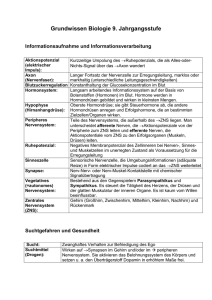
Quiz-Fragen
Werbung

Biofragen Apel Kapitel 3, Wasser und die Umwelt 1. Bestimmen Sie die Molarität von 2l Wasser (Molar Gewicht von Wasser = 18g/mol) 2l = 2000g 1 Mol = 18g 2000/18 = 111,11 mol (d.h.: 55,56 mol/l) 2. Wieviele mol Wasser befinden sich in 5l Wasser? 5l = 5000g 1 Mol = 18g 5000/18 = 277,78 mol 3. Bestimmen Sie die molare Konzentration von Protonen in einer wässrigen Lösung mit dem pH 1? Wie hoch ist die Hydroxidionen Konzentration in der gleichen Lösung? pH = -log[H+] [H+] = 10-pH = 10-1 pH = 14 – pOH [OH-] = 10-pOH = 10-13 4. Durch eine Membran werden zwei zelluläre Kompartimente getrennt, die einen pH von 7 bzw. 4 besitzen. Wie hoch ist der Unterschied der Protonenkonzentration zwischen beiden Kompartimenten? pH = 7 [H+] = 10-7 pH = 4 [H+] = 10-4 10-4/10-7 = 103 1000 x mehr H+ Ionen bei pH 4 im Vergleich zu pH 7 Antworten Quiz Fragen 7. We can be sure that a mole of table sugar and a mole of vitamin C are equal in their c.number of molecules 8. How many grams of acetic acid (C2H4O2) would you use to make 10l of a 0.1 M aqueous solution of acetic acid? (Note: The atomic weights, in Daltons, are approximately 12 for carbon, 1 for hydrogen and 16 for oxygen.) d. 60 g 9. Acid precipitation has lowered the pH of a particular lake to 4.0. What is the hydrogen ion concentration of the lake? c. 10-4 M 10. What is the hydroxide ion concentration of the lake described in question 9? c. 10-10 M Kapitel 7, die Zelle 1. Nenne Unterschiede zwischen prokaryotischen und eukaryotischen Zellen Die beiden Zelltypen unterscheiden sich deutlich im inneren Aufbau. Prokaryoten: Eukaryoten: - Typisch für die Organismenreiche der - typisch für die Organismenreiche der Archae und Eubakterien. Protisten, vielzelligen Pflanzen, Pilze Und Tiere - keinen Zellkern ->Nucleoid = Bereich, - echter Zellkern, der von einer Membran in dem sich das genetische Material umschlossen ist. Konzentriert, ist aber von keiner Membran Umschlossen und nicht von der übrigen Zelle getrennt. - haben kein endoplasmatisches System - Cytoplasma (Bereich zwischen Zellkern und Zellmembran) bestehend aus halbflüssigem Cytosol, in das die Organellen mit ihrer spezialisierten Form und Funktion eingelagert sind (die meisten von ihnen fehlen in Prokaryoten). - keine Kompartimentierung - Kompartimentierung der Zelle führt dazu, dass verschiedene Reaktionen gleichzeitig ablaufen können, die sich eigentlich nicht vertragen. Prokaryotische Zelle Klein (0.1-10 Mikrometer Durchmesser) Kein Zellkern, die DNA liegt im Nucleoid (einer bestimmten Region) Eukaryotische Zelle Gross (10-100 Mikrometer Durchmesser) Zellkern, der durch eine Membran vom Rest der Zelle abgeschlossen ist und wird von keiner Membran umschlossen Die meisten Organellen der eukaryotischen Zelle kommen nicht vor Keine membrangebundenen Organellen Kein Membranfluss Bereich zwischen Zellkern und Zellmembran nennt man Cytoplasma (aus halbflüssigem Cytosol, in das Organellen eingelagert sind) Membrangebundene Organellen Mitose, Meiose Pendelströmung gesteuert durch Kontraktion des Plasmas 2. Warum sind Zellen mikroskopisch klein? Zellen sind mikroskopisch klein, da das Verhältnis von Volumen und Oberfläche stimmen muss, damit die lebenswichtigen Vorgänge ablaufen können. Die Plasmamembran grenzt die Zelle nach Aussen ab. Sie ist eine Schranke, die Atemgase, Nährstoffe und Abfallstoffe passieren müssen, um die Lebensvorgänge in der ganzen Zelle aufrecht zu halten. Durch jeden Quadratmikrometer kann aber pro Sekunde nur eine begrenzte Stoffmenge hindurchtreten, deshalb muss die Oberfläche im Verhältnis zum Volumen sehr gross sein um die Aufgabe zu meistern. Dies ist der Grund, weshalb die meisten Zellen mikroskopisch klein sind. Die Oberfläche muss im Verhältnis zum Volumen gross genug sein, um genügend Stoffaustausch über die Plasmamembran zu ermöglichen. Ein zu grosses Volumen würde Stoffmengen zum Austausch bereitstellen, die von der kleinen Oberfläche nicht schnell genug ausgetauscht werden könnten. Deshalb besitzen grössere Organismenarten in der Regel keine grösseren, sondern mehr Zellen als kleinere Lebewesen. 3. Beschreibe den Aufbau und die Funktion der Plasmamembran. Die Plasmamembran besteht aus einer Phospholipiddoppelschicht, in die eine Vielfalt von Proteinen eingelagert ist. Phospholipide sind amphipathisch ( sie haben einen hydrophoben und hydrophilen Teil), wobei der hydrophobe Teil ins Innere der Schicht und der hydrophile Teil (Kopf) nach aussen schaut. An der Aussenseite der Plasmamembran sind noch Kohlenhydrate angeheftet. Die Plasmamembran grenzt die Zelle gegen aussen ab. Sie steuert den Stoffaustausch zwischen der Zelle und ihrer Umgebung. Sie verfügt über selektive Permeabilität (sie erlaubt bestimmten Substanzen den Durchtritt leichter als anderen). Die Plasmamembran besteht aus einer Doppelschicht aus Phospholipiden, in die eine Vielfalt von Proteinen eingelagert ist. An der Aussenseite der Plasmamembran sind Kohlenhydrate und Proteine angeheftet. Die Schwänze der Phospholipide im Innern der Doppelschicht sind hydrophob, die Köpfe hydrophil wie auch die Enden der Proteine, die aus der Membran herausschauen. Membranen sind flüssig, das heisst, sie sind nicht starr. Die Proteine und Lipide haben eine Eigenbewegung (Drehung um eigene Achse oder Seitwärtsdriften). Je höher die Temperatur ist, desto mehr Bewegung entsteht, desto flüssiger ist die Membran. Die speziellen Funktionen der Plasmamembran sind davon abhängig, welche Phospholipide und Makromoleküle jeweils vorhanden sind. Abgrenzung der Zelle Selektive Barriere, die den Stoffaustausch ermöglicht 4. Nenne die Zusammensetzung und Funktion des Endomembransystems. Zusammensetzung und Funktion des Endomembransystems: Das Endomembransystem besteht aus vielen der verschiedenen Membranen einer Eukaryotenzelle. Diese sind entweder unmittelbar verbunden oder es erfolgt einen Austausch via Vesikel (winzige membranumhüllte Bläschen). Aus der Vernetzung folgt aber nicht, dass sich alle inneren Membranen in Form und Funktion gleichen; Dicke, molekulare Zusammensetzung, sowie Stoffwechselfunktion einer Membran sind nicht festgelegt, sondern können sich im Laufe ihrer Existenz mehrfach ändern. Das Endomembransystem besteht aus: - der Kernhülle - dem endoplasmatischen Retikulum (ER) - dem Golgi-Apparat - den Lysosomen - verschiedenartigen Vakuolen - der Plasmamembran ( die nach ihrer Lage keine eigentliche innere Membran ist, aber mit dem ER und anderen Membranen über Vesikel in Verbindung steht.) Kernhülle Endoplasmatisches Retikulum (glattes und raues) Golgi- Apparat Lyosomen Vakuolen Vesikel Funktion: Glattes endoplasmatisches Retikulum: o Wirkt bei vielfältigen Stoffwechselvorgängen mit, unter anderem beim Kohlenhydratstoffwechsel und bei der Beseitigung von Giften und Arzneimitteln o Synthese von Fettsäuren, Phospholipiden, Steroiden und anderen Lipiden o Steuerung der Kalziumionen in Muskelzellen Raues endoplasmatisches Retikulum: o Proteinsynthese o Membranproduktion Golgi- Apparat: o Hier werden die Produkte des ER abgewandelt, gespeichert und dann zu anderen Bestimmungsorten weiterbefördert (Fertigungs-, Lager-, Sortier- und Versandzentrale) o Erzeugt manche Markomoleküle selbst Lyosomen: o Dient zur intrazellulären Verdauung von Makromolekülen (Magen und Mülleimer der Zelle) o In Lyosomen befinden sich Enzyme für die Hydrolyse von Proteine, Polysacchariden, Fetten und Nukleinsäuren Vakuolen: o Nahrungsvakuole o Kontraktile Vakuole (pumpt überschüssiges Wasser aus der Zelle) o Zentralvakuole der Pflanzen (Speicherung von organischen Verbindungen) Vesikel: o Verteilen Produkte der ER 5. Was sind cis- und trans-Zisternen des Golgi-Apparates, welche Rolle spielen sie? Der Golgi-Apparat besitzt eine eindeutige Polarität, das heisst die Zisternen an seiner konvexen und konkaven Seite unterscheiden sich in ihrer Struktur und Funktion. Die beiden Pole des Golgi-Apparats werden als cis- und trans-Seite bezeichnet: Die (konvexe) cis-Seite ist in der Regel dem ER und dem Zellkern zugewandt und nimmt Substanzen auf, welche an der (konkaven) trans-Seite, die zur Plasmamembran zeigt, den Golgi-Apparat wieder verlassen. Die Vesikel kommen auf der cis-Seite an, die enthaltene Substanz wird vom Golgi-Apparat aufgenommen, chemisch abgewandelt, gespeichert und auf der trans-Seite wieder in Vesikel verpackt und ins Cytosol abgegeben. Cis-Zisternen (dem ER und dem Zellkern zugewandt) empfangen Material vom ER durch Transportvesikel, die mit der Membran des Golgi- Apparates verschmelzen Trans-Zisternen (zeigt zur Plasmamembran) schnüren Transportvesikel ab, die Stoffe vom Golgi- Apparat wegzutransportieren Sie verbinden den Golgi- Apparat mit dem restlichen Endomembransystem und erlauben so den Stoffaustausch zwischen den Organellen der Zelle (Transport). 6. Aufgaben und Besonderheiten des Lysosoms Lysosom: ein membrangebundener Sack von hydrolytischen Enzymen, welche die Zelle braucht um Makromoleküle zu verdauen. Die intrazelluläre Verdauung durch Lysosome wir auf verschiedene Arten ausgeführt: Phagozytose: dabei werden kleine Organismen oder andere Esspartikel umschlossen, die so entstehende Vakuole fusioniert mit einem Lysosom Autophagie: dabei wird zelleigenes organisches Material recyclet. Wenn das Lysosom ein anderes Organell oder eine kleine Menge Cytosol umschliesst, die lysosomalen Enzyme das umschlossene Material abbrechen, dann werden die organischen Monomere zum Cytosol zur Wiederverwendung retourniert. Die lysosomalen Enzyme arbeiten am besten bei pH5. Indem die lysosomale Membran H+Ionen vom Cytosol ins Innere des Lysosoms pumpt, wird dieser pH aufrecht erhalten. Die Lysosomen sind Membransäcke, die hydrolytische Enzyme zur Verdauung von Makromolekülen (Proteine, Säuren, Fette, Polysaccharide) enthalten. Die H+Membranpumpen dieser Lysosomen schaffen ein pH-Wert von 5. Bei solchen Bedingungen sind die Enzyme am effizientesten. Im Cytosol haben sie keine Wirkung, da pH 7 herrscht. Transportvesikel mit inaktiven Enzymen werden vom rauhen ER gebildet und fliessen zum Golgi-Apparat, der die Enzyme aktiviert und sie an die Lysosomen weitergibt. Bei der Phagozytose (z.B.: Essvorgang der Amöben) fusionieren die entstandenen Vakuolen mit den Lysosomen und die Enzyme verdauen den Vakuloeinhalt. Bei der Autophagie werden zelleigene Organellen/Baustoffe zerlegt und die entstandene Monomere ins Cytosol zum recyceln zurückgegeben. Das Fehlen solcher Lysosomen führt zu seltene, schwere Krankheiten. 7. Wichtigkeit der zellulären Kompartimentierung Arbeitsteilung, Zellschutz, örtliche und zeitliche Trennung Lysosom: arbeitet nur bei pH5. Der Rest der Zelle braucht aber pH7 -> Abgrenzung nötig Nucleus: hält Erbmaterial zusammen -> erleichtert die Zellteilung und schützt die Gene durch Abtrennung von den anderen Organellen vor Schädigung und Mutationen Mitochondrium: Oberflächenvergrösserung im Innern erlaubt Energieumsetzung im Grossen Stil ohne Verlust an die Umgebung Ribosomen: schwimmen nicht nutzlos im Cytosol herum, sondern sind dort konzentriert, wo man sie braucht (z.b. rER) Zentralvakuole (bei Pflanzen): Tonoplast ist selektiv im Transport von löslichen Stoffen und ermöglicht in Innern eine andere Zusammensetzung der Flüssigkeit (cell sap) als ausserhalb (cytosol); disposel site for metabolic by-products that would endanger the cell if they accumulated in the cytosol; bewirkt Oberflächenvergrösserung Durch die Kompartimentierung sind verschiedene pH-Millieu möglich innerhalb einer einzigen Zelle. So können die Enzymeigenschaften am besten genutzt werden und verschiedene inkompatible Reaktionen können in einer Zelle gleichzeitig stattfinden. Nucleus und Ribosome, Endomembranes System, Chloroplasten, Mitochondrien, Perixome, 8. Arten von Vakuolen Food Vakuole (Essvakuole): Durch Phagocytose gebildet Kontraktile Vakuole: In vielen Frischwasserprotisten Pumpen überschüssiges Wasser aus der Zelle Zentralvakuole Durch eine Membran (Tonoplast) abgeschlossen Speicherung, Abfallentsorgung, Schutz und Wachstum Nahrungsvakuolen: In Amöben Phagozytose, Zwischenspeicherung der Nahrung Kontraktionsvakuolen: In süsswasser Protisten zum Schutz vor dem Platzen Zentralvakuole: bei Pflanzen, wird vom Tonoplasten umschlossen, zur Speicherung von Nährstoffen, Abfallentsorgung, zum Schutz (enthalten Alkaloide, Duftstoffe) und für das Wachstum 9.membranöse Organelle, die nicht zum Endomembransystem gehören Mitochondrien, Chloloplasten, Peroxysomen Warum nicht? Weil ihre Membranproteine nicht vom ER, sondern von freien Ribosomen im Cytosol (und in Mitochondrien und Chloroplasten: in ihnen enthaltenen Ribosomen) hergestellt werden, weil sie Ribosomen und DNA enthalten, semiautonom sind, unabhängig wachsen und sich fortpflanzen Beispiele und deren funktionelle Besonderheiten: convert energy to forms that the cells can use for work: Mitochondria: site of cellular respiration, the catabolic process that generates ATP by extracting energy grom sugars, fats, and other fuels with help of oxygen Chloroplasts: site of photosynthesis: convert solar energy to chemical energy by absorbing sunlight an using it to drive the synthesis of organic compounds from carbon dioxide and water haben andere Ribosomen Peroxysomen: Fettspaltung Mitochondrien + Chloroplasten: Sie besitzen eigene DNA, welche die Synthese von Proteinen durch die eigenen Ribosomen steuert. Sie sind semiautonom: wachsen und vermehren sich in der Zelle ohne Einfluss der anderen Organellen. Mitochondrien: sind durch 2 Membranen umgeschlossen. Die Äussere ist glatt und die Innere ist sehr stark gefaltetCristae, Innenraum ist dadurch in 2 Teile geteilt: Zwischenraum und Matrix. In der Matrix befinden sich Enzyme, welche der Zellatmung (Energieherstellung) dienen. Chloroplasten: nur bei Pflanzen. Chloroplasten enthalten das Pigment Chlorophyll und andere Enzyme und Moleküle, die zur Photosynthese benötigt werden. Ebenfalls in 2 Membranen umschlossen. Im Innern gibt es Thylakoiden, die gestapelt die Granas ausbilden. Die Flüssigkeit ausserhalb der Thylakoiden heisst Stroma. Perixome: ist ein spezialisiertes, metabolisches Kompartiment. Nur von 1 Membran umschlossen. Enthält Enzyme, die H2 mit O2 reagieren lassen, es entsteht das toxische H2O2, das mit einem weiteren Enzym zu Wasser neutralisiert wird. Einige Perixome zerlegen Moleküle in kleinere (für die Zellatmung), andere (in der Leber) machen Alkohole und andere Giftstoffe unschädlich. 