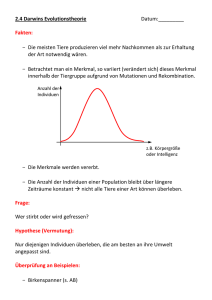
Untersuchungen zur Obliteration der Nähte des Schädels
Werbung

Altersschätzung von Skelettmaterial – Untersuchungen zur Obliteration der Nähte des Schädels Ronny Bindl, Rainer Nitsche und Horst Bruchhaus Zusammenfassung – Das Ziel der vorliegenden Studie ist die Untersuchung der Relationen zwischen der Obliteration der Schädeldachnähte und den alterspezifischen Veränderungen am Postcranium am Skelettmaterial des Gräberfeldes Briesnitz (Dresden; 10. bis 13. Jahrhundert). Des Weiteren sollten unter Berücksichtigung der Obliteration Hinweise zum Sterbealter der einzelnen Skelettfunde erarbeitet werden. Die Informationen zum Sterbealter spielen im vorliegenden Fall neben der Geschlechtsbestimmung und der Relation der Knochen zueinander eine wichtige Rolle, da bei dem Gräberfeld die Gräber z. T. in stark gestörtem bzw. verwühltem Zustand vorliegen. Anschließend kann mit Hilfe der gewonnenen Ergebnisse die Gräberfeldbereinigung abgeschlossen werden. In die vorliegende Untersuchung gingen 234 Skelettindividuen des Altersbereiches Juvenil und älter ein. Die Beurteilung der Obliteration der Nähte des Schädeldaches erfolgte nach BROCA (1861) und NEMESKÉRI et al. (1966) sowie nach dem Verfahren von MEINDL/LOVEJOY (1985) an der Tabula interna sowie der Tabula externa. Die Obliterationsgrade der jeweiligen Nahtabschnitte bzw. vollständig erhaltener Nähte wurden danach mit den Angaben zum Sterbealter, der für die Altersschätzung verwendeten Knochen, des postcranialen Skeletts verglichen und in Kreuztabellen gegenübergestellt. Erwartungsgemäß sind die Ergebnisse an der Tabula interna bessere als an der Tabula externa. An allen Abschnitten der Sutura sagittalis (T. interna) wurden deutlich signifikante Zusammenhänge zum Skelettalter ermittelt. Die unter Auslassung der Abschnitte C3 und L3 berechneten Obliterationsgrade führten zu einer deutlich besseren Übereinstimmung zwischen den altersspezifischen Veränderungen am Postcranium und dem Nahtverschluss. Mit dem Verfahren von MEINDL/LOVEJOY (1985) konnte keine Verbesserung der Ergebnisse am vorliegenden Material erreicht werden. Außerdem hat sich gezeigt, dass sich das Verfahren am historischen Skelettmaterial mit schlechtem Erhaltungszustand kaum anwenden lässt. Schlüsselwörter – Altersschätzung, Cranium, Obliteration Abstract – The aim of this study is the research of the relation between the obliteration and the age specific changes of the postcranial skeleton based on the material of the graveyard of Briesnitz (Dresden; 10th – 13th century). Furthermore, the obliteration should be used to estimate age at death of the single skeletons. In the present case the informations about age at death, sex estimation and the relation of the bones to each other are very important because of the bad state of preservation and the confused situation of the burials. Following this identification results the revision of the graveyard could be finished. In the present study 234 skeletal individuals with juvenile age or older were implicated. The evaluation of the obliteration of the skullsutures occurred according to BROCA (1861) and NEMESKÉRI et al. (1966) as well as to the method of MEINDL/LOVEJOY (1985) on the Tabula interna and the Tabula externa. The degree of obliteration at the sutures was compared with the estimated age of the postcranial skeleton. This results were compared in cross tabulations. It could be shown that, as expected, the results at the Tabula interna were better then the results of the Tabula externa. All parts of the Sutura sagittalis (T. interna) showed significant high correlations with age. The degrees of obliteration, which were calculated without the parts C3 and L3, lead to an explicit better correlation between age at death and the obliteration. The method of MEINDL/LOVEJOY (1985) was not able to upgrade the results at the present material. Furthermore this method was hardly applicable to the historic material with a bad state of preservation. Keywords – age estimation, crania, obliteration Das Sterbealter ist neben dem Geschlecht einer der wichtigsten Parameter zur Individualanalyse von Skelettfunden und somit ein wesentlicher Bestandteil der Rekonstruktion prähistorischer und historischer Populationen. Zur Ermittlung des Sterbealters von Skelettindividuen stehen eine Vielzahl von morphologischen, röntgenologischen sowie histologischen und chemischen, für subadulte Individuen zusätzlich auch metrische Verfahren zur Auswahl. Die Anwendung der einzelnen Methoden zur Altersschätzung richten sich dabei vor allem nach der Erfahrung des Bearbeiters und den technischen Möglichkeiten des jeweiligen Institutes. Im Hinblick auf eine verhältnismäßig schnelle und damit auch kostenarme Bearbeitung spielen morphologische Verfahren zur Altersschätzung eine wichtige Rolle. Dabei fanden vor allem die Veränderungen am Os coxae (Facies symphysialis, Facies auricularis und Acetabulum) sowie am Cranium (Obliteration der Nähte des Schädeldaches, wenn möglich in Verbindung mit der Abrasion der Zähne) Eingang in zahlreiche Studien. Die Ungenauigkeit der morphognostischen inkl. röntgenologischen Methoden, bedingt durch die große Variabilität der altersspezifischen Veränderungen, wurde und wird dabei häufig kritisiert. Weiterführende histologische Untersuchungen am Skelett - insbesondere die Wurzelzementannulation an den Zähnen - werden im Gegensatz zu den morphologischen Methoden als wesentlich genauer eingeschätzt, sind aber mit einem höheren technischen und zeitlichen Aufwand verbunden. Die Voraussetzung für den Einsatz morphognostischer, röntgenologischer und histologischer Archäologische Informationen 30/1, 2007, 25-33 Bulletin de la Société Suisse25d‘Anthropologie 13 (1), 2007 Bulletin der Schweizerischen Gesellschaft für Anthropologie 13 (1), 2007 Ronny Bindl, Rainer Nitsche und Horst Bruchhaus Verfahren sind dabei genaue Kenntnisse, der in der Literatur beschriebenen Verfahren. Insbesondere im ersten Schritt sollte bei der Graduierung, d.h. der Einteilung der altersspezifischen Veränderungen in Stadien, die praktische Erfahrung die Basis für jegliche Anwendung sein. Grundsätzlich sollten im Vorfeld solcher Studien Skelettmaterial mit bekanntem Alter und Geschlecht in genügend hoher Individuenanzahl untersucht werden. Somit wird der subjektive Faktor bei der Materialaufnahme aber vor allem auch bei der Interpretation der Ergebnisse durch die erhöhte Genauigkeit deutlich minimiert. Im zweiten Schritt können die aus den Untersuchungen am rezenten Material gewonnenen Erkenntnisse auf die Sterbealtersschätzung des (prä-)historischen Skelettmaterials angewendet werden. Obliterationsgrad der Suturae craniales - geschätztes Alter Nahtabschnitt Tabula externa Sutura coronalis 1 (R / L) 2 (R / L) 3 (R / L) Sutura sagittalis 1 2 3 4 Sutura lambdoidea 1 (R / L) 2 (R / L) 3 (R / L) Tabula interna Sutura coronalis 1 (R / L) 2 (R / L) 3 (R / L) Sutura sagittalis 1 2 3 4 Sutura lambdoidea 1 (R / L) 2 (R / L) 3 (R / L) Die Untersuchungen zu den alterspezifischen Veränderungen am Skelett sowie an den Zähnen haben seit jeher einen hohen Stellenwert am Institut für Humangenetik und Anthropologie der FSU Jena und sind Bestandteil zahlreicher Arbeiten (u. a. LISS 2001, FORSTER 2002, BIEDERMANN 2003, GOTTSCHALD 2003, VOIGT 2006, SEEBER 2007, KAISER 2007, VOLKMANN 2007, NITSCHE in Vorb.). Auch die Untersuchungen zur Obliteration der Nähte des Schädeldaches insbesondere am alters- und geschlechtsbekanntem Skelettmaterial fand Eingang in zahlreiche Arbeiten (u. a. HILDEBRAND/WUTSCHKE 1987, VOIGT et al. 2006). Die Bedeutung der Nahtobliteration als ein wichtiges Hilfsmittel zur Sterbealtersschätzung von Skelettmaterial wurde dabei grundsätzlich bestätigt. Auf den Einsatz dieses Verfahrens kann insbesondere bei der Bearbeitung von (prä-) historischem Skelettmaterial mit z. T. sehr schlecht erhaltenem, d. h. unvollständigem und fragmentiertem Skelettmaterial zur Unterstützung der Altersschätzung nicht verzichtet werden. Aufbauend auf die Arbeiten an rezentem Material (vgl. Tab. 1a-c), mit dessen Hilfe der Einblick in die Beurteilung der alterspezifischen Veränderungen erarbeitet wurde, ist nun das Ziel der vorliegenden Studie die Untersuchung der Relationen zwischen der Obliteration der Schädeldachnähte und den alterspezifischen Veränderungen am Postcranium. Des Weiteren soll am Skelettmaterial des Gräberfeldes Briesnitz (Dresden; 10. bis 13. Jahrhundert) unter Berücksichtigung der Obliteration ein weiterer Beitrag zur Altersschätzung erarbeitet werden. Die Informationen zum Sterbealter spielen eine besonders große Rolle, da bei dem vorliegenden Gräberfeld die Gräber z. T. in stark gestörtem bzw. verwühltem Zustand aktuell Obliterationsgrad 0 4 Perzentile Perzentile 90% 50% 50% 75% 75% 90% Alter (unter) * Alter (über)* 40 42 35 55 58 45 74 73 59 66 66 59 48 59 50 40 56 37 42 35 29 36 56 46 38 51 69 61 57 64 57 55 54 57 43 43 40 43 37 37 34 36 39 36 46 51 51 59 68 61 71 62 67 62 52 47 38 39 32 28 27 39 36 34 52 49 46 55 55 56 44 43 42 36 35 38 29 26 25 26 36 35 35 32 51 55 52 56 54 56 54 55 43 45 46 43 36 36 35 37 33 30 41 43 40 52 55 52 63 59 53 61 46 42 46 39 37 40 * ( R / L ): Der jeweils ungünstigere Wert wurde verwendet. Tab. 1a Altersverteilung nach den Obliterationsgraden der einzelnen Nahtabschnitte der rezenten Sammelserie (VOIGT et al. 2006). Altersklasse in Jahren ES Punkte 20-29 30-39 40-49 50-59 60-69 1 0 - 1,4 51,0 2 1,5 - 9,4 23,0 5,0 6,0 3 9,5 - 19,4 9,0 7,0 2,0 4 19,5 - 29,4 6,0 20,0 6,0 5 29,5 - 39,4 6,0 20,0 6 39,5 - 46,4 3,0 12,0 7 46 < 2,0 24,0 n 16,0 19,0 12,0 4,0 - >70 n - - 11,0 - - - 11,0 2,0 3,0 - 6,0 4,0 - 6,0 4,0 21,0 15,0 3,0 6,0 7,0 18,0 35,0 34,0 7,0 12,0 43,0 44,0 60,0 81,0 22,0 20,0 21,0 17,0 7,0 ES - Entwicklungsstadien; n - Anzahl Tab. 1b Prozentuale Verteilung der einzelnen Kalotten der rezenten Sammelserie entsprechend des Obliterationsgrades ihrer Schädelnähte innerhalb festgelegter Altersklassen über definierte Entwicklungsstadien (VOIGT et al. 2006). vorliegen, und daher alle Informationen im Hinblick auf die Individualanalyse, insbesondere die Ergebnisse der Altersschätzung und Geschlechtsbestimmung notwendig sind, um die einzelnen Skelettelemente sicher zuordnen zu können. In die vorliegende Untersuchung gingen 234 Skelettindividuen des Gräberfeldes Briesnitz 26 Altersschätzung von Skelettmaterial – Untersuchungen zur Obliteration der Nähte des Schädels Altersklasse in Jahren ES Punkte 50-59 60-69 >70 Alter in Jahren Vmin Vmax 20-29 30-39 40-49 1 0 - 1,4 76,0 17,9 6,0 - - - 20 43 2 1,5 - 9,4 67,6 (16,2) (16,2) - - - 20 48 - 25 60 3 9,5 - 19,4 39,1 30,4 (10,1) (10,1) (10,1) 4 19,5 - 29,4 14,3 47,6 14,4 9,5 (7,1) (7,1) 27 75 5 29,5 - 39,4 8,5 28,2 29,6 21,1 (6,3) (7,1) 27 72 6 39,5 - 46,4 2,8 11,0 16,5 32,1 32,1 6,4 28 81 7 46 < 0,8 9,4 16,9 17,3 23,6 31,9 24 81 ES - Entwicklungsstadien; n - Anzahl Tab. 1c Prozentuale Verteilung der einzelnen Kalotten der rezenten Sammelserie entsprechend des Obliterationsgrades ihrer Schädelnähte innerhalb festgelegter Entwicklungsstadien über aufeinander folgende Altersklassen (VOIGT et al. 2006). (Dresden, Sachsen) des Altersbereichs Juvenil und älter ein. Aufgrund der schwierigen Fundsituation konnten mit der notwendigen Sicherheit zum aktuellen Stand lediglich zu 138 Individuen Angaben zum Sterbealter des postcranialen Skeletts getroffen werden. Die Sterbealtersschätzung am Postcranium erfolgte nach den Verfahren am Os coxae nach MCKERN/STEWART 1957, OWINGS 1981 und OWINGS-WEBB/SUCHEY 1985 an der Crista iliaca, nach TODD 1923, MCKERN/STEWART 1957, NEMESKÉRI et al. 1960, GILBERT/MCKERN 1973, SUCHEY 1979, BROOKS/SUCHEY 1990 und BRUCHHAUS et al. 2003 an der Facies symphysialis sowie nach LOVEJOY et al. 1985 und BUCKBERRY/CHAMBERLAIN 2002 an der Facies auricularis und nach SZILVÁSSY 1977 und OWINGS 1981 an den sternalen Gelenkfläche der Claviculae. Die Individuen wurden in die anthropologischen Altersgruppen JuvenilAdult (J/A), Spätadult-Matur (SA/M) und Spätmatur-Senil (SM/S) eingeteilt. Dabei wurden die Gruppen so gewählt, dass sich die Randbereiche aufgrund der Streuung der morphologischen Merkmale deutlich überlappen, die zentralen Bereiche der Altersgruppen sich aber deutlich unterscheiden. Die Aufnahme der Obliteration der Nähte des Schädeldaches erfolgte nach dem allgemein verwendeten Schema nach BROCA (1861) und nach der Stadieneinteilung nach NEMESKÉRI et al. (1966) von 0 (Naht vollständig offen) bis 4 (Naht vollständig verstrichen) an der Tabula interna sowie der Tabula externa. Aus den Stadien der Obliteration der untersuchten Nahtabschnitte wurden die Mittelwerte (Obliterationskoeffizienten) für verschiedene Nähte gebildet (vgl. Abb. 1; Tab. 2). Des Weiteren wurde die Obliteration nach dem Verfahren von MEINDL/LOVEJOY (1985) aufgenommen und ausgewertet. Bei diesem Verfahren erfolgt die Einteilung in vier Stadien (0: Naht vollständig offen bis 3: Naht vollständig verstrichen). Die Obliteration wird an 16 ein cm großen Abb. 1 Übersicht über die Nähte des Schädeldaches nach BROCA (1867; verändert). Arealen am Schädel aufgenommen (vgl. Abb. 2.). Obwohl die Methode nur eine Aufnahme an der Tabula externa verlangt, wurde in Bezug auf eine vollständige Materialaufnahme auch die Veränderungen an der Tabula interna aufgenommen. Die Auswertung der Obliteration nach MEINDL/ LOVEJOY (1985) erfolgt durch die Bildung von S. coronalis ( C ) links C3 C2 rechts C1 C2 C3 MW C1+2+3 MW C1+2+3 MW links C1+2 rechts C1+2 MW links C1+2+3 rechts C1+2+3 S. saggitalis ( S ) S1 S2 S3 S4 MW S1+2+3+4 S. lambdoidea ( L ) links L3 L2 rechts L1 MW L1+2 MW L1+2+3 L1 L2 Tabula externa Tabula interna (ohne C3+L3) C1 MW C1+2 Tabula externa (ohne C3+L3) Tabula interna MW C1+2 L3 MW L1+2 MW L1+2+3 MW links L1+2 rechts L1+2 MW links L1+2+3 rechts L1+2+3 MW Tabula interna (ohne C3+L3) + Tabula externa (ohne C3+L3) MW Tabula interna + Tabula externa S - Sutura MW - Mittelwert Tab. 2 Übersicht über die im Rahmen der vorliegenden Arbeit untersuchten und ausgewerteten Nahtabschnitte. 27 aktuell Ronny Bindl, Rainer Nitsche und Horst Bruchhaus Nahtabschnitt Tabula interna Tabula externa 0,156 0,802 0,246 0,849 0,141 0,844 0,187 0,523 0,054 0,842 0,214 0,731 0,097 0,669 0,070 0,484 0,078 0,158 0,445 0,127 S. coronalis 0,177 0,222 0,274 0,270 0,136 0,189 0,054 0,554 0,150 0,111 0,261 0,234 0,088 0,563 Gesamt 0,619 0,131 0,354 Gesamt ohne C3 0,049 1 0,091 0,537 2 0,010 0,472 3 0,003 0,128 S. sagittalis 4 0,011 0,047 0,013 0,478 Gesamt 0,014 links 0,046 0,156 rechts 0,067 0,423 1 MW 0,026 0,163 MW+li+re* 0,008 0,163 links 0,064 0,283 rechts 0,017 0,160 2 MW 0,004 0,292 MW+li+re* 0,016 0,076 links 0,090 0,234 rechts 0,208 0,528 L. lamdoidea 3 MW 0,046 0,542 MW+li+re* 0,426 0,231 links 0,216 1,000 links L1+L2 0,021 0,094 rechts 0,200 0,374 rechts L1+L2 0,012 0,192 0,220 0,314 Gesamt 0,034 0,011 0,217 Gesamt ohne L3 0,020 0,044 0,591 Gesamt ohne C3+L3 0,257 0,407 Gesamt 1,000 links rechts 3 MW MW+li+re* links rechts 2 MW MW+li+re* links rechts 1 MW MW+li+re* links links C1+C2 rechts rechts C1+C2 Abb. 2 Übersicht über die Nähte des Schädeldaches nach MEINDL/LOVEJOY (1985). Punktsummen einerseits für die Nahtabschnitte des Schädeldachsystems und andererseits für die Nahtabschnitte des vorderen Nahtkomplexes (vgl. Tab. 3). Die Obliterationsgrade der jeweiligen Nahtabschnitte bzw. Nähte wurden danach mit den Altersgruppen vom postcranialen Skelett verglichen und in Kreuztabellen gegenübergestellt. Die statistischen Zusammenhänge wurden mit Hilfe des Chi-Quadrat-Tests geprüft und die Grenze des Signifikanzniveaus bei p≤0,1 festgelegt. Aufgrund der verhältnismäßig wenigen Individuen zu denen postcraniale Angaben zum Sterbealter vorliegen wurden die Obliterationsgrade in drei Gruppen zusammengefasst (0; 1-2; 3-4). Die Zusammenfassung der Stadien nach MEINDL/ LOVEJOY (1985) erfolgte in die Gruppen 0, 1-2 und 3 und die Zusammenfassung der Punktsummen für das Schädeldachsystem in 0-7, 8-14 und 15-21 Schädeldachsystem * (Zusammenfassung der Mittelwerte der zusammengehörigen linken und rechten sowie der einzeln vorliegenden linken und einzelnen rechten Nahtabschnitte) Vorderer Nahtkomplex (1) Sutura lambdoidea (Mitte) (6) Sutura coronalis (Mitte) (2) Lambda (7) Pterion (3) Obelion (8) Sutura sphenofrontalis (4) Sutura sagittalis (anterior) (9) Sutura sphenotemporalis inferior (5) Bregma (6) Sutura coronalis (Mitte) (7) Pterion Tab. 4 Vergleich der anthropologischen Altersgruppen mit den jeweiligen Nähten und Nahtabschnitten – Signifikanzwerte (p) der Kreuztabellen. sowie für den vorderen Nahtkomplex in die Gruppen 0-5, 6-10 und 11-15. (10) Sutura sphenotemporalis superior Die Ergebnisse der Signifikanztests für das Verfahren nach NEMESKÉRI et al. (1966) wurden in Tabelle 4 zusammengefasst. Dabei zeigen sich erwartungsgemäß an der Tabula interna bessere Ergebnisse als an der Tabula externa. An der Ta- Tab. 3 Nahtabschnitte nach MEINDL/LOVEJOY (1985). aktuell 28 Altersschätzung von Skelettmaterial – Untersuchungen zur Obliteration der Nähte des Schädels Tabula interna S. coronalis (C3) (li/re+li+re) 0 J/A SA/M SM/S S 1-2 3-4 S 4 0 2 6 (9,52) (0,00) (4,76) (14,29) 3 12 4 (9,52) 3 (7,14) 11 (7,14) (28,57) 3 11 (7,14) (26,19) 6 25 19 (45,24) 17 (40,48) 42 Tab. 9 Vergleich der Altersgruppen mit den Obliterationsgraden der Sutura saggitalis 1. S 0 1-2 3-4 S 6 0 2 8 (4,44) (17,78) 12 19 (13,33) (0,00) 5 2 (11,11) (4,44) (26,67) 4 1 13 (8,89) (2,22) (28,89) 15 3 27 (42,22) 18 (40,00) S 0 1-2 3-4 5 0 2 (11,90) (0,00) (4,76) 6 2 10 (14,29) (4,76) (23,81) 4 2 11 (9,52) (4,76) (26,19) 15 4 0 1 (13,33) (0,00) (3,33) 3 1 9 S 5 (16,67) 13 (10,00) (3,33) (30,00) (43,33) 3 0 9 12 (10,00) (0,00) (30,00) (40,00) 10 1 19 30 (33,33) (3,33) (63,33) (100,00) SM/S 0 1-2 3-4 S 6 1 1 8 (2,44) (19,51) 9 17 (14,63) (2,44) 5 3 (12,20) (7,32) (21,95) (41,46) 3 (7,32) 14 3 10 16 (7,32) (24,39) (39,02) 7 20 41 (34,15) (17,07) (48,78) (100,00) (...)-Angaben in Prozent Tab. 10 Vergleich der Altersgruppen mit den Obliterationsgraden der Sutura saggitalis 2. 4 23 S 7 Cranium - Obliterationsgrade Tabula interna S. sagittalis (S2) J/A (16,67) Altersgruppen Altersgruppen SM/S SA/M (33,33) (6,67) (60,00) (100,00) Tabula interna S. coronalis (C)1 (li/re+li+re) SA/M J/A S Cranium - Obliterationsgrade J/A 3-4 Tabula interna S. sagittalis (S1) 45 (...)-Angaben in Prozent Tab. 7 Vergleich der Altersgruppen mit den Obliterationsgraden der Sutura coronalis 1 (Mittelwert der paarigen linken und rechten sowie der einzelnen linken und einzelnen rechten). SM/S 1-2 Cranium - Obliterationsgrade Altersgruppen Altersgruppen SM/S SA/M 0 (...)-Angaben in Prozent Tabula interna S. coronalis (C2) (li/re+li+re) SA/M J/A (26,19) (14,29) (59,52) (100,00) Cranium - Obliterationsgrade J/A Tabula interna S. coronalis (C1+C2) S (...)-Angaben in Prozent Tab. 6 Vergleich der Altersgruppen mit den Obliterationsgraden der Sutura coronalis 2 (Mittelwert der paarigen linken und rechten sowie der einzelnen linken und einzelnen rechten). Cranium - Obliterationsgrade Altersgruppen Tab. 8 Vergleich der Altersgruppen mit den Obliterationsgraden der Sutura coronalis 1 und 2 (Mittelwert der paarigen linken und rechten sowie der einzelnen linken und einzelnen rechten). Cranium - Obliterationsgrade Altersgruppen Tab. 5 Vergleich der Altersgruppen mit den Obliterationsgraden der Sutura coronalis 3 (Mittelwert der paarigen linken und rechten sowie der einzelnen linken und einzelnen rechten). 18 (42,86) 17 (40,48) 42 SA/M SM/S S (35,71) (9,52) (54,76) (100,00) (...)-Angaben in Prozent 0 1-2 7 0 (16,67) (0,00) 5 3 3-4 S 1 8 (2,38) (19,05) 9 17 (11,90) (7,14) (21,43) (40,48) 3 2 12 17 (7,14) (4,76) (28,57) (40,48) 15 5 22 42 (35,71) (11,90) (52,38) (100,00) (...)-Angaben in Prozent bula interna der Sutura coronalis zeigt sich kein homogenes Verteilungsmuster (vgl. Tab. 5-8). Dies betrifft zum einen etwaige Seitenunterschiede und zum andern die zusammenfassenden Berechnungen mit bzw. ohne den Nahtabschnitt C3, der meist in Bezug auf die Verstreichung der Nähte eine höhere biologische Variabilität zeigt. 29 aktuell Ronny Bindl, Rainer Nitsche und Horst Bruchhaus Tabula interna S. sagittalis (S3) Altersgruppen J/A SA/M SM/S S 0 1-2 3-4 S 7 1 1 9 (16,67) (2,38) (2,38) 3 5 8 (21,43) Tab. 11 Vergleich der Altersgruppen mit den Obliterationsgraden der Sutura saggitalis 3. Cranium - Obliterationsgrade Tabula interna S. lambdoidea (L1) (li/re+li+re) J/A Altersgruppen Cranium - Obliterationsgrade 16 (7,14) (11,90) (19,05) (38,10) 2 3 12 17 (4,76) (7,14) (28,57) (40,48) 12 9 21 42 Tabula interna S. sagittalis (S4) S 0 1-2 3-4 S 7 0 1 8 (16,28) (0,00) (2,33) 2 11 Tab. 12 Vergleich der Altersgruppen mit den Obliterationsgraden der Sutura saggitalis 4. J/A (18,60) 18 2 12 17 (6,98) (4,65) (27,91) (39,53) 15 4 24 43 S 1-2 3-4 S 6 0 1 7 (16,67) (0,00) 0 (2,78) (19,44) 9 15 (16,67) (0,00) (25,00) 1 11 (5,56) (2,78) (30,56) 14 1 21 Tab. 13 Vergleich der Altersgruppen mit den Obliterationsgraden der Sutura saggitalis. 5 5 7 17 (13,51) (13,51) (18,92) (45,95) 15 6 16 37 (40,54) (16,22) (43,24) (100,00) 0 1-2 3-4 S 6 0 1 7 (15,79) (0,00) (2,63) 2 3 8 (18,42) 13 (5,26) (7,89) (21,05) (34,21) 1 11 18 (15,79) (2,63) (28,95) (47,37) 14 4 20 38 Tab. 15 Vergleich der Altersgruppen mit den Obliterationsgraden der Sutura lambdoidea 2 (Mittelwert der paarigen linken und rechten sowie der einzelnen linken und einzelnen rechten). (36,84) (10,53) (52,63) (100,00) Cranium - Obliterationsgrade Tabula interna S. lambdoidea (L3) (li/re+li+re) J/A Altersgruppen Altersgruppen SM/S 0 2 (18,92) (...)-Angaben in Prozent Tabula interna S. sagittalis 6 (0,00) (16,22) (2,70) 6 SM/S (34,88) (9,30) (55,81) (100,00) Cranium - Obliterationsgrade SA/M SA/M S (...)-Angaben in Prozent J/A 7 Tabula interna S. lambdoidea (L2) (li/re+li+re) (11,63) (4,65) (25,58) (41,86) 3 S 0 Cranium - Obliterationsgrade Altersgruppen Altersgruppen SM/S 3-4 1 (...)-Angaben in Prozent Cranium - Obliterationsgrade SA/M SM/S (28,57) (21,43) (50,00) (100,00) 5 1-2 6 4 0 9 13 SA/M (10,81) (0,00) (24,32) (35,14) S (...)-Angaben in Prozent J/A 0 Tab. 14 Vergleich der Altersgruppen mit den Obliterationsgraden der Sutura lambdoidea 1 (Mittelwert der paarigen linken und rechten sowie der einzelnen linken und einzelnen rechten). (41,67) 14 (38,89) 36 SA/M SM/S S (38,89) (2,78) (58,33) (100,00) 0 1-2 5 0 (15,63) (0,00) 4 1 3-4 1 6 (3,13) (18,75) 6 (12,50) (3,13) (18,75) 6 1 8 (18,75) (3,13) (25,00) 15 2 S 15 11 (34,38) 15 (46,88) 32 Tab. 16 Vergleich der Altersgruppen mit den Obliterationsgraden der Sutura lambdoidea 3 (Mittelwert der paarigen linken und rechten sowie der einzelnen linken und einzelnen rechten). (46,88) (6,25) (46,88) (100,00) (...)-Angaben in Prozent (...)-Angaben in Prozent An allen Abschnitten der Sutura sagittalis (T. interna) wurden deutlich signifikante Zusammenhänge zun den Altersgruppen des Postcraniums ermittelt (vgl. Tab. 9-13). Bei der Betrachtung der Sutura lambdoidea (T. interna) wurden an den Nahtabschnitten L1 und L2 signifikante Zusammenhänge ermittelt. Der Nahtabschnitt L3 zeigt keine Signifikanz. Die aktuell 30 Altersschätzung von Skelettmaterial – Untersuchungen zur Obliteration der Nähte des Schädels Tab. 20 Vergleich der Altersgruppen mit den Punktsummen des Schädeldachsystems nach MEINDL/LOVEJOY (1985). Cranium - Obliterationsgrade SA/M SM/S S 1-2 3-4 S 2 0 0 2 (0,00) (11,76) 5 8 (11,76) (0,00) 1 2 (5,88) (11,76) (29,41) (47,06) 2 1 4 7 (11,76) (5,88) (23,53) (41,18) 5 3 9 Altersgruppen SM/S S SM/S (29,41) (17,65) (52,94) (100,00) 0-7 8-14 15-21 S 4 0 0 4 (44,44) (0,00) (0,00) 1 1 (11,11) (11,11) (0,00) 2 0 (44,44) 0 2 (22,22) 1 3 (22,22) (0,00) (11,11) (33,33) 7 1 1 9 (77,78) (11,11) (11,11) (100,00) (...)-Angaben in Prozent Tab. 21 Vergleich der Altersgruppen mit den Punktsummen des Vorderen Nahtkomplex nach MEINDL/ LOVEJOY (1985). Tabula interna S. lambdoidea (L1+L2) SA/M SA/M S Cranium - Obliterationsgrade J/A J/A 17 (...)-Angaben in Prozent Tab. 18 Vergleich der Altersgruppen mit den Obliterationsgraden der Sutura lambdoidea 1 und 2 (Mittelwert der paarigen linken und rechten sowie der einzelnen linken und einzelnen rechten). Schädeldachsystem 0 1-2 3-4 S 4 0 0 4 (0,00) (16,67) (16,67) (0,00) 1 1 7 9 (4,17) (4,17) (29,17) (37,50) 2 2 7 11 (8,33) (8,33) (29,17) (45,83) 7 3 14 24 Cranium - Punktsummen Vorderer Nahtkomplex J/A Altersgruppen J/A 0 Cranium - Punktsummen Altersgruppen Tabula interna S. lambdoidea Altersgruppen Tab. 17 Vergleich der Altersgruppen mit den Obliterationsgraden der Sutura lambdoidea (Mittelwert der paarigen linken und rechten sowie der einzelnen linken und einzelnen rechten). SA/M 6-10 11-15 S 4 0 0 4 (33,33) (0,00) (0,00) 2 1 0 (16,67) (8,33) (0,00) 4 SM/S S (29,17) (12,50) (58,33) (100,00) 0-5 0 1 (33,33) (0,00) (8,33) 10 1 1 (33,33) 3 (25,00) 5 (41,67) 12 (83,33) (8,33) (8,33) (100,00) (...)-Angaben in Prozent (...)-Angaben in Prozent Hand des Obliterationsgrades der Tabula externa liefert ungenauere Ergebnisse als die der Tabula interna und sollte daher nur mit der nötigen Vorsicht zur Altersschätzung eingesetzt werden. Besonders gut geeignet erscheint die Sutura sagittalis an der Tabula interna, da sie relativ unanfällig für mechanische Einflüsse sein soll (HERSKOVITZ et al. 1997). Die Sutura lambdoidea besitzt besonders im Abschnitt L3 eine sehr komplexe Struktur und weist des Weiteren oft Nahtknochen auf, welche den Obliterationsverlauf zusätzlich beeinflussen. Daher sollte auch die S. lambdoidea nur mit großer Vorsicht für die Altersschätzung verwendet werden. Das Verfahren von MEINDL/LOVEJOY (1985) – Verwendung von vier Stadien, Beurteilung der Obliteration an 16 ein cm großen Nahtabschnitten, Berechnung von Punktsummen – konnte am vorliegenden Material zu keiner Verbesserung der Ergebnisse beitragen. Die Vergleiche an den Abschnitten der Nähte des Schädeldaches zeigen zusammenfassenden Berechnungen der S. lambdoidea unter Auslassung der Abschnitte L3 zeigen eine deutliche Verbesserung der Ergebnisse (vgl. Tab. 14-18). Die Berechnung der Obliterationsgrade unter Auslassung der Abschnitte C3 und L3 führten zu einer deutlich besseren Übereinstimmung zwischen den Altersgruppen des Postcraniums und dem Nahtverschluss. Die Ergebnisse für die Obliteration nach MEINDL/LOVEJOY (1985) wurden in Tabelle 19 zusammengefasst. Die Verteilungen der Obliterationsgrade im Vergleich zum Skelettalter finden sich für das Schädeldachsystem in Tabelle 20 und für den vorderen Nahtkomplex in Tabelle 21, wobei auf die sehr geringe Individuenanzahl hingewiesen werden muss. Die vorliegenden Ergebnisse zeigen, wie schwierig die Verwendung der Schädelnähte für die Sterbealtersschätzung ist. Die Altersschätzung an 31 aktuell Ronny Bindl, Rainer Nitsche und Horst Bruchhaus Nahtabschnitt links Sutura sphenotemporalis superior rechts Sutura sphenotemporalis inferior Sutura sphenofrontalis Pterion Sutura coronalis (Mitte) - 1,000 1,000 1,000 MW 0,333 1,000 links - 1,000 rechts - - MW - 1,000 links - 0,491 rechts 1,000 1,000 MW 1,000 0,381 links - 1,000 rechts 1,000 1,000 MW 0,333 0,725 links 0,052 0,459 rechts 0,139 0,645 MW 0,026 0,410 Bregma 0,048 0,154 Sutura sagittalis (anterior) 0,044 0,241 Obelion 0,002 0,144 Lambda 0,049 0,397 links 0,060 0,311 rechts 0,021 0,554 MW 0,030 0,321 Schädeldachsystem 1,000 0,194 Vorderer Nahtkomplex - 0,545 Sutura lambdoidea (Mitte) Die vorliegenden Ergebnisse geben Hinweise zu den alterspezifischen Veränderungen am Skelett, es sollte jedoch berücksichtigt werden, dass aufgrund der schwierigen Fundsituation und des z. T. schlechten Erhaltungszustandes die Anzahl der in die jeweiligen Vergleiche eingegangenen Individuen verhältnismäßig gering waren. Tabula interna Tabula externa In weiteren Untersuchungen ist geplant etwaige Geschlechtsunterschiede bei der Obliteration der Nähte, wie sie von einigen Autoren (u. a. HAJNIS/NOVAK 1976, KEMKES-GROTTENTHALER 1996) beschrieben werden, zu untersuchen. Literatur ACSÁDI, G./J. NEMESKÉRI (1970): History of human life span and mortality. Budapest 1970. BIEDERMANN, U. (2003): Zur Altersschätzung anhand von Schädelknochen und Schädelknochenfragmenten aus dem mittelalterlichen Gräberfeld DresdenBriesnitz. Diss. Jena 2003. BROCA, P. (1861): Sur le volume et ala forme du cerveau suivant les individus et suivant les races. Bull Soc Anthropo Paris 2, 1861, 139–207. - (1861): Sur les rapports anatomiques des divers points de la surface du crâne et des diverses parties des hémisphères cérébraux. Bull. Soc. d‘Anthropol. 2, 1861, 340–357. Tab. 19 Vergleich der anthropologischen Altersgruppen mit den jeweiligen Nähten und Nahtabschnitten nach MEINDL/LOVEJOY (1985)- Signifikanzwerte (p) der Kreuztabellen BROOKS, S./J. SUCHEY (1990): Skeletal age determination base on the os pubis: A comparison of the AcsádiNemeskéri and Suchey-Brooks Methods. Human Evolution 5, 1990, 227–238. ähnliche Übereinstimmungen zu den Altersgruppen des Postcraniums wie bei dem Verfahren nach NEMESKÉRI et al. (1966). Von der Verwendung der Abschnitte nach MEINDL/LOVEJOY (1985; vgl. Abb. 2) an der Sutura sphenofrontalis sowie der Sutura sphenotemporalis inferior und superior muss deutlich abgeraten werden. Des Weiteren kann das Verfahrens am (prä-) historischen Skelettmaterial nur selten angewendet werden, da ein nahezu vollständiges Calvarium erforderlich ist um die 16 Nahtabschnitte zu beurteilen. Zusammenfassend lässt sich sagen, das die Obliteration der Nähte insbesondere bei schlecht erhaltenem Skelettmaterial nützliche Hinweise zur Altersschätzung liefern kann. Es ist aber davon abzuraten, die Obliteration als alleinigen Parameter zur Altersschätzung zu verwenden (vgl. NEMESKÉRI et al. 1960). aktuell BRUCHHAUS, H./LUDWIG, M./C. MÜLLER (2003): Zur Altersschätzung am Os pubis. Beiträge zur Archäozoologie und prähistorischen Anthropologie 4, 2003, 133–140. BUCKBERRY, J. L./A. T. CHAMBERLAIN (2002): Age estimation from the auricular surface of the ilium: a revised method. Am. J. Phys. Anthropol. 119(3), 2002, 231–239. FOSTER, J. (2002): Zur Altersschätzung an Knochenfunden am Beispiel des Hüftbeines unter besonderer Berücksichtigung der Ossa pubica. Diss. Jena 2002. GOTTSCHALD, U. (2003): Anthropologische Untersuchungen zur Geschlechtsbestimmung adulter Individuen sowie zur Altersschätzung subadulter Individuen. Diss. Jena 2003. 32 Altersschätzung von Skelettmaterial – Untersuchungen zur Obliteration der Nähte des Schädels GILBERT, B. M./T. W. MCKERN (1973): A method for aging the female os pubis. Am. J. Phys. Anthropol. 38, 1973, 31–38. OWINGS, P. A. (1981): Epiphyseal union of the anterior iliac crest and medial clavicle in a modern multiracial sample of males and females. Fullerton 1981. HAJNIS, K./J. T. NOVAK (1976): Die Verwachsung der Nähte am Schädeldach. Anthropologie 14(1,2), 1976, 89–92. OWINGS-WEBB, P. A./J. M. SUCHEY (1985): Epiphyseal union of the anterior iliac crest and medial clavicle in an modern multiracial sample of American males and females. Am. J. Phys. Anthropol. 68, 1985, 457–466. HERSHKOVITZ, I./LATIMER, B./DUTOUR, O./JELLEMA, L. M./WISH-BARATZ, S./ROTHSCHILD, C./B. M. ROTHSCHILD (1997): Why do we fail in aging the skull from the sagittal suture? Am. J. Phys. Anthropol. 103(3), 1997, 393–399. SUCHEY, J. M. (1979): Problems in the aging of females using the Os pubis. Am. J. Phys. Anthropol. 51(3), 1979, 467–470. SZILVÁSSY, J. (1977): Altersschätzung an den sternalen Gelenkflächen der Schlüsselbeine. Beitr. Gerichtl. Med. 35, 1977, 343–345. HILDEBRAND, T./C. WUTSCHKE (1987): Zur Altersbestimmung von Skelettindividuen anhand des Verknöcherungsgrades der Schädelnähte. Diplomarbeit. Jena 1987. TODD, T. W. (1923): Age changes in the pubic symphysis. VII. The anthropoid strain in human pubic symphysis of the third decade. J. Anat. 57, 1923, 274–294. KEMKES-GROTTENTHALER, A. (1996): Sterbealterbestimmung anhand des ektokranialen Nahtverschlusses: Eine Evaluierung der Meindl-LovejoyMethode. Rechtsmedizin 6(6), 1996, 177–184. VOIGT, S./UERLINGS, H./H. BRUCHHAUS (2006): Möglichkeiten und Grenzen der Altersschätzung an Schädekalotten. Beiträge zur Archäozoologie und Prähistorischen Anthropologie, 5, 2006, 143–147. LISS, M. (2001): Rekonstruktion der Altersstruktur subadulter Individuen anhand von Lang- und Beckenknochen aus dem mittelalterlichen Gräberfeld Dresden-Briesnitz. Diss. Jena 2001. LOVEJOY, C. O./MEINDL, R. S./PRYZBECK, T. R./R. P. MENSFORTH (1985): Chronological metamorphosis of the auricular surface of the ilium. A new method for the determination of adult skeletal age at death. Am. J. Phys. Anthropol. 68, 1985, 15–28. Ronny Bindl M. Sc., Rainer Nitsche, Dr. Horst Bruchhaus Institut für Humangenetik und Anthropologie Friedrich-Schiller Universität Jena Kollegiengasse 10 07740 Jena [email protected] [email protected] MCKERN, S. W./T. D. STEWART (1957): Skeletal age changes in young american males: Analyzed from the standpoint of age identification. Headquarters QM. Res. and Dev. Command, Tech. Rep., EP-45, 1957. NEMESKÉRI, J./HARSÁNYI, L./G. ACSÁDI (1960): Methoden zur Diagnose des Lebensalters von Skelettfunden. Anthrop. Anz. 24, 1960, 70–95. 33 aktuell Das linearbandkeramische Gräberfeld von Schwetzingen Claudia Gerling und Michael Francken Zusammenfassung – Das Gräberfeld von Schwetzingen (Baden-Württemberg) ist mit 203 identifizierbaren Gräbern eine der größten bekannten Nekropolen der frühneolithischen Linearbandkeramik. Es befand sich südlich der Neckarmündung am Rande der Rheinebene. Im Großen und Ganzen weist Schwetzingen die Merkmale bekannter Nekropolen dieser Kultur auf: Die Toten legte man weitgehend in linker Hockerlage in NO-SW-orientierten engen Grabgruben nieder. Ungefähr der Hälfte der Bestatteten wurden Beigaben in Form von Keramikgefäßen, Geräten aus Felsgestein, Silex, Knochen und Geweih sowie Schmuck mitgegeben. Die unterschiedlichen Verzierungsmotive der Keramik legen eine Datierung in die jüngere Linearbandkeramik, in die Zeit um 5200/5000 v. Chr., nahe. Neben den knapp 200 Körpergräbern beinhaltet das Gräberfeld auch eine Reihe von Brandbestattungen und so genannten Leergräbern. Eine vollständige anthropologische Auswertung fand bisher nicht statt, grundlegende Arbeiten wurden aber bereits durchgeführt. Damit werden erste vorläufige Aussagen zur Alter- und Geschlechtszusammensetzung möglich. Die Zahl der Frauen innerhalb der Population übersteigt die der Männer, während eine auffällig große Zahl an Kindern zu verzeichnen ist. Im Übrigen ist das Gräberfeld in seiner Zusammensetzung mit anderen Fundstellen dieser Periode vergleichbar. Schlüsselwörter – Anthropologie, Baden-Württemberg, Demographie, Gräberfeld, Linearbandkeramik, Neolithikum, Rheintal. Abstract – The cemetery of Schwetzingen (Baden-Württemberg) is with its 203 remaining graves one of the largest known necropolises of the early Neolithic Linear Bandceramic culture (LBK). It was located south of the mouth of the Neckar into the Rhine, at the edge of the Rhine valley. On the whole Schwetzingen shows the characteristics of the known cemeteries of this culture. The dead were mostly laid down on left side in a flexed position in NE-SW-orientated, narrow grave pits. Approximately 50 % of the dead were buried with grave gifts such as pottery and tools out of stone, silex, bone and antler and trinket. The findings in the grave, especially the ceramics’ varying ornamentation, suggest a dating around 5200/5000 BC. Beside nearly 200 body burials some cremated remains and some so called empty graves were found at the cemetery. A complete anthropological analysis hasn´t been carried out but fundamental informations exist. Based on these results some preliminary statements concerning the composition of age and gender are possible. Within the population the women exceed the men while a large number of children is recorded. As for the rest the momentary anthropological results of the cemetery are comparable with other sites of this period. Keywords – Anthropology, Baden-Württemberg, cemetery, Demography, Early Neolithic, Linear Bandceramic culture, Rhine valley. Einleitung Bereits im September 1988 war das Gräberfeld von Schwetzingen im Zusammenhang mit Baumaßnahmen entdeckt worden und wurde in den folgenden Monaten ausgegraben. Nach mehreren kurzen vom Ausgräber Dr. R.