Wert der Maxima der unterschiedlichen Selenkonzentrationen .............................................................................. 38 Tabelle 4: Zeitpunkt und Proteinkonzentration der Maxima der unterschiedlichen Selenkonzentrationen .............................................................................. 39 Tabelle 5: Vergleich der berechneten und mittels ICP-MS gemessenen Selenitanfangskonzentrationen ............................................................... 40 Tabelle 6: Durchschnittliche Partikeldurchmesser der verschiedenen Ansätze ....... 50 Tabelle 7: Inkubationstemperaturen mit dazugehörigem durchschnittlich gemessenem Durchmesser ...................................................................... 59 Einleitung 1 Seite |1 EINLEITUNG Ein schwerwiegendes Problem für die Umwelt ist die Verschmutzung durch umweltschädigende Substanzen, welche immer mehr auf Aktivitäten der Menschen zurückzuführen ist. Die Quellen dafür sind meistens landwirtschaftlich oder industriell verursachte Eintragspfade. Umweltbiologen und ökologische Risikobewerter beschäftigen sich mit Schadstoffen, die eine hohe Gefährdung der Umwelt aufweisen, wobei häufig Stoffe, wie Selen, vernachlässigt werden. Doch gerade Selen weist ernste langfristige Risiken speziell für die aquatischen Lebensräume und die damit verbundenen Nahrungspfade auf. Werden diese Lebensräume mit Selen verschmutzt, so folgt daraus eine Anreicherung in allen mit dem Wasserkreislauf verbundenen Organismen. Durch Aufnahme von Nahrung, wie zum Beispiel Pflanzen und Tieren, die auf selenhaltigen Boden vorkommen, nimmt auch der menschliche Körper unbewusst Selen zu sich. Eine erhöhte Selenkonzentration hat einen negativen Einfluss auf die menschliche Gesundheit und auch auf die Landwirtschaft. Durch Aufnahme und Bioakkumulation in den Organismen in den verschiedenen Nahrungsebenen ist ein gezielter Eingriff für die Unterbrechung der Aufnahme von Selen nicht gewährleistet (Lemly 2004). Selen ist in der Natur Bestandteil eines biogeochemischen Zyklus, bei dem mit Hilfe von mikrobiell ablaufenden Redoxreaktionen jede Oxidationsstufe des Selens erreicht werden kann. Einige Bakterien nutzen während der Respiration des Kohlenstoffs Selenat und Selenit als Elektronenakzeptor in ihrem Energiestoffwechsel. Dabei ist zwischen Bakterien zu unterscheiden, die zum Einen anaerob und zum Anderen aerob die Oxyanionen reduzieren (Fernández-Martínez 2009). Somit besitzen beispielsweise einige Bakterien die Fähigkeit Selenat zu ungiftigen elementarem Selen zu reduzieren (Zhang et al. 2003). Herbel et al. (2003) beschreiben, dass einige Bakterienstämme elementares Selen zu Selenid reduzieren oder Selenid zu elementaren Selen oxidieren können. Andere wiederum können elementares Selen oder Selenid zu Selenit oder Selenat oxidieren (Sarathchandra & Watkinson 1981, Levanony & Bashan 1991, Dowdle & Oremland 1998). Bis zum heutigen Tage sind 16 Spezies von Bakterien und Archaeen bekannt, bei denen unter anaeroben Bedingungen organisches Material oder Wasserstoff oxidiert wird, um Selenoxyanionen zu reduzieren (Stolz & Oremland 1999). Einleitung Seite |2 Eine Besonderheit einiger Bakterienstämme ist es, dass sie in der Lage sind, das giftige Selenit oder Selenat in Form von Nanopartikeln umzusetzen. Dieser Prozess der Entgiftung wird als Bioremediation bezeichnet, bei dem Bakterien zur Sanierung des kontaminierten Bodens oder selenhaltiger Gewässer beitragen können. Es beschreibt die durch Organismen generierte biologische Umwandlung, um kontaminierte biologische Systeme zu sanieren (Frankenberger & Arshad 2001). Die Nanopartikel können sich je nach Bakterienstamm entweder innerhalb oder außerhalb der Zellwand anlagern (Oremland et al. 2004). In der vorliegenden Arbeit wird das Bakterium Azospirillum brasilense auf dessen Fähigkeit untersucht, die toxischen Selenoxyanionen Selenat und Selenit zum weniger toxischen elementaren Selen zu reduzieren (Abbildung 1). In der Publikation von Tugarova et al. (2013) wird beschrieben, dass das Bakterium A. brasilense unter aeroben Bedingungen Selenit zu elementaren Selen unter Bildung von Nanopartikeln reduzieren kann. Aufbauend auf diesen Aspekt wird in dieser Arbeit untersucht, inwiefern eine Reduktion von Selenat durch das Bakterium A. brasilense stattfindet. Des Weiteren wird die Selentoleranz der Bakterien analysiert, indem unterschiedliche Konzentrationen an Selenit eingesetzt werden. Durch eine Vielzahl angewendeter Methodik werden die von den Zellen isolierten Nanopartikel charakterisiert. Lösliches Se (toxisch) SeO42- Unlösliches Se (weniger toxisch) Se0 SeO32- (Selenat) (Selenit) Mikrobielle Reduktion durch A.brasliense (Elementares Selen) Mikrobielle Reduktion durch A.brasliense Charakterisierung der Nanopartikel: • Zetapotential • Dynamische Lichtstreuung • Thermostabilität • UV/Vis-Spektroskopie • Totale Kohlenstoffgehalt Abbildung 1: Reduktion von Selenat zu Selenit und elementaren Selen Theoretische Grundlagen 2 Seite |3 THEORETISCHE GRUNDLAGEN 2.1 DAS BAKTERIUM AZOSPIRILLUM BRASILENSE Die Bakterien der Gattung Azospirillum (Abbildung 2 und Abbildung 3) gehören zu der phylogenetischen Gruppe der α-Proteobacteria, sind gram-negativ und zählen zu den Plant growth-promoting rhizobacteria (PGPR) (Kloepper et al. 1991), da die Bakterien die Fähigkeit besitzen das Wachstum und den Ertrag von Pflanzen zu fördern. Dies geschieht durch die Ausscheidung von Phytohormonen (z.B. Indolessigsäure) durch das Bakterium und bewirkt bei Pflanzen eine morphologische Veränderung der Wurzeloberfläche, in dem mehr Wurzelhaare gebildet werden. Es kommt zu einer Vergrößerung der Oberfläche, welche zu einer besseren Aufnahme von Nährstoffen und Wasser aus dem Boden führt und schließlich ein erhöhtes Streckwachstum verursacht (Bloemberg & Lugtenberg 2001). In Folge dessen kann es zu einer Steigerung der Ertragsausbeute von 5 bis 30 % kommen. Aufgrund dieser Ertragssteigerung setzt der Mensch in der Landwirtschaft Azospirillen als Bestandteil in ökologischen Düngern ein (Okon & Labanderagonzalez 1994). Abbildung 2: Bakterium A. brasilense auf festem Nährmedium Abbildung 3: Mikroskopische Aufnahme von A. brasilense Bakterien der Gattung Azospirillum, insbesondere A. brasilense, gehören zu der am meisten studierten Rhizobakterien und sind dafür bekannt Symbiose mit höheren Pflanzen einzugehen (Bashan & de-Bashan 2010). A. brasilense gilt als fakultativ endophytisches Bakterium, da es auch außerhalb der Wurzel leben kann. Einige Bakterienstämme dieser Gattung konnten von Pflanzenwurzeln aus gemäßigten bis subtropischen Gegenden isoliert werden (Rothballer et al. 2003). Theoretische Grundlagen Seite |4 Dabei ist besonders A. brasilense von großer Bedeutung, da das Bakterium vermehrt an den Wurzeln von Nutzpflanzen, wie Mais (Gafny et al. 1986), Weizen (Bashan et al. 1986), Reis (Murty & Ladha 1987), Tomaten, Pfeffer, Baumwolle und Sojabohnen (Bashan et al. 1991, Levanony & Bashan 1991), vorkommt. Eine weitere wichtige Charakteristik der Rhizobakterien ist es, dass sie die Fähigkeit besitzen atmosphärischen Stickstoff zu binden. Dies spielt jedoch eine untergeordnete Rolle bei Azospirillum brasilense (Patriquin et al. 1983). 2.2 SELEN 2.2.1 Allgemeines Selen wurde als neues Element im Jahre 1817 von Jöns Jacob Berzelius entdeckt. Bei dem Versuch Schwefel aus einem schwefelhaltigen Felsen zu isolieren, entstand eine ungewöhnliche Rotfärbung. Berzelius benannte den Stoff „Selenium“, abgeleitet von der griechischen Mondgöttin Selene, in Anlehnung an das schon früher entdeckte Tellur („Erde“) (Berzelius 1817, Berzelius 1818, Lide 1994). Selen ist ein Nichtmetall mit der Ordnungszahl 34 und einer relativen Atommasse von 78,96 u. Selen ist chemisch verwandt mit anderen Stoffen der 16. Gruppe. Diese Gruppe der Chalkogene enthält neben Selen auch Sauerstoff, Schwefel, Tellur und Polonium. In der Natur sind vier verschiedene Oxidationsstufen vorzufinden: (-II), (0), (IV) und (VI). Dabei liegen Selenit (SO32-) und Selenat (SO42-) als lösliche Oxyanionen (AxOyz−) vor. Die Oxyanionen zeigen eine erhöhte Löslichkeit und Mobilität mit steigendem pH-Wert, im Gegensatz zu Metallen, die ein entgegengesetztes Verhalten zeigen (Chapman et al. 2010). Neben diesen anorganischen Formen des Selens gibt es auch nicht-flüchtige Selenide (Se (–II)), wie zum Beispiel Selenocystein (21. Aminosäure), die am meisten vorkommende Selenspezies im mensch-lichen Körper (Fernández-Martínez 2009). Das Auftreten der verschiedenen Spezies des Selens leitet sich von den physikalischchemischen Eigenschaften ab. Die Oxyanionen Selenit und Selenat kommen vorwiegend in wässrigen Lösungen oder in belüfteten basischen Böden vor. Im Gegensatz dazu finden sich unlöslichen Selenide und das elementare Selen meist in minder belüfteten sauren Böden vor, welche reich an organischem Material sind. Theoretische Grundlagen Seite |5 Selen besitzt sowohl einen toxischen als auch einen nützlichen Charakter. Dabei ist die Konzentrationsspanne zwischen Selenmangel (< 40 µg/Tag) und einer Selenvergiftung (> 400 µg/Tag) sehr gering (Levander & Burk 2006). Ein Selendefizit steht in Verbindung mit Leber,- Muskel- und Herzerkrankungen. Somit stellt das Spurenelement Selen in der Ernährung einen essentiellen Nährstoff, aber auch ein Gift dar (Schwarz & Foltz 1957). 2.2.2 Vorkommen Sowohl in Steinen, verschiedenen Böden, als auch in Wasser und Luft kommt Selen vor und gilt somit als weitverbreitete Komponente. Der Umweltqualitätsstandard für die Wasserqualität besagt, dass ≤ 0,01 mg/l Selen enthalten sein dürfen. Im Gegensatz dazu befindet sich in Industrieabwässern ein Selengehalt von ≤ 0,1 mg/l (McNeal & Balistrieri 1989, Soda 2009). Weitere anthropogene Quellen des Seleneintrags in die Atmosphäre sind die Kohle- und Erdölverbrennung, aber auch während der Extraktion und Verarbeitung verschiedener Elemente (Kupfer, Zink, Uran, etc.) entsteht Selen (FernándezMartínez 2009). Erhöhte Selenwerte im Boden wurden in den Ländern China (Wang & Gao 2001), Indien (Dhillon & Dhillon 2003), Irland (Seby et al. 1997) und den USA (Presser 1994) festgestellt. Durch die zunehmende Verwendung von Selen in den industriellen und medizinischen Bereichen gelangt es in das Abwassersystem und somit in dem gesamten Wasserkreislauf. Dies stellt einen weiteren Eintragspfad von Selen in die Umwelt dar. Die primäre natürliche Selenquelle stellt die Verwitterung von Gesteinen dar, insbesondere tonhaltige Gesteine, die eine höhere Konzentration an Selen enthalten. Ein weiterer Eintragspfad ergibt sich durch vulkanische Aktivitäten, bei dem Selen in die Atmosphäre freigegeben wird (McNeal & Balistrieri 1989). Im Pflanzenreich kommt Selen besonders in Pflanzen, beispielsweise im Knoblauch vor, bei den es als Se-Methylselenocystein vorliegt. Pflanzen nehmen Selen in Abhängigkeit vom Selengehalt des Standortes des Organismus auf. Wenige Pflanzen, beispielsweise der Gattung Astragalus, besitzen sogar die Fähigkeit Selen zu akkumulieren, weshalb sie auch als „Selen-Hyperakkumulatoren“ bezeichnet werden (Virupaks & Shrift 1965). Theoretische Grundlagen 2.2.3 Seite |6 Verwendung Selen ist von großem Interesse für die Photovoltaikindustrie aufgrund der halbleitenden Eigenschaften bei Raumtemperatur (RT) und der ausgeprägten Photoleitung. Folge dessen kommt Selen als hochreine Chemikalie bei der Entwicklung von Solarzellen zum Einsatz (Kausch & Matschullat 2005, Neukirchen 2009). Aufgrund der unterschiedlichen Farbgebung von Selen wird es in der Glasindustrie zum Färben von Fenstergläsern und zum Entfärben des Grüntons von Flaschenglas verwendet (Kausch & Matschullat 2005). Neue Studien belegen, dass sich Selennanopartikel positiv in der Pharmakotherapie auswirken. Von der Verwendung von sogenannten „Subnanopartikeln“ (0,4 – 1 nm) wurde viel in medizinischen Bereichen berichtet. So rufen diese Anti-Tumor-Effekte hervor (Miyagi et al. 2003), werden als Anti-Leukämie-Mittel (Sieber 2003) oder als Antioxidans (Gao et al. 2002) verwendet. Material und Methoden 3 3.1 1. 1. Seite |7 MATERIAL UND METHODEN CHEMIKALIEN Stoff Hersteller; Lot.-Nr. Agar (Difco™) BD; 2363441 Ammoniumsulfat (NH4)2SO4 Merck; A0080411044 Biotin (C10H16N2O3S) Sigma; 019K1769 Bovine Serum Albumin (BSA) Merck; K2881118645 Calciumchlorid-Dihydrat Merck; A862982745 (CaCl2 · 2 H2O) Eisen(II)-Sulfat-Heptahydrat (FeSO4 · 7 H2O) Merck; TA886265046 D-(+)-Glucose, wasserfrei (C6H12O6) Roth; 290158205 Glutaraldehyd (C5H8O2) Serva; PO90341 Hefeextrakt BD; 0344732 Hydroxylamin-Hydrochlorid (NH4OCl) Merck; S22427741 Kaliumchlorid (KCl) Roth; 071167621 di-Kaliumhydrogenphosphat (K2HPO4) Merck; A880404803 Kalium-dihydrogenphosphat (KH2PO4) Calbiochem; B32851 Kupfersulfat-Heptahydrat (CuSO4 · 5 H2O) Merck; A866453618 Magnesiumsulfat-Heptahydrat (MgSO4 · 7 H2O) Merck; A972786844 Mangan(II)-Sulfat-Monohydrat (MnSO4 · H2O) Merck; 3837841634 Natriumcarbonat, wasserfrei (Na2CO3) Roth; 042178778 Natriumchlorid (NaCl) Roth; 202186415 Natriumhydrogenphosphat (NaHCO3) Roth; 192185982 Natriumhyrdogenphosphat-Dihydrat (NaHCO3 · 2 H2O) Roth; 44837953 Natriumhydroxid (NaOH) Merck; B0160998743 di-Natrium-Molybdat-Dihydrat (Na2MoO4 · 2 H2O) Merck; A854221520 Natriumtartrat-Dihydrat (C₄H₄Na₂O₆ · 2H₂O) Aldrich; 06619TH-259 Trishydroxymethylaminomethan (TRIS) (C4H11NO3) Roth; 172182199 Salzsäure (HCl) Roth; 211867465 Natriumselenit (Na2SeO3) Applichem; 1Y008059 Natriumselenat (Na2SeO4) Sigma; 13555558 Material und Methoden 3.2 Seite |8 BAKTERIENSTAMM Für diese Arbeit wurde der Bakterienstamm Azospirillum brasilense (DSMZ 1843) verwendet. 3.3 KULTURMEDIEN UND LÖSUNGEN Alle unter sterilen Bedingungen stattfindenden Arbeiten wurden an der Sterilbank Heraeus Instruments des Typs „Hera Safe“ durchgeführt. Die für die Kulturmedien oder anderen Lösungen abzuwiegenden Mengen wurden entweder an der Feinwaage „LA 129 S“ der Firma Sartorius oder an der Waage (ab 1 g) der Firma Kern (Typ 510) abgewogen. Für das Einstellen der pH-Werte wurde das pH-Meter der Firma WTW des Typs pH 540, MultiCal verwendet. Mit Hilfe verschiedener Konzentrationen einer verdünnten Natriumhydroxidlösung (0,01 M; 0,1 M; 1 M; 10 M) und einer verdünnten Salzsäure (0,01 M; 0,1 M; 1 M; 10 M) wurde der gewünschte pH-Wert eingestellt. 3.3.1 Azo-Medium zur Kultivierung von Azospirillum brasilense Das Medium für das Bakterium A. brasilense wird nach der Vorschrift der Deutschen Sammlung von Mikroorganismen und Zellkulturen (DSMZ) (DSMZ 2007) hergestellt. Dieses Medium ist speziell auf das verwendete Bakterium abgestimmt und wird in der vorliegenden Arbeit weitergehend als Azo-Medium bezeichnet. Stammlösung 950 ml Spurenelementlösung (10-fach konzentriert) 1 ml Na-Malat-Lösung (20%-ig) 25 ml Glucose-Lösung (20%-ig) 25 ml Biotin 500 µl pH 7,1 Für das Medium wurde ein Volumen von 1 l angesetzt. Dazu wurde zunächst die Stammlösung (Kapitel 3.3.1.1) hergestellt und nach vollständigem Lösen der Substanzen in vollentsalztem Wasser (VE-Wasser) bei 121 °C 30 Minuten autoklaviert (Hiclave™, HV-25). Material und Methoden Seite |9 Nach Abkühlen der Stammlösung wurde die sterile Spurenelementlösung mit dem sterilen Biotin zu dem Medium gegeben. Die Natrium-Malat-Lösung und die Glucose-Lösung wurden sterilfiltriert und zu dem Medium hinzugeführt. 