neuronale Kommunikation
Werbung

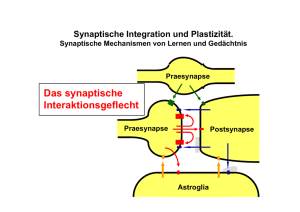
Ruhepotenzial (t2) Potenzialdifferenz zwischen Zellinnerem und Extrazellulärraum =Membranpotenzial, da Potenzialdifferenz über gut isolierender Membran auftritt Konstanter Wert bei ruhender Zelle( zwischen -55 und -100 mV-75 mV) Messung mit Mikroelektroden(intrazellulär) und Bezugselektroden ( extrazellulär) Ursache: Überschuss an neg. Ionen in der Zelle (im Vgl. zu außen) Verschiebung relativ weniger K+ Ionen ( 6 von 100000) im Verhältnis zu Eiweiß-Anionen ( A-) Sehr unterschiedliche Verteilung von Anionen und Kationen innen und außen K+ - Gleichgewichtspotenzial: K+ können Membran gut passieren, da innen mehr K+ Ionen als außen, Wahrscheinlichkeit größer das nach außen diffundieren, als nach innen Nettoausstrom von K+ - Ionen größer Anionen bleiben zurück ( Konzentrationsgefälle von innen nach außen) Dadurch Bildung von Gegen (Halte –) Kraft durch inneres, neg. Potenzial für K+ - Ionen (Ladungsgefälle von außen nach innen) K+ Gleichgewichtspotenzial Instabilität durch Na+-Ionenpermeabilität Zellmembran für Na+ - Ionen weniger durchlässig als für K+ ( 1:25) Konzentrationsgefälle für Na+ von außen nach innen (10:1) + neg. Anziehung von inne Na+ strömt ein ( passiv) Ruhpotenzial wird verringertK+ Ionen strömen nach außen Ständiger Austausch von K+ und Na+ Kein Konstantes Ruhepotenzial durch passive Wege Na+-K+ Pumpe Führt aktiv zur Aufrechterhaltung d. Ruhepotenzialsdynamisches Gleichgewicht von aktiven und passiven Ionen - Strömen Aktionspotenzial Wenn Neuron aktiv werdenkurze, impulsartige positive Änderung des Membranpotenzial Kommunikationsmittel des Nervensystems Amplitude : ca. 110 mV (auf +30mV) Dauer : Skelettmuskulatur : 1ms, Herzmuskel > 200ms einförmige Impulse Kodierung der Information allein durch Frequenz der Aktionspotenziale Ablauf : 1.) Erreichen der Schwelle (-60mV) Alles oder Nichts Prinzip 2.) Aufstrich/ Depolarisationsphase : schnelle, positive Potenzialänderung o Öffnung der Na+ - KanäleEinstrom von Na+ durch kurzzeitige Erhöhung der Membranleitfähigkeit für Na+ 3.) Overshoot/Überschuss : positive Anteil von 1.) 4.) Repolarisation: Rückkehr d. Aktionspotenzials zum Ruhepotenzial o K+ Kanäle öffnen sich vollständig Ausstrom von K+ aus Zelle 5.) Nachpotenziale: Nachschwankungen d. Potenzials 6.) Refrakträzeit: 1-2 ms, in der Neuron nicht erregbar ist Fortleitung des Aktionspotenzials Aktionspotenzial entsteht i.d.R am Übergang von Soma und Axon (=Axonhügel) Leitung in marklosen Axonen/Nervenfasern Elektrischer Spannungsunterschied zwischen erregter und unerregter MembranstelleStrom fließt aus depolarisiertem Gebiet in polarisiertes Gebiet Nachbarbezirk wird ebenfalls zur Schwelle depolarisiertusw. Ausbreitung Richtung Synapse: orthodrom Ausbreitung Richtung Axon : antidrom Ausbreitungsgeschwindigkeit abhängig von Durchmesserje größer Durchmesser, desto schneller (1m/s) Leitung in markhaltigen Axonen/Nervenfasern 100x schneller als bei marklosen Axonen Depolarisierung nur an den Ranvier - Schnürringen springt von Ring zu Ring : saltatorische Erregungsleitung“ Dazwischen liegender Bereich (Internodien) durch Myelin isoliert Synaptische Erregung und Hemmung Chemische Synapsen Verdickung des Axonspräsynaptische Endigung Enthält synaptische Bläschen (Vesikel), die Überträgerstoff ( Transmitter)enthalten Zwischen Synapse und Nachbarzelle: Synaptischer Spalt Gegenüber von Präsynaptischer Endigung: subsynaptische Membran(der postsynaptischen Zelle) mit Rezeptoren für die Überträgersubstanz Beispiel: Motoneuron Soma + Dendriten vollständig von Synapsen bedeckt Axone der präsynaptischen Neurone teilweise Markhaltig Motoaxon markhaltig, endet auf Skelettmuskelfaser Erregendes postsynaptisches Potential ( EPSPs) lokale, graduelle Änderung des Membranpotentials an der postsynaptischen Membran von Nervenzellen, welche ein Aktionspotential im postsynaptischen Element auslöst oder zu dessen Auslösung beiträgt. Reizung von afferenten Nervenfasern vorübergehende Depolarisation des Membranpotenzials Amplitude der Positivierung von Zahl der erregten Afferenzen abhängiggleichzeitige Aktivierung vieler Synapsen nötig um Schwelle zu erreichen Bei Erreichen der Schwelle Aktionspotenzial Entstehung/Zeitverlauf des EPSPs ( am Beispiel des Motonneuron) Anstieg 2ms, Abfall 10-15msunabhängig von Amplitude 1.) Einlaufen des Aktionspotenzials in präsynaptische Endigung 2.) Ca+ Einstrom in präsynaptische Endigung ( durch Depolarisation öffnen der Ca+ Kanäle) 3.) Präsynaptische Glutamat – Freisetzung (Überträgersubstanz/Transmitter) aus Vesikeln (durch Einströmen der Ca+ Ionen) 4.) Glutamat diffundiert durch synaptischen Spalt und verbindet sich mit postsynaptischen GlutamatrezeptorenAktivierung 5.) Öffnen der postsynaptischen IonenkanäleEinstrom von Na, Ca entlang des Diffusionsgradientenfließen von synaptischem Strom 6.) Depolarisation der postsynaptischen MembranEPSP 7.) Beendigung der Transmitterwirkung durch Diffusion ( Transportproteine „pumpen“ Glutamat aus synaptischen Spalt zurück) und Wiederaufnahme des Transmitters(Glutamat) in präsynaptische Endigung EPSP auch an anderen Neuronen im ZNS, dort oft längerer Zeitverlauflangsamere EPSP evtl. an Lernprozess beteiligt Hemmende chemische Synapsen Passiv: in Refraktärphase nach Aktionspotenzial keine Erregung möglich Aktiv: Verminderung der Erregbarkeit eines Neurons :Hemmung, Inhibition 1.) Postsynaptische Hemmung: Herabsetzen der Erregbarkeit der subsynaptischen Soma – und Dendritenmembran der Neurone an axosomatischen und axodendritischen Synapsen 2.)Präsynaptische Hemmung: Reduzierung/Verhinderung der Transmitterfreisetzung an präsynaptischen Endigungen bei axoaxonischen Synapsen Wenn übertragende Synapse vor Übertragung selbst aktiviert wirdReduktion der Übertragung da weniger Trsanmitter Funktion: einzelne afferente Zuflüsse können gezielt gehemmt werden für Empfindlichkeitsverstellung der afferenten Kanäle oder zur Unterdrückung „unerwünschter“ Informationen bzw. Auswahl „erwünschter“ Informationen 1.) Inhibitorische postsynaptische Potenziale (IPSP) Enstehung/Verlauf Reizung von Muskelspindelafferenzen erregt nicht nur eigene Motoneurone sondern hemmt gleichzeitig Motoneurone des Gegenspielers ( Antagonist) Reiz löst hyperpolarisierende Potenzialverschiebung an subsynaptischer Membran aus Verlauf spiegelbildlich zu EPSP Membranpotenzial wird von Schwelle entferntHemmung des Neurons Inhibitorische Transmitter bewirken Öffnung von CI- Kanälen (oder Kaliumkanälen)Einstrom von CI – Ionen (o. Kalium)Erhöhung des Ruhepotenzials ( Zellinnere wird negativer) An anderen Neuronen längere Zeitverläufe möglich Wirkweise 1.) durch HyperpolarisationEntfernung von der Schwelle 2.) während Anstieg des IPSP erhöhte Membranleitfähigkeit durch Öffnung der CIIonenkanäle, die erregenden Strom des EPSP ( Einstrom von Na+) kompensieren ( durch gleichzeitigen CI- Einstrom (oder erhöhten K+ Ausstrom) Neurotransmitter: Entdeckung chemischer Überträgerstoffe in 20er Jahre Benennung der Synapse anhand Substanz, die freigesetzt wird ( bsp Glutamat glutamaterg) Kleine Moleküle Werden im Neuron selbst synthetisiert/hergestellt Sind in Vesikeln gespeichert Bei Aktionspotenzial Freisetzung in synaptischen Spalt Durch Spaltung und/oder Wiederaufnahme (Reuptake) in präsynaptische Endigung inaktiviert, entfernt. Bsp. Azetylcholin (Ach) o Meistens erregender Natur o 10% alles Synapsen im ZNS sind cholinerg ( =benutzen ACh als Transmitter) o Im ANS an parasympathischen und sympathischen Nerven wichtig Bsp. Aminosäuren o Glutamat: Häufigste erregende Transmitter im ZNS o GABA ( Gamma – amino – Buttersäure): häufigster hemmender Transmitter o Glyzin: verbreiteter hemmender Transmitter im Rückenmark und Hirnstamm Bsp. biogene Amine als Transmitter o Katecholamine ( Adrenalin, Noradrenalin und Dopanin) chemisch end miteinander verwandt durch Biosynthese) o =andrenerge Überträgersubstanzen Neuromodulatoren Bei Ausschüttung der Transmitter auch weitere Substanz augeschüttet ( = Kotransmitter Meistens Peptide Aufgabe : Langzeitverstellung der Erregbarkeit ( zu- oder Abnahme) synaptische Modulation Modifikation von Intensität und Dauer der Wirkung von Transmittern Speicherung in Vesikeln (größer als bei kleinmolekularen Transmittern) Freisetzung Kalziumgesteuert, benötigt aber stärkere Aktivierungmehrere Aktionspotenziale in kurzem Abstand Synaptische Interaktion und Plastizität Kurzzeitig: synaptische Bahnung Zeitliche Bahnung: kurz hintereinander folgende EPSPs, möglich, da Dauer der EPSP (15ms) länger als Refraktärzeit (1-2ms) Räumliche Bahnung: gleichzeitige Reizung mehrerer Axone führt zum Überschreiten der Schwelle, was alleine jeweils nicht gereicht hätte Heretrosynaptische Bahnung: Effektivität einer Synapse durch Koaktivierung einer anderen verstärkt Langfristig: Neuronale Plastizität Veränderung der synaptischen Effizienz Tetanische Potenzierung: wiederholte Benutzung einer Synapse führt zur Steigerung der Effizienzschon während der tetanischen Reizung LTP ( Long – term potentiation): Erhöhung der postsynaptischen Glutamat – Rezeptoraffinität + vermehrte präsynaptische Aussschüttung von Glutamat LTD ( long term depression): Verminderung der synaptischen Effizienz