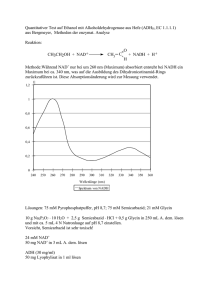
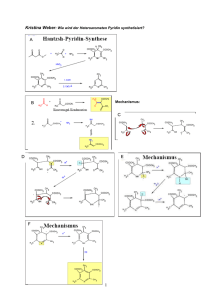
Der Elektronentransfer der Escherichia coli NADH:Ubichinon
Werbung

Der Elektronentransfer der Escherichia coli NADH:Ubichinon Oxidoreduktase sowie die ESR-spektroskopische Charakterisierung weiterer Metalloproteine INAUGURALDISSERTATION zur Erlangung des Doktorgrades der Fakultät für Chemie, Pharmazie und Geowissenschaften der Albert-Ludwigs-Universität Freiburg vorgelegt von Diplom-Chemikerin Katerina Henrike Dörner Freiburg im Breisgau, 2010 Vorsitzender des Promotionsausschusses: Prof. Dr. Rolf Schubert Referent: Prof. Dr. Thorsten Friedrich Korreferent: Prof. Dr. Oliver Einsle Tag der Verkündigung der Prüfung: 16.12.2010 Ein Teil der in dieser Arbeit vorgestellten Ergebnisse wurde in folgenden Artikeln veröffentlicht: Pohl, T., Bauer, T., Dörner, K., Stolpe, S., Sell, P., Zocher, G. & Friedrich, T. (2007) Ironsulfur cluster N7 of the NADH:ubiquinone oxidoreductase (complex I) is essential for stability but not involved in electron transfer. Biochemistry 46, 6588-6596. Kohlstädt, M., Dörner, K., Labatzke, R., Koc, C., Hielscher, R., Schiltz, E., Einsle, O., Hellwig, P. & Friedrich, T. (2008) Heterologous production, isolation, characterization and crystallization of a soluble fragment of the NADH:Ubiquinone Oxidoreductase (Complex I) from Aquifex aeolicus. Biochemistry 47, 13036-13045. Pohl, T., Schneider, D., Hielscher, R., Stolpe, S., Dörner, K., Kohlstädt, M., Böttcher, B., Hellwig, P. & Friedrich, T. (2008) Nucleotide-induced conformational changes in the Escherichia coli NADH:ubiquinone oxidoreductase (complex I). Biochem. Soc. Trans. 36, 971-975. Schneider, D., Pohl, T., Walter, J., Dörner, K., Kohlstädt, M., Berger, A., Spehr, V. & Friedrich, T. (2008) Assembly of the Escherichia coli NADH:ubiquinone oxidoreductase (complex I). Biochim. Biophys. Acta 1777, 735-739. Kung, J. W., Löffler, C., Dörner, K., Heintz, D., Gallien, S., Van Dorsselaer, A., Friedrich, T. und Boll, M. (2009) Identification and characterization of the tungsten-containing class of benzoyl-coenzyme A reductases. Proc. Natl. Acad. Sci. U.S.A. 106, 17687–92. Krätzer, C., Welte, C., Dörner, K., Friedrich, T. und Deppenmeier, U. (2010) Characterization of methanoferrodoxin: a novel superoxide reductase. FEBS J., im Druck. Folgender Artikel wurde eingereicht: Morina, K., Schulte, M., Hubrich, F., Dörner, K., Steimle, S., Stolpe, S. und Friedrich, T. (2010) Engineering the respiratory complex I to an energy-converting NADPH:ubiquinone oxidoreductase. INHALTSVERZEICHNIS 1. Zusammenfassung ..............................................................................................................1 2. Einleitung ...........................................................................................................................3 2.1. Das aerobe Elektronentransportsystem aus E. coli ........................................................3 2.2. Die NADH:Ubichinon Oxidoreduktase (Komplex I) ....................................................5 2.2.1. Elektronentransfer im Komplex I ........................................................................ 11 2.2.2. Überproduktion des Komplex I und seiner Varianten in E. coli ........................... 15 2.3. Beteiligung aromatischer Aminosäuren am Elektronentransport ................................. 16 2.4. Die Microsecond Hyper-Quenching (MHQ)-Technik ................................................. 18 2.5. Weitere untersuchte Metalloproteine ..........................................................................22 2.5.1. Die Superoxid Reduktase aus Methanosarcina mazei ..........................................22 2.5.2 Die Benzoyl-CoA Reduktase aus Geobacter metallireducens ............................... 23 2.6. Ziel der Arbeit ...........................................................................................................24 3. Material und Methoden ..................................................................................................... 26 3.1 Bakterienstämme und Vektoren .................................................................................. 26 3.2 Oligonukleotide ..........................................................................................................29 3.3 Chemikalien................................................................................................................ 31 3.4 Medien, Puffer und Lösungen ..................................................................................... 34 3.5 Mikrobiologische Methoden ....................................................................................... 38 3.5.1 Bakterienanzucht .................................................................................................. 38 3.5.2. Herstellung von Glycerindauerkulturen ............................................................... 39 3.5.3. Herstellung elektrokompetenter Zellen ................................................................ 39 3.6. Molekularbiologische Methoden ................................................................................ 40 3.6.1. Plasmidpräparation .............................................................................................. 40 3.6.2. Restriktion von DNA .......................................................................................... 41 3.6.3. Polymerase Kettenreaktion (PCR) ....................................................................... 41 3.6.4. Transformation durch Elektroporation ................................................................. 43 3.6.5. λ-Red vermittelte Rekombination ........................................................................ 44 3.6.6. Agarose Gelelektrophorese.................................................................................. 44 3.7 Proteinchemische Methoden ....................................................................................... 45 3.7.1. Zellaufschluß und Präparation von Membranen ................................................... 45 3.7.2 Zuckergradientenzentrifugation ............................................................................ 45 3.7.3. Proteinpräparation ............................................................................................... 46 3.7.4. Größenausschluss-Chromatographie .................................................................... 47 3.7.5 Aktivitätsbestimmungen ....................................................................................... 48 3.7.6. SDS-Polyacrylamid-Gelelektrophorese (SDS-PAGE) ......................................... 49 3.7.7. Proteinbestimmung ............................................................................................. 49 3.7.8. Microsecond Freeze-hyperquenching (MHQ) Technik ........................................ 50 3.7.9 ESR-Spektroskopie .............................................................................................. 51 4. Ergebnisse ........................................................................................................................ 52 4.1 Isolierung des Komplex I und des NADH Dehydrogenase Fragmentes ....................... 52 4.1.1. Isolierung des Komplex I .................................................................................... 52 4.1.2 Isolierung des NADH Dehydrogenase Fragmentes ............................................... 55 4.2. Die Reduktion der Fe-S-Zentren durch NADH ........................................................... 56 4.3 Die Bedeutung des Fe-S-Zentrums N1a ......................................................................65 4.3.1 Mutagenese des Bindemotivs des Fe-S-Zentrums N1a..........................................65 4.3.2. Komplex I vermittelte Aktivität der cytoplasmatischen Membranen aus den Mutanten ANN0221/pBADnuo nuoFHis nuoECXA ....................................................... 68 4.3.3. Isolierung und Charakterisierung des Komplex I aus den Mutanten ANN0221/pBADnuo nuoFHis nuoECXA ......................................................................71 4.3.4. Isolierung und Charakterisierung des Komplex I aus den Mutanten ANN0221/pBADnuo nuoFHis nuoE CXS ......................................................................74 4.3.5 Eignung des Stammes AN387 Δnuo als Expressionsstamm .................................. 76 4.3.6. Eignung des Stammes BW25113Δnuo als Expressionsstamm.............................. 78 4.3.7. Charakterisierung der Alanin- und Serin-Varianten aus dem Stamm BW25113Δnuo/pBADnuo nuoFHis................................................................................ 83 4.4 Die Bedeutung aromatischer Aminosäuren für den Elektronentransport im E. coli Komplex I ........................................................................................................................ 86 4.4.1 Mutagenese zum Austausch der aromatischen Aminosäuren ................................ 88 4.4.2 Komplex I vermittelte Aktivitäten der cytoplasmatischen Membranen aus den Mutanten mir den ausgetauschten aromatischen Aminosäuren ......................................89 4.4.3 Stabilität der Komplex I Varianten W221YNuoG, W221YNuoG und F515NuoCD ........ 94 4.5 ESR-Spektroskopische Charakterisierung weiterer Proteine ....................................... 98 4.5.1. ESR-spektroskopische Charakterisierung einer Superoxid Reduktase aus M. mazei ................................................................................................................. 98 4.5.2. ESR-spektroskopische Charakterisierung von BamBC aus Geobacter metallireducens ........................................................................................................... 100 5. Diskussion ...................................................................................................................... 103 5.1 Die MHQ-Technik zur Beobachtung des Elektronentransfers innerhalb des Komplex I aus E. coli ............................................................................................... 103 5.2. Die Bedeutung des Fe-S-Zentrum N1a ..................................................................... 105 5.3 Die mögliche Beteiligung aromatischer Aminosäuren am Elektronentransport im Komplex I ...................................................................................................................... 108 5.4 Der Überexpressionsstamm BW25113 Δnuo pBADnuo nuoFHis ............................... 111 5.5 ESR-Spektroskopische Charakterisierung zwei weiterer Proteine .............................. 111 6. Literatur.......................................................................................................................... 113 Zusammenfassung 1. ZUSAMMENFASSUNG Die NADH:Ubichinon Oxidoreduktase, auch respiratorischer Komplex I genannt, ist ein energiewandelnder Enzymkomplex der Atmungskette. Der Komplex I koppelt den Elektronentransfer von NADH auf Ubichinon mit einer Protonentranslokation über die Cytoplasmamembran in Prokaryoten und die innere Membran der Mitochondrien in Eukaryoten. Der Komplex I aus Escherichia coli setzt sich aus 13 Untereinheiten zusammen und ist aus einem in der Membran eingebetteten, hydrophoben Arm und einem ins Cytoplasma ragenden, peripheren Arm aufgebaut. Alle bisher bekannten Kofaktoren, ein Flavinmononukleotid (FMN) und die neun Eisen-Schwefel (Fe-S)-Zentren N1a, N1b, N2, N3, N4, N5, N6a, N6b und N7, liegen im peripheren Arm. Sieben der neun Fe-S-Zentren bilden eine Elektronentransportkette zwischen dem FMN, das die Elektronen vom NADH aufnimmt, und dem Elektronenakzeptor Ubichinon. Das zweikernige Fe-S-Zentrum N1a ist aufgrund seiner Lage im Komplex nicht Teil der Elektronentransportkette. Es kann Elektronen ausschließlich vom FMN aufnehmen und wird durch NADH reduziert. Seine Funktion im Komplex I ist bisher unbekannt. In dieser Arbeit wurde durch Mutagenesestudien gezeigt, dass N1a essentiell für die Stabilität des Komplex I ist. Der Austausch der Cysteine des Bindemotivs von N1a gegen Alanine oder Serine führte jeweils zu einem instabilen Komplex I. Der Elektronentransfer innerhalb des E. coli Komplex I war das zentrale Thema dieser Arbeit. Mittels der Microsecond-Hyperquenching Technik und Tieftemperatur ESR-Spektroskopie wurde die Reduktion der einzelnen Fe-S-Zentren während der Reaktion des Komplex I mit NADH in einem Zeitfenster von 100 µs bis 6 ms verfolgt. Ein Apparatur-bedingtes Artefakt im spektralen Bereich der vierkernigen Fe-S-Zentren lässt nur die detaillierte Analyse der Reduktionskinetik der zweikernigen Fe-S-Zentren N1a und N1b zu. Die Ergebnisse zeigen, das N1a an der physiologischen Reaktion des Komplex I beteiligt ist. Sie unterstützen die Vermutung, dass N1a durch die zeitweilige Aufnahme eines Elektrons eine mögliche Radikalbildung am FMN vermindert und somit zum Schutz des Organismus vor gefährlichen Radikalen beiträgt. Die Vermutung, dass das erste Elektron vom FMN über die Elektronentransportkette auf das nahe der Ubichinonbindestelle gelegene N2 und das zweite Elektron auf N1a übertragen wird, wurde in dieser Arbeit unterstützt. 1 Zusammenfassung Desweiteren sollte die Beteiligung aromatischer Aminosäuren am Elektronentransfer im Komplex I untersucht werden. Ist der Abstand zwischen zwei Redoxzentren kleiner als 14 Å, wird eine direkte Elektronenübertragung mit ausreichend hohen Transferraten angenommen. Bei größeren Abständen sind nach der hopping Theorie aromatische Aminosäuren über ihr лElektronensystem direkt am Elektronentransfer beteiligt. Im Komplex I liegen die Fe-SZentren N5 und N6a mehr als 14 Å voneinander entfernt und sind auf zwei verschiedenen Modulen lokalisiert, aus denen der Komplex I evolutionär entstanden ist. Die hoppingTheorie schlägt drei konservierte, aromatische Aminosäuren vor, die an diesem Transferschritt beteiligt sein können. In dieser Arbeit wurden die konservierten, aromatischen Aminosäuren im E. coli Komplex I gegen Leucin und gegen die jeweils anderen aromatischen Aminosäuren ausgetauscht. Der Einfluss dieses Austausches auf die Aktivität des Komplex I wurde bestimmt. Die Ergebnisse unterstützen die hopping Theorie und lassen die Beteiligung aromatischer Aminosäuren an dem Transferschritt zwischen N5 und N6a vermuten. Weitere konservierte, aromatische Aminosäuren, die am Elektronentransfer zwischen Fe-S-Zentren mit Abständen kleiner als 14 Å beteiligt sein sollen, wurden in dieser Arbeit ausgetauscht. Eine Beteiligung am Elektronentransfer konnte aber experimentell nicht bestätigt werden. Zwei weitere Metalloproteine wurden in der vorliegenden Arbeit ESR-spektroskopisch charakterisiert. Das Protein MM0632 aus Methanosarcina mazei wird als Prototyp einer neuen Klasse von Superoxid Reduktasen, die in anaeroben Organismen am Abbau von reaktiven Sauersoffspezies beteiligt sind, vorgeschlagen. Durch ESR-Spektroskopie wurden neben einem für die Enzymfamilie typischen [Fe(His)4(Cys)]-Zentrum erstmalig ein vierkerniges Fe-S-Zentrum in einer Superoxid Reduktase nachgewiesen. Das Fragment BamBC aus dem Proteinkomplex BamB-I aus Geobacter metallireducens ist ein Vertreter einer neuen Klasse von Benzyl-CoA Reduktasen, die in anaeroben Organismen am Abbau von aromatischen Verbindungen beteiligt sind. Durch ESR-Spektroskopie wurden in dieser Arbeit neben einem Wolfram-Kofaktor ein dreikerniges und drei vierkernige Fe-S-Zentren nachgewiesen. 2 Einleitung 2. EINLEITUNG 2.1. Das aerobe Elektronentransportsystem aus E. coli Bei der oxidativen Phosphorylierung wird die im Katabolismus gewonnene Energie für den Organismus in Form von ATP nutzbar gemacht. Die bei Glykolyse, Citratzyklus und Fettsäureoxidation entstandenen energiereichen Reduktionsäquivalente NADH und FADH2 werden durch die Enzyme der Atmungskette oxidiert und die Elektronen in einer Reihe von Redoxreaktionen auf Sauerstoff übertragen. Die membranständigen Enzyme der Atmungskette koppeln den Elektronentransfer mit dem Aufbau eines Protonengradienten über die Membran. Die bei den Redoxreaktionen frei werdende Energie wird so in Form eines Protonengradienten, auch protonenmotorische Kraft (PMK) genannt, gespeichert. Die durch den Abbau des Gradienten gewonnene Energie wird für die Synthese von ATP durch die ATP-Synthase und für andere energieverbrauchende Prozesse genutzt (Mitchell & Moyle, 1967). Die gemeinsame Arbeit der Enzyme der Atmungskette und der ATP-Synthase wird als oxidative Phosphorylierung bezeichnet. Bei Eukaryoten sind die Atmungskettenkomplexe und die ATP-Synthase in der inneren Membran der Mitochondrien, bei Prokaryoten in der Cytoplasmamembran lokalisiert. Escherichia coli ist ein gram-negatives, fakultativ anaerobes Enterobakterium (griech.: „enteron“ Darm). Es ist in der Lage, seinen Energiebedarf über aerobe und anaerobe Elektronentransportsysteme zu decken. Das aerobe System hat für die Wissenschaft große Bedeutung, da Enzymkomplexe des bakteriellen Systems mit denen der Mitochondrien verwandt sind (Calhoun et al., 1994; Friedrich & Scheide, 2000). Erkenntnisse über die einfacher aufgebauten und im Labor leichter zugänglichen bakteriellen Proteine lassen sich auf die mitochondrialen Enzyme übertragen und helfen, durch die humanen Enzyme hervorgerufenen Dysfunktionen zu verstehen. Das aerobe Elektronentransportsystem aus E. coli besteht im Wesentlichen aus zwei primären Dehydrogenasen, der NADH:Ubichinon Oxidoreduktase (NADH Dehydrogenase Typ I, NDH I) und der alternative NADH Dehydrogenase (NADH Dehydrogenase Typ II, NDH II) und den terminalen ChinolOxidasen, der Cytochrom bo3-Oxidase und der Cytochrom bd-Oxidase. Die Elektronen werden dabei von NADH erst auf ein Chinon (Q) und dann auf Sauerstoff übertragen (Abb. 2-1). Die in Analogie zum mitochondrialen Enzym auch als Komplex I bezeichnete NDH I und die beiden terminalen Oxidasen sind energiewandelnde Enzyme, dabei ist die 3 Einleitung Anzahl der pro Elektron transportierten Protonen bei der bo3-Oxidase doppelt so hoch wie bei der Cytochrom bd-Oxidase (Anraku & Gennis, 1987; Weiss et al., 1991; Puustinen et al., 1991; Walker, unterschiedlicher 1992; Abb. 2-1). Energieausbeute Die Kombination ermöglichen E. von coli Enzymkomplexen mit abhängig den von Wachstumsbedingungen einerseits eine optimierte ATP-Synthese und andererseits eine schnelle Reoxidation des NADH. Unter aeroben Bedingungen sind hauptsächlich die alternative Dehydrogenase und die bo3-Oxidase aktiv, unter mikroaeroben Bedingungen der Komplex I und die bd-Oxidase (Unden et al, 2002). Abbildung 2-1: Schematische Darstellung der aeroben Elektronentransportkette aus E. coli. Die blauen Kästen stellen die primären Dehydrogenasen, die NADH:Ubichinon Oxidoreduktase (NDH I) und die alternative NADH Dehydrogenase (NDH II), die roten Kästen die beiden terminalen Oxidasen dar. In gelb ist das lipophile Ubichinon gezeigt. Die Pfeile geben die Richtung der Elektronenübertragung von NADH auf Sauerstoff (O2) an. In grün ist das für die NDH I spezifische künstliche Substrat Desamino-NADH gekennzeichnet. Die alternative NADH Dehydrogenase besitzt eine zehnfach geringere Affinität zu NADH als der Komplex I und eine tausendfach geringere Affinität zu dem künstlichen Substrat Desamino-NADH (d-NADH). Bei enzymatischen Tests mit E. coli Membranen bietet d-NADH die Möglichkeit die Aktivitäten der beiden NADH Dehydrogenasen zu unterscheiden (Matsushita et al, 1987). 4 Einleitung 2.2. Die NADH:Ubichinon Oxidoreduktase (Komplex I) Die membranständige NADH:Ubichinon Oxidoreduktase ist der Haupteintrittspunkt von Elektronen in die Atmungskette. Das Enzym katalysiert die Elektronenübertragung von NADH auf Ubichinon (Q) und koppelt diesen Prozess mit dem Aufbau eines Protongradienten. Pro übertragenenden Elektronenpaar werden vier Protonen von der negativen Innenseite auf die positive Außenseite der Membran transportiert (Weiss et al, 1991; Walker, 1992). Gleichung 2-1 zeigt die Reaktionsgleichung des Komplexes, dabei stehen bzw. für die Protonen auf der negativen Innenseite bzw. auf der positiven Außenseite der Membran. 21 Die eukaryotische NADH:Ubichinon Oxidoreduktase, auch Komplex I genannt, besteht aus bis zu 45 Proteinuntereinheiten, hat eine Masse von etwa 1 MDa und enthält als Kofaktoren ein FMN und acht Fe-S-Zentren (Hirst et al, 2003; Carroll et al, 2006). Die bakterielle NADH:Ubichinon Oxidoreduktase (Komplex I) setzt sich in den meisten Organismen aus 14 Untereinheiten zusammen, hat eine Masse von etwa 500 kDa und enthält ein FMN und bis zu zehn Eisen-Schwefel (Fe-S)-Zentren (Friedrich et al, 1995; Sazanov & Hinchliffe, 2006). Homologe der 14 sogenannten Kernuntereinheiten wurden in bisher allen Organsimen mit einer funktionellen NADH:Ubichinon Oxidoreduktase nachgewiesen (Friedrich & Scheide, 2000). Zudem besitzen der bakterielle und der mitochondriale Komplex I die gleichen Kofaktoren für den Elektronentransport und werden durch die gleichen Hemmstoffe wie z.B. Rotenon und Piericidin A gehemmt (Friedrich et al, 1994; Degli Esposti, 1998). Der für Mutagenesestudien zugängliche bakterielle Komplex I stellt ein strukturell einfacheres Modell für den mitochondrialen Komplex I dar. Dysfunktionen im Komplex I führen beim Menschen zu neurodegenerativen Erkrankungen wie zum Beispiel der Parkinson´schen und Alzheimerschen Erkrankung. (Lin & Beal, 2006; Rhein et al., 2009). Zudem ist der Komplex I eine Quelle von reaktiven Sauerstoffspezies (engl.: reactive oxygen species; ROS) und somit vermutlich am Alterungsprozess beteiligt (Galkin & Brandt, 2005; Kussmaul & Hirst, 2006; Lin & Beal, 2006). Der Komplex I aus E. coli ist auf dem 15„020 bp großen nuo-Operon (von NADH:Ubichinon Oxidoreduktase) durch die Gene nuoA bis nuoN kodiert (Weidner et al., 1993). Die Gene 5 Einleitung nuoC und nuoD sind fusioniert, so dass sich der E. coli Komplex I aus 13 Proteinuntereinheiten mit einer Gesamtmasse von 535 kDa zusammensetzt (Friedrich, 1998). Aus elektronenmikroskopischen Aufnahmen ist bekannt, dass der Komplex I eine L-förmig Überalles-Struktur besitzt: ein Arm ragt ins Cytoplasma, der andere ist in die Membran eingebettet (Guenebaut et al., 1998). Dabei bilden die hydrophilen Untereinheiten NuoB, NuoCD, NuoE, NuoF, NuoG und NuoI den peripheren Arm und die hydrophoben Untereinheiten NuoA, NuoH, NuoJ, NuoK, NuoL, NuoM und NuoN den Membranarm. Alle bisher bekannten Kofaktoren, das FMN und die neun Fe-S-Zentren N1a, N1b, N2, N3, N4, N5, N6a, N6b und N7 liegen im peripheren Teil des Enzyms (Friedrich, 1998). Erst 2010 wurden die ersten Kristallstrukturen des gesamten Komplex I veröffentlicht, die Struktur des Komplex I aus Thermus thermophilus wurde bei einer Auflösung von 4.5 Å bestimmt (Efremov et al, 2010; Abb. 2-2). In Übereinstimmung mit den elektronenmikroskopischen Daten zeigt die Struktur einen etwa 180 Å langen Membranarm aus 63 transmembranen Helices und einen senkrecht dazu angeordneten peripheren Arm, der etwa 130 Å über die Membranebene herausragt. Die Struktur des peripheren Armes des Enzyms aus dem gleichen Organismus wurde bereits 2006 mit einer Auflösung von 3.3 Å gelöst (Sazanov & Hinchliffe, 2006, Abb. 2-2) und die Auflösung durch weitere Arbeiten auf 3.1 Å verbessert (Berrisford & Sazanov, 2009). Sie zeigt die sieben hydrophilen Untereinheiten des bakteriellen Komplex I, die nach der T. themophilus Nomenklatur als Nqo1 bis Nqo6 und Nqo9 bezeichnet werden. Die Struktur gibt Aufschluss über die Lage der NADH-Bindestelle, des nicht-kovalent gebundenen FMN und der neun Fe-S-Zentren. Dabei bilden sieben Fe-S-Zentren in der Reihenfolge N3 → N1b → N4 → N5 → N6a → N6b → N2 eine Elektronenübertragungskette zwischen dem FMN in der NADH Bindestelle und einer Position in der Nähe des Membranarms, an der die Ubichinonbindestelle vermutet wird. Die beiden Fe-S-Zentren N1a und N7 liegen abseits dieser Kette (Kap. 2.2.1.). Die Struktur enthält zusätzlich die hydrophile Untereinheit Nqo15. Homologe dieser Untereinheit sind jedoch nur in sehr nahen Verwandten von T. thermophilus enthalten (Sazanov & Hinchliffe, 2006). Nqo15 gehört zu der Familie der Frataxine, die vermutlich die Assemblierung von FeS-Zentren unterstützen (Stemmler et al., 2010). Das Frataxin-Ortholog in E. coli ist CyaY und kein struktureller Bestandteil des Komplex I. Die Deletion von cyaY hat in E. coli keinen Effekt auf die Assemblierung und die Aktivität des Komplex I (Pohl et al., 2007a). Die Verbindung des peripheren Arms mit dem Membranarm erfolgt über die Untereinheiten Nqo4 (NuoD-Domäne der Untereinheit NuoCD) und Nqo6 (NuoB) im hydrophilen Teil und 6 Einleitung hauptsächlich durch Nqo8 (NuoH) im Membranarm. Die Untereinheiten Nqo7, 10 und 11 (NuoA, J, K) des Membranarms liegen in enger Nachbarschaft und sind vermutlich mit dem N-Terminus von Nqo4 in Kontakt. Sie tragen damit auch zur Verbindung von peripheren Arm und Membranarm bei. Die Elektronendichten in diesem Bereich waren jedoch nicht ausreichend, um die Kontaktfläche strukturell zu bestimmen. Die Struktur enthält kein gebundenes Ubichinon, die genaue Lage der Ubichinonbindestelle ist also unbekannt. Vermutlich liegt sie in einem Hohlraum im Bereich der Kontaktfläche zwischen der Untereinheit Nqo6 (NuoB) und Nqo4 (NuoD-Domäne der Untereinheit nuoCD). Diese Vermutungen werden unterstützt durch die veränderte Inhibitorsensivität von Mutationen in diesem Bereich des Komplex I aus Rhodobacter capsulatus und Yarrowia lipolytica (Prieur et al., 2001; Fendel et al., 2008). Aus Abstandsmessungen mit einem Spin-markierten Komplex I aus E. coli und einem ebenfalls markierten Decyl-Ubichinon mittels ESR-Spektroskopie ergab sich ein Abstand von etwa 15 Å zwischen dem hydrophoben Alkylrest des Ubichinons und des Fe-S-Zentrums N2 (Spatzal et al., 2010). Dies steht im Einklang mit der vermuteten Position der Ubichinonbindestelle. A) B) Nqo1 (NuoF) Nqo3 (NuoG) Nqo2 (NuoE) Nqo5 (NuoC) Nqo4 (NuoD) Nqo2 Nqo3 FMN Nqo1 Nqo15 Nqo9 (NuoI) Nqo15 Nqo5 Nqo6 (NuoB) Nqo9 Nqo8 (NuoH) Nqo6 Nqo12 (NuoL) Nqo13 (NuoM) Nqo14 (NuoN) Nqo7, 10, 11 (NuoA, J, K) Nqo4 Abbildung 2-2: Kristallstruktur des Komplex I aus T. thermophilus. A) zeigt die Struktur des gesamten Komplexes mit einer Auflösung von 4.5 Å (Efremov et al., 2010) und B) zeigt die Struktur des hydrophilen Arms mit einer Auflösung von 3.3 Å (Sazanov & Hinchliffe, 2006). Die Untereinheiten sind farblich unterschiedlich gekennzeichnet und nach der T. thermophilus Nomenklatur (E. coli Nomenklatur) benannt. Die Fe-S-Zentren sind als gelb-rote Sphären gezeigt, das FMN in magenta, die vermutete Lage der Ubichinonbindestelle ist mit einem Q gekennzeichnet. 7 Einleitung Neben der Gesamtstruktur des T. thermophilus Komplex I wurde auch die Struktur des Membranarms aus E. coli mit einer Auflösung von 3.9 Å gelöst (Efremov et al, 2010; Abb. 2-3). Die E. coli Struktur wurde benutzt, um durch molecular replacement den hydrophoben Teil der Gesamtstruktur aus T. themophilus zu lösen. Im Folgenden wird der Membranarm anhand der E. coli Struktur beschrieben. Sie zeigt die sechs hydrophoben Untereinheiten NuoA, NuoJ, NuoK, NuoL, NuoM, und NuoN. Die ebenfalls zu dem Membranarm zählende Untereinheit NuoH fehlt in dem Kristall. Die Auflösung von 3.9 Å lässt die Anordnung von insgesamt 55 transmebranen α-Helices erkennen, die sich über eine Länge von 160 Å erstrecken. Die Höhe des Arms beträgt 40 Å, was der Dicke der cytoplsmatischen Membran entspricht. Die Untereinheiten NuoL, M und N sind homolog zueinander und werden jeweils aus 14 transmembranen Helices aufgebaut, vier Helices bilden dabei einen Kern, der von den übrigen zehn Helices ringförmig umgeben ist. An den homologen Positionen dieser Untereinheiten befinden sich jeweils zwei Helices, die auf halber Höhe durch einen kurzen loop unterbrochen sind. Diese diskontinuierlichen Helices werden als funktionell wichtiger Bestandteil für Transporterproteine und Kanäle angenommen (Screpanti & Hunte, 2007). Es wird deshalb diskutiert, dass diese drei Untereinheiten für den Protonentransport verantwortlich sind. Das steht im Einklang zu früheren Erkenntnissen, dass NuoL, M und N homolog zu Untereinheiten der H+/Na+ bzw. H+/K+-Antiportern sind (Friedrich & Scheide, 2000; Mathiesen & Hägerhäll, 2002). Die homologen Untereinheiten wurden in der Struktur aufgrund von elektronenmikroskopischen Aufnahmen von Subkomplexen ohne die Untereinheiten NuoL und NuoM zugeordnet (Baranova et al., 2007). Die Aufnahmen zeigen, dass diese beiden Untereinheiten am distalen Ende des Membranarms liegen. Auffallend ist eine lange α-Helix, die als HL bezeichnet wird. Sie erstreckt sich über 110 Å entlang des Membranarms und zeigt in der Mitte einen Knick. Sie wurde anhand von Sequenzvergleichen der C-terminalen Domäne der distal gelegenen Untereinheit NuoL zugeordnet und endet auf Höhe der Untereinheit NuoN mit einer weiteren transmembranen Helix. Dieses ungewöhnliche Strukturmerkmal koppelt vermutlich den Elektronentransfer im peripheren Arm mit der in mehr als 100 Å entfernt gelegenen Protonentranslokation im Membranarm. Diese Vermutung wird durch eine E. coli Komplex I Variante, der die Untereinheit NuoL fehlt, unterstützt (S. Steimle, pers. Mitteilung). Die Variante enthält alle ESR-spektroskopisch detektierbaren Fe-S-Zentren, die NADH:Decyl-Ubichinon Oxidoreduktase Aktivität ist unverändert, aber die Protonentranslokation ist in etwa halbiert. 8 Einleitung A) NuoL NuoM NuoN NuoA, J, K B) Abbildung 2-3: Kristallstruktur des Membranarms des Komplex I aus E. coli (Efremov et al., 2010). A) zeigt die Seitenansicht in der Membranebene, B) die Aufsicht aus Richtung des peripheren Arms. Die einzelnen Untereinheiten sind farblich markiert, die hydrophobe Untereinheit NuoH fehlt in der Struktur. Die diskontinuierlichen Helices jeder Untereinheit sind orange und bei Kontakt mit der HL-Helix rot dargestellt. Nur wenige Monate nach der Veröffentlichung des Gesamtstruktur des bakteriellen Komplex I wurde die Gesamtstruktur des mitochondrialen Komplex I aus Y. lipolytica mit einer Auflösung von 6.3 Å veröffentlicht (Hunte et al., 2010; Abb. 2-4). Auch in diesem Organismus liegt der Komplex L-förmig vor. Der Membranarm hat eine Länge von 180 Å und insgesamt 71 transmembrane α-Helices wurden modelliert. Der periphere Arm enthält alle bekannten Fe-S-Zentren, deren Lage und deren Abstände denen im bakteriellen Komplex ähneln. Im Membranarm wurden ebenfalls diskontinuierliche, transmembrane Helices beschrieben. Außerdem wurde parallel zum Membranarm auf einer Länge von 60 Å eine Elektronendichte beobachtet, die vermutlich einer α-Helix zuzuordnen ist und der langen HLHelix der Untereinheit NuoL des bakteriellen Komplex I entspricht. Das unterstützt die Vermutung, dass diese Helix eine entscheidende Rolle bei der Kopplung von Elektronentransfer und Protonentranslokation spielt. Aufgrund dieser Daten wurde der Mechanismus des Komplex I mit einer Dampflok verglichen (Efremov et al, 2010). 