Nervensystem - Institut für Biologie
Werbung

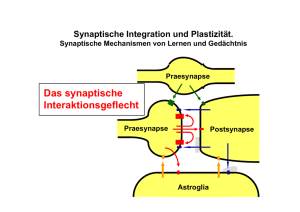
Die Erregung im Nervensystem: - Ruhepotential - Aktionspotential Representative Verlauf der Erregung in einem neuron Neuron ein in Soma Soma “cell body” postsynaptische Seite präsynaptische Seite ou aus t Ionenkanäle geschlossen offen Das Membranpotential hängt von der K+ Konzentration ab E= Ion 58 log [Ion]a [Ion]i hypothetisch (nach Nernst Gleichung) Neuron Elektrische Eigenschaften von Nervenzellen Ruhepotenzial (Membranpotenzial bei Ruhe) Wenn man mit einer Glasmikroelektrode in Zellen einsticht, zeigen sie eine gegenüber dem äußeren Milieu negativ geladene elektrische Spannung (Membranspannung VM, die in Ruhe auch Ruhepotential genannt wird.) Grund: * Zwischen dem Zellinneren und dem Zelläußeren bestehen Unterschiede in den Konzentrationen verschiedener Ionen, insbesondere der Kalium- und Natriumionen [K+]i > [K+]o und [Na+]i < [Na+]o, K+ und Na+ sind also unterschiedlich verteilt. * Die Zellmembran (eine semipermeable-Membran) ist für diese Ionen unterschiedlich durchlässig (z.B. für Kalium recht gut, für Natrium sehr gering) Die Ionen können nur durch entsprechende und für eine Ionensorte spezifische „Poren“, die Ionenkanäle, durch die Membran fließen. Ionenkanäle sind in der Membran der Zelle sitzende Proteinmoleküle, welche dank ihrer hydrophilen Eigenschaften eine solche Porengröße aufweisen, dass das entsprechende Ion mit seiner Hydrathülle genau durchpasst. Man spricht vom: Kaliumkanal, Natriumkanal, Calciumkanal, Chloridkanal, oder von unspezifischen Kationenkanälen (positiv geladene Ionen), Anionenkanälen (negativ geladene Ionen) Für jedes Ion kann der Spannungswert ermittelt werden, bei dem die Ionenflüße im Gleichgewicht sind (Fließgleichgewicht, d.h. es fließen beim Spannungswert des Gleichgewichts, genannt Gleichgewichtspotenzial, genau so viele Ionen in die Zelle oder aus der Zelle; es besteht also keine Nettobewegung von Ionen bzw. Ladungen Berechnung über die von Walter Nernst (1889) aufgestellte Gleichung: Nernst-Gleichung Eion = RT / zF x ln [Ion+]a / [Ion+]i R = allgemeine Gaskonstante, 8,314 JK-1mol-1 T = absolute Temperatur in oK z = Wertigkeit des Ions (1 bei einwertigen Ionen, 2 bei Metallionen wie Ca2+) F = Faraday-konstante, 96500 Cmol-1 ln = Logarithmus naturalis [Ion] = Konzentration des Ions außen (a oder o(ut)) oder innen (i) Bei Zusammenfassung dieser Konstanten und einer Raumtemperatur von 20oC sowie der Ersetzung des natürlichen durch den dekadischen Logarithmus ergibt sich folgende Vereinfachung: Eion = 58 mV x log [Ion+]a / [Ion+]i Membranpotential Vm in mV Gleichgewichtspotenziale der wichtigsten Ionen Na+ ENa = 56 mV 20 0 -20 ClECl = -61 mV K+ EK = -93 mV Zeit, t Aus der Nernst-Gleichung berechnet: EK = 58 mV x log ([Ion+]a ) / ([Ion+]i ) [K+]a / [K+]i [Na+]a / [Na+]i [Cl-]i / [Cl-]a = 10 mM / 400 mM = 460 mM / 50 mM = 40 mM / 450 mM EK = -93 mV ENa = 56 mV ECl = -61 mV Die NatriumKalium Pumpe - Energieverbrauch ATP A T P - 3 Na + raus, 2 K + rein A D ADP + Pi P + P i INNEN - Aufrechterhaltung des Em AUSSEN - (fast) elektrisch neutral Zusammenfassung: Ruhepotenzial Bestimmend für das Ruhepotenzial einer Nervenzelle: * Unterschiedliche Verteilung der Ionen innen und außen (Natrium, Kalium, Chlorid, große organische Anionen können nicht aus dem Zellinnern) * Semipermeable Eigenschaften der Membran (für die Ionen bestehen