3.3.1.1 Stammlösung für das Azo-Medium Die Stammlösung für das Azo-Medium (Kapitel 3.3.1) setzt sich wie folgt zusammen. CaCl2 · 2 H2O 0,02 g Hefeextrakt 0,05 g K2HPO4 0,25 g MgSO4 · 7 H2O 0,20 g NaCl 0,10 g (NH4)2SO4 1,00 g VE-H2O 950 ml 3.3.1.2 Spurenelementlösung (10-fach) Die Spurenelementlösung (10-fach) für das Azo-Medium wurde aus folgenden Substanzen zusammengesetzt: FeSO · 7 H2O 0,1 g MnSO · H2O 0,02 g Na2MoO4 · 2 H2O 0,01 g VE-H2O ad 10 ml 4 4 3.3.2 Reagenzien für die Proteinbestimmung Für die Proteinbestimmung wurde die Lösung C erst kurz vor der Anwendung frisch hergestellt. Diese Lösung wurde in einem Verhältnis von 50:1 aus den Lösungen A und Lösung B angesetzt. Material und Methoden S e i t e | 10 3.3.2.1 Lösung A BCA 1g Na2CO3 1,49 g C4H4Na2O6 · 2 H2O 0,19 g NaOH 0,4 g NaHCO3 0,96 g VE-H2O 50 ml pH 11,25 mit 5 N NaOH VE-H2O ad 100 ml CuSO4 · 5 H2O 1g VE-H2O 25 ml 3.3.2.2 Lösung B 3.3.3 Salzlösung zur Isolierung der Nanopartikel NaCl 17,5 g KCl 0,74 g MgSO4 · 7 H2O 12,3 g TRIS 0,15 g VE-H2O ad 1000 ml pH 7,5 Material und Methoden S e i t e | 11 3.4 ZELLAUFSCHLUSSPUFFER/LAGERUNGSPUFFER 3.4.1 Kaliumphosphatpuffer (0,1 M) Kaliumphosphat-Lösung (mono-Kalium-Salz) 16 ml KH2PO4 2,72 g VE-H2O 100 ml Kaliumphosphat-Lösung (di-Kalium-Salz) K2HPO4 3,48 g VE-H2O 100 ml VE-H2O 84 ml 100 ml pH 7,5 3.4.2 PBS-Puffer NaCl 0,8 g KCl 0,02 g NaHCO3 · 2 H2O 0,18 g KH2PO4 0,024 g VE-H2O ad 100 ml pH 7,4 3.4.3 Lysispuffer NaCl 9g NaOH 12 g VE-H2O ad 1000 ml Material und Methoden 3.5 METHODEN 3.5.1 Erhaltung der Stammkultur S e i t e | 12 Die Stammkultur von A. brasilense wurde auf einem festen Nährboden, bestehend aus Azo-Medium (Kapitel 3.3.1) mit 1,5 % Agar, kultiviert. Zur Erhaltung dieser, wurde einmal monatlich 1 bis 2 cm der Kultur mit einer sterilen Impföse aufgenommen und auf eine frische Kulturplatte ausgestrichen. Die Kulturplatte wurde anschließend bei Raumtemperatur gelagert. 3.5.2 Kultivierung von A. brasilense Um eine Vorkultur des Bakteriums A. brasilense herzustellen wurden 1 bis 2 cm der Kultur mit einer sterilen Impföse von dem festen Nährboden aufgenommen und in einem mit 30 ml flüssigen Azo-Medium befüllten Erlenmeyerkolben überführt. Die Inkubation erfolgte über Nacht im Wärmeraum bei 30 °C auf dem Schüttler der Firma Sartorius (Typ Certomat MO II) mit 100 Umdrehungen pro Minute (rpm). Die Kultivierung von A. brasilense erfolgte in Erlenmeyerkolben mit 100 ml Azo-Medium ebenfalls bei 30 °C, 100 rpm unter aeroben Bedingungen. Der Wachstumsverlauf der Zellkultur wurde durch Messung der optischen Dichte (OD) bei einer Wellenlänge von 600 nm mittels Photometer der Firma Pharmacia Biotech (Typ: Ultrospec 1000) verfolgt. Die Berechnung der Wachstumskonstanten (μ) in der exponentiellen Phase des Wachstums erfolgt nach Formel 1 (F. 1) und die der Verdopplungszeit der Zellmasse (td) nach der Formel 2 (F. 2). F. 1 F. 2 Material und Methoden 3.5.3 S e i t e | 13 Chemische Reduktion von Selenit und Selenat Um Selenat bzw. Selenit chemisch zu reduzieren wurden 100 µl einer 100 mM-Selenitund 100 µl einer 100 mM-Selenatlösung in je ein Reaktionsgefäß gegeben und mit je 600 µl VE-Wasser verdünnt. Anschließend wurden je 200 µl Hydroxylamin (100 mM) hinzu pipettiert. Das durch die chemische Reduzierung entstandene elementare Selen wird mit dem mikrobiell reduzierten verglichen. Dabei wird auf Farbe und Form der Partikel und Absorption, welches mit Hilfe der UV/Vis-Spektroskopie (Kapitel 3.5.11) untersucht wird, eingegangen. 3.5.4 Mikrobielle Reduktion von Selenit und Selenat Zur Untersuchung der mikrobiellen Reduktion von Selenat und Selenit durch A. brasilense erfolgte die Kultivierung der Zellen mit Selen, wie in Kapitel 3.5.2 beschrieben. Die Ansätze wurden aus der Vorkultur so angeimpft, bis sich eine OD600nm von 0,25 einstellt. Je nach zu untersuchender Selenat- (1 mM) oder Selenitkonzentration (1 mM bis 5 mM) wurde ein entsprechendes Volumen aus den 100 mM-Stammlösungen in das Kulturmedium überführt. Insgesamt gab es je 2 Ansätze mit Selenat und je 2 mit Selenit. Zusätzlich wurden 3 Kontrollansätze mitgeführt: Kontrolle 1: Medium + Bakterien Kontrolle 2: Medium + Selenit Kontrolle 3: Medium + Selenat Die Probenentnahme (zweimal täglich) erstreckte sich über einen Zeitraum von 7 bis 10 Tagen. Der Wachstumsverlauf während der Kultivierung wurde durch Messung der Optischen Dichte bei 600 nm, sowie der Bestimmung des Gesamtproteingehalts verfolgt. Außerdem wurde der pH-Wert im Medium zu den Probezeitpunkten gemessen. Der Selengehalt wurde mittels ICP-MS (Kapitel 3.5.8) und HG-AAS (Kapitel 3.5.9) ermittelt. Dabei wurden 5 ml der jeweiligen Kultur abgenommen und bei 10.000 x g für 10 min zentrifugiert. Jeweils 2 ml des Überstandes wurden auf zwei Reaktionsgefäße aufgeteilt. Die Probe, die für die Messung mittels ICP-MS bestimmt war, wurde mit 20 µl konzentrierter Salpetersäure versetzt. Die andere Probe wurde weiterverwendet für die HG-AAS. Die Lagerung bis zur Messung erfolgte im Tiefkühlschrank. Material und Methoden 3.5.5 S e i t e | 14 Lichtmikroskopie Für die optische Untersuchung wurde das Lichtmikroskop Olympus (BX 61) mit einer 1000-fachen Vergrößerung verwendet. Hierbei wurden die Bakterien mittels Phasenkontrastaufnahme untersucht, während die Nanopartikel im Hellfeld sichtbar gemacht wurden. 3.5.6 Zellaufschluss mittels basischer Verseifung Eine weitere Kontrolle des Zellwachstums ist die Ermittlung der Gesamtzellproteinkonzentration. Dafür müssen die Zellen der entnommen Proben zunächst mittels basischer Verseifung aufgeschlossen werden. Dazu wurden die Zellen durch Zentrifugation 10 min bei 10.000 x g (Centrifuge 5804 R, Eppendorf) vom Medium getrennt. Das Zellpellet wurde mit 500 µl Lysispuffer (Kapitel 3.4.3) resuspendiert (Scientific Industries, Vortex-Genie 2), für 10 min im Heizblock bei 90 °C inkubiert und erneut zentrifugiert. Vom Überstand wurde 50 µl entnommen und im Anschluss die Proteinkonzentration durch das Bicinchoninic Acid (BCA) Protein Assay (Kapitel 3.5.7) bestimmt. 3.5.7 Proteinbestimmung Für die Proteinbestimmung wurde das Prinzip des Bicinchoninic Acid (BCA) Protein Assays verwendet, welches auf der Bildung eines Kupfer2+-Proteinkomplexes unter basischen Bedingungen beruht (Biuret-Reaktion). Nachdem sich der Komplex gebildet hat, kommt es zur Reduktion von Cu2+ zu Cu+. Durch die anschließende Chelatbildung von zwei Molekülen BCA mit einem Cu+-Ion verfärbt sich die Lösung blau-violett (Abbildung 4). Die Konzentration des gebildeten Farbstoffes wurde durch Messung der Absorption bei 562 nm mit dem Plattenreader µQuant der Firma Bio-Tek Instruments, inc. gemessen. Die Höhe der Reduktion des Kupfers ist dabei proportional zu der Menge an Protein (Smith et al. 1985). Material und Methoden S e i t e | 15 Abbildung 4: Farbgebung nach 1 h Inkubation Nachdem jeweils 50 µl des Überstandes der Probe entnommen wurde (Kapitel 3.5.6), konnten 950 µl der Reagenz C (Kapitel 3.3.2) hinzugegeben werden. Als Blank- bzw. Referenzlösung wurden bei jeder Messung Lysispuffer und Rinderserumalbumin (1 mg/ml) in einem Konzentrationsbereich von 100 bis 800 µg/ml mitgeführt. Die Proben wurden für eine Stunde bei RT inkubiert und für die Messung (Doppelbestimmung) der Absorption im Plattenreader jeweils 200 µl in ein Well der Mikrotiterplatte (Greiner, bio-one-PS Microplate) überführt. Anhand der Eichgerade (Abbildung 5) wurde die Proteinkonzentration ermittelt. 1150 BSA-Konzentration [µg/ml] 950 y = 3884,5x - 912,9 R² = 0,9972 750 550 350 150 -50 0,22 0,27 0,32 Eichgerade 0,37 OD562nm 0,42 0,47 Linear (Eichgerade) Abbildung 5: BSA-Eichkurve zur Proteinbestimmung mit dem BCA-Reagenz Material und Methoden 3.5.8 S e i t e | 16 Massenspektrometrie mit induktiv gekoppeltem Plasma (ICP-MS) Die Massenspektrometrie mit induktiv gekoppeltem Plasma (ICP-MS, inductivelycoupled-plasma massspectrometry) ist ein Verfahren der Spurenelementanalytik, bei dem der Gesamtgehalt an Selen in den Proben ermittelt wird. Die flüssigen Proben werden über ein Zerstäubersystem als feine Tropfen in das induktiv gekoppelte Argonplasma überführt und bei einer Temperatur von ca. 5000 °C ionisiert. Dieses Plasma wird durch einen hochfrequenten Strom in ionisiertem Argon induziert. Die geladenen Molekülfraktionen werden beschleunigt und über ein Linsensystem in das Hochvakuum des Massenspektrometers überführt. Die Ionenoptik fokussiert den Ionenstrahl, der in einem Magnetfeld, dem Quadrupol-Massenspektrometer, eingebracht ist und die Moleküle abhängig von ihrem Masse-Ladungs-Verhältnis auftrennt. Dem entsprechend erfolgt eine Einteilung der Probemoleküle nach Molekülmasse. Die Ionen werden mit Hilfe eines ladungssensitiven Detektors analysiert. Da jedes Element mindestens ein Isotop aufweist und dessen Masse bei keinem natürlichen Isotop eines anderen Elements auftritt, ist die Masse eine charakteristische Eigenschaft der Elemente (Schmidt & Gebel 2000, Less et al. 2008). Die Messung der Proben wurde an dem Institut für Ressourcenökologie mit dem Gerät der Firma Perkin Elmer des Typs SCIEX Elan 9000 durch Frau Aline Ritter durchgeführt. 3.5.9 Hydrid-Generation (Hydridtechnik) Atomabsorptionsspektrometrie (HG-AAS) Mit Hilfe der Hydridtechnik können Metalle analysiert werden, die flüchtige Hydride bilden, wozu Arsen, Bismuth, Antimon, Eisen und auch Selen zählen. Bevor die Probe analysiert werden kann, muss sie in ionogener Form vorliegen. Um dies zu realisieren, wurde die Probe chemisch vorbehandelt durch Verdünnung mit 3 %-iger Salzsäure, welche auch gleichzeitig zum Verdünnen der Kalibrierlösung verwendet wurde. Diese Salzsäure dient als Trägerlösung. Als Reduktionsmittel dient ein Gemisch aus 0,2 %-igem Natriumbortetrahydrid und 0,05 %-iger Natriumhydroxidlösung in Wasser. Zuerst wurde der Selenstandard (10 µg/ml) 1:100 verdünnt und je 10 µl, 50 µl, 100 µl, 150 µl und 200 µl entnommen und jeweils auf 10 ml mit 3 %-iger Salzsäure aufgefüllt. Material und Methoden S e i t e | 17 Anschließend wurden die Proben präpariert, indem sie 1:10.000 mit 3 %-iger Salzsäure verdünnt wurden. Nachdem die Zugabe des Reduktionsmittels erfolgte, reagiert es mit der Säure und setzte Wasserstoff frei. Dieser Wasserstoff reduziert die Metallionen zum gasförmigen Hydrid. Der Inertgasstrom aus Argon spült das Hydrid in die bis ca. 1000 °C aufheizbare Quarzküvette, in der Selen chemisch zersetzt wird. Bei der Hydridtechnik ist darauf zu achten, dass Störsubstanzen und Begleitstoff als gasförmiges Hydrid mit Hilfe von Argon abgetrennt werden. Die Atomabsorptionsspektrometrie wurde am Institut für Ressourcenökologie des HZDRs mit dem „Atomic Absorption Spektrometer 4100“ der Firma Perkin Elmer durch die Mitarbeiterin Aline Ritter durchgeführt. 3.5.10 Isolierung der Nanopartikel Zur Isolierung der Se(0)-Nanopartikel wurde eine modifizierte Methode nach Oremland (Oremland et al. 2004) verwendet. Von den Kulturansätzen, bei denen mikroskopisch Nanopartikel sichtbar waren, wurden jeweils 40 ml entnommen und in ein 50 ml Polypropylen-Röhrchen gegeben. Diese Zellsuspension wurde mittels Ultraschallfinger (Branson, Digital Sonifier W-250 D) bei 50 %-iger Intensität zwei Minuten homogenisiert. Anschließend wurde die Probe 30 Minuten (Eppendorf, Centrifuge 5804 R) bei 1500 x g zentrifugiert. Nach dem Zentrifugieren folgten drei Waschschritte, bei denen der Überstand verworfen wurde und das Pellet jeweils mit 0,5 M Natriumchloridlösung, 0,5 M Saccharoselösung und einer Salzlösung (Kapitel 3.3.3) gewaschen und anschließend nach jedem Schritt erneut mit dem Ultraschallfinger behandelt und zentrifugiert wurde. Nach der Zentrifugation wurden 35 ml, der mit Eiweißlysozym versetzten Salzlösung, hinzugegeben und bei Raumtemperatur 19 h inkubiert. Am folgenden Tag wurden die lysierten Zellen erneut mit dem Ultraschallfinger behandelt und bei 9000 x g für 15 min zentrifugiert. Die Trennung der Selennanopartikel von den Zellen bzw. Zellbestandteilen erfolgte durch weitere 4 Waschschritte (Salzlösung (Kapitel 3.3.3), 0,25 M NaOH, 0,1 M NaOH und 10 mM Na2HPO4) bei denen der Überstand verworfen wurde und das Pellet jeweils mit der entsprechenden Lösung resuspendiert wurde. Material und Methoden S e i t e | 18 Nach jedem dieser Waschschritte erfolgte eine Behandlung mit dem Ultraschallfinger, eine Inkubation von 1,5 h und die Zenrifugation. Die gereinigten Nanopartikel wurden in VEWasser resuspendiert. 3.5.11 UV/Vis-Spektroskopie Die UV/Vis-Spektroskopie beruht auf dem Prinzip, dass Elektronen in einem Molekül durch elektromagnetische Strahlung auf höhere, im Grundzustand des Moleküls, nicht besetzte Energieniveaus angehoben werden. Dabei wird die elektromagnetische Strahlung absorbiert und im Wellenbereich des ultravioletten Lichts (UV-Bereich, 400 nm) oder im Bereich des sichtbaren Lichts (Vis-Bereich, =170 bis =400 nm bis 800 nm) gemessen (Hädener & Kaufmann 2006, Less et al. 2008). Bei der UV/Vis-Spektroskopie wird die Absorption der UV- oder der Vis-Strahlung durch einem in Lösung vorliegenden Analyten bestimmt. Dabei wird die Probe mit polychromatischen Licht beleuchtet, welches transmittiert und schließlich spektral aufgespalten wird. Die Absorption berechnet sich dann aus der Differenz zwischen den Werten der Referenz (in diesem Fall VE-Wasser) und der Probe. Die UV/Vis–Spektren wurden mit Hilfe des Spektrometers des Typs Lambda 750 der Firma Perkin Elmer unter Verwendung von Quarzküvetten mit Schichtdicke 1 cm aufgenommen. 