Kapitel 14
Werbung

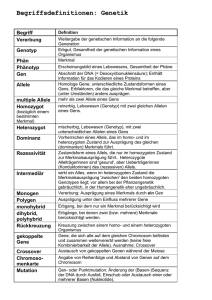
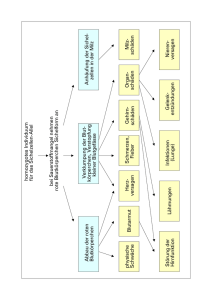
Kapitel 14 Mendel und die Gen Hypothese homozygot = reinerbig heterozygot = mischerbig Mendel führte das Experiment und die quantitative Auswertung in die Genetik ein Mendel führte im 19. Jahrhundert Versuche mit Erbsen durch. Er entdeckte die grundlegenden Prinzipien der Vererbung, indem er in sorgfältig geplanten Experimenten Gartenerbsen züchtete. Er fand dabei die 3 Mendelregeln: Spaltungsregel, Unabhängigkeitsregel und Uniformitätsregel (ist nur von untergeordneter Bedeutung und hat nicht den vollwertigen Status eines genetischen „Gesetzes“). Mendel wählte wahrscheinlich die Gartenerbse als Versuchspflanze, weil man viele Erbsensorten bekommen konnte. Eine Sorte hatte purpurfarbene Blüten, während eine andere weisse Blüten besass. Genetiker benutzen den Begriff Merkmal für eine erbliche Eigenschaft. Jede Variante dieses Merkmals, wie z. B. purpurfarbene oder weisse Blüten, bezeichnet man als Merkmalsform. Bei der Gartenerbse konnte Mendel die Befruchtung kontrollieren. Normalerweise bestäuben die Pollenkörner gleich die benachbarte Narbe in derselben Blüte Selbstbefruchtung. Mendel beabsichtigte aber Fremdbefruchtung (Befruchtung durch eine andere Pflanze) und entfernte deshalb die unreifen Staubblätter bevor sie Pollen produzierten und bestäubte die von Staubblättern befreite Blüte mit dem Pollen einer anderen Pflanze. So war er sich stets sicher über die Herkunft der Erbsensamen. Zudem verliess er sich nur auf eindeutige Merkmale („entweder oder“ nicht „mehr oder weniger“). Er achtete darauf, dass er seine Experimente mit reinerbigen Rassen begann; bei solchen weist nach Selbstbefruchtung die ganze Nachkommenschaft dieselben Merkmalsformen auf. Campbell deutsch: S. 262/ 263 Abb. 13.1 + 13.2, engl.: S. 248/ 249 Abb. 14.1 + 14.2 Die Paarung oder Kreuzung zweier Varietäten wird Hybridisierung genannt. Die daraus resultierenden mischerbigen Individuen sind Hybriden. Bei einer monohybriden Kreuzung geht es nur um die Vererbung eines einzigen Merkmals. Die reinerbigen Elternpflanzen werden als PGeneration bezeichnet, die hybriden Nachkommen als F1-Generation. Wenn man bei den F1Hybriden Selbstbefruchtung zulässt, entsteht die F2-Generation. Mendel verfolgte die Vererbung einer Merkmalsform über drei aufeinanderfolgende Generationen: die P-, die F1- und die F2-Generation. Hätte er seine Experimente nach der F1-Generation abgebrochen, wären ihm die grundlegenden Gesetzesmässigkeiten verborgen geblieben. Nach der „Spaltungsregel“ gelangen die beiden Allele für ein bestimmtes Merkmal in getrennte Gameten Mendel zog grosse Mengen an Versuchspflanzen und protokollierte seine Experimente sehr genau. Dabei ergab sich ein Verhältnis von 3:1 in der F2-Generation (75% purpurfarbene und 25% weisse Blüten). In den Hybriden beeinflusste ausschliesslich der Faktor für purpurfarbene Blüten die Blütenfarbe, der Erbfaktor für weisse Blüten prägte sich nicht aus. Daraus folgt, dass purpurfarben dominant ist und weiss rezessiv. Mendel beobachtete das gleiche Vererbungsmuster bei sechs weiteren Merkmalsformen. Campbell deutsch: S. 264 Tab. 13.1, engl. S. 250 Tab. 14.1 Mendels Interpretation der Ergebnisse: 1. Alternative Zustandsformen (Allele) eines Gens bedingen die genetische Variabilität bei Erbmerkmalen. z.B. traten zwei Versionen von Blütenfarben auf, eine violette und eine weisse. Solche alternative Versionen werden als Allele eines Gens bezeichnet. 