Thematischer Schwerpunkt 1: Stoffwechsel und seine
Werbung

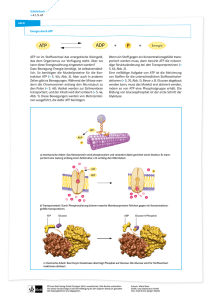
Thematischer Schwerpunkt 1: Stoffwechsel und seine Regulation 1. Zellatmung und Gärung Bau und Funktion von Mitochondrien Kompartimentierung: Einteilung des Mitochondriums in zwei verschiedene Reaktionsräume, einmal zwischen innerer und äußerer Membran (nicht-plasmatisches Kompartiment) und Matrix (plasmatisches Kompartiment), die durch die innere Membran begrenzt ist. ATP-Bildung an der der Matrix zugewandten inneren Membran StrukturDie Struktur der Mitochondrien durch die Kompartimentierung gibt Funktionsbeziehung: zugleich die Funktion vor. Durch die gefalteten Membranen erfolgt zudem eine Oberflächenvergrößerung. Die innere Membran bildet dabei Einstülpungen in die Matrix; diese werden als Cristae bezeichnet. Prinzipielle Reaktionen in Glykolyse, Citratzyklus und Endoxidation a) Anaerober Abbau Glykolyse: findet im Cytoplasma statt, Umwandlung eines C6-Körpers von Glucose: (Glucose) in zwei C3-Körper (Pyruvat) Milchsäuregärung: Pyruvat übernimmt die 2 H+, so dass wieder 2 NAD+ entstehen. Dabei wird Pyruvat zu Milchsäure (Lactat), wodurch das Milieu der Zelle immer saurer wird. Alkoholische Gärung: Hierbei entsteht statt Lactat Ethanol, wird von Hefen betrieben b) Aerober Abbau oxidative Decarboxylierung: Bindeglied zwischen Glykolyse und von Glucose: Citratzyklus, erfolgt in der Mitochondrien-Matrix. Der C3-Körper Pyruvat wird in einem Multienzymkomplex in den C2-Körper Acetyl-CoA umgewandelt Citratzyklus: Der C2-Körper Acetyl-CoA wird in den C6-Körper Citrat umgewandelt. Dieser wird zu einem C5-Körper oxidiert, danach zu einem C4Körper. Nach einigen weiteren Umwandlungen entsteht am Ende ein C4Körper, der sich wieder mit Acetyl-CoA verknüpft. Dies ist ein sich wiederholender Kreisprozess Der Citratzyklus steht mit vielen Stoffwechselwegen sowohl direkt als auch indirekt in Verbindung. So gelangen z.B. Fettsäuren bei ihrer Zerlegung über die Zwischenstufe Acetyl-CoA in den Citratzyklus. Elektronentransport Findet in der inneren Mitochondrienmembran statt. Dabei werden der Atmungskette Elektronen von NADH+H+ und FADH2 über mehrere Enzymkomplexe über Redox-Systeme: transportiert. Diese wechseln zwischen reduziertem und oxidiertem Zustand, um diesen Transport zu ermöglichen. Am Ende werden die Elektronen auf Sauerstoff übertragen, der dann zu Wasser reagiert. Allgemeines Bei der Redox-Reaktion werden Elektronen auf einen Stoff übertragen, Redox-Prinzip: der dadurch reduziert wird. Dieser Stoff kann die Elektronen auch wieder abgeben, er wird oxidiert. Laufen Reduktion und Oxidation parallel voneinander ab, so findet eine Redox-Reaktion statt. Dabei werden die Elektronen zum elektronegativeren Atom übertragen. Da dieses Atom die Elektronen näher an sich heran zieht, verlieren diese potentielle Energie, die in anderen Prozessen verwendet werden kann. chemiosmotische Durch die bei dem Transport der Elektronen frei werdende Energie werden Bildung von ATP Protonen in den perimitochondrialen Raum (zwischen innerer und äußerer durch Membran) transportiert. Diese Protonen fließen dann wieder entlang des Protonengradient: Gradienten zurück in die Matrix. Dies geschieht am Enzymkomplex ATPSynthase, der die in den Protonen gespeicherte Energie nutzt um aus ADP und P das ATP zu bilden. Dabei entstehen aus einem Mol NADH+H+ 3 Mol ATP und aus einem Mol FADH2 2 Mol ATP. Regulation von Stoff- Die Reaktion eines Substrats an einem Enzym kann durch zweierlei Wege wechselvorgängen: verhindert und dadurch reguliert werden. Bei der kompetitiven Hemmung allosterische und konkurriert ein Inhibitor mit dem Substrat um das aktive Zentrum eines kompetitive Enzymregulation: Enzyms. Bei der allosterischen Hemmung bindet der Inhibitor an das allosterische Zentrum des Enzyms, so dass das aktive Zentrum seine Konformation ändert. Das Substrat kann dann nicht mehr an das aktive Zentrum binden. Sauerstoffaufnahme, Die Sauerstoffaufnahme erfolgt über die Atmungsorgane (z.B. Lunge). -transport und In der Lunge, in der ein relativ hoher Partialdruck herrscht, wird der -abgabe: Sauerstoff auf das Hämoglobin übertragen, welches in den Erythrozyten enthalten ist. Der Transport erfolgt über das Blut in alle möglichen Gewebe. Der Partialdruck in den Geweben ist geringer als in der Lunge. Da Hämoglobin Sauerstoff bei sinkendem Partialdruck weniger gut bindet, gibt es diesen im Gewebe wieder ab. Die Aufnahmefähigkeit des Hämoglobins hängt ebenfalls von der Säurekonzentration im Blut ab. Wenn viel CO2 im Gewebe vorhanden ist löst sich der Sauerstoff vom Hämoglobin. Dies wird auch als Bohr-Effekt bezeichnet. Sauerstoffbindungs- Myoglobin besitzt bei niedrigen Partialdrücken eine höhere O2-Sättigung kurven von als Hämoglobin. Myoglobin kommt in der Muskulatur vor. Durch die Hämoglobin und höhere Sättigung wird der komplette Sauerstoff vom Hämoglobin auf das Myoglobin: Myoglobin übertragen, so dass die Muskeln bestmöglich mit Sauerstoff versorgt werden Stoff- und EnergieGlykolyse: bilanz des anaeroben Glucose + 2 NAD+ + 2 ADP + 2 P Æ 2 Pyruvat + 2 NADH+ 2 H+ + 2 ATP und aeroben Abbaus: oxidative Decarboxylierung: 2 Pyruvat + 2 NAD+ + 2 CoA-SH + 2 H+ Æ 2 Acetyl-CoA + 2 NADH + 2 H+ + 2 CO2 Citratzyklus: 2 Acetyl-CoA + 6 NAD+ + 2 FAD + 6 H2O + 2 GDP + 2 P + 2 H+ Æ 4 CO2 + 6 NADH + 6 H+ + 2 FADH2 + 2 CoA-SH + 2 H+ + 2 GTP Atmungskette: 10 NADH + 10 H+ + 2 FADH2 + 6 O2 + 34 ADP + 34 P Æ 10 NAD+ + 2 FAD + 34 ATP + 12 H2O Bilanz: 38 ATP – 2 ATP für Transportkosten beim Elektronentransport = 36 ATP, beim anaeroben Abbau 4 ATP (Glykolyse + Gärung) Wirkungsgrad: 1 Mol ATP speichert 30,5 kJ. 36 * 30,5 = 1098 kJ/Mol Knallgasreaktion (Glucose + Sauerstoff): 2870 kJ/Mol Wirkungsgrad: 38%, Rest wird als Abwärme frei (u.a. zur Aufrechterhaltung einer konstanten Körpertemperatur) 2. Spezielle Aspekte des Energieumsatzes Energetische Koppelung ATP als Bindeglied Wird vom ATP eine Phosphatgruppe abgespalten, so wird Energie frei zwischen Energie (30,5 kJ/Mol). Die Reaktion ist exergonisch. Die Bindung der freisetzenden und Phosphatgruppen ist relativ schwach, so dass ADP und P stabiler sind als Energie benötigenden ATP. Die Energie aus der Spaltung kann mit Hilfe von Enzymen an Prozessen: endergonische Prozesse übertragen werden. Um ATP wieder zu synthetisieren wird Energie benötigt, um die Phosphatgruppe wieder anzulagern. Energieentwertung Freisetzung von Abwärme als typische Begleiterscheinung aller Stoffwechselprozesse: Temperaturregulation bei einem Säugetier: Der Wirkungsgrad der Zellatmung beträgt 38%, der Rest der Energie wird als Wärme frei. Bei allen Stoffwechselprozessen wird nicht die