10. Struktur und Funktion der Bestandteile des Cytoskeletts Mikrotubuli Eine "Röhre" aus Tubulibproteinen im Cytoplasma aller Eukaryonten, in Cilien, Flagellaten und in Cytoskelett Stabilisierung von Zellen, Zellteilung -Tubuli Mikrofilament "Schnur" aus Actinproteinen im Cytoplasma der meisten eukaryontischen Zellen, Teil des Cytoskeletts, alleine oder zusammen mit Myosin (-> Zellkontraktion) Cytoplasmaströmung Bewegungsabläufe der Zelle -> Motorproteine nötig Intermediäre Filamente Komponente des Cytoskeletts, dass alle Filamente beinhaltet, die in der Grösse zwischen Mikrotubuli und Mikrofilamenten liegen Aus Teratinproteinen Cytoskelett: Netzwerk aus Fibrillen (Mikrotubuli, Mikrofilamente und intermediäre Filamente), die im ganzen Cytosol sind. Mikrotubuli (25nm) sind röhrenförmig und bestehen aus Tubulin (globuläres Protein). Fkt.: Zellbewegung, Bewegung der Chromosomen während der Zellteilung, Bewegung von anderen Organellen, Erhaltung der Zellform. Mikrofilamente (7nm) bestehen aus Aktin (auch globuläres Protein), genauer genommen aus 2 ineinander verdrehte Aktinketten. Fkt.: Erhaltung und Veränderungen der Zellform, Muskelkontraktion, Cytoplasmafluss, Zellbewegung und Zellteilung Mikrotubuli und Mikrofilamente interagieren mit den Motorproteine durch ATP-Verbrauch und ermöglichen somit Zellmobilität. Intermediären Filamente (10nm) bestehen aus verschiedenen Arten von Keratinen. Fkt.: Fixierung der Zellgestalt und der Organellen, Erhaltung der Zellform, Verankerung von Kern und anderen Organellen und die Formation der Kernlamina. 11. abschliessende Zelloberfläche und Kontakt zur Nachbarstelle bei Tieren und Pflanzen Tierzelle: Abgrenzung: Plasmamembran Kontakt: Festelterbindung (Gürtel) Demosomen (Vernetzung) Gap junction Pflanzenzelle: Abgrenzung: Zellwand Kontakt: Zellwandverdickungen an dieser Stelle ausgespart Bei Pflanzenzellen: Zellwand, besteht aus Mikrofibrillen von Zellulose, dient zum Schutz, und der Formerhaltung, sowie vor ungehinderten Wasseraufnahme. Junge Pflanzen bilden eine primäre Zellwand, danach entsteht eine Zwischenlamelle (reich an Pektin), zum Schluss bildet sich noch die sekundäre Zellwand. Bei den meisten anderen tierischen Zellen wird die Grenze durch die Zellmembran geschaffen. ECM (Extrazelluläre Matrix) besteht aus Glycoproteinen (vor allem Collagen). Diese Proteine befinden sich in einem Netzwerk aus Proteoglykagenen (auch Glycoproteine). Beide Proteinarten sind mit Fibronektine gebunden, die wiederum an Rezeptoren; den Integrinen angemacht sind, welche über die Zelloberfläche verteilt sind. Somit können Veränderungen vom ECM zum Cytoskelett übertragen werden und umgekehrt. Interzelluläre Verbindungen: Die benachbarten Zellen besitzen kleine Löcher mit Kanälen, den sogenannten Plasmodesmaten. Cytoplasma kann hierdurch ausgetauscht werden. 3 verschiede Arten: Tight junction: Membranen zweier benachbarten Zellen sind fusioniert, Austauschmöglichkeit der Zellflüssigkeit. Desmosomen: befestigen Zelle in starken Blättern, werden durch intermediären Filamente verstärkt. Gap junction: Cytoplasmatunnel, Poren sind offen für Salzionen, AS, Zucker-Austausch. Quiz-Fragen 1. Die Motormoleküle, die jedes Mikrotubulidoppel mit dem nächsten verbinden, bestehen aus dem, Protein Dynein. Diese Dynein-Arme sind zuständig für die Fortbewegung der Cilien und Flagellaten. Wenn durch eine Mutation diese Dynein-Arme nicht mehr funktionieren, dann können sich die Spermien nicht mehr fortbewegen und die Cilien in der Lunge funktionieren auch nicht mehr. 2. Gebundene Ribosome synthetisieren Membran- und Sekretionsproteine. -> Machen Proteine für die Einsetzung in die Membrane, für das Hineinpacken in gewisse Organelle wie Lysosome, oder für den Export aus der Zelle (Sekretion) 3. Chloroplasten (Photosyntheseorganell) gehören nicht zum Endomembransystem (Zellmembran, ER, Golgi, Lysosome, Plasmamembran) 4. ER->Golgi->Vesikel, die mit der Plasmamembran fusionieren. Im Golgi-Apparat werden die Produkte des ER modifiziert und gespeichert und dann in Vesikeln an andere Stellen geschickt. 5. Chloroplasten, Zellwand aus Cellulose, und Tonoplast nur in Pflanzenzellen, Centriolen nur in Tierzellen, Mitochondrien in beiden. 6. Ribosome; Mitochondrien und Chloroplaste sind prokaryontische Zellen, ER kommen nur in eukaryontischen Zellen vor. 7. Lysosome (phagocytische weisse Blutzelle): Verdauung von Proteinen, Polysaccharide,... 8. In Prokaryonten ist keine Cytoplasmaströmung vorhanden 9. Mikrofilamente und nicht Mikrotubuli sind für die Zell-/Muskelkontraktionen zuständig 10. In Mitochondrien, weil die Zellatmung stattfindet (aus Glucose und Fetten wird mit O2 ATP hergestellt) Kapitel 8, Membranstruktur und Funktion 1. Welche Beobachtungen/Experimente führten zum Modell einer Membranstruktur? Die ersten Modelle des molekularen Aufbaus von Biomembranen entwickelte man schon Jahrzehnte bevor man diese Gebilde in den fünfziger Jahren erstmals am Elektronenmikroskop sehen konnte. Bereits 1895 vermutete C. Overton, Membranen müssten aus Lipiden bestehen; dies schloss er aus seiner Beobachtung, dass fettlösliche (lipophile) Substanzen viel einfacher in Zellen eindringen können als solche, die sich in Fett lösen (lipophobe). Zwanzig Jahre später analysierte man dann Membranen roter Blutzellen und stellte fest, dass diese aus Lipiden und Proteinen zusammengesetzt sind. 2. Welche Beobachtungen/Experimente unterstützen das „Fluid mosaic“Modell einer Membran? 1917 Herstellung von künstlichen Membranen durch I. Langmuir. Dafür löste er Phospholipide in Benzol und gab anschliessend das Gemisch in Wasser. Nach dem Verdunsten des Benzols fand er als Rückstand einen dünnen Film Phospholipide auf der Wasseroberfläche. Er beobachtete dabei, dass nur die hydrophile Köpfe der Phospholipide in Wasser eingetaucht waren. 1925 E. Gorter und F. Grendel meinen erkannt zu haben, dass Zellmembranen in der Tat Lipid-Doppelschichten sind, deren Dicke zwei Moleküle beträgt. Eine solche Doppelschicht könnte eine stabile Abgrenzung zwischen zwei wässrigen Kompartimenten bilden, weil die hydrophoben Schwänze der Phospholipide - durch die Molekülanordnung - gegen das Wasser abgeschirmt sind, während die hydrophilen Köpfe freiliegen. Das Experiment dazu: Gorter und Grendel massen den Phospholipidgehalt der aus den roten Blutzellen isolierten Membran und stellten fest, dass er gerade ausreichte, um die Zelle mit zwei Molekülschichten zu umgeben. 1935 Davson-Danielli-Modell: Die Membran ist wie ein Sandwich aufgebaut, mit einer Lipid-Doppelschicht zwischen zwei Schichten globulärer Proteine. In den 50er Jahren: Betrachtung von Membranen am Elektronenmikroskop Die Hypothese über die Dicke der Membranen wurde bestätigt. Das Davson-Danielli-Modell besitzt 2 Schwachpunkte: Man zweifelt, dass alle Membranen der Zelle gleich aufgebaut sind Lage der Proteine im Sandwichmodell. Membranproteine sind ebenfalls amphipathisch. Wenn sie als durchgehende Schicht auf der Membran liegen würden, käme auch der hydrophobe Bereich mit Wasser in Berührung. 1972 S. J. Singer und G. Nicolson schlagen ein verändertes Membranmodell vor. Die Phospholipid-Doppelschicht ist in diesem Modell nicht von festen Proteinschichten bedeckt, sondern die Membranproteine sind einzeln in die Phospholipid-Doppelschicht eingelagert und ragen nur mit ihren nur mit ihren hydrophilen Bereichen in das umgebende Wasser. Diese Molekülanordnung würde den hydrophilen Regionen von Phospholipiden und Proteinen den grösstmöglichen Kontakt mit dem Wasser gestatten und ihren hydrophoben Bereichen gleichzeitig eine nichtwässrige (lipophile) Umgebung bieten. Membranen als Mosaik aus Proteinmolekülen, die in einer flüssig Doppelschicht aus Phospholipiden liegen. 3. Wie lässt sich die Beweglichkeit von Proteinen innerhalb der Membran experimentell nachweisen? Belege für die Bewegung der Membranproteine: Fusioniert man im Labor eine menschliche Zelle mit einer Mauszelle, sind die Proteine der beiden Arten in der Membran der Hybridzelle nach kaum einer Stunde völlig gemischt. 4. Erkläre die folgenden Begriffe: Turgor, Osmose, semipermeable Membran, Osmoregulation, Plasmolyse Turgor Im Innern einer pflanzlichen Zelle herrscht ein bestimmter Druck (Wanddruck), mit dem der Protoplast an die aus Zellulose bestehende Zellwand gedrückt wird. Für diesen Turgor sind osmotische Vorgänge verantwortlich, indem durch den semipermeablen, wandständigen Protoplasten Wasser in die salz- und zuckerhaltige Vakuolenflüssigkeit diffundiert. Turgeszent nennt man solche Zellen, deren Turgor gross ist. Von deturgeszenten Zellen spricht man dagegen, wenn durch Wasserabwanderung nach aussen der Turgor klein wird. Osmose Ist eine Form der Diffusion. Wasser diffundiert durch die Membran aus einer hypotonischen in eine hypertonische Lösung in dem Bestreben, diese zu Verdünnen. Diese Diffusion von Wasser durch eine selektiv permeable Membran ist ein Sonderfall des passiven Transports. Semipermeable Membran Membran, die für bestimmte, meist kleine Moleküle oder Ionen durchlässig sind, für andere, meist grössere jedoch nicht. Osmoregulation ist die Steuerung des Wasserhaushalts eines Lebewesens. Plasmolyse Die Membran einer Zelle kann als semipermeable Membran, die osmotisch wirksam ist, aufgefasst werden. Legt man Pflanzenzellen ein eine Lösung, deren Konzentration höher ist als die des Vakuolensafts, beginnt Wasser aus der Vakuole nach aussen zu diffundieren. Damit nimmt der Turgor ab, und das Protoplasma beginnt sich von der Zellwand abzulösen. Den Beginn dieses Vorgangs nennt man Grenzplasmolyse, den gesamten Vorgang Plasmolyse. 5.Gemeinsamkeiten und Unterschiede zwischen Transportproteinen und Enzymen Gemeinsamkeiten zwischen Transportproteinen und Enzymen: Gemeinsamkeiten Unterschiede Wie ein Enzym, das für ein Substrat spezifisch ist, so ist auch ein Transportprotein auf eine gelöste Substanz spezialisiert und besitzt eine Bindungsstelle, die dem aktiven Zentrum des Enzyms entspricht. Wie Enzyme, so kann man auch Transportproteine absättigen. Jeder Typ von Transportprotein ist nur mit einer begrenzter Zahl von Kopien in der Plasmamembran vertreten, und wenn alle diese Moleküle ihre Fracht mit der maximal möglichen Geschwindigkeit binden und durch die Membran schläusen, verläuft der Transport mit Maximalgeschwindigkeit. Wie Enzyme können auch Transportproteine durch Moleküle gehemmt werden, die dem normalen Substratmolekül ähneln. In einem solchen Fall konkurriert der Hemmstoff mit dem Substrat um die Bindung am Transportprotein. Anders als Enzyme katalysieren Transportproteine in der Regel keine chemischen Reaktionen, sondern ihre Aufgabe besteht darin, einen physikalischen Vorgang zu beschleunigen: Die Diffusion von Molekülen durch eine Membran, die für die betreffende Substanz ansonsten nahezu undurchlässig wäre. 6. Erkläre den Unterschied zwischen erleichterter Diffusion und aktivem Transport. Erleichterte Diffusion Polare Moleküle und Ionen, die von der Lipid-Doppelschicht gestoppt werden, diffundieren mithilfe kanalbildender Transportproteine durch die Membran. Aktiver Transport ist eine Art des Stoffaustausches. Manche Transportproteine können gelöste Stoffe entgegen deren Konzentrationsgefälle durch die Plasmamembran befördern, also von der Seite, auf der sie weniger konzentriert sind, auf die Seite mit höherer Konzentration. Dieser Transport verläuft somit „bergauf“, gegen das Bestreben der Substanzen, ihrem Konzentrationsgefälle entsprechend zu diffundieren, und deshalb erfordert er Energie. Um eine Substanz gegen deren Konzentrationsgradienten durch die Membran zu pumpen, muss die Zelle Energie aus ihrem Stoffwechsel aufwenden; deshalb bezeichnet man diese Art des Stoffaustausches auch als aktiven Transport. 7. Beschreibe Gemeinsamkeiten und Unterschiede zwischen einer Natrium/Kalium-Pumpe und einer Protonpumpe. Gemeinsamkeiten und Unterschiede zwischen einer Natrium/Kalium-Pumpe und einer Proton-Pumpe. Natrium/KaliumPumpe Proton-Pumpe Gemeinsamkeiten Ist ein Membranprotein, das in tierischen Zellen aktiv Ionen transportiert und dadurch ein Membranpotential erzeugt. Indem diese Arten von Pumpen eine Spannung an Membranen aufbauen, speichern sie Energie, die für die Aktivitäten der Zelle nutzbar gemacht werden kann. Sie transportiert die Natrium- und Kaliumionen nicht im Verhältnis 1:1 für jeweils drei Natriumionen, die sie aus der Zelle hinausbefördert, transportiert sie nur zwei Kaliumionen hinein Es ist die bedeutenste elektrogene Pumpe der Pflanzen, Bakterien und Pilze. Sie befördert aktiv Protonen (Wasserstoffionen) aus der Zelle hinaus. Durch das Pumpen von H+ werden positive Ladungen aus dem Cytoplasma in das die Zelle umgebende Milieu verschoben. Dieser Vorgang führt zur Speicherung von Energie in Form eines Membranpotentials. Es handelt sich um Transportproteine, die eine elektrische Spannung zwischen zwei Membranseiten aufbauen „elektrogene Pumpen“ ( elektrogener Transport) Quizfragen: 1.Wie unterscheiden sich die verschiedenen Membranen eukaryotischer Zellen? Jede Membran hat eine für sie charakteristische Proteinausstattung Zellen haben die Fähigkeit, verschiedene Typen von Nachbarzellen zu unterscheiden. Diese gegenseitige Erkennung erfolgt durch Moleküle (verzweigte Oligosaccharide) auf der Oberfläche der Zellmembran. Oligosaccharide haben je nach Typ oder Zelle unterschiedliche Struktur und unterschiedliche Positionen auf der Zelloberfläche und dienen deshalb gut als Unterscheidungsmerkmal für Zellen. 2. Anordnung der Membranproteine: Die meisten Proteine „driften“ innerhalb der Membran umher (wie Eisberge im Wasser). Sie drehen sich um die Längsachse oder verschieben sich seitwärts. Es gibt zwei Hauptgruppen von Membranproteinen: Integrale Membranproteine (eingelagert in Lipiddoppelschicht); periphere Membranproteine (an die Oberfläche gebunden) 3. Welche Faktoren beeinflussen die Fluidität von Membranen? Temperatur: Die Membran verfestigt sich gelartig bei sinkender Temperatur Lipidzusammensetzung: Je mehr Phospholipide mit ungesättigten Fettsäuren vorhanden sind, desto kälter kann es sein, ohne dass der Festigungspunkt erreicht wird. Cholesterol: Ist ein Steroid, das zwischen den Phospholipidmolekülen der Membran eingelagert wird und zur Stabilisierung der Membranfluidität beiträgt. 4. Passiver Transport: Diffusion (Bsp.: Lunge: Austausch Sauerstoff und Kohlendioxid) Erleichterte Diffusion = mit Hilfe von Carriermolekülen (Bsp.: Transport der Glucose in die Zelle) Osmose (Bsp.: Austausch in den Kapillaren) Filtration (Bsp.: Harnentstehung in den Nieren) Ionentransport entlang eines elektrischen Gradienten (Bsp.: Ruhepotential: Zellinneres negativ, Zelläusseres positiv, Kaliumionenkanäle offen: Kaliumionen bestreben nach aussen zu diffundieren) 5. Wird der extrazelluläre pH erniedrigt (= mehr Wasserstoffionen extrazellulär), wird der Sucrosetransport in die Zelle gefördert: Das Membranprotein Sucrose-WasserstoffionenCotransporter kann Sucrose gegen einen Konzentrationsgradienten transportieren. Dies aber nur, wenn das Sucrose-Molekül zusammen mit einem Wasserstoffion auftritt. 6. Warum tendieren Phospholipide dazu, sich in einer Doppelschicht zu organisieren, wenn sie sich in wässriger Umgebung befinden? Diese Struktur schirmt die hydrophoben „Schwänze“ der Phospholipide vom Wasser ab. Gleichzeitig befinden sich die hydrophilen „Köpfe“ im Wasser. 7. Kohlenwasserstoffe, die an einige der Membranproteine- und lipide gebunden sind, werden zugefügt, wenn die Membran im Golgi-Apparat erneuert wird. Die neue Membran bildet dann Transportvesikel, die zur Zelloberfläche wandern. Auf welcher Seite der Vesikelmembran befinden sich die Kohlenwasserstoffe jetzt? Sie sind auf der Innenseite der Vesikelmembran, denn Golgi-Vesikel werden durch Einschnürung der Membran gebildet. Moleküle, die sich auf der Aussenseite der Membran befanden, sind dann neu auf der Innenseite des Vesikels (und umgekehrt). 8. Das Hormon Epinephrin bewirkt, dass eine Leberzelle sein gespeichertes Glycogen hydrolysiert und Zucker frei wird, ohne dass es dabei selber in die Zelle eindringt. Erklärung: Epinephrin bindet an einen Rezeptor der Leberzelloberfläche. Dadurch wird ein Signal innerhalb der Zelle aktiviert, das zur Zuckerfreigabe führt. 9. Wie hilft das 2. thermodynamische Gesetz, die Diffusion einer Substanz durch eine Membran zu erklären? Das 2. Gesetz besagt, dass eine Tendenz zur Zufälligkeit besteht. Gleiche Konzentrationen einer Substanz auf beiden Seiten einer Membran ist eine zufälligere Verteilung als ungleiche Konzentrationen. Die Diffusion einer Substanz in eine anfangs weniger konzentrierte Region erhöht somit laut dem 2. thermodynamischen Gesetz die Entropie. 10. Erkläre warum es nicht genügt, zu sagen, dass eine Lösung hypoton ist. Hyperton und hypoton sind relative Ausdrücke. Eine Lösung, die hyperton zu Hahnenwasser ist, kann hypoton zu Meerwasser sein. Man muss also immer sagen, mit was die Lösung verglichen wird. 11. In welchem Sinn ist der aktive Transport ein endogener Prozess? Aktiver Transport bedeutet energetisch „aufwärts“ im Sinn, dass Energie aufgewendet werden muss (in Form von ATP). 12. Was ist der Beitrag der Transportproteine zur selektiven Permeabilität einer Membran? Transportproteine sind spezifisch für die Substanz, die sie transportieren. Die Anzahl und der Typ der verschiedenen in der Membran eingebetteten Transportproteine beeinflusst daher die Permeabilität der Membran für verschiedene Substanzen. 13. Wie kann eine proteinausschüttende Zelle ihr Produkt synthetisieren und ausschütten, ohne dass das Protein eine Membran passieren muss? Vom Zeitpunkt weg, wo das Protein vom rauen ER gebildet wird, ist es eigentlich schon „ausserhalb“ der Zelle: zuerst im Innern des ER, dann im Golgi-Apparat und den Transportvesikeln und zum Schluss ausserhalb der Plasmamembran, nachdem der Vesikel seinen Inhalt durch Exocytose abgegeben hat. 14. Eine künstliche Zelle, die 0.03M Sucrose und 0.02M Glucose in wässriger Lösung enthält und durch eine semipermeable Membran abgeschlossen wird, gibt man in ein Becherglas, dessen wässrige Lösung jeweils 0.01M Sucrose, Glucose und Fructose enthält. Die Membran ist permeabel für Wasser und die Einfachzucker (Glucose und Fructose) aber impermeabel für das Disaccharid (Sucrose). Welches Solvans wird in die Zelle diffundieren? Fructose 15. Welches Solvans wird aus der Zelle diffundieren? Glucose 16. Welche Lösung – die, des Zellinnern oder die, des Zelläussern – ist hyperton zur anderen? Die, des Zellinnern 17. In welche Richtung wird eine osmotische Bewegung des Wassers erfolgen? In das Zellinnere 18. Wird die Zelle im Becherglas sich füllen oder sich entleeren? Sie wird sich mehr füllen. Kapitel 12, der Zellzyklus 1. Beschreiben sie den Zellzyklus, nenne die wichtigsten Schritte vor, während und nach der Mitose. Nenne Gemeinsamkeiten und Unterschiede bei tierischen und pflanzlichen Zellen. Interphase: - G1: Zelle wächst - S: Zelle wächst, Verdoppelung der Chromosomen - G2: Zelle wächst, Vorbereitung auf Zellteilung -Tiere: 2 Centromere mit 2 Centriolen-Paaren -Pflanzen: 2 Centromere ohne Centriolen Mitotische Phase: - Prophase: -im Nucleus: - DNA windet sich spiralförmig auf - X-förmige 2-Chromatiden-Chromosomen bilden sich - Kernmembran/-körperchen lösen sich auf - im Cytoplasma: - Spindelfasern bilden sich - Prometaphase . Mikrotubuli binden an die Kinetochoren an der CentromerRegion jedes Chromosoms - Metaphase: - Spindelfasern sind komplett ausgebildet - Chromosomen ordnen sich in der Äquatorialebene an - Anaphase: - Chromosomen trennen sich -> es entstehen 2 identische 1-Chromatiden-Chromosomen - 1-Chromatiden-Chromosomen werden von Spindelfasern zu den Zellpolen gezogen - Telophase: - DNA der Chromatiden entspiralisiert sich - Kernmembran wird gebildet - Spindelapparat bildet sich zurück - Cytokinase: Zellplasma teilt sich -Tiere. Teilung erfolgt durch Bildung einer Furche -Pflanzen: es wächst eine neue Membran im Bereich der Äquatorialebene 2. Welche experimentellen Befunde sprechen für ein Zell-ZyklusKontrollsystem? Kultivierte Säugetierzellen können dazu gebracht werden, zu fusionieren -> Bildung 1 Zelle mit zwei Kernen Das Resultat der Verschmelzung zweier Zellen in verschiedenen Phasen des Zellzyklus deutet darauf hin, dass chemische Stoffe den Verlauf der Phase kontrollieren. Bsp: Wenn eine Zelle in M-Phase mit einer Zelle in G1-Phase verschmolzen wird, beginnt die Zelle in der G1-Phase sofort mit der Mitose. 3. G1, G2, Go, S, M: Nenne Unterschiede und die Bedeutung dieser Begriffe für den Ablauf des Zellzyklus G1: Wichtigster Kontrollpunkt: startet den Prozess: Wachstum, Vorbereitung auf S-Phase S: Wachstum und Verdopplung der DNA: Sehr wichtig (ohne S-Phase hätten die Tochterzellen nur die Hälfte der Chromosomen nach der Mitose) Fehler in dieser Phase sind fatal! G2: Kontrollpunkt: Verhindert Mitose im Falle eines Fehlers (z.B wenn S nicht funktioniert hat) Wachstum, Vorbereitung auf Zellteilung G0: Wenn die Zelle kein „grünes Licht“ vom G1-Kontrollpunkt erhalten hat, endet hier der Zyklus und wechselt in den sich nicht teilenden Zustand. Dies verhindert Fehler. M: Kontrollpunkt, Zellteilung: Sehr schwer, da beide Tochterzellen die genau gleichen Bestandteile brauchen 4. Was sind Wachstumsfaktoren (Growth factors) und welche physiologische Bedeutung haben sie? Externe Signale, die helfen den Zellzyklus zu regulieren Definition: Ein Protein, dass in der extrazellulären Umgebung der wachsenden Zelle ist und andere Zellen zur Teilung stimuliert Bsp: Platelet-derived growth factors (PDGF) Von Blutzellen produziert; verantwortlich für die Teilung von Blutplättchen (Wundverschluss) 5. Wodurch unterscheiden sich Krebszellen von normalen Zellen? - Stören Zellzyklus: - keine Wachstumsfaktoren - keine Kontrollpunkte - kein Endpunkt - Hören nicht auf, sich zu teilen, wenn Wachstumsfaktoren nicht mehr da sind (brauchen diese nicht oder stellen sie selber her) - Müssen nicht an Substrat angemacht sein: Sie teilen sich schnell ->bilden Klumpen sich überlappender Zellen - Stoppen die Teilung an zufällig ausgewählten Punkten im Zellzyklus - Können sich unendlich oft teilen (normale Zelle altern und sterben) - Weichen dem Immunsystem aus - können Tumore bilden: Bösartig: dringt in Nachbarzellen ein, verteilt sich durch Lymph- und Blutgefässe in anderen Teilen des Körpers (Metastasen) Challenge-Fragen: a) 20 b) Interphase:16h, Metaphase: 1h c) G1 ist 50% der Interphase= 8h S ist 30% der Interphase = 288min G2 ist 20% der Interphase =192min Kapitel 13, Meiose und sexuelle Lebenszyklen 1. Beschreibe die Lebenszyklen eines Haplonten, Diplonten, Haplodiplonten (alternativer Generationswechsel) Der Wechsel zwischen Meiose und Befruchtung findet bei allen sich sexuell fortpflanzenden Organismen statt, er unterscheidet sich aber im Zeitablauf der Ereignisse von Art zu Art. Lebenszyklus eine DIPLONTEN Dieser Zyklus ist für Menschen, die meisten Tiere und auch für manche niedere Pflanzen und Protisten charakteristisch. Bei Diplonten sind die Gameten die einzigen haploiden Zellen. Die Meiose findet während der Bildung der Gameten statt, die sich dann bis zur Befruchtung nicht mehr teilen. Die nach der Befruchtung entstandene diploide Zygote teilt sich mitotisch und entwickelt sich zu einem vielzelligen diploiden Organismus; zu eine DIPLONTEN. Lebenszyklus eines HAPLONTEN Dieser Zyklus ist für viele Pilze, niedere Pflanzen und ein paar Algen charakteristisch. Nach der Fusion der Gameten (Befruchtung) bildet sich eine diploide Zygote und es findet sofort eine Meiose statt. Es entstehen haploide Zellen, die sich mitotisch teilen und einen vielzelligen haploiden Organismus bilden; einen HAPLONTEN. Dieser bildet seine Gameten durch Mitosen, nicht durch Meiosen!! Bei den Haplonten ist das einzig diploide Stadium die Zygote. Die Entwicklungszyklen der Diplonten und der Haplonten nennt man einfache Generationsfolgen. Lebenszyklus eines HAPLODIPLONTEN Dieser Entwicklungszyklus nennt man Generationswechsel, oder genauer diplohaploider Generationswechsel. In diesem Entwicklungszyklus gibt es sowohl diploide als auch haploide vielzellige Stadien. Das diploide vielzellige Stadium nennt man Sporophyt (2n). Im Sporophyten entstehen durch Meiose haploide Zellen, die Sporen. Im Gegensatz zu den Gameten entwickelt sich eine Spore zu einem vielzelligen Individuum ohne dass sie vorher mit einer anderen Zelle fusioniert. Die Spore teilt sich mitotisch und bildet ein vielzelliges haploides Stadium, den Gametophyten (n). Im Gametophyten entstehen durch Mitose Gameten. Das Ergebnis der Befruchtung ist eine diploide Zygote, die sich dann durch Mitosen zum Sporophyten der nächsten Generation entwickelt. Bei diesem Zyklus geht der Sporophyt aus dem Gametophyt und der Gametophyt aus dem Sporophyt hervor der Organismus ist ein Haplo-Diplont! 2. Karyotyp, homologe Chromosomen, Autosomen, Sexchromosomen: Definiere diese Begriffe und grenze sie gegeneinander ab. KARYOTYP Jede somatische Zelle (das sind alle Zellen ausser Spermien und Eier) besitzen 46 Chromosomen. Diese kann man an ihrer Grösse und der Lage ihrer Centromere unterscheiden. Auch kann man sie daran unterscheiden, dass jedes Chromosom nach Behandlung mit bestimmten Farbstoffen ein typisches Bandenmuster zeigt. Die 46 menschlichen Chromosomen liegen paarweise vor. Wenn man die 46 Chromosomen nach Grösse und Gestalt zu Paaren anordnet, bekommt man eine Darstellung, die man Karyotyp nennt. HOMOLOGE CHROMOSOMEN Chromosomen, die ein Paar bilden, also dieselbe Länge, dieselbe Centromerposition und dasselbe Bandenmuster besitzen, nennt man homologe Chromosomen. Diese zwei Chromosomen eines Paares tragen Gene für dieselben Erbmerkmale (Z.B. Das Gen für die Augenfarbe liegt auf beiden Chromosomen am gleichen Ort) SEXCHROMOSOMEN/GESCHLECHTSCHROMOSOMEN Es gibt eine Ausnahme der Regel der homologen Chromosomen bei menschlichen Somazellen: Die X und Y Chromosomen. Frauen besitzen zwei X-Chromosomen (XX) und Männer ein Y und ein X Chromosom (XY). Da diese Chromosomen geschlechtsbestimmend sind, nennt man X- und Y-Chromosomen auch Geschlechtschromosomen. Alle anderen Chromosomen nennt man AUTOSOMEN. 