3.5.12 Rasterelektronenmikroskopie in Kombination mit der energiedispersiven Röntgenspektroskopie (REM-EDX) Mit Hilfe der Rasterelektronenmikroskopie kann eine plastische dreidimensionale Darstellung von Oberflächen sichtbar gemacht werden. Das Prinzip dieser Art der Mikroskopie beruht darauf, dass ein Elektronenstrahl auf die zu untersuchende Probe fokussiert wird, der durch Wechselwirkung mit der Probe sogenannte sekundäre Elektronen an der Oberfläche des Präparates erzeugt. Dabei geben Löcher und Spalten weniger sekundäre Elektronen ab und erscheinen dunkel. Runde Erhebungen hingegen geben mehr sekundäre Elektronen ab und wirken dem entsprechend heller (Raven et al. 2000). Material und Methoden S e i t e | 19 Von den Zellkulturen in denen lichtmikroskopisch Nanopartikel sichtbar waren, wurden zweimal je 1 ml entnommen und bei 10.000 x g für 5 min zentrifugiert. Der Überstand wurde verworfen und mit dem Pellet wurde weitergearbeitet. Die Probenpräparation erfolgte nach zwei unterschiedlichen Methoden. Dabei wurde zum Einen mit Glutaraldehyd gearbeitet, wo durch Vernetzung der Proteine Zellstrukturen fixiert wurden. Somit lassen sich die Zellen besser abbilden. Zum Anderen verwendete man eine Methode ohne Glutaraldehyd, da hierbei ein „Verkleben“ der Zellen vermieden wird und somit eine gute Darstellung der Nanopartikel erhält. Bei der ersten Methode wurde das Pellet zweimal mit PBS (Phosphatgepufferte Salzlösung) gewaschen, zentrifugiert und der Überstand verworfen. Danach folgte die Fixierung in 1,5 ml 2 % Glutaraldehyd in PBS für 1 h. Nach dem Zentrifugieren, wird es erneut zweimal mit Reinstwasser gewaschen, zentrifugiert, der Überstand verworfen und in 1,5 ml Reinstwasser resuspendiert und im Ultraschallbad für 3 min behandelt. Bei der zweiten Methode wurde auf das Waschen mit PBS und die Fixierung mit Glutaraldehyd verzichtet. Die Waschschritte mit Reinstwasser wurden in gleicher Weise durchgeführt wie bei der 1. Methode. Zum Aufbringen der Proben wurden Siliziumplättchen (0,5 x 0,5 cm) mit Hilfe von Leitsilber, dem Methylisobutylketon, auf einem Stiftprobenteller fixiert. Danach wurden 10 µl der jeweiligen Probe auf das Plättchen gegeben und über Nacht bei Raumtemperatur getrocknet. Zusätzlich zum REM wurde eine energiedispersive Röntgenspektroskopiemessung (EDX) durchgeführt, welche eine Identifizierung der Elementzusammensetzung erlaubt. Dabei wurde die Probe mit Elektronen beschossen, wodurch die Probenatome angeregt werden. Die Elektronen der inneren Schalen werden auf ein höheres Energieniveau angehoben. Die bei dem Zurückfallen der Elektronen in den Grundzustand entstandene Energiedifferenz tritt in Form von Röntgenstrahlung auf. Diese ist charakteristisch für Elemente. Die REM-EDX-Untersuchungen wurden vom Institut für Ionenstrahlphysik des HZDRs mit dem Mikroskop „REM Zeiss EVO 50“ durch die Mitarbeiterin Elfi Christalle durchgeführt. Material und Methoden S e i t e | 20 3.5.13 Transmissionselektronenmikroskop in Kombination mit der energiedispersiven Röntgenspektroskopie (TEM-EDX) Das Prinzip des Transmissionselektronenmikroskops beruht auf einem ähnlichen Prinzip wie des Lichtmikroskops, jedoch wird statt einer Lichtquelle eine Strahlungsquelle verwendet, die das Objekt mit schnellen Elektronenstrahlen durchstrahlt. Aus der Strahlungsquelle, einer beheizten Feldemissionskathode, treten Elektronen aus, die durch Anlegen einer Beschleunigungsspannung ihre kinetische Energie erhalten. Während ein Teil der Elektronen ohne Beeinflussung durch die Probe hindurch tritt, kommt ein anderer Teil in Wechselwirkung mit dem Kern der Probenatome. Die Elektronenstrahlen, die das Objekt passieren, hinterlassen auf einem darunter liegenden Fluoreszenzschirm Kontrastspuren. Dabei erscheinen „elektronentransparente Gebiete“ hell, „elektronendichte Gebiete entgegen dunkel. Bei biologischen Proben liegt die Auflösung bei 2 nm und ist damit 100 mal besser als bei einem Lichtmikroskop (Raven et al. 2000) Zur Probenpräparation wurden 5 µl einer Nanopartikelsuspension (resuspendiert in VE-Wasser) auf ein Kupfernetz pipettiert, welches mit einem Formvar/Kohle-Film beschichtet war. Zusätzlich zur Bildgebung wurde die Elementzusammensetzung mittels EDX bestimmt (Kapitel 3.5.12). Die TEM-EDX-Untersuchung wurde durch Herrn Dr. René Hübner (Institut für Ionenstrahlphysik des HZDRs) an dem Gerät der Firma FEI des Typs „Titan 80-300“ durchgeführt. 3.5.14 Bestimmung des Zetapotentials Das Zetapotential gibt Informationen über die physikalische Stabilität von Dispersionen und bestimmt die Wechselwirkung von dispergierten Partikeln mit Substanzen aus dem Dispersionsmedium. Jedes Teilchen ist von einer flüssigen Schicht umgeben, die aus zwei Phasen besteht. Zum Einen eine innere Region, die Sternschicht und zum Anderen eine äußere, diffuse Schicht. Bei der Sternschicht sind die Ionen stark gebunden, wo hingegen die Ionen bei der diffusen Schicht weniger fest gebunden sind. Aufgrund der elektrophoretischen Bewegung der Material und Methoden S e i t e | 21 Teilchen und den dabei entstehenden Reibungskräften kommt es zum Abstreifen der diffusen Schicht. Dadurch kommt es zur Entfernung der Gegenionen, womit das Teilchen seine Neutralität verliert und nun in einem geladenen Zustand vorliegt und sich zur entgegengesetzt geladenen Elektrode bewegt. Durch Messen der Partikelgeschwindigkeit kann die Nettoladung des Partikels und die Abhängigkeit seines Oberflächenpotenzials zur umgebenden Phase berechnet werden. Diese Nettoladung wird als Zetapotential gemessen. Wenn alle Teilchen in der Suspension ein großes negatives oder positives Zetapotential besitzen, dann neigen sie dazu, sich gegenseitig abzustoßen, in Folge dessen kommt es nicht zum Ausflocken. Das Zetapotential ist abhängig vom pH-Wert. Daher stellt der pH-Wert einen wichtigen Faktor bei der Bestimmung des Zetapotentials dar. Je basischer die Suspension, desto eher neigt das Partikel dazu eine negative Ladung anzunehmen. Durch Zugabe der Säure wurde erreicht, dass ein Punkt erreicht wird, an dem die negative Ladung neutralisiert ist. Die Konzentrationsverhältnisse, bei denen das Zetapotential null ist, bezeichnet man als isoelektrischen Punkt. Um diese Abhängigkeit darzustellen wurde die Nanopartikelsuspension 1:100 mit einer 0,1 M NaCl-Lösung verdünnt. Anschließend erfolgte die Einstellung der pH-Werte in dem Bereich 1 bis 7. Um eine statistische Sicherheit zu erzielen, wurden 10 Messungen pro pH-Wert gemessen. Daraus wurde ein Mittelwert mit der jeweiligen Standardabweichung berechnet. Beim Auftragen des Zetapotentials gegen den pH-Wert entsteht eine Kurve, vergleichbar mit Abbildung 6. Die Zetapotential-Messungen wurden am Zetasizer der Firma Malvern Instruments (Typ: Nano-ZS) mit der Kapillarzelle „TDS 1060“ ebenfalls von der Firma Malvern Instruments gemessen. Material und Methoden S e i t e | 22 60 Zetapotential (mV) 40 20 0 -20 -40 IEP -60 2 4 6 8 10 12 pH Abbildung 6: Typische Kurve nach Auftragen des Zetapotentials gegen den pH-Wert (Malvern Instruments 2007) 3.5.15 Dynamische Lichtstreuung (DLS) Um die Partikelgröße zu identifizieren bzw. eine Partikelgrößenverteilung aufzustellen wurde die Methode der dynamischen Lichtstreuung (DLS) verwendet, welche auch häufig als Photonenkorrelationsspektroskopie (Photon Correlation Spectroscopy, PCS) bezeichnet wird. Bei der dynamischen Lichtstreuung wird das Streulicht eines Lasers an einer suspendierten Probe analysiert. Durch die Bewegung der Teilchen im Lichtstrahl kommt es bei der Intensität des Lichtes zu Fluktuationen. Mit Hilfe dieser Fluktuation kann der Diffusionskoeffizient der Teilchen, welcher Aufschluss über den hydrodynamischen Radius bzw. den Stokes-Teilchendurchmesser gibt. Das heißt, es wird der Durchmesser einer hypothetischen Kugel gemessen, die mit der gleichen Geschwindigkeit wie das zu analysierende Partikel diffundiert. Je größer ein Teilchen ist, desto langsamer bewegt es sich und desto kleiner ist sein Diffusionskoeffizient. Das gemessene Signal bei der Fluktuation zeigt ein charakteristisches Rauschverhalten, aus dessen zeitlichen Verlauf die Bewegungsgeschwindigkeit des Teilchens ermittelt werden kann. Dies wird in einer Autokorrelationsfunktion der Intensität dargestellt. In einem gewissen Zeitintervall werden Bilder von dieser Bewegung aufgenommen, womit die Geschwindigkeit des Partikels und die damit verbundene Größe berechnet werden konnte (Pingoud & Urbank 1997). Material und Methoden S e i t e | 23 3.5.16 Raman-Spektroskopie der Nanopartikel Die Raman-Spektroskopie ist eine Methode um die Molekülschwingungen und -rotationen eines Stoffes zu analysieren. Durch Bestrahlen der Flüssigkeit mit monochromatischem Licht (z.B. Argon-Laser) mit einer Wellenlänge von 488 nm kommt es zu einer Durchstrahlung der Probe. Nur ein geringer Teil des Lichtes wird ungerichtet gestreut. Ein noch geringerer Anteil des eingestrahlten Lichtes tritt ebenfalls in alle Richtungen aus, besitzt jedoch eine Frequenzverteilung. Diese entsteht durch Absorption und Reemission, welche mit Schwingungsanregung oder -löschung verbunden sind. Mit Hilfe eines photoelektronischen Detektors kann die spektral zerlegte Streustrahlung gemessen werden. Als Folge der Wechselwirkung zwischen Materie und elektromagnetischer Strahlung folgt somit der Raman-Effekt. Das Raman-Spektrum ist ein Emissions-Spektrum (Hesse et al. 2005). Bei der Raman-Spektroskopie wird in einem Wellenzahlbereich von 0 cm-1 bis 8000 cm-1 gemessen. Nachdem 3 µl der Nanopartikelsuspension auf den Quarzobjektträger aufgetragen wurde, konnte das Raman-Spektrum mit Hilfe der Mitarbeiterin Sophia Kostudis des HelmholtzInstituts Freiberg für Ressourcentechnologie des HZDR mit dem Gerät RFS100/S der Firma Bruker aufgenommen werden. 3.5.17 Thermostabilitätstest Bei diesem Testverfahren wurde die Thermostabilität der isolierten Nanopartikel untersucht. Um vergleichende Aussagen treffen zu können, wurde vor und nach dem Erhitzen ein UV/Vis-Spektrum (Kapitel 3.5.11) aufgenommen und die Partikelgrößenverteilung mittels dynamischer Lichtstreuung (Kapitel 3.5.15) untersucht. Des Weiteren wurde der TOC-Gehalt der Nanopartikel bestimmt (Kapitel 3.5.18). Für dieses Testverfahren wurde ein Edelstahlautoklav verwendet, in dem ein Teflonbecher eingesetzt ist (Abbildung 7). Nach Zugabe von 5 ml Nanopartikelsuspension (Nanopartikel in VE-Wasser) wurde der Autoklav mittels Schraubdeckel fest verschlossen, so dass keine Flüssigkeit verloren gehen kann. Anschließend wurde der Stahlautoklav bei 50 °C, 100 °C und 250 °C für 24 h im Muffelofen erhitzt und die Proben nach dem Abkühlen erneut mit den oben genannten Methoden untersucht. Die Temperierung für 24 h bei 35 °C wurde mittels eines temperierten Wasserbades realisiert. Material und Methoden S e i t e | 24 Abbildung 7: Stahlautoklav mit eingesetzten Teflonbecher 3.5.18 Bestimmung des Totalen organischen Kohlenstoffgehalts (TOC) Zur Bestimmung des Kohlenstoffgehalts (TC-Total Carbon) werden sogenannte Summenparameter verwendet. Darunter zählen der gesamte anorganische Kohlenstoffgehalt (TIC - Total Inorganic Carbon) und der gesamte organische Kohlenstoffgehalt (TOC - Total Organic Carbon). Dabei wurde beim TOC zwischen dem Anteil an gelösten organischen Kohlenstoff (DOC -Dissolved Organic Carbon), dem partikulären organischen Kohlenstoff (POC - Particulate Organic Carbon) und dem flüchtigen organisch gebundenen Kohlenstoff (VOC - Volatile Organic Carbon) unterschieden (Abbildung 8). Der DOC-, POC- und VOC-Wert spielen jedoch eine untergeordnete Rolle, da nur der gesamte organische Kohlenstoff betrachtet wird. Die Bestimmung des TOC-Gehaltes, der in VE-Wasser suspendierten Nanopartikel, soll Aufschluss über das Vorhandensein organischer Substanzen liefern. Material und Methoden S e i t e | 25 TC (Gesamtkohlenstoff) TIC (Gesamter anorganischer Kohlenstoff) TOC (Gesamter organischer Kohlenstoff) DOC (Gelöster organischer Kohlenstoff) POC (Partikulärer organischer Kohlenstoff) VOC (Organisch gebundener Kohlenstoff) Abbildung 8: Bestimmungsformen des Kohlenstoffs Der TOC-Gehalt wird mit Hilfe des Differenzverfahrens bestimmt. Dabei wird zunächst die Gesamtheit aller Kohlenstoffverbindungen (TC) gemessen und anschließend den Anteil an anorganischer Kohlenstoffverbindungen (TIC) ermittelt. Den TOC-Gehalt erhält man durch Subtraktion des TICs vom TC. Die TOC-Untersuchungen wurden vom Institut für Ressourcenökologie mit dem „multi N/C 2100 S“ der Firma Analytik Jena durchgeführt. durch die Mitarbeiterin Carola Eckardt Ergebnisse 4 4.1 S e i t e | 26 ERGEBNISSE WACHSTUMSKURVE VON A. BRASILENSE Durch Aufnehmen der Wachstumskurve des Bakteriums A. brasilense wird das Wachstum und die Vermehrung der Zellkultur beobachtet. Dabei wird das Wachstumsverhalten unter normalen Wachstumsbedingungen charakterisiert. Somit kann später, bei Einwirkung von Stressfaktoren, ein Vergleich zum Wachstumsverhalten gezogen werden. Da die Anzahl der Bakterien durch Zellteilung ansteigt, kommt es zu einer Trübung der Kultur, welche mittels Messung der Optischen Dichte bei 600 nm erfasst und quantitativ bestimmt werden kann. In Abbildung 9 ist eine repräsentative Wachstumskurve unter aeroben Bedingungen dargestellt. 10 0 5 10 15 Stationäre Phase 0 Übergangsphase 0 Exponentielle Phase 1 lag-Phase Logarithmischer OD600nm-Wert Bestimmung von µ und td 20 25 Zeit (h) 30 35 40 45 50 Bakterienkultur Abbildung 9: Logarithmische Darstellung der Wachstumskurve von A. brasilense Am Anfang der Kultivierung ist eine geringe lag-Phase bis 2 h erkennbar. Diese Anlaufphase wird von den Bakterien benötigt, um ihren Stoffwechsel an die veränderten Umgebungsbedingungen anzupassen. In dem Zeitraum 2 bis 10 h ist ein exponentielles Wachstum der Zellen zu erkennen, wobei hier eine Teilung der Bakterienzellen in gleichen Zeitabständen stattfindet. Nach der Ergebnisse S e i t e | 27 exponentiellen Phase folgt eine Übergangsphase, bevor die stationäre Phase bei 23,5 h beginnt. In der stationären Phase bleibt der Biomassegehalt konstant. Anhand der Werte der exponentiellen Phase der Wachstumskurve kann die Wachstumsrate µ und die Verdopplungszeit td berechnet werden. Die Ergebnisse sind in Tabelle 1 dargestellt. Bis zu einem Zeitpunkt von ca. 30 h ist ein stetiges Wachstum zu erkennen und erreicht hier das Maximum mit einer Optischen Dichte von etwa 3,4. Tabelle 1: OD-Werte und die jeweiligen Zeiten am Anfang und Ende der exponentiellen Phase, Wachstumsrate und Verdopplungszeit Parameter Bakterienkultur x0 0,110 xt 0,810 t 9,67 h t0 2,00 h µ 0,26 h-1 td 2,67 h Im folgenden Kapitel wird der Einfluss der Selenoxyanionen auf das Bakterium A. brasilense untersucht. Dazu wurde eine Anzahl von Methoden verwendet, um das Wachstumsverhalten in Gegenwart von Selenit und Selenat zu beschreiben und die Veränderungen der Selenkonzentrationen im Medium zu verdeutlichen. Mit Hilfe des Lichtmikroskops wurden Bakterien und Nanopartikel sichtbar gemacht. Zur besseren Charakterisierung, der durch mikrobielle Reduktion von Selenit gebildeten Nanopartikel, wurden diese, wie in Kapitel 3.5.10 beschrieben, von den Bakterienzellen isoliert. Bei den Nanopartikeln wurden die Partikelgrößenverteilung, die Oberflächenladung und die Thermostabilität untersucht. Dazu wurde die Oberflächenladung durch das Zetapotential bestimmt, die Größenverteilung durch die dynamische Lichtstreuung gemessen, und die Thermostabilität der Nanopartikelsuspension nach Erhitzen mittels UV/Vis und TOC-Messung näher beschrieben. Ergebnisse 4.2 S e i t e | 28 REDUKTION VON SELENAT UND SELENIT DURCH DAS BAKTERIUM A. BRASILENSE Bei diesem Versuch wurde das Bakterium A. brasilense auf dessen Fähigkeit Selenit und Selenat zu elementarem Selen zu reduzieren, untersucht. Um den Einfluss der toxischen Selenoxyanionen auf das Wachstum von A. brasilense verfolgen zu können, wurden die optische Dichte bei 600 nm, die Gesamtzellproteinkonzentration sowie der pH-Wert gemessen. Um den zeitlichen Verlauf der Selenit- und Selenatkonzentration und den daraus resultierenden produzierten Anteil