gesamte verfügbare Energie für weitere Prozesse als verwertbare Energie gespeichert. Ein Rest wird immer als Wärme freigesetzt. Säugetiere sind homoiotherme / endotherme (gleichwarme) Tiere, d.h. sie halten eine konstante Körpertemperatur aufrecht. Dies geschieht durch die Stoffwechselprozesse, bei denen Abwärme frei wird. Der erhöhte Energie- haushalt hat zur Folge, dass mehr Nährstoffe aufgenommen werden müssen. Übersteigt die Temperatur einen bestimmten Wert, so weiten sich die Blutgefäße damit sich die Wärme auf einen größeren Bereich verteilt. Die größere Fläche sorgt für eine Abkühlung. Zudem kann über Schweißdrüsen Wärme abgegeben werden. Im Gegenzug können die Blutgefäße bei Unterschreiten einer Temperatur verengt werden, so dass weniger Wärme aufgrund einer kleineren Fläche verloren geht. Zudem kommt es zur Kontraktion der Muskulatur, dem Zittern. Bei der entstehenden Gänsehaut kontrahiert die Hautmuskulatur, wodurch es zu einem Aufstellen der Haare kommt. Es lassen sich zudem zwei Regeln formulieren: Bergmann’sche Regel: Bei Homoiothermen nimmt die Größe nah verwandter Arten oder Rassen von den warmen Zonen zu den Polen hin zu. Bei einer Größenzunahme steigt das Volumen schneller als die Oberfläche, wodurch die Wärmeabgabe verringert wird. Allen’sche Regel: Bei Homoiothermen sind Körperanhänge (z.B. Ohren) nahe verwandter Arten in kälteren Gebieten kleiner. Bei Körperanhängen geht Wärme verloren, da die Oberfläche im Vergleich zum Volumen sehr groß ist. 3. Gene und Merkmalsbildung Ursachen für die Merkmalsausprägung Erbliche Stoffwechselerkrankungen Genwirkkette: Innerhalb einer Genwirkkette arbeiten viele Enzyme zusammen, die jeweils von einem oder mehreren entsprechenden Genen codiert werden. Jedes Glied in der Kette ist dabei auf das Produkt des vorangegangenen Gliedes angewiesen und stellt wiederum selbst ein Produkt her, das für nachfolgende Glieder benötigt wird. Fällt aufgrund eines Gendefekts ein Teil der Kette aus, kann es zu unterschiedlichen Krankheitsbildern kommen. (Bsp.: Phenylketonurie, keine Umsetzung von Phenylalanin) Polygenie: Mehrere Gene haben Einfluss auf die Ausprägung des Merkmals. Dabei hat jedes Gen eine Auswirkung auf den Phänotyp. (Bsp.: Xeroderma pigmentosum) Polyphänie: Ein einzelnes mutiertes Gen hat Auswirkung auf viele Merkmale, ein Beispiel hierfür ist die Phenylketonurie, bei der ein einzelnes Gen, das ein entsprechendes Enzym codiert, durch eine Veränderung alle weiteren Stoffwechselschritte stört. Genetische Variabilität multiple Allelie: Liegen für ein Gen mehr als zwei verschiedene Allele (Zustände) vor, so spricht man von multipler Allelie. Ein Beispiel hierfür ist der Erbgang der Blutgruppen, bei der es neben einer Konstellation aus dominantem und rezessivem Allel (A0) auch zu kodominanten Allelen kommen kann (AB). Von den drei möglichen Allen besitzt jeder Mensch nur zwei. Mutagene Überblick zu Mutagenen und deren Wirkung: chemische Stoffe: Teerstoffe: - wirken krebserregend - schieben sich zwischen Nucleotide - zusätzliche, vorgetäuschte Base, an der eine andere angelagert wird - der DNA-Strang wird verlängert Bromuracil: - Ähnlichkeit mit normalen Basen (Basenanaloga) - vertritt diese oder bildet Basenpaare - Thymin wird durch Bromuracil ersetzt, Adenin paart mit Bromuracil. Dieses ist instabil und wird durch Guanin ersetzt Mutationstypen: DNA-Schäden durch UV-Strahlung und deren Reparatur: - das Basenpaar A-T wird durch G-C ersetzt - bei der Replikation können so Mutationen entstehen salpetrige Säure: - Cytosin wird zu Uracil (Desaminierung) - Uracil ist komplementär zu Adenin - das Basenpaar C-G wird so zu U-A - Replikationsfehler, Ablesefehler, Protein kann wirkungslos werden Akridinfarbstoffe: - schieben sich zwischen Nucleotide - Veränderung der Basensequenz physikalische Einflüsse: kurzwellige UV-Strahlen: - Verknüpfung benachbarter Thymin-Basen zu Thymindimeren - es erfolgt keine Paarung mit Adenin, die Informationen werden ungenau abgelesen Radioaktive Strahlung / Röntgenstrahlung: - bilden in den Zellen reaktionsfreudige Radikale - diese gehen chem. Reaktionen mit der DNA ein - evtl. Brüche im Strang, dadurch Basentausch oder der Ausfall eines Nucleotids Genmutationen: - Basenaustauschmutation: Austausch einer Base, wodurch die Möglichkeit besteht, dass ein Protein nicht mehr funktionsfähig ist (Bsp.: Sichelzellanämie) - Rastermutationen: Basen werden eingeschoben oder weggelassen, dadurch wird der gesamte folgende Strang verändert, das Protein oder Enzym somit wirkungslos Chromosomenmutation: - Duplikation (Verdopplung eines Abschnitts [ABCDCDE]) - Deletion (Endstück-, Zwischenstückverlust [BCDE], [ABE]) - Inversion (Drehung [ABDCE]) - Translokation (Austausch nichthomologer Abschnitte [XBCD]) Genommutation: Veränderung der Gesamtzahl der Chromosomen im Zellkern (Bsp.: Trisomie 21, Polyploidie beim Weizen) Durch UV-Strahlung kann es zur Bildung von Thymin-Dimeren kommen, so dass keine Paarung mit Adenin erfolgt und die Informationen ungenau abgelesen werden. Diese Schäden können durch die NucleotidExcisionsreparatur (NER) wieder behoben werden: Der defekte Abschnitt wird durch die Endonuclease aufgeschnitten, die Exonuclease entfernt darauf hin den Abschnitt. Anhand des Komplementärstranges synthetisiert die DNA-Polymerase I wieder den korrekten Strang. Abschließend werden der reparierte und normale Strang wieder durch die Ligase verknüpft. 4. Regulation der Genaktivität Induktion, Repression Operonmodell im Zusammenhang mit Stoffwechselaktivitäten von Bakterien: Ein Operon wird in drei Abschnitte eingeteilt: Den Promotor, die Bindungsstelle für die RNA-Polymerase, den Operator und die Strukturgene, die Enzyme codieren. Der Operator kann mit einem angelagerten Repressor das Wandern der RNA-Polymerase und somit die weitere Synthese eines bestimmten Enzyms verhindern. Bei der Enzyminduktion ist der Operator von einem Repressor besetzt, die Enzymsynthese ist verhindert. Wird nun ein Stoff, z.B. Lactose zugeführt, bindet dieser an eine spezifische Bindungsstelle am Repressor, der dadurch inaktiv wird. Die RNA-Polymerase kann an den Strukturgenen entlang wandern und Enzyme synthetisieren. Dies erfolgt häufig dann, wenn ein bestimmter Stoff, z.B. Lactose abgebaut werden muss. Die Induktion bestimmter Strukturgene ist demnach spezifisch für das vorhandene Substrat, man spricht von der Substratinduktion. Bei der Enzymrepression ist der Repressor in einem inaktiven Zustand und durch die Strukturgene werden Enzyme synthetisiert, die z.B. Tryptophan bilden. Sollte Tryptophan allerdings vorhanden sein, muss das Ablesen der Strukturgene verhindert werden. Tryptophan bindet dann an die spezifische Bindungsstelle am Repressor, wodurch dieser aktiviert wird. Dadurch wird die Enzymsynthese unterbrochen. Das Endprodukt Tryptophan unterbindet somit seine eigene Synthese, daher spricht man auch von der Endproduktrepression. 