3. Beschreibe die Unterschiede zwischen einer mitotischen und einer meiotischen Teilung, Zu welchen unterschiedlichen Resultaten führen die beiden Teilungswege? Campbell englisch: p.242 Campbell deutsch: p.24 Die Mitose unterscheidet sich als erstes in der Zahl der Zellteilungen von der Meiose. In der Mitose gibt es eine Zellteilung, die aus Prophase, Metaphase, Anaphase und Telophase besteht. Die Meiose besteht aus zwei Zellteilungen, die ebenfalls aus Prophase, Metaphase, Anaphase und Telophase bestehen. In der Prophase I der Meiose beginnen sich die Chromosomen zu verdichten und die homologen Chromosomen finden sich zu Paaren zusammen (Synapsis). Jedes Chromosomenpaar ist als Tetrade angeordnet, die aus vier parallel angeordneten Chromatiden besteht. An mehreren Stellen haben sich die Chromatide homologer Chromosomen gekreuzt; das Crossing over findet statt. Diesen Vorgang findet man nur in der Meiose. Das Produkt der Mitose sind zwei Tochterzellen, die beide diploid (2n) sind und genetisch identisch mit der Mutterzelle. Das Produkt der Meiose sind vier Tochterzellen. Jede besitzt halb so viele Chromosomen (n) wie die Mutterzelle. Die Tochterzellen sind genetisch weder mit der Mutterzelle noch untereinander identisch. (Dies deshalb, weil sich in der Anaphase I homologe Chromosomen trennen und in der Anaphase II dann die Schwesterchromatide) Die Bedeutung der Mitose ist die Entwicklung eines vielzelligen Organismus aus einer Zygote, die Produktion der Zellen für das Wachstum und die Gewebeheilung. Die Bedeutung der Meiose ist die Produktion der Gameten, die Reduktion der Chromosomenzahl auf die Hälfte und sie sorgt auch für die genetische Variabilität der Gameten. 4. Welche drei Faktoren tragen zur genetischen Variabilität bei der geschlechtlichen Fortpflanzung bei? FREIE REKOMBINATION In der Metaphase der Meiose I liegen die homologen Chromosomen gepaart in der Metaphaseplatte. Die Orientierung in Bezug auf die Pole ist zufällig, es bestehen für jedes Paar zwei Möglichkeiten. Für die Tochterzelle besteht also eine Chance von 50 % das mütterliche oder das väterliche Chromosom eines homologen Chromosomenpaares zu bekommen. Jeder Gamet repräsentiert also eine von allen möglichen Rekombinationen. Die Zahl der möglichen Chromosomenkombinationen in den Gameten durch freie Kombination ist 2n , wobei n die haploide Chromosomenzahl darstellt. Die Anzahl der möglichen Rekombinationen der mütterlichen und väterlichen Chromosomen in den Gameten beim Menschen (n=23) beträgt 223 also ungefähr 8 Millionen!! CROSSING OVER Aufgrund der freien Rekombination der Chromosomen bei der Meiose produziert jeder von uns Gameten mit unterschiedlichen Kombinationen von Chromosomen beider Eltern. Bis jetzt haben wir angenommen, dass jedes einzelne Chromosom eines Gameten nur mütterliches oder väterliches Genmaterial enthält. Dem ist aber nicht so. In den Chromosomen sind Gene mütterlichen und väterlichen Ursprungs kombiniert; dies geschieht durch Crossing over, welches in der Prophase der Meiose I stattfindet. Hier paaren sich homologe Chromosomen (Synapsis). Diese Paarung ist sehr präzise, homologe Gene liegen genau Seite an Seite. Das Crossing over findet statt, wenn sich homologe Anschnitte zweier NICHTSChwesterchromatide berühren und Genmaterial austauschen. Dies findet beim Mensch im Durchschnitt 2-3 Mal pro Chromosomenpaar statt. ZUFÄLLIGKEIT DER BEFRUCHTUNG Ein weibliches Ei enthält eine von 223 möglichen Kombinationen von Chromosomen und wird von einen Spermium befruchtet, das ebenfalls eine von 223 möglichen Kombinationen von Chromosomen enthält. Es entsteht eine Zygote, die eine von 223 x 223 (64 Billionen!) möglichen diploider Rekombinationen darstellt. Dies alles ohne das Crossing over miteinzubeziehen!! Kapitel 14, Mendel und die Genidee 1. Warum eigneten sich Erbsen besonders gut für Mendels Erbversuche? Erbsen sind in vielen Variationen erhältlich: Violette, weisse Blüten, grüne, gelbe Bohnen usw., des weiteren ist es einfach sicher zu stellen, dass nur die Individuen miteinander gekreuzt werden, die er kreuzen will: Erbsen sind Selbstbestäber, das heisst sie sind zwittrig und geben ihre Pollen auf ihre eigene Narbe. Mendel hat nun die Staubblätter entfernt, bevor sie Pollen produziert haben und die Pollen einer anderen Pflanze auf diese „einsame“ Narbe gebracht. Ein bisschen Glück war natürlich auch dabei, hätte er nicht Merkmale genommen die dominant/rezessiv sind, sondern intermediär, hätte er die Gestze wohl nie gefunden. i. ii. die Generationszeit ist relativ kurz relativ viele Nachkommen iii. iv. v. vi. es lassen sich mit relativ geringem Aufwand grosse Mengen an Versuchspflanzen züchten. es gibt viele verschiedene Erbsensorten (purpurfarbene/weisse Blüten, runde/runzelig Samen, gelbe/grüne Samen....) mit Erbsen kann man gezielte Kreuzungsexperimente machen man kann die Befruchtung kontrollieren (Fremdbefruchtung) Mendel verliess sich bei seinen Experimenten immer nur auf eindeutige Merkmale (entweder oder) und begann seine Züchtungsexperimente immer mit reinerbigen Erbsenrassen. 2. Die vier Mendelschen Aussagen zur Wirkungsweise eines Gens 1. Die verschiedenen Allele eines Gens sind die Erklärung für die Variationen in vererbten Eigenschaften. 2. Für jede Eigenschaft erbt der Organismus zwei Allele, eines von jedem Elternteil. 3. Sind die zwei Allele verschieden, zeigt sich eines, das Dominante vollständig in der Erscheinung des Organismus, während das andere, rezessive vollständig verschwindet (phänotypisch) 4. Die zwei Allele für jede Eigenschaft trennen sich während der Gametebildung. 3. Beschreibe die drei Mendelschen Erbregeln. Welche experimentellen Voraussetzungen waren notwendig, um diese Regeln zu finden? Welche dieser drei Regeln musste nachfolgend korrigiert werden? vii. viii. ix. x. xi. Alternative Zustandsformen (Allele) eines Gens bedingen die genetische Variabilität bei Erbmerkmalen. z.B. traten zwei Versionen von Blütenfarben auf, eine violette und eine weisse. Solche alternative Versionen werden als Allele eines Gens bezeichnet. Für jedes Merkmal besitzt ein Organismus zwei Allele, je eines von jedem Elternteil. Wenn die beiden Allele unterschiedlich sind, dann wird eines, und zwar das dominante Allel voll exprimiert; das andere, das rezessive Allel, zeigt keinerlei Ausprägung. Man spricht vom dominant rezessiven Erbgang. Eine Eizelle oder eine Spermazelle erhält nur eines der beiden Allele, die in zwei Kopien in den Somazellen des betroffenen Organismus vorliegen. Auf die Chromosomen bezogen entspricht diese Trennung oder Segregation der Reduktion der Chromosomenzahl vom diploiden zum haploiden Satz während der Meiose. 1.°Mendel’sche Regel (Uniformitätsregel, Reziprozitätsregel): In seinen ersten Experimenten kreuzte Mendel reinerbige Erbsenlinien, die sich in einem Merkmal unterschieden, z. B. große und zwergwüchsige Linien. Als Nachkommen erhielt er Hybride, die keine Mischung beider Eigenschaften aufwiesen, sondern äußerlich dem großwüchsigen Elternteil entsprachen. Als Erklärung postulierte er Erbeinheiten, die wir heute Gene nennen und die häufig in unterschiedlichen Zustandsformen (Allelen) auftreten. Man unterscheidet dominante (A) und rezessive (a) Zustandsformen eines Gens, wobei das dominante Allel die Wirkung des rezessiven Allels unterdrückt und äußerlich in Erscheinung tritt. Mendel erkannte, dass Gene in normalen Körperzellen gewöhnlich paarweise vorkommen, sich aber bei der Entstehung der Geschlechtszellen (Ei- und Samenzellen) aufteilen. Jedes Gen aus einem solchen Paar gelangt dabei in eine andere Geschlechtszelle. Bei der Vereinigung von Ei- und Samenzelle entsteht wieder ein Genpaar, in dem das dominante Allel (in dem genannten Fall für die Großwüchsigkeit) die Wirkung des rezessiven (für Zwergwuchs) überdeckt. Diese Ergebnisse liefern die Grundlage für die 1. Mendel’sche Regel, nach der eine Kreuzung zweier reinerbiger Eltern, die sich in einem oder mehreren Merkmalen unterscheiden, eine gleichförmige (uniforme), mischerbige (Aa) Tochtergeneration hervorbringt. Die Uniformität der Tochtergeneration wird nicht beeinflusst, wenn der jeweils andere Elter das betreffende Merkmal aufweist (reziproke Kreuzung). 2.°Mendel’sche Regel (Spaltungsregel, Dominanzregel): Um zu beweisen, dass es solche Erbeinheiten gibt, kreuzte Mendel die erste Generation der großwüchsigen Hybriderbsen (Aa×Aa) untereinander. Wie sich dabei herausstellte, tauchten in der ersten Tochtergeneration wieder kleinwüchsige Erbsenpflanzen (aa) auf, und zwar kleinwüchsige und großwüchsige im Verhältnis eins zu drei. Daraus zog er den Schluss, dass sich die Gene zu den Paaren AA, Aa und aa zusammengefunden hatten. Wie er bei weiteren Kreuzungsexperimenten feststellte, gingen aus den reinerbigen AA-Pflanzen bei Selbstbestäubung nur große Nachkommen hervor, und die Nachkommen der aa-Exemplare waren stets klein. Bei der Kreuzung der Aa-Hybride fand sich unter den Nachkommen wieder das gleiche Zahlenverhältnis von 3 : 1. Aufgrund dieser Versuchsergebnisse beschrieb Mendel die 2. Mendel’sche Regel, nach der die Nachkommen einer Kreuzung mischerbiger Individuen nicht mehr gleichförmig sind, sondern ihr äußeres Erscheinungsbild in einem bestimmten Zahlenverhältnis aufspalten. Dieses Zahlenverhältnis wird sowohl durch die Anzahl der Merkmale (Genorte), in denen sich die Eltern unterscheiden, als auch durch den Erbgang beeinflusst. Man unterscheidet einen dominant-rezessiven Erbgang (das dominante Allel unterdrückt die Wirkung des rezessiven) von einem intermediären Erbgang (die Wirkung beider Allele ist erkennbar; ein mischerbiges Individuum nimmt eine mittlere Erscheinungsform an). Bei einem dominant-rezessiven Erbgang spaltet sich das äußere Erscheinungsbild der Tochtergeneration im Verhältnis 3 : 1 auf, wenn nur ein Merkmal betrachtet wird, sowie bei einem intermediären Erbgang im Verhältnis 1 : 2 : 1. 3. Mendel’sche Regel (Regel von der unabhängigen Aufspaltung der Allelenpaare): Wie weitere Kreuzungsexperimente mit Elterngenerationen zeigten, die sich in zwei oder mehreren Merkmalen unterschieden, werden die einzelnen Genorte und damit die Merkmalsausprägungen unabhängig voneinander weitergegeben und sind frei miteinander kombinierbar. Allerdings gilt die 3. Mendel’sche Regel nur für Gene, die auf verschiedenen Chromosomen liegen. Zufälligerweise waren die sieben Merkmale der Erbsenpflanzen, die Mendel untersuchte, auf verschiedenen Chromosomen lokalisiert. Ansonsten hätte er keine statistische Verteilung der Merkmalskombinationen erhalten. Die Mendel’schen Regeln wurden zur theoretischen Grundlage der modernen Genetik. Die letzte der drei Regeln musste nachfolgend korrigiert werden, was aber in der Beschreibung oben schon gesagt wurde: Die Regel gilt nur für Merkmale welche auf verschiedenen Chromosomen liegen, das hat Mendel noch nicht gewusst. Er hatte Glück, dass seine Merkmale auf verschiedenen Chromosomen lagen. Voraussetzungen um diese Regeln zu finden: Experimente mit reinerbigen Rassen beginnen sich nur auf eindeutige Merkmale verlassen (entweder oder) Vererbung einer Merkmalsform über drei aufeinanderfolgende Generationen untersuchen: P-, F1- und F2-Generation (hätte Mendel seine Experimente nach der F1-Generation abgebrochen, wären ihm die grundlegenden Gesetzesmässigkeiten verborgen geblieben.). 4. Was ist ein dominantes Allel (Dominanz, Codominanz, Intermediär)? Erkläre am Beispiel der Blutgruppen 0, A, B, AB Blutgruppe 0 ist ein rezessives Allel, während A und B gegenüber 0 dominant sind, das heisst, wenn ein Organismus von einem Elter das Allel 0 und vom anderen das Allel A (oder B) erbt, hat der Organismus Blutgruppe A (oder B). Bekommt ein Fötus aber vom Vater ein Allel A (oder B) und von der Mutter ein Allel b (oder A), dann hat das Kind Blutgruppe AB, das heisst, es prägen sich beide Allele unabhängig voneinander aus. Das ist Codominanz. Bei den Blutgruppen gibt es keinen intermediären Typen, es gibt weder eine Mischung aus 0 und A (oder B), also weniger Anti-B(oder A)-Körperchen, noch eine Mischung aus A und B. 5. Durch welche Besonderheiten zeichnet sich ein quantitatives Merkmal aus? Wie lässt sich der Einfluss der Gene nachweisen? Beschreibe den Erbgang Ein quantitatives Merkmal zeichnet sich dadurch aus, dass es nicht nur von einem Gen, sondern von mehreren Genen beeinflusst wird. Zum Beispiel die menschliche Hautfarbe. ? Die menschliche Hautfarbe wird (der Einfachheit halber) von drei Genen beeinflusst: A, B, C. A, B und C stehen für dunkle Hautfarbe, a, b und c für helle. Ein Mensch mit Genotyp AABBCC hätte also sehr dunkle Hautfarbe, während einer mit Genotyp aabbcc sehr helle Hautfarbe hat. Dazwischen liegen AaBbCc (gleiche Hautfarbe wie AABbcc) und AABbCc (gleiche Hautfarbe wie AABBcc) usw. i. Dominantes Allel: Allel, dessen Merkmal ausgeprägt wird, wenn ein zweites anderes Allel vorhanden ist. (Ein rezessives Allel wird nicht ausgeprägt, wenn ein dominantes Allel vorhanden ist, sondern nur wenn zwei rezessive Allele aufeinander treffen.) ii. Kodominanz: Beide Merkmale werden nebeneinander auf dem selben Organismus ausgeprägt. iii. Intermediärer Erbgang: Die F1-Hybriden zeigen ein Erscheinungsbild, das zwischen den beiden elterlichen Phänotypen liegt. Blutgruppen: Die meisten Gene kommen in mehr als zwei allelen Formen vor. Das AB0Blutgruppensystem des Menschen ist ein Beispiel für multiple Allele eines einzigen Gens. Es gibt vier Phänotypen für dieses Merkmal. Es existieren drei Allele des Ab0-Blutgruppensystems. Weil jedes Individuum zwei Allele trägt, gibt es sechs versch. Genotypen. Die Allele IA (für die Ausbildung des Kohlenhydrats A) und IB (für die Ausbildung des Kohlenhydrats B) sind dominant über das Allel i (führt weder zur Bildung von A noch B). IAIA, IAi IBIB, IBi Blutgruppe A Blutgruppe B Rezessive Homozygote, ii, produzieren weder das Molekül A noch das Molekül B und besitzen die Blutgruppe 0. ii Blutgruppe 0 Die Allele IA und IB sind kodominant. In einem heterozygoten Individuum des Genotyps IAIB werden beide Allele exprimiert, dies führt zur Ausbildung der Blutgruppe AB. IA IB Blutgruppe AB 5. quantitatives Merkmal Es gibt viele Merkmale (z.B. Körpergrösse, Hautfarbe), die denen eine EntwederOder-Klassifizierung nicht möglich ist, weil die Merkmale in einer Population ein Kontinuum bilden. Solche Merkmale werden als quantitative