2. Für jedes Merkmal besitzt ein Organismus zwei Allele, je eines von jedem Elternteil. 3. Wenn die beiden Allele unterschiedlich sind, dann wird eines, und zwar das dominante Allel voll exprimiert; das andere, das rezessive Allel, zeigt keinerlei Ausprägung. Man spricht vom dominant rezessiven Erbgang. 4. Eine Eizelle oder eine Spermazelle erhält nur eines der beiden Allele, die in zwei Kopien in den Somazellen des betroffenen Organismus vorliegen. Auf die Chromosomen bezogen entspricht diese Trennung oder Segregation der Reduktion der Chromosomenzahl vom diploiden zum haploiden Satz während der Meiose. Wegen der Dominanz oder der Rezessivität von Allelen enthüllt das Erscheinungsbild eines Organismus nicht immer seine genetischen Anlagen. Es muss immer zw. dem Phänotyp (Erscheinungsbild) und dem Genotyp (genetische Grundlage) eines Organismus unterschieden werden. Rückkreuzung: Um feststellen zu können, ob es sich um einen reinerbigen oder mischerbigen Organismus handelt, wird eine Testkreuzung eines rezessiven Homozygoten mit einem Organismus mit dominantem Phänotyp, aber unbekanntem Genotyp durchgeführt, eine sogenannte Rückkreuzung. Sie wurde von Mendel eingeführt und ist immer noch eine wichtige Methode der Genetik. Campbell deutsch: S. 267 Abb. 13.6, engl.: S. 252 Abb. 14.6 Mendel leitete seine Spaltungsregel aus monohybriden Kreuzungen ab. Nach der „Unabhängigkeitsregel“ segregieren bei der Gametenbildung die verschiedenen Allelpaare unabhängig voneinander Dihybride Kreuzung: Was wäre das Ergebnis einer Kreuzung, bei der sich die Elternrassen in zwei Merkmalen unterscheiden – einer dihybriden Kreuzung? Hypothese 1: die Merkmale werden gemeinsam vererbt Hypothese 2: die Merkmale werden unabhängig vererbt Mendel studierte sieben Erbmerkmale von Erbsen in versch. dihybriden Kombinationen und erhielt in der F2-Generation stets die Aufspaltung 9:3:3:1. Jedes Merkmal wird unabhängig vom anderen vererbt. Campbell deutsch: S. 268 Abb. 13.7, engl.: S. 253 Abb. 14.7 Was die einzelnen Erbmerkmale betrifft, gleicht das Segregationsverhalten der Gene der monohybriden Kreuzung. Die Unabhängigkeitsregel ist das 2. Mendelsche Gesetz. Auch „Regel von der Neukombination der Gene“ genannt, da die unabhängige Segregation eine genetische Neukombination ist. Die Mendelsche Genetik beruht auf den Gesetzen der Wahrscheinlichkeit Mendels genetische Gesetze der Spaltung und der unabhängigen Segregation sind spezielle Anwendungen der allg. Regeln der Statistik. Die Wahrscheinlichkeit für ein bestimmtes Ereignis variiert zw. 0 und 1. Ein Ereignis, das mit absoluter Sicherheit abläuft, hat die Wahrscheinlichkeit 1, ein Ereignis, das mit Sicherheit nicht stattfindet die Wahrscheinlichkeit 0. Das Ergebnis eines Versuchs ist unabhängig von den Ergebnissen früherer Versuche unabhängige Ereignisse. Die Multiplikationsregel: Man muss die Wahrscheinlichkeiten für jedes Ereignis berechnen und dann die Einzelwahrscheinlichkeiten miteinander multiplizieren um die