Uebungsaufgaben II, Lo%CC%88sungsblatt, 2012
Werbung

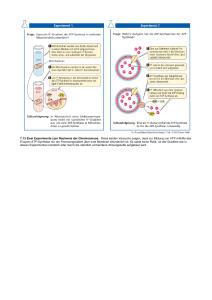
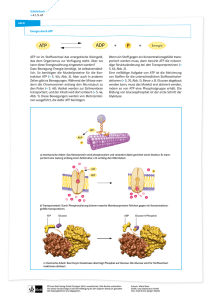
Uebungsaufgaben mit Lösungen zum Vorlesungsteil Metabolismus (Citronensäurezyklus, Oxidataive Phosphorylierung, Glycogenstoffwechsel) 1) Malat-dehydrogenase katalysiert die Regenerierung von Oxalacetat im Citronensäurezyklus. Das G0’ dieser Reaktion ist allerdings mit + 29,7 kJ mol-1 energetisch ungünstig, und darum ist die Konzentration von Oxalacetat verglichen mit Malat relativ niedrig. Warum wird kann dieser Schritt in der Zelle trotzdem ablaufen? Begründen Sie Ihre Antwort. Der Citratcyklus wird durch die stark exergone Citrat-synthase reaktion (G0’ = - 31,5 kJ mol-1, Thioesterspaltung) getrieben. 2) Wo laufen die Reaktionen des Citronensäurezyklus ab? Die Reaktionen des Citronensäurezyklus laufen im der mitochondrialen Matrix ab. Daher muss Pyruvat in Mitochondrien transportiert werden. 3) Nennen Sie die zwei mobilen Elektronenträger der mitochondrialen Elektronentrabsportkette. Als mobile Elektronenträger funktionieren membranlösliches Ubiquinon (Coenzym Q) und der periphere Elektronenträger Cytochrom C. 4) Wie wird der mitochondriale Protonengradient für die ATPSynthese genutzt? Beschreiben Sie kurz den Vorgang. Der Protonenfluss treibt Konformationsänderungen der ATPsynthase an. In der offenen Konformation der ATP-synthase wird ATP freigesetzt, während in der L-form ADP und Phosphat binden können. Die Konformationsänderung zur T-form erlaubt dann die eigentliche ATP-katalyse. Die zum Antrieb der koordinierten Konformationswechsel erforderliche Energieübertragung erfolgt über die Protonen-getriebene Rotation. 5) Organismen mit aeroben Stoffwechsel benötigen Superoxiddismutase. Warum? Obwohl die Uebertragung von Elektronen auf Sauerstoff in der Cytochrom C-oxidase mit erstaunlicher Geschwindigkeit und Genauigkeit abläuft, können als Nebenreaktion trotzdem gefährliche Sauerstoffradikale austreten. Diese werden von der Superoxiddismutase zu H202 umgewandelt und anschliessend von der Katalase als Sauerstoff und Wasser freigesetzt. 6) Warum liefert NADH aus der Glykolyse weniger ATP als NADH aus dem Zitronensäurezyklus? Da die Glykolyse im Cytoplasma abläuft, muss NADH aus der Glyceraldehydphosphat-dehydogenase Reaktion zuerst in die mitochondriale Membran eingeschleust werden. Dies geschieht mittels Glyceraldehyd-3-phosphat-shuttle, der die Elektronen in drei Schritten auf FADH2 in Mitochondrien überträgt. Dadurch treten die Elekronen aber erst durch die Succinat-Q-reduktase in die Atmungskette ein, wodurch die Protonenpumpe des Komplex I umgangen wird, und daher weniger ATP durch indirekte Synthese gewonnen werden kann. 7) Wie erhalten Mitochondrien den Protonengradienten unter anaeroben Bedingungen aufrecht? Die ATP-synthase kann unter aeroben Bedingungen dank dem Protonenfluss ATP-synthese betreiben. Die ATP-synthase kann aber auch in umgekehrter Richtung arbeiten, d.h. unter ATP Verbrauch Protonen durch die mitochondriale Membran pumpen. Dadurch kann der Protonengradient auch unter anaeroben Bedingungen aufrechterhalten werden. 