unterschiedliche Permeabilitäten durch die Kanäle aufgrund selektiver Ionenkanäle) * Natrium-Kalium-Pumpe (Na-K-ATPase) schafft dauernd eingelecktes Natrium aus der Zelle (bei Vergiftung der Pumpe kein Ruhepotenzial möglich) * dadurch Aufrechterhaltung eines negativ geladenen Ruhepotenzials Neuronale Aktivität eines Säugerneurons - intrazelluläre Ableitung - außen innen gK Ströme während des Aktionspotentials Ruhestrom: gleich starker Ein- und Ausstrom von K+ Depolarisation: Na+ EinStrom g Na gK gK Repolarisation: verzögerter K+ Ausstrom Hyperpolarisation: anhaltend stärkere K+ Leitfähigkeit Zusammenfassung Aktionspotenzial Während eines Aktionspotenzials kommt es also zu folgenden Ionenflüssen (Strömen): * Öffnen eines spannungsabhängigen Natriumkanals (beim Schwellenwert hohe Wahrscheinlichkeit des Öffnens) und dadurch große Leitfähigkeit für Natriumionen * Umpolarisation des Membranpotenzials bis zum Erreichen des Natriumgleichgewichtspotentials (ENa). (Dadurch wird das Natrium-gleichgewichtpotenzial (ENa) für das Membranpotenzial bestimmend) * Inaktivierung (Schließen) der spannungsabhängigen Natriumkanäle (dadurch Leitfähigkeit für Natriumionen wieder sehr gering) und Anstieg der Kaliumleitfähigkeit (ein langsamer Prozess). Dadurch wird wieder das Kalium-Gleichgewichtspotenzial (EK) für die Zelle bestimmend. * Aktionspotenziale treten erst auf, wenn das Membranpotenzial einen bestimmten Schwellenwert (immer positiver als das Ruhepotenzial) überschritten hat. * Für einen kurzen Moment polarisiert das Membranpotzenzial um, das heisst das Innere wird positiv * Die Spitze des Aktionspotenzials liegt bei Spannungswerten, die in etwa denen für das Natrium-Gleichgewichtspotenzial entsprechen (ENa) * Danach repolarisiert das Membranpotenzial sofort und fällt ín etwa auf den Wert des Ausgangspotenzials (Ruhepotenzials) ab. * Nach einem Aktionspotential kann nicht sofort ein neues ausgelöst werden (Absolute Refraktärperiode). * Nach Beendigung dieser absoluten Refraktärperiode können zwar wieder Aktionspotenziale ausgelöst werden, ihre Amplitude ist aber geringer (das heisst die Spitze des AP liegt bei geringeren Spannungswerten) (relative Refraktärperiode) Fortleitung von Aktionspotenzialen Aktionspotenziale werden "aktiv" fortgeleitet - schnell - ohne Abschwächung - ohne "Informationsverlust" - in eine Richtung Fragen: - Welche Konzentrationen der K+, Na+ und Cl- Ionen herrschen in Neuronen während des Ruhepotentials? Welche Membranspannung Herrscht während des Ruhepotentials? - Was versteht man unter dem Gleichgewichtspotentials eines Ions? - Welche Art von Ionenkanälen gibt es? - Was sind die Voraussetzungen für die Ausbildung des Ruhepotentials? -Welche Ionenkanäle sind für das Aktionspotentials verantwortlich? -Welche Ströme fließen während des Aktionspotentials? -Erklären Sie die Ursache für die absolute und relative Refraktärzeit. -Wie wirkt sich die Refraktärzeit auf die maximal mögliche Frequenz von Aktionspotentialen aus? Synapsen und synaptische Integration: Wie rechnet das Gehirn? Kontaktstellen zwischen Neuronen, oder zwischen Neuronen und Muskel (neuromuskuläre Synapse) Entsprechend der Art ihrer Übertragung unterscheidet man elektrische oder chemische Synapsen. Eine Synapse besteht aus einem präsynaptischen Teil (Präsynapse) und einem postsynaptischen Teil (Postsynapse). Neurone kommunizieren über Synapsen Synapse Chemische