Gesamtwahrscheinlichkeit für die Kombination der Ereignisse zu ermitteln. Nach der Multiplikationsregel ist die Wahrscheinlichkeit, dass zwei Münzen mit dem Kopf nach oben landen ½ * ½ = ¼. Eine monohybride Kreuzung der F1-Generation entspricht diesem Beispiel mit den Münzen. Die Multiplikationsregel kann auch auf dihybride Kreuzungen angewendet werden. Campbell deutsch: S.270 Abb. 13.9, engl.: S. 254 Abb. 14.8 Die Additionsregel: Wie gross ist die Wahrscheinlichkeit für eine F2-Pflanze aus einer monohybriden Kreuzung heterozygot zu sein? Es gibt zwei Möglichkeiten für F1-Gameten heterozygot zu werden. Das dominante Allel kann aus der Eizelle und das rezessive Allel aus dem Pollenkorn stammen, oder umgekehrt. Die Wahrscheinlichkeit, dass ein Ereignis durch zwei oder mehr unterschiedliche Ereignisse eintreten kann, ist die Summe der Wahrscheinlichkeiten dieser beiden Ereignisse. So ist die Wahrscheinlichkeit für einen Heterozygoten in der F2-Generation ¼ + ¼ = ½. Wie man die Statistikregeln benutzt, um genetische Probleme zu lösen: Campbell deutsch: S. 271 Beispiel, engl.: S. 255 Wie Mendel entdeckte verhalten sich Erbmerkmale wie diskrete Teilchen Die beiden Mendelschen Gesetze, Spaltungsregel und Unabhängigkeitsregel, erklären die erbliche Variabilität durch alternative Versionen (Allele) von „Erbteilchen“ (Genen); diese werden nach einfachen statistischen Regeln von Generation zu Generation weitergegeben. Diese Teilchentheorie der Vererbung, zuerst an der Gartenerbse entdeckt, gilt genauso für Feigen, Fliegen, Fische, Vögel und Menschen. Die Beziehung zwischen Genotyp und Phänotyp ist in der Regel nicht einfach Unvollständige Dominanz: Für einige Merkmalsformen gilt unvollständige Dominanz. Hier zeigen die F1-Hybriden ein Erscheinungsbild, das zw. den beiden elterlichen Phänotypen liegt, also intermediär ist intermediärer Erbgang. Kreuzt man beispielsweise ein rotes Löwenmäulchen mit einem weissen, so haben alle F1-Hybriden rosa Blüten ( unvollständige Dominanz). Die Kreuzung von F1Hybriden führen zu einer F2-Nachkommenschaft, in der die Phänotypen rot zu rosa zu weiss im Verhältnis 1:2:1 auftreten. Das Verhältnis der Genotypen und der Phänotypen ist in der F2Generation das gleiche. Campbell deutsch: S. 272 Abb. 13.10, engl.: S. 256 Abb. 14.9 Was ist ein dominantes Allel?: Was ist ein dominantes Alles, wann ist ein Allel nicht dominant? Im Falle der unvollständigen Dominanz (Mendel) unterscheiden sich die Phänotypen der Heterozygoten nicht von den dominanten Homozygoten. Bei der Kodominanz exprimieren beide Allele unabhängig voneinander ihren Phänotyp. Ein Beispiel dafür sind die drei Blutgruppen des Menschen, die man mit M, N und MN bezeichnet (das MNSystem hat nichts mit dem AB0-System zu tun). Der MN-Phänotyp ist kein Zwischending zw. den Phänotypen M und N, sondern beide Phänotypen sind unabhängig voneinander verwirklicht. Die Beziehung von Allelen kann sich in vollständiger Dominanz, Kodominanz oder versch. Graden unvollständiger Dominanz äussern. Diese Variationen spiegeln sich in den Phänotypen der Heterozygoten wider. Beispiel: Der für die Tay-Sachs-Krankheit (Erbkrankheit des Menschen) typische Enzymdefekt ist auch bei den Heterozygoten festzustellen. Diese zeigen eine Aktivität des