5. Gentechnik Isolierung und Veränderung genetischer Strukturen nach Plan Vervielfältigung von Die Polymerase-Kettenreaktion ermöglicht es, geringe DNA-Mengen in DNA-Abschnitten kurzer Zeit zu vervielfältigen. Das Prinzip ist dass der DNA-Replikation. durch PCR: Der Doppelstrang mit dem Abschnitt, der die benötigten Informationen enthält, wird auf 94° erhitzt, damit die DNA denaturiert, d.h. sich in beide Einzelstränge aufspaltet. Dann erfolgt eine Abkühlung auf 50°, so dass synthetisierte Primer an die Einzelstränge binden können. Die Primer sind dabei rückläufig komplementär zu dem Abschnitt, der kopiert werden soll. Daher muss für die Primer-Herstellung die Basensequenz an den Enden bekannt sein. Dann kann bei 72° die Taq-Polymerase die Synthese des Komplementärstranges an den Primern beginnen. Dieser Prozess wird nun wiederholt, bis eine ausreichende Menge DNA vervielfältigt wurde. Verfahrensschritte Die DNA-Sequenz für das Insulin-Gen ist bekannt, daher kann das Gen der genetischen synthetisiert werden. Dabei wird anhand des codogenen Stranges die Herstellung von mRNA gebildet, an der das Insulin-Gen nun synthetisiert werden kann. Insulin: Zudem wird das Gen an beiden Enden noch mit einem Restriktionsenzym geschnitten. Dies wird auch bei dem Plasmid von Bakterien durchgeführt, die eine Ampicillin und Tetracyclin-Resistenz besitzen. Hierbei wird das Gen für die Tet-Resistenz mit dem gleichen Restriktionsenzym geschnitten, so dass „sticky-ends“ entstehen. Durch ein Ligase-Enzym werden nun beide Stränge miteinander verbunden, so dass das Insulin-Gen in das Bakterienplasmid eingebaut wird. Es sind jedoch auch Plasmide ohne das eingebaute Gen und Insulin-Gene, die nicht eingebaut worden sind, vorhanden. Die Plasmidringe werden nun wieder in Bakterien eingebaut und diese vermehrt. Um nun die Bakterien mit dem Insulin-Gen zu finden erfolgt das „screening“. Dabei werden die Bakterien auf Nährböden mit Ampicillin oder Tetracyclin gegeben. Auf den Ampicillin-Nährböden wachsen die Bakterien, die das unveränderte Plasmid und das mit dem Insulin-Gen eingebaut haben. Das nicht eingebaute Insulin-Gen vermehrt sich nicht. Auf den Tetracyclin-Nährböden wachsen nur die Bakterien mit dem unveränderten Plasmid, da die Tet-Resistenz bei den anderen durch das InsulinGen zerstört ist. Also enthalten die Bakterien, die auf dem ApicillinNährboden überleben und auf dem Tetracyclin-Nährboden absterben das Insulin-Gen. Anhand des Gens wird nun in den Bakterien mRNA gebildet, die an den Ribosomen das fertige Insulin synthetisiert, das nun isoliert und als Medikament verwendet werden kann. Herstellung und Gesonden werden spezifisch für ein Gen hergestellt, daher muss die Anwendungsbereiche Basensequenz des Gens bekannt sein, damit eine komplementäre Sonde von Gensonden: hergestellt werden kann. Mithilfe einer Sonde, die radioaktiv markiert wird, lassen sich z.B. gezielt Gene innerhalb einer Genbibliothek finden. Bei der Insulinherstellung kann eine Sonde verwendet werden, um die Plasmide zu finden, die das Insulin-Gen eingebaut haben. Thematischer Schwerpunkt 2: Informationsverarbeitung 1. Hormonelle Regulation Kontrolle der Hormonausschüttung Hierarchie der Allen hormonellen Vorgängen vorgeschaltet steht der Hypothalamus. HormondrüsenDieser setzt Hormone frei, die ihm untergestellte Hormondrüsen, z.B. die funktionen im Hypophyse stimulieren. Diese kann dann wieder untergeordnete HormonÜberblick: drüsen wie die Schilddrüse anregen. Zudem können die Produkte auch jeweils über eine negative Rückkopplung die übergeordneten Hormondrüsen hemmen. So kann z.B. das Hormon TSH, welches von der Hypophyse gebildet wird, eine hemmende Wirkung auf den Hypothalamus haben. Exemplarische Behandlung am Beispiel der Stressphysiologie Allgemeines System, welches bei längerfristigen Beeinträchtigungen einsetzt. AnpassungsHypothalamus Æ Neurosekretion von CRF Æ Stimulation der Hypophyse Æ Syndrom (AAS): vermehrte Bildung von ATCH Æ Ausschüttung von Glucocorticoiden: Unterstützung der Adrenalinwirkung Steigerung des Fettsäuregehaltes im Blut Abbau von Muskelproteinen Abschwächung der Proteinbiosynthese Æ Unterdrückung von Entzündungsprozessen und Abwehrfunktionen Der Organismus soll möglichst lange in einer „Stand-by-Position“ gehalten werden, Herz-Kreislaufsystem in erhöhter Bereitschaft, übriger Stoffwechsel gedrosselt. Fight-or-FlightSetzt bei kurzfristiger Einwirkung von Stressoren ein. Syndrom (FFS): Hypothalamus aktiviert Sympathicus Æ Anregung des Nebennierenmark Æ Ausschüttung von Adrenalin und Noradrenalin Æ FFS Æ Steigerung von: Herz-Kreislauf-Tätigkeit Æ erhöhten O2 und Glucose-Bedarf decken Durchblutung von Gehirn und Skelettmuskulatur Mobilisierung von Glykogenreserven Unwichtige Körperfunktionen wie Verdauung oder Sexualfunktionen werden reduziert. Regenerierung durch vom Parasympathicus vermittelte Erholungsphase. Das FFS wird zur blitzschnellen Reaktion bei Gefahren benötigt neuronale und Das Vegetative Nervensystem lässt sich in das sympathische und das hormonelle parasympathische NS unterteilen. Die Wirkungen sind anhand des FFS Verschränkung: bekannt. Somit gibt eine neuronale Erregung den Ausschlag für die Ausschüttung von Hormonen, so z.B. im Nebennierenmark hormonelle Regulation Der Blutzuckergehalt im Blut liegt normalerweise bei einem konstantem des KohlenhydratSollwert. Durch Nahrungsaufnahme wird dieser Wert jedoch überschritten stoffwechsels: und muss wieder abgesenkt werden. Hierfür wird in den β-Zellen innerhalb der Bauchspeicheldrüse Insulin freigesetzt. Insulin fördert die Aufnahme von Glucose durch Zellmembranen, stimuliert die Glykogensynthese in Leber- und Muskelzellen, die Energiegewinnung durch Glucoseabbau und die Synthese von Fetten und Eiweißen aus Glucose. Zudem hemmt es den Abbau von Glykogen. Der Blutzuckerspiegel wird daher durch Bildung von Glykogen und dessen Speicherung und durch erhöhten Verbrauch abgesenkt. Körperliche Tätigkeiten und der Grundumsatz jedoch verbrauchen Glucose und senken somit den Blutzuckerspiegel. Es wird kein Insulin mehr abgegeben, so dass die hemmende Wirkung auf das Glukagon, welches in den α-Zellen der Bauchspeicheldrüse gebildet wird, verringert wird. Glukagon fördert den Glykogenabbau in der Leber und somit die Glucoseneubildung. Eiweiße und Fette werden zur Glucosesynthese abgebaut. Der Blutzuckerspiegel steigt wieder. Zelluläre Wirkungsweise von Hormonen Molekulare