Synapsen * Synaptischer Spalt etwa 20 - 40 nm, * Präsynaptische Zelle mit Vesikeln setzt Botenstoff (Neurotransmitter) frei, der durch den synaptischen Spalt zur postsynaptischen Zelle diffundiert und dort an spezifische Rezeptormoleküle bindet. Menge des freigesetzten Transmitters abhängig von der Amplitude des Membranpotenzials! * Chemische Synapsen sind gleichrichtend (Leitung nur in einer Richtung) und arbeiten mit einer Zeitverzögerung (delay) von etwa 1 ms. •Die postsynaptische Zelle enthält auf der Membran zum synaptischen Spalt Rezeptormoleküle, an denen die Transmitter binden. Durch die Bindung ändern sich diese Rezeptormoleküle und es kommt zu einem Ionenstrom. Dieser führt zu einem postsynaptischen Potenzial. Vesikel fusionieren mit der präsynaptischen Membran an aktiven Zonen. Für die Fusion ist ein Einstrom von Ca2+ nötig. Diese enthalten Komplexe von Proteinen und spanungsabhängige Ca-Kanäle Reaktionskaskade bei der synaptischen Übertragung 1) In der Präsynapse (Axonterminal) ankommendes AP führt zu Calciumionen-Einstrom, damit Fusion der Vesikel mit der Zellmembran in der aktiven Zone möglich. 2) Vesikel enthalten Transmitter (Durchmesser etwa 50 nm, mit 10 000 – 50 000 Transmittermolekülen), Exocytose der Transmittermoleküle und Diffusion durch den etwa 30 nm breiten, mit extrazellulärer Matrix gefüllten synaptischen Spalt. 3) Bindung der Transmittermoleküle an spezifische Rezeptormoleküle der postsynaptischen Zelle, dadurch Aufbau eines postsynaptischen Potenzials (erregend: EPSP oder hemmend (inhibitorisch): IPSP). 4) Wirkung des Transmitters wird durch raschen enzymatischen Abbau begrenzt, Aufnahme der Abbauprodukte oder auch ganzer Moleküle durch Gliazellen oder in die präsynaptische Zelle durch spezifische Transportmoleküle (Transporter) 5) Vesikel werden recycelt (Bildung eines Endosoms in der Präsynapse, Bildung neuer Vesikel) (Würde das nicht geschehen, würde die Grösse der Präsynapse dauernd anwachsen, da ja ständig Vesikel mit der präsynaptischen Zellmembran verschmelzen). Neurotransmitter Acetylcholin (neuromuskuläre Synapsde der Wirbeltiere, autonomes Nervensystem Biogene Amine Noradrenalin, Adrenalin (Catecholamine), Serotonin (5-Hydroxytryptamin, 5-HT), wichtige Transmitter im Gehirn Aminosäuren (Gamma) g-Aminobuttersäure (GABA), Glycin, Glutamat (neuromuskuläre Synapse der Wirbellosen Tiere, und wichtiger Transmitter im Gehirn) Peptide FMRF-amid, Opioide: Opiocortine, Enkephaline, Dynorphin (endogene, körpereigene Opiate) Peptide der Neurohypophyse: Vasopressin, Oxytocin, Neurophysine Tachykinine: Substanz P, Insuline: Insulin, insulinähnliche Wachstumsfaktoren I und II Somatostatine: Somatostatin, Polypeptide der Bauchspeicheldrüse Gastrine: Gastrin, Cholecystokinin Gasförmige Transmitter Stickoxid (NO) Kohlenmomoxid (CO) Ionotroper Rezeptor z.B. nikotinischer ACh-rezeptor Metabotroper Rezeptor z.B. muskarinischer ACh-rezeptor Der nikotinische Acteylcholin Rezeptor (nAChR) besteht aus 5 Einheiten (α – γ), jede Einheit aus 4 in der Membran befindliche α Helices (TM1 – TM4). Die ACh Bindungsstelle befindet sich in jeder der beiden α Einheiten Pore für Kationen durchlässig Der wichtigste erregende Rezeptor im Wirbeltiergehirn ist der Glutamat Rezeptor. Es gibt mehrere Typen, ionotrope und metabotrope. Die Rezeptortypen werden nach den Substanzen benannt, die als Antagonisten wirken, z.B. NMDA: N-Methyl-D-Aspartat