Einführung in die Evolution
Werbung

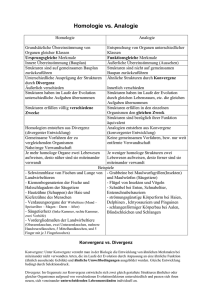
Einführung in die Evolution 1. Block WS 2010/2011 O.Univ.Prof.Dr. Hannes F.Paulus Department für Evolutionsbiologie www.geocities.com/capecanaveral/lab/2948/originoflife.html Evolution ist Veränderung Veränderungen in der Zeit im Weltall: Evolution des Weltalls Kosmologie Veränderungen in der Zeit im Laufe eines Individuallebens Ontogenie, Altern Veränderung der Organismen in der Zeit durch Selektion ist Biologische Evolution Die großen Themen 5 1 2 3 1. 2. 3. 4. 5. Entstehung des Kosmos Die präbiotische Erde Die frühe Evolution des Lebens Die Entfaltung des Lebens Die Zukunft 6. Mechanismen der Evolution 4 Astronomie Molekulare Biologie Biologie/Paläobiologie Phylogenie/Ökologie angewandte Biologie kausale Evolutionsforschung Stoffplan 1. Nachweise für Evolution: Paläontologie Homologie und Analogie 2. Prinzipien und Methoden der Stammesgeschichte und Systematik 3. Die Entstehung des Lebens und der Zelle 4. Kausalfaktoren: Selektion 5. Evolutionsökologie 6. Arten und Artenvielfalt 7. Artbildung 8. Biogeographie 9. Evolution des Menschen Evolution = Veränderung in der Zeit Manchmal ist es schwierig zu entscheiden, in welcher Richtung gefundene oder rekonstruierte Veränderungsreihen (in der Stammesgeschichte) gelesen werden müssen ! ? oder so ? Grundfragen der Evolutionsforschung • • Evolution ist die zentrale These der Biologie Zwei wichtige Grundfragen sind: – Wie sind die Organismen entstanden ? – Warum sind Organismen zweckmäßig gebaut ? • Zentrale Aussagen der Evolutionstheorie sind: – Die Mannigfaltigkeit der Organismen ist das Produkt einer stammesgeschichtlichen Entwicklung, die sich im Laufe der Jahrmillionen aus einer Stammart heraus gebildet hat. – Alle Organismen sind daher miteinander verwandt. – Dies war und ist eine klare Neuposition gegenüber „Schöpfungsmythen“ (Bibel: Genesis oder zahlreiche weitere weltweite Mythen), die meist von einmaligen Schöpfungsakten und anschließender Konstanz der Arten ausgehen. Carl von Linné 1707-1778 Grundfragen der Evolutionsforschung • Der Evolutionsgedanke ist bereits früh entstanden und wurde in zunehmenden Maß durch die Paläontologie deutlich. • Daraus folgte, dass im 18. und 19. Jahrhundert der Evolutionsgedanke weit verbreitet war. • Lamarck (1809) entwickelte die erste geschlossene Evolutionstheorie. Er konnte jedoch nicht sagen, welches die Mechanismen einer evolutiven Veränderung sind. • Der Durchbruch kam durch Charles Darwin (1859) (und in Grenzen auch Wallace), der nicht nur aufzeigen konnte, dass es Evolution gegeben hat, sondern vor allem eine Theorie über die Kausalfaktoren anbieten konnte: Selektionstheorie. Darwin: Der Begründer der Selektionstheorie 1809 - 1889 Darwinjahr: 2009=200 Jahre alt 1859 Vor 150 Jahren ! Darwin und die Folgen Drei wesentliche Grundaufgaben der Evolutionsforschung: • 1. Nachweise dafür, dass es Evolution gibt und gegeben hat. • 2. Wie ist die Stammesgeschichte abgelaufen ? Phylogenese • 3. Welches sind die Evolutionsfaktoren (Kausalanalysen) ? Selektion Drei wesentliche Grundaufgaben der Evolutionsforschung: 1. Nachweise dafür, dass es Evolution gibt und gegeben hat. • Direkte Nachweise – Fossilien (Paläobiologie) – Kurzzeitbeobachtungen evolutiver Änderungen in Populationen – Langzeitbeobachtungen in Populationen – Selektionsexperimente im Labor Indirekte Nachweise – Homologien- und Analogienforschung anhand rezenter und fossiler Organismen – Daraus ergeben sich Argumente für die Rekonstruktion der Stammesgeschichte Fossilgeschichte = Rekonstruktion der Funde in den Schichtfolgen Fossilien: Zustandekommen der Schichten Fossilien und ihre Rekonstruktion Verbreitungsgeschichte des Hauspferdes Fossilgeschichte als Belege für Evolution: Evolution der Wale Ambulocetus natans (50 Mill.) Rudiment des Beckens Dorudon atrox (40 Mill.) rezente Wale Stellung der Wale im System der Säuger Die Wale sind eine Schwestergruppe der Paarhufer. = Paarhufer und Wale haben einen letzten gemeinsamen Vorfahren Stammbaum nach morphol. Daten Wale entstammen den Vorfahren der Paarhufer Die Wale sind eine Schwestergruppe der Flußpferde und damit eine Teilgruppe der Paarhufer. Doch die Unterschiede der Wale zu ihren Landvorfahren sind gewaltig ! Stammbaum nach molekularen Daten Wo sind die Übergänge? Direkte Nachweise für Evolution Fossilien Die Suche nach den „missing links“ „Missing links“ werden besser „connecting links“ bezeichnet, da sie an den basalen Verzweigungen der Stammesgeschichte den Übergang von einer Grundkonstruktion zu einer neuen dokumentieren können/sollen. H.Paulus: Einführung in die Evolution WS 2010-2011: Walvorfahren Haldanodon Seymouria Ichthyostega Archaeopteryx Archaeopteryx – ein „connecting link“ zu den Vögeln Vogel Die Schemazeichnung stellt einen typischen Vogel und ein typisches Reptil dem Archaeopteryx gegenüber. Vogelmerkmale: Archaeopteryx Reptil • Vogelschädel • Federkleid • Schultergürtel mit Stütze der Flugmuskulatur • Form der Armknochen (Flügel) • Vogelbeine (Mittelfußknochen gehört zum Laufbein) • Vogelfüße (1. Zehe nach hinten gerichtet) Vogel Reptilienmerkmale: Reptilienmerkmale • Kegelzähne • lange Schwanzwirbelsäule • freie Finger- und Mittelhandknochen • Schien- und Wadenbein nicht verwachsen (hier leider nicht gut sichtbar) • Beckenknochen nur durch Bindegewebe verbunden • Krallen an Vorder- und Hinterextremitäten Archaeopterix Reptil Fossilfunde von Ichthyostega – das Bild hier ist Rekonstruktion (ein Nachbau) – kennen wir aus den Devon-Schichten Grönlands. Dieser Urlurch gilt heute als das erste bekannte Landwirbeltier. Er besitzt Merkmale sowohl von den Fischen als auch von den Amphibien Stammbaum der Wirbeltiere Fischmerkmale: Amphibienmerkmale: fischähnlicher Schwanz schmale Rückenflosse schuppenbedeckter Körper vier Beine mit Knochenskelett fünf Zehen keine Kiemen (Lungen- und Hautatmung) „Connecting links“ oder Brückentiere sind für ein Verständnis des Evolutionsgeschehens von großer Bedeutung, da sie als Zwischenformen die stammesgeschichtliche Verwandtschaft zweier Tiergruppen zeigen. Das bedeutet, dass die These von der allmählichen Entwicklung der Lebewesen (Evolution) sehr gut belegt ist. Dass sich zum Beispiel Landwirbeltiere aus Fischen oder die Vögel aus Reptilien fortentwickelt haben, ist damit sehr gut belegt. In diesen beiden Fällen gelten die Brückentiere Quastenflosser (oder auch Ichthyostega) und Archaeopteryx als besonders prominente Belege. Lebende Fossilien sind im System der Organismen isoliert stehende und im Vergleich zu ihren weit verbreiteten Vorfahren oft auf Schrumpf- oder Reliktareale beschränkte Arten. Altertümliche Merkmale bestätigen, dass