Merkmale bezeichnet. Variabilität in der Quantität deutet in der Regel auf polygene Vererbung hin, d.h. zwei oder mehr Gene wirken zusammen um einen Phänotyp hervorzubringen (Gegenteil von Pleiotropie (eine Reihe von phänotypischen Erscheinungen werden durch ein einzelnes Gen beeinflusst)). Beispiel: Die Hautpigmentierung beim Menschen wird von mind. Drei unabhängigen Genen kontrolliert. AaBbCc AaBbCc aabbcc Aabbcc AaBbcc AaBbCc AABbCc AABBCc AABBCC Kapitel 15, Die chromosomale Basis der Vererbung 1. Was sind die Grundlagen für die Chromosomentheorie der Vererbung? Wie verhält sich diese Theorie zu den von Mendel gefundenen Erbgesetzen? Welche Beobachtung gibt es, die nicht mit den Mendelschen Erbgesetzen erklärt werden können? Die Grundlagen der Chromosomentheorie sind einerseits die Entdeckung/Beobachtung der Mitose und Meiose, die man mit verbesserten Mikroskopen Ende des 19.Jh langsam zu verstehen begann, und andererseits die Gesetze von Mendel. Sie entstand und wurde weiterentwickelt, als die Biologen die Parallelen zwischen dem Verhalten der Chromosomen und Mendels Erbfaktoren erkannten: Chromosomen und Gene kommen in diploiden Zellen beide als Paare vor, homologe Chromosomen und Allele trennen sich während der Meiose und die Befruchtung führt wieder zu gepaarten Chromosomen und Genen. -> Gene haben bestimmte Plätze auf den Chromosomen und diese werden getrennt und unabhängig verteilt. Die Chromosomen ordnen sich zufällig in der Methaphaseplatte der Meiose 1 an -> Unabhängigkeitsregel Eine weitere Grundlage sind die Ergebnisse von Morgans Versuchen: Lokalisierung eines Gens auf einem Geschlechtschromosom (Versuch: Drosophila, Männchen mit weissen Augen) ->geschlechtsgebundene Gene Bestimmte Gene werden in Kombination weitergegeben. (Versuch Drosophila, Rückkreuzung, nicht erwartete Aufspaltung 1:1:1:1, sondern parentale Nachkommen vermehrt, aber auch Rekombinanten kamen vor) ->gekoppelte Gene, liegen auf gleichem Chromosom, Kopplung aber nicht absolut -> Crossing-Over. Die Choromosentheorie verbindet/kombiniert sie mit den Beobachtungen der Meiose/Mitose. Geschlechtsgebundene Gene und gekoppelte Gene können mit den Mendelschen Gesetzen nicht erklärt werden. (Ausserdem auch plasmatische Erbfaktoren). 2. Zwei genetische Loci haben auf einer Koppelungsgruppe einen Abstand von 110 Centimorgan. Wie kann man beweisen, dass diese Gene auf einem und nicht auf zwei Chromosomen lokalisiert sind? 2 Loci, die 50 Centimorgan entfernt sind, haben eine Wahrscheinlichkeit von 50%, dass zwischen ihnen eine ungerade Zahl an Crossing-overs stattfindet. Das ist derselbe Wert, wie wenn sie auf zwei verschiedenen Chromosomen wären. Somit kann nicht direkt gesagt werden, ob Loci, die 50 oder mehr Centimorgan voneinander entfernt sind, auf demselben Chromosom liegen. Erstellt man aber mit Hilfe von Rekombinationsdaten mit anderen Genen eine Kopplungskarte, kann man beweisen, dass die Loci auf dem gleichen Chromosom liegen. Die 110 Centiomorgan ergeben sich dann durch Addieren der einzelnen Centimorganabstände (zwischen Genen, die weniger als 50cM voneinander entfernt und somit auf dem gleichen Chromosom liegen). Der Beweis erfolgt also schrittweise, liegen sowohl A und B als auch B und C auf dem gleichen Chromosom, dann liegen auch die Gene A und C auf dem gleichen Chromosom. 3. Wie unterscheidet sich der Erbgang eines Gens, das auf einem Geschlechtschromosom liegt (XY-Chromosom), von dem eines normalen Gens? Die Gene auf dem Y-Chromosom können nur von Vater zu Sohn weitergegeben werden. Die Gene auf dem X-Chromosom werden vom Vater nur an die Tochter, von der Mutter an Tochter und Sohn weitergegeben. Bei XY-Chromosomen ist nur teilweise ein crossing-over möglich. Bei rezessiven Allelen hat eine Frau nur den entsprechenden Phänotyp, wenn sie homozygot ist. Bei einem Mann spricht man nicht von homo- oder heterozygot, sondern hemizygot, bei ihm wird jedes rezessive Allel, das er von seiner Mutter erbt, exprimiert. Deshalb sind bei Männern geschlechtsgebundene Krankheiten viel häufiger als bei Frauen. 4. Wie lassen sich plasmatische Erbfaktoren von „mendelnden“ Genen unterscheiden? Plasmatische Erbfaktoren gibt es, da Mitochondrien und bei den Pflanzen zusätzlich Plastide und Chloroplasten eigene Gene besitzen (extrachromosomale Gene) . Diese Gene werden den Tochterorganellen weitergeben. Sowohl bei Pflanzen als auch bei Säugetieren werden Zellorganellen von der Mutter/mütterlichen Pflanze weitergegeben, da bei beiden die Zygote das Zytoplasma und somit die Organellen nur von der Eizelle erhält (nicht vom Sperma/Pollen) . Plasmatische Erbfaktoren weisen eine sogenannte maternale Vererbung auf und lassen sich dadurch von den „mendelnden“ Genen unterscheiden. Bsp: gelbe Flecken auf einer sonst grünen Pflanzen hängt nur von der „Mutter“ (nicht vom Pollen) ab, denn die Gene dafür liegen auf Plastiden. Genetische Probleme (S.276, 277) Nr. 1, 2, 3, 4, 5, 7, 10, 12 Kapitel 16, die molekulare Basis der Vererbung 1. mit welchen Experimenten liess sich die stoffliche Identität von Genen aufklären? - - - Die Eigenschaft der DNA eines pathogenen Bakterienstammes, harmlose Bakterien in Krankheitserreger umzuwandeln, lieferte den ersten Hinweis, dass DNA die Erbsubstanz ist. Die Befähigung von Phagen, Bakterienzellen durch Injektion ihrer DNA zur Produktion von Phagen umzuprogrammieren, war ein weiterer Beweis für die DNA als genetisches Material. Auch die Korrelation des DNA –Gehalts einer Zelle mit der Replikation der Chromosomen vor der Zellteilung sprach für die Rolle der DNA als Erbmaterials 2. Der experimentelle Nachweis der semikonservativen DANN-Replikation, Meselson- Strahl-Experiment Das Meselson- Strahl-Experiment testete drei Modelle von DNA Replikation. Meselson und Stahl züchteten E . coli Bakterien während mehreren Generationen in einem schweren Medium mit einem schweren Stickstoffisotop, 15N. Die Bakterien bauten den schweren Stickstoff in ihre Nucleotide und in ihre DNA. Die Wissenschaftler transferierten dann die Bakterien in ein Medium mit 14N, den leichteren, gewöhnlicheren Stickstoffisotop. Jede neue DNA, die die Bakterien synthetisierten würde leichter sein als die “alte” DNA, welche im 15N Medium synthetisiert wurde. Meselson und Stahl konnten die DNA an der unterschiedlichen Dichte beim Zentrifugieren von DNA Extrakten der Bakterien unterscheiden. Die Zentrifugenröhrchen in dieser Zeichnung zeigen das Ergebnis, welches von jedem der drei Modelle vorhergesagt wurde. Die erste Replikation im 14N Medium produzierte ein Band von Hybrid (15N-14N) DNA. Dieses Ergebnis eliminierte das konservative Model. Eine zweite Replikation produzierte beides, ein leichtes Band und ein HybridDNA, ein Ergebnis, welches das dispersive Model eliminiert und das semikonservative unterstützt. 3. Beschreibe den Prozess der DNA-Replikation: Welche Rolle spielen DNA-Polymerase, Ligase, Primase, Helicase, Single-strand binding proteins, Okazaki-Fragmente? 4. Welche Aufgaben haben Telomere? Das Replikationsende- Problem. Wenn ein lineares DNA- Molekül repliziert, entsteht eine Lücke am 5’- Ende bei jedem neuen Strang, da die DNA Polymerase nur Nucleotide an das 3’-Ende anhängen kann. Als Folge wird das DNA Molekül mit jeder Replikation kürzer. Prokaryoten verhindern dieses Problem, indem sie kreisförmige DNA- Moleküle haben, aber was machen Eukaryoten? Eukaryotisch- chromosomale DNA- Moleküle haben spezielle Nucleotid- Sequenzen, Telomere genannt, an ihren Enden. Telomere enthalten keine Gene; stattdessen besteht die DNA aus vielen Wiederholungen einer kurzen Nucleotid- Sequenz. Im menschlichen Körper ist dies die TTAGGG – Sequenz. Somit wird verhindert, das die Gene weggefressen werden. Kapitel 17, Vom Gen zum Protein 1. Beschreibe Experimente, mit denen die „Ein Gen – ein Enzym“Hypothese von Beadle und Tatum unterstützt wurde. In den 30er Jahren untersuchten Beadle und Ephrussi verschiedene Mutationen welche einen Einfluss auf die Augenfarbe der Drosophila haben. Sie stellten dann die Spekulation auf, dass verschiedene Mutationen die Pigmentsynthese störten, indem sie die Produktion von Enzymen verhinderten, welche bei der Pignmentsynthese beteiligt sind. „Auxotrophs“ sind Mutanten welche unfähig sind mittels der normalen Minimalernährung zu entwickeln. Durch Zugabe von Arginin konnte das normale Wachstum erreicht werden, je nach Zugabe der Vorgänger-Aminsosäuren (Citrullin, Lysin) konnte Arginin synthetisiert werden oder nicht; daraus schlossen Beadle und Tatum, dass unterschiedliche Enzyme blockiert werden, welche bei der Synthese des Arginins aus Vorläufern notwendig sind und dass je eine Mutation eines betroffenen Genes zum Ausfall des entsprechenden Enzyms führen musste. Beadle und Tatum experimentierten mit verschiedenen Stoffwechselmutanten des roten Brotschimmels Neurospora crassa. Der Wildtypstamm benötigt zum Wachsen nur ein Minimalmedium aus Saccharose, anorganischen Salzen u. einem Vitamin. Sie verteilten Pilzgeflecht auf verschiedene Gefässe, welches je Minimalmedium und ein verschiedenen zusätzlichen Stoff enthielt. Wenn der Pilz nur in dem Medium wuchs der mit der Aminosäure Arginin angereichert war konnten die Forscher davon ausgehen, dass diese Mutante einen Defekt auf Argininsynthesestoffwechselweg besass. In weiteren Experimenten konnte der Stoffwechselweg genauer detailliert werden: Arginin wird in 3 Schritten synthetisiert: aus einer Vorläufersubstanz entsteht Ornithin, welches zu Citrullin wird, das zu Arginin umgesetzt wird. Beadle u. Tatum fanden heraus, dass einige Mutanten Arginin, andere entweder Arginin o. Citrullin, andere nur einen von den 3 Stoffen zum Wachstum brauchten. Sie schlossen daraus, dass diese Mutanten in versch. Schritten der Argininsynthese blockiert waren. Offenbar fehlte jeder Mutante ein spezifisches Enzym. Unter Annahme, alle Mutanten seien jeweils in einem Gen defekt, formulierten sie ihre Hypothese wonach die Rolle eines Gens darin besteht, die Produktion eines spezif. Enzyms zu bewirken. 2. Beschreibe Experimente, mit denen der genetische Code entschlüsselt wurde. Nierenberg gelang die künstliche Synthese einer mRNA poly U, er fügte Aminosäuren, Ribosomen und andere Komponenten der Proteinsynthese dazu, und es entstand die Aminosäure Polyphenylalanin. Das Triplett UUU codiert für die Aminosäure Phe. 1961 entschlüsselte Marshall Nirenberg das erste Codon indem er eine künstliche mRNA herstellte, indem er lauter RNA-Nucleotide mit der Stickstoffbase Uracil aneinanderhängte (UUU....). Diese Poly(U)-RNA gab er zusammen mit einer Mischung von Aminosäuren, Ribosomen und anderen f. Proteinsynthese nötigen Komponenten in ein Reagenzglas. Sein künstliches System translatierte Poly(U) in ein Polypeptid welches nur aus eine Sorte von Aminosäure enthielt, nämlich Phenylalanin (Phe).Daraus schloss er, dass das m RNA-Codon UUU müsse Phenylalanin codieren. Die Identifizierung von AAA, GGG, CCC erfolgte analog. Für gemischte Tripletts mussten kompliziertere Techniken eingesetzt werden. 3. Beschreibe die wichtigsten Schritte der Translation. Welche RNA-Typen sind beteiligt und wie unterscheiden sie sich? Translation: Je ein ein Triplett der mRNA codiert für eine Aminosäure welche durch die tRNA vermittelt wird. Die Aminoäure wird auf die tRNA mit einer spezifischen Aminoacyl tRNA Synthetase übergeben. Die Transkription wird in Initiation, Elognation und Termination gegliedert. mRNA: Messenger-RNA transportiert die genetische Botschaft v. d. DNA zur Proteinsynthese-Maschinerie. 1strangig. tRNAs: TransferRNAs binden spezifische Aminosäuren u. bauen durch die Wechselwirkung ihrer Anticodon-tripletts mit den komplementären Codons der mRNA am Ribosom eine Polypeptidkette auf. Kleeblattstruktur aus 4 basengepaarten Regionen, drei Schleifen, Aminosäureanheftungsstelle rRNA: ist Bestandteil des Ribosoms (60%), gliedert sich in drei Moleküle, ist der häufigste RNA-Typ in der Zelle Die 3 Hauptschritte der Translation sind: Initiation: die kleine Untereinheit des Ribosoms bindet an die spezif. Basensequenz am 5´Ende einer mRNA. Unmittelbar danach erfolgt der Initiationscodon AUG an dem die Translation beginnt. Initiator-tRNA, welche die Aminosäure Met trägt bindet an Initiationscodon. Die grosse Untereinheit des Ribosoms kommt hinzu. Elongation: Aminosäure nach der anderen wird anschliessend an die Startaminosäure angeheftet. Termination: Wenn Terminationscodon UAA , UAG, UGA an a-Stelle des Ribosoms kommt, führt diese zu einem Stoppsignal. Der Freisetzung oder Releasefaktor besetzt Terminationscodon welcher das fertige Polypeptid unter Addition eines Wassermoleküls von der letzten tRNA abspaltet. Das Ribosom dissoziert wiederum in 2 Teile und setzt mRNA u. Polypeptidkette frei. 4. Erkläre den Unterschied zwischen „missense“, „nonsense“ und „frame shift“ Mutationen. Auf welche Weise können bei diesen Mutationen Merkmalsänderungen ausgelöst werden? Eine „frameshift mutation“ entsteht, wenn die zugefügten oder gelöschten Basen kein Triplett bilden. Eine „missense mutation“ wird hervorgerufen durch die Mutagenese in eine andere Aminosäure. Bei einer nonsense uation wird eine Aminosäure in ein Stopcodon umgewandelt. Punktmutationen: Ist der Ersatz eines Nucleotids und seines Partners im komplementären DNA-Strang durch ein anderes Nucleotidpaar sind gewöhnlich Missense-Mutationen=Fehlsinn-Mutationen: das veränderte Triplett codiert immer noch eine Aminosäure, ist ein sinnvolles Codon, aber nicht im richtigen Sinne manchmal auch Nonsense-Mutationen: wenn dadurch ein Aminosäurecodon in ein Stoppcodon umgewandelt wird endet die Translation verfrüht, und das normale Polypeptid ist kürzer als das vom normalen Gen codierte. Führt meist zu einem untauglichen Protein. Insertionen oder Deletionen: Entstehen durch Einfügung oder Verlust eines oder mehrerer Nucleotidpaare in einem Gen. Rasterschub- o. Frameshift-Mutationen Entstehen, wenn die Zahl der eingefügten o. deletierten Nucleotide kein Vielfaches von 3 ist => Leseraster ändert sich Führt früher o. später zu Nonsense-Triplett => frühzeitige Termination der Translation. Wenn nicht sehr nahe am Ende der Translation liegt, führt Rasterschub mit Sicherheit zu nicht funktionellen Protein. 