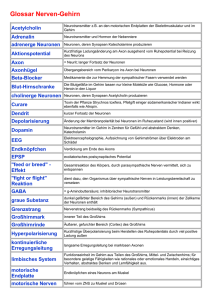
Brain Modelling
Werbung

BRAIN MODELLING physikalische Modelle über das Gehirn BRAIN MODELLING physikalische Modelle für das Gehirn und Gedächtnis nach einer Vorlesung von W.Gruber im Jahr 2000/01 Inhaltsverzeichnis: Historischer Überblick Biologische Neuronen Elektrische Potentiale in biologischen Netzwerken Die chemische Übertragung zwischen zwei Neuronen Technische neuronale Netzwerke Lernen - synaptische Plastizität Repräsentation von Objekten Gestaltspsychologie Entwicklung des Nervensystems Sehsystem Geräuschlokalisation bei der Schleiereule Das Riechsystem Die Großhirnrinde Sprache und Sprachverständnis Steuerung und Regelung Der Thalamus Das Arbeitsgedächtnis Der Schlaf Das Gedächtnis Neurotransmitter Der kranke Geist Künstliche Intelligenz Synthetische Psychologie Spieltheorie 4 12 16 32 37 49 55 62 64 69 78 80 85 90 98 100 101 102 105 113 114 120 121 123 Literaturverzeichnis 125 "Die vielleicht letzte wissenschaftliche Grenze - die ultimative Herausforderung - besteht darin, die biologische Basis des Bewußtseins und der geistigen Vorgänge, durch die wir wahrnehmen, handeln, lernen und uns erinnern, zu verstehen." Eric Kandel Das menschliche Gehirn stellt sicher eines der komplexesten Systeme in der Natur dar. Es gibt verschiedene Ansätze, dieses System zu beschreiben. Die Übergänge zwischen den einzelnen Fachgebieten und Methoden sind kontinuierlich. Das Gebiet der Hirnforschung ist äußerst interdisziplinär und somit ist es notwendig, auf die jeweiligen Fachgebiete einzugehen und die einzelnen Fakten und Theorien aus verschiedenen Gebieten gegeneinander aufzuwiegen und zu überprüfen. Wie kommt die Physik dazu sich mit dem Gehirn zu beschäftigen ? Nun, zum einen ist die Physik eine der elementarsten Naturwissenschaften, die schon in vielen anderen Arbeitsbereichen interessante Themenbeiträge liefern konnte (Erwin Schrödinger; Was ist Leben ?). Zum anderen ist das klassisch physikalische Gebiet der komplexen dynamischen Systeme (Chaos- und Systemtheorie) geradezu prädestiniert, Phänomene der Selbstorganisation des Gehirns zu beschreiben. Leider gibt es noch keine konsistente physikalische Theorie, die das Gehirn, Bewußtsein, Lernen und so weiter beschreibt. In das Gebiet der Neurowissenschaft fließen Ergebnisse aus den verschiedensten Wissenschaftsdisziplinen. Natürlich ist es notwendig, die jeweiligen Fachsprachen zu sprechen, um das arbeiten mit unterschiedlicher Literatur zu erleichtern. Dieses Skriptum sollte helfen, ein fundiertes Wissen aus den verschiedensten Bereichen aufzubauen. Dort wo es möglich ist, werden auch die jeweiligen physikalischen Modelle, beziehungsweise die Grenzen verdeutlicht. Die Physik konnte sehr viele interessante Gedanken zur Gehirnforschung beitragen. Der Physiker Gottfried Wilhelm Leibniz wollte Denken und Wissen überhaupt auf das Rechnen zurückführen, um dann alle wissenschaftlichen Probleme letztlich durch Rechenmaschinen lösen zu können. John von Neumann hat später auf die Analogie zwischen den binären Computern und der Signalübertragung im Nervensystem hingewiesen. Aufgrund der raschen Computerentwicklung in den letzten Jahrzehnten entstand ein neues Teilgebiet der Physik: die computative Physik. Es wurden neue numerische Verfahren entwickelt und dadurch neue physikalische Phänomene (Chaos, neuronale Netzwerke, seltsame Attraktoren und so weiter) entdeckt beziehungsweise neue Lösungen gefunden. Viele dieser Experimente unterliegen dem Selbstorganisationsprinzip, wie etwa Bénard-Zellen oder Spingläser. Durch verschiedenste physikalische Methoden ist es möglich, Erklärungen zu liefern und Vorhersagen zu treffen. Nachdem Christoph von der Malsburg eine Theorie zur Informationsverarbeitung im Gehirn aufgestellt hat, die dem Selbstorganisationsprinzip zugrunde liegt, hat es viele interessante Ansätze gegeben, um diese Theorie zu beweisen beziehungsweise zu widerlegen. Erst 1984 ist es Wolf Singer gelungen, dieses Prinzip in medizinischen Studien zu beweisen. Durch verschiedenste Ansätze der physikalischen Beschreibungsweise versucht man dieses Modell zu verbessern, um mehr Erkenntnisse über das menschliche Verhalten zu gewinnen. Ein Ansatz besteht darin, einzelne Neuronen als "integrate-and-fire"-Oszillatoren zu betrachten. Durch eine sinnvolle Kopplung zwischen diesen Oszillatoren ist es möglich, viele Phänomene aus dem Bereich der Neurowissenschaft physikalisch zu beschreiben. Interessanterweise können die Ergebnisse der Neurophysik auch in anderen Gebieten, wie der Festkörperphysik oder Hydrodynamik, angewendet werden. Die heutige Neurowissenschaft entwickelte sich aus der klassischen Medizin und der Psychologie. Es ist wichtig zu wissen, welches Gebiet für welche Probleme, bzw. Lösungen verantwortlich ist. So ist einem Mediziner die Wirkung einer Behandlung wichtiger, als die exakte mathematische Formel, die die Erkrankung beschreibt. Heute sind folgende ___________________________________________________________________________________ Brain Modelling 1 Wissenschaften mit folgenden Problemen beschäftigt: Die Biologie beschäftigt sich unter anderem mit den einzelnen Zellen, deren Aufbau und der Evolution unterschiedlicher Strukturen des menschlichen Gehirns. Der Bereich der Informatik ist wichtig um verschiedenen Modelle auszutesten. Durch die Informatik sollte die Software "intelligenter" werden. Die Mathematiker stellen Verfahren zur Berechnung komplexer Probleme zur Verfügung. Ohne das Wirken von Henry Poincaré wäre es nicht möglich gewesen, die Selbstorganisation im Gehirn zu beschreiben. Die Medizin versucht organische Schäden zu heilen. Bei unmittelbaren Problemen, die spontan entstehen und eine nachweislich organische Ursache haben, können sie durch verschiedene Therapien helfen. Die Neurologie im speziellen beschäftigt sich unter anderem mit den chemischen Vorgängen im menschlichen Gehirn. Die beiden französischen Psychiater Jean Delay und Pierre Deniker deckten die antischizophrene Eigenschaft von Chlorpromazin auf, Julius Axelrod entdeckte, auf welchem Weg die meisten Botenstoffe im Gehirn inaktiviert werden. Heute versucht eine Vielzahl von Biochemikern, die chemischen Vorgänge im Gehirn zu verstehen und neue Medikamente zu entwickeln. Damit wird es zum Beispiel möglich, sinnvolle Therapien gegen die Abhängigkeit von Suchtmittel durchzuführen. Die Biochemiker beschäftigen sich aber auch immer mehr mit den biochemischen und genetischen Vorgängen in den Neuronen. Dies führte zu äußerst interessanten Erkenntnissen auf dem Gebiet des zellulären Lernens. Natürlich werden die Mediziner kräftig von den Pharmazeuten unterstützt. Die Pharmazie stellt verschiedenen Medikamente bereit. Eine wesentliche Frage beschäftigt sich vor allem damit, wie bringt man das Medikament durch die Blut-Hirn Schranke. Die Psychologie versucht bei persönlichen Problemen des Lebens mit der Theorie des menschlichen Verhaltens zu helfen. Viele Verhaltensweisen sind stark strukturiert und unterliegen einer eigenen Dynamik. Leider sind die Regeln des menschlichen Verhaltens noch nicht naturwissenschaftlich herleitbar. Durch die Technik im Allgemeinen entstehen bessere Computer die leistungsfähigere Programme erlauben. Aber auch der Zusammenhang zwischen Technik und Biologie wird immer wichtiger. So seien hier die Neuroprothesen erwähnt, die teilweise schon ganz excelente Leistungen erbringen (Cochlea-Implantat). Die Physik, wie schon oben beschrieben versucht komplexe dynamische Systeme zu erklären und zu berechnen. Und gibt es ein komplexers und dynamischeres System als das menschliche Gehirn ? Natürlich können die einzelnen Gebiete nicht wirklich voneinder getrennt werden. Jedes Gebiet profitiert von den Ergebnissen anderer. ___________________________________________________________________________________ Brain Modelling 2 Die Neurowissenschaften können aber nicht nur in verschiedene Teilbereiche untergliedert werden. Es existieren verschiedene Ebenen der Betrachtung: Körper Immunsystem ZNS PNS Systeme von Neuronen 10 cm Karten von Neuronen 1 cm Gruppen von Neuronen 1 mm Neuronen 100 m Synapsen 1 m Moleküle 1 Å Die Pharmazeuten beschäftigen sich mit den Molekülen, die Biologen beschäftigen sich mit den Synapsen und für den Mediziner ist es wichtig, wie die Moleküle auf die Synapsen einen Einfluß nehmen. Dadurch, daß die Synapsen die elektrischen Eigenschaften der Membran des Neurons verändern, kommt es zu einer veränderten Reizweiterleitung. Diese Reizweiterleitung läßt sich sehr gut durch die Diffusionsgleichung (Hodgkin-Huxley) beschreiben. Wenn mehrere Neuronen - durch Synapsen verbunden sind - dann spricht man von einer Gruppe. Diese Gruppe (rund 1000-10.000 Neuronen) ist schon in der Lage einfache Probleme zu lösen. Auf dieser Ebene ist das Phänomen der Synchronisation von wesentlicher Bedeutung. Wenn mehrere Gruppen von Neuronen benachbart sind, spricht man von einer Karte. Eine Karte analysiert Muster einer Sinnes- oder Assoziationsmodalität. Diese Karten sind für die Sinnes-Physiologie und Psychologie von zentraler Bedeutung. Sie entstehen in sensiblen Phasen der Entwicklung von den Individuen. Wenn mehrere Karten (Neuronen auf der Großhirnrinde) mit Kernen (moduliernde Strukturen im Inneren des Gehirns) sich mit einer Aufgabe beschäftigen, so spricht man von einem System. Das motorische System umfaßt verschiedene Strukturen: primär motorischer, prämotorischer und supplemäntär motorischer Cortex, die Basalganglien, Teile des Thalamus, und die Sustantia nigra - ein modulierender Kern. Alle Systeme gehören dem Zentralnervensystem (ZNS) an. Man sollte aber nicht vergessen, daß auch das Rückenmark zum Zentralnervensystem gehört. Ein Nerv des Zentralnervensystems, der durchtrennt wurde, wächst nicht mehr zusammen. Für Nerven des peripheren Nervensystems (PNS) gilt dies nicht. Im Bereich des peripheren Nervensystem findet in der Regel auch keine Reizverarbeitung statt. Das Nervensystem dient dazu, rasch Signale aus der Umwelt und dem Körper zu verarbeiten und in geeigneter Weise zu reagieren. Ein anderes informationsverarbeitendes System ist das Hormonsystem. Die Verarbeitungsgeschwindigkeit ist etwas langsamer und die Zielorientierng nicht so präzise. Aber einzelne Hormone, die als Reaktion auf äußere Einflüße ausgeschüttet werden, wirken wiederum auf das Gehirn zurück. So können Synapsen durch einzelne Hormone absterben und der Kreis schließt sich wieder. ___________________________________________________________________________________ Brain Modelling 3 Meist sprechen Laien beim Gehirn nur von der Großhirnrinde. Aber die Großhirnrinde ist nur ein Teil. Unter der Rinde im Zentrum des Schädels befinden sich viele kleine anatomische Strukturen. Diese Strukturen werden als Kerne bezeichnet. Sie steuern den Informationsfluß, während die Rinde die Information verarbeitet und speichert. 1.1 Historischer Überblick Die Medizin beschäftigte sich schon sehr lange mit dem Gehirn, um den Menschen Linderung von seinen Qualen zu bringen. So sind uns schon Symptome, Diagnosen und Prognosen über Kopfverletzungen aus dem alten Ägypten überliefert. Im Edwin Smith Surgical Papyrus aus dem 17. Jahrhundert vor Christus erschienen zum ersten Mal die alten ägyptischen Hieroglyphen für das Wort »Gehirn«. Im alten Griechenland versuchten die Philosophen religiöse Definitionen durch wissenschaftliche zu ersetzen: Anaximenes (585-525 v.Chr): Heraklith (550-480 v.Chr.): Die Seele besteht aus Luft. Die Seele besteht aus Feuer. Etwas konkreter formulierte Empedokles (490-430 v.Chr.) seine Gedanken: das Herz ist Zentrum des Denkens, Fühlens und Handelns. Eine gegenteilige Vermutung äußerte Alkmaion von Kroton (500 v.Chr.), der das Gehirn als zentrales Denkorgan ansah. So wurden zwei Hypothesen geboren: die Herz- und Hirnhypothese. Aristoteles (384-322 v.Chr.), ein Anhänger der Herzhypothese, vermutete, daß das Gehirn zur Kühlung des Blutes dient. So versuchte Aristoteles einen Zusammenhang zwischen der Körpergröße und dem Gehirngewicht herzustellen. Tatsächlich gibt es diesen Zusammenhang - aber aus anderen Gründen. Wenn ein Organismus komplex genug wird, benötigt er auch ein besseres Informationsverarbeitungssystem. Aber aus der Größe oder dem Gewicht des Gehirns läßt sich nur sehr wenig über die Funktionsfähigkeit beziehungsweise über die Komplexität ___________________________________________________________________________________ Brain Modelling 4 aussagen. Zum Beispiel haben Frauen ein leicht geringeres Gewicht des Gehirns. Man stellte aber fest, daß die Zahl der Neuronen praktisch gleich sind - nur daß die Neuronen der Frauen eine Spur kleiner und damit auch leichter sind. Hirngewicht [g] 104 Elephant Mensch Blauwal 103 Australopithecus Säuger 102 10 Strauß Alligator Primaten Kolibri Reptilien 1 Aal Maulwurf 10-3 10-2 10-1 1 10 Körpergewicht 102 103 104 [kg] Relation zwischen Körpergewicht und Hirngewicht: In Griechenland glaubte man, daß bei epileptischen Anfällen Götter und Dämonen in den Körper einfahren. Deshalb betrachtete man diese Krankheit als heilig, denn nur wenige "Auserwählte" wurden von den Göttern besucht. Auch die Araber verehrten die Betroffenen von geistigen Erkrankungen, denn sie seien in der Gnade Gottes bevorzugt. Der berühmte Gladiatorenarzt Galen (129-199 v.Chr.) konnte zeigen, daß die Nerven zum Gehirn und nicht zum Herzen führen. Er konnte zeigen, daß ein Druck auf bestimmte Regionen des Gehirns dazu führt, daß Bewegungen reversibel unterbrochen sind. Hingegen ein Druck auf das Herz führt zwar zu massiven Schmerz, aber es gibt keinen Einfluß auf willentliches Verhalten. Nur noch in der Literatur und dergleichen ist das Herz, das Zenrum der Emotionen und des Fühlens und Denkens. Der Philosoph Rene Descartes (1596-1650) warf ein interessantes Problem auf: Der Körper ist materiell und hat eindeutig eine begrenzte räumliche Ausdehnung. Er reagiert reflexiv auf sensorische Reize aufgrund der Aktivität des Gehirns. Der Geist beziehungsweise die Seele wirkt auf den Körper und erzeugt so ein Verhalten. So stellen stellen sich einige Fragen: Gibt es einen Geist beziehungsweise eine Seele ? Wie interagiert der Leib und die Seele miteinander ? Descartes meinte der Geist sei unteilbar. Deswegen können auch die Gehirnfunktionen nicht unterteilt werden und die Analyse des Körpers kann nicht die Funktionalität des Geistes erklären. Nach heutigen Erkenntnissen ergeben sich neue Aspekte. Unser Gehirn besteht aus zwei Hirnhälften, die größtenteils symmetrisch zueinander sind. Früher war es bei Reflex für eine Hitzereiz, der einerseits Epilepsiepatienten manchmal notwendig, die ein Zurückziehen des betroffenen beiden Hirnhälften anatomisch voneinander zu Körperteils bewirkt und auch die trennen. Es zeigte sich, daß sich die beiden Aufmerksamkeit durch die Augen Hälften nach der Operation individuell steuert. ___________________________________________________________________________________ Brain Modelling 5 entwickelten. Man kann durchaus von zwei Gehirnen in einem Körper sprechen. Hat der Mensch nun zwei Seelen ? So ist für Descartes die Zirbeldrüse (Epiphyse) der Sitz der Seele, denn die Zirbeldrüse ist der einzige Bereich im Gehirn, der nicht bilateral vorhanden ist. Eine Schädigung der Epiphyse führt aber nicht zu offenkundigen Verhaltensänderungen. Man vermutet, daß sie für jahreszeitliche Rhythmen verantwortlich ist. Descartes beschäftigte sich aber auch mit "einfacheren" Problemen. So prägte er den Begriff des Reflexes. In der aktuellen Forschung beschäftigt man sich mit dem körperlichen Anteil des Erlebens. Über das scheinbar "geistige" ist es schwierig Aussagen zu treffen. Wäre das Geistige etwas immaterielles, wie könnte es dann auf materielles einen Einfluß haben. Manche Forscher setzen den Geist mit dem Gehirn und all den damit verbundenen Aktivitäten gleich. J.Locke (1632-1704) meinte, daß das Bewußtsein eine leere Tafel (tabula rasa) sei, auf der unsere Wahrnehmungen ihre Spuren hinterlassen. Damit stellen sich einige interessante Fragen. Wie starke sind die Gene beziehungsweise die Umwelt an der Intelligenz und dem Bewußtsein beteiligt. Eine Frage, die bis heute äußerst kontroversiel diskutiert wird. Interessanterweise hat Locke auf die Entwicklung des Menschen insbesonders des Nervensystems angesprochen: der Mensch ist noch nicht voll entwickelt, wenn er auf die Welt kommt, sondern viele seiner Vorlieben, Ekelgefühle und auch die Sprache entstehen durch die Sozialisation. Die Wissenschafter Leibniz, Pascal und Schickard versuchten eine Mechanisierung des Denkens herbeizuführen. Es ging darum intelligente Handlungen durch ein mechanisches System nachzubilden. Damals wurde das Rechnen mit Zahlen als eine intelligente Handlung angesehen (wer konnte damals schon rechnen ?). So versuchte man des Rechnen mit Zahlen durch Maschinen zu bewerkstelligen. Heute bezeichnet man einen guten Schachspieler als intelligent (zumindest machte man dies solange er nicht von einem Computer geschlagen wurde). Der Versuch eine Rechenmaschine zu bauen führte zu der Einführung des Dualsystems. Wenn zwei Zahlen verknüpft werden sollen, dann müssen zuerst die einzelnen Ziffern verknüpft werden. Betrachten wir die Aufgabe: 15 + 28 = __. Als erstes müssen die Ziffern 5 und 8 zusammengezählt werden und dann die Ziffern 1 und 2, wobei auf den Übertrag nicht vergessen werden darf. Also ist es notwendig für alle Ziffern (1; 2; 3; 4; 5; 6; 7; 8; 9; 0) alle Elementaroperationen (Addition, Subtraktion, Multiplikation und Division) zu definieren. Dies sind über einige hundert Rechenoperationen. Durch die Einführung des Dualsystems läßt sich der Rechenaufwand drastisch reduzieren. Die Zahlen, mit denen gearbeitet werden muß, werden in das Dualsystem umgewandelt und dort gibt es gibt nur mehr zwei Ziffern (0 und 1) und für die Addition ergeben sich 4 Rechenoperationen: 0+0= 0 0+1= 1 1+0= 1 1 + 1 = 10 Dies führt zu einer drastischen Vereinfachung der Rechnungen. Die ersten Maschinen konnten dann tatsächlich rechnen, wenngleich die Mechanik ziemlich klobig war. Dem Nachbau des Gehirns war man aber nur einem kleinen Schritt näher gekommen. Erst durch die Einführung der Elektronik war es möglich, leistungsfähigere Maschinen zu bauen, mit denen es sogar gelungen ist, einzelne Bereiche des Gehirns zu simulieren. ___________________________________________________________________________________ Brain Modelling 6 Gregor Reisch, Prior der Kartause in Freiburg, vermutete daß die Seele in den 3 Gehirnkammern (Ventrikel) ansäßig sei. So soll in der ersten Kammer der Sitz für den Gemeinsinn, die Phantasie und das Imaginationsvermögen sein. Die 2. Kammer ist für das Denken und Urteilen wichtig und die dritte Kammer für das Erinnern. Zwischen der Kammer 1 und der Kammer 2 befindet sich eine Schleuse. Dadurch können die unterschiedlichen "Seelenbereiche" Informationen austauschen. Beindruckt durch die Pneumatik, der damaligen Spitzentechnologie, entstanden die Ballontheorien. Das Gehirn ist nichts anderes als ein großer Blasebalg und eine Flüssigkeit wird über die Neuronen in die Muskeln geblasen. So können Muskeln kontrahieren und erschlaffen. Diese Theorie war auch von Blutkreislauf motiviert, wo Blut durch den ganzen Körper strömt. Erst 1677 konnte Francis Glisson mit einem einfachen Experiment zeigen, daß die Ballontheorie falsch ist. Man braucht nur den Oberarm in Wasser eintauchen und die Veränderung des Wasserspiegels messen, wenn der Muskel erschlafft beziehungsweise angespannt ist. Wenn die Pneumatheorie richtig wäre, dann müßte sich der Wasserspiegel ändern - was er aber nicht tut. Auch der Philosoph Immanuel Kant (1724-1804) äußerte sich zum Thema der Sensorik: "Wahrnehmung sei kein passiver Empfang von Sinneseindrücken, sondern es handle sich dabei um einen aktiven Prozeß, der in spezifischer Weise durch den menschlichen Wahrnehmungsapparat hergestellt und organisiert wird." Damit wird ein wesentlicher Aspekt der menschlichen Organisation der Sensorik charakterisiert. Durch das Aufmerksamkeitssystem werden die einlangenden Informationen gefiltert und gesteuert. Die Steuerung scheint durch wesentliche Bereiche der Formatio Reticularis durchgeführt zu werden. Während dieser ganzen Zeit wurden Personen mit einer Geisteserkrankung durch fahrende Chirurgen behandelt. Sie entfernten sogenannte "Narrensteine" aus dem Kopf des Betroffenen. Praktisch wurde aber nur die Kopfhaut eingeritzt und mit etwas schauspielerischem Geschick ein Stein hervorgeholt. Geholfen wurde nur dem Chirurgen, für den diese Operationen eine lukrative Einnahmequelle darstellten. All die bisher genannten Wissenschafter haben sich "nur" mit Teilsaspekten des menschlichen Gehirns beschäftigt. Wichtig ist aber eine umfassende Theorie des menschlichen Gehirns. Diese Theorie wurde von Franz J. Gall (1758-1828) und Johannes C. Spurzheim (1776-1832) in Wien geschaffen. Beide konnten zeigen, daß das Gehirn aus lebenden Zellen besteht und daß manche Zellen des Gehirns in das Rückenmark projezieren. Beide stellten fest, daß Studenten mit gutem Gedächtnis große hervortretende Augen haben. Also - so die neue Idee müsse sich das Gehirn hinter den Augen besonders gut entwickelt haben und drücke nun auf die Augen. Wenn also manche Gebiete besonders gut entwickelt sind, dann drücken sie auch auf die Schädeloberfläche und sorgen so für Erhöhungen und Vertiefungen des Schädelknochens. Damit war die Phrenologie geboren. Diese damalige Idee führt zu einigen interessanten Schlußfolgerungen. Eine der Wichtigsten war die Lokalisation von Denkleistungen und daß aus geistigen Denkleistungen biologische Korrelate folgen. ___________________________________________________________________________________ Brain Modelling 7 Durch Vermessung des Schädelknochens glaubte man nun auf bestimmte "geistige" Denkprozesse rückschliessen zu können. Münze des Phrenologen F.J. Gall (links) und eine Darstellung der verschiedenen Areale des Schädelknochens, die mit Denkprozessen verbunden sind (links). Aber es gab einige Probleme mit dieser Theorie. Zum einen wurden nicht eindeutige Begriffe für die unterschiedlichen Denkleistungen verwendet. Glaube, Selbstliebe, Verehrung sind nur sehr schwer meßbar und es gibt keine eindeutigen Definitionen dafür. Zum anderen wurde auch nicht überprüft, ob die Schädeloberfläche tatsächlich ein Abdruck der Innenseite sei. Den meisten Spott mußten die beiden aber durch die Annahme, daß es keinen Geist gäbe, sondern daß alles angeboren sei, hinnehmen. Heute wissen wir, daß sich die Phrenologie als falsch herausgestellt hat. Aber es war die erste in sich konsistente Theorie und die Annahme, daß Denken ein naturwissenschaftlicher Prozeß sei, daß es besondere spezifische Gebiete des Gehirns gibt, war revolutionär. Dadurch, daß die gesamte Theorie massive Mängel aufwies und in Verruf geriet, hatten nachfolgenden Wissenschaftler die tatsächlich Lokalisationen im Gehirn (nicht auf der Schädeloberfläche) festgestellt hatten, beträchtliche Probleme. Pierre Flourens (1794-1867) versuchte die Phrenologie zu demontieren. So entwickelte er die Läsionstechnik. Tauben wurde der Schädel geöffnet und ein kleines Stück des Gehirns entfernt. Danach beobachtete man die jewiligen Verhaltensstörungen. So fand er ein Zentrum für die Atemtätigkeit und stellte fest, daß das Kleinhirn für die Bewegungskoordination verantwortlich ist. In den Untersuchungen fand man, daß nur rund 10% des Gehirns wichtig sind. Aber diese Ausssage ist falsch. Man beobachtete die Tiere nicht in einer natürlichen Umgebung. Man konnte feststellen, ob die Tauben und Hühner in der Lage waren selbstständig zu fressen und ob sie mit den Flügeln schlagen konnten. Heute wissen wir, daß jeder Kubikmilimeter Gehirn wichtig sind. Aber manche Areale des Gehirns lassen sich nur durch ausgefeilte psychologische Tests austesten. So stellte Flourens trotz mangelhafter Tests folgende Dogmen auf: 1) Der Cortex ist nicht erregbar. 2) Der Cortex spielt keine Rolle bei der Erzeugung von Bewegung. 3) Einzelne Funktionen sind nicht lokalisiert. Ein besonders Areal konnte Paul Broca (1824-1880) entdecken. Broca interssierte sich für die Sprechfähigkeit von Menschen. Er fand eine Gruppe von Patienten, die etwas gemeinsam hatten. Die Patienten hatten zwar ein gutes Wortverständnis und der Stimmapparat war in Ordnung, trotzdem konnten sie nicht sprechen. So nahm er Autopsien an dieser Patientengruppe vor. Es zeigte sich, daß praktisch alle Patienten in einer speziellen Region der Großhirnrinde eine Läsion aufwiesen. Dieses Zentrum ist für die motorische ___________________________________________________________________________________ Brain Modelling 8 Sprachsteuerung notwendig. Mit diesen Befunden wurde Broca in das Eck der Phrenologen gestellt. Aber spätere Studien bestätigten seine Befunde und das von ihm gefundene Areal wird ihm zu Ehren als Broca-Areal bezeichnet. Luigi Galvani (1737-1798) versuchte das Fluidums des Lebens zu ergründen. So entdeckte er, daß Froschschenkel durch Elektrizität kontrahiert werden können. Also könnte die Elektrizität für Verhaltensweisen wichtig sein. Gustav T. Fritsch (1838-1929) und Eduard Hitzig (18381907) griffen diesen Gedanken auf und veröffentlichten 1870 eine aufsehenserregende Arbeit mit dem Titel "Über die elektrische Erregbarkeit des Cerebrum". Damit war die Elektrophysiologie geboren. Es konnte gezeigt werden, daß elektrische Reizungen des Gehirn in unterschiedlichen Arealen zu unterschiedlichen Bewegungen und Zuckungen bestimmter Körperteile verursacht. Somit wurde die Lokalisationstheorie auch von einer anderen Seite her bestätigt. Damit wurden die 3 Dogmen von Flourens gestürzt. Trotzdem sprachen sich mehrere Wissenschafter gegen die Lokalisation aus. Zum Beispiel meinte Friedrich L. Goltz (1834-1902), daß es unterschiedliche Arten des Essens, des Gehens gäbe. Also kann es nicht ein Gebiet für unterschiedliche Verhaltensweisen geben. Hughlings und Jackson konnten zeigen, daß das Geirn hierarchisch organisiert ist. Die Information gelangt über das Rückenmark zum Hirnstamm und dann weiter zur Großhirnrinde. Im Rückenmark findet ein einfaches Reiz-Reaktions Verhalten statt. Je höher die Verarbeitungsebene ist, umso komplexer werden die Verhaltensweisen. Camillo Golgi (1843-1926) entwickelte eine neue Färbungstechnik (KaliumBichromat-Fixierung mit anschließender Silber-Imprägnierung), die nur wenige Nervenzellen in einem mikroskopischen Feld anfärbte und dadurch die cytologische Untersuchung von Neuronen wesentlich erleichterte. So fand man unglaublich verschlungene Strukturen, Faserzüge und Kerne. Der Beweis, daß Nervenzellen klar abgegrenzte Einheiten mit einem rezeptiven Ende (Dendriten), einem leitenden Teil Camillo Golgi und Santiago Ramón y Cajal. (Axon) und einem übertragenden Ende (Synapse) sind, gelang Ramón y Cajal. Er zeigte, daß das Nervensystem kein durchgehendes Netzwerk ist, sondern aus einzelnen Nervenzellen - Neuronen genannt - besteht. Ihm war bewußt, daß eine exakte Kenntnis der Gehirnstruktur für den Aufbau einer rationalen Psychologie von überragender Bedeutung ist. Beide bekamen 1906 den Nobelpreis. Mit den heutigen neuen bildgebenden Verfahren ist es möglich, eine gute Kartographie des menschlichen Gehirns durchzuführen. Mit physikalischen Meßmethoden (Computertomographie CT, Magnetspintomographie NMR, Positronen-Emissions-Tomographie PET, Elektroencephalographie EEG) können nicht nur Verletzungen genau lokalisiert werden, sondern neuerdings ist es auch möglich, dem Gehirn unmittelbar beim Denken zuzusehen. Durch die Arbeiten von Golgi und Cajal entstand die Neuronenhypothese. Neuronen sind die elementarsten Einheiten. Sie können miteinander interagieren und dadurch entsteht die gesamte Bandbreite menschlichen Verhaltens. So stellen sich 3 Fragen: 1) Wie werden Signale im Nervensystem weitergeleitet ? 2) Wie ist das Nervensystem aufgebaut ? 3) Wie sind die einzelnen Neuronen untereinander und mit der Muskulatur verbunden ? ___________________________________________________________________________________ Brain Modelling 9 Charles S. Sherington (1857-1952) untersucht im Besonderen die Verbindung zwischen dem Neuron und dem Muskel. So fand er eine wesentliche Untereinheit des Neurons: die Synapse. 1952 veröffentlichten A.L. Hodgkin und A.F. Huxley eine Arbeit, die die Reizweiterleitung im Neuron erklärt: "A quantative description of membrane curent and its application to conduction and excition in nerve." Mit dieser Differentialgleichung ist es möglich die elektrischen und chemischen Eigenschaften der Membran zu beschreiben. Dadurch konnte erklärt werden, wie manche Gifte im Körper wirkten. Für ihre Arbeiten wurden sie 1963 mit dem Nobelpreis ausgezeichnet. Der Begriff Psychologie leitet sich aus dem Griechischen ab (psyché = Seele) und bedeutet die Lehre von seelisch-geistigen Erscheinungen. Die Aufgabe der wissenschaftlichen Psychologie ist es, Gesetzmäßigkeiten des Erlebens und Verhaltens zu erforschen. So wird zum Beispiel das Lernverhalten von Menschen analysiert. Die Psychologie besitzt zwar einen weniger naturwissenschaftlichen Zugang zu den Denkprozessen, aber es gibt sehr viele Modelle für verschiedene geistige Phänomene. 1902 prägte Wundt den Begriff der Biopsychologie. Er hatte große Problem damit sich durchzusetzen. Allerdings wurden große Problem in den letzen 20-30 Jahren gelöst und nun hat auch die Psychologie vermehrt Interesse an den Naturwissenschaften. Auch Simund Freud war ein Anhänger einer naturwissenschaftlichen Theorie des menschlichen Verhaltens. Er mußte aber bald feststellen, daß die Naturwissenschaft noch nicht so weit ist, um die geistigen Phänomene erklären zu können. So schaffte er ein psychologisches Modell vom Verhalten. Er stellte verschiedene Postulate über das Seelenleben auf: 1) Das Seelenleben ist, ähnlich wie in der Physik determiniert, auch wenn die Ursachen nicht immer gleich erkennbar sind. 2) Große Bereiche des Denkens sind unbewußt und emotional gesteuert. 3) Konflikte werden in einem eigenem Bereich (dem Unbewußten) gespeichert. Es treten Widerstände bei der Bewußtwerdung auf, die aber im Traum herabgesetzt Ein Netzwerk, das unterschiedliches Verhalten werden können. beschreibt (Handschrift von S.Freud). 4) Ursachen für Konflikte reichen bis in die Kindheit zurück. Die Kybernetik ist eine übergreifende Wissenschaftsdisziplin, die sich mit der formalen mathematischen Beschreibung und modellartigen Erklärung von dynamischen Systemen beschäftigt. Sie wurde von Norbert Wiener gegründet. Als besonderes Kennzeichen dieser Disziplin gilt das Prinzip der selbsttätigen Regelung und Steuerung durch Übertragung und Verarbeitung sowie Rückübertragung von Informationen in wenigstens einem Rückkopplungssystem. Norbert Wiener ging von Analogien zwischen organischen und technischen Systemen aus. So eignet sich die Kybernetik hervorragend zur mathematischen Modellierung von Gehirnprozessen und den damit verbundenen Verhaltensprozessen. Nicht ___________________________________________________________________________________ Brain Modelling 10 die materiellen Eigenschaften stehen im Vordergrund, sondern die Möglichkeit des Verhaltens. Die Eigenschaften des Systems stehen im Vordergrund. Um das menschliche Gehirn und das damit verbundene Denken, Wahrnehmen und Lernen naturwissenschaftlich beschreiben zu können, bedarf es einer mathematisch-physikalischen Formulierung basierend auf medizinischen, physikalischen, biologischen und chemischen Grundlagen, um psychologische und psychiatrische Modelle zu überprüfen, zu ergänzen oder auch neu zu entwickeln. Wie man sieht gibt es verschiedene Möglichkeiten das Gehirn zu erforschen. Im ersten Teil dieses Skriptums beschäftigen wir uns mit den mikroskopischen lokalen Effekten. Es wird erläutert was ein Neuron ist, wie es funktioniert, wie man es nachbauen kann und so weiter. Diese Phänomene sind die Basis für das weitere Verstehen des Gehirns. Nur wenn man diesen Bereich wirklich verstanden hat, dann ist man in der Lage die makroskopischen globalen Phänomene zu verstehen. Alle makroskopschen Effekte können in der Regel direkt an Patienten beobachtet werden. Meist sind mehrere Subsysteme des Gehirns beteiligt, die das Betrachten noch verkomplizieren. Eine Trennng in diese beiden Bereiche ist natürlich willkürlich und die Übergänge sind fließend. ___________________________________________________________________________ In den Neurowissenschaften gibt es zwei wesentliche Arbeitshypothesen: 1) Gehirnhypothese: Der Ursprung allen Denkens, Verhaltens und Fühlens liegt im Gehirn (geht auf die Ägypter zurück) 2) Neuronenhypothese: Das Neuron ist die kleinste funktionelle Einheit des menschlichen Gehirns. Alle beobachtbaren Phänomene des Verhaltens, Empfindens und Reagierens lassen sich auf Neuronen zurückführen (Santiago Ramon ý Cajal) ___________________________________________________________________________________ Brain Modelling 11 2.0 Biologische Neuronen Nervenzellen, auch Neuronen genannt, sind die wesentlichen informationsvermittelnden Bausteine des Gehirns. Diese Zellen sind nach denselben Grundsätzen aufgebaut wie die übrigen Zellen im ganzen Körper. Sie zeigen aber einige Besonderheiten wie die Zellform, die Art der Zellmembran und die Möglichkeiten, chemische wie elektrische Signale zu empfangen, zu modulieren und weiterzugeben. Der Empfang von solchen Signalen geschieht in der Regel durch Synapsen, die Modulation durch den Zellkörper, und die Weitergabe von Signalen wiederum durch Synapsen. Als zweite Besonderheit von Neuronen ist die fehlende Zellteilung zu erwähnen. Nervenzellen teilen sich nicht mehr nach der embryonalen Entwicklung, das heißt der bis zur Geburt entstandene Vorrat von Zellen muß ein Leben lang ausreichen. Die Abbildung zeigt ein Golgi-Präparat (1911) mit Pyramidenzellen (links) und Purkinjezellen (rechts). Nervenzellen kann man in drei wesentliche Teile aufgliedern: den Zellkörper, die Dendriten und die Nervenfaser oder auch Axon. Der Zellkörper (Perikaryon, Soma), kugelförmig oder auch pyramidenförmig, enthält den Zellkern und den biochemischen Apparat für die Synthese von Enzymen und anderen zellnotwendigen Substanzen. Besonders wichtig ist die Synthese von Membranproteinen. Diese Proteine lassen sich in fünf Gruppen einteilen: Pumpenproteine sorgen unter Energieverbrauch dafür, daß Ionen und andere Moleküle die Membran entgegen einem Konzentrationsgefälle passieren können. Kanalproteine ermöglichen es Ionen und Moleküle, die für sie normalerweise undurchlässige Membran entsprechend einem Konzentrationsgefälle zu passieren. Rezeptormoleküle reagieren mit Molekülen, wie zum Beispiel mit Neurotransmittern oder Neuropeptiden, die dann das Verhalten der Zelle beeinflussen. Zellenzyme beschleunigen die chemischen Reaktionen der Zellmembran und Strukturproteine erhalten die Feinstruktur der Membran aufrecht. Die Wirkung und der Einfluß von einzelnen Proteinen kann übergreifend sein. Die Dendriten sind röhrenförmige Fortsätze des Zellkörpers, die sich vielfach verästeln. Vom Feinbau her gesehen unterscheiden sich die Dendritenfortsätze eines Neurons nicht vom Somabereich, außer daß ihnen das rauhe endoplasmatische Reticulum zur eigenen Proteinsynthese fehlt. Die Dendriten dienen der Vergrößerung der rezeptiven Oberfläche einer Nervenzelle und sind deshalb mit besonders vielen Synapsen übersät. Über diese Synapsen nimmt die Nervenzelle ankommende Signale auf und leitet sie über die Dendriten zum Zellkörper weiter. Dort werden die Signale summiert und bewertet. Zentrale Neuronen haben oft 20 bis 40 Hauptdendriten, die sich in feinere Zweige aufspalten. Auf jedem Zweig gibt es zwei Arten von bevorzugten synaptischen Kontaktstellen: den Hauptschaft, wo direkt Synapsen von anderen Neuronen Informationen übermitteln (meist handelt es sich dabei um hemmende Signale); weiters die Dornen, die selbst Synapsen sind und sich auf dem Hauptschaft oder sich meist auf weiteren Verästelungen befinden (Die dort ankommenden Signale sind in der Regel erregend). Durch die Weiterleitung über die Dendriten kommt es zu einer zusätzlichen Gewichtung von elektrischen Signalen. Sie werden entweder abgeschwächt oder verstärkt. ___________________________________________________________________________________ Brain Modelling 12 Das Axon (Neurit oder Nervenfaser) ist wie ein Dendrit ein Ausläufer des Zellkörpers. Es dient zur Informationsweiterleitung zu anderen Neuronen. Die Übermittlung findet meistens über größere Distanzen statt. Üblicherweise ist das Axon bedeutend länger und dünner als die Dendriten. Das Neurit verzweigt sich erst dort, wo die Teiläste mit anderen Neuronen über Synapsen wieder in Verbindung treten. Diese Verästelungen werden Neuritenbaum, Seitenzweige oder Kollaterale genannt. Schwann-Zellen, eine Sonderform der Gliazellen, umgeben das Axon mit einer Myelinhülle. Jede Schwannzelle umhüllt der Länge nach jeweils rund einen Millimeter des Axons. So ist die Myelinhülle ungefähr alle ein bis zwei Millimeter von einem feinen Spalt, dem Ranvierischen Schnürring, unterbrochen. Eine Pyramidenzelle mit verästelten Fortsätzen, die mit Tausenden von signalaufnehmenden Dornen übersät ist. Rechts sieht man eine erregende Synapsenverbindung (oben) und unten eine hemmende Synapsenverbindung. ___________________________________________________________________________________ Brain Modelling 13 Nervenzellen unterscheiden sich natürlich auch in ihrer Funktion und lassen sich in drei Arten unterteilen: sensorische Neuronen, motorische Neuronen und Interneuronen. Sensorische (oder afferente) Neuronen übermitteln dem Nervensystem Information, die der Wahrnehmung wie auch der motorischen Koordination dienen. Motorische Neuronen, oder auch Motoneuronen, übermitteln Signale an Muskeln und Drüsen. Interneuronen bilden die größte Menge an Neuronen im Nervensystem und sind nicht spezifisch sensorisch oder motorisch. Sie verarbeiten Informationen in lokalen Schaltkreisen oder vermitteln Signale über weite Entfernungen. Die Pyramidenzellen, die zur Klasse der Interneuronen zählen, befinden sich in der Großhirnrinde und besitzen einen apikalen (an der Spitze gelegen, nach oben gerichteten) Hauptfortsatz des Zellkörpers, der sich in zahlreiche Dendriten verzweigt, und ein Axon. Weitere Dendriten entspringen dem Pyramidenmantel. Das Axon kann eine Länge von bis zu einem Meter erreichen. Sternzellen sind der zweitwichtigste Typ von Neuronen im Gehirn. Ihren Namen haben sie von der sternförmigen Struktur der Dendriten. Die Ausläufer bleiben in der Regel in der näheren Umgebung des Zellkörpers. Man unterscheidet zwischen dorntragenden und dornlosen Sternzellen. Die dornlosen Zellen emitieren meist einen Neurotransmitter, der eine hemmende Wirkung auf die Pyramidenzellen hat. Purkinjezellen, wie sie häufig im Kleinhirn vorkommen, sind bipolar gebaute dendritische Neuronen. An ihrem spitzwärtigen Pol entspringt ein verzweigter Riesendendrit, an der Basis der Neurit. a) b) c) d) Die strukturelle Vielfalt der Nervenzellen trägt dazu bei, daß das Gehirn sowohl Informationen aufnehmen, filtern, sortieren, interpretieren, speichern, abrufen, nutzen und mitteilen als auch Gefühle erleben und Bewegungen kontrollieren kann. (a) An einer Stelle, dem Nucleus reticularis lateralis sind die Dendriten kurz und buschig und enden in signalaufnehmenden "Sammelstellen". (b) In der Nähe des Nucleus hypoglossi sind die Dendriten lang, leicht gekrümmt und relativ wenig verzweigt (c) An einer anderen Stelle, dem Nucleus reticularis gigantocellularis einer neugeborenen Katze, ziehen die Dendriten in alle Richtungen und sind dicht mit dendritischen Dornen besetzt. (d) Wenn die Katze fünf Monate alt ist, haben die Dendriten die meisten ihrer Dornen verloren. Die Zellen haben jetzt eine eindrucksvolle Größe. ___________________________________________________________________________________ Brain Modelling 14 Eine dreidimensionale Darstellung eines Neurons, das sechsmal um etwa 10° um die vertikale weitergedreht wurde. Um das Neuron dreidimensional zu sehen, führt man die Nasenspitze zu einer Nahtstelle zwischen zwei Bildern. Danach vergrößert man wieder - langsam - den Abstand und versucht dabei einen imaginären Punkt - weit entfernt - anzuvisieren (viel Glück und es funktioniert wirklich). Im Nervensystem werden die Informationen durch elektrische Impulse, worauf wir noch näher eingehen werden, verarbeitet. Diese Impulse entstehen durch eine elektro-chemische Reizung der Zellmembran. Es entstehen unterschiedliche Pulsfolgen (Aktivitätsmuster) auf die selbe Reizung, da die Struktur der Zellmembran variieren kann. Die Klassifizierung basiert auf drei generellen Variablen: (1) Die Charakteristik des individuellen Aktionspotentials (eines einzelnen Pulses) und das unmittelbar folgende elektrische Verhalten (Hyperpolarisation, Depolarisation usw.). (2) Das Antwortverhalten auf einen intrazellulären Spannungsimpuls in der Größe nahe dem Schwellwert. (3) Das wiederholte Antwortverhalten auf einen länger andauernden intrazellulären Reiz. ___________________________________________________________________________ ___________________________________________________________________________________ Brain Modelling 15 3.0 Elektrische Potentiale in biologischen Neuronen U [mV] t Aktionspotential Schwellwertspannung Ruhemembranpotential U [mV] t Depolarisation (Anhebung des Membranpotentials) Hyperpolarisation (Erniedrigung des Membranpotentials) An der Membran eines Neurons (überall) herrscht ein Ruhemembranpotential. Dies liegt bei -55 bis -90 mV. Der Überträgerstoff aus der Synapse erzeugt eine Potentialänderung ( 0.04-1 mV) an der Membran des nachgeschaltenen Neurons. Das heißt das Ruhemembranpotential kann erhöht (Depolarisation) oder erniedrigt (Hyperpolarisation) werden. Die Ströme aus den Synapsen gelangen über den Dendritenbaum zum Axonhügel und werden dort nichtlinear addiert. Wenn eine gewisse Schwelle (liegt rund 10-20 mV höher als das Ruhemembranpotential) am Axonhügel überschritten wird, dann wird ein Aktionspotential ausgelöst, über das Axon weitergeleitet und am Neuritenbaum des betreffenden Neuron schütten die Synapsen wiederum Überträgersubstanz an andere Neuronen aus. Die Stärke der Ausschüttung von Überträgersubstanz kann verändert werden. Wenn ein Neuron stark gereizt wird, dann feuert es öfters. Es werden mehr Aktionspotentiale ausgelöst. Aufgrund einer unterschiedlichen Membranbeschaffenheit, beziehungsweise einem unterschiedlichen Metabolismus im Zellkern ergeben sich unterschiedliche Aktivitätsmuster bei der Reizung des Neurons: reguläres, schnelles und salvenartiges Aktivitätsmuster. Reguläre Aktivitätsmuster treten bei vielen Neuronen auf. Das Aktionspotential zeigt eine ausgeprägte Phase an Hyperpolarisation und Depolarisation nach dem Aktionspotential. Die Repolarisation findet langsam statt. Bei elektrischer Stimulation des Neurons beim Schwellwert kommt es zu einem Aktionspotential. Bei Neuronen, die ein schnelles Aktivitätsmuster zeigen, findet eine geringere Hyperpolarisation und Depolarisation unmittelbar nach dem Aktionspotential statt. Das Neuron feuert für einige hundert Millisekunden mit 500-600 Hertz bei einer starken Stimulation. Die temporären Eigenschaften des Inputs bleiben im Output sehr gut über einen großen Frequenzbereich erhalten. Neuronen mit salvenartigen Aktivitätsmustern gibt es relativ selten. Wird ein solches Neuron durch einen einzelnen Reiz stimuliert, dann antwortet es mit einer Salve von Aktionspotentialen, deren Amplituden abnehmen. Einzelne Aktionspotentiale zeigen eine markante Nach-Depolarisation, sind aber ähnlich den regulären Potentialen. Die Salven treten rhythmisch mit einer Frequenz von 5-15 Hertz auf. ___________________________________________________________________________________ Brain Modelling 16 elektrisches Antwortverhalten Schwelle Reizspannung (U10 mV) t t t Darstellung eines regulären, eines schnellen und eines salvenartigen Aktivitätsmusters. 3.1 Entstehung des Ruhemembranpotentials Die Membran besteht aus einer Doppelschicht aus Phospholipiden. Sie trennt den intrazellulären vom extrazellulären Bereich (innen und außen). Die Membran ist von ionenspezifischen Kanälen durchsetzt. Das heißt es können nur bestimmte Ionen durch diese Kanäle. Im inneren und äußeren Bereich befinden sich gelöste Ionen: Na+, K+, Cl-. Die organischen Ionen A- (Aminosäuren) befinden sich nur im Inneren des Neurons. ionenspezifischer Kanal außen Doppellipidschicht innen Diese Ionen verteilen sich nun gemäß ihres Konzentrationsgradienten (bis dieser gleich Null wird) im intra- und extrazellulären Raum. Überall sollte die gleiche Konzentration vorherrschen. Es muß auch der elektrostatische Gradient ausgeglichen werden. Wasser Betrachten wir - als den einfachsten Fall - ein Glas Wasser und geben wir eine lösliche Substanz dazu. Durch verschiedenste thermodynamische Effekte verteilt sich diese Substanz die Konzentration ist überall gleich groß - der Konzentrationsgradient ist Null. Jedes System versucht einen Zustand möglichst geringer Energie einzunehmen. Wäre unsere lösliche Substanz elektrisch geladen, so würde sie sich genauso gleichverteilen - der elektrostatische Gradient der Ladungsdichte wäre in dem Behälter gleich Null. ___________________________________________________________________________________ Brain Modelling 17 Teilen wir nun den Behälter durch eine semipermeable Membran und nehmen wir zwei lösliche elektrisch geladene Substanzen. Zum Beispiel stellen die Gliazellen ein so einfaches System dar. Die Gliazellen umhüllen die Neuronen, versorgen sie mit Nährstoffen und sie stellen die Blut-Hirnschranke dar. Dieser Zelltyp verfügt nur über K+-Ionenkanäle. K+ kann durch die Membran diffundieren, während die Aminosäuren A– im Inneren der Zelle bleiben müssen - sie sind zu groß um durch die Ionenkanäle diffundieren zu können. Die Membran läßt nur eine Substanz K+ durch. Durch das Konzentrationsgefälle wird das Kalium versuchen sich sich überall auszubreiten - es diffundiert durch die Membran. Dabei bleiben die negativ geladenen Aminosäuren zurück. Es ergibt sich ein positiver Ladungsüberschuß auf der Außenseite und ein negativer innerhalb der Zelle. Der elektrostatische Gradient führt dazu, daß die Kalium-Ionen außerhalb der Zelle wieder zur Membran getrieben werden. Das System versucht den elektrischen Gradienten auszugleichen. Das K + strömt also dem Konzentrationsgefälle folgend so lange nach außen, bis die elektrostatische Anzeihung so groß ist, daß Rückwanderung und Auswanderung gleich sind und durch den Ausstrom gestoppt wird. So entsteht ein Gleichgewicht zwischen dem elektrischen Gradienten und dem Konzentrationsgradienten. + + + + + außen Aufgrund der Ladungsdifferenz wandern einige Ionen zurück. + Doppellipidschicht innen + + + + - + - + + + + - + + + - außen Doppellipidschicht - ++ + innen - + + + -- - + - + + + + + - - + + + Betrachten wir die K+ Ionen: Da K+ -Ionen im Zellinneren in höherer Konzentration als im extrazellulären Raum vorliegen, diffundieren sie entlang ihres Konzentrationsgefälles vom Zellinneren nach außen. An der Außenseite entsteht eine positive Ladung wegen des leichten Überschusses an positiven Ionen, die Innenseite wird negativer. Weil sich ungleichnamige Ladungen anziehen, sammeln sich die entsprechenden Ionen an den jeweiligen Seiten der Membran: Die Positiven außen, die Negativen innen. Die Diffusion der K + Ionen hört auf, wenn die elektrostatische Kraft und die Kraft aus dem Konzentrationsunterschied sich die Waage halten. Je mehr K+ ausströmt, desto größer wird die Ladungstrennung und damit auch die Potentialdifferenz. Die elektrische Kraft, die sich aufgrund dieses Potentials aufbaut, wirkt der Kraft aus dem Konzentrationsgradienten entgegen. Bei einem bestimmten Potential (Nernst-Potential) befindet sich der Konzentrationsgradient und der elektrostatische Gradient im Gleichgewicht: E k R T K außen ln zF K innen ___________________________________________________________________________________ Brain Modelling 18 EK+ ist das K+ -Gleichgewichtspotential, R (=8.314 J.K-1.mol-1) die allgemeine Gaskonstante, T die absolute Temperatur in Kelvin, z die Wertigkeit von K + (also z=1) und F (=9.648.104.C.mol-1 ) die Faraday-Konstante. RT/F beträgt bei 25°C rund 25 mV. - Na+ Cl Na+ - Cl K Na+ + - A- Cl - Na+ A K+ K+ A- K+ K+ A- Cl - Cl Na+ - Na+ Cl - Na+ - Cl Na+ K+ - Cl ++++ ++++ ---- ---- AK+ K+ A- K+ K+ AA- Für die Membran des Neurons ist der Sachverhalt noch etwas komplizierter. Wir haben nicht nur eine oder zwei sondern mehrere Ionensorten. Das K+-Ion versucht rauszuwandern, das Na+-Ion versucht reinzuwandern - aufgrund des Konzentrationsgefälles (links). Beide Ionentypen werden aber aufgrund des elektrostatischen Gradienten daran gehindert. Wesentlich mehr Chlor befindet sich im Extrazellulärraum, da im Inneren die negativen Ionen überwiegen (A-). Das Chlor kann sich im wesnetlichen relativ frei durch alle Ionenkanäle bewegen und so hat das Chlor keinen Einfluß auf das Ruhemembranpotential. An der Membran bauen sich lokale Wölkchen von geladenen Teilchen auf. Diese Verteilung wird aber durch die Thermodynamik gestört. So werden manche Ionen in den weiteren Extrazellulärraum getrieben. Damit ist das Gleichgewicht zwischen dem Konzentrationsgefälle und dem elektrischen Gradieneten wieder gestört. Deshalb strömt wieder etwas Kalium in den Extrazellulärraum. Auf Dauer würden sich das Membranpotential abbauen. Aus diesem Grund gibt es ein Pumpensystem, daß Kalium Querschnitt durch einen Dendriten. in, und Natrium aus der Zelle pumpt. So kann das Gleichgewicht aufrecht erhalten werden. Die Ladungsverteilung muß überall gleich groß sein. Mehrere Prozesse versuchen dies zu verhindern: 1) Die Membran ist undurchlässig für Aminosäuren A-, weil diese zu groß für die Kanäle sind. 2) Die Membran ist semipermeabel für Na+ und K+. Normalerweise passiert K+ die Membran leichter als Na+. 3) Die Membran enthält ein Pumpensystem, das intrazelluläres Na+ mit extrazellulärem K+ austauscht. ___________________________________________________________________________________ Brain Modelling 19 Für die unterschiedlichen Ionenarten ergeben sich folgende Werte: Ionenart K+ Na+ ClA- intrazellulär [mM] 400 50 32 385 extrazellulär [mM] 20 440 560 - Gleichgewichtspot. [mV] -75 mV +55 mV -60 mV - Die Natrium-Kalium Pumpen bringen rund 2 K+-Ionen nach innen und 3 Na+-Ionen nach außen. Dies ist ein aktiver Prozeß, der Energie benötigt. Die Ionenkanäle arbeiten nicht perfekt. Das heißt es werden auch Ionen einer anderen Art durchgelassen. Dadurch kommt es zu Leckströmen J L, das heißt die Werte werden sich noch etwas verändern. Das Verhalten eines kleinen Stückchens der Membran eines Neurons im unterschwelligen Spannungsbereich kann man zum Groteil mit einer einfachen Schaltung aus Widerständen, Kondensatoren und Batterien beschreiben. + Jm Jc Jk intrazellulär JNa JL GL Cm GK(Vm,t) VK + – GNa(Vm,t) VNa — – + VL + – extrazellulär Die Membran besitzt eine gewisse Kapazität Cm. Ein Ionenkanal wird durch eine Batterie und einen Widerstand dargestellt. Der Kanal stellt den Widerstand dar. Die Physiologen geben aber lieber die Leitfähigkeit G (Kehrwert des Widerstandes) an, da sie ein direktes Maß für die Effizienz eines Ionenkanals ist. Das dort - durch Diffusion von Ionen - auftretende Potential - das Nernstpotential - entspricht einer Batterie. ___________________________________________________________________________________ Brain Modelling 20 3.2 Elektrische Eigenschaften der Membran Im Gegensatz zum Ruhemembranpotential ist das Membranpotential Vm ein allgemeinerer Begriff, der jede Art von Potential in jedem Augenblick an der Membran folgendermaßen definiert: Vm=Vi - Va wobei Vi das Potential im Inneren und Va das Potential im Zelläußeren ist. Vaussen Vinnen Membran Ändert sich das Gleichgewicht der Ionen im intra- und extrazellulären Raum durch das Anlegen einer Spannung - der elektrische Gradient wird verändert - so wird sich das Membranpotential ändern. Es depolarisiert. Die Größe der Depolarisation, die als Reaktion auf die Strominjektion, gegeben durch ein Experiment oder chemische Einflüße auf das Axon, entsteht, bestimmt den Eingangswiderstand R einer Zelle. Nach dem Ohm'schen Gesetz ist die Größe der Depolarisation U gegeben durch U = IR Wenn es bei zwei Neuronen zu identischen synaptischen Reizungen im elektrischen Sinne kommt, tritt bei der Zelle mit dem größeren Eingangswiderstand eine größere Änderung des Membranpotentials auf. Bei einer idealisierten kugelförmigen Nervenzelle ohne Ausläufer hängt der Eingangswiderstand sowohl von der Dichte der Ruhemembrankanäle als auch von der Größe des Neurons ab. Je größer ein Neuron ist, desto größer ist seine Membranoberfläche und desto geringer ist sein Eingangswiderstand, weil mehr Ruhemembrankanäle zur Ionenleitung vorhanden sind. Um die Widerstände von unterschiedlich großen Nervenzellen vergleichen zu können, sprechen Elektrophysiologen oft vom spezifischen Membranwiderstand Rm, also dem Widerstand einer Flächeneinheit der Membran, der in Ohm mal Quadratzentimeter [cm2] angeben wird. Um den Gesamtwiderstand zu berechnen muß der spezifische Membranwiderstand durch die Membranoberfläche der Zelle dividiert werden und so erhalten wir für ein kugelförmiges (idealisiertes) Neuron: R in Rm 4a 2 wobei a der Radius des Neurons ist. Im realistischeren Fall eines Neurons mit ausgedehnten Dendriten und Axonen hängt der Eingangswiderstand sowohl vom Membranwiderstand der Fortsätze als auch vom Widerstand des Cytoplasmas zwischen dem Zellkörper und seinen Ausläufern ab. Ein unterschwelliges Spannungssignal an Dendriten und Axonen nimmt mit zunehmender Entfernung von seinem Entstehungsort ab. Postsynaptische Potentiale, die an ___________________________________________________________________________________ Brain Modelling 21 Dendriten entstehen, werden in Richtung Zellkörper und Axonhügel geleitet. Das Cytoplasma eines Dendriten setzt dem längsgerichteten Strom einen signifikanten Widerstand entgegen, weil es eine relativ geringe Querschnittsfläche hat. Je länger ein Dendrit ist, desto größer ist auch der Widerstand, weil sich die Querschnittswiderstände addieren. Will man darstellen, wie der Widerstand über die Gesamtlänge eines Dendriten kontinuierlich zunimmt, kann man sich den Dendriten als eine Reihe von identischen, mit Cytoplasma gefüllten Membranzylindern vorstellen. Jeder Zylinder hat seine eigene Membrankapazität und seinen cytoplasmatischen Längswiderstand. Die Membrankapazität entsteht dadurch, daß die Proteine, aus denen sich die Membran zusammensetzt, einen Isolator darstellen. In dieser Kapazität können elektrische Ladungen gespeichert werden. Der Axial- oder Längswiderstand des cytoplasmatischen Innenraums ra, pro Längeneinheit (1 cm) in /cm angegeben, hängt sowohl vom spezifischen Widerstand des Cytoplasmas an-gegeben in cm, als auch von der Querschnittsfläche des Dendriten mit dem Radius a ab: ra a 2 Der Membranwiderstand rm hängt sowohl vom spezifischen Widerstand einer Flächeneinheit der Membran Rm als auch vom Umfang des Dendriten ab und wird pro Längeneinheit des Zylinders in .cm angegeben. rm Rm 2a Wie ändert sich nun das Membranpotential entlang des Dendriten mit der Entfernung, wenn man an einer Stelle Strom injiziert ? Sei vorausgesetzt, der kapazitive Strom sei Null und das Membranpotential hat ein konstantes Niveau erreicht, dann hängt die Potentialveränderung durch eine Strominjizierung ausschließlich von den relativen Werten von rm und ra ab. rm cm rm ra cm rm ra cm rm ra cm cm rm ra cm rm ra ra Der injizierte Strom fließt in den aufeinanderfolgenden Membranzylindern auf mehreren Wegen durch die Membran nach außen. Jeder dieser Strompfade besteht aus zwei Widerständen in Serie: Den Gesamtlängswiderstand rx und dem Membranwiderstand rm eines Membranzylinders. Weil sich Widerstände in Serie summieren, gilt rx = rax, wobei x die Entfernung in cm entlang des Dendriten vom Injektionsort ist. Durch die Membran eines Zylinders in der Nähe der Injektionsstelle fließt mehr Strom als an entfernteren Orten, (weil Strom immer den Weg des geringsten Widerstands folgt) und der Gesamtlängswiderstand rx, mit zunehmender Entfernung vom Injektionsort zunimmt. Wegen Um = Imrm wird die Änderung des Membranpotentials Um(x), die der Strom durch die Membran erzeugt, kleiner, wenn man sich auf dem Denriten von der Stromelektrode entfernt. Die Abnahme mit wachsender Entfernung erfolgt exponentiell und wird durch folgende Gleichung ausgedrückt: U m x U 0e x wobei die Längskonstante der Membran, x die Entfernung von der Strominjektionsstelle und U0 die Änderung des Membranpotentials ist, die vom Strom am Ort der Stromelektrode (x=0) ___________________________________________________________________________________ Brain Modelling 22 hervorgerufen wird. Die Längskonstante ist die Entfernung von der Strominjektionsstelle zu dem Ort auf dem Dendriten, an dem Um auf 1/e oder auf 37% seines Ursprungswertes abgenommen hat. Sie wird folgendermaßen berechnet: rm ra Je besser die Isolierung der Membran ist - je größer rm ist - und je besser die Leitungseigenschaften des Dendriteninneren sind - je niedriger ra ist - desto größer ist die Längskonstante des Dendriten. Weil rm in umgekehrtem Verhältnis zum Radius steht, wogegen ra indirekt proportional zum Quadrat des Radius ist, ist die Längskonstante proportional zur Quadratwurzel des Radius. Deswegen haben Axone mit größerem Durchmesser eine größere Längskonstante als dünne Axone. Eine Myelinisierung führt ebenso zu einer größeren Längskonstante. 3.3 Entstehung eines Aktionspotentials Es existieren nicht nur spannungsunabhängige Ionenkanäle. Es gibt auch spannungsabhängige Ionenkanäle. Diese Kanäle öffnen sich ab einer gewissen Spannung. Die Na+ spannunggesteuerten Kanäle arbeiten wesentlich schneller als die K+ -spannungs-gesteuerten Kanäle. Bei der Signalweiterleitung im Axon werden Natrium- und Kalium-Kanäle primär durch die Membranspannung gesteuert. Diese Kanäle werden nach dem "Alles oder Nichts"Prinzip ein oder ausgeschaltet. Mg2+ Im linken Bereich sehen wir einen offenen Ionenkanal. Mitte Links gibt es eine lokale Konformationsänderung. Mitte rechts gibt es eine Änderung der Struktur. Rechts ist ein ganz spezifischer Ionenkanal dargestellt. Ein Teilchen blockiert den Ionenkanal und nur durch einen speziellen Mechanismus kann dieses Teilchen den Ionenkanal öffnen. Glaspipette I (Strom durch einen einzelnen Ionenkanal) geschlossen 2 pA offen 0 20 40 60 Zellmembran t [ms] Ionenkanal Der Strom durch einen einzelnen Ionenkanal kann mir der Patch-Clamp-Technik gemessen werden. An eine Mikropipette (1 m Durchmesser) die mit der Membranoberfläche Kontakt hat, wird ein Unterdruck erzeugt. So wird der Ionenkanal elektrisch von der Umgebung separiert. Im Inneren der Pipette ist eine Salzlösung, die in etwa den elektrischen ___________________________________________________________________________________ Brain Modelling 23 Eigenschaften der extrazellulären Flüssigkeit entspricht. Mit dieser Methode konnten 3 verschiedenen Typen von Ionenkanälen identifiziert werden: spannungs-, transmitter- und mechanisch gesteuerte Kanäle. Die Signale, die den Kanal steuern, kontrollieren die Wahrscheinlichkeit mit der ein Kanal offen oder geschlossen ist. Es ist nicht so, daß ein Kanal die ganze Zeit ununterbrochen offen bleibt, wenn ein geeignetes Signal auf den Ionenkanal wirkt. Das Öffnen und Schließen erfolgt fast augenblicklich. Na+ und K+ Ionenkanäle Betrachten wir die spannungsabhängigen Ionenkanäle. Wird die Membran lokal 50 vorüber-gehend auf etwa -50 mV Aktionspotential depolarisiert, dann ist der Schwellwert überschritten und es werden die schnellen spannungsgesteuerten Kanäle für 0 Natrium-Ionen geöffnet und diese Ionen Natriumleitfähigkeit können in die Zelle einströmen. Aufgrund der so vergröerten Kaliumleitfähigkeit Membranpermeabilität für Na+ + -50 überschreitet der Na -Einstrom den K+ Ausstrom. Der resultierende Nettoeinstrom positiver Ladungen verursacht eine weitere Depolarisation und als Ergebnis kommt es zu einer totalen Depolarisation der Membran. Dieser positive Rückkopplungs-mechanismus entwickelt sich explosionsartig. Das Potential bewegt sich in Richtung +55 mV. Membranpotential (mV) U t U t U t ___________________________________________________________________________________ Brain Modelling 24 Danach schalten sich die Na+ -Kanäle langsam ab und die langsameren K+ -Kanäle werden aktiviert. Nach einem gewissem Zeitpunkt kehrt das System wieder in den Ruhezustand zurück. Die Depolarisation der Membran, das Ungleichgewicht der Ladungsverteilung, ist nicht lokal begrenzt. Wenn es zu einer Depolarisation kommt, dann werden auch die benachbarten Membranareale davon beinflußt. Meist wird ein Aktionspotential am Axonhügel ausgelöst. Von dort bewegt sich die Depolarisation in zwei Richtungen: entlang des Axons und entlang der Dendriten. Es können mehrere Ersatzschaltkreise miteinander gekoppelt werden. Damit ist es möglich die Ausbreitung eines Aktionspotentials beziehungsweise eine Strominjektion in den Dendriten zu beschreiben (Achtung: Die Membran ist überall gleich gebaut. Es existiert kein Unterschied zwischen einer "Axon-" oder "Dendritenmembran"). Das Hodgkin Huxley Modell Das Hodgkin-Huxley Modell bringt die Membranstromdichte und die Membranspannung in einen Zusammenhang (siehe das Ersatzschaltbild). Die J's sind Stromdichten [A/cm2], die V's sind Spannungen oder auch Potentiale [V]; Cm ist die Kapazität der Membran pro Flächeneinheit [F/cm2] und die G's bezeichnen die spezifischen Ionenleitfähigkeiten [S/cm2]. Es gibt vier Zweige im Schaltplan (siehe oben), aus denen sich der resultierende Gesamtstrom ergibt. Einer davon berücksichtigt die Kapazität der Membran, während die anderen drei die Ionenströme (spannungsabhängige Natrium- und Kaliumströme und die Leckströme, verursacht durch die spannungsunabhängigen Kanäle, aus denen auch das Ruhepotential resultiert) beschreiben. Der Natrium- und Kaliumzweig ist repräsentiert durch einen Widerstand in Serie mit einem Nernst-Gleichgewichtspotential (Batterie) für das jeweilige Ion. Wichtig in diesem Modell ist, daß die Natrium- und Kaliumleitfähigkeiten vom Membranpotential und der Zeit abhängen. Das Leckpotential beziehungsweise die damit verbundene Leitfähigkeit wird als konstant betrachtet. Unter Anwendung der Kirchhoff'schen Regeln und dem Ersatzschaltbild ergibt sich: J m J C Jion 1.1 und die Stromdichten für die verschiedenen Ionen J ion J K J Na J L 1.2 was geschrieben werden kann als: J m J C J K J Na J L 1.3 ___________________________________________________________________________________ Brain Modelling 25 Durch Anwendung des Ohm'schen Gesetzes (I=U/R=UG) und (I= Q , Q=C∙U) erhalten wir: J m Cm Vm G K (Vm , t ) (Vm VK ) G Na (Vm , t ) (Vm VNa ) G L (Vm VL ) 1.4 t VNa und VK stellen das Nernst'sche Gleichgewichtspotential für Natrium und Kalium dar, definiert durch VNa ce RT log iNa F c Na und VK ce RT log iK F cK 1.5 wobei R die molare Gaskonstante, T die absolute Temperatur und F die Faradaykonstante (F=9.6487104 Cmol-1) darstellen. Die molaren Konzentrationen für intrazelluläre beziehungsweise extrazelluläre Natriumionen und Kaliumionen ist durch c iNa , c eNa , c iK und c eK gegeben. Für den Zusammenhang von Strom und Spannung entlang eines Zylinders, das heißt in Richtung z, ergibt sich mit a als Radius der Querschnittsfläche eines Axons und r a als axoplasmatischer Längswiderstand. Man kann zeigen, daß gilt: 2 Vm z 2 2 a ra J m 1.6 Unter Verwendung von 1.4 und 1.6 ergibt sich 1 2 Vm V Cm m G K (Vm , t ) (Vm VK ) 2 2ara z t G Na (Vm , t ) (Vm VNa ) G L (Vm VL ) 1.7 Diese Gleichung beschreibt die Ausbreitung des elektrischen Potentials entlang einer Axonmembran. Das Membranpotential, gegeben durch Gl. 1.7 kann nun berechnet werden, wenn die Funktionen GK(Vm,t) und GNa(Vm,t) bekannt sind. Um diese Leitfähigkeiten bestimmen zu können, muß das Membranpotential systematisch variiert werden, wobei man gleichzeitig die resultierenden Veränderungen der Na +- und K+Leitfähigkeiten mißt. Das ist experimentell schwierig durchzuführen, weil das Membranpotential und das Öffnungsverhalten der Na+- und K+- Kanäle stark voneinander abhängen. Im Jahr 1949 entwickelte Cole eine Technik, die als Spannungsklemme (voltage clamp) bezeichnet wird. Mit Hilfe dieser Apparatur lassen sich die Leitfähigkeiten bestimmen. Strommeßgerät Stromelektrode Spannungsklemme Strom Stromelektrode Axon ___________________________________________________________________________________ Brain Modelling 26 Wenn das Membranpotential eines Axons "geklemmt" wird, dann öffnen oder schließen sich aufgrund der aufgezwungenen Potentialänderungen hin zwar immer noch die spannungsgesteuerten Ionenkanäle; die Spannungsklemme verhindert jedoch wirkungsvoll, daß die dabei entstehenden Ströme durch die Membran das vorgegebene Membranpotential beinflussen. Auf diese Weise kann man die Veränderungen der Membranleitfähigkeit für einzelne Ionenarten bei verschiedenen Membranpotentialen messen. Die Apparatur besteht aus einer Stromquelle, die mit zwei Elektroden, jeweils für den intra- beziehungsweise extrazellulären Bereich, verbunden ist. Man kann das Membranpotential schnell auf einen vorbestimmtem Depolarisationswert springen lassen, indem man Spannung an der Membran anlegt. Damit die gemessene Strom-Spannungs-Beziehung der Membran auswertbar ist, muß das Membranpotential über der gesamten Membranoberfläche konstant sein. Man kann diese Bedingung dadurch erhalten, daß ein sehr gut leitender strominjizierender Draht den axoplasmatischen Widerstand kurzschließt und daher den Längswiderstand auf Null reduziert. Aufgrund der vorgegebenen Depolarisation öffnen sich die Na+- und K+-Kanäle . Die so entstehenden Na+- und K+-Ströme würden normalerweise das Membranpotential verändern, aber die Spannungsklemme hält es auf dem vorgegebenen Wert fest. Wenn sich die Na +Kanäle nach einem mäßig depolarisierenden Spannungsprung öffnen, entwickelt sich normalerweise ein Einwärtsstrom, weil Na+-Ionen, angetrieben von der elektrochemischen Potentialdifferenz, durch diese Kanäle in die Zelle fließen. Dieser Na +-Einstrom depolarisiert die Membran, indem er die positive Ladung an der Membraninnenseite erhöht und die an der Außenseite erniedrigt. Die Spannungsklemme greift hier ein, indem sie gleichzeitig positive Ladungen aus der Zelle in die externe Lösung pumpt. Der Voltage-Clamp-Stromkreis erzeugt also einen gleich großen, aber entgegengerichteten Strom und steuert so automatisch jedem Strom durch die Membran entgegen, die zu einer Abweichung des Membranpotentials vom vorgegebenen Wert führen würde. Im Endergebnis findet keine Nettoänderung der Ladungsmenge über der Membran statt und damit auch keine signifikante Verschiebung des Membranpotentials. Betrachten wir jetzt den kapazitiven Strom. Das Membranpotential V m ist zu jeder Zeit proportional zur Ladung Qm des Membrankondensators. Wenn sich Vm nicht ändert, ist die Ladung Qm konstant und es fließt kein kapazitiver Strom. Wenn auf die Sollspannung ein Rechteckimpuls gelegt wird, fließt nur am Anfang und am Ende dieses Impulses ein kapazitiver Strom. Dieser Strom ist sehr kurzlebig und er unterscheidet sich deutlich vom ionischen Strom. Damit läßt sich der Ionenstrom isolieren und entsprechen analysieren. Wenn man nun ein um zehn Millivolt depolarisierendes Potential (Sollspannung) anlegt, beobachtet man zuerst, daß ein sehr kurzer Auswärtsstrom den Membrankondensator augenblicklich mit der Strommenge entlädt, die für eine Depolarisation um 10 Millivolt notwendig ist. Diesem kapazitiven Strom JC folgt ein kleiner Ionenausstrom, der für die Dauer des Spannungssprungs anhält. Am Ende des Spannungssprungs beobachtet man einen kurzen, einwärts gerichteten kapazitiven, Strom und der gesamte Membranstrom kehrt zu Null zurück (Figur 3.3 links). Der ionische Gleichgewichtsstrom, den man während der Dauer der Reizung messen kann, fließt durch die Ruhemembrankanäle und wird als Leckstrom J L bezeichnet. Diese Ruhemembrankanäle, die immer geöffnet bleiben, sind für den Aufbau des Ruhemembranpotentials verantwortlich. ___________________________________________________________________________________ Brain Modelling 27 Vm [mV] Vm [mV] 0 -50 t -60 Jm t -60 Jm JC JC JL JL auswärts 0 einwärts t 0 t JC Messungen mit der Spannungsklemme am Riesenaxon des Tintenfisches. In der linken Darstellung wird die Schwelle für das Aktionspotential im Gegensatz zur rechten Darstellung nicht ausgelöst. Wenn man eine größere depolarisierende Sollspannung anlegt, werden die Stromkurven komplizierter. Die Amplituden der kapazitiven Ströme und der Leckströme steigen an. Außerdem entstehen kurz nach dem Ende des kapazitiven Stroms und nach dem Beginn des Leckstroms ein Einwärtsstrom. Dieser erreicht innerhalb weniger Millisekunden sein Maximum, fällt dann wieder ab und wird durch einen Auswärtsstrom ersetzt. Dieser Auswärtsstrom erreicht ein Plateau, das für die Dauer des Spannungsprungs bestehen bleibt. Diese Ergebnisse lassen sich am einfachsten so erklären, daß die depolarisierende Spannung nacheinander Kanäle für zwei unterschiedliche Ionen aktiviert: Einen Kanaltyp für den einwärts und einen Kanaltyp für den auswärts gerichteten Strom. Weil sich diese entgegengerichteten Ströme teilweise zeitlich überlappen, ist die schwierigste Aufgabe bei der Analyse solcher Voltage-Clamp-Experimente, die einzelnen Zeitverläufe getrennt voneinander zu bestimmen. Wenn man die K+-Kanäle mit TEA (Tetraethylammonium) in der Axonmembran blockiert, besteht der Membrangesamtstrom J m aus dem kapazitiven Strom, dem Leckstrom und dem Natriumstrom (JC, JL, JNa). Die Leckleitfähigkeit ist konstant; sie variiert praktisch nicht mit dem Membranpotential oder mit der Zeit. Daher kann JL leicht bestimmt werden und vom Membrangesamtstrom abgezogen werden, wodurch JNa und JC übrigbleiben. Der Strom verursacht durch die Membrankapazitäten tritt nur kurz am Anfang und am Ende des Pulses auf, man kann ihn leicht erkennen und eliminieren. So bleibt der reine Natriumstrom JNa übrig. Um den Strom durch die spannungsgesteuerten Na+-Kanäle zu messen, injiziert man mehrere verschiedene Ladungsmengen und klemmt das Membranpotential dadurch auf verschiedene Werte. In ähnlicher Weise kann man den Kaliumstrom JK messen, wenn man die Na+-Kanäle durch TTX (Tetrodotoxin) blockiert. ___________________________________________________________________________________ Brain Modelling 28 Jm [mA/cm2] Jm [mA/cm2] Kalium 0.4 Natrium 2.5 0 0 -0.4 0 5 10 Zeit [ms] 15 0 5 10 Zeit [ms] 15 20 Verschiedene Messungen von Kalium- und Natriumströmen unter Voltage-ClampBedingungen. Das Ruhepotential wurde auf -70 mV eingestellt und für Kalium wurde das Potential von -60 bis +40 mV, für Natrium von -40 bis +80 mV mit Schritten von 10 mV variiert. Die Temperatur betrug 4-5 °C. Um die Leitfähigkeiten aus den Voltage-Clamp-Strömen zu berechnen, wird das Ersatzschaltbild herangezogen. Aus dem Ohmschen Gesetz kann man den Strom durch jede Klasse der spannungsgesteuerten Kanäle errechnen. J K Vm , t G K (Vm , t ) Vm VK und J Na Vm , t G Na (Vm , t ) Vm VNa 1.8 Die Umformung und Auflösung nach G ergibt zwei Gleichungen, nach denen sich die Leitfähigkeiten berechnen lassen: J (V , t ) J (V , t ) 1.9 G K (Vm , t ) k m und G Na (Vm , t ) Na m Vm VK Vm VNa Um diese Gleichungen zu lösen, muß man Vm, VK, VNa, JK und JNa kennen. Die unabhängige Variable Vm wird durch das Experiment festgelegt. Die abhängigen Variablen J K und JNa können aus den Messungen der Voltage-Clamp-Experimente abgeleitet werden. Die verbleibenden Konstanten VK und VNa können bei bekannten Ionenkonzentrationen nach der Nernst-Gleichung berechnet werden. Hodgkin und Huxley haben die Kaliumleitfähigkeit durch folgende Gleichungen beschrieben: G K (Vm , t ) G K n 4 (Vm , t) 1.10 dn n (1 n) n n dt 1.11 dabei ist G K der konstante maximale Leitwert und n ist spannungs- und zeitabhängig. Die Formulierung geht von der Annahme aus, daß die Fähigkeit der K+-Ionen, durch die Membran zu fließen, vom Zustand oder der Position von geladenen Molekülen in der Membran abhängt. Im Zustand können die K+-Ionen passieren, während sie im Zustand für K+ undurchlässig ist. Die Funktion n beschreibt dann den Anteil der Moleküle, die sich im Zustand befinden, und 1-n ist der Anteil im Zustand . Die Reaktionsgeschwindigkeit des Überganges vom Zustand in den Zustand wird mit n bezeichnet und die des Übergangs von mit n. Die beiden Parameter n und n sind potential- aber nicht zeitabhängig. Beim Ruhepotential ist n groß und n klein. Bei der Depolarisation wächst n potentialabhängig, und n fällt, folglich steigt n und damit GK. Nach ausreichend langer ___________________________________________________________________________________ Brain Modelling 29 Depolarisation ist n größer als n und das Gleichgewicht stellt sich so ein, daß die meisten Moleküle im Zustand sind. Die Leitfähigkeit GK hat somit einen hohen Wert und behält ihn solange die Depolarisation andauert. Die Funktion n erhielt die vierte Potenz, weil nur so der verzögerte, aber dann steile Anstieg der Kaliumleitfähigkeit nachgebildet werden konnte. Die Natriumleitfähigkeit der Membran wird durch ein ähnliches Gleichungssystem beschrieben. Es muß aber der Komplikation Rechnung getragen werden, daß GNa nach einer Depolarisation der Membran nach rund einer Millisekunde wieder auf den Ruhewert abfällt. Kurz gesagt, das Natriumsystem wird nach einer Depolarisation deaktiviert. Der Anstieg der Na+-Leitfähigkeit wird durch die Variable m beschrieben, die sich ähnlich wie n bei der Natriumleitfähigkeit verhält, und zusätzlich wird die Variable h eingeführt, die die Inaktivierung beschreibt. G Na (Vm , t ) G Na m 3 (Vm , t) h (Vm , t) 1.12 dm m (1 m) m m dt 1.13 dh h (1 h) h h dt 1.14 Hier ist analog zum Kaliumsystem G Na der konstante maximale Natriumleitwert und m und h beschreiben zwei Mechanismen, die die Natriumleitfähigkeit regulieren, wobei die Reaktionsgeschwindigkeiten m, m, h und h nur potentialabhängig sind. Die Funktionen m und m zeigen qualitativ dieselbe Potentialbahängigkeit wie n und n , das heißt sie beschreiben einen Prozeß, der bei der Depolarisation die Membran für Na+-Ionen öffnet. Dagegen haben h und h eine spiegelbildlich verkehrte Potentialabhängigkeit, sind aber absolut kleiner als m und m. Bei Depolarisation nimmt deshalb h langsamer ab, als m zunimmt. Folglich steigt die Leitfähigkeit schnell an, inaktiviert aber nach kurzer Zeit die Membrandurchläßigkeit. Die numerischen Werte für m, m, h und h könne aus den Zeitverläufen von GNa bestimmt werden. Zeitverlauf für n, n4, m, m3, h und m3h während eines Depolarisierungspulses von -70 bis 0 mV für die Dauer von 3 ms. Die Funktionen n und m folgen dem (1-e-t/) Verlauf, im Gegensatz zu h, das den e-t/ Verlauf nimmt. ___________________________________________________________________________________ Brain Modelling 30 Hodgkin und Huxley bestimmten die Geschwindigkeitskonstanten in Abhängigkeit des Membranpotentials und paßten die analytische Ausdrücke an: n m 0.01(Vm 50) e 0.1( Vm 50) 1 0.1Vm 35 e 0.1Vm 35 1 h 0.07e (Vm 60) / 20 n 0.125 e -(Vm 60)/80 1.15 m 4e (Vm 60) / 18 1.16 h 1 e 0.1( Vm 30) 1 1.17 Das Hodgkin Huxley Modell umfaßt die Gleichungen 1.7 und 1.10-1.17 mit folgenden numerischen Parametern: c eNa = 491 mmol/L 36 mS/cm2 c iNa = 50 mmol/L G L = 0.3 mS/cm2 c eK = 20 mmol/L VL = -49 mV c iK = 400 mmol/L G Na = 120 mS/cm2 GK = Cm = 1 F/cm2 Diese Daten gelten bei 6.3 °C. Unter Anwendung obiger Formeln ist es nun möglich, die Weiterleitung von Signalen nichtmyelinisierter Axone hervorragend zu beschreiben. ___________________________________________________________________________________ Brain Modelling 31 4.0 Die chemische Übertragung zwischen zwei Neuronen Ein biologisches Neuron empfängt Informationen von anderen Nervenzellen über synaptische Verbindungsstellen und überträgt sie, zum Beispiel in der Großhirnrinde, an Tausende andere Neuronen weiter. Die Synapse verstärkt oder schwächt ein Signal ab, das von einem Neuron zu einem anderen übertragen wird. Die meisten Nervenzellen besitzen zwischen tausend und zehntausend Synapsen. Man unterscheidet drei verschiedene Synapsentypen: a) Effektorsynapsen regen mit den Kollateralen verschiedene Drüsen oder Muskelzellen an. b) Rezeptorsynapsen dienen der sensiblen Innervation. c) Interneuronale Synapsen stellen den Kontakt zwischen Nervenzellen auf unterschiedlichste Weise her. Dieser Typ ist am häufigsten in unserem Gehirn vorhanden. Die 1 bis 2 m groen Synapsen sind vergleichbar mit kleinen Dornen (Endknöpfchen), die auf den Dendriten oder am Ende des Axons sitzen. Erreicht ein Nervensignal die Synapse am Axonende (=Neurit), dann wird eine Überträgersubstanz in den Spalt zwischen den einzelnen Synapsen ausgeschüttet, durchquert ihn und wird von den Rezeptoren der Synapse des Dendrits gebunden. Dadurch kommt es zu einer Änderung des elektrischen Zustandes der nachgeschaltenen Nervenzelle. Es gibt zwei häufige morphologische Synapsentypen im Gehirn, Gray-Typ I und Gray-Typ II. Synapsen vom Typ I sind erregend (99% von ihnen arbeiten mit Glutamat), während Synapsen vom Typ II hemmend (oft GABAerg) sind. In der Gesamtzahl der synaptischen Stärke sind beide gleich häufig vertreten. Die interneuralen Synapsen können wiederum unterteilt werden. Axosomatische Synapsen verbinden die Kollaterale mit einer Postsynapse, die direkt am Zellkörper einer nachgeschaltenen Nervenzelle liegt. Axodendritische Synapsen münden mit den Axonendigungen an einem Dendriten, wo sie einen Dornfortsatz umgreifen können. Axoaxonale Synapsen stellen den Kontakt zwischen einer Präsynapse und dem Nachbarneurit her. Dendrodendritische Synapsen koppeln zwei unterschiedliche Dendriten. Die 1 bis 2 mm großen Synapsen sind vergleichbar mit kleinen Dornen (Endknöpfchen), die auf den Dendriten oder am Ende des Axon sitzen. Diese Dornen enthalten kleine Bläschen, auch synaptische Vesikel genannt, in denen eine Überträgersubstanz gespeichert ist. Erreicht ein Nervensignal die Synapse im Axonende, wird die Überträgersubstanz in den Spalt zwischen den einzelnen Synapsen ausgeschüttet, durchquert ihn und wird von den Rezeptoren der Synapse des Dendrits gebunden. Dadurch kommt es zu einer Änderung des chemischen beziehungsweise elektrischen Zustandes der nachgeschalteten Nervenzelle. Es gibt zwei häufige morphologische Synapsentypen im Gehirn: Gray-Typ I und Gray-Typ II. Synapsen vom Typ I sind oft glutamerg und, wie wir noch in Kapitel 1.5 sehen werden, daher erregend, während Synapsen vom Typ II oft GABAerg und somit hemmend sind. Bei Synapsen vom Typ I ist der synaptische Spalt leicht erweitert auf 30 nm, und die präsynaptische aktive Zone hat eine Fläche von 1 - 2 m2. Bei Synapsen vom Typ II hat der Spalt eine Breite von 20 nm. Somit ist die aktive Zone etwas kleiner. ___________________________________________________________________________________ Brain Modelling 32 Die Synapse ist eine spezialisierte Struktur, die der Kommunikation zwischen zwei Neuronen beziehungsweise zwischen einem Neuron und Vesikel einer Muskelzelle dient. Beide Rezeptor gesteuerVesikelZellen sind durch den ter Na+-Ionenkanal depot sogenannten synaptischen Spalt voneinander getrennt. Das präsynaptische Neuron schüttet einen Neurotransmitter aus, der an Rezeptoren auf dem Freisetzungsstellen Rezeptor postsynaptischen Neuron gebunden wird. Dadurch wird synaptischer Spalt das nachgeschalten Neuron in U [mV] ~20 - 40 nm breit den elektrischen Eigenschaften beinflußt. Die chemische 0.4 Signalübertragung lät sich in EPSP zwei Schritte unterteilen: einen Transmitter-Schritt, bei dem die präsynaptische Zelle einen t [ms] Botenstoff freisetzt, und einen Rezeptor-Schritt, bei dem der IPSP Transmitter an den Rezeptoren der postsynaptischen Zelle Die Werte können stark variieren. gebunden wird. Trifft ein ausreichend starkes elektrisches Signal auf die Präsynapse, dann werden spannungsgesteuerte Calciumkanäle geöffnet. Der Anstieg der Calcium-Ionen-Konzentration bewirkt eine Verschmelzung der synaptischen Bläschen (Vesikel) mit der Membran und der Neurotransmitter kann aus den Bläschen durch den synaptischen Spalt zu den Rezeptoren der Postsynapse diffundieren. Bei einem Aktionspotential werden rund 1-10 Vesikel verschmolzen. Ein Vesikel enthält rund 2000 Moleküle (bezieht sich auf ACTH). Die Transmittermoleküle reagieren mit den Rezeptoren der postsynaptischen Membran. Diese Rezeptoren veranlassen daraufhin Ionenkanäle dazu, sich zu öffnen oder zu schlieen was eine Änderung des Membranpotentials des nachgeschaltenen Neurons zur Folge hat. An der Postsynapse entsteht ein Potential, wenn ein Neurotransmittermolekül an einem Rezeptor bindet. Präsynapse Postsynapse ausgeschütteter Neurotransmitter Man unterscheidet zwei verschiedene Arten von postsynaptischen Potentialen. Zum einen kann eine der Neurotransmitter verbunden mit dem Rezeptor eine Depolarisation der Membran des nachgeschaltenen Neurons bewirken. Dies führt zu einem Excitatorischen PostSynaptischen Potential (EPSP). Wird die Membran hyperpolarisiert, dann spricht man von einem Inhibitorischem PostSynaptischem Potential (IPSP). Die Signalzeitverzögerung zwischen dem Eintreffen des Aktionspotentials an der Präsynapse und dem postsynaptischen Potential beträgt mindestens 0.3 ms, normalerweise aber 1-5 ms. Es gibt einen speziellen Typ von Synapsen - die NMDA-Synapsen (eigentlich Rezeptoren). Die Signale werden genauso übertragen, wie oben beschrieben. Dies geschieht durch die gewöhnlichen Glutamatrezeptoren (AMPA - -amino-3-hydroxy-5-methyl-4-isoxazolepropionate). Die Ionenkanäle steuern direkt das Membranpotential. Die NMDA-Rezeptoren (N-methyl-D-aspartate), die genauso auf Glutamat ansprechen, müssen durch einen eigenen Mechanismus geöffnet werden (Tetanisierung). Sie steuern nur indirekt das Membranpotential. Wenn sie aktiviert werden, beginnt im inneren der Synapse eine molekulare Kaskade (second messenger), die unter anderem einen Einfluß auf das Membranpotential hat. Eine Aktivierung von NMDA-Rezeptoren kann zu einer langanhaltenden Modifikation der Effizienz von glutamergen Synapsen führen. ___________________________________________________________________________________ Brain Modelling 33 Bei der Signalübertragung an der Synapse können mehrere Schritte unterschieden werden: 1. Synthese: Der Neurotransmitter wird entweder im Zellkörper hergestellt oder Vor-stufen des Neurotransmitter werden zur Synapse über das Axon transportiert und in der Synapse selbst hergestellt. 2. Speicherung: Der Neurotransmitter wird meist in Vesikel gespeichert, in denen er für die Ausschüttung zur Verfügung steht. 3. Ausschüttung: Beim Feuern des Neurons verschmelzen die Vesikeln mit der präsynaptischen Membran und der Neurotransmitter wird in den synaptischen Spalt abgegeben. 4. Rezeption: Der Neurotransmitter diffundiert durch den Spalt und bindet sich schwach an den Rezeptoren an der postsynaptischen Membran. Es kommt zu einer Depolarisation oder Hyperpolarisation des nachgeschaltenen Neurons. 5. Inaktivierung: Bei manchen Synapsen wird der Neurotransmitter im synaptischen Spalt inaktivert. 6. Wiederaufnahme: Bei anderen Synapsen wird der Neurotransmitter wieder in das präsynaptische Neuron aufgenommen. Es kann sowohl zur Inaktivierung als auch zur Wiederaufnahme kommen. 7. Abbau: Der freie Neurotransmitter innerhalb der Endigung kann abgebaut werden, um die Transmitterkonzentration im Neuron zu regulieren. Jedes Neuron empfängt ständig die synaptischen Signale andere Neuronen. Einige sind excitatorisch, einige inhibitorisch, einige effektiver, andere weniger effektiv. Manche Synapsen liegen auf den Spitzen der apikalen Dendriten, einige auf dem Dendritenschaft andere auf den Dendritendornen. Die verschiedenen Inputs können einander verstärken oder auch auslöschen. Die Potentiale die von einem einzelnen postsynaptischen Neuron erzeugt werden sind klein und nicht in der Lage eine postsynaptische Zelle so stark zu depolarisieren, da die Schwelle für das Aktionspotential erreicht wird. Der Nettoeffekt der Inputs an jeder excitatorischen oder inhibitorischen Synapse hängt daher von verschiedenen Faktoren ab: Vom Ort der Synapse, von ihrer Gröe und Form, sowie von der Nähe und relativen Stärke anderer erregenden oder hemmenden Synapsen. Die Effektivität von chemischen Synapsen kann für kürzere oder auch längere Zeit verändert werden. Die Regulierbarkeit oder synaptische Plastizität wird von zwei Arten von Vorgängen gesteuert: 1) Vorgänge innerhalb des Neurons, wie Änderung des Membranpotentials oder das Abfeuern von Aktionspotentialen. 2) Extrinsische Prozesse wie die synaptischen Eingänge von anderen Neuronen. ___________________________________________________________________________________ Brain Modelling 34 Für einen Neurotransmitter müssen folgende Bedingungen erfüllt sein: 1. Er muß im Neuron synthetisiert werden. 2. Er muß in den präsynaptischen Endigungen vorhanden sein. 3. Exogen verabreicht, wird die selbe Wirkung erzielt. 4. Es ist ein spezifischer Mechanismus vorhanden, um den Transmitter abzubauen. Ob ein Neurotransmitter erregend oder hemmend wirkt, ist von den Rezeptoren der Postsynapse abhängig. Praktisch gesehen ist Glutamat meist bei erregenden (Erhöhung der Membranspannung) und GABA (-Aminobuttersäure) bei hemmenden (Erniedrigung der Membranspannung) Prozessen beteiligt. Wir unterscheiden zwischen Neurotransmitter, die praktisch nur der Signalweiterleitung dienen und Transmitter, die verschiedene Prozesse im Neuron oder an der Synapse modulieren. Signalweiterleitung: Glutamat: meist erregend, dient der Informationsweitergabe, sehr häufig GABA: meist hemmend, 25-40% der Synapsen Modulierende Wirkung: Insgesamt mit nur ein paar Prozent im Gehirn vorhanden: Acetylcholin: Steuerungsaufgaben im Vorderhirn, Hippocampus, Septum Catecholamine: Dopamin, Noradrenalin, Adrenalin; modulierende Aufgaben Serotonin: Modulierend im Vorderhirn, Cerebellum und Rückenmark Die letztgenannten Neurotransmitter finden sich im gesamten Gehirn. Von einzelnen zentralen Schaltstationen (für jeden Neurotransmitter gibt es in ein bis drei Kernen) ziehen Faserstränge in verschiedenste Gebiete des Gehirns und modulieren dort verschiedenste Prozesse. Die Neurotransmitter Serotonin, Dopamin und Noradrenalin haben häufig bei wichtigen geistigen und neurologischen Fehlfunktionen eine wesentliche Bedeutung, wie sie zum Beispiel bei Depression, Schizophrenie, Autismus, Rauschgiftsucht und bei der Parkinson-Krankheit auftreten. Die Prozesse im Inneren einer Synapse können durchaus kompliziert werden. Zum Teil können aber auch Neurotransmitter auf die regulativen Prozesse einen Einfluß haben. ___________________________________________________________________________________ Brain Modelling 35 Überblick zwischen verschiedenen Signalen, die an Neuronen auftreten können, und den damit verbundenen Ionenkanälen: Ionenkanäle Signal Typ Mechanismus Signaleigenschaften Ruhepotential meist K+- und Cl-- überwiegend Leck- variiert in verschiedenen Kanäle einige Na+- kanäle (auch span- Zellen zwischen -45 Kanäle nungsgesteuerte K+- und -90 [mV] Kanäle) Aktionspotential separate Na+und K+-Kanäle spannungsgesteuerte Alles oder Nichts Prinzip Kanäle, Na+- und K+- Amplitude: 100 [mV] Kanäle werden nach- Dauer: 1-10 [ms] einander aktiviert Rezeptorpotential unspezifischer sensorischer Reiz Kationen-kanal für Na+- und K+-Ionen graduiert, schnell, Dauer: mehrere Millisek. Amplitude: einige [mV] ___________________________________________________________________________________ Brain Modelling 36 5.0 Technische neuronale Netzwerke Es gibt eine Vielzahl von unterschiedlichen Modellen, die die Aktivitäten von Neuronen beschreiben. Hodgkin und Huxley waren die ersten, die mit Hilfe einer Differentialgleichung die elektrische Erregung in einem Neuron über den Natrium- und Kaliumhaushalt sinnvoll beschrieben haben. Dieses Modell war nur der Beginn einer langen Reihe von verschiedenen mathematischen Beschreibungsweisen. So ist das Bonhoeffer-Van der Pol als auch das FitzHugh Nagumo Modell als Weiterentwicklung zu erwähnen. Dadurch ist es gelungen, die Wirkungsweise von Medikamenten und Nervengiften wesentlich besser verstehen zu können. So ist es wichtig, nicht nur die chemischen Eigenschaften von Neuronen, sondern auch das mathematische Konstrukt zur Beschreibung der biochemischen Eigenheiten zu kennen. Dieses Kapitel liefert eine kurze Einführung in die Modellbildung und physikalische Beschreibungsweise von einzelnen Neuronen. 5.1 Formale Neuronen U [mV] 0 Schw elle t Entladungsfrequenz des Neurons t Entladungsfrequenz [Hz] 20 Im Unterschied zur Biologie beziehungsweise Neurologie können auch andere Modelle benutzt werden, bei denen nicht alle Aspekte eines Neurons exakt beschrieben werden. Stattdessen wird ein Modell verwendet, daß eine sehr grobe Verallgemeinerung darstellt. Die sich ergebenden Netze sind somit keine Neuronen-Netze, sondern nur "neuronale" (neuronenähnliche) Netze. In Hinkunft werden alle neuronenähnliche Verarbeitungseinheiten, als formale oder technische Neuronen bezeichnet. Beschäftigen wir uns einmal mit der Kodierung der Information in technischen neuronalen Netzen und deren biologischer Begründung. Wie vorher dargelegt, muß eine elektrische Schwelle überschritten 15 werden, damit es zur Auslösung eines Aktionspotentials kommt. Steigt die 10 b Reizstromstärke weiter an, so feuert das b2 1 Neuron öfters. Obwohl das 5 resultierende Axonsignal von Natur aus binär ist (feuern oder nicht feuern), läßt sich scheinbar diese binäre Information 1 2 3 4 5 6 7 8 in eine nicht-binäre Form übertragen, Reizstromintensität (log.Einheiten) wenn man die Feuerfrequenz eines Neurons betrachtet. Wie wir sehen können ist die Feuerfrequenz in weiten Grenzen in guter Näherung stückweise linear bezogen auf den Logarithmus der Stärke des angelegten Reizes. Bei hohen Reizstärken geht die Zellaktivität in eine Sättigung über, die durch eine mindestens 1 ms lange Regenerationszeit nach einem Aktionspotential bestimmt wird. Damit ist die maximale Taktfrequenz der Neuronen auf einen relativ niedrigen Wert von 1 kHz beschränkt. Sättigung ___________________________________________________________________________________ Brain Modelling 37 Das Grundmodell eines technischen Neurons stützt sich im wesentlichen auf die Vereinfachungen von McCulloch und Pitts, die ein Neuron als eine Art Addierer mit Schwellwert betrachteten. Die Synapsen eines Neurons nehmen Aktivierungen xi, vergleichbar den EPSP's, mit bestimmten Stärken wi (Gewichtungen, synaptischen Stärken) von anderen Neuronen auf, summieren diese und lassen dann am Ausgang y des Neurons (entspricht in der Biologie dem Axonhügel) eine Aktivität entstehen, sofern die Summe vorher einen Schwellwert b überschritten hat. Ein technisches (formales) Neuron: x1 w1 x2 w2 xi wi xn wn Aktivierung Synapse Eingabe Gewichtung Ausgabe y z Schwellwert b Axonhügel Summation Axon Ausgabe x1w1 + x2w2 +...+ xiwi +...+ xnwn = z McCulloch und Pitts zeigten, daß mit diesen einfachen Elementen jeder finite logische Ausdruck berechnet werden kann. Fassen wir die Eingabeaktivitäten x1, ... , xn zum Eingabevektor x =(x1, ... , xn)T und die Gewichte w1, ... , wn zum Gewichtsvektor w =(w1, ... , T wn) zusammen, so läßt sich die resultierende interne Aktivität z formal als Skalarprodukt beider Vektoren schreiben: z( x , w ) w j x j w T x 3.19 j Die biologische Aktivitätsfunktion "sammelt" die Ladungen der dendritischen Eingänge zu einem elektrischen Potential und wird deshalb auch als "Potentialfunktion" bezeichnet. Sehr oft muß die Aktivität erst eine Schwelle b (bias) überschreiten, bevor es zu einer Aktivität am Ausgang kommt. Dies läßt sich durch die Minderung der Aktivität um einen Schwellwert modellieren: z( x , w ) w T x b 3.20 Den zusätzlichen Term b kann man allerdings in der Notation wieder zum Verschwinden bringen, indem man eine Erweiterung der Vektoren um eine Zusatzkomponente x =(x1, ... , T T xn, 1) und w =(w1, ... , wn, -b) vornimmt. Die Aktivität y am Neuronenausgang wird durch die Ausgabefunktion S(z), abhängig von der internen Aktivität z, beschrieben: y=S(z) 3.21 Der Wertebereich der verwendeten Variablen ist je nach Modellvariante und Anwendungsbereich unterschiedlich. Meist ist xi zwischen Null und Eins definiert, manchmal ___________________________________________________________________________________ Brain Modelling 38 sind aber andere normierungen notwendig und sinnvoll. Man kann folgende Klassifizierung für die Ausgabefunktionen treffen: Binäre Ausgabefunktion: Die Werte für den Input xi und dem Output y sind binär (aktiv oder nicht aktiv). Die Gewichte wi können aber reell sein. Eine positive Aktivität y ergibt sich erst nach dem Überschreiten einer Schwelle: 1 y S BIN (z) 0 z0 3.22 z0 Anstelle vom Wert "1" kann für spezielle Anwendungen auch eine beliebige andere Zahl verwendet werden. Begrenzt-lineare Ausgabefunktion: Wie oben ersichtlich, ist die Entladungsfrequenz in weiten Bereichen stückweise linear bezogen auf den Logarithmus der Stärke des angelegten Reizes (Reizstromstärke). Dieser Sachverhalt läßt sich durch eine begrenzt-lineare Ausgabefunktion modellieren. Damit ist es möglich die Frequenzmodulation zu berücksichtigen. Aus dieser Graphik geht hervor, daß es zwei wichtige Werte für die Eingabeaktivität gibt: der untere Schwellwert b1, der überschritten werden muß, um die Membran zu depolarisieren, und den Wert b2, nach dessen Überschreitung es zu einer Sättigung kommt. So erfolgt die Ausgabe zwischen den zwei Schwellwerten und dem Sättigungswert ymax: 0 y max z b1 y S LIN (z) b 2 b1 y max y 1 0 z b1 b1 z b 2 3.23 b2 z y Zmax SBIN(z) SLIN(z) b1 z 0 b2 z Darstellung einer binären (links) und einer begrenzt-linearen Ausgabefunktion (rechts). ___________________________________________________________________________________ Brain Modelling 39 Sigmoide Ausgabefunktion: Bei der analytischen Verwendung der Ausgabefunktion ist es manchmal nötig, auch die Ableitung der Funktion selbst zu kennen. Somit sollte diese Funktion glatt sein im Gegensatz zu den bisher besprochenen Ausgabefunktionen. Die als sigmoiden Funktionen bekannten Ausgabe-funktionen sind dabei praktischer als obige Stufenfunktionen. Meist wird die Fermi-Funktion verwendet: y S Fermi (z) 1 3.24 1 e kz y SFermi(z) 1 z 0 Darstellung der Fermi-Funktion als Ausgabefunktion. Im Unterschied zu biologischen Neuronen ist es aber bei formalen Neuronen möglich, völlig beliebige und für eine bestimmte Problemlösung günstige Neuronen neu zu definieren. Beispielsweise kann die Aktivität auch über den Abstand definiert sein: z x w , sodaß S(z) dann eine Bewertungsfunktion der Eingabe x bezüglich des Abstandes vom Punkt w darstellt. x2Körpergröße Neuron 1 alternative Neuronen x1Körpergewicht d x1 1 w1 x2 w2 b y Formale Neuronen lassen sich sehr effektiv beim Problem der Mustererkennung verwenden. Mehrere ähnliche Muster sollen zu einer Gruppe zugeordnet werden. Man spricht dann von Musterklassen. Betrachten wir dabei zwei Musterklassen "Übergewicht" und "Untergewicht". Jedes Muster hat (in diesem Beispiel) zwei Eigenschaften x1 (Körpergewicht) und x2 (Körpergröße), und jede Eigenschaft kann einen Wert zwischen 0 und 1 annehmen. Somit läßt sich jedes Muster in einer Ebene als Punkt mit x =(x1, x2) darstellen. Aufgabe einer Klassifizierung ist es, zwischen den beiden Klassen eine Trennungsline zu finden, die die beiden Mengen trennt. Für die Beurteilung eines neuen unbekannten Musters reicht es dann aus, die Lage bezüglich der Klassengrenze (diesseits oder jenseits) zu bestimmen. ___________________________________________________________________________________ Brain Modelling 40 Betrachten wir ein formales Neuron: x1w1 + x2w2 + x3w3 = 0 Im Term x3w3 ist der Schwellwert versteckt, x3 = –b und w3 = 1: x1w1 + x2w2 – b = 0 Führen wir ein paar Umformungen durch: x2w2 = –x1w1 + b und w b x 2 x 1 1 w2 w2 Durch Neubezeichnung der Terme (–w1/w2) = k und (b/w2) = d ergibt sich: x2 = x1 k + d Diese Gleichung ist gerade die Geradengleichung: y = k x + d. Das heißt ein Neuron trennt über eine Gerade die beiden Muster. Die Geradengleichung beschreibt die Menge aller Punkte die die Gleichung erfüllen. Was passiert, wenn Punkte, die oberhalb oder unterhalb der Gleichung liegen, eingesetzt werden. Der Term x1w1 + x2w2 + x3w3 wird jetzt nicht Null gesetzt, sondern das Ergebnis berechnet: x1w1 + x2w2 + x3w3 = E x2Körpergröße Neuron 1.0 E>0 E<0 x1Körpergewicht Das Ergebnis E entspricht dem Normalabstand des jeweiligen Punktes zur Geraden. Liegt ein Punkt, also eine Person mit geringem Körpergewicht und eher großer Körpergröße oberhalb der Geraden, dann gilt E > 0. Hat eine Person ein großes Körpergewicht und ist von kleiner Statur, so liegt der Punkt unterhalb der Geraden und es gilt E < 0. 1.0 Ein solchermaßen definiertes formales Neuron stellt also einen linearen Klassifikator dar. Jedes Muster wird in eine durch die Gewichte und den Schwellwert definierten Klasse eingeordnet. Bei zwei Eingängen kann man das Problem ganz leicht graphisch darstellen. Wenn ein technisches Neuron über mehrere Eingänge verfügt, dann trennt nicht eine Gerade, sondern eine Hyperebene den Raum. ___________________________________________________________________________________ Brain Modelling 41 Natürlich können auch mehrere Geraden - technische Neuronen - zur Mustertrennung herangezogen werden. x2 g3 x2 g1 g2 g1 x1 x1 x1 x1 y1 x2 y1 y2 y2 x2 5.2 Das Perzeptron Dieses Modell löste unter den damaligen Gehirnforschern großen Enthusiasmus aus. Endlich schien ein Modell vorhanden zu sein, das nicht nur bestimmte Reize lernen und wiedererkennen konnte, sondern auch von den ursprünglichen Eingabedaten leicht abweichende Reize richtig zuordnen konnte. Informationsfluß w1 w2 wn-1 y wn Eingabeschicht vorverarbeitende Einheiten Ausgabeneuron Ein Perzeptron besteht in der Regel aus einer endlichen gitterartigen Eingabeschicht, die in einzelne Zellen unterteilt ist. Weiters gibt es eine Vielzahl vorverarbeitender Einheiten, die nur einen Teilausschnitt des Gitters beobachten können. Sie werden gewöhnlich als lokale Detektoren bezeichnet. Diese Detektoren sind nicht vergleichbar mit den formalen Neuronen aus dem vorigen Kapitel. Sie werden zwischen der Eingabeschicht und der Neuronenschicht eingefügt. Diese ersten Schritte der Bild-Vorverarbeitung werden oft als low-level oder early vision bezeichnet. Es findet auf dieser Ebene noch keine eigentliche Erkennung statt. Die Abbildung von der Eingabeschicht zur Vorverarbeitungsschicht wird mit bezeichnet. Diese vorverarbeitenden Einheiten schicken dann ihre Daten an die eigentliche Neuronenschicht. Dort wird dann eine Entscheidung über das "gesehene" Objekt getroffen. ___________________________________________________________________________________ Brain Modelling 42 Jedes dieser vorverarbeitenden Einheiten untersucht eine bestimmte Teilmenge der Eingabeschicht und teilt das Ergebnis den Neuronen, kompetenteren Entscheidungsträgern mit. In diesem Fall beschränken wir uns nur auf ein Neuron in der Neuronenschicht (o.B.d.A.). Jede vorverarbeitende Einheit führt einen Vergleich von intern gespeicherten Werten (Mustern) mit der zu beobachtenden Teilmenge durch. Sobald eines der gespeicherten Muster ident ist mit dem zu untersuchenden Bereich, dann sendet diese Einheiten ein Signal an das Neuron. Dieses Neuron ist ident mit den zuvor im Kapitel beschriebenen formalen Neuronen. Sonst verhält es sich still. Das Indizien Neuron multipliziert jedes erhaltene Signal mit die für das einer Zahl (Gewichtung) und bildet schließlich Vorliegen die Summe über alle so erhaltenen Zahlen. einer Treppe Wenn die Summe einen gewissen Schwellwert sprechen : übersteigt, antwortet das finale Neuron mit 1,sonst mit 0. Betrachten wir in der linken Abbildung eine Eingabeschicht und die dazugehörigen Indizien, die auf eine Treppe hinweisen. Wesentlich ist, daß die 44 Felder die beobachtet werden, überlappen. Ein wesentlich kompliziertere Aufgabe kann das nachfolgende Perzeptron lösen. Als Beispiel sei hier ein Perzeptron vorgestellt, das erkennen kann ob sich in seinem Blickfeld Rechtecke (Null inbegriffen) beliebiger Form und Größe befinden, oder ob sich Rechtecke an Seiten oder Kanten berühren. vorverarbeitene Einheit Indizien, die für das Vorliegen mehrerer getrennter Rechtecke sprechen: Indizien, die gegen das Vorliegen mehrerer getrennter Rechtecke sprechen: Eingabeschicht (links) und interne Muster (Gewichte) für die Vorverarbeitung (rechts). Für die Mustererkennung kann man entweder die Indizien die für beziehungsweise die Indizien die gegen das Vorliegen von Rechtecken sprechen in den vorverarbeitenden Einheiten abspeichern. Zu jedem 22-Quadrat im Eingabebereich wird eine Einheit positioniert. Weiters werden dann die Muster, die für das Vorliegen von Rechtecken sprechen, in alle diese vorverarbeitenden Einheiten übertragen (intern gespeicherte Werte). Schließlich erhalten alle Gewichte des eigentlichen Neurons den Wert +1 und der Schwellwert wird auf d, die Anzahl aller vorverarbeitenden Einheiten gesetzt. Wenn also alle Einheiten einen positiven Vergleich zwischen den intern gespeicherten Repräsentationen und dem zu beobachtenden Bereich machen, schicken sie alle ein Signal zum beurteilenden Neuron, die Summe ist d und das Neuron setzt den internen Wert auf "1". ___________________________________________________________________________________ Brain Modelling 43 Das Perzeptron hat erkannt, daß nur voneinander getrennte Rechtecke in der gitterartigen Eingabeschicht vorliegen. Stimmt mindestens ein zu beobachtender Teilbereich nicht mit den internen Werten überein, dann bleibt mindestens eine Einheit stumm und die Schwelle wird nicht erreicht. Folglich bleibt das Neuron im Zustand "0". Für eine 22 Teilmenge auf einer nn Matrix gilt: Da es (n-1)2 lokale Einheiten gibt lautet der Schwellwert: T=(n-1)2-1 Wenn alle lokalen Einheiten ein Indiz entdeckt haben, daß für die Rechtecke spricht, dann wird als Summe (n-1)2 herauskommen; also (n-1)2 > T = (n-1)2 - 1. Wenn nur eine einzige lokale Einheit kein positives Signal weiterleitet, dann gilt: (n-1)2 -1 > T falsche Aussage. Also werden keine Rechtecke gesehen. Mit diesen Perzeptronen konnten tolle Dinge durchgeführt werde. Man glaubte, daß man das Rätsel Gehirn gelöst hatte. Ein Neuron wurde nach dem Vorbild der Natur nachgebildet und es konnte "scheinbar" komplizierte Strukturen erkennen. Doch dann kamen die Neurowissenschafter Papert und Minsky. Sie zeigten in einem einfachen und eindrucksvollen Beweis, daß einfache Perzeptrone nicht in der Lage sind komplizierte Dinge zu erkennen. Betrachten wir ein Perzeptron, das erkennen soll, ob eine Figur zusammenhängend ist oder nicht. Trennen wir die Eingabeschicht in drei Bereiche, den Linken L, den Mittleren A und den Rechten. L A R Graphik A Graphik B Graphik C Graphik D Indirekt angenommen, das Perzeptron erkennt die erste Graphik (A) als durchgehende Linie, dann wäre es schon gescheitert. Also muß der Schwellwert kleiner als 3 - da es drei Teilbereiche gibt - sein. Bei der zweiten Graphik (B) müssen alle drei Bereiche den Schwellwert erreichen. Praktisch ist aber nur der linke Bereich unterschiedlich zur Graphik A. Dasselbe gilt natürlich auch für Graphik C - nur ist diesmal der rechte Bereich unterschiedlich zur Graphik A - aber auch hier muß wieder der Schwellwert erreicht werden. Bekommt das Perzeptron die Graphik D vorgesetzt, dann kommen der linke Bereich von Graphik B und der rechte Bereich von Graphik C zum Tragen - daraus setzt sich die Graphik D zusammen. Damit würde der Schwellwert überschritten und auch Graphik D würde aus einer zusammenhängenden Figur bestehen, was falsch ist: Graphik AL + A + R < 3 Graphik BL´+ A + R = 3 Graphik CL + A + R´= 3 Graphik DL´+ A + R´< 3 (0.5+1.0+0.5 < 3) (1.5+1.0+0.5 = 3) (0.5+1.0+1.5 = 3) (1.5+1.0+1.5 = 4 < 3) WID. Damit wäre gezeigt, daß ein Perzeptron gewisse Muster nicht sinnvoll erkennen kann. Dies führte zu einer schlimmen Krise innerhalb der Neurowissenschaften. Viele Wissenschafter suchten sich andere Arbeitsgebiete. Der Glaube an eine naturwissenschaftliche Erklärung des Gehirns war erschüttert. ___________________________________________________________________________________ Brain Modelling 44 Aber versuchen Sie doch selbst einmal mit einem raschen Blick zu erkennen, welche der nachfolgenden Graphiken zusammenhängend sind. Mit einem sehr kurzen Blick werden Sie es wahrscheinlich nicht schaffen. Erkennen ist ein sehr komplexes Verhalten - dabei sind viele Neuronen beteiligt - nicht nur ein einziges, wie beim Neuron. Erst einige Jahre später erkannte man, daß sich mit mehr Neuronen praktisch alle mathematischen beziehungsweise formalisierbaren Probleme lösen lassen. Dafür gibt es auch einen Beweis. 5.3 Mehrschichtnetzwerke Wenn wir unsere Klasse (Gewicht zu Größe) in drei Gruppen unterteilen wollen, übergewichtig, normalgewichtig und untergewichtig, benötigen wir 2 Neuronen in der Hidden-Layer. Diese Neuronen unterteilen die Gruppe. x2Körpergröße Diese Problem entspricht dem Untergewicht 1.0 XOR-Problem. XOR-Problem: x1 0 0 1 1 x2 0 1 0 1 Ausgabe 0 1 1 0 x1Körpergewicht Natürlich müssen die beiden Neuronen ihre Information über den jeweiligen Abstand zu Muster gemeinsam an ein anderes Muster weitergeben. Dieses Neuron entscheidet dann über den Output. Diese Gruppierung von Neuronen werden als Netzwerke bezeichnet. Da die Informationsrichtung eindeutig vorgegeben ist, bezeichnet man diesen Typ als "feed-forwardNetzwerk". Das Netz besitzt 3 Schichten, allerdings ist die Bezeichnung der Schichten in der Literatur nicht immer eindeutig. Idealgewicht x1 Übergewicht 1.0 3-Schicht feed-forward Netzwerk b1 w1 w5 w2 b3 w3 x2 1.Schicht Eingabeschicht Eingabeschicht w4 w6 b2 2.Schicht 1.Schicht Hidden-Layer 3.Schicht 2.Schicht Ausgabeschicht ___________________________________________________________________________________ Brain Modelling 45 Damit das Netzwerk obiges Problem (XOR beziehungsweise das Gewichtsproblem) lösen kann, müssen die Gewichte wi und Schwellwerte bi folgendermaßen gesetzt werden: w1=w4=w5=w6=0.5 w2=w3=-0.5 b1=b2=b3=0.3 Durch entsprechende Algorithmen lassen sich auch in komplizierteren Fällen die Koeffizienten wi und damit die Klassengrenzen berechnen. Rechnen wir ein gesamtes Netzwerk durch. Für die einzelnen Neuronen in der jeweiligen Schicht ergeben sich für vier verschiedene Input's ( 0-0 / 0-1 / 1-0 / 1-1 ) folgende Werte: 1.Neuron-2.Schicht: 1.0 00 01 10 00 1.0 1.0 2.Neuron-2.Schicht: 00 01 10 00 1.0 1.0 y1 < 0 0 y1 > 0 1 y1 < 0 0 y1 < 0 0 y1 > 0 1 y1 > 0 1 y1 < 0 0 y1 > 0 1 1.Neuron-3.Schicht: 0001 0111 1000 0001 y1 > 0 1 y1 < 0 0 y1 < 0 0 y1 > 0 1 Mit der Anwendung von mehreren Neuronen auf ein Problem ist eine neue Idee geboren. Jedes Neuron der 2.Schicht (Hidden-Layer) - natürlich auch jedes Neuron der 3.Schicht beschäftigt sich mit einem kleinen Teilproblem. Es wird nicht das Muster als Ganzes (ein Symbol) betrachtet, sondern es werden kleine Strukturen betrachtet (Sub-Symbole). Wodurch ist ein Elefant definiert ? Ein Elefant ist durch einen Rüssel, große Schlappohren und vier große Beine definiert. Diese Sub-Symbole stellen in ihrer Gesamtheit ein Symbol ___________________________________________________________________________________ Brain Modelling 46 dar: den Elefant. Wenn manche Sub-Symbole verändert werden (zum Beispiel die Füße), oder neue Sub-Symbole hinzugefügt werden, dann entsteht ein Phantasiewesen, das mit dem ursprünglichen Symbol nur mehr sehr wenig zu tun hat. Die Grundidee der sub-symbolischen Informationsverarbeitung ist das Auflösen der Symbole zur Beschreibung der Anwendungswelt in Mikrostrukturen auf der Basis primitiver Verarbeitungseinheiten. Dies können z.B. Neuronen sein. Damit stellt sich die Frage nach der Bestimmung der Gewichte. 5.4 Bestimmung der Gewichte Es gibt verschiedene Möglichkeiten die Gewichte der einzelnen Neuronen zu bestimmen. Das jeweilige Verfahren hängt sehr stark vom Problem ab. Raten gutes Raten Gradientenverfahren genetische Algorithmen Hybride (Trial and Error) (Sukzessive Approximierung) (Backpropagation) Gutes Raten (sukzessive Approximation): 0 12 25 50 100 6...gesuchte Zahl Am Anfang eines Lernalgorithmuses steht eine geeignete Definition des Ziels, welches erreicht werden soll. Dies kann eine Minimierung des Fehlers, den das Netzwerk beim "Erlernen" macht, sein. Der Fehler entsteht als Differenz zwischen der tatsächlichen Ausgabe und der gewünschten Ausgabe (Lehrervorgabe). y Endwerte Energie x Startwerte Diese Energielandschaft ist von zwei Variablen, x und y, abhängig: In Abhängigkeit vom Startwert findet das Gradientenverfahren unterschiedliche Lösungen (in diesem Fall). Gradientenverfahren: Man bestimmt den Gradienten [k = f '(x*)] einer Funktion f(x) an der Stelle x*. Die Steigung k gibt an, ob die Differenz zwischen gewünschtem und tatsächlichem Wert besser oder schlechter wird, wenn x* erhöht oder erniedrigt wird. Der Wert x* wird proportional zu Steigung erhöht oder erniedrigt (je nachdem was besser ist). Das heißt der aktuelle Fehler wird kleiner. In der neuen Position wird wiederum der Gradient bestimmt. Und so weiter ... ___________________________________________________________________________________ Brain Modelling 47 ACHTUNG: Dieses Verfahren liefert nicht immer den besten möglichen Wert da es lokale Minima geben kann ! f(x) f(x) a) f(x) b) c) Steigung k x* x x x Die Steigung bestimmt die Schrittweite, zusätzlich kann die Schrittweite über den Lernfaktor eingestellt werden. Ist der Lernfaktor zu groß kann das Optimum meist nicht erreicht werden (Abb. b). Ist der Lernfaktor zu klein, dann dauert es sehr lange bis das Optimum oder ein lokales Extremum erreicht wird (Abb. c). (2) (1) (1) (1) w ij(1) ( x ) (2) k w ki (1 S Fermi ( z i )) S Fermi (z i ) x j k (2) (2) w ij(2) ( x ) ( y i(2) L i ) (1 S Fermi (z i(2) )) S Fermi (z i(2) ) x (2) j i x j Eingabe 1.Schicht x(1) 2.Schicht y(1) = x(2) hidden units Ausgabe y(2) Ausgabeeinheiten (1) (2) x(1) ...Eingabevektor, y(2) ...Ausgabevektor Beim Gradientenverfahren bestimmt Gewichtsveränderung zurück ((1),(2)). man den Fehler, und rechnet auf die genetischer Algorithmus: Die Gewichte aller Neuronen werden als Gencode betrachtet. Das heißt: Gen1:=(1w1, 1w2, 1w3,..., 1wn) Gen2:=(2w1, 2w2, 2w3,..., 2wn) usw. y Energie x Mit genetische Algorithmen werden unterschiedliche Bereiche (dunkle Kreise) der Energielandschaft betrachtet. Die Bereiche, die gut sind, dürfen überleben und können sich vermehren. Dabei verringert sich meistens der Abstand zum globalen Maximum. Nach Bestimmung der jeweiligen Fitness (Fehler den das Netz macht), treten folgende Operationen auf: ___________________________________________________________________________________ Brain Modelling 48 Überkreuzung: Zwei Tupel werden an einer Stelle auseinandergeschnitten und überkreuzt zusammengesetzt (Crossing-over): Gen1=(1w1, 1w2, 1w3,..., 1wn) Gen2=(2w1, 2w2, 2w3,..., 2wn) Gen1=(2w1, 2w2, 2w3, 1w4,..., 1wn) Gen2=(1w1, 1w2, 1w3 ,2w4,.., 2wn) Invertierung (Inversion): Ein Tupel wird in der Reihenfolge der Gene invertiert. Genk=(kw1, kw2, kw3,..., kwn) Mutation: Genk=(kw3, kw2, kw1,..., kwn) Einige Gene (Gewichte) werden zufällig geändert. Genk=(kw1, 0.134, kw3,..., kwn) Genk=(kw1, 0.671, kw3,..., kwn)Backpropagation: Nach Durchlaufen (propagation) des Eingabesignals x durch die Netzschichten wird der Fehler :=y(x)-L(x) des Ausgabesignals y bezüglich der gewünschten Ausgabe L ermittelt und mit Hilfe eines Gradientenverfahrens durch alle Schichten zurückgeführt (error backpropagation). 6.0 Lernen - synaptische Plastizität Implizites Gedächtnis: Inhalt wird nicht bewußt als Information verarbeitet. Oft-maliges Üben führt zu einer Verbesserung (motorisches Lernen). Explizites Gedächtnis: Tatsachen, Ereignisse, Vorstellungen Alle Lernvorgänge können auf die Veränderung von synaptischen Stärken zurückgeführt werden. Die Zeichnung von Dendriten von hippocampalen Neuronen in einer Phase hoher (links) und geringer (rechts) Östrogenkonzentration. Auch beim Lernen verändern sich die Dornen (spines) auf den Dendriten in der Größe. ___________________________________________________________________________________ Brain Modelling 49 Priming: Es wird ein Reiz (das Wort "Kerze") präsentiert. Nach einer kurzen Zeit-spanne wird ein Teil des ursprünglichen Reizes ("Ker") präsentiert und das Wort kann vervollständigt werden (funktioniert auch bei Personen die sich nichts mehr merken können). Nicht assoziatives Lernen: Es wird etwas über die Eigenschaft eines EINZELNEN Reizes gelernt. Meist geschieht dies über Wiederholungen. Auf synaptischer Ebene entspricht dies der Habituation und Sensibilisierung. Assoziatives Lernen: Klassische Konditionierung: Es werden 2 unabhängige Reize verknüpft. Operante Konditionierung: Es wird ein Reiz und ein Verhalten verknüpft. Mantelrand Atemröhre Siphon Kieme Die meisten Experimente zur synaptischen Plastizität - dem Lernen auf zellulärem Niveau wurden an Aplysia californica (kalifornischer Seehase) durchgeführt. Bei dieser Schnecke sind die Neuronen genetisch determiniert und leicht zugänglich. Habituation: Die Reaktion auf einen harmlosen Reiz schwächt sich ab, bis der Reiz vollständig ignoriert wird. Dies geschieht durch monotone Wiederholung. visuelle Fixierung Herzschlagfrequenz Atemfrequenz Bei Aplysia führt eine 10 malige Reizung des Siphons (normalerweise wird die Kieme dann zurückgezogen) zu einer Vermeidung des Reflexes für rund 10-15 Minuten. Die synaptische Verbindung wird geschwächt, es sind weniger Vesikel an der Präsynapse angedockt. (Kurzzeithabituierung) ___________________________________________________________________________________ Brain Modelling 50 An der Synapse sind weniger Vesikel angedockt. Die Zahl der Rezeptoren ändert sich bei der Synapse nicht. Dadurch, daß weniger Vesikel angedockt sind, wird das EPSP kleiner. Vesikel Vesikeldepot U [mV] ursprüngliches EPSP t [ms] habituiertes EPSP Bei mehreren Reizungen über mehrere Tage lang, wird der Reflex bis zu 3 Wochen lang vermieden. (Langzeithabituation). Die Zahl der Synapsen nimmt ab (von 1300 auf 840). Was nicht benötigt wird, wird abgebaut. Sensitivierung: Bei der Sensitivierung wird etwas über einen schädlichen Reiz gelernt. Zum Beispiel wird der Kiemenrückziehreflex durch Sensitivierung deutlich ge-stärkt. Sensitivierung moduliert die Synapse über folgende Schritte: Ein Schwanz-schock aktiviert sensorische Neuronen im Schwanz der Aplysia. Diese Neuronen akti-vieren ihrerseits modulatorische Interneuronen. Diese Interneuronen steuern die Transmitterfreisetzung im Kiemenrückziehschaltkreis. Der modulierender Neuro-transmitter (Serotonin) der Interneuronen erhöht die Vesikelanzahl. ___________________________________________________________________________________ Brain Modelling 51 Präsynapse des Interneurons aus der Schwanzregion modulierender Neurotransmitter "informationsübertragender" Neurotransmitter Erhöhung von cAMP Präsynapse des Siphonneurons Postsynapse des Kiemenneurons Die Wirkungsweise einer kurzfristigen Sensibilisierung beruht auf der Veränderung des Calciumstroms durch die Membrane der Synapsen. Die Konzentration freier CalciumIonen bestimmt, wieviele Vesikel mit der präsynaptischen Membran ver-schmelzen. Serotonin aktiviert die Adenylat-Cyclase. Dies führt zur Bildung von cyclischem Adenosinmonophosphat(cAMP) worauf unter anderem mehr Calciumkanäle gebildet werden. Veränderungen an der Synapse im Überblick: Klassische Konditionierung: In der Regel kann klassische Konditionierung die Antwortbereitschaft eines Reflexes effektiver verstärken als eine Senstitivierung. Der ursprüngliche Reiz (unbedingte), der angeboren ist (Speichelfluß beim Fressen) wird in Verbindung mit einem neutralen (zu konditionierenden) Reiz gebracht (Pfleger mit Futter). Es müssen 2 elementare Regeln erfüllt sein: 1. zeitliche Nähe 2. Es muß eine hohe Wahrscheinlichkeitsbeziehung zwischen dem neutralen Reiz und dem unbedingten Reiz geben (Vorhersagbarkeit der Umwelt). Der neutrale Reiz muß kurz vor dem unbedingten Reiz angeboten werden, beide sollten aber zur gleichen Zeit enden. Der Unterschied zur Sensitivierung besteht in der Zeitabhängigkeit. Aufgund der besseren Zeitabstimmung können die chemischen Mechanismen der modulierenden Interneuronen viel nachhaltiger wirken. Langzeitpotenzierung (LTP) Sie verursacht eine längerfristige Änderung der Effizienz von Synapsen. Donald O. Hebb sagte bereits 1949 voraus, daß es auf dem Niveau von Synapsen eine einfache Lernregel gibt: "Wenn die Aktivität eines präsynaptischen Neurons wiederholt mit der Entladung eines postsynaptischen Neurons verbunden ist, wenn also beide Neuronen gleichzeitig feuern, dann wird die Effizienz zwischen der Prä- und Postsynapse erhöht." ___________________________________________________________________________________ Brain Modelling 52 w AB (t ) w AB (t 1) w x A y B w (t ) (t ) x y (t)...Lernrate Ein paar Jahre später wurde eine länger andauernde Erhöhung der synaptischen Stärke nach wiederholten Aktivierungen von exzitatorischen Synapsen im Hippocampus beobachtet. Diese exzitatorischen Synapsen arbeiten mit Glutamat als Transmitter, welches von zwei verschiedenen Rezeptortypen - AMPA und NMDA -, die in den einzelnen Synapsen gleichzeitig vorhanden sind, erkannt wird. Die LTP kann entweder durch gleichzeitiges Feuern des prä- und des post-synaptischen Neurons, als auch durch Aktivierung der Synapse mit hoher Frequenz ausgelöst werden. Wesentlich dabei ist aber, daß die postsynaptische Membran depolarisiert ist, während neue EPSP's das Neuron erreichen. Eine übliche Voraussetzung für das Zustandekommen von LTP ist das gleichzeitige Feuern von mehreren innervierenden Fasern. Wenn zwei unterschiedliche erregende Signale - ein schwaches und ein starkes - im gleichen Bereich eines Dendriten einer Pyramidenzelle ankommen, wird der schwache Impuls nur dann verstärkt, wenn er in Assoziation mit dem starken Impuls eintrifft. LTP bedarf also einer simultanen Erregung von prä- und postsynaptischem Neuron. Bei der normalen synaptischen Signalübertragung dominieren die AMPA-Rezeptoren, da die Membrankanäle der NMDA-Rezeptoren durch Mg2+-Ionen blockiert werden. Die Blockade wird nur dann aufgehoben - das heißt die NMDA-Kanäle werden nur dann aktiviert - wenn die postsynaptische Zelle gleichzeitig von anderen starken Aktionspotentialen vieler präsynaptischer Neuronen depolarisiert wird. Diese multiple Depolarisation bewirkt, daß sich die positiv geladenen Mg2+-Ionen vom Eingang der NMDAKanäle lösen und damit den Einstrom von Na+-Ionen und besonders Ca2+-Ionen in die Zelle ermöglichen. Der Ca2+-Einstrom ist dann das Signal zur Induktion zur Langzeitpotenzierung. Für die Änderung der Membranleitfähigkeit durch NMDA-Rezeptoren gibt es zwei Bedingungen: Glutamat muß am Rezeptor gebunden und die Membran muß gleichzeitig depolarisiert sein. Zwischen diesen beiden Bedingungen dürfen maximal 100 Millisekunden an Zeit vergehen, damit es zu einer LTP kommt. Die entscheidende Depolarisierung wird normalerweise durch die Aktivierung zahlreicher AMPA-Rezeptoren aufgrund der Aktionspotentiale zahlreicher präsynaptischer Neuronen erzeugt. Dadurch kommt es zu einem vermehrten Ca2+-Einstrom durch die nun freien NMDARezeptoren. Dies initiiert eine anhaltende Verstärkung der synaptischen Signalübertragung, indem es zwei calciumabhängige Serin-Theonin-Proteinkinasen - die Ca2+/Calmodulin-Kinase und die Proteinkinase C - sowie eine Tyrosinkinase aktiviert. Nicht assoziative LTP: Zustand der Postsynapse spielt keine Rolle !!! 1) Tetanische Reizung der Präsynapse. 2) Dadurch kommt es zu einem massiven Ca2+-Einstrom in der Präsynapse. 3) Calmodulin-Ca2+-Komplex bindet an Enzym Adenylatcyclase (Typ I). 3) cAMP-Spiegel steigt. 4) cAMP-Proteinkinase wird aktiviert. 5) Mehr Vesikel werden gebildet !!! ___________________________________________________________________________________ Brain Modelling 53 assoziative LTP: 1) AMPA, -Amino-3-hydro-5-methyl-4- isoazolpropionsäure-Rezeptor (nonNMDA). 2) NMDA, N-Methyl-D-Aspartat-Rezeptor. NMDA-Rezeptoren werden aktiviert durch eine starke Depolarisation der postsynaptischen Membran durch: a) Tetanus bzw. b) Glutamat aus Präsynapse und Aktionspotential aus Postsynapse treffen zusammen. Dadurch wird die Blockade der Mg2+-Ionen inaktiviert: starker Einstrom von Ca2+-Ionen a) Calcium-Calmodulin-Proteinkinase II, b) Proteinkinase C, c) Tyrosinkinase Dadurch wird der retrograde Messenger NO zur Postsynapse geleitet: mehr Transmitterausschüttung ! LERNEN t=0 assoziatives Lernen Konditionierung t ungenau Habituierung Sensibilisierung nicht assoziatives Lernen LTP assoziative LTP nicht assoziative LTP Tetanische Reizung t ungenau mod ulierender Neurotransmitter Es existieren 2 prinzipielle Lernmechanismen: 1. Hebbsche Lernregel:"Wenn die Aktivität eines prä-synaptischen Neurons wiederholt mit der Entladung eines postsynaptischen Neurons verbunden ist, wenn also beide Neuronen gleichzeitig feuern, dann wird die Effizienz zwischen der Prä- und Postsynapse erhöht." 2. Aktivitätsabhängige Bahnung: "Koinzidenz zwi-schen vorgeschaltetem und modulatorischem Neuron." modulierendes Neuron ___________________________________________________________________________________ Brain Modelling 54 7.0 Repräsentation von Objekten Wie werden die Informationen schließlich gespeichert und bei einer späteren Assoziation wieder vervollständigt (rekombiniert) ? Wie kommt es überhaupt zur Trennung (Segmentierung) eines Objekts vom Hintergrund oder von anderen Objekten ? Man unterscheidet zwischen der lokalen und der verteilten Repräsentation. Bei der lokalen Repräsentation wird eine Einzeleigenschaft oder eine gesamte Wahrnehmung durch ein einzelnes Neuron dargestellt, das zum Beispiel durch eine erhöhte Aktivität deren Vorhandensein anzeigt. Im Gegensatz dazu werden Eigenschaften bei einer verteilten Repräsentation stets durch viele Neuronen angezeigt, wobei das Aktivitätsmuster als Ganzes ein Perzept darstellt. Ein anderes Aktivitätsmuster der gleichen Neuronen kann für eine andere Eigenschaft stehen. Barlow hat hierzu den Begriff der Großmutterzelle geprägt, die Zelle, die genau dann feuert, wenn eine Großmutter in das visuelle Aufmerksamkeitszentrum trifft. Die immense Zahl von Neuronen des menschlichen Gehirns reicht nicht aus, um für alle relevanten Kombinationen von Reizeigenschaften, die sich für natürliche Objekte ergeben, ein eigenes Neuron zur Verfügung zu stellen. Es kommt zu einer kombinatorischen Explosion der Neuronenanzahl, wenn man alle möglichen Reize betrachtet, die gespeichert werden sollen. Was passiert wenn sich ein gespeicherter Reiz sich verändert ? Was passiert, wenn das Neuron, das meine Großmutter repräsentiert, abstirbt - verliere ich die Erinnerung an meine Großmutter ? Zusätzlich müßten Neuronen reserviert werden für mögliche neue Reizkombinationen. (a) o u o u Neuron ist aktiv Leinwand (b) o Neuron ist passiv o u u Rosenblatt's Bindungsmaschine: Vier Neuronen detektieren unterschiedliche Eigenschaften von Mustern, die auf eine Leinwand projeziert werden. Die ersten beiden reagieren auf den Ort des Musters (oben oder unten), die anderen beiden auf dessen Form (Quadrat oder Dreieck). Tritt nun ein Muster auf (a), so kann anhand des Aktivitätsmusters die Situation auf der Leinwand eindeutig dargestellt werden. Werden zwei Muster gleichzeitig auf der Leinwand dargestellt (b), so lassen sich Ort und Form nicht mehr eindeutig zuordnen - die Maschine ist mit dem Bindungsproblem beschäftigt. Die Zusammengehörigkeit der durch neuronale Aktivität repräsentierten Merkmale zu bestimmten Objekten soll durch wiederholtes, synchrones Feuern der jeweiligen Neuronen zum Ausdruck gebracht werden, während Merkmale, die zu anderen gleichzeitig darzustellenden Objekten gehören, mit einer anderen Frequenz beziehungsweise um eine Phase verschoben feuern. ___________________________________________________________________________________ Brain Modelling 55 Aktionspotentiale o o u u t Bindung wird also über die zeitliche Korrelation neuronaler Antworten dargestellt. Dadurch können die Vorteile der Assemble-Codierung wie Robustheit und sparsamer Umgang mit den vorhandenen Neuronen beibehalten werden. Gleichzeitig ist es möglich, in hierarchischer Art und Weise den strukturellen Aufbau eines Objektes oder eines gesamten Bildes darzustellen, da Assembles nun auch in einem Hirnareal koexistieren können. Die Kreise symbolisieren veschiedene Neuronen. Es kann zu unterschiedlichen und dennoch gleichzeitigen Aktivierungen kommen. Die Aktivierungen sind als Stern, Quadrat, bemalter Kreis dargestellt. Die Verwendung einer Zeitstruktur ermöglicht es, ein Assemble von konstituierenden Neuronen über die Synchronisation zu definieren. Durch subtile Änderung der zeitlichen Relationen können Neuronen schnell zwischen verschiedenen Assembles umschalten. Neuronen sind Koinzidenzdetektoren ! Einige wenige Spikes von verschiedenen Neuronen können kaum ein nachgeschaltenes Neuron erregen, es sei denn, sie kommen gleichzeitig (Gleichzeitigkeit als Bindemittel). Ursache für eine Synchronisation können gemeinsame Stimuli sein, oder aber starke synaptische Kopplungen. Zur Vermeidung des nutzlosen und trivialen Zustandes totaler Synchronisation dienen inhibitorische Subsysteme. Wiederholen sich die synchronen Zustände in regelmäßigen Abständen, so ergeben sich Oszillationen. Solch ein rythmisches Feuerverhalten kennt man auch schon von einzelnen Neuronen. In diesem Sinne kann man folgende Vorteile von Oszillationen vermuten: 1. Durch die Periodizität sollte sich auch bei langen Verbindungswegen (wie Kopplungen zwischen den Hemisphären) sukzessive eine verschwindende Phasendifferenz einstellen. 2. Über intermediäre Oszillatoren können auch nicht direkt gekoppelte Oszillatoren synchronisieren. Dies ist wichtig beim Binden verschiedener, nicht a priori verschalteter, sensorischer Modalitäten (Hören-Sehen). 3. Auch bei stark gestreuten Übertragungszeiten, hervorgerufen durch axonale und synaptische Delays, können Oszillatoren zuverlässig mit verschwindender Phasendifferenz synchronisieren. ___________________________________________________________________________________ Brain Modelling 56 Modell eines "integrate and fire neurons" Zur Definition eines "integrate-and-fire"-Oszillators: Der Hauptteil des Modell-Neurons, der sogenannte "integrator", beschreibt im unterschwelligen Bereich, in dem noch kein Aktionspotential ausgelöst wird, die Eigenschaften der Zellmembran eines Neurons. Eine solche Membran, die ein nichtidealer Isolator zwischen zwei elektrisch leitfähigen Flüssigkeiten ist, wird durch die Parallelschaltung eines Kondensators C ("integrator") und eines Widerstandes R repräsentiert. In dieser Anordnung stellen die beiden Bauelemente einen einfachen Tiefpaß-Filter (RC-Glied) dar. Über den Eingang, den Synapsen, dargestellt durch Widerstände werden die EPSP's mit der jeweiligen Höhe an der Zellmembran integriert. Wenn am RC-Glied eine ausreichende Spannung anliegt, dann wird am Axonhügel eine Schwelle überschritten und ein elektrischer Impuls ausgelöst. Dieser Impuls kann ein Dirac'scher -Impuls sein. Mit einem zusätzlichen RCAP-Glied ist es möglich, den -Impuls in ein realistischeres Aktionspotential umzuformen. Wenn die Aktivierungsvariable x eines puls-gekoppelten Oszillators den Schwell-wert K erreicht, dann wird ein Aktionspotential ausgelöst und, und die Variable x wird auf Null zurückgesetzt. Die Zeit, die die Aktivierung benötigt, um von Null zum Schwellwert zu kommen, ist die Periode P. Die Frequenz des nichtlinearen Oszillators ist definiert durch P-1. Aktionspotential x(t) Schwelle t 0 T Zeitverlauf eines "integrate-and-fire"-Oszillators mit konstanter externer Erregung. x Kj Ki xj=f(j(t)) xi=f(i(t)) Tj Ti ___________________________________________________________________________________ Brain Modelling 57 Die Oszillatoren interagieren nur durch eine einfache Form der Impulskopplung. Wenn der ite Oszillator zum Zeitpunkt ti feuert, dann werden die Aktivierungen der anderen Oszillatoren j um einen Betrag angehoben. Die Kopplungskonstanten sind vergleichbar mit den EPSP's, >0, beziehungsweise den IPSP's, <0. Die Phase gibt die aktuelle Aktivierung den Zustand des Neurons, an. Das heißt: x (t ) 0 xi (t i ) K i i i x j (t i ) min( K j , x j (t i ) ) j i Die Oszillatoren haben haben individuelle Schwellwerte Ki (auf der y-Achse) beziehungsweise Ti (auf der x-Achse). Erreicht oder überschreitet eine Aktivierung xi den Schwellwert, dann wird seine Aktivierung infintesimal später auf Null gesetzt und alle anderen Aktivierungen werden um ein angehoben. Verhalten von zwei puls-gekoppelten "integrate and fire"-Neuronen: a) Zum Zeitpunkt t0+ gilt: Oszillator 1 hat gerade gefeuert: 1(t0+) = 0. Die Phase von Oszillator 2 liegt innerhalb der zwei zu bestimmenden Return-Grenzen und : 0 < < 2(t0+) < < T2. b) Als nächstes wird zum Zeitpunkt t1- Oszillator 2 den Schwellwert T2 erreichen: 2(t1-)=T2. Da sich Oszillator 1 ebenfalls gleichmäßig bewegt (für beide Oszillatoren vergeht gleich viel Zeit) ergibt sich 1(t1-) = T2 - 2(t0+). c) Infinitesimal später feuert Oszillator 2, da er seine Schwelle erreicht hat: 2(t1+)=0. Durch das Feuern wird Oszillator 1 um ein angehoben. Daraus folgt: 1(t1+) = g(+f(T2 - 2(t0+))). Es hätte passieren können, daß Oszillator 1 während der Bewegung (also zwischen den Zeitpunkten t0+ und t1-) von Oszillator 2, beziehungsweise durch das Feuern von Oszillator 2, also zu t1+, selbst über die eigene Schwelle T1 gehoben worden wäre. Das heißt beide Oszillatoren würden daraufhin gemeinsam feuern. Dies steht im Widerspruch zu den Anfangsbedingungen. Also muß gelten: 1(t1+) = g(+f(T2 - 2(t0+))) < T1. Um zu bestimmen wie groß 2(t0+) beim Start des Zyklus sein mußte, damit es zu keiner Synchronisation in diesem Schritt kommt, muß man die Ungleichung g(+f(T2-2(t0+))) < T1 nach 2(t0+) auflösen (Umkehrfunktion) und man erhält: T2- g[f(T1) - ] < 2(t0+) Damit haben wir die untere Grenze für 2(t0+) gefunden. d) Der Oszillator 1 wird als nächstes feuern (er hat ja seine Schwelle zum Zeitpunkt t 1+ noch nicht erreicht) Also ergibt sich für die Phase von Oszillator 1: 1(t2-) = T1. Da sich Oszillator 2 ebenfalls bewegt, lautet seine Phase dann: 2(t2-) = T1 - g(+f(T2 2(t0+))). e) Oszillator 1 feuert, seine Phase springt somit auf Null zurück und die Aktivität von Oszillator 2 wird um ein angehoben. Also gilt: 1(t2+) = 0 und 2(t2+) = g( + f(T1 g(+f(T2 - 2(t0+))))). Der Zyklus ist beendet. Aber wiederum wäre es möglich, daß diesmal Oszillator 2 über seine eigene Schwelle, durch das Feuern von Oszillator 1 angeregt, gehoben wird und es somit zum gleichzeitigen Feuern kommt. Damit dies nicht geschieht muß folgendes gelten: 2(t2+)=g(+f(T1-g(+f(T2-2(t0+))))) < T2. Durch Auflösung dieser Ungleichung nach dem Startwert 2(t0+) erhalten wir: 2(t0+) < T2 - gf(T1 - g[f(T2) - ]) - Mit diesem Zyklus ist es möglich zu zeigen unter welchen Bedingungen sich 2 Neuronen synchronisieren. ___________________________________________________________________________________ Brain Modelling 58 K1 K2 a) Os.2 Os.2 b) Os.1 1(t0+ t -)= T2-2(t0) 2(t0) Os.1 T1 Frequenzbedingung: g[f(T2)-] + g() < T1 T1 T2 Os.1 c) mit T1<T2 T2 1(t0+ t -)= T2-2(t0) Os.2 T1 T2 1(t0+ t+)=g(+f(T2-2(t0))) Was ist Synchronisation ? Phase-locking: Die Oszillatoren feuern zu unterschiedlichen Zeiten, aber die Phasendifferenz bleibt konstant, ist in der Regel aber nicht Null. Synchronisation: Die Oszillatoren feuern gleichzeitig, der Phasenunterschied ist nur beim Feuern Null. Supersynchronisation: Die Oszillatoren feuern gleichzeitig, der Phasenunterschied ist immer Null. Von was hängt Synchronisation ab ? a) Die Schwellwertfunktion muß nichtlinear sein. b) Die Frequenzen sollten in der selben Größenordnung sein. Das heißt die Schwellwerte sollten ungefähr gleich groß sein. c) Die EPSP´s müssen positiv sein. Wie kann man Synchronisation biologisch relevant beinflussen ? a) Eine Anhebung des Ruhemembranpotentials führt zu einem geringeren Schwellwert. Es erhöht sich die Frequenz,es kann schneller zu Synchronisation kommen. b) Wenn die EPSP´s erhöht werden, ist der Einfluß von einem Neuron auf andere Neuronen stärker. Es kann leichter zur Synchronisation kommen. Assemble 1 U Assemble 2 t a) b) Beispiel einer visuellen Szene (a). Die Bindung von Objektmerkmalen erfolgt durch zeitliche Korrelationen zwischen den neuronalen Antworten (b). Im hier gezeigten Fall - durch schwarze und weiße Kreise angedeutet - würden die beiden Personen durch jeweils ein unterschiedliches neuronales Assemble dargestellt. Die grauen Kreise stellen unkorrelierte Neuronen dar. ___________________________________________________________________________________ Brain Modelling 59 Ein typisches Experiment, in dem zwei Mikroelektroden aus dem visuellen Kortex eines Versuchstieres elek-trische Signale ableiten (a). Die Neuronen können nun mit verschiedenen Reizkonfigurationen aktiviert werden. Bietet man ein ein-ziges kohärentes Objekt an, in diesem Fall ein durchgehender vertikaler Lichtbalken, der über die rezeptiven Felder bewegt wird (b), so sind die Zellen an den beiden Ableiteorten synchron aktiv. Stimuliert man die gleichen Neuronen hingegen mit verschiedenen Objekten, etwa zwei klei-neren balkenförmigen Licht-reizen, die sich in verschiedene Richtungen bewegen (c), so sind die neuralen Impulse nicht mehr synchronisiert. a) 1 2 b) U t c) U t Hemmung von Assemblies: Hemmung durch Körnerzellen: Hemmung durch modulierende Neuronen: Muster erregende Neuronen hemmende Neuronen Verknüpfung von 2 Assemblies: ___________________________________________________________________________________ Brain Modelling 60 Das Liebesleben der Glühwürmchen Männliche Glühwürmchen blinken um Weibchen anzulocken - ein kurzer Lichtimpuls gefolgt von ein paar Sekunden der Dunkelheit. In den Weiten des Amazonas hat es ein einziges männliches Glühwürmchen schwer ein Weibchen anzulocken, zumal diese rund 50-100 Meter über dem Boden fliegen. Also versammeln sich die Männchen auf einem Baum. Damit haben es die Weibchen leichter das schwache Licht zu sehen. Aber die Glühwürmchen dürfen nicht durcheinander blinken. Die Weibchen reagieren nur auf ein regelmäßiges Blinken. Also müssen alle Glühwürmchen gleichmäßig blinken. Den ersten Berichten nach, die von diesem Phänomen berichteten, glaubte man an eine optische Täuschung. Man konnte sich nicht vorstellen, wie tausende und abertausende Glühwürmchen gleichzeitig blinkten. Zuerst glaubte man, daß es ein sogenanntes Chef-Glühwürmchen gibt, daß den Rhythmus vorgibt. Diese Annahme stellte sich als falsch heraus. Im Prinzip läßt sich dieses Problem durch Demokratie lösen. Zwei Würmchen entscheiden sich für eine Frequenz. Wenn ein neues hinzukommt, dann paßt sich das eine an den Rhythmus der anderen beiden an und umgekehrt und so weiter. Dieses Problem ist mit dem obigen Problem der Synchronisation verwandt, wenn nicht sogar ident. Die Ein Baum voller Glühwürmchen. mathematische Formel, mit der die optische Aktivierung beschrieben werden kann, ist ident mit der Gleichung, die die elektrische Aktivität im unterschwelligen Bereich eines Neurons beschreibt. Kein Glühwürmchen ist wichtiger, als die anderen. Der einzige Unterschied besteht darin, daß jedes Glühwürmchen mit jedem anderen Glühwürmchen die Information austauschen kann. Bei den Neuronen ist dies nicht gegeben, da die Anzahl der Synapsen begrenzt ist. Über die synaptische Stärke entsteht dann ein geometrisches Muster, das mit dem einlangenden Signal in irgendeiner Form korreliert. ___________________________________________________________________________________ Brain Modelling 61 8.0 Gestaltspsychologie "Das Ganze ist mehr als die Summe der Teile, auf gut Deutsch: das System ist nichtlinear." Eine Anwendung der Synchronisation (Figur-Hintergrund Trennung): Aus einem schlechten Bild wird das Muster vervollständigt. Es werden nur wenige Neuronen angeregt (die den Hund darstellen). Durch Synchronisation werden zusätzliche Neuronen angeregt, bis das Bild eines Hundes entsteht. Beim Kaniza-Dreieck (links) entsteht ein Dreieck vor dem geistigen Auge. Das Gehirn erkennt einige Strukturen und versucht diese zu vervollständigen. Am Beispiel des RubinPokals läßt sich das Umkippen aus einem Bild (Darstellung eines Pokals) in ein anderes (2 Gesichter sehen sich an) wiedergeben. Gruppierungsgesetze: räumliche Nähe Ähnlichkeit abgeschlossener Umriß gute Fortsetzung gemeinsamer Bereich Zusammenhang ___________________________________________________________________________________ Brain Modelling 62 Es kommt zu einer Konstruktion der Wahrnehmung (Scheinbewegung beim Zug). Wenn wir zewnig Information haben, dann fügen wir die am einfachsten erscheinenden Teile hinzu. Man vermutet, daß es sich ein Kreissegment hinter der Abdeckung befindet (links), aber es handelt sich um eine estwas verworrene Figur. Ein Rätsel aus der Gestaltspsychologie: Verbinden Sie die 9 Punkte durch 4 (3) Linien ohne abzusetzen. ___________________________________________________________________________________ Brain Modelling 63 9.0 Entwicklung des Nervensystems Verschiedene Stadien bei der Entwicklung des Nervensystems: [1] Induktion der Neuralplatte. [2] Vermehrung von Zellen in bestimmten Regionen (Die Vermehrungsrate ist nicht überall gleich groß. Daraus folgt das Relief der Hirnstrukturen). [3] Wanderung der Zellen vom Ort ihrer Entstehung zu den Stellen, an denen sie schließlich bleiben. [4] Bildung anatomisch identifizierbarer Zellverbände. [5] Ausreifung der einzelnen Nervenzellen. [6] Bildung von Verbindungen zwischen den Nervenzellen. [7] Selektiver Tod einzelner Nervenzellen (rund 50% sterben ab). [8] Umstrukturierung und Stabilisierung (Lebenslanges LERNEN !). ad 7) Bei den Nervenzellen werden offenbar alle diejenigen wieder eliminiert, die während der Ausdifferenzierung [ad 6] keinen Kontakt zu anderen Neuronen oder Erfolgsorganen bekamen. Somit ist eine ausreichende Reizlage während der frühen Entwicklung eines Menschen notwendig, um bestmögliche Innervierung frühzeitig zu gewährleisten. Nervenwachstumsfaktor unreifes Neuron Zellen des Zielgewebes Wachstumszone Nervenfaser (Axon) Innervierung des Zielgewebes Durchtrennung des Axon Im linken Bereich wirkt kein Nervenwachstumsfaktor auf das neurale Gewebe - im Gegensatz zur rechten Abbildung. ___________________________________________________________________________________ Brain Modelling 64 Topologische Abbildung des Großhirnrinde: somatosensorischen und motorischen Bereiches der Wie gelingt es dem Gehirn eine exakte Abbildung von der Körperoberfläche auf dem somatosensorischen Cortex durchzuführen ? Es müssen nachbarschaftserhaltende Beziehungen hergestellt werden. Es werden mehr Neuronen zur Verfügung gestellt als notwendig. Wenn nun zwei Neuronen gleichzeitig aktivert sind, wenn die beiden benachbarten (z.B. Haut-, Retina-) Areale aktiviert sind, dann ist zwischen den beiden Neuronen im (z.B. somatosensorischen, primären visuellen) Cortex eine nachbarschaftliche Beziehung hergestellt und diese Neuronen werden weiter benötigt. Sie sterben nicht ab. Es sterben nur die Neuronen ab, denen es nicht gelungen ist an der nachbarschaftserhaltenden Abbildung teilzunehmen. Die richtige Verschaltung (Bildung/Entstehung) setzt eine bestimmte Stimulierung des Gehirns voraus. Die einzelnen Gebiete müssen zeitlich-räumlich korrekt aktiviert werden. Dies geschieht in sensiblen Phasen während der Gehirnreifung. Wenn die Grobverschaltung abgeschloßen ist, müssen externe Reize aus der Umwelt die Feinabstimmung vornehmen. Damit kann sich das Gehirn auf die gegebene Umwelt optimal anpassen. ___________________________________________________________________________________ Brain Modelling 65 Entstehung der nachbarschaftserhaltenden Beziehungen (Topologie): t=2 t=3 t=1 t=4 t=5 t=6 t=7 t=8 t=9 Die spontane elektrische Aktivität in der sich entwickelnden Netzhaut ist lokal synchronisiert. In jedem Diagramm ist die Verteilung der Aktionspotential-Salven einzelner Ganglienzellen in der Retina eingetragen. Die Signale werden über den Thalamus zum primären visuellen Cortex weitergeleitet. Dort entstehen Signale die räumlich und zeitlich koordiniert sind. Retina Thalamus Gehirnrinde R Von der rechten Hälfte der Retina des rechten Auges innervieren Neuronen das rechte primäre visuelle Rindenfeld. Zwischen diesen Zielgebieten innervieren die Neuronen, kommend aus dem rechten Teil der Retina des linken Auges die Großhirnrinde. Es gibt ein Konkurenzverhalten. Damit beide Augen gleich stark vertreten sind, bedarf es eines externen Stimulus. Zum Zeitpunkt der Geburt überlappen die Neuronen im Zielgebiet LR L ___________________________________________________________________________________ Brain Modelling 66 Nach einer gewissen Zeit entstehen sog. Augendominanzsäulen im primären visuellen Areal. Die Säulen sind die kleinen weißen Rechtecke vom rechten Auge (untere Graphik). Dazwischen sind schwarze Säulen von linken Auge. In der oberen Graphik ist die Trennung noch nicht vollzogen. Die Schichtung des Cortex ist gut zu beobachten. Großhirnrinde Man kann sehr schön sehen, wie sich die Säulen durch die einzelnen Neuronen ausbilden. L R L R Thalamus von der Retina Wenn von einem Auge (in der linken unteren Graphik das linke Auge) keine Signale mehr an die Großhirnrinde weitergeleitet wird, dann verkümmern die dazugehörigen Neuronen. Wenn die Signale nicht koordiniert sind, dann wuchern sie und es bilden sich keine Augendominanzsäulen aus (rechte untere Graphik). Großhirnrinde L Großhirnrinde R L R Thalamus von der Retina L R L R Thalamus von der Retina ___________________________________________________________________________________ Brain Modelling 67 Training führt zu einer Ausdehnung solcher Flächen des Cortex, in denen sensorische Afferenzen eintreffen. Ein Affe wurde trainiert eine Aufgabe durchzuführen, bei der er häufig die Spitzen des zweiten und dritten, gelegentlich auch des vierten, Fingers einsetzen mußte. Die Repräsentation der Fingerspitzen vor Trainingsbeginn im Vergleich zur Phase nach der starker Stimulation. Die Entwicklung des menschlichen Gehirns ist erst nach der Pupertät abgeschlossen. Manche Gebiete werden erst sehr spät in das Netzwerk Gehirn integriert. Zum Beispiel der Hippocampus. Diese Struktur ist für unser deklaratives Lernen verantwortlich. Die meisten Menschen erinnern sich praktisch kaum an erlebte Fakten vor dem 3. Lebensjahr - der Hippocampus wird erst zu diesem Zeitpunkt angeschlossen. Die Myelinisierung der Axone gibt über den Entwicklungsstand der einzelnen Rindenareale Auskunft. Die dunklen Areale werden früh, die hellgrauen später und die weißen sehr spät (bis in die Pupertät hinein) myelinisiert: Synapsenwachstum im visuellen Cortex Synapsenwachstum im Stirnlappen 0 2 4 6 7 10 Zeit in Jahren 20 Entwicklung der Großhirnrinde auf zellulären Niveau: ___________________________________________________________________________________ Brain Modelling 68 Prägung:Als Prägung wird ein artspezifisch bedeutsamer Informationserwerb während sensibler Entwicklungsphasen bezeichnet, der irreversible Veränderungen bedingt. Die Prägung befähigt ein Individuum zum dauerhaften Wiedererkennen der gleichen Merkmalsgestaltung, die später immer gleichartige Verhaltensreaktionen auslöst. (Hospitalismus) 10.0 Das visuelle System Das menschliche Auge ist nicht einfach mit einer Videokamera oder einem Fotoapparat zu vergleichen. So betrachtet das Auge einzelne Ausschnitte des Gesichtsfeldes öfters als andere. Lederhaut Netzhaut Aderhaut Hornhaut Sehgrube Sehnerv Iris Linse Bipolarzelle Zäpfchen Darstellung des Auges (oben) und der neuralen Verschaltungen in der Retina. Licht Rückseite des Augapfels retinale Ganglienzelle Stäbchen ___________________________________________________________________________________ Brain Modelling 69 Es gibt zwei Arten von Photorezeptoren: Stäbchen und Zapfen. Die Stäbchen können sehr schwaches Licht detektieren. Dies liegt vorallem an der neuralen Nachverarbeitung, die für eine sehr gute Verstärkung sorgt. Sie dienen der Schwarz-Weiß Verarbeitung eines Bildes. Der Konvergenzgrad auf die nachgeschaltenen Bipolarzellen ist sehr groß. Die Zapfen dienen dem Farbsehen.Durch sie sehen wir schärfer und dieser Rezeptortyp besitzt eine bessere zeitliche Auflösung. Die drei Zapfen der menschlichen Retina antworten bevorzugt , aber nicht ausschliesslich auf die jeweilige Wellen-länge, wie hier durch die Absorptionsspektren der Photopigmente aller drei Zapfentypen dargestellt ist. Die Kurve mit dem Maximum bei 496 nm zeigt das Absorptionsspektrum von Stäbchen. Es gibt ungefähr 20 mal mehr Stäbchen als Zapfen. Zwischen den Photorezeptoren und den Ganglienzellen liegen drei Klassen von Interneuronen: Bipolar-, Horizontal- und Amakrinzellen. Sie kombinieren die Signale von mehreren Photorezeptoren, so daß die Antwort der Ganglienzellen stark von räumlichen und zeitlichen Lichtmustern abhängt. Die Axone dieser Zellen bilden den Sehnerv. Ganglienzellen besitzen ein rezeptives Feld, das einen bestimmten Retinabereich überwacht. Zwei wesentliche Merkmale einer Ganglienzelle: 1) Das rezeptive Feld ist kreisförmig. 2) Das rezeptive Feld teilt sich in zwei Bereiche: a) einer kreisförmigen Zone im Inneren und b) dem Umfeld Ganglienzelle mit ON-Zentrum Ganglienzelle mit OFF-Zentrum ON-Bereich OFF-Bereich OFF-Bereich ON-Bereich Lichtpunkt auf das Zentrum Lichtpunkt auf das Zentrum Lichtpunkt in das Umfeld Lichtpunkt in das Umfeld Beleuchtung des gesamten Zentrums Beleuchtung des gesamten Zentrums Beleuchtung des gesamten Umfelds Beleuchtung des gesamten Umfelds diffuse Beleuchtung diffuse Beleuchtung ___________________________________________________________________________________ Brain Modelling 70 Farbwahrnehmung: Farbantagonismus: Bestimmte Farben können nie in Kombination gesehen werden. Es existiert kein rötliches Grün oder bläuliches Gelb. Farbsimultankontrast: Dieser Effekt tritt dann auf, wenn Gegenfarben von räumlich benachbarten Stellen ausgehen. So hebt sich ein grünes Objekt besser von einem roten Hintergrund besser ab, als vor einem blauen. Farbkonstanz: Die Farbe eines Objektes bleibt relativ konstant, obwohl enorme Schwankungen in der Spektralverteilung der Umgebungsbeleuchtung auftreten. Farbe wird in der Retina und im seitlichen Kniehöcker des Thalamus durch Gegenfarbzellen codiert. G+;R+ G-;R- G-;R- G+;R+ G+ G- R- R+ konzentrische Breitbandzellen konzentrische einfache Gegenfarbenzellen R+ R- G- G+ B+ G-;R- BG+;R+ coextensive einfache Gegenfarbenzellen Konzentrische Breitbandzelle: Die Ein-gangssignale von den G- und R-Zapfen wer-den im rezeptiven Feld unabhängig vonein-ander aufsummiert. Sie reagieren auf den Helligkeitskontrast innerhalb ihres Feldes und leisten keinen Beitrag zum Farbsehen. Konzentrische einfache Gegenfarbzellen: Ein Zapfentyp (R oder G) überwiegt im Zentum des rezeptiven Feldes und führt zu einer Reaktion, die zu der des im Umfeld überwiegenden Zapfentyps entgegengesetzt ist. Coextensive einfache Gegenfarbzellen: Sie besitzen ein undifferenziertes rezeptives Feld in dem die Wirkung der B-Zapfen der kombinierten Wirkung von G- und R-Zapfen gegenübersteht. tatsächliche Intensität Machbänder beobachtete Intensität Durch die laterale Hemmung in der Retina werden Kanten mit einer anderen Helligkeit gesehen, als es tatsächlich ist. Dadurch wird der Kontrast erhöht. ___________________________________________________________________________________ Brain Modelling 71 intensives Licht schwaches Licht Das rechte (linke) Halbbild der Retina beider Augen wird über den Sehnerv zum rechten (linken) seitlichen Kniehöcker (Teil des Thalamus - Corpus geniculatum laterale) abgebildet. Der seitliche Kniekörper liefert eine vollständige Repräsentation der jeweiligen Gesichtshälften. Er besitzt sechs Zellkörperschichten. Die Fasern von der kontra-lateralen Retinahälfte enden in den Schichten 1, 4 und 6, die von der ipsilateralen Hälfte in den Schichten 2, 3 und 5. Die Schichten 1 und 2 stellen die magno-zelluläre Bahn dar, im Gegensatz zu den Schichten 3 bis 6 die die parvozelluläre Bahn repräsentieren. Beide Bahnen projezieren zum primären visuellen Cortex. Aus diesen Bahnen entstehen die 3 Hauptkanäle für die Wahr-nehmung von Form, Farbe und Bewegung. Von den Ganglienzellen der Netzhäute ziehen ausgedehnte Nervenfasern (Sehnerv) zu den äußeren Kniekörpern, deren Zellen ihrer-seits durch Nervenfasern unmittelbar mit den Zellen des primären Sehfeldes ver-bunden sind. Die sechs Schichten der Kniekörper sind durch je eine Nervenzelle angedeutet und es läßt sich erkennen, daß diese Schichten jeweils nur mit Signalen entweder vom linken oder vom rechten Auge versorgt werden. ___________________________________________________________________________________ Brain Modelling 72 Durch eine spezielle Färbetechnik (Cytochron-Oxidase) zeigen sich charakteristische Zellsäulen in der V1 Region, die von der Oberfläche der grauen Hirnrinde bis zur weißen Substanz - den darunterliegenden Nervenfasern - verlaufen. Auf einem Schnitt parallel zur Oberfläche erscheinen sie als stark angefärbte Flecken, die nach dem entsprechenden englischen Wort als Blobs bezeichnet werden; dazwischen liegen weniger stark gefärbte Interblobregionen. 1) Streifen in A18 2) Blobs in A17 V2 V1 Es existieren 3 verschiedene Bahnen für das Wahrnehmen von Bildern. 1) Parvo-Blob Bahn: parvozelluläre Schicht des seitlichen Kniehöckers Blob-Region von V1 Gebiet der dünnen Streifen in V2 Cortexareal V4 (Wahrnehmung von Farbe) 2) Parvo-Interblob Bahn: parvozelluläre Schicht des seitlichen Kniehöckers Interblobregion von V1 Gebiet mit blassen Streifen in V2 Cortexareal V4 (Wahrnehmung von Formen und Farbe) 3) Magno-Interblob Bahn: magnozelluläre Schichten des seitlichen Kniehöckers Interblobregion von V1 Gebiet mit dicken Streifen von V2 Cortexareal V5 und V3 (Wahrnehmung von Tiefe und Bewegung) Die Neuronen im visuellen Cortex können (funktionell) in zwei Klassen unterteilt werden: einfache Zellen und komplexe Zellen. In V2, ein dem primären visuellen Cortex benachbartes Areal, bilden die stark färbenden Regionen zwei Typen von dunklen Streifen - dicke und dünne, die durch blasse InterstreifenRegionen voneinander getrennt sind. ___________________________________________________________________________________ Brain Modelling 73 Einfache Zellen:Ihre rezeptiven Felder erhalten mehr Input als die Ganglienzellen. Sie sind auch nicht kreisrund, sondern länglich, meist sogar fast rechteckig. Das rezeptive Feld gesteht aus einer rechteckigen On-Zone mit einer eindeutig orientierten Längsachse, flankiert mit Off-Zonen von beiden Seiten. Um optimal wirksam zu sein, muß der Stimulus genau die gleiche Orientierung haben, wie das rezeptive Feld der Zelle. Es können Reize mit einem Orientierungs bereich von rund 10° erkannt werden. Der Reiz darf nur die excitatorische Region des rezeptiven Feldes bedecken und nicht in die inhibitorische Region hineinreichen. Komplexe Zellen:Auch sie besitzen rezeptive Felder mit bestimmter Orientierung. Ihre rezeptiven Felder sind jedoch größer als die der einfachen Zellen und sie besitzen keine klar umgrenzten On- und Off-Zonen. Neben Kontrasten und schnellen Belichtungsänderungen analysiert unser Sehsystem auch andere Aspekte, wie Farbe, Form und Bewegung: Die Neuronen des primären visuellen Cortex haben lineare rezeptive Felder mit diskreten excitatorischen und inhibitorischen Bereichen. rezeptives Feld: OFF-Bereich Eine einfache Zelle antwortet auf einen Lichtbalken einer Orientierung, die zur Orientierungsachse ihres rezeptiven Feldes paßt. ON-Bereich Augendominanzsäule: Blobs I II III IV V VI vom rechten - linken Auge Ein Modul aus unterschiedlichen funktionalen Kolumnen im visuellen Cortex enthält alle neuralen Strukturen, die zur Analyse einer diskreten Region des gesichtsfeldes benötigt wird. Jedes Modul, das auch Hyperkolumne genannt wird, enthält ein vollständiges Set an Orientierungssäulen, die 360° repräsentieren, eine linke und eine rechte Augendominanzsäule und mehrere Blobs (wichtig für die Farbverarbeitung). Das gesamte Gesichtsfeld ist im Cortex durch ein regelmäßiges Muster aus Hyperkolumnen repräsentiert. ___________________________________________________________________________________ Brain Modelling 74 Modell für die visuelle Informationsverarbeitung: Verschaltungen im visuellen System: Das Geheimnis der visuellen Wahrnehmung liegt in der komplizierten Arbeitsteilung verborgen. Anatomisch zeigt sich darin, daß bestimmte Regionen und Teilregionen der Hirnrinde auf einzelne visuelle Funktionen spezialisiert sind. Anatomischer Schnitt durch die Sehrinde von einem Makaken. Auf dem Querschnitt ist ein Teil der primären Sehrinde (V1) und einige Areale im prästriären visuellen Cortex (V2-V5) markiert. Die Areale V1 und V2, fungieren als eine Art Postamt um die verschiedenen Signale auf die richtigen Areale zu verteilen. Diese beiden Gebiete analysieren stückweise das Gesichtsfeld. Es wurde herausgefunden, daß in den Blobs von V1 farb- und in den Interblob-Regionen formspezifische Zellen konzentriert sind. Besonders stark ausgeprägt sind die Zellsäulen in der zweiten und dritten Schicht von V1, deren Input von den parvozellulären Schichten des seitlichen Kniehöckers stammen. Die Zellen in diesen Kniehöcker-Schichten reagieren mit einer hohen, lang anhaltenden Aktivität auf visuelle Reize und sind größtenteils farbempfindlich. ___________________________________________________________________________________ Brain Modelling 75 parietale Areale Bewegung V5 V3 V4 Farbe V1 V2 Form temporale Areale In dieser Seitenansicht sind die wichtigsten Verbindungen eingetragen. Die Information wird aber nicht nur von einem Areal zum nächsten Areal vorwärtsvermittelt, sondern es gibt auch starke Rückwärtsverbindungen. Ein Areal vergleicht seine neu gebildetes neurales Assemble mit den ursprünglichen Informationen. Dies kann zum Beispiel zu einer Kontrastverstärkung führen (Rückverbindung von V1 und CGL - corpus geniculatum laterale - seitlicher Kniehöcker). Stark vereinfachend sind hier die drei Objektmerkmale Form, Farbe und Bewegung einzelnen Arealen zugeordnet. Auf jeder Verarbeitungsstufe hat jede Zelle eine höhere Abstraktionsfähigkeit als die Zellen der vorangegangenen Stufen. Sehen erfordert Aufmerksamkeit ! Welche Merkmale sorgen dafür, daß ein Objekt aus dem Hintergrund hervortritt ? Deutlich wahrnehmbare Objektgrenzen beruhen auf elementaren visuellen Eigen-schaften wie Helligkeit, Farbe und Orientierung von Linien. Wo ist das Gebiet mit den T´s ? Folgerung: Es existieren zwei (un)abhängige Wege der Verarbeitung von visueller Information. [1] Ein schnell ablaufender nicht aufmerksamkeitsgesteuerter Prozeß, der nur für das Erkennen der wichtigsten Umrisse eines Objektes sorgt (Hauptmerkmale wie globale Textur eines Objekts). [2] Der aufmerksamkeitsgesteuerte Prozeß führt zu einer Fokussierung auf feinere Merkmale eines Objektes. Hierbei werden Merkmale ausgewählt und hervorgehoben, die in verschiedenen Merkmalskarten (nachgeschalteten Cortexbereichen) verzeichnet sind (the winner takes it all). ___________________________________________________________________________________ Brain Modelling 76 Neokognitron: Das Neokognitron versucht stark deformierte Buchstaben wiederzuerkennen. In diesem Netzwerk gibt es keine vollständige Translations-, Rotations- und Skalierungsinvarianz, sondern nur die Fähigkeit, begrenzte Veränderungen der gelernten Muster zu tolerieren. Wie man sieht, nimmt jedes Neuron der Schicht 1 nur die Ausgabe einer lokal begrenzten Menge von Neuronen der Schicht 0 wahr. An jeder Stelle der Schicht 1 antworten nur diejenigen Neuronen besonders stark, deren rezeptive Felder in ihrer Orientierung mit der lokalen Orientierung der Striche des "A" übereinstimmen. In der nächsten Schicht werden schon komplexere Merkmale erkannt (Ecken, Enden). Beispiele für deformierte mit dem Neocognitron richtig erkannte Zahlen. ___________________________________________________________________________________ Brain Modelling 77