9 Einleitung Abbildung 2-4: Kristallstruktur des Komplex I aus Y. lipolytica (Hunte et al., 2010). A) zeigt die Elektronendichtekarte des Komplexes, die Lage der Fe-S-Zentren ist vergrößert dargestellt. B) zeigt das helikale Model des Membranarms mit Blick auf die Membran, C) die seitliche Ansicht. Zur Orientierung ist die Lage der Fe-S-Zentren angegeben. Die beiden Domänen des mitochondrialen Komplex I sind farblich unterschieden, die lange Helix entlang des Arms ist in violett gezeigt. Phylogenetische Studien zeigen, dass der Komplex I evolutionär aus einem NADH Dehydrogenase Modul, einem Hydrogenasemodul und einem Transportermodul entstanden ist (Friedrich & Weiss, 1997; Abb. 2-5). Durch Erhöhung des pH-Wertes und Wechsel des Detergenz zerfällt der Komplex in drei Fragmente, das NADH Dehydrogenase Fragment, das verbindende Fragment und das Membranfragment. Die Fragmente entsprechen in ihrer Untereinheiten-Zusammensetzung in etwa derer der drei Module (Leif et al., 1995). Das NADH Dehydrogenasemodul, das mit dem NADH Dehydrogenase Fragment identisch ist, setzt sich aus den Untereinheiten NuoE, NuoF und NuoG zusammen und bildet den Elektroneneingang des Enzyms. Es enthält die NADH-Bindestelle, das FMN und das vierkernige Fe-S-Zentrum N3 auf der Untereinheit NuoF, das zweikernige Fe-S-Zentrum N1a auf der Untereinheit NuoE sowie das zweikernige Zentrum N1b und die vierkernigen Fe-SZentren N4, N5 und N7 auf der Untereinheit NuoG. Homologe dieser Untereinheiten sind in vielen, nicht energiewandelnden NAD(P)H abhängigen Hydrogenasen und Dehydrogenasen zu finden (Friedrich et al., 1995; Tersteegen & Hedderich, 1999). Das Hydrogenase-Modul setzt sich aus den Untereinheiten NuoB, NuoCD, NuoH, NuoI und NuoN zusammen, es enthält die vierkernigen Fe-S-Zentren N6a, N6b und N2 und die Ubichinonbindestelle. Es ist homolog zu multimeren, membranständigen Ni/Fe-Hydrogenasen, die als redoxgetriebene 10 Einleitung Ionenpumpen arbeiten (Friedrich & Scheide, 2000). Die Untereinheiten NuoA, NuoJ, NuoK, NuoL und NuoM bilden das Transportermodul, der Name ergibt sich aus der Homologie der Untereinheiten NuoL und NuoM zu Untereinheiten der H+/Na+ bzw. H+/K+-Antiportern (Friedrich & Scheide, 2000; Mathiesen & Hägerhäll, 2002). Abbildung 2-5: Modularer Aufbau des Komplex I aus E. coli. Das NADH Dehydrogenase Modul ist in gelb gezeigt, das Hydrogenase Modul in rot und das Transportemodul in blau. Die Lage der NADH- und Ubichinonbindestelle und der Fe-S-Zentren und die Wege der Protonen sind angedeutet. Die Membran ist schematisch in grau gezeigt. 2.2.1. Elektronentransfer im Komplex I Seit der Veröffentlichung der Struktur des peripheren Arms aus T. thermophilus ist die Lage der Fe-S-Zentren im Komplex I bekannt (Abb. 2-6). Die aus der Struktur ersichtlichen Abstände zwischen den Kofaktoren lassen vermuten, dass die Fe-S-Zentren eine Elektronentransportkette zwischen dem FMN, das die Elektronen vom NADH aufnimmt, und der Ubichinonbindestelle bilden. Das Fe-S-Zentrum N7 liegt 20.5 Å von den anderen Zentren entfernt und ist nicht Teil der Kette. Aus ESR-spektroskopischen Experimenten ist bekannt, dass N7 durch das Substrat NADH nicht reduziert wird, was eine Beteiligung am 11 Einleitung Elektronentransport unwahrscheinlich macht. Nur durch chemische Reduktion mit Dithionit wurden ESR-Signale dieses Zentrums detektiert (Tab. 2-1). Mutagenesestudien am Bindemotiv auf der Untereinheit NuoG haben gezeigt, dass N7 wichtig für die Stabilität des Komplex I ist (Pohl et al., 2007b). Abbildung 2-6: Lage der Fe-S-Zentren im peripheren Arm des Komplexes I aus T. thermophilus (PDB ID: 2FUG; Sazanov und Hinchliffe, 2006). Die Zentren sind als sphärische Kugeln dargestellt und anhand ihrer ESR-Spektren zugeordnet. Angegeben sind die Abstände zwischen den Mittelpunkten bzw. zwischen den Kanten (Werte in Klammern) der Fe-S-Zentren in Å. Die Pfeile markieren den angenommenen Elektronenübertragungsweg. Tabelle 2-1 gibt eine Übersicht über die durch ESR-Spektroskopie nachgewiesenen Fe-SZentren und ihrer Mittenpunktspotentiale des Komplex I aus E. coli. Die Signale der zweikernigen Fe-S-Zentren N1a und N1b sind bei 40 K im ESR-Spektrum sichtbar, die Signale der schneller relaxierenden vierkernigen Fe-S-Zentren werden bei 13 K detektiert. Das auf der Untereinheit NuoG koordinierte Fe-S-Zentrum N5 wurde bisher nur in einem rekombinanten Fragment des Komplexes nachgewiesen und ist vermutlich ein sehr schnell relaxierendes Zentrum und damit ESR-spektroskopisch schwierig nachzuweisen (Nakamaru12 Einleitung Ogiso et al., 2008). Im Komplex I aus Paracoccus denitrificans besitzt N5 ein Mittenpunktspotential von -250 mV (Yano et al., 2003). Die beiden auf der Untereinheit NuoI koordinierten Fe-S-Zentren N6a und N6b wurden im E. coli Komplex I UV-vis spektroskopisch nachgewiesen, die Mittenpunktspotentiale liegen jeweils bei -270 mV (Rasmussen et al., 2001). ESR-Spektren des verbindenden Fragmentes aus den Untereinheiten NuoB, NuoCD und NuoI zeigen ein breites, axiales Signal (gx = 1.89, gy = 1.91 und gz = 2.09; Rasmussen et al., 2001), das den Fe-S-Zentren N6a oder N6b zugeordnet wird (Ohnishi & Nakamura-Ogiso, 2008). In einer weiteren Veröffentlichung wurde dieses Signal im gesamten Komplex I aus E. coli nachgewiesen, die zugehörige Präparation ist jedoch nach Angaben der Autoren mit der Succinat Dehydrogenase und einem weiteren Fe-S-Zentrum haltigen Protein verunreinigt, die ebenfalls zu Signalen in dem betreffenden spektralen Bereich führten (Belevich et al., 2007). Tabelle 2-1: ESR-spektroskopisch nachgewiesene Fe-S-Zentren im Komplex I aus E. coli. Die Werte der Mittenpunktspotentiale und die g-Werte wurden folgenden Veröffentlichungen entnommen: Leif et al, 1995; Uhlmann & Friedrich 2005; Pohl et al., 2007b). . Fe-SZentrum Koordinierende Untereinheit Mittenpunktpotential [mV] Feldposition gx gy gz N1a NuoE -330 1.92 1.96 2.00 N1b NuoG -230 1.94 1.94 2.03 N2 NuoB -220* 1.91 1.91 2.05 N3 NuoG -270 1.88 1.92 2.04 N4 NuoG -270 1.89 1.93 2.09 N7 NuoG ~ -250 1.89 1.95 2.05 *Das Mittenpunktspotential von N2 ist pH-abhängig, dieser Wert wurde bei pH 7.0 gemessen (Flemming et al., 2006). Die Zuordnung der Signale zu den einzelnen Fe-S-Zentren in dieser Arbeit und auch die Benennung der Zentren in der Kristallstruktur erfolgten nach Ohnishi (Ohnishi, 1998; Ohnishi & Nakamura-Ogiso, 2008). Seit einigen Jahren wird die Zuordnung intensiv diskutiert. Hirst und Mitarbeiter schließen aus dem Vergleich der ESR-Signale der isolierten E. coli Untereinheit NuoG im Vergleich zum Gesamtkomplex und aus Doppel-ElektronenElektronen-Resonanz (DEER)-spektroskopischen Experimenten, dass das dem auf NuoG liegenden N4 zugeordnete Signal eigentlich dem auf NuoI liegenden N6a zuzuordnen ist. Das 13 Einleitung nur in einem Fragment detektierte und dem Fe-S-Zentrum N5 auf der Untereinheit NuoG zugeordnete Signal, ordnet diese Arbeitsgruppe dem Fe-S-Zentrum N4 zu (Yakovlev et al., 2007; Roessler et al., 2010). Die genaue Zuordnung der Signale zu den einzelnen Fe-SZentren muss noch geklärt werden. Aus der Lage und den Mittenpunktspotentialen der Kofaktoren lässt sich der wahrscheinliche Weg der Elektronen durch den Komplex I ableiten. Das FMN nimmt zwei Elektronen vom NADH auf und bildet einen Schalter zwischen dieser Zwei-Elektronenübertragung und den Ein-Elektronenübertragungen auf die Fe-S-Zentren. Für die Abgabe des ersten Elektrons (FMNH·/ FMNH2) liegt das Mittenpunktpotential bei -300 mV (Sled et al., 1994; Kussmaul et al., 2006), damit wird das erste Elektron vermutlich auf N3 (-270 mV) und dann weiter über die Kette auf N2 (-220mV) übertragen, eine Übertragung auf N1a ist wegen seines niedrigen Mittenpotentials (-330 mV) unwahrscheinlich. Diese ist erst bei der Übertragung des zweiten Elektrons möglich, da das Mittenpunktspotential des FMN/FMNH·-Paares bei -390 mV liegt (Sled et al., 1994; Kussmaul et al., 2006). Vermutlich wird das zweite Elektron dann von N1a über das FMN auf N3 übertragen, nachdem das Zentrum N3 reoxidiert wurde, und durchläuft so auch die Kette bis N2. Diese zeitweilige Übertragung des Elektrons auf N1a könnte die Bildung eines Radikals am FMN vermeiden. Die Funktion des N1a wäre also die Verminderung der ROS-Bildung am Komplex I und damit hätte das Fe-S-Zentrum eine wichtige Schutzfunktion für den Organismus gegen die Bildung der gefährlichen Sauerstoffradikale. Es ist bekannt, dass der Komplex I für die Bildung von ROS verantwortlich ist und das solvensexponierte FMN wird als ein Ort der Entstehung diskutiert (Kussmaul & Hirst, 2006; Esterhazy et al., 2008; Berrisford & Sazanov, 2009). Die Abstände zwischen den Kanten der einzelnen Fe-S-Zentren sind alle kleiner 14 Å und liegen damit in einem Bereich, der eine direkte Elektronenübertragung zulässt (Page et al., 1999). Eine Ausnahme bildet der Abstand zwischen den Zentren N5 und N6a (14 Å von Kante zu Kante, 16.9 Å zwischen den Mittelpunkten), er liegt damit an der Grenze, die eine direkte Elektronenübertragung wahrscheinlich macht (Kap. 2.3.). Grund für diesen großen Abstand ist vermutlich die Tatsache, dass die beiden Zentren auf unterschiedlichen Modulen liegen und dieser Übertragungsweg evolutionär aus der Anlagerung zweier Proteine entstanden ist (Friedrich & Weiss, 1997). 14 Einleitung Während dieser Arbeit wurden Kinetiken über die Elektronenübertragung innerhalb des Komplex I aus E. coli veröffentlicht (Verkhovskaya et al., 2008). Es wurden Zeitkonstanten von 90 µs für die Elektronenübertragung auf das Fe-S-Zentrum N1a und das am Ende der Elektronentransportkette liegende N2 bestimmt, die einer Übertragung der Elektronen vom ersten gebundenen NADH Molekül zugeordnet werden. Außerdem wurden Zeitkonstanten von 1.2 ms für das zweikernige Fe-S-Zentrum N1b und das vierkernige Fe-S-Zentrum N6b bestimmt. Diese wurden der Übertragung von einem zweiten NADH Molekül zugeordnet, das gebunden wurde, nachdem NAD+ die Bindestelle verlassen hatte. Da aber keine Übertragung der Elektronen auf ein in der Präparation vorliegendes Ubichinon detektiert wurde, ist fraglich, ob tatsächlich die physiologische Reaktion beobachtet wurde. 2.2.2. Überproduktion des Komplex I und seiner Varianten in E. coli Das Plasmid pBADnuo nuoFHis enthält das gesamte nuo-Operon und kodiert zudem ein Hexahistidin-Affinitätspeptid (His-Tag) N-terminal an der Untereinheit NuoF (Pohl et al., 2007c). Im Expressionstamm ANN0221 wurde auf dem Genom in das Gen nuoB eine nptIKassette, die eine Kanamycin Resistenz vermittelt, insertiert. Zudem ist das Gen ndh, das die alternative NADH Dehydrogenase kodiert, durch eine tet-Kassette, die eine Tetracyclin Resistenz vermittelt, ausgetauscht. Dieser Stamm ist also weder in der Lage, den Komplex I noch die alternative Dehydrogenase zu produzieren (Spehr, 1996). Die Transformation des Stammes ANN0221 mit dem Plasmid pBADnuo nuoFHis liefert ein System zur Überproduktion des Komplex I, der anschließend effizient mittels Affinitätschromatographie gereinigt werden kann. Durch Einfügen von Mutationen in das Plasmid pBADnuo nuoFHis ist es mit diesem System möglich, Varianten des Komplex I zu produzieren (Pohl et al., 2007c). Aufgrund seiner Größe von mehr als 20 kbp ist es nicht möglich in das Plasmid pBADnuo nuoFHis mittels der QuikChange Methode direkt Punktmutationen einzufügen. Aus diesem Grund werden Mutationen erst in die einzelnen nuo-Gene die auf Plasmiden mit einer Größe von 4-7 kbp liegen, eingefügt und die mutierten Gene dann mittels λ-Red vermittelter Rekombination (Murphy, 1998; Datsenko & Wanner, 2000) in pBADnuo nuoFHis eingebracht. Dafür werden Zellen des Stammes DH5αΔnuo (Pohl, 2008) verwendet. Diesen Zellen fehlt das gesamte nuo-Operon im Genom, bei der Rekombination findet also nur ein Strangaustausch mit der Zielsequenz auf dem Plasmid statt. Für die Selektion wird das nptI15 Einleitung sacR-sacB Selektionssystem genutzt (Ried und Collmer, 1987). Das nptI Gen vermittelt eine Kanamycin Resistenz und ist somit ein positiver Selektionsmarker. Das sacB-Gen aus Bacillus subtilis kodiert für die Levansucrase, ein Enzym, das Saccharose in Levan umbaut. Levan akkumuliert in den Zellen und führt zur Zelllyse (Gay et al., 1985). Gram-negative Bakterien, die diesen negativen Selektionsmarker zusammen mit seinem Regulator sacR enthalten, können nicht auf Saccharose-haltigen Medien wachsen. Nutzt man dieses Selektionssystem, um die Ausbeute an gewünschten Mutationen pro Absatz zu erhöhen, sind zwei Rekombinationsschritte notwendig: Im ersten Schritt wird das entsprechende nuo-Gen gegen die nptI-sacB-Kassette ausgetauscht und positive Klone auf Kanamycin haltigem Medien selektiert. Im zweiten Rekombinationsschritt wird die nptI-sacB-Kassette durch das mutierte Gen ersetzt, die Selektion erfolgt auf Saccharose-haltigem Medium. 2.3. Beteiligung aromatischer Aminosäuren am Elektronentransport Der Elektronentransport in Proteinen ist für viele energiewandelnde, biologische Prozesse, wie zum Beispiel der aeroben Atmung oder der Photosynthese, von essentieller Bedeutung. Welche Rolle das Protein selbst dabei spielt, wird seit Mitte des 20. Jahrhunderts diskutiert (Gray & Winkler, 2003). Prinzipiell unterscheidet man zwischen dem through-bond Transfer, bei dem die Elektronen über б-Bindungen zwischen den Redoxzentren wandern, und dem through-space Transfer, bei dem die Elektronen von Redoxzentrum zu Redoxzentrum tunneln. Die Theorie des through-space Transfers hat sich zur Beschreibung der Elektronenübertragung in Proteinen allgemein durchgesetzt. Ihr liegt die Marcus-Theorie zu Grunde, die den Transfer von Elektronen zwischen einem Akzeptor und einem Donor beschreibt, die nicht über eine chemische Bindung miteinander verbunden sind. Die Wechselwirkungen der relevanten Molekülorbitale der einzelnen Kofaktoren sind also so schwach, dass die Elektronen zum Tunneln gezwungen werden, das Protein bildet dabei lediglich die Tunnelbarriere. Die Arbeitsgruppe von Dutton hat diese Theorie so weit vereinfacht, dass die Übertragungsrate nur noch vom Abstand zwischen den beiden Redoxzentren beschrieben wird. Die Elektronen werden nach diesen Berechnungen unabhängig von der Proteinumgebung über eine Entfernung von bis zu 14 Å zwischen den Redoxpartnern übertragen (Page et al., 1999). Ergänzend zu dieser Theorie wird diskutiert, dass auch aromatische Aminosäuren der Proteine am Elektronentransport beteiligt sind. Diese als hopping bezeichnete Theorie nimmt an, dass die Elektronen nicht nur auf den im Protein 16 Einleitung koordinierten Redoxzentren lokalisiert sind, sondern zusätzlich in den л-Elektronensystemen aromatischer Aminosäuren. Dadurch werden auch Elektronenübertragungen zwischen Redoxzentren mit einem Abstand von mehr als 14 Å wahrscheinlich (Stubbe et al., 2003; Shih et al., 2008; Wittekindt et al., 2009). In der Literatur sind mehrere Beispiele zu finden, in denen die Beteiligung aromatischer Aminosäuren am Elektronentransport diskutiert wird. In der Klasse I Ribonukleotid Reduktase aus E. coli aktiviert ein Cystein auf der Untereinheit α2 ein Tyrosylradikal auf der Untereinheit β2, das 35 Å entfernt liegt. Die Elektronen werden über diese große Distanz vermutlich über ein Tryptophan und drei Tyrosinreste übertragen (Rova et al., 1999; Stubbe et al., 2003; Seyedsayamdost et al., 2007; Bollinger, 2008). Ein weiteres Beispiel ist ein modifiziertes Azurin aus Pseudomonas aeruginosa, in dem durch Einfügen eines Tryptophanrestes die Elektronenübertragungsrate um das 300-fache gesteigert wurde (Shih et al., 2008). Auch in der bakteriellen Cytochrom c Peroxidase weisen strukturelle und spektroskopische Untersuchungen in Verbindung mit Mutagenese-Studien daraufhin, dass ein Tryptophan am Elektronentransport zwischen zwei Häm-Gruppen beteiligt ist (Dias et al., 2004; De Smet et al., 2006). Die Elektronentransferraten der einzelnen Übertragungsschritte im Komplex I wurden sowohl mit der Theorie des Tunnelns (Moser et al., 2006), als auch mit der Theorie des hoppings (Wittekindt et al., 2009) berechnet. Nach Moser et al. erlauben die Abstände der einzelnen Fe-S-Zentren ein direktes Tunneln der Elektronen mit Transferraten, die mit der experimentell bestimmten Gesamtrate des Proteins im Bereich von 100 s-1 übereinstimmen. Nach Wittekindt et al. lässt sich die experimentell bestimmte Transferrate des Gesamtprozesses nur unter Beteiligung von aromatischen Aminosäuren erklären. Berechnungen unter Annahme des through-space Mechanismus führten zu Transferraten zwischen den Fe-S-Zentren N5 und N6a von 2.2 * 10-4 s-1, die nicht mit dem experimentellen Wert in Einklang zu bringen sind. Werden jedoch konservierte, aromatische Aminosäuren als hopping-Partner zwischen den FeS-Zentren in die Rechnung miteinbezogen, erhöht sich die Transferrate für diesen Schritt auf 2 * 103 s-1. Aufgrund der Berechnungen und mit Hilfe der Struktur des hydrophilen Teils des Komplex I aus T. thermophilus werden sieben, konservierte aromatische Aminosäuren als hopping Partner vorgeschlagen (Wittekindt et al., 2009; Abb. 2-7), vier davon liegen zwischen den Fe-S-Zentren N5 und N6a. Experimentelle Mutagenesestudien liegen zu diesem Thema bisher nicht vor. Der in einem anderen Zusammenhang untersuchte Austausch des konservierten Histidins zwischen N5 und N6a gegen ein Alanin im Komplex I aus Y. 17 Einleitung lipolytica führte zu einer 50%igen Erniedrigung der Aktivität des Komplex I im Vergleich zum unveränderten Protein (Kashani-Poor et al., 2001). Abbildung 2-7: Die Lage der Fe-S-Zentren des Komplex I und der nach der hopping Theorie möglicherweise am Elektronentransport beteiligten aromatischen Aminosäuren. Die Fe-S-Zentren sind als sphärische Kugeln dargestellt und die Aminosäuren nach der T. thermophilus Nomenklatur benannt (PDB ID: 2FUG; Sazanov und Hinchliffe, 2006). Die Pfeile markieren den angenommenen Elektronenübertragungsweg. 2.4. Die Microsecond Hyper-Quenching (MHQ)-Technik Zum Verständnis biologischer Prozesse ist neben der Kenntnis der Struktur der beteiligten Enzyme deren Reaktionskinetik von großer Bedeutung. Eine wichtige Voraussetzung für die Untersuchung dieser Kinetik sind Versuchsaufbauten, die einen definierten Startzeitpunkt der Reaktion durch schnelles Mischen von Enzym und Substrat garantieren und eine zum Reaktionsstart zeitnahe Detektion oder ein Beenden der Reaktion zu einem definierten 18 Einleitung Zeitpunkt erlauben. Bei den sogenannten Strömungsverfahren unterscheidet man grundsätzlich zwischen der Continous-Flow-, der Stopped-Flow- und Quenched-FlowMethode. Beim Continous-Flow Verfahren, dass im Jahre 1923 von Hartridge und Roughton entwickelt wurde, treffen das Enzym und das Substrat in einer Mischungskammer aufeinander und fließen anschließend in ein Beobachtungsröhrchen. Bei bekannter Fließgeschwindigkeit entspricht die Ortskoordinate im Beobachtungsröhrchen der Zeitkoordinate der Reaktion (DuBois, 1941). Bei dem zuerst von Chance (1943) beschriebenen und von Gibson und Milnes (1964) weiterentwickelten Stopped-flow Verfahren werden die zu mischenden Substanzen mit zwei Spritzen in die Mischungskammer und anschließend in die Beobachtungskammer eingebracht. Eine dritte Spritze hinter der Beobachtungskammer, die Stopp-Spritze, wird dadurch bis zu einem definierten Anschlag zurückgeschoben, daraufhin bricht der Fluss ab und die Detektion wird gestartet (Chance, 1943; Gibson & Milnes, 1964). Dieses Verfahren wurde über die Jahre weiterentwickelt, ist mit der UV/vis-, CD-, FTIR- und Fluoreszens-Spektroskopie kombinierbar und findet heute eine breite Anwendung. Die Totzeit der Apparaturen liegt bei etwa 1 ms, die eingesetzten Volumina bei 50 µL bis 1 mL. Das Stopped-Flow Verfahren ist jedoch nur in Verbindung mit Detektionsverfahren mit kurzer Ansprechzeit und Aufzeichnung der Daten in Echtzeit einsetzbar. Bei langsameren Detektionsmethoden wie zum Beispiel der ESR-Spektroskopie findet das Quenched-Flow Verfahren Anwendung. Die zu beobachtende Reaktion wird dabei chemisch, optisch oder durch abruptes Einfrieren beendet und die Proben anschließend analysiert. Das Ansetzen mehrerer Proben mit unterschiedlichen Reaktionszeiten ermöglicht die Verfolgung der Intermediatsbildung während der Reaktion. Zur Charakterisierung der Reaktionskinetik von Enzymen mit paramagnetischen Kofaktoren wie zum Beispiel Fe-S-Zentren bildet das Freeze Quench Verfahren in Verbindung mit der ESR-Spektroskopie eine wichtige Methode, da sich der Redoxzustand dieser Kofaktoren ESR-spektroskopisch nur bei Temperaturen unterhalb 80 K bestimmen lässt. Die erste Apparatur für das so genannte Rapid Freeze Quenching wurde in den sechziger Jahren von Bay entwickelt. Am Ende eines dem continous flow ähnlichem System werden die gemischten Substanzen durch eine Düse in eine Kryoflüssigkeit (flüssiges Isopentan, -140°C) gespritzt und dadurch eingefroren. Durch Weiterentwicklung dieses Verfahrens wurde ein zu beobachtender Reaktionszeitraum von 5 ms bis 5 s erreicht (Ballou et al., 1974). Der Nachteil des Verfahrens ist die hohe Totzeit (5 ms) und die starke Verdünnung der Probe in der Kryoflüssigkeit. Durch Veränderung zum Beispiel der Mikromixergeometrie, der Flussgeschwindigkeit oder des Düsendurchmesser wurden die Totzeiten weiter gesenkt werden (Shastry et al., 1998, Tanaka et al., 2003, Cherepanov & de 19 Einleitung Vries 2004). Auch durch das Einfrieren der Probe auf mit flüssigem Stickstoff gekühlten Metalloberflächen konnte die Totzeit weiter verringert werden (Tanaka et al., 2003; Lu et al., 2005). Dieses cold-block freezing wurde schon im Rahmen der Elektronenmikrokopie als die schnellste Methode zum Einfrieren von Proben diskutiert, innerhalb von einer Millisekunde wird die Temperatur eines Tropfens mit mehr als 10 µM Durchmesser um 100 K gesenkt (Robards and Steytr, 1985; Bald 1985). Zudem wurde durch das Einfrieren ohne die Verwendung von Kryoflüssigkeit die Probenkonzentration gesteigert (Lu et al, 2005). Diese Modifikationen führten zu Apparaturen mit Totzeiten unterhalb von 200 µs und wurde aus diesem Grund Microsecond Hyperquenching (MHQ)-Technik genannt (Cherepanov & de Vries, 2004). Die in dieser Arbeit verwendete Apparatur wurde im Labor von Prof. Simon de Vries, Technische Universität Delft, Niederlande, entwickelt und aufgebaut (Cherepanov & de Vries, 2004; Wiertz et al., 2004; Abb. 2-8). In einer Unterdruckkammer befinden sich ein Mikromixer und ein rotierender, mit Wolfram überzogener Aluminiumzylinder zum Sammeln der Probe, der vor Versuchsbeginn mit flüssigen Sticksoff auf 78 K gekühlt wird. Die gesamte Apparatur wird vor Beginn der Experimente mit einem mit Argon begasten Puffer gespült, um die Sauerstoffkonzentration zu minimieren. Das Enzym und das Substrat werden mittels zweier HPLC-Pumpen in den Mikromixer eingebracht, verlassen diesen durch eine Düse, treffen auf den rotierenden Zylinder und werden dort eingefroren. Der Abstand zwischen Mixerausgang und der Wolframoberfläche des Zylinders und damit die Flugzeiten kann variiert werden und so die Reaktionszeiten der Probe verändert werden. Unmittelbar nach dem Einfrieren der Probe auf der Zylinderoberfläche wird der Unterdruck in der Kammer durch Einlassen von Sticksoff entfernt und der Zylinder mit flüssigem Stickstoff gefüllt. Anschließend wird die Probe mit einem gewöhnlichen Holzlöffel in flüssigem Sticksoff vom Zylinder gesammelt. Für die ESR-Spektroskopie wird die Probe in flüssigem Stickstoff in ein speziell präpariertes ESR-Röhrchen überführt. Der Boden des Röhrchens ist ein Filter aus Polypropylen, durch den der flüssige Stickstoff entweichen kann. 20 Einleitung Abbildung 2-8: Aufbau des Apparates der MHQ-Technik. Gezeigt sind der Mikromixer und der rotierende Zylinder in der Unterdruckkammer. Während des Experiments sinkt der Mixer in den Rotor, so dass die Probe auf die Zylinderwände trifft, diese Bewegung ist durch den Pfeil angedeutet. Außerhalb der Kammer befinden sich die beiden HPLC-Pumpen, der mit Argon gespülte Puffervorrat, die loops für das Enzym (E) und sein Substrat (S) und die Vakuumpumpe. Abbildung 2-9 zeigt den Aufbau des tangentialen Mikromixers mit Kreuzgeometrie. Aus zwei gegenüberliegenden Kanalausgängen strömt die Lösung mit dem Enzym und trifft in der sogenannten pre-mixing Kammer auf die Lösung mit dem Substrat, die ebenfalls über zwei gegenüberliegenden Kanälen in den Mixer gelangt. Die Probe wird durch eine trichterförmige Düse mit einem Durchmesser von 20 oder 100 µm, abhängig von der angestrebten Reaktionszeit, in die Unterdruckkammer mit dem gekühlten Zylinder entlassen und dabei weiter durchmischt. Das pre-mixing-Volumen liegt bei 1 nL, das Mischungsvolumen in der Düse bei 10 nL. Weitere spiralförmige Aufsätze auf dem Mixer können den Weg zwischen Mixerausgang und Zylinder verlängern und ermöglichen somit unterschiedliche Reaktionszeiten im ms Bereich. 21 Einleitung Abbildung 2-9: Aufbau des tangentialen Mikromixers mit Kreuzgeometrie (Cherepanov & de Vries, 2004). 1) zeigt den Mixer, 2) die Düse mit 20 µM Durchmesser und 3) die Messingschraube, die den Deckel des Mixers bildet. Die zu mischenden Substanzen gelangen über Kanal A bzw. B in einer ABAB-Anordnung in den Mixer. Die Totzeit des Aufbaus wurde durch die Reaktion von Metmyoglobin mit Azid bei einer Reaktionstemperatur von 10°C bestimmt. Die Mischungszeit beträgt weniger als 20 µs, die minimale Transportzeit der Probe zwischen Mixer und der kalten Wolframoberfläche 25 µs und die Einfrierzeit 40 µs. Daraus ergibt sich eine Totzeit von 85 µs. Die Reaktionszeiten können in einem Bereich von 97 µs bis 60.9 ms mit definierten Zeitpunkten variiert werden (Cherepanov & de Vries, 2004; Wiertz et al., 2004). 2.5. Weitere untersuchte Metalloproteine 2.5.1. Die Superoxid Reduktase aus Methanosarcina mazei In aeroben Organismen besitzt die Superoxid Dismutase (SOD) eine Schlüsselrolle im Abbau von ROS. Seit einigen Jahren ist bekannt, dass in anaeroben und mikroanaeroben Bakterien, in denen keine SOD nachgewiesen werden konnte, die Superoxid Reduktase (SOR) am Abbau von ROS beteiligt ist (Jenney et al., 1999). Die SOR katalysieren die Reduktion von Superoxid zu Wasserstoffperoxid und wurden bisher in zwei Klassen unterteilt, die sich insbesondere durch ihre Kofaktoren unterscheiden. Die Enzyme beider Klassen besitzen das katalytisch aktive Zentrum [Fe(His4)(Cys)], während die Klasse I SOR zusätzlich ein 22 Einleitung [Fe(Cys)4] binden. Das Gen mm0632 des strikt anaeroben und methanogenen Archaea M. mazei kodiert für ein Protein, dass Superoxid Reduktase Aktivität zeigt (Krätzer et al., 2010). Die Sequenz des Proteins MM0632 enthält das konservierte Bindemotiv für das [Fe(His4)(Cys)]-Zentrum, während ein Bindemotiv für das [Fe(Cys) 4]-Zentrum fehlt. Zusätzlich existieren ein Bindemotiv der Form Cys(X) 7CysXXCys und ein viertes Cystein in dem im Vergleich zu den Klasse I und II SOR verlängerten C-Terminus des Proteins. Der Gehalt an Eisen und anorganischem Schwefel im Protein lässt vermuten, dass neben dem [Fe(His4)(Cys)]-Zentrum ein vierkerniges Fe-S-Zentrum im Protein koordiniert ist. Das Protein MM0632 ist vermutlich der Vertreter einer dritten Klasse der SOR (Krätzer et al., 2010). 2.5.2 Die Benzoyl-CoA Reduktase aus Geobacter metallireducens Viele aerobe Bakterien nutzen aromatische Verbindungen als Kohlenstoffquelle, dabei wird das stabile Ringsystem durch Oxygenasen unter Sauerstoffverbrauch abgebaut. Auch anaerobe Organismen katabolisieren Aromaten, bei der Destabilisierung des aromatischen Systems ist Benzyl-CoA (BCoA) ein häufig auftretendes Intermediat. In fakultativ anaeroben Bakterien wie zum Beispiel Thauera aromatica katalysieren BCoA Reduktasen den Elektronentransfer von Ferredoxin auf BCoA unter gleichzeitiger ATP-Hydrolyse. Diese Reaktion verläuft vermutlich ähnlich wie eine Birch-Reduktion und führt zur Dearomatisierung des stabilen Ringsystems (Wischgoll et al., 2005; Kung et al., 2009). In obligatorisch anaeroben Bakterien wird ein ATP unabhängiger Abbau von Aromaten vermutet. Im Genom des anaeroben Bakteriums Geobacter metallireducens wurden die durch Benzoat induzierbaren Gene bamB-I identifiziert und das Genprodukt, der BamBCDEFGHI Komplex, als eine bisher nicht charakterisierte BCoA Reduktase vorgeschlagen (Wischgoll et al., 2005). Der Komplex setzt sich aus verschiedenen Proteinkomponenten zusammen: BamGHI zeigt Ähnlichkeit zu dem NADH Dehydrogenase Fragment des Komplex I, BamC und BamF zu einzelnen Untereinheiten von Hydrogenasen und BamDE zu Heterodisulfid Reduktasen. BamB zeigt hohe Sequenzähnlichkeiten zu Wolfram- oder Molybdän-haltigen Aldehyd:Ferredoxin Oxidoreduktasen und trägt vermutlich das katalytische Zentrum. Die anderen Proteine des Komplexes sind wahrscheinlich an der Aufnahme der Elektronen von einem bisher unbekannten Donor und der Übertragung von Elektronen zu BamB beteiligt (Wischgoll et al., 2005). Ein BamBC genanntes Fragment mit einer Größe von etwa 185 kDa aus den Genprodukten BamB (73 kDa) und BamC (21 kDa) wurde anaerob isoliert und liegt 23 Einleitung vermutlich in einer α2β2-Form vor. Die im BamB-I Komplex für die Aufnahme der Elektronen zuständigen Proteine sind nicht mit dem isolierten Enzym assembliert, aus diesem Grund wird die Aktivität des Enzyms über die vermutete Rückreaktion bestimmt. BamBC katalysiert die Oxidation des Cyclohexa-1,5-dien-1-carboyl-CoA (Dienly-CoA) zu BCoA in Anwesenheit eines Elektronenakzeptors und stellt den Prototyp einer neuen Klasse von BcoA Reduktasen dar (Kung et al., 2009). 2.6. Ziel der Arbeit Die Geschwindigkeit der NADH:Decyl-Ubichinon Oxidoreduktase Aktivität des isolierten E. coli Komplex I beträgt etwa 50 s-1. Die Detektion des Redoxzustandes der einzelnen Fe-SZentren gelingt nur mittels Tieftemperatur ESR-Spektroskopie. Aus diesem Grund liegen bisher kaum experimentelle Ergebnisse zur Kinetik des Elektronentransfers zwischen den FeS-Zentren des Komplex I vor. In dieser Arbeit sollte diese Kinetik mit Hilfe der MHQTechnik und Tieftemperatur-ESR-Spektroskopie im ms- bis µs-Bereich erfasst werden. In der Literatur wird die Beteiligung aromatischer Aminosäuren am Elektronentransport in Proteinen diskutiert. Berechnungen zur Folge soll auch im Komplex I nur unter Beteiligung konservierter aromatischer Aminosäuren die experimentell bestimmte Transferrate des Gesamtprozesses der Elektronenübertragung von NADH auf Ubichinon erreicht werden. In dieser Arbeit sollten diese konservierten Aminosäuren im E. coli Komplex I gegen andere aromatische Aminosäuren und Leucin ausgetauscht und der Einfluss dieses Austausches auf die Aktivität des Enzym detektiert werden. Die Funktion des Fe-S-Zentrums N1a für den Mechanismus des Komplex I ist bisher nicht geklärt. Eine direkte Beteiligung an der Elektronentransportkette zwischen dem FMN und der Ubichinonbindestelle ist aufgrund seiner Lage ausgeschlossen. Eine Vermutung ist, dass es die Lebensdauer eines möglichen Flavinsemichinon-Radikals verkürzt und somit die Produktion schädlicher ROS am Komplex I vermindert. In dieser Arbeit sollten die Eigenschaften des Zentrums N1a durch Mutagenese seines Bindemotivs im E. coli Komplex I verändert und die entsprechenden Varianten charakterisiert werden. 24 Einleitung Ein weiteres Thema dieser Arbeit sollte die ESR-spektroskopische Charakterisierung zweier Proteine sein, die vermutlich Vertreter einer jeweils neuen Klasse von Enzymen darstellen. Das durch die Gene bamB und bamC kodierte Fragment BamBC aus G. metallireducens ist Teil einer Benzoyl-Coenzym A Reduktase und damit an dem Abbau aromatischer Verbindungen beteiligt. Das durch das Gen mm0632 kodierte Protein aus M. mazei ist vermutlich eine Superoxid Reduktase und somit am Abbau von ROS beteiligt. 25 Material und Methoden 3. MATERIAL UND METHODEN 3.1 Bakterienstämme und Vektoren Die Tabellen 3-1 und 3-2 zeigen die in dieser Arbeit verwendeten E. coli Stämme bzw. die verwendeten Vektoren. Tabelle 3-1: Verzeichnis der verwendeten E. coli Stämme. Stamm Genotyp AN387 Referenz Wallace & Young, 1977 AN387Δnuo AN387/Δnuo Pohl, 2008 ANN0221 AN387/ ndh::tet nuoB::nptI Spehr, 1996 BL21(DE3) F– ompT hsdSB (rB – mB–) gal dcm (DE3) Studier & Moffatt, 1986 BW25113 lacIq, rrnBT14, ΔlacZWj16, hsdR514, Datsenko & Wanner, 2000 araBADAH33, ΔrhaBADLD78 BW25113Δnuo BW25113/Δnuo Vranas, M. unveröffentl. Ergebnisse DH5α (Electromax) F– Φ80lacZΔM15 Δ lacZY A-argF) U169 Invitrogen recA1 endA1 hsdR17 (rk – mk +) gal– phoA supE44 λ– thi-1 gyrA96 relA1 DH5αΔnuo DH5α/Δnuo Pohl et al., 2007b Tabelle 3-2: Verzeichnis der verwendeten Vektoren. Vektor Genotyp Referenz pBADnuo nuoFHis camR, pACYC184 origin, araCParaBAD, nuoA-N, His6-nuoF Pohl et al., 2007c pBADnuo nuoFHis nuoCD::nptI-sacB pBADnuo nuoFHis nuoCD::nptI-sacB Morina, 2009 pBADnuo nuoFHis nuoCD F515L pBADnuo nuoFHis, nuoCD F515L Morina, 2009 pBADnuo nuoFHis nuoCD F515W pBADnuo nuoFHis, nuoCD F515W Morina, 2009 pBADnuo nuoFHis nuoCD F515Y pBADnuo nuoFHis, nuoCD F515Y Morina, 2009 26 Material und Methoden Tabelle 3-2, Fortsetzung. pBADnuo nuoFHis nuoCD H514L pBADnuo nuoFHis, nuoCD H514L diese Arbeit pBADnuo nuoFHis nuoE::nptI-sacB pBADnuo nuoFHis nuoE::nptI-sacB diese Arbeit pBADnuo nuoFHis nuoE C92A pBADnuo nuoFHis, nuoE C92A diese Arbeit pBADnuo nuoFHis nuoE C97A pBADnuo nuoFHis, nuoE C97A diese Arbeit pBADnuo nuoFHis nuoE C97S pBADnuo nuoFHis, nuoE C97S diese Arbeit pBADnuo nuoFHis nuoE C133A pBADnuo nuoFHis, nuoE C133A diese Arbeit pBADnuo nuoFHis nuoE C133S pBADnuo nuoFHis, nuoE C133S diese Arbeit pBADnuo nuoFHis nuoE C137A pBADnuo nuoFHis, nuoE C137A diese Arbeit pBADnuo nuoFHis nuoGbeg::nptI-sacB pBADnuo nuoFHis nuoGbeg::nptI-sacB diese Arbeit pBADnuo nuoFHis nuoG F35L pBADnuo nuoFHis, nuoG F35L diese Arbeit pBADnuo nuoFHis nuoG F35W pBADnuo nuoFHis, nuoG F35W diese Arbeit pBADnuo nuoFHis nuoG F35Y pBADnuo nuoFHis, nuoG F35Y diese Arbeit pBADnuo nuoFHis nuoG::nptI-sacB pBADnuo nuoFHis nuoG::nptI-sacB Pohl, 2008 pBADnuo nuoFHis nuoG W221L pBADnuo nuoFHis, nuoG W221L Morina, 2009 pBADnuo nuoFHis nuoG W221F pBADnuo nuoFHis, nuoG W221F diese Arbeit pBADnuo nuoFHis nuoG W221Y pBADnuo nuoFHis, nuoG W221Y diese Arbeit pBADnuo nuoFHis nuoI::nptI-sacB pBADnuo nuoFHis, nuoI::nptI-sacB Morina, 2009 pBADnuo nuoFHis nuoI F101L pBADnuo nuoFHis, nuoI F101L diese Arbeit pBADnuo nuoFHis nuoI F101W pBADnuo nuoFHis, nuoI F101W diese Arbeit pBADnuo nuoFHis nuoI F101Y pBADnuo nuoFHis, nuoI F101Y diese Arbeit pCA24N nuoE camR, ColE1 origin, lacI-PT5-lac His6-nuoE Kitagawa et al., 2005 pCA24N nuoE C92A pCA24N nuoE C92A diese Arbeit 27 Material und Methoden Tabelle 3-2, Fortsetzung. pCA24N nuoE C97A pCA24N nuoE C97A diese Arbeit pCA24N nuoE C97S pCA24N nuoE C97S diese Arbeit pCA24N nuoE C133A pCA24N nuoE C133A diese Arbeit pCA24N nuoE C133S pCA24N nuoE C133S diese Arbeit pCA24N nuoE C137A pCA24N nuoE C137A diese Arbeit pCA24N nuoCDGFP camR, ColE1 origin, lacI-PT5-lac His6-nuoCDGFP Kitagawa et al., 2005 pCA24N nuoCDGFP H514L pCA24N nuoCDGFP H514L diese Arbeit pCA24N nuoCDGFP F515L pCA24N nuoCDGFP F515L Morina, 2009 pCA24N nuoCDGFP F515W pCA24N nuoCDGFP F515W Morina, 2009 pCA24N nuoCDGFP F515Y pCA24N nuoCDGFP F515Y Morina, 2009 pCA24N nuoI camR, ColE1 origin, lacI-PT5-lac His6-nuoI Kitagawa et al., 2005 pCA24N nuoI F101L pCA24N nuoI F101L Morina, 2009 pCA24N nuoI F101W pCA24N nuoI F101W Morina, 2009 pCA24N nuoI F101Y pCA24N nuoI F101Y Morina, 2009 pET11a nuoB-G nuoFSTII ampR pBR322 origin, lacI-PT7-lac nuoB-G, nuoF-StrepTagII Bungert et al., 1999 pkD46 ampR , R101 origin, araC-ParaBAD exo, bet, gam Datsenko & Wanner, 2000 pUC25 nuoE´-G ampR, pUC origin, nuoE´-G, nuoFStrepTagII Kohlstädt, 2009 pUC25 nuoE´-G nuoG F35L pUC25 nuoE´-G nuoG F35L diese Arbeit pUC25 nuoE´-G nuoG F35W pUC25 nuoE´-G nuoG F35W diese Arbeit pUC25 nuoE´-G nuoG F35Y pUC25 nuoE´-G nuoG F35Y diese Arbeit pUC25 nuoE´-G nuoG W221L pUC25 nuoE´-G nuoG W221L Morina, 2009 pUC25 nuoE´-G nuoG W221F pUC25 nuoE´-G nuoG W221F diese Arbeit pUC25 nuoE´-G nuoG W221Y pUC25 nuoE´-G nuoG W221Y diese Arbeit 28 Material und Methoden 3.2 Oligonukleotide Die Tabellen 3-3, 3-4 und 3-5 zeigen die in dieser Arbeit verwendeten DNA-Oligonukleotide. Tabelle 3-3: Verzeichnis der verwendeten DNA-Oligonukleotide zur Einführung von Punktmutationen mit der QuikChange Methode. Die neu generierten Codons sind fett gedruckt, die ausgetauschten Basen sind kursiv und die neu eingefügten Restriktionsschnittstellen sind unterstrichen dargestellt. Das für die Mutagenese benötigte zweite DNA-Oligonukleotid hat die entsprechende revers-komplementäre Sequenz. Neue Oligonukleotid Sequenz Schnittstelle BauI nuoCD H514L fwd 5´-GAAACCCTGATCACCCTGTTCCTGCAAGTCTC GTGGGGTCC-3´ nuoCD F515L fwd 5´-GAAACCCTGATCACCCACTTGCTGCAAGTCTC BauI GTGGGGTCC-3´ nuoCD F515W fwd 5´-CTGATCACCCACTGGCTGCAAGTCTCGTGGG BauI GTC-3´ nuoCD F515Y fwd 5´-GAAACCCTGATCACCCACTATCTGCAAGTCTC BauI GTGGGGTCC-3´ nuoE C92A fwd 5´-GGTCGCCATGTGATCAGATATGCTGACAGCGT BclI GGTCTGTC-3´ nuoE C97A fwd 5´-GTGACAGCGTGGTCGCTCATATTAATGGTTATC VspI AGGGTATTCAGGCG-3´ nuoE C133A fwd 5´-CGCTGCTGCCAACTTGTGCACTGGGGAACTGT Alw44I GATAAAGGGC-3´ nuoE C137A fwd 5´-CTTGCTGCCTGGGGAACGCTGATAAGGGCCCA ApaI AACATGATGATCG-3´ nuoE C97S fwd 5´-GTGACAGCGTGGTCTCTCATATTAATGGTTAT VspI CAGGGTATTCAGGCG- 3´ nuoE C133S fwd 5´-CGCTGCTGCCAACTTGCTCCCTTGGGAACTGT Eco130I GATAAAGGGC- 3´ nuoG F35L fwd 5´-GTCTGTCTCTGGGCCTTGATATCCCTTACCTG EcoRV TGCTGGCATC-3´ nuoG F35W fwd 5´-GTCTGTCTCTGGGCCTTGATATCCCTTACTGG EcoRV TGCTGGCATC-3´ nuoG F35Y fwd 5´-GTCTGTCTCTGGGCCTTGATATCCCTTACTA EcoRV TTGCTGGCATC-3´ nuoG W221L fwd 5´-GCACTCCGAGCGTTATAACCGTAAACTGGATA PsiI TGCAGTTTGCGCCGAGC-3´ 29 Material und Methoden Tabelle 3-3 Fortsetzung. nuoG W221F fwd 5´-GCACTCCGAGCGTTATAACCGTAAATTCGAT PsiI ATGCAGTTTGCGCCGAGC-3´ nuoG W221Y fwd 5´-GCACTCCGAGCGTTATAACCGTAAATACGA PsiI TATGCAGTTTGCGCCGAGC-3´ nuoI F101L fwd 5´-CTCACGCTGCATTTTGTGTGGTCTCTGCG Eco31I AAGAAGCCTGTC-3´ nuoI F101W fwd 5´-CTCACGCTGCATTTGGTGTGGTCTCTGCGA Eco31I AGAAGCCTGTC-3´ nuoI F101Y fwd 5´-CTCACGCTGCATTTACTGTGGTCTCTGCGAA Eco31I GAAGCCTGTC-3´ Tabelle 3-4: Verzeichnis der verwendeten DNA-Oligonukleotide zur Amplifikation von DNA Fragmenten. Oligonukleotid Sequenz nuoDfwd 5´-TGGGTGGTGGCGTTTCTGAC-3´ nuoDrev 5´-ATCGCCGCCGGAATTTGCTG-3´ nuoEfwd 5´-CACGAGAATC AACAACCACA AACCG-3´ nuoErev 5´-TTTATACCGC TCCAGCAGTT CAGG-3´ nuoGfwd 5´-GCGCACGACAACGTCTACTTC-3´ revBC2xxA (nuoGrev) 5´-GGGTAATGAAATCATCGCCACGAC -3´ nuoGbegfwd 5´-CGATTAACGCTCAGTCTC-3´ nuoGbegrev 5´-TTACGGTGGGTACGTTTG-3´ nuoIfwd 5´-TCTGGATGATCGGCCTGCAC-3´ nuoIrev 5´-TCGCCCTTATCTTTGCCGTC-3´ 30 Material und Methoden Tabelle 3-5: Verzeichnis der verwendeten DNA-Oligonukleotide zur Amplifikation von Selektionskassetten für die λ-Red vermittelte Rekombination. Die zur Zielsequenz homologen Sequenzen sind unterstrichen. Oligonukleotid Sequenz nuoE::nptI-sacB fwd 5´-GCGTGAAGCGATCGAGCACGAGATGCACCACTACGAAG ACCCGCGTGCGGTACCGGATCCGTCGACCTG-3´ nuoE::nptI-sacB rev 5´-GAGTGTCCTCATCGATCATCATGTTTGGCCCTTTATCACA GTTCCCCAGGGAATTCCCCGGGGGATCCG-3´ nuoD::nptI-sacB fwd 5´-TCACCCGCTGACCACGCCGCCGCCGAAAGAGCGCA CGCTGCACGTTGTGTCTCAAAATCTCTGATG-3´ nuoD::nptI-sacB rev 5´-CTTTGGTCGCCTCAATCATCTGGAAAGATTCATTGG CAGGCATCACCGAATTCCCCGGGGGATCCG-3´ nuoGbeg::nptI-sacB fwd 5´-GAATACGAGGTCAACGGAGCGGACAACCTGCTGGA AGCTTCACGTTGTGTCTCAAAATCTCTGATG-3´ nuoGbeg::nptI-sacB rev 5´-GAAATAAAGGTGCCATCGGAAGCCGGTGTCATACAGGA CATCACCAGAATTCCCCGGGGGATCCG-3´ nuoI::nptI-sacB fwd 5´-CAGAAACCAAAGACGGTCGCTGGTACCCGGAATTTT TCCGCACGTTGTGTCTCAAAATCTCTGATG-3´ nuoI::nptI-sacB rev 5´-GTATTCCCCCATTTCGAAATCCGGGGTTAACTGAAT CGCCGTGGTCGGAATTCCCCGGGGGATCCG-3´ 3.3 Chemikalien Tabelle 3-6 zeigt die in dieser Arbeit verwendeten Chemikalien. Tabelle 3-6: Verzeichnis der Chemikalien. Chemikalie Hersteller Acrylamid Gerbu Agar AppliChem Agarose low EEO AppliChem Ammoniumchlorid (NH4Cl) Grüssing Ammoniumperoxodisuldat (APS) Merck Ammoniumsulfat ((NH4)2SO4) Grüssing Ampicillin (AMP) AppliChem Antifoam 204 Sigma L(+)-Arabinose Roth Bromphenolblau Sigma 31 Material und Methoden Tabelle 3-6, Fortsetzung. BSA (Rinderserumalbumin) Sigma Chloramphenicol (CAM) AppliChem Coomassie Brilliant Blue R250 Merck L-Cystein Sigma D-Desthiobiotin IBA 2,3-Dimethoxy-5-methyl-6-decyl-1,4-benzochinon (Decyl-Ubichinon) Sigma Dimethylsulfoxid (DMSO) Aus Institutdbeständen 1,4-Dithiothreitol (DTT) Gerbu DNAse I AppliChem dNTP-Mix (je 10 mM dATP, dCTP, dGTP, dTTP) Fermentas n-Dodecyl-β-D-maltopyranosid (DDM) AppliChem E. coli polare Lipide (E. coli polar lipid extract) Avanti Eisen-Ammonium-Citrat Merck Essigsäure Aus Institutsbeständen Ethanol Aus Institutsbeständen Ethidiumbromid Fluka Ethylendiamintetraessigsäure, Dinatriumsalz (EDTA) Sigma Glucose Monohydrat Grüssing Glycerin Roth Glycin Riedel-de-Haën Hefeextrakt Hefewerke HH HEPES (N-(2-Hydroxyethyl)-piperazin-N´-(2-ethansulfonsäure)) Sigma Imidazol AppliChem Isopropyl-β-D-thiogalactopyranosid (IPTG) Gerbu Kaliumacetat Roth Kaliumchlorid (KCl) Merck Kaliumcyanid (KCN) Merck Kaliumdihydrogenphosphat-Diydrat (KH2PO4·2H2O) Merck Kaliumhexacyanoferrat(III) (K3 [Fe(CN)]6 ) Fluka Kaliumhydroxid (KOH) Sigma Kaliumiodid (KI) Aus Institutsbeständen Kanamycindisulfat (KAN) Merck Kupfersulfat (CuSO4) Merck Lysozym aus Hühnereiweiß Fluka Magnesiumchlorid-Hexahydrat (MgCl2·6H2O) Merck Magnesiumsulfat-Heptahydrat (MgSO4·7H2O) Merck 32 Material und Methoden Tabelle 3-6, Fortsetzung. D-Mannitol Sigma 2-Mercaptoethanol Roth 2-Methybutan Merck Methylcyclohexan Merck 2-(N-Morpholino)-ethansulfonsäure (MES) AppliChem Natriumchlorid (NaCl) Grüssing Natriumdithionit (Na2S2O4) Merck Natriumdodecylsulfat (SDS) USB Di-Natriumhydrogenposphat (Na2HPO4) Merck Natriumhydroxid (NaOH) Riedel-de-Haën Natrium-Kalium-Tartrat Merck Natriumphosphat Dodecahydrat (Na 3PO4·12 H2O) Merck Natriumsulfat (Na2SO4) Merck Nicotinamidadenin-Dinukleotid, Dinatriumsalz (NADH) Roche Desamino-Nicotinamidadenin-Dinukleotid, Dinatriumsalz (d-NADH) Sigma Pepton aus Casein (pankreatischer Verdau) AppliChem Phenylmethansulfonsäurefluorid (PMSF) Merck Piericidin A Sigma 2-Propanol (Isopropanol) Aus Institutsbeständen Riboflavin Sigma RNase A Fermentas D(+)-Saccharose Roth Salzsäure (HCl) Riedel-de-Haën N,N,N,N„- Tetramethyldiamin (TEMED) Sigma Trichloressigsäure (TCA) Merck Tris-(hydroxymethyl)-aminomethan (TRIS) AppliChem TRIS-Borat Merck N-[Tris(hydroyxmethyl)-methyl]-glycin (Tricine) Merck 33 Material und Methoden 3.4 Medien, Puffer und Lösungen Die Tabellen 3-7 bis 3-12 zeigen die in dieser Arbeit verwendeten Nährmedien, Puffer und Lösungen. Tabelle 3-7: Verzeichnis der Medien für die Zellzucht. Medium Zusammensetzung LB-Medium LB-Platten Pepton 1% (w/v) NaCl 1% (w/v) Hefeextrakt 0.5% (w/v) LB-Medium mit Agar LB*-Medium YP/SUC-Platten Fermentermedium Konzentration 2% (w/v) LB-Medium mit L-Cystein 0.5 mM Eisen-Ammonium-Citrat 10 mg/L Riboflavin 50 mg/L (NH4)2SO4 100 mg/L Pepton 1% (w/v) Hefeextrakt 0.5% (w/v) D(+)-Saccharose 10% (w/v) Agar 2% (w/v) Pepton 1% (w/v) Hefeextrakt 0.5% (w/v) Na3PO4 15 mM Na2HPO4 10 mM KH2PO4 25 mM NH4Cl 50 mM Na2SO4 5 mM MgSO4 2 mM Riboflavin 50 mg/mL Eisen-Ammonium-Citrat 30 mg/mL L-Cystein 0.5 mM 34 Material und Methoden Tabelle 3-7, Fortsetzung. Autoinduktionsmedium SOC-Medium Pepton 1% (w/v) Hefeextrakt 0.5% (w/v) Na2HPO4 25 mM KH2PO4 25 mM NH4Cl 50 mM Na2SO4 5 mM MgSO4 2 mM D-Mannitol 0.5% (w/v) L(+)-Arabinose 0.2% (w/v) Glucose 0.05% (w/v) Pepton 2% (w/v) Hefeextrakt 0.5% (w/v) NaCl 10 mM KCl 2.5 mM MgSO4 10 mM MgCl2 10 mM Glucose 20 mM Tabelle 3-8: Verzeichnis der Antibiotika. Antibiotikum Stammlösung Endkonzentration Ampicillin (AMP) 50 mg/mL in H2O 100 µg/mL Chloramphenicol (CAM) 34 mg/mL in Ethanol 170 µg/mL (flüssige Medien) 20 µg/mL (feste Medien) Kanamycin (KAN) 50 mg/mL in H2O 50 µg/mL 35 Material und Methoden Tabelle 3-9: Verzeichnis der Puffer für die Molekularbiologie. Puffer Zusammensetzung Konzentration P1 TRIS/HCl 50 mM, pH 8.0 EDTA 10 mM RNase A 100 µg/mL NaOH 0.2 M SDS 1% (w/v) P3 Kaliumacetat/Essigsäure 3 M, pH 5.5 TBE TRIS-Borat 89 mM, pH 8.3 EDTA 1 mM P2 Tabelle 3-10: Verzeichnis der Puffer für die Proteinchemie. Puffer Zusammensetzung A-Puffer MES/NaOH 50 mM, pH 6.0 NaCl 50 mM ADDM-Puffer B-Puffer BDDM-Puffer A-Puffer mit DDM 0.1% (w/v) MES/NaOH 50 mM, pH 6.0 NaCl 350 mM B-Puffer mit DDM * A -Puffer MES-Puffer Bindepuffer Elutionspuffer 0.1% (w/v) A-Puffer mit MgCl2 5 mM MES/NaOH 50 mM, pH 6.6 NaCl 50 mM MES/NaOH 50 mM, pH 6.3 NaCl 200 mM Imidazol 20 mM DDM 0.1% (w/v) MES/NaOH 50 mM, pH 6.3 NaCl 200 mM Imidazol 500 mM DDM 0.1% (w/v) 36 Material und Methoden Tabelle 3-11: Verzeichnis der Puffer und Lösungen für die SDS-PAGE nach Laemmli (1970). Puffer Zusammensetzung Konzentration Probenpuffer TRIS/HCl 0.4 M, pH 6.8 SDS 4% (w/v) Glycerin 12% (v/v) Bromphenolblau 1% (w/v) EDTA 2.5 mM DTT 3.5 mM TRIS/HCl 3% (w/v), pH 8.3 Glycin 14% (w/v) SDS 0.5% (w/v) TRIS/Acetat 124 mM, pH 6.8 SDS 0.1% (w/v) TRIS/Acetat 175 mM, pH 8.8 SDS 0.04% (w/v) Anoden- und Kathodenpuffer Sammelgelpuffer Trenngelpuffer Tabelle 3-12: Verzeichnis der Puffer für die SDS-PAGE nach Schägger & von Jagow (1987). Puffer Zusammensetzung Konzentration Probenpuffer TRIS/Acetat 0.4 M, pH 6.8 SDS 8% (w/v) Glycerin 20% (w/v) Coomassie Brilliant Blue R250 0.01% (w/v) Mercaptoethanol 2% (w/v) Anodenpuffer TRIS/HCl 0.2 M, pH 8.9 Kathodenpuffer TRIS/HCl 0.1 M, pH 8.25 Tricine 0.1 M SDS 0.1% (w/v) TRIS/HCl 750 mM, pH 8.45 SDS 0.1% (w/v) TRIS/HCl 1 M, pH 8.45 SDS 0.1% (w/v) Glycerin 10.5% (w/v) Sammelgelpuffer Trenngelpuffer 37 Material und Methoden 3.5 Mikrobiologische Methoden 3.5.1 Bakterienanzucht Anzucht von Vorkulturen: Zur Plasmidpräparation oder als Vorkultur wurde LB-Medium mit den entsprechenden Antibiotika versetzt und mit einer Einzelkolonie oder in einer 1:10´000 (v/v) Verdünnung aus einer Glycerindauerkultur angeimpft und bei 37°C für 14 - 16 h inkubiert. Stämme mit temperatursensitiven Plasmiden wurden bei 30°C kultiviert. Anzucht für die Membranpräparation: Für die Membranpräparation wurden Zellen des Stammes BW25113Δnuo/pBADnuo nuoFHis in 0.4 L Autoinduktionsmedium mit dem entsprechenden Antibiotikum in 1 L Erlenmeyerkolben mit Schikane bei 37°C und 160 Upm kultiviert. Das Wachstum wurde durch Messung der optischen Dichte bei 600 nm (OD600) verfolgt. Am Ende der exponentiellen Wachstumsphase wurden die Zellen durch Zentrifugation bei 10´810·g (8´000 Upm, 10 min, 4°C, Rotor FiberLite F-10, RC-5C, Sorvall Instruments) geerntet, in flüssigem Stickstoff schockgefroren und bei -80°C gelagert. Wurden Zellen der Stämme ANN0221/pBADnuo nuoFHis, AN387Δnuo/pBADnuo nuoFHis oder AN387nuo:nptI/pBADnuo nuoFHis verwendet, wurde diese in LB-Medium kultiviert und episomal kodierte Gene bei einer OD600 von 0.1 – 0.3 mit 0.2% (w/v) L-Arabinose induziert. Anzucht für die Komplex I-Präparation: Um ausreichend Zellen für eine Komplex I-Präparation zu erhalten, wurden Zellen des E. coli Stammes BW25113Δnuo/pBADnuo nuoFHis in einem 10 L Fermenter (Braun Biotech, Melsungen) aerob kultiviert. Hierzu wurden 10 L Autoinduktionsmedium mit 400 mL einer Vorkultur angeimpft und bei 37°C unter Rühren (200 Upm) inkubiert. Die Zellen wurden am Ende der exponentiellen Wachstumsphase durch Zentrifugation bei 10´810·g (8´000 Upm, 10 min, 4°C, Rotor FiberLite F-10, RC-5C, Sorvall Instruments) geerntet, in flüssigem Stickstoff schockgefroren und bei -80°C gelagert. Wurden Zellen der Stämme ANN0221/pBADnuo nuoFHis oder AN387Δnuo/pBADnuo nuoFHis verwendet, wurden diese in Fermentermedium kultiviert und episomal kodierte Gene bei einer OD600 von 0.1 – 0.3 mit 0.2% (w/v) L-Arabinose induziert. 38 Material und Methoden Größere Mengen des E. coli Stamms ANN0221/pBADnuo nuoFHis wurden durch Anzucht im 200 L Maßstab erhalten (Institut für Biologie, Albert-Ludwigs-Universität Freiburg). Hierzu wurde 200 L Fermentermedium mit 200 mL 34 mg/mL Chloramphenicol in Ethanol versetzt, mit 5 L einer Vorkultur angeimpft und bei 37°C unter Rühren (180 Upm) inkubiert. Die Luftzufuhr wurde während der Anzucht linear von 10 auf 180 L/min erhöht. Nach 40 min wurde die Expression des nuo-Operons durch Zusatz von 0.2% (w/v) L-Arabinose induziert. Am Ende der exponentiellen Wachstumsphase wurde die Temperatur auf 6°C abgesenkt und die Zellen in einer Durchlaufzentrifuge bei 17´000·g (20´000 Upm, 1.5 L/min, 4°C, Typ Z41, CEPA, Lahr) geerntet, in flüssigem Stickstoff schockgefroren und bei -80°C gelagert. Anzucht für die Präparation des NADH Dehydrogenase Fragmentes: Der E. coli Stamm BL21(DE3)/pET11anuoB-G nuoFSTII wurde zur Überproduktion des NADH Deyhydrogenase Fragments in einem Fermenter (Institut für Biologie, AlbertLudwigs-Universität Freiburg) aerob kultiviert. 200 L LB*-Medium wurden mit 400 mL 50 mg/mL Ampicillin versetzt, mit 5 L einer Vorkultur angeimpft und bei 37°C unter Rühren (180 Upm) inkubiert. Die Luftzufuhr wurde während der Anzucht linear von 10 auf 180 L/min erhöht. Bei einer OD600 von 0.5 wurde mit 200 mL 0.4 M IPTG die Expression von nuoB-G induziert. Am Ende der exponentiellen Wachstumsphase wurde die Temperatur auf 6°C abgesenkt und die Zellen in einer Durchlaufzentrifuge bei 17´000·g (20´000 Upm, 1.5 L/min, 4°C, Typ Z41, CEPA, Lahr) geerntet, in flüssigem Stickstoff schockgefroren und bei -80°C gelagert. 3.5.2. Herstellung von Glycerindauerkulturen Für die Herstellung von Glycerindauerkulturen wurden 150 μL 80% (v/v) steriles Glycerin vorgelegt, mit 250 μL einer E. coli Vorkultur vermischt und sofort in flüssigem Stickstoff schockgefroren. Die Dauerkulturen wurden bei -80°C gelagert. 3.5.3. Herstellung elektrokompetenter Zellen Zellen für die Elektroporation von Plasmid-DNA : Für die Elektroporation von Plasmid-DNA wurde der zu transformierende Stamm bei 37°C in LB-Medium aerob angezogen. Bei einer OD600 von 0.4 - 0.5 wurde die Kultur für 15 min auf Eis gestellt und anschließend bei 4´500·g zentrifugiert (5´000 Upm, 10 min, 4°C, Rotor A-444, Centrifuge 5804R, Eppendorf). Das Sediment wurde in dem gleichen Volumen wie die Ausgangskultur eiskaltem MilliQ H2O resuspendiert und erneut bei 4´500·g zentrifugiert 39 Material und Methoden (5´000 Upm, 10 min, 4°C, Rotor A-4-44, Centrifuge 5804R, Eppendorf). Dieser Waschschritt wurde zweimal wiederholt. Die Zellen wurden in 10% (v/v) eiskaltem Glycerin entsprechend einer OD600 von 35 - 45 resuspendiert und direkt verwendet oder in flüssigem Stickstoff schockgefroren und bei -80°C gelagert. Zellen für die λ-Red vermittelte Rekombination: Für die λ-Red vermittelte Rekombination wurde eine Vorkultur eines mit dem Plasmid pKD46 transformierten E. coli Stamms angesetzt. 0.5 - 1.0 mL dieser Vorkultur wurde auf 100 mL LB-Medium mit Ampicillin übergeimpft und in einem 300 mL Erlenmeyerkolben mit Schikane bei 30°C und 200 Upm inkubiert. Bei einer OD600 von 0.2 - 0.25 wurde die Expression des Red-Operons durch Zusatz von 0.2% L-Arabinose induziert, die Kultur für weitere 30 min inkubiert und für 15 min auf Eis gestellt. Anschließend wurde bei 4´500·g zentrifugiert (5´000 Upm, 10 min, 2°C, Rotor A-4-44, Centrifuge 5804R, Eppendorf), das Sediment in 50 mL eiskaltem 10% (v/v) Glycerin resuspendiert und nochmals bei 4´500·g zentrifugiert (5´000 Upm, 10 min, 4°C, Rotor A-4-44, Centrifuge 5804R, Eppendorf). Dieser Waschschritt wurde zweimal wiederholt. Die Zellen wurden in 10% (v/v) eiskaltem Glycerin entsprechend einer OD600 von 35 - 45 resuspendiert und direkt verwendet. 3.6. Molekularbiologische Methoden 3.6.1. Plasmidpräparation Analytischer Maßstab: Für die analytische Plasmid-Präparation wurden 2 - 4 mL einer E. coli Übernachtkultur bei 9´300·g zentrifugiert (10´000 Upm, 1 min, 20°C, Rotor F45-24-11, Centrifuge 5415D, Eppendorf) und die Zellen in 250 μL Puffer P1 resuspendiert. Zur Lyse wurde die Suspension mit 250 μL Puffer P2 versetzt und für 4 min bei Raumtemperatur inkubiert. Das Lysat wurde mit 250 μL Puffer P3 neutralisiert, 15 min auf Eis inkubiert und bei 20´800·g zentrifugiert (14´000 Upm, 15 min, 4°C, Rotor F45-30-11, Centrifuge 5804R, Eppendorf). Der Überstand wurde mit 750 μL Isopropanol versetzt und für 20 min bei -20°C inkubiert. Nach Zentrifugation bei 20´800·g (14´000 Upm, 15 min, 4°C, Rotor F45-30-11, Centrifuge 5804R, Eppendorf) wurde der Überstand verworfen, das Sediment bei 37°C getrocknet und in 25 μL 40 Material und Methoden MilliQ H2O durch Schütteln gelöst (50°C, 1200 Upm, 15 min, Thermomixer compact, Eppendorf). Präparativer Maßstab: Für die Präparation von mehr als 2 μg Plasmid-DNA wurde das Wizard® Plus SV Minipreps Kit (Promega) nach Herstellerangaben verwendet. Zur Elution der DNA von der Säule wurde auf 55°C erwärmtes MilliQ H2O verwendet. 3.6.2. Restriktion von DNA Für die Restriktionsanalyse von Plasmiden wurden 200 - 500 ng DNA mit der benötigten Menge an Restriktionsenzym (Fermentas) unter Zusatz des entsprechenden Puffers bei 37°C inkubiert. Das Gesamtvolumen des Ansatzes betrug in der Regel 20 μL. Die eingesetzte Menge an Enzym und die Inkubationsdauer ergaben sich hierbei aus der Definition von 1 Unit, das der Menge an Enzym entspricht, die 1 μg λ-DNA in 1 h vollständig verdaut. Bei einem präparativem Verdau von DNA wurde darauf geachtet, dass die DNA-Konzentration im Ansatz 100 ng/μL nicht überstieg. Der Ansatz wurde über ein präparatives Agarosegel aufgetrennt und mit dem Wizard® SV Gel and PCR Clean-Up (Promega) nach Herstellerangaben aufgereinigt. Bei der Restriktion zur Trennung von Konkatameren wurde der Ansatz nach der Inkubation mit dem Wizard® SV Gel and PCR Clean up Kit (Promega) gereinigt. 3.6.3. Polymerase Kettenreaktion (PCR) Die PCR Ansätze wurden in dünnwandigen Reaktionsgefäßen (0.2 mL dünnwandige PCR Gefäße, Biozym) auf Eis vermischt und nach Erreichen der initialen Denaturierungstemperatur in den Thermocycler (Mastercycler, Eppendorf) gestellt. DNAOligonukleotide „Primer“ einer Länge von mehr als 35 Nukleotiden wurden vom Hersteller HPLC gereinigt bzw. bei einer Länge von mehr als 60 Nukleotiden PAGE gereinigt bestellt (Operon). Es wurde darauf geachtet, dass die Schmelztemperatur der Primer mindestens 60°C betrug (Primer Designer 5, Scientific & Educational Software) und dass die Primer am 3´Ende ein G oder C besaßen. 41 Material und Methoden Präparative PCR : Die Selektionskassetten für die λ-Red vermittelte Rekombination wurden mit einer kommerziellen Mischung aus Taq und Pfu DNA-Polymerase hergestellt (Long PCR Enzyme Mix, Fermentas). Ein 50 μL Ansatz enthielt 5 μL Long PCR Buffer + MgCl2, 10 - 20 ng Templat-DNA, je 200 μM dNTPs, je 500 nM Primer und 2.5 U Long PCR Enzyme Mix. Das PCR-Programm bestand aus einer initialen Denaturierung (95°C, 3 min), einem Zyklus (95°C, 30 s; 58°C, 30 s; 68°C, 1.5 min/kb; 25 Wiederholungen) und einer abschließenden Extension (68°C, 10 min). Der PCR Ansatz wurde mit dem Wizard® SV Gel and PCR Clean up Kit (Promega) nach Herstellerangaben aufgereinigt, wobei mit zusätzlichen 200 μL Membrane Wash Solution gewaschen wurde und die DNA mit 25 μL MilliQ H2O (55°C) von der Säule eluiert wurde. Die DNA-Fragmente mit neu eingefügter Punktmutation wurden für die λ-Red vermittelte Rekombination mittels Phusion-DNA-Polymerase (5 U/μL, Finnzymes) vervielfältigt. Ein 50 μL Ansatz enthielt 10 μL 1× Phusion HF-Puffer + MgCl2 (Finnzymes), 20-40 ng TemplatDNA, je 200 μM dNTPs, je 2 µM Primer und 2,5 U Phusion DNA Polymeras. Das PCRProgramm bestand aus einer initialen Denaturierung (94°C, 3 min), einem Zyklus (94°C, 20 s; 60°C, 30 s; 72°C, 20 s /kb; 25 Wiederholungen) und einer abschließenden Extension (72°C, 5 min). Der PCR-Ansatz wurde mit DpnI verdaut, um die methylierte Vorlage-DNA zu zerstören. Hierfür wurde der Ansatz mit 10 μL Puffer Tango und 1 μL DpnI versetzt und 2 h bei 37°C inkubiert. Anschließend wurde der Ansatz mit dem Wizard® SV Gel and PCR Clean up Kit (Promega) nach Herstellerangaben gereinigt, wobei mit zusätzlichen 200 μL Membrane Wash Solution gewaschen wurde und die DNA mit 25 μL MilliQ H2O (55°C) von der Säule eluiert wurde. Ortsgerichtete Mutagenese: Zur Mutagenese von Vektor-DNA wurde die QuikChange® Methode angewandt. Für die Mutagenese mittels Phusion DNA Polymerase (Finnzymes) enthielt der 50 μL Ansatz 10 μL Phusion HF Puffer (Finnzymes), 20-50 ng Templat-DNA, je 200 μM dNTPs, je 160 nM Primer und 1 U Phusion DNA Polymerase (Finnzymes). Das PCR-Programm bestand aus einer initialen Denaturierung (98°C, 30 s) und einem Zyklus (98°C, 20 s; 55 - 72°C, 10 s; 72°C, 20 s/kb; 18 Wiederholungen). 42 Material und Methoden Für die Mutagenes mittels Pfu-DNA-Polymerase (Promega) wurde ein Ansatz mit 40-45 ng Vorlage-DNA, je 240 nM DNA-Oligonukleotide, 0.2 mM dNTP-Mix, 1×Pfu-Puffer und 3 U Pfu-DNA-Polymerase pipetiert. Das PCR-Programm bestand aus einer initialen Denaturierung (95°C, 120 s) und einem Zyklus (95°C, 60 s; 55 - 58°C, 30 s; 72°C, 120 s/kb; 18 Wiederholungen). Nach der Reaktion wurden 10 μL des Ansatzes zur Kontrolle entnommen und auf ein Agarosegel aufgetragen. Der Ansatz wurde mit dem Wizard® SV Gel and PCR Clean-Up System (Promega) gereinigt und mit 30 μL MilliQ H2O eluiert. Das Eluat wurde mit DpnI verdaut um die methylierte Templat-DNA zu zerstören. Hierfür wurde die DNA-Lösung mit 2.5 μL Puffer Tango und 1 μL DpnI versetzt. Der Ansatz wurde 2 h bei 37°C inkubiert, erneut gereinigt und mit 25 μL MilliQ H2O eluiert. Zur Kontrolle wurden 4 μL dieser Lösung auf ein Agarosegel aufgetragen. Mit 1 - 3 μL der gereinigten Probe wurden elektrokompetente DH5α Zellen transformiert. 3.6.4. Transformation durch Elektroporation 50 μL frisch hergestellte oder auf Eis aufgetaute elektrokompetente Zellen (Kap. 3.5.3.) wurden in einem 1.5 mL Reaktionsgefäß mit 0.5 - 5.0 μL DNA (0.1 - 20 ng) vermischt und in eine eisgekühlte Elektroporationsküvette (d = 1 mm, Eppendorf) überführt. Die Zellen wurden einem Spannungspuls mit einer Amplitude von 1„700 V ausgesetzt (Electroporator 2510, Eppendorf). Sollte die Dauer des Pulses 4.5 ms unterschreiten, wurde der Ansatz verworfen. Andernfalls wurde der Ansatz sofort in 1 mL SOC-Medium aufgenommen, in ein 1.5 mL Reaktionsgefäß überführt und bei 37°C inkubiert (1 h, 750 Upm, Thermomixer compact, Eppendorf). Zellen, die mit einem Vektor mit temperatursensitivem Replikon transformiert wurden, wurden bei 30°C inkubiert (1 h, 750 Upm). Je nach Stamm wurden 100 μL des unverdünnten bzw. 1:10 - 1:10´000 (v/v) in LB-Medium verdünnten Ansatzes auf LBPlatten mit dem entsprechenden Antibiotikum ausgestrichen und für 16 h bei 37°C bzw. 20 24 h bei 30°C inkubiert. 43 Material und Methoden 3.6.5. λ-Red vermittelte Rekombination 50 μL frisch hergestellte elektrokompentente und induzierte Zellen (Kap. 3.5.3.) wurden in einem eisgekühlten 1.5 mL Reaktionsgefäß mit bis zu 500 ng linearer DNA (bei etwa 4 kb) und mit 25 - 50 ng Plasmid-DNA (bei etwa 21 kb) vermischt, für 1 min auf Eis inkubiert und in eine eisgekühlte Elektroporationsküvette (d = 1 mm, Eppendorf) überführt. Das molare Verhältnis von Zellen zu Plasmid-DNA und zu linearer DNA sollte demnach 1:1:100 betragen. Die Zellen wurden einem Spannungspuls mit einer Amplitude von 1´700 V ausgesetzt (Electroporator 2510, Eppendorf). Sollte der Spannungspuls 5.0 ms unterschreiten, wurde der Ansatz verworfen. Andernfalls wurde der Ansatz sofort in 1 mL SOC-Medium aufgenommen, in ein 1.5 mL Reaktionsgefäß überführt und bei 37°C inkubiert (1 h, 750 Upm, Thermomixer compact, Eppendorf). Sollte auf Antibiotika-resistente Klone selektioniert werden, so wurde der Transformationsansatz bei 9´300·g zentrifugiert (10´000 Upm, 1 min, 20°C, Rotor F45-24-11, Centrifuge 5415D, Eppendorf), der Überstand verworfen und das Sediment in 100 μL LB-Medium resuspendiert. Die Zellsuspension wurde auf LB-Platten mit dem entsprechenden Antibiotikum ausgestrichen und für 16 h bei 37°C inkubiert. Sollte hingegen auf Saccharose-resistente Klone selektioniert werden, so wurden 200 μL des Transformationsansatzes auf YP/SUC-Platten mit dem entsprechenden Antibiotikum ausgestrichen und für 20 - 24 h bei 30°C inkubiert. 3.6.6. Agarose Gelelektrophorese Die DNA-Lösung wurde mit 6X Loading Dye Solution (Fermentas) versetzt und auf das Gel aufgetragen. Die DNA-Fragmente wurden in einem Gel aus 0.8% (w/v) Agarose in TBE Puffer bei 6 - 9 V/cm für 30 - 40 min getrennt. Zur Visualisierung der DNA Banden enthielt das Gel 200 ng/mL Ethidiumbromid. Die Fluoreszenz des DNA/Ethidiumbromid Komplexes wurde auf einem Leuchtschirm bei 254 nm angeregt. Über einen Vergleich der Intensität der DNA-Banden mit einem Standard wurde die Größe und Menge an DNA abgeschätzt. Dabei wurden immer 500 ng DNA-Längenstandard (Lambda DNA/Eco130I (StyI) Marker, 16, Fermentas) aufgetragen. 44 Material und Methoden 3.7 Proteinchemische Methoden 3.7.1. Zellaufschluß und Präparation von Membranen Sämtliche Arbeitschritte wurden bei 4°C bzw. auf Eis ausgeführt. Mehr als 5 g noch gefrorener E. coli Zellen wurden im Mixer (Stufe: High, 3 min, Waring) in A-Puffer (2-5 mL/g Bakterien-Feuchtmasse) mit 100 μM PMSF, 10 μg/mL DNase und 100 μg/mL Lysozym zerkleinert und in einem Teflon-in-Glas-Homogenisator resuspendiert. Weniger als 5 g gefrorener E. coli Zellen wurden in 25 mL A-Puffer mit 100 μM PMSF, 10 μg/mL DNAse und 100 μg/mL Lysozym direkt in einem Teflon-in-Glas-Homogenisator resuspendiert. Die Zellen wurden in einer French Press bei 110 MPa (17´000 Psi, French Pressure Cell, SLM Aminco) aufgeschlossen, nicht-aufgeschlossene Zellen und Zelltrümmer durch Zentrifugation bei 30´590·g (10´000 Upm, Rotor A8.24, 20 min, RC-5 Centrifuge, Sorvall) sedimentiert und der Überstand bei 250´000·g zentrifugiert (50´000 Upm, Rotor 60Ti, 1 h, Coulter-Optima LE80K Ultracentrifuge, Beckman). Der Überstand der Zentrifugation wurde als Cytosol und das Sediment als Membranfraktion weiterverwendet. 3.7.2 Zuckergradientenzentrifugation Sämtliche Arbeitschritte wurden bei 4°C bzw. auf Eis ausgeführt. Die Membranen wurden in 1 g/mL A*-Puffer, 50 μM PMSF mittels eines Teflon-in-Glas-Homogenisators resuspendiert. 1.5 mL der Membransuspension wurden mit 263 μL einer DDM-Stammlösung (20% (w/v) in H2O) versetzt, der Ansatz für 15 min auf Eis inkubiert und dabei alle 5 min mit einem Teflonin-Glas-Homogenisator durchmischt. Anschließend wurde der Extrakt bei 150´000·g (30 Psi, 15 min, Rotor A-100, Airfuge, Beckman) zentrifugiert. 0.4 - 1.0 mL des Überstandes wurden auf einen 12 mL Saccharosegradienten (5 - 30% (w/v) Saccharose in A*DDM mit 50 μM PMSF) aufgetragen und bei 160´000·g zentrifugiert (36´000 Upm, 18 h, Rotor SW 41, Coulter-Optima LE-80K Ultracentrifuge, Beckman . Der Gradient wurde in 600 μL Portionen fraktioniert und die NADH/Ferricyanid Oxidoreduktaseaktivität der Fraktionen bestimmt. 45 Material und Methoden 3.7.3. Proteinpräparation Sämtliche Arbeitschritte wurden bei 4°C bzw. auf Eis ausgeführt. Die Puffer für die Chromatographie wurden direkt vor ihrer Verwendung mit 30 μM PMSF versetzt. Isolierung des überproduzierten NADH-Dehydrogenase Fragments : Das Cytosol aus 30-60 g Zellen des Stamms BL21(DE3)/pET11anuoB-G nuoFSTII wurde bei einer Flussgeschwindigkeit von 8 mL/min auf eine in A-Puffer mit 5 mM DTT und 30 μM PMSF äquilibrierte 60 mL Anionenaustausch-Chromatographiesäule (110 x 26 mm, Fractogel EMD TMAE Hicap(M), Merck) aufgetragen und die Säule mit 60 mL des gleichen Puffers gewaschen. Die gebundenen Proteine wurden mit einem linearen 300 mL Gradienten von 50 300 mM NaCl bei einer Flussgeschwindigkeit von 5 mL/min eluiert. Das Eluat wurde in 8 mL Fraktionen gesammelt und die NADH/Ferricyanid Oxidoreduktaseaktivität bestimmt. Fraktionen mit einer Aktivität von mehr als 10 μmol·min-1·mL-1 wurden vereinigt, durch Ultrafiltration (100 kDa MWCO, Amicon Ultra-15, Millipore) auf das halbe Ausgangsvolumen konzentriert und anschließend der pH-Wert mit 0.2 M NaOH auf 6.6 eingestellt. Das Konzentrat wurde mit 0.8 mL/min auf eine 10 mL AffinitätsChromatographiesäule (39 x 18 mm, Strep-Tactin Sepharose, IBA), die in MES-Puffer mit 5 mM DTT und 30 μM PMSF äquilibriert worden war, aufgetragen. Die Säule wurde bei der gleichen Flussgeschwindigkeit mit MES-Puffer gewaschen, bis die Absorbanz des Eluats bei 280 nm auf den Wert des Puffers gesunken war. Das NADH Dehydrogenase Fragment wurde bei einer Flussrate von 1 mL/min mit 2.5 mM D-Desthiobiotin in A-Puffer, 5 mM DTT, 30 μM PMSF eluiert. Das Protein wurde durch Ultrafiltration (100 kDa MWCO, Amicon Ultra15, Millipore) auf eine Konzentration von mehr als 10 mg/mL gebracht (4´000·g, 4°C, Rotor A-4-44, Centrifuge 5804R, Eppendorf), in flüssigem Stickstoff schockgefroren und bei -80°C gelagert. Isolierung des Komplex I: Die Membranen von etwa 25 - 35 g Zellen der Stämme ANN0221/pBADnuo nuoFHis oder BW25113Δnuo/pBADnuo nuoFHis wurden in A*-Puffer (1 mL/g Membranen) mit 50 μM PMSF mittels eines Teflon-in-Glas-Homogenisators resuspendiert. Mit einer DDMStammlösung (20% (w/v) in H2O) wurde eine DDM-Endkonzentration von 3% (w/v) eingestellt, der Ansatz für 15 min auf Eis inkubiert und dabei alle 5 min mit einem Teflon-inGlas-Homogenisator durchmischt. Anschließend wurde der Extrakt mit A*-Puffer auf 64 mL verdünnt und mit 250´000·g zentrifugiert (50´000 Upm, Rotor 60Ti, 15 min, Coulter-Optima 46 Material und Methoden LE-80K Ultracentrifuge, Beckman). Der Überstand wurde mit 6 mL/min auf eine mit ADDMPuffer äquilibrierte 60 mL Anionenaustausch-Chromatographiesäule (110 x 26 mm, Fractogel EMD TMAE Hicap(M), Merck) aufgetragen. Die Säule wurde bei einer Flussgeschwindigkeit von 6 mL/min mit 60 mL ADDM-Puffer und 60 mL 33% BDDM-Puffer (150 mM NaCl) gewaschen. Gebundene Proteine wurden mit einem linearen 75 mL Gradienten von 150 - 350 mM NaCl bei einer Flussgeschwindigkeit von 6 ml/min eluiert. Das Eluat wurde in 4 mL Fraktionen gesammelt und die NADH/Ferricyanid Oxidoreduktaseaktivität bestimmt. Fraktionen mit einer Aktivität von mehr als 10 μmol·min-1·mL-1 wurden vereinigt, mit Elutionspuffer auf eine Imidazol-Endkonzentration von 20 mM eingestellt und mit 1 mL/min auf eine in Bindepuffer äquilibrierte 15 mL Ni2+-IDA Affinitäts-Chromatographiesäule (20 x 48 mm, ProBond, Invitrogen) aufgetragen. Die Säule wurde bei einer Flussgeschwindigkeit von 1 mL/min mit Bindepuffer gewaschen, bis die Absorbanz des Eluats bei 280 nm auf den Wert des Puffers gesunken war. Die gebundenen Proteine wurden bei 1 mL/min mit einem Stufengradienten von 25, 50, 75 und 100% Elutionspuffer (140, 260, 380 und 500 mM Imidazol) eluiert. Fraktionen mit NADH/Ferricyanid Oxidoreduktaseaktivität, die ab einer Imidazolkonzentration von 260 mM eluierten, wurden vereinigt und aufkonzentriert (100 kDa MWCO, Amicon Ultra-15, Millipore, 4´000·g, 4°C, Rotor A-4-44, Centrifuge 5804R, Eppendorf). Das Konzentrat wurde dreimal 1:10 mit ADDM-Puffer verdünnt und wieder aufkonzentriert (100 kDa MWCO, Amicon Ultra-15, Millipore, 4´000·g, 4°C, Rotor A-4-44, Centrifuge 5804R, Eppendorf), um die Imidazolkonzentration auf etwa 1 mM zu erniedrigen. Anschließend wurde das Protein in flüssigem Stickstoff schockgefroren und bei -80°C gelagert. 3.7.4. Größenausschluss-Chromatographie Um die Homogenität der Komplex I-Präparationen zu überprüfen, wurden mindestens 100 μL Komplex I (300 µg in ADDM-Puffer) bei einer Flussrate von 0.5 mL/min auf eine mit ADDMPuffer äquilibrierte 24 mL Größenausschluss-Chromatographiesäule (10 x 300 mm, Superose 6 10/300 GL, GE Healthcare) aufgetragen und eluiert. Um die molekulare Masse der Präparationen zu bestimmen, wurde die Säule mit Thyroglobulin (669 kDa), Ferritin (440 kDa), BSA (67 kDa), and RNase A (13.7 kDa) geeicht. 47 Material und Methoden Für eine präparative Größenausschluss-Chromatographie wurden 250 µL Komplex I (1.5–2.5 mg in ADDM-Puffer) aufgetragen, 0.5 mL Fraktionen gesammelt und von diesen die NADH/Ferricyanid Oxidoreduktase Aktivität bestimmt. Die vereinigten Fraktionen wurden konzentriert (100 kDa MWCO, Amicon Ultra-15, Millipore, 4´000·g, 4°C, Rotor A-4-44, Centrifuge 5804R, Eppendorf), in flüssigem Stickstoff schockgefroren und bei -80°C gelagert. 3.7.5 Aktivitätsbestimmungen Bestimmung der (d-)NADH Oxidase Aktivität: Die NADH und die desamino (d-)NADH Oxidase Aktivität cytoplasmatischer Membranen wurde mit einer Clarke-Sauerstoffelektode (RE K1-1, Oxytec) bei 30°C gemessen. 5 - 10 μL Membransuspension (200 - 600 μg Protein) wurden in 2 mL A*-Puffer vorgelegt und die Reaktion durch Einspritzen von 5 μL 0.5 M (d-)NADH gestartet. Durch Zugabe von 20 μM Piericidin A (10 mM in Ethanol) wurde die Reaktion gehemmt. Für die Berechnung der Aktivität wurde von einer Sauerstoffkonzentration im Puffer von 237 μM (Weiss, 1970) ausgegangen. Zur Eichung der Elektrode wurde das Signal des Sauerstoff-gesättigten Puffers vor und nach Reduktion mit gesättigter Na2S2O4-Lösung aufgezeichnet. Bestimmung der NADH/Ferricyanid Oxidoreduktaseaktivität: Zur Bestimmung der artifiziellen NADH/Ferricyanid Oxidoreduktaseaktivität wurde etwa 20 pmol Enzym zum Testpuffer (50 mM MES/NaOH, 50 mM NaCl, 1 mM K3[Fe(CN)6], 0.2 mM NADH, pH 6.0) gegeben und die Änderung der Absorbanz bei 410 nm, hervorgerufen durch die Abnahme der [Fe(CN)6]3- Konzentration, verfolgt. Zur Berechnung der Aktivität wurde ein molarer Extinktionskoeffizient von ε410nm = 1 mM-1·cm-1 verwendet (Friedrich et al., 1989). Bestimmung der NADH:Decyl-Ubichinon Oxidoreduktaseaktivität: Zur Messung der physiologischen Elektronentransferreaktion von Komplex I wurden 5 μg Enzym (5 - 15 mg/mL in ADDM-Puffer) 1:1 (w/w) mit E. coli polaren Lipiden (10 mg/mL in A-Puffer, 2% (w/v) DDM) vermischt und für 1 min auf Eis inkubiert (Stolpe & Friedrich, 2004). Das in Lipiden rekonstituierte Enzym wurde zu 60 μM Decyl-Ubichinon in 985 μL APuffer (30°C) gegeben und der Ansatz für 1 min unter Rühren inkubiert. Anschließend wurde die Reaktion durch Zugabe von 15 μL 10 mM NADH gestartet und bei Bedarf durch Zugabe von 20 μM Piericidin gehemmt. Die Aktivität wurde aus der zeitlichen Änderung der 48 Material und Methoden Absorbanz bei 340 nm (ε340nm= 6.2 mM-1·cm-1, Hellma QS Küvetten, d = 1 cm; TIDAS II, J&M) berechnet (Friedrich et al., 1994). Zur Bestimmung der Michaelis-Konstante KM und der maximalen Aktivität wurde bei einer konstanten Decyl-Ubichinon Konzentration von 60 µM die NADH-Konzentration im Bereich von 5 bis 200 µM variiert und die Aktivitäten gegen die NADH-Konzentration nach Hanes (1932) aufgetragen. 3.7.6. SDS-Polyacrylamid-Gelelektrophorese (SDS-PAGE) Die Zusammensetzung einer Proteinprobe wurde mittels SDS-PAGE untersucht. Für eine schnelle Auftrennung der Polypeptide (1h bei 30 mA) wurde ein diskontinuierliches Gelsystem mit 4.9%igem Sammelgel und 16%igem Trenngel nach Laemmli (1970) verwendet. Für eine verbesserte Auftrennung der Polypeptide wurde das diskontinuierliche Gelsystem mit 3%igem Sammelgel und 10%igem Trenngel nach Schägger & von Jagow (1987) verwendet. Hierbei ist jedoch die längere Laufzeit von etwa 2.5 h bei 30 mA zu berücksichtigen. Zur Probenvorbereitung wurden 50 - 100 μg Protein mit dem entsprechenden Laufpuffer versetzt, für 30 min bei 30°C denaturiert und auf das Gel aufgetragen. Höhere Temperaturen führen zur Aggregation der hydrophoben Proteine. Die Polypetide wurden bei konstanten 35 mA (EPS 600, Pharmacia Biotech) aufgetrennt, bis die Farbstoffbande die untere Gelkante erreicht hatte. Die Proteinbanden wurden mit heißer Coomassie-Lösung (0.1 % (w/v) Coomassie Brilliant Blue R250, 25% (v/v) Ethanol, 10% (v/v) Essigsäure) gefärbt und durch anschließende Entfärbung des Hintergrunds mit 30% (v/v) Ethanol, 10% (v/v) Essigsäure sichtbar gemacht. 3.7.7. Proteinbestimmung Biuret-Methode: Der Proteingehalt konzentrierter Proben wie z.B. der Membransuspensionen wurde nach der Biuret-Methode (Gornall et al., 1949) bestimmt. Hierzu wurde die Proteinlösung mit 1 mL 6% (w/v) Trichloressigsäure versetzt und bei 20´800·g zentrifugiert (14´000 Upm, 1 min, Rotor 45-30-11, Centrifuge 5804R, Eppendorf). Der Überstand wurde verworfen und das Sediment mit 1 mL Biuret-Reagenz (0.2 M NaOH, 0.5% (w/v) Na-K-Tartrat, 0.3% (w/v) CuSO4, 0.5% (w/v) KI) versetzt und für 30 min geschüttelt. Die Absorbanz wurde bei 546 nm 49 Material und Methoden gemessen. Zur Korrektur gegen Störungen durch Lipide und Detergentien wurde der mit einigen Körnchen KCN entfärbte Ansatz vermessen und die Differenzabsorbanz berechnet. Die Eichgerade wurde mit 0 - 1 mg BSA ermittelt und jede Probe dreifach vermessen. UV-spektroskopische Methode: Es wurde die gegen reinen Puffer korrigierte Absorbanz des verdünnten aufgereinigten Proteins bei 280 nm minus der bei 310 nm gemessen (TIDAS II, J&M). Die Korrektur wurde aufgrund der Lichtstreuung durch das Protein und der Detergenzmizellen eingeführt. Zur Berechnung der Konzentration gemäß des Lambert Beer´schen Gesetzes wurde der molare Absorptionskoeffizient ε280nm und die molare Masse M aus der Aminosäuresequenz abgeleitet (Gill & von Hippel, 1989). Für das NADH Dehydrogenase Fragment ergab sich ein ε280nm = 190 mM-1·cm-1und M = 170 kDa bzw. ε280nm = 781 mM-1·cm-1und M = 535 kDa für den Komplex I. 3.7.8. Microsecond Freeze-hyperquenching (MHQ) Technik Für die Aufnahme von Tieftemperatur ESR-Spektren zur Verfolgung der Reduktion des Komplex I durch NADH im µs Bereich wurden die Proben mit der Microsecond Freezehyperquenching (MHQ) Technik angesetzt. Dazu wurde ein im Labor von Prof. Simon de Vries (Department of Biotechnology, TU Delft, Niederlande) selbst entworfener Versuchsaufbau in Zusammenarbeit mit Herrn Marc Strampraad (Department of Biotechnology, TU Delft) genutzt (Kap. 2.4). Für diese Untersuchungen wurden der Komplex I oder das NADH Dehydrogenase Fragment mehrerer Präparationen jeweils vereinigt, die Proteinkonzentration bestimmt, Portionen mit 1.0 bis 2.8 mg Protein in flüssigem Stickstoff eingefroren und die Proben bis zu ihrer Verwendung bei -80°C gelagert. Zur Vorbereitung der Proben für die MHQ Technik wurden die frisch aufgetauten Proteine bei 20´000·g zentrifugiert (13´000 Upm, 2 min, 4°C, Rotor 45-30-11, Centrifuge 5804R, Eppendorf), um eventuell denaturiertes Protein abzutrennen. 275 µL Komplex I (7-10 mg/mL, 14 -19 µM in ADM-Puffer) oder 275 µL NADH Dehydrogenase Fragment (3.4 mg/mL, 20 µM in A-Puffer) und 600 µL NADH (4 - 200 mM in ADM- bzw. A-Puffer) wurden auf zwei Injektionsschleifen aufgetragen. Sollte der Komplex I in Lipide rekonstituiert werden, wurde das Enzym vor dem Auftrag 1:1 (w/w) mit E. coli polaren Lipiden (20 mg/mL in A-Puffer, 2% (w/v) DDM) vermischt und 1 min auf Eis inkubiert. Die für diese Proben verwendete NADH-Lösung enthielt ebenfalls E. coli polare Lipide in der gleichen Konzentration. Zusätzlich wurde zu den Komplex I Proben für eine weitere Messreihe 60 µM Piericidin A (10 mM in Ethanol) 50 Material und Methoden gegeben, um die Kinetik des Elektronentransfers zu bestimmen. Um eine Probe vollständig zu reduzieren, wurde die Proteinlösung vor dem Auftrag mit 5 µL 1 M NADH versetzt. Um eine vollständig oxidierte Probe zu erhalten, wurden statt NADH 600 µL ADM- bzw. A-Puffer aufgetragen. Durch Hochdruck Injektionsventile wurden die Protein- und die NADHLösungen mit zwei HPLC Pumpen bei einer Geschwindigleit von 1.8 bis 12 mL/min in einen tangentialen Mikromixer in einer Niedrigdruckkammer (< 100 mbar) im Verhältnis 1:1 eingespritzt. Nach Austritt aus dem Mixer trafen die Proben in der Niedrigdruckkammer auf die Wolframoberfläche eines mit flüssigem Stickstoff auf 77 K gekühlten rotierenden Aluminiumzylinders und wurden so eingefroren. Die Flugzeit wurde dabei zwischen 63 µs und 20.1 ms variiert, somit ergaben sich Reaktionszeiten von 97 µs bis 21.1 ms. Die Niedrigdruckkammer wurde mit Stickstoff geflutet und der Zylinder sofort mit flüssigem Stickkstoff gefüllt. Das Pulver wurde in flüssigem Stickstoff mit einem Holzlöffel von der Zylinderoberfläche gesammelt und in speziell präparierten Plastikgefäßen in flüssigem Stickstoff gelagert. Zur Herstellung von ESR-Proben wurde das Pulver in flüssigem Stickstoff mit einem Trichter in ESR-Röhrchen überführt, deren Boden aus einem Polyethylenfilter bestand. 3.7.9 ESR-Spektroskopie Zur Charakterisierung von Fe-S-Zentren des isolierten Komplex I (5 mg/mL) wurden 300 µL mit einigen Körnchen Na2S2O4 und 3 µL 1 M NADH reduziert und luftblasenfrei in ein EPR Röhrchen überführt. Die Proben wurde in einer Kältemischung (2-Methylbutan/ Methylcyclohexan 6:1 (v/v)) bei 150 K eingefroren und in flüssigen Stickstoff gelagert. Die anderen Proben wurden wie oben beschrieben mittels MHQ hergestellt. Die Spektren wurden bei 40 und 13 K mit einem EMX 6/1 ESR-Spektrometer (Bruker) und einem ESR-9 HeliumKryostaten (Oxford Instruments) aufgenommen. Die Mikrowellenfrequenz betrug 9.462 GHz, die Modulationsamplitude 6 G und die Zeitkonstante 164 ms. Die mit der MHQ-Technik angesetzten Proben wurden unter den gleichen Bedingungen vermessen. Wurden die Spektren im Labor von Prof. Simon de Vries aufgenommen, betrug die Mikrowellenfrequenz 9.457 GHz und die Modulationsamplitude 10 G. Weitere Bedingungen der Messungen sind bei den jeweiligen Abbildungen der Spektren vermerkt. 51 Ergebnisse 4. ERGEBNISSE 4.1 Isolierung des Komplex I und des NADH Dehydrogenase Fragmentes 4.1.1. Isolierung des Komplex I Aus 35 g Zellen des E. coli Stammes ANN0221/pBADnuo nuoFHis wurden Membranen präpariert und die darin befindlichen Proteine mit Dodecylmaltosid extrahiert. Mittels Anionenaustausch-Chromatographie an Fractogel EMD Hicap und Affinitätschromatographie an probond Ni2+-IDA wurde der Komplex I gereinigt, Abbildung 4-1 zeigt die zugehörigen Elutionsprofile. Die Tabelle 4-1 gibt eine Übersicht über den Verlauf einer typischen Präparation. Die Banden des SDS-Gel der gereinigten Proteine wurden anhand ihrer apparenten molekularen Massen den Untereinheiten des Komplex I zugeordnet. Es wurden sämtliche Untereinheiten in der Präparation nachgewiesen (Abb. 4-1). Tabelle 4-1: Isolierung des E. coli Komplex I aus 35 g Zellen (Feuchtgewicht) des Stammes ANN0221/pBADnuo nuoFHis. Präparations- Volumen Protein schritt NADH/Ferricyanid Ausbeute Oxidoreduktase Aktivität gesamt spezifisch [mL] [mg] [µmol·min-1] [µmol·min-1·mg-1] [%] Membranen 22 1020 5106 6 100 Detergenzextrakt 29 980 4626 5 91 Fractogel EMD 71 241 2663 11 52 Probond Ni-IDA 14 7 784 121 15 52 Ergebnisse A) NaCl Konzentration [mM] 250 300 250 200 200 150 150 100 100 50 50 0 NADH/Ferricyanid Oxido-1 -1 reduktaseaktivität [µmol min ml ] 300 350 50 100 150 200 app. mol. Masse [kDa] 85 1 2 NuoG NuoCD 50 0 0 C) 250 Volumen [mL] NuoF NuoL NuoM, N 25 B) NuoH NuoB, I NuoE, J Imidazol Konzentration [mM] 250 400 200 300 150 200 100 100 50 0 NADH/Ferricyanid Oxido-1 -1 reduktaseaktivität [µmol min ml ] 300 500 NuoA NuoK 10 0 0 50 100 150 200 Volumen [mL] Abbildung 4-1: Isolierung des Komplex I aus Zellen des E. coli Stammes ANN0221/pBADnuo nuoFHis. Gezeigt sind die Elutionsprofile der Anionenaustausch-Chromatographie an Fractogel EMD Hicap (A) und Affinitäts-Chromatographie an Pro Bond Ni2+ IDA (B) mit der Absorbanz bei 280 nm in schwarz, der NADH/Ferricyanid Oxidoreduktaseaktivität in blau und der Lage des NaCl- bzw. Imidazol-Gradient in rot. C zeigt das SDS-Gel nach Schägger & von Jagow (1987) der Präparation (Spur 2) mit dem Proteinstandard (Spur 1). Die Banden wurden anhand ihrer apparenten molekularen Massen den Komplex I-Untereinheiten zugeordnet. 300 µg des präparierten Proteins wurden auf eine Größenauschluß-Chromatographiesäule aufgetragen, um die Homogenität der Präparation zu überprüfen. Das zugehörige Elutionsprofil zeigt nur einen Gipfel im Massenbereich des Komplex I (Abb. 4-2). Einzelne Präparationen zeigten jedoch einen zusätzlichen Elutionsgipfel im Bereich niedrigeren Molekulargewichtes (Abb. 4-2). Die Fraktionen dieses Gipfels zeigten ebenfalls NADH/Ferricyanid Oxidaseaktivität und nach einer SDS-PAGE hauptsächlich die Banden des NADH Dehydrogenase Fragmentes (Daten nicht gezeigt). Das NADH Dehydrogenase Fragment aus den Untereinheiten NuoE, NuoF und NuoG ist ein Zerfallsprodukt des Komplex I. Es enthält die NADH-Bindestelle und das FMN und zeigt somit ebenfalls NADH/Ferricyanid Oxidoreduktaseaktivität. Da sich der His-tag des Komplex I N-terminal an NuoF befindet, bindet auch das Fragment an die Affinitätssäule. In diesem Fall wurde die Präparation durch eine präparative Größenausschluß-Chromatographie gereinigt und nur die 53 Ergebnisse Fraktionen des Gipfels im höher molekularen Bereich vereinigt. Eine nachfolgende analytische Größenausschluß-Chromatographie der vereinigten und konzentrierten Fraktionen bestätigte die Homogenität des so gereinigten Komplex I (Daten nicht gezeigt). B) 25 20 Absorbanz bei 280 nm [mAU] Absorbanz bei 280 nm [mAu] A) 20 15 10 5 15 10 5 0 0 0 2 4 6 8 10 12 14 16 18 20 Volumen [mL] 0 2 4 6 8 10 12 14 16 18 20 Volumen [mL] Abbildung 4-2: Homogenität der Komplex I Präparation. Gezeigt sind die Elutionsprofile der Größenauschluss-Chromatographie einer homogenen Präparation (A) und einer Präparation, die teilweise in das NADH Dehydrogenase Fragment zerfallen ist (B). Die maximale NADH:Decyl-Ubichinon Oxidoreduktase Aktivität der Präparation wurde zu 2.6 ± 0.1 µmol·min-1·mg-1 bestimmt. Die Reaktion wird zu 90 % durch Piericidin A inhibiert. Die Michaelis-Konstante KM zu NADH lag bei 13 ± 2 µM (Schulte, 2009). Die enzymkinetischen Parameter liegen damit im Bereich der Literaturwerte (Stolpe, 2007; Pohl et al., 2007c). Im Verlaufe der Arbeit wurde der Expressionstamm gewechselt und der Komplex I im Stamm BW25113Δnuo/pBADnuo nuoFHis überproduziert und aus diesem isoliert. Der Verlauf der Präparationen entsprach dem Verlauf der Präparationen des Komplex I aus dem Stamm ANN0221/pBADnuo nuoFHis (Kap. 4.3.6.). 54 Ergebnisse 4.1.2 Isolierung des NADH Dehydrogenase Fragmentes Das NADH Dehydrogenase Fragment des Komplex I aus E. coli setzt sich aus den Untereinheiten NuoE, NuoF und NuoG zusammen, enthält die NADH-Bindestelle, das FMN und die Fe-S-Zentren N1a, N1b, N3, N4, N5 und N7. Es bildet damit den Elektroneneingang des Komplexes. In dem Stamm BL21(DE)/pET11a nuoB-G nuoFSTII lässt sich dieses hydrophile Fragment homolog überproduzieren und mittels Affinitätschromatographie an StrepTactin aus dem Cytosol isolieren (Braun et al., 1998; Bungert et al., 1999). Aufgrund seiner geringen Masse und seiner Löslichkeit ist das Fragment einfacher zu isolieren, im Labor leichter zu handhaben und bietet somit ein gutes System, um das Elektroneneintrittsmodul des Komplex I zu charakterisieren. Aus 35 g Zellen des Stammes BL21(DE3)/pET11a nuoB-G nuoFSTII wurde das Cytosol präpariert und das NADH Dehydrogenase Fragment mittels Anionenaustausch- chromatographie an Fractogel EMD Hicap und Affinitätschromatographie an StrepTactin Sepharose aufgereinigt. Die Tabelle 4-2 gibt eine Übersicht über den Verlauf der Präparation, Abbildung 4-3 zeigt die zugehörigen Elutionsprofile und das SDS-Gel des gereinigten Proteins. Die Proteinbanden des Gels wurden aufgrund ihrer apparenten molekularen Massen den Untereinheiten NuoE, NuoF und NuoG zugeordnet. Die zusätzliche Bande unterhalb von NuoG wird durch ein Abbauprodukt von NuoG hervorgerufen (Bungert et al., 1999). Tabelle 4-2: Isolierung des NADH Dehydrogenase Fragmentes aus 35 g Zellen (Feuchtgewicht) des Stammes BL21(DE3)/pET11a nuoB-G nuoFSTII. Präparations- Volumen Protein schritt NADH/Ferricyanid Ausbeute Oxidoreduktase Aktivität gesamt spezifisch [mL] [mg] [µmol·min-1] [µmol·min-1·mg-1] [%] Cytosol 145 1595 10295 7 100 Fractogel EMD 110 440 5610 13 54 StrepTactin 17 6 984 164 10 55 Ergebnisse A) NaCl Konzentration [mM] 1000 250 800 200 600 150 400 100 200 50 NADH/Ferricyanid Oxido-1 -1 reduktaseaktivität [µmol min ml ] 1200 300 C) app. mol. Masse [kDa] 100.5 NuoG 49.5 NuoF 0 0 0 100 200 300 400 500 Volumen [mL] B) Absorbanz bei 280 nm [mAU] 1000 2000 800 1500 600 1000 400 500 200 0 NADH/Ferricyanid Oxido-1 -1 reduktaseaktivität [µmol min ml ] 1200 2500 18.6 NuoE 0 0 20 40 60 80 100 Volumen [mL] Abbildung 4-3: Isolierung des NADH Dehydrogenase Fragmentes aus Zellen des E. coli Stammes BL21(DE3)/pET11a nuoB-G nuoFSTII. Gezeigt sind die Elutionsprofile der Anionenaustausch- Chromatographie an Fractogel EMD Hicap (A) und Affinitäts-Chromatographie an StrepTactin Sepharose (B) mit der Absorbanz bei 280 nm in schwarz, der NADH/Ferricyanid Oxidoreduktaseaktivität in blau und der Lage des NaCl-Gradients in rot. C) zeigt das SDS-Gel nach Laemmli (1970) des gereinigten Fragments, die Banden wurden anhand ihrer apparenten molekularen Massen NuoE, NuoF und NuoG zugeordnet. 4.2. Die Reduktion der Fe-S-Zentren durch NADH Die katalytische Reaktion des Komplex I ist mit einer Elektronentransferrate von NADH auf Ubichinon mit etwa 100 s-1 zu schnell, um die Elektronenübertragungsprozesse zwischen den Fe-S-Zentren innerhalb des Enzyms durch Standardmessmethoden zu erfassen. Eine weitere Schwierigkeit ist, dass die Reduktion der einzelnen Fe-S-Zentren nur durch ESRSpektroskopie und nur bei Temperaturen von weniger als 40K eindeutig bestimmt werden kann. Aus diesen Gründen liegen bisher kaum Erkenntnisse über die Kinetik des Elektronentransfers zwischen den Fe-S-Zentren vor. In dieser Arbeit sollte die im Labor von Prof. Simon de Vries (Department of Biotechnology, TU Delft, Niederlande) entwickelte 56 Ergebnisse Microsecond Freeze-hyperquenching (MHQ) Technik genutzt werden, die ein schnelles Mischen von Enzym und Substrat in einem Mikromixer und ein augenblickliches Beenden der Reaktion durch Einfrieren der Probe ermöglicht. Zur Bestimmung der Reduktion der Fe-S-Zentren nach einem definierten Zeitpunkt mit der MHQ-Technik werden mindestens 2 mg Komplex I benötigt. Die große Proteinmenge resultiert daraus, dass etwa die Hälfte während des Verfahrens verloren geht (Prof. Simon de Vries, pers. Mitteilung) und nur so ausreichend hohe Proteinkonzentrationen für die ESRSpektroskopie erreicht werden. Aus diesem Grund wurden mehrere Komplex I Präparationen vereinigt, aliquotiert schockgefroren und bis zu ihrer Verwendung bei – 80 °C gelagert. Für eine erste Datenreihe wurden jeweils 275 µL Komplex I (7.4 mg/ml in ADDM, 14 µM) und 600 µL NADH (4 mM in ADDM) in die beiden loops des MHQ-Aufbaus injiziert und insgesamt acht Proben mit Reaktionszeiten zwischen 97 µs und 21.1 ms angesetzt. Zusätzlich wurden ebenfalls mit dem MHQ-System eine oxidierte und eine vollreduzierte Probe eingefroren. Die ESR-Spektren wurden in Zusammenarbeit mit Prof. S. de Vries in Delft aufgenommen. Zum Vergleich wurde ein Aliquot mit NADH reduziert und vermessen, ohne die MHQ-Technik anzuwenden (Abb. 4-4). 57 Ergebnisse A) B) oxidiert oxidiert 97 µs 97 µs 131 µs 131 µs 209 µs 209 µs 395 µs 395 µs 794 µs 794 µs 2.2 ms 6.0 ms 2.2 ms 21 ms 6.0 ms reduziert reduziert N1a N1b N4 N1a N1b N2 N3 N1a 330 335 340 345 Magnetfeld [mT] 350 N2 355 320 330 340 350 N4 N3 360 Magnetfeld [mT] Abbildung 4-4: ESR-Spektren des Komplex I zu verschiedenen Zeitpunkten nach Reduktion mit NADH bei 40 K (A) und 13 K (B). Beim Ansetzen der Proben wurden Komplex I (14 µM) in ADDM-Puffer mit 4 mM NADH gemischt und nach den angegebenen Reaktionszeiten eingefroren. Zum Vergleich sind die Spektren der vollständig reduzierten Proben gezeigt, die mit bzw. ohne Verwendung der MHQ-Technik angesetzt wurden. Die Spektren bei 40 K wurden mit einer Mikrowellenleistung von 5 mW und die Spektren bei 13 K mit einer Leistung von 10 mW aufgenommen. Die Mikrowellenfrequenz betrug 9.457 GHz, die Modulationsamplitude 1 mT, die Zeitkonstante 164 ms und die Aufnahmegeschwindigkeit 17.9 mT/min. Pro Spektrum wurden zwei Scans aufgenommen. Das intensive Signal im Bereich von g = 2.0 ist ein Artefakt, es tritt in allen Spektren der mit der MHQ Technik angesetzten Proben einschließlich der oxidierten Probe auf. Dieses Signal wurde auch in den Spektren anderer mit der MHQ Technik angesetzten Proteinproben beobachtet. Der Ursprung dieses Signals ist nicht bekannt (Prof. S. de Vries, pers. Mitteilung). Dieses Artefakt lässt eine Auswertung der gz-Werte der Signale der Fe-S-Zentren des Komplex I nicht zu. Die Spektren der mit der MHQ Technik angesetzten, voll reduzierten Probe zeigen im g x und gy Bereich jedoch die gleichen Signale wie das Vergleichsspektrum der voll reduzierten Probe. Die gx und gy Werte der Zentren stimmen bei beiden Temperaturen mit den Literaturwerten überein (Leif et al, 1995; Uhlmann & Friedrich 2005). Bei 40 K sieht man das Signal von N1a (gx = 1.92 und gy = 1.96) und N1b (gx, gy = 1.94), bei 13 K zusätzlich die Signale von N2 (gx, gy = 1.91), N3 (gx, gy = 1.88, 1.92) und N4 (gx, gy = 1.89, 1.93). Die bei 40 K aufgenommen Spektren zeigen ab einer Reaktionszeit von 794 µs die Signale der Fe58 Ergebnisse S-Zentren N1a und N1b, die Intensitäten der Signale nehmen mit steigender Reaktionszeit zu und erreichen im Spektrum der Probe nach einer Reaktionszeit von 6 ms die Intensitäten der Signale des voll reduzierten Spektrums. Die bei 13 K aufgenommen Spektren zeigen erst ab dem Zeitpunkt von 2.2 ms die Signale der vierkernigen Fe-S-Zentren. Die Intensitäten der Signale der vollreduzierten Probe wurden nach 6 ms Reaktionszeit erreicht. Zusätzlich wurde eine Messreihe mit dem NADH Dehydrogenase Fragment aufgenommen, dieses hydrophile Fragment enthält die NADH-Bindestelle, das FMN und die Fe-S-Zentren N1a, N1b, N3, N4 und N5. Jeweils 275 µl des präparierten Fragmentes (3.4 mg/mL, 20 µM in A-Puffer) und 600 µL NADH (5 mM in A-Puffer) wurden injiziert und zu insgesamt sieben Zeitpunkten im Bereich zwischen 97 µs und 60.9 ms Proben angesetzt. Außerdem wurden eine voll reduzierte und eine oxidierte Probe mit der MHQ-Technik eingefroren. Die ESRSpektren wurden in Zusammenarbeit mit Prof. S. de Vries in Delft bei 40 K (Abb. 4-5) und 13 K (Daten nicht gezeigt) aufgenommen. oxidiert 97 µs 131 µs 198 µs 404 µs 794 µs 1.2 ms 2.2 ms 6 ms 21 ms 60.9 ms reduziert N1a N1b 300 320 N1a 340 360 380 Magnetfeld [mT] Abbildung 4-5: ESR-Spektren des NADH Dehydrogenase Fragmentes zu verschiedenen Zeitpunkten der Reduktion mit NADH bei 40K. Das NADH Dehydrogenase Fragment (20 µM) in A-Puffer wurde mit 5 mM NADH gemischt und nach den angegebenen Reaktionszeiten eingefroren. Zum Vergleich ist das Spektrum der vollständig reduzierten Probe gezeigt, die unter Verwendung der MHQ-Technik angesetzt wurde. Die Spektren wurden mit einer Mikrowellenleistung von 5 mW aufgenommen. Die Mikrowellenfrequenz betrug 9.44 GHz, die Modulationsamplitude 0.6 mT, die Zeitkonstante 164 ms und die Aufnahmegeschwindigkeit 17.9 mT/min. Pro Spektrum wurden vier Scans aufgenommen. 59 Ergebnisse Auch bei den Proben des NADH Dehydrogenase Fragmentes tritt das Artefakt im Bereich von g = 2.0 auf. Somit ist auch in diesem Fall eine Auswertung der g z-Werte nicht möglich. Das Spektrum der voll reduzierten Probe zeigt die Signale der beiden zweikernigen Zentren N1a und N1b im gx und gy Bereich, die g-Werte stimmen mit den Literaturwerten überein (Braun et al., 1998; Bungert et al, 1999). Die bei 13 K aufgenommenen Spektren des NADH Dehydrogenase Fragments zeigten keine Signale. Die schon geringe Intensität der Signale bei 40 K lassen vermuten, dass die Proteinkonzentration der Proben zu gering war, um die Signale bei 13 K zu detektieren. Im Gegensatz zu den Komplex I Proben zeigen die Spektren des NADH Dehydrogenase Fragmentes schon ab einer Reaktionszeit von 131 µs Signale der Fe-S-Zentren N1a und N1b und die Intensität dieser Signale ist zum Reaktionszeitpunkt von 404 µs schon deutlich erhöht. Auch in dieser Datenreihe nimmt mit steigender Reaktionszeit die Intensität der Signale zu und nach 6 ms wird die Intensität der voll reduzierten Probe erreicht. Aus den relativen Intensitäten des Signals bei g = 1.92 (N1a) und bei g = 1.94 (N1b) lässt sich ablesen, dass die Reduktion des Fe-S-Zentrums N1a früher stattfindet als die des FeS-Zentrums N1b. Die Komplex I Präparation enthält pro Enzym drei Moleküle Ubichinon (M. Kaufenstein, pers. Mitteilung). Es ist also möglich, dass bei der Reaktion die Elektronen auf Ubichinon übertragen werden. Das NADH Dehydrogenase Fragment enthält die Elektronentransportkette nur bis zum Fe-S-Zentrum N5, die übrigen Fe-S-Zentren sowie die Ubichinonbindestelle fehlen. Das Fragment nimmt also Elektronen vom NADH auf, kann diese im Gegensatz zum Komplex I aber nicht auf einen Elektronenakzeptor abgeben. Möglicherweise ist das der Grund für den unterschiedlichen Reduktionsgrad der Fe-S-Zentren im Komplex I und im NADH Dehydrogenase Fragment zu gleichen Reaktionszeitpunkten. Um zu vermeiden, dass die Elektronen im Komplex I über das Ubichinon abgegeben werden und somit ein „steady state“ gemessen wird, sollten für eine weitere Datenreihe die Komplex I Proben mit dem Inhibitor Piericidin A versetzt werden. Piericidin A bindet im Komplex I an der Ubichinonbindestelle und verhindert damit die Bindung und die Reduktion des Ubichinons (Friedrich et al., 1994). Damit wäre gewährleistet, dass die Elektronen des ersten gebundenen NADH detektiert werden. Vor der Aufnahme einer neuen Datenreihe sollte überprüft werden, ob eine Erhöhung der NADH Konzentration Einfluss auf die Intensität der Signale der Fe-S-Zentren hat. Der KMWert zu NADH liegt für den E. coli Komplex I bei 10 µM (Stolpe, 2007; Pohl, 2008; vgl. 60 Ergebnisse Kap. 4.1.1.). Die NADH Konzentration von 4 mM lag bei einem 400-fachen des KM-Wertes. Damit sollte die Reaktion unabhängig von der NADH Konzentration sein. Bei den während der Entstehung dieser Arbeit veröffentlichten Experimenten mit dem E. coli Komplex I in einem ähnlichen Versuchsaufbau wurden bei gleicher Proteinkonzentration 200 mM NADH eingesetzt (Verkhovskaya et al., 2008). Daher wurden das NADH Dehydrogenase Fragment mit 4 mM und 200 mM NADH reduziert und Proben nach Reaktionszeiten von 97 µs und 794 µs eingefroren und vermessen. Dabei war die Intensität der Signale der mit 200 mM NADH reduzierten Probe nach einer Reaktionszeit von 97 µs um das 4-fache gegenüber der Probe mit 4 mM NADH erhöht. Die Probe nach 794 µs zeigte mit 200 mM NADH eine 1.5fach größere Signalintensität (Daten nicht gezeigt). Alle weiteren Proben wurden aus diesem Grund mit 200 mM NADH angesetzt. Um den Einfluß des Piericidin A auf die Kinetik des Komplex I zu messen, wurde je eine Datenreihe von Proben mit und ohne Piericidin A angesetzt. Zusätzlich sollte der Komplex I durch Rekonstitution in E. coli polare Lipide stabilisiert werden. Dazu wurde der Komplex I im Verhältnis 1:1 (w/w) mit E. coli polaren Lipiden (20 mg/mL in A-Puffer, 2 % DDM (w/v)) inkubiert, so dass sich eine Proteinkonzentration von 8.4 mg/mL (16 µM) ergab. Die NADHLösung (Endkonzentration: 200 mM in ADDM) wurde auf die gleiche Lipidkonzentration eingestellt. Vor dem Mischen der Proben in der MHQ-Apparatur wurden der rekonstituierte Komplex I gegebenenfalls mit Piericidin A (Endkonzentration: 60 µM) versetzt und 1 min auf Eis inkubiert. 