Klassische Konditionierung Warum soll
Werbung

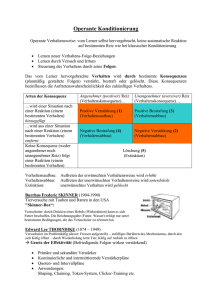
Dr. Götz Fabry Vorlesung Medizinische Psychologie 30.04.2004 Psychologische Lerntheorien: Klassische Konditionierung Warum sollten sich angehende Ärzte mit psychologischen Lerntheorien auseinandersetzen, abgesehen davon, daß sie als Grundlagenwissen der Medizinischen Psychologie im Gegenstandskatalog aufgeführt werden? Aus der Vielzahl guter Gründe seien hier nur einige exemplarisch heraus gegriffen: 1) Es gibt praktisch keine therapeutische Maßnahme, die nicht auch eine Verhaltensänderung umfaßt (z.B. mit dem Rauchen aufhören, sich häufiger bewegen, Tabletten einnehmen). Mit vielen dieser Verhaltensänderungen sind auch Lernvorgänge verbunden, z.B. weil neue Bewegungsabläufe erlernt werden müssen oder bestimmte Gewohnheiten verlernt werden müssen. Auf der Grundlage lerntheoretischer Erkenntnisse wird der Arzt den Patienten in diesem Prozeß besser unterstützen und anleiten können als ohne dieses Wissen. 2) Lernprozesse können die Entstehung und den Verlauf verschiedener Erkrankungen beeinflussen. Insbesondere für die Angststörungen wurden verschiedene Modelle vorgeschlagen, die aus lerntheoretischen Überlegungen hervor gegangen sind und auf deren Grundlage effektive therapeutische Strategien entwickelt wurden. 3) Auch unter Selbsterfahrungsperspektive ist die Auseinandersetzung mit psychologischen Lerntheorien gewinnbringend, wenn man etwa an die im Medizinstudium zu bewältigenden Stoffmassen denkt. Folie 1 Vorlesung Medizinische Psychologie SS 2004 Psychologische Lerntheorien Klassische Konditionierung Vorlesungsskripte unter: http://www.medizinische-psychologie.de [lehre] [vorlesung] Am Beispiel der klassischen Konditionierung als einer der einfachsten Formen assoziativen Lernens (assoziativ = verbindend, hier: Verknüpfen von zwei Ereignissen) soll gezeigt werden, wie ein zunächst nur auf der Verhaltensebene beobachtetes Phänomen mittlerweile bis auf seine zellulären Grundlagen hinunter verstanden werden konnte und damit grundlegende Erkenntnisse über die bei der Gedächtnisbildung beteiligten neurobiologischen Mechanismen möglich wurden. Das Konzept der Klassischen Konditionierung ist untrennbar mit dem russischen Physiologen Iwan Petrowitsch Pawlow (1849 – 1936) verbunden, der bei seinen Untersuchungen zur Speichel- und Magensaftsekretion von Hunden zufällig eine verblüffende Entdeckung machte. Läutete man kurz bevor man dem Hund einen vollen Freßnapf präsentierte, was natürlich massiven Speichelfluß auslöste, eine Glocke, so war nach einigen solcher Durchgänge der Glockenton allein ausreichend, um ebenfalls einen massiven Speichelfluß auszulösen. Der ursprünglich neutrale Stimulus (NS) war durch die enge zeitliche Kopplung an den unkonditionierten (engl.: unconditioned) Stimulus (UCS) zu einem konditionierten (engl.: conditioned) Stimulus (CS) geworden, der allein in der Lage ist, eine konditionierte Reaktion (CR) auszulösen, die sich nicht von der unkonditionierten Reaktion (UCR) unterscheiden läßt (Folie 2). Folie 2 Klassische Konditionierung UCS UCR „Futter“ „Speichel“ NS „Glocke“ CS CR „Glocke“ „Speichel“ Dieses erstaunliche, von Pawlow erstmals beschriebene Phänomen konnte durch zahlreiche andere Untersuchungen bestätigt werden und führte zu der in Folie 3 beschriebenen Charakterisierung. Demnach werden bei der klassischen Konditionierung neue Reize in ein bestehendes Reflexsystem integriert, wobei die ehemals neutralen Reize zu konditionierten Reize werden, wenn eine unmittelbare zeitliche Nähe von neutralem Reiz und unkonditioniertem Reiz gegeben ist und zwar dergestalt, daß der neutrale Reiz dem unkonditionierten Reiz unmittelbar vorausgeht. Die durch den schließlich konditionierten Reiz ausgelöste konditionierte Reaktion muß der unkonditionierten weitgehend ähnlich sein. Folie 3 Klassische Konditionierung (traditionelles Verständnis) • Integration neuer Reize in ein bestehendes Reflexsystem • neutrale Reize werden zu konditionierten Reizen • unmittelbare zeitliche Nähe von neutralem Reiz und unkonditioniertem Reiz (Kontinguität) • konditionierte Reaktion muß der unkonditionierten weitgehend ähnlich sein Dieses traditionelle Verständnis von klassischer Konditionierung hat sich unter dem Einfluß neuer Forschungsergebnisse gewandelt. Betrachten wir dazu die in Folie 4 schematisch dargestellten Versuchsanordnungen. Folie 4 Konditionierungsphänomene CR + 0 – (+) + = neutraler Reiz = unkonditionierter Reiz In der ersten Zeile ist eine klassische Konditionierung wie in Pawlows Versuch dargestellt. Nach einigen Paarungen von neutralem und unkonditioniertem Reiz löst der nun konditionierte Reiz die konditionierte Reaktion aus. Die Versuchsanordnung in der zweiten Zeile unterscheidet sich von der ersten nur dadurch, daß der unkonditionierte Stimulus zwischen den gepaarten Durchgängen auch ohne den neutralen Stimulus gegeben wurde. Mit dieser Anordnung ist es nicht möglich eine Konditionierung zu erzeugen, der neutrale Reiz bleibt neutral und löst allein keine Reaktion aus. In der dritten Zeile erfolgt die Präsentation von neutralem und unkonditioniertem Reiz vollkommen willkürlich, das Ergebnis scheint zunächst das gleiche zu sein, wie in Zeile 2. Wird jedoch im gleichen Experiment ein „klassischer“ Durchgang angeschlossen, dann zeigt sich, daß es jetzt schwieriger ist, den neutralen Reiz zu einem konditionierten Reiz zu machen. Der neutrale Reiz scheint also gar nicht mehr neutral zu sein, sondern mit der Information aus dem vorangegangenen Durchgang aufgeladen, daß er eben in keinem relevantem Zusammenhang mit dem unkonditionierten Reiz steht, daß ihm quasi einen „Nicht-Bedeutung“ zukommt. In Zeile 3 ist also ebenfalls etwas gelernt worden (nämlich eine „Nicht-Assoziation“), was insofern überrascht, als der enge zeitliche Zusammenhang wie er in der traditionellen Form des klassischen Konditionierens beschrieben ist, hier nicht gegeben ist. Auch mit den in Folie 5 dargestellten Versuchsanordnungen wurden Erkenntnisse gewonnen, die das Konzept des klassischen Konditionierens erweitert haben. Statt einem neutralen Reiz wurde hier mit zwei verschiedenen Reizen (z.B. Glockenton und Lichtblitz) gearbeitet. In Experiment 1 wurde zunächst eine herkömmliche Konditionierung à la Pawlow durchgeführt, aus der ein konditionierter Reiz resultierte. In einem zweiten Durchgang wurde ein neuer neutraler Reiz hinzugefügt, so daß der konditionierte Reiz aus dem ersten Durchgang und der neue neutrale Reiz gleichzeitig vor dem unkonditionierten Reiz gegeben wurden. Merkwürdigerweise gelang es in diesem zweiten Durchgang aber nur schlecht, den zweiten neutralen Reiz ebenfalls zu einem konditionierten Reiz zu machen. Dies ist umso erstaunlicher, als – wie in Experiment 2 gezeigt – die zeitgleiche Präsentation von zwei neutralen Reizen vor einem unkonditionierten Reiz beide zu gleichstarken konditionierten Reizen werden läßt. Offensichtlich blockiert der erste Durchgang in Experiment 1 weitere Konditionierungen, daher wird der gesamte Vorgang auch als „Blockade-Phänomen“ bezeichnet. Folie 5 „Blockade-Phänomen“ CR + 1 (+) 2 + = neutrale Reize = unkonditionierter Reiz Diese Erkenntnisse führten schließlich zu einem erweiterten Konzept des klassischen Konditionierens (Folie 6), das demnach als der Prototyp assoziativen Lernens angesehen werden kann, eines Lernens also, bei dem verschiedene Ereignisse aber auch Sinnesqualitäten, Wahrnehmungen und Erinnerungen, Wahrnehmungen und Gefühle u.a. miteinander verknüpft werden. Schon diese kurze Aufzählung macht deutlich, daß damit ein ganz grundlegender Mechanismus des Lernens überhaupt angesprochen ist. Folie 6 Klassische Konditionierung (modernes Verständnis) • Verhaltensänderung durch Konfrontation mit Ereignisrelationen (Zusammenhänge zwischen Reizen) • Lernvorgang, der Voraussagen über Zusammenhänge zwischen Ereignissen ermöglicht Mittlerweile ist es auch gelungen, die neurobiologischen Grundlagen der klassischen Konditionierung am Tiermodell aufzuzeigen. Großen Anteil an diesen Entdeckungen hatte der amerikanischen Neurowissenschaftler Eric Kandel (geb.: 1929), der für seine bahnbrechenden Forschungen auf diesem Gebiet im Jahr 2000 mit dem Medizin-Nobelpreis ausgezeichnet wurde. Folie 7 zeigt eine schematische Darstellung der Meeresschnecke Aplysia californica, an deren relativ einfach gebautem Nervensystem, das aus „nur“ 20.000 Nervenzellen besteht, die neurobiologischen Grundlagen des Lernens erforscht wurden. Im Zentrum des Interesses stand dabei die Steuerung des Kiemenrückzugsreflexes, der durch taktile Stimulation des Mantels oder des sogenannten Syphons (einem Ausscheidungsorgan) ausgelöst werden kann. Folie 7 Aplysia californica Kopf Mantel Kiemen Syphon Elektroschock taktile Stimulation Schwanz n. Kandel, 1991 Folie 8 zeigt stark vereinfacht und schematisiert den neuronalen Aufbau des Reflexbogens, der in Wirklichkeit allerdings noch über Interneurone moduliert wird, die hier der Einfachheit halber weggelassen sind. Eine taktile Stimulation des Syphons führt zu einer Erregung im sensorischen Neuron, die auf an einer Synapse auf das motorische Neuron umgeschaltet wird, das dann den Rückzug des Kiemens auslöst. Folie 8 Reflexbogen (Kiemenrückzugsreflex bei Aplysia) Sensorisches Neuron Syphon Motorisches Neuron Kiemen Syphon taktile Stimulation Kiemen n. Kandel, 1991 Stimuliert man den Syphon mehrmals hintereinander so schwächt sich der Reflex ab, bis schließlich gar keine Reaktion mehr erfolgt. Nach einem Durchgang mit zehnmaliger Stimulation bleibt der Reflex für einige Minuten unterdrückt, nach mehreren solcher Durchgänge bleibt der Reflex gar für einige Wochen unterdrückt bzw. abgeschwächt. Dieses Phänomen bezeichnet man als Habituation. Der kurzfristigen, für einige Minuten anhaltenden Habituation liegt eine verminderte Transmitterausschüttung an der Synapse zugrunde, während die Ursache der langfristigen Habituation in einer verminderten Synapsendichte zwischen sensorischen und motorischen Neuronen zu suchen ist (siehe unten). Folie 9 Kiemenrückzugsreflex bei Aplysia Sensorisches Neuron Habituation Syphon Motorisches Neuron Kiemen Kiemen Syphon taktile Stimulation n. Kandel, 1991 Der Kiemenrückzugsreflex kann aber nicht nur abgeschwächt werden, er kann auch verstärkt werden (Folie 10). Dieses Phänomen ist dann zu beobachten, wenn der Schwanz der Schnecke etwa durch einen leichten Elektroschock stimuliert wird, was allein noch zu keiner Reaktion am Kiemen führt. Wird danach allerdings der Syphon gereizt, so kann am motorischen Neuron eine im Vergleich zu einem Kontrolltier viel größere Erregung festgestellt werden. Der Grund für dieses als Sensitivierung bezeichnete Phänomen sind fördernde Interneurone, die durch die Reizung des Schwanzes aktiv werden und deren Aktivität an der Synapse zwischen sensorischem und motorischem Neuron dazu führt, daß mehr Transmitter ausgeschüttet wird. Auch bei der Sensitivierung lassen sich solche kurzfristig wirksamen von langfristig wirksamen Mechanismen unterscheiden. Letztere umfassen wiederum eine Veränderung in der Zahl der Synapsen, die sich diesmal jedoch vermehren (siehe unten). Folie 10 Kiemenrückzugsreflex bei Aplysia Sensorisches Neuron Sensitivierung Syphon Förderndes Interneuron Motorisches Neuron Schwanz Kiemen Sensorisches Neuron Versuchstier Kontrolle Syphon taktile Stimulation Kiemen Elektroschock n. Kandel, 1991 Neben dem Nachweis der neuronalen Grundlagen von Habituation und Sensitivierung ist es auch gelungen, die Mechanismen aufzuklären, die der klassischen Konditionierung zugrunde liegen. Dazu wählte man als konditionierten Stimulus (CS) die Stimulation des Mantels, mit der ebenfalls zuverlässig ein Kiemenrückzugsreflex ausgelöst werden kann. (Dieses Beispiel zeigt die Problematik des Begriffes „neutraler“ Stimulus in der „traditionellen“ Terminologie, der hier natürlich keinen Sinn macht. Vielmehr geht es wie oben bereits angedeutet wurde, um die Verbindung zweier Ereignisse, wobei als konditionierter Stimulus derjenige Reiz bezeichnet wird, der nach dem Lernvorgang zu anderen Konsequenzen führt als vorher.) Als unkonditionerten Stimulus wählte man die Stimulation des Schwanzes (wie im Beispiel der Sensitivierung), die als unkonditionierte Reaktion eine Aktivität der fördernden Interneurone zur Folge hat. Reizt man zuerst den Mantel (CS) und kurz darauf den Schwanz (UCS) so ist die Erregung des nachgeschalteten Motoneurons bei einer darauffolgenden alleinigen Reizung des Mantels höher, als ohne die vorhergehende Konditionierung. Dies kann leicht überprüft werden, indem man den Syphon stimuliert, der über eine analoge Verschaltung ja ebenfalls den Kiemenrückzugsreflex auslösen kann. Die verstärkte Übertragung, die hier als Konvergenz bezeichnet wird, läßt sich tatsächlich nur dann beobachten, wenn (wie für die klassische Konditionierung nicht anders zu erwarten) der CS dem UCS unmittelbar vorausgeht. Im Unterschied zur Sensitivierung, die überall dort, wo die Synapsen der fördernden Interneurone enden, die Übertragung verstärkt, beschränkt sich die Verstärkung bei der klassischen Konditionierung auf die Synapsen, die durch den vorausgegangenen konditionierten Stimulus aktiviert wurden. In „technischer“ Hinsicht könnte man die für die klassische Konditionierung verantwortlichen Mechanismen also als eine Art ausgefeiltere „Weiterentwicklung“ der Sensitivierungsmechanismen sehen, was den Gedanken nahe legt, daß komplexere Lernmechanismen auf einfachen aufbauen. Folie 11 Klassische Konditionierung (Kiemenrückzugsreflex bei Aplysia) Sensorisches Neuron (CS) Konvergenz Mantel förderndes Interneuron (UCS) Sensorisches Neuron Schwanz Kiemen Motorisches Neuron Syphon (Kontrolle) (Kontrolle) Sensorisches Neuron n. Kandel, 1991 Wie mehrfach bereits erwähnt wurde erfolgt die kurzfristige Abschwächung bzw. Verstärkung der synaptischen Übertragung über eine Regulation der Transmitterausschüttung, die langfristige Regulation dagegen über eine Reduktion bzw. Verstärkung der Synapsendichte (Folie 12), die auch morphologisch nachweisbar ist, wie Folie 13 zeigt. Übrigens ist diese synaptische Plastizität nicht auf so einfache Nervensysteme wie das von Aplysia beschränkt. Auch im ZNS des Menschen spielt sie eine große Rolle. So konnte z.B. gezeigt werden, daß die Alzheimersche Krankheit, bei der ja ein Verlust vor allem von Gedächtnisfunktionen im Vordergrund steht, mit einer rapiden Abnahme der Synapsendichte einhergeht. Überhaupt scheint das Gehirn, entgegen langjährigen Annahmen, zumindest in bestimmten Regionen, ein äußerst veränderliches Gebilde zu sein, in dem in Abhängikeit von den körperlichen und geistigen Aktivitäten seines Trägers bis ins hohe Alter ständig synaptische Umbauprozesse stattfinden. Folie 12 Zelluläre Grundlagen des Lernens kurzfristig langfristig Veränderung der Transmitterausschüttung v.a. durch Modulation von Ionenkanälen Proteinsynthese und Neusprossung von Synapsen Folie 13 synaptische Plastizität sensorisches Neuron motorisches Neuron LangzeitHabituation LangzeitSensitivierung Bailey & Chen, Science (220): 91-93, 1983 Literatur: - Kandel ER, Schwartz JH, Jessel TM: Principles of Neural Science. (Mc Graw Hill) 42000. - Lachnit H: Assoziatives Lernen und Kognition. Heidelberg (Spektrum Akademischer Verlag) 1993. - Margraf J: Lehrbuch der Verhaltenstherapie. Bd. 1: Grundlagen, Diagnostik Verfahren, Rahmenbedingungen. Berlin, Heidelberg, New York (Springer) 22000.