an elementarem Selen zu ermitteln, wurden ICP-MS- und HG-AAS-Messungen durchgeführt. Weitere Untersuchungen fanden mittels Lichtmikroskopie statt. Während der Kultivierung gab es farbliche Auffälligkeiten zwischen den Selenit-, Selenatund Kontrollansätzen, dessen farbliche Verläufe in Abbildung 10, Abbildung 11 und Abbildung 12 zusammenfassend dargestellt sind. Kontrolle Selenat Selenit 1.Tag 6.Tag 7.Tag Abbildung 10: Farbverlauf der Kontrolle Abbildung 11: Farbverlauf der Selenatkultur Abbildung 12: Farbverlauf der Selenitkultur Zusätzlich zu der normalen Kontrolle, in der lediglich die Bakterien ohne Zusatz von Selenit oder Selenat enthalten waren, wurden zwei weitere Kontrollen angesetzt, die lediglichSelenat aus Medium und Selenit oder Selenat bestanden. Diese Kulturen blieben während der gesamten Kultivierung klar und farblos. Ergebnisse S e i t e | 29 Am Ende der Kultivierung mit Bakterien wurden diese Unterschiede in der farblichen Entwicklung gut sichtbar (Abbildung 13). Medium + Bakerien Medium + Se(IV) Kontrollen Medium + Se(VI) Selenitansätze Selenatansätze Abbildung 13: Farbliche Entwicklung der Kontroll-, Selenat- und Selenitkultur nach sieben Tagen Kultivierung Es ist zu erkennen, dass die Farbverläufe bei der Kontrollkultur und bei dem SelenatAnsatz identisch sind. Nach Zugabe des Selenats bleibt die Selenatkultur weiß und trüb. Erst am 2. Tag beginnen beide Kulturen leicht orange-rot zu werden. Am Ende des Versuches ist bei beiden eine kaminrote Färbung wahrzunehmen. Unterschiede in der Farbentwicklung sind bei der Selenitkultur (Abbildung 12) zu beobachten. Schon kurze Zeit (10 min) nach Zugabe des Selenits ist ein Farbumschlag zu orange-rot erkennbar. Am 6. Tag nimmt die Selenitkultur einen intensiven dunklen Orangeton an. Diese Farbe verändert sich bis zum 7. Tag insofern, dass diese in einen orange-roten Farbton übergeht. Die Bakterien wurden mit Hilfe der Lichtmikroskopie auf die Morphologie der Zellen mit und ohne Selen analysiert. Aufgrund der Farbänderung wurden die Veränderungen einer möglichen Bildung von Nanopartikeln während der Kultivierung verfolgt. Beim Betrachten der Kulturen wurden sowohl in Selenit- und Selenatkulturen, als auch in der Kontrollkultur Bakterien gut sichtbar. Dabei wurde kein Unterschied zwischen den Selenit-, Selenat- und Kontrollkulturen bezüglich der Zellform und -größe der Bakterien festgestellt. Bei der Phasenkontrastaufnahme (Abbildung 14, Abbildung 15 und Abbildung 16) konnten die Nanopartikel nur bei den Selenitansätzen festgestellt werden. Bei den Selenitansätzen wurden sphärische Partikel wahrgenommen, die laut Literatur, aufgrund der Färbung der Kultur, Selennanopartikel sein könnten (Oremland et al. 2004). Mittels Hellfeldaufnahme wurden diese Partikel hervorgehoben (Abbildung 17). In den Aufnahmen der Kontrolle (Abbildung 14) und der Selenatkultur (Abbildung 15) kann ausschließlich das Vorhandensein der Bakterien beobachtet werden. Ergebnisse S e i t e | 30 Abbildung 14: Mikroskopische Aufnahme von A. brasilense im Kontrollmedium (ohne Nanopartikel) Abbildung 15: Mikroskopische Aufnahme von A. brasilense mit Nanopartikeln im Selenatansatz (ohne Nanopartikel) Abbildung 16: Mikroskopische Aufnahme von A. brasilense mit Nanopartikeln im Selenitansatz Abbildung 17: Hellfeldaufnahme der Nanopartikel in der Selenitkultur 4.2.1 Wachstumsverlauf in Gegenwart von Selen Die Wachstumskurven von A. brasilense in Gegenwart von Selenat und Selenit sind in Abbildung 18 dargestellt. Dabei wurden jeweils 2 Kulturen mit 1 mM Selenit und mit 1 mM Selenat versetzt und eine Kontrolle mitgeführt. In der Darstellung wird sichtbar, dass die Bakterienkulturen mit Selenat einen identischen Wachstumsverlauf zeigen, wie die Kontrollkultur. Ergebnisse S e i t e | 31 OD 600nm (logarithmisch) 25 1 0 0 1 2 3 4 5 6 7 Zeit (d) Selenit (Ansatz 1) Selenat (Ansatz 1) Selenit (Ansatz 2) Selenat (Ansatz 2) Kontrolle Abbildung 18: Logarithmische Darstellung der OD-Werte der Kontroll-, Selenit- und Selenatkultur Die OD-Werte der Kontrollkultur und der Kulturen mit Selenat steigen stetig bis zum Beenden des Versuchs nach sieben Tagen. Nach Überwinden der Anlaufphase gehen diese in die exponentielle Phase über, in der die Kulturen ähnliche Wachstumsraten besitzen. Beim Betrachten des Verlaufs der OD-Werte der Selenitansätze ist zu erkennen, dass nach einer geringen Wachstumsphase bis 4 h eine lange lag-Phase im Zeitraum von vier Stunden bis drei Tagen folgt. Anschließend beginnt die exponentielle Phase, welche bis zum Versuchsende andauert. Tabelle 2 zeigt die Mittelwerte (MW) des maximalen OD-Wertes zu dem entsprechenden Zeitpunkt. Dabei wird sichtbar, dass alle Ansätze ähnliche Maxima aufweisen. Die Kulturen mit Selen erreichen das Maximum nach 6,5 Tagen mit einer OD von 4,3 bei dem Selenatansatz und 4,7 bei dem Selenitansatz. Die Kontrolle weist 12 h zuvor ein Maximum bei 4,7 auf. Tabelle 2: Zeitpunkt und OD-Wert des Maximums der Kontroll-, Selenat- und Selenitkultur Kontrolle Selenat Selenit Zeit Zeit Ansatz 1 Ansatz 2 (d) (d) 5,85 5,85 6,85 5,85 6,85 MW der Zeit (d) 5,85 6,35 6,35 ODODWert Wert Ansatz 1 Ansatz 2 4,69 3,80 4,72 5,17 4,14 MW ODWert 4,7 4,3 4,7 Ergebnisse S e i t e | 32 Die optische Dichte bei 600 nm erfasst sowohl die Absorption durch Bakterienzellen als auch die gebildeten Nanopartikel. Dies kann zu einer Überschätzung der Zelldichte in den Selenitkulturen führen. Um zusätzliche Informationen über das Zellwachstum zu erlangen, wurde der Gesamtzellproteingehalt bestimmt. In der Abbildung 19 ist der nichtlogarithmische Verlauf der Gesamtzellproteinkonzentration gegenüber der Zeit dargestellt. Beim Vergleich der Selenit- und Selenatansätze wird sichtbar, dass die Bakterienkulturen mit 1 mM Selenat im zeitlichen Verlauf eine ähnliche Proteinkonzentration aufweisen wie die Kontrollkultur. Die Verläufe der Proteinkonzentration zeigen Gemeinsamkeiten mit den OD-Werten in Abbildung 18. Große Unterschiede werden bei den Selenitansätzen sichtbar, die erst nach einer längeren lag-Phase einen Anstieg in der Gesamtzellproteinkonzentration und somit in der Zelldichte zeigen. Die Selenitansätze weisen im Vergleich zu der Selenat- und Kontrollkultur eine höhere Proteinkonzentration auf und dem entsprechend auch eine höhere Zellzahl. 2480 Proteinkonzentration [µg/ml] 1980 1480 980 480 -20 0 20 40 Selenit (Ansatz 1) Selenit (Ansatz 2) 60 80 100 Zeit (h) Selenat (Ansatz 1) Selenat (Ansatz 2) 120 140 160 180 Kontrolle Abbildung 19: Darstellung der Gesamtzellproteinkonzentration der Kontroll-, Selenit- und Selenatkultur Die Proteinkonzentration der Kontrollkultur steigt ab der ersten Messung bis 147 h (sechs Tage) auf eine Konzentration von 868 µg/ml an und nimmt von diesem Zeitpunkt an ab. Ergebnisse S e i t e | 33 Der erste Selenatansatz weist im gesamten Verlauf eine niedrigere Proteinkonzentration als Ansatz 2 auf. Aufgrund der geringen Unterschiede und des gleichen Verlaufes der Proteinkonzentration werden die folgenden Konzentrationen als Durchschnittswert aus beiden Ansätzen angegeben. Der Anfangswert beider Selenatansätze von durchschnittlich 157 µg/ml liegt höher als bei der Kontrollkultur. Von da an nimmt die Konzentration bis 68,5 h einen Wert von durchschnittlich 762 µg/ml an. Die Gesamtzellproteinkonzentrationen der beiden 1 mM-Selenitansätze bleiben bis 74,5 h konstant mit einem Wert von 167 µg/ml. Ab diesem Zeitpunkt weisen die Kulturen eine unterschiedliche Proteinkonzentration auf. Der erste Selenitansatz zeigt eine erhöhte Gesamtzellproteinkonzentration (bei sechs Tagen eine Proteinkonzentration von 2348 µg/ml). Die zweite 1 mM-Selenitkultur beginnt ab 140,5 h zu steigen bis auf eine Proteinkonzentration von 967 µg/ml bis zum Beenden der Versuchsreihe. Weitere Untersuchungen wurden durch Messen des pH-Wertes während des gesamten Versuches durchgeführt. Dabei wurde ein Zusammenhang zwischen dem Zellwachstum und dem pH-Wert sichtbar. Mit steigendem pH-Wert erhöhten sich die OD600nm-Werte und die gemessene Gesamtzellproteinkonzentration. Die Kontroll- und Selenatkultur begannen bei einem pH-Wert von 7,6 ± 0,2, welcher innerhalb von 74,5 h auf 8,6 ± 0,2 anstieg. Bei den Selenitansätzen startete der pH-Wert bei 7,8 und erreichte nach 52 h sein Maximum bei 8,7. Die gemessenen erhöhten pH-Werte korrelieren mit den Werten der optischen Dichte und spiegeln somit ebenfalls den Wachstumsverlauf der Kulturen wieder. 4.2.2 Messung der Selenit- und Selenatkonzentration Der Einfluss des Wachstums von A. brasilense auf den Selenat- und Selenitgehalt in den Zellkulturen wurde mit Hilfe der ICP-MS und HG-AAS bestimmt. Dabei kann mittels der HG-AAS ausschließlich der Selenitgehalt bestimmt werden, weshalb keine Proben der Selenatkultivierung gemessen wurden. Aus Abbildung 20 geht hervor, dass die Konzentration an Selenit im Laufe der Kultivierung von A. brasilense abnimmt. Dabei werden die Ergebnisse der HG-AAS und ICP-MS der Proben der Selenitansätze gegenübergestellt. Ergebnisse S e i t e | 34 90 80 Konzentration [mg/l] 70 60 50 40 30 20 10 0 0 1 2 3 4 5 6 7 Zeit (d) Selenit (1) (HG-AAS) Selenit (1) (ICP-MS) Selenit (2) (HG-AAS) Selenit (2) (ICP-MS) Abbildung 20: Vergleich der Messungen der Selenitkonzentrationen der HG-AAS und der ICP-MS Die HG-AAS-Ergebnisse verdeutlichen, dass am Anfang beim ersten Selenitansatz eine Konzentration von 81,5 mg/l und beim zweiten eine von 80 mg/l im Überstand aufzufinden ist. Dies entspricht in etwa 100 % der theoretisch eingesetzten Selenit-konzentration. Bei beiden Selenitansätzen ist ab dem 3. Tag eine Abnahme der Selenitkonzentration zu verzeichnen, sodass man nach sieben Tagen bei dem ersten Selenitansatz einen Wert 0,3 mg/l und bei dem zweiten Ansatz eine Endkonzentration von 4 mg/l erhält. Dies entspricht einer Abnahme des Selenits im Überstand nahe 100 %. Zur Überprüfung, dass Selenit von Bakterien umgesetzt wurde, wurde eine weitere Kontrolle angesetzt, die ebenfalls eine Woche untersucht wurde. Diese Kontrolle (ohne Zusatz von Bakterien), besteht lediglich aus Selenit und Medium. Während des Messungszeitraumes wurde eine konstante Selenitkonzentration im Medium gemessen. Somit ist eine Oxidation oder Reduktion des Selenits ausschließlich durch Bakterien möglich. Vergleicht man die Ergebnisse der Selenitansätze der HG-AAS mit denen der ICP-MS, so kann festgestellt werden, dass die Werte fast identisch sind. Damit wird bestätigt, dass die in der ICP-MS gemessene Selenkonzentration der theoretischen Selenitkonzentration entspricht. Am Ende des 2. Tages und am Anfang des 3. Tages wurden bei der HG-AAS Werte gemessen, die nicht zu erklären sind. Diese Fehlwerte können nun durch Bestätigung der Selenkonzentration bei der ICP-MS vernachlässigt werden. In Abbildung 21 ist die zeitliche Änderung der Gesamtselenkonzentration des Überstandes der Bakterienkultur dargestellt. Ergebnisse S e i t e | 35 Die berechnete Selenkonzentration im Überstand am Anfang des Versuchsansatzes beträgt 79 mg/l. Die Werte der Selenatansätze liegen über diesem berechneten Wert und haben im Durchschnitt eine Selenkonzentration von 85 mg/l. Diese Konzentration bleibt bei beiden Selenatansätzen über den gesamten Messzeitraum relativ konstant. Lediglich bei den Selenitansätzen sind Unterschiede in Bezug auf die berechnete Selenkonzentration und den Selenatansätzen zu verzeichnen. Im Durchschnitt beginnen beide Ansätze mit einer Konzentration von 77 mg/l, die drei Tage gehalten wird. Ab diesem Zeitpunkt beginnt die Selenkonzentration abzunehmen. Dabei verläuft die Abnahme des Selens aus dem Überstand beim 1. Selenitansatz, schneller als beim 2. Ansatz. Ein extremer Abfall der Selenkonzentration wurde beim 2. Ansatz ab dem 6. Tag gemessen. Am Ende der Messreihe wurde beim 1.Selenitansatz eine Abnahme des Selens von 97,4 % und beim zweiten von 93,9 % verzeichnet. 100 90 Konzentration [mg/l] 80 70 60 50 40 30 20 10 0 0 1 2 3 4 5 6 7 Zeit (d) Selenitansatz (1) Selenatansatz (1) Selenitansatz (2) Selenatansatz (2) Konzentration 1 mM Abbildung 21: Zeitlicher Verlauf der Selenkonzentration im Selenit- und Selenatansatz Aufgrund dessen, dass bei der Messung mittels HG-AAS ausschließlich die Selenitkonzentration und bei der ICP-MS der Gesamtselengehalt bestimmt wurde, kann daraus die Se(0)-Konzentration in Form von Nanopartikeln berechnet werden. Dieser Zusammenhang wird in Abbildung 22 dargestellt. Ergebnisse S e i t e | 36 In der Darstellung wurde der Mittelwert aus beiden Selenat- und der Selenitansätzen der ICP-MS-Messung, aufgrund ihres ähnlichen Verlaufes, verwendet. Nach ungefähr drei Tagen, beginnen die Bakterien das Selenit aus dem Überstand zu reduzieren und die Nanopartikel zu produzieren. Dies stimmt sehr gut mit den lichtmikroskopischen Beobachtung (Kapitel 4.2) überein. Diese Nanopartikel bestehen vermutlich aus elementarem Selen. Nach etwa sieben Tagen entstand ein prozentualer Prozentualer Anteil des Se-Gehalts (%) Anteil von 95,6 % der Se(0)-Nanopartikel durch Reduktion des Selenits aus dem Medium. 