5. Beschreibe den Ames-Test. Der Ames Test wird vor allem eingesetzt, um die Wirksamkeit von Carcinogenen zu testen. Die Mutagene werden einem Medium zugefügt das Bakterien enthält, welche Histidin nicht selbst herstellen können. Damit nun die Bakterien überleben können müssen sie eine Rückmutation gemacht haben, welche ihnen erlaubt Histidin selbst herzustellen. Der Ames-Test misst das mutagene Potential verschiedener Chemikalien. Dazu wird a) ein potentielles Mutagen und Rattenleberextrakt mit einer Bakterienkultur gemischt und auf Histidinfreien Medium ausplattiert. (Das Rattenleberextrakt enthält Enzyme, welche bestimmte nichtmutagene Chemikalien in Mutagene umwandeln. Eigentlich sollten sie toxische Substanzen verstoffwechseln und diese so unschädlich machen.) Als Testbakterium wird eine Mangelmutante von Salmonella eingesetzt, die nicht in der Lage ist die Aminosäure Histidin zu produzieren. Diese Bakterien können nicht überleben, wenn sie auf einem Histidinfreien Medium ausplattiert werden. Einige dieser Bakterien machen jedoch eine Rückmutation durch. Durch ein Mutagen wird die Frequenz zur Rückmutation erhöht. b) Zum Vergleich wird eine Kultur von Histidinmangelmutante von Salmonella mit Rattenleberextrakt ohne Mutagen ausplattiert. => Durch auszählen der Kolonien rückmutierter Bakterien kann die mutagene Potenz des zu testenden Stoffes ermittelt werden. Biofragen Amrhein Kapitel 10, Photosynthese 1. Begründen Sie, weshalb die Gleichung 6CO2 + 6H2O → C6H12O6 + 6O2 für die Photosynthese inkorrekt ist. Wie lautet die korrekte Gleichung? Durch radioaktive Markierung des Sauerstoffs in H2O konnte nachgewiesen werden, dass das O2 ausschliesslich aus dem H2O entsteht. Die Gleichung 6CO2 + 6H2O → C6H12O6 + 6O2 ist daher nicht korrekt, da nach ihr nur 6 Sauerstoffatome aus dem H2O stammen können. Die restlichen 6, die es für die Bildung von 6O2 braucht, kämen laut dieser Gleichung vom CO2. Folglich lautet die korrekte Gleichung: 6CO2 + 12H2O → C6H12O6 + 6O2 + 6H2O 2. Schwefelpurpurbakterien produzieren keinen Sauerstoff bei der Photosynthese. Geben Sie die Gleichung für die Photosynthese dieser Organismen an. Was sind die Vor- und Nachteile dieser Form der Photosynthese? - Gleichung der anoxygenen Photosynthese: 6CO2 + 12H2S → C6H12O6 + 12S + 6H2O -II 0 + - H2S – 2e → S + 2H E0’ = -0.24V → H2S gutes Reduktionsmittel Vorteil: energetisch günstige Reaktion Nachteil: beschränkte Verfügbarkeit von H2S in der Natur 3. Welche Organismen verfügen über ein, und welche über zwei Photosysteme? Wie unterscheiden sich grundsätzlich die Reaktionen der Photosynthese in diesen beiden Gruppen? Photosystem I: Photosystem II: P-700 P-680 4. Wie erstellen Sie ein Aktionsspektrum der Photosynthese und welche Informationen entnehmen Sie ihm? -Das Aktionsspektrum sagt aus, in welchem Lichtwellenbereich die Photosynthese aktiv ist. -Engelmann’s Experiment: Man beleuchtet ein Filament einer Alge mit Licht, welches von einem Prisma in die verschiedenen Farben, also Wellenlängen, aufgeteilt wurde. Nun werden verschiedene Teile der Alge mit Licht unterschiedlicher Wellenlänge bestrahlt. Man benutzt noch aerobe Bakterien, welche sich in sauerstoffreichen Gebieten anlagern und vermehren. Lässt man die Alge so Photosynthese betreiben, sieht man nach einer gewissen Zeit, wo sich die Bakterien angehäuft haben. Dies ist bei rotem und blauem Licht der Fall. → rotes und blaues Licht ist am besten für die Photosynthese wird absorbiert Grünes Licht wird reflektiert → dient nicht so gut der Photosynthese. Das ist der Grund, dass die Chloroplasten grün sind. 5. Beschreiben Sie die Struktur und Funktion einer „Antenne“ (light harvesting center) in der Thylakoidmembran. - LHC (light harvesting center) = lichtsammelnder Antennenkomplex aus mehreren hundert Chlorophylla-, Chlorophyllb- und Carotinoidmolekülen - Der Vorteil dieses Komplexes ist eine grössere Aufnahmeoberfläche und ein breiterer Wellenbereich. Absorbiert ein Antennenpigment (AP) ein Photon, wird die Energie zum nächsten AP weitergegeben bis sie ein bestimmtes Chlorophylla-Molekül erreicht, das sich im Reaktionszentrum (RZ) befindet. Im RZ findet die erste chemische Reaktion der Photosynthese statt. Photon Reaktionszentrum Energieübertragung Daneben befindet sich im Reaktionszentrum der primäre Elektronenakzeptor (pEA). An ihn verliert das Chlorophylla eines seiner Elektronen, da der pEA das angeregte Elektron des Chlorophylls bindet, bevor es wieder in den Grundzustand fällt. Man kann sich den pEA als Schranke vorstellen, die dem energiereichen Elektron den Rückweg in den Grundzustand versperrt. 6. Wo in einem Chloroplasten findet die Photophosphorylierung, d.h. ATPBildung, statt? Woher stammt unmittelbar die Energie für die Bildung des ATP? Die Photophosphorylierung findet in der Thylakoidmembran des Chloroplasten statt. Die Energie für die Bildung des ATP stammt unmittelbar vom Licht (dessen Energie in der Chemiosmose in chemische Energie umgewandelt wird). Chemiosmose (siehe S.212 Campbell, dt. Ausgabe): Die Photophosphorylierung in den Chloroplasten findet an der Thylakoidmembran statt. Der ATP-Synthase-Komplex koppelt die protonenmotorische Kraft, die durch den Rückfluss der Protonen vom Lumen ins Stroma entsteht für die Synthese von ATP (Chemiosmotische Theorie) (Die Protonen ihrerseits wurden durch die Energie aus der Elektronenkette gegen ihren Gradienten ins Lumen transportiert) 7. Welche Einheiten sind im sogenannten „Z-Schema“ des photosynthetischen Elektronentransports auf der Ordinate aufgetragen? Geben Sie in einer Skizze die Position von H2O/O2 und NADP+/NADPH + H+ und die Richtung des Elektronen-transports an. Auf der y-Achse ist die Energie der Elektronen in Volt aufgetragen (E`0). H20/02 liegen am Anfang der Elektronenkette, da aus der Oxidation von 2 Wasser-Molekülen am Photosystem II ½ Sauerstoff + 2 Elektronen + 2Protonen (Protonentransport für ATP-Synthese) resultieren. Die Elektronen werden für den Elektronen-Transport gebraucht. Ganz am Schluss der Elektronentransport-Kette wird NADP zu NADPH reduziert (Reduktionsmittel im CalvinZyklus). Wie es der Name schon sagt, werden die Elektronen z-förmig über zwei Photosysteme (II dann I) weitergegeben. Z-Form deshalb, weil die Elektronen an Energie gewinnen (Photosystem I ist energetisch höher gelegen als PS II). 8. Geben Sie die Reaktionen (mit Strukturformeln!) an, in denen die Produkte der Lichtreaktion der Photosynthese, ATP und NADPH+H+, verbraucht werden. Diese 2 Reaktionen finden im Calvin-Zyklus,2. Phase (Reduktion), statt: PhosphoglyceratKinase 6x Phosphoglycerat 6x ATP 6x Bisphosphatglycerat 6x ADP Reduktion 6x Bisphosphatglycerat 6x (NADPH+H+) Glyceralphos phatDehydrogena se 6x Glycerinaldehyd-3-Phosphat 6x (Pi + NADP+) NADPH und ATP, die während der Photosynthese (Lichtreaktion) gebildet wurden, werden im Calvin-Zyklus (Dunkelreaktion im Stroma) wieder verbraucht. Im Calvin-Zyklus wird Glycerinaldehyd-3-Phosphat gebildet. Zur Bildung von 6 Molekülen des Aldehyds braucht es 6 ATP und 6 NADPH. ATP wird verbraucht bei der Uebertragung einer Phosphat-Gr. aufs 3-Phosphoglycerat, was zu 1,3-Bisphosphoglycerat führt. NADH jedoch wird in der folgenden Reduktion zu Glycerinaldehyd-3-Phosphat verwendet 9. Was passiert mit dem Gycerinaldehyd-3-Phosphat, das in der reduzierenden Phase des Calvin-Zyklus entsteht? In den letzten Schritten der Photosynthese werden die Kohlenstoffgerüste der G3P (Glycerinaldehyd-3-Phosphat) – Moleküle in einer komplizierten Reaktionsfolge so umgeordnet, dass wieder drei Moleküle Ribulose-1,5-bis-phosphat (RuBP) entstehen. Und zwar werden 5 der 6 G3P dafür verwendet und zudem wird noch einmal ein ATP verbraucht. Anschliessend kann das RuBP erneut CO2 aufnehmen, und der Zyklus läuft ein weiteres Mal ab. Glycerinaldehyd-3-Phosphat, das während dem Reduktionsschritt entstanden ist, wird für die Bildung von Glucose, Kohlenhydrate oder Synthese von Zellbestandteilen weiterverwendet. Glucose kann bsw weiter zur Stärke-Synthese (Energiespeicher) gebraucht werden. 10. Mit welchem Experiment hat M. Calvin das erste, nach der Fixierung von CO2 in der Photosynthese entstehende Produkt nachgewiesen? Was ist die chemische Struktur dieses Produkts? Durch Autoradiographie: Calvin gab einer Algensuspension (einzellige Grünalgen) radioaktiv markiertes Kohlenstoffdioxid zu (NaH14CO3). Diesen Ansatz tötete er durch eingiessen in siedenden Alkohol ab, wobei die Kohlenstoffdioxid-Fixierungsperiode variiert wurde. Die löslichen Inhaltsstoffe wurden extrahiert und durch Chromatographie getrennt. Das Chromatogramm brachte er im Dunkeln in engen Kontakt mit einem unbelichteten Röntgenfilm. Die hochempfindliche Schicht des Röntgenfilmes wurde durch die radioaktive Strahlung des 14C geschwärzt. Nach 2s Fixierungszeit war der grösste Teil des radioaktiv markierten Kohlenstoffs in Form von Phosphoglycerinsäure festgelegt. Es war das erste stabile Zwischenprodukt. Calvin machte einen Versuch mit Chlorella, einer einzelligen Grünalge (1 Thylakoid): Er hat die Algen, die vorher mit einem 14C radioaktiv markiert wurden, mit Licht bestrahlt (nur 2sek.!!) Danach hat er durch Zugabe von Alkohol die Alge abgetötet. Im weiteren Schritt extrahierte er die Metaboliten. Nach erfolgter Chromatographie konnte er auf dem Autoradiogramm einen schwarzen Fleck erkennen, was auf das 1. Produkt 3-Phosphoglycerinsäure hinwies 11.Wie ist Stärke chem. aufgebaut, wie lässt sie sich nachweisen, und in welchen Organellen einer Pflanzenzelle kommt sie vor? Der in den Chloroplasten erzeugte Zucker wird enzymatisch weiter zu Saccharose und Stärke abgebaut. Dabei ist Stärke eine Speicherform und wird somit in Blättern, Wurzeln, Knollen, Samen und Früchten gespeichert. Sie kommt natürlich auch in den Chloroplasten der Blätter vor. Die Stärke ist ein Kohlenhydrat (Polysaccharid; 1-4 Verbindung), aus Glucose aufgebaut. Unverzweigte Stärke färbt sich mit elementarem Iod blau, verzweigte Stärke lediglich braun od. rotbraun. Ein Polymer, das vollständig aus Glucosemonomeren besteht, ist die Stärke, ein Speicherpolysaccharid der Planzen (Campbell, page 66/ 5.6a). Die meisten dieser Monomere sind über 1,4-Bindungen miteinander verknüpft (Kohlenstoff Nummer 1 mit Kohlenstoff Nummer 4), genau wie die Glucoseeinheit der Maltose (siehe Abbildung page 65/ 5.5a). Der Winkel zwischen diesen Bindungen macht das Polymer helikal (gewendelt). Die einfachste Form der Stärke, die Amylose, ist unverzweigt, während das Amylopektin, eine komplexere Stärkeform, ein verzweigtes Polymer ist, das 1,6-Bindungen an den Verzweigungspunkten aufweist. Pflanzen speichern Stärke als Granula in zellulären Strukturen, die Plastiden genannt werden und zu denen auch die Chloroplasten gehören (siehe Abbildung page 66/ 5.6a). Indem sie Stärke synthetisieren, können Pflanzen überschüssigen Zucker lagern. Da Glucose einer der Hauptbetriebsstoffe der Zelle ist, stellt Stärke gespeicherte Energie dar. Aus dieser „Kohlenhydratbank“ kann der Zucker dann später durch Hydrolyse, welche die Bindungen zwischen den Glucosemonomeren aufbricht, entnommen werden. Die meisten Tiere und auch der Mensch besitzen ebenfalls Enzyme zur Hydrolyse von Pflanzenstärke, was die Glucose den Zellen als Nährstoff zugänglich macht. Getreidekörner, also die Früchte von Weizen, Mais, Reis und anderen Gräsern, sind neben Kartoffelknollen die hauptsächlichen Stärkequellen. 12.Im Calvin-Zyklus wird CO2 in ein Kohlenhydrat umgewandelt. Welches ist die tatsächliche Verbindung, die reduziert wird? Geben sie die Strukturformel der oxidierten und der reduzierten Form an. Welche Kofaktoren werden für diese Reaktion benötigt? In der 1.Phase des CZ wird CO2 an die Ribulose-1,5-biphosphat angelagert. Das Produkt dieser Reaktion ist ein instabiles Molekül mit 6 C-Atomen das sofort in 3Phosphoglycerat zerfällt (3 C-Atome). In der 2.Phase wird an 3-Phosphoglycerat noch eine Phosphatgruppe von ATP angelagert. Es entsteht 1,3-Biphosphoglycerat welches von einem e-Paar des NADPH zu Glycerinaldehyd-3-Phosphat reduziert wird. Strukturen von 1,3-Biphosphoglycerat und Glycerinaldehyd-3-Phosphat habe ich nirgens genauer als auf Seite 215(dt. Campbell) gefunden. Kofaktoren sind ATP und NADPH Der Zyklus beginnt, indem CO2 aus der Luft mit organischen Molekülen reagiert, die bereits in Chloroplasten vorhanden sind. Diesen ersten Einbau von Kohlenstoff in organische Verbindungen bezeichnet man als Kohlenstoff-Fixierung. Der so gebundene Kohlenstoff wird dann im Calvin-Zyklus durch Aufnahme von Elektronen zum Kohlenhydrat reduziert. Das Reduktionsmittel ist NADPH, das seine energiereichen Elektronen in den Lichtreaktionen aufgenommen hat. Es wird dabei zu NADP+ oxidiert. Um CO2 zum Kohlenhydrat umzusetzen, braucht der Calvin-Zyklus auch chemische Energie in Form von ATP, und dieses wird ebenfalls in den Lichtreaktionen gebildet. Der Calvin-Zyklus ist also derjenige Prozess, der die Zuckermoleküle erzeugt, aber das ist nur mit Hilfe von NADPH und ATP aus den Lichtreaktionen möglich. Die Stoffwechselreaktionen des Calvin-Zyklus bezeichnet man manchmal auch als Dunkelreaktionen, weil keine davon unmittelbar Licht benötigt. Dennoch läuft der Calvin-Zyklus in den meisten Pflanzen bei Tageslicht ab, denn nur dann können die Lichtreaktionen jene NADPH- und ATP-Moleküle regenerieren, die bei der Reduktion von CO2 zu Zucker verbraucht werden. Im Endeffekt nutzt der Chloroplast die Lichtenergie zur Zuckersynthese, indem er die beiden Abschnitte der Photosynthese miteinander koppelt. 