-H. Behrends verfassten Vorberichten und Artikeln (u.a. BEHRENDS 1989; ders. 1990; ders. 1993) lag der Friedhof „brach“. Fast 15 Jahre später erfolgten nun noch unabgeschlossene Bearbeitungen auf archäologischer und anthropologischer Seite (u.a. GERLING 2006, FRANCKEN 2006), deren Zwischenergebnisse an dieser Stelle vorgestellt werden sollen. Archäologie … Die bandkeramische Nekropole von Schwetzingen aus der Zeit der jüngeren Linearbandkeramik (um 5200-5000 v. Chr.) befand sich am Rande der Rheinebene, südlich der Mündung des Neckars in den Rhein. 203 Gräber bzw. 211 Bestattungen konnten auf unregelmäßig langovalen Nekropole identifiziert werden (Abb. 1). 194 davon können als Körper-, neun als Brandgräber angesprochen werden. Wahrscheinlich erhaltungsbedingt wurden Brandgräber großteils im südlichen Gräberfeldareal gefunden, wo sich die Grabgruben tiefer im Boden erhielten. Des Weiteren wurden 16 grabähnliche Gruben gegraben. Vier dieser Gruben enthielten teilweise Skelettreste und beigabenähnliche Artefakte und fungierten aufgrund dessen als Kenotaphe. Meist langovale oder annähernd eckige, 1 m bis 2 m lange Grabgruben fassten die Körperbestattungen. Ins Grab gelegt wurden die Toten, wie für die Bandkeramik üblich, vornehmlich in linksseitiger Hockerlage (Abb. 2). Indessen lag nur ein Fünftel der Skelette in rechter Hockerposition. Die Ausnahme bildet die gestreckte Lage. Ferner stellen Mehrfach- und Nachbestattungen Sonderfälle dar. Obwohl fast die Hälfte der Toten (48 %) nordost-südwestlich orientiert lag, wirkt die Nekropole in Bezug auf die Skelettausrichtung verhältnismäßig uneinheitlich. Die zweithäufigste Orientierung war die in die Gegenrichtung Südwest-Nordost (21 %). Davon abweichende Ausrichtungen spielen eine untergeordnete Rolle. Die karge Ausstattung der Gräber sticht ins Auge, denn nur die Hälfte der Bestatteten bekam Beigaben ins Grab. Diese „Beigabenarmut“ konn- Archäologische Informationen 30/1, 2007, 43-50 Bulletin de la Société Suisse43d‘Anthropologie 13 (1), 2007 Bulletin der Schweizerischen Gesellschaft für Anthropologie 13 (1), 2007 Claudia Gerling und Michael Francken Abb. 1 Gesamtplan des Gräberfeldes. RPK 25. waren diese auch in die Grubenverfüllung eingestreut. In Bezug auf die anthropologische Auswertung interessante Gefäße sind die nahezu identisch dekorierten Kümpfe aus den Gräbern 21 und 43. Bestand eine Beziehung zwischen den beiden Toten? Einem Kind wurde ein mit organischem Material überzogener, verzierter Kumpf mitgegeben. Welche Funktion hatte die Ummantelung? Welche Rolle spielte der oder die Tote? Neben der Keramik wurde den Toten vor allem Silex in Form von Klingen und Pfeilspitzen mitgegeben. Auffallend ist die große Anzahl an sehr langen und unbearbeiteten Klingen, die typisch für ältere Männerbestattungen zu sein scheinen (Abb. 2). Jüngere Tote erhielten hingegen weitestgehend fragmentierte Klingen. Pfeilspitzen lagen primär im Nordwesten des Friedhofes, dort auch in größeren Ensembles, wohingegen im Südteil zwar Gräber mit Pfeilspitzen ausgestattet waren, diese aber auf eine Anzahl von ein bis zwei Spitzen pro Grab beschränkt waren. Wurde nun der te auch bei den beiden anderen LBK-Friedhöfen auf dem Gebiet des heutigen Baden-Württembergs (Stuttgart-Mühlhausen und Fellbach-Oeffingen) verzeichnet werden (NIESZERY 1995, 104). Keramik ist die Beigabe, die den Toten am häufigsten mitgegeben wurde (Abb. 3). Meist handelt es sich um ein, maximal zwei, Keramikgefäße, die vorwiegend in Kopfnähe abgestellt wurden. Die Verzierungen – beliebteste Dekoration waren beidseitig von Einstichreihen bzw. schräg gesetzten kurzen Strichen gerahmte plastische Leisten – weisen auf eine überwiegende Belegung der Nekropole in der jüngeren Bandkeramik hin. Nur wenige Gefäßverzierungen – Winkelbänder mit vertikalen Einstichreihen in den Zwickeln – datieren in die mittlere LBK und nur ein Gefäß trägt ein flombornartiges Dekor, das an den Beginn der mittleren Linearbandkeramik zu setzen ist. Neben vollständigen Kümpfen, Flaschen und Schalen fanden sich vereinzelte Scherben unterschiedlicher Gefäßeinheiten direkt am Skelett, oftmals aktuell 44 Das linearbandkeramische Gräberfeld von Schwetzingen Abb. 2 Grab 106. RPK 25. Abb. 3 Keramikgefäße der Gräber 160 und 201. CG. 45 aktuell Claudia Gerling und Michael Francken nen sekundär durchlochten, flachen Dechsel und einen doppelschneidigen, durchlochten Dechsel – scheinen u.a. aufgrund ihrer Rarität neben ihrer Primärfunktion als Werkzeug einen hohen Prestigewert besessen zu haben. Sie waren nicht allein auf Männergräber beschränkt, sondern der doppelschneidige Dechsel („Zeremonialaxt“ (BEHRENDS 1990, 47)) lag in einem Kindergrab (Abb. 5 unten). Was war das für ein Kind, dem ein solches Statussymbol in verbranntem Zustand mitgegeben wurde? Generell wurden die Schwetzinger Kinder nicht beigabenlos bestattet, sondern ihre Gräber waren teilweise reich mit Beigaben versehen. Des Weiteren lagen zu einem nicht geringen Teil Tierknochen und Geweihe als Rohmaterialien für die Geräteherstellung vor. Hervorzuheben ist die große Anzahl an dreieckigen Knochenspitzen (21 Exemplare), deren zahlreiches Vorkommen im Gegensatz zum ansonsten raren Auftreten dieser Artefakte steht (Abb. 2). Da sie bei nur einer anthropologisch sicher weiblich bestimmten Toten lagen, können sie in Schwetzingen als männerspezifisch gelten. Form und Gestaltung sowie ihre Lage, die sich auf Kopf- und Fußbereich konzentrierte, machen eine Deutung als Pfeilspitzen wahrscheinlich. Dies wird durch die Verteilung Zugang zum Rohmaterial Silex knapper und/ oder änderte sich die Beigabentradition? Hervorzuheben ist Grab 133 mit zehn Silexspitzen, von denen acht am Oberarm des Bestatteten beieinander lagen (Abb. 4). Die großteils gleiche Ausrichtung spricht für eine Anordnung in einem Köcher aus organischem Material. Der Tote wurde auch mit weiteren Beigaben großzügig ausgestattet und in selten vorkommender gestreckter Rükkenlage bestattet, weshalb eine herausragende Stellung des Mannes innerhalb der Schwetzinger Gemeinschaft anzunehmen ist. Ein Werkzeug aus Felsgestein befand sich in ungefähr jedem zehnten Grab. Unterscheiden lassen sich hohe, flache und Miniaturdechsel sowie in geringer Zahl vorliegende durchlochte Steingeräte (Abb. 5). Dechsel scheinen eine typische Beigabe für Männer mittleren Alters gewesen zu sein, denn in nur einem Fall lag ein Exemplar bei einer anthropologisch sicher weiblich bestimmten Bestatteten. Da nur wenige Artefakte Gebrauchsspuren aufweisen, kann bei den meisten Geräten von einer Herstellung speziell als Grabbeigabe ausgegangen werden. Die vier durchlochten Steingeräte, die in Schwetzingen geborgen werden konnten, – es handelt sich dabei um zwei Scheibenkeulen (eine davon ist verschollen), ei- Abb. 4 Grab 133. RPK 25. aktuell 46 Das linearbandkeramische Gräberfeld von Schwetzingen Abb. 5 Geräte aus Felsgestein, Gräber 220 und 21. CG. auf unterschiedliche Friedhofsareale unterstrichen. Während im Nordteil Silexspitzen dominieren, sind im Südosten Knochenspitzen gehäuft und teilweise in mehrfachen Ensembles vertreten. Dies lässt sich zeitlich und/oder mit unterschiedlichen Ausstattungstraditionen erklären. Für weitere anthropologische Analysen interessant erscheinen acht lange, undurchlochte Knochenspitzen, die einer älteren Frau in die Hand gelegt wurden. Sie weisen Ähnlichkeiten zu Flachskämmen aus Feuchtbodengrabungen auf. Ebenfalls bei den Knochen- und Geweihartefakten ist ein Kindergrab hervorzuheben. In Grab 129 lag eine Geweihhacke in einem sonst beigabenlosen Grab eines Kindes. Erfüllte das Artefakt allein repräsentative Zwecke. In diesem Fall stellt sich die Frage nach der Stellung der Kindes und seiner Familienzugehörigkeit. Die prestigeträchtigen Spondylusmuscheln schmückten die Toten in Form von Röhrenperlen, durchlochten Scheiben und Armreifen. Als gering zu erachten ist der Anteil von nur 2 % spondylusführenden Gräbern (WILLMS 1985, 339 f.). Dieser Schmuck scheint kein bestimmtes Geschlecht oder Altersgruppe sondern eher einzelne Individuen aus anderen Gründen hervorzuheben. Eventuell können anthropologische Untersuchungen diese Gründe erkennbar werden lassen. Was nun zeichnet Schwetzingen, eines der größten bekannten LBK-Gräberfelder, in archäologischer Hinsicht aus? Im Großen und Ganzen ist es „normal unnormal“. Auffällig ist die ärmliche Beigabensitte, die auch für die am Neckar gelegenen Nekropolen belegt ist (NIESZERY 1995, 104). In Hinsicht auf die Keramik lässt sich die Nekropole regional jedoch an die Pfalz angliedern. Nur wenige „reich“ ausgestattete Gräber erscheinen einzeln verteilt auf dem Friedhof. Das Gräberfeld von Schwetzingen spiegelt folglich eine Gesellschaft wieder, die nicht als egalitär zu bezeichnen ist sondern aus der sich einzelne Individuen hervorheben. … und Anthropologie Obwohl das Gräberfeld in den Jahren 1988/89 ausgegraben wurde und eine ganze Reihe spezieller Fragestellungen am anthropologischen Material überprüft wurden, ist die Auswertung der Knochen zum momentanen Zeitpunkt noch nicht abgeschlossen. Dementsprechend handelt es sich 47 aktuell Claudia Gerling und Michael Francken bei den vorgestellten Ergebnissen um eine Zwischenbilanz, die in nächster Zeit weiter ergänzt werden soll. Der Erhaltungszustand des Skelettmaterials aus dem Gräberfeld von Schwetzingen kann als überwiegend gut charakterisiert werden. Insgesamt sind 194 Körpergräber geborgen worden, außerdem wurden 16 Brandbestattungen identifiziert (TRAUTMANN & WAHL 2005), von denen sieben innerhalb von Körpergräbern gefunden wurden. Zusätzlich wurden 15 Grabbefunde als Leergräber klassifiziert, die in jüngster Vergangenheit im Rahmen einer Magisterarbeit von M. Wenzel (WENZEL 2007) untersucht wurden. Zu den bereits ausgeführten Basisarbeiten gehört die Präparation des Knochenmaterials, außerdem eine erste Alters- und Geschlechtsbestimmung durch J. Wahl. Die osteometrische Erfassung sowie eine pathologische Sichtung des Materials wurde ebenfalls durch J. Wahl durchgeführt, eine Interpretation der Daten steht allerdings noch aus. Unabhängig von der eigentlichen Analyse wurden die Skelette zur Untersuchung von speziellen Fragestellungen herangezogen. Darunter fallen die Auswertung von Sonderbestattungen in Form von Brand- und möglichen Leergräbern, die Analyse des Strontiumgehalts (BENTLEY ET AL. 2002) und ein Vergleich von Al- Basistabelle für Geschlechtsverteilung (n=211) ♂♂ ♀♀ Unbestimmt 5,3 11,4 61,1 Erwachsene 50,5 62,4 19,9 Gesamt 55,8 73,8 81,0 Kinder & Jugendliche Tab. 1 Vorläufige Geschlechterverteilung aufgeteilt nach NichtErwachsenen und Erwachsenen (n = 211). tersbestimmungsmethoden bei erwachsenen Individuen unter besonderer Berücksichtigung der Zahnzementannulation (FRANCKEN 2006). Dank der vorläufigen Geschlechtsbestimmung lassen sich erste Aussagen zur Zusammensetzung innerhalb des Gräberfeldes machen. Insgesamt wurde bei 211 Individuen eine Geschlechtsdiagnose versucht, eine Bestimmung war bei 130 Individuen möglich, während 81 Skelette nicht genauer klassifiziert werden konnten (Tab. 1). Innerhalb der Gruppe der geschlechtsbestimmten Individuen vereinen sich sowohl sichere als auch noch fragliche Skelette, bei denen eine definitive Entscheidung noch aussteht. Zusätzlich zu den Erwachsenen wurde, soweit möglich, das Geschlecht der Kinder und Jugendlichen bestimmt. Die Ergebnisse sind ebenfalls in der Auflistung berücksichtigt worden und haben einen großen 30 Anzahl der Gestorbenen 25 20 15 10 5 0 0-4 5-9 10 - 14 15 - 19 20 - 24 25 - 29 30 - 34 35 - 39 40 - 44 45 - 49 50 - 54 55 - 59 Altersgruppen in Jahren Dx(ƃƃ) Dx(ƂƂ) Dx(Unbestimmt) Abb. 6 Vorläufige Altersverteilung (n = 211). aktuell 48 60 - x Das linearbandkeramische Gräberfeld von Schwetzingen Einfluss auf das Segment der unbestimmten Individuen. Konnte das Alter bei der Bestimmung nicht in die vorgegebenen 5-Jahres-Kategorien eingegrenzt werden, ist das Individuum gleichmäßig auf alle betroffenen Altersgruppen verteilt worden. Individuenzahlen mit Dezimalstellen (wie in Tab. 1) werden so möglich. 61 (75,3%) der insgesamt 81 Skelette dieser Gruppe gehören zu Kindern und Jugendlichen. Dies entspricht einem Anteil von 79,2% (bezogen auf 77 Fälle) aller Individuen dieser Altersgruppen, während lediglich bei den übrigen 20,8% eine Bestimmung möglich war (6,5% (n≈5): ♂ / 14,3% (n≈11): ♀). Dagegen stehen die Ergebnisse der Geschlechtsdiagnose bei den Erwachsenen im umgekehrten Verhältnis, hier konnten 85% der erwachsenen Individuen bestimmt werden (38,4% (n≈51):♂/ 46,6% (n≈62):♀). Insgesamt liegt der Anteil der Frauen mit insgesamt 74 über dem der Männer mit 56 Personen. Berücksichtigt man den großen Anteil von Kinder und Jugendlichen zeigt sich bei der vorläufigen Altersverteilung ein wenig überraschendes Bild (Abb. 6). Die Gruppen der 0-4jährigen, bzw. der 5-9jährigen stellen bei der Anzahl der Gestorbenen (Dx) die meisten Individuen, während ihre Zahl in den folgenden subadulten Gruppen stark abnimmt. Der im Vergleich zur ersten Gruppe höhere Wert bei den 5-9jährigen könnte möglicherweise durch eine zunehmende Mobilität der Kinder und einer damit verbundenen Einbindung in den Arbeitsalltag erklärt werden, bei denen ein mögliches Verletzungsrisiko bestand. Eine Bestätigung durch Analyse pathologischer Veränderungen und damit möglicher Hinweise auf Mangelsituationen, Krankheiten oder Verletzungen steht allerdings noch aus. Aufgrund der vorliegenden Daten errechnet sich eine Kindersterblichkeit in den ersten zehn Jahren von 24%. Insgesamt starben 37% der Individuen von Schwetzingen bevor sie älter als 20 Jahre waren. Mit dem Eintritt ins Erwachsenenalter (20+ Jahre) steigt die Anzahl der gestorbenen Individuen wieder annähernd auf den Stand der ersten zwei Altersgruppen und erhält diesen über die nächsten 20 Jahre fast konstant. Dabei überwiegt die Zahl an weiblichen Individuen die der männlichen. Bei den frühmaturen Gruppen sinkt die Zahl der Gestorbenen im Vergleich zu den adulten Gruppen um gut die Hälfte, während sich bei der Geschlechtszusammensetzung eine Umkehrung bemerkbar macht. Die Sterbewahrscheinlichkeit der Frauen sinkt unter die der Männer, was durch eine Verringerung der Anzahl Gestorbener und des relativen Anteils an Gestorbenen gut dokumentiert wird. Möglicherweise spielt dabei das Ende der Gebärfähigkeit bei Frauen eine Rolle. Ähnliche Beobachtungen sind auch bei anderen bandkeramischen Gräberfeldern gemacht worden (NIESZERY 1995, 94-97). In der spätmaturen Phase steigt die Anzahl der Gestorbenen nochmals, bevor sie im senilen Stadium auf das Minimum innerhalb dieser Altersverteilung absinkt. Auch in diesem Fall folgt die Verteilung dem Trend, dass Frauen durchschnittlich ein höheres Alter erreichen als Männer. Nur drei Prozent (mathematisch 1,7 Personen) der Männer aus dem Gräberfeld von Schwetzingen erreichten ein Alter jenseits von 60 Jahren, während bei den Frauen 4,6% (mathematisch 3,4 Personen) ein seniles Alter erreichten. Aufgrund der momentan vorliegenden Altersverteilung errechnet sich für die Schwetzinger Gesamtpopulation eine durchschnittliche Lebenserwartung von 27,5 Jahren, die aufgetrennt nach Männer und Frauen eine Lebenserwartung von 36,8 bzw. 33,7 Jahren ergibt. Die starken Schwankungen der Geschlechter im Vergleich zur durchschnittlichen Lebenserwartung erklären sich durch die große Zahl der unbestimmten Subadulten. Da sie in der aufgetrennten Sterbetafel keinen Einfluss haben, steigt die Lebenserwartung der Geschlechter beträchtlich im Vergleich zum Gesamtbild. Bei der Interpretation der hier aufgeführten Ergebnisse muss stets bedacht werden, dass es sich nur um eine Zwischenbilanz handelt. Die ausführliche Bearbeitung und Auswertung des Materials aus Schwetzingen steht noch aus und soll innerhalb der nächsten Jahre erfolgen. Ein abschließendes Fazit wäre dementsprechend an dieser Stelle verfrüht, doch fällt als Besonderheit des Gräberfeldes von Schwetzingen schon jetzt sein hoher Anteil an Subadulten in Verbindung mit einer großen Individuenanzahl auf. Dr. B. Rabold und Dr. G. Wieland, RP Karlsruhe, Dr. R.-H. Behrends und Dr. J. Wahl, RP Stuttgart, danke ich für Ihre Hilfestellung. Literatur BEHRENDS, R.-H. (1989): Ein Gräberfeld der Linienbandkeramik in Schwetzingen, Rhein-NeckarKreis, in: Arch. Deutschland 3/1989, 1989, 39. – (1990): Ein Gräberfeld der Bandkeramik von Schwetzingen, Rhein-Neckar-Kreis, in: Arch. Ausgr. Baden-Württemberg 1989, 1990, 45-48. – (1993): Gräber der ersten Bauern, in: Arch. Nachr. Baden 50, 1993, 38-39. 49 aktuell Claudia Gerling und Michael Francken BENTLEY, A.R. (2001): Human migration in Early Neolithic Europe: Strontium and lead isotope analysis of archaeological skeletons. Ph. D. diss. University of Wisconsin, Madison, Wis., 2001. BENTLEY, A.R., PRICE, T.D., LÜNING, J., GRONENBORN, D., WAHL, J., & P. FULLAGER (2002): Prehistoric migration in Europe: strontium isotope analysis of early Neolithic skeletons. Current Anthropology 43, 2002, 799–804. FRANCKEN, M. (2006): Zahn um Zahn – Die Zahnzementannulation als Methode zur Altersdiagnose. (Unpubl. Magisterarbeit Tübingen 2006). GERLING, C. (2006): Das linienbandkeramische Gräberfeld von Schwetzingen, Rhein-Neckar-Kreis. (Unpubl. Magisterarbeit Würzburg 2006). NIESZERY, N. (1995): Linearbandkeramische Gräberfelder in Bayern. Internationale Archäologie 16, 1995. TRAUTMANN, I., J. WAHL (2005): Leichenbrände aus linearbandkeramischen Gräberfeldern Südwestdeutschlands – Zum Bestattungsbrauch in Schwetzingen und Fellbach-Oeffingen. Fundberichte Baden-Württemberg 28, 2005, 1–13. WENZEL, M. (2007): Leergräber auf linearbandkeramischen Friedhöfen. Kenotaphe, geräumte Primärbestattungen oder Gruben mit anderer Funktion? (Unpubl. Magisterarbeit Tübingen 2007). WILLMS, C. (1985): Neolithischer Spondylusschmuck. Hundert Jahre Forschung. Germania 63, 1985, 331–343. Claudia Gerling M.A. Universität Würzburg Institut für Vor- und Frühgeschichte Siligmüllerstraße 9, 97072 Würzburg [email protected] Michael Francken M.A. Universität Tübingen Arbeitsbereich Paläoanthropologie Rümelinstr. 23, 72074 Tübingen [email protected] aktuell 50 Leichenbrand erzählt vom Umgang mit den Toten – Die interdisziplinäre Rekonstruktion ritueller Handlungen am Beispiel eines Urnengräberfelds der Lausitzer Kultur Birgit Großkopf und Alexander Gramsch Zusammenfassung – Knochen können auch dann noch „sprechen“, wenn sie verbrannt sind. Leichenbrand ist eines der wichtigsten Quellenmaterialien für die historisch-archäologische Forschung und kann, wie dieser Beitrag darlegt, nicht nur biologische Daten liefern, sondern auch Informationen zu kulturhistorischen Fragestellungen. Die anthropologischen Grundlagen der Leichenbrandbearbeitung werden kurz dargestellt, um zum einen deutlich zu machen, welches Potenzial im anthropologisch-archäologischen Quellenmaterial Leichenbrand über die biologischen Grunddaten hinaus verborgen ist. Zum anderen wird erkennbar, dass bereits die Erhebung der Daten von der Fragestellung abhängt und deshalb Leichenbrand nur dann „sprechen“ kann, wenn er in interdisziplinärer Zusammenarbeit zwischen Anthropologie und Archäologie analysiert wird. Am Beispiel des bronzezeitlichen Urnengräberfeldes Cottbus Alvensleben-Kaserne (Brandenburg) wird die interdisziplinäre methodische Herangehensweise beschrieben und die dadurch ermöglichte Rekonstruktion von Funeralpraktiken diskutiert. Auf der Basis dieser Rekonstruktion der verschiedenen rituellen Handlungen wird interpretiert, wie soziale Identitäten im Bestattungsritual dargestellt und transformiert werden. Schlüsselwörter – Leichenbrand, Bestattungsritual, interdisziplinäre Datenerhebung, kommunikative Handlungen, soziale Identität Abstract – Cremation remains talk about the treatment of the dead – The interdisciplinary reconstruction of ritual action: An example from an urnfield of the Lusatian Culture Bones “speak” – even when they are burned. Cremation remains are one of the major source materials for archaeological research; moreover, as this paper explains, they do not only yield biological data but also provide us with information for particular historical questions. An overview of the essentials of the anthropological examination of cremation remains displays that there is considerable potential inherent in this source material beyond biological data. Furthermore, it becomes clear that already the collection of data relies upon the research questions that are asked; thus, cremation remains only do “speak” if they are analysed in interdisciplinary cooperation between anthropology and archaeology. Using the Bronze Age urnfield from Cottbus Alvensleben-Kaserne (Brandenburg, Germany) as a starting point the interdisciplinary approach is explicated, which allows the detailed reconstruction of funeral practices. This reconstruction of the sequence of ritual actions finally serves as the basis for interpreting how social identities are presented and transformed in burial ritual. Keywords – cremations, burial ritual, interdisciplinary data collection, communicative actions, social identity Auf ihrer gemeinsam veranstalteten Tagung forderten DGUF und SGA dazu auf „Knochen sprechen“ zu lassen. Diese Metapher, die wir im Titel unseres Beitrages aufgreifen, erinnert an einen Ansatz der postprozessualen Archäologie der 1980er und frühen 1990er Jahre. Man versuchte, materielle Kultur als „Text“ zu verstehen und als solchen zu lesen (z.B. HODDER 1989, 1991). Insbesondere von der symbolischen Bedeutung dieses Textes sollte materielle Kultur erzählen. Mittlerweile gibt man jedoch zu bedenken, dass Text ein anderes Zeichensystem als das der materiellen Kultur ist, und dass dieses „Text“-Paradigma materielle Kultur zu wenig im Kontext der vergangenen Alltagswelt und Praxis und zu sehr in der symbolischen Ebene verankert (HODDER/ Hutson 2003, 204; HODDER 2005). Darüber hinaus impliziert es aber, dass wir diesen „Text“ oder das „Sprechen“ der materiellen Kultur oder der Knochen unmittelbar lesen und verstehen könnten. Das Wort vom „Erzählen“ bzw. „Sprechen lassen“ verstehen wir in unserem Beitrag als sprachliches Bild, nicht als Paradigma. Weder Knochen noch andere Elemente der materiellen Kultur erzählen von sich aus oder können in dem Sinne zum Sprechen gebracht werden, dass sie „ihre Sicht der Dinge“ übermitteln, also sozusagen emisch aus ihrer ehemaligen Lebenswelt berichten. Archäologen sind nicht die „Detektive der Vergangenheit“, die aus materiellen Indizien das Geschehene objektiv rekonstruieren (s. HOLTORF 2003). Die „Erzählung der Knochen“ entsteht vielmehr aus den Fragen, die wir an sie stellen, und den Daten, die wir erheben; dies werden wir im Folgenden verdeutlichen. Diese „Erzählung“ ist aber dennoch nicht völlig willkürlich, sie wird nicht allein von der Gegenwart, sondern durch den Kontext bestimmt, d.h. durch das Zusammenwirken der Vielzahl von Fragen und Daten, in dem sich Knochen und andere Objekte befinden. Das Beachten dieses archäologischen Kontextes ermöglicht, „dass wir systematisch Kontrollen einbauen (oder einbauen sollten), die darauf zielen eine falsche oder unangemessene ‚Lesung‘ der Objekte zu eliminieren“ (VEIT 2003, 24). Die Metapher von den sprechenden Knochen impliziert noch etwas anderes: Verstehen wir Knochen als historische Quellen? Können Archäologische Informationen 30/1, 2007, 71-80 Bulletin de la Société Suisse71d‘Anthropologie 13 (1), 2007 Bulletin der Schweizerischen Gesellschaft für Anthropologie 13 (1), 2007 Birgit Großkopf und Alexander Gramsch sie selbst dann etwas über rituelle Handlungen, über Bestattungspraktiken „erzählen“, wenn sie verbrannt sind? Als historische Quellen werden üblicherweise Schriftzeugnisse verstanden. Die Quellen der archäologischen Fächer dagegen sind Geräte und Schmuck, Gräber und Siedlungen usw. – das komplette Spektrum materieller Kultur. Auch diese ermöglichen historische Erkenntnisse. Die unmittelbaren Überreste menschlicher Individuen jedoch wurden bislang primär als biologische Quelle gesehen und weniger als Teil der materiellen Kultur und damit als gleichwertige historische Quelle (an)erkannt. Knochen können Auskunft geben über Alter, Geschlecht und verschiedene Pathologien des Individuums. Doch sie können noch mehr berichten – selbst wenn sie verbrannt wurden. Welche (prä)historischen Aussagen möglich sind, was also Leichenbrand uns „erzählen“ kann, wollen wir im Folgenden zunächst grundsätzlich und dann anhand eines Beispiels vorstellen. Dabei wird deutlich, dass für eine historische, genauer: sozialgeschichtliche Auswertung dieses Materials eine interdisziplinäre Arbeitsweise notwendig ist. Nicht nur das Auswerten der Daten nach biologischen und historischen Gesichtspunkten, sondern bereits die Datenerhebung darf nicht getrennt stattfinden; sie muss von den historisch-archäologischen und biologisch-anthropologischen Disziplinen gemeinsam formuliert und durchgeführt werden. Erst durch die enge Zusammenarbeit können die Handlungen, die die Bestattungssitten ausmachen, rekonstruiert und schließlich sozialgeschichtlich interpretiert werden. Obwohl grundsätzlich dieselben Kriterien für die Alters- und Geschlechtsdiagnose gültig sind wie bei der Bestimmung von intakten Skeletten, erfordern doch die Spezifika verbrannter Knochen ihre Berücksichtigung. Die Schrumpfung und Fragmentierung sowie die teilweise thermisch bedingte Verformung von Skelettelementen erfordert sehr grundlegende Kenntnisse im Umgang mit dem Material Leichenbrand. Gerade im Hinblick auf die Bestimmung des biologischen Geschlechts zeigt sich nämlich, dass die Anwendung metrischer Methoden sehr kritisch beurteilt werden muss. Bei HERRMANN ET AL. (1990) ist z.B. der Trennwert für das transversale Maß des Dens axis mit 9 mm angegeben (Frauen < 9 mm < Männer). Bei morphologisch robusten Populationen ist jedoch auch bei weiblichen Individuen ein Maß von > 9 mm wiederholt zu beobachten. Eine populationsübergreifende Anwendung publizierter Daten bzw. Trennwerte kann zu ungenauen oder gar falschen Ergebnissen führen (VAN VARK ET AL. 1996). Daher sollten Trennwerte, anhand derer „weiblich“ und „männlich“ unterschieden werden, nur aus der zu untersuchenden Stichprobe selbst ermittelt werden. Zudem kann auch nicht in jedem Fall der gleiche Schrumpfungsfaktor zugrunde gelegt werden (GROSSKOPF 2004). Als die derzeit bewährteste Methode für die Geschlechtsdiagnose von verbrannten Individuen muss daher die morphologische Bestimmung gelten, obwohl sie letztendlich auf einer in mancher Hinsicht subjektiven, da auf dem Erfahrungshorizont des Anthropologen basierenden Zuordnung qualitativer Kriterien basiert (GROSSKOPF 2004). Dabei werden die Unterschiede in Form, Robustizität und Oberflächenstruktur jeweils unter Berücksichtigung der populationsspezifischen Varianz beurteilt. Auch bei der Altersdiagnose ist das Spektrum der beurteilbaren Merkmale gegenüber komplett überlieferten Skeletten materialbedingt eingeschränkt. Oftmals ist die morphologische Diagnose erwachsener Individuen nur auf die Beurteilung des Schädelnahtverschlusses reduziert. Somit kommt der histologischen Altersdiagnose eine erhebliche Bedeutung zu. Für die knochenhistologische Untersuchung werden Dünnschnitte von Knochenfragmenten angefertigt und mikroskopisch untersucht. Die Knochenbinnenstruktur unterliegt altersabhängigen Veränderungen und erlaubt Rückschlüsse auf das Sterbealter des Verstorbenen. Bei konsequenter Beurteilung der einzelnen Komponenten über den gesamten (vorliegenden) Knochenquerschnitt lassen sich die Strukturen meist noch in früh-, mittel- und Anthropologische Untersuchungen des Leichenbrands Leichenbrand wird oftmals sogar in seinem Potential als biologische Quelle unterschätzt, da er gegenüber einem gut erhaltenen Skelett vermeintlich wenig mehr ist als Brösel und Bruch. Die Verbrennung bewirkt jedoch nicht nur Fragmentierung und Schrumpfung, sondern auch eine Modifikation des Knochenapatits, so dass verbrannter Knochen deutlich stabiler ist und somit wesentlich bessere Überlieferungsaussichten als unverbrannter Knochen aufweist. Wichtiger noch: Da die Leichenverbrennung im Alten Europa für ca. 1500 Jahre die vorherrschende Bestattungsform war, stellt Leichenbrand das wesentliche Quellenmaterial für zahlreiche Generationen (prä)historischer Populationen dar. aktuell 72 Leichenbrand erzählt vom Umgang mit den Toten ... spätadult, bzw. -matur differenzieren (GROSSKOPF 2004). Liegen geeignete Zahnwurzeln vor, werden zudem Zahndünnschnitte angefertigt und die Zuwachsringe im Zahnzement gezählt, die sich im circaannualen Rhythmus bilden. Durch Addition des entsprechenden Zahndurchbruchsalters kann das kalendarische Alter eines Individuums auf wenige Jahre genau bestimmt werden (GROSSKOPF 1990). Grundsätzlich gilt für die Alters- und Geschlechtsdiagnose, nicht nur am Leichenbrand, dass das Ergebnis optimiert werden kann, je mehr Daten berücksichtigt werden bzw. je vielfältiger die methodischen Zugänge genutzt werden. Regel nicht eingesammelt werden, da er sich nicht vom Sediment separieren lässt. Zum anderen ist aber auch ein (intentionales) unvollständiges Einsammeln oder Teil-Deponieren möglich. Auch eine beabsichtigte oder zufällige Vermischung mit Überresten eines weiteren Individuums ist möglich. Hier kann anhand der mengenmäßigen Anteile und der repräsentierten Skelettregionen in einer Leichenbrandpartie in der Regel eine Differenzierung zwischen Doppelbestattung und Beimengung erfolgen. Beimengungen können z.B. durch unvollständige Reinigung eines wiederholt genutzten Verbrennungsplatzes auftreten. Auch die Art der Deponierung lässt Rückschlüsse auf Praktiken während der Bestattung zu. Lassen sich z.B. zahlreiche Holzkohlefragmente in einer Urne finden, so ist der Leichenbrand offensichtlich nicht gezielt aus den Scheiterhaufenresten ausgelesen, sondern eher gemeinsam mit diesen in die Urne gefüllt worden. Die Art der Deponierung der Knochen in der Urne weist ebenfalls auf verschiedene Vorgänge rund um das Bestattungsritual hin, z.B. auf die Scheiterhaufengröße, die Lage des Verstorbenen auf dem Scheiterhaufen und die Mindestdauer des Rituals. So ist ein experimentell verbrannter Schweinekadaver in peripheren Bereichen erst nach 24 Stunden so weit abgekühlt, dass die Knochen ausgelesen werden können (LEINEWEBER 2002). Leichenbrand stellt neben dem biologischen Überrest eines Individuums auch ein „Produkt“ menschlichen Handelns dar, welches durch verschiedene Vorgänge rund um die Verbrennung in seinem „Ergebnis“ (d.h. Fragmentgröße, Menge und Färbung) beeinflusst sein kann. Ein Ablöschen des Scheiterhaufens bewirkt z.B. eine deutliche Verkleinerung der Knochenfragmente. Auch eine mechanische Belastung vor dem kompletten Auskühlen, z.B. durch ein Zusammenschieben der Scheiterhaufenüberreste, bedingt eine zusätzliche Fragmentierung, da Leichenbrand erst im ausgekühlten Zustand seine stabile Konsistenz aufweist (HERRMANN 1988). Eine ungenügende Temperaturhöhe und/oder Dauer der Temperatureinwirkung bewirken eine unvollständige Verbrennung, welche durch graue oder sogar schwarze Verfärbungen im Knocheninnern erkennbar ist. Dieses sind Reste primären Kohlenstoffs, der nicht durch die Umwandlung in CO2 aus dem Knochen entwichen ist. So kann ein Auftreten von primären Kohlenstoffverfärbungen in bestimmten Skelettregionen Hinweise auf deren Lage an der Peripherie des Scheiterhaufens geben, in der die Hitze für eine vollständige Verbrennung nicht ausgereicht hat. Bei Brandbestattungen ist davon auszugehen, dass ein Teil des ursprünglichen Skelettmaterials nicht überliefert ist. Auch bei einer Körperbestattung sind Verluste einzelner Knochen z.B. durch eine anthropogene Grabstörung oder die Verlagerung von kleinen Knochen z.B. durch Mäuse möglich; es kann in der europäischen Prähistorie aber davon ausgegangen werden, dass regelhaft der intakte Körper in das Grab gelegt wurde. Bei der Deponierung von Leichenbrand tritt jedoch zum einen ein natürlicher Verlust von Material auf, der sich in der Entstehung von Knochengruß durch die Verbrennung erklärt. Dieser kann in der Das Fallbeispiel: Das Gräberfeld von Cottbus Alvensleben-Kaserne Leichenbrand ist also nur im wörtlichen Sinne eine trockene Materie; er ist eine besondere Form materieller Kultur1 und kann Daten liefern, die für kulturhistorische Fragestellungen genutzt werden können. Diese Daten zu erheben und dann kulturhistorisch zu interpretieren erfordert, wie eingangs betont, interdisziplinäres Arbeiten (GROSSKOPF/GRAMSCH 2004, GRAMSCH/GROSSKOPF 2005). Die Möglichkeiten einer solchen interdisziplinären Leichenbrand-Untersuchung wollen wir im Folgenden am Beispiel des Urnengräberfelds Cottbus Alvensleben-Kaserne (Brandenburg) verdeutlichen. Die Funde und Befunde dieses Gräberfelds haben wir im Rahmen eines Forschungsprojektes am Lehrstuhl für Ur- und Frühgeschichtliche Archäologie der Universität Leipzig gemeinsam bearbeitet unter Leitung von Prof. S. Rieckhoff2 (GROSSKOPF 2004, GRAMSCH 2006, in Vorb.). Bei Umbauarbeiten in der ehemaligen Cottbuser Kaserne in den Jahren 1997-1998 wurden neue Leitungen verlegt, Straßen neu angelegt und 73 aktuell Birgit Großkopf und Alexander Gramsch rend der früheisenzeitlichen Billendorfer Gruppe in Ha C1 angelegt. Im Mittelpunkt sollen nun die endbronzezeitlichen und hallstattzeitlichen Befunde stehen. Neben Keramikpackungen, Gefäßdepots und Scherbenstreuungen (GRAMSCH in Vorb.) liegen insgesamt 74 Gräber mit 105 brandbestatteten Individuen vor. Alle Bestatteten waren verbrannt und ganz überwiegend in Urnen in Grubengräbern oder Rechteck- bzw. Kammergräbern beigesetzt worden, wie sie auch von anderen Niederlausitzer Fundorten bekannt sind (vgl. BÖNISCH 1990, 1995; BUCK 1986, 2007). In den einfachen Grubengräbern wurde meist nur eine Urne beigesetzt, es kommen hier aber auch Doppelbestattungen vor. Die zehn aus Steinsetzungen und vermutlichen Holzeinbauten bestehenden Rechteckbzw. Kammergräber wurden für Doppel- und Mehrfachbestattungen mit bis zu acht Individuen angelegt. Von den 105 überlieferten Individuen des Gräberfelds Cottbus Alvensleben-Kaserne sind 38 Kinder unter 13 Jahren (infans I und II) (= 36,2%), 4 Jugendliche und 1 juvenil bis frühadultes Individuum (4,8%), sowie 62 Erwachsene (59%) (Abb. 2). Von den erwachsenen Individuen sind 27 sicher oder wahrscheinlich weiblich bestimmt (43,5%) und 19 sicher oder wahrscheinlich männlich (30,7%); 16 Erwachsene sind nicht geschlechtsbestimmt (25,8%). Abb. 1 Abb. 1 Skizze der Alvensleben-Kaserne mit ungefährer Ausdehnung des Gräberfelds der Lausitzer Kultur. ein Gebäude neu errichtet (Abb. 1). Da seit 1885 die Existenz von Gräbern der Lausitzer Kultur bekannt war, wurden die Bauarbeiten archäologisch begleitet (GAIDA 1999, GRAMSCH 1999). 