betreffenden Enzyms, die zw. der von gesunden und kranken Individuen liegt. Heterozygote weisen die Symptome der Krankheit nicht auf. Die Beziehung von Dominanz und Rezessivität ist selten so klar und einfach wie in Mendels Beispielen. Es ist auch wichtig, zu verstehen, dass ein Allel nicht deshalb als dominant bezeichnet wird, weil es das rezessive in irgendeiner Weise „unterdrückt“ oder verändert. Allele stellen einfach Variationen in der Nucleotidsequenz eines Gens dar. Wenn wir nach den biochem. Grundlagen forschen, erscheint das Konzept von Dominanz und Rezessivität gar nicht mehr so geheimnisvoll. Wenn ein Allel dominant ist, bedeutet das nicht notwendigerweise, dass es in einer Population häufiger auftritt als das rezessive Allel für dieses Merkmal. Die natürliche Selektion wirkt sich auf die relativen Allelhäufigkeiten in einer Population aus. Zusammenfassung der drei wichtigsten Punkte über die Beziehung zw. Dominanz und Rezessivität: 1. Diese Beziehungen reichen von der vollständigen Dominanz über versch. Grade der unvollständigen Dominanz bis hin zur Kodominanz. 2. Dominanz und Rezessivität ergeben sich erst auf der Ebene des Phänotyps; sie beruhen nicht etwa darauf, dass ein Allel das andere auf dem Niveau der DNA unterdrückt. 3. Die Häufigkeit eines Allels in einer Population ist von seiner Dominanz oder Rezessivität unabhängig. Multiple Allele: Die meisten Gene kommen in mehr als zwei allelen Formen vor. Das AB0-Blutgruppensystem des Menschen ist ein Beispiel für multiple Allele eines einzigen Gens. Es gibt vier versch. Phänotypen für dieses Merkmal. Für Bluttransfusionen ist es wichtig, die richtige Blutgruppe zu wählen. Wenn das Spenderblut einen Faktor (A oder B) trägt, der für den Empfänger fremd ist, so besitzt dieser Antikörper, welche die fremden Moleküle binden und die Blutzellen des Spenders zum Agglutinieren (Verklumpen) bringen. Durch diese Verklumpung kann der Empfänger sterben. Campbell deutsch: S. 273 Abb. 13.11, engl.: S. 257 Abb. 14.10 Pleiotropie: Bei der Mendelschen Genetik sind wir bislang davon ausgegangen, dass nur ein Gen für einen bestimmten Phänotyp verantwortlich ist. Die meisten Gene aber zeigen multiple phänotypische Ausprägungen. Die Befähigung eines Gens, den Phänotyp eines Organismus in vielfacher Weise zu beeinflussen, wird als Pleiotropie (griech. pleion = mehr) bezeichnet. Ein Genprodukt hat viele Wirkorte im Organismus. Epistase: In einigen Fällen verändert ein Gen die phänotypische Ausprägung eines anderen – ein Vorgang, der als Epistase bezeichnet wird (griech. epistateo = „die Aufsicht haben“). Beispiel: Bei Mäusen und vielen anderen Säugetieren ist schwärze Fellfarbe dominant über braune. Die beiden Allele für dieses Merkmal werden mit B und b bezeichnet. Der Genotyp einer Maus mit braunem Pelz muss also bb sein. Aber ein zweites Gen regelt, ob in den Haaren Pigment eingelagert wird oder nicht. Das Produkt eines Gens bestimmt, ob das Fell schwarz oder braun wird, abhängig von den Allelen B und b. Wenn die Maus homozygot rezessiv ist für das zweite Gen (cc), so ist die Fellfarbe weiss, die Maus ist dann ein „Albino“, unabhängig vom jeweiligen Allel für die Fellfarbe (B oder b). Die beiden Gene erden getrennt vererbt dihybride Kreuzung der