120 100 80 60 40 20 0 0 1 -20 MW Selenat (ICP-MS) 2 3 4 5 6 7 Zeit (d) MW Selenit (ICP-MS) Nanopartikel Abbildung 22: Zeitlicher Verlauf der Selenat- und Selenitintensität und den daraus resultierenden Selennanopartikelanteil 4.2.3 Einfluss der Konzentration auf die mikrobielle Reduktion von Selenit und das Wachstum von A. brasilense Wie in Kapitel 4.2 beschrieben, besitzt das Bakterium A. brasilense die Fähigkeit Selenit zu elementaren Se(0) zu reduzieren. Da in bisherigen Versuchen ausschließlich mit 1 mM-Selenitansätzen gearbeitet wurde, wurde in einem weiteren Versuch die Toleranz des Bakteriums gegenüber unterschiedlich hohen Selenitkonzentrationen (1 mM, 2,5 mM und 5 mM) untersucht. In Abbildung 23 sind die Kulturen mit unterschiedlichen Selenitkonzentrationen nach sieben Tagen dargestellt. Das Wachstumsverhalten wurde mittels OD600nm und der Gesamtzellproteinkonzentration (OD562nm) bestimmt. Ergebnisse S e i t e | 37 Mögliche Änderungen der Selenkonzentrationen wurden durch die ICP-MS und HG-AAS untersucht. Kontrolle 1 mM-Selenitsätze 2,5 mM-Selenitansätze 5 mM-Selenitansätze Abbildung 23: Farbentwicklung nach sieben Tagen bei unterschiedlichen Selenitkonzentrationen 4.2.4 Wachstumsverlauf des Bakteriums A. brasilense bei unterschiedlichen Selenitkonzentrationen Das Wachstum der Bakterien in den unterschiedlich hoch konzentrierten Selenitkonzentrationen (1 mM, 2,5 mM und 5 mM) wurde mittels Photometer bei einer Wellenlänge 600 nm verfolgt. Die zeitlich aufgenommenen Wachstumskurven sind in Abbildung 24 dargestellt. Logarithmischer OD600nm-Wert 20 1 0 0 2 Kontrolle 4 1 mM MW Zeit (d) 2,5 mM MW 6 8 5 mM (Ansatz 1) 10 5 mM (Ansatz 2) Abbildung 24: Wachstumskurven der Kontrollkultur und den Ansätzen mit unterschiedlichen Selenitkonzentrationen Die Kontrollkultur (Bakterienkultur ohne Selenit) zeigt das bekannte Wachstumsverhalten. Am 5. Tag wird bei der Kontrolle ein OD600-Maximum von 5 gemessen, welches bis zum Beenden des Versuches konstant bleibt. Ergebnisse S e i t e | 38 Da die beiden Ansätze der 1 mM- und 2,5 mM-Selenitkonzentration einen ähnlichen Verlauf aufwiesen, wurden diese Wachstumskurven gemittelt. Bei den beiden 5 mM-Selenitansätzen gab es große Unterschiede bezüglich des Wachstumsverhaltens, weshalb auf eine Mittelung der Werte verzichtet wird. Dies gilt ebenfalls für die Kurven zur Bestimmung der Gesamtzellproteinkonzentration in Abhängigkeit von der Zeit. Im Gegensatz zu der Kontrollkultur gibt es bei allen Selenitansätzen eine lag-Phase von drei Tagen. Ab dem 3. Tag beginnen die Bakterien, mit Ausnahme des zweiten 5 mM-Selenitansatzes, mit dem Wachstum. Beim Betrachten der Maxima der Kurven (Tabelle 3) wurde ermittelt, dass im Durchschnitt die Bakterien im 1 mM-Ansatz schneller wachsen und zwischen dem 6. und 7. Tag ein Maximum von durchschnittlich 3,1 (OD600nm) erreichen. Mit steigender Selenitkonzentration wird das Maximum zu einem späteren Zeitpunkt mit einem geringeren OD-Wert erreicht. So wurde bei einer Konzentration von 2,5 mM bei 10 Tagen ein durchschnittlicher OD-Wert von 2,9 und bei der 5 mM-Selenitkonzentration ebenfalls bei 10 Tagen eine Optische Dichte von 1,10. Tabelle 3: Zeitpunkt und OD-Wert der Maxima der unterschiedlichen Selenkonzentrationen Selenitkonzentration Kontrolle 1 mM 2,5 mM 5 mM Zeit Ansatz 1 (d) Zeit Ansatz 2 (d) 4,91 6,17 9,97 9,97 6,91 9,97 9,97 MW der Zeit (d) 4,9 6,5 10 10 ODODWert Wert Ansatz Ansatz 1 2 3,30 4,08 2,07 2,70 3,17 2,06 0,14 MW ODWert 3,3 3,1 2,9 1,1 In Abbildung 25 wurden der zeitliche Verlauf der Gesamtzellproteinkonzentration bei unterschiedlich hohen Selenitkonzentrationen gegenübergestellt. Die Maxima der verschiedenen Ansätze und deren Proteinkonzentration wurden in Tabelle 4 angegeben. Ergebnisse S e i t e | 39 2400 Konzentration [µg/ml] 1900 1400 900 400 -100 0 2 Kontrolle 4 1 mM-Selenitansatz Zeit (d) 2,5 mM-Selenitansatz 6 8 10 5 mM Selenit (Ansatz 1) 5 mM Selenit (Ansatz 2) Abbildung 25: Zeitlicher Verlauf der Gesamtzellproteinkonzentration der Kontrollkultur und den Ansätzen mit unterschiedlichen Selenitkonzentrationen Tabelle 4: Zeitpunkt und Proteinkonzentration der Maxima der unterschiedlichen Selenkonzentrationen Zeit Ansatz 1 (d) 4,91 Kontrolle 1 mM 2,5 mM 5 mM Zeit Ansatz 2 (d) 6,91 9,97 9,97 6,91 9,97 9,97 MW der Zeit (d) 4,9 6,9 10 10 c Ansatz 1 [µg/ml] c Ansatz 2 [µg/ml] 1030,4 1775,6 918,0 0 706,3 1082,5 704,4 MW c [µg/ml] 1030 1241 1000 321 Die Kontrollkultur startet mit einer Gesamtzellproteinkonzentration von 31 µg/ml und erhöht sich kontinuierlich bis diese am 5. Tag ein Maximum bei 1030 µg/ml erreicht Danach nimmt die Konzentration bis auf einen Wert von 788 g/ml am 10. Tag ab. Die 1 mM-Selenitkulturen beginnen am 5. Tag mit der Erhöhung der Proteinkonzentration, die bis zum 7. Tag ansteigt. Hierbei wird eine Gesamtzellproteinkonzentration von 1241 µg/ml erreicht. Bei den 2,5 mM-Ansätzen ist die lag-Phase nach ca. 3-4 Tagen beendet. Von da an steigt die Proteinkonzentration bis zum 10. Tag auf einen Wert von 1000 g/ml an. Die Verläufe der Gesamtzellproteinkonzentrationen der 5 mM-Selenitkulturen unterscheiden sich wesentlich im Verlauf. Während der eine 5 mM-Ansatz vom 3. bis zum 7. Tag ansteigt und dort sein Maximum erreicht, ist bei dem anderen 5 mM-Ansatz keine Zunahme der Proteinkonzentration zuerkennen. Ergebnisse S e i t e | 40 Daraus ist zu schlussfolgern, dass keine wachstumsfähigen Bakterien mehr im Medium enthalten sind. Aufgrund dessen wurde für weitere Untersuchungen nur mit dem zweiten 5 mM-Selenitansatz gearbeitet. 4.2.5 Messung der unterschiedlichen Selenitkonzentrationen Um die Selenitkonzentrationen während der gesamten Kultivierung analysieren zu können wurde zum Einen der Selenitgehalt mittels HG-AAS und zum Anderen der Gesamtselengehalt mit Hilfe der ICP-MS gemessen. Wie in Kapitel 4.2.2 gezeigt werden konnte, entspricht der gemessene Gesamtselengehalt dem der mittels HG-AAS ermittelte Selenitgehalt. Daher wird auf die Darstellung der HG-AAS-Ergebnisse verzichtet. Tabelle 5 zeigt die mittels ICP-MS gemessenen Anfangskonzentrationen der unterschiedlichen Selenitkonzentrationen im Vergleich zu den theoretischen Selenitkonzentrationen. Diese stimmen bei allen Selenitkonzentrationen annähernd überein. In Abbildung 26 sind die Messergebnisse der ICP-MS dargestellt. Tabelle 5: Vergleich der berechneten und mittels ICP-MS gemessenen Selenitanfangskonzentrationen Berechnete Konzentration [mg/l] Gemessene Anfangskonzentration [mg/l] 1 mM 79,0 70,8 2,5 mM 197,4 199,5 5 mM 394,8 404,0 450 Selen-Konzentration [mg/l] 400 350 300 250 200 150 100 50 0 0 2 4 -50 1 mM Konzentration 1 mM 6 Zeit (d) 2,5 mM Konzentration 2,5 mM 8 5 mM Konzentration 5 mM Abbildung 26: Zeitlicher Verlauf der Gesamtselenkonzentration der unterschiedlichen Selenitkonzentrationen 10 Ergebnisse S e i t e | 41 Bei dem 1 mM-Selenitansatz wurde am Anfang ein Durchschnittswert von 71 mg/l ermittelt. Diese Konzentration bleibt bei beiden Ansätzen bis zum 3. Tag konstant. Zwischen dem 3. und 5. Tag beginnt die Abnahme der Gesamtselenkonzentration, welche bis zum 7. Tag auf einen durchschnittlichen Wert von 34 mg/l sinkt. Dies entspricht einer Verringerung von 52 %. Von diesem Zeitpunkt an bleibt die Konzentration konstant. Bei den 2,5 mM-Selenitansätzen bleiben die Konzentration bei beiden Ansätzen bis zum 7. Tag konstant. Von da an sinkt die Gesamtselenkonzentration und verringert die Konzentration um durchschnittlich 23,4 %. Bei dem 5 mM-Selenitansatz wurde ein Anfangswert von 404 mg/l gemessen, welcher über den gesamten Messungszeitraum relativ konstant blieb. Die Ergebnisse zeigen, dass mit steigender Selenitkonzentration die Reduzierung des Selenits aus dem Medium verzögert wird und ein vollständiger Umsatz im untersuchten Zeitraum somit nicht beobachtet werden konnte. 4.3 CHARAKTERISIERUNG DER NANOPARTIKEL 4.3.1 Farbgebung Nanopartikelsuspension nach der Isolierung Nach der Isolierung der Nanopartikel entsteht nach Zugabe von VE-Wasser eine kaminrote Suspension (Abbildung 27). Wenn es große Unterschiede im Wachstumsverhalten und dem damit verbundenen Fortschritt der Selenitreduktion gab, wie zum Beispiel beim ersten und zweiten Ansatz der 1 mM-Selenitkultur, spiegelte sich das auch in den isolierten Suspensionen wieder. Die Ergebnisse der HG-AAS sowie der ICP-MS lieferten Aussagen darüber, dass bei dem ersten 1 mM-Selenitansatz eine schnellere Abnahme des Selenits aus dem Überstand und damit eine erhöhte Bildung von Nanopartikel zu beobachten war (Kapitel 4.2.2). Dies wurde auch nach der Isolierung sichtbar. Der zweite 1 mM-Selenitansatz zeigt gegenüber dem 2. Ansatz eine schwach rote Farbe, was auf eine geringe Anzahl an Nanopartikel zurückzuführen ist. Die Farbgebung und die Messwerte bei den 2,5 mM-Selenitansätzen (Abbildung 29) ähnelten sich sehr mit dem ersten 1 mM-Selenitansatz. Ergebnisse S e i t e | 42 Ein ähnlicher Farbton wie bei dem 5 mM-Ansatz (Abbildung 30) ist bei dem 2. Ansatz der 1 mM-Selenitkultur (Abbildung 28) zu sehen, welcher ebenfalls aufgrund der Vorversuche zwar ein Bakterienwachstum und eine Zunahme des Proteingehaltes aufwies, jedoch laut HG-AAS und ICP-MS der Selengehalt nicht signifikant abnahm und nur eine geringe Anzahl an Nanopartikeln produziert wurde. Abbildung 27: Suspension der isolierten Nanopartikel (1 mM-Selenitansatz (Ansatz 1)) Abbildung 28: Suspension der isolierten Nanopartikel (1 mM-Selenitansatz (Ansatz 2)) Abbildung 29: Suspension der isolierten Nanopartikel (2,5 mM-Selenitansatz) Abbildung 30: Suspension der isolierten Nanopartikel (5 mM-Selenitansatz) Zum Vergleich der mikrobiell erzeugten Nanopartikel wurde auch chemisch reduziertes Se(0) hergestellt. Die dafür eingesetzte Selenitlösung zeigt sofort nach Zugabe des Reduktionsmittels Hydroxylamin eine Rotfärbung und nimmt nach circa fünf Minuten eine intensivere Rotfärbung an. Nach circa 20 Minuten fällt ein roter Niederschlag aus, es bilden sich rote Flocken am Boden des Reaktionsgefäßes und der Überstand wird klar (Abbildung 31). Diese instabile Lösung ist ein weiterer Unterschied zu den durch Bakterien reduzierten Nanopartikeln, die eine stabile Suspension mit den enthaltenen Nanopartikeln aufweisen. Bei der Selenatlösung sind auch nach über 100 Tagen keine farblichen Änderungen zu erkennen. Somit fand keine Reduktion zu Selenit oder elementaren Selen unter den gewählten Versuchsbedingungen statt (Abbildung 32). Ergebnisse S e i t e | 43 Abbildung 31: Chemischen Reduktion von Selenit (Bildung von rotem Niederschlag) Abbildung 32: Chemische Reduktion von Selenat (kein Niederschlag) Nach einigen Monaten schlug der Niederschlag von rot zu grau um, welches mit der Oxidation des Selens zu begründen ist. Des Weiteren waren kleinere Partikel zu erkennen, als bei dem Niederschlag der frisch hergestellten chemischen Reduktion. Mittels UV/Vis wurde zum Einen der Unterschied zwischen einer neuen chemischen Reduktion und einer älteren dargestellt und zum Anderen wurde die mikrobielle mit der chemischen Reduktion verglichen. Die UV/Vis-Spektren sind in der Abbildung 33 dargestellt. Hierbei ist deutlich bei beiden Proben ein Peak bei 567 nm zu erkennen, der jedoch bei der neuen chemischen Reduktion stärker ausgeprägt ist. neu alt Absorption 0,05 0,00 500 550 600 650 Wellenlänge (nm) Abbildung 33: UV/Vis Spektren des chemisch reduzierten Selenits Ergebnisse 4.3.2 S e i t e | 44 Elektronenmikroskopie in Kombination mit der energiedispersiven Röntgenspektroskopie (EDX) Die bei der Lichtmikroskopie wahrgenommenen Nanopartikel der Selenitkultur werden mittels Elektronenmikroskopie erneut untersucht. Zum Einen wird dabei REM verwendet, um die dreidimensionale Struktur der Bakterien und der Nanopartikel darzustellen. Mittels TEM-Aufnahmen konnten ebenfalls Aussagen über Form, Größe und Lokalisation der Nanopartikel getroffen werden. Mittels EDX konnte zusätzlich ermittelt werden, aus welchen Elementen die Nanopartikel bestehen. Die Bilder der Rasterelektronenmikroskopie zeigen Bakterien mit Nanopartikeln der Selenitansätze, die schon bei der Lichtmikroskopie beobachtet wurden (Abbildung 34 und Abbildung 35). Zur besseren Darstellung der Bakterien wurden teilweise einige Proben mit Glutaraldehyd fixiert (Abbildung 36). Abbildung 34: A. brasilense mit Nanopartikeln aus dem Selenitansatz (Methode ohne Fixierung) Abbildung 35: Nanopartikel aus dem Selenitansatz (Methode ohne Fixierung) Bei der Untersuchung der Nanopartikel der Selenitkultur mittels energiedispersiver Röntgenspektroskopie wurde bestätigt, dass die Partikel aus Selen bestehen. Der Hauptpeak im EDX-Spektrum (Abbildung 37), bei einer Energie von 1,4 keV, ist dem Selen zuzuordnen. Der Peak bei 0,5 keV zeigt das Vorhandensein von Sauerstoff und der Peak bei 1,75 keV entspricht Silizium. Ergebnisse S e i t e | 45 Diese beiden Stoffe wurden detektiert, da die Proben auf einem Plättchen aus Siliziumdioxid fixiert wurden. Des Weiteren gibt es ein Schwefelsignal bei einer Energie von 2,3 keV. Energie (keV) Abbildung 36: A. brasilense mit Nanopartikeln aus dem Selenitansatz (Fixierung mit Glutaraldehyd) Abbildung 37: EDX-Spektrum der Nanopartikel Vergleicht man die REM-Bilder der Selenitansätze mit denen der Selenatansätze, so wird sichtbar, dass in den Selenatansätzen ausschließlich die Bakterien und keine Nanopartikel (Abbildung 38) zu erkennen sind. Durch das EDX-Spektrum (Abbildung 39) wird bestätigt, dass die Ablagerungen neben den Bakterienzellen im Selenatansatz kein partikuläres Selen enthalten. Das Auftreten von Sauerstoff und Silizium ist durch den Träger zu begründen. Ergebnisse S e i t e | 46 Energie (keV) Abbildung 38: Bakterien ohne Nanopartikel aus dem Selenatansatz (Fixierung mit Glutaraldehyd) Abbildung 39: EDX-Spektrum aus dem Selenatansatz Vergleicht man die beiden angewendeten Methoden untereinander (Abbildung 34 und Abbildung 36), ist zu erkennen, dass wenn die Bakterien mit Glutaraldehyd fixiert wurden, es zu einer besseren Visualisierung führt. Wird der Fokus jedoch auf die Nanopartikel gelegt, dann empfiehlt sich die Methode ohne Glutaraldehyd, da hierbei die Nanopartikel besser dargestellt werden können. Auf dem TEM-Bild (Abbildung 40) sind sphärische Nanopartikel mit einem Durchmesser von 400 nm zu erkennen. Der von den Nanopartikeln umgebene graue „Schatten“ und die Nanopartikel wurden ebenfalls durch EDX analysiert. Ergebnisse S e i t e | 47 Abbildung 40: TEM-Aufnahme der Nanopartikel Dabei wurde bestätigt (Abbildung 41), dass die Nanopartikel größtenteils aus Selen (Peak bei 1,4 keV, 11,5 keV und 12,5 keV) und Schwefel (Peak bei 2,3 keV) bestehen. Neben Selen und Schwefel wurden mittels EDX auch Kohlenstoff, Sauerstoff, Silizium und Kupfer detektiert, welche durch die Probenpräparation auf einem Kupfernetzchen mit einem Kohle/Formvarfilm erklärbar sind. Die Selen-Peaks wurden mit Hilfe der Literatur bestätigt (Oremland et al. 2004). Se Se Abbildung 41: EDX-Spektrum der Nanopartikel Bei der Untersuchung des grauen „Schleiers“ mittels TEM-EDX (Abbildung 42) wurden Stoffe wie Silizium, Sauerstoff, Chlor, Natrium und Magnesium identifiziert. Diese Stoffe sind Rückstände aus der Isolierung. Ergebnisse S e i t e | 48 Abbildung 42: EDX-Spektrum der von Nanopartikel umgebenden Substanz 4.3.3 Bestimmung der Oberflächenladung durch Zetapotentialmessungen Durch Messen des Zetapotentials in Abhängigkeit von dem pH-Wert in dem Bereich 1 bis 7 konnten die Oberflächenladung und der isoelektrische Punkt der Selennanopartikeln ermittelt werden. In Abbildung 43 werden vier Messungen dargestellt, die von zwei unterschiedlichen Ansätzen stammen. Zusätzlich wurde aus allen Messungen der Mittelwert gebildet. Diese Kurve ist ebenfalls mit der jeweiligen Standardabweichung (Std.Abw.) im Diagramm dargestellt. pH-Wert 0 -2 0 1 2 3 4 5 6 7 8 Zeta-Potential (mV) -4 -6 -8 -10 -12 -14 -16 -18 -20 Ansatz 2 vom 1. Exp. (1:30) Ansatz 2 vom 1. Exp. (1:100) MW Zeta-Potential Ansatz 1 vom 2. Exp (1:30) Ansatz 2 vom 2. Exp (1:100) Abbildung 43: Zetapotentiale aller Nanopartikelsuspensionen der Selenitansätze mit Mittelwert aller Zetapotentialmessungen mit Standardabweichung Ergebnisse S e i t e | 49 Alle Kurven besitzen in dem untersuchten pH-Wertbereich ein negatives Zetapotential. Bei pH 1 liegt das Zetapotential in dem Bereich von -0,6 bis -6,3 mV. Mit steigendem pH-Wert verschiebt sich das Oberflächenpotential immer mehr ins Negative. So wurde bei einem pH-Wert von 7 ein Zetapotential in dem Bereich von -15 bis -18 mV gemessen. Da bei keinem pH-Wert ein positives Zetapotential auftrat, konnte kein isoelektrischer Punkt mit einem Zetapotential von Null bestimmt werden. 4.3.4 Dynamische Lichtstreuung Durch das Messen der dynamischen Lichtstreuung der Nanopartikelsuspension wurden der Durchmesser der Selennanopartikel und die dazugehörige Partikelgrößenverteilung ermittelt. Das Auftragen der Partikelgröße nach ihrem prozentualen Anteil ist in der Abbildung 44 dargestellt. Dabei wurden zwei 1 mM-Selenitkulturen verwendet, bei dem sich die Ansätze farblich unterschieden, um zu untersuchen, inwiefern die Farbänderung in Beziehung zu der Partikelgröße steht. 