8) Was unterscheidet langsame (Typ I) und schnelle (Typ II) Muskelfasern? Was bedeutet dies für den Energiehaushalt der zwei Muskeltypen? Begründen Sie Ihre Antwort. Diese Muskelfasern unterscheiden sich vor allem durch die Anzahl Mitochondrien. Schnelle Muskelfasern enthalten wenige Mitochondrien, da sie ATP möglichst schnell mittels Glykolyse anaerob herstellen müssen. Pyruvat wird nachher vor allem zu Lactat reduziert, und ausgeschieden. Langsame Muskelfasern verbrennen Zucker meist aerob, d.h. Pyruvat wird zu Acetyl-CoA abgebaut , das dann im Citronensäurezyklus veratmet wird. Die gewonnenen Elektronen werden durch die Atmungskette als Protonengradient in Mitochondrien gespeichert, der dann wiederum für die indirekte ATP synthese ausgenutzt wird. Dadurch erzielen langsame Muskelfasern eine höhere ATPAusbeute, allerdings auf Kosten der Geschwindigkeit. 9) Krebszellen leiden oft unter Sauerstoffmangel. Wie decken sie trotzdem den ATP-Bedarf? Warum verbrauchen sie dabei viel mehr Glukose als normale Zellen mit genügend Sauerstoff. Krebszellen decken ihren ATP-Bedarf vorallem durch anerobe Glykolyse. Da dieser Stoffwechselweg aber nur die Synthese von zwei ATP-Molekülen pro Glucose erlaubt, muss etwa 100 mal mehr Zucker verbraucht werden als durch die aeroben Abbauwege. 10)Der Andersenkrankheit liegt ein genetischer Defekt der Amylo(1,4 -> 1,6)- Transglykosylase (Verzweigungsenzym) zu Grunde. Dagegen haben Patienten mit McArdle-Krankheit einen Mangel an Muskel-phosphorylase. Beschreiben Sie kurz die erwartete Glykogenstruktur in den Muskelzellen der zwei Patienten. Was für Auswirkungen erwarten Sie bei erhöhtem ATP-Bedarf durch Muskelanstrengung in den beiden Patienten? Begründen Sie kurz Ihre Antworten. Anderson-Krankheit: Durch den Defekt des Verzweigungsenzyms weist die Glykogenstruktur viele unverzweigte Ketten auf. Die Löslichhkeit des Glykogens ist daher herabgesetzt, und es kann dadurch weniger Zuckermoleküle in dieser Form gespeichert werden. Bei erhöhter Muskelanstrengung können durch die Phosphorylase weniger Glucoseeinheiten abgespalten werden, was sich in einem ATP-Mangel äussert. McArdle-Krankheit: Die Glykogenstruktur von McArdle Patienten erscheint normal, da die benötigten Enzyme für den Glykogensynthese effizient arbeiten können. Allerdings kann der ATP-Bedarf bei Anstrengung nur ungenügend gedeckt werden, da die defekte Phosphorylase Glucoseeinheiten nicht entsprechend von den nicht-reduzierenden Enden des Glycogens abspalten kann. 11)Welches Trägermolekül wird gebraucht um Glucose für die Glykogensynthese zu aktivieren? Wie wird die dazu nötige Energie gewonnen? Die Glycogensynthese erfordert aktivierte Glucose in Form von Uridin-diphosphat-glucose. Die benötige Energie wird durch die Abspaltung und anschliessende Hydrolyse von Pyrophosphat erzeugt. 12) Welche aktivierten Trägermoleküle übertragen welche chemische Gruppe in hochenergetischer Verknüpfung? a. b. c. d. e. f. a–6 b–4 c–2 d–5 e–3 f-1 Acetyl-CoA S-adenosinemethionin ATP Carboxyliertes-Biotin NADH, NADPH, FADH2 Uridine diphosphate glucose 1. Glucose 2. Phosphat 3. Electronen und Wasserstoff 4. Methyl group 5. Carboxyl-Gruppe 6. Acetyl-Gruppe