Excitatorische Postsynaptische Potentiale von ionotropen Rezeptoren (EPSP) schnell, direkt und von metabotropen Rezeptoren (EPSP) Erinnerung: Bildung des Rezeptorpotentials im Auge: Rhodopsin! langsam G-Protein vermittelt * Ob ein Neurotransmitters erregend oder hemmend wirkt, hängt ausschliesslich von der Art der postsynaptischen Rezeptormoleküle ab: erregend: in der postsynaptischen Zelle wird ein EPSP (erregendes postsnaptisches Potential) gebildet hemmend: es wird ein IPSP (inhibitorisches postsynaptisches Potential) gebildet * Eine Nervenzelle kann mehr als einen Transmitter freisetzen (oft klassischer Transmitter und ein bis mehrere Co-Transmitter). Verrechnung (Integration) an Synapsen: Addition und Subtraktion erregende Synapse Axonhügel (Aktionspotential auslösende Zone) Laborjargon: Spike-generierende-Zone spike initiating zone hemmende Synapse Membranpotential (mV) In diesem Beispiel heben sich Erregung und Hemmung gerade auf ! 20 0 -20 EPSP IPSP Zeit (ms) Verrechnung (Integration) an Synapsen: Zeitliche Summation Verrechnung (Integration) an Synapsen: Räumliche Summation Verrechnung (Integration) an Synapsen: Räumliche Summation EPSPs/IPSPs verschiedener Synapsen, die z. B. an einem Dendritenbaum ansetzen, werden In der postsynaptischen Zelle zu jedem Zeitpunkt addiert Zeitliche Summation Die in einer Präynapse zeitlich kurz aufeinanderfolgenden Aktionspotentiale lösen in der postsynaptischen Zelle EPSPs/IPSPs aus, welche addiert werden. Für die Integration sind die passiven elektrischen Eigenschaften (Kabeleigenschaften) des postsynaptischen Neurons sehr wichtig (Konstanten τ und λ). Präsynaptische Hemmung an Synapsen Das präsynaptische Neuron wird durch Freisetzung eines Transmitters (vorwiegend GABA oder Glycin) gehemmt, und damit wird die Freisetzung des Transmitters aus der Präsynapse verhindert). Ein Mechanismus, der z.B. die Axonterminale von Sinneszellen differenziert hemmen kann, und damit die sensorischen Signale unterdrückt. Präsynaptisches Neuron (Axonterminal) Präsynaptische Hemmung Inhibitorisches (Hemm) Neuron Postsynaptische Hemmung Postsynaptisches Neuron Bahnung (Facilitation) Modulation an Synapsen Eines von beiden, oder beide, das präsynaptische und das postsynaptische Neuron werden durch einen von einem dritten Neuron freigesetzten Transmitter beeinflusst. Dieser als Neuromodulator bezeichnete Botenstoff hat selbst keine rasche Wirkung auf die Neurone, sondern die normale Übertragung zwischen den beiden Neuronen wird verändert (z.B. effizienter). Eine Modulation kann auch dadurch stattfinden, dass das präsynaptische Neuron Co-transmitter (beispielsweise nach hochfrequenter AP-Salve) freisetzt, die dann die weitere synaptische Übertragung beeinflussen, oder im postsnaptischen Neuron Signalkaskaden aktiviert werden, die dann zu langanhaltenden Veränderungen der neuronalen Eigenschaften bis hin zur Genexpression führen (Lernen, Gedächtnis) Fragen: -Welche Transmitter kennen Sie? -Wie steuert die präsynaptische Erregung die Transmitterausschüttung? -Welche postsynptischen Rezeptormoleküle kennen Sie? -Wie bewirket die Bindung des Transmitters an seinen Rezeptor die -Postsynaptische Erregung? -Welche Arten von postsynaptischer Erregung gibt es und welche Ionenflüsse -Sind dafür verantwortlich? -Wie werden postsynaptische Potentiale integriert? -Wie breiten sich postsynaptische Potentiale im Bereich der Dendriten aus?