20 18 16 Ansatz 1 Ansatz 2 Intensität (%) 14 12 10 8 6 4 2 0 200 300 400 500 600 700 Log. Partikelgrößenverteilung (d.nm) Abbildung 44: Partikelgrößenverteilung der 1 mM-Selenitansätze 800 Ergebnisse S e i t e | 50 Die orangefarbene Kultur besitzt Nanopartikel mit einem hydrodynamischen Durchmesser in dem Bereich von 220 nm bis 825 nm. Der größte prozentuale Anteil ist bei einer Partikelgröße von 396 nm mit 18,1 % zu finden. Bei der rot-orangenen Kultur ist das Maximum der Partikelgröße ebenfalls bei 396 nm mit einer Intensität von 19,4 %. Die Partikelgrößenverteilung dieses Ansatzes liegt im Bereich von 220 nm und 712 nm. Die berechneten Durchschnittswerte der Partikelgrößen der beiden Ansätze sind in der Tabelle 6 zusammengefasst. Tabelle 6: Durchschnittliche Partikeldurchmesser der verschiedenen Ansätze Selenitansatz Durchschnittlicher Durchmesser (nm) 1 443 2 407 Der orangene Ansatz besitzt im Durchschnitt größere Partikel mit einem Durchmesser von 443 nm. Der rot-orangene Ansatz hat mit 407 nm einen geringeren durchschnittlichen Partikeldurchmesser. Zwar befinden sich im 2. Ansatz mehr kleinere Partikel, jedoch sind die Unterschiede zu gering um die Aussage zu bestätigen, dass die Partikelgrößenverteilung mit der Farbgebung der Kulturen zusammenhängt. Neben der Partikelgrößenverteilung lässt sich aus den Messwerten der DLS zusätzlich eine Autokorrelationsfunktion (Abbildung 45) aus den gemessenen Werten darstellen. Ergebnisse S e i t e | 51 1,0 Korrelationskoeffizient Ansatz 1 Ansatz 2 0,5 0,0 0,1 1 10 100 1000 10000 100000 1000000 Verzögerungszeit [µs] Abbildung 45: Autokorrelationsfunktion Mit Hilfe der Autokorrelationsfunktion können Probleme, beispielsweise zu kleine oder zu große Partikel oder eine kontaminierte Probe identifiziert und interpretiert werden. Der Verlauf der Funktion der beiden Ansätze zeigt eine typische Kurve, die repräsentativ für große Partikel ist. Diese liegen mit einem Durchmesser von durchschnittlich 420 nm noch im messbaren Bereich zwischen 3 nm und 3 µm. Somit stellt diese Größe kein Problem für die Messung dar. Da verschiedene Ansätze mit unterschiedlichen Selenitkonzentrationen verwendet wurden sind die Ergebnisse der 1 mM-Selenitkulturen in Abbildung 46 mit denen der 2,5 mM-Kulturen in Abbildung 47 zum Vergleich gegenübergestellt. Dafür wurde aus der Partikelgrößenverteilung der Durchschnittswert der Partikelgröße der verschiedenen Ansätze berechnet. Durchschnittliche Partikelgröße (d.nm) Ergebnisse S e i t e | 52 1 2 3 4 5 600 Exp. 1; 1 mM Exp. 2; 1 mM (1) Exp. 2; 1 mM (2) Exp. 3; 1 mM (1) Exp. 3; 1 mM (2) 500 MW (443,2 d.nm) 400 300 200 100 0 1 2 3 4 5 Proben Durchschnittliche Partikelgröße (d.nm) Abbildung 46: Durchschnittliche Partikelgrößen der 1 mM-Selenitansätze 1 2 3 4 400 Exp. 3; 2,5 mM Exp. 4; 2,5 mM (1) Exp. 4; 2,5 mM (2) Exp. 4; 2,5 mM (3) MW (341,7 d.nm) 300 200 100 0 1 2 3 4 Proben Abbildung 47: Durchschnittliche Partikelgrößen der 2,5 mM-Selenitansätze Beim Vergleich der beiden Diagramme wird ersichtlich, dass bei einer höheren Selenitkonzentration eine geringere Partikelgröße mit einem Durchschnitt von 341 nm zu erkennen ist. Im Gegensatz dazu besitzen die 1 mM-Selenitkulturen einen durchschnittlichen Durchmesser von 443 nm. Inwiefern die Höhe der Selenitkonzentration (Mittelwert beider 1 mM-Ansätze, 2,5 mMund 5 mM-Ansatz) in der Kultur mit der Partikelgröße der gebildeten Nanopartikel zusammenhängt, wurde in Abbildung 48 dargestellt. Ergebnisse S e i t e | 53 Dabei wurde festgestellt, dass die 2,5 mM-Selenitkultur eine geringe Partikelgrößenverteilung besitzt als die anderen Konzentrationen. Die 2,5 mM-Selenitkultur weist die meisten Nanopartikel mit der Größe 396 nm mit einem prozentualen Anteil von 30,5 % auf. Die höchste Partikelgröße in einer Größenklasse von 459 nm mit einer Intensität von 22 % zeigt hingegen die 1 mM-Selenitkultur. Die Nanopartikel der 5 mM-Selenitkultur besitzen den größten prozentualen Anteil mit 18 % bei einem Partikeldurchmesser von 396 nm. 1 mM 2,5 mM 5 mM 30 Intensität (%) 25 20 15 10 5 0 200 400 600 800 1000 Log. Partikelgrößenverteilung (d.nm) Abbildung 48: Partikelgrößenverteilung der Nanopartikel bei unterschiedlichen Selenitkonzentrationen Werden alle mit dynamischer Lichtstreuung gemessenen Partikelgrößen verglichen (Abbildung 49), so kommt man zu dem Ergebnis, dass die Nanopartikel eine Größe von durchschnittlich 443 nm aufweisen, welche in dem Bereich von 301 nm bis 603 nm variieren. Ergebnisse S e i t e | 54 Durchschnittliche Partikelgröße (d.nm) 600 500 MW (403,1 d.nm) 400 1 2 3 4 5 6 7 8 9 10 Exp. 1; 1 mM Exp. 2; 1 mM (1) Exp. 2; 1 mM (2) Exp. 3; 1 mM (1) Exp. 3; 1 mM (2) Exp. 3; 2,5 mM Exp. 3; 5 mM (2) Exp. 4; 2,5 mM (1) Exp. 4; 2,5 mM (2) Exp. 4; 2,5 mM (3) 300 200 100 0 1 2 3 4 5 6 7 8 9 10 Proben Abbildung 49: Darstellung der durchschnittlichen Partikelgröße aller 1 mM-Selenitkulturen und Ermittlung des durchschnittlichen Durchmessers 4.3.5 Raman-Spektroskopie Zur weiteren Charakterisierung der mikrobiell gebildeten Selennanopartikel wurden Ramanspektren im Wellenzahlenbereich von 150 bis 450 cm-1 aufgenommen. In Abbildung 50 werden die gemessen Spektren zweier Nanopartikelsuspensionen aus den 2,5 mM-Selenitansätzen verglichen. Dabei zeigen beide Proben ein identisches Spektrum, womit die Reproduzierbarkeit bewiesen wird. Bei einer Raman-Verschiebung von 257 cm-1 besitzen beide Proben den ersten Peak. Dieser Peak wird, laut Literatur, einer Selenkette oder einem Se8-Ring zugeordnet (Poborchii et al. 1997). Ein zweiter Peak wird bei beiden Ansätzen bei 355 cm-1 sichtbar und entspricht laut Schmidt und Wilhelm (1970) einer S-Se-Valenzschwingung. Ergebnisse S e i t e | 55 Probe 1 Probe 2 250 Raman-Intensität 200 150 100 50 0 200 250 300 350 400 450 -1 Raman-Verschiebung (cm ) Abbildung 50: Raman-Spektrum der Nanopartikelsuspension 4.3.6 Thermostabilitätstest Bei der Charakterisierung der Nanopartikel wurde die Stabilität gegenüber höheren Temperaturen untersucht. Dabei wurden die isolierten Nanopartikel nach 24-stündiger Erwärmung im Temperaturbereich von 35 °C bis 250 °C mittels UV/Vis und dynamischer Lichtstreuung analysiert. In der Abbildung 51 wird die farbliche Änderung der Nanopartikelsuspensionen nach der Erwärmung bei den jeweiligen Temperaturen deutlich. Dabei wird sichtbar, dass die bei 35 °C-inkubierte Probe ein intensiveres rot besitzt als die Ausgangslösung. Nachdem die Probe bei 50 °C erhitzt wurde, ist zu beobachten, dass die Partikel, nicht wie gewohnt homogen verteilt vorliegen, sondern sich Aggregate gebildet haben, die aufgrund des Gewichtes schneller sedimentieren. Die 100 °C-Probe hat eine blass-rote Färbung, jedoch ist am Boden feines rötliches Sediment erkennbar. Bei dieser Probe konnte außerdem im Teflonbecher des Stahlautoklaven ein rotes Sediment beobachtet werden. Dies war bei den anderen Temperaturen nicht der Fall. Die 250 °C-Probe stellt eine klare farblose Flüssigkeit dar, die optisch keine Nanopartikel mehr enthält. Ergebnisse S e i t e | 56 Ausgangslösung 35 °C 50 °C 100 °C 250 °C Abbildung 51: Aufnahme der Ausgangslösung und der bei unterschiedlich hohen Temperaturen inkubierten Nanopartikelsuspension 4.3.6.1 UV/Vis-Spektroskopie Zur weiteren Charakterisierung der Nanopartikelsuspensionen nach der Inkubation bei den unterschiedlichen Temperaturen (35 °C, 50 °C, 100 °C, 250 °C) wurden Absorptionsspektren im Wellenlängenbereich von 200 bis 1200 nm aufgenommen. Durch Auftragen der Absorption über die Wellenlänge entsteht das Absorptionsspektrum (Abbildung 52) der Nanopartikelsuspension der Ausgangslösung vor und nach der Inkubation bei unterschiedlichen Temperaturen (35 °C, 50 °C, 100 °C, 250 °C). Ergebnisse S e i t e | 57 Ausgangslösung 35 °C 50 °C 100 °C 250 °C 1,0 0,8 Absorption 0,6 0,4 0,2 0,0 -0,2 200 400 600 800 1000 1200 Wellenlänge (nm) Abbildung 52: UV/Vis-Spektren der Nanopartikelsuspension bei unterschiedlichen Inkubationstemperaturen Die Absorptionsspektren der Partikelsuspensionen der Ausgangslösung und der bei 35 °C inkubierten Probe zeigen keine signifikanten Unterschiede. Beide Spektren besitzen einen Peak bei 600 nm und eine Schulter bei 780 nm. Diese Peaks können dem Element Selen zugeordnet werden (Oremland et al. 2004). Bei der 50 °C-inkubierten Probe kommt es zu einer Veränderung der Verhältnisse, wobei der Hauptpeak und die Schulter nun eine Wellenlänge zwischen 600 nm und 800 nm aufweisen. Die Lösung bei 100 °C und 250 °C weisen kaum eine Absorption auf. Ein vergrößerter Ausschnitt der zweier UV/Vis-Spektren ist in Abbildung 53 dargestellt. Ergebnisse S e i t e | 58 100 °C 250 °C Absorption 0,03 0,00 -0,03 600 800 Wellenlänge (nm) Abbildung 53: UV/Vis-Spektren der Nanopartikelsuspension bei einer Inkubation von 100 °C und 250 °C Hierbei ist zu sehen, dass bei der 100 °C Probe eine geringe Absorption von 0,02 bei 620 nm zu erkennen ist. Dies bestätigt die Beobachtung, dass bei dieser Probe nach der Inkubation rotes Sediment im Teflonbecher aufzufinden und nach dem Resuspendieren eine schwach rote Lösung zu erkennen war. Bei der 250 °C Probe wurde hingegen eine klare Lösung wahrgenommen. 4.3.6.2 Dynamische Lichtstreuung Mit Hilfe der dynamischen Lichtstreuung wurde nach der Inkubation der Nanopartikelsuspension der Einfluss von unterschiedlichen Temperaturen auf die Partikelgrößenverteilung (Abbildung 54) untersucht. Ergebnisse S e i t e | 59 Ausgangslösung 35 °C 50 °C 100 °C 35 30 Intensität (%) 25 20 15 10 5 0 1000 Log. Partikelgrößenverteilung (nm) Abbildung 54: Partikelgrößenverteilung der Ausgangslösung und der Proben, die bei 35 °C, 50 °C und 100 °C inkubiert wurden Mittels dieser Ergebnisse wurde aus der Partikelgrößenverteilung der jeweiligen Probe ein Partikelgrößendurchschnitt bestimmt (Tabelle 7). Tabelle 7: Inkubationstemperaturen mit dazugehörigem durchschnittlich gemessenem Durchmesser Inkubationstemperatur Durchmesser (nm) Ausgangslösung (RT) 301,2 35 °C 370,1 50 °C 2329,5 100 °C 380,7 250 °C - Ergebnisse S e i t e | 60 Dabei ist gut zu erkennen, dass die Ausgangslösung und die Proben, die bei 35 °C und bei 100 °C inkubiert wurden, einen ähnlichen Partikeldurchmesser besitzen. Bei der 250 °C-Probe konnte keine Partikelgrößenverteilung bestimmt werden. Dies war zu vermuten, da die Nanopartikelsuspension nach der Inkubation keine rötliche Färbung mehr besaßen, sondern eine farblose Flüssigkeit im Teflonbecher des Stahlautoklaven zurückblieb. Nach einer Wärmebehandlung bei 50 °C wiesen die Partikel mit 2329 nm einen fast 8-mal größeren Durchmesser als die Partikel in der Ausgangslösung auf. 4.3.6.3 Bestimmung des TOC-Gehaltes Die Bestimmung des Kohlenstoffgehalts soll Aufschluss über das Vorhandensein organischer Substanzen liefern. Die Ergebnisse sind in der Abbildung 55 grafisch dargestellt. TOC-Gehalt 30 28,4 25 15 Ausgangslösung TOC [mg/l] 20 10 5 10,2 3,1 2,5 1,9 0 0 50 100 150 200 250 Temperatur (°C) Abbildung 55: Grafische Darstellung des TOC-Gehaltes der Nanopartikelsuspension vor und nach der Inkubation bei unterschiedlichen Temperaturen Zunächst ist ein eindeutiger Trend, angefangen bei der Ausgangslösung (RT) bis zu den höher temperierten Nanopartikelsuspensionen, zu erkennen. Die Ausgangslösung besitzt einen Kohlenstoffgehalt von 1,9 mg/l. Danach ist eine Steigerung der Konzentration zu Ergebnisse S e i t e | 61 beobachten. Bei der Probe, die 24 h bei 35 °C erhitzt wurde, ist ein Wert von 2,5 mg/l Kohlenstoff gemessen worden. Eine weitere Erhöhung der Konzentration ist auch bei der 50 °C-Probe zu erkennen. Eine Ausnahme mit 3,1 mg/l bildet die Probe, die bei 100 °C erwärmt wurde, denn diese besitzt einen geringeren Kohlenstoffgehalt als die 50 °C-Probe. Den höchsten Gehalt an TOC mit 28,4 mg/l besitzt die bei 250 °-inkubierte Probe. Je mehr Nanopartikel durch die Inkubation zerstört wurden, desto höher ist auch der gemessene TOC-Gehalt. Diskussion 5 5.1 S e i t e | 62 DISKUSSION REDUKTION VON SELENIT/SELENAT In dieser Arbeit wurden signifikante Unterschiede zwischen den Wachstumskurven der Selenit- und Selenatkulturen ersichtlich. Da die Wachstumskurven der Kontroll- und Selenatkultur einen fast identischen Verlauf aufweisen, kann davon ausgegangen werden, dass das Bakterium A. brasilense das im Medium enthaltene Selenat nicht in seinem Stoffwechsel aufnimmt und Selenat somit nicht toxisch bei der eingesetzten Selenatkonzentration von 1 mM auf seinen Metabolismus wirkt. Im Gegensatz dazu wirkt sich Selenit bereits ab einer Konzentration von 1 mM negativ auf den Wachstumsverlauf der A. brasilense-Kulturen aus. Sowohl der Verlauf der optischen Dichte, des Gesamtproteingehalts und auch des pH-Wertes spiegelten während der Kultivierung eine starke Verlängerung der lag-phase im Zellwachstum wieder. Dies spricht für die hohe Toxizität des löslichen Oxyanions Selenit. Nach der lag-Phase folgt ebenfalls bei den Selenitkulturen eine Steigerung der Zelldichte, welches auf eine Anpassung der Bakterien an die Kultivierungsbedingungen zurückzuführen ist. Diese Wachstumsverzögerung der Bakterien in Gegenwart von Selenit wurde bereits in der Literatur beschrieben (Sarret et al. 2005). Es konnte durch die Ergebnisse der HG-AAS und der ICP-MS (Kapitel 4.2.2) gezeigt werden, dass Selenit mikrobiell zu elementarem Selen reduziert wurde. Innerhalb von sieben Tagen konnte das Bakterium A. brasilense 1 mM Selenit vollständig zu Selennanopartikel umwandeln. Dagegen konnte Selenat durch A. brasilense nicht reduziert werden. Die Selenatkonzentration blieb während der gesamten Kultivierung konstant. Dies bestätigt die Aussage, dass Selenat nicht in den Metabolismus des A. brasilense aufgenommen wird. Studien belegen, dass unterschiedliche Arten von Bakterien nach der Reduktion von Selenit rote, amorphe Allotrope von Se(0) bilden (Kessi et al. 1999, Oremland et al. 2004). Dabei konnte ein Zusammenhang zwischen der Farbintensität der suspendierten Nanopartikel (Kapitel 4.3.1) und dem Selengehalt im Medium (Kapitel 4.2.5) hergestellt werden. Die blass-rötlich gefärbten Suspensionen, zeigten bei der HG-AAS und bei der ICP-MS eine geringere Abnahme des Selenits aus dem Medium und dementsprechend auch eine geringere Bildung der Selennanopartikel. Diskussion S e i t e | 63 Dieser Zusammenhang kann auch mit dem Wachstumsverhalten und der Ermittlung der Gesamtzellproteinkonzentration hergestellt werden. Je schneller die lag-Phase überwunden wird, desto eher beginnen die Bakterien mit der Reduktion des elementaren Selens. Da nur ein Ansatz bei den 5 mM-Kulturen Bakterienwachstum aufwies, konnte hier schon vermutet werden, dass nur geringe Mengen an Nanopartikel gebildet wurde. Dies spiegelt sich auch bei der blass-rosa Farbe der isolierten suspendierten