sich »lebende Fossilien« im Gegensatz zu verwandten Formen seit Jahrmillionen nicht oder kaum verändert haben. Lebende Fossilien Lebende Fossilien Nautilus Mehrere Typen gibt es: 1. Letzte Überlebende einer früher florierenden, heute aber ausgestorbenen Gruppe Latimeria, Nautilus, Brückenechse, Gingko-Baum Sphenodon 2. Ursprünglich gebliebene Teilgruppe Triops einer heute florierenden Gruppe „Urzeitkrebse“, Neopilina 3. Arten, die sich bis heute kaum verändert haben, da sich ihre Umwelt nicht verändert hat Limulus Limulus: Chelicerata, Xiphosura Neopilina Nautilus (Perlboote) Vertreter der Mollusca, Cephalopoda, 6 rezente Arten Sphenodon (Brückenechsen)(2 Arten)(Neuseeland) seit der Trias (200 Mill.Jahre) „Urzeitkrebse“ (Crustacea: Anostraca, Notostraca), leben in „astatischen Gewässern“ (Triops, Artemia etc.) Limulus (Pfeilschwanzkrebse) Chelicerata (!), Xiphosura Gingko biloba Silberaprikose, Ginkgobaum, Fächerblattbaum Superdivision Spermatophyta Division/phylum Ginkgophyta Klasse Ginkgoopsida Ordnung Ginkgoales Familie Ginkgoaceae Genus Ginkgo L. Einziger lebender Vertreter der Ordnung Reliktgattung von vor 180 Mill. Jahren Der in der chinesischen Provinz Sichuan heimische Ginkgo gilt als lebendes Fossil, da die Ginkgopflanzen (manchmal auch Ginkgoartigen) schon seit 250 Millionen Jahren existieren, aber alle anderen Arten ausgestorben sind. Der bis zu 40 m hohe Baum wurde ab ca. 1000 n. Chr. in ganz Ostasien als Tempelbaum verbreitet und auch auf die Koreanische Halbinsel sowie nach Japan exportiert. Ginkyō (jap: Gin ist "Silber" und Kyō ist "Frucht") Ginkgo = vermutlich Schreibfehler ! Coelacanthiformes Comoren-Quastenflosser (Latimeria chalumnae) (J. L. B. Smith, 1939) Manado-Quastenflosser (Latimeria menadoensis) (Pouyaud, Wirjoatmodjo, Rachmatika, Tjakrawidjaja, et al., 1999) alle anderen Gattungen sind nur fossil bekannt ! Diese Art wurde erst 1938 entdeckt. Bisher ist sie nur von etwa 100 Exemplaren aus einem kleinen Gebiet bei Madagaskar und den Comoren bekannt. Seit September 1998 ist eine 2. Population bei Sulawesi (Indonesien) bekannt. Ihre nächsten Verwandten lebten im Erdmittelalter. Devonische Quastenflosser sind als Vorfahren der Landwirbeltiere stammesgeschichtlich von großer Bedeutung. Während die meisten Quastenflosser vor etwa 300 Millionen Jahren im Erdaltertum gelebt haben, sind nach unserem heutigen Wissen die letzten Arten in der Kreidezeit vor 70 Millionen Jahren am Ende des Erdmittelalters ausgestorben. Übriggeblieben ist offensichtlich nur die Gattung Latimeria. Das sensationelle an der Entdeckung von Latimeria war nicht nur, dass sie der längst für ausgestorben geglaubten Tiergruppe der Quastenflosser angehört, sondern dass es zwischen den ältesten fossilen Vertretern des Devons und der rezenten (noch lebenden) Latimeria kaum einen morphologischen Unterschied gibt, ihr Bauplan sich in den Jahrmillionen also kaum verändert hat. Die Schwanzflosse ist stets dreigeteilt, wobei der mittlere Teil wie ein Pinsel oder eine Quaste herausragt, was dieser Fischgruppe den deutschen Namen Quastenflosser eingebracht hat. Latimeria kann deshalb mit Recht als ein "lebendes Fossil" bezeichnet werden. Homologien und Analogien sind indirekte Beweise für Evolution Pierre Belon (1555): Vergleich Vogel- und Menschenskelett Die vergleichende Betrachtung zeigte schon früh eine auffällige „Formverwandtschaft“ Die Formverwandtschaft konnte zunächst nicht erklärt werde, bis Darwin (1859): Formulierung der Evolutionstheorie als Selektionstheorie Der „Plan“ aller Organismen ist in ihnen als ERBINFORMATION enthalten. Diese stammt von ihren gemeinsamen AHNEN (VORFAHREN). Damit konnte die bisherige Formverwandtschaft als „Blutsverwandtschaft“ (= GENEALOGIE) erklärt werden. Stammesgeschichtliche Forschung versucht mit Hilfe vieler sehr unterschiedlicher Methoden diese genealogische Verwandtschaft aufzuklären. Die verbreitetste Form der Darstellung der Resultate dieser Forschungen sind STAMMBÄUME (Dendrogramme, Kladogramme). Stammbäume sind HYPOTHESEN der vermuteten verwandtschaftlichen Beziehungen. Sie können aufgrund neuer Daten verändert werden ! Pierre Belon (1555): Vergleich Vogel- und Menschenskelett Diese Formverwandtschaft besteht deshalb, weil beide (hier Mensch und Vogel) diese Ähnlichkeiten von einem gemeinsamen Vorfahren geerbt haben. Dieser war der erste „Tetrapode“. Stammbaum der Wirbeltiere TETRAPODA 1. Nachweise für Evolution: Homologienforschung Das Phänomen Homologie kann man 1. operational Beschreiben 2. Definieren Homologie-Definition Homologie bedeutet im Vergleich zweier oder mehrerer Arten das Vorliegen von Übereinstimmungen von Strukturen, Verhaltensweisen, physiologische Prozesse, Makromoleküle etc., die auf für diese Strukturen spezifischer gemeinsamer genetischer Information aus dem Informationsspeicher Genom beruhen und durch den Prozess der Vererbung von Eltern auf ihre Kinder entstanden und erhalten sind. Ihre Übereinstimmung beruht daher nicht auf Zufall. Homologie-Kriterien: Homologien können anhand von Kriterien erkannt werden: 1. Lage und Lagebezug 2. Spezifische Qualität 3. Übergangsreihen (Kontinuität) Homologie ist im Vergleich zweier Arten eine solche Strukturähnlichkeit, die identischen genetischen Ursprungs ist, weil sie einem gemeinsamen Bauplan oder Grundplan entstammt, also gemeinsamer Abstammung ist. Die Feststellung von Homologien bei verschiedenen Arten bedeutet, dass sie diese von einem gemeinsamen Vorfahren ererbt haben müssen. Homologien: 1. Kriterium der Lage 1. Kriterium der Lage Festlegung von korrekten Benennungen eines Teiles im Verband anderer Teile: z.B.: Der Radius-Knochen (rot) befindet sich stets zwischen Humerus (blau) und Carpus-Knochen (gelb) Menschen Hund Robbe 2. Kriterium der spezifischen Qualität Isolierte Teile von Tieren können an ihrem „spezifischen Aussehen“ dem ganzen Tiere zugeordnet werden. Australopithecus afarensis Pfauenfeder: Spitzenteil der langen Rückenfeder Homologie: 3. Kriterium der Übergangsreihen Wichtigste Methode der vergleichenden Anatomie Säugetiere besitzen ein sogenanntes „sekundäres Kiefergelenk“ Das primäre (ursprüngliche der Wirbeltiere) befand sich zwischen den Schädel-Kieferknochen Quadratum (rot) und dem Articulare (gelb): so noch bei Fischen und Reptilien. Durch Abwandlung des Schädels, Stabilisierung der gelenkigen Verbindung des Unterkiefer mit dem Oberkiefer verband sich das Zähne tragende Dentale (grau) mit dem Knochen des Zähne tragenden Oberkiefers (Quadratomaxillare) zu einem neuen Gelenk (=„sek. KieferGelenk“). Die ursprünglichen Kieferknochen Quadratum und Articulare wurden zu den 2. und 3. Gehörknöchelchen. • Cynognathus crateronotus = Reptil-Säuger • • • • Reptilienmerkmale: Schwanz, Körperbau: z.B. noch primäres Kiefergelenk Säugermerkmale: Zähne, Milchdrüsen, Fell Unter- bis Mitteltrias 245 bis 237 Mio. Jahre Gondwana Homologie - Analogie H. Paulus Einführung in die Evolution WS 2010-2011 Homologie-Definition Homologie bedeutet im Vergleich zweier oder mehrerer Arten das Vorliegen (nicht zufälliger) Übereinstimmungen von Strukturen, Verhaltensweisen, physiologische Prozesse, Makromoleküle etc., die auf für diese Strukturen spezifischer gemeinsamer genetischer Information aus dem Informationsspeicher Genom beruhen und durch den Prozess der Vererbung von Eltern auf ihre Kinder entstanden und erhalten sind. Ein anderer Informationsspeicher ist das Gehirn ! Über dieses kann ebenfalls Information (=Gelerntes) an Nachkommen weitergegeben werden (= Tradition, Kultur). Homologie-Kriterien: Homologien können anhand von Kriterien erkannt werden: 1. Lage und Lagebezug 2. Spezifische Qualität 3. Übergangsreihen (Kontinuität) Homologie ist im Vergleich zweier Arten eine solche Strukturähnlichkeit, die identischen genetischen Ursprungs ist, weil sie einem gemeinsamen Bauplan oder Grundplan entstammt, also gemeinsamer Abstammung ist. Die Feststellung von Homologien (homologen Merkmalen) bei verschiedenen Arten bedeutet, dass sie diese von einem gemeinsamen Vorfahren geerbt haben müssen. Homologienforschung Homologie gilt auf allen Ebenen ! • • • • • Strukturen physiologischen Prozessen Verhaltensweisen Molekülen etc. Bedingung: Die betreffenden Strukturen müssen erblich sein ! Einsichtig ist dies bei Verhalten: Gleiches Verhalten kann auch erlernt sein. Homologien bei Makromolekülen: Cytochrom C Ritualisiertes Verhalten in der Entenbalz Stockenten benutzen 8 Verhaltenstypen Löffelenten nutzen 1 bis 2 Verhaltenstypen Strukturen aus genetisch fixierten Verhaltenskoordinationen = Erbkoordination SchmetterlingsRaupenköcher Spinnennetze Köcher von Köcherfliegen (Trichoptera) Strukturen, die aus erb-homologen Verhalten stammen, sind ebenfalls als homolog zu betrachten Weitere verwandte Begriffe für den Vergleich von Strukturen: Analogie → Strukturähnlichkeiten, die bei zwei oder mehr Arten konvergent, also unabhängig aus ungleichen Ursprüngen entstanden sind: Funktionsähnlichkeit durch parallele Selektion (= Homoplasie) Beispiel: Vogelflügel - Insektenflügel Homoiologie → Analogien auf homologer Basis (Parallelismen) Beispiele: Vogelflügel – Fledermausflügel, Fangbeine bei Mantis (Fangschrecken) und Mantispa (Netztflügler) Homonomie → Strukturidentische Bauteile an ein und demselben Individuum. Die Differenzen werden als Divergenzen aus primär identischen Grundformen angesehen (seriale Homologie besser seriale Homonomie bzw. bei Abwandlungen als seriale Heteronomie zu bezeichnen). Beispiel (vor allem bei segmentalen Organismen) für Homonomie: gleichgestaltete Beine bei Arthropoda Heteronomie: unterschiedlich gestaltete Beine bei Arthropoda Weitere verwandte Begriffe für den Vergleich von Strukturen: Analogie → Strukturähnlichkeiten, die bei zwei oder mehr Arten konvergent, also unabhängig aus ungleichen Ursprüngen entstanden sind: Funktionsähnlichkeit durch parallele Selektion (= Homoplasie) Beispiel: Vogelflügel - Insektenflügel Homoiologie → Analogien auf homologer Basis (Parallelismen) Beispiele: Vogelflügel – Fledermausflügel, Fangbeine bei