F1Generation. Campbell deutsch: S.274 Abb. 13.12, engl.: S. 258 Abb. 14.11 Polygene Vererbung: Es gibt eine grosse Zahl von Merkmalen, inklusive der Hautfarbe und der Körpergrösse des Menschen, bei denen eine Entweder – Oder – Klassifizierung (Mendel) nicht möglich ist, weil die Merkmale einer Population ein Kontinuum bilden (also graduell sind). Man bezeichnet solche Merkmale als quantitative Merkmale. Variabilität in der Quantität deutet in der Regel auf polygene Vererbung hin: zwei oder mehr Gene wirken zusammen, um einen Phänotyp hervorzubringen. Polygene Vererbung ist das Gegenteil von Pleiotropie. Die Hautpigmentierung beim Menschen wird von mindestens drei unabhängigen Genen kontrolliert. Umweltfaktoren, wie z. B. die Sonneneinstrahlung beeinflussen den Phänotyp der Haut natürlich auch. Erbanlagen und Umwelt; der Beitrag der Umwelt zum Phänotyp: Der Phänotyp ist sowohl vom Genotyp als auch von der Umwelt abhängig. Ein Baum, dessen Erscheinungsbild im Prinzip von seinem Genotyp bestimmt ist, zeigt dennoch Blätter unterschiedlicher Grösse, Gestalt und Grüntönung, je nach Exposition gegenüber Sonne und Wind. Beim Menschen beeinflusst die Ernährung di Körpergrösse, Training wirkt sich auf Körperbau aus, Sonneneinstrahlung dunkelt die Haut und durch Übung steigert man die körperliche und geistige Leistungsfähigkeit. Selbst eineiige Zwillinge, die genetisch identisch sind, zeigen phänotypische Unterschiede, die auf ihre individuellen Lebensumstände zurückzuführen sind. Aus einem bestimmten Genotyp lässt sich nicht ein genau definierter Phänotyp ableiten, sondern es existiert eine phänotypische Bandbreite, die von Umwelteinflüssen abhängig ist. Diese Bandbreite des Phänotyps wird als Reaktionsnorm des Genotyps bezeichnet. Es gibt Fälle, bei denen die Reaktionsnorm gleich null ist; dann lässt der Genotyp nur einen ganz bestimmten Phänotyp zu. Im Allg. zeigen polygen vererbte Merkmale die breiteste Reaktionsnorm. Genetiker bezeichnen solche Merkmale als multifaktorell (viele Faktoren, genetische und umweltbedingte, wirken zusammen auf den Phänotyp ein). Vererbung und Variabilität aus der Sicht Mendels: Der Begriff Phänotyp hat zwei Bedeutungen. Er beschreibt nicht nur spezifische Erbmerkmale sondern beschreibt auch einen Organismus in seiner Gesamtheit (alle Aspekte des äusseren Erscheinungsbildes, der Anatomie, der Physiologie und des Verhaltens). Gleichermassen steht der Genotyp für das gesamte Genom eines Organismus und nicht nur für die Allele eines einzelnen Genortes. In den meisten Fällen wird die phänotypische Ausprägung eines Gens durch andere Gene und die Umwelt beeinflusst. Der Phänotyp eines Organismus spiegelt seinen gesamten Genotyp und seine Umweltsituation wider. Campbell deutsch: S. 276 Abb. 13.14, engl.: S.259 Abb. 14.13 Stammbaumanalysen bestätigen Mendelsche Erbgänge beim Menschen Erbsen sind günstige Versuchsobjekte für genetische Forschung, Menschen jedoch nicht. Die menschl. Generationszeit beträgt etwa 20 Jahre und Menschen haben relativ wenig Nachkommen (verglichen mit Erbsen und anderen Organismen). Ausserdem sind gezielte Kreuzungsexperimente aus ethischen Gründen nicht durchführbar. Aber die Gesetzmässigkeiten der Mendelschen Genetik sind nach wie vor die Grundlage