Nanopartikel wieder. In Kapitel 4.2 wurde beschrieben, dass Auffälligkeiten in Bezug auf die Farbentwicklung der Kulturen mit Selenit zu beobachten sind. Während der Kultivierung wurde eine schwach rote Farbe der Selenitkulturen wahrgenommen, die bis zum Ende der Kultivierung intensiver wurde. In anderen Publikationen wurde während des Kultivierungszeitraumes der Selenitansätze eine Bandbreite von verschiedenen Rottönen angegeben. Dabei wurde ebenfalls bei A. brasilense, in Gegenwart von Selenit, eine Veränderung der Kulturen, beobachtet. Dies äußerte sich in durch eine schwache Rotfärbung, die sich bis zum Ende der Kultivierung ebenfalls intensivierte. Dieser Effekt wurde bei Tugarova et al. (2012) bis zu einer Selenitkonzentration von 1 mM beschrieben. Im Kapitel 4.2.3 wurde beschrieben, wie tolerant das Bakterium A. brasilense gegenüber unterschiedlich hohen Selenitkonzentrationen ist. Dabei war eine Tendenz zu erkennen, dass mit steigender Selenitkonzentration im Kulturmedium mehr Zeit benötigt wird, um das Maximum der Zelldichte zu erreichen. Die maximale wachstumsinhibierende Selenitkonzentration konnte nicht eindeutig bestimmt werden, da sich die 5 mM-Ansätze untereinander sehr unterschieden. Tugarova et al. (2013) untersuchte die Wirkung von höheren Selenitkonzentration auf A. brasilense. Bei der Kultivierung auf festem Medium wurde eine Wachstumshemmung bei einer Selenitkonzentration von 10 mM festgestellt. Bei der Charakterisierung der Wachstumsverläufe der Kulturen bei unterschiedlichen Selenitkonzentrationen besitzen alle ein lag-Phase, die bis zum 3. Tag andauerte. Die Ergebnisse der HG-AAS und der ICP-MS hingegen verdeutlichen, dass mit steigender Selenitkonzentration im Kultivierungsmedium die Selenitabnahme und die damit verbundene Nanopartikelbildung später einsetzt. Bei einer Konzentration von 1 mM beginnt die Abnahme der Selenitkonzentration zwischen dem 3. und dem 6. Tag, hingegen die Abnahme bei den 2,5 mM erst am 7. Tag beginnt. Diskussion S e i t e | 64 Die 5 mM-Selenitkultur, bei der ein Wachstum sichtbar wurde, zeigte innerhalb von 10 Tagen keine Reduzierung der Selenitkonzentration. Dabei war die Selenkonzentration zum Schluss höher als am Anfang. Diese Erhöhung könnte dadurch zu begründen sein, dass aufgrund der Kultivierung im Wärmeraum ein geringer Teil des Mediums verdampft und das Selenit somit höher konzentriert ist. Um eine eindeutige Aussage über die Höhe der Selenitkonzentration, die zum Hemmung des Wachstums führt, machen zu können, sind weitere Versuche notwendig. Damit kann eindeutig ein Zusammenhang zwischen Wachstumsverhalten von A. brasilense und Abnahme des Selenits im Medium und der damit verbundenen Bildung der elementaren Selennanopartikel bewiesen werden. Je schneller sich die Bakterien in Gegenwart von Selenit erholen, desto eher beginnt die Reduktion des Selenits zu elementarem Selen. Neben den mikrobiell erzeugten Nanopartikel wurden zum Vergleich Untersuchungen mit chemisch reduziertem Selen durchgeführt. Bei dem Vergleich der UV/Vis-Spektren, wurde beobachtet, dass die chemisch reduzierte Selenitlösung einen Peak bei 667 nm aufwies und demnach einen Peak bei einer ähnlichen Wellenlänge besitzte wie die mikrobiell erzeugten Nanopartikel. Somit kann davon ausgegangen werden, dass es sich bei beiden um elementares Selen handelt. Die geringe Verschiebung des Peaks könnte mit dem Schwefel in Verbindung stehen, welches in mikrobiell erzeugten Partikeln nachzuweisen ist. Durch das in der Arbeit verwendete Reduktionsmittel Hydroxylamin konnte ausschließlich Selenit chemisch reduziert werden, welches durch Bildung eines roten Niederschlags zu beobachten war (Kapitel 4.3.1). Dies deutet auf eine Reduktion von Selenit zu elementarem Selen hin. Es entstand im Gegensatz zum mikrobiell erzeugten Nanopartikeln keine stabile Suspension. Beim Selenat hingegen wurde optisch keine Veränderung sichtbar. Diese Beobachtungen wurden in der Literatur ebenfalls mit anderen reduzierenden Reagenzien beschrieben. Viele Publikationen beschreiben bei der Verwendung verschiedenster Bakterien, dass bei der chemischen Reduktion mit unterschiedlichen Reduktionsmitteln keine Selennanopartikel, sondern ein roter flockiger Niederschlag entsteht (Bruggeman et al. 2005, Bruggeman et al. 2007, Scheinost & Charlet 2008). Diskussion S e i t e | 65 Selenit- und Selenat reduzierende Reagenzien, die in der Literatur beschrieben werden, sind beispielsweise Hydrazin (Buckley 1976), Ascorbinsäure (Shaker 1996) und Natriumascorbat (Mees et al. 1995). 5.2 CHARAKTERISIERUNG DER NANOPARTIKEL In der vorliegenden Arbeit konnte gezeigt werden, dass das Bakterium A. brasilense die Fähigkeit besitzt unter aeroben Bedingungen das giftige Selenit in das weniger gefährliche elementare Selen in Form von Nanopartikeln umzusetzen. Nachdem die Nanopartikel isoliert (Kapitel 3.5.10) und mit VE-Wasser resuspendiert wurden, blieb eine kaminrote Lösung zurück, welche auf die Farbe der Nanopartikel zurückzuführen ist. Die Nanopartikel wurden über mehrere Monate in VE-Wasser gelagert. Dabei war zu beobachten, dass im Gegensatz zu den chemisch reduzierten Partikeln, eine stabile Suspension entstand. In dieser Zeit lösten sich die Nanopartikel nicht in Wasser und agglomerierten auch nicht. Diese Beobachtung wird durch Oremland et al. (2004) bestätigt. Durch Lagerung der Nanopartikel in destilliertem Wasser über einen Zeitraum von mehreren Monaten behielten diese die Struktur bei und können somit als stabil angesehen werden. Die unter dem Lichtmikroskop beobachteten Nanopartikel konnten, ebenfalls mittels REM und TEM visualisiert und über EDX als ein Selen-Schwefel-Gemisch identifiziert werden. Dabei lagen die Nanopartikel außerhalb der Zelle vor. Nur durch TEM-Dünnschnitte der Bakterien kann eindeutig bestimmt werden, ob die Nanopartikel innerhalb und/oder außerhalb der Bakterienzelle vorkommen. In der Veröffentlichung von Tugarova et al. (2013) wird ein Versuch mit ähnlichen Bedingungenbeschrieben. Die Kultivierung des A. brasilense erfolgte sieben Tage bei aeroben Bedingungen und wurde während des Versuches nicht geschüttelt. Die dabei entstandenen Partikel lagen intrazellulär vor (Tugarova et al. 2013). Andere in der Literatur erwähnten Bakterien besaßen ebenfalls die Fähigkeit Nanopartikel in Gegenwart von Selenit zu bilden waren sowohl intrazellulär als auch extrazellular lokalisiert (Oremland et al. 2004) Mittels EDX konnte ebenfalls gezeigt werden, dass der von Nanopartikel umgebene „Schatten“, weitestgehend aus Spurenelementen besteht und dies auf Rückstände aus der Isolierung der Nanopartikel zurückzuführen ist. Diskussion S e i t e | 66 Durch Bestimmen der Oberflächenladung (Kapitel 4.3.3) wurde bei einem pH-Wert von 7 ein Oberflächenpotential in dem Bereich -15 bis -18 mV gemessen. Während der Bestimmung des Zetapotentials konnte in dem untersuchten pH-Bereich von 1 bis 7 kein isoelektrischer Punkt ermittelt werden. Diese starke Ladung der Partikel ist für die Stabilität und Ausbildung der Nanopartikelsuspension notwendig, da sich gleichgeladene Partikel voneinander abstoßen und nicht agglomerieren. Für Nanopartikel, die durch Bakterium Bacillus cereus gebildet wurden, konnte eine Ladung von -46,86 mV bestimmt werden (Dhanjal & Cameotra 2010). Mit Hilfe der dynamischen Lichtstreuung (Kapitel 4.3.4) wurde die Partikelgrößenverteilung ermittelt. Für die durch A. brasilense gebildeten Selennanopartikel konnte daraus ein durchschnittlicher Partikeldurchmesser von 400 nm berechnet werden. Dieser Wert stimmt mit der Ausmessung der isolierten Nanopartikel der TEM-Aufnahme (Kapitel 4.3.2) überein. Bei den 1 mM-Selenitkulturen wurde häufig die Auffälligkeit beobachtet, dass sich die Ansätze schon kurz nach Zugabe des Selenits sich farblich unterscheiden. So färbten sich einige Ansätze orange und andere hingegen rot. Durch die Analyse mittels DLS sollte untersucht werden, inwiefern ein Zusammenhang zwischen Farbe und Partikelgröße bzw. deren Verteilung besteht. Die orangefarbene Kultur besaß Partikel mit einem durchschnittlichen Durchmesser von 443 nm, die rötliche hingegen 407 nm. Da die Unterschiede so gering sind, kann nicht eindeutig gesagt werden, ob eine Abhängigkeit zwischen Partikelgröße und Färbung der Kultur besteht. Bei der Messung der Optischen Dichte und der Bestimmung der Gesamtzellproteinkonzentration konnten hingegen große Unterschiede festgestellt werden. Ansätze, die sich während der Kultivierung orange färbten, wiesen ein starkes Wachstum auf (ODmax bei 4 bis 5) und eine hohe Proteinkonzentration (cmax = 1780 µg/ml bis 2350 µg/ml) auf. Die rötlichen Kulturen hingegen besaßen durchschnittlich eine OD von 3 und eine Gesamtzellproteinkonzentration von 800 µg/ml. Nach circa zwei Wochen im Wärmeraum konnte beobachtet werde, dass sich die orangene Färbung der Kultur sich zu Rot änderte. Dieser Farbwechsel wird auch von Lin und Wang (2005) beschrieben, wobei diese die Änderung von gelb zu rot als einen Beweis für die Vergrößerung der Partikelgröße der Selennanopartikel angeben. Diskussion S e i t e | 67 Andere, in der Literatur angegebene, Bakterien produzieren in Gegenwart von Selenit ebenfalls sphärische Partikel, bei denen von dem Pseudomonas sp. gebildeten Nanopartikeln ein Durchmesser von 400 nm und bei Bacillus sp. die Partikel mittels REM-Aufnahme auf 200 bis 300 nm geschätzt wurden (Kashiwa et al. 2000, Hunter & Manter 2009). Bei dem Vergleich der durchschnittlichen Partikelgröße der unterschiedlich hohen Selenitkonzentrationen ist eine Tendenz erkennbar. Je höher die Selenitkonzentration im Medium, desto kleiner ist die durchschnittliche Partikelgröße. Jedoch sind die Unterschiede so gering, dass nicht eindeutig bestimmt werden kann, dass die Partikelgröße von der Höhe der Selenitkonzentration abhängig ist. Betrachtet man die Verteilung der Partikel, so ist zu erkennen, dass sich die Partikel des 5 mM-Ansatzes in dem Bereich 190 bis 712 nm aufteilen, wo hingegen die Partikel des 1 mM-Ansatzes sich zwischen 295 und 1106 nm befinden. Dies kann mit Länge der lag-Phase zu begründen sein. Die 1 mM-Kulturen besitzen eine kürzere lag-Phase und beginnen dementsprechend eher mit Wachstum als die anderen Kulturen. Somit haben die Bakterien bei einer geringeren Selenitkonzentration mehr Zeit, um das Selenit aus dem Medium zu Nanopartikel zu reduzieren, welches wiederrum Einfluss auf die Nanopartikelgröße hat. Der in dieser Arbeit detektierten Peaks bei 257 cm-1 und 355 cm-1 werden auch in der Literatur als Strukturen von Selen beschrieben. Die Raman-Verschiebung bei 257 cm-1 wird in Publikationen einer Selenkette und einem Se8-Ring zugeordnet (Poborchii et al. 1997). Der kleine Peak bei 355 cm-1 könnte vor dem Hintergrund der TEM-EDX-Ergebnisse als Selen-Schwefelverbindung interpretiert werden. In Schmidt und Wilhelm (1970) wird beschrieben, dass Banden, die um 360 cm-1 liegen, der S-Se-Valenzschwingung zugeordnet werden. Dies würde mit den Aussagen der EDX-Analyse übereinstimmen, bei der die Nanopartikel als ein Selen-Schwefel-Gemisch identifiziert wurden. In Oremland et al. (2004) wurden ebenfalls die Nanopartikel dreier Bakterienstämme beschrieben, die mittels Raman-Spektren charakterisiert wurden. Ein ähnliches Spektrum, wie das in dieser Arbeit gemessene, besitzt einen Peak mit einer RamanVerschiebung von 240 cm-1 zeigen die Nanopartikel des Bakteriums. Die Nanopartikel des Bakterienstamm S. shriftii wiesen Se8 strukturelle Einheiten auf. In Kohara et al. (1998) wird die typische strukturelle Anordnung der Selen-Atome beschrieben. Diese liegt in verschiedenen Oxidationsstufen als Ringe oder Ketten vor. Dabei variiert die molekulare Zusammensetzung oder die Kettenlänge, beginnend von Se3 bis S12. Diskussion S e i t e | 68 Durch Inkubation der Nanopartikelsuspension bei unterschiedlich hohen Temperaturen (Kapitel 4.3.6) wurde die Thermostabilität der Nanopartikel untersucht und anschließend mittels dynamischer Lichtstreuung und UV/Vis charakterisiert. Des Weiteren wurde der TOC-Gehalt der Lösungen gemessen. Schon nach dem Erhitzen wurden bereits rein optische Unterschiede sichtbar (Abbildung 51), die sich auch in den UV/Vis-Spektren zeigten. Da die Spektren der Ausgangslösung identisch (Peak bei 600 nm) mit der bei 35 °C inkubierten Probe sind, kann davon ausgegangen werden, dass hier keine Änderung in der Atomanordnung zu verzeichnen ist. Bei einer Temperatur von 50 °C beginnen die Partikel ihre Größe zu ändern oder zu agglomerieren, sodass die roten Nanopartikel nach einer gewissen Zeit sedimentieren. Dies spiegelt sich auch bei der Messung der DLS wieder. Die durchschnittliche Partikelgröße nimmt mit zunehmender Inkubationstemperatur zu. Eine „Ausnahme“ bildet sich 50 °C-inkubierte Probe, bei dem ein hydrodynamischer Durchmesser von 2330 nm gemessen. Dies ist ein 7-fach größerer Durchmesser als die Partikel bei den anderen unterschiedlichen Inkubationstemperaturen. Die UV/Vis Spektren der 100 °C- und 250 °C-Probe wiesen kaum eine Absorption auf. Jedoch wurde bei der 100 °C-inkubierten Probe ein kleiner Peak bei 620 nm detektiert werden. In Lin und Wang (2005) werden die Absorptionsspektren für eine Folge verschiedener Größen der Selennanopartikel dargestellt. Die Partikel, die ebenfalls bei einer Wellenlänge von 600 nm einen Peak aufwiesen, besitzen einen Partikeldurchmesser von 182,8 ± 33,2 nm. Je größer die Partikel werden, desto mehr verschiebt sich der Peak in höhere Wellenlängen. Das Absorptionsspektrum der isolierten Selennanopartikel des Bakteriums Bacillus cereus unterstützt ebenfalls die Aussage, dass Selen bei einer Wellenlänge von 600 nm aufzuweisen ist (Dhanjal & Cameotra 2010). Dieser Unterschied, zwischen den in dieser Arbeit bestimmten Partikelgröße und deren Partikeldurchmesser, könnte mit der von Dhanjal und Cameotra (2010) durchgeführten chemischen Reduktion zu begründen sein. Die TOC-Messungen der unterschiedlich temperierten Proben sollte Aufschluss über das Vorhandensein organischer Substanzen liefern, welche an der Nanopartikelbildung beteiligt sind. Dazu wurde der Gehalt an organisch gebundenen Kohlenstoff bestimmt. Es kann gesagt werden, dass mit steigender Temperatur mehr organisch gebundener Kohlenstoff vorhanden ist. Grund dafür könnte sein, dass mit steigender Temperatur Diskussion S e i t e | 69 zunehmend Nanopartikel zerstört werden und dementsprechend auch der TOC-Ghealt erhöht wird. Die Ausnahme stellt die 100 °C-Probe dar, die einen geringeren TOC-Gehalt aufwies. Ein Grund könnte das bei der Erhitzung gebildete rote Sediment sein, da dieses mittels Pipette nicht resuspendieren und somit nicht aufgenommen werden konnte. Daher fehlt ein gewisser Teil an Nanopartikel, der mittels TOC-Messung nicht mit erfasst werden konnte. Ausblick 6 S e i t e | 70 AUSBLICK Die in der vorliegenden Arbeit untersuchten Aspekte gaben Aufschlüsse über die Verträglichkeit von Selenat und Selenit gegenüber des Bakteriums A. brasilense. Die Untersuchungen zeigten, dass nur das Oxyanion Selenit toxisch auf das Bakterium wirkt. Durch Variation der Selenitkonzentration konnte unter gewählten Bedingungen und Konzentrationen gezeigt werden, inwieweit das Bakterium A. brasilense in der Lage ist, eine bestimmte Selenitkonzentration zu tolerieren, sodass es lebensfähig bleibt. Aufgrund der Reduktion von Selenit zu elementaren Selen bildeten sich extrazellulär Nanopartikel, die mit Hilfe verschiedener Methoden charakterisiert wurden. Die Erkenntnis, dass die Nanopartikel bei einer 24-stündigen Inkubation bei 50 °C einen größeren Partikeldurchmesser annehmen kann für die Isolierung der Nanopartikel in großem Maßstab von Nutzen sein. Diese könnten leichter mittels Sieb abgetrennt werden als kleinere Partikel. Für die Betrachtung des Bakteriums in natürlicher Umgebung wären weitere Untersuchungen von großem Interesse. Dabei könnte ermittelt werden, ob das Bakterium in der Natur ebenfalls das Verhalten äußert Selenit zu reduzieren und Nanopartikel zu bilden. Demnach könnte Wasserproben aus regionalen Flüssen oder Seen entnommen werden und beispielsweise durch Zugabe von Selenit das Verhalten beobachtet werden. Des Weiteren kann sowohl die Sorption von Selennanopartikel an Mineralen untersucht werden als auch die Sorption von nuklearen Stoffen, beispielsweise Europium und Uran, an Selennanopartikel und wäre somit für die Endlagerforschung von großem Interesse. Auch die Sorption an Metallen, beispielsweise Kalzium, Magnesium und Eisen, könnte in nachfolgenden Experimenten analysiert werden. Bei der Kultivierung des A. brasilense wurden einige Auffälligkeiten bezüglich des aeroben Wachstums festgestellt, welche durch weitere Experimente untersucht werden können. Die Bakterienkulturen, die bei der Kultivierung, aufgrund der unterschiedlichen Größen der Erlenmeyerkolben, eine größere Oberfläche und damit verbundenen größeren Kontakt mit Sauerstoff, besaßen, begannen schneller mit der Produktion der Selennanopartikel. Diese Aerotaxis wird in einigen Publikationen in Gegenwart einer geringen Sauerstoffkonzentration beschrieben (Barak et al. 1982) und die Bakterien wandern zur optimalen Sauerstoffkonzentration (Zhulin et al. 1996). Ausblick S e i t e | 71 In einigen Publikationen wird davon berichtet, dass das Bakterium A. brasilense in der Lage ist, das Phytohormon Indolessigsäure zu produzieren. Inwieweit ein Zusammenhang zwischen der Produktion dieser Indolessigsäure und Bildung der Nanopartikel besteht, könnte in weiterführenden Experimenten untersucht werden. Literaturverzeichnis 7 S e i t e | 72 LITERATURVERZEICHNIS Ardenne M, Musiol G, Klemradt U (2005) Effekte der Physik und ihre Anwendungen, Vol 3. Wissenschaftlicher Verlag Harri Deutsch, Frankfurt am Main Barak R, Nur I, Okon Y, Henis Y (1982) Aerotactic Response of Azospirillum brasilense. J Bacteriol 152:643-649 Bashan Y, de-Bashan LE (2010) How the Plant Growth-Promoting Bacterium Azospirillum Promotes Plant Growth—A Critical Assessment. In: Advances in Agronomy. Elsevier Inc Bashan Y, Levanony H, Klein E (1986) Evidence for a Weak Active External Adsorption of Azospirillum brasilense Cd to Wheat Roots. J Gen Microbiol 132:3069-3073 Bashan Y, Levanony H, Whitmoyer RE (1991) Root Surface Colonization of Non-Cereal Crop Plants by Pleomorphic Azospirillum brasilense Cd. J Gen Microbiol 137: 187-196 Berzelius JJ (1817) Lettre de M. Berthollet sur deux metaux nouvaux. Annales Chimie et de Physique 7:199-207 Berzelius JJ (1818) Recherches sur un nouveau corps minera truove dans le souffre fabrique a Fahlun. Annales Chimie et de Physique 9:160-180 (225-267, 337-365) Bloemberg GV, Lugtenberg BJJ (2001) Molecular basis of plant growth promotion and biocontrol by rhizobacteria. Biotic interactions: 343-350 Bruggeman C, Maes A, Vancluysen J (2007) The interaction of dissolved Boom Clay and Gorleben humic substances with selenium oxyanions (selenite and selenate). Appl Geochem 22:1371-1379 Bruggeman C, Maes A, Vancluysen J, Vandenmussele P (2005) Selenite reduction in Boom clay: Effect of FeS2, clay minerals and dissolved organic matter. Environmental pollution 137:209-221 Buckley DA (1976) Red amorpous selenium. US Patent 3:951-954 Chapman PP, Adams WJ, Brooks ML, Delos CG, Luoma SN, Maher WA, Ohlendorf HM, Presser TS, Shaw DP (2010) Ecological Assessment of Selenium in the Aquatic Environment, Vol 1. Crc Pr Inc, Pensacola Dhanjal S, Cameotra SS (2010) Aerobic biogenesis of selenium nanospheres by Bacillus cereus isolated from coalmine soil. Microb Cell Fact 9:52 Dhillon KS, Dhillon SK (2003) Distribution and management of seleniferous soils. Adv Agron 79:119-184 Dowdle PR, Oremland RS (1998) Microbial oxidation of elemental selenium in soil slurries and bacterial cultures. Environ Sci Technol 32:3749-3755 DSMZ (2007) 221. AZOSPIRILLUM MEDIUM. URL: http://www.dsmz.de/ microorganisms/medium/pdf/DSMZ_Medium221.pdf [Stand: 8.April 2013] Fernández-Martínez A, Charlet, L. (2009) Selenium environmental cycling and bioavailability: a structural chemist point of view. Rev Environ Sci Biotechnol 8:81-110 Literaturverzeichnis S e i t e | 73 Frankenberger WT, Jr., Arshad M (2001) Bioremediation of selenium-contaminated sediments and water. Biofactors 14:241-254 Gafny R, Okon Y, Kapulnik Y, Fischer M (1986) Adsorption of Azospirillum brasilense to Corn Roots. Soil Biol Biochem 18:69-75 Gao XY, Zhang JS, Zhang L (2002) Hollow sphere selenium nanoparticles: Their in-vitro anti hydroxyl radical effect. Adv Mater 14:290-293 Hädener A, Kaufmann H (2006) Grundlagen der organischen Chemie, Birkhäuser Verlag, Basel, Bosten, Berlin Herbel MJ, Blum JS, Oremland RS, Borglin SE (2003) Reduction of elemental selenium to selenide: Experiments with anoxic sediments and bacteria that respire Se-oxyanions. Geomicrobiol J 20:587-602 Hesse M, Meier H, Zeeh B (2005) Spektroskopische Methoden in der organischen Chemie, Vol 7. Georg Thieme Verlag, Stuttgart Hunter W, Manter D (2009) Reduction of Selenite to Elemental Red Selenium by Pseudomonas sp. Strain CA5. Curr Microbiol 58:493-498 Kashiwa M, Nishimoto S, Takahashi K, Ike M, Fujita M (2000) Factors affecting soluble selenium removal by a selenate-reducing bacterium Bacillus sp SF-1. J Biosci Bioeng 89:528-533 Kausch P, Matschullat J (2005) Rohstoffe der Zukunft - Neue Basisstoffe und neue Energien, Vol 1. Frank & Timme GmbH - Verlag für wissenschaftliche Literatur, Berlin Kessi J, Ramuz M, Wehrli E, Spycher M, Bachofen R (1999) Reduction of selenite and detoxification of elemental selenium by the phototrophic bacterium Rhodospirillum rubrum. Appl Environ Microbiol 65:4734-4740 Kloepper J, Zablotowicz R, Tipping E, Lifshitz R (1991) Plant growt promoting mediated by bacterial rhizosphere colonizers. The rhizosphere and plant growth:315-326 Kohara S, Goldbach A, Koura N, Saboungi ML, Curtiss LA (1998) Vibrational frequencies of small selenium molecules. Chem Phys Lett 287:282-288 Lemly AD (2004) Aquatic selenium pollution is a global environmental safety issue. Ecotox Environ Safe 59:44-56 Less RL, Eckardt S, Kettner M, Schmitt F, Walter B (2008) Die handlungsorientierte Ausbildung für Laborberufe, Vol 2. Vogel Services GmbH, Würzburg Levander OA, Burk RF (2006) Update a human dietary standards for selenium. In: Hatfield DL, Berry MJ, Gladyshev VN (eds) Selenium its molecular biology and role in human health. Springer, New York Levanony H, Bashan Y (1991) Active Attachment of Azospirillum brasilense to Root Surface of Non-Cereal Plants and to Sand Particles. Plant Soil 137:91-97 Lide D (1994) CRC handbook of chemistry and physics: a ready-reference book of chemical and physical data, CRC Pr Inc, Boca Raton Lin ZH, Wang CRC (2005) Evidence on the size-dependent absorption spectral evolution of selenium nanoparticles. Mater Chem Phys 92:591-594 Malvern Instruments (2007) Zetasizer Nano-User Manual. Worcestershire Literaturverzeichnis S e i t e | 74 McNeal JM, Balistrieri L (1989) Geochemistry and Occurrence of Selenium: An Overview. In: L.W. J (ed) Selenium in Agriculture and the Environment. Soil Science Society of America, Madison, Winsconsin Mees DR, Pysto W, Tarcha PJ (1995) Formation of Selenium Colloids Using Sodium Ascorbate as the Reducing Agent. J Colloid Interf Sci 170:254-260 Miyagi K, Sampson RW, Tsujino I, Gunther WHH, Krieg M, Sieber F (2003) Proteinated subnanoparticles of elemntal selenium potentiate the anti-tumor effect of ionizing radiation and selected chemotherapeutic agents. Proc Am Assoc Res 44:385 Murty MG, Ladha JK (1987) Differential Colonization of Azospirillum Lipoferum on Roots of 2 Varieties of Rice (Oryza-Sativa-L). Biol Fert Soils 4:3-7 Neukirchen D (2009) Chemikalien für die Photovoltaik. URL: http://www.laborpraxis. vogel.de/laborbedarf/chemikalien/articles/228147/. In: Laborpraxis [Stand: 6.August 2013] Okon Y, Labanderagonzalez CA (1994) Agronomic Applications of Azospirillum - an Evaluation of 20 Years Worldwide Field Inoculation. Soil Biol Biochem 26:15911601 Oremland RS, Herbel MJ, Blum JS, Langley S, Beveridge TJ, Ajayan PM, Sutto T, Ellis AV, Curran S (2004) Structural and spectral features of selenium nanospheres produced by se-respiring bacteria. Appl Environ Microbiol 70:52-60 Patriquin DG, Dobereiner J, Jain DK (1983) Sites and Processes of Association between Diazotrophs and Grasses. Can J Microbiol 29:900-915 Peitzsch M (2008) Speziation mikrobilogisch alkylierter, leichtflüchtiger Selenverbindungen in Abhängigkeit der geochemischen Verfügbarkeit des Selens. Doktor der Naturwissenschaften, Johannes-Gutenberg-Universität Mainz, Mainz Pingoud A, Urbank C (1997) Arbeitsmethoden der Biochemie, de Gruyter, Berlin, New York Poborchii VV, Kolobov AV, Oyanagi H, Romanov SG, Tanaka K (1997) Structure of selenium incorporated into nanochannels of mordenite: dependence on ion exchange and method of incorporation. Chem Phys Lett 280:10-16 Presser TS (1994) The Kesterson Effect. Environ Manage 18:437-454 Raven PH, Evert RF, Eichhorn SE (2000) Biologie der Pflanzen, Vol 3. de Gruyter, Berlin, New York Rothballer M, Schmid M, Hartmann A (2003) In situ localization and PGPR-effect of Azospirillum brasilense strains colonizing roots of different wheat varieties. Symbiosis 34:261-279 Sarathchandra SU, Watkinson JH (1981) Oxidation of Elemental Selenium to Selenite by Bacillusn Megaterium. Science 211:600-601 Sarret G, Avoscan L, Carriere M, Collins R, Geoffroy N, Carrot F, Coves J, Gouget B (2005) Chemical forms of selenium in the metal-resistant bacterium Ralstonia metallidurans CH34 exposed to selenite and selenate. Appl Environ Microbiol 71:2331-2337 Literaturverzeichnis S e i t e | 75 Scheinost AC, Charlet L (2008) Selenite reduction by mackinawite, magnetite and siderite: XAS characterization of nanosized redox products. Environ Sci Technol 42: 1984-1989 Schmidt KH, Gebel A (2000) Massenspektroskopie mit induktiv gekoppeltem Plasma. Institut für Mineralogie und mineralische Rohstoffe der TU Clausthal. URL: http://www.icp-ms.de/ [Stand: 18.Juni 2013]. Schmidt M, Wilhelm E (1970) Diselenacyclooctasulfur and Triselenacyclooctasulfur. Z Naturforsch Pt B B 25:1348 Schwarz K, Foltz CM (1957) Selenium as an Integral Part of Factor-3 against Dietary Necrotic Liver Degeneration. Journal of the American Chemical Society 79: 3292-3293 Seby F, Gautier MP, Lespes G, Astruc M (1997) Selenium speciation in soils after alkaline extraction. Sci Total Environ 207:81-90 Shaker AM (1996) Kinetics of the reduction of Se(IV) to Se-sol. J Colloid Interf Sci 180:225-231 Sieber F (2003) Selenium in oxidation state zero is a potent and selective antileukemia/lymphoma agent. Exp Hematol 31:119-120 Smith PK, Krohn RI, Hermanson GT, Mallia AK, Gartner FH, Provenzano MD, Fujimoto EK, Goeke NM, Olson BJ, Klenk DC (1985) Measurement of protein using bicinchoninic acid. Analytical Biochemistry 150:76-85 Soda S (2009) Biotechnology for Recovery of Rare Metals in Wastewater. Graduate School of Engineering, Osaka University, Junba Stolz JF, Oremland RS (1999) Bacterial respiration of arsenic and selenium. Fems Microbiol Rev 23:615-627 Tugarova AV, Vetchinkina EP, Loshchinina EA, Shchelochkov AG, Nikitina VE, Kamnev AA (2013) The ability of the rhizobacterium Azospirillum brasilense to reduce selenium(IV) to selenium(0). Microbiology 82:352-355 Virupaks TK, Shrift A (1965) Biochemical Differences between Selenium Accumulator and Non-Accumulator Astragalus Species. Biochim Biophys Acta 107:69 Wang ZJ, Gao YX (2001) Biogeochemical cycling of selenium in Chinese environments. Appl Geochem 16:1345-1351 Zhang YQ, Zahir ZA, Frankenberger WT (2003) Factors affecting reduction of selenate to elemental selenium in agricultural drainage water by Enterobacter taylorae. J Agr Food Chem 51:7073-7078 Zhulin IB, Bespalov VA, Johnson MS, Taylor BL (1996) Oxygen taxis and proton motive force in Azospirillum brasilense. J Bacteriol 178:5199-5204 Eidesstattliche Erklärung 8 S e i t e | 76 EIDESSTATTLICHE ERKLÄRUNG Hiermit erkläre ich eidesstattlich, dass die vorliegende Masterarbeit ohne unzulässige Hilfe Dritter und ohne Benutzung anderer als der von mir angegeben Hilfsmittel angefertigt habe. Die aus fremden Quellen direkt oder indirekt übernommenen Gedanken sind als solche kenntlich gemacht. Dresden, den 30.September 2013 Anika Maffert Danksagung 9 S e i t e | 77 DANKSAGUNG Als erstes möchte ich Herrn Prof. Dr. rer. nat. habil. Thomas Wiegert für die Betreuung meiner Masterarbeit sowie sein Interesse am Fortgang der Arbeit danken. Die vorliegende Masterarbeit wurde in der Abteilung Biogeochemie am Institut für Ressourcenökologie des Helmholtz-Zentrums Dresden-Rossendorf e.V. angefertigt. An dieser Stelle möchte ich mich bei allen Personen bedanken, die durch ihre tatkräftige Unterstützung zum Entstehen dieser Arbeit beigetragen haben. Dabei gilt mein ganz besonderer Dank meinen Betreuern Frau Dr. Manja Vogel, Herr Dr. Robin Steudtner und Frau Dr. Carola Franzen, bei denen jederzeit die Tür für mich offen stand, um Fragen zu beantworten und Probleme zu lösen. Außerdem danke ich ihnen für die lehrreichen Gespräche, die anregenden Diskussionen, die fachliche Kompetenz und die mitreißende „wissenschaftliche Neugier“ am zu bearbeitenden Thema. Weiterhin möchte ich mich bei meinen Bürokolleginnen Julia Berger, Maria Berger, Sandra Köppchen und Dorina Köpke für die vielen wissenschaftlichen und privaten Diskussionen, die entspannte Arbeitsatmosphäre und die stete Hilfsbereitschaft aller bedanken. Des Weiteren möchte ich mich herzlich für die tatkräftige Hilfe beim Korrigieren der grammatischen Fehler bei meinen Freundinnen Julia Berger und Maria Berger bedanken, die sich bereit erklärt haben, meine Arbeit gegenzulesen. Nicht zuletzt gilt mein besonderer und tiefer Dank meinen Eltern, meiner Oma und meinem Freund, die mir immer bei Seite standen, mich unterstützten und immer ein offenes Ohr für mich hatten.