132 Befunde der Bronze- und frühen Eisenzeit wurden ergraben. Wenige Befunde datieren an das Ende der Stufe Bronze D mit Inventaren der sogenannten „Fremdgruppenzeit”, die Zahl der Befunde nimmt in der Stufe Hallstatt A deutlich zu, die meisten Bestattungen wurden aber in der späten Urnenfelderzeit in Stufe Ha B und wäh- Abb. 2 Alters- und Geschlechtsverteilung der Individuen des Gräberfelds Cottbus Alvensleben-Kaserne. aktuell 74 Leichenbrand erzählt vom Umgang mit den Toten ... Abb. 3 Bergung der Urnen und Beigefäße während der Grabung mithilfe elastischer Binden. Abb. 4 Zur zeichnerischen Dokumentation der Lage der verbrannten Knochen und Kleinfunde in der Urne wurde ein Zeichengitter verwendet. Ziel des Leipziger Projektes war es, die Handlungen des Bestattungsrituals möglichst detailliert zu rekonstruieren. Ausgehend von der These, dass diese Handlungen eine Form der Kommunikation mithilfe materieller Kultur sind, sollte untersucht werden, wie die einzelnen Verstorbenen im Ritual behandelt und bestattet worden waren und wie dadurch die soziale Identität der Bestatteten dargestellt und transformiert worden war; dazu weiter unten mehr. Eine Quelle zur Beantwortung dieser Fragen waren die Leichenbrände selbst. Sie wurden mit den aufgeführten Methoden alters- und geschlechtsbestimmt; darüber hinaus waren folgende Arbeitsschritte notwendig, um den Ablauf der Funeralpraktiken rekonstruieren zu können: nachlässigt wurden, zum anderen diese Daten interdisziplinär auszuwerten, d.h. in enger Zusammenarbeit zwischen Anthropologie und Archäologie. Wie eingangs erwähnt sind Datenerhebung und Datenauswertung abhängig von den sozialgeschichtlichen Fragestellungen, die wir im folgenden Abschnitt vorstellen. Die Darstellung sozialer Identitäten im Bestattungsritual Gräber standen im Mittelpunkt der Forschung seit der Entstehung der bürgerlichen Archäologie als „vaterländische Altertumskunde“ im 19. Jahrhundert, während Siedlungen vergleichsweise spät erforscht wurden (GRAMSCH 2007a). Mit ihrer Hilfe wurden nicht nur relative Chronologien erarbeitet, sondern auch anhand von Unterschieden in Beigaben und Grabbau die Sozialstruktur analysiert. Dabei hat sich die traditionelle Gräberfeldarchäologie vornehmlich auf vertikale soziale Strukturen konzentriert, also auf Rangordnung und Klassen, und hierbei insbesondere auf mögliche Eliten. Die sozialgeschichtliche Fragestellung bei der Untersuchung des Gräberfelds Cottbus Alvensleben-Kaserne dagegen richtete sich auf die horizontale Sozialstruktur. Die horizontale Sozialstruktur wird insbesondere durch Alter und soziales Geschlecht der Individuen geprägt. Diese Aspekte bestimmen wesentlich die soziale Identität des Individuums. Ziel unserer Analyse war deshalb die Beantwortung der Frage, ob und wie im Bestattungsritual Unterschiede in der sozialen Identität, d.h. in Gender und Alter3 der Individuen dargestellt werden durch den Umgang mit • Während der Grabung waren alle vollständig erhaltenen Urnen und Beigefäße mithilfe elastischer Binden geborgen worden, darunter auch solche Gefäße, die in situ zerbrochen waren (Abb. 3). • Dies ermöglichte die „Werkstattuntersuchungen“, d.h. das kleinschrittige schichtweise Entnehmen des Leichenbrands in z. T. mehr als 10 Schichten. • Bei dieser Bergung des Leichenbrands wurden die Schichten und die Lage und Größe der verbrannten Knochen und von Beigaben fotografisch und z.T. zeichnerisch dokumentiert (Abb. 4). • Die geborgenen Knochenfragmente wurden nach Schichten getrennt aufbewahrt und entsprechend anthropologisch untersucht. Diese detaillierte Werkstattuntersuchung ermöglichte zweierlei: zum einen Daten in einem großen Umfang zu erheben, die bisher eher ver- 75 aktuell Birgit Großkopf und Alexander Gramsch 1989). Wie A. VAN GENNEP und V. TURNER zeigten, ist die Stufe der Liminalität die komplexeste und die bedeutsamste für die Sozialanalyse. Die bisherige Identität des Individuums wird aufgelöst und seine „Wiedergeburt“ in einem neuen Status vorbereitet. Der transformative Charakter von Ritualen, besonders Bestattungsritualen, ist besonders während der Liminalität stark ausgeprägt (vgl. HUNTINGTON/METCALF 1979, 122). Von den Handlungen, die den Umgang mit dem Leichnam betreffen, können wir vor allem jene erschließen, die während dieser Phase der Auflösung, Umkehrung und Transformation ausgeführt wurden: Hierzu rechnen wir • die Platzierung des Leichnams auf dem Scheiterhaufen, • den Verbrennungsvorgang und • die anschließende Handhabung der Überreste beim Einbetten in die Urne4. Ausgehend vom Konzept der kommunikativen Handlungen betrachten wir mögliche Unterschiede in der Behandlung der oder des Toten in dieser Phase als Hinweise auf unterschiedliche soziale Identitäten (GRAMSCH 2007b, in Vorb.). Deshalb ist es wichtig, die Handlungen zu rekonstruieren, die uns „erzählen“, wie mit dem Leichnam umgegangen wurde – damit kommen wir also wieder zurück zu den Daten. den Toten, also durch die rituellen Handlungen (GRAMSCH 2007b). Dies bedeutet gegenüber traditionellen Gräberfeldanalysen eine zweifache Veränderung: Zum einen werden die archäologischen Daten nicht als „Spiegel des Lebens“ verstanden, die unmittelbar Rang oder Status eines Individuums erschließen lassen, sondern wir betrachten das Grab als das Resultat einer Reihe von Aktionen. Zum anderen verstehen wir diese rituellen Handlungen als Mittel der Darstellung und Transformation der sozialen Identität der Bestatteten. Die Abfolge dieser Handlungen ist durch das Bestattungsritual mehr oder weniger stark festgelegt; dennoch liegen ihnen Entscheidungen zugrunde: Welche Handlungen durchgeführt werden und in welcher Reihenfolge ist weder eine willkürliche Entscheidung noch Zufall oder reiner Pragmatismus. Kommunikative Handlungen Deshalb können die Handlungen etwas über die Entscheidungen und die Intentionen der Handelnden verraten. Die Praktiken, aus denen das Ritual besteht, sind also kommunikative Handlungen. Während zweckrationales Handeln erfolgsorientiert ist, d.h. auf Beeinflussung eines Handlungspartners ausgerichtet, um eine Absicht, ein erstrebtes Ergebnis zu erreichen, ist kommunikatives Handeln verständigungsorientiert, d.h. auf Kommunikation und Sozialisation ausgerichtet (HABERMAS 1981a, 384f.). Wir können unserer Analyse Habermas’ Definition kommunikativen Handelns zugrunde legen: „Unter dem funktionalen Aspekt der Verständigung dient kommunikatives Handeln der Tradition und der Erneuerung kulturellen Wissens; unter dem Aspekt der Handlungskoordinierung dient es der sozialen Integration und der Herstellung von Solidarität; unter dem Aspekt der Sozialisation schließlich dient kommunikatives Handeln der Ausbildung von personalen Identitäten“ (HABERMAS 1981b, 208). Der Umgang mit dem Körper des Verstorbenen kann also etwas darstellen und vermitteln: dessen individuelle soziale Identität und deren Transformation. Bestattungsrituale sind rites de passage und begleiten den Übergang von einer Lebensphase in eine andere, d.h. die Transformation des verstorbenen Individuums. Wie alle Übergangsrituale folgen sie einer dreiteiligen Struktur: Es gibt eine Phase der Loslösung (Separation), der Umkehrung (Liminalität) und der Wiedereingliederung (Reintegration) (VAN GENNEP 1986, TURNER aktuell Die Rekonstruktion ritueller Handlungen Wir haben eingangs auf bestimmte Daten hingewiesen, durch die diese funeralen Handlungen rekonstruiert werden können: • Die Repräsentanz des Leichenbrandes gibt an, ob die verbrannten Knochen aller Skelettregionen eingesammelt und in die Urne deponiert wurden. • Die primären Kohlenstoffverfärbungen zeigen, ob alle Körperregionen gleichmäßig dem Scheiterhaufenfeuer ausgesetzt waren. • Die Fragmentgröße lässt z.B. Rückschlüsse zu, ob die Knochen mechanisch ungestört auskühlen konnten oder zerkleinert wurden bzw. durch ein Ablöschen des Scheiterhaufens zersprungen sind. • Die Art der Deponierung kann auf den Vorgang der Bergung der Knochen aus dem Scheiterhaufen hinweisen. Zentral war vor allem die Frage, ob die verbrannten Knochen auf eine bestimmte Art und Weise in die Urne eingeschichtet bzw. in der Urne 76 Leichenbrand erzählt vom Umgang mit den Toten ... Abb. 5 Planumszeichnung einer Knochenschicht in Urne 4 aus Grab 56 Abb. 6 Große Abschnitte annähernd parallel ausgerichteter Diaphysen ließen sich z.B. in mehreren Schichten in Gefäß 1 aus Grab 53 beobachten (hier: Schicht 8) angeordnet worden waren. Die detaillierte Dokumentation der Lage der Skelettreste in der Urne und ihre Bergung und anthropologische Untersuchung nach Schichten getrennt zeigte eine erstaunlich große Regelhaftigkeit in der Behandlung der Leichenbrände. Bei fast allen Leichenbränden weisen die Repräsentanz der überlieferten Körperregionen und die Vorkommen und Verteilung primärer Kohlenstoffverfärbungen darauf hin, dass der Körper (annähernd) ausgestreckt auf dem Scheiterhaufen gelegen haben muss. Bis auf wenige Ausnahmen sind alle Körperregionen recht gleichmäßig verbrannt und etwa in gleichem Maße erhalten. Auch die Extremitätenknochen und selbst die kleinen Hand- und Fußknochen sind in großer Zahl und meist vollständig verbrannt vorhanden. Wir schließen daraus, dass die Scheiterhaufen ausreichend groß angelegt worden waren, um das Individuum vollständig zu verbrennen. Außerdem erkennen wir hieraus, dass die verbrannten Überreste der Verstorbenen meist sehr gründlich und recht vollständig aus dem heruntergebrannten Scheiterhaufen herausgelesen worden waren; es fand sich auch fast keine Holzkohle in den Urnen. In der Urne sind die Knochenfragmente oft bis zu einer Länge von 10 cm erhalten, selten auch darüber. Die Knochen sind also nicht zerkleinert worden sind, bevor sie in die Urnen kamen. Vor allem aber ist der Scheiterhaufen nicht abgelöscht worden, sondern die Reste des Brandes konnten auskühlen, bevor sie eingesammelt wurden, da sonst die Knochen viel kleinteiliger zersprungen wären. Das Fotografieren und Zeichnen der Knochen in der Urne zeigte, dass viele Fragmente ursprünglich recht groß waren, als sie in die Urnen gelegt wurden, und erst während der Liegezeit weiter zerbrachen. Die verbrannten Knochen müssen zum Zeitpunkt der Deponierung in der Urne soweit ausgekühlt gewesen sein, dass man sie mit Händen anfassen konnte. Aufgrund von experimentalarchäologischen Untersuchungen (z.B. LEINEWEBER 2002) können wir deshalb davon ausgehen, dass Verbrennung und Knochenbergung mindestens zwei Tage umfassten. Am auffälligsten ist die Einschichtung des Leichenbrands: In fast allen Fällen waren die verbrannten Knochen so in die Urne geschichtet worden, dass Fragmente der unteren Extremitäten zuunterst lagen, darüber jene des Torsos, oben die oberen Extremitäten und zuletzt die Schädelknochen. Dass das postkraniale Skelett unten und die Schädelknochen oben in den Urnen liegen, war bereits früher beobachtet worden (z.B. KEILING 1962, BUCK 1977, GEBÜHR ET AL. 1989)5. Jedoch gehen unsere Erkenntnisse weiter. Die Dokumentation der Lage der Knochen (Abb. 5 u. 6) zeigt in fast allen Urnen eine sorgfältige Deponierung der verschiedenen Skelettregionen entsprechend ihrer anatomischen Abfolge; Vermischungen lassen sich nur feststellen, wo kleinere Fragmente in tiefere Regionen der Urne rutschten. Auch Fuß- und Handphalangen, die beim Einschichten leicht zu verwechseln gewesen wären, fanden sich ganz überwiegend an der anatomisch „richtigen“ Stelle. Gelegentlich waren Wirbelknochen in einer Reihe zueinander gelegt worden; häufiger 77 aktuell Birgit Großkopf und Alexander Gramsch ihre ehemalige soziale Identität ist aufgelöst und sie wurden vereinheitlicht zu „Ahnen“. waren Diaphysen parallel angeordnet und Extremitätenknochen paarig nebeneinander oder die Gelenkköpfe einander gegenüber platziert worden (Abb. 6). Diese sorgfältige Niederlegung der verbrannten Knochen in der Urne erweckt den Eindruck einer Wiederherstellung der Form des menschlichen Körpers; wir haben diese rituellen Handlungen als eine „Anthropomorphisierung“ der Urne bezeichnet (GRAMSCH & GROSSKOPF 2005; GRAMSCH 2007c). Darüber hinaus ließ sich rekonstruieren, dass in der Regel jedes Individuum einzeln verbrannt und in einer eigenen Urne bestattet wurde. In fünf Fällen jedoch waren zwei Individuen gemeinsam in einer Urne bestattet. Hier scheint das gemeinsame Deponieren in die Urne wichtiger gewesen zu sein als die Darstellung von Individualität6. Doch wurde auch hier, soweit erkennbar, die Idee der „menschengestaltigen“ Urne gewahrt. Durch die interdisziplinäre Zusammenarbeit von Anthropologie und Archäologie können also die Knochen auf eine bestimmte Weise zum „Sprechen“ gebracht werden. Sie „erzählen“ dann nicht nur etwas über die biologischen Grundlagen der Individuen, sondern geben auch Antworten auf sozial- und kulturhistorische Fragen. Im hier vorgestellten Beispiel vermitteln sie uns ein Bild davon, wie im Verbrennungsritual die individuellen menschlichen Körper behandelt wurden, wie die Körper verändert wurden parallel zur Veränderung des verstorbenen Individuums, d.h. sie „erzählen“ von der Transformation in eine neue, einheitliche Identität, die wir als „Ahnen“ interpretieren können. Werden Gräber also als Resultat komplexer, vieldeutiger und kommunikativer Handlungen aufgefasst, vermitteln sie kein statisches Bild, aus dem soziale Verhältnisse direkt abzulesen wären, sondern sie liefern kontextabhängige Daten zu Übergangsritualen und deren Rolle in der Darstellung und Transformation sozialer Identitäten. Die sozialgeschichtliche Interpretation Nahezu alle derart untersuchten Individuen wurden auf die gleiche Weise behandelt, seien es Neugeborene oder über 60jährige Greise, Männer oder Frauen. Soziale Unterschiede wurden demnach nicht bei der Behandlung der individuellen Körper gemacht. Gewisse Unterschiede sind jedoch erkennbar in der Gabe von Schmuck und Keramik und in der Beisetzung in einem Einzelgrab oder gemeinsam mit weiteren Verstorbenen in einem Doppel- oder Mehrfachgrab (GRAMSCH in Vorb.). Die Rekonstruktion und Interpretation der rituellen Handlungen lassen also die Knochen davon „erzählen“, dass die Transformation der Verstorbenen durch Verbrennung und anschließende Anthropomorphisierung unabhängig von Alter und Geschlecht durchgeführt wurde. Ebenso „erzählen“ sie, dass meist ein derart transformierter Toter auch durch EINE Urne repräsentiert wird, da in der Regel jede Urne den Leichenbrand nur EINES Individuums aufnahm. Die Gemeinschaft, die das Bestattungsritual durchführte, transformierte somit die Verstorbenen alle auf die gleiche Weise. Die Körperbehandlung als Kommunikation über soziale Identitäten entwirft ein idealisiertes Bild, das möglicherweise bestehende Unterschiede ignoriert oder gar überspielt – gerade der Umgang mit dem individuellen Körper und seine Transformation stellt alle Menschen gleich dar, als Teil der Gemeinschaft. Am Ende dieses Prozesses sind alle Toten gleich, aktuell Anmerkungen 1 Für die Untersuchung materieller Kultur unterschied Heinrich HÄRKE (1993) zwischen funktionalen und intentionalen Daten. Da menschliche Skelettreste zunächst funktionale, biologische Daten liefern, die nicht von den Trägern der prähistorischen Gesellschaft beabsichtigt wurden, z.B. zu körperlichen Belastungen, aber auch intentionale Daten liefern können, die von absichtsvollen Handlungen wie z.B. Modifikationen am lebenden oder toten Körper zeugen, können auch sie zur materiellen Kultur gerechnet werden. 2 Unser Dank gilt der Unterstützung durch Prof. S. Rieckhoff und dem Sächsischen Ministerium für Wissenschaft und Kultur für die finanzielle Unterstützung des Projekts „Herrschaft und Geschlechterdifferenz im 1. Jahrtausend v.Chr. – Spätbronzezeitliche Gesellschaften in der Niederlausitz (Brandenburg) aus Sicht der Genderforschung“. 3 Soziales Geschlecht und soziales Alter beruhen auf der biologischen Grundlage, sind aber nicht zwingend identisch mit dieser. 4 Auch die Errichtung des Grabbaus und möglicherweise das Auswählen und Präsentieren der wegzugebenden Grabkeramik können Teil der Phase der Liminalität gewesen sein. Zur Parallelisierung der Bestattung und insbesondere der Kremation mit den Phasen der rites de passage s. GRAMSCH 2004, 2007b, 2007c. 78 Leichenbrand erzählt vom Umgang mit den Toten ... 5 Häufig wird nur beobachtet, dass Schädelfragmente oben auf dem Leichenbrand liegen. In Fällen, in denen dies nicht zu beobachten ist, wird im Umkehrschluss gefolgert, dass keine Schichtung vorliegt und der Leichenbrand daher nicht aufwändig in Schichten geborgen werden muss. Cottbus Alvensleben-Kaserne (Brandenburg). Arch. Nachrichtenblatt 11, 2006, 328–334. – (2007a): Ein Abriss der Geschichte der Prähistorischen Archäologie in Deutschland: Genese, Entwicklung und Institutionalisierung. Das Altertum 52, 2007, in Druck. 6 In zwei Urnen waren jeweils zwei Kinder gemeinsam bestattet worden. Aus einer Urne, deren Inhalt nicht schichtweise untersucht werden konnte, stammt Leichenbrand eines Mannes und einer Frau. In zwei Fällen lagen die Knochen von neugeborenen oder perinatalen Kindern zwischen denen von erwachsenen Frauen; hier ist eine Interpretation als „Mutter-KindBestattungen“ möglich. – (2007b): A microarchaeological approach to the social significance of Late Bronze Age burial practices. In: CORNELL, P./F. FAHLANDER (Hrsg.): Encounters – Materialities – Confrontations: Archaeologies of Social Space and Interaction. Newcastle 2007, 83–99. – (2007c): Rekonstruierte Körper – Körperinszenierung in der rituellen Kommunikation der Lausitzer Kultur. In: VEIT, U. et al. (Hrsg.): Körperinszenierung, Objektsammlung, Monumentalisierung. Totenritual und Grabkult in frühen Gesellschaften. Tübinger Arch. Taschenbücher Bd. 6. Münster 2007, in Druck. Literatur – (in Vorb.): Ritual und Gemeinschaft. Die Darstellung von Altersklassen und Geschlechterdifferenz in der rituellen Kommunikation der Lausitzer Kultur. UPA. Bonn. BÖNISCH, E. (1990): Das jungbronzezeitliche Gräberfeld der Lausitzer Kultur Saalhausen 2, Kr. Senftenberg. Veröff. Mus. Potsdam 24, 1990, 63–169. – (1995): Holzkistengräber, Steinkreise und urgeschichtliche Grabstörungen auf jüngstbronzezeitlichen Bestattungsplätzen in der Niederlausitz. Arbeits- u. Forsch.ber. z. sächs. Bodendenkmalpfl. 37, 1995, 105–116. GRAMSCH, A./B. GROSSKOPF (2005): Altersklassen und Geschlechterdifferenz – „making the difference“ in der Genderarchäologie. In: FRIES, J. E./J. KOCH (Hrsg.): Ausgegraben zwischen Materialclustern und Zeitscheiben. Perspektiven zur archäologischen Geschlechterforschung. Frauen-ForschungArchäologie, Bd. 6. Münster 2005, 75–94. BUCK, D.-W. (1977): Die Billendorfer Gruppe. Teil 1, Katalog. Veröff. Mus. Ur- u. Frühgesch. Potsdam 11. – (1986): Hallstattzeitliche Kammergräber der Lausitzer Kultur und ihr sozialökonomischer Hintergrund. In: Hallstatt Kolloquium Veszprém 1984. Budapest 1986, 19–23. GROSSKOPF, B. (1990): Individualaltersbestimmung mit Hilfe von Zuwachsringen im Zement bodengelagerter menschlicher Zähne. Zeitschr. f. Rechtsmed. 103, 1990, 351–359. – (2007): Zur Anlage und Gestaltung von Gräberfeldern der Lausitzer Kultur am Beispiel von Klein Lieskow. Veröff. Museum Westlausitz 27, 2007, 3–58. - (2004): Leichenbrand – Biologisches und kulturhistorisches Quellenmaterial zur Rekonstruktion vor- und frühgeschichtlicher Populationen und ihrer Funeralpraktiken. Dissertation Universität Leipzig. Online-Publikation http://dol.dl.uni-leipzig.de/ receive/DOLDissHabil_disshab_00000356. GAIDA, J. (1999): Urnengräber der Billendorfer Kultur aus der General-von-Alvensleben-Kaserne, Stadt Cottbus. Archäologische Begleituntersuchungen beim Neubau einer Kfz-Halle 1997. Einsichten Archäologische Beiträge für den Süden des Landes Brandenburg 2, 1998. Cottbus 1999, 31–40. GROSSKOPF, B./A. GRAMSCH (2004): Leichenbrand – biologische Quelle für Genderforschung. Anthropologischer Anzeiger 62/3, 2004, 281–289. HABERMAS, J. (1981a): Theorie des kommunikativen Handelns, Bd. 1. Frankfurt a.M. 1981. GEBÜHR, M./HARTUNG, U./H. MEIER (1989): Das Gräberfeld von Neubrandenburg. Beobachtungen zum anthropologischen und archäologischen Befund. Hammaburg NF 9, 1989, 85–107. – (1981b): Theorie des kommunikativen Handelns, Bd. 2. Frankfurt a.M. 1981. HÄRKE, H. (1993): Intentionale und funktionale Daten. Ein Beitrag zur Theorie und Methodik der Gräberarchäologie. Arch. Korrbl. 23, 1993, 141–146. GRAMSCH, A. (1999): Gräber im Kasernenhof. Die Lausitzer Nekropole der Alvensleben-Kaserne in Cottbus. Archäologie in Berlin und Brandenburg 1998. Stuttgart 1999, 55–57. HERRMANN, B. (1988): Behandlung von Leichenbrand. In: KNUSSMANN, R. (Hrsg.): Anthropologie. Handbuch der vergleichenden Biologie des Menschen. Stuttgart/ New York 1988, 576–585. – (2006): Ritual und Gemeinschaft. Die Darstellung von Altersklassen und Geschlechterdifferenz in der rituellen Kommunikation der Lausitzer Kultur (12.-8. Jh. v.Chr.) anhand des Gräberfeldes 79 aktuell Birgit Großkopf und Alexander Gramsch TURNER, V. (1989): Das Ritual. Struktur und AntiStruktur. Frankfurt/M. 1989. HERRMANN, B. et al. (1990): Prähistorische Anthropologie. Leitfaden der Feld- und Labormethoden. Berlin u.a. VAN GENNEP, A. (1986): Übergangsriten. Frankfurt/M. 1986. HODDER, I. (1989): This is not an article about material culture as text. Journal of Anthropological Archaeology 8, 1989, 250–269. VAN VARK, G. N./W. H. M. AMESZ-VOORHOEVE/ A. G. F. M. CUIJPERS (1996): Geschlechtsdiagnose an menschlichem Leichenbrand mit Hilfe mathematischstatistischer und datenanalytischer Methoden. Homo 47, 1996, 305–338. – (1991): Reading the Past. Current Approaches to Interpretation in Archaeology, 2nd ed. Cambridge 1991. – (2005): Post-processual and interpretive archaeology. In: RENFREW, C./P. BAHN (Hrsg.): Archaeology. The Key Concepts. London 2005, 207–212. VEIT, U. (2003): Menschen – Objekte – Zeichen: Perspektiven des Studiums materieller Kultur. In: VEIT, U. ET AL. (Hrsg.): Spuren und Botschaften: Interpretationen materieller Kultur. Tübinger Archäologische Taschenbücher Bd. 4. Münster 2003, 17–28. HODDER, I./S. HUTSON (2003): Reading the Past. Current Approaches to Interpretation in Archaeology, 3rd ed. Cambridge 2003. HOLTORF, C. (2003): Archäologie als Fiktion – Anmerkungen zum Spurenlesen. In: VEIT, U. ET AL. (Hrsg.), Spuren und Botschaften: Interpretationen materieller Kultur. Tübinger Archäologische Taschenbücher Bd. 4. Münster 2003, 531-544. Birgit Grosskopf Johann Friedrich Blumenbach Institut für Zoologie und Anthropologie Historische Anthropologie und Humanökologie Bürgerstraße 50 37073 Göttingen HUNTINGTON, R./P. METCALF (1979): Celebrations of death. The anthropology of mortuary ritual. Cambridge 1979. KEILING, H. (1962): Ein Bestattungsplatz der jüngeren Bronze- und vorrömischen Eisenzeit von Lanz, Kr. Ludwigslust. Der Leichenbrand. Jahrbuch Bodendenkmalpflege in Mecklenburg 62, 1962, 36-38. Alexander Gramsch Museum der Westlausitz Macherstraße 140 LEINEWEBER, R. (2002): Brandneu. Verbrennung auf dem Scheiterhaufen. Studie über branddeformierte Beigaben aus Brandgräbern der römischen Kaiserzeit Innergermaniens. Experimentelle Archäologie in Europa, Bilanz 2002. Oldenburg 2002, 159–171. aktuell 80 Paläoanthropologie – Standortbestimmung einer innovativen Disziplin Winfried Henke Zusammenfassung – Die Paläoanthropologie zählt unter den öffentlichkeitsorientierten biologischen Wissenschaften zu den faszinierendsten Disziplinen, hat aber auch gleichzeitig den diskreditierenden Vorwurf zu gegenwärtigen, keine sehr glaubwürdige Disziplin zu sein. Zielsetzung dieses Beitrags ist es, dieses Vorurteil als einseitige Außensicht zu entkräften und die paläoanthropologische Forschung als eine innovative Wissenschaft zu kennzeichnen, deren Aufgabe es ist, den Prozess der Menschwerdung als adaptive Entwicklung in der Primaten-Evolution zu verstehen. Neben den Prinzipien werden die wichtigsten Methoden, mit denen es gelingt, die in Homininenfossilien verschlüsselten Informationen akribisch zu dekodieren, exemplarisch aufgezeigt. ’Begründete’ Paläoanthropologie, um die es hier geht, ist stets theoriegeleitete und hochgradig inter- und multidisziplinär vernetzte Stammesgeschichtsforschung. Nur mit einem komplexen vergleichend-primatologischen Ansatz kann es gelingen, den Prozess und die Etappen der Menschwerdung zu rekonstruieren und langfristig gültige Hominisationsmodelle und phylogenetische Stammbäume zu erstellen. Mehr als Modelle (Jeweilsbilder sensu Heberer 1972), die aufgrund neuer Methoden und Fakten ständig überprüft werden müssen, werden es jedoch niemals sein. Wir modellieren nur! Schlüsselwörter – Fossilien, Hominini, Morphologie, Systematik, Modelle Abstract – Palaeoanthropology is as a publicity-oriented biological science one of the most fascinating disciplines which has to deal simultaneously with the discrediting allegation of a low credibility of her results. This paper aims to invalidate this prejudice as a singleedged exterior view and to characterise palaeoanthropological research as an innovative science which aims to explain the process of hominisation as evolutionary adaptations within the order Primates. The principles and methods of theory-guided studies of fossil remains are exemplarily explained. Palaeoanthropological approaches meticulously try to decode the information hidden in hominid fossils, to reconstruct the process and the phylogenetical steps of hominisation and to conceptualize a substantiated – but always hypothetical – pedigree of our origin. We are just modelling. Keywords – fossils, hominini, morphology, systematics, models Einführung Seit der Entwicklung der Deszendenz- und Selektionstheorie durch Charles Darwin vor fast 150 Jahren sind wir mit der Vorstellung konfrontiert, dass sich unsere Existenz aus tierischen Vorformen ableitet und wir mit anderen Organismen real-historisch in einem phylogenetischen Kontinuum stehen. Die anfänglich helle Aufregung über Darwins Hauptwerk „On the Origins of Species by Means of Natural Selection“ (DARWIN 1859) sowie die beiden anthropologisch ausgerichteten Bände “The Descent of Man, and Selection in Relation to Sex (DARWIN 1871) und „The Expressions of Emotions in Man and Animals“ (DARWIN 1872) ist längst verflogen, aber die „Darwinsche Kränkung“ sitzt offenbar immer noch tief, zumal „Darwin’s Dangerous Idea“ sensu Daniel Dennett (1996) sich wie eine „ätzende Säure“ in die Erklärung aller Lebensbereiche hineingefressen hat. Die Gegenreaktion auf das Paradigma der Evolutionsbiologie kommt heute nicht mehr von den christlichen Kirchen [von wenigen aparten Ausnahmen wie dem überholten Vorstoß von Christoph Kardinal Schönborn (2007) abgesehen], sondern insbesondere von Kreationisten und Theoretikern des Intelligent Design, die nicht nur in den U.S.A. verstärkt Anhänger finden, sondern auch in Europa, wo der Nährboden für ihr wissenschaftliches Analphabetentum statistischen Umfragen zufolge ebenfalls breiter geworden ist. Da die von kreationistischer Seite geäußerten Zweifel an den Darwinschen Theorien nicht wissenschaftlich begründet und daher haltlos sind (vgl. auch JUNKER UND HOSSFELD 2001; KUTSCHERA 2006), muss gefragt werden, warum der abendländische aufklärerische Bildungsanspruch versagt hat. In diesem Kontext gilt es nachdrücklich zu betonen, dass der Darwinismus keine Weltanschauung ist, sondern eine wissenschaftliche Theorie, die zentrale Theorie der Biologie, weshalb der Populationsgenetiker Theodosius DOBZHANSKY (1973) mit Recht formulierte: „Nichts macht Sinn in der Biologie, außer im Lichte der Evolution.“ Eines Lückenbüßer-Gottes, wie die Kreationisten ihn postulieren, bedarf es nicht. Grundsätzlich sollten Glaubens- und Wissenskategorien strikt auseinander gehalten werden, aber offenbar ist das nicht einmal in allen Kultusministerien anerkannt (siehe die durch die KM Karin Wolff entfachte curriculare Diskussion in Hessen). Theologische Erklärungen dieser Welt, d.h. Schöpfungstheorien, religiöse Glaubensvorstellungen, bedürfen keines wissenschaftlichen Beweises, da die Gläubigen die Gewissheit über die Schöpfungskraft ihres Schöpfers in sich tragen Archäologische Informationen 30/1, 2007, 1-23 Bulletin de la Société Suisse1d‘Anthropologie 13 (1), 2007 Bulletin der Schweizerischen Gesellschaft für Anthropologie 13 (1), 2007 Winfried Henke (vgl. LÜKE ET AL. 2004; SÖLING 2004; HENKE 2006c). Es bleibt damit fraglich, ob es überhaupt ein gegenseitiges Anerkennen von Glauben und Wissenschaft geben kann, aber, so betonen STORCH ET AL. (2007) wohl mit Recht, ein kritischer Dialog über die jeweils andere Art des Umgangs mit der Wirklichkeit muss möglich sein – und sie fügen hinzu: „Beiden Bereichen, Glauben und Wissenschaft, ist gemeinsam, dass sie – auf unterschiedliche Weise – in einem Spannungsfeld zwischen Gewissheit und Zweifel stehen.“ Der Biochemiker Erwin CHARGAFF (2002, S. 9) fasste die Feststellung, dass Naturwissenschaft immer konjektural bleiben wird, bestechend feinsinnig: „Ich kenne einige Vorgänge, die zum Leben beitragen, aber ich kenne sie nur insoweit, als die von mir verwendeten Methoden zugänglich sind.“ Wir modellieren nur! – und die in der Darwinschen Evolutionstheorie formulierten generellen Evolutionsprinzipien sind das Fundament der biologischen Modellbildungen (Abb. 1), die sowohl unseren Ursprung als auch unsere phylogenetische Entwicklung betreffen. Der durch den Darwinismus induzierte Paradigmenwechsel liegt im Verwerfen der Vorstellung „…einer auf Vollkommenheit angelegten und geplanten, gütigen Natur“ (vgl. MAYR 1994) zugunsten der Konzeption des ‚struggle for life’. Nach Günter ALTNER (1982, S. 5) besteht die hochgradig desillusionierende Wirkung der ‚Darwinschen Revolution’ gegenüber Schöpfungsmythen darin, dass der „…Mensch auf sich und sein Werden zurückgeworfen“ wurde. Natürliche Schöpfung ohne Schöpfer, das scheint uns, die wir die Natur als „zweckmäßig“ erfahren und unser Leben bewusst planen – oder zumindest einem Lebensentwurf folgen – , kaum begreifbar; es brauchte deshalb auch lange, um die Teleologie (Lehre der ziel- und zweckbestimmten Ordnung) gänzlich aus der empirischen Biologie zu verbannen, wie Konzepte der ‚idealistischen Morphologie’, des ‚Vitalismus’ oder Teilhard de Chardins‚naturwissenschaftlich-theologisches Mischkonzept’ zeigen (vgl. VOGEL 1982). Das offenbar für Viele Unvorstellbare ist, die evidente Zweckmäßigkeit der Welt durch ‚reinen Zufall’ zu erklären. Um deutlich zu machen, dass die Evolution im Sinne Darwins, stochastisch betrachtet, nicht ‚rein’ zufällig und ‚ungerichtet’ verläuft, sondern selbstorganisatorisch ‚kanalisiert’, sprach der Freiburger Genetiker Carsten BRESCH (1977) pointiert vom „schmutzigen“ Zufall, „… weil jede via anpassende Selektion erzwungene „Entscheidung“ auf dem Wege einer organismischen genealogischen Stammlinie zugleich bestimmte Begrenzungen und Vorgaben aktuell Abb. 1 Systemansatz zur Analyse der Stammesgeschichte des Menschen [aus HARDT et al. (2006) nach FOLEY (1987), modifiziert). für zukünftige ‚Entscheidungsspielräume’ setzt“ (vgl. VOGEL 1982, S. XV). Evolution ist offen, ihre Produkte erweisen sich nur im Nachhinein betrachtet als zweckmäßig. Die evolutionsbiologische Herausforderung besteht daher darin, die Entwicklung der Organismen als selbstorganisatorischen Anpassungsprozess via Mutations- und Selektionsmechanismen zu verstehen, und zwar ohne inkonsistente Spekulationen aller Art. Das Hominisationsproblem ist damit nicht die Frage nach dem ‚großen Entwurf’ der Natur, denn die Anthropogenese verlief nicht zielgerichtet. Bereits Charles DARWIN hatte in seinem Erklärungsansatz zur Zweckmäßigkeit der Natur die ‚äußere’ Sinngebung aus der Verklammerung mit der ‚inneren’ Sinngebung gelöst, was PITTENDRIGH (1958) schließlich in dem Begriff Teleonomie in Abgrenzung zur Teleologie aristotelischer Tradition zum Ausdruck brachte. Auf den Menschen bezogen heißt das aber auch, sich nicht nur auf die Analyse und Interpretation des morphologischen Formenwandels zu beschränken, sondern auch unsere spezifische psycho-physische Konstitution evolutionsbiologisch zu erklären und die besonderen Rahmenbedingungen der Menschwerdung zu erfassen. Es gilt nicht nur, die den Menschen kennzeichnende Morphologie, wie z.B. die habituelle Bipedie so- 2 Paläoanthropologie – Standortbestimmung einer innovativen Disziplin wie die omnivore Ernährungsweise und exzessive Cerebralisation, am Fossilmaterial nachzuweisen und selektionstheoretisch zu erklären, sondern auch die Kulturfähigkeit, wie z.B. komplexe Werkzeugherstellung und -verwendung, gesteigertes tradigenetisches Verhalten, Symbolsprache, Geschichtlichkeit und soziale Verantwortung, als Adaptationsmuster zu verstehen (OSCHE 1983; VOGEL 1975, 1983, 2000; HENKE UND ROTHE 1999a; ROTHE UND HENKE 2005). Dabei kommt hinsichtlich der Erklärung der Hominisation das Problem der Objekt-Subjekt-Identität erschwerend hinzu, denn der Mensch ist sowohl erklärendes Subjekt als auch erklärtes Objekt. Das erfordert den unverstellten Geist der Aufklärung und birgt aufgrund zu großer Nähe zum ‚Objekt Mensch’ die Gefahr der subjektiven Fehleinschätzung, was bereits Thomas H. Huxley als erkenntnistheoretisches Hindernis erkannte und deshalb vorschlug, sich „… einmal in die Stelle wissenschaftlich gebildeter Bewohner des Saturns zu versetzen, die hinreichend mit solchen Tieren, wie sie die Erde bewohnen, bekannt sind“ (HUXLEY 1863, s. Nachdruck 1963, S. 110), um die Verwandtschaftsbeziehungen unvoreingenommen und leidenschaftslos aus der Distanz zu bewerten. Außenstehende den Eindruck gewinnen, dass die Paläoanthropologie eine prinzipienlose Wissenschaft sei. In der Tat besitzen phylogenetische Hypothesen aufgrund neu entdeckter Fossilfunde und innovativer Methodenansätze eine kurze ‚Halbwertszeit’ respektive hohe Verfallsrate. Es wäre aber völlig verfehlt, die Paläoanthropologie bei voller Ausschöpfung der methodischen Möglichkeiten sowie Respektierung der Grenzen des Methodeninventars deshalb als wenig seriöse Disziplin abzuwerten. Man könnte den raschen Wandel der Modelle ja auch als Folge und Ausdruck der besonderen Innovativität und Dynamik des Faches werten, was aber blauäugig wäre, denn es ist nicht zu bestreiten, dass Fossilfunde und paläoanthropologische Befunde bisweilen journalistisch voreilig ‚vermarktet’ werden, dass es sich um eine hochgradig fossil and media driven science handelt. Schlagzeilen in Der Spiegel wie: „Großer Kampf um kleine Kerle. War der berühmte „Hobbit“ von der Insel Flores ein Krüppel oder der Vertreter einer neuen Menschenart? Der Forscherstreit geht in die nächste Runde“ – oder: „Fossilien lassen menschlichen Stammbaum wackeln“ […]. Nicht direkt aus Afrika, wie zuvor angenommen, sondern über Asien wanderten die ersten Europäer ein“, belegen angebliche oder tatsächliche Fehleinschätzungen, ständigen Wandel und harte Kontroversen. Letztere prägen das Image einer ‚flüchtigen’ Wissenschaft und diskreditieren die ‚Glaubwürdigkeit’ (engl. credibility) der Paläoanthropologie. Sie vermitteln in der Öffentlichkeit den Eindruck, die Paläoanthropologen hätten – in wissenschaftlicher Hinsicht – „Prinzipien wie Segelschiffe“, soll heißen, dass sie sich mit jedem neuen Fossil und jeder neuen Datierung in die Richtung bewegen, in die der „öffentliche Wind“ bläst. „Simple curiosity“ sensu Gaylord G. Simpson (vgl. WHITE 2000) ist keine hinreichende Basis für anerkannte paläoanthropologische Wissenschaft, wohl aber für Karrieristen, die dieses Fach offenbar stärker anzieht als andere Disziplinen. Deren maximale persönliche Bedrohung lautet offenbar: „Du kommst nicht ins Fernsehen!“ Die gegenwärtigen Wissenschaftsstrukturen sind an dieser Entwicklung nicht ganz schuldlos, denn die Gratwanderung zwischen Elfenbeinturm und Öffentlichkeit wird zunehmend schwieriger, zumal die die Forschung finanzierenden Institutionen die intensive mediale Umsetzung wissenschaftlicher Befunde als evaluationsrelevant werten. Im Informationszeitalter ist es deshalb nicht unerwartet, dass die „Ökonomie der Aufmerksamkeit“ sensu Stephan FRANCK (1998) diejenige biologische Disziplin, die unser eigenes Werden und Sein betrifft, Seitdem uns bewusst ist, dass auch der Mensch Geschichte hat, lauten die Kernfragen der Paläoanthropologie nach wie vor: • wer sind unsere nächsten lebenden Verwandten in der Primatenordnung, • wann und wo, d. h. an welcher Stelle im Primatenstammbaum, zweigte die zum Menschen führende Stammlinie ab, • welche speziellen evolutionsökologischen Rahmenbedingungen ermöglichten den Prozess der Menschwerdung; • wie viele fossile hominine Vorläuferformen gab es, und • wie verlief die evolutive Entstehung unseres spezifisch menschlichen Merkmalgefüges (insbesondere Geschichtlichkeit, Sprache, Moral)? Zielsetzung des vorliegenden Beitrags ist es, in aller gebotenen Kürze einige Prinzipien und Methoden der Paläoanthropologie darzulegen und das Natur-Kultur-Wesen Mensch als „another unique species“ sensu Robert FOLEY (1987) – eine andere einzigartige Spezies der Ordo Primates – zu kennzeichnen. Aufgrund des rapiden Methodenfortschritts der letzten Jahrzehnte und des Wechsels des stammesgeschichtlichen ‚Jeweilsbildes’ könnten 3 aktuell Winfried Henke besonders prägt; es verwundert nicht, dass die Paläoanthropologie aufgrund übermäßigen populärwissenschaftlichen Interesses und der Neigung seiner Vertreter, dieses intensiv zu bedienen, von außen häufig als das wahrgenommen wird, was sie bis zur Mitte des letzten Jahrhunderts auch weitestgehend war, eine ideographische, narrative Wissenschaft (vgl. auch WHITE 2000; HENKE UND ROTHE 2006; HENKE 2007). Eine detaillierte ‚Innenschau’ des Faches kommt jedoch zu einem ganz anderen Befund, denn der wissenschaftliche Fortschritt der letzten Dezennien kennzeichnet die Paläoanthropologie als eine faktenreiche, höchst innovative, theoriengeleitete Wissenschaft, als „begründete“ Stammesgeschichtsforschung, was nachfolgend verdeutlicht werden soll (HENKE UND ROTHE 1994, 1999a, 2003; HENKE UND TATTERSALL 2007). sensalamanders herausstellte, ferner die für die Konstituierung der Paläoanthropologie so folgenschwer fehldiagnostizierte Piltdown-Fälschung (‚Eoanthropus dawsoni’; vgl. SPENCER 1990a, b) oder Noel Boaz’ Fehlinterpretation einer Delfinrippe aus Nordwest-Afrika als hominoide Clavicula, von Tim White hämisch als „Flipperpithecus“ bezeichnet. Phylogenese ist also nie an den Funden selbst abzulesen, und auch der Zuwachs an Fossilien bringt nicht automatisch mehr Klarheit über stammesgeschichtliche Abläufe, sondern bewirkt bisweilen sogar das Gegenteil, d.h. lange Zeit für ‚wahr’ gehaltene phylogenetische Vorstellungen mussten aufgrund neuer Funde und Befunde verworfen werden. Menschen sind ständig nach „Wahrheiten“ suchende, auch in der Paläoanthropologie. Wer als Paläoanthropologe etwas über die Menschwerdung erfahren will, hat im Rahmen der Darwinschen Evolutionstheorie bzw. des seither stetig fortentwickelten Gebäudes der Systemtheorie der Evolution Hypothesen zu formulieren, und er hat auf der Basis geeigneten Methodeninventars den Versuch zu unternehmen, die Hypothesen zu verifizieren oder zu falsifizieren. Selbstverständlich ist auch der quantitative Aspekt, die Anzahl fossiler Fundstücke, für die phylogenetische Rekonstruktion wesentlich. Man kann aber zuverlässig prognostizieren, dass der Fossilreport stets defizitär bleiben wird; darüber hinaus ist die Suche nach dem ‚missing link’, dem Bindeglied zwischen Menschenaffen und Mensch, ein überaus fragwürdiges Konzept, und zwar in der Hinsicht, als es paläoanthropologisch nach kladistischen Vorstellungen nur um den „most recent common ancestor“ (MRCA), den letzten gemeinsamen Vorfahren, und die Bestimmung des Gabelungspunktes der zu den Schwestertaxa (Adelphotaxa) führenden Stammlinien von afrikanischen Menschenaffen und Menschen gehen kann (ROTHE UND HENKE 2006). Der rapide Anstieg von Hominidenfossilien ist nicht zufällig; er geht auf eine präzise geologische und archäologische Exploration und Planung von Feldstudien in fundträchtigen Regionen zurück. Dennoch hängt der Erfolg jeder Grabungsexpedition, insbesondere das Auffinden von Homininenfossilien, d. h. der ‚Nuggets’ jeder Grabung, von vielen Zufallsfaktoren ab. Grabungskompetenz, Fleiß und Ausdauer sind zwar wichtige Voraussetzungen für den Erfolg, aber keineswegs eine Erfolgsgarantie. Was die wissenschaftliche Analyse und Interpretation von Fossilien betrifft, so ist das Vorurteil zu widerlegen, dass Laborarbeit die gegenüber Feldstudien entschieden 2. Prinzipielles und Methodisches 2.1 Fachliche Vernetzung und innovative Trends Wissenschaftshistorische Trends in der Paläoanthropologie sind der sprunghafte Anstieg neuer Fossilfunde aufgrund systematisch geplanter und durchgeführter Grabungen (Stichwort GPS – Global Positioning System), die zunehmend fächerübergreifende Bearbeitung stammesgeschichtlicher Probleme (Multi- und Interdisziplinarität) mit positiven Konsequenzen für die Methodologie und empirische Forschung sowie die sprunghaften Innovationstrends und die wachsende Bedeutung zahlreicher Disziplinen wie Taphonomie, phylogenetische Systematik, Funktionsund Evolutionsmorphologie, Paläoökologie, Soziobiologie, Paläogenetik und Archäometrie für die Lösung evolutionsbiologischer Fragen (vgl. HENKE UND ROTHE 1994, 2003; HENKE 2005, 2006a, b, 2007; HENKE UND TATTERSALL 2007). Entgegen landläufiger Meinung ist die Stammesgeschichte an Fossilien, also an versteinerten Überresten früherer Lebewesen, nicht direkt erfahrbar. Fossilien sind zwar – trotz molekularbiologischer Befunde – unerlässliche Belege für stammesgeschichtliche Prozesse, sie liefern jedoch keine unmittelbare faktische Information über den Ablauf der Evolution. Das wird insbesondere dadurch deutlich, dass Fossilien in der Vergangenheit häufig verkannt oder eklatant missinterpretiert wurden; man denke nur an den ‚Homo diluvii testis’, Johann Jakob Scheuchzers ‚armen Sünder’, der sich als Skelett eines Rie- aktuell 4 Paläoanthropologie – Standortbestimmung einer innovativen Disziplin Modellen kommen, die über eine sog. Mittlertheorie aufgrund empirischer oder quasi-empirischer Tests „Fenster zur Vergangenheit“ öffnen. Da die Ansprüche der Paläoanthropologie hoch gesteckt sind, ist das Fach auf die intensive Kooperation mit Nachbardisziplinen (Abb. 2) angewiesen. Eine intensive interdisziplinäre Vernetzung kennzeichnet deshalb die wachsende Attraktivität des Faches, das weit über eine reine Fossilkunde hinausgeht (HENKE UND ROTHE 2006; HENKE 2007). Was die aktuellen Wissenschaftstrends betrifft, die innovativen Felder der Paläogenetik (HUMMEL 2003, 2007; BURGER 2007) und Archäometrie (WAGNER 2007b), so befindet sich die gegenwärtige Forschung in einem sehr dynamischen Prozess. Es bleibt jedoch abzuwarten, ob es uns gelingen wird, taxonomische Fragen der Homininen-Evolution mittels alter DNA (aDNA) zweifelsfrei zu lösen, das Ticken der „Molekularen Uhr“ zuverlässig zu erfassen oder den „Speiseplan“ und die Abb. 2 VENN-Diagramm, das die Beziehungen jener Disziplinen aufzeigt, die an der Erforschung der Stammesgeschichte mitwirken [nach HENKE und ROTHE (1994), und HENKE und TATTERSALL (2007), modifiziert]. weniger anspruchsvolle Art der Forschung sei; sicherlich ist sie meist die weniger spektakuläre. Wer als Paläoanthropologe Ruhm ernten will, muss offenbar graben – nur dann erlangt er die gewünschte Aufmerksamkeit, die „unwiderstehlichste aller Drogen“ (FRANCK 1998). Um es salopp zu formulieren: „Adventure sells“; – aber bei näherem Hinsehen nehmen sich die Abenteuer im 3. Jahrtausend als kalkulierbar und risikofrei aus, es sei denn, man kommt – wie zu Goldgräberzeiten – seinen Kollegen in den Claims in die Quere (vgl. WHITE 2000; KALB 2001). Offenbar bedarf es sowohl höchst effizienter Feldstudien als auch exzellenter Laborstudien, um dem Prozess der Menschwerdung nachzuspüren. Der Systemansatz zur Analyse der Stammesgeschichte des Menschen (vgl. Abb. 1) verdeutlicht, dass wir, ausgehend von einer Kontrolltheorie, d. h. den allgemeinen Evolutionsprinzipien, über eine operationale Theorie zu biologischen 5 aktuell Winfried Henke Klassifikation der zu ordnenden Organismen (oder Objekte) voraus, d. h. die genaue Analyse ihres Erscheinungsbildes. Diese fußt auf Merkmalen, also auf gesondert erfassbaren, abgrenzbaren Eigentümlichkeiten oder Eigenschaften, die ihren Träger kennzeichnen und ihn zu beschreiben erlauben. Eine fundamentale Methode biologischer Forschung ist der Vergleich anhand deskriptiver oder messbarer morphologischer, anatomischer, physiologischer, serologischer, molekularbiologischer oder ethologischer Eigenschaften oder „Merk-Male“, die man 1. bemerkt, 2. sich merkt und auf die man 3. auch andere aufmerksam macht – mit anderen Worten: eine Einheit, die man beobachtet, festlegt und mitteilt (WERNER 1970). Erst die Kenntnis der Verwandtschaftsverhältnisse zwischen Organismen bzw. systematischen Gruppen ermöglicht die Konstruktion von Stammbäumen bzw. phylogenetischen Verwandtschaftsdiagrammen (ROTHE UND HENKE 2006; WIESEMÜLLER ET AL. 2003). Da die unüberschaubare Mannigfaltigkeit der pflanzlichen und tierischen Organismen in der Phylogenese über die fortlaufende Spaltung von Arten entstand, ist die Vielfalt der Organismen Ausgangspunkt der Systematik. Grundeinheit der Evolution ist die Art. Die kontroverse Diskussion über Artbegriff und Evolutionsvorstellungen kann hier nicht vertieft werden, ebenso wenig wie Ansätze der unterschiedlichen Schulen biologischer Klassifikation (WÄGELE 2000; ROTHE UND HENKE 2001, 2006; WIESEMÜLLER ET AL. 2003). Sie sind aber essentiell, um die Vielfalt der hypothetisierten Speziationsprozesse während der Hominisation zu verstehen, denn erst auf dieser Ebene werden die gravierenden Diskrepanzen in den Stammbaummodellen transparent und nachvollziehbar (HARDT UND HENKE 2007). Lästerzungen behaupten, Stammbäume seien wie Blumensträuße – schön anzusehen, aber schnell verwelkt. Recht haben sie – und das wusste schon Darwin (1871, dt. Übersetzung 1982, S. 262): „Falsche Tatsachen sind äußerst schädlich für die Wissenschaft, denn sie erhalten sich oft lange; falsche Theorien dagegen, die einigermaßen durch Beweise gestützt werden, tun keinen Schaden, denn jedermann bestrebt sich mit löblichem Eifer ihre Unrichtigkeit zu beweisen. Und wenn die Arbeit getan ist, so ist der Weg zum Irrtum gesperrt, und der Weg zur Wahrheit ist oft in demselben Moment eröffnet.“ Der Ansatz ist wegweisend, aber es bedarf unter Wissenschaftlern keines Hinweises, dass das Wort Wahrheit als philosophischer Begriff in den Registern der Werke aller Philosophen viel Raum einnimmt (vgl. hierzu auch CHARGAFF 2002). Erkrankungsmuster fossiler Homininen mittels molekularbiologischer, biochemischer und biophysikalischer Verfahren präzise zu rekonstruieren (KRINGS ET AL. 1997; TEMPLETON 2006; KRAUSE ET AL. 2007). Die Konkurrenz, die die molekularbiologischen High-Tech-Laboratorien den bisweilen etwas verstaubt anmutenden osteologischen Laboren machen, ist begrüßenswert und stimulierend – denn Konkurrenz belebt das Geschäft. Die Leistungsfähigkeit „klassischer“ Disziplinen wie die Morphologie – man denke nur an moderne bildgebende Verfahren, z. B. RasterelektronenMikroskopie (REM), 3D-Computertomographie (CT), Magnetresonanztomographie (MRT), Röntgen-Kinematographie, Virtuell Imaging (VI) (ULHAAS ET AL. 2005; ZOLLIKOFER UND PONCE DE LEON 2005; ULHAAS 2007), – aber auch die Systematik mit ihren komplexen verwandtschaftsanalytischen Methoden (Phylogenetische Systematik, Evolutionäre Taxonomie, Numerische Taxonomie) sollte man jedoch in ihrer Bedeutung nicht unterschätzen (WIESEMÜLLER ET AL. 2003; HARDT ET AL. 2006; HENKE UND TATTERSALL 2007; ROTHE UND HENKE 2001a). Ferner eröffnet nur der Vergleich mit rezenten Primaten, die den Vorzug der allseitigen Erforschbarkeit haben, Möglichkeiten, die stammesgeschichtlichen Adaptationen des Menschen adäquat zu interpretieren. Ethologische Freiland- und Laborforschungen sind essentiell, um die Wechselbeziehungen zwischen den Komponenten der Organismen und des Lebensraumes und damit letztlich auch die hypothetische ökologische Nische der frühen Homininen zu rekonstruieren (BOESCH UND BOESCH-ACHERMANN 2000; KAPPELER 2005; MEDER 2007). Um ein stammesgeschichtliches Szenario zu erstellen und evolutionsökologisch relevante Aussagen über Vorläufer unserer Spezies zu treffen, gilt es, diese zunächst überhaupt erst einmal aufgrund ihrer morphologischen Eigenheiten zu identifizieren. Damit erlangt die älteste Disziplin der Biologie, die Systematik, die in den 1960er Jahren schon fast abgeschrieben schien, wieder erhebliche Bedeutung (WÄGELE 2000; WIESEMÜLLER ET AL. 2003). 2.2 Zur Rekonstruktion phylogenetischer Beziehungen Systematik ist zunächst ein Mittel zur „Beherrschung der Mannigfaltigkeit der Organismen“ (aber auch unbelebter Dinge, wie z. B. Mineralien) oder – bezogen auf die Biologie – die Theorie und Praxis in der Aufdeckung und Wiedergabe der Ordnung der lebenden Natur. Systematik setzt aktuell 6 Paläoanthropologie – Standortbestimmung einer innovativen Disziplin Die phylogenetische Rekonstruktion nimmt auf die definierten (jedoch in der Paläoanthropologie stark umstrittenen) Arten und deren Merkmalsausprägungen Bezug und folgt einer Serie logischer Schritte; einige sind in der Abb. 3 festgehalten. Zunächst werden Morphoklines, d. h. Gradienten der Merkmalsentwicklung, ermittelt. Dann wird die Richtung des Gestaltwandels, die Polarität, bestimmt. Dabei wird davon ausgegangen, dass sich die Ausprägung verschiedener Merkmale von einem ursprünglichen Merkmalsstatus über Zwischenstadien zu abgeleiteten Ausprägungen entwickelt. Alle Artbildungsmodelle setzen für die Entstehung neuer Arten die Entwicklung evolutiver Neuheiten – abgeleitete Merkmale oder Apomorphien – voraus, weshalb die Stammbaumrekonstruktion allein auf diesen – im Gegensatz zu den ursprünglichen Merkmalen, den Plesiomorphien – aufbaut. In einem weiteren Schritt erfolgt die Konstruktion eines Kladogramms, d. h. einer Baumgrafik, wobei am einfachsten von einem polarisierten Morphokline ausgegangen wird und jene Arten, die ein oder mehrere Merkmale gemeinsam haben, auf demselben Ast des Kladogramms angeordnet werden. Damit wird angedeutet, dass sie miteinander näher verwandt sind als mit Spezies, mit denen sie keine Neuerwerbungen teilen. Dass die Dinge in praxi weit komplexer sind, zeigt sich immer dann, wenn sich mehrere Kladogramme ergeben, d. h. widersprüchliche Lösungen, wobei dann nach dem sog. Sparsamkeits- oder Parsimonieprinzip die einfachste Lösung als die wahrscheinlichste angenommen wird. Das „Ockham’s razzor“ nur ein methodischer Kompromiss in Ermangelung besser geeigneter Vorgehensweisen ist, versteht sich von selbst. Der nächste Schritt ist die Konstruktion des Stammbaumes. Kladogramme sind keine Stammbäume! Um einen Stammbaum zu erhalten, bedarf es weiterer Analyseschritte. Das dabei anzuwendende Prinzip basiert auf der maximalen Nutzung von Informationen, die nicht aus dem Kladogramm selbst abzulesen sind. Dazu gehören Angaben zur Stratigraphie, Chronologie und geographischen Verbreitung eines Taxon. Diejenige Stammbaum-Hypothese, die die meisten Informationen kompatibel miteinander vereint, hat den höchsten Wahrscheinlichkeitsgrad. Aufgrund neuer Informationen „wackelt“ ein Stammbaum nicht selten schon nach kurzer Zeit – das ist im Rahmen der Erwartung. Abschließend erfolgt die Erstellung eines Szenarios. Die Zeichnung eines umfassenden Lebensbildes unter Berücksichtigung des „life-history“Konzepts (HEMMER 2007) erfordert die konzertierte Aktion aller paläoanthropologisch relevanten Disziplinen: So wird uns nur der Vergleich mit heute lebenden Primaten zuverlässige Modelle über die Fortbewegungsweise unserer Vorfahren ermöglichen; nur der ethnoarchäologische Vergleich mit rezenten Wildbeutern kann uns hinreichende Informationen zur Rekonstruktion früherer Lebensweisen liefern; nur mittels der Methoden der Taphonomie – im Deutschen häufig etwas unzutreffend als „Begräbniswissenschaft“ bezeichnet – werden wir die biostratinomischen Prozesse der Fossilisation und Fossildiagenese nachvollziehen können; d. h. mit elaborierten Verfahren wird der Übergang eines Organismus von der Biosphäre in die Lithosphäre, also in die Versteinerung, rekonstruiert. Nur die Paläogeologie und -geographie Abb. 3 Von der Merkmalsanalyse zum Szenario; Schritte der phylogenetischen Rekonstruktion: a Bestimmung eines Morphokline, b Konstruktion eines Kladogramms, c Konstruktion eines Stammbaums, d Erstellung eines Szenarios (nach HARDT et al. 2006, aus HENKE u. ROTHE 1994, modifiziert). 7 aktuell Winfried Henke Das Alter eines Fossils, das mit relativen (z.B. Biostratigraphie; Fluormethode) und absoluten Datierungsverfahren (radiometrische und nichtradiometrische Methoden; vgl. WAGNER 2007a, b) bestimmt werden kann, ist eine zusätzliche Information gegenüber einem rezenten Fund; es erlaubt die Korrektur der relativen Lage der Verzweigungen von Stammbäumen (RIEDL 1975). Molekularbiologische Verfahren haben seit den 1960er Jahren (GOODMAN 1962, 1963; SARICH 1971; JOBLING ET AL. 2004) unsere chronologischen Vorstellungen stark verändert (man denke nur an die late divergence- versus early divergence-Kontroverse um die Abzweigung der zum Menschen führenden Stammlinie; vgl. HENKE und ROTHE 1994). Nach heutigen Befunden der Molekularen Uhr wird das Alter des letzten gemeinsamen Vorfahren von Menschenaffen und Homininen auf 5,5 bis 8 Millionen Jahre geschätzt, erfolgte gegenüber alternativen Vorstellungen (RamapithecusModell) also erst relativ spät. Die bislang ältesten Fossilien, die – wenn auch höchst umstritten – der Homininen-Linie zugeordnet werden, haben ein Alter von ca. 6 Mill. Jahren und stammen aus Afrika, wo übrigens schon Darwin aufgrund der verwandtschaftlichen Nähe des Menschen mit den Gattungen Pan und Gorilla den Ursprung vermutete. Die schon viele Dezennien währende Suche nach dem ‚Urahnen’ der Stammlinien der Hominini und Panini/Gorillini und die zahlreichen Propagierungen der ältesten Vertreter der zum Menschen führenden Linie, angefangen 1924 beim Taung-Kind (Australopithecus africanus) über „Lucy“ (Australopithecus afarensis) sowie Australopithecus garhi und Australopithecus anamensis bis hin zu Ardipithecus ramidus kadabba, Orrorin tugenensis oder dem höchst umstrittenen Sahelanthropus tchadensis (vgl. Abb. 4) demonstrieren einerseits, wie lückenhaft der Fossilreport ist, und andererseits, wie uneinheitlich das verfügbare Methodeninventar zur Kennzeichnung fossiler Taxa und zur Rekonstruktion phylogenetischer Prozesse appliziert wird (VRBA 2007). Auch hinsichtlich der molekularbiologisch ermittelten zeitlichen Tiefe unseres Eigenwegs besteht offenbar aufgrund neuerer paläontologischer Daten Unsicherheit, wie die mit den molekulargenetischen Gabelungshypothesen nicht im Einklang stehende Interpretation des jüngst entdeckten miozänen Gorilla-Verwandten Chororapithecus abyssinicus (SUWA ET AL. 2007) zeigt. Während die Paläoanthropologen den meisten Australopithecus-Taxa heute nur noch eine marginale Rolle im Rahmen der Hominisation zuschreiben, d.h. sie als aufrecht gehende Men- können die in vor- und frühgeschichtlicher Zeit existierenden Migrationswege der Homininen in Zusammenarbeit mit der Paläontologie und Zoogeographie aufzeigen, und in jüngerer Zeit schicken sich auch Molekularbiologen an, diese Aufgabe tatkräftig mit populationsgenetischen Vergleichsanalysen zu unterstützen (RICHARDS 2002; JOBLING ET AL. 2004; BURGER 2007). Nur in der Paläopathologie und Epidemiologie Geschulte werden in der Lage sein, über die Krankheitsbelastungen unserer stammesgeschichtlichen Vorfahren fundiert zu urteilen und erste Hinweise auf soziale und medizinische Hilfe sowie auf Seuchen und ihre selektiven Wirkungen finden; nur mittels ausgefeilter Verfahren der Archäometrie, d. h. mikro- und makromorphologischer sowie biophysikalischer und -chemischer Ansätze, werden wir die Fossilien differentialdiagnostisch richtig deuten und Fehlurteile minimieren können. Noch viele weitere Disziplinen wären hier zu kennzeichnen (vgl. Abb. 2), eines sollte aber bereits deutlich geworden sein: Nur durch das Zusammenwirken unterschiedlicher Disziplinen der Feld- und Laborforschung werden wir der Herausforderung der Evolutionsbiologie gewachsen sein, den einmalig abgelaufenen, realhistorisch-genetischen Prozess der Menschwerdung angemessen, d. h. nachhaltig und dauerhaft, zu rekonstruieren. 2.3 Raumzeitliche Verortung homininer Fossilien Die Verortung des Hominisationsprozesses in Raum und Zeit kann auf zwei Wegen erfolgen, einerseits durch die vergleichende Analyse lebender Primaten, andererseits durch die Analyse und Interpretation von Hominidenfossilien. Da Primaten in den ihnen angestammten Lebensräumen relativ selten fossilisierten, gehören Sie zu den seltenen paläontologischen Funden. Die Wahrscheinlichkeit für den Übergang von der Biosphäre in die Lithosphäre hängt von der Sequenz biologischer, chemischer und physikalischer Transformationsbzw. Dekompositionsvorgänge ab, die Gegenstand der Taphonomie sind (SHIPMAN 1981; ROTHE und HENKE 2001b; HARDT ET AL. 2007). Die systematische Dekodierung umfasst alle postmortalen biostratinomischen und fossildiagenetischen Veränderungen des Individuums von seiner definitiven Einbettung bis zur Entdeckung. Die Paläoökologie analysiert all jene Prozesse, die einen Organismus während seiner Lebensspanne beeinflusst haben; sie rekonstruiert die ökologische Nische, wobei die Taphonomie als integraler Bestandteil der Paläoökologie verstanden wird (PIANKA 1988; ETTER 1994; NENTWIG 2007a, b). aktuell 8 Paläoanthropologie – Standortbestimmung einer innovativen Disziplin Abb. 4 SplitterStammbaummodell aus HENKE und ROTHE (2003), modifiziert (vgl. HARDT ET AL. 2006). schenaffen betrachten (z.B. TATTERSALL 2005), schließen andere diese und selbst die Habilinen (Homo habilis, Homo rudolfensis) mit hoher Wahrscheinlichkeit (WOOD 1996, 2006; WOOD und COLLARD 1999a, b, 2001; WOOD und CONSTANTINO 2006; COLLARD und WOOD 2007) oder gänzlich aus unserer Vorfahrenschaft aus (vgl. SENUT 2007). Dabei hatte Tobias (1989a, b) so zahlreiche und z.T. auch sehr glaubwürdige Argumente für den Homininenstatus von Homo habilis geliefert, bis hin zur Annahme „that he could say and do“. Aber die credibility von Homo habilis bröckelte schon seit Langem (STRINGER 1986) und auch der Eigenweg innerhalb des Genus Homo wird sehr kontrovers diskutiert (Übersicht in HARDT und HENKE 2007). Eine nähere Betrachtung der den Analysen zugrunde liegenden Verfahren zeigt, dass sehr häufig unterschiedliche Artbegriffe (Biospezies, evolutionäre Spezies, Morphospezies, Chronos- pezies) verwendet werden (vgl. u.a. HENNIG 1950, 1966; MAYR 1975, 1991; TATTERSALL 1986; MCHENRY 1996; WOLPOFF und CASPARI 1997; WHEELER und MEIER 2000; FUTUYAMA 2007), die voneinander abweichende kladistische und gradualistische Modelle prägen (Methodenübersicht in WÄGELE 2000; WIESEMÜLLER ET AL. 2003; ROTHE und HENKE 2001a, 2006). Während die archäometrischen Datierungverfahren bei hinreichenden materiellen Voraussetzungen eine präzise zeitliche Zuordnung von Fossilien erlauben, bieten die phylogenetischen Rekonstruktionsverfahren eine erhebliche Breitseite für kritische Einwände. Dabei handelt es sich keineswegs nur um semantische Probleme, wie manche meinen, sondern um massive Schwierigkeiten, die „Ordnung des Lebendigen“ aufgrund morphologischer und selbst molekularbiologischer Merkmalsmuster (z.B. mtDNA) zu rekonstruieren. 9 aktuell Winfried Henke Die Merkmalsanalyse von Fossilien erfasst sog. Form-Funktion-Komplexe (Fakultäten) und deren Beziehungen untereinander, aus denen jedoch nur hypothetisch auf die biologische Rolle geschlossen werden kann. In der praktischen Arbeit ergeben sich erhebliche Schwierigkeiten, die Anpassungen von Merkmalen zweifelsfrei nachzuweisen. Hierzu ist immer der Abgleich mit lebenden Organismen notwendig (Abb. 5), da über die Adaptationen (Prozesse und Zustände) nur Hypothesen formuliert werden können. Einige Beispiele: So lässt sich aus dem auffällig tiefen Handwurzelkanal von Australopithecus anamensis auf eine extrem starke Greiffunktion schließen. Da diese pliozäne Spezies bereits habituell biped 2.4 Funktions- und evolutionsmorphologische Kennzeichnung fossiler Hominini Ziel paläoanthropologischer Forschung ist es, die Hominisation als evolutive Anpassung großwüchsiger, terrestrisch lebender, omnivorer Primaten an die ökologischen Herausforderungen saisonaler tropischer Landschaften im trans-, inter- und intraspezifischen Wettbewerb zu verstehen. Nach den fossilen Quellen zeichnet sich eine Mosaikevolution ab: zunächst erfolgte – offenbar mehrfach – die Entwicklung der habituellen Bipedie, dann die Umgestaltung des Kauapparates auf Omnivorie und schließlich die exzessive Hirnentfaltung (Cerebralisation). Abb. 5 Vergleichendmorphologischer Ansatz der Paläoanthropologie, illustriert am Beispiel der Analyse der Kranialmorphologie fossiler und rezenter Homininen (nach HARDT und HENKE 2007, modifiziert). Homo ergaster aktuell 10 Paläoanthropologie – Standortbestimmung einer innovativen Disziplin war, ist zu vermuten, dass anamensis noch teilweise arborikol lebte. – Verhaltensfossilien wie die Fußspuren in der vulkanischen Asche von Laetoli lassen Rückschlüsse auf Schrittlänge und Körpergewicht von Australopithecus zu. – Computertomographisch gemessene Kompaktadicken der Extremitätenknochen bei Neandertalern belegen deren enorme körperliche Stärke. – Außer der Makromorphologie (Kauflächen, Höckerstruktur) geben z.B. mikromorphologische Befunde an Zähnen (z.B. Abkauungsspuren, Zahnschmelzdicken) Aufschluss über Ernährungsweisen (HENKE und ROTHE 1997; ULHAAS ET AL. 2004; ULHAAS 2007) und Habitatpräferenzen. – Sexualdimorphismen erlauben Rückschlüsse auf Verhaltensmuster der Geschlechter und Vergesellschaftungsformen früher Hominini. – Ausgeheilte Pathologien und intentionale Bestattungen lassen auf entwickelte Kooperationsstrukturen und soziale Kompetenz schließen. – 3D-analytische Verfahren und das Arsenal bildgebender Diagnostik (ZOLLIKOFER und PONCE DE LEON 2005; ULHAAS 2007) eröffnen die Möglichkeit, wie Preziosen gehütete, rare Fossilien durch digitale Modelle einem breiteren Untersucherkreis zugänglich zu machen. Aufgrund der damit gegebenen Kontrolle sollte das kein Desiderat bleiben! Ansätze wie das Verbundsystem TNT (The Neanderthal Tools) inkl. NESPOS - Neanderthal Studies Professional Online Service sind aussichtsreiche Perspektiven, um zu einer intersubjektiven Beurteilung des Fossilreports zu gelangen (http://www.the-neanderthal-tools.org/ ?page_id=13). In jüngerer Zeit gewinnt in der Paläoanthropologie die integrierte Betrachtung von ‚evolution and development“ (Abk. Evo-Devo) verstärkt Beachtung. So genannte Life-history-Analysen können potenzielle Ursachen für Fitnessunterschiede detektieren. Die Erfassung ontogenetischer Variablen (z.B. Größe bei Geburt, Wachstumsverlauf und -geschwindigkeit, Alter und Größe bei Fortpflanzung, Sexualproportionen, Mortalitätsmuster) ermöglicht die Ermittlung artspezifischer Parameter und diachroner Trends. In diesem Kontext stellt sich die Frage, ob z.B. die Reifung von Homo erectus in ähnlichem Tempo oder deutlich akzelerierter als bei Homo sapiens verlief (MINUGHPURVIS und MCNAMARA 2002). Die Synopsis aller paläoanthropologischen Befunde führt schließlich zur Rekonstruktion der ökologischen Nische früher Hominini, zu der alle paläobiologischen Disziplinen mit ihren Analysen, z.B. Faunen- und Florenzusammensetzung, Landschafts- und Klimarekonstruktion (u. a. Paläosole; ALVERSON 2007; RETALLACK 2007), Nahrungsres- sourcen (Abkauungsmuster, Dentolithen, stabile Isotope; RICHARDS ET AL. 2000; GRUPE 2007; HARDT ET AL. 2007; SPONHEIMER und LEE-THORP 2007), Vergesellschaftungsformen (Sexualdimorphismus; SUSSMAN und HART 2007) beizutragen vermögen, jedoch sollte man stets L. P. Hartleys Warnung eingedenk sein: „… the past is a foreign country, they do things differently there“ (zit. nach FOLEY 1987, S. 78). 3. Stammbäume mit baldigem Verfallsdatum - Jeweilsbilder 3.1. „Zuwachs an Kenntnis ist Zuwachs an Unruhe“ (J. W. Goethe) Aufgrund intensiver Feld- und Laborforschung wurde das Jeweilsbild unseres phylogenetischen Eigenwegs stetig modifiziert. Das gilt insbesondere für die Anzahl der Speziationsereignisse. Während Splitter über 20 hominine Arten annehmen, gehen extreme Lumper von weitaus weniger Spezies aus, ja einige schließen Artspaltungen in der pleistozänen Homo-Linie sogar aus (Übersicht in WOLPOFF 1996-1997, 1999; HENKE 2003a, b, 2005, 2006; HENKE und ROTHE 1999a, b, 2005). Das Splitter-Stammbaummodell (Abb. 4; TATTERSALL 2006) macht deutlich, daß Sahelanthropus tchadensis (BRUNET ET AL. 2002) eng an der Basis der Homininen-Linie steht; aufgrund des Merkmalsmosaiks ist die Einbeziehung in unsere Stammlinie fraglich, denn nach WOLPOFF ET AL. (2002) liegen wenig überzeugende Befunde vor. Die Klassifikation von Orrorin tugenensis (SENUT ET AL., 2001) an der Homininenbasis „dehumanisiert“ dagegen die Australopithecus-Spezies. Selbst die Glaubwürdigkeit des generischen Status von Homo habilis (L.S.B. LEAKEY ET AL., 1964) und Homo rudolfensis (ALEXEEV 1986; WOOD, 1992) steht in Frage (WOOD und COLLARD 1999a, b), so dass H. ergaster (GROVES und MAZÁK, 1975) als derzeit ältester, anerkannter Vertreter unserer Gattung gilt. Dass mit dem stark gewachsenen Umfang der Hypodigmata die paläoanthropologischen Probleme nicht kleiner werden, belegt das Neandertaler-Problem. Weder morphologisch noch aufgrund der mtDNA-Befunde ist endgültig geklärt, ob die Neandertaler eine eigene Art waren – und dass sie „eigenartig“ waren, ist hinlänglich bekannt (HENKE und ROTHE 1994, 1999b TATTERSALL 1995a, b, 1999, 2007; HENKE ET AL. 1996; RELETHFORD 2001; HENKE 2005; ZILHÃO 2006). Molekulargenetische Befunde haben seit den 1960er Jahren die paläoanthropologische Modellbildung erheblich beeinflusst (s.o.), aber erst mit den mtDNA-Analysen an rezenten Populatio- 11 aktuell Winfried Henke nen (CANN ET AL. 1987; STONEKING ET AL. 1989) und schließlich mit der erfolgreichen Beprobung der Fossilien aus der Kleinen Feldhofer Grotte (KRINGS ET AL. 1997) und weiterer Neandertaler sowie jungpaläolithischer Skelette (u.a. OVCHINNIKOV ET AL. 2000; SERRE ET AL. 2004) gelang mit der Analyse alter DNA ein entscheidender methodischer Durchbruch (RELETHFORD 1999, 2001a, b; JOBLING ET AL. 2004; TEMPLETON 2002, 2006). Die ‚klassische’ CellArtikel von 1997 mit dem Titel „Neandertals were not our ancestors“ belegt jedoch nur die Divergenz der mt-DNA-Linien, schließt entgegen der vermeintlich klaren Aussage des zitierten Covertitels die Neandertaler nicht gänzlich aus unserer Vorfahrenschaft aus (zur Diskussion siehe RELETHFORD 2001a; HENKE 2005). Aufgrund der Degradation und Dekomposition alter DNA (BURGER ET AL. 1997; BURGER 2007) blieben weitere spektakuläre Befunde an noch älteren Homo-Fossilien bislang aus, aber man darf gespannt sein, ob zukünftig Befunde an der heidelbergensis- und erectus-Gruppe vorgelegt werden. Die Wahrscheinlichkeit dafür ist jedoch minimal. Gegenwärtig ist das Neandertaler-Genom-Projekt mit ersten Befunden auf dem ‚Wissenschaftsmarkt’ und liefert aufgrund des Nachweises der Identität des FoxP2-Gens beim anatomisch-modernen Menschen und beim Neandertaler die ersten als spektakuläre paläoanthropologische Erkenntnisse publizierten Befunde (KRAUSE ET AL. 2007). Die Erwartung, dass genomische Analysen nukleärer DNA möglicherweise eine mangelnde Sprachfähigkeit der Neandertaler auf der Basis plesiomorpher FoxP2-Varianten bescheinigen würden (ENARD 2005), konnten durch jüngere Befunde nicht gestützt werden, denn die abgeleitete FoxP2-Variante des modernen Menschen besaßen auch schon die Neandertaler (KRAUSE ET AL. 2007) – aber wer glaubte eigentlich daran, dass Neandertaler noch nicht einer Sprache mächtig waren, wenn schon der Homo heidelbergensis, den einige als archaischen Homo sapiens, andere als Ante-Neandertaler betrachten, schon aufwändige Holzspeere für die Distanzjagd fertigte (THIEME 2005). Zur jüngeren Diskussion kognitiver Leistungen der Gattung Homo siehe auch CONARD (2006a, b, 2007) und HAIDLE (2006, 2007). POFF ET AL. 1994a, b; WOLPOFF und CASPARI 2000) gehen davon aus, dass sich die Gattung Homo vor knapp 2 Mill. Jahren in Afrika entwickelte und dann sukzessive den asiatischen und europäischen Kontinent besiedelte. Ende des letzten Jahrhunderts entdeckte Fossilien aus Dmanisi (Georgien) belegten erstmals unzweifelhaft eine sehr frühe Auswanderung in die nördlichen Breiten Westasiens und deutlich später auch nach Europa (BRÄUER ET AL. 1995; ULLRICH 1998; SCHWARTZ und TATTERSALL 2002; VEKUA ET AL. 2002; TATTERSALL und SCHWARTZ 2003; RIGHTMIRE ET AL. 2005). Damit stellte sich die Frage nach der Anzahl der Speziationsprozesse innerhalb der Gattung Homo neu (FOLEY 1991; TATTERSALL 2000b; TATTERSALL und SCHWARTZ 2000). Die Antworten fielen sehr vielfältig aus. Die einfachste Lösung bieten nach wie vor die „Multiregionalisten“ und begründen das wie folgt: Da die Mehrzahl der Homo ergaster/ erectus-Kennzeichen auch H. sapiens teilt und die wenigen Merkmalsmuster die H. sapiens nicht mit H. ergaster/erectus gemeinsam hat, als Reaktionen auf Entwicklungstendenzen in Richtung auf eine zunehmende Cerebralisation (Hirnentwicklung) und kulturelle Vielfalt, d.h. fortschreitende tradigenetische Entwicklung, zu interpretieren ist, lässt sich dem ‚multiregionalen Entwicklungsmodell’ zufolge nur ein einziger Speziationsprozess annehmen. H. ergaster/erectus geht in der Art H. sapiens auf, da es keine deutliche Grenze, weder räumlich noch zeitlich, zwischen beiden Spezies gibt. Ständiger Genfluß und Migrationen verhinderten Speziationen, so dass die offensichtlichen zeitlichen und regionalen Bevölkerungsunterschiede der polytypischen Art H. sapiens anagenetisch als intraspezifische Adaptationen erklärt werden (vgl. auch Abb. 6a). H. heidelbergensis wäre in diesem extremen gradualistischen Modell nur eine Chrono- oder Morphospezies, eine mittelpleistozäne Entwicklungsstufe, eine regionale Variante oder Subspezies des H. sapiens (vgl. WOLPOFF 1999). 3.2.2 Out-of-Africa-Modell mit Hybridisierung Aufgrund der nachweislich graduellen Veränderungen in der europäischen Linie vom frühen Präneandertaler zum klassische Neandertaler (vgl. Abb. 6b) - sowie der von BRÄUER (1984, 1994, 2006, 2007) als Protagonist der out-of-Africa-Hypothese immer wieder an neuen Fossilien mit neuen vergleichend-morphologischen Befunden nachgewiesenen stufenweisen Modernisierung innerhalb der afrikanischen Linie (grades vgl. Abb. 7) und eventuell auch spät-archaischer Gruppen in Chi- 3.2 Die Phylogenie des Genus Homo – ein ewiges Puzzle? 3.2.1 Multiregionales Evolutionsmodell Extreme ‚Lumper’ (THORNE und WOLPOFF 1981, 1992; FRAYER ET AL. 1993; WOLPOFF 1992, 1999; WOL- aktuell 12 Paläoanthropologie – Standortbestimmung einer innovativen Disziplin Abb. 6 Alternative Evolutions- und Migrationsszenarien: a Multiregionales Evolutionsmodell (n. WOLPOFF 1999, modifiziert, vgl. HARDT und HENKE 2007) b Out-of-Africa-Modell mit Hybridisierung nach BRÄUER (2006) c Splitter-Modell mit einer afrikanischen Linie zum H. sapiens über H. rhodesiensis und einer europäischen Linie über H. heidelbergensis zum H. neanderthalensis (n. STRINGER 2003) d „gemäßigtes“ Splitter-Modell mit H. heidelbergensis als afro-europäischer Vorläufer der afrikanischen und europäischen Linien (RIGHTMIRE 1998). H. sapiens und der europäischen und nahöstlichen Neandertaler-Populationen vertreten hat, ist die taxonomische Kennzeichnung der letzten als H. sapiens neanderthalensis nur konsequent, wenn auch die deutliche Annäherung an die Multiregionalisten gerade im Hinblick auf die seitens der Archäometrie und Molekulargenetik eingebrachten Argumente etwas irritiert, denn ein Hybrisierungsnachweis konnte bislang nicht glaubwürdig erbracht werden. Da aber nach Auffassung vieler Paläoanthropologen die mtDNA-Befunde hinsichtlich ihrer taxonomischen Aussagekraft weit überschätzt wurden (pro: KRINGS ET AL. 1997; PÄÄBO ET AL. 2004; SERRE ET AL. 2004; contra: RELETHFORD 2001a, b) stimmt Bräuers Modell sehr gut mit dem na (Dali, Dingcun, Maba; vgl. BRÄUER 2006, 2007; BRÄUER und SMITH 1992) erscheint die Annahme gut begründet, für die letzten 700 000 Jahre nur eine einzige Speziation zwischen H. erectus und H. sapiens anzunehmen. In diesem Modell (vgl. Abb. 6b) würde das Taxon H. heidelbergensis gänzlich “geschluckt” werden, denn innerhalb der polytypischen Spezies H. sapiens käme ihm kein Status einer Biospezies oder evolutionären Spezies zu. Als mittelpleistozäner „früh-archaischer“ Typus nimmt er nur eine marginale Übergangsrolle in der gradualistischen Entwicklung an. Da BRÄUER (1984) stets die Hypothese einer potenziellen Hybridisierung der sich graduell in Afrika entwickelnden Stammlinie zum anatomisch-modernen 13 aktuell Winfried Henke Abb. 7 Wichtige afrikanische Funde in ihrer zeitlichen Abfolge und Zuordnung zu bestimmten Taxa bzw. den graduellen Stadien des „archaischen“ H. sapiens nach BRÄUER (1984, 2006). von dem Molekulargenetiker RELETHFORD (2001b) propagierten mostly-out of Africa-Modell überein (vgl. auch SMITH 2002). hatte. Während sich der europäische Zweig sukzessive zum Neandertaler entwickelte (AccretionModell; DEAN ET AL. 1998 HUBLIN 1998; HARVATI 2007; contra: HAWKS und WOLPOFF 2001), bildete sich in Afrika – ebenfalls gradualistisch – der H. sapiens heraus. Dieser gelangte im Spätpleistozän via Naher Osten nach Europa und löste die neanderthaliden Populationen ab. In diesem Modell nimmt H. heidelbergensis eine zentrale Rolle als anzestrale afrikanisch-europäische Stammart der Neandertaler und des modernen Menschen ein (vgl. auch Abb. 8). 3.2.3 Gemäßigte und extreme Splitter-Modelle In dem von RIGHTMIRE (1998, 2001a, b, 2007) vertretenen ‚gemäßigten’ Splitter-Modell (Abb. 6c) entstand H. heidelbergensis durch Speziation aus H. ergaster/erectus in Afrika und wanderte nach Europa und eventuell auch in den Fernen Osten aus, den H. erectus schon seit langem besiedelt aktuell 14 Paläoanthropologie – Standortbestimmung einer innovativen Disziplin Abb. 8 Kartierung der wichtigsten europäischen Fossilfunde in ihrer zeitlichen Abfolge und Zuordnung zu bestimmten Taxa bzw. Stufen (grades) des Accretion-Modells nach DEAN ET AL. (1998) und BRÄUER (1998), aus HARDT und HENKE (2007). nach kladistischen Prinzipien äußerst problematisch, da es keine Artspaltung, sondern sukzessive Artbildungen in Europa und Afrika annimmt (STRINGER 1982, 1986, 1992, 1996; 2001, 2002; STRINGER und GAMBLE 1993; ARSUAGA ET AL. 1997). Schlösse man diese aus und nähme für die europäische Entwicklungslinie einen anagenetischen Prozess an, so wäre es nur konsequent, den europäischen H. heidelbergensis als H. neanderthalensis zu führen Entsprechend wäre auch in Afrika H. rhodesiensis nicht zu halten, sondern in das Taxon H. sapiens einzubeziehen. Ein extremes Splitter-Modell (vgl. Abb. 6d) geht von weiteren Speziationen in Afrika aus. Danach soll sich die bislang nur in Spanien nachgewiesene, umstrittene Spezies H. antecessor schon in Afrika vom H. ergaster abgespalten haben und sehr bald nach Europa migriert sein. Diese hypothetische Art wird als Stammform von zwei Entwicklungszweigen angenommen: einerseits dem H. heidelbergensis, der in Afrika evolvierte und kurz darauf nach Europa gelangte, wo durch einen weiteren Speziationsprozess H. neanderthalensis entstand, und andererseits dem H. rhodesiensis, der eine ausschließlich afrikanische Spezies war und aus dem H. sapiens speziiert sein soll, der dann als anatomisch-moderner Mensch alle Kontinente besiedelte und die archaischen Populationen der Alten Welt ablöste. Das von STRINGER (2002) kürzlich vorgestellte Speziationsmodell ist 3.3. Resümee Wie der wissenschaftsgeschichtliche Abriss zur Taxonomie und Systematik der Gattung Homo (vgl. auch HENKE 2005, 2006a; HARDT und HENKE 15 aktuell Winfried Henke rer Analyse bereitsteht, sollte sich die „moderne Fossilienkunde“ ihrer Möglichkeiten – aber auch ihrer Grenzen – bewusst sein und die neue Sachlichkeit und Glaubwürdigkeit ihrer Befunde nicht als fossil- und journalism-driven science gefährden (WHITE 2000). Es geht nämlich um uns und unsere Wurzeln und den Imperativ gnôthi seautón. Fossilarchive der Hominisation bieten diesbezüglich eine einzigartige Chance. Stammesgeschichtliche Beweise im engeren Sinne gibt es jedoch nicht – es bleiben stets Modelle, so dass nur die aphoristische Homo-Definition der Times Kontinuität hat: „Humans are animals who wonder intensively and endlessly about their origin“. Die Paläoanthropologie – soviel sollte hier deutlich geworden sein – fragt nicht nur nach dem Erscheinungsbild der fossilen Homininen, danach, wann und wo wir entstanden sind, sondern sucht nach Antworten auf die fundamentale Frage, wie wir wurden, was wir sind. Sie ist theoriegeleitetes Hypothesentesten im Rahmen eines Forschungsansatzes, dessen Ziel eine dynamische Darwinsche Erklärung der Menschwerdung in ihren evolutionären, geschichtlichen und ontogenetischen Dimensionen ist (vgl. VOGEL 1983; HENKE & ROTHE 1994, 2001, 2003, 2006; Übersicht in HENKE und TATTERSALL 2007). 2007) zeigt, bestehen auch heute noch sehr unterschiedliche Auffassungen bezüglich der phylogenetischen Rollen der ihr zugeschriebenen Spezies. Es ist jedoch offensichtlich, dass diese zu einem nicht unwesentlichen Anteil durch die angenommenen Prinzipien (grade- versus clade-Konzept) und methodischen Ansätze (Evolutionäre Taxonomie versus Phylogenetische Systematik) verursacht werden – oder aber bedauerlicherweise keinen erkennbaren systematischen Grundsätzen folgen, was die Modelle unnötig kompliziert. Ingesamt zeichnet sich aber nach den harten Diskussionen der out-of-Africa-Protagonisten versus Multiregionalisten der beiden vergangenen Dezennien eine unerwartete Annäherung ab, bei der H. heidelbergensis als grade im europäischen und/oder afrikanischen H. sapiens aufgeht – ein Modell, dem ‚Splitter’ nach wie vor heftig widersprechen. Dass der Fossilreport immer wieder Herausforderungen bereithält, zeigt die umstrittene Interpretation von 18000 Jahre alten Fossilien aus Flores (Indonesien). Während BROWN ET AL. (2004) sie als Homo floresiensis klassifizierten, halten andere den mit 417 cm³ extrem kleinhirnigen Homininen für einen nanowüchsigen Mikrozephalen. HERSHKOVITZ ET AL. (2007) bekräftigten letztere Auffassung aufgrund der Diagnose des LaronSyndroms. Träfe das wirklich zu, würde jede Spekulation über einen langen Eigenweg der Flores-Linie seit Homo erectus oder gar Homo habilis (BROWN ET AL. 2005; FALK ET AL. 2005; MARTIN ET AL. 2007) kippen. Somit bleibt als generelles Resümee: „We don´t know where the gene flow barriers were among the sampled populations, nor do we know about unsampled populations/lineages. It is evident that new fossils continue to illuminate these evolutionary processes more successfully than poorly constructed phenetic analyses of individual specimens.“ (GILBERT ET AL. 2003, S. 259) Literatur ALEXEEV, V.P. (1986): The Origin of the Human Race. Moscow 1986. ALTNER, G. (1982, Hrsg.): Der Darwinismus. Die Geschichte einer Theorie. Darmstadt 1982. ALVERSON, K. (2007): Paleoclimate. In: HENKE, W., TATTERSALL, I. (Eds.), Handbook of Paleoanthropology, Volume I. Berlin, Heidelberg, New York 2007, 357–382. ARGUE, D., DONLON, D., GROVES, C., R. WRIGHT (2007): Homo floresiensis: Microcephalic, pygmoid, Australopithecus, or Homo? Journal of Human Evolution 51(4): 360–374. 4. Ausblick Obwohl die Paläoanthropologie in den vergangenen Dezennien ihr Theorie-Defizit weitgehend abgelegt und sich von einer vorwiegend deskriptiv-narrativen Disziplin zu einer theoriegeleiteten Wissenschaft entwickelt hat, bestehen hochgradig kontroverse Stammbaummodelle der Hominisation. Da Homininen-Fossilien aufgrund ihrer Seltenheit jeden paläontologischen und archäologischen Fundplatz „adeln“ und ein innovatives, höchst effizientes Methodeninventar zu ih- aktuell ARSUAGA, J.L., MARTÍNEZ, I., GRACIA, A., C. LORENZO (1997): The Sima de los Huesos crania (Sierra de Atapuerca, Spain). A comparative study. Journal of Human Evolution 33: 219–281. ARSUAGA, J.L., MARTÍNEZ, L.C., GRACIA, A., MUÑOZ, A., ALONSO, O., J. GALLEGO (1999): The human cranial remains from Gran Dolina Lower Pleistocene site (Sierra de Atapuerca, Spain). Journal of Human Evolution 37: 431–457. 16 Paläoanthropologie – Standortbestimmung einer innovativen Disziplin CANN, R.L., STONEKING, M., A.C. WILSON (1987): Mitochondrial DNA and human evolution. Nature 325: 31–36. BERMUDEZ DE CASTRO, J.M., ARSUAGA, J.L., CARBONELL, E., ROSAS, A., MARTÍNEZ, I., M. MOSQUERA (1997): A hominid from the Lower Pleistocene of Atapuerca, Spain. Possible ancestor to Neandertals and modern humans. Science 276, 1392–1395. CHARGAFF, E. (2002): Abscheu vor der Weltgeschichte. Fragmente vom Menschen. Stuttgart 2002. BOESCH, C., H. BOESCH-ACHERMANN (2000): The Chimpanzees of the Tai Forest: Behavioural Ecology and Evolution. Oxford 2000. COLLARD, M., B.A. WOOD (2007): Defining the Genus Homo. In: HENKE, W., TATTERSALL, I. (Eds.), Handbook of Paleoanthropology, Volume III. Berlin, Heidelberg, New York 2007, 1575–1610. BRÄUER, G. (1984): The “Afro-European sapiens hypothesis” and hominid evolution in East Asia during the late middle and upper Pleistocene. In: ANDREWS, P., J.L. FRANZEN (Eds.): The Early Evolution of Man, with Special Emphasis on Southeast Asia and Africa. Courier Forschungsinstitut Senckenberg 69, Forschungsinstitut Senckenberg, Frankfurt am Main, 145–165. CONARD, N.J. (2006a, Hrsg.): Woher kommt der Mensch? Tübingen 2006. – (2006b): Die Entstehung der modernen Kulturalität, In: CONARD, N. J. (Ed.): Woher kommt der Mensch? Tübingen 2006, 197–228. – (2006): Das Out-of-Africa-Modell und die Kontroverse um den Ursprung des modernen Menschen. In: CONARD, N.J. (Ed.), Woher kommt der Mensch? Tübingen 2006, 171–196. – (2007): Cultural Evolution in Africa and Eurasia during the Middle and Late Pleistocene. In: HENKE, W., TATTERSALL, I. (Eds.), Handbook of Paleoanthropology, Volume III. Berlin, Heidelberg, New York 2007, 2001–2038. – (2007): Origin of Modern Humans. In: HENKE, W., TATTERSALL, I. (Eds.), Handbook of Paleoanthropology, Volume III. Berlin, Heidelberg, New York 2007, 1749–1780. CORBEY, R., W. ROEBROEKS (2001, Eds.): Studying human origins: Disciplinary history and epistemology. Amsterdam 2001. BRÄUER, G., HENKE, W., M. SCHULTZ (1995): Der hominide Unterkiefer von Dmanisi: Morphologie, Pathologie und Analysen zur Klassifikation. Jahrbuch des Römisch-Germanischen Zentralmuseums Mainz 42, 143–203, Tafel 57–64. COUTURE, C., J.-J. HUBLIN (2005): Les Néandertaliens. In: DUTOUR, O., HUBLIN, J.-J. VANDERMEERSCH, B. (Eds.), Origine et Evolution des Populations Humaines. Comité des Travaux Historiques et Scientifiques, Paris 2005, 129–150. BRÄUER, G., F. SMITH (1992, Eds.). Continuity or Replacement. Controversies in Homo sapiens Evolution. A.A. Balkema, Rotterdam 1992. DARWIN, C. (1859): Origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. London 1859. BRESCH, C. (1977): Zwischenstufe Leben - Evolution ohne Ziel? München 1977. DARWIN, C., (1871): The Descent of Man and Selection in Relation to Sex. Reprinted 1981 by Princeton University Press ed. London 1871. BROWN, P., SUTIKNA, T., MORWOOD, M.J., SOEJONO, R.P., JATMIKO, WAYHU SAPTOMO, E., ROKUS AWE DUE (2004): A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia 2004. Nature 431: 1055–1061. – (1872): The expressions of emotions in man and animals. London 1872. – (1982): Abstammung des Menschen. Mit einem Vorwort von C. Vogel. Stuttgart 1982. BRUNET, M., GUY, F. PILBEAM, D., MACKAYE, H.T., LIKIUS, A., AHOUNTA, DJ, BEAUVILAIN, A., BLONDEL, C. BOCHERENS, H., BOISSERIE, J.-R., DE BONIS, L., J. COPPENS ET AL. (2002): A new hominid from the upper Micocene of Chad, central Africa. Nature 418, 145–151. DEAN, D., HUBLIN, J.-J., HOLLOWAY, R., R. ZIEGLER (1998): On the phylogenetic position of the pre-Neanderthal specimen from Reilingen, Germany. Journal of Human Evolution 17, 715–718. DENNETT, D.C. (1996): Darwin’s dangerous idea. Evolution and the meaning of life. New York 1996. BURGER, J. (2007): Palaeogenetik. In: WAGNER, G.A. (Ed.), Einführung in die Archäometrie.Berlin, Heidelberg, New York 2007, 279–301. DOBZHANSKY, TH. (1973): Nothing in biology makes sense except in the light of evolution. Am. Biol. Teach. 25, 125–129. BURGER, J., HUMMEL, S., HERRMANN, B., W. HENKE (1999): DNA preservation: A microsatellite-DNA study on ancient skeletal remains. Electrophoresis 20, 8, 1722–1728. ENARD, W. (2005): Mensch und Schimpanse – ein genetischer Vergleich. In: KLEEBERG, B., WALTER, T., 17 aktuell Winfried Henke HAIDLE, M. (2006): Menschenaffen? Affenmenschen? Menschen! Kognition und Sprache im Altpaläolithikum. In: CONARD, N.J. (Ed.), Woher kommt der Mensch? 74–103. CRIVELLARI, F. (Hrsg.), Urmensch und Wissenschaften. Eine Bestandsaufnahme. Darmstadt 2005, S. 126–135. ETTER, W. (1994): Palökologie. Basel, Boston, Berlin 1994. – (2007): Archeology. In: HENKE, W., TATTERSALL, I. (Eds.), Handbook of Paleoanthropology, Volume I. Berlin, Heidelberg, New York 2007, 261–289. EXCOFFIER, L. (2002): Human demographic history: redefining the recent African origin model. Current Opinion in Genetics & Development 12, 675–682. HARDT, T., HARDT, B., P. MENKE (2007): Paleoecology. An Adequate Window on the Past? In: HENKE, W., TATTERSALL, I. (Eds.), Handbook of Paleoanthropology, Volume I. Berlin, Heidelberg, New York 2007, 503–554. FALK, D. (2007): Evolution of the Primate Brain. In: HENKE, W., TATTERSALL, I. (Eds.), Handbook of Paleoanthropology, Volume III. Berlin, Heidelberg, New York 2007, 1133–1162. FALK, D., HILDEBOLT, C., SMITH, K., MORWOOD, M. J., SUTIKNA, T., BROWN, P., JATMIKO, SAPTOMO, E. W., BRUNSDEN, B., F. PRIOR (2005): The brain of LB1, Homo floresiensis. Science 308, 242–245. HARDT, T., W. HENKE (2007): Zur stammesgeschichtlichen Stellung des Homo heidelbergensis. In: WAGNER, G. A., RIEDER, H., ZÖLLER, L. MICK, E. (Hrsg.), homo heidelbergensis. Schlüsselfund der Menschheitsgeschichte. Stuttgart 2007, 184–202. FOLEY, R.A. (1987): Another Unique Species: Patterns in Human Evolutionary Ecology. Longman Scientific & Technical, Harlow, New York 1987. HARDT, T., MENKE, P., W. HENKE (2006): Innovative morphologische Ansätze der primatologischen Anthropologie. Mainzer naturwiss. Archiv 44, 87–149. – (1991): How many hominid species should there be? Journal of Human Evolution 20, 413–427. HARVATI, K. (2007): Neanderthals and Their Contemporaries. In: HENKE, W., TATTERSALL, I. (Eds.), Handbook of Paleoanthropology, Volume III. Berlin, Heidelberg, New York 2007, 1717–1748. FRANCK, G. (1998): Ökonomie der Aufmerksamkeit. Ein Entwurf. München 1998. HAWKS, J.D., M.H. WOLPOFF (2001): The Accretion Model of Neandertal Evolution. Evolution 55, 1474–1485. FRAYER, D.W., WOLPOFF, M.H., THORNE, A.G., SMITH, F.H., G. POPE (1993): Theories of modern human origins: The paleontological test. American Anthropologist 95, 14–50. FUTUYAMA, D. J. (2007): Evolution. Heidelberg 2007. HEBERER, G. (1972): Der Ursprung des Menschen. Stuttgart 1972. GILBERT, W.H., WHITE, T.D., B. ASFAW (2003): Homo erectus, Homo ergaster, Homo “cepranensis”, and the Daka cranium. Journal of Human Evolution 45, 255– 259. HEMMER, H. (2007): Estimation of Basic Life History Data of Fossil Hominoids. In: HENKE, W., TATTERSALL, I. (Eds.), Handbook of Paleoanthropology. Volume I, Berlin, Heidelberg, New York 2007, 587–620. GOODMAN, M. (1962): Immunochemistry of the primates and primate evolution. Annals New York Academy of Sciences 102, 219–234. HENKE, W. (2003a): Evaluating human fossil finds. In: GRUPE, G., PETERS, J. (Eds.), Decyphering ancient bones of bioarchaeological collections. Documenta Archaeobiologicae Volume 1, Rahden/Westf. 2003, 59–76. – (1963): Man’s place in the phylogeny of primates as reflected in serum proteins. In: WASHBURN, S.L. (Ed.), Classification and Human Evolution. Aldine, Chicago 1963, 204–234. – (2003b): Population dynamics during the European Middle and Late Pleistocene – smooth or jumpy? In: BURDUKIEWICZ, M., FIEDLER, L., HEINRICH, W.-D., JUSTUS, A., BRÜHL, E. (Hrsg.), Erkenntnisjäger. Kultur und Umwelt des frühen Menschen. Festschrift für Dietrich Mania. Veröffentlichungen des Landesamtes für Archäologie Sachsen-Anhalt, Band 57/I, Halle: Landesmuseum für Vorgeschichte, 247–258. GROVES, C.P., V. MAZÁK (1975): An approach to the taxonomy of the Hominidae: gracile Villafranchian hominids in Africa. Casopis pro Mineralogii a Geologii. 20, 225–247. GRUPE, G. (2007): Taphonomic and Diagenetic Processes. In: HENKE, W., TATTERSALL, I. (Eds.), Handbook of Paleoanthropology, Volume I. Berlin, Heidelberg, New York 2007, 241–260. – (2005): Human biological evolution. In: WUKETITS, F.M., AYALA, F.J. (Ed.), Handbook of Evolution, Volume 2, Wiley – VCH, Weinheim, New York 2005, 117–222. – (2006a): Evolution und Verbreitung des Genus Homo – Aktuelle Befunde aus evolutionsökologischer Sicht. aktuell 18 Paläoanthropologie – Standortbestimmung einer innovativen Disziplin floresiensis and Patients with Primary Growth Hormone Insensitivity (Laron Syndrome). American Journal of Physical Anthropology 134, 198–208. In: CONARD, N.J. (Hrsg.), Woher kommt der Mensch? Tübingen 2006, 104–142. – (2006b): Ursprung und Verbreitung des Genus Homo – paläobiologische Anmerkungen zum evolutiven Erfolg unserer Gattung. In: Rheinisches LM Bonn, Gabriele Uelsberg (Hrsg.), Roots//Wurzeln der Menschheit. Mainz 2006, 33–52. HUBLIN, J.J. (1998): Climatic changes, paleogeography, and the evolution of the Neandertals. In: AKAZAWA, T., AOKI, K., BAR-YOSEF, O. (Eds.), Neandertals and modern humans in Western Asia. Plenum, New York, 295–310. – (2006c): Religiöses Verhalten – eine soziobiologische Annäherung an den Glauben. Forum TTN (Technik, Theologie, Naturwissenschaften) 18, 2–18. HUMMEL, S. (2003): Ancient DNA typing: Methods, strategies and applications. Berlin, Heidelberg, New York 2003. HENKE, W. KIESER, N., W. SCHNAUBELT (1996): Die Neanderthalerin – Botschafterin der Vorzeit. Edition Archaea, Gelsenkrichen und Schwelm. – (2007): Ancient DNA. In: HENKE, W., TATTERSALL, I. (Eds.): Handbook of Paleoanthropology. Volume I: Principles, Methods and Approaches. Berlin, Heidelberg, New York 2007, 643–671. HENKE, W., (2007): Historical Overview of Paleoanthropological Research. In: HENKE, W, TATTERSALL, I. (Eds.), Handbook of Paleoanthropology, Volume I. Berlin, Heidelberg, New York 2007, 1–56. HUXLEY, T.H. (1863): Zeugnisse für die Stellung des Menschen in der Natur. Braunschweig 1863. HENKE, W., H. ROTHE (1994): Paläoanthropologie. Berlin, Heidelberg, New York, London, Paris, Tokyo, Hong Kong, Barcelona, Budapest 1994. JOBLING, M.A., HURLES, M. E., C. TYLER-SMITH (2004): Human Evolutionary Genetics. Origins, Peoples & Disease. New York 2004. – (1997): Zahnphylogenese der Hominiden. In: ALT, K. W., TÜRP, J. C. (Hrsg.), Die Evolution der Zähne. Phylogenie, Ontogenie, Variation. Berlin 1997: Quintessenz, 279–360. JUNKER, T., U. HOSSFELD (2001): Die Entdeckung der Evolution – Eine revolutionäre Theorie und ihre Geschichte. Darmstadt 2001. – (1999a): Stammesgeschichte des Menschen. Eine Einführung. Berlin, Heidelberg, New York 1999. KALB, J. (2001): Adventures in the Bone Trade: The Race to Discover Human Ancestors in Ethiopia‘s Afar Depression. New York 2001. – (1999b): Die phylogenetische Stellung des Neandertalers. Biologie in unserer Zeit, 29, 6, 320–329. – (2001): Entstehung des Genus Homo. Ein Quantensprung in der Evolution des Menschen. Praxis der Naturwissenschaften. Biologie in der Schule. 50, 29–33. KAPPELER, P. (2005): Verhaltensbiologie. Berlin, Heidelberg, New York 2005. KRAUSE, J. LALUEZA-FOX, C., ORLANDO, L., ENARD, W., GREEN, R. E., BURBANO, H. E., HUBLIN, J.-J., HÄNNI, C.FORTEA, J. DE LA RASILLA, M., BERTRANPETIT, J., ROSAS, A. , S. PÄÄBO (2007): The Derived FOXP2 Variant of Modern Humans Was Shared with Neandertals. Current Biology, 17, 1908–1912. – (2003): Menschwerdung. Franfurt a. M 2003. – (2005): Ursprung, Adaptation und Verbreitung der Gattung Homo. In: KLEEBERG, B., WALTER, T., CRIVELLARI, F. (Hrsg.), Urmensch und Wissenschaften. Festschrift Dieter Groh zum 70sten Geburtstag. Darmstadt 2005, 89–123. KRINGS, M., STONE, A., SCHMITZ, R.W., KRAINITZKI, H., STONEKING, M., S. PÄÄBO (1997): Neandertal DNA sequences and the origin of modern humans. Cell 90, 1–20. – (2006): Zur Entwicklung der Paläoanthropologie im 20. Jahrhundert – von der Narration zur hypothetik deduktiven Forschungsdisziplin. In: BREIDBACH, O., HOSSFELD, U. (Eds.): Anthropologie nach Haeckel. Stuttgart 2006, 46–71. KUTSCHERA, U. (2006): Evolutionsbiologie. 2. Auflage. Stuttgart 2006. HENKE, W., I. TATTERSALL (2007, Eds.): Handbook of Paleoanthropology. Volume I: Principles, Methods and Approaches. Volume II: Primate Evolution and Human Origins. Volume III: Phylogeny of Hominids. Berlin, Heidelberg, New York 2007. 2173 S. LEAKEY, L.S.B., TOBIAS, P.V., J.R. NAPIER (1964): A new species of the genus Homo from Olduvai Gorge. Nature 2002, 7–9. LÜKE, U., SCHNAKENBERG, J., G. SOUVIGNIER (2004, Hrsg.): Darwin und Gott. Das Verhältnis von Evolution und Religion. Darmstadt 2004. HENNIG, W. (1950): Grundzüge einer Theorie der phylogenetischen Systematik. Berlin 1950. - (1966): Phylogenetic Systematics. Urbana, Ill. HERSHKOVITZ, I., KORNREICH, Z., Z. LARON (2007): Comparative Skeletal Features between Homo 19 aktuell Winfried Henke RELETHFORD, J.H. (1999): Genetics of modern human origins and diversity. Annual Review of Anthropology 27: 1–23. MARTIN, R.D., MACLARNON, A.M., PHILLIPS, J.L., W.B. DOBYNS (2007): Flores hominid: New species or microcephalic dwarf? The Anatomical Record Part A: Discoveries in Molecular, Cellular, and Evolutionary Biology 288A (11): 1123–1145. – (2001a): Genetics and the search for modern human origins. A. New York, Weinheim 2001. – (2001b): Ancient DNA and the origins of modern humans. Proceedings of the National Academy of Sciences 98, 390–391. MAYR, E. (1975): Grundlagen der zoologischen Systematik. Hamburg 1975. – (1991): One Long Argument: Charles Darwin and the Genesis of Modern Evolutionary Thought. Cambridge, Mass. 1991 RETALLACK, G. (2007): Paleosols. In: HENKE, W., TATTERSALL, I. (Eds.), Handbook of Paleoanthropology, Volume I. Berlin, Heidelberg, New York 2007, 383–408. – (1994): und Darwin hat doch recht: Charles Darwin, seine Lehre und die moderne Evolutionsbiologie. Aus dem Engl. von Inge Leipold. München; Zürich 1994. RICHARDS, M. (2002): The Neolithic Invasion of Europe. Annual Review of Anthropology 32: 135–162. MCHENRY, H.M. (1996): Homoplasy, clades, and hominid phylogeny. In: MEIKLE, W.E., HOWELL, F.C., JABLONSKI, N.G. (Eds.), Contemporary Issues in Human Evolution. Wattis Symposium Series in Anthropology, California Academy of Sciences Memoir 21, San Francisco CA, 77–92. RICHARDS, M.P., PETTIT, P. B. TRINKAUS, E., SMITH, F. H., PAUNOVIC, I. KARAVANIC (2000): Neanderthal diet at Vindija and Neanderthal predation: The evidence from stable isotopes. Proceedings of the National Academy of Sciences (PNAS) 97, 13. 7663–7666. MEDER, A. (2007): Great Ape Social Systems. In: HENKE, W., TATTERSALL, I. (Eds.), Handbook of Paleoanthropology, Volume II. Berlin, Heidelberg, New York 2007, 1235–1272. RIEDL, R. (1975): Die Ordnung des Lebendigen. Systembedingungen der Evolution. Hamburg, Berlin 1975. RIGHTMIRE, G.P. (1990): The evolution of Homo erectus: Comparative anatomical studies of an extinct human species. Cambridge, Mass 1990. MINGH-PURVIS, N., K.J. MCNAMARA (2002, Eds.): Human Evolution through Developmental Change. Baltimore, Maryland 2002. – (1998): Human Evolution in the Middle Pleistocene: The Role of Homo heidelbergensis. Evolutionary Anthropology 6, 218–227. NENTWIG, W. (2007a): Human Environmental Impact in the Paleolithic and Neolithic. In: HENKE, W., TATTERSALL, I. (Eds.), Handbook of Paleoanthropology, Volume III. Berlin, Heidelberg, New York 2007, 1881–1900. – (2007b): Humanökologie. 2. Auflage. Berlin, Heidelberg, New York 2007. – (2001a):. Morphological Diversity in Middle Pleistocene Homo. In: TOBIAS, P.V., RAATH, R., MOGGICECCHI, J. DOYLE, G. (Eds.), Humanity from African Naissance to Coming Millenia. Firenze, Johannesburg 2001, 135–140. OSCHE, G. (1983): Die Sonderstellung des Menschen in evolutionsbiologischer Sicht. Nova Acta Leopoldina NF 55 (235), 57–72. – (2001b): Comparison of the Middle Pleistocene hominids from Africa and Asia. In: BARHAM, L., ROBSON-BROWN, K. (Eds.), Human roots: Africa and Asia in the Middle Pleistocene. Bristol 2001, 123–133. – (2007): Later Middle Pleistocene Homo. In: HENKE, W., TATTERSALL, I. (Eds.), Handbook of Paleoanthropology, Volume III. Berlin, Heidelberg, New York 2007, 1695–1716. OVCHINNIKOV, I.V., GÖTHERSTRÖM, A., ROMANOVA, G.P., KHARITONOV, V.M., LIDÉN, K., W. GOODWIN (2000): Molecular analysis of Neanderthal DNA from the northern Caucasus. Nature 404, 490–493. RIGHTMIRE, G.P., LORDKIPANIDZE, D., A. VEKUA (2005): Anatomical descriptions, comparative studies and evolutionary significance of the hominin skulls from Dmanisi, Republic of Georgia. Journal of Human Evolution 50, 115–141. PÄÄBO, S., POINAR, H., SERRE, D., JAENICKE-DESPRÉS, V., HEBLER, J., ROHLAND, N., KUCH, M., KRAUSE J., VIGILANT L., M. HOFREITER (2004): Genetic analyses from ancient DNA. Annual Review of Genetics 38, 645–679. PIANKA, E.R. (1988): Evolutionary Ecology. 3rd ed., New York 1988. ROTHE, H., W. HENKE (2001a): Methoden der Klassifikation und Systematisierung. Praxis der Naturwissenschaften. Biologie in der Schule 50, 1–7. PITTENDRIGH, C. S. (1958): Adaptation, Natural selection, and Behavior. In: ROE, A., SIMPSON, G.G. (Eds.), Behavior and Evolution. New Haven 1958, 390–416. aktuell – (2001b): Vom Lebewesen zum Fossil: Grundzüge der Taphonomie. Praxis der Naturwissenschaften. Biologie in der Schule 50, 8–12. 20 Paläoanthropologie – Standortbestimmung einer innovativen Disziplin – (2005): Machiavellistische Intelligenz bei Primaten. Sind die Sozialsysteme der Menschenaffen Modelle für frühmenschliche Gesellschaften. In: KLEEBERG, B., WALTER, T., CRIVELLARI, F. (Hrsg.), Urmensch und Wissenschaften. Eine Bestandsaufnahme. Darmstadt 2005, 161–194 SPONHEIMER, M., LEE-THORP, J. (2007): Hominin Paleodiets: The Contribution of Stable Isotopes. In: HENKE, W., TATTERSALL, I. (Eds.): Handbook of Paleoanthropology, Volume I. Berlin, Heidelberg, New York 2007, 555–586. – (2006): Stammbäume sind wie Blumensträuße – hübsch anzusehen, doch schnell verwelkt. In: PREUSS, D., HOSSFELD, U., BREIDBACH, O. (Hrsg.), Anthropologie nach Haeckel. Stuttgart 2006, 149–183. STONEKING, M., R.L. CANN (1989): African origin of human mitochondrial DNA. In: MELLARS, P., STRINGER, C.B. (Eds.), The Human Revolution. Behavioural and Biological Perspectives on the Origins of Modern Humans. Edinburgh 1989, 17–30. SARICH, V. M. (1971): A molecular approach to the question of human origins. In: DOLHINOW, P., SARICH, V. M. (Eds.), Background for Man. Little, Brown Comp., Boston, 60–81. STORCH, V., WELSCH, U., WINK, M. (2007): Evolutionsbiologie. 2. Aufl., Berlin, Heidelberg, New York 2007. SCHÖNBORN, C., Kardinal (2007): Ziel oder Zufall? Schöpfung und Evolution aus der Sicht eines vernünftigen Glaubens. Freiburg 2007. STRINGER, C.B. (1982): Towards a solution of the Neanderthal problem. Journal of Human Evolution 2, 431–438. SCHWARTZ, J.H., I. TATTERSALL (2002): The Human Fossil Record. Volume 1: Terminology, Craniodental Morphology of Genus Homo (Europe). New York 2002. – (1986): The credibility of Homo habilis. In: WOOD, B.A., MARTIN, L.B., P. ANDREWS (Eds.), Major Topics in Primate and Human Evolution. Cambridge 1986, 266–294. – (2003): The Human Fossil Record. Volume II: Craniodental Morphology of Genus Homo (Africa and Asia). New York 2003. – (1992): Replacement, continuity and the origin of Homo sapiens. In: BRÄUER, G., SMITH, F.G. (Eds.), Continuity or Replacement. Controversies in Homo sapiens Evolution. A.A. Balkema, Rotterdam, 9–24. SENUT, B. (2007): The Earliest Putative Hominids. In: HENKE, W., TATTERSALL, I. (Eds.). Handbook of Paleoanthropology, Volume III. Berlin, Heidelberg, New York 2007, 1519–1538. – (1996): Current issues in modern human origins. In: MEIKLE, W., HOWELL, F.C., JABLONSKI, N. (Eds.), Contemporary issues in human evolution. Wattis Symposium Series in Anthropology. California Academy of Sciences Memoir 21, San Francisco, CA, 115–134. SENUT, B., PICKFORD, M., GOMMERY, D., MEIN, P. CHEBOI, K., Y. COPPENS (2001): First hominid from the Miocene (Lukeino Formation, Kenya). CR Acad Sci Paris332: 137–144. – (2001): Modern human origins: distinguishing the models. African Archaeological Review, 18, 67–75. – (2002): Modern human origins: progress and prospects. Philosophical Transactions of the Royal Society of London (B), 357, 563–579. SERRE, D., LANGANEY, A., CHECH, M., TESCHLER-NICOLA, M., PAUNOVIC, M., MENNECIER, P., M. HOFREITER (2004): No Evidence of Neandertal mtDNA Contribution to Early Modern Humans. PLoS Biology 2(3), 0313–0317. STRINGER, C.B., GAMBLE, C. (1993): In Search of the Neanderthals. Solving the Puzzle of Human Origins. London 1993. SHIPMAN, P. (1981): Life History of a Fossil. An Introduction to Taphonomy and Palaeoecology. Cambridge, MA 1981. SUSSMAN, R.W., D. HART (2007): Modelling the Past; The Primatological Approach. In: HENKE, W., TATTERSALL, I. (Eds.): Handbook of Paleoanthropology, Volume I. Berlin, Heidelberg, New York 2007, 701–722. SMITH, F. (2002): Migrations, radiations and continuity: Patterns in the evolution of Middle and Late Pleistocene humans. In: HARTWIG, W.C. (Ed.), The Primate Fossil Record. Cambridge 2002, 437- 456. SUWA, G., KONO, R.T., SHIGEHIRO KATOH, ASFAW, B., Y. BEYENE (2007): A new species of great ape from the late Miocene epoch in Ethiopia. Nature 448, 921–924. SÖLING, C. (2004): Der Gottesinstinkt. Diss. FB 08, Biologie, Chemie, Geowissenschaften der Universität Gießen, Gießen 2004. SPENCER, F. (1990a): The Piltdown Papers 1908 – 1955. Natural History Museum, Oxford 1990 TATTERSALL, I. (1986): Species recognition in human paleontology. Journal of Human Evolution 15, 165–175. – (1995a): The fossil trail. How we know what we think we know about human evolution. New York, San Diego, London 1995. – (1990b): Piltdown – A Scientific Forgery. Natural History Museum, Oxford 1990. – (1995b): The last Neanderthal: The rise, success, and mysterious extinction of our closest human relatives. New York 1995. 21 aktuell Winfried Henke VEKUA, A., LORDKIPANIDZE, D., RIGHTMIRE, G.P., AGUSTÍ, J., FERRING, R., MAISURADZE, G., MOUSKHELISHVILI, A., NIORADZE, M., PONCE DE LEON, M., TAPPEN, M., TVALCHRELIDZE, M., C. ZOLLIKOFER (2002): A New Skull of Early Homo from Dmanisi, Georgia. Science 297, 85–89. – (1999): Neandertaler. Der Streit um unsere Ahnen. Basel: Birkhäuser 1999. – (2000a): Paleoanthropology: The last half-century. Evolutionary Anthropology 9, 2–16. – (2000b): Once we were not alone. Scientific American 2, 56–63. – (2005): Patterns of Innovation in Human Evolution. Nova Acta Leopoldina NF 93, Nr. 345: 145–157. VOGEL, C. (1975): Praedispositionen bzw. Praeadaptationen der Primaten-Evolution im Hinblick auf die Hominisation. In: KURTH, G., EIBL-EIBESFELDT (Hrsg.), Hominisation und Verhalten. Stuttgart 1975, 1–31. – (2007): Homo ergaster and its contemporaries. In: HENKE, W., TATTERSALL, I. (Eds.), Handbook of Paleoanthropology, Volume III. Berlin, Heidelberg, New York 2007, 1633–1654. – (1982): Charles Darwin, sein Werk „Die Abstammung des Menschen“ und die Folgen. In: DARWIN, C. , Die Abstammung des Menschen. Deutsche Übersetzung von H. Schmidt. Stuttgart 1982, VII–XLII. TATTERSALL, I., SCHWARTZ, J.H. (2000): Extinct Humans. Boulder, Colorado 2000. TEMPLETON, A.R. (2002): Out of Africa again and again. Nature 416: 45–51. – (1983): Biologische Perspektiven der Anthropologie: Gedanken zum Theorie-Defizit der biologischen Anthropologie in Deutschland. Zeitschrift für Morphologie und Anthropologie 73, 225–236. – (2006): Population genetics and microevolutionary theory. New York 2006. – (2000): Anthropologische Spuren. Zur Natur des Menschen. Herausgegeben von Volker Sommer. Stuttgart 2000, Leipzig. THIEME, H. (2005): Die ältesten Speere der Welt – Fundplätze der frühen Altsteinzeit im Tagebau Schöningen. Archäologisches Nachrichtenblatt 10, 409–417. VRBA, E. (2007): Role of Environmental Stimuli in Hominid Origins. In: HENKE, W., TATTERSALL, I. (Eds.), Handbook of Paleoanthropology, Volume III. Berlin, Heidelberg, New York 2007, 1441–1482. THORNE, A.G., M.H. WOLPOFF (1981): Regional continuity in Australasian Pleistocene hominid evolution. American Journal Physical Anthropology 55, 337–349. – (1992): Multiregionaler Ursprung des modernen Menschen. Spektrum der Wissenschaft H. 6, 80–87. WÄGELE, J.-W. (2000): Grundlagen der phylogenetischen Systematik. München 2000. TOBIAS, P.V. (1989a): The status of Homo habilis in 1987 and some outstanding problems. In: GIACOBINI, G. (Ed.), Hominidae. Proceedings of the 2nd International Congress on Human Paleontology, Turin, 1987. Mailand 1989, 141–149. WAGNER, G.A. (2007a): Chronometric Methods in Paleoanthropology. In: HENKE, W., TATTERSALL, I. (Eds.), Handbook of Paleoanthropology, Volume I. Berlin, Heidelberg, New York 2007, 311–338. – (2007b, Hrsg.): Einführung in die Archäometrie. Berlin, Heidelberg, New York 2007. – (1989b): A gradual appraisal of Homo habilis. In: GIACOBINI, G. (Ed.): Hominidae. Proceedings of the 2nd International Congress of Human Paleontology, Turin 1987. Mailand 1989, 151–154. WERNER, F. C. (1970): Die Benennung der Organismen und Organe nach Größe, Form, Farbe und anderen Merkmalen. Halle 1970. ULHAAS, L. (2007): Computer-based Reconstruction. Technical Aspects and Applications. In: HENKE, W., TATTERSALL, I. (Eds.): Handbook of Paleoanthropology. Volume I: Principles, Methods, and Approaches. Berlin, Heidelberg, New York 2007, 787–814. WHEELER, Q.D., R. MEIER (2000, Eds.): Species Concept and Phylogenetic Theory. A Debate. New York 2000. WHITE, T.D. (2000): A view on the science: Physical anthropology at the millennium. American Journal of Physical Anthropology 113, 287–292. ULHAAS, L., KULLMER, O., SCHRENK, F., W. HENKE (2004): A new 3-d approach to determine functional morphology of cercopithecoid molars. Annals of Anatomy 186, 487–493. – (2001) Once Were Cannibals. Scientific American, August, 2001, 58–65. WIESEMÜLLER, B., ROTHE, H., W. HENKE (2003): Einführung in die Phylogenetische Systematik. Berlin, Heidelberg, New York 2003. ULLRICH, H. (1998): Zur Diskussion um die Erstbesiedlung Europas und den ältesten Europäer. Mitteilungen der Berliner Gesellschaft für Anthropologie, Ethnologie und Urgeschichte Bd. 19, 57–74. aktuell WOLPOFF, M.H. (1992): Theories of modern human origins. In: BRÄUER, G., SMITH, F. (Eds.), Continuity or 22 Paläoanthropologie – Standortbestimmung einer innovativen Disziplin Replacement. Controversies in Homo sapiens Evolution. A.A. Balkema, Rotterdam, 25–63. WOOD, B.A., M. COLLARD (1999a): The Human Genus. Science 284, 65–71. – (1996-1997): Human Evolution. 1996-1997 Edition. The McGraw-Hill Companies, Inc., College Custom Series, New York. – (1999b): The Changing Face of Genus Homo. Evolutionary Anthropology 8, 195–207. – (2001): Evolving Interpretations of Homo. In: TOBIAS, P.V., RAATH, R., MOGGI-CECCHI, J. DOYLE, G. (Eds.), Humanity from African Naissance to Coming Millenia. Firenze, Johannesburg 2001, 141–146. – (1999): Paleoanthropology, 2nd ed., McGraw-Hill, Boston. WOLPOFF, M.H., R. CASPARI (1997): Race and Human Evolution. A fatal attraction. New York 1997. WOOD, B., CONSTANTINO, P. (2006): Human origins: life at the top of the tree. In: CRACRAFT, J., DONOGHUE, M. J. (Eds.): Assembling the Tree of Life. Oxford 2006, 517–535. – (2000): The many species of humanity. Prezglad Anthropologiczny - Anthropological Review 63, 2–17. WOLPOFF, M.H., SENUT, B., PICKFORD, M., J. HAWKS (2002): Sahelanthropus or ‘Sahelpithecus’? Nature 419, 581–582. ZILHÃO, J. (2006): Neandertals and Modern Mixed, and It Matters. Evolutionary Anthropology 15, 183–195. WOLPOFF, M.H., THORNE, A.G., JELINEK, J., ZHANG YINYUN (1994a). The case of sinking Homo erectus. 100 years of Pithecanthropus is enough! In: FRANZEN, J. L. (Ed.), 100 years of Pithecanthropus. The Homo erectus problem. Courier Forschungsinstitut Senckenberg 171, Forschungsinstitut Senckenberg, Frankfurt am Main, 341–361. ZOLLIKOFER, C., M. PONCE DE LEON (2005): Virtual Reconstruction: A Primer in Computer-assisted Paleontology and Biomedicine. Weinheim 2005. Prof. Dr. rer. nat. habil. Dr. h.c. Winfried Henke Institut für Anthropologie (1050) Fachbereich 10 Biologie Johannes Gutenberg-Universität Mainz 55099 Mainz phone: (0049) 06131/ 39-22398 fax: (0049) 06131/ 39-23799 Email: [email protected] WOLPOFF, M.H., THORNE, A.G., SMITH, F.H., FRAYER, D.W., G.G. POPE (1994b): Multiregional Evolution: a world-wide source for modern human populations. In: NITECKI, M.H., NITECKI, D.V. (Eds.), Origins of Anatomically Modern Humans. New York 1994, 175–199. WOOD, B.A. (1996): Origin and Evolution of the Genus Homo. In: MEIKLE, W.E., HOWELL, F.C., JABLONSKI, N.G. (Eds.): Contemporary Issues in Human Evolution. Wattis Symposium Series in Anthropology, California Academy of Sciences Memoir 21, San Francisco, CA, 105–113. 23 aktuell Die Pest als Interpretationsproblem mittelalterlicher und frühneuzeitlicher Massengräber Simone Kahlow Zusammenfassung – Die Gründe für die Anlegung von Massengräbern sind äußerst vielgestaltig. Darum verwundert es zum Teil, dass als Interpretation fast immer auf die „Wahrscheinlichkeit“ einer Seuche oder auf die Pest selbst zurückgegriffen wird. Die kurze Einführung in die Problematik des Themas dürfte zeigen, dass andere Seuchen, Hungersnöte oder auch Feuersbrünste, die im Mittelalter und der Frühen Neuzeit allesamt als pestis bezeichnet wurden, eine Ursache darstellen können, ebenso wie klimatische Auswirkungen und Kriege. Die Unsicherheit hinsichtlich der Interpretation könnte durch ein Verfahren gemindert werden, mit welchem es möglich ist, eine spezifische DNA-Sequenz nachzuweisen, die dem Pestbazillus zugeordnet werden kann (PRECHEL 2002, 247). Schlüsselwörter – Abstract – The definition of mass or common graves from 14th to 17th century as well as the interpretation of the detected structures remains problematic. Almost all discussed examples are referred to as „pest-graves“ by their authors, inspite of the frequent lack of any adequate indices. The very fact of being faced with a grave revealing several burried individuals seems to be sufficiently for such an interpretation. But there are various reasons for the usage of common graves, which are evidence of an state of emergency of affected populations. Historical documents attest to multiple disasters causing the death of many people all over Europe. They testify a famine in consequence of crop failures (1315-1317) resulting from dramatic climate changes. Contemporaries reported on another remarkable high mortality in the years 1339/1340. An epidemic infectious disease like anthrax appears imaginable. Seventeenth century records document the interdependence of wars and the appearance of epidemic plagues. During the Thirty Years’ War (1618-1648) many more people died from typhus or pest than from injuries caused by the military conflict, because hunger and misery have a subtantial impact on the immune system. But of course mass graves with war victims are well-known like the battlefields of Visby and Towton. These disasters are mostly closely related to each other. A clear distinction between them is hardly possible if not impossible with archaeological means. Keywords – Einleitung Auch in jüngster Zeit werden immer wieder Gräber aus der Zeit des 14. bis 17. Jahrhunderts freigelegt, die mehrere Individuen beinhalten und somit als Massengräber angesprochen werden. Problematisch ist nicht nur die Definition dieses Begriffes, sondern auch die Interpretation des Inhaltes der besagten Gräber. Eine stattliche Anzahl der betreffenden Befunde wurde von den Bearbeitern als Pestgräber angesprochen, auch wenn häufig jegliche Indizien hierzu fehlen. Oft scheint allein die Tatsache, dass sie mehrere Individuen beinhalteten, für die genannte Auslegung auszureichen. Die Pest Angaben zur Pest des 14. Jhs. sind sich im Allgemeinen recht ähnlich. Diese kontinentübergreifende Seuchenwelle kam aus Zentralasien, traf 1342 in Genua ein und verbreitete sich von dort aus über fast alle Regionen Europas. In den Jahren 1349 bis 1351 wurde schließlich auch der deutschsprachige Raum von ihr heimgesucht, wo sie die Bevölkerung möglicherweise um ein Drittel dezimierte, was 20 bis 25 Millionen Tote bedeuten würde (GECKELER 1961).1 Zahlreiche Schriftquellen berichten von einem unter der Bevölkerung herrschenden Ausnahmezustand. Zusehends wurde auf jegliches christliches Protokoll verzichtet und die Toten in Massengräbern bestattet. Giovanni Boccaccio schreibt 1348 im Decameron: „Da für die große Menge Leichen … der geweihte Boden nicht langte ... machte man … sehr tiefe Gruben und warf die Hinzukommenden in diese zu Hunderten. Hier wurden die Leichen angehäuft wie Waren in einem Schiff und von Schicht zu Schicht mit ein wenig Erde bedeckt, bis die Grube bis zum Rand voll war.“ (Zit. nach WITTE 2001, 22-23). Der Strassburger Chronist Fritsche Closener führt 1349 weiterhin an, wo diese Gruben angelegt worden sind: „Daz sterbent [Pest 1349] war so gros, das gemeintlich alle tage ein ieglichem kirspel liche worent 7 oder 8 oder 9 oder 10, oder noch danne me, one die man in klöstern begrub un ene[ohne] die, die man in den spital drug: der was unzelich vil, das man die spitelgrube, die bi der kirchen stunt, muste in einen witen garten setzen, wann die alte grub zu enge und zu klein was.“ (Zit. nach ILLI 1992, 58). Archäologische Informationen 30/1, 2007, 97-104 Bulletin de la Société Suisse97d‘Anthropologie 13 (1), 2007 Bulletin der Schweizerischen Gesellschaft für Anthropologie 13 (1), 2007 Simone Kahlow Im jüngeren Horizont wurden zwei Gruben geöffnet (Bef.-Nr. 4528 und 4529), die, trotz einer Störung, insgesamt 696 Individuen erbrachten und als Pestopfer angesprochen wurden (PRECHEL 1996, 325). Die Körper lagen dicht an dicht in fünf bis sechs Schichten übereinander. Ebenfalls in diesem Horizont befanden sich zwei kleinere Massenbeisetzungen (Bef.-Nr. 4562 und 4571) mit insgesamt 120 Bestattungen. Zwischen den einzelnen Körperlagen des „Pestgrabes“ soll Erde gestreut worden sein, da aus den einzelnen Skelettschichten Keramikscherben geborgen werden konnten. Hier deckt sich der archäologische Befund mit den schriftlichen Quellen, beispielsweise der Erwähnung Boccaccios. Die Keramikscherben des „Pestgrabes“ sollen in das 13./14. Jahrhundert datieren, was zunächst mit dem C14-Ergebnis des Knochenmaterials von 1260 bis 1390 übereinstimmt. Prechel äußert daraufhin, dass „die Bestattungsgruben tatsächlich im Jahre 1350 ausgehoben wurden“ (PRECHEL 1996, 325). Diese Sicherheit ist keineswegs gegeben, da Problemstellung Um die Problematik bezüglich der Interpretation von Massengräbern zu verdeutlichen, sollen im Folgenden die vom Heiligen-Geist-Hospital zu Lübeck (Abb. 1) diskutiert werden (PRECHEL 1996; 2002).2 Ihren Bekanntheitsstatus dürften sie erlangt haben, da auf dem untersuchten Areal zum einen gleich mehrere Massengräber zutage traten, diese weiterhin eine enorme Anzahl von mehr als 600 Individuen aufgenommen hatten und zudem anthropologische Untersuchungen vorgenommen worden sind. Die besagten Gräber, die im Jahr 1990 an der Südwand des besagten Hospitals freigelegt werden konnten, gehören stratigraphisch zwei unterschiedlichen Zeitstufen an. Aus mehreren Grabgruben des älteren Bestattungshorizontes konnten insgesamt 158 Skelette geborgen werden. Eine C14-Datierung des Knochenmaterials ergab einen Zeitraum der Niederlegung zwischen 1280 und 1390 (PRECHEL 1996, 325). Abb. 1 Heiligen-Geist-Hospital zu Lübeck: Massengrab, Befund-Nr. 4529 (aus PRECHEL 1996, Taf. 11,2). aktuell 98 Die Pest als Interpretationsproblem mittelalterlicher und frühneuzeitlicher Massengräber jedes einzelne Jahr des angegebenen Zeitraums, das „gesuchte“ sein könnte. Aus den Chroniken, so PRECHEL (1996, 325), ist wohl tatsächlich überliefert, dass die Pest 1349/1350 in Lübeck grassiert und ca. 5000 Menschen ihr Leben lassen mussten. Auch der absolute Sterbegipfel bei den Jungerwachsenen (Abb. 2), der bei den Individuen aus dem „Pestgrab“ nachgewiesen wurde, könnte auf den ersten Blick für Pestopfer sprechen. So schrieb schon Gisela Grupe (GRUPE 1986, 32): „Sicheres Zeugnis dafür, daß die Pest eine neue Krankheit war, die eine Bevölkerung ohne Immunitätsschutz traf, ist der Befund, daß bevorzugt die jüngeren Erwachsenen starben. Dafür gibt es physiologische Gründe. Nach dem Abklingen dieser ersten Welle war die überlebende Bevölkerung hinreichend immunisiert, so daß die folgende Welle erst nach etwa 12 Jahren wieder viele Opfer forderte. Doch dieses Mal wurden bevorzugt die nachgewachsenen Kinder und Jugendlichen hinweggerafft...“. Leider ist diese Feststellung hinsichtlich der Interpretation keine Hilfe. Zum einen ist es nicht möglich, die Skelette auf das Jahr genau zu datieren, um sie dann mit den schriftlichen Erwähnungen einzelner Pestwellen abzugleichen. Zum anderen scheint sich die Pest nicht überall gleich verhalten zu haben. Es gibt zahlreiche Regionen, die von dieser Krankheit fast unberührt geblieben sind, wie ein „Großteil der Auvergne oder des nördlichen Flandern, Teile Frankens und weitere Gebiete Europas“ (RUFFIÉ/SOURNIA 2000, 36). Warum dies so war, kann nur vermutet werden. Bereits Erkrankte sind möglicherweise wieder genesen, da sie „jenes Genmaterial schon in sich trugen, das unerlässlich ist, um dem Bazillus zu widerstehen..., oder weil sie es erwarben, indem sie später an einer regelrechten natürlichen Imp- fung teilnahmen. Diese widerstandsfähige Gruppe wurde zum Bollwerk der Gemeinschaft gegen die wiederholten Attacken der Seuche“ (ebd). Weiterhin wird von Ruffié und Sournia vermutet, dass sogar die Nachkommen der Betroffenen eine genetische Resistenz geerbt haben könnten. Jedoch dürften die Gene nicht der alleinige Grund für das Fernbleiben dieser Seuche gewesen sein, abgesehen davon, dass diese „natürliche Impfung“ nur gut 12 Jahre anhielt. Es scheint doch so, dass mehrere Faktoren dafür verantwortlich sind, ob ein Land oder eine Stadt von der Pest heimgesucht wurde. In diesem Fall beziehe ich mich nicht auf Äußerungen, nach welchen bestimmte Blutgruppen für den Erreger als bevorzugt angesehen wurden, sondern auf die nüchterne Erkenntnis, dass ein vermehrtes Rattenvorkommen, unhygienische Zustände und klimatische Einflüsse eine große Rolle spielten, so wie es auch VASOLD (2003) völlig überzeugend darlegen konnte. Jedoch halte ich auch eine Variation bzw. Mutation des Erregers nicht für unwahrscheinlich. Es ist anzunehmen, dass sich der Erreger verändern musste, um wirksam zu bleiben. Schließlich kam der Floh ursprünglich aus einer warmen Zone, bevorzugte 20 bis 25 Grad Celsius und traf in Europa schließlich auf ein kühl-gemäßigtes Klima. Zudem musste die Luftfeuchtigkeit stimmen (VASOLD 2002, 174). Die Annahme einer Variation des Erregers könnte auch von Aussagen mittelalterlichen Chronisten gestützt werden, welche eine andere Sterbeverteilung, als die eben angesprochene, innerhalb der einzelnen Pestwellen beobachteten. Diepold Schilling schilderte für Bern die kurz aufeinanderfolgenden Epidemiewellen des 15. Jhs. „Allein die drei schweren Epidemien zwischen 1478 und 1493 forderten über 2000 Opfer, beim ersten Seuchenzug vor allem Kinder“ (ULRICH-BOCHSLER 1999, 105), beim zweiten Seuchenzug 1482 „vor allem Frauen, und Hunderte von Kranken“ (GERBER 1999, 100). Als ein Jahr später wieder ein Seuchenzug durch das Land ging, scheinen alle Bevölkerungsteile betroffen gewesen zu sein (ebd.). Es kann sich in diesem Fall jedoch nicht um besagte die „Kinderpest“ gehandelt haben, hierfür müsste der vorherige Pestausbruch ca. 12 Jahre zurückliegen. Laut den Schriftquellen hatte dieser jedoch 1439 stattgefunden, was eine Differenz von 39 Jahren bedeuten würde. Angesichts dieser Beobachtungen muss die Frage gestellt werden, inwiefern bestimmte Sterbemaxima der Skelette aus Massengräbern auf Pestgräbern deuten sollen? So ist der maximale Sterbegipfel der „robusten“ (PRECHEL 1996, 327) Jungerwachsenen auch anderorts nachweisbar, so Abb. 2 Heiligen-Geist-Hospital zu Lübeck: Relative Anzahlen der Verstorbenen nach Altersklasse (aus PRECHEL 1996, Abb. 2). 99 aktuell Simone Kahlow z.B. auf dem Heidelberger Spitalfriedhof (Abb. 3), auf dem keine Massengräber ergraben wurden (WAHL 1993, 481). Als modernes Beispiel wäre die Spanische Grippe von 1918 heranzuziehen, bei der vorrangig diese Altersgruppe gestorben ist (SCHUH 2003). Nach neuesten Schätzungen sollen ihr 25 bis 40 Millionen Menschen zum Opfer gefallen sein. Bei dieser Größenordnung scheint es nicht verwunderlich, dass auch in diesem Fall Massengräbern angelegt worden sind, so beispielsweise in dem Dorf Brevig in Alaska (ebd.). Grab 506 (Abb. 4) ist für diese Arbeit besonders relevant, da es von dem münzdatierten Grab 525 mit den Denaren des Markgrafen Waldemars (1305-1319) überlagert wurde. Nach LANGE (1997, 116-117) muss das Massengrab 506 somit noch ins frühe 14. Jh. datiert werden, wobei es wohl zu Zeit der Pest angelegt worden sein kann, da diese im Jahr 1348 auch Berlin heimsuchte. Diese Bestätigung findet sich in den Schriftquellen jedoch nicht. So äußerte auch schon Christian Popp 2006 als These seiner Disputation die Vermutung, dass Berlin nicht von der Pest heimgesucht wurde, was möglicherweise auch auf das angrenzende Brandenburg zutrifft. Denn trotz „dichter Urkunden- oder Stadtbuchüberlieferung wie [in] Stendal und Berlin/Cölln lassen sich keine Belege für ein Übergreifen der Pest finden“.3 Da der Friedhof des Berliner Heilig-Geist-Spitals bereits im 13. Jh. angelegt worden ist, könnte auch Grab 506 durchaus noch in diesen Zeitraum datieren und die Opfer einer anderen Seuche beinhalten. Hesse verwies hierzu auf den sogenannten Schneiderbrief von 1288 aus dem Berliner Stadtbuch4; welches von Martin Ohm (OHM 1954, 76-77) übersetzt wurde: „Jetzt wo der Tod anstürmt, sich nicht beruhig sondern tobend alles zum Einsturz bringt ist es nützlich, vorsichtig aufzuzeichnen solche zutreffenden Tatbestände und Schriftstücke (darüber) anzufertigen, die zu gegebener Zeit die Wahrheit mit Sicherheit erweisen sollen, wodurch sich Zweifelsfragen widerlegen lassen.“ Ohm merkt hierzu an: „Der Vorspruch zu dieser Urkunde läßt erahnen, daß um 1288 schwere Seuchen in Berlin herrschten“. Im allgemeinen wird die These einer hier zur Sprache gebrachten Seuche jedoch, aufgrund der Wortwahl, eher verworfen. Dies ist jedoch kein Beweis dafür, dass nicht doch die Opfer einer Seuche in das besagte Massengrab gelangt sind.5 Respekt wurde ihren sterblichen Überresten offenkundig nicht entgegengebracht. So wurden alle 20 Individuen in die Gruben geworfen, so dass sie zum Teil mit völlig verrenkten Gliedern liegen geblieben sind. Tatsächlich ist dies ein eher seltenes Phänomen bei Massengräbern, und konnte beispielsweise auch am Stadelhof in Paderborn (Abb. 5) beobachtet werden (WITT-STUHR/WIEDMANN/SPIONG 2007). Insgesamt scheint jedoch ein pietätvoller Umgang häufiger. Als Beispiel sollen die Gruppenbestattungen an der Bartholomäuskirche in Erfurt genannt werden (Abb. 6). Hier lagen bis zu 10 Individuen in „sorgsamer Verschränkung“ übereinander. „Die beiden größten Individuen bildeten stets die unterste Lage auf der Sohle der Alternative Interpretationen Dieses Beispiel soll als Überleitung zu anderen Interpretationsmöglichkeiten dienen. Viele mir bekannte Nachweise von Massengräbern, werden von den Bearbeitern entweder direkt als Pestgräber oder aber als Gräber, die zu Epidemiezeiten angelegt worden sind, gedeutet. Zu diesen Epidemien könnten auch die Grippe, der Milzbrand, die Pocken oder Typhus gehören. Möglicherweise bezeugen auch Schriftquellen Epidemien noch vor der Pandemie des „Schwarzen Todes“. Im Jahr 1995 wurden auf dem Gelände des ehemaligen Heilig-Geist-Spitals in Berlin mehrere Massengräber freigelegt, die in die Zeit des 14. bis 17. Jahrhundert datiert werden (LANGE 1996) und von den Bearbeitern Heinrich Lange (LANGE 1996) und Henrike Hesse (HESSE 1996) zunächst allgemein als Pestgräber interpretiert wurden. Abb. 3 Heidelberger Spitalfriedhof: Relative Altersverteilung der verstorbenen Individuen (aus WAHL 1993, 481). aktuell 100 Die Pest als Interpretationsproblem mittelalterlicher und frühneuzeitlicher Massengräber Abb. 4 Heilig-Geist-Spital Berlin: Grab 506 (aus LANGE 1997, 117). Grabgrube. Daraufhin lagen mit den Köpfen etwa auf Schulterhöhe die nächstgrößeren Individuen. Extrem stark belegte Grabgruben wiesen eine dritte Lage mit ebenso sorgsam niedergelegten Kleinkindern bzw. Säuglingen auf.“ (WULF 2005, 54). Die Unregelmäßigkeit der Grabgrube, aber auch eine geringe Tiefe dieser, könnten darauf hindeuten, dass ein Massengrab im Winter angelegt worden ist. In diesem Fall sind neben der Deutung als Seuchenopfer auch solche von klimatischen Einflüssen vorstellbar. Dass besonders die ärmere Bevölkerung unter den strengen Wintern, ganz besonders in der Frühen Neuzeit während der kleinen Eiszeit, leiden musste, ist ebenfalls durch Schriftquellen bewiesen. Ferner ist bekannt, dass im 12./13. Jahrhundert eine Knappheit an Holz herrschte (GRUPE 1986, 29). Somit wäre es durchaus denkbar, dass die Opfer des Winters, sei es, dass sie an der Kälte, an Unterernährung oder an daraus resultierenden Erkrankungen gestorben sind, in diesen Gräbern bestattet wurden. In dem Fall ist sicherlich auch der Begriff Armengräber angebracht. Spuren von Mangelerscheinungen wie Cribra orbitalia, transversale Schmelzhypoplasien oder Harris-Linien, um nur einige zu nennen, wären dann ein zu erwar- Abb. 5 Paderborn „Am Stadelhof”, Gruppenbestattungen (aus WITT-STUHR/WIEDMANN/SPIONG 2007, Abb. 1). 101 aktuell Simone Kahlow rum ein solcher Hungerstod nicht auch auf die anderen Individuen der Lübecker Massengräber zutreffen sollte. Einmal abgesehen von der unterschiedlichen Sterbeverteilung, die dadurch erklärt werden könnte, dass zu Beginn die schwachen und dann erst die stärkeren Individuen verstarben, konnte PRECHEL nämlich feststellen, dass der Gesundheitszustand der Individuen aus allen Gruben „als ungewöhnlich schlecht bezeichnet werden muss“ (PRECHEL 2002, 284). Nur die Toten aus den kleineren Gräbern des oberen Bestattungshorizontes wiesen weniger Gelenkerkrankungen und Mangelerscheinungen auf als die übrigen Untersuchten. Im Vergleich zu einer normalen Lübecker Population des Mittelalters war jedoch auch dieser Grad an pathologischen Veränderungen zu hoch (ebd.). Als ein an dieser Stelle letzter Lösungsvorschlag ist an die Opfer gewalttätiger Auseinandersetzungen und Kriege zu denken. Sicherlich wären in diesem Fall mehrheitlich Verletzungserscheinungen anzutreffen, wie sie beispielsweise von den Toten aus der Schlacht von Visby 1361 (THRODEMAN 1939) oder der von Towton 1461 (Abb. 7) bekannt sind (FIORATO/BOYLSTON/KNÜSEL 2000). Doch auch hier sind Parallelen zu anderen Ursachen für die Anlage von Massengräbern möglich. Zur Zeit des Dreißigjährigen Krieges (1618-1648) beispielsweise, starben die Menschen weniger an Verwundungen, als an Fleckenfieber oder an der Pest. Abb. 6 Erfurt; Anger 53: Gruppenbestattung (aus WULF 2005, 54). tendes Bild, sollte ein Zustand aus adäquater und unzureichender Ernährung über einige Zeit angehalten haben. An einer Hungersnot könnten auch die Individuen aus dem unteren Bestattungshorizont des Lübecker Heiligen-Geist-Hospitals gestorben sein. So nimmt es Prechel an, die eine hohe Anzahl an Mangelerscheinungen feststellen konnte. Zudem ist für die Jahre 1315/1317 eine Hungersnot in Lübeck überliefert, ebenso wie der Ort, an dem die Opfer bestattet wurden: „Binnen den twen Jaren starf to dene hilghen gheste 2300 volkis unde de hunger was so grot“ (PRECHEL 1996, 327-328). Es stellt sich jedoch dem Betrachter die Frage, wa- aktuell 102 Die Pest als Interpretationsproblem mittelalterlicher und frühneuzeitlicher Massengräber Abb. 7 Towton (England): Massengrab (FIORATO/BOYLSTON/KNÜSEL 2000, Fig. 4.14). Anmerkungen Literatur 1 Diese, in der Literatur häufig anzutreffende Angabe wird jedoch öfter als übertrieben bezeichnet. So beispielsweise Johannes Nohl (NOHL 1924, 40) sowie in jüngerer Zeit Manfred Vasold. Seiner Meinung nach kann die Pest nicht solch hohe Verluste verursacht haben. Die Zahlen müssen entweder zu hoch gegriffen sein oder es handelte sich um eine oder mehrere andere Seuchen, unter der selbstverständlich auch die Pest grassiert haben könnte (VASOLD 2003, 11, 116, 118, 123). FIORATO, V./BOYLSTON, A./C. KNÜSEL (Hrsg.) (2000): Blood Red Roses. The Archaeology of a Mass Grave from the Battle of Towton AD 1461. Oxford 2000. GECKELER, C. (2007): Kieler Erinnerungstage: 14. Februar 1961. Mittelalterlicher Kieler Pestfriedhof gefunden: http://www.kiel.de/kultur/stadtarchiv/ erinnerungstage1.php?id=49[12.05.2007]. GERBER, R. (1999): Rückgang und Stagnation. Die Bevölkerungsentwicklung im 15. Jahrhundert. In: Berns grosse Zeit. Das 15. Jahrhundert neu entdeckt. Bern 1999, 97-102. 2 Die Vorstellung weiterer mir bekannter Massengräber, die eine Interpretation als Seuchengräber erfuhren, kann hier nicht vorgenommen werden, was jedoch an anderer Stelle in Kürze nachgeholt wird. 3 GRUPE, G. (1986): Umwelt und Bevölkerungsentwicklung im Mittelalter. In: B. Herrmann (Hrsg.), Mensch und Umwelt im Mittelalter. Stuttgart 1986, 24-34. Frdl. schriftliche Mitteilung vom 12.07.2007. 4 H. Hesse, Die anthropologischen Untersuchungen des Friedhofs des Heiliggeist-Spitals. Vortrag zum Landesgeschichtlichen Kolloquium von Prof. Winfried Schich am 6.12.2005 in Berlin. HESSE, H. (1998): Der Friedhof des Berliner HeiligGeist-Spitals – ein anthropologischer Zwischenbericht. Mitteilungen der Berliner Gesellschaft für Anthropologie, Ethnologie und Urgeschichte 19, 1998, 75-86. 5 Auch der Nachweis einer Feuersbrunst in Berlin bringt bei Grab 506 keine weitere Interpretationsmöglichkeit. Dieses Feuer hatte erst im Jahr 1380 innerhalb einer Nacht fast ganz Berlin zerstört: „Alle öffentlichen Gebäude, zahlreiche Bürgerhäuser und alle Kirchen, außer der Klosterkirche, wurden zerstört. Wieviele Einwohner den Flammen zum Opfer fielen, ist unbekannt.“ (RIBBE/SCHMÄDEKE 1994, 1). Die Anlage von Massengräbern wäre aber somit auch in diesem Fall möglich, wenngleich mir archäologische Nachweise hierzu bisher fehlen. ILLI, M. (1992): Wohin die Toten gingen. Begräbnis und Kirchhof in der vorindustriellen Stadt. Zürich 1992. KAHLOW, S. (2005): Krankheiten im Mittelalter und der Frühen Neuzeit im archäologischen Befund. Unpublizierte Magisterarbeit, Philosophische Fakultät I der Humboldt-Universität zu Berlin, 2005. 103 aktuell Simone Kahlow THORDEMAN, B. (1939): Armor from the Battle of Wisby. Stockholm 1939. LANGE, H. (1997): Massengräber und mittelslawische Siedlung. Archäologie in Berlin und Brandenburg 1995-1996. Stuttgart 1997, 116-118. ULRICH-BOCHSLER, S. (1999): Krankheit und Tod – im Spiegel des Siechenfriedhofs am Klösterlistutz. In: Berns grosse Zeit. Das 15. Jahrhundert neu entdeckt. Bern 1999, 102-107. NOHL, J. (1924): Der Schwarze Tod. Potsdam 1924. OHM, M. (1954): Das mittelalterliche Stadtbuch von Berlin. Neu bearbeitet zu Studienzwecken auf Anregung der Verwaltungsakademie Berlin in den Jahren 1952-1954, Privatsonderdruck o.O. 1954. VASOLD, M. (2003): Die Pest. Ende eines Mythos. Stuttgart 2003. PRECHEL, M. (1996): Anthropologische Untersuchungen der Skelettreste aus einem Pestmassengrab am Heiligen-Geist-Hospital zu Lübeck. Lübecker Schriften zur Archäologie und Kulturgeschichte 24, 1996, 323-339. WAHL, J. (1993): Der Heidelberger Spitalfriedhof. In: Stadtluft, Hirsebrei und Bettelmönch. Die Stadt um 1300. Katalog zur Ausstellung. Stuttgart 1993, 470-485. WITTE, K. (2001): Giovanni Boccaccio. Das Dekameron. Düsseldorf 2001. – (2002): Eine Lübecker Population von 1350. Krankheiten und Mangelerscheinungen. In: M. GLÄSER (Hrsg.), Archäologische Untersuchungen auf dem Lübecker Stadthügel: Befunde und Funde. Lübecker Schriften zu Archäologie und Kulturgeschichte 26, 2002, 245-286. WITT-STUHR, M./WIEDMANN, B./S. SPIONG (2007): Verscharrt und vergessen – Die Toten vom Stadelhof. Die Warte, Jg. 68,Nr. 133, 2007, 8-14. WULF, D. (2005): Seuchenopfer unter der Kirche? Archäologie in Deutschland, Heft 4, 2005, 54-55. RIBBE, W./J. SCHMÄDEKE (1994): Kleine BerlinGeschichte. Berlin 1994. Simone Kahlow M.A. Rehfelder Str. 24 D-15566 Schöneiche b. Berlin [email protected] RUFFIE, J./J.-C. SOURNIA (2000): Die Seuchen in der Geschichte der Menschheit. Stuttgart 2000. SCHUH, H. (2007): Grippen, Gräber und Gelehrte. Die Zeit 16.10.2003, Nr. 43. http://wissenschaft-online.de/ artikel/689843 [10.05.2007]. aktuell 104 Gesichtsrekonstruktionen von drei alamannischen Individuen aus dem Gräberfeld Basel–Gotterbarmweg (5./6. Jh. n. Chr.) Fabian Link Zusammenfassung – Der vorliegende Aufsatz behandelt drei Gesichtsrekonstruktionen von frühmittelalterlichen, alamannischen Individuen aus dem Gräberfeld Basel–Gotterbarmweg (alter Gotterbarmweg, heute Schwarzwaldallee, BS), das vom 5. bis ins beginnende 6. Jh. n. Chr. datiert. Drei der 34 Individuen weisen als einzige einen mehr oder weniger kompletten Schädel auf. Bis dahin konnte von zwei der Individuen nicht mit Sicherheit gesagt werden, ob es sich um eine Frau oder um einen Mann handelt. Der dritte Schädel wurde als eindeutig weiblich identifiziert, was sowohl die neueste anthropologische Geschlechtsbestimmung, als auch die Gesichtsrekonstruktion untermauern. Die zweite anthropologisch interessante Frage ist mit der erstaunlichen Tatsache verbunden, dass das vermutete weibliche Individuum (Individuum 1) ein äusserst kleines Schädelvolumen aufweist. Zudem konnte an den Individuen 1 und 2 eine auffällige alveolare Prognathie beobachtet werden. Eine ikonographische und eine morphologische Frage stehen demnach in der vorliegenden Untersuchung im Vordergrund: 1) eine mögliche Geschlechtsbestimmung und 2) eine Erklärung der auffälligen Prognathie und des kleinen Schädelvolumens des erwähnten Individuums. Die drei Gesichtsrekonstruktionen wurden mit der Methode Skultety gefertigt, die sich in ihren Hauptcharakteristika nach der Methode von Michail M. Gerassimow und den Weichteildickenmessungen von Gerassimow, Kollmann und Skultety richtet. Schlüsselwörter – Gesichtsrekonstruktion – Frühmittelalter – Alamannen – Prognathie – Bevölkerungsstruktur Abstract – The present article discusses three facial reconstructions of 5th and early 6th century alamanic individuals of the graveyard at Gotterbarmweg (old Gotterbarmweg, today Schwarzwaldallee, BS) in Basel, which I would like to discuss. Only three of 34 individuals have a more or less complete skull. Up until now it could not be defined if two of the alamanic individuals were females or males, whereas the third skull is definitely female. The second anthropologically interesting question is the observation that one individual has an unusual cerebral volumina and a conspicuous Prognathism (individual 1). Resumtive there is an icongraphical and a morphologically interesting question in the foreground: 1) the definition of the sex and 2) an explanation of the conspicuous Prognathism and of the »small grown« skull of the one individual. The three facial reconstructions have been done using the Skultety method which is based on the method that developed by Michael M. Gerassimow and the measurings of the soft tissues by Gerassimow, Kollmann and Skultety. Keywords – Facial reconstruction – Early medieval age – Alamanics – Prognathism – Population structure Einleitung Das Gebiet um Basel hat eine bewegte und dynamische Geschichte im Frühmittelalter gehabt. Dies bezeugen die zuerst vorwiegend alamannischen1, später dann fränkischen Gräberfelder, die auf Archäologen und interessierte Laien schon seit dem 18. und vor allem dann im 19. und beginnenden 20. Jh. grosse Faszination ausübten (vgl. FINGERLIN [A] 1997, 45; 46). Doch verursachte die archäoanthropologische Laientätigkeit – und die drei rekonstruierten Individuen aus dem Gräberfeld Basel–Gotterbarmweg (alter Gotterbarmweg, heute Schwarzwaldallee, BS) scheinen paradigmatisch hierfür zu sein – ein aus heutiger Sicht unzureichendes Mass an ungesichertem und nur teilweise nachvollziehbarem Wissen (vgl. WAHL ET AL. 1997, 337). Sind die Grabbeigaben und die Lokalisierung der Gräber für den damaligen Forschungsstand des frühen 20. Jh. recht fortschrittlich publiziert (vgl. VOGT 1930), so wurden die menschlichen Überreste des Gräberfelds mehr geplündert als sorgfältig ausgegraben. Wichtige Knochenreste und ganze Schädel gingen auf diese Weise verloren und wurden nicht dokumentiert. Erfreulicherweise wurde dieses frühe rechtsrheinische Gräberfeld neu von Lic. Phil. Stefan Leh- mann im Rahmen eines Dissertationsprojekts bearbeitet und sollte in absehbarer Zeit publiziert werden. Soviel als kritische Vorbemerkung. Drei der 34 Individuen weisen als einzige einen mehr oder weniger kompletten Schädel auf, bei Individuum 3 konnten die teils ausgebrochenen oberen und unteren Augenbögen mit Bienenwachs ergänzt werden. Bis dahin, trotz früherer anthropologischer Untersuchungen, konnte von zwei der Individuen (Individuum 1 und 3) nicht mit Sicherheit gesagt werden, ob es sich dabei um eine Frau oder um einen Mann handelt. Der dritte Schädel (Individuum 2) wurde als eindeutig weiblich identifiziert, was sowohl die neueste anthropologische Geschlechtsbestimmung, als auch die Gesichtsrekonstruktion untermauern. Die zweite anthropologisch interessante Frage ist mit der erstaunlichen Tatsache verknüpft, dass das vermutete weibliche Individuum (Individuum 1) ein äusserst kleines Schädelvolumen aufweist. Da eine Verwechslung im Naturhistorischen Museum Basel (NMB) ausgeschlossen werden kann, muss das Phänomen anderweitig erklärt werden. Bei den Individuen 1 und 3 konnte zudem eine auffällige alveolare Prognathie (nach KROMER 1938, 9, vorstehende Zahnhälse, die zu einer ausladenden Gesichtsbildung der Mundpartie führt) Archäologische Informationen 30/1, 2007, 81-96 Bulletin de la Société Suisse81d‘Anthropologie 13 (1), 2007 Bulletin der Schweizerischen Gesellschaft für Anthropologie 13 (1), 2007 Fabian Link festgestellt werden, was zur Frage nach der sogenannten, in der frühen historischen und archäoanthropologischen Forschung vermeintlich festgestellten »alamannischen Prognathie« führt; ein angeblich anthropologisches Phänomen, das die zwei Schädel des Gräberfelds Basel–Gotterbarmweg augenscheinlich betonen. Eine ikonographische und eine morphologische Frage bezüglich dieser alamannischen Individuen stehen also in der vorliegenden Untersuchung im Vordergrund: a) die eindeutige Geschlechtsbestimmung und b) eine Erklärung der auffälligen Prognathie der Individuen 1 und 3 und des auffällig kleinen Schädelvolumens des Individuums 1. Diese Fragen möchte ich mittels folgender Gliederung des Textes diskutieren: Ein erster und kurzer Überblick soll das Gräberfeld Basel–Gotterbarmweg in die frühmittelalterliche Besiedlungsgeschichte des süddeutschen und nordwestschweizerischen Raums eingliedern. Eine darauf folgende forschungsgeschichtliche Skizze bettet die Gesichtsrekonstruktion als wissenschaftlich–artifizielles Betätigungsfeld in die Anthropologie ein, das daran anschliessende Kapitel legt in grober Weise die Grundzüge der angewandten Methode (Methode Skultety) ad exemplum der drei alamannischen Individuen dar. Die Ergebnisse, die aus der Untersuchung resultieren, werden in einem abschliessenden Teil in Verbindung mit den formulierten Fragestellungen erläutert. rigen Forschungen ist zu entnehmen, dass es sich bei den sogenannten Alamannen um eine freie Vereinigung germanischer Stämme und Splittergruppen handelte (MOOSBRUGGER–LEU 1982, 10). Es darf aber davon ausgegangen werden, dass sich Unterschiede in der Grabbeigabenkultur von alamannischen zu z. B. fränkischen Gräberfeldern feststellen lassen (vgl. MARTIN 1976). Demnach muss aus anthropologisch–demographischer Perspektive von einer heterogenen Bevölkerungszusammensetzung im Gebiet des Basler Rheinknies im 5. und frühen 6. Jh. n. Chr. gesprochen werden, das sich aus einer romanischen Restbevölkerung, aus alamannischen Bevölkerungsteilen rechts des Rheins und der zunächst schwachen, mit der Zeit aber dominanter werdenden fränkischen Bevölkerung zusammensetzte (GIESLER 1981, 92). Nach der Niederlage der losen alamannischen Stammensverbände im Bodenseeraum gegen den Frankenkönig Chlodwig 496 n. Chr. strömten diese Leute langsam in das schweizerische Mittelland ein (MOOSBRUGGER–LEU 1982, 10; 11). Eine alamannische Besiedlungswelle grösserer Bevölkerungsgruppen in der Nordwestschweiz lässt sich aber erst ab dem 6. und verstärkt im 7. Jh. feststellen (WINDLER 1997, 261–268; vgl. CHRISTLEIN 1978, 22; 23). Erstmals von Karl Stehlin und dem Basler Kantonsingenieur Moor im Jahr 1915 ausgegraben und später dann von Emil Vogt in verdankenswerter Weise publiziert, liegt das behandelte Gräberfeld auf der rechtsrheinischen Seite oberhalb des heutigen Kleinbasels, nordwestlich von der linksrheinischen Einmündung der Birs in den Rhein, etwa 250 m vom Flussufer entfernt. Erste Knochenreste wurden 1915 bei der Neugestaltung des Gehsteigs am alten Gotterbarmweg in einem zur Baumbepflanzung ausgehobenen Loch entdeckt (VOGT 1930: 145; 146). Die vorgefundenen Skelette lagen in Gräbern von etwa 70 cm Breite, die offenbar ohne Steinsetzung konstruiert waren. Holzreste oder sonstige Aufschlüsse über die Bestattungsart wurden nicht beobachtet. Die Toten waren nordwestlich ausgerichtet, die Schädel lagen im Westen (VOGT 1930; 147). Das kleine, unvollständig ausgegrabene Gräberfeld Basel–Gotterbarmweg wies insgesamt 34 Gräber auf. Das Gräberfeld gehört zu den ältesten alamannischen Bestattungsplätzen und war reichlich ausgestattet mit Beigaben, die von grosser Bedeutung für die Chronologie der frühen Merowingerzeit sind. Für die Datierung sehr aufschlussreich sind die Ergebnisse von Ursula Kochs Korrespondenzanalyse für den süddeutschen Raum, in der 13 Gräber zur statistischen Auswertung herangezogen wur- Siedlungsgeschichtliche Hintergründe Seit dem Jungpaläolithikum wurde die Region um das heutige Basel von verschiedenen Menschengruppen als Durchgangs– und später als Siedlungsgebiet genutzt (MOOSBRUGGER–LEU 1982, 2; 3). Erstmals im beginnenden 3. Jh. n. Chr. in der Mainzer Gegend erwähnt, darf die erste alamannische Besiedlungswelle ins späte 3. und 4. Jh. n. Chr. datiert werden, als sich die römische Besatzungsmacht langsam aufzulösen begann (MOOSBRUGGER–LEU 1982, 4; 5; 10)2. Ob das Grenzgebiet zwischen römischem Limes und Rhein infolge einer grösseren Expansionswelle besetzt wurde oder ob verschiedene alamannische Gruppen erst allmählich ihre Siedlungsplätze gegen die römische Reichsgrenze hin vorgeschoben hatten, kann aus Sicht des heutigen Forschungsstands nicht sicher bestimmt werden (FINGERLIN 1997 [B], 125). Die Frage, in welcher Weise von einer alamannischen Ethnie gesprochen werden kann, ist bis anhin noch nicht zureichend geklärt. Den bishe- aktuell 82 Gesichtsrekonstruktionen von drei alamannischen Individuen aus dem Gräberfeld Basel ... 1981, 99)4, wird wohl siedlungsgeschichtlich nicht abschliessend beantwortet werden können. Zu Beginn des 6. Jhs. wird die zum Gräberfeld Basel–Gotterbarmweg zugehörige Siedlung aufgegeben (GIESLER 1997, 211). Im späteren 6. und im 7. Jh. verlagerte sich der Hauptbestattungsplatz der rechtsrheinisch lebenden Bevölkerungsteile auf den heutigen Theodorskirchplatz (GIESLER 1981, 116; vgl. BING 1993: 100). den. Das Frauengrab 15 gehört in die SD–Phase 1, die Frauengräber 6, 24, 13, 18, 10 und 20 sowie die Männergräber 9, 32 und das Kindergrab 22 in die SD–Phase 2. Die Frauengräber 33 und 12 sowie das Männergrab 34 datieren in die SD–Phase 3. Grab 6 enthielt eine geöste Siliqua des Jovianius aus der Zeit von 411–413 n. Chr. (KOCH 2001, 37). Nach der Korrespondenzanalyse würde die SD–Phase 1 die erwähnten Gräber in die Jahre von ca. 430–460 n. Chr. datieren, die SD–Phase 2 setzt die Bestattungen in die Zeit von ca. 460–480 n. Chr. und die SD–Phase 3 datiert in die Zeit von ca. 480–510 n. Chr. (KOCH 2001, 70–73; 80–85). Zusammenfassend darf also mit einer Belegungszeit des Gräberfeldes von etwa 430 bis etwa um 510 n. Chr. gerechnet werden. Zum Gräberfeld gehörige Siedlungsreste wurden entweder nicht entdeckt oder es handelt sich dabei um einen derart kleinen Weiler, dass im Boden keine Siedlungsbefunde mehr beobachtet werden konnten (VOGT 1930, 146). Weitere frühe alamannische und somit vergleichbare Gräberfelder sind Herten im Landkreis Lörrach (D, GIESLER 1997, 209; vgl. KUHN 1963) und das Gräberfeld Basel–Kleinhüningen BS (GIESLER 1997, 209; GIESLER 1981, 96)3. Strategische und wehrtechnische sowie ökonomische und infrasktrukturelle Überlegungen werden wohl bei der Wahl des jeweiligen Siedlungsplatzes ausschlaggebend gewesen sein (GIESLER 1981, 99), wobei heute nicht gesagt werden kann, welcher Gesichtspunkt bei welcher Siedlung wohl überwogen hat (FINGERLIN [B] 1997, 125). Anhand der reichen Grabbeigaben kann vermutet werden, dass es sich bei den bestatteten Alamanninnen und Alamannen um Angehörige einer (adligen) Oberschicht handelte. Die Grabbeigaben, die auch bei den anderen beiden frühen Gräberfeldern (Herten und Basel–Kleinhüningen) häufig aus spätantiken Formen und Motiven bestehen, bezeugen, dass die Alamannen wohl mit der romanischen Restbevölkerung in einem Handelskontakt standen (GIESLER 1981, 103). Nach der Art der Tracht und sonstiger Beigaben dürfen die alamannischen Leute des Gräberfeldes Basel–Gotterbarmweg aus dem südwestdeutschen Raum zugezogen sein. Es könnte sich um eine heterogene Gruppe gehandelt haben, die immer wieder kleinere Verbände aufgenommen hat. Ob die Vermutung sich als zutreffend erweist, dass die rechtsrheinisch lebenden Alamannen ursprünglich der Vinařicer–Gruppe aus Nordböhmen, Thüringen und Mähren angehört haben könnten, die vermutlich vor den eindringenden Hunnen geflohen war (vgl. GIESLER 1997, 209; GIES- LER Forschungsgeschichtlicher Abriss der Gesichtsrekonstruktion Es soll hier nicht der Platz für eine Gesamtdarstellung der Forschungsgeschichte der Gesichtsrekonstruktion eingeräumt, sondern die wichtigsten Arbeiten ab der 2. Hälfte des 19. Jhs. und v. a. im 20. Jh. kurz vorgestellt werden. Totenmasken oder mit Lehm übermodellierte Schädel können ab neolithischer Zeit in verschiedenen Gesellschaften beobachtet werden (PRAG/ NEAEVE 1999, 14)5. Grundlegend für die spätere Technik der Gesichtsrekonstruktion sind die anatomischen Modelle neuzeitlicher Mediziner, die eng mit erfahrenen Bildhauern zusammenarbeiteten, so z. B. die Italiener Gaetano Giulio Zumbo und Ercole Lelli im späten 17. und im 18. Jh. sowie der Brite Joseph Towne im 19. Jh. (PRAG/NEAEVE 1999, 14). Gemeinhin wird dem Anatom Wilhelm His die erste, auf wissenschaftlicher Methodik beruhende Gesichtsrekonstruktion (1895) des 1894 gefundenen Johann Sebastian Bach (1685–1750) zugeschrieben6. Mehr als eine intuitive als wissenschaftliche Arbeit muss indes die vom Jenaer Anatom Schaaffhausen 1884 geschaffene Rekonstruktion eines Gesichts auf einem weiblichen Schädel angesprochen werden (PRAG/NEAEVE 1999, 15). Durch die Zusammenarbeit des Basler Anatoms Julius Kollmann mit dem Schweizer Bildhauer Büchly konnte 1899 die berühmte „Pfahlbauerin von Auvergnier“ rekonstruiert werden (GERASSIMOW 1968, 9)7. Kollmann wendete dabei eigene, auf empirisch ermittelten Messwerten der Weichteildicken der menschlichen Gesichtsmuskeln und –haut beruhende Daten an, die er mit den Weichteildicken von His und Welcker statistisch auswertete (PRAG/NEAEVE 1999, 16). In der Folgezeit erlitt die Wissenschaftlichkeit der Gesichtsrekonstruktion allerdings einen Einbruch, denn der Anatom Solger »rekonstruierte« einen Erwachsenen auf einem Neandertalerschädel eines Knaben aus der Höhle von Le Moustiers (1910) und der berühmte Anthropologe Martin sowie der Anatom Eggeling schufen unabhängig 83 aktuell Fabian Link voneinander ein sehr unterschiedliches Gesicht desselben Schädels aus La Chapelle–aux–Saints (1913)8. Auch die schönen und zu ihrer Zeit beliebten Porträts des talentierten Bildhauers Louis Masquet und des belgischen Anthropologen Rutot müssen aus wissenschaftlicher Sicht verworfen werden (PRAG/NEAEVE 1999, 16). Methodologisch besser scheint die La Chapelle–Rekonstruktion des französischen Anthropologen Boule zu sein (GERASSIMOW 1968, 10). Interessant ist dabei die früh gestellte Frage nach der Identifikation, die für die Gesichtsrekonstruktion gerade im Bereich der Forensik in der Folgezeit von grösster Wichtigkeit sein würde, denn der Anatom Welcker vermochte z. B. die Echtheit des Schädels von Raffael (1833) aufgrund dessen bekannter Ikonographie zu verifizieren, wie dies auch His mit den Schädeln von Bach, Kant und Haydn gelang. Das Bildnis Johann Sebastian Bachs wurde von dem deutschen Bildhauer Sefner 14 Jahre nach der Identifikation von Bachs Schädel von 1894 geschaffen. Dabei wurde dieselbe Methode angewendet, die bereits Kollmann vorgeschlagen hatte (GERASSIMOW 1968, 11). In der nachfolgenden Zeit bürgerte sich die Gesichtsrekonstruktion in der kriminalistischen Praxis ein, doch wurde von den massgebenden Anatomen und Anthropologen noch immer bezweifelt, dass der Zusammenhang zwischen Schädelknochen und Weichteilen im Gesicht so eng wäre, dass eine Gesichtsrekonstruktion wirklich authentisch sein könnte (GERASSIMOW 1968, 12; 13). Erst dem russischen Anatom und Anthropologen Michail M. Gerassimow sollte die Entwicklung einer glaubwürdigen und plausiblen Methode gelingen (1924), die eine Authentizitätsgarantie gewährleisten konnte und in der Folge von seinen Schülern evaluiert wurde (PRAG/NEAEVE 1999, 17)9 . Dabei führte er die empirischen Messungen von Kollmann weiter und suchte auf diese Weise »sein Gesicht des Urmenschen«. Mit über 200 rekonstruierten Gesichtern – darunter so prominenten Fällen wie z. B. Iwan IV. »der Schreckliche« (1530–84) – bürgerte sich seine Methode als die »Russische Methode« in der Anthropologie ein und führte 1950 zur Gründung des heute noch existierenden Labors für Plastische Rekonstruktion am Institut für Ethnologie und Anthropologie der Russischen Akademie der Wissenschaften in Moskau (PRAG/NEAEVE 1999, 17). Das etwa 600 Seiten umfassende Werk über Gerassimows anthropologische Erkenntnisse wurde leider nie aus dem Russischen übersetzt. Abb. 1 Basel, Gotterbarmweg. Schädel des alamannischen Individuums 1) mit starker Prognathie und kleinem Schädelvolumen. Das Sterbealter liegt zwischen 18–25 Jahren, die Geschlechtsbestimmung ist unklar, Frontalansicht. (Gyula Skultety/Fabian Link) Abb. 2 Basel, Gotterbarmweg. Schädel des alamannsichen Individuums 1), Dreiviertelansicht. (Gyula Skultety/Fabian Link) aktuell 84 Gesichtsrekonstruktionen von drei alamannischen Individuen aus dem Gräberfeld Basel ... Abb. 3 Basel, Gotterbarmweg. Schädel des alamannischen Individuums 2). Das Sterbealter liegt zwischen 18–29 Jahren, das Geschlecht ist eindeutig weiblich, Frontalansicht. (Gyula Skultety/Fabian Link) Abb. 4 Basel, Gotterbarmweg. Schädel des alamannischen Individuums 2), Dreiviertelansicht. (Gyula Skultety/Fabian Link) In deutscher Sprache ist lediglich ein kleines Büchlein erschienen, das einen schönen Eindruck von der Gesichtsrekonstruktion vermittelt, aber nicht als eigentliches Lehrbuch zu betrachten ist. Glücklicherweise konnte mein Lehrer Dr. Gyula Skultety als gebürtiger Ungare die Grundzüge der Methode erfassen und durch eigene Messungen und Techniken vervollständigen und erweitern (vgl. SKULTETY 1987–2000). Rekonstruktionsmethode Skultety am Beispiel der drei alamannischen Individuen Die drei vorliegenden, einzig vollständig erhaltenen Schädel alamannischer Individuen aus dem Gräberfeld Basel–Gotterbarmweg konnten nicht mehr mit aller Sicherheit den einzelnen Gräbern zugeordnet werden. Die aktuelle Alters– und Geschlechtsbestimmung wurden am 7. 5. 2007 von Dr. Gerhard Hotz, Lic. Phil. Liselotte Meyer und dem Autor vorgenommen. Die Geschlechtsbestimmung richtet sich nach den Richtlinien bei FEREMBACH ET AL. 1979. Das erste Individuum (Individuum 1, Abb. 1 und 2)10 wurde auf ein Sterbealter von 18–25 Jahren geschätzt. Das Geschlecht konnte nicht sicher bestimmt werden, tendiert aber mit einem Index von –0,4 gegen weiblich. Bei der zweiten, eindeutig weiblichen Alamannin (Individuum 2, Abb. 3 und 4, Geschlechtsindex von –1)11, wurde ein Alter von 18–29 Jahren angesetzt. Einzig der dritte Schädel (Individuum 3, Abb. 5 und 6)12 kann vermutlich dem Grab 3 zugeordnet werden. Der Geschlechtsbestimmungsindex des Schädels liegt bei 0, das Sterbealter wurde auf 45–59 Jahre geschätzt. Es liegen keine weiteren Knochenreste aus dem Inventar des Gräberfeldes Basel–Gotterbarmweg vor, die sicher zu den drei Schädeln gehörten. Die einzelnen, zusammengefassten Schritte der Methode Skultety werden im unten stehenden Kapitel am Beispiel der drei alamannischen Individuen des Gräberfeldes Basel–Gotterbarmweg erörtert. Erweiternd zu diesen methodischen Prinzipien erweisen sich die verdankenswerten Werke von Mehmet Yasar Iscan und Richard P. Helmer (vgl. ISCAN/HELMER 1993) sowie vom britischen Gesichtsrekonstrukteur Richard Neave (vgl. PRAG/NEAEVE 1999), dessen Rekonstruktionen von hoher analytischer und künstlerischer Qualität sind. 85 aktuell Fabian Link Abb. 5 Basel, Gotterbarmweg. Schädel des alamannischen Individuums 3) mit starker Prognathie. Das Sterbealter liegt zwischen 45–59 Jahren, das Geschlecht ist unbestimmt, Frontalansicht. (Gyula Skultety/Fabian Link) Abb. 6 Basel, Gotterbarmweg. Schädel des alamannischen Individuums 3), Dreiviertelansicht. (Gyula Skultety/Fabian Link) dings nur aufgrund allgemeiner Beobachtungen rekonstruiert, nicht aber deren genaues Aussehen wiedergegeben werden (SKULTETY 1987–2000, 3). Bei der Gesichtsrekonstruktion nach der Methode Skultety wird nicht jeder einzelne Gesichtsmuskel rekonstruiert, sondern die für das Gesicht ausschlaggebenden Muskeln werden besonders hervorgehoben oder kleine Muskeln zu Muskelgruppen zusammengefasst und modelliert. Bevor die detaillierte Vorgehensweise bei der Gesichtsrekonstruktion besprochen wird, müssen einige methodische Einschränkungen formuliert werden13 . Entgegen der Annahme Gerassimows, findet Skultety keine Anhaltspunkte an den Schädelknochen, um Rückschlüsse auf die Fülle des Gesichts bzw. den Ernährungszustand der jeweiligen Person machen zu können. Nur wenn andere Knochen der verstorbenen Person vorhanden sind und für die anthropologische Untersuchung beigezogen werden können, ist es möglich, allenfalls Hinweise auf den Ernährungszustand der Person zu Lebzeiten zu erhalten. Verletzungen oder Entstellungen (z. B. durch eine Hautkrankheit), die nur die Gesichtsweichteile betreffen, können in der Regel nicht eruiert werden14. Ausnahmen stellen Personen dar, über deren Leiden medizinhistorische Dokumente existieren. Die Augen– und die Hautfarbe sowie die Haare lassen sich ebenfalls nicht ableiten, ihre Gestaltung kann nur durch eventuelle ikonographische Analogieschlüsse vermutet werden. So erscheint es z. B. sehr unwahrscheinlich, dass Menschen afrikanischer Herkunft blonde und glatte, europide Menschen hingegen dicht gekrauste und schwarze Haare besitzen. Die Ausrichtung der Ohren wird von der allgemeinen Ausrichtung des Gehörgangs abgeleitet, Höhe und Breite können aller- aktuell Schritt 1 Der Gesichtsrekonstruktion vorausgehend muss der vorliegende Schädel als erstes anthropologisch untersucht werden. Falls andere Knochen des Skeletts noch vorhanden sind, sollten auch sie in die Analyse miteinbezogen werden. Wenn möglich, so muss das jeweilige Geschlecht des Menschen bestimmt, der Zustand des Gebisses beurteilt, spezielle Merkmale beobachtet, und die Weichteildicken der insgesamt 29 Weichteildickenpunkte bestimmt werden. Die jeweilige Stärke der Wichteildicken wird anhand der Stärke und Ausbildung des Schädel– und Gesichtsknochenwuchses des zu rekonstruierenden Individuums festgesetzt. Die Weichteildicken variieren erheblich zwischen männlichen und weiblichen Individuen und zwischen robusteren oder grazileren Individuen. Als anthropometrische Grund- 86 Gesichtsrekonstruktionen von drei alamannischen Individuen aus dem Gräberfeld Basel ... Abb. 7 Basel, Gotterbarmweg. Rekonstruktionsschritt 4/1 am Beispiel des alamannischen Individuums 2), Frontalansicht. (Gyula Skultety/Fabian Link) Abb. 8 Basel, Gotterbarmweg. Rekonstruktionsschritt 4/1 am Beispiel des alamannischen Individuums 2), Dreiviertelansicht. (Gyula Skultety/Fabian Link) Abb. 9 Basel, Gotterbarmweg. Rekonstruktionsschritt 4/2 am Beispiel des alamannischen Individuums 2), Frontalansicht. (Gyula Skultety/Fabian Link) Abb. 10 Basel, Gotterbarmweg. Rekonstruktionsschritt 4/2 am Beispiel des alamannischen Individuums 2), Dreiviertelansicht. (Gyula Skultety/Fabian Link) 87 aktuell Fabian Link Für den nächsten Schritt muss der Schädel zuerst präpariert werden. Weil für den zu erstellenden Gipsabguss ein flüssiger Silikonkautschuk auf den Schädel aufgetragen werden muss, ist es unbedingt erforderlich, alle Hohlräume des Schädels (z. B. Augenhöhlen, Nasenöffnung, Gebiss etc.) mit Bienenwachs abzudichten. Es lässt sich leicht und ohne Rückstände wieder entfernen. Auf diese Weise kann der füssige Gummi nicht in die hohlen Partien des Schädels hineinfliessen. Zugleich kann der Unterkiefer am Oberkiefer sicher platziert und befestigt werden. Es werden zwei Gipsabgüsse des Schädels benötigt. Auf dem einen Abguss wird das Gesicht aus Plastilin15 rekonstruiert, der andere Abguss dient der ständigen Kontrolle beim Modellieren. Der Vorteil des Plastilins gegenüber Ton ist, dass er nicht gebrannt werden muss, somit nicht an Volumen verlieren kann und ein Material mit langer Haltbarkeit ist. Wie Richard Neave zu Recht schreibt, sollte aus mehreren Gründen nicht auf dem originalen Schädel rekonstruiert werden (PRAG/NEAEVE1999, 22). Einerseits kann durch die Herstellung eines Abgusses der Originalschädel für weitere Untersuchungen verwendet werden und ist somit nicht während der Rekonstruktionsphase blockiert. Andererseits sind gerade Schädel in fragilem und oft beschädigtem Zustand nicht dazu geeignet, dem Gewicht des Plastilins und dem Druck des Auftrags durch den Rekonstrukteur Stand zu halten. Als drittes Argument muss der ethische Standpunkt vertreten werden: Aus Respekt vor der/dem Toten sollte eine Rekonstruktion auf dem Originalschädel vermieden werden. Ein Gipsabguss wird mit Hilfe von reissfestem Silikonkautschuk16 hergestellt. Dabei wird auf den Originalschädel zuerst Vaseline aufgetragen, sodass am Ende von Schritt 2 sich die Silikonmaske ohne Schäden zu hinterlassen wieder Abb. 11 Basel, Gotterbarmweg. Rekonstruktionsschritt 5/1 am Beispiel des alamannischen Individuums 2), Frontalansicht. (Gyula Skultety/Fabian Link) Abb. 12 Basel, Gotterbarmweg. Rekonstruktionsschritt 5/1 am Beispiel des alamannischen Individuums 2), Dreiviertelansicht. (Gyula Skultety/Fabian Link) lagen der Weichteildicken liegen bei der Methode Skultety die Messungen von Kollmann, Gerassimow und Skultety sowie von Richard Neave vor (vgl. PRAG/NEAEVE 1999, 26; Tab.1). Die Stärke der Gesichtsmuskeln und der darüber liegenden Haut kann bei einem lebenden Menschen je nach Klima, nach Alkoholgenuss und durch andere Faktoren leicht variieren (bis zu 1 mm). Die Nasenform, die sich aus der allgemeinen Richtung des Nasenknochens (Os nasale) und des Fortsatzes des Nasenstachels (Spina nasaslis anterior) ergibt, wird mittels der Gerassimow’schen Tabelle abgeleitet. Erweiternd zu dieser Tabelle können die vier von Richard Neave ermittelten Nasentypen beigezogen werden (vgl. PRAG/NEAEVE 1999, 29; Tab. 3). Schritt 2 aktuell 88 Gesichtsrekonstruktionen von drei alamannischen Individuen aus dem Gräberfeld Basel ... Abb. 13 Basel, Gotterbarmweg. Rekonstruktionsschritt 5/2 am Beispiel des alamannischen Individuums 2), Frontalansicht. (Gyula Skultety/Fabian Link) Abb. 14 Basel, Gotterbarmweg. Rekonstruktionsschritt 5/2 am Beispiel des alamannischen Individuums 2), Dreiviertelansicht. (Gyula Skultety/Fabian Link) vom Schädel ablösen lässt. Vier bis fünf Schichten des flüssigen, mit Verdickungsmittel und Härter angereicherten Silikonkautschuks werden anschliessend auf den Schädel mit einem Pinsel aufgetragen, wobei die einzelne Schicht nach dem Auftrag zuerst aushärten muss, bevor die nächste aufgepinselt werden kann. Weil die vom Schädel abgelöste Silikonmaske nicht selbsttragend ist, d. h. sie verformt sich beim Eingiessen des Gipses, werden auf dem mit Silikonkautschuk ummantelten Schädel vier sich voneinander abtrennende Gipsschalen aufgetragen, die mit Gipsbinden verfestigt sind. Die Trennnähte müssen mit der wasserabweisenden und nicht bindenden Vaseline eingestrichen werden. Wenn die Gipsschalen ausgetrocknet sind, werden sie behutsam von der Silikonkautschukmaske gelöst. Daraufhin wird die Silikonkautschukmaske sorgfältig vom höchsten Punkt des Schädels (Vertex) entlang der Sutura sagitalis aufgeschnitten, sodass der Schädel nicht beschädigt wird und sich der Silikonkautschuk leicht ablösen lässt. Die Maske kann nun in die Gipsschalen passend eingelegt werden. Die Schalen werden mit einem Gummiband zusammengehalten, denn eine Verschiebung kann eine Deformation des Abgusses bewirken. Drei bis vier Gipsschichten werden nun in den Hohlkörper eingegossen, wobei eine Schicht ca. 5–10 Minuten lang austrocknen muss, bevor die nächste folgen kann. Zum Schluss werden die Gipsschalen abgenommen und der fertige Gipsabguss von der Silikonkautschukmaske befreit (SKULTETY 1987–2000, 2–4). Einer der Gipsabgüsse wird auf einem Skulpturen– oder Büstenständer in der Lage der sogenannten »Frankfurter Horizontalen« aufgesetzt und mit Gips befestigt. Der vom Silikonkautschuk befreite Schädel muss zum Schluss von Schritt 2 unbedingt gründlich gereinigt werden, sodass möglichst keine Rückstände der Bienenwachsverkleidung, der Vaseline oder des Silikonkautschuks mehr zurückbleiben. Bevor zum nächsten Schritt übergegangen werden kann, muss der Gipsschädel lackiert werden, sodass der Plastilin auf der Oberfläche besser haftet. Schritt 3 Als nächstes sind insgesamt 29 Punkte, nämlich die für die Modellierung des Gesichts ausschlaggebenden Wichteildickenmarkierungen auf der Knochenstruktur des Abgusses anzuzeichnen, einzubohren und mit kleinen Metallstiften zu versehen. Dabei werden die Weichteildicken, die von 89 aktuell Fabian Link dem vorliegenden Schädel bei Schritt 1 bestimmt wurden, an den Stiften abgemessen und zurechtgeschnitten. Die Stifte helfen bei der Modellierung der Muskelformen und der Haut des Gesichts, die jeweiligen plastischen Volumina nicht zu verlieren. Die Mundwinkel liegen in der Regel unmittelbar hinter dem rechten und dem linken Eckzahn (Dens caninus) des Oberkiefers, weshalb auch diese Stellen mit eingebohrten, grösseren Nadeln gekennzeichnet werden (SKULTETY 1987–2000, 2; 3). Mit denselben grösseren Nadeln wird auch der Okklusionsgrad der Augen angegeben. Der innere Ansatzpunkt der Augenlieder (Musculus orbicularis oculi, Pars palpebrale) ist das Ligamentum palpebrale mediale, das etwa in der Mitte des Os lacrimale liegt. Der äussere Ansatzpunkt lässt sich anhand eines kleinen Höckers erkennen, der sich auf dem äusseren Margo supraorbitalis befindet (SKULTETY 1987–2000, 2–4). Schritt 4 Abb. 16 Basel, Gotterbarmweg. Rekonstruktionsschritt 6, alamannisches Individuum 2). (Gyula Skultety/Fabian Link) Nun werden die Augen eingesetzt und der Nasenknorpel in gelbem Plastilin modelliert (Abb. 7 und 8). Die Augen sind aus gläsernen und innen naturgerecht bemalten Halbkugeln hergestellt. Anschliessend werden die Halsmuskeln, also der hintere Musculus trapezius (grösster Nackenmuskel) und der in Richtung Brust– und Schlüssel- bein (Sternum und Clavicula) ziehende Musculus sternocleidomastoideus sowie die seitlichen Kopf– und die Kaumuskeln (Musculus temporalis und Musculus masseter) geformt. Die Ansätze der Ohren um das Ohrloch (Porus acusticus externus) herum müssen in diesem Stadium bereits markiert und andeutungsweise modelliert werden. Der vordere Halsbereich wird ebenfalls in diesem Schritt bereits ausgestaltet (Musculus sternohyoideus und Musculus omohyoideus, SKULTETY 1987–2000, 11; 12). Der Musculus sternocleidomastoideus setzt direkt am Processus mastoideus an, die Ansatzstelle am Knochen des hinteren Musculus trapezius gibt sich durch die deutlich zu erkennende Lineae nuchalis inferior. Der seitliche, flächige Muskel (Musculus temporalis) zieht von der Lineae temporalis superior hinunter zum Arcus zygomaticus und vermengt sich mit dem Musculus Masseter, dessen Ansatzstelle wiederum am unteren Grad des Unterkiefers liegt (Abb. 9 und 10). Im selben Zug können bereits schon die Haut und die darunter liegende Muskulatur (Musculus orbicularis oculi, Pars orbitalis et Pars palpebralis, und Musculus depressor supercilii) der Augenlieder aufgesetzt und die Öffnung nach dem zuvor markierten Okklusionsgrad ausmodelliert werden. Zuletzt wird die Ohrspeicheldrüse (Glandula parotidea) aufgesetzt, die sich auf dem Musculus Abb. 15 Basel, Gotterbarmweg. Rekonstruktionsschritt 6, alamannisches Individuum 1). (Gyula Skultety/Fabian Link) aktuell 90 Gesichtsrekonstruktionen von drei alamannischen Individuen aus dem Gräberfeld Basel ... sternocleidomastoideus im unteren Bereich der Mandibula und vor der Ohrmuschel befindet. Schritt 5 Dem Gesicht gibt der nächste Schritt bereits eine Idee von seinem Bildnis, denn nun wird die Mundpartie im Zusammenspiel mit der Kinnmuskulatur modelliert (Abb. 11 und 12). Es brauchen an dieser Stelle nicht vollständig alle zum Mund gehörigen Muskeln aufgezählt werden, wichtig sind v. a. die Muskelgruppen des oberen und unteren Mundbereichs. Zur ersten Gruppe gehört der vom oberen Augenbereich zum Mund ziehende Musculus levator labii superioris und der Musculus zygomaticus minor und major. Die Gruppe der unteren Mundmuskeln besteht aus dem Musculus levator anguli oris, dem Musculus orbicularis oris, Pars marginalis und dem Musculus risorius. Wichtig für die Dicke der Ausstülpungen der oberen und unteren Mundmuskulatur, also der Lippen, ist, wie weit vorn oder hinten sich der Unterkieferinnenwinkel am unteren Grad der Mandibula befindet. Je weiter vorn, also zum Kinn hin, der Winkel liegt, desto aufgeworfener sind die Lippen gestaltet. Der vom Mund zum Hinterkopf ziehende Musculus buccinator wird als letzter zum Mund gehörender Muskel modelliert. Eng mit der Mundpartie verbunden Abb. 18 Basel, Gotterbarmweg. Rekonstruktionsschritt 7 alamannisches Individuum 1). (Claudia Link) ist die Muskelgruppe der Kinnmuskeln, die aus dem Musculus depressor labii inferioris, dem Musculus mentalis und dem Musculus depressor anguli oris besteht (SKULTETY 1987–2000, 13–17). Nun muss nur noch die letzte Schicht auftragen werden, die Haut. Dabei werden Plastilinstreifen von etwa 1 cm Breite und ca. 10–15 cm Länge, die eine Stärke von 0,3–0,5 cm aufweisen, zurechtgeschnitten und auf den Kopf gelegt. Wenn die ganze Fläche des Gesichts und der oberen Schädelpartie bedeckt sind, werden die Hautstreifen miteinander verbunden und ausgearbeitet. Weil der Musculus nasalis sehr dünn und mit der Haut eng verwachsen ist, wird er nicht separat geformt, sondern mit dem Auftragen der Haut modelliert. Gleiches gilt für die grosse, von der Kinn– und Mundmuskulatur über die Halsmuskulatur ziehende Muskelplatte des Platysmas (Abb. 13 und 14). Schritt 6 Was nun folgt ist die eigentlich kritische Phase des gesamten Rekonstruktionsablaufs, nämlich die Harmonisierung des Gesichts, besser des bis dahin Modellierten zu einem Gesicht (Abb. 15–17). Abb. 17 Basel, Gotterbarmweg. Rekonstruktionsschritt 6, alamannisches Individuum 3). (Gyula Skultety/Fabian Link) 91 aktuell Fabian Link Gerade an dieser Stelle wird oft die Kritik geäussert, dass eine Gesichtsrekonstruktion ja lediglich der artistische Ausdruck eines wissenschaftlichen Künstlers sei und niemals das einstige Gesicht »tatsächlich« wiedergeben könne. Dieser Kritik können folgende zwei Argumente entgegengehalten werden: Kein vernünftiger Anthropologe würde davon ausgehen, eine Gesichtsrekonstruktion sei ein tatsächliches Abbild des ehemals lebendigen Menschen, schliesslich können wir nicht bestimmen und nur schätzen, welche Haarfarbe und –form der Mensch hatte, wie beleibt sie/er war, welche Augen– und Hautfarbe sie/er hatte und ob sie/er im Gesicht und auf den Knochen nicht sichtbar in irgendeiner Weise entstellt oder verletzt war. Insofern kann eine Gesichtsrekonstruktion lediglich eine Annäherung an die Ikongraphie eines vergangenen Menschen sein, dies soll an dieser Stelle nicht bestritten werden. Aber wir wissen die ungefähre Weichteildicke eines menschlichen Gesichts und ein verständiger Anatomiekünstler ist sich der Form der Muskeln, der Haut und der Gesichtsausprägungen insgesamt bewusst und kann diese auf eine authentische Weise wiedergeben. Das zweite Argument kann ganz allgemein formuliert werden: Keine archäologische Fundstätte könnte jemals eine gelebte Realität widerspiegeln – ein gewisses Rest–, teils auch Übermass an Unwissen/Nichtwissen bleibt bei jeder histo- Abb. 20 Basel, Gotterbarmweg. Rekonstruktionsschritt 7 alamannisches Individuum 3). (Claudia Link) risch–anthropologischen Untersuchung bestehen. oder archäologischen Schritt 7 Zwei Möglichkeiten für den Umgang des fertigen Plastilinkopfes stehen nun zur Verfügung, nämlich die Fertigung eines vollrealistischen, d. h. panoptischen Kopfes, oder eines statuarischen Gipskopfes, der mit einer hypothetischen Frisur versehen werden kann (Abb. 18–21). Die statuarische Variante birgt den Vorteil, dass die nicht sicher bestimmbaren Faktoren als Hypothese sicher erkennbar dargestellt werden können. Eindrücklicher, zugleich aber auch weit hypothetischer, ist die Fertigung eines Panoptikums. Dabei wird für die Herstellung des Gipsabgusses dieselbe Methode wie bei Schritt 2 angewendet (vgl. Schritt 2). Für ein Panoptikum kann der Gips mit einem Kunststoff wie z. B. Polyurethan ersetzt werden, der anschliessend bemalt und mit Haaren versehen wird. Ein Abguss des fertigen Kopfes empfiehlt sich immer, denn der Plastilin ist zwar lange, aber nicht ewig haltbar (SKULTETY 1987–2000, 4; 5). Abb. 19 Basel, Gotterbarmweg. Rekonstruktionsschritt alamannisches Individuum 2). (Claudia Link) aktuell 92 Gesichtsrekonstruktionen von drei alamannischen Individuen aus dem Gräberfeld Basel ... te bereits 1863 A. Ecker fest, der bei zwei alamannischen Schädeln dieses Phänomen feststellen konnte – der eine davon stammte aus den sogenannten »Hünengräbern« bei Freiburg i. Br. Dabei handelte es sich um langschädlige, junge und männliche Alamannen, die von S. Schultze – auf dem Stand der damaligen Anthropologie – als »mit der äthiopischen Rasse verwandt« beschrieben wurden, weil sie einen ausgeprägten Prognathie– Winkel von 81° aufwiesen (KROMER 1938, 23; 24). KROMER (1938, 24; 25) muss allerdings zugeben, dass ihm aus dem Raum Basel kein ähnlicher Fall bekannt geworden sei. Anders hingegen liegt der Fall bei den vier stark prognathen Schädeln von Individuen aus dem Gräberfeld in Herten. Eine Bestätigung der früheren Hypothese, dass die Prognathie ein Merkmal von alamannischen Individuen sei, konnte Kurt Kromer nicht erbringen. Vorstehende Alveolen der Mundpartie lassen sich bei vielen Bevölkerungsgruppen über alle Zeiten hinweg beobachten, ohne dass bestimmte Gruppen zusammengefasst werden könnten (vgl. KROMER 1938, 17–26). Die Frage, ob es sich bei der Proganthie der alamannischen Individuen aus dem Gräberfeld bei Herten und denjenigen aus dem Gräberfeld Basel–Gotterbarmweg um ein regional gebundenes, morphologisches Phänomen handelt, kann daher nicht beantwortet werden. Prognathie – ein »alamannisches Phänomen«? Vorweg muss eine kurze Definition des heute meist in der Zahnmedizin verwendeten Begriffs der Prognathie17 (KROMER 1938, 12) gegeben werden. Der Begriff wurde der Anthropologie entnommen und lässt sich mit »Vorkiefrigkeit« übersetzen. Darunter wird eine Kieferbildung verstanden, bei der die Profillinie des Gesichts von der Stirn über den Nasenrücken bis zu den Zähnen zur horizontal verlaufenden Schädelebene (sogenannte »Frankfurter Horizontale«) einen Winkel von ca. 80° bildet. Im Gegensatz zu dieser Kieferbildung steht in der Anthropologie der Begriff der »Orthognathie«, also der »Geradkiefrigkeit« (ASCHER 1951, 7). Ob die Prognathie tatsächlich als Merkmal einer bestimmten Bevölkerungsgruppe zu betrachten ist, das durch eine bestimmte Ernährungstechnik entstanden ist, muss offen gelassen werden. Ascher unterscheidet hierbei zwischen einem Prognathismus des Kiefers, der bei europiden Menschen weitgehend unbekannt ist und vor allem bei negriden und australoiden wie auch papuanischen Menschen zu beobachten ist, und dem alveolaren Prognathismus, der auch bei Europäern festgestellt werden kann (ASCHER 1951, 8). Aus diesen Gründen verwirft Ascher aus kieferorthopädischer Sicht den Begriff der »alveolaren Prognathie«, sondern definiert das Phänomen auf dem Boden pathologischer Veränderungen eines »normal« angelegten Oberkiefers, der vergesellschaftet sei mit einem in Distillage befindlichen Unterkiefer (Rachitis, erschwerte Nasenatmung, Lutschen u. a., vgl. ASCHER 1951, 8). Wie dem auch sei, die kieferorthopädische Betrachtungsweise muss hier nur ansatzweise interessieren. Die Prognathie der beiden vorliegenden alamannischen Individuen (Individuen 1 und 3) lässt sich als alveolare Prognathie definieren, weil die Alveolen (Zahnhälse) sowohl beim Unterkiefer wie auch beim Oberkiefer relativ gleichmässig vorstehen (ASCHER 1951, 8). Als einzigen wissenschaftlichen Beitrag, der sich mit dem Thema der Prognathie bei Alamannen, nicht aber mit dem Phänomen der Kleinschädligkeit befasst, konnte die kurze Publikation der Inaugural–Dissertation von Kurt Kromer ausfindig gemacht werden (vgl. KROMER 1938, 7; 8). Für diesen Beitrag zum Glück, konzentriert sich der Autor auf das süddeutsche Gebiet. Den Ausschlag für diese Untersuchung gaben die Ausgrabungen des alamannischen Gräberfeldes in Herten (D), bei dem vier Alamannenschädel gefunden wurden, die eine auffällige Prognathie aufwiesen. Hinweise auf eine alamannische Prognathie stell- Ergebnisse und Schlussbemerkung Schwierig und nach wie vor nicht mit völliger Klarheit zu beantworten bleibt die Frage nach der ethnischen Deutung und Bestimmbarkeit der vorliegenden alamannischen Individuen. Im Zuge der belegbaren These von der alamannischen Stammesgemeinschaft (JENTGENS 2001, 121; 122), darf aber angenommen werden, dass es sich bei alamannischen Ethnien um sehr heterogene Gruppen gehandelt haben muss, dies untermauern zumindest die sehr unterschiedlichen morphologischen Ausprägungen der vorliegenden drei wohl weiblichen Individuen. Abschliessend ist zu bemerken, dass das Phänomen der alveolaren Proganthie – bei der Kieferprognathie liegt der Fall anders – grundsätzlich bei allen bisher bekannten Menschentypen, sowohl der Vorzeit, als auch beim Homo sapiens sapiens von der Antike bis über das frühe und hohe Mittelalter wie auch in der Neuzeit immer wieder beobachtet werden konnte, ohne dass damit verknüpfte morphologische Merkmale einer bestimmten Menschengruppe hätten beobachtet werden können (KROMER 1938, 27). So wird der 93 aktuell Fabian Link 4 Die vermutete demographische Zuordnung ergibt aus gewissen Eigentümlichkeiten der Frauentracht, aus bestimmten Merkmalen in den Keramikfundkomplexen und aus dem gelegentlichen Auftreten künstlicher Schädeldeformationen. Fall auch bei den rekonstruierten drei Alamanninnen liegen, denn sowohl die Prognathie als auch die ausgesprochene Kleinschädligkeit des Individuums 1 müssen wohl einfach als physiologisches Phänomen beurteilt werden, ohne dass dabei eine genauere typologische und damit geo-demographische Zuordnung möglich wäre. Ein ähnlich morphologisch diffuses Bild ergibt sich bei der als Vergleich herangezogenen, anthropologischen Betrachtung des alamannischen Gräberfeldes von Elgg im Kanton Zürich: Neben den für die Zeit des 6. Jhs. angeblich typischen schmalen und hohen Schädelformen liegen auch Individuen mit breiten und niedrigen Schädeln vor (WINDLER 1994, 180). Die drei rekonstruierten Alamanninnen unterstreichen das Bild einer morphologisch sehr gemischten alamannischen Bevölkerung, die sich allenfalls durch die Analyse der Grabbeigaben in einzelne Gruppen oder gar Stammesverbände einteilen lässt. 5 Aus Jericho sind neolithische Schädel der Zeit von ca. 7’500–5‘500 v. Chr. bekannt, die mit Lehm übermodelliert wurden und auf diese Weise das Gesicht auf dem Knochen sozusagen »rekonstruiert« wurde (PRAG/NEAEVE 1999, 12; 13). Ein ähnlicher Umgang mit Verstorbenen lässt sich aus dem frühen 20. Jh. in Papua Neuguinea und der Insel Malakula (Vanuatu), Melanesien, beobachten (SPEISER 1996, Taf. 81; Abb. 7; 11). Steht bei diesen Beispielen nicht die wissenschaftliche Rekonstruktion eines Gesichts im Vordergrund, so bezeugen sie doch den besonderen Umgang mit Schädeln von verstorbenen Menschen. 6 1883 publizierte der Anatom Welcker bereits Daten von empirisch ermittelten Weichteildicken (PRAG/ NEAEVE 1999, 14; 15). His wendete dabei ermittelte Weichteildickendaten von 24 männlichen und 4 weiblichen Leichen an. Dank 7 Die neoklassizistische Frisur muss selbstverständlich verworfen werden (vgl. PRAG/NEAEVE 1999, 16; Abb. 3). Zum Schluss soll Dr. Gerhard Hotz, Leiter der anthropologischen Sammlungen des Naturhistorischen Museums Basel (NMB), der uns die drei Schädel zur Verfügung gestellt hat, sowie seiner Mitarbeiterin Lic. Phil. Liselotte Meyer einen herzlichen Dank ausgesprochen werden. Weiteren Dank gebührt Lic. Phil. Stefan Lehmann für wertvolle bibliographische Hinweise. 8 Der Jenaer Anatom Eggelin postulierte daraufhin, dass die Rekonstruktion eines Gesichts auf Basis des Schädels seinen Erfahrungen nach nicht möglich sei (PRAG/NEAEVE 1999, 17). 9 Anmerkungen Dabei wurde Gerassimows Methode durch verschiedentlich durchgeführte Blindversuche getestet, indem die Ergebnisse mit Fotographien verglichen wurden, die dem Gesichtsrekonstrukteur nicht bekannt waren. 1 10 Nr. 1435 (NMB). 11 Nr. 1443 (NMB). 12 VII. 73/1 (NMB). Die Im Text verwendete Terminologie »Alamannen« richtet sich nach der um 300 n. Chr. erstmals in den römischen Schriftquellen überlieferten Bezeichnung »Alamannia« für ein Land, das grösstenteils das heutige Bundesland Baden–Württemberg, Teile des heutigen Bayerns und Hessens umfasst (vgl. NUBER 1997, 59). 13 Die im Text verwendeten anatomischen Fachausdrücke richten sich nach PUTZ/PABST (2006, 32–117). 2 Zur Besiedlungsgeschichte der heutigen Schweiz im Frühmittelalter vgl. MARCHAL 2004, 109–214, zu archäologischen Fundstellen und Befunden vgl. WINDLER ET AL. 2005. 14 Eine Ausnahme stellt z. B. die Knochensyphilis dar, die an den Gesichtsknochen mehr oder weniger deutlich zu erkennen ist. 3 »Kleinhüningen« ist dabei der einzige alte alamannische Ortsname, der bis heute überlebte. Aus dem Gräberfeld von Herten sind 335 Gräber, aus demjenigen von Basel–Kleinhüningen 450 Gräber überliefert, also bedeutend mehr als aus dem Gräberfeld Basel–Gotterbarmweg. aktuell 15 Für die Rekonstruktion des Nasenknorpels und des inneren Augenkarnats wird ein gelber, wächserner Plastilin verendet: PAN.PONGO SCULT. 12 (Italien). Die übrigen Partien werden mit einem leicht zäheren, grauen Plastilin modelliert: OMYACOLOR PLASTICOLOR. 12 (Frankreich). Alle bei der 94 Gesichtsrekonstruktionen von drei alamannischen Individuen aus dem Gräberfeld Basel ... JENTGENS, G. (2001): Die Alamannen. Methoden und Begriffe der ethnischen Deutung archäologischer Funde und Befunde, Freiburger Beiträge zur Archäologie und Geschichte des ersten Jahrtausends 4. Rahden/ Westfalen 2001. Gesichtsrekonstruktion verwendeten Materialien stammen aus dem Geschäft Troller–Kunststoffe AG, Höllstrasse 20, CH–4629 Fulenbach. 16 SILASTIC 3481 BASE. Als Härtungsmittel wird der dazugehörige SILASTIC Härter 81R im Verhältnis 100:5, als Verdickungsmittel der Accelerateru PC37 Thixo LOT N. 4270701 (Shell Aseol AG, 3000 Bern 5) im Verhältnis 100:1 der Silikonkautschukmasse beigegeben. KOCH, U. (2001): Das alamannisch–fränkische Gräberfeld bei Pleidelsheim. Forsch. U. Ber. Vor– u. Frühgesch. Baden–Württemberg 60. Stuttgart 2001. Der Ausdruck »Prognathie« wurde vom britischen Physiologen I. C. Prichard geprägt (KROMER 1938, 9). KROMER, K. (1938): Zur Prognathie einiger Alamannen von Herten. Inaug.–Diss. zur Erlangung der Doktorwürde der Zahnheilkunde der medizinischen Fakultät der Universität Basel. Winterthur 1938. Literatur KUHN, F. (1963): Aus der Frühgeschichte von Herten, Sonderdruck aus der Festschrift anlässlich der Einweihung der neuen Volksschule Herten 1963. Lörrach 1963. 17 ASCHER, F. (1951): Prognathie. Ihre kieferorthopädische, chirurgische und prothetische Behandlung. Kieferorthopädie – Kinderzahnheilkunde 6. München 1951. MARCHAL, G. P. (2004): Die Ursprünge der Unabhängigkeit (401–1394), in: ULRICH IM HOF ET AL., Geschichte der Schweiz und der Schweizer. Basel 2004. 109–214. BING, C. (1993): Neue Befunde im Friedhofsareal St. Theodor Theodorskirchplatz 5 (A). JberBasel 15. Basel 1993. 100–110. MARTIN, M. (1976): Das fränkische Gräberfeld von Basel–Bernerring, Basler Beitr. Ur– u. Frühgesch. 1. Basel 1976. CHRISTLEIN, R. (1978): Die Alamannen. Archäologie eines lebendigen Volkes. Stuttgart 1978. MOOSBRUGGER–LEU, R. (1982): Die frühmittelalterlichen Gräberfelder von Basel, Führer durch das Historische Museum Basel 3. Basel 1982. GERASSIMOW, M. M. (1968): Ich suchte Gesichter. Schädel erhalten ihr Antlitz zurück. Wissenschaft auf neuen Wegen. Gütersloh 1968. NUBER, H. U. (1997): Zeitenwende rechts des Rheins. Rom und die Alamannen, in: Archäologisches Landesmuseum Baden–Württemberg (Hrsg.), Die Alamannen. Stuttgart 1997. 59–68. FEREMBACH, D., ET AL. (1979): Empfehlungen für die Alters– und Geschlechtsdiagnose am Skelett. Homo 30, Anhang. Amsterdam 1979. 1–32. PRAG, J./R. NEAVE (1999): Making Faces. Using Forensic and Archaeological Evidence. London 1999. FINGERLIN, G. (1997): Vom Schatzgräber zum Archäologen. Die Geburt einer Wissenschaft, in: Archäologisches Landesmuseum Baden–Württemberg (Hrsg.), Die Alamannen. Stuttgart 1997. 45–51. PUTZ, R./R. PABST (Hrsg.) (2006): Sobotta. Atlas der Anatomie des Menschen. Kopf, Hals, obere Extremität, Bd. 1, 22., neu bearbeitete Auflage. München 2006. – (1997): Siedlungen und Siedlungstypen. Südwestdeutschland in frühalamannischer Zeit, in: Archäologisches Landesmuseum Baden–Württemberg (Hrsg.), Die Alamannen. Stuttgart 1997. 125–134. GIESLER, U. (1997):Völker am Hochrhein. Das Basler Land im frühen Mittelalter, in: Archäologisches Landesmuseum Baden–Württemberg (Hrsg.), Die Alamanne. Stuttgart 1997. 209–218. SKULTETY, G. (1987-2000): Handarbeit/Kopfarbeit. Gesichtrekonstruktionen auf Grund des Schädels, unpubliziertes Manuskript. Basel 1987–2000. [Das zitierte Werk beinhaltet auch die Ergebnisse eines Lehrgangs in der Technik der manuell gefertigten, plastischen Gesichtsrekonstruktion des Autors, Anm. d. Verf.] – (1981): Die römische Zeit am rechten Rheinufer zwischen Augst und Kembs, in: R. Dehn et al., Lörrach und das rechtsrheinische Vorland von Basel, Führer vor– und frühgesch. Denkmäler 47. Mainz 1981. 92–125. SPEISER, F. (1996): Ethnology of Vanuatu. An early twentieth century study, transl. by D. Q. Stephenson. Hong Kong 1996. ISCAN, M. Y./R. P. HELMER (Hrsg.) (1993): Forensic Analysis of the Skull. Craniofacial Analysis, Reconstruction and Identification. New York 1993. VOGT, E. (1930): Das alamannische Gräberfeld am alten Gotterbarmweg in Basel. Anz. Schweiz. Altkde 32. Zürich 1930. 145–164. 95 aktuell Fabian Link WAHL, J., ET AL. (1997): Zwischen Masse und Klasse. Alamannen im Blickfeld der Anthropologie, in: Archäologisches Landesmuseum Baden–Württemberg (Hrsg.), Die Alamannen. Stuttgart 1997. 337–348. WINDLER, R. (1994): Das Gräberfeld von Elgg und die Besiedlung der Nordostschweiz im 5.–7. Jh. Zürcher Denkmalpflege, Archäologische Monographien 13. Zürich/Elgg 1994. – (1997): Franken und Alamannen in einem Land. Besiedlung und Bevölkerung der Nordschweiz im 6. und 7. Jahrhundert, in: Archäologisches Landesmuseum Baden–Württemberg (Hrsg.), Die Alamannen. Stuttgart 1997. 261–268. WINDLER, R., ET AL. (2005): Frühmittelalter. Die Schweiz vom Paläolithikum bis zum frühen Mittelalter, Vom Neandertaler bis zu Karl dem Grossen. SPM 6. Basel 2005. Fabian Link Wasgenring 125 4055 Basel Tel.: 00 41/76/4 29 02 44 E–Mail: [email protected] aktuell 96 Virtuelle Rekonstruktion und stereolithographisches Modell eines jungneolithischen Schädelfundes aus der Blätterhöhle in Hagen, Nordrhein-Westfalen Jörg Orschiedt, Flora Gröning und Thorsten M. Buzug Zusammenfassung – Die Anwendung neuer computergestützter Verfahren wie der Computertomographie und 3D-Bildbearbeitung eröffnet neue Möglichkeiten in der Rekonstruktion fragmentarischer Skelettfunde. Gegenüber herkömmlichen Methoden sind diese Verfahren nichtinvasiv und damit zerstörungsfrei. Im Folgenden wird die virtuelle Rekonstruktion eines jungneolithischen Schädels aus der Blätterhöhle bei Hagen, Nordrhein-Westfalen beschrieben. Von diesem Schädel sind Fragmente des Calvariums und des Gesichtsschädels vorhanden. Durch die virtuelle Spiegelung der vorhandenen Stücke konnte der Schädel fast vollständig rekonstruiert werden. Auf der Basis dieser virtuellen Rekonstruktion wurde anschließend ein stereolithographisches Modell erstellt. Die virtuelle Rekonstruktion dieses Fundes bietet nicht nur Möglichkeiten für die weitere wissenschaftliche Bearbeitung des Fundes, sondern verbessert vor allem die Attraktivität und Anschaulichkeit des Fundes in der musealen Präsentation. Schlüsselwörter – Virtuelle Rekonstruktion, Stereolithographie, Jungneolithikum, menschlicher Schädel Abstract – The application of new computer-based techniques like computed tomography and 3D image processing provides new possibilities for the reconstruction of fragmentary skeletal remains. In contrast to conventional methods, these techniques are non-invasive and thus non-destructive. In this article, the virtual reconstruction of a Late Neolithic cranium from the cave site Blätterhöhle close to Hagen, North Rhine-Westphalia is described. Neurocranial as well as facial fragments of this skull are preserved. The cranium could be almost completely reconstructed by mirror imaging of the preserved parts. On the basis of this virtual reconstruction, a stereolithographic model has been produced. The virtual reconstruction of this find provides not only possibilities for the additional scientific examination of the cranium, but also makes the find more attractive and comprehensible for museum visitors. Keywords – virtual reconstruction, stereolithography, Late Neolithic, human skull Fundkontext Die Blätterhöhle befindet sich im „Weißenstein“, einem 189,4 m ü. NN hohen Kalkmassiv im Tal der Lenne, einem Seitenfluss der Ruhr, in unmittelbarer Nähe der Ortschaft Holthausen, innerhalb des Stadtgebietes von Hagen. Der Eingang der Blätterhöhle am Südhang dieser Felsformation liegt unmittelbar an der Mündung eines engen Seitentals der Lenne. Die weißen Kalkfelsen bilden eine weithin sichtbare Landmarke im unteren Lennetal. Sie formen den Anfang eines sich von Hohenlimburg nach Süden zu einem tiefen Gebirgstal verengenden Flussbereichs der Lenne. Am „Weißenstein“ öffnet sich das Lennetal zu einer weiten Terrassenlandschaft, die im Norden durch das Ruhrtal und die südlichen Ausläufer des Ardeygebirges sowie von dem beherrschenden Syberg (Hohensyburg) abgeschlossen wird. Am östlichen Fuß des Weißensteins befindet sich eine Karstquelle, der „Barmer Teich“, die den Großteil des Karstgebietes entwässert. Seit den 1920’er Jahren ist die Umgebung des „Weißenstein“ durch verschiedene archäologische Funde bekannt. In verschiedenen Höhlen und auf Äckern der Umgebung wurden in der Vergangenheit paläolithische, mesolithische und neolithische Artefakte, bronze- und eisenzeitliche Trachtbestandteile und Waffen sowie mittelalterliche Keramik entdeckt. Im Rahmen einer hydrologischen Untersuchung, die im Auftrag der Stadt Hagen durchgeführt wurde, konnte die Blätterhöhle im April 2004 erstmalig speleologisch durch den Arbeitskreis Kluterthöhle e.V. untersucht werden. Dabei wurde die Höhle als Kriechgang begehbar gemacht und bis auf eine Länge von 65 m vermessen. Die Höhle selbst besteht in ihrer heute bekannten Form aus einem schräg nach unten führenden, ca. 10 m langen, röhrenförmigen Schacht von ca. 60 x 70 cm Durchmesser, der in einem waagerechten gewunden Kriechgang mündet. Die Höhle ist bis auf den hinteren Bereich nur kriechend zu befahren. Teilweise liegen Engstellen vor, die einem Fall auch nur auf der Seite liegend durchquert werden können. Bei der Ausräumung des Kriechganges konnte eine Vielzahl menschlicher und tierischer Skelettreste, die in ungeordneter Fundsituation angetroffen wurden, geborgen werden. Aufgrund dieser Funde wurde auf eine weitere Ausräumung von Sediment verzichtet. Da an keiner Stelle der gewachsene Felsboden erreicht ist, kann erwartet werden, dass der ursprüngliche Zugang deutlich größer gewesen sein muss. Die Fundstelle wurde nach der Entdeckung der archäologischen Funde durch ein alarmgesicher- Archäologische Informationen 30/1, 2007, 35-41 Bulletin de la Société Suisse35d‘Anthropologie 13 (1), 2007 Bulletin der Schweizerischen Gesellschaft für Anthropologie 13 (1), 2007 Jörg Orschiedt, Flora Gröning und Thorsten M. Buzug tes Tor verschlossen. Seit 2006 wird sowohl die Höhle selbst als auch der Vorplatz archäologisch untersucht. In den bislang zwei Grabungskampagnen wurden weitere Funde menschlicher und tierischer Reste sowie Steingeräte und Keramikfragmente geborgen. Ein Vorbericht über erste Ergebnisse zur Fundstelle befindet sich derzeit im Druck (ORSCHIEDT et al. im Druck). diglich isolierte menschliche Reste vorliegen. Insgesamt lassen sich zwei Kinder der Altersstufen infans I und II, eine juvenile Frau, ein erwachsener Mann, eine erwachsene Frau und ein nicht geschlechtsbestimmtes erwachsenes Individuum nachweisen. Datierung Es liegen keine stratigraphischen Beobachtungen vor, die für eine zeitliche Einordnung der Knochenfunde aus dem Höhleninneren nutzbar wären. Da die Schichten, in denen die Funde gemacht wurden, zum größten Teil gestört waren, wie bei der Nachgrabung ermittelt werden konnte, kann bisher nur auf 14C-Datierungen zurückgegriffen werden. Unmittelbar nach der Entdeckung der Skelettreste wurde eine AMS-Datierung eines Schädeldaches veranlasst. Diese Messung ergab ein frühmesolithisches Alter des Schädels. In der Folge wurden weitere menschliche Skelettreste in den 14C-Laboratorien von Kiel und Oxford datiert. Die Untersuchung des Kollagengehaltes der Knochen deutet auf eine sehr gute bis gute Erhaltung hin. Die Ergebnisse lassen erkennen, dass die menschlichen Reste aus der Blätterhöhle aus zwei verschiedenen Zeitphasen stammen, dem Frühmesolithikum und dem Jungneolithikum. Unter den datierten menschlichen Resten befindet sich auch ein fragmentiertes menschliches Calvarium (04/007), dessen Datierung ein Alter von 4833 ± 37 BP (3610 ± 60 calBC, OxA-14464), bzw. 4835 ± 30 BP (3610 ± 50 calBC, KIA-26263a) ergab. Weitere Datierungen menschlicher Reste fallen ebenfalls in einen jungneolithischen Zeitabschnitt zwischen 3.600 und 3.000 calBC. Menschenreste Menschliche Reste wurden in einem Bereich unmittelbar hinter der Mündung des Schachtes im Bereich des waagerechten Kriechganges entdeckt. Anatomische Zusammenhänge konnten weder bei den menschlichen, noch bei den tierischen Skelettresten beobachtet werden. Die bei der Bergung gemachte Beobachtung einer verstreuten Lage der Skelettreste sowie das teilweise sehr lose Sediment ist mit einer Störung der ursprünglichen Lage durch grabende Tiere in Verbindung zu bringen. Die in den Faunenresten sehr häufig vertretenen Dachse dürften zu dieser Störung beigetragen haben. Die menschlichen Skelettreste stellen die größte Fundgattung im bislang geborgenen archäologischen Material dar. Insgesamt liegen ca. 200 menschliche Reste vor, die in der Mehrzahl bei der Ausräumung des Kriechganges entdeckt wurden aber auch bei den seit 2006 stattfindenden Grabungen in der Höhle und auf dem Vorplatz geborgen wurden. Bisher liegen Skelettreste von mindestens sieben menschlichen Individuen vor (ORSCHIEDT/GRÖNING 2007). Die Erhaltung der Skelettreste kann generell als gut bis sehr gut bezeichnet werden. Einige Skelettreste weisen sowohl alte als auch frische Brüche auf, die auf die Bergung zurückzuführen sind. Die gute qualitative und quantitative Erhaltung ist vor allem auf die Beschaffenheit der weitgehend trockenen Kalkhöhle zurückzuführen. Innerhalb des Skelettmaterials sind alle Regionen des menschlichen Skelettes vertreten. Allerdings sind kleinere Skelettreste wie Hand- und Fußknochen, Wirbel und Rippenfragmente deutlich unterrepräsentiert. Einige dieser kleineren Skelettelemente konnten jedoch aus den Faunenresten und beim Schlämmen des Sedimentes ausgelesen werden. Die Alters- und Geschlechtsbestimmung wurde nach den üblichen anthropologischen Standards durchgeführt (FEREMBACH et al. 1979; RÖSING et al. 2005; SJØVOLD 1988; SZILVÁSSY 1988). Die Bestimmung wird durch die Tatsache erschwert, dass keine vollständigen Individuen, sondern le- aktuell Anthropologische Bearbeitung des Schädels Das jungneolithische Calvarium wurde aus zahlreichen Fragmenten rekonstruiert und weist wie die übrigen Skelettreste aus der Blätterhöhle einen sehr guten qualitativen Erhaltungszustand auf. Es lässt nur einige Beschädigungen erkennen, die aufgrund der hellen Bruchränder eindeutig mit der Bergung bzw. mit der unmittelbaren Entdeckung des Stückes in Zusammenhang stehen. Defekte Bereiche sind im Bereich des linken Os parietale und des rechten Os frontale zu erkennen (Abb. 1). Neben dem Calvarium ist dem Individuum ein Fragment des Gesichtsschädels zuzuordnen. Bei diesem Fragment sind die linke Gesichtshälfte 36 Virtuelle Rekonstruktion und stereolithographisches Modell eines jungneolithischen Schädelfundes ... einem grazilen Erscheinungsbild die Bestimmung als weibliches Individuum. Computertomographische Erfassung des Schädels Bei der Computertomographie (CT) handelt es sich um ein Verfahren zur Erzeugung von Schnittbildern eines Objektes mit Hilfe von Röntgenstrahlen. Ein wesentlicher Vorteil der Computertomographie gegenüber herkömmlichen Röntgenverfahren besteht darin, dass das Bild nicht auf eine zweidimensionale Fläche projiziert wird, sondern schichtweise und überlagerungsfrei in eine dreidimensionale Matrix rekonstruiert wird (BUZUG 2004). Auf der Basis des resultierenden Bildstapels kann daher anschließend eine dreidimensionale Rekonstruktion, eine sogenannte sekundäre Rekonstruktion, des gescannten Objektes erstellt werden. Die Schädelreste des spätjuvenilen Individuums wurden mit einem Philips CT Secura Scanner vermessen (Abb. 2). Hierfür wurde eine Spannung von 100 kV und eine Stromstärke von 80 mA verwendet. Das Calvarium und der Gesichtschädel wurden einzeln eingescannt. Vom Calvarium wurden koronale Schichtbilder aufgenommen, vom Gesichtsschädel axiale. Der Schichtabstand betrug in beiden Scans 2 mm. Im Falle des Calvariums wurde eine Pixelgröße der Schichtbilder von 0,34 mm rekonstruiert. Im Falle des Gesichtsschädels lag die Pixelgröße bei 0,23 mm. Abb. 1 Schädel der jungen Frau aus der Blätterhöhle nach Zusammensetzung der vorhandenen Fragmente (Orschiedt/Gröning) sowie beide Hälften der Maxilla fast komplett erhalten. Das Os zygomaticum befindet sich jedoch nicht mehr in seiner ursprünglichen Position sondern ist nach posterior gedreht und verschoben. Im Bereich des rechten Os frontale liegt ein isoliertes Fragment des rechten Orbitadaches vor, das nicht direkt angepasst werden konnte. Das Schädelfragment wurde aus zahlreichen Fragmenten rekonstruiert und weist wie die übrigen Skelettreste aus der Blätterhöhle einen sehr guten qualitativen Erhaltungszustand auf. Die Schädelfragmente repräsentieren ein juveniles Individuum, da die Sphenobasilarfuge noch nicht verschlossen ist, was üblicherweise bis zum ca. 23. Lebensjahr erfolgt, und der 3. Molar des Oberkiefers noch nicht durchgebrochen ist (FEREMBACH et al. 1979; RÖSING 2005; SZILVÁSSY 1988). Das Geschlecht kann anhand der vorliegenden Schädelmerkmale übereinstimmend als weiblich bestimmt werden. So liegen neben einer gering ausgeprägten Glabella und dem Arcus superciliarus, einem scharfkantigen Margo supraorbitale, deutliche Tuber parietale sowie ein schwach ausgeprägter Processus mastoideus vor (FEREMBACH et al. 1979; RÖSING 2005; SJØVOLD 1988). Die Bestimmung wird durch die Messung des Austrittswinkels des Meatus acusticus internus in der Pars petrosa von ca. 90° bestätigt (GRAW et al. 2005). Die Ansatzstellen der Muskulatur im Bereich des Os parietale und Os occipitale sind nur schwach ausgeprägt. Dies unterstützt ebenfalls zusammen mit Virtuelle Rekonstruktion Die virtuelle Rekonstruktion fragmentarischer Funde gewinnt in der Anthropologie zunehmend an Bedeutung. Sie bietet zwei wesentliche Vorteile gegenüber konventionellen Verfahren: Sie ist nicht-invasiv und reversibel. Bei einer herkömmlichen Rekonstruktion ist es schwierig, die verklebten Teile wieder voneinander zu lösen ohne den Fund zu beschädigen. Eine virtuelle Rekonstruktion erlaubt dagegen, das Stück beliebig oft zusammenzusetzen und wieder zu zerlegen. In den vergangenen Jahren wurden zahlreiche fragmentarische Fossilfunde virtuell rekonstruiert (z. B. KALVIN et al. 1995; ZOLLIKOFER et al. 1995; THOMPSON/ILLERHAUS 1998; GUNZ et al. 2004). Ein gängiges Verfahren ist dabei das Ersetzen fehlender Fragmente durch das Spiegeln vorhandener Teile. Bei dem jungneolithischen Schädel aus der Blätterhöhle ist die linke Gesichtshälfte komplett 37 aktuell Jörg Orschiedt, Flora Gröning und Thorsten M. Buzug Abb. 3 Screenshot der virtuellen Rekonstruktion mit Ergänzung der fehlenden anatomischen Bereiche (Gröning). Gespiegelte Fragmente: grau, ungespiegelte Fragmente: violett Abb. 2 Einrichten der Vermessung des Schädels mit einem Philips CT Secura Scanner (Orschiedt) re Rekonstruktionsmethode darstellt, da das menschliche Skelett im wesentlichen bilateral symmetrisch ist, kann es im Detail Probleme geben, da einzelne Knochen durchaus auf der rechten und linken Körperhälfte eine unterschiedliche Größe oder Gestalt besitzen können. Auch in der hier beschriebenen Rekonstruktion war es an einigen Stellen nicht möglich die Fragmente perfekt aneinander zu passen. Es wurden dann jeweils die bestmöglichen Anpassungen gewählt und die Kanten manuell korrigiert. Eine besondere Behandlung erforderte das Os zygomaticum. Durch eine Beschädigung ist es im Orginalfund nicht mehr in seiner ursprünglichen Position, sondern leicht nach posterior gedreht und verschoben. Um diese Beschädigung zu korrigieren, wurde das Os zygomaticum als ein separates Objekt segmentiert und anschließend durch manuelles Drehen und Verschieben in seine ursprüngliche, anatomisch korrekte Position gebracht. Die virtuelle Anpassung der einzelnen Fragmente geschah auf der Basis von Abbildungen kompletter Schädel in verschiedenen Ansichten, dem Verlauf der Schädelnähte und Messungen, z. B. des Durchmessers des Foramen magnums. Im Allgemeinen ist das dreidimensionale Zusammensetzen von Fragmenten auf einem zweidimensionalen Computerbildschirm keine einfache Aufgabe, da die dreidimensionale Position der einzelnen Objekte nur unzureichend wiedergegeben wird. Dies kann durch eine stereoskopische Darstellung verbessert werden. Die vorläufige Rekonstruktion wurde daher im HIVE-Zentrum der vorhanden. Darüber hinaus liegt auch die rechte Hälfte des Calvariums fast vollständig vor. Der Fund bietet sich daher besonders für eine Rekonstruktion durch Spiegelung erhaltener Teile an. Vor der eigentlichen Rekonstruktion wurden daher die beide Scans des Calvariums und des Gesichtsschädels gespiegelt, sodass insgesamt vier Datensätze zur Verfügung standen. Die Spiegelung wurde mit Hilfe der freien Bildbearbeitungssoftware ImageJ durchgeführt. Die eigentliche Rekonstruktion erfolgte mit der 3D-Bildbearbeitungssoftware AMIRA 3.1.1®. Im ersten Schritt wurden die Schichtbilddatensätze eingelesen und knöcherne Strukturen von der umgebenden Luft durch das Festlegen eines minimalen Dichtewertes getrennt. Auf diese Weise wurde von jedem Fragment und seiner gespiegelten Version ein 3D-Oberflächenmodell erstellt. Anschließend wurden die Fragmente durch manuelles Drehen und Verschieben aneinander angepasst (Abb. 3). Da die Fragmente an einigen Stellen über die Mediansagittale hinaus reichten, entstanden beim Zusammensetzen einige Überlagerungen. Diese wurden manuell korrigiert, indem gespiegelte Bereiche, die die Originalstücke überlappten, entfernt wurden. Dies betrifft vor allen Dingen die Maxilla. Da sie fast vollständig erhalten ist, wurde darauf verzichtet, eine Spiegelung entlang der Mediansagittalen vorzunehmen. Obleich die Spiegelung vorhandener Fragmente desselben Individuums eine sehr siche- aktuell 38 Virtuelle Rekonstruktion und stereolithographisches Modell eines jungneolithischen Schädelfundes ... Universität Hull (HIVE - Hull Immersive Visualization Environment), das über einen interaktiven Virtual Reality Room verfügt, überprüft. Hierbei wurden einige Bereiche entdeckt, in denen die Fragmente nicht optimal aneinander passten, was anschließend korrigiert wurde. Der letzte Schritt der Rekonstruktion bestand darin, die einzelnen Oberflächenmodelle zu einem einzigen Oberflächenmodell zusammenzufassen. weile wurde es jedoch schon mehrfach angewendet, um Repliken von archäologischen Funden und Fossilien zu erstellen (ZUR NEDDEN et al. 1994, ZOLLIKOFER et al. 1995, SEIDLER et al. 1997). Um eine Stereolithographie des rekonstruierten neolithischen Schädels erstellen zu können, war es erforderlich, die Daten weiter zu bearbeiten. Mit der freien Software MeshLab wurde das Modell von sich überschneidenden oder isolierten Flächen bereinigt und anschließend geglättet. Eine solche Bereinigung ist für den Aufbau eines stereolithographischen Modells zwingend erforderlich. Darüber hinaus wurde mit der Glättung eine deutliche Reduzierung der Dateigröße ereicht. Das endgültige Modell wurde schließlich als STL-Datei exportiert. Das Stereolithographische Modell wurde von der österreichischen Firma Zumtobel-Werkzeugbau mit Hilfe einer Stereolithographieanlage des Typs SLA 7000 erstellt. Die Schichtdicke betrug dabei 0,1 mm und als Material wurde Epoxydharz verwendet (Abb. 4). Erstellung des stereolithographischen Modells Um von einer virtuellen Rekonstruktion ein greifbares Modell zu erstellen, bietet sich die Technik der Stereolithographie an. Bei diesem Verfahren wird ein zunächst flüssiger Kunststoff mit Hilfe eines Lasers schichtweise ausgehärtet. Die Lenkung des Lasers erfolgt computergesteuert auf der Basis des virtuellen Modells und ist bei hoch auflösenden Geräten auf Mikrometer genau. Ursprünglich wurde dieses Verfahren für die Herstellung von Prototypen in der Industrie entwickelt. Mittler- Abb. 4 Die fertig gestellte Stereolithographie des Schädels (Orschiedt) 39 aktuell Jörg Orschiedt, Flora Gröning und Thorsten M. Buzug Fazit Literatur Nach den ersten Grabungsergebnissen sprechen die Funde für eine Nutzung des Vorplatzes und der Blätterhöhle während des ausgehenden Spätpaläolithikums, älteren Frühmesolithikums sowie während des Jungneolithikums. In den letzten beiden Zeitperioden ist die Höhle als Bestattungsplatz genutzt worden. Parallelen für die Niederlegung menschlicher Körper in Höhlen in der Zeit zwischen 3.600 und 3.000 calBC. liegen unter anderem aus dem benachbarten Belgien vor (ORSCHIEDT et al. im Druck). Der Einsatz computergestützter Verfahren hat sich bei der Rekonstruktion des spätjuvenilen neolithischen Schädels als äußerst lohnenswert erwiesen. Aus den beiden Fragmenten, die zuvor nicht aneinander passten, ließ sich allein durch die Spiegelung vorhandener Reste ein fast vollständiger Schädel rekonstruieren. Dies ermöglicht z. B. metrische Vergleiche mit anderen Funden, die vorher durch den fragmentarischen Zustand nur eingeschränkt möglich waren. Neben dem Nutzen für eine weitere wissenschaftliche Bearbeitung bietet die virtuelle Rekonstruktion und Erstellung eines stereolithographischen Modells vor allen Dingen Vorteile für die museale Präsentation des Fundes. Der zuvor fragmentarische Fund kann nun dem Besucher als fast vollständiger Schädel präsentiert werden, was den Fund für die Besucher attraktiver und anschaulicher macht. Das stereolithographische Modell wird derzeit in der Dauerausstellung des Museums für Ur- und Frühgeschichte Wasserschloss Werdringen zusammen mit dem Originalfund gezeigt. Es ist beabsichtigt auf der Basis des Modells eine Gesichtsrekonstruktion der Person anfertigen zu lassen. BUZUG, T. M. (2004): Einführung in die Computertomographie. Mathematisch-physikalische Grundlagen der Bildrekonstruktion. Berlin, Heidelberg 2004. FEREMBACH, D./SCHWIDETZKY, I./M. STLOUKAL (1979): Empfehlungen für die Alters- und Geschlechtsdiagnose am Skelett. Homo 30, 1979, 1–32. GRAW, M./WAHL, J./M. AHLBRECHT (2005): Course of the meatus acusticus internus as criterion for sex differentiantion. For. Scie. Internat. 147, 2005, 113–117. GUNZ, P./MITTEROECKER, P./BOOKSTEIN, F. L./G. W. WE(2004): Computer aided reconstruction of incomplete human crania using statistical and geometrical estimation methods. BAR Int. Series 1227. Oxford 2004, 92–94. BER KALVIN, A. D./DEAN, D./J.-J. HUBLIN (1995): Reconstruction of human fossils. IEEE Comp. Graphics Applications 15, 1995, 12–15. ZUR NEDDEN, D. /KNAPP, R. /WICKE, K./JUDMAIER, W./ MURPHY, W. A. /SEIDLER, H./W. PLATZER (1994): Skull of a 5,300-year-old mummy: reproduction and investigation with CT-guided stereolithography. Radiology 193, 1994, 269–272. ORSCHIEDT, J./F. GRÖNING (2007): Die menschlichen Skelettreste aus der Blätterhöhle, Stadt Hagen, In: ANDRASCHKO, F./KRAUS, B. /B. MELLER, (Hrsg.): Archäologie zwischen Befund und Rekonstruktion. Ansprache und Anschaulichkeit. Festschrift für Prof. Dr. Renate Rolle zum 65. Geburtstag. Hamburg 2007, 349–361. ORSCHIEDT, J./KEGLER, J./GEHLEN, B./SCHÖN, W./F. GRÖNING (im Druck): Die Blätterhöhle in HagenHolthausen (Westfalen). Vorbericht über die ersten archäologischen Untersuchungen. Arch. Korrbl. (im Druck). Danksagung RÖSING, F. W./GRAW, M./MARRÉ, B./RITZ-TIMME, S./ROTHSCHILD, M. A./RÖTZSCHER, K./SCHMELING, A./SCHRÖDER, I./G. GESERICK (2005): Arbeitsgemeinschaft für Forensische Altersdiagnostik der Deutschen Gesellschaft für Rechtsmedizin. Empfehlungen für die forensische Geschlechts- und Altersdiagnose am Skelett. Rechtsmedizin 15, 2005, 32–38. Wir danken dem HIVE-Zentrum der Universität Hull für die Nutzung des Virtual Reality Rooms, die eine wesentliche Verbesserung der Rekonstruktion ermöglichte. Ebenso möchten wir uns bei Herrn Bruno Kuen von der Firma ZumtobelWerkzeugbau für die gute Zusammenarbeit bedanken. aktuell SEIDLER, H./FALK, D./STRINGER, C./WILFING, H./MÜLLER, G. B./ZUR NEDDEN, D./WEBER, G. W./RECHEIS, W./J.-L. ARSUAGA (1997): A comparative study of stereolithographically modelled skulls of Petralona and Broken Hill: implications for future studies of middle Pleistocene hominid evolution. Jour. Hum. Evol. 33, 1997, 691–703. 40 Virtuelle Rekonstruktion und stereolithographisches Modell eines jungneolithischen Schädelfundes ... SJØVOLD, T. (1988): Geschlechtsdiagnose am Skelett. In: KNUSSMANN, R. (Hrsg.): Vergleichende Biologie des Menschen. Lehrbuch der Anthropologie und Humangenetik. Stuttgart 1988, 444–480. SZILVÁSSY, J. (1988): Alterdiagnose am Skelