The Role of the FIS Polycomb Group Complex - ETH E
Werbung

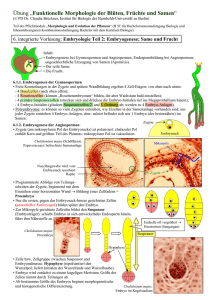
DISS. ETH Nr. 19788 The Role of the FIS Polycomb Group Complex during Seed Development A dissertation submitted to the ETH ZÜRICH For the Degree of DOCTOR OF SCIENCES Presented by ELISABETH CHRISTINE HEHENBERGER Diplom-Ingenieurin University of Natural Resources and Applied Life Sciences, Vienna Born on 8th December 1980 Citizen of Austria Accepted on the recommendation of Prof. Claudia Köhler, examiner Prof. Samuel C. Zeeman, co‐examiner Dr. Bruno Müller, co‐examiner 2011 Summary The eukaryotic genome is packaged into a highly ordered structure called chromatin. The basic unit of the chromatin, the nucleosome, is composed of DNA that is wrapped around histone proteins, resulting in a beads-on-a-string configuration. Interaction of neighboring nucleosomes determines the condensation status of the chromatin. Highly condensed chromatin prevents access of the transcriptional machinery to the DNA, thereby regulating gene expression. The ability of chromatin to condense can be regulated by modifications of the histones at their exposed N-terminal tails. Modifications can influence internucleosomal interactions, thus leading to the opening or condensation of local chromatin regions. Polycomb Group (PcG) proteins maintain specific gene-expression patterns by histone tail modification of their target genes. PcG proteins are conserved in animals and in plants and function by forming large protein complexes. In the model plant Arabidopsis thaliana PcG proteins form at least three complexes with distinct functions. The FERTILIZATION INDEPENDENT SEED (FIS) PcG complex acts during reproductive development by suppressing seed formation without fertilization and by promoting proper seed development after fertilization. Lack of the FIS PcG complex subunit FERTILIZATION INDEPENDENT SEED2 (FIS2) leads to initiation of seed development without fertilization and, if fertilization occurs, to strongly disturbed endosperm differentiation, arrest of embryo development and eventually seed abortion. In my work, I first addressed the question if the failure of the endosperm to cellularize in the fis2 mutant can be correlated with the developmental arrest of the embryo. For this purpose I investigated seeds also displaying a non-cellularization phenotype caused by loss of ENDOSPERM DEFECTIVE1 (EDE1). Thereby I could demonstrate that a failure of the endosperm to cellularize is connected with an arrest in embryo development. Transcript profiling of fis2 seeds revealed a strong deregulation of genes involved in carbohydrate metabolism, suggesting a disturbed nutrient content in fis2 seeds. I could show that hexose levels are indeed increased in developing fis2 seeds. Localization of the hexoses within the seed revealed that the accumulation of hexoses in fis2 seeds is - 11 - probably the consequence of an early developmental arrest. Furthermore, by comparing fis2 and ede1, I found a correlation between increased hexose levels and endosperm cellularization failure. I investigated the underlying reasons for elevated hexose levels in fis2 seeds and discovered a deregulation of invertase activity and expression. Manipulation of invertase expression affected the hexose levels only mildly, but overexpression of one invertase caused abnormal embryo development, introducing a new role for this specific class of invertases in seed development. The transcription factor AGL62 was described as repressor of endosperm cellularization. I found that reduction of AGL62 levels in the fis2 mutant restores endosperm cellularization and allows completion of seed development. Subsequent interaction analysis showed that AGL62 is a direct target of the FIS PcG complex. Finally I discovered that the exact timing of endosperm cellularization initiation decides over the fate of the embryo, probably in correlation with the levels of AGL62. Thus, the obtained results implement AGL62 as a key target of the FIS PcG complex and a central regulator of endosperm development. - 12 - Zusammenfassung Das eukaryotische Genom ist in einer hochkomplexen Struktur verpackt, die als Chromatin bezeichnet wird. Die Grundeinheit des Chromatins, das Nucleosom, besteht aus DNA die um Histon Proteine gewickelt ist, wodurch eine Perlenkettenformation ensteht. Durch die Interaktion benachbarter Nucleosome wird der Kondensierungsstatus des Chromatins bestimmt. Stark kondensiertes Chromatin verhindert den Zugang der Transkriptionsmaschinerie zur DNA, wodurch die Genexpression reguliert werden kann. Durch Modifizierung der exponierten Enden der Histone kann die Kondensierungsfähigkeit des Chromatins gesteuert werden. Diese Modifizierungen können die Interaktionen zwischen den Nucleosomen beeinflussen und auf diese Weise zu einer Öffnung beziehungsweise einer Verdichtung bestimmter Chromatinregionen führen. Polycomb group (PcG) Proteine kontrollieren die Aufrechterhaltung spezifischer Genexpressionsmuster indem sie die Histon Enden ihrer Zielgene modifizieren. PcG Proteine sind in Tieren und Pflanzen evolutionär konserviert und ihre Funktion beruht auf der Bildung von grossen Proteinkomplexen. In der Modellpflanze Arabidopsis thaliana bilden die PcG Proteine mindestens drei verschiedene Komplexe mit unterschiedlichen Funktionen. Der FERTILIZATION INDEPENDENT SEED (FIS) PcG Komplex agiert in der reproduktiven Phase der Pflanzenentwicklung indem er vor der Befruchtung die Bildung von Samen verhindert und nach der Befruchtung für eine korrekte Samenentwicklung sorgt. In Abwesenheit des FIS PcG Komplexes kommt es auch ohne Befruchtung zu einer beginnenden Samenentwicklung. Im Falle einer Befruchtung führt die Abwesenheit vom FIS PcG Komplex zu einer stark gestörten Endospermentwicklung, Abbruch der Embryoentwicklung und schlussendlich zum Samenabort. In meiner Arbeit habe ich mich zuerst mit der Frage befasst ob der Endospermzellularisierungdefekt in der fis2 Mutante mit dem Entwicklungsabbruch des Embryos in Zusammenhang steht. Zu diesem Zweck habe ich Mutanten untersucht, die ebenfalls einen Endospermzellularisierungdefekt aufweisen, in diesem Fall verursacht durch Abwesenheit von ENDOSPERM DEFECTIVE1 (EDE1). Auf diese Weise konnte ich - 13 - zeigen dass eine erfolglose Endospermzellularisierung mit einem Entwicklungsabbruch des Embryos korreliert. Die Transkriptionsanalyse von fis2 Samen hat gezeigt, dass vor allem Gene die im Kohlenhydratmetabolismus eine Rolle spielen, in dieser Mutante dereguliert sind. Diese spezifische Anreicherung deutet auf einen gestörten Nährstoffgehalt in fis2 Samen hin. Ich konnte zeigen, dass der Hexosezuckergehalt in fis2 Samen während ihrer Entwicklung tatsächlich erhöht ist. Die Lokalisierung der Hexosezucker im Samen hat ergeben, dass die Anreicherung von Hexosezuckern in fis2 Samen wahrscheinlich durch einen frühen Entwicklungsabbruch verursacht wird. Durch den Vergleich der fis2 und der ede1 Mutanten konnte ich ausserdem einen Zusammenhang zwischen erhöhtem Hexosezuckergehalt und einer erfolglosen Endospermzellularisierung herstellen. Die Suche nach den Gründen für den erhöhten Hexosezuckergehalt in fis2 Samen hat zur Entdeckung von zwei deregulierten Invertasen geführt. Durch Manipulierung der Invertaseexpression konnte ich den Hexosegehalt nur wenig beeinflussen, aber die Überexpression einer Invertase führte zu einer abnormalen Embryoentwicklung. Auf diese Weise konnte ich eine neue Rolle für diese spezielle Klasse von Invertasen identifizieren. Der Transkriptionsfaktor AGL62 dient zur Unterdrückung einer frühzeitigen Endospermzellularisierung. Durch Reduktion der AGL62 Level in der fis2 Mutante konnte ich eine Wiederherstellung der Endospermzellularisierung und eine vollständige Samenentwicklung erreichen. Mit der anschliessenden Interaktionsanalyse konnte ich ausserdem zeigen, dass AGL62 ein Zielgen vom FIS PcG Komplex ist. Schliesslich habe ich herausgefunden, dass präzise zeitliche Koordination der Endospermzellularisierung entscheidenden Einfluss auf die Embryoentwicklung hat. Der Beginn der Endospermzellularisierung hängt wahrscheinlich vom Expressionslevel von AGL62 ab. Die gewonnenen Resultate implementieren somit den Transkriptionsfaktor AGL62 als eines der wichtigsten Zielgene des FIS PcG Komplexes sowie als zentralen regulierenden Faktor während der Endospermentwicklung. - 14 -