Wirkmechanismen von: Steroidhormonen: Steroidhormone sind lipidlöslich, d.h. sie können die Zellmembran durchdringen. Im Inneren der Zelle binden sie an einen spezifischen Rezeptor, dessen Konfirmation sie ändern, so dass ein Hormon-Protein-Komplex entsteht. Dieser kann in den Zellkern diffundieren und bestimmte Gene aktivieren. Proteohormone: Proteohormone (auch Peptidhormone) können nicht durch die Zellmembran in die Zelle eindringen. Sie binden daher an einen Rezeptor an der Außenseite der Membran. Es entsteht ein Hormon-Rezeptor-Komplex, der ein an der Innenseite befindliches Enzym, z.B. Adenylatcyclase aktiviert. Dieses setzt ATP zu cAMP um, welches an ein bestimmtes Enzym bindet und dieses aktiviert. Das Enzym bewirkt dann die besondere Zellantwort, so z.B. den Glykogenabbau in Leberzellen. Signalverstärkung: Ausgehend von einem einzelnen Rezeptor werden viele Folgeprozesse in Gang gesetzt, die sich in jedem Schritt verstärken. Z.B. aktiviert ein Rezeptor ca. 10 Moleküle eines G-Proteins, welche jeweils ein cAMPbildendes Enzym aktivieren. Jedes Enzym bildet nun ca. 100 cAMPMoleküle, von denen wiederum jedes ein Molekül der Proteinkinase aktiviert. Jedes Proteinkinasemolekül aktiviert ca. 100 Moleküle eines Enzymproteins, von denen wieder jeweils eins ca. 100 Substratmoleküle umsetzt. Die Verstärkung ist in diesem Fall 10*100*100*100=107fach. 2. Reizaufnahme, Erregungsbildung und –weiterleitung Bau von Sinnes- und Nervenzellen Nervenzelle und Sekundäre Sinneszellen (Bsp. Geschmackssinneszellen) menschliche besitzen kein Axon, AP wird an der nachgeschalteten Nervenzelle gebildet, Lichtsinneszelle: Informationsübertragung auf elektrischem Wege Primäre Sinneszelle (Bsp. Geruchs-, Lichtsinneszellen) Rezeptorpotenzial breitet sich bis zum Axonhügel aus. Wird dort der Schwellenwert erreicht, werden AP gebildet, die sich über das Axon ausbreiten Sinnesnervenzelle Die Dendritenregion geht ins Axon über, der Zellkörper spielt keine Rolle. Die AP werden bereits am Dendriten erzeugt Struktur- und Die Einteilung der Lichtsinneszelle in Innen- und Außensegment ermöglicht Funktionsbeziehung: es, unterschiedliche Funktionen durchzuführen. So sind im Außensegment übereinander geschichtete Membranzisterne (Discs) zu finden. Bei der Nervenzelle erfolgt ebenfalls eine Einteilung, das Soma verrechnet die Informationen und erhält die Zelle am Leben, die Dendriten dienen zur Informationsaufnahme und sind in viele Richtungen ausgeprägt, während das Axon eine gezielte entfernte Verbindung für Informationsweiterleitung darstellt. Umwandlung von Reiz in Erregung (Rezeptorpotenzial) Adäquater Reiz: Reiz mit einer für das Sinnesorgan passenden Energieform (Auge Æ Licht) Schwellenwert: gibt an, wie hoch der elektrische Impuls in der Zelle sein muss, damit eine Reaktion in der Zelle stattfindet. Reize, die unter dem Schwellenwert liegen, lösen keine Aktionspotenziale aus. Wird es jedoch ausgelöst, ist es immer gleich stark, die Amplitude ist gleich hoch (Alles-oder-NichtsPrinzip) Codierung: Durch die Frequenz der Aktionspotenzial kann trotz des Alles-oder-NichtsPrinzips die Intensität eines Reizes weitergegeben werden. Je stärker der Reiz ist, desto höher ist die Anzahl der Aktionspotenziale pro Zeiteinheit (Frequenz). Es gibt zudem drei verschiedene Typen an Rezeptorzellen: Rezeptorpotenzial: Molekulare Grundlagen der Entstehung des Rezeptorpotenzials am Beispiel der Lichtsinneszellen: Phasisch-tonische Rezeptorzellen: Bei gleich bleibender Dauerreizung ist die Impulsfrequenz zunächst hoch, fällt nach einigen Sekunden oder Minuten auf einen niedrigen, konstanten Wert ab. Phasische Rezeptorzellen: Bei gleich bleibender Dauerreizung fällt die Impulsfrequenz auf Null ab. Sie reagieren sehr empfindlich auf Reizänderungen. Tonische Rezeptorzellen: Die Impulsfrequenz wird bei einer Dauerreizung nicht geändert Die Abweichung vom Ruhepotenzial an einer Rezeptorzelle, die durch einen Reiz ausgelöst wird, bezeichnet man als Rezeptorpotenzial. Je stärker der Reiz ist, desto stärker ist auch das Rezeptorpotenzial. Überschreitet das Rezeptorpotenzial den Schwellenwert, wird ein Aktionspotenzial ausgelöst. Dabei können auch mehrere unterschwellige Rezeptorpotenziale, die sich summieren, ein Aktionspotenzial bilden. Wird Licht durch Rhodopsin absorbiert, verändert der Retinalanteil seine Struktur vom cis-Retinal zum trans-Retinal und löst sich vom Opsin. Dadurch wird ein in der Discmembran liegendes G-Protein, das Transducin aktiviert, welches eine Phosphodiesterase (PDE) aktiviert. Diese hydrolisiert den sekundären Botenstoff der Stäbchen, zyklisches Guanosinmonophosphat (cGMP) zu 5’GMP. Das cGMP hält die Na+-Kanäle offen. Diese schließen sich, wenn es hydrolisiert wird. So entsteht das Rezeptorpotenzial der Stäbchen. Erregungsleitung (Ruhepotenzial, Aktionspotential) Ruhepotenzial: Die Konzentration der K+-Ionen ist auf der Innenseite der Membran 40fach Ionenverteilung: größer als auf der Außenseite. Aufgrund des K+-Ausstroms ist der Intrazellularraum relativ zum Extrazellularraum negativ geladen. Da sich die unterschiedlichen Ladungen anziehen lagern sich an der Membraninnenseite negativ geladene Ionen an, an der Außenseite hingegen positivgeladene Ionen. Hierdurch entsteht ein elektrisches Feld. Es wirken zwei Kräfte auf die K+-Ionen: Das Konzentrationsgefälle (Gradient) zwischen innen und außen, wodurch der Austritt der K+-Ionen durch Ionenkanäle begünstigt wird. Ein durch Ladungstrennung bedingtes elektrische Feld, das den Einstrom von K+-Ionen begünstigt. GleichgewichtsBeide Vorgänge stehen im Gleichgewicht. Die durch die K+-Ionen Potenzial: entstandene Spannung, das Kaliumgleichgewichtspotenzial, bildet die Grundlage für das Membranpotenzial im Ruhezustand, also das Ruhepotenzial. Die Na+ und Cl--Ionen beeinflussen dieses nur in geringem Maße. Aktionspotenzial Das Aktionspotenzial lässt sich in Phasen einteilen: (Ionenströme): A: Ruhezustand Die Nervenzelle ist innen negativ geladen, die Natriumkanäle sind geschlossen und nur einige Kalium-Ionen können zum Erhalt des Ruhepotenzials diffundieren. B: Depolarisation Ein elektrischer Impuls öffnet die Na+-Kanäle. Na+ wird von der negativen Ladung im inneren angezogen, so dass die Membran depolarisiert wird und das Zellinnere positiv geladen wird. C: Repolarisation Im Inneren befinden sich viel K+ und Na+-Ionen. Die Na+-Kanäle werden geschlossen und es öffnen sich die K+-Kanäle. Durch die gleiche Ladung stoßen sich diese ab und die K+-Ionen bewegen sich nach außen. Zudem werden durch die Na+-K+-Pumpe Na+-Ionen aus der Zelle transportiert. Die Innenseite verliert positive Ladung und die Membran wird repolarisiert. Na+-K+-Pumpe: Refraktärphase: Kontinuierliche: und saltatorische Fortleitung: D: Hyperpolarisation Nachdem das Ruhepotenzial erreicht ist strömen trotzdem noch einige K+Ionen nach außen, so dass das Zellinnere negativer als zuvor wird. Dies wird jedoch wieder ausgeglichen (K+-Ströme) und die Membran erreicht wieder das Ruhepotenzial. Das Ruhepotenzial wird durch Ionenleckströme verändert, bei denen Na+Ionen nach innen diffundieren und K+-Ionen veranlassen, nach außen zu diffundieren. Dem wirkt die Na+-K+-Pumpe entgegen, die unter ATPVerbrauch 3 Na+-Ionen wieder nach außen und 2 K+-Ionen wieder nach innen transportiert. Sie stellt auch bei der Repolarisation das Ruhepotenzial wieder her. Nach den Vorgängen bei der Entstehung eines Aktionspotenzials folgt die Refraktärphase, in der die Membran nicht erregbar ist oder die Erregbarkeit verringert ist. Der Schwellenwert ist dann höher, auch die Amplitude ist kleiner als gewöhnlich. Die bei einem Aktionspotenzial ins innere strömenden Na+-Ionen sorgen an dieser Stelle des Axons für eine positive Ladung. Der Bereich nebenan ist jedoch noch negativ geladen, so dass es zu einem Stromfluss zu den Seiten kommt, der elektrotonischen Ausbreitung. Der neue Membranbereich wird auch depolarisiert. Wird der Schwellenwert erreicht, entsteht ein neues Aktionspotenzial. Somit induziert ein Aktionspotenzial ein neues in der benachbarten Region bis das Endknöpchen erreicht ist. Die Fortleitung erfolgt nur in eine Richtung, da sich die Region, in der vorher ein Aktionspotenzial ausgelöst wurde, noch in der Refraktärphase befindet. Im Gegensatz zur kontinuierlichen Fortleitung sind die Axone hier mit einer Myelinscheide umgeben. Da diese elektrisch isolieren, erfolgt eine Auswirkung des Stromflusses erst am nächsten Schnürring, an dem wieder die Na+-Kanäle geöffnet werden. Die Aktionspotenziale bewegen sich also von Schnürring zu Schnürring, weswegen diese Form der Fortleitung schneller ist als die kontinuierliche. Die saltatorische Fortleitung erfolgt bei Wirbeltieren. Zudem wird hier weniger Energie verbraucht, da die Na+-K+Pumpe weniger Ionen transportieren muss. Synapsenvorgänge und Verrechnung Bau und Funktion Synapsen lassen sich in drei Bereiche einteilen: von erregenden und präsynaptische Endigung des Neurons, dass die Information überträgt, die hemmendem präsynaptische Zelle. Die präsynaptische Endigung enthält synaptische Synapsen: Vesikel mit den Neutotransmittern (Übertragungssubstanzen) synaptischer Spalt zwischen den beiden Neuronen, befestigt die präsynaptische Endigung mit dem nachgeschalteten Neuron postsynaptische Zelle, die Zellmembran des nachgeschalteten Neurons. Trifft ein Aktionspotenzial in der präsynaptischen Endigung ein, wird die Zellmembran depolarisiert. Spannungsabhängige Ca2+-Kanäle öffnen sich. Durch den Anstieg der Ca2+-Konzentration verschmelzen die Vesikel mit der Zellmembran und geben die Transmitter in den synaptischen Spalt ab. An der postsynaptischen Membran befinden sich transmittergesteuerte Ionenkanäle, die sich nur öffnen, wenn der passende Transmitter – zwei Moleküle pro Kanal – an sie bindet. Die Öffnung bleibt nur so lange bestehen wie die Transmitter angelagert sind. Diese werden von bestimmten Enzymen der postsynaptischen Membran gespalten. Durch die geöffneten Kanäle strömen Ionen ein, die das Membranpotenzial ändern. Das so genannte postsynaptische Potenzial (PSP) ist der Transmittermenge proportional. Wird die postsynaptische Membran durch das PSP depolarisiert, entsteht ein neues Aktionspotenzial. Die Spaltung der Transmitter verhindert dabei eine Dauererregung. Daher muss der Transmitter in der präsynaptischen Endigung neu synthetisiert werden. Daher werden die Produkte der Spaltung recycelt. Im Falle des Transmitters Acetylcholin, der in Cholin und Essigsäure gespalten wird, diffundiert das Cholin wieder in die präsy- naptische Endigung, in der ein Essigsäurerest wieder auf das Cholin übertragen wird. Acetylcholin entsteht wieder und wird in synaptische Vesikel transportiert und steht wieder zur Verfügung. Synapsen, die eine Depolarisation bewirken werden als erregende Synapsen bezeichnet. Die von ihnen ausgehenden PSP werden als exzitatorische postsynaptische Potenziale (EPSP) bezeichnet. Wird jedoch eine Hyperpolarisation durch die Öffnung von Cl-Ionenkanälen bewirkt, so spricht man von einer hemmenden Synapse. Die entstehenden PSP werden inhibitorische postsynaptische Potenziale (IPSP) genannt. Ein Neuron kann seine Informationen von vielen tausend erregenden und hemmenden Synapsen erhalten. Dieser Vorgang wird als Konvergenz bezeichnet. Werden die Aktionspotenziale durch Verzweigungen eines Axons auf viele Folgeneurone verteilt, so spricht man von Divergenz. Motorische Endplatte: Ort der Informationsübertragung zwischen Nervenzelle und Muskelfaser. Die Endplatte liegt an der Oberfläche der Muskelfaser an. Durch Abgabe von Acetylcholin in den synaptischen Spalt wird die Muskelfaser durchlässig für Na+- und K+-Ionen. Dadurch entsteht ein Aktionspotenzial an der Muskelfaser, das Endplattenpotenzial. Endplattenpotenzial: Wird auf das sarkoplasmatische Retikulum übertragen und erreicht in kürzester Zeit jedes Sarkomer der Muskelfaser. Aus dem sarkoplasmatischen Retikulum strömen dann Ca2+-Ionen in das Cytoplasma, die einen Komplex aus zwei Proteinen, Troponin und Tropomyosin bewirken. Beide sind an das Actin-Filament gebunden. Im entspannten Zustand blockiert Tropomyosin die Bindungsstelle für Myosin, Troponin bindet Ca2+-Ionen. Durch die Bindung wird die Konformation beider Proteine verändert, das Tropomyosin gibt die Myosin-Bindungsstellen nun frei. Verrechnung: Treffen an einem Axonhügel mehrere EPSP, die von verschiedenen räumliche erregenden Synapsen ausgehen, ein, die allein den Schwellenwert nicht überschreiten würden (unterschwellig sind), können sie durch die räumliche Summation dennoch ein Aktionspotenzial auslösen. Es können auch mehrere EPSP und IPSP zusammentreffen, so dass ein Rezeptorpotenzial das den Schwellenwert erreichen würde kein Aktionspotenzial auslöst. und zeitliche Treffen innerhalb kurzer Zeit mehrere unterschwellige EPSP der gleichen Summation: erregenden Synapse am Axonhügel ein können diese durch die Summation trotzdem ein Aktionspotenzial auslösen. Auch hier kann es wieder zu einer hemmenden Wirkung durch IPSP kommen. 