13.Welche Doppelfkt. Hat Plastochinon in der Thylakoidmembran? Aufgrund welcher Eigenschaften kann es diese Fkt. wahrnehmen? Ich habe leider nur eine Funktion des Plastochinons gefunden. Bei der Nichtzyklischen Phosphorylierung in der Lichtreaktion spielt es eine Rolle beim Elektronentransport vom Photosystem 2 zum Photosystem 1. Die Elekronen fliessen vom primären Elektronenakzeptor des Photosystems 2 über Plastochinon (Elektronenakzeptor) dann Cytochromkomplex und Plastocyanin bis es dann beim Photosystem 1 ankommt. Nichtzyklischer Elektronentransport: Licht versetzt die beiden in die Thylakoidmembran eingelagerten Photosysteme in einen energiereichen Zustand und treibt so die Synthese von NADPH und ATP an. Das Entscheidende bei dieser Energieumwandlung ist der Elektronenfluss durch die Photosysteme und die anderen molekularen Bestandteile in der Thylakoidmembran. In den Lichtreaktionen stehen den Elektronen 2 Wege offen, ein zyklischer und ein nichtzyklischer Elektronentransport. Die angeregten Elektronen fliessen über eine Elektronentransportkette vom primären Elektronenakzeptor des Photosystems II zum Photosystem I. Diese Kette ähnelt stark der Atmungskette. In den Chloroplasten besteht sie zunächst aus einem Elektronenüberträger namens Plastochinon, dann folgt ein Komplex aus zwei Cytochromen und schliesslich ein kupferhaltiges Protein namens Plastocyanin. Das Plastochinon verschiebt Elektronen auf den Cytochromkomplex, und gleichzeitig werden Protonen durch die Membran transportiert. Dies bedeutet, wenn die Elektronen die Energiekaskade der Transportkette passieren, nutzt die Thylakoidmembran ihren exergonischen Fluss zur Produktion von ATP. Diese Art der ATP-Synthese nennt man Photophosphorylierung, weil sie von Lichtenergie angetrieben wird. Genauer bezeichnet man die Produktion von ATP durch nichtzyklischen Elektronentransport als nichtzyklische Photophosphorylierung. 14.Was versteht man unter zyklischer Phosphorylierung und welche Produkte entstehen? Unter bestimmten Bedingungen (NADPH-Konz.) ist nur das Ph 1 am Elektronenfluss beteiligt. Dabei kehren die Elektronen über die Transportkette Fd, Cytochromkomplex, Pc zum Ausgangschlorophyll zurück. Bei diesem Vorgang entsteht kein NADPH. Es wird aber ATP (über chemiosmotische Synthese) erzeugt. Der Calvin-Zyklus verbraucht mehr ATP als NADPH und darum gleicht die zyklische Phosphorylierung das aus. Wie man auf page 187/ 10.14 erkennen kann ist der zyklische Elektronentransport ein Kurzschluss: Die Elektronen kehren über die gleiche Transportkette, die auch am nichtzyklischen Vorgang beteiligt ist, zum Chlorophyll-Paar P700 zurück. Dabei wird weder NADPH produziert noch Sauerstoff freigesetzt. Der zyklische Elektronentransport erzeugt aber ATP. Man spricht deshalb auch von zyklischer Photophosphorylierung im Gegensatz zur nichtzyklischen Photophosphorylierung. Beim nichtzyklischen Prozess entstehen ATP und NADPH in ungefähr gleichen Mengen, aber der Calvin-Zyklus verbraucht mehr ATP als NADPH. Den Unterschied gleicht der zyklische Elektronentransport aus. Wird das ATP für den Calvin-Zyklus knapp, sammelt sich NADPH an, weil der Calvin-Zyklus seinen Durchsatz verlangsamt. Dieser Anstieg der NADPH-Menge dürfte vorübergehend für eine Verschiebung vom nichtzyklischen zum zyklischen Elektronentransport sorgen, bis der Nachschub an ATP wieder dem Bedarf entspricht. Es spielt keine Rolle von welchen Elektronentransport die Photophosphorylierung angetrieben wird, denn der eigentliche Mechanismus der ATPSynthese bleibt stets derselbe. 15.Die Überführung eines Mol CO2 auf dieRedoxstufe des Kohlenhydrates in der Photosynthese erfordert 3 Mol ATP. Bei welchen Reaktionen wird dieses ATP verbraucht? 6 Mol ATP werden verbraucht um jeweils 1 Phosphat auf die 6 Mol neu entstandenen 3Phosphoglycerate zu übertragen, welche aus der Fixierung von 3 Mol CO2 entstanden sind. Um aus 5 Mol Glycerin-3-phosphat (1 Mol wird als Etrag aus dem Zyklus ausgeschleust) wieder Ribulose-1,5-biphosphat herzustellen, braucht es nochmals 3 Mol ATP. Somit braucht es für 3 Mol CO2 9Mol ATP, d.h. für 1 Mol CO2 braucht es 3 Mol ATP. Bei dem Kohlenhydrat, das im Calvin-Zyklus unmittelbar entsteht, handelt es sich nicht um Glucose, sondern um Glycerinaldehyd-3-phosphat (G3P), einen Zucker mit 3 Kohlenstoffatomen. Damit unter dem Strich 1 Molekül dieses Zuckers synthtisiert wird, muss der Zyklus 3-mal ablaufen, sodass 3 CO2-Moleküle fixiert werden. Wenn man die Schritte des Zyklus nachzeichnet, muss man daran denken, dass man in Wirklichkeit den Weg dreier CO2Moleküle durch die Reaktionen verfolgt. 16. Welche Produkte der Lichtreaktionen der Photosynthese werden im Calvinzyklus verbraucht und welche Produkte werden nicht weiter benötigt (Abfallprodukt)? Das gebildete ATP und NADPH + H+ werden im Calvinzyklus weiter benötigt, O2(Sauerstoff) ist hingegen ein „Abfallprodukt“. 17. Wie wird die Redoxenergie des photosynthetischen Elektronentransports in die „proton-motive-force“ umgewandelt? (siehe Folie 26 der Powerpointpräsentation Photosynthese) Mit Hilfe von Plastoquinon (PQ), das mit den 2 Elektronen vom primären Akzeptor des Photosystems II auch 2 H+ vom Stroma aufnimmt und so reduziert wird. Bei der anschliessenden Oxidation werden die 2 H+ ins Lumen (= Thylakoidinnenraum) abgegeben und so wird ein Protonengradient erzeugt, der dann die ATP Synthase antreibt. 18. Zeichnen Sie ein topographisches Modell der Thylakoidmembran, das erkennen lässt, wie die Elektronen vom Sauerstoff des Wassers zum NADP+ gelangen. Habe leider keinen Scanner, aber die Abbildung im Buch ist sehr übersichtlich: im englischen Buch: Seite 189 und im deutschen Buch: Seite 213. Die Folie 26 der Powerpointpräsentation zeigt das Ganze noch etwas genauer(aber nicht unbedingt verständnisvoller). Bei der Skizze würde ich aber die ATP-Synthase weglassen. 19. Weshalb fluoresziert Chlorophyll in intakten Chloroplasten weniger als in Lösung? In Lösung absorbiert Chlorophyll Licht bestimmter Wellenlängen genauso wie in der Thylakoidmembran, es fehlt aber an Photosystemen, die die Energie des vom angeregten in den Grundzustand zurückfallenden Elektrons auffangen und weiterleiten können. So fällt das Elektron in jedem Fall unter Ausstoss von Wärme und energieärmerem Licht (d.h. die Wellenlänge ist grösser) in seinen Grundzustand zurück. Das abgegebene, energieärmere Licht nehmen wir als Fluoreszenz wahr. 20. Warum findet die Photosynthese in grünem Licht am wenigsten effektiv statt? Die Thylakoidpigmente absorbieren vorwiegend im blauen und roten Bereich des sichtbaren Lichtes (bei bestimmten Wellenlängen, wo die Lichtreaktionen ablaufen können). -> Grünes Licht wird meistens an der Thylakoidmembran reflektiert oder durchquert sie(so genannte Transmission). (-> Deshalb erscheinen die Chloroplasten grün!) Und wenn keine Photonen absorbiert werden, kann auch die Kettenreaktion in den Photosystemen nicht starten, was wiederum keine Photosynthese bedeutet. Kapitel 29, Diversität der Pflanzen I, wie die Pflanzen das Land kolonisierten Kapitel 30, Diversität der Pflanzen II, die Evolution der Samenpflanzen 1. Aus welchen Bestandteilen besteht ein Samen, und aus welchen Strukturen entstehen diese? a) Embryo (2n) → entsteht aus der Zygote Nährgewebe: i) Bei Gymnospermae: Megagametophyt legt Nährgewebe an (primäres Endosperm, 1n). ii) Bei Angiospermae: doppelte Befruchtung → triploides sekundäres Endosperm (aus Fusion des Zellkerns einer generativen Zelle mit den 2 Polkernen). b) Samenschale (=Testa): Entsteht aus dem oder den Integumenten, die die Samenanlage abschliessen. Samenanlage besteht aus Nucellus (entspricht Megasporangium) + Integument(en). 2. Wie werden das (primäre) Endosperm der Gymnospermen und das (sekundäre) Endosperm der Angiospermen gebildet? Primäres Endosperm: Meiose der Megasporenmutterzelle (2n) im Nucellus → 4 haploide Zellen. Nur eine überlebt und wird zur Megaspore. Sie teilt sich mehrmals und bildet den unreifen weiblichen Gametophyten. (Schliesslich ist der Pollenschlauch durch den Nucellus zum weiblichen Gametophyten gewachsen und befruchtet dort eine oder mehrere Eizellen.) Der weibliche Gametophyt umgibt und ernährt den Embryo. Sekundäres Endosperm: Der Embryosack ist der Megagametophyt der Angiospermen, entstanden aus der Megaspore durch 3 Mitosen → 8 Zellen. Mitotische Teilung des generativen Kerns → 2 generative Kerne. Aus der Fusion eines Kerns der generativen Spermazelle mit den 2 Polkernen entsteht das sekundäre, triploide Endosperm. 3. Geben Sie den Ploidiegrad und die jeweilige Herkunft des Genoms / der Genome an für 1.) die Integumente, 2.) den Embryo, 3.) das sekundäre Endosperm einer angiospermen Pflanze. 1.) 2.) 3.) 2n, aus sporophytischem Gewebe 2n. 2n, 1n aus der Eizelle + 1n eines Spermakerns 3n, 2n aus den beiden Polkernen der zentralen Zelle im weiblichen Gametophyten (=Embryosack) + 1n aus einer generativen Zelle (1 Spermakern, männlicher Gamet). 4. Ordnen Sie die nachfolgenden Begriffe jeweils zu Paaren von Homologen: Megasporophyll – Fruchtblatt Megasporangium – Nucellus Mikrosporangium – Pollensack Mikrosporophyll – Staubblatt Spermazelle (Spermatozoid) – generativer Kern Megagametophyt – Embryosack 5. Welche Strukturen / Komponenten beinhaltet der reife, trockene Zapfen einer Konifere? Weiblicher Zapfen: - Samenschuppe (Megasporophyll) mit Samenanlage(Megasporangium): o Nucellus (Sporangium) o Megasporenmutterzelle (2n) o Integumente o Mikropyle - Deckschuppe Männlicher Zapfen: - Mikrosporophyll mit Pollensäcken (Mikrosporangium): diese bilden Mikrosporen → Bestäubung 6. Was bedingt die besondere Stellung des Ginkgo Baumes (Ginkgophyta) innerhalb der Gymnospermen? Sie haben fächerförmige Blätter, welche sich im Herbst golden Färben und abfallen ungewöhnliche Eigenschaft für Gymnospermen) (eine Die Ginkgoatae sind mit einer Ausnahme, eben Ginkgo biloba, nur fossil bekannt → lebendes Fossil. 7. In welchen Pflanzengruppen kommen Tracheiden, und in welchen kommen Tracheen vor? Was ist der Unterschied zwischen diesen beiden Typen? (Beim Übergang der Pflanzen vom Wasser- zum Landleben hat sich das Leitgewebe entwickelt, um den Stofftransport und die Festigung der Pflanzen an Land zu ermöglichen. Im Siebteil (Phloem) erfolgt der Transport von Stoffwechselprodukten der Pflanze von den Blättern wurzelwärts. Im Gefässteil (Xylem) werden Wasser und Nährsalze von den Wurzeln zu den Blättern geleitet. Die Leitelemente des Xylems sind verholzt. (→Festigung)) Kormophyta (Gefässpflanzen) = Pteridophyta (Farnpflanzen) und Spermatophyta (Samenpflanzen) Tracheiden kommen als Leitgefässe des Xylems in den Kormophyten vor: Sie sind röhrenförmige, englumige, aneinandergereihte Gefässzellen. Sie erfüllen eine Stützfunktion und eine Leitfunktion (Wasser und Nährstoffe). Bei den meisten Angiospermen haben sich zusätzlich zu den Tracheiden die Tracheen entwickelt. Sie sind kürzer und weitlumiger als die Tracheiden und werden Ende an Ende zu einer offenen Röhre aneinandergereiht. Tracheen haben sich mehr auf den Wassertransport als auf die Stützfunktion spezialisiert. 8. Zeichnen Sie die Idealisierte Blüte einer angiospermen Pflanze im Längsschnitt und bezeichnen Sie die modifizierten Blätter und ihre Funktionen. Funktionen der modifizierten Blätter: Abgewandelte sterile Blätter: Kelchblatt: Umhüllung der geschlossenen Blüte Kronblatt: meist bunt gefärbt zur Anlockung von bestäubenden Insekten Abgewandelte fertile Blätter (=Sporophylle): Staubblatt (männliches Sporophyll): Produktion von Mikrosporen (1n), welche zu Pollenkörnern (männlichen Gametophyten) reifen. Fruchtblatt (weibliches Sporophyll): Produktion von Megasporen (1n), welche sich jeweils zu einem Embryosack (weiblicher Gametophyt = Megagametophyt) entwickeln. Aus welchen Blättern entsteht die Frucht, und woraus entstehen die Samen? Die Samenanlage besteht aus dem Megagametophyten, welcher von sporophytischem Gewebe (Nucellus und einem oder mehreren Integumenten) umgeben ist. Wenn die Eizelle im Megagametophyt von der Spermazelle eines Pollenkorns befruchtet wird, entwickelt sich die Samenanlage zu einem Samen. Die Frucht entsteht aus dem Fruchtknoten, welcher sich aus einem oder mehreren Fruchtblättern zusammensetzt und die Samenanlage umgibt. 9. Wo in einer angiospermen Pflanze findet die Meiose statt? Was sind die Produkte der Meiose? In den Pollensäcken des Staubbeutels (Anthere): Aus einer Pollenmutterzelle (2n) entstehen durch Meiose vier haploide Mikrosporen (aus welchen sich nach einer ersten Pollenmitose vier zweizellige Pollenkörner entwickeln.) Im Nucellus: Aus einer Embryosackmutterzelle (2n) entstehen durch Meiose vier haploide Makrosporen, von welchen drei verkümmern und eine (=Megaspore) sich zum Embryosack entwickelt. 