der Humangenetik. Ein Stammbaum stellt die verwandtschaftlichen Beziehungen zw. Kindern und Eltern über einige Generationen hinweg dar. Wenn man sich durch solch einen Stammbaum hindurch arbeitet, sollte immer wieder die Mendelsche Genetik angewendet werden, auf diese Weise kann man den Genotyp der meisten Individuen ermitteln. Aus einem Stammbaum können wir nicht nur die Genotypen unserer Vorfahren ableiten, sondern auch Voraussagungen machen für unsere Nachkommen. Viele menschliche Erbkrankheiten folgen den Mendelschen Regeln der Vererbung Auch bei Krankheiten, die wie einfache Mendelsche Merkmale vererbt werden, kann man die Technik der Stammbaumanalyse anwenden. Rezessiv vererbte Krankheiten: Man kennt mehrere Tausend genetisch bedingte Krankheiten, die rezessiv vererbt werden. Gene codieren Proteine mit spezifischen Funktionen. Ein Allel, das eine Erbkrankheit bedingt, codiert entweder ein nichtfunktionierendes Protein oder überhaupt kein Protein mehr. Bei rezessiven Erbkrankheiten sind die Heterozygoten gesund, weil sie noch ein zweites „normales“ Allel besitzen, das die Bildung einer ausreichenden Menge des betreffenden Proteins veranlasst. Eine rezessive Erbkrankheit manifestiert sich nur in einem homozygoten Individuum, das von jedem Elternteil ein rezessives Allel erhalten hat. Die Heterozygoten, die phänotypisch gesund sind, werden als Träger bezeichnet, da sie das rezessive Allel an ihre Nachkommen weitergeben können. Die meisten Patienten mit rezessiven Erbkrankheiten sind Kinder „normaler“ Eltern, die beide Träger des rezessiven Gens sind. Die Vereinigung zweier Träger entspricht einer MendelschenF1Kreuzung, bei der die Zygote die Wahrscheinlichkeit von ¼ besitzt zwei rezessive Allele zu erhalten. Ein Kind mit normalem Phänotyp ist mit einer Wahrscheinlichkeit von 2/3 ein Träger. Deshalb sind zwei von drei Individuen mit normalem Phänotyp heterozygote Träger. Falls die Krankheit vor Eintritt der Geschlechtsreife zum Tode führt oder steril macht, pflanzen sich die rezessiv homozygoten Individuen nicht fort. Doch auch wenn sich rezessiv Homozygote fortpflanzen können, bilden solche Individuen stets einen erheblich kleineren Prozentsatz der Population als heterozygote Träger. Im Allg. treten Erbkrankheiten nicht in allen Bevölkerungsgruppen im gleichen Umfang auf. Diese Unterschiede sind auf die unterschiedliche genetische Vorgeschichte zurückzuführen, als die Populationen noch stärker geographisch und daher auch genetisch isoliert waren. Cystische Fibrose: Die häufigste letale genetisch bedingte Krankheit in Europa ist die cystische Fibrose von der jedes 2000ste Neugeborene betroffen ist. Einer von 20 Europäern (5%) ist Träger (heterozygot). Die abnorm hohe extrazelluläre Chlorid-Konzentration macht die Schleimschicht bestimmter Zellen ungewöhnlich dick und zäh. Der Schleim sammelt sich in Pankreas, Lunge, Verdauungstrakt und anderen Organen, wodurch die Gefahr einer Lungenentzündung oder anderer Infektionen steigt. Ohne Behandlung würden Kinder mit cystischer Fibrose vor Ablauf des fünften Lebensjahrs sterben. Eine spezielle Diät, tägliche Antibiotikagaben und andere vorbeugende Massnahmen können die Prognose deutlich verbessern. Heute beträgt die durchschnittliche Lebenserwartung