3. Nervensystem Bau und Funktion von ZNS, peripherem Nervensystem, vegetativem Nervensystem Bau: Die Neuronen lassen sich je nach „Richtung“ in zwei verschiedene Gruppen einteilen: afferente Neuronen sind sensorische Neuronen, sie leiten Signale zum ZNS efferente Neuronen sind motorische Neuronen, sie leiten Signale vom ZNS Zudem können die Nervenfasern ebenfalls in sensorische, die von einem Rezeptor, und motorische, die zu einem Muskel oder einer Drüse führen, einteilen Hierarchische ZNS (Zentralnervensystem): Gehirn und Rückenmark Struktur des peripheres Nervensystem: (Gesamtheit der Nerven, die die einzelnen Nervensystems: Organe versorgen), unterteilt in afferent (Peripherie Æ Zentrum) und efferent (Zentrum Æ Peripherie) vegetatives Nervensystem: (autonomes Nervensystem), versorgt die Eingeweide, unterliegt nicht der willentlichen Kontrolle, unterteilt in: sympathisches NS: Erhöht Bereitschaft zum Angriff, Verteidigung, Flucht parasympathisches NS: Vorgänge, die der Erholung dienen neuronale und hormonelle Verschränkung im vegetativen Nervensystem: Das vegetative Nervensystem hat über den Sympathicus und Parasympathicus Einfluss auf die Hormone. So sorgt der Sympathicus bei Gefahr für Adrenalinausschüttung im Nebennierenmark (FFS). Willkürliche und unwillkürliche Reaktionen Reflex: schnelle, stereotype nervös ausgelöste Reaktion des Organismus auf einen Reiz; wird beim Überschreiten der Reizschwelle unausweichlich. Reflexe laufen unbewusst ab und treten erst nach der Reaktion ins Bewusstsein. Reflexbogen: Die Verschaltung von Neuronen, die einem Reflex zu Grunde liegt, wird als Reflexbogen bezeichnet. Feinbau von Muskeln Ein Muskel besteht aus vielen Muskelfaserbündeln, die mehrere Muskelbis auf die Ebene fasern enthalten. Der Großteil des Cytoplasmas ist von zylindrischen Zellder Filamente: bestandteilen durchzogen, den Myofibrillen. Die Myofibrille besteht aus gleichen Untereinheiten, den Sarkomeren, die in der Mitte dunkle ABanden und an den Enden helle I-Banden aufweisen. Die Grenze zwischen den Sarkomeren bilden die Z-Scheiben. Sarkomere sind aus Filamenten aufgebaut, den dickeren Myosin-Filamenten und den dünneren ActinFilamenten. Actin ist ein kugelförmiges Protein, mehrere dieser Proteine sind wie Perlen aneinandergereiht. Zwei solcher Perlenketten bilden ein Actin-Filament. Myosin ist ein lang gestrecktes Molekül, an dessen „Stiel“ eine Art „Köpfchen“ sitzt (ähnlich wie ein Golfschläger). Sie kommen Paarweise vor. 100 solcher Paare bilden ein Myosin-Filament. Actin-Filamente sind an den Z-Scheiben befestigt, in der Mitte des Sarkomers, der A-Bande, überlappen sie mit den Myosin-Filamenten. Dabei ist jedes Myosin-Filament von 6 Actin-Filamenten umgeben. Im Bereich der IBande befinden sich nur Actin-Filamente. Muskeln arbeiten nach dem Antagonismus-Prinzip. Energetische Bei der Muskelkontraktion wird ATP durch die Myosinköpfe wieder in ADP Prozesse bei der und P gespalten. Um die Kontraktion aufzuheben und eine erneute zu Muskelkontraktion: ermöglichen muss wieder ATP bereitgestellt werden. Phasen der Energie- Der ATP-Speicher ist bereits nach ca. 2-3 Muskelkontraktionen verbraucht, bereitstellung im daher muss der Speicher ständig aufgefüllt werden. Zunächst wird mit Muskel bei Belastung: Hilfe von Kreatinphosphat (KP) ADP wieder in ATP umgewandelt. Der KPSpeicher ist jedoch auch nach ca. 8-9 Sekunden aufgebraucht. Ist die Hälfte des Speichers verbraucht erfolgt die Energiebereitstellung durch die anaerobe Oxidation, die nach ca. 40 Sekunden durch die effektivere aerobe Oxidation abgelöst wird. Molekulare GrundOhne Anwesenheit von ATP sind die Myosinköpfe an das Actin gebunden. lagen der MuskelUnter Anwesenheit von ATP beginnt die Muskelkontraktion. Der Myosinkontraktion kopf bindet das ATP und löst sich vom Actin. Das ATP wird gespalten, so (Gleitfilamentdass der Myosinkopf wieder an das Actin bindet. Hierbei wird das Phosphat Modell): freigesetzt. Nun knickt der Myosinkopf ab und zieht das Myosin-Filament in Richtung der Z-Scheibe. Das ADP wird freigesetzt und die Kontraktion kann von neuem beginnen. 4. Verhalten Genetische Disposition und erfahrungsbedingte Formung des Verhaltens Ultimate und Soziobiologen untersuchen, warum sich die Verhaltensweisen eines Tieres in der Evolution durchsetzen konnten. Es wird versucht, die zu Grunde liegenden Gesetzmäßigkeiten der Evolution zu ergründen. Dies sind die ultimaten Ursachen (Wozu-Fragen). proximate Ursachen: Untersucht man die im Tier ablaufenden Mechanismen, die eine Entscheidung zu einer Reaktion bewirken, spricht man von den proximaten Ursachen (Wie-Fragen). Man sucht nach den genetisch festgelegten Nervenschaltungen und hormonellen Zuständen, die zu Grunde liegen. Angepasstheit von Verhalten Kosten-NutzenBerechnung, ob es sich lohnt eine bestimmte Aktion auszuführen, z.B. Analyse: Kampf in der Fortpflanzungszeit. Es wird abgewogen, wie viel Aufwand betrieben werden muss (Kraftaufwand, evtl. Verletzungen) und ob ein Nutzen daraus gezogen werden kann (Nachkommen). Erst wenn der Nutzen die Kosten überwiegt oder verhältnismäßig ist wird ein Kampf eingegangen. Reproduktive Fitness: Die relative Anzahl an Genen eines Individuums, die in die nächste Generation weitergegeben wird. Dies kann direkt oder indirekt erfolgen. r- und K-Strategien: r-Strategen (Vermehrungsrate) zeugen sehr viele Nachkommen, betreiben danach jedoch keine Brutpflege. Der Erfolg wird durch die Masse an Nachkommen erhöht. Meist kommen in dem Lebensraum wenige Konkurrenten vor. K-Strategen (Kapazität) zeugen nur wenige Nachkommen, die lange großgezogen werden. Durch das Betreiben einer ausführlichen Brutpflege soll der Nachkomme bestmöglich für Konkurrenzkämpfe vorbereitet werden, so dass die eigenen Gene weitergegeben werden. Der Druck kann auch durch eine erhöhte Zahl an Konkurrenten im Lebensraum verstärkt werden. Paarungs- und Monogamie: Paarbindung zwischen einem Männchen und einem Weibchen über eine Fortpflanzungsperiode oder bis zum Tod des Partners Polygamie: Paarung mit mehreren Partnern des anderen Geschlechts. Polygynie: ein Männchen, mehrere Weibchen, Brutpflege Weibchen Polyandrie: ein Weibchen, mehrere Männchen, Männchen übernehmen teilweise die Brutpflege Polygynandrie: mehrere Weibchen und Männchen, mehrfache Paarung mit verschiedenen Partnern. Beide Geschlechter betreiben Brutpflege Sozialsysteme bei Primaten bilden Rangordnungen aus, bei dem ein Männchen das rangPrimaten: höchste Tier ist. Es wird als Alpha-Tier bezeichnet. Die Stellung hat es durch einen Kampf errungen, und kann diese genauso wieder verlieren, wenn es durch einen stärkeren Kontrahenten besiegt wird. Bei der linearen Rangordnung dominieren die ranghöheren Tiere jeweils alle rangniedrigeren Tiere. Bei Weibchen sind Rangordnungen meist nicht ausgeprägt. Uneigennütziges Bei altruistischem (uneigennützigem) Verhalten setzt ein ausführendes Verhalten: Tier Kosten ein, um einem empfangendem Tier einen Nutzen zuzuführen. Sind die Tiere verwandt entsteht trotzdem ein Nutzen, da durch erhöhte Fortpflanzungschancen auch eigene Gene weitergegeben werden. Neben Kosten und Nutzen spielt auch der Verwandtschaftsgrad eine Rolle. Die Hamilton-Ungleichung liefert eine Formel hierfür: K < r * N. Die Kosten (K) für den Altruisten müssen geringer sein als der Nutzen (N) für den Empfänger multipliziert mit dem Verwandtschaftsgrad (r). Ein Beispiel für Altruismus ist die Hilfe bei der Aufzucht der Kinder von Geschwistern, wenn z.B. eigene Nachkommen nicht möglich sind. Der genetische Verwandtschaftsgrad liegt bei eigenen Kindern bei 0,5, bei Neffen oder Nichten bei 0,25. 