10. Vergleichen Sie die Übertragung der männlichen Gameten (Spermazellen) zur Eizelle bei den Farnen und Koniferen. [Gymnospermen →Unterklasse Pinidae (=Coniferae) → Ordnung Pinales (=Koniferen im engeren Sinne)] Bei den Farnen entwickeln sich die männlichen (Antheridien) und die weiblichen Organe (Archegonien) auf dem meist flach und blattartigen Prothallium (Gametophyt). In den Antheridien werden bewegliche männliche Gameten gebildet, die Spermatozoiden. Diese können bei Anwesenheit von Wasser (Regen, Tau) zu den Archegonien schwimmen und dort die reife Eizelle befruchten. Bei den Koniferen gibt es weibliche und männliche Blüten: In der weiblichen Blüte sitzt die Samenanlage offen auf einem Träger (Samenschuppe). [Dieser ist in der Achsel eines Tragblatts (Deckschuppe) angeordnet. Viele Blüten bilden vielblütige Blütenstände (Zapfen)] In der männlichen Blüte sitzen dicht gedrängt zahlreiche schuppenförmige Staubblätter, in denen sich zwei bis viele Pollensäcke befinden. In ihnen entwickeln sich die Pollen - Im Gegensatz zu den Spermatozoiden der Farne, erlauben die Pollen den Koniferen die Windbestäubung. (Die Pollen sind von einer schützenden Hülle umgeben, die es erlaubt, an der Luft weite Strecken zurückzulegen ohne auszutrocknen.) - Die Pollen der Koniferen besitzen keine Flagellen wie die Spermatozoiden der Farne und sind daher selber unbeweglich. (Sie sind bei einigen Arten beflügelt, was den Windtransport erleichtert) - Bei den Koniferen gelangen die Spermazellen der Pollen über einen ausgebildeten Pollenschlauch zur Samenanlage, während die Spermatozoiden der Farne selber in das Flaschen- oder Krugförmige Archegonium schwimmen und zur Eizelle gelangen. 11. Welche Struktur der angiospermen Pflanze entspricht dem Megagametophyten? Wie wird sie gebildet? Der Embryosack entspricht dem Megagametophyten. Er entwickelt sich aus einer Megaspore (Siehe Frage 9). Die Megaspore (1n) teilt sich durch Mitosen in acht gleiche Kerne (1n): Je drei Kerne wandern zu den Polen des Embryosacks und umgeben sich mit eigenem Plasma und einer Membran. Die drei Zellen in der Basis nennt man Antipoden, die oberen drei Zellen bilden den Eiapparat, welcher aus einer Eizelle und zwei Synergidien (Gehilfinnen) besteht. Die restlichen beiden Kerne verschmelzen im Zentrum des Embryosacks zu einer Zentralzelle (2n). 12. Bei welchen Pflanzen findet die „doppelte Befruchtung“ statt, wie läuft sie ab, und was sind die Produkte und deren Ploidiegrad? Die doppelte Befruchtung ist vor allem charakteristisch für Angiospermen, sie kommt aber auch bei einigen Gymnospermen der Klasse Gnetatae vor. Ein Pollenkorn besteht aus einer kleinen generativen Zelle und einer grossen vegetativen Zelle, (Siehe Frage 9) welche von einer widerstandsfähigen Schicht umgeben sind. Der Kern der generativen Zelle teilt sich durch eine Mitose in zwei befruchtungsfähige Spermazellen (1n). Wenn das Pollenkorn auf die Narbe eines Fruchtblattes fällt, so bildet die vegetative Zelle einen Pollenschlauch aus, welcher durch Narbe, Stempel und Mikropyle (Öffnung der Integumente) bis in den Embryosack vordringt. Die Spermakerne vollziehen nun die doppelte Befruchtung: Generative Befruchtung: Ein Spermakern (1n) verschmilzt mit dem Eikern (1n) zu einer Zygote (2n) (wird später zum Embryo) Vegetative Befruchtung: Der andere Spermakern verschmilzt mit der diploiden Zentralzelle zum Endospermkern (3n) (Wird später zum Nährgewebe (=Endosperm)) Kapitel 35, Pflanzenstruktur und –wachstum 1. Nennen Sie fünf wesentliche morphologische und anatomische Unterschiede zwischen monokotylen und dikotylen Angiospermen. Monocotyledonen Dicotyledonen Keimblätter 1 Keimblatt 2 Keimblätter Blätter meist parallelnervig meist netznervig Leitbündel zerstreut angeordnet und ringförmig angeordent und geschlossen (ohne Kambium) offen (Kambium) fasriges Wurzelsystem Pfahlwurzelsystem (taproot s.): (fibrous root system) lange vertikale Wurzel mit vielen Wurzeln kleinen seitlichen W. Blüten meist 3zählig oft 4- oder 5zählig 2. Zeichnen Sie Querschnitte durch die Stengel monokotyler und dikotyler Pflanzen und machen Sie eine Aussage zum jeweiligen Dickenwachstum. Zeichnung: FIGURE 35.18, p 733 Monocotyledonen: sekundäres Dickenwachstum selten Dicotyledonen: Bei den meisten Dicotyledonen findet sekundäres Dickenwachstum statt. Dabei spielen zwei laterale Meristeme eine Rolle: das Leitgewebe-Kambium bildet sekundäres Xylem und Phloem und das Korkkambium bildet eine widerstandsfähige, dicke Schutzschicht für Stengel oder Stamm und Wurzeln. 3. Vergleichen Sie die Entstehung von Seitensprossen und Seitenwurzeln (Zeichnung). Eine etablierte Wurzel kann Seitenwurzeln bilden, die aus der äussersten Schicht der Stele (zentraler Gefässzylinder), dem Perzykel hervorgehen. Seitenwurzeln werden im Perzykel angelegt, strecken sich und durchstossen den Cortex bis sie als Primärwurzeln hervortreten. Das Leitgewebe ist in der Wurzel ein durchgehendes System. Zeichnung: FIGURE 35.16, p 732 Seitensprosse entstehen aus Achselknospen,die auf der Oberfläche des Hauptsprosses angesiedelt sind. Die Leitbündel einer Sprossachse befinden sich nahe der Oberfläche. Daher können sich Seitenzweige mit einer Verbindung zum Leitgewebe entwickeln, ohne aus der Tiefe des Hauptsprosses hervorgehen zu müssen. Zeichnung: FIGURE 35.17, p 732 4. Der Spross von Angiospermen hat eine modulare Konstruktion (“Phytomere“). Zeichnen Sie ein Phytomer und benennen Sie seine Bestandteile. Ein Phytomer besteht aus Blatt, Achselknospe, Nodium und Internodium. Der Spross bestheht aus Phytomeren -> Modularer Aufbau. Zeichnung: Vorlesung!, (FIGURE 35.2, p 722) 5. Wo und wie entsteht ein Blatt an einer Pflanze? (Zeichnung) Blätter entstehen aus Blattprimordien an den Seiten des Apikalmeristems. Achselknospen entwickeln sich aus Inseln meristematischer Zellen, die vom Apikalmeristem an der Basis der Blattprimordien zurückgelassen werden. Zeichnung: FIGURE 35.17, p 732 6. Vergleichen Sie die Entstehung einer Seitenwurzel mit derjenigen eines Wurzelhaars. (Zeichnung) Etablierte Wurzeln können Seitenwurzeln bilden, die aus der äussersten Schicht der Stele (zentraler Gefässzylinder), dem Perzykel hervorgehen. Seitenwurzeln werden im Perzykel angelegt, strecken sich und durchstossen den Cortex bis sie als Primärwurzeln hervortreten. Zeichnung: FIGURE 35.16, p732 Wurzelhaare bilden sich an den Spitzen der Wurzeln und sind Ausstülpungen der Epidermis. Zeichnung: FIGURE 35.3, p 722 7. Wie ist ein Leitbündel aufgebaut? Was sind die wesentlichen funktionellen Unterschiede seiner Bestandteile? Wie unterscheiden sich die Leitbündel von monokotylen und dikotylen Pflanzen? Leitbündel bestehen aus Leitgewebe: Xylem und Phloem. Das Xylem besteht aus Tracheiden und Gefässen, deren langgezogene Zellen bei ihrer funktionalen Reife tot sind. Sie hinterlassen verdickte Zellwände, die einen Kanal bilden, durch den Wasser fliessen kann. Das Phloem besteht aus sog. Siebröhren, die aus lebenden Zellen gebildet werden. Das Xylem transportiert Wasser und Mineralien von den Wurzeln bis in den Trieb. Das Phloem transportiert Nährstoffe (Assimilationsprodukte) von den Blättern zu den Wurzeln und Teilen des Sprosssystems ,die keine Photosynthese machen, z.B. Früchte. In den meisten Dicotyledonen sind die Leitbündel ringförmig angeordnet, mit Markparenchym innerhalb und Rindenparenchym (Cortex) ausserhalb des Rings. Beide sind Teile des Grundgewebesystems. Das Xylem liegt jeweils innen, zum Mark hin gerichtet, das Phloem verläuft entlang der Rinde. Die Leitbündel sind offen, d.h. sie besitzen ein Kambium. In den Monocotyledonen liegen die Leitbündel im Grundgewebe über den Querschnitt verstreut und sind geschlossen (ohne Kambium). Kapitel 38, Reproduktion der Pflanzen 1. Wie haben unterschiedliche Vertreter der Spermatophyten das Problem der Reservestoffspeicherung in den Samen gelöst? Meistens ist es so, dass die Nährstoffe, die reich im Endosperm gespeichert sind, dem sich entwickelnden Embryo geliefert werden. MONOCOTYLEDONEN: Das Endosperm der Monocotyledonen speichert auch Nährsoffe, die vom Keimling nach der Keimung genutzt werden können. Sie besitzen ein einzelnen Keimblatt (Scutellum), das sehr dünn ist, aber eine grosse Oberfläche hat, was die Absorption von Nährstoffen während der Keimung deutlich verbessert. DICOTYLEDONEN: Bei den Dicotyledonen werden die Nahrungsreserven des Endosperms zu den Keimblättern transportiert, bevor der Samen seine Entwicklung vollendet. Daher fehlt dem reifen Samen ein Endosperm. Einige von ihnen halten ihre Nahrungsreserven im Endosperm zurück und haben sehr dünne Keimblätter.Diese übernehmen die Nährstoffe erst dann aus dem Endospoerm und leiten sie zum Embryo, wenn der Samen keimt. 2. Wie erfolgt die Endospermbildung bei a) den Gymnospermen b) den Angiospermen? Was ist der jeweilige Ploidiegrad? Woher stammen die jeweiligen haploiden Chromosomensätze? ANGIOSPERMEN: Nachdem der Pollenschlauch seine beiden haploiden Spermakerne aus dem Pollenkorn in den Embryosack entlassen hat, befruchtet der eine Spermakern die Eizelle und bildet die Zygote, der andere Spermakern vereinigt sich mit den beiden Polkernen und bildet im Zentrum der Zentralzelle des Embryosacks einen triploiden Kern (3n). Der triploide Kern teilt sich und bildet eine vielkernige "Superzelle" von milchiger Konsistenz. Diese Masse ,das Endosperm, wird vielzellig und fester, wenn sich durch Cytokinese zwischen den Kernen Membranen und Wände bilden. 3. Wie entsteht aus der Samenanlage ein Samen? Die Zygote teilt sich in eine Terminal- und eine Basalzelle. Aus der Terminalzelle entseht durch gleichmässige Teilung der kugelförmige Proembryo. Dieser entwickelt sich weiter zu einem oder zwei Cotyledonen (Keimblättern). Die Basalzelle teilt sich längs weiter und bildet den Suspensor, welcher den Embryo am Integument der Samenanlage befestigt. DICOTYLEDONEN: Nach der Ausbildung der Hügel, sieht man zwischen den Hügeln den Keimspross. Beim Suspensor sieht man nach der Befestigung die Keimwurzel. COTYLEDONEN: Man unterscheidet noch das Protoderm, Grundmeristem und Procambium. Diese entwickeln sich zu Abschlussgewebe, Grundgewebe und Leitgewebe. Während der Embryo wächst, lagert der Samen Proteine, Fette und Stärke solange ein, bis der Embryo keimt. Vor der Keimung ist der Embryo von einer aus dem Integument der Samenanlage entstandenen Samenschale umgeben. 4. Wo in einer angiospermen Pflanze findet Meiose statt? Welche Produkte werden gebildet? - Diploide Pflanzen (Sporophyt) produzieren meiotisch haploide Sporen. - Die Pollenmutterzellen teilen sich in den Pollensäcken der Antheren zu vier haploiden Mikrosporen aus jeder Mutterzelle. - Die Megasporenmutterzelle teilt sich meiotisch in vier haploide Megasporen. 5. Wie und wo entsteht ein Pollenkorn? Zeichnen sie ein reifes Pollenkorn und geben sie die Zahl und Funktionen der Zellen/Kerne an. In den Pollensäcken, die sich in den Antheren an der Spitze der Staubblätter befinden, gibt es zahlreiche Mikrosporenmutterzellen (diploide Zellen, aus denen die Pollenkörner hervorgehen). Jede Mikrosporenmutterzelle teilt sich meiotisch in vier haploide Megasporen. Nur eine der vier Mikrosporen entwickelt sich durch Mitose zum Pollenkorn (männlicher, unreifer Gametophyt). Er besteht aus einem generativen Kern und einer Schlauchzelle und hat eine dicke, feste Wand. Der Pollen wird zum männlichen, reifen Gametophyten, wenn sich der generative Kern in die beiden Spermakerne teilt. Dies geschieht meist, nachdem der Pollen auf der Narbe des Fruchtblattes gelandet ist und der Pollenschlauch zu wachsen beginnt. 6. Beschreiben sie die Entwicklung des Embryos einer dikotylen Pflanze. (Zeichnung) In der ersten Mitose teilt sich die Zygote durch Querteilung in eine basale und eine terminale Zelle. Durch weitere Teilung der Basalzelle entsteht der Suspensor, der den Embryo an den Integumenten der Samenanlage verankert und die Nährstoffe von der Mutterpflanze an ihn weiterleitet. Durch Teilung der Terminalzelle entsteht der am Suspensor befestigte Embryo. Die Cotyledonen bilden sich als zunächst kleine Schwellungen auf dem Proembryo. Sowohl zwischen ihnen, als auch auf der gegenüberliegenden Seite der Embryonenachse, liegt je ein Apikalmeristem. Die drei primären Meristeme Protoderm, Grundmeristem und Procambium sind ebenfalls bereits im Embryo vorhanden. 7. Was versteht man unter Apomixis? Apomixis ist ein Mechanismus, mit dem Pflanzen (v.a. Korbblütengewächse wie Löwenzahn oder Habichtskraut) ohne Befruchtung ihrer Blüten Samen bilden. In der Samenanlage entsteht aus einer diploiden Zelle ein Embryo, und die Samenanlage reift zu einem Samen. Beim Löwenzahn wird die Frucht (Achäne) durch den Wind davongetragen und der Samen auf diese Weise verbreitet -> Vorteil der Samenverbreitung trotz ihrer ungeschlechtlichen Vermehrung. 8. Beschreiben sie die Stadien der Keimung und der frühen Keimlingsentwicklung einer Erbse. (Zeichnung) Die Radicula, die Wurzelanlage des Embryos, tritt zuerst aus dem Samen aus. Dann durchbricht der Spross die Bodenoberfläche, indem das über den Keimblättern liegende Epicotyl einen Haken bildet. Die Keimblätter verbleiben im Boden. 9. Warum ist die Bezeichnung „Samen“ für ein Maiskorn inkorrekt? Mais ist eine monokotyle Pflanze; welche Struktur entspricht ihrem Keimblatt? Frage 1: Da Mais zu den Poaceen gehört, besteht das Maiskorn aus Pericarp (Fruchtwand) und Samen; Pericarp und Samenschale sind dabei verwachsen. Es handelt sich also um eine Frucht (Schliessfrucht-Nussfrucht) --> Karyopse (da oberständiger Fruchtknoten). Frage 2: Als monokotyle Pflanze besitzt Mais nur ein einzelnes Keimblatt, das Scutellum. Dieses ist sehr dünn, hat aber eine grosse Oberfläche (, was die Absorption von Nährstoffen während der Keimung erheblich verbessert). Der Embryo ist von einer Scheide eingeschlossen. Diese Scheide besteht aus einer Coleorrhiza, welche die Wurzel bedeckt und einer Coleoptile, die den embryonalen Spross einschliesst. 10 Wie sehen die Blüten einer Maispflanze aus und wie findet die Bestäubung statt? Blüten: Mais ist eine monözische Spezies mit Blütenständen aus männlichen und weiblichen Blüten auf demselben Individuum. Der männliche Blütenstand ist als aufgerichtete Rispe sichtbar. Der Maiskolben ist eine Ansammlung von Maiskörnern (einsamige Früchte), der sich aus einem Blütenstand befruchteter weiblicher Blüten entwickelt. Bestäubung: Die männlichen Blüten produzieren den Pollen, der vom Wind weitergetragen wird und die weiblichen Blüten befruchtet, die im unteren Stengelteil sitzen.