in den Industrieländern ungefähr 27 Jahre (In diesen Durchschnittswert geht die hohe Sterblichkeit im Kindesalter mit ein, der einzelne Betroffene kann wesentlich älter als 27 Jahre werden.). Tay-Sachs-Krankheit: Diese Krankheit wird durch ein funktionsuntüchtiges Enzym verursacht, das nicht in der Lage ist, eine bestimmte Klasse Gehirnlipiden abzubauen. Die Symptome der Tay-SachsKrankheit manifestieren in der Regel einige Monate nach der Geburt. Der Säugling leidet an Krampfanfällen, erblindet und die motorischen und mentalen Fähigkeiten gehen verloren. Der Tod tritt unausweichlich nach einigen Jahren ein. Sichelzellen-Anämie: Die Sichelzellen-Anämie beruht auf einem einzigen Aminosäureaustausch im Hämoglobin der roten Blutzellen. Wenn der Sauerstoffpartialdruck im Blut eines Sichelzellen-Patienten niedrig ist (in extremer Höhenlage oder bei körperlicher Anstrengung), kann das Sichelzellen-Hämoglobin die Gestalt der Blutzellen in eine Sichelform umwandeln. Die Sichelzellenbildung der Blutzellen zieht dann andere Symptome nach sich. Die multiplen Effekte des Sichelzellen-Allels sind auch ein Beispiel für Pleiotropie. Campbell deutsch: S. 279 Abb. 13.16, engl.: S. 262 Abb. 14.15 Für das Sichelzellen-Allel sind heterozygote Individuen Merkmalsträger. Sie sind normalerweise gesund, obwohl manche der Heterozygoten einige Symptome der Sichelzellenkrankheit zeigen, wenn sie unter starkem Sauerstoffmangel leiden. Die Ursache für das gehäufte Auftreten des Sichelzellen-Allels könnte sein, dass nur die Homozygoten tatsächlich Krankheitssymptome haben, die Heterozygoten aber in bestimmten Gegenden einen Vorteil gegenüber gesunden Homozygoten haben. Eine einzelne Kopie des Sichelzellen-Allels erhöht nämlich die Resistenz gegen Malaria. Dominant vererbte Krankheiten: Obwohl die meisten schädlichen Allele rezessiv sind, gibt es doch einige dominante Erbkrankheiten. Achondroplasie: Eine Form des Zwergwuchses mit einer Häufigkeit von 1:10’000 Heterozygoten zeigen Zwergwuchs. Dominant letale Allele sind viel seltener als rezessiv letale. Eine Ursache dafür ist, dass die Effekte dominant letaler Allele nicht durch den heterozygoten Zustand unterdrückt werden. Viele dominant letale Allele sind das Ergebnis von Mutationen in einem Gen des Spermiums oder der Eizelle, durch welche die neuentstehende Nachkommenschaf bereits sehr früh abgetötet wird. Ein Individuum, welches das fortpflanzungsfähige Alter nicht erreicht, kann die mutierte Form des Gens auch nicht weitergeben. Dies steht im Gegensatz zu rezessiv letalen Mutationen, die durch die Fortpflanzung heterozygoter Träger mit normalem Phänotyp von Generation zu Generation weitergegeben wird. Ein dominant letales Allel kann der Eliminierung entgehen, wenn es erst im vorgerückten Alter zum Tode führt. Chorea Huntington: Es handelt sich hierbei um eine generative Nervenerkrankung. Sie wird durch ein dominant letales Allel hervorgerufen, das bis zum 35sten bis 45sten Lebensjahr keine erkennbaren Krankheitssymptome hervorruft. Wenn aber die Zerstörung des Nervensystems beginnt, ist der Krankheitsverlauf irreversibel und führt unweigerlich zum Tode. Jeder Mensch, der das Allel für Chorea Huntington trägt, gibt mit einer 50%-igen Wahrscheinlichkeit das krankmachende Allel an seine Nachkommen weiter. Es ist möglich die Krankheit vor dem Ausbrechen zu diagnostizieren. Für Familien, in denen Chorea Huntington vorgekommen ist, bedeutet dies ein Dilemma: Ist es für einen bislang gesunden Menschen überhaupt wünschenswert zu wissen, dass er eine tödliche, heute noch unheilbare Erbkrankheit hat? Multifaktorielle Krankheiten: - Herzkrankheiten - Diabetes - Krebs - Alkoholismus - Schizophrenie - manisch-depressive Psychose Unser Lebensstil ist von entscheidender Bedeutung in der Balance zwischen genetischen Umwelteinflüssen, insbesondere bei Herz-Kreislauf-Problemen und anderen multifaktoriell bedingten Krankheiten. Sportliches Training, gesunde Ernährung, wenig Alkohol, Abstinenz gegenüber Nikotin und eine positive Bewältigung von Stress-Situationen reduzieren das Risiko für Herzerkrankungen und einige Krebsarten. Die Gentechnik entwickelt neue Methoden für genetische Tests und Familienberatung Eine Verhütung von Erbkrankheiten ist manchmal möglich, weil in einigen Familien das Risiko abgeschätzt werden kann, mit dem ein geplantes Kind ein bestimmtes schädliches Allel erben wird oder ein Fötus dieses Allel trägt. Campbell deutsch: S. 281 Beispiel, engl.: S.264 Diagnose von Trägern: Weil die meisten Kinder mit rezessiven Erbkrankheiten von gesunden Partnern gezeugt werden, ist es zur Ermittlung des genetischen Risikos notwendig festzustellen, ob die zukünftigen Eltern heterozygote Träger des rezessiven Allels sind. Bei einigen Erbkrankheiten lässt sich durch einen Test der dominante Homozygote vom Heterozygoten unterscheiden. Die moderne Biotechnologie eröffnet Möglichkeiten, menschliches Leid zu mindern, aber zuerst müssen einige ethische Probleme gelöst werden. Pränatale Diagnose: - Amniocentese - Chorionzotten-Biopsie - Ultraschall-Untersuchung - Fötoskopie In ungefähr einem Prozent der Fälle verursachen Amniocentese und Fötoskopie Komplikationen, wie etwa Blutungen der Mutter oder gar den Tod des Fötus. Falls durch diese Untersuchungsmethoden eine ernstzunehmende Krankheit diagnostiziert wird, stehen die Eltern vor der schweren Entscheidung zwischen einem Schwangerschaftsabbruch und der Geburt eines Kindes mit einem erblichen Defekt. Untersuchung des Neugeborenen: Einige Erbkrankheiten lassen sich nach der Geburt mit Hilfe recht einfacher Untersuchungsmethoden diagnostizieren, die heute in den meisten Krankenhäusern routinemässig durchgeführt werden. Phenylketonurie: rezessive Erbkrankheit, von der etwa jedes 10’000ste Neugeborene in Europa betroffen ist Kinder mit dieser Krankheit können die Aminosäure Phenylalanin nicht in ausreichendem Mass abbauen. Diese Verbindung und ihr Abbauprodukt können sich in toxischen Mengen im Blut konzentrieren, was zu geistiger Retardation führt. Wenn diese Krankheit aber beim Neugeborenem erkannt wird, kann durch spezielle Diät mit phenylalaninarmen Lebensmitteln die geistige Retardation verhindert und die normale Entwicklung gewährleistet werden. Daher ist die Untersuchung der Neugeborenen auf Phenylketonurie und andere therapierbare Krankheiten besonders wichtig. Leider lassen sich nur sehr wenige Erbkrankheiten erfolgreich behandeln. Wichtig! Der Phänotyp ist sowohl vom Genotyp als auch von der Umwelt abhängig.