5. Wirkung von Medikamenten und Drogen Wirkungsmechanismen Wirkung von Neurotoxinen und psychoaktiven Stoffen an Synapsen: ausgewählter Medikamente und Drogen Neurotoxine können verschieden an Synapsen wirken. So führt eine Hemmung der Acetylcholinesterase durch E 506 zu Dauererregungen und Krämpfen, da der Transmitter nicht abgebaut wird. Bei Curare werden die Acetylcholin-Rezeptoren blockiert, so dass es zu einer Lähmung kommt. Möglich ist auch eine Entleerung aller Vesikel in den synaptischen Spalt, so dass es zu einer Übererregung kommt. Suchtmechanismus auf zellulärer Ebene am Beispiel der Morphinsucht: Psychoaktive Stoffe wie z.B. Opiate sind ähnlich aufgebaut wie die körpereigenen Endorphine und erzeugen Glücksgefühle. Sie reagieren mit dem Enzym Adenylatcyclase, dass daraufhin ATP nicht zu cAMP umsetzt. Die Ca2+-Kanäle können nicht geöffnet werden. Die Erregungsweiterleitung ist unterbrochen und Schmerzen werden nicht wahrgenommen. Da die Opiate die Rezeptoren der Adenylatcyclase blockieren und somit keine Umsetzung von ATP zu cAMP erfolgt bildet der Körper weitere Rezeptoren, damit die Erregungsleitung wieder möglich ist. Da nun mehr Rezeptoren vorhanden sind, empfindet der Konsument nicht mehr ein so starkes Rauschgefühl wie beim ersten Mal und erhöht die Dosis, so dass wieder alle Rezeptoren blockiert sind. Dieser Vorgang wiederholt sich in einer Art Kreislauf. Wird ein Entzug begonnen liegen relativ viele Rezeptormoleküle frei und wahrgenommene Empfindungen werden viel stärker als im normalen Zustand weitergeleitet. Diese sorgen für die Schmerzen bei einem Entzug. Mit der Zeit baut der Körper die Rezeptoren wieder ab so dass der Ausgangszustand wiederhergestellt wird. Thematischer Schwerpunkt 3: Evolution 1. Ursachen der Evolution Evolutionsfaktoren und ihr Zusammenwirken Mutationen: Zufällige, spontane Veränderung der Erbsubstanz. Kann an Nachkommen vererbt werden, wenn sie in den Geschlechtszellen stattfindet. Können durch Ablesefehler oder durch äußere Einflüsse entstehen. Rekombination: Neuverteilung der Erbinformation, bei sexueller Fortpflanzung durch die Kombination der beiden elterlichen Erbinformationen. Isolation: Voraussetzung für die Aufspaltung einer Art in zwei Tochterarten. Räumliche Trennung verhindert den Genaustausch und genetische Unterschiede können entstehen. Selektion: Selektion fungiert als richtender Faktor, der Mutationen und Rekombinationen nach der Eignung in der Umwelt sortiert. Dabei wird anhand des Phänotyps sortiert. Gendrift: Veränderung des Genpools einer Population aufgrund von Zufall. Flaschenhalseffekt: Die genetische Variabilität wird aufgrund einer Umweltkatastrophe stark verringert Gründereffekt: Durch Neubesiedlung eines Lebensraums mit wenigen Gründerindividuen wird der Genpool stark verändert Lamarckismus: Dem Lamarckismus nach erfolgt zunächst eine Änderung der Umwelt, an die sich Lebewesen anpassen. Gebrauchte Organe werden verbessert, nicht gebrauchte verkümmern. So entwickelte die Giraffe einen langen Hals, damit sie an die höher gelegenen Blätter gelangt. Darwinismus: Jede Art zeugt mehr Nachkommen als aufgrund der zur Verfügung stehenden Rohstoffe überleben können. Es kommt zu einem Kampf ums Dasein (struggle for life). Es überleben im natürlichen Wettbewerb um Nahrung, Lebensraum und Geschlechtspartner diejenigen, die am besten an die Umweltbedingungen angepasst sind (survival of the fittest). Durch natürliche Zuchtwahl (natural selection) kommt es zu einer stetigen Veränderung der Arten, bei der nur die am besten angepassten überleben. Synthetische Vereinigt Darwins Theorie mit modernen Erkenntnissen der Genetik. Evolutionstheorie: Demnach liefern Zufallsprozesse (Mutation, Rekombination, Gendrift) das Rohmaterial, an dem dann als zweiter Prozess die Selektion greift, die keine Zufallsergebnisse liefert. Durch reproduktive Isolation entstanden neue Arten und die biologische Vielfalt. Trotzdem bleibt es eine Theorie, die sich auf vergangenes bezieht und nicht durch Experimente bestätigt oder widerlegt werden kann. Genetische und Durch sexuelle Fortpflanzung erhalten Individuen Erbinformationen von zwei verschiedenen Individuen. Die Neukombination des Erbgutes sorgt für genetische Variabilität modifikatorische Trotz gleichen Erbguts stimmen Lebewesen nicht in allen äußeren Variabilität: Merkmalen überein. Umweltfaktoren beeinflussen bestimmte Perioden der Entwicklung und können den Phänotyp verändern. Ursachen genetischer Rekombinationen und Mutationen sorgen für genetische Variabilität, die Variabilität: sich in einer phänotypischen Vielfalt äußert. Allelfrequenzen: Geben die relative Allelhäufigkeit in einer Population an Genpool: Der Gesamtbestand der Gene einer Population Populationsgenetik Für Berechnungen nach Hardy-Weinberg geht man von einer Idealmit einfachen population aus, in der keine Mutationen, keine Selektion und Panmixie Berechnungen (d.h. alle Individuen paaren sich gleich häufig miteinander) vorherrschen. nach HardyDie Allele werden durch die Buchstaben p und q beschrieben, wobei gilt, Weinberg: dass p + q = 1 und p² + 2pq + q² = 1 Präadaptation: Zufällige Mutation, die durch eine Umweltveränderung von Vorteil sein kann. Individuen verfügen über eine zufällige Anpassung an zukünftige Umweltsituationen. Kann auch als rezessive Mutation lange in heterozygoter Form überdauern, bei einer Umweltveränderung profitieren dann die homozygoten Träger. Selektionsfaktoren: abiotische Faktoren: Klima, Bodenverhältnisse, geografische Lage etc. biotische Faktoren: ▪ Schrecktracht: Tagpfauenauge ▪ Warntracht: Wespe ▪ Tarntracht: Chamäleon ▪ Mimese (Nachahmungstracht): Spannerraupe ▪ Mimikry (Scheinwarntracht): Schwebfliege Selektionstypen: natürliche Selektion: Individuen, die aufgrund ihrer erblichen Anlagen besser mit der veränderten Umwelt zurechtkommen haben höhere Überlebenschancen gerichtete (transformierende) Selektion: Präadaptation, ein Extrem des Merkmalsspektrums hat einen Selektionsvorteil. Häufig bei Umweltveränderungen stabilisierende