Mantis (Fangschrecken) und Mantispa (Netztflügler) Homonomie → Strukturidentische Bauteile an ein und demselben Individuum. Die Differenzen werden als Divergenzen aus primär identischen Grundformen angesehen (seriale Homologie besser seriale Homonomie bzw. bei Abwandlungen als seriale Heteronomie zu bezeichnen). Beispiel (vor allem bei segmentalen Organismen) für Homonomie: gleichgestaltete Beine bei Arthropoda Heteronomie: unterschiedlich gestaltete Beine bei Arthropoda Ähnlichkeiten nach Vergleich von Strukturen (Merkmalen) müssen nicht unbedingt homolog sein ! Alternative Erklärungen: Analogie Strukturähnlichkeiten, die bei zwei oder mehr Arten konvergent, also unabhängig aus ungleichen Ursprüngen entstanden sind: Funktionsähnlichkeit durch parallele Selektion (= Homoplasie) Beispiel: Vogelflügel - Insektenflügel Spezialfall der Analogie: Homoiologie Analogien auf homologer Basis (Parallelismen) Beispiele: Vogelflügel – Fledermausflügel, Fangbeine bei Mantis (Fangschrecken) und Mantispa (Netztflügler) Analogie, Konvergenz 3 Wirbeltiergruppen haben aus ihren Vorderextremitäten Flügel entwickelt. Sind diese homolog oder analog ? Flugsaurier Vogel Fledermaus Feststellungen: 1. Die Konstruktionen sind im Detail verschieden. 2. Die jeweils gemeinsamen Vorfahren hatten noch keine Flügel. Homoiologie = Konvergenz auf homologer Basis Flugsaurier Vogel Fledermaus Feststellungen: 1. Kriterium der Lage: Alle sind Vorderextremitäten 2. Kriterium der spezifischen Qualität: Die Konstruktionen sind im Detail verschieden. 3. Kriterium der Übergangssreihen: Die jeweils gemeinsamen Vorfahren hatten noch keine Flügel. Homoiologie = Konvergenz auf homologer Basis Analogie – Homologie der Flügel ? • Sie sind alle analog, da konvergent (unabhängig) entstanden ! • Ihre jeweiligen Vorfahren hatten nämlich noch keine Flügel, wohl aber Vorderextremitäten. Konvergenz: Grabbeine Grabbeine bei verschiedenen Insekten Grabbeine sind sind also mehrfach konvergent entstanden ! Mantis (Gottesanbeterin) Ordnung MANTODEA Homoiologie = Parallelismus Der Bau der Vorderbeine beider Tiere ist zwar außerordentlich ähnlich. Dennoch sind sie als Fangbeine nicht homolog ! Ihre jeweiligen Vorfahren hatten nämlich jeweils noch keine Fangbeine. Mantispa (Fanghafte) Ordnung NEUROPTERA „Hemimetabola“ Die Gruppe der Geier enthält Arten, die konvergent entstanden sind Mönchsgeier (Europa) „Altweltgeier“ Truthahngeier (S.Amerika) „Neuweltgeier“ Schmutzgeier Afrik.Palmgeier Kondor Truthahngeier Gänsegeier Bartgeier Königsgeier Wespenbussard Kalif. Kondor Schlangenadler Mönchsgeier Auch Konvergenz ist Beweis für Evolution In Anpassung an ähnliche Umweltbedingungen entstehen aus unterschiedlichem Ausgangsmaterial gleichartige Formen Euphorbia canariensis (Wolfsmilchgewächs) Kanarische Inseln Kaktus (Cactaceae) der Neuen Welt Verwandte Artengruppen zeigen eine parallele Entwicklungsgeschichte, wenn ihre Mitglieder in getrennten Regionen unter parallelen ökologischen Bedingungen leben. Beispiel: Säugetiere (Plazentalia) und Beuteltiere (Marsupialia) Man spricht von einer „Ökologischen Stellenäquivalenz“ Ökologische Stellenäquivalenz Ähnliche Umwelten bieten vergleichbare Lizenzen für ähnliche ökologische Rollen Stellenäquivalenz und adaptive Radiation: Madagaskar Ableitung aus einer frühen Besiedlung durch einen Vertreter der Insectivora