275 µL des Komplex I Ansatzes und 600 µL der NADH-Lösung wurden injiziert und insgesamt jeweils acht Proben mit und ohne Piericidin A angesetzt. Die Reaktionszeiten wurden von 97 µs bis 21.1 ms variiert. Zusätzlich wurden eine oxidierte und eine vollreduzierte Probe mit der MHQ-Technik eingefroren. Von den Proben wurden ESRSpektren bei 33 K (Abb. 4-6) und bei 11 K (Abb. 4-8) in Zusammenarbeit mit Prof. S. de Vries in Delft aufgenommen. 61 Ergebnisse B) A) oxidiert oxidiert 97 µs 97 µs 131 µs 131 µs 198 µs 198 µs 404 µs 404 µs 794 µs 794 µs 1.2 ms 1.2 ms 2.2 ms 2.2 ms 6.0 ms 6.0 ms 21 ms 1.94 340 350 360 Magnetfeld [mT] 21 ms 1.92 370 1.94 340 350 360 1.92 370 Magnetfeld [mT] Abbildung 4-6: ESR-Spektren des Komplex I in Lipiden zu verschiedenen Zeitpunkten nach der Reduktion mit NADH bei 40 K. A) zeigt die Spektren der Proben mit Piericidin A, B) die Spektren der Proben ohne den Hemmstoff. In E. coli polare Lipide rekonstituierter Komplex I (16 µM) wurde mit 200 mM NADH gemischt und nach den angegebenen Reaktionszeiten sind eingefroren. Die Spektren wurden mit einer Mikrowellenleistung von 5 mW aufgenommen. Die Mikrowellenfrequenz betrug 9.457 GHz, die Modulationsamplitude 1 mT, die Zeitkonstante 164 ms und die Aufnahmegeschwindigkeit 17.9 mT/min. Pro Spektrum wurden drei Scans aufgenommen. Die bei 40 K aufgenommenen ESR-Spektren der Komplex I Proben mit und ohne Piericidin A zeigen die für die Fe-S-Zentren N1a und N1b erwarteten Signale. Die Amplituden der Signale bei gx = 1.92 (N1a) und gx,y = 1.94 (N1b) wurden jeweils gegen die Reaktionszeit aufgetragen und an eine exponentielle Kurve angepasst (Abb. 4-7). Aus dieser Anpassung ergeben sich für die Reduktion der Fe-S-Zentren die in der Tabelle 4-3 zusammengefassten Zeitkonstanten τ, also die Zeitpunkte an denen die Fe-S-Zentren zu 63.2 % reduziert vorliegen. 62 Ergebnisse B) 1,0 1,0 0,8 0,8 Grad der Reduktion Grad der Reduktion A) 0,6 0,4 0,2 0,0 0,6 0,4 0,2 0,0 0 5000 10000 15000 20000 0 5000 10000 Zeit in µs 15000 20000 Zeit in µs Abbildung 4-7: Kinetik der Reduktion der Fe-S-Zentren N1a und N1b. Gezeigt ist der Grad der Reduktion der Fe-S-Zentren abgelesen aus den Amplituden der ESR-Signale zu den jeweiligen Zeitpunkten bei gx = 1.92 (N1a) in rot und bei gx,y = 1.94 (N1b) in schwarz. A) zeigt die Daten der Proben mit Piericidin A und B) zeigt die Proben ohne Hemmstoff. Tabelle 4-3: Zeitkonstanten τ der Reduktion der Fe-S-Zentren N1a und N1b. Die Werte ergeben sich aus der Abbildung 4-7. τ [ms] Komplex I mit Piericidin A N1a (g = 1.92) 3.0 N1b (g = 1.94) 4.3 Komplex I ohne Piericidin A 2.5 4.4 Die Zeitkonstanten τ geben Aufschluss über die Reihenfolge und die Geschwindigkeit der Reduktion der einzelnen Fe-S-Zentren. Sowohl bei der Reaktion mit als auch ohne Piericidin A wird zuerst das Fe-S-Zentrum N1a reduziert. N1a ist nicht der Teil der Elektronentransportkette zwischen dem FMN und der Ubichinonbindestelle, seine Lage im Protein lässt nur eine Elektronenübertragung vom und zum FMN zu. Die ESR-Spektren zeigen, dass N1a während der physiologischen Reaktion reduziert wird. Die Zeitkonstanten der Reduktion des N1b, welches nach dem vierkernigen Fe-S-Zentrum N3 an der zweiten Position der Elektronentransportkette liegt, sind unter beiden Bedingungen gleich. Die Zeitkonstante der Reduktion des N1a ist in Anwesenheit des Inhibitors um 20% erhöht. Ob es sich dabei um eine echte Abweichung handelt, ist nicht eindeutig, da die Bestimmung des Reduktionsgrades der Fe-S-Zentren über die Amplituden der Signale stark fehlerbehaftet ist. 63 Ergebnisse Um dies zu weiter zu klären, ist die Kenntnis der Zeitkonstanten der Reduktion des Fe-SZentrums N2, welches die Elektronen auf das Ubichinon überträgt, notwendig (siehe unten). Die in dieser Arbeit bestimmten Zeitkonstanten der Reduktion der beiden zweikernigen Fe-SZentren des Komplex I sind deutlich höher, als die in einem ähnlichen Experiment bestimmten Zeitkonstanten mit ~ 90 µs für N1a und ~ 1.2 ms für N1b (Verkhovskaya et al., 2008). Nach Angaben der Autoren wurde die verwendete Mischungstechnik dem im Labor von Prof. S. de Vries entwickelten Aufbau nachgebildet. Die Einfriertechnik wurde einem dritten Freeze-quench Aufbau entnommen (Lin et al., 2003). Eine Charakterisierung der Apparatur, also die Berechnung der Mischungszeit, der Flugzeit der Probe und der Einfrierzeit liegt nicht vor. Es lässt sich also nicht beurteilen, ob die unterschiedlichen Zeitkonstanten auf das Experiment an sich oder auf die unterschiedlichen Aufbauten zurückzuführen sind. Das Abweichen der Werte um mehr als 2 ms aufgrund unterschiedlicher Präparationen ist unwahrscheinlich. B) A) oxidiert oxidiert 97 µs 97 µs 131 µs 131 µs 198 µs 198 µs 404 µs 404 µs 794 µs 794 µs 1.2 ms 1.2 ms 2.2 ms 2.2 ms 6.0 ms 6.0 ms 21 ms 21 ms N2 340 345 350 355 N2 N3, N4 360 Magnetfeld [mT] 365 340 345 350 N3, N4 355 360 365 Magnetfeld [mT] Abbildung 4-8: ESR-Spektren des Komplex I in Lipiden zu verschiedenen Zeitpunkten nach Reduktion mit NADH bei 11 K. A) zeigt die Spektren der Proben mit Piericidin A, B) zeigt die Spektren der Proben ohne den Hemmstoff. In E. coli polare Lipide rekonstituierter Komplex I (16 µM) wurde mit 200 mM NADH gemischt und nach den angegebenen Reaktionszeiten eingefroren. Die Spektren wurden mit einer Mikrowellenleistung von 5 mW aufgenommen. Die Mikrowellenfrequenz betrug 9.457 GHz, die Modulationsamplitude 1 mT, die Zeitkonstante 164 ms und die Aufnahmegeschwindigkeit 17.9 mT/min. Pro Spektrum wurden drei Scans aufgenommen. 64 Ergebnisse Eine kinetische Analyse der Reduktion der vierkernigen Fe-S-Zentren N2, N3 und N4 ist aufgrund der Überlagerung der Signale und der fehlenden g z-Werte schwierig. Die bei 11 K aufgenommen Spektren der Proben mit Piericidin A (Abb. 4-8) zeigen ab einem Reaktionszeitpunkt von 404 µs ein Signal, dieses wird dem Fe-S-Zentrum N2 zugeordnet (gx,y = 1.91). Die Spektren der Proben ohne den Hemmstoff zeigen das entsprechende Signal erst ab einem Zeitpunkt von 794 µs. In den Spektren der Proben sowohl mit als auch ohne Hemmstoff treten nach 1.2 ms zusätzliche Signale auf, die vermutlich durch N3 (g x = 1.88, gy = 1.92) und N4 (gx = 1.89 gy = 1.93) hervorgerufen werden. Die späte Reduktion des N2 in den Proben ohne Hemmstoff resultiert möglicherweise aus einer Übertragung der Elektronen von N2 auf Ubichinon. Insgesamt ist aber der Verlauf der Reduktion der vierkernigen Fe-SZentren bei den Proben mit Hemmstoff ähnlich zu dem Verlauf der Proben ohne Hemmstoff. Das lässt vermuten, dass der Unterschied der Zeitkonstanten der Reduktion des N1a der Proben mit und ohne Piericidin A (Tab. 4-3) nicht signifikant ist. Auch die Ergebnisse zum zeitlichen Verlauf der Reduktion des Fe-S-Zentrums N2 unterscheiden sich bezüglich der Zeitskala deutlich von den bisher veröffentlichten Ergebnissen. Die in einem Experiment mit ähnlichem Aufbau bestimmte Zeitkonstante der Reduktion des N2 betrug ~ 90 µs und nach Angaben der Autoren wurde ein geringer Anteil erst nach 1 - 2 ms reduziert. Für die Reduktion des in dieser Veröffentlichung dem Fe-SZentrum N6b zugeordneten Signal wurde eine Zeitkonstante von ~ 1.2 ms bestimmt (Verkhovskaya et al., 2008). Allerdings wurde dieses Fe-S-Zentrum bislang ESRspektroskopisch nicht eindeutig beschrieben (Rasmussen et al., 2001; Belevich et al., 2007; Ohnishi & Nakamura-Ogiso, 2008). Die Reduktion der Fe-S-Zentren N3 und N4 wurde in der Veröffentlichung von Verkhovskaya et al. nicht betrachtet. 4.3 Die Bedeutung des Fe-S-Zentrums N1a 4.3.1 Mutagenese des Bindemotivs des Fe-S-Zentrums N1a Das Fe-S-Zentrum N1a wird im Komplex I aus E. coli von den vier Cysteinen C92, C97, C133 und C137 auf der Untereinheit NuoE koordiniert. Um einen Komplex I ohne N1a oder mit einer veränderten Umgebung des Fe-S-Zentrums zu erhalten, sollten diese Cysteine jeweils gegen Alanine oder Serine ausgetauscht werden. Die Charakterisierung der daraus resultierenden Varianten sollte Aufschluss über die Funktion des Fe-S-Zentrums geben. Es 65 Ergebnisse wird vermutet, dass N1a durch zeitweilige Aufnahme eines Elektrons die Radikalbildung am FMN verhindert und somit die Bildung von für den Organismus schädlichen reaktiven Sauerstoffspezies am Komplex I vermindert. Alternativ könnte das Fe-S-Zentrum N1a ähnlich wie das Fe-S-Zentrum N7 ein evolutionäres Relikt darstellen (Kap. 2.2.1.) und aus strukturellen Gründen weiterhin im Komplex I vorliegen. Das Plasmid pBADnuo nuoFHis, welches das gesamte nuo-Operon enthält, ist aufgrund seiner Größe von mehr als 20 kbp für die Mutagenese nach der QuikChange Methode nicht direkt zugänglich. Aus diesem Grund wurden die Punktmutationen in den Subklon pCA24NnuoE eingefügt, der lediglich das Gen nuoE enthält. Das mutierte Gen wurde dann mittels λ-Red vermittelter Rekombination in das Plasmid pBADnuo nuoFHis eingebracht. Dazu wird der Stamm DH5Δnuo/pkD46 als Wirt benötigt. In diesen Zellen ist das nuo-Operon im Genom deletiert, das Plasmid pkD46 kodiert für das Rekombinasesystem. Als Selektionsmarker wurde die nptI-sacB-Kassette verwendet, die einerseits eine Kanamycin Resistenz vermittelt, andererseits in saccharosehaltigen Medien zum Zelltod führt (Kap. 2.2.2). Die bei der QuikChange Mutagenese mittels Phusion DNA Polymerase eingesetzten Oligonukleotide sind in Tabelle 3-3 aufgeführt. Als Vorlage-DNA wurde das Plasmid pCA24NnuoE genutzt. DH5α Zellen wurden mit dem Ansatz transformiert, einzelne Kolonien auf LB-Platten mit Chloramphenicol kultiviert und die Plasmide isoliert. Der Erfolg der ortsgerichtenen Mutagenese wurde durch Restriktionsverdau und Sequenzierung bestätigt. Mit Hilfe des Restriktionsenzyms BamHI wurde ein, das Gen nuoE enthaltendes, Fragment aus dem Plasmid herausgeschnitten und nach präparativer Gelelektrophorese isoliert. Dieses Fragment diente als Vorlage-DNA für die nachfolgende PCR zur Vervielfältigung des mutierten Gens, das so für die λ-Red vermittelte Rekombination zur Verfügung stand. Die Restriktion mit BamHI war notwendig, da das Plasmid pCA24NnuoE dieselbe Resistenz vermittelt wie das Plasmid pBADnuo nuoFHis und somit bei der anschließenden Rekombination stört. Bei PCR Ansätzen mit pCA24NnuoE als Vorlage-DNA war es nicht gelungen, nur durch DpnI-Verdau den Ansatz vom Plasmid vollständig zu befreien. Für die Rekombination wurde zuerst die Resistenzkassete nptI-sacB mit den Oligonukleotiden nuoE::nptI-sacB fwd und nuoE::nptI-sacB rev und dem Plasmid pVO1100 als Vorlage-DNA vervielfältig. In einem ersten Rekombinationsschritt wurde die Resistenzkassette in pBADnuo nuoFHis eingefügt. Dazu wurden Zellen des Stammes DH5Δnuo/pkD46 mit 20 ng pBADnuo 66 Ergebnisse nuoFHis transformiert und anschließend einzelne Kolonien auf LB-Platten mit Chloramphenicol kultiviert. Aus 2 mL Vorkultur wurde die Plasmid-DNA isoliert. Der Erfolg der Transformation wurde durch Restriktionsverdau bestätigt. Mit derselben Vorkultur wurde LB-Medium angeimpft und elektrokompetente Zellen für die Rekombination hergestellt. Die elektrokompetenten Zellen DH5Δnuo/pkD46 pBADnuo nuoFHis wurden mit 400 ng nptIsacB transformiert, einzelne Kolonien auf LB-Platten mit Kanamycin selektiert und die Plasmid-DNA isoliert. Das Gel nach Restriktion des Ansatzes mit PvuI und CpoI (Abb. 4-9A) zeigte die Banden sowohl des Plasmids pBADnuo nuoFHis nuoE::nptI-sacB und als auch die des Plasmids pBADnuo nuoFHis (Tab. 4-4). Vermutlich bildeten sich während der Rekombination lineare und zirkuläre Multimere aus den Sequenzen beider Plasmide, diese Multimere wurden häufig nach dem Rekombinationsschritt zum Einfügen der Resistenzkassette beobachtet (Pohl, 2008). Um diese zu trennen, wurden 320 ng des Gemisches mit 8 U des Restriktionsenzyms SacI, das in beiden Plasmiden jeweils nur einmal schneidet, verdaut. DH5α Zellen wurden mit dem Ansatz transformiert, einzelne Kolonien kultiviert, davon Stammkulturen angelegt und die Plasmid-DNA isoliert. Nach Restriktionsverdau zeigte das Agarosegel nur die Banden des Plasmids pBADnuo nuoFHis nuoE::nptI-sacB (Abb. 4-9B, Tab. 4-4). Tabelle 4-4: Erwartete Fragmentlängen nach Restriktion der Plasmide pBADnuo nuoFHis nuoE::nptIsacB und pBADnuo nuoFHis mit PvuI und CpoI. pBADnuo nuoFHis nuoE::nptI-sacB pBADnuo nuoFHis [bp] [bp] 12´690 12´690 8´903 5´898 1´078 1´078 844 844 613 365 365 343 343 67 Ergebnisse A) 1 2 [kbp] 19329 6223 1489 B) 1 2 [kbp] 19329 6223 1489 Abbildung 4-9.: Restriktionsanalysen zum Nachweis des Plasmids pBADnuo nuoFHis nuoE::nptI-sacB. Gezeigt ist die Plasmidpräparation nach der Rekombination (A, Spur 1) mit den Banden des Plasmidgemischs (Tab. 4-4) und die Plasmidpräparation nach SacI-Verdau (B, Spur 1) mit den erwarteten Banden des pBADnuo nuoFHis nuoE::nptI-sacB (Tab. 4-4). Spur 2 zeigt jeweils den DNA Längenstandard. Im zweiten Rekombinationsschritt wurden elektrokompetente DH5αΔnuo/pKD46 Zellen mit 50 ng pBADnuo nuoFHis nuoE::nptI-sacB und 400 ng des mutierten Gens nuoE kotransformiert, einzelne Kolonien auf YP/SUC-Platten mit Chloramphenicol kultiviert und die Plasmid-DNA isoliert. Der Erfolg der Rekombination wurde durch Restriktionsverdau und Sequenzierung bestätigt. 4.3.2. Komplex I vermittelte Aktivität der cytoplasmatischen Membranen aus den Mutanten ANN0221/pBADnuo nuoFHis nuoECXA Bei der Arbeit zu Teilen dieses Kapitels wurde Frau Elvira Friedrich im Rahmen ihrer Diplomarbeit (2008) angeleitet. Elektrokompetente Zellen des Stammes ANN0221 wurde mit den in Kapitel 4.3.1 beschriebenen Plasmiden transformiert und in LB-Medium mit Chloramphenicol kultiviert. Die so generierten Überexpressionstämme wurden als ANN0221/pBADnuo nuoFHis nuoEC97A, nuoEC133A und nuoEC137A bezeichnet. Die Mutanten und der Ausgangsstamm ANN0221/pBADnuo nuoFHis wurden in 400 mL LBMedium mit Chloramphenicol angezogen und episomal kodierte Gene bei einer OD 600 von 0.1 durch Induktion des araBAD-Promotors exprimiert (Abb. 4-10). 68 Ergebnisse 5 4 OD600 3 2 1 0 0 50 100 150 200 250 300 Zeit [min] Abbildung 4-10: Wachstumskurven der Mutante ANN0221/pBADnuo nuoFHis nuoEC97A (-■-) und des Ausgangsstammes ANN0221/pBADnuo nuoFHis (-▲-). Bei einer OD600 von 0.1 wurde mit Arabinose induziert. Die Wachstumskurven der anderen Alanin-Mutanten zeigten den gleichen Verlauf. Die Wachstumskurven der Mutanten und des Ausgangsstammes sind sehr ähnlich, es wird jeweils eine OD600 von etwa 4.5 erreicht. Im Genom des Stammes ANN0221 ist im Genom das Gen ndh der alternativen NADH Dehydrogenase inaktiviert, außerdem ist in das Gen nuoB eine Kanamycin Resistenzkassette insertiert (Kap. 2.2.2.). Ohne das entsprechende Plasmid ist dieser Stamm also weder in der Lage die alternative Dehydrogenase noch den Komplex I zu produzieren, was zur Folge hat, dass die Zellen auch nach 300 min eine OD 600 von weniger als 0.3 erreichen (Spehr, 1996). Das Wachstum der Mutanten bis zu einer OD 600 von 4.5 zeigt an, das die episomal kodierten Gene exprimiert wurden und in den Mutanten ein aktiver Komplex I vorliegt. 69 Ergebnisse Tabelle 4-5: Komplex I-vermittelte Aktivitäten der cytoplasmatischen Membranen der Alanin-Mutanten und des Ausgangsstammes ANN0221/pBADnuo nuoFHis. Um den unterschiedlichen Komplex I-Gehalt in der Membran zu berücksichtigen, wurde die NADH Oxidase Aktivität auf die NADH/Ferricyanid Oxidoreduktaseaktivität normiert. PicA steht für Piericidin A. Stamm NADH/Ferricyanid Oxidoredukatseaktivität NADH Oxidase Aktivtät Inhibition durch PicA Normierte NADH Oxidase Aktivität [µmol·min-1·mg-1] [µmol·min-1·mg-1] [%] [µmol·min-1·mg-1] ANN0221/pBADnuo nuoFHis 5.6 0.36 92 0.36 ANN0221/pBADnuo nuoFHis nuoE C97A 3.3 0.27 94 0.46 ANN0221/pBADnuo nuoFHis nuoE C133A 4.8 0.24 86 0.28 ANN0221/pBADnuo nuoFHis nuoE C137A 3.8 0.23 88 0.34 Die Membranen der Mutanten wurden präpariert und deren NADH Oxidase und NADH/Ferricyanid Oxidoreduktase Aktivität bestimmt (Tab. 4-5). Die Membranen jeder Mutante zeigten eine NADH Oxidase Aktivität, die durch Piericidin A fast vollständig gehemmt wurde. Da die vom Komplex katalysierte Reaktion der geschwindigkeitsbestimmende Schritt bei der Elektronenübertragung in der aeroben E. coli Atmungskette ist und da diesen Stämmen die alternative NADH Dehydrogenase fehlt, gibt die NADH Oxidase Aktivität der Membranen Aufschluss über die Aktivität der verschiedenen Komplex I Varianten. Die NADH/Ferricyanid Oxidoreduktaseaktivität ist ein Maß für die Menge an Komplex I in den Membranen. Berechnet man die NADH Oxidase Aktivität unter Berücksichtigung der Menge des Komplex I in der Membran, erhält man die normierte Aktivität, die für einen direkten Vergleich der verschiedenen Stämme geeignet ist. Die normierten Aktivitäten der Membranen aus den Mutanten weichen um 6 bis 25 % von dem Wert des Ausgangstammes ab, diese Abweichung liegt im Fehlerbereich der NADH/Ferricyanid Oxidoredukatse Aktivität. Die Elektronentransferrate des Komplex I aus den rekombinanten Stämmen entspricht also in etwa der des unveränderten Enzyms. 70 Ergebnisse 4.3.3. Isolierung und Charakterisierung des Komplex I aus den Mutanten ANN0221/pBADnuo nuoFHis nuoECXA Aus 25-50 g Zellen der Mutanten wurden die Membranen präpariert und die Proteine durch Dodecylmaltosid aus den Membranen extrahiert. Durch Anionenaustausch-Chromatographie und Affinitätschromatographie wurden die Varianten aufgereinigt. Der Verlauf der Präparationen entsprach dem der Präparation aus dem Ausgangsstamm ANN0221/pBADnuo nuoFHis und die SDS-Gele der einzelnen Präparationen zeigten das gleiche Bandenmuster (Kap. 4.1.1.). Tabelle 4-6 gibt beispielhaft eine Übersicht der Präparation aus dem Stamm ANN0221/pBADnuo nuoFHis nuoE C137A. Tabelle 4-6: Präparation des E. coli Komplex I aus 26 g Zellen (Feuchtgewicht) des Stammes ANN0221/pBADnuo nuoFHis nuoE C137A. Präparations- Volumen Protein schritt NADH/Ferricyanid Ausbeute Oxidoreduktase Aktivität gesamt spezifisch -1 [mL] [mg] [µmol·min ] [µmol·min-1·mg-1] [%] Membranen 11 668 1725 3 100 Detergenzextrakt 30 634 1679 3 97 Fractogel EMD 28 99 851 9 49 Probond Ni-IDA 8 1.4 188 134 11 Um die Präparationen miteinander zu vergleichen, wurden die Gesamtaktivitäten in den Membranen und die Ausbeute an Protein auf eine Ausgangsmenge von 30 g Zellen (Feuchtgewicht) berechnet. Damit ergaben sich für die Mutanten eine Gesamtaktivität in den Membranen von 1600 bis 2000 µmol·min-1 mit einer spezifischen Aktivität von 2.5 bis 3.5 µmol·min-1·mg-1. Für den Ausgangsstamm liegen die Werte im Bereich von 3500-4000 µmol·min-1 mit einer spezifischen Aktivität von 3.0 bis 6.0 µmol·min-1·mg-1 (Kap. 4.1.1.). Die Präparationen lieferten eine Ausbeute von 1.6 bis 2.9 mg Protein mit einer spezifischen Aktivität von 80-100 µmol·min-1·mg-1. Der Ausgangsstamm liefert 4.5 bis 6.0 mg Protein mit einer spezifischen Aktivität von 100 bis 200 µmol·min-1·mg-1. Die Ausbeute der Präparation aus den Mutanten ist also bei ähnlicher Aktivität in etwa halbiert. 71 Ergebnisse Um einen möglichen Einfluss des mutierten Bindemotivs auf die Eigenschaften des Fe-SZentrums N1a nachzuweisen, wurden die mit NADH reduzierten Präparationen der Varianten ESR-spektroskopisch charakterisiert. Abbildung 4-11: ESR-Spektren der Komplex I Präparationen aus den Stämmen ANN0221/pBADnuo nuoFHis nuoE C97A (A), nuoE C133A (B) und nuoE C137A (C) und aus dem Ausgangsstamm (D). Die Präparationen wurden mit NADH reduziert und die Spektren bei 40 K und 2 mW aufgenommen. Die Mikrowellenfrequenz betrug 9.462 GHz, die Modulationsamplitude 0.6 mT, die Zeitkonstante 164 ms und die Aufnahmegeschwindigkeit 17.9 mT/min. Die Abbildung 4-11 zeigt die bei 40 K aufgenommenen Spektren des aus den Mutanten isolierten Komplex I im Vergleich mit dem Spektrum des unveränderten Proteins. Alle Spektren zeigen deutlich die Signale des Fe-S-Zentrums N1a bei den g-Werten gx,y,z = 1.92, 1.94 und 2.00. Auch das Verhältnis des N1a Signals zum Signal des Fe-S-Zentrums N1b (gx.y,z = 1.94, 1.94, 2.03) ist bei allen Spektren gleich. Die unterschiedlichen Intensitäten der einzelnen Spektren sind lediglich auf die unterschiedliche Proteinkonzentration der Proben 72 Ergebnisse zurückzuführen. Auch die bei 13 K aufgenommen Spektren gleichen einander und zeigen die Signale der im E. coli Komplex I nachweisbaren Fe-S-Zentren (Daten nicht gezeigt). Aus diesen Resultaten ergeben zwei Möglichkeiten: Entweder führt der Austausch der einzelnen Cysteine zu Alaninen zu keiner Veränderung des ESR-Signals von N1a oder das Bindemotiv der aus den Mutanten isolierten Proteine ist unverändert und besteht weiterhin aus den vier Cysteinen C92, C97, C133 und C137. Die folgenden Begründungen sollen zeigen, dass vermutlich die zweite Möglichkeit zutreffend ist. Mit dem Stamm ANN0221 und dem Plasmid pBADnuo nuoFHis liegt ein Expressionsystem vor, in dem eine E. coli Komplex I-Variante überproduziert werden kann (Pohl, 2008). Ist die Variante nicht stabil, ist dieses Expressionsystem für die Produktion der Varianten aber vermutlich nicht geeignet. Im Laufe dieser Arbeit erhärtete sich der Verdacht, dass im Stamm ANN0221/pBADnuo nuoFHis eine in trans Komplementation auftritt. Nicht funktionelle episomal kodierte Proteinuntereinheiten werden durch chromosomal kodierte und somit unveränderte, stabile Untereinheiten ersetzt. Dies ist für alle Untereinheiten mit Ausnahme von NuoB möglich. Dieser Verdacht wurde durch das folgende Experiment bestätigt. Mit dem Expressionsystem ANN0221/pBADnuo nuoF::nptI-sacB wurde ein stabiler Komplex I produziert, der aus episomal und chromosomal kodierten Untereinheiten zusammengesetzt sein muss, da im Genom nuoB und auf dem Plasmid nuoF inaktiviert wurde (Pohl, 2008). In diesem Stamm stammt die Untereinheit NuoB von dem episomalen Gen und NuoF von dem chromosomalen Gen. Das bedeutet, dass eine Mutation in nuoE zu einer instabilen Untereinheit NuoE führen kann, die vermutlich bei der Produktion im Stamm ANN0221 durch die genomisch kodierte, unveränderte Untereinheit NuoE ersetzt werden würde. Die ESR-Spektren zeigen in diesem Fall das Signal des N1a von der genomisch kodierten Untereinheit NuoE mit dem nicht veränderten Bindemotiv. Die geringeren Ausbeuten der Komplex I Präparationen aus den Mutanten im Vergleich zu der Präparation aus dem Ausgangsstamm stehen im Einklang mit dieser Annahme. Die Produktion des Komplex I ist abhängig vom Maß der Expression des Gens nuoE unter seinem natürlichen Promotor im Genom und wird nicht durch die Expression über den effizienten araBAD Promoter auf dem Plasmid bestimmt. Die Möglichkeit, dass das mutierte Gen nuoE durch Rekombination mit der genomischen DNA auf dem Plasmid gegen das nicht-mutierte Gen ausgetauscht wurde, wurde 73 Ergebnisse ausgeschlossen. Während der Zellzucht wurden nach der Induktion und vor der Zellernte Zellen entnommen und die Plasmid-DNA isoliert. Durch Restriktionsverdau wurde bestätigt, dass weiterhin das mutiert Gen im Plasmid vorlag (Daten nicht gezeigt). Auch der Vergleich mit der Literatur unterstützt die Vermutung, dass die ESR-Signale einem N1a in nicht veränderter Umgebung zuzuordnen ist. Yano et al. (1994) haben im Bindemotiv des Fe-S-Zentrums N1a des Komplex I in P. denitrificans die Cysteine einzeln gegen Alanine ausgetauscht. Nach Überproduktion und Isolierung der hydrophilen Untereinheit Nqo2 wurden ESR-Spektren aufgenommen. Jeder Austausch eines einzelnen Cysteins führte zu einer geringen Verschiebung der g-Werte, zu einer starken Verbreiterung der Signale und teilweise zu zusätzlichen Signalen im Spektrum (Yano et al., 1994). Allerdings muss berücksichtigt werden, dass es sich dabei um die Produktion einer einzelnen Untereinheit handelt, deren Faltung sich von der im Gesamtkomplex deutlich unterscheiden kann. 4.3.4. Isolierung und Charakterisierung des Komplex I aus den Mutanten ANN0221/pBADnuo nuoFHis nuoE CXS Wie im Kapitel 4.3.3. beschrieben, war es nicht gelungen, durch den Austausch der einzelnen Cysteine des Bindemotivs des Fe-S-Zentrums N1a gegen Alanine stabile Komplex I Varianten zu erhalten, die Untersuchungen über die Funktion von N1a ermöglichten. Aus diesem Grund sollten die Cysteine einzeln gegen Serine ausgetauscht werden. Serin ist durch die Hydroxylgruppe in der Lage, sich an der Koordination von Fe-S-Zentren zu beteiligen. In der Literatur sind Proteinvarianten beschrieben, in denen durch Mutation eingefügte Serine zweikernige Fe-S-Zentren koordinieren. Dieser Austausch führte zu veränderten spektroskopische Eigenschaften der Fe-S-Zentren und hatte eine Verschiebung des Mittenpunktspotentials zur Folge (Fujinaga et al 1993, Werth et al, 1990). Es sollten Komplex I Varianten hergestellt werden, die zwar noch das Fe-S-Zentrum N1a enthalten und somit stabil sind, aber in denen das Mittenpunktspotential von N1a verschoben ist. Das könnte zur Folge haben, dass N1a nicht mehr in der Lage ist, Elektronen vom FMN aufzunehmen und dass diese Varianten Aufschluss darüber geben, ob N1a für die Verminderung der Radikalbildung am FMN verantwortlich ist. 74 Ergebnisse Bei der Arbeit zu Teilen dieses Kapitels wurde Herr Thomas Bayer im Rahmen seiner Diplomarbeit (2008) angeleitet. Elektrokompetente Zellen des Stammes ANN0221 wurde mit den in Kapitel 4.3.1 beschriebenen Plasmiden transformiert und in LB-Medium mit Chloramphenicol kultiviert. Die so hergestellten Überexpressionstämme wurden als ANN0221/pBADnuo nuoFHis nuoE C97S und nuoE C133S bezeichnet. Analog zu den im Kapitel 4.3.2 beschriebenen Präparationen der Alanin-Varianten wurde der Komplex I aus diesen Mutanten isoliert. Die Präparationen zeigten den gleichen Verlauf wie die der AlaninVarianten (Daten nicht gezeigt). Sie wurden mit NADH reduziert und ESR-Spektren bei 40 K und 13 K aufgenommen (Daten nicht gezeigt). Auch diese Spektren zeigen deutlich das Signal des Fe-S-Zentrums N1a bei g x,y,z = 1.92, 1.94 und 2.00. Das Verhältnis des N1a Signals zum Signal des Fe-S-Zentrums N1b (gx,y,z = 1.94, 1.94, 2.03) entspricht dem des unveränderten Komplex I. Auch die bei 13 K aufgenommen Spektren zeigen die Signale der im E. coli Komplex I detektierbaren Fe-S-Zentren in den üblichen Verhältnissen und den unveränderten Positionen. Die unveränderten ESR-Signale des Fe-S-Zentrums N1a weisen darauf hin, dass die aus den Mutanten isolierten Proteine das unveränderte Bindemotiv mit den vier Cysteinen C92, C97, C133 und C137 enthalten und kein Serin an der Koordination beteiligt ist. Der Austausch eines Thiol-Liganden gegen einen Hydroyxl-Liganden an dem zweikernigen Fe-S-Zentrum sollte zur einer deutlichen Veränderung des ESR Signals führen (Fujinaga et al 1993, Werth et al, 1990). Vermutlich führt der Austausch der Cysteine C97 und C133 jeweils gegen Serin zu nicht stabilen Komplex I Varianten und das hat wahrscheinlich eine in trans Komplementation in dem Stamm ANN0221 zur Folge. Es wird also statt einer Variante ein Komplex I mit einer unveränderten, chromosomal kodierten Untereinheit NuoE produziert (Kap. 4.3.3.). Um diese Fragestellung eindeutig beantworten zu können, muss ein Expressionsystem gefunden werden, in dem eine in trans Komplementation ausgeschlossen ist (Kap. 4.3.5 und 4.3.6). 75 Ergebnisse 4.3.5 Eignung des Stammes AN387 Δnuo als Expressionsstamm Um die Problematik der in trans Komplementation im Stamm ANN0221 zu umgehen, sollte die Eignung des Stammes AN387Δnuo (Pohl, 2008) als Expressionsstamm überprüft werden. In diesem Stamm wurde das gesamte nuo-Operon aus dem Genom entfernt, bei Überproduktion des Komplex I mit Hilfe des Plasmids pBADnuo nuoFHis ist eine in trans Komplementation ausgeschlossen. Das Gen für die alternative Dehydrogenase ndh ist jedoch vorhanden. Ein Wachstum der Zellen ist also auch möglich, wenn ein episomal kodierter, nicht funktioneller Komplex I vorliegt. Bei den Arbeiten zu diesem Kapitel wurde Herr Thomas Bayer im Rahmen seiner Diplomarbeit (2008) angeleitet. Zellen des Stammes AN387Δnuo wurden mit dem Plasmid pBADnuo nuoFHis transformiert, einzelne Kolonien in LB-Medium mit Chloramphenicol kultiviert und der Erfolg der Transformation durch Restriktionsanalyse bestätigt. Zellen der Stämme AN387, AN387Δnuo und AN387Δnuo/pBADnuo nuoFHis wurden in 400 mL LB-Medium angezogen, die Expression episomal kodierter Gene durch Zugabe von Arabinose induziert und die Zellen bei Eintritt in die stationäre Phase geerntet. Dabei erreichten die Stämme AN387 und AN387Δnuo/pBADnuo nuoFHis eine OD600 von 5 und der Stamm AN387Δnuo eine OD600 von 2.5. Die Membranen der drei Stämme wurden isoliert und deren NADH/Ferricyanid Oxidoreduktase Aktivität sowie (d-)NADH Oxidase Aktivität gemessen (Tab. 4-7). Zu der NADH Oxidase Aktivität trägt neben dem Komplex I auch die alternative NADH Dehydrogenase bei, während die d-NADH Oxidase Aktivität spezifisch vom Komplex I vermittelt wird (Matsushita et al., 1987, Kap. 2.1). 76 Ergebnisse Tabelle 4-7: (d-)NADH-abhängige Aktivitäten der cytoplasmatischen Membranen der Stämme AN387, AN387Δnuo und AN387Δnuo/pBADnuo nuoFHis. Die d-NADH Oxidase Aktivtät wird größtenteils vom Komplex I verursacht. Das Verhältnis der Oxidase Aktivitäten gibt den Anteil des Komplex I an der NADH Oxidase Aktivität an. Stamm NADH/Ferricyanid Oxidoredukatseaktivität NADH Oxidase Aktivtät d-NADH Oxidase Aktivität d-NADH/NADH Oxidase Aktivität [µmol·min-1·mg-1] [%] AN387 14.1 0.35 0.19 54 AN387Δnuo 14.5 0.33 0.00 0 AN387Δnuo/ pBADnuo nuoFHis 12.4 0.34 0.13 38 Die NADH/Ferricyanid Oxidoreduktase Aktivität und die NADH Oxidase Aktivität der Membranen ist in allen drei Stämmen in etwa gleich. Wie erwartet, ist die Komplex I spezifische d-NADH Oxidase Aktivität im Stamm AN387Δnuo nicht messbar, da in diesem Stamm sämtliche Komplex I Gene fehlen. Die Membranen des Stammes AN387Δnuo/pBADnuo nuoFHis zeigen eine ähnlich hohe d-NADH Oxidase Aktivität wie der Ausgansstamm. Der episomal kodierte Komplex I wird auf dem gleichen Niveau produziert wie der chromosomal kodierte Komplex I im Ausgangsstamm AN387. Um die Stabilität und die Assemblierung des im Stamm AN387Δnuo/pBADnuo nuoFHis produzierten Komplex I zu überprüfen, wurden die Proteine aus den Membranen mit Dodecylmaltosid extrahiert und mittels Zuckergradientenzentrifugation aufgetrennt. Zum Vergleich wurde auch der Membranextrakt aus dem Stamm AN387 aufgetragen. Die NADH/Ferricyanid Oxidoreduktase Aktivität der Fraktionen des Gradienten wurden bestimmt. Abbildung 4-12 zeigt die Aktivitätsprofile. 77 NADH/Ferricyanid Oxido-1 -1 reduktaseaktivität [µmol min ml ] Ergebnisse 10 8 6 4 2 0 2 4 6 8 10 12 14 16 18 Fraktion Abbildung 4-12.: Zuckergradientenzentrifugation der Detergenzextrakte aus Cytoplasmamembranen der Stämme AN387 (-▲-) und AN387Δnuo/pBADnuo nuoFHis (-■-). Die Gradienten wurden mit 30 - 45 mg Protein beladen und die NADH/Ferricyanid Oxidoreduktase Aktivitäten auf 10 mg Proteinauftrag normiert. Das Aktivitätsprofil des Zuckergradienten der Membranproteine aus dem Stamm AN387 zeigt zwei Aktivitätsmaxima. Das Maximum im hinteren Drittel ist dem Komplex I zu zuordnen, die Aktivitäten im ersten Drittel des Gradienten der alternativen NADH Dehydrogenase. Im hinteren Drittel des Gradienten des Membranextraktes aus dem Stamm AN387Δnuo/pBADnuo nuoFHis wurde kaum Aktivität gemessen, es konnte also kein stabiler Komplex I aus den Membranen herausgelöst werden. Dies steht im Einklang mit der erhöhten Aktivität im ersten Drittel des Gradienten, die von der alternativen NADH Dehydrogenase und vermutlich von Bruchstücken des Komplex I stammen. Somit ist der Stamm unter diesen Bedingungen nicht als Überproduzent für den Komplex I geeignet. 4.3.6. Eignung des Stammes BW25113Δnuo als Expressionsstamm Um zu untersuchen, ob der Stamm BW25113Δnuo (Vranas 2009, unveröffentliche Ergebnisse), in dem ebenfalls das gesamte nuo-Operon auf dem Genom deletiert wurde, ein geeignetes Expressionsystem darstellt, wurde er mit dem Plasmid pBADnuo nuoFHis 78 Ergebnisse transformiert. Durch die vollständige Deletion des nuo-Operons auf dem Genom ist eine in trans Komplementation, wie sie im Stamm ANN0221 auftrat (Kap. 4.3.3.) nicht möglich. Bei den Arbeiten zu diesem Kapitel wurden Herr Marius Schulte und Frau Klaudia Morina im Rahmen ihrer Diplomarbeiten (2009) angeleitet. Zellen des Stammes BW25113Δnuo wurden mit dem Plasmid pBADnuo nuoFHis transformiert, einzelne Kolonien in LB-Medium mit Chloramphenicol angezogen und der Erfolg der Transformation durch Restriktionsanalyse bestätigt. Um eine ausreichende Zellmenge für die Isolierung des Komplex I zu erhalten, wurden verschiedene Nährmedien für die Zellzucht ausprobiert. Die höchsten Ausbeuten wurden mit dem Autoinduktionsmedium nach Studier et al. (2005) erreicht (Schulte 2009; Morina 2009). Für die Charakterisierung des Stammes BW25113Δnuo/pBADnuo nuoFHis wurden Zellen in 400 mL Autoinduktionsmedium gezogen und bei Eintritt in die stationäre Phase geerntet. Zum Vergleich wurden auch Zellen der Stämme BW25113 und BW25113Δnuo angezogen. Dabei erreichten die Zellen des Stammes BW25113 eine OD600 von 9 und Zellen des Stammes BW25113Δnuo und BW25113Δnuo/pBADnuo nuoFHis eine OD600 von 6.5. Die Membranen wurden präpariert und deren NADH/Ferricyanid Oxidoreduktase Aktivität sowei (d-)NADH Oxidase Aktivität gemessen (Tab. 4-8). Tabelle 4-8: (d-)NADH-abhängige Aktivitäten der cytoplasmatischen Membranen der Stämme BW25113, BW25113Δnuo und BW25113Δnuo/pBADnuo nuoFHis. Die d-NADH Oxidase Aktivtät wird größtenteils vom Komplex I verursacht. Das Verhältnis der Oxidase Aktivitäten gibt den Anteil des Komplex I an der NADH Oxidase Aktivität an. Stamm NADH/Ferricyanid Oxidoredukatseaktivität NADH Oxidase Aktivtät d-NADH Oxidase Aktivität [µmol·min-1·mg-1] dNADH/NADH Oxidase Aktivität [%] BW25113 8.9 0.50 0.08 16 BW25113Δnuo 18.0 1.55 0.01 1 BW25113Δnuo/ pBADnuo nuoFHis 4.6 0.56 0.34 61 79 Ergebnisse Wie erwartet, zeigen die Membranen des Stammes BW25113Δnuo keine Komplex I gekoppelte Aktivität. Die NADH Oxidase Aktivität und die NADH/Ferricyanid Oxidoreduktaseaktivität dieser Membranen ist aber deutlich höher als in den anderen Stämmen, das Fehlen des Komplex I wird vermutlich durch eine erhöhte Produktion der alternativen NADH Dehydrogenase ausgeglichen. Diese besitzt eine wesentlich höhere Umsatzrate als der Komplex I, was die hohe Aktivität erklärt. Die Membranen aus dem Stamm BW25113Δnuo/pBADnuo nuoFHis zeigen eine vierfach erhöhte d-NADH Oxidase Aktivität im Vergleich zum Ausgangsstamm BW25113. Auch ist der Anteil des Komplex I an der NADH Oxidase Aktivität im Stamm BW25113Δnuo/pBADnuo nuoFHis deutlich höher als im Ausgangsstamm. Das deutet darauf hin, dass der episomal kodierte Komplex I produziert wird. Um die Stabilität und die Assemblierung des im Stamm BW25113Δnuo/ pBADnuo nuoFHis produzierten Komplex I zu überprüfen, wurden die Proteine aus den Membranen die Proteine mit Dodecylmaltosid extrahiert und mittels Zuckergradientenzentrifugation aufgetrennt. Zum Vergleich wurden auch die Membranextrakte der Stämme BW25113 und BW25113Δnuo aufgetragen. Die NADH/Ferricyanid Oxidoreduktase Aktivität der Fraktionen der Gradienten wurden bestimmt. Abbildung 4-13 zeigt die Aktivitätsprofile. NADH/Ferricyanid Oxido-1 -1 reduktaseaktivität [µmol min ml ] 5 4 3 2 1 0 2 4 6 8 10 12 14 16 18 Fraktion Abbildung 4-13.: Zuckergradientenzentrifugation der Detergenzextrakte aus Cytoplasmamembranen der Stämme BW25113 (-▲-), BW25113Δnuo (-■-) und BW25113Δnuo/pBADnuo nuoFHis (-●-). Die Gradienten wurden mit 30 - 45 mg Protein beladen und die NADH/Ferricyanid Oxidoreduktase Aktivitäten auf 10 mg Proteinauftrag normiert. 80 Ergebnisse Das Aktivitätsprofil des Zuckergradienten des Membranextraktes aus dem Stamm BW25113 zeigt das Maximum im Bereich um die Fraktion 14 des Komplex I sowie das Aktivitätsmaximum im Bereich um die Fraktion 5 der alternativen NADH Dehydrogenase. Das durch den Komplex I vermittelte Aktivitätsmaximum im hinteren Drittel des Gradienten fehlt in den Membranen des Stammes BW25113Δnuo. Lediglich das Aktivitätsmaximum in den Fraktionen der alternativen NADH Dehydrogenase ist nachzuweisen. Das Aktivitätsprofil des Zuckergradienten des Stammes BW25113Δnuo/pBADnuo nuoFHis zeigt beide Aktivitätsmaxima. Die hohen Aktivitäten im Bereich um die Fraktion 14 zeigen, dass ein vollständig assemblierter Komplex I überproduziert und stabil aus den Membranen extrahiert wurde. Die erhöhte Aktivität in den Fraktionen 17-19 wird vermutlich durch eine Aggregation des Komplex I hervorgerufen, die durch das eingefügte Affinitätspeptid hervorgerufen wird (Pohl, 2008). Um eine ausreichende Menge Zellen für eine Präparation zu erhalten, wurde der Stamm BW25113Δnuo/pBADnuo nuoFHis in 10 L Autoinduktiosmedium angezogen. Die Ausbeute betrug 85 g Zellen (Feuchtgewicht). Aus 38 g Zellen wurden die Membranen präpariert und die Proteine mit Dodecylmaltosid aus den Membranen herausgelöst. Mittels Anionenaustausch-Chromatographie und Affinitätschromatographie wurde der Komplex I aufgereinigt. Die Elutionsprofile zeigten den gleichen Verlauf wie die der in Kapitel 4.1.1. beschriebenen Präparation des Komplex I aus dem Stamm ANN0221/pBADnuo nuoFHis. Tabelle 4-9 gibt eine Übersicht über die Präparation, die Abbildung 4-14 zeigt das SDS-Gel der gereinigten Proteine. Die Banden des Gels wurden anhand ihrer apparenten molekularen Massen den Untereinheiten des Komplex I zugeordnet. Es wurden sämtliche Untereinheiten in der Präparation nachgewiesen. 81 Ergebnisse Tabelle 4-9: Isolierung des E. coli Komplex I aus 25 g Zellen (Feuchtgewicht) des Stammes BW25113 Δnuo/pBADnuo nuoFHis. Präparations- Volumen Protein NADH/Ferricyanid schritt Ausbeute Oxidoreduktase Aktivität gesamt spezifisch [mL] [mg] [µmol·min-1] [µmol·min-1·mg-1] [%] Membranen 15 1248 2911 2 100 Detergenzextrakt 59 845 2790 3 96 Fractogel EMD 47 247 1857 8 64 Probond Ni-IDA 8 5 490 98 17 app. mol. Masse [kDa] 1 2 NuoG 85 NuoCD 50 NuoF NuoL NuoM, N 25 NuoH NuoB, I NuoE, J NuoA NuoK 10 Abbildung 4-14: SDS-Gel der Präparation des Komplex I aus dem Stamm BW25113Δnuo/pBADnuo nuoFHis. Die Banden wurden auf einem Gel nach Schägger & von Jagow (1987) aufgetrennt und anhand ihrer apparenten molekularen Massen den Komplex I-Untereinheiten zugeordnet. Mittels Größenausschlußchromatographie wurde die Homogenität der Präparation bestätigt (Daten nicht gezeigt, Kap. 4.1.1.). Die maximale NADH:Decyl-Ubichinon Oxidoreduktase Aktivität wurde zu 3.0 ± 0.1 µmol·min-1·mg-1 bestimmt und wird zu 90 % mit Piericidin A inhibiert. Der KM-Wert zu NADH betrug 11 ± 1 µM (Horn, 2010). Die Werte entsprechen den Werten der Präparation des Komplex I aus dem Stamm ANN0221/pBADnuo nuoFHis und denen in der Literatur (Stolpe, 2007; Pohl et al., 2007b). Aus dem Stamm BW25113Δnuo/pBADnuo nuoFHis wurde also ein stabiler und katalytisch aktiver Komplex I 82 Ergebnisse isoliert, somit steht ein Expressionssystem zur Verfügung mit dem Komplex I Varianten überproduziert werden können, da nur episomale nuo-Gene in diesem Stamm vorliegen. 4.3.7. Charakterisierung der Alanin- und Serin-Varianten aus dem Stamm BW25113Δnuo/pBADnuo nuoFHis Elektrokompetente Zellen des Stammes BW25113Δnuo wurden mit den in Kapitel 4.3.1 beschriebenen Plasmiden transformiert und in LB-Medium mit Chloramphenicol angezogen. Die Überexpressionstämme wurden als BW25113Δnuo/pBADnuo nuoFHis nuoEC92A, nuoEC97A, nuoEC133A, nuoEC137A sowie BW25113Δnuo/pBADnuo nuoFHis nuoEC97S und C133S bezeichnet. Die Mutanten und der Ausgangsstamm BW25113Δnuo/pBADnuo nuoFHis wurden in 400 mL Autoinduktionsmedium kultiviert und bei Eintritt in die stationäre Phase geerntet. Der Verlauf der Wachstumskurven der Mutanten und des Ausgangsstammes waren gleich, es wurde jeweils eine OD600 von etwa 5 erreicht. Die Membranen wurden präpariert und deren Komplex I spezifische d-NADH Oxidase Aktivität (Kap. 2.1) an der Sauerstoffelektrode bestimmt. Zudem wurden die dNADH/Ferricyanid Oxidoreduktase Aktivität als Maß für die Menge an Komplex I in den Membranen gemessen (Tab. 4-10). 83 Ergebnisse Tabelle 4-10: d-NADH vermittelte Aktivitäten der cytoplasmatischen Membranen der Alanin- und SerinMutanten und des Ausgangstammes BW25113Δnuo/pBADnuo nuoFHis. Plasmid in BW25113Δnuo d-NADH Oxidase Aktivität d-NADH/Ferricyanid Oxidoredukatseaktivität [µmol·min-1·mg-1] pBADnuo nuoFHis 0.30 1.6 pBADnuo nuoFHis nuoE C92A 0.08 0.5 pBADnuo nuoFHis nuoE C97A 0.15 0.7 pBADnuo nuoFHis nuoE C133A 0.09 0.4 pBADnuo nuoFHis nuoE C137A 0.03 0.4 pBADnuo nuoFHis nuoE C97S 0.10 0.5 pBADnuo nuoFHis nuoE C133S 0.16 0.7 Da die vom Komplex katalysierte Reaktion der geschwindigkeitsbestimmende Schritt bei der Elektronenübertragung in der E. coli Atmungskette ist, gibt die d-NADH Oxidase Aktivität der Membranen Aufschluss über die Aktivität der verschiedenen Komplex I Varianten. Sowohl die d-NADH Oxidase Aktivität als auch die d-NADH/Ferricyanid Oxidoreduktase Aktivität der Membranen sind mit Ausnahme der Mutanten BW25113Δnuo/pBADnuo nuoFHis nuoE C97A und C133S im Vergleich zum Ausgangsstamm jeweils um mehr als zwei Drittel erniedrigt (Tab. 4-10). Das deutet darauf hin, dass in diesen Mutanten nur wenig oder kein funktioneller Komplex I in den Membranen vorliegt. Die Membranen der Mutanten in der das Cystein auf der Position C97 gegen ein Alanin bzw. das Cystein auf der Position C133 gegen ein Serin ausgetauscht wurde, zeigen jeweils halb so große Aktivitäten wie der Ausgangstamm. Vermutlich liegt ein funktioneller Komplex I in geringen Mengen vor. Um die Stabilität und die Assemblierung der NuoE Varianten zu überprüfen, wurden die Proteine aus den Membranen Zuckergradientenzentrifugation mit aufgetrennt. Dodecylmaltosid Die extrahiert NADH/Ferricyanid und mittels Oxidoreduktase Aktivität der Fraktionen des Gradienten wurde bestimmt. Abbildung 4-15 zeigt die zugehörigen Aktivitätsprofile am Beispiel der Mutanten BW25113Δnuo/pBADnuo nuoFHis 84 Ergebnisse nuoE C137A und BW25113Δnuo/pBADnuo nuoFHis nuoE C133S im Vergleich zum Ausgansstamm BW25113Δnuo/pBADnuo nuoFHis. NADH/Ferricyanid Oxido-1 -1 reduktaseaktivität [µmol min ml ] 2,5 2,0 1,5 1,0 0,5 0,0 5 10 15 20 Fraktion Abbildung 4-15.: Zuckergradientenzentrifugation der Detergenzextrakte aus Cytoplasmamembranen der Stämme BW25113Δnuo/pBADnuo nuoFHis nuoE C137A(-▲-), BW25113Δnuo/pBADnuo nuoFHis nuoE C133S (-●-) und zum Vergleich BW25113Δnuo/pBADnuo nuoFHis (-■-). Die Gradienten wurden mit 15 - 25 mg Protein beladen und die NADH/Ferricyanid Oxidoreduktase Aktivitäten auf 10 mg Proteinauftrag normiert. Die Aktivitätsprofile der Zuckergradienten der Membranextrakte aus allen drei Stämmen zeigen das Aktivitätsmaximum im Bereich von Fraktion 5 und 6, das der alternativen Dehydrogenase zu zuordnen ist. Der Membranextrakt aus dem Ausgangsstamm BW25113Δnuo/pBADnuo nuoFHis zeigt zusätzlich ein Maximum um die Fraktion 14, das dem Komplex I zu zuordnen ist. Das Aktivitätsprofil des Zuckergradienten des Membranextraktes aus dem Stamm BW25113Δnuo/pBADnuo nuoFHis nuoE C137A zeigt ein sehr kleines Aktivitätsmaximum im hinteren Drittel des Gradienten (Abb. 4-15), die Profile der Extrakte aus den Stämmen BW25113Δnuo/pBADnuo nuoFHis nuoE C92A bzw. C133A zeigten den gleichen Verlauf (Daten nicht gezeigt). Aus den Membranen dieser drei Stämme konnte kein stabiler Komplex I extrahiert werden. Das Aktivitätsprofil des Extraktes aus der Mutante BW25113Δnuo/pBADnuo nuoFHis nuoE C133S (Abb. 4-15) zeigt den gleichen Verlauf wie die Extrakte aus den Stämmen BW25113Δnuo/pBADnuo nuoFHis nuoE C97A bzw. C97S (Daten nicht gezeigt). Die geringe NADH/Ferricyanid Oxidoreduktase Aktivität in den Fraktionen 12 bis 19 deutet darauf hin, dass die geringen Mengen der Varianten in den Membranen nach der Extraktion nicht mehr stabil waren. Die aus der Zuckergradientenzentrifugation gewonnen Ergebnisse stehen im Einklang mit den 85 Ergebnisse Aktivitätsmessungen der Membranen der Mutanten und bestätigen die Vermutung, dass der Austausch der das Fe-S-Zentrum N1a koordinierende Cysteine zu einem Stabilitätsverlust des Komplex I führt. Diese Ergebnisse zeigen, dass das Fe-S-Zentrum N1a essentiell für die Stabilität des Komplex I aus E. coli ist. Der Austausch der N1a koordinierenden Cysteine gegen Alanine oder Serine behindert entweder die Assemblierung des Komplex I oder führt zu instabilen Komplex I Varianten. 4.4 Die Bedeutung aromatischer Aminosäuren für den Elektronentransport im E. coli Komplex I Im Komplex I bilden die sieben Fe-S-Zentren N3, N1b, N4, N5, N6a, N6b und N2 eine Elektronentransportkette zwischen dem FMN, das die Elektronen vom NADH aufnimmt, und der Ubichinonbindestelle (Sazanov & Hinchliffe, 2006). Die Elektronen werden über eine Distanz von 80 Å übertragen und die experimentell bestimmte Elektronentransferrate für diese Reaktion liegt bei mehr als 50 s-1. Der Übertragungsmechanismus der zu diesen Transferraten führt, wird kontrovers diskutiert. Allgemein akzeptiert ist die Theorie von Moser et al. (2006). Nach dieser Theorie sind die Elektronen ausschließlich auf den Fe-SZentren lokalisiert und tunneln von Zentrum zu Zentrum, das Protein bildet dabei nur eine Tunnelbarriere. Mit ihren Berechnungen unter Einbezug der aus der Kristallstruktur des Komplex I aus T. themophilus bekannten Abständen zwischen den Redoxzentren ergeben sich Transferraten die mit experimentell bestimmten Werten in Einklang zu bringen sind. Eine andere Theorie entwickelten Wittekindt et al. (2009) in Zusammenarbeit mit unserer Arbeitsgruppe. Nach diesen Berechnungen sind die hohen Elektronentransferraten im Komplex I nur mit der so genannten hopping Theorie zu erklären. Dabei sind die Elektronen nicht nur auf den Fe-S-Zentren lokalisiert, sondern zusätzlich auf zwischen den Zentren liegenden konservierten, aromatischen Aminosäuren. Die direkte Beteiligung des лElektronensystems aromatischer Aminosäuren am Elektronentransfer wurde auch für andere Elektronentransportwege in der Literatur beschrieben (Kap. 2.3). Im Komplex I ist insbesondere der Abstand zwischen den Mittelpunkten der Fe-S-Zentren N5 und N6a mit mehr als 14 Å zu groß, als dass nach Berechnungen von Wittekindt et al. eine direkte Elektronenübertragung zwischen diesen Zentren wahrscheinlich ist. Mit Hilfe der Struktur des Komplex I aus T. thermophilus ergaben sich aus den Berechnungen sieben konservierte 86 Ergebnisse Aminosäuren als hopping Kandidaten, vier davon liegen zwischen den Fe-S-Zentren N5 und N6a. Im Komplex I aus E. coli sind sechs dieser konservierten aromatischen Aminosäuren zu finden, die siebte liegt im Komplex I aus T. thermophilus auf der Proteinuntereinheit Nqo15, die nicht konserviert ist (Sazanov & Hinchliffe, 2006; Pohl et al., 2007c; Kap. 2.2). In dieser Arbeit sollten im E. coli Komplex I die konservierten aromatischen Aminosäuren (Tab. 4-11) jeweils gegen andere aromatische Aminosäuren oder gegen Leucin ausgetauscht werden, um ihren Einfluss auf die katalytische Reaktion des Komplex I zu detektieren. Leucin besitzt im Gegensatz zu den aromatischen Aminosäuren kein л-Elektronensystem, dass am Elektronentransport beteiligt sein kann, kommt den aromatischen Aminosäuren aber strukturell am nächsten. Tabelle 4-11: Aromatische Aminosäuren im E. coli Komplex I, die direkt am Elektronentransfer beteiligt sein könnten. Angegeben sind der Transferschritt, die Aminosäuren in der Ein-Buchstaben-Schreibweise und die Proteinuntereinheit, auf der die jeweilige Aminosäure liegt. Das Phenylalanins F397 ist nicht konserviert und an der Position 101NuoI befindet sich in anderen Organismen ein F oder ein Y, alle anderen genannten Aminosäuren sind konserviert. Elektronentransferschritt Aminosäure N3→N1b F397NuoF N1b→N4 F35NuoG N5→N6a W221NuoG H514NuoCD F515NuoCD N6b→N2 F101NuoI 87 Ergebnisse 4.4.1 Mutagenese zum Austausch der aromatischen Aminosäuren Bei der Arbeit zu Teilen dieses Kapitels wurde Frau Klaudia Morina im Rahmen ihrer Diplomarbeit (2009) angeleitet. Das Plasmid pBADnuo nuoFHis, welches das gesamte nuoOperon enthält, ist aufgrund seiner Größe von mehr als 20 kbp für die Mutagenese nach der QuikChange Methode nicht direkt zugänglich. Aus diesem Grund wurden die Punktmutationen zum Austausch der aromatischen Aminosäuren erst in Subklone mit einer Größe von 5 bis 7 kbp, die jeweils nur ein bzw. drei Gene des Komplex enthalten, eingefügt. Von diesen Subklonen wurden die mutierten Gene durch PCR vervielfältigt und mittels λ-Red vermittelter Rekombination in das Plasmid pBADnuo nuoFHis eingebracht. Für die Rekombination wurden Zellen des Stammes DH5Δnuo/pkD46 benutzt. In diesen Zellen ist das nuo-Operon im Genom deletiert, das Plasmid pkD46 kodiert für das Rekombinasesystem. Als Selektionsmarker wurde die nptI-sacB-Kassette verwendet, die einerseits eine Kanamycin Resistenz vermittelt, andererseits in saccharosehaltigen Medien zum Zelltod führt (Kap. 2.2.2). Die bei der QuikChange Methode eingesetzten Oligonukleotide sind der Tabelle 3-3 zu entnehmen, als Vorlage-DNA dienten pUC25 nuoE´-G, pCA24NnuoCD oder pCA24NnuoI. Bei der QuikChange zum Austausch der Aminosäure W221NuoG wurde die Phusion DNA Polymerase eingesetzt, bei allen anderen Ansätzen die Pfu-DNA-Polymerase. DH5α Zellen wurden mit dem jeweiligen Ansatz transformiert, einzelne Kolonien kultiviert und die Plasmide isoliert. Der Erfolg der ortsgerichtenen Mutagenese wurde durch Restriktionsverdau und Sequenzierung bestätigt. Durch eine PCR mit den so erhaltenen Subklonen als VorlageDNA und den in der Tabelle 3-4 aufgeführten Oligonukleotiden wurden die mutierten Gene vervielfältig und standen somit für die Rekombination zur Verfügung. In einem ersten Rekombinationsschritt wurden die zu mutierenden Gene auf dem Plasmid pBADnuo nuoFHis gegen die Resistenz Kassette nptI-sacB ausgetauscht. Dazu wurden durch PCR mit dem Plasmid pVO1100 als Vorlage-DNA und mit den in Tabelle 3-5 aufgeführten Oligonukleotiden die Resistenzkassette vervielfältig und dabei mit homologen Bereichen zu den entsprechenden Genen versehen. Elektrokompetente Zellen des Stammes DH5Δnuo/pkD46 wurden mit 50 ng pBADnuo nuoFHis und 300 ng nptI-sacB transfomiert, einzelne Kolonien auf LB-Platten mit Kanamycin selektiert und die Plasmid-DNA isoliert. Der Erfolg der Rekombination wurde durch Restriktionsanalyse bestätigt. Zeigte die Analyse, 88 Ergebnisse dass ein Gemisch aus dem pBADnuo nuoFHis mit und ohne nptIsacB Kassette vorlag, wurde zur Trennung der Multimere ein SacI-Verdau durchgeführt und DH5α Zellen mit der geschnittenen DNA transformiert (Kap. 4.3.1). So wurden die shuttle-Plasmide pBADnuo nuoFHis nuoCD::nptI-sacB, pBADnuo nuoFHis nuoGbegin::nptI-sacB und pBADnuo nuoFHis nuoI::nptI-sacB dargestellt. Das Plasmid pBADnuo nuoFHis nuoG::nptI-sacB stand bereits zur Verfügung (Pohl, 2008). Im zweiten Rekombinationsschritt wurden elektrokompetenter DH5Δnuo/pkD46 mit 50 ng des jeweiligen shuttle-Plasmids und 300-500 ng der amplifizierten, mutierten Gene transfomiert, einzelne Kolonien auf YP/SUC-Platten mit Choramphenicol selektiert und die Plasmid-DNA isoliert. Der Erfolg der Rekombination wurde durch Restriktionsanalyse und Sequenzierung bestätigt. 4.4.2 Komplex I vermittelte Aktivitäten der cytoplasmatischen Membranen aus den Mutanten mir den ausgetauschten aromatischen Aminosäuren Bei der Arbeit zu Teilen dieses Kapitels wurde Frau Annemarie Horn im Rahmen ihrer Diplomarbeit (2010) angeleitet. Elektrokompetente Zellen des Stammes BW25113Δnuo wurde mit den in Kapitel 4.3.1 beschriebenen Plasmiden transformiert, einzelne Kolonien in LB-Medium mit Chloramphenicol kultiviert und Stammkulturen angelegt. Die Mutanten und der unveränderte Stamm BW25113Δnuo/pBADnuo nuoFHis wurden in 400 mL Autoinduktionsmedium mit Chloramphenicol kultiviert und bei Eintritt in die stationäre Phase geerntet. Die Wachstumskurven der Mutanten und des Ausgangsstammes zeigten den gleichen Verlauf, es wurde eine OD600 von 4.0 bis 6.0 erreicht. Die Membranen der Mutanten wurden präpariert und deren (d-)NADH Oxidase Aktivität und (d-)NADH/Ferricyanid Oxidoreduktase Aktivität, zur Bestimmung der Menge an Komplex I, gemessen. Wird die d-NADH Oxidase Aktivität auf die Menge des Komplex I in den Membranen bezogen, ermöglicht die so berechnete normierte d-NADH Oxidase Aktivität einen direkten Vergleich der Komplex I Aktivitäten in den Mutanten. Die Tabellen 4-12 bis 414 zeigen die Ergebnisse eingeteilt nach den Transferschritten, an denen die jeweilige aromatische Aminosäure beteiligt sein könnte. Zwischen den Fe-S-Zentren N1b und N4 ist nach der hopping Theorie nur eine konservierte, aromatische Aminosäure, das Phenylalanin F35NuoG, ein Kandidat für die Beteiligung am Elektronentransport. Im T. thermophilus Komplex I liegt das Phenylalanins 6.4 Å von N1b 89 Ergebnisse und 7.7 Å von N4 entfernt, während der Abstand zwischen den Kanten der beiden Fe-SZentren 10.7 Å (Abstand zwischen den Mittelpunkten: 13.9 Å) beträgt. Tabelle 4-12: Komplex I-vermittelte Aktivitäten der cytoplasmatischen Membranen der Mutanten F35NuoG. Zum Vergleich wurden Membranen aus dem Ausgangsstamm BW25113Δnuo/pBADnuo nuoFHis vermessen. In die normierte d-NADH Oxidase Aktivität, auf die sich auch die Abweichung vom Ausgansstamm bezieht, ist die d-NADH/Ferricyanid Oxidoreduktaseaktivität, ein Maß für die Menge an Komplex I in den Membranen, miteinbezogen. Plasmid in BW25113Δnuo d-NADH Oxidase Aktivität d-NADH/ Ferricyanid Oxidoredukatseaktivität Normierte d-NADH Oxidase Aktivität [µmol·min-1·mg-1] pBADnuo nuoFHis Vergleich zum Ausgangsstamm [%] 0.25 1.6 0.25 - pBADnuo nuoFHis nuoG F35L 0.27 1.8 0.24 -4 pBADnuo nuoFHis nuoG F35W 0.18 1.8 0.16 -36 pBADnuo nuoFHis nuoG F35Y 0.23 1.2 0.31 +24 Wurde das Phenylalanin gegen ein Leucin ausgetauscht, ist die normierte d-NADH Oxidase Aktivität der Membranen aus Mutante und Ausgangsstamm gleich. Der Austausch des Phenylalanins gegen ein Tyrosin führt zwar zu einer Steigerung der normierten d-NADH Oxidase Aktivität um 24%, diese liegt aber nur knapp oberhalb der Fehlergrenze der in diesen Wert miteinbezogenen d-NADH/Ferricyanid Oxidoreduktase Aktivität. Der Austausch des Phenylalanins gegen ein Tryptophan führt zu einer deutlichen Erniedrigung der normierten dNADH Oxidase Aktivität. Da der Austausch gegen Leucin, welches kein л--System zur Aufnahme eines Elektrons besitzt, keinen Einfluss auf die Aktivität des Komplex I zeigt, wird die verringerte Aktivität der Tryptophan-Variante vermutlich durch strukturelle Änderungen hervorgerufen. Vergleicht man die Sequenzen der Untereinheit NuoG und der homologen Untereinheit Nqo3 aus T. thermophilus, fällt auf, dass in E. coli im Gegensatz zu T. thermophilus ein Tyrosin in direkter Nachbarschaft zum Phenylalanin F35 liegt (Abb. 4-16). Im T. thermophilus Komplex I liegt an dieser Position ein Leucin mit einem Abstand von 4.1 Å zu N1b und 12.5 Å zu N4. 90 Ergebnisse Es besteht also die Möglichkeit, dass in E.coli das Tyrosin an dieser Position am Elektronentransferschritt beteiligt ist und somit der Austausch des F35 gegen ein Leucin zu keiner messbaren Verminderung der d-NADH Oxidase Aktivität führt 25 E. coli T. thermophilus 30 35 40 EACLSLGLDIPYFCWHPALGSVGAC DAVFHAGYDVPLFCSEKHLSPIGAC Abbildung 4-16: Sequenzvergleich von E. coli und T. thermophilus. Das als hopping Kandidat vorgeschlagene Phenylalanin ist fett markiert, das zusätzliche Tyrosin in E. coli ist unterstrichen. Die Nummerierung bezieht sich auf die E. coli Sequenz. Ein weiterer Grund für die unveränderte Aktivität der Leucin-Mutante könnte auch die relativ hohe Übertragungsrate zwischen den beiden Fe-S-Zentren auch ohne Beteiligung einer aromatischen Aminosäure sein. Für die direkte Übertragung N1b → N4 wurde eine Tranferrate von 4.7 s-1 berechnet, für die Übertragung N1b → F35 →N4 liegt sie nach der hopping Theorie bei 6.5 x 105 s-1 (Wittekindt et al., 2009). Die maximale NADH:DecylUbichinon Oxidoreduktaseaktivität des in dieser Arbeit isolierten Komplex I aus E. coli beträgt 3.0 µmol·min-1·mg-1, damit ergibt sich für den Gesamtprozess der Elektronenübertragung von NADH auf Decyl-Ubichinon eine Elektronentransferrate von 54 s-1. Die Transferrate des Übertragungsprozesses liegt ohne Beteiligung der aromatischen Aminosäure in der gleichen Größenordnung, wie die experimentell bestimmte Transferrate der Gesamtreaktion. Es ist möglich, dass die unterschiedlichen Transferraten mit oder ohne Beteiligung des Phenylalanins im Rahmen dieser Aktivitätsmessung nicht erfasst werden können. Der Transferschritt zwischen den Fe-S-Zentren N5 und N6a Der Abstand zwischen den Fe-S-Zentren N5 und N6a beträgt 14.0 Å von Kante zu Kante und 16.9 Å zwischen den Mittelpunkten. Zwischen diesen Zentren liegen im E. coli Komplex I drei konservierte aromatische Aminosäuren, die nach der hopping Theorie am Elektronentransport beteiligt sein können. Das sind das Tryptophan W221 NuoG, das Phenylalanin F515NuoCD und das Histidin H514NuoCD. Den Berechnungen zur Folge kann der Elektronentransport dabei entweder über das W221NuoG und/oder das F515NuoCD erfolgen und/oder nacheinander über das F515NuoCD und das H514NuoCD. Die Tabelle 4-13 zeigt die 91 Ergebnisse Komplex I vermittelten Aktivitäten der Membranen, die aus den Mutanten isoliert wurden, in denen diese Aminosäuren jeweils ausgetauscht wurden. Tabelle 4-13: Komplex I-vermittelte Aktivitäten der cytoplasmatischen Membranen der Mutanten mit Austausch zwischen den Fe-S-Zentren N5 und N6a. Zum Vergleich wurden auch Membranen aus dem Ausgangsstamm BW25113Δnuo pBADnuo nuoFHis vermessen. In die normierte d-NADH Oxidase Aktivität, auf die sich die Abweichung vom Ausgangsstamm bezieht, ist die Menge der Komplex I Varianten in den Membranen über die d-NADH/Ferricyanid Oxidoreduktase Aktivität miteinbezogen. Plasmid in BW25113Δnuo d-NADH Oxidase Aktivität d-NADH/ Ferricyanid Oxidoreduktaseaktivität Normierte d-NADH Oxidase Aktivität [µmol·min-1·mg-1] pBADnuo nuoFHis Vergleich zum Ausgangsstamm [%] 0.25 1.6 0.25 - pBADnuo nuoFHis nuoCD H514L 0.22 1.5 0.23 -8 pBADnuo nuoFHis nuoCD F515L 0.13 1.2 0.17 -32 pBADnuo nuoFHis nuoCD F515W 0.19 1.2 0.25 0 pBADnuo nuoFHis nuoCD F515Y 0.17 1.1 0.25 0 pBADnuo nuoFHis nuoG W221L 0.15 1.4 0.17 -32 pBADnuo nuoFHis nuoG W221F 0.26 1.8 0.23 -8 pBADnuo nuoFHis nuoG W221Y 0.18 1.6 0.18 -28 Wurden die Positionen F515 und W221 gegen ein Leucin ausgetauscht, führte das jeweils zu einer deutlichen Verminderung der normierten d-NADH Oxidase Aktivität. Die Verminderung der Komplex I Aktivität wird also durch den Austausch des F515 bzw. W221 gegen das Leucin hervorgerufen. Das ist ein Hinweis darauf, dass diese Aminosäuren am Elektronentransport beteiligt sind. Der Austausch des F515 gegen eine andere aromatische Aminosäure und der Austausch des W221 durch Tryptophan hat keinen Einfluss auf die normierte d-NADH Oxidase Aktivität. Die Membranen der Mutante in der das W221 gegen 92 Ergebnisse ein Tyrosin ausgetauscht wurde, zeigen eine verminderte normierte d-NADH Oxidase Aktivität. Die Abweichung von -28% liegt an der Fehlergrenze der dNADH/Ferricyanid Oxidoreduktaseaktivität. Die normierte d-NADH Oxidase Aktivität der Membranen aus der Mutante, in denen das Histidin H514 gegen ein Leucin ausgetauscht wurde, ist unverändert im Vergleich zum Ausgangsstamm. Das kann einerseits bedeuten, dass das Histidin nicht am Elektronentransport im Komplex I beteiligt ist. Eine andere Möglichkeit ist, dass das Histidin im unveränderten Enzym am Elektronentransport beteiligt ist, aber dass das Fehlen des Histidins in der Mutante zu keiner messbaren Beeinflussung des Elektronentransfers führt. Die berechneten Elektronentransferraten der möglichen Wege N5 → F515 →H514 → N6a und N5 → F515 → N6a liegen beide in der Größenordnung von 1„000 s -1 (Wittekindt et al., 2009), das heißt auch ohne Beteiligung des H514 werden nach der Theorie ausreichend hohe Übertragungsraten erreicht. Zwischen den Fe-S-Zentren N6b und N2 ist nach der hopping Theorie nur eine konservierte, aromatische Aminosäure, das Phenylalanin F101, ein Kandidat für die Beteiligung am Elektronentransport. Im T. thermophilus Komplex I liegt das Phenylalanins 8.6 Å von N6b und 5.6 Å von N2 entfernt, während der Abstand zwischen den beiden Fe-S-Zentren 10.5 Å von Kante zu Kante (14.2 Å zwischen den Mittelpunkten) beträgt. Die normierte d-NADH Oxidase Aktivität der Membranen ist jedoch in allen Mutanten, in denen dieses Phenlyalanin ausgetauscht wurde, unverändert gegenüber dem Ausgangsstamm (Tab. 4-14). Entweder ist diese Aminosäure nicht an der Elektronenübertragung beteiligt oder die Differenz der Transferraten zwischen der direkten Elektronenübertragung zwischen den beiden Fe-SZentren und der Übertragung mit Beteiligung des F101 ist zu gering, um durch diese Messungen erfasst zu werden. 93 Ergebnisse Tabelle 4-14: Komplex I-vermittelte Aktivitäten der cytoplasmatischen Membranen der Mutanten F101NuoI. Zum Vergleich wurden auch Membranen aus dem Ausgangsstamm BW25113Δnuo/pBADnuo nuoFHis vermessen. In die normierte d-NADH Oxidase Aktivität, auf die sich die Abweichung vom Ausgangsstamm bezieht, ist die Menge der Komplex I Varianten in den Membranen über die dNADH/Ferricyanid Oxidoreduktase Aktivität miteinbezogen. Plasmid in BW25113Δnuo d-NADH Oxidase Aktivität d-NADH/ Ferricyanid Oxidoredukatseaktivität Normierte d-NADH Oxidase Aktivität Vergleich zum Ausgangsstamm [µmol·min-1·mg-1] [%] - pBADnuo nuoFHis 0.25 1.6 0.25 pBADnuo nuoFHis nuoI F101L 0.22 1.1 0.32 +28 pBADnuo nuoFHis nuoI F101W 0.23 1.5 0.25 0 pBADnuo nuoFHis nuoI F101Y 0.21 1.3 0.26 +4 Für den Übertragungsschritt N6b → N2 ohne Beteiligung einer aromatischen Aminosäure wurde eine Tranferrate von 100 s-1 berechnet, für die Übertragung N6b → F101 →N2 liegt sie nach der hopping Theorie bei 3.6 x 105 s-1. Die experimentell bestimmte Elektronentransferrate des Gesamtprozesses des in dieser Arbeit isolierten Komplex I aus E. coli beträgt 54 s-1. Die Transferrate des Übertragungsprozesses ohne Beteiligung der aromatischen Aminosäure liegt somit in der gleichen Größenordnung wie die experimentell bestimmte Transferrate. 4.4.3 Stabilität der Komplex I Varianten W221YNuoG, W221Y NuoG und F515NuoCD Der Austausch der Aminosäuren W221NuoG und F515NuoCD jeweils gegen Leucin führte zu einer deutlichen Verminderung der Komplex I vermittelten Aktivtäten in den Membranen. Um zu Überprüfen, ob diese Aktivitätsabnahme somit durch eine Instabilität der Komplex I Varianten hervorgerufen wurde, wurden diese Varianten isoliert und charakterisiert. 94 Ergebnisse Aus jeweils 40 g Zellen der Stämme BW25113Δnuo/pBADnuo nuoFHis nuoG F515L und BW25113Δnuo/pBADnuo nuoFHis nuoG W221L wurden die Membranen präpariert und die Proteine durch Dodecylmaltosid aus den Membranen extrahiert. Mittels AnionenaustauschChromatographie und Affinitätschromatographie wurden analog zur Präparation aus dem Ausgangstamm BW25113Δnuo/pBADnuo nuoFHis die Komplex I Varianten gereinigt (Kap. 4.3.6.). Die Ausbeute der Präparation der Variante F515LNuoCD lag bei 2.5 mg mit einer spezifischen NADH/Ferricyanid Oxidoreduktase Aktivität von 155 µmol·min-1·mg-1 und die der Variante W221LNuoG bei 1.7 mg mit einer spezifischen Aktivität von 138 µmol·min-1·mg-1. Im Vergleich zur Präparation aus dem Ausgangstamm waren die Ausbeuten um 50 bzw. 70% geringer, während die spezifische NADH/Ferricyanid Oxidoreduktase Aktivität in der gleichen Größenordnung lag. Die Homogenität der Präparationen wurde mittels analytischer Größenausschluß-Chromatographie überprüft. Das Elutionsprofil der Variante F515LNuoCD zeigte neben dem Gipfel, der dem Komplex I zugeordnet wird, einen weiteren Gipfel, der dem NADH Dehydrogenase Fragment entspricht (Daten nicht gezeigt). Das NADH Dehydrogenase Fragment aus den Untereinheiten NuoE, NuoF und NuoG enthält neben der NADH-Bindestelle und dem FMN auch den His-tag N-terminal an NuoF. Wenn der Komplex I während der Präparation teilweise in seine Fragmente zerfällt, bindet auch das NADH Dehydrogenase Fragment an die Affinitätssäule und trägt ebenfalls zur NADH/Ferricyanid Oxidoreduktaseaktivität bei. Auch die Präparation des unveränderten Komplex I ist häufig durch das NADH Dehydrogenase Fragment verunreinigt. Dieses Fragment kann aber einfach mittels GrößenausschlußChromatographie abgetrennt werden (Kap. 4.1.1.). Aus diesem Grund wurde die gesamte Präparation der Variante F515LNuoCD auf eine präparative Größenausschluß- Chromatographiesäule aufgetragen und von den Fraktionen die NADH/Ferricyanid Oxidoreduktaseaktivität bestimmt (Abb. 4-17). 95 Ergebnisse A) Absorbanz bei 280 nm [mAu] 60 150 50 40 100 30 20 50 10 0 0 5 10 15 20 25 30 Absorbanz bei 280 nm [mAu] 70 200 NADH/Ferricyanid Oxidoreduktaseaktivität -1 -1 [µmol·min ·mg ] B) 25 20 15 10 5 0 Volumen [mL] 5 10 15 20 25 Volumen [mL] Abbildung 4-17: Größenausschlußchromatographie der Präparation der Komplex I Variante F515LNuoCD. Gezeigt sind die präparative Größenausschlußchromatographie (A) und die anschließende analytische Chromatographie (B). Die Absorbanz bei 280 nm ist als durchgezogene Linie und die NADH/Ferricyanid Oxidoreduktaseaktivität der Fraktionen als Punkt/Strich Linie gezeigt. Wie erwartet, zeigt das Elutionsprofil zwei Gipfel, die beide NADH/Ferricyanid Oxidoreduktaseaktivität aufwiesen. Die Fraktionen des Gipfels, der dem Komplex I zugeordnet wird, wurden vereinigt und mittels analytischer Größenausschlußchromatographie analysiert (Abb. 4-17). Das Elutionsprofil zeigt nur einen Gipfel im Größenbereich des Komplex I, die gereinigte Variante F515LNuoCD ist also stabil und homogen. Das Elutionsprofil der analytischen Größenausschluß-Chromatographie der präparierten Variante W221LNuoG enthält ebenfalls die dem Komplex I und dem NADH Dehydrogenase Fragment zugeordneten Gipfel (Abb. 4-18). Da der Gipfel im Größenbereich des NADH Dehydrogenase Fragmentes aber wesentlich intensiver ist, ist die Komplex I Variante vermutlich nicht stabil und zerfällt während der Präparation. Die Fraktionen des Gipfels im Größenbereich des Komplex I wurden vereinigt und wieder auf eine GrößenauschlußChromatographiesäule aufgetragen, das zugehörige Elutionsprofil zeigte jedoch ebenfalls den Gipfel des Zerfallsprodukt (Daten nicht gezeigt). 96 Ergebnisse Absorbanz bei 280 nm [mAu] 18 16 14 12 10 8 6 4 2 0 5 10 15 20 25 Volumen [mL] Abbildung 4-18: Größenausschlußchromatographie der Präparation der Komplex I Variante W221LNuoG. Gezeigt ist die Absorbanz bei 280 nm. Der Austausch des Tryptophans W221 gegen ein Leucin führt zur Destabilisierung des Komplex I, diese Variante gibt keinen Aufschluss über eine mögliche Beteiligung des Tryptophans am Elektronentransport. Da die Membranen der Mutante BW25113Δnuo/pBADnuo nuoFHis nuoG W221Y, in der das Tryptophan gegen ein Tyrosin ausgetauscht wurde, ebenfalls verminderte d-NADH Oxidase Aktivität im Vergleich zum Ausgangsstamm zeigten, sollte die Stabilität der Variante W221Y NuoG überprüft werden. Dazu wurden aus den Membranen der Mutante BW25113Δnuo pBADnuo nuoFHis nuoG W221Y die Membranproteine extrahiert und mittels Zuckergradientenzentrifugation aufgetrennt. Zum Vergleich wurde auch ein Membranextrakt des Ausgangsstammes aufgetragen. Die NADH/Ferricyanid Oxidoreduktase Aktivität der Fraktionen des Gradienten wurden bestimmt. Abbildung 4-19 zeigt die Aktivitätsprofile. NADH/Ferricyanid Oxido-1 -1 reduktaseaktivität [µmol min ml ] 2,5 2,0 1,5 1,0 0,5 0,0 2 4 6 8 10 12 14 16 18 Fraktion Abbildung 4-19.: Zuckergradientenzentrifugation der Detergenzextrakte aus Cytoplasmamembranen der Stämme BW25113Δnuo/pBADnuo nuoFHis nuoGW221Y (▲) und BW25113Δnuo/pBADnuo nuoFHis (■). Die Gradienten wurden mit 15 - 25 mg Protein beladen und die NADH/Ferricyanid Oxidoreduktase Aktivitäten auf 10 mg Proteinauftrag normiert. 97 Ergebnisse Die Membranextrakte beider Stämme zeigen im Aktivitätsprofil des Zuckergradienten die bekannten zwei Aktivitätsmaxima. Das Maximum um die Fraktionen 5 und 6 wird der alternativen NADH Dehydrogenase zugeordnet, das Maximum um Fraktion 13 dem Komplex I. Der Membranextrakt der Mutante enthält also die stabile Komplex I Variante W221Y NuoG. Die Fraktionen 8-11 des Gradienten zeigen leicht erhöhte Aktivität im Vergleich zum Ausgangstamm, die von dem NADH Dehydrogenase Fragment hervorgerufen werden könnte. Es kann also nicht ausgeschlossen werden, dass ein Teil des aus den Membranen herausgelösten Komplex I Variante zerfällt. 4.5 ESR-Spektroskopische Charakterisierung weiterer Proteine 4.5.1. ESR-spektroskopische Charakterisierung einer aus M. mazei Superoxid Reduktase Das Protein MM0632 aus M. mazei hat eine Größe von 20 kDa, zeigt Superoxid Reduktase (SOR) Aktivität und enthält pro mol Protein 5.7 mol Fe und 4.5 mol anorganischen Schwefel. Die Sequenz des Proteins zeigt das Bindemotivs eines [Fe(His)4(Cys)]-Zentrums, dass in den bisher bekannten SOR das aktive Zentrum bildet. Zusätzlich existieren ein Bindemotiv der Form Cys(X)7CysXXCys und ein viertes Cystein in dem im Vergleich zu den bisher bekannten SOR verlängerten C-Terminus des Proteins (Krätzer et al., 2010). In dieser Arbeit sollte in Zusammenarbeit mit Christian Krätzer aus dem Arbeitskreis Prof. Deppenmeier (Institut für Mikrobiologie und Biotechnologie, Universität Bonn) die Kofaktoren von MM0632 durch ESR-Spektroskopie charakterisiert werden. Die ESR-Proben wurden anaerob von Christian Krätzer angesetzt. Es wurden ESR-Spektren des isolierten Enzyms (10 mg/mL in 50 mM TRIS, 150 mM NaCl, 2.5 mM Desthiobiotin, 2.5 mM Dithioerythritol, pH 8) und des mit einigen Körnchen Dithionit reduzierten Proteins aufgenommen (Abb. 4-20). 98 Ergebnisse A 2.047 B 2.03 1.93 4.3 110 120 130 140 150 160 170 180 190 200 280 300 Magnetfeld [mT] 320 340 360 380 400 Magnetfeld [mT] Abbildung 4-20: ESR-Spektren des Proteins MM0632. A zeigt das Differenzspektrum der Spektren der mit Dithionit reduzierten Probe und des isolierten Enzyms aufgenommen bei 6 K und 10 mW. Pro Spektrum wurden fünf scans aufgenommen, die Modulationsamplitude betrug 10 G. B Zeigt das Spektrum der mit Dithionit reduzierten Probe aufgenommen bei 13 K und 10 mW mit einer Modulationsmaplitude von 0.6 mT. Die Mikrowellenfrequenz betrug 9.46 GHz, die Zeitkonstante 0.164 s und die Aufnahmegeschwindigkeit 17.9 mT/min. Die angegeben Zahlen stehen für die entsprechenden g-Werte. Das bei 13 K aufgenommene Spektrum der reduzierten Probe zeigt ein axiales Signal mit gx,y= 1.93 und gz = 2.047 (Abb. 4-20, B), dessen Intensität mit steigender Temperatur abnimmt und das nicht in den Spektren der oxidierten Probe beobachtet wurde (Daten nicht gezeigt). Dieses Signal wird einem vierkernigen high-spin Fe-S-Zentrum zugeordnet. Es ist bei g = 2.03 überlagert von einem zweiten Signal unbekannter Herkunft. Abbildung 4-20 (A) zeigt das Differenzspektrum aus einem bei 6 K aufgenommenen Spektrum des isolierten, im oxidierten Zustand vorliegenden Proteins und aus einem bei 6 K aufgenommen Spektrum der reduzierten Probe. Es zeigt ein typisches Signal eines high spin (S = 5/2) Fe 3+-Zentrums bei g = 4.3. Dieses Signal wurde dem [Fe(His)4(Cys)] Zentrum zugeordnet und wurde ebenfalls in den SOR aus Pyrococcus furiosus (Clay et al., 2002) und Desulfovibrio vulgaris (Clay et al., 2003) nachgewiesen. In Verbindung mit der Aminosäuresequenz und den Gehalt an Eisen und anorganischem Sulfat wurden ein [Fe(His)4(Cys)]-Zentrum und ein vierkerniges Fe-SZentrum durch ESR-Spektroskopie als Kofaktoren in MM0632 nachgewiesen. Ein zusätzliches Signal unbekannter Herkunft wurde detektiert und stammt möglicherweise von einem low-spin S = ½ Fe3+, vermutlich lokalisiert im [Fe(His)4(Cys)]-Zentrum. 99 Ergebnisse 4.5.2. ESR-spektroskopische Charakterisierung von BamBC aus Geobacter metallireducens Das sauerstoffempfindliche Protein BamBC aus dem obligativ anaeroben Bakterium G. metallireducens hat eine Größe von etwa 180 kDa und eine Untereinheitenzusammensetzung der Form α2β2. Es ist am Abbau von Aromaten beteiligt, indem es die Reaktion von Benzoyl-CoA zu Dienyl-CoA katalysiert. Die Katalyse der Rückreaktion, also die Umsetzung des Dienyl-CoA, wurde in Anwesenheit eines Elektronenakzeptors nachgewiesen. Pro mol einer αβ-Einheit wurden 15.2 ± 0.6 mol Eisen und 12.7 ± 1.4 mol anorgansicher Schwefel kolorimetrisch bestimmt und eine massenspektrometrische Analyse ergab pro Einheit 0.9 W, 1.2 Zn und 2.1 Ca. UV-vis Spektren des isolierten Proteins zeigen die Anwesenheit von Fe-S-Zentren (Kung et al., 2009). Die Sequenz von BamB enthält ein Bindemotive für ein Wolfram-Kofaktor und ein vierkerniges Fe-S-Zentrum und die Sequenz von BamC zeigt ebenfalls Bindemotive für Fe-S-Zentren (Wischgoll et al., 2005). In dieser Arbeit sollte das Protein in Zusammenarbeit mit Herrn Johannes Kung aus der Arbeitsgruppe von Prof. M. Boll (Institut für Biochemie, Universität Leipzig) durch ESR-Spektroskopie charakterisiert werden. Die Proben wurden anaerob von Herrn Johannes Kung angesetzt. Es wurden Spektren des isolierten Proteins (300 µM in 20 mM Triethanolamin/HCl, 5 mM MgCl2, pH 7.8 und 5% PEG 4000 (w/v)) und zum Vergleich Spektren des mit Dienyl-CoA (4.6 mM) oder Dithionit (4 mM) reduzierten Proteins vermessen. Die bei 10 K aufgenommenen Spektren der Dienyl-CoA reduzierten Proben zeigen mehrere überlagerte Signale (Abb. 4-21, A3), deren Intensität mir Erhöhung der Temperatur abnimmt und welche nicht in den Spektren der oxidierten Probe detektiert wurden (Daten nicht gezeigt). Diese Signale werden mehreren vierkernigen Fe-S-Zentren zugeordnet. Das bei 40 K aufgenommene ESR-Spektrum des isolierten Proteins zeigt ein axiales Signal mit g x = 1.893 und gy,z = 1.986 (Abb. 4-21, B1) und das bei 80 K aufgenommene Spektrum der selben Probe zeigt ein rhombisches Signal mit den gleichen gx und gy-Werten und einem g z-Wert von 2.013 (Abb. 4-21, B2). Diese Signale werden einem W(V)-Kofaktor zugeordnet und wurden nicht in den Spektren der mit Dienyl-CoA reduzierten Probe beobachtet (Abb. 4-21, B3). 100 Ergebnisse Abbildung 4-21: ESR-Spektren von BamBC. A1 zeigt ein Differenzspektrum aus einem bei 10 K und 20 mW aufgenommen Spektrum und dem Spektrum B1 des isolierten Proteins, A2 zeigt das bei 20 K und 5 mW aufgenommene Spektrum der mit Dithionit versetzten Probe und A3 zeigt das bei 10 K und 20 mW aufgenommene Spektrum der mit Dienyl-CoA reduzierten Probe. B1 und B2 zeigen Spektren des isolierten Enzyms aufgenommen bei 40 K bzw. 80 K und 5 mW. B3 zeigt das bei 40 K und 5 mW aufgenommene Spektrum der mit Dienyl-CoA reduzierten Probe. Die angegeben Zahlen stehen für die entsprechenden g-Werte. Die Mikrowellenfrequenz betrug 9.46 GHz, die Modulationsamplitute 0.6 mT, die Zeitkonstante 0.164 s und die Aufnahmegeschwindigkeit 17.9 mT/min. Die Abbildung wurde Kung et al. (2009) entnommen und modifiziert. Das Differenzspektrum aus einem bei 10 K aufgenommen Spektrum und dem bei 40 K gemessenen Spektrum der oxidierten Probe zeigt zusätzlich das Signal eines dreikernigen FeS-Zentrums (Abb. 4-21, A3). Dieses Signal wurde nicht in den Spektren der mit Dienly-CoA versetzten Probe beobachtet (Daten nicht gezeigt). In Verbindung mit den Informationen aus den Aminosäuresequenzen und den Metallanalysen lässt sich aus den ESR-spektroskopsichen Daten ableiten, das BamB einen Wolfram-Kofaktor und ein vierkerniges Fe-S-Zentrum koordiniert, während auf BamC zwei vierkernige und ein dreikerniges Fe-S-Zentrum gebunden sind. Die Spektren der mit Dithionit reduzierten Probe, zeigen das Signal des W(V)-Kofaktors und in geringer Intensität das der vierkernigen Fe-S-Zentren (Abb. 4-21, 101 Ergebnisse A2). Das Signal, das dem dreikernigen Fe-S-Zentrum zugeordnet wurde, wurde nicht detektiert. Demnach wird das dreikerinige Fe-S-Zentrum durch Dithinoit reduziert, während das Wolfram-Zentrum weiterhin oxidiert vorliegt. In der mit Dithionit versetzten Probe sind die vierkernigen Fe-S-Zentren im Vergleich zu der mit Dienyl-CoA versetzten Probe nur zu einem geringen Maß reduziert. 102 Diskussion 5. DISKUSSION 5.1 Die MHQ-Technik zur Beobachtung des Elektronentransfers innerhalb des Komplex I aus E. coli Zu verschiedenen Zeitpunkten der physiologischen Reaktion des isolierten Komplex I aus E. coli wurde die Reduktion einzelner Fe-S-Zentren erfasst. Dazu wurde mit der MHQ-Technik die Reaktion des Komplex I mit NADH zu Zeitpunkten im µs Bereich beendet und die Proben durch Tieftemperatur ESR-Spektroskopie analysiert. Zunächst wurden durch Vorversuche mit dem Komplex I und dem NADH Dehydrogenase Fragment die geeigneten Reaktionsbedingungen ermittelt. Die ESR-Spektren zeigen jeweils ein Apparatur-bedingtes Artefakt im Bereich von g = 2.0, das die Auswertung der Signale der Fe-S-Zentren im gzBereich nicht zulässt. Gerade in diesem Bereich sind die Signale der vierkernigen Zentren N2, N3 und N4 aufgelöst, somit ist die Analyse der Reduktionskinetik nur eingeschränkt möglich (Kap. 4.2). In der vorliegenden Arbeit wird die Zuordnung der ESR-Signale zu den einzelnen Fe-SZentren nach der Nomenklatur von Ohnishi vorgenommen (Ohnishi, 1998). Für die zweikernigen Fe-S-Zentren wurden die Zeitkonstanten der Reduktion des N1a zu ~ 3.0 ms und für die der Reduktion des N1b zu ~ 4.3 ms bestimmt (Tab. 4-3). Die Signale der vierkernigen Fe-S-Zentren zeigen die Reduktion von N2 ab einem Reaktionszeitpunkt von 404 µs, während die Reduktion der Fe-S-Zentren N3 und N4 erst ab einem Reaktionszeitpunkt von 1.2 ms detektiert wurde (Abb. 4-8). Diese Erkenntnisse stehen im Einklang mit Ergebnissen einer Arbeit, die den Elektronentransfer innerhalb des Komplex I aus E. coli mit der gleichen Methode untersucht hat (Verkhovskaya et al., 2008). Der Aufbau zum Ansetzen der Proben unterscheidet sich hauptsächlich in der Einfriertechnik von der in dieser Arbeit verwendeten Apparatur. Die Analyse der ESR-Signale ergab kürzere Zeitkonstanten für die Reduktion der Fe-S-Zentren N1a und N2, während die Reduktion der Fe-S-Zentren N1b und N6b langsamer verlief. Die Daten für die Reduktion des Fe-SZentrums N6b wurden durch Auswertung des Signals bei g x,y = 1.88 und gz = 2.09 erhalten. Beiträge weiterer Fe-S-Zentren zu den Signalen der bei 10 K und 10 mW aufgenommenen Spektren wurden von den Autoren ausgeschlossen (Verkhovskaya et al. 2008; Belevich et al., 2007). Das Signal der bei 11 K und 10 mW aufgenommenen Spektren bei g = 1.88 wird den Fe-S-Zentren N3 und N4 und das Signal bei g = 2.09 ebenfalls dem Zentrum N4 zugeordnet 103 Diskussion (Leif et al., 1995; Kap. 2.2.1). Das Signal mit dem g-Wert 2.09 ist aufgrund des oben genannten Artefakts jedoch nicht auswertbar. In beiden Arbeiten wird sowohl eine frühe Reduktion des nahe des FMN und abseits der übrigen Fe-S-Zentren gelegenen N1a als auch des nahe der Ubichinonbindestelle gelegenen N2 beobachtet. Auffallend ist jedoch der große Unterschied der Zeitkonstanten der Reduktionen in beiden Experimente. Die von Verkhovskaya et al. veröffentlichten Spektren der Proben zeigen ab dem ersten aufgenommenen Reaktionszeitpunkt von 90 µs die Signale der Fe-S-Zentren N1a, N1b und N2 und die Zeitkonstanten der Reduktion von N1a und N2 wurden zu ~ 90 µs bestimmt. In der vorliegenden Arbeit zeigen die Spektren der Proben zu den Zeitpunkten von 97 µs und 131 µs keine Signale, erst ab 404 µs werden Signale des N1a bzw. N2 detektiert. Die Zeitkonstante für die Reduktion von N1a wurde zu ~3.0 ms bestimmt. Der Nachweis eines Semichinonradikals ist in beiden Arbeiten nicht gelungen. Radikale führen in ESR-Spektren zu Signalen im Bereich von g = 2.0, genau in diesem Bereich wird das oben genannte Artefakt beobachtet. Die Identifizierung eines Radikalsignals war in dieser Arbeit somit nicht möglich. Auch in den Spektren der Arbeiten von Verkhovskaya et al. wurde nach Angaben der Autoren ein Artefakt im gleichen Bereich beobachtet (Belevich et al., 2009). Dies ist eine mögliche Erklärung, warum auch bei diesen Experimenten kein Radikal detektiert wurde. Um sicher zu stellen, dass die Kinetik des Elektronentransfers des Komplex I und nicht ein steady-sate erfasst wird, wurden in der vorliegenden Arbeit Proben mit einem durch den Hemmstoff Piericidin A inhibierten Komplex I angesetzt. Das Piericidin A blockiert die Ubichinonbindestelle und verhindert eine Übertragung der Elektronen auf das in der Präparation vorliegende Ubichinon (Friedrich et al., 1994). Auf diese Weise wurde sicher gestellt, dass die Elektronen der ersten Umsatzrate beobachtet wurden. Dabei unterscheiden sich die aufgenommenen Spektren der Proben mit und ohne Hemmstoff jedoch kaum. Auffällig ist, dass das Signal des Fe-S-Zentrums N2 in den Spektren der Proben ohne Hemmstoff (794 µs) später detektiert wurde als in den Spektren der Proben mit dem Hemmstoff (404 µs). Die spätere Detektion ist vermutlich auf die Tatsache zurückzuführen, dass in diesen Proben eine Übertragung auf das Ubichinon stattfindet. In Anwesenheit des Hemmstoffes wird der Reduktionsgrad des N2 nur durch die Aufnahme von Elektronen bestimmt, ohne Hemmstoff wird der Reduktionsgrad zusätzlich durch die Abgabe der Elektronen an Ubichinon beeinflusst. 104 Diskussion Die in dieser Arbeit gezeigten Spektren geben Aufschluss über die physiologische Reaktion des Komplex I. Die Daten zeigen, dass das Fe-S-Zentrum N1a an der physiologischen Reaktion beteiligt ist. Die von Verkhovskaya et al. getroffene Aussage, dass das erste Elektron, eines von NADH auf das FMN übertragenen Elektronenpaares, auf N2 übertragen wird und das zweite Elektron auf N1a, wird unterstützt. Die beinahe simultane Reduktion des nahe des FMN gelegenen N1a und des am Ende der Elektronentransportkette gelegene N2 lassen diesen Schluss zu. Die große Abweichung in den Reduktionszeiten wird vermutlich durch apparative Unterschiede hervorgerufen. Die in dieser Arbeit ermittelten Geschwindigkeiten des Elektronentransports sind deutlich niedriger, liegen aber in einem Bereich, der mit der experimentell bestimmten Geschwindigkeit des Gesamtprozesses vereinbar ist. Weitere Experimente zu dieser Thematik sind in der Literatur nicht beschrieben. Diese Arbeit zeigt auch die Grenzen der eingesetzten Methode zur Analyse der Reaktionskinetik der Fe-S-Zentren des Komplex I auf. Die Auswertung der ESR-Spektren des Komplex I ist aufgrund der Anzahl und der Überlappung der Signale nicht trivial und wird durch das Artefakt in den Spektren der MHQ-Proben deutlich erschwert. Aufgrund der benötigten großen Menge Komplex I ist eine breite Anwendung der Methode zur Charakterisierung von Komplex I Varianten nur eingeschränkt möglich. 5.2. Die Bedeutung des Fe-S-Zentrum N1a In dieser Arbeit wurde erstmals gezeigt, dass das zweikernige Fe-S-Zentrum N1a auf der Untereinheit NuoE im Komplex I aus E. coli für die Stabilität des Proteins essentiell ist. Der Austausch der einzelnen Cysteine gegen Alanine in den drei Positionen C92, C133 und C137 des Bindemotivs des Fe-S-Zentrums verhindert die Assemblierung des Komplex I in der cytoplasmatischen Membran. Der Austausch auf der vierten Position C97 führt zwar zu einem aktiven Komplex I in der Membran, dieser zerfiel jedoch nach der Extraktion des Proteins aus der Membran. Der Austausch der Cysteine C97 und C133, jeweils eines an jedem Fe-Atom von N1a, gegen Serin führte zum gleichen Effekt. Es wurden geringe Mengen an aktivem Komplex I in der Membran detektiert, dieser war jedoch nach Detergenzextraktion nicht stabil (Kap. 4.3.7.). Aufgrund dieser Erkenntnisse wurde in einem weiteren Experiment in unserem Arbeitskreis versucht, ein Fragment aus den Untereinheiten NuoE und NuoF aus A. aeolicus ohne das Fe-S-Zentrum N1a heterolog in E. coli überzuproduzieren. Dafür wurde ein Plasmid genutzt, in dem der Teil des Gens nuoE fehlt, der die C-terminale und damit die Fe-S105 Diskussion Zentrum N1a koordinierende Domäne trägt. Das produzierte Protein bestand jedoch nur aus der Untereinheit NuoF (Yao, 2010) und mittels ESR-Spektroskopie wurde gezeigt, dass das Fe-S-Zentrum N3 in der Präparation fehlte (unveröffentliche Ergebnisse). Diese Daten lassen vermuten, dass auch im Komplex I aus A. aeolicus die intakte N1a koordinierende Cterminale Domäne wichtig für die Stabilität des gesamten Komplex I ist. Sie unterstützt vermutlich die Interaktion der N-terminalen Domäne der Untereinheit NuoE mit den anderen Untereinheiten des Komplex I. In der Literatur liegen bisher wenige Erkenntnisse zur strukturellen Bedeutung des Fe-S-Zentrums N1a vor. In der zu NuoE homologen Untereinheit Nqo2 aus P. denitrificans wird N1a über das gleiche Bindemotiv wie in E. coli (CX4CX35CX3C) koordiniert, die Cysteine dieses Motivs wurden ebenfalls gegen Alanine und Serine ausgetauscht (Yano et al., 1994). Die varianten Untereinheiten wurden heterolog in E. coli überproduziert, isoliert und die Menge an Eisen im Protein sowohl chemisch als auch durch Spinquantifizierung mittels ESR-Spektroskopie bestimmt. Die chemisch bestimmte Menge des Eisens pro mg Protein lag lediglich bei 12-27% und die durch Spinquantifizierung bestimmte Menge bei 2 bis 5 % im Vergleich zum unveränderten Protein. Das Signal des N1a in den ESR-Spektren war bei den varianten Proteinen verbreitert, die g-Werte leicht verschoben und zum Teil traten zusätzliche Signale auf. Der Austausch der einzelnen Cysteine im Bindemotiv hatte anscheinend keinen Einfluss auf die Stabilität der Untereinheit, die Menge an koordinierten Eisen im Protein und die ESR-Spektren der Fe-S-Zentren waren jedoch verändert (Yano et al., 1994). Die [2Fe-2S] Ferredoxine aus Clostridium pasteurianum und A. aeolicus sind homolog zur C-terminalen, N1a tragenden Domäne der Untereinheit NuoE und koordinieren das zweikernige Fe-S-Zentrum mit dem Bindemotiv CX12CX32CX3C (Meyer, 2001). Der Austausch zwei dieser Cysteine gegen Alanine bzw. drei dieser Cysteine jeweils gegen Serine im Ferredoxin aus C. pasteurianum führt zu einem stabilen Protein mit dem gebunden Fe-S-Zentrum (Fujinaga et al., 1993; Golinelli et al., 1998). Zwei der entsprechenden Serin-Varianten aus A. aeolicus wurden kristallisiert. Die Struktur mit einer Auflösung von 1.25 Å bestätigte die Beteiligung der Serine an der Koordination des Fe-SZentrums und die Gesamtstruktur der varianten Proteine war gegenüber des unveränderten Proteins nahezu unverändert (Yeh et al., 2002). Weitere Mutageneseexperimente haben gezeigt, dass die Cysteine C24 und C60 in einem strukturell flexibleren Bereich des Proteins liegen, während die übrigen, C11 und C56, von einem eher starren Proteinbereich umgeben sind (Golinelli et al., 1998). Es wird vermutet, dass ein loop in der Nähe des Fe-S-Zentrums das Zusammenspiel von flexibler und starrer Proteinumgebung und der starren Geometrie des Fe-S-Zentrums ermöglicht und damit die Stabilität des Proteins gewährleistet (Yeh et al., 106 Diskussion 2002). Dieser loop hat keine Entsprechung in der Untereinheit NuoE und seinen Homologen (Yeh et al., 2000). Möglicherweise führt der Austausch der Cysteine im Bindemotiv zu größeren strukturellen Veränderungen, die zwar die Produktion der Untereinheit zulassen, die bei einer Überproduktion des gesamten Komplex I aus P. denitrificans die Assemblierung der Untereinheiten aber verhindern würden. Der Sequenzvergleich von NuoE aus P. denitrificans, A. aeolicus und E. coli zeigt, dass der C-Terminus von NuoE aus P. denitrificans um 69 Aminosäuren verlängert ist. Diese zusätzlichen Aminosäuren könnten die strukturelle Flexibilität und damit die Stabilität der Varianten begünstigen. Interessant wäre in diesem Zusammenhang, ob bei Mutagenese des Bindemotivs auch die Produktion des gesamten P. denitrificans Komplex I möglich ist. Ein Austausch der Cysteine C92 und C137 gegen Serine in der E. coli Untereinheit NuoE wurde nicht vorgenommen. Die Ergebnisse der in dieser Arbeit charakterisierten Mutanten im Vergleich mit den Varianten aus P. denitrificans lassen vermuten, dass auch diese Komplex I Varianten nicht stabil wären. Im Komplex I bilden die sieben Fe-S-Zentren N3, N1b, N4, N5, N6a, N6b und N2 eine Elektronentransportkette zwischen dem FMN, das die Elektronen vom NADH aufnimmt, und dem Ubichinon als Elektronenakzeptor. Aufgrund seiner aus der Struktur des hydrophilen Teils des Komplex I aus T. thermophilus bekannten Lage ist das Fe-S-Zentrums N1a nicht Teil dieser Kette (Sazanov & Hinchliffe, 2006). ESR-Spektren des mit NADH reduzierten Komplex I aus E. coli zeigen, dass Elektronen auf dieses Zentrum übertragen werden (Leif et al., 1995; Verkhovskaya et al., 2008) Die in dieser Arbeit aufgenommenen Spektren zur Verfolgung der Elektronenübertragung innerhalb des Komplex I zeigen, dass N1a auch während der physiologischen Reaktion Elektronen aufnimmt (Kap. 5.1.). Es wird vermutet, dass das Fe-S-Zentrum, durch die zeitweilige Aufnahme eines Elektrons, die Radikalbildung beim Umschalten zwischen der Zweielektronenübertragung des NADH auf die Einelektronenübertragung der Fe-S-Zentren am FMN vermeidet. Ob dies der Fall ist, lässt sich durch den gewählten Ansatz nicht klären. Die aus den MHQ-Experimenten gewonnen Ergebnisse lassen aber vermuten, dass N1a nicht nur strukturelle Bedeutung im Komplex I hat. 107 Diskussion 5.3 Die mögliche Beteiligung Elektronentransport im Komplex I aromatischer Aminosäuren am In dieser Arbeit wurde erstmals experimentell untersucht, ob aromatische Aminosäuren an der Elektronenübertragung zwischen den Fe-S-Zentren im Komplex I beteiligt sind. Dazu wurden aromatische Aminosäuren an fünf Positionen im Komplex I aus E. coli gegen die jeweils anderen aromatischen Aminosäuren und gegen Leucin ausgetauscht und der Einfluss des Austausches auf die Aktivität des Komplex I in den cytoplasmatischen Membranen ermittelt. Die Positionen wurden aufgrund einer theoretischen Arbeit ausgewählt, in der für den Elektronentransport im Komplex I ein hopping Mechanismus und in der unter Einbeziehung der Struktur des Komplex I aus T. thermophilus sieben konservierte, aromatische Aminosäuren als mögliche hopping Partner vorgeschlagen werden (Wittekindt et al., 2009). Im E. coli Komplex I führte der Austausch des Phenylalanins F515NuoCD zwischen den Fe-SZentren N5 und N6a gegen Leucin zu einer stabilen Komplex I Variante, deren Aktivität in der Membran um ein Drittel reduziert war, während ein Austausch gegen ein Tryptophan oder ein Tyrosin die Aktivität nicht beeinflusste (Tab. 4-14 und Kap. 4.4.3.). Diese Messungen unterstützen die Vermutung, dass die konservierte aromatische Aminosäure F515 am Elektronentransport beteiligt ist. Der Austausch des Tryptophans W221 NuoG zwischen diesen beiden Fe-S-Zentren gegen ein Tyrosin führte ebenfalls zu einer stabilen Komplex I Variante, deren Aktivität in der Membran um knapp ein Drittel vermindert war. Der Austausch des Tryptophans gegen Leucin führte zu einem instabilen Komplex I, während der Austausch gegen ein Phenylalanin zu einem stabilen Komplex I führte und keinen Einfluss auf die Aktivität des Komplex I zeigte (Tab. 4-14 und Kap. 4.4.3.). Die Eigenschaften der Varianten W221LNuoG und W221FNuoG stehen im Einklang mit Ergebnissen zum E. coli Komplex I, die in einem anderen Kontext veröffentlichen wurden. Der Austausch des W221NuoG gegen ein Alanin führte zu keinem stabilen Komplex I, während der Austausch gegen ein Phenylalanin die Stabilität und die Aktivität des Komplex I nicht beeinflusste (Nakamaru-Ogiso et al., 2008). Eine Beteiligung der konservierten aromatischen Aminosäure W221 NuoG am Elektronentransport kann demnach nicht ausgeschlossen werden. Nach der hopping Theorie sind diese beiden Aminosäuren entweder seriell am Elektronentransport N5→ F515 → W221 → N6a beteiligt, oder die Elektronen werden alternativ über die Wege N5 → W221 → N6a oder N5→ F515→N6a übertragen (Wittekindt et al., 2009). Die Ergebnisse aus dieser Arbeit unterstützen die serielle Beteiligung beider Aminosäuren oder die Annahme des 108 Diskussion Phenylalanins F515 als alleinigen hopping Partner. Nach der Theorie ist die Beteiligung einer dritten Aminosäure, des Histidin H514NuoCD, am Elektronentransport zwischen den beiden Zentren möglich. Der Austausch dieser Aminosäure gegen Lecuin hatte jedoch keinen Einfluss auf die Aktivität des Komplex I (Tab. 4-14). In Y. lipolytica wurde das zu H514NuoCD homologe H384 gegen Alanin ausgetauscht. Dies führte zu einem um 70% geringeren Gehalt des Komplex I in den Membranen und zu einer Halbierung der Komplex I vermittelten Aktivität der Membranen (Kashani-Poor et al., 2001). Da im Komplex I aus Y. lipolytica an den zu W221NuoG und F515NuoCD homologen Positionen ebenfalls ein Tryptophan und ein Phenylalanin liegen, ist die verminderte Komplex I Aktivität dieser Alanin-Mutante vermutlich auf strukturelle Gründe zurückzuführen. Der Austausch einzelner Aminosäuren kann immer zu strukturellen Veränderungen führen. Eine verminderte Aktivität der Varianten kann somit auch durch Änderung der Lage der beiden Fe-S-Zentren zueinander hervorgerufen werden. Um diese Möglichkeit auszuschließen, wird die Kristallstruktur der Varianten benötigt. Die ist jedoch nicht trivial, da schon die Struktur des unveränderten, hydrophilen Teils des Komplex I aus E. coli bisher nicht verfügbar ist. Für eine Beteiligung aromatischer Aminosäuren an dem Transferschritt zwischen den Fe-S-Zentren N5 und N6a spricht der große Abstand zwischen den Zentren von 16.9 Å zwischen den Mittelpunkten und 14.0 Å zwischen den Kanten. Nach der allgemein anerkannten through-space Theorie tunneln Elektronen innerhalb einer Elektronenübertragungskette in Proteinen zwischen Redoxzentren mit einem Abstand kleiner als 14 Å (Page et al., 1999; Moser et al., 2006). Nach theoretischen Berechnungen von Koslowski und Mitarbeitern liegt die berechnete Elektronentransferrate für die direkte Übertragung zwischen den Fe-S-Zentren N5 und N6a bei 10-4 s-1 und ist damit um einige Größenordnungen zu niedrig, um die experimentelle Elektronentransferrate von etwa 50 s -1 erklären zu können. Durch Beteiligung der aromatischen Aminosäuren werden nach der hopping Theorie Transferraten im Bereich von 10 3 s-1 erreicht (Wittekindt et al., 2009.). Die Beteiligung einer aromatischen Aminosäure an genau diesem Transferschritt ist zudem wahrscheinlich, da die beiden Zentren auf zwei verschiedenen Modulen des Komplex I lokalisiert sind. Die Elektronentransportkette entstand vermutlich evolutionär durch die Anlagerung zweier Module (Kap. 2.2.), eine aromatische Aminosäure kann somit zur Überbrückung des ungewöhnlich langen Elektronenübertragungsweg zwischen den beiden Modulen dienen. 109 Diskussion In dieser Arbeit wurden außerdem das konservierte Phenylalanin F35NuoG zwischen den Fe-SZentren N1b und N4 (Mittelpunktsabstand: 13.9 Å, Kantenabstand: 10.7 Å) und das Phenylalanin F101NuoI zwischen den Fe-S-Zentren N6b und N2 (Mittelpunktsabstand: 14.2 Å, Kantenabstand: 10.5 Å) ausgetauscht. Der Austausch des Phenylalanins F101NuoI gegen Leucin, Tyrosin oder Tryptophan führte bei keiner Variante zu einer Veränderung der Aktivität im Vergleich zum unveränderten Komplex I. Das gleiche gilt für den Austausch des Phenylalanins F35NuoG gegen Leucin oder Tyrosin. Der Austausch des F35NuoG gegen ein Tryptophan führte zu einer Aktivitätsverminderung, diese wird aber möglicherweise durch strukturelle Veränderungen des größeren Restes hervorgerufen (Tab. 4-13 und 4-15). Im E. coli Komplex I liegt in direkter Nachbarschaft zum F35NuoG ein Tyrosin, im Komplex I aus T. thermophilus liegt an dieser Position ein Leucin. Möglicherweise ist im E. coli Komplex I das Tyrosin anstelle des F35NuoG an der Elektronenübertragung beteiligt. Die Aktivitätsminderung der Variante F35WNuoG wäre dann auf eine veränderte Position des Tyrosins aufgrund des eingefügten Tryptophans an der Nachbarposition zu erklären. Die Ergebnisse zu den Transferschritten N1b → N4 und N6b → N2 lassen zwei Möglichkeiten zu: Entweder sind diese aromatischen Aminosäuren nicht am Elektronentransport beteiligt oder der Einfluss der Beteiligung wird durch die in dieser Arbeit durchgeführten Messungen nicht erfasst. Die nach Wittekindt et al. berechneten Elektronentransferraten für die direkte Übertragung zwischen N1b und N4 (5 s-1) bzw. N6b und N2 (100 s-1) liegen nahe bzw. im Bereich der experimentell bestimmten Elektronentransferrate für den Gesamtprozess des in dieser Arbeit präparierten Komplex I (50 s-1). Auch ohne Beteiligung einer aromatischen Aminosäure können möglicherweise ausreichend hohe Geschwindigkeiten erreicht werden. Die Transferraten der direkten Übertragung werden in der theoretischen Arbeit als nicht ausreichend bezeichnet, da dort für den Gesamtprozess von einer höheren Übertragungsrate (170 s-1) ausgegangen wird. Dieser Wert ergab sich aus der Mittelung der Übertragungsraten des Komplex I aus unterschiedlichen Präparationen aus verschiedenen Organismen, auch des mitochondrialen Komplex I. Die Beteiligung der aromatischen Aminosäure F35NuoG und F101NuoCD führen bei beiden Übertragungsschritten zu berechneten Elektronentransferraten im Bereich von 10 5 s-1. Sowohl die berechneten Transferraten mit als auch ohne Beteiligung der aromatischen Aminosäuren können mit der in dieser Arbeit gemessene Aktivität erklärt werden. Es bleibt somit offen, ob aromatische Aminosäuren an diesen Übertragungsschritten beteiligt sind bzw. ob eine Beteiligung zwischen den relativ nah beieinander liegenden Fe-S-Zentren notwendig ist. 110 Diskussion Die nach der hopping Theorie am Übertragungsschritt zwischen den Fe-S-Zentren N3 und N1b (Mittelpunktsabstandstand: 14.2 Å, Kantenabstand: 11.0 Å) beteiligte Aminosäure F397NuoF wurde in dieser Arbeit nicht ausgetauscht. Diese Aminosäure ist nicht konserviert, im Komplex I einiger Organismen befindet sich an dieser Position ein Isoleucin. Die Abstände zwischen den Fe-S-Zentren lassen vermuten, dass auch in diesem Fall ein möglicher Effekt des Austausches nicht durch die Aktivitätsmessung erfasst werden würde. 5.4 Der Überexpressionsstamm BW25113 Δnuo pBADnuo nuoFHis Der Stamm ANN0221, der mit dem Plamsmid pBADnuo nuoFHis transformiert wurde, bietet ein gutes System zu Überproduktion des Komplex I und seiner Varianten (Pohl et al., 2007b). Da der Stamm ANN0221 aber zur in-trans-Komplementation fähig ist (Pohl, 2008), ist dieses System für eine Produktion von nicht stabilen oder inaktiven Komplex I Varianten nicht geeignet. In dieser Arbeit wurde im Rahmen der Mutagenesexperimente am Bindemotiv des Fe-S-Zentrums N1a gezeigt, dass im Stamm ANN0221 die episomal kodierte und nicht stabile Untereinheit NuoE durch eine chromosomal kodierte, stabile Untereinheit NuoE ersetzt wurde (Kap. 4.3.3. und 4.3.7.). Die Kombination des Stamm BW25113Δnuo (M. Vranas, unveröfftl. Ergebnisse) mit dem Plasmid pBADnuo nuoFHis als Produktionssystem für den Komplex I wurde in dieser Arbeit charakterisiert und steht als System zur Charakterisierung von Mutanten unabhängig der Eigenschaften der resultierenden Komplex I Varianten zur Verfügung (Kap. 4.3.6.). 5.5 ESR-Spektroskopische Charakterisierung zwei weiterer Proteine Das Protein MM0632 aus M. mazei ist der Vertreter einer neuen Klasse von Superoxid Reduktasen (SOR), die in anaeroben Organsimen an dem Abbau von reaktiven Sauerstoffspezies beteiligt ist (Krätzer et al., 2010). In dieser Arbeit wurde durch ESRSpektroskopie ein [Fe(His)4(Cys)]-Zentrum und eine vierkerniges Fe-S-Zentrum in dem isolierten Enzym nachgewiesen. Das [Fe(His)4(Cys)]-Zentrum ist in allen bisher untersuchten SOR der Klasse I und II enthalten, während das vierkerniges Fe-S-Zentrum erstmals in einem Protein mit SOR-Aktivität nachgewiesen wurde (Krätzer et al., 2010; Clay et al., 2002; Clay et al., 2003). Das Signal des vierkernigen Fe-S-Zentrums der Spektren des reduzierten 111 Diskussion Enzyms ist von einem Signal unbekannter Herkunft überlagert. Eine Simulation des Signals des Fe-S-Zentrums war aus diesem Grund bisher nicht möglich. Die Charakterisierung des zusätzlichen Signals, welches möglicherweise durch eine low-spin S = ½ Fe3+ Spezies hervorgerufen wird, steht noch aus. Das Protein BamBC aus G. metallireducens katalysiert die Reaktion von Dienyl-CoA zu Benzoyl-CoA in Anwesenheit eines Elektronenakzeptors (Kung et al., 2009). In einer kürzlich erschienen Veröffentlichung ist auch der Nachweise der Katalyse der Hinreaktion, also die Reduktion des Benzyol-CoA, gelungen (Kung et al., 2010). Das BamBC stellt somit den Vertreter einer neuen Klasse von Benzoyl-CoA Reduktasen dar, die eine ATPunabhängige Birch ähnliche Reduktion katalysieren (Wischgoll et al., 2005; Kung et al., 2009). Die Ergebnisse der ESR-spektroskopischen Charakterisierung des Proteins in dieser Arbeit in Verbindung mit Sequenzinformationen zeigen, dass dieses Protein einen WolframKofaktor, drei vierkernige Fe-S-Zentren und ein dreikerniges Fe-S-Zentrum koordiniert. Die Reduktion der Kofaktoren durch Dienyl-CoA lassen eine Beteiligung an der physiologischen Reaktion vermuten. Die Signale der vierkernigen Fe-S-Zentren sind in den Spektren überlagert, eine Auflösung der einzelnen Signale ist bisher nicht gelungen. 112 Literatur 6. LITERATUR Anraku, Y. & Gennis, R.B. (1987) The aerobic respiratory chain of Escherichia coli. Trends Biochem. Sci. 12, 262-266. Bald, W. D. (1985) The relative merits of various cooling methods. J. Microsc. 140, 17–40. Ballou, D.P. & Palmer, G. A. (1974) Practical rapid quenching instrument for the study of reaction mechanisms by electron paramagnetic resonance spectroscopy. Anal. Chem. 46, 1248-1253. Baranova, E.A., Morgan, D.J. & Sazanov, L.A. (2007) Single particle analysis confirms distal location of subunits NuoL and NuoM in Escherichia coli complex I. J. Struct. Biol. 159, 238-242. Bayer, T. (2008) Untersuchung von Expressionstämmen zur Überproduktion von Varianten der Escherichia coli NADH:Ubichinon Oxidoreduktase. Diplomarbeit, Albert-LudwigsUniversität Freiburg. Belevich, G., Euro, L., Wikström, M. & Verkhovskaya, M. (2007) Role of the conserved arginine 274 and histidine 224 and 228 residues in the NuoCD subunit of complex I from Escherichia coli. Biochemistry 46, 526-533. Belevich, G., Verkhovskaya, M. und Verkhovsky M.I. (2009) Electron transfer in respiratory complexes resolved by an ultra-fast freeze-quench approach. Meth. in Enzym. 456, 75-93. Berrisford, J. M. & Sazanov, L. A. (2009) Structural basis for the mechanism of respiratory complex I. J. Biol. Chem. 284, 29773-29783. Bollinger, J. M. (2008) Electron relay in proteins. Science 320, 1730-1731. 113 Literatur Braun, M., Bungert, S. & Friedrich, T. (1998) Characterization of the overproduced NADH dehydrogenase fragment of the NADH:ubiquinone oxidoreductase (complex I) from Escherichia coli. Biochemistry 37, 1861-1867. Bungert, S., Krafft, B., Schlesinger, R. & Friedrich, T. (1999) One-step purification of the NADH dehydrogenase fragment of the Escherichia coli complex I by means of Strep-tag affinity chromatography. FEBS Lett. 460, 207-211. Calhoun, M. W., Thomas, J. W. & Gennis, R. B. (1994) The cytochrome oxidase superfamily of redox-driven proton pumps. Trends Biochem. Sci. 19, 325-330. Carroll, J., Fearnley, I.M., Skehel, J.M., Shannon, R.J., Hirst, J. & Walker, J.E. (2006) Bovine complex I is a complex of 45 different subunits. J. Biol. Chem. 281, 32724-32727. Chance, B. (1943) J. Biol. Chem. 151, 553–577. Cheperanov, A. V. & de Vries, S. (2004) Microsecond freeze-hyperquenching: development of a new ultrafast micro-mixing and sampling technology and application to enzyme catalysis. Biochim. Biophys. Acta 1656, 1-31. Clay, M. D., Jenney, F. E., Hagedoorn, P. L., George, G. N., Adams, M. W. und Johnson, M. K. (2002) Spectroscopic studies of Pyrococcus furiosus superoxide reductase: Implications for active-site structures and the catalytic mechanism. J Am Chem Soc 124, 788-805 Clay, M. D., Emerson, J. P., Coulter, E. D., Kurtz, D. M. und Johnson, M. K. (2003) Spectroscopic characterization of the [Fe(His)4(Cys)] site in 2Fe-superoxide reductase from Desulfovibrio vulgaris. J Biol Inorg Chem 8, 671-682. Datsenko, K.A., und Wanner, B.L. (2000) One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. U.S.A.. 97, 6640-6645. Degli Esposti, M. (1998). Inhibitors of NADH-ubiquinone reductase: an overview. Biochim. Biophys. Acta 1364, 222-235. 114 Literatur De Smet, L., Savvides, S. N., Van Horen, E., Pettigrew, G. und Van Beeumen, J. J. (2006) Structural and mutagenesis studies on the Cytochrome c Peroxidase from Rhodobacter capsulatus provide new insights into structure-function relationships of bacterial Di-heme Peroxidases. J. Biol. Chem. 281, 4371-4379. Dias J. M., Alves, T., Bonifacio, C., Pereira, A.S., Trincao, J., Bourgeois, D., Moura, I. und Romaao, M.J. (2004) Structural basis for the mechanism of Ca2+ activation of the Di-Heme Cytochrome c Peroxidase from Pseudomonas nautica 617. Structure 12, 961–973. DuBois, D. (1941) An apparatus for the study of rapid chemical reactions. J. Biol. Chem.137, 123-138. Efremov, R. G., Baradaran R. und Sazanov, L.A. (2010) The architecture of respiratory complex I. Nature 465, 441-447. Esterhazy, D., King, M. S., Yakovlev, G. and Hirst, J. (2008) Production of reactive oxygen species by complex I (NADH:ubiquinone oxidoreductase) from Escherichia coli and comparison to the enzyme from mitochondria. Biochemistry 47, 3964–3971. Fendel,U., Tocilescu, M.A., Kerscher, S. & Brandt, U. (2008). Exploring the inhibitor binding pocket of respiratory complex I. Biochim. Biophys. Acta 1777, 660-665. Flemming, D., Hellwig, P., Lepper, S., Kloer, D. & Friedrich, T. (2006). Catalytic importance of acidic amino acids on subunit NuoB of the Escherichia coli NADH:Ubiquinone Oxidoreductase (Complex I) J. Biol. Chem. 281, 24781-24789. Friedrich, T., Hofhaus, G., Ise, W., Nehls, U., Schmitz, B. & Weiss, H. (1989) A small isoform of NADH:ubiquinone oxidoreductase (complex I) without mitochondrially encoded subunits is made in chloramphenicol-treated Neurospora crassa. Eur. J. Biochem. 180, 173180. 115 Literatur Friedrich, T., van Heek, P., Leif, H., Ohnishi, T., Forche, E., Kunze, B., Jansen, R., Trowitzsch-Kienast, W., Höfle, G., Reichenbach, H. & . Weiss, H. (1994). Two binding sites of inhibitors in NADH: ubiquinone oxidoreductase (complex I). Relationship of one site with the ubiquinone-binding site of bacterial glucose:ubiquinone oxidoreductase. Eur. J. Biochem. 219, 691-698. Friedrich, T., Steinmüller, K. & Weiss, H. (1995). The proton-pumping respiratory complex I of bacteria and mitochondria and its homologue in chloroplasts. FEBS Lett. 367, 107-111. Friedrich, T. & Weiss, H. (1997). Modular evolution of the respiratory NADH:ubiquinone oxidoreductase and the origin of its modules. J. Theor. Biol. 187, 529-540. Friedrich, T. (1998). The NADH:ubiquinone oxidoreductase (complex I) from Escherichia coli. Biochim. Biophys. Acta 1364, 134-146. Friedrich, T. & Scheide, D. (2000). The respiratory complex I of bacteria, archaea and eukarya and its module common with membrane-bound multisubunit hydrogenases. FEBS Lett. 479, 1-5. Friedrich, E. (2008) Die Bedeutung des [Fe-S]-Zentrums N1a der NADH:Ubichinon Oxidoreduktase aus Escherichia coli für deren Mechanismus. Diplomarbeit, Albert-LudwigsUniversität Freiburg. Fujinaga, J., Gaillard, J. und Meyer, J. (1993) Mutated forms of a [2Fe-2S] ferredoxine with serine ligands to the iron-sulfur-cluster. Biochem. Biophys. Res. Commun. 194, 104-111. Galkin, A. & Brandt, U. (2005). Superoxide Radical Formation by Pure Complex I (NADH:Ubiquinone Oxidoreductase) from Yarrowia lipolytica. J. Biol. Chem. 280, 3012930135. Gay, P., Le Coq, D., Steinmetz, M., Bekelman, T., und Kado, C.I. (1985) Positive selection procedure for entrapment of insertion sequence elements in gram-negative bacteria. J. Bacteriol. 164, 918-921. 116 Literatur Gibson, Q.H. & Milnes, L. (1964) Apparatus for rapid and sensitive spectrophotometry. Biochem. J. 91, 161-171. Gill, S. C. & von Hippel, P. H. (1989) Calculation of protein extinction coefficients from amino acid sequence data. Anal. Biochem. 182, 319-326. Golinelli, M. P., Chatelet, C., Duin, E. C., Johnson, M. K., and Meyer, J. (1998) Extensive ligand rearrangements around the [2Fe-2S] cluster of Clostridium pasteurianum ferredoxin. Biochemistry 37, 10429–10437. Gornall, A. G., Bardawill, C. J. & David, M. M. (1949) Determination of serum proteins by means of the biuret reaction. J. Biol. Chem. 177, 751-766. Gray, H. B. & Winkler J. R. (2003) Electrontunneling through proteins. Q. Rev. Biophys. 36, 341-372. Grigorieff, N. (1999). Structure of the respiratory NADH:ubiquinone oxidoreductase (complex I). Curr. Opin. Struct. Biol. 9, 476-483. Guenebaut, V., Schlitt, A., Weiss, H., Leonard, K. & Friedrich, T. (1998) Consistent structure between bacterial and mitochondrial NADH:ubiquinone oxidoreductase (complex I). J. Mol. Biol. 276, 105-112. Hanes, C. S. (1932). Studies on plant amylases: The effect of starch concentration upon the velocity of hydrolysis by the amylase of germinated barley. Biochem. J. 26, 1406-1421. Hirst, J., Carroll, J., Fearnley, I.M., Shannon, R.J. & Walker, J.E. (2003) The nuclear encoded subunits of complex I from bovine heart mitochondria. Biochim. Biophys. Acta 1604, 135150. Horn, A. (2010) Über die Bedeutung der Positionen NuoG W221 und NuoCD F515 für den Elektronentransfer desKomplex I. Diplomarbeit, Albert-Ludwigs-Universität Freiburg. 117 Literatur Hunte, C., Zickermann, V. und Brandt, U. (2010) Functional modules and structural basis of conformational coupling in mitochondrial Complex I. Science 329, 448-451. Jenney, F.E., Verhagen, M. F. J. M., Cui, X. und Adams, M. W. W. (1999) Anaerobic microbes: Oxygen detoxification without Superoxide Dismutase. Science 286, 306-309. Kashani-Poor, N., Zwicker, K., Kerscher, S. und Brandt, U. (2001) A Central Functional Role for the 49-kDa Subunit within the Catalytic Core of Mitochondrial Complex I. J. Biol. Chem. 276, 24082–24087. Kitagawa, M., Ara, T., Arifuzzaman, M., Ioka-Nakamichi, T., Inamoto, E., Toyonaga, H. & Mori, H. (2005) Complete set of ORF clones of Escherichia coli ASKA library (a complete set of E. coli K-12 ORF archive): unique resources for biological research. DNA Res. 12, 291299. Kohlstädt, M. (2009) Funktionelle und strukturelle Untersuchungen zur NADH-Bindung der bakteriellen NADH:Ubichinon Oxidoreduktase. Dissertation, Albert-Ludwigs-Universität Freiburg. Krätzer, C., Welte, C., Dörner, K., Friedrich, T. und Deppenmeier, U. (2010) Characterization of methanoferrodoxin: a novel superoxide reductase. FEBS J., im Druck, online verfügbar. Kung, J. W., Löffler, C., Dörner K., Heintz, D., Gallien, S., Van Dorsselaer, A., Friedrich, T. und Boll, M. (2009) Identification and characterization of the tungsten-containing class of benzoyl-coenzyme A reductases. Proc. Natl. Acad. Sci. U.S.A. 106, 17687–92. Kung, J. W., Baumann, S., von Bergen, M., Müller, M., Hagedoorn, P-L., Hagen, W. R. und Boll, M. (2010) Reversible biological Birch reduction at an extremely low redox potential. J. Am. Chem. Soc. 132, 9850–9856. Kussmaul, L. & Hirst, J. (2006) The mechanism of superoxide production by NADH:ubiquinone oxidoreductase (complex I) from bovine heart mitochondria Proc. Natl. Acad. Sci. U.S.A. 103,7607-7612. 118 Literatur Laemmli, U. K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680-685. Leif, H., Sled, V. D., Ohnishi, T., Weiss, H. & Friedrich, T. (1995). Isolation and characterization of the proton-translocating NADH: ubiquinone oxidoreductase from Escherichia coli. Eur. J. Biochem. 230, 538-548. Lin, Y., Gerfen, G.J., Rousseau, D.L. und Yeh S. R. (2003) Ultrafast microfluidic mixer and freezequenching device. Anal Chem 75, 5381–5386. Lin, M. T. & Beal, M. F. (2006). Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature 443, 787-795. Lu, S., Wiertz, F. G. M., de Vries, S. und Moenne-Loccoz, P. (2005) Resonance Raman characterization of a high-spin six-coordinate iron(III) intermediate in metmyoglobin–azido complex formation trapped by microsecond freeze-hyperquenching (MHQ). J. Raman Spectrosc. 36: 359–362. Mathiesen, C. & Hägerhäll, C. (2003). The 'antiporter module' of respiratory chain complex I includes the MrpC/NuoK subunit - a revision of the modular evolution scheme. FEBS Lett. 549, 7-13. Matsushita, K., Ohnishi, T., und Kaback, H.R. (1987) NADH-Ubiquinone oxidoreductases of Escherichia coli aerobic respiratory chain. Biochemistry 26, 7732-7737. Meyer, J. (2001) Ferredoxins of the third kind FEBS Lett. 509, 1–5. Mitchell, P. & Moyle, J. (1967). Chemiosmotic hypothesis of oxidative phosphorylation. Nature 213, 137-139. Morina, K. (2009) Mutagenese aromatischer Aminosäuren des respiratorischen Komplex I. Diplomarbeit, Albert-Ludwigs-Universität Freiburg. 119 Literatur Moser, C. C., Farid, T. A., Chobot, S. E. & Dutton, P. L. (2006). Electron tunneling chains of mitochondria. Biochim. Biophys. Acta 1757, 1096-1109. Murphy, K.C. (1998) Use of bacteriophage lambda recombination functions to promote gene replacement in Escherichia coli. J. Bacteriol. 180, 2063-2071. Nakamaru-Ogiso, E., Matsuno-Yagi, A., Yoshikawa, S., Yagi, T. & Ohnishi, T. (2008) Ironsulfur cluster N5 is coordinated by an HXXXCXXCXXXXXC motif in the NuoG subunit of Escherichia coli NADH:quinone oxidoreductase (complex I). J. Biol. Chem. 283, 2597925987. Ohnishi, T. (1998). Iron-sulfur clusters/semiquinones in complex I. Biochim. Biophys. Acta 1364, 186-206. Ohnishi, T. & Nakamaru-Ogiso, E. (2008) Were there any “misassignments” among iron– sulfur clusters N4, N5 and N6b in NADH-quinone oxidoreductase (complex I)? Biochim. Biophys. Acta 1777, 703-710. Page, C. C., Moser, C. C., Chen, X. & Dutton, P. L. (1999). Natural engineering principles of electron tunnelling in biological oxidation-reduction. Nature 402, 47-52. Pohl, T., Walter, J., Stolpe, S., Soufo, J.H.D., Grauman, P.L. & Friedrich, T. (2007a) Effects of the deletion of the Escherichia coli frataxin homologue CyaY on the respiratory NADH:ubiquinone oxidoreductase. BMC Biochem. 8, 13. Pohl, T., Bauer, T., Dörner, K., Stolpe, S., Sell, P., Zocher, G. & Friedrich, T. (2007b) Ironsulfur cluster N7 of the NADH:ubiquinone oxidoreductase (complex I) is essential for stability but not involved in electron transfer. Biochemistry 46, 6588-6596. Pohl, T., Uhlmann, M., Kaufenstein, M. & Friedrich, T. (2007c) Lambda Red-mediated mutagenesis and efficient large scale affinity purification of the Escherichia coli NADH:ubiquinone oxidoreductase (complex I). Biochemistry 46, 10694-10702. 120 Literatur Pohl, T. (2008) Lambda-Red vermittelte Mutagenese des nuo-Operons zur Herstellung von Protein-Varianten für die biophysikalische und zellbiologische Charakterisierung der Escherichia coli NADH:Ubichinon Oxidoreduktase. Dissertation, Albert-Ludwigs-Universität Freiburg. Prieur, I., Lunardi, J. und Dupuis, A. (2001) Evidence for a quinone binding site close to the interface between NuoD and NuoB subunits of complex I. Biochim. Biophys. Acta 1504 173178. Puustinen, A., Finel, M., Haltia, T., Gennis, R.B., und Wikström, M. (1991) Properties of the two terminal oxidases of Escherichia coli. Biochemistry 30, 3936-3942. Rasmussen, T., Scheide, D., Brors, B., Kintscher, L., Weiss, H. & Friedrich, T. (2001) Identification of two tetranuclear FeS clusters on the ferredoxin-type subunit of NADH:ubiquinone oxidoreductase (complex I). Biochemistry 40, 6124-6131. Rhein, V., Song, X., Wiesner, A., Ittner, L.M., Baysang, G., Meier, F., Ozmen, L., Bluethmann, H., Drösee, S., Brandt, U., Savaskan, E., Czech, C., Götz J. und Eckert, A. (2009) Amyloid-β and tau synergistically impair the oxidative phosphorylation system in triple transgenic Alzheimer‟s disease mice. Proc. Natl. Acad. Sci. U.S.A. 106, 20057-20062. Ried, J.L., und Collmer, A. (1987) A nptI-sacB-sacR cartridge for constructing directed, unmarked mutations in gram-negative bacteria by marker-exchange-eviction mutagenesis. Gene 57, 239-246. Robards, A. W. and U. B. Steytr. (1985) Low temperature methods in biological electron microscopy. Practical Method in Electron Microscopy, 10, 5–133. Roessler, M. M., King, M. S., Robinson, A. J., Armstrong, F. A., Harmer, J. und Hirst J. (2010) Direct assignment of EPR spectra to structurally defined iron-sulfur clusters in complex I by double electron–electron resonance. Proc. Natl. Acad. Sci. U.S.A. 107, 1930– 1935 121 Literatur Rova, U., Adrait, A., Pötsch, S., Gräslund, A. und Thelander L. (1999) Evidence by mutagenesis that Tyr370 of the mouse Ribonucleotide Reductase R2 protein is the connecting link in the intersubunit radical transfer pathway. J. Biol. Chem. 274, 23746–23751. Sazanov, L. A. & Hinchliffe, P. (2006) Structure of the hydrophilic domain of respiratory complex I from Thermus thermophilus. Science 311, 1430-1436. Schägger, H. & von Jagow, G. (1987) Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal. Biochem. 166, 368-379. Schulte, M. (2009) Über eine mögliche Beteiligung aromatischer Aminosäuren am Elektronentransport in der NADH:Ubichinon Oxidoreduktase. Diplomarbeit, Albert-LudwigsUniversität Freiburg. Screpanti, E. & Hunte, C. (2007) Discontinuous membrane helices in transport proteins and their correlation with function. J. Struct. Biol. 159, 261-267 Seyedsayamdost, M.R., Xie, J., Chan, C.T., Schultz, P.G., und Stubbe, J. (2007) Site-specific insertion of 3-aminotyrosine into subunit α2 of E. coli ribonucleotide reductase: direct evidence for involvement of Y730 and Y731 in radical propagation. J. Am. Chem. Soc. 129, 15060-15071. Shastry, M. C. R., Luck, S. D. und Roder H. (1998) A continuous-flow capillary mixing method to monitor reactions on the microsecond time scale. Biophys. J. 74, 2714–2721. Shih, C., Museth, A. K., Abrahamsson, M., Blanco-Rodriguez, A. M., Di Bilio, A. J., Sudhamsu, J., Crane, B. R., Ronayne, K. L., Towrie, M., Vlcek, A., Jr., Richards, J. H., Winkler, J. R. & Gray, H. B. (2008). Tryptophan-accelerated electron flow through proteins. Science 320, 1760-1762. Sled, Y. D., Rudnitzky, N.I., Hatefi, Y. und Ohnishi T. (1994) Thermodynamic analysis of flavin in mitochondrial NADH:ubiquinone oxidoreductase (Complex I) Biochemistry 33, 10069-10075. 122 Literatur Spatzal, T., Pohl, T., Aksoyoglu, M., Schleicher, E., Rostas, A. M., Lay, H., Glessner, U., Boudon, C., Hellwig, P., Weber, S. und Friedrich, T. (2010) Spin labeling of the Escherichia coli NADH ubiquinone oxidoreductase (complex I). Biochim Biophys Acta. 1797, 1894-1900. Spehr, V. (1996). Spezifische Mutagenese der NADH:Ubichinon Oxidoreduktase in Escherichia coli. Dissertation, Heinrich-Heine-Universität Düsselorf. Stemmler, T. L., Lesuisse, E., Pain, D. und Dancis, A. (2010) Frataxin and Mitochondrial FeS Cluster Biogenesis. J. Biol. Chem. 285, 26737-26743. Stolpe, S. (2007) Über den Mechanismus der NADH:Ubichinon Oxidoreduktase aus Escherichia coli. Dissertation, Albert-Ludwigs-Universität Freiburg. Stubbe, J., Nocera, D. G., Yee, C. S., und Chang, M. C. Y. (2003) Radical Initiation in the class I ribonucleotide reductase: Long-range proton-coupled electron transfer? Chem. Rev. 103, 2167-2201. Studier, F. W. & Moffatt, B. A. (1986) Use of bacteriophage T7 RNA polymerase to direct selective high-level expression of cloned genes. J. Mol. Biol. 189, 113-130. Studier, F. W., (2005). Protein production by auto-induction in high-density shaking cultures. Science 41, 207-234. Tanaka, M., Matsuura, K., Yoshioka, S., Takahashi, S., Ishimori, K., Hori, H. und Morishima, I. (2003) Activation of hydrogen peroxide in horseradish peroxidase occurs within ~200 ms observed by a new freeze-quench device. Biophys. J. 84, 1998–2004. Tersteegen, A. & Hedderich, R. (1999) Methanobacterium thermoautotrophicum encodes two multisubunit membrane-bound [NiFe] hydrogenases. Transcription of the operons and sequence analysis of the deduced proteins. Eur. J. Biochem. 264, 930-943. Uhlmann, M. & Friedrich, T. (2005). EPR signals assigned to Fe/S cluster N1c of the Escherichia coli NADH:ubiquinone oxidoreductase (complex I) derive from cluster N1a. Biochemistry 44, 1653-1658. 123 Literatur Unden, G., Achebach, S., Holighaus, G., Tran, H. G., Wackwitz, B. & Zeuner, Y. (2002). Control of FNR function of Escherichia coli by O2 and reducing conditions. J. Mol. Microbiol. Biotechnol. 4, 263-268. Verkhovskaya, M. L., Belevich, N., Euro, L., Wikström, M. & Verkhovsky, M. I. (2008). Real-time electron transfer in respiratory complex I. Proc. Natl. Acad. Sci. U. S. A 105, 37633767. Walker, J.E. (1992) The NADH:ubiquinone oxidoreductase (complex I) of respiratory chains. Q. Rev. Biophys. 25, 253-324. Wallace, B. J. & Young, I. G. (1977) Role of quinones in electron transport to oxygen and - - nitrate in Escherichia coli. Studies with a ubiA menA double quinone mutant. Biochim. Biophys. Acta 461, 84-100. Weidner, U., Geier, S., Ptock, A., Friedrich, T., Leif, H. & Weiss, H. (1993). The gene locus of the proton-translocating NADH:ubiquinone oxidoreductase in Escherichia coli. Organization of the 14 genes and relationship between the derived proteins and subunits of mitochondrial complex I. J. Mol. Biol. 233, 109-122. Weiss, R. F. (1970) Helium Isotope Effect in Solution in Water and Seawater. Science 168, 247-248. Weiss, H., Friedrich, T., Hofhaus, G. & Preis, D. (1991) The respiratory-chain NADH dehydrogenase (complex I) of mitochondria. Eur. J. Biochem. 197, 563-576. Werth, M.T., Cecchini, G., Manodori, A., Ackrell, B.A.C., Schröder, I., Gunsalus, R.P. und Johnson, M.K. (1990) Site-directed mutagenesis of conserved cysteine residues in Escherichia coli fumarate reductase: Modification of the spectroscopic and electrochemical properties of the [2Fe-2S] cluster. Proc. Natl. Acad. Sci. U.S.A. 87, 8965-8969. Wiertz, F.G.M., Richter O-M. H., Cherepanov A.V., MacMillan, F., Ludwig, B., und de Vries, S. (2004) An oxo-ferryl tryptophan radical catalytic intermediate in cytochrome c and quinol oxidases trapped by microsecond freeze-hyperquenching (MHQ). FEBS Lett. 575, 127130. 124 Literatur Wischgoll, S., Heintz, D., Peters, F., Erxleben, A., Sarnighausen, E., Reski, R., Van Dorsselaer, A. und Boll, M. (2005) Gene clusters involved in anaerobic benzoate degradation of Geobacter metallireducens. Mol. Microbiol. 58, 1238–1252. Wittekindt, C., Schwarz, M., Friedrich, T. & Koslowski, T. (2009). Aromatic amino acids as stepping stones in charge transfer in respiratory complex I: an unsual mechanism deduced from atomistic theory and bioinformatics. J. Am. Chem. Soc. 131, 8134-8140. Yakovlev, G., Reda, T., and Hirst J. (2007) Reevaluating the relationship between EPR spectra and enzyme structure for the iron–sulfur clusters in NADH:quinone oxidoreductase. Proc. Natl. Acad. Sci. U.S.A. 104, 12720–12725. Yano, T., Sled, V. D., Ohnishi, T. & Yagi, T. (1994) Identification of amino acid residues associated with the [2Fe-2S] cluster of the 25 kDa (NQO2) subunit of the proton-translocating NADH-quinone oxidoreductase of Paracoccus denitrificans. FEBS Lett. 354, 160-164. Yano, T., Sklar, J., Nakamaru-Ogiso, E., Takahashi, Y., Yagi, T. & Ohnishi, T. (2003). Characterization of cluster N5 as a fast-relaxing [4Fe-4S] cluster in the Nqo3 subunit of the proton-translocating NADH-ubiquinone oxidoreductase from Paracoccus denitrificans. J. Biol. Chem. 278, 15514-15522. Yao, C. (2010) Die Rolle des Eisen-Schwefel-Zentrums N1a bei der Komplex I-vermittelten Bildung reaktiver Sauerstoffspezies. Diplomarbeit, Albert-Ludwigs-Universität Freiburg. Yeh, A.P., Chatelet, C., Soltis, S.M., Kuhn, P., Meyer, J. and Rees, D.C. (2000) Structure of a thioredoxin-like [2Fe-2S] ferredoxin from Aquifex aeolicus. J. Mol. Biol. 300, 587-595. Yeh, A. P., Ambroggio, X. I., Andrade S., L., A., Einsle, O., Chatelet, C., Meyer, C. und Rees, D. C. (2002) High resolution crystal structures of the wild type and Cys-55 → Ser and Cys-59 → Ser variants of the thioredoxin-like [2Fe-2S] ferredoxin from Aquifex aeolicus. J. Biol. Chem. 277, 34499–34507. 125 Danksagung Bei meinem Doktorvater Herrn Prof. Dr. Friedrich bedanke ich mich für die Bereitstellung des interessanten Themas und die vielen Möglichkeiten bei der experimentellen Arbeit auch über das eigene Labor hinaus. Ein herzliches Dankeschön für die angenehme Arbeitsatmosphäre und die geduldige Einführung in die ESR-Spektroskopie! Bei Herrn Prof. Dr. Simon de Vries bedanke ich mich für die freundliche und hilfreiche Betreuung im Labor in Delft und das Nutzen der MHQ-Apparatur. Bei Herrn Marc Strampraad für die wertvolle Hilfe beim Sprayen. Mein Dank gilt allen Kooperationspartnern Prof. Dr. Matthias Boll, Prof. Dr. Uwe Deppenmeier, Prof. Dr. Georg Fuchs, Nasser Gad„on, Christian Krätzer und Johannes Kung. Bei Frau Helga Lay bedanke ich mich für die stets verlässliche Hilfe bei den vielen Alltagsproblemen im Labor und Herrn Helmut Hamacher für die technische Unterstützung, beide hatten immer eine Lösung parat. Bei Frau Dörte Thiel und Herrn Antonio Espin möchte ich für die unermüdliche Hilfe nicht nur bei der langwierigen Proteinpräparation bedanken. An Frau Linda Williams und Frau Angelika Massler vielen Dank für die freundliche Unterstützung bei allerlei bürokratischen Dingen. Ein herzliches Dank auch an die im Rahmen dieser Arbeit betreuten Diplomanden Thomas Bayer, Elvira Friedrich, Annemarie Horn, Klaudia Morina und Marius Schulte sowie die Bachelorstudentin Isabella Straub und die studentische Hilfskraft Vanda Barros Pires. Gedankt sei auch den vielen während dieser Arbeit betreuten Mitarbeiterpraktikanten. Meinem Arbeitskreis danke ich für die freundschaftliche Atmosphäre und die schöne Zeit nicht nur im Labor! Meinen langjährigen Mitstreitern Heiko Erhardt, Markus Kohlstädt, Thomas Pohl und Daniel Schneider danke ich für die tatkräftige Unterstützung und stets heiteren Stunden bei der Arbeit. Zwar keine langjährigen, aber natürlich ebenso angenehme Mitdoktoranden waren Udo Glessner, Ramona Labatzke, Klaudia Morina, Marius Schulte, Stefan Steimle und Marta Vranas. Meinen lieben Eltern kann ich nicht genug danken für ihre ausnahmslose Unterstützung in allen Lebenslagen! Ich danke von Herzen Nele, Fabian, Annika, Leila, Christine und vor allem für die Unterstützung im Endspurt Thomas.