Selektion: Häufig bei gleich bleibender Umwelt, Individuen die nahe am Mittelwert (Durchschnitt) liegen werden bevorzugt disruptive Selektion: Die extremen Phänotypen an den Enden des Merkmalsspektrums haben einen Selektionsvorteil. Isolationsreproduktive Isolation, die zu der sympatrischen Artbildung gehört, also zu mechanismen: der Artbildung die ohne eine räumliche Isolation stattfindet. Darunter: präzygotisch (bevor eine Befruchtung erfolgt): zeitliche Isolation: Paarung erfolgt zu unterschiedlichen Zeiten mechanische Isolation: Geschlechtsverkehr ist aufgrund der Anatomie nicht möglich Verhaltensisolation: Signale (Balzrituale) die den Partner anlocken gametische Isolation: Ei- und Samenzelle verschmelzen nach dem Schlüssel-Schloss-Prinzip Habitatisolation: Bewohnung unterschiedlicher Lebensräume (Wasser – Land) postzygotisch (nach der Befruchtung): Bastardsterblichkeit: Die Hybriden sind nicht lebensfähig und beenden ihre Entwicklung nicht Bastardsterilität: Die Hybride sind steril und es erfolgt kein weiterer Genfluss Bastardzusammenbruch: Die Hybriden sind lebensfähig und fruchtbar, die Nachkommen jedoch sind schwach oder steril Artbildung Art und Population: allopatrische Artbildung: Veränderung des Genpools: Adaptive Radiation: Co-Evolution: Gruppe von Individuen, die durch gemeinsame Abstammung miteinander verbunden sind und (bei zweigeschlechtlicher Fortpflanzung) fruchtbare Nachkommen miteinander Zeugen können. Individuen einer Art, die zur gleichen Zeit im gleichen Raum leben und eine Fortpflanzungsgemeinschaft bilden. Form der Isolation, bei der Populationen durch geografische Barrieren nicht mehr miteinander in Kontakt stehen und der Genfluss so unterbrochen wird. So können zwei verschiedene Arten entstehen. Einige Barrieren wären z.B. große Entfernungen, klimatische oder tektonische Veränderungen; aber auch durch Sturm in weit entfernte Gebiete transportierte Arten, die Gründer neuer Populationen werden. Die Allele in einem Genpool treten in unterschiedlicher Häufigkeit (Allelfrequenz) auf. Durch Mutationen wird die Vielfalt des Genpools ständig erhöht. Es ergibt sich eine ständige Tendenz zur Veränderung des Genpools. Form der sympatrischen Artbildung, bei der eine ökologische Isolation erfolgt. Vertreter einer Art besetzen aufgrund mangelnder Konkurrenz verschiedene ökologische Nischen und entwickeln sich mit der Zeit zu eigenständigen Tochterarten. Einige Arten sind zudem endemisch, d.h. sie kommen nur in diesem Lebensraum vor. Prozess der wechselseitigen Einflussnahme zweier stark interagierender Arten aufeinander, der sich über sehr lange Zeiträume in der Stammesgeschichte beider Partner erstreckt. (Bsp.: Weil die Gazelle immer schneller wird muss auch der Gepard immer schneller werden.) 2. Verlauf der Evolution Belege für den Verlauf der Evolution Divergenz und Auseinanderentwicklung zweier Tochterpopulationen aufgrund von Isolation, bis schließlich eine genetische Isolation vorliegt und zwei neue Arten entstehen. Konvergenz: Ausbildung funktionsgleicher Organe, die in der Stammesgeschichte unabhängig voneinander entstanden sind. (Bsp.: Körperform von Fischen) Konvergenz bezeichnet dabei den Prozess, das Endstadium ist die Analogie Homologiekriterien: Kriterium der Lage: Strukturen sind dann homolog, wenn sie in vergleichbaren Gefügesystemen gleich angeordnet sind. Kriterium der spezifischen Qualität: Ähnliche Strukturen können auch ohne Berücksichtigung ihrer Lage homologisiert werden, wenn sie in zahlreichen Sondermerkmalen übereinstimmen. Kriterium der Stetigkeit: Verknüpfung durch Zwischenformen; die verschiedenen Ausbildungsformen homologer Organe treten nicht regellos auf, sondern sie bilden Entwicklungsreihen, deren Endpunkte durch Zwischenformen miteinander verbunden sind. Zwischenformen können in der Embryonalentwicklung oder als Fossilfunde auftreten. Morphologische und Progressionsreihen: Reihen aus Vertretern einer Gruppe mit unterschiedanatomische lich weit fortgeschritten Entwicklungen. Bei einer Regressionsreihe lassen Verwandtschaftssich unterschiedlich weit fortgeschrittene Rückbildungen feststellen. Beibelege: spiel für eine Progressionsreihe ist der Blutkreislauf verschiedener Wirbeltiere. Atavismus: Das Wiederauftreten eines Merkmals, das im Laufe der Evolution bereits verschwunden war. Sie beweisen dass die Gene noch vorhanden sind, jedoch nicht mehr phänotypisch ausgebildet werden. (Bsp.: Griffelbeine beim Pferd oder Schweif beim Menschen) Rudimente: Reste ehemals funktionstüchtiger Organe der Vorfahren, die im Laufe der Evolution aufgrund einer Veränderung der Lebensweise ihre Funktion verloren haben und sich zurückbildeten. (Bsp.: Beckenknochen beim Grönlandwal) Biogenetische Grundregel nach Haeckel: Die Ontogenese (Individualentwicklung) ist eine kurze und schnelle Wiederholung der Philogenese (Stammesentwicklung). Es werden jedoch nur einzelne, nicht vollständig ausgebildete Anlagen eines Organs ausgebildet, nicht die adulte Form. Analyse bzw. Erstellung eines Stammbaumes Methoden für biochemische und molekularbiologische Verwandtschaftsbelege und Stammbäume: DNA-Sequenz: Vergleich von Nucleotidsequenzen von DNA oder RNA. Geeignet sind Gene, die für Proteine codieren, die überall vorkommen; z.B. Proteine der Atmungskette. Aminosäure-Sequenz Vergleich der Aminosäuren-Abfolge in einem Stoffwechselprotein, das bei von Proteinen: vielen Arten vorkommt. Je näher verwandt die Arten sind, desto ähnlicher ist der Bau der Proteine und demnach die DNA (Ein-Gen-ein-PolypeptidHypothese) Ein Beispiel wäre der Vergleich von Cytochrom-c, ein Enzym, das im Mitochondrium bei der Zellatmung benötigt wird. Anhand solcher Informationen lassen sich Molekülstammbäume erstellen. DNA-Hybridisierung: Die DNA zweier Arten wird isoliert, durch Enzyme in Fragmente gespalten und die DNA der Art A radioaktiv markiert. DNA der Art B wird in 1000facher Menge mit Art-A-DNA gemischt. Das Gemisch wird auf 95° erhitzt, so dass die H-Brücken der Stränge gespalten werden. Danach wird auf 60° abgekühlt und die Temperatur gehalten. Es kommt zur Neubildung der HBrücken zwischen den Basen und es entstehen neukombinierte DNADoppelstränge. Übrige Einzelstränge werden ausgewaschen. Nun wird die Temperatur stufenweise erhöht, so dass es erneut zum Schmelzen der DNA kommt. Die neu entstandenen Einzelstränge jeder Stufe werden wieder ausgewaschen. In den Auffanggefäßen wird die Radioaktivität gemessen, wobei nur die Hybrid-DNA radioaktiv ist. Der Schmelzpunkt, bei dem 50% der Hybrid-Doppelstränge geschmolzen sind, wird als T50-H-Wert bezeichnet. Übersicht über den Fische Æ Amphibien Æ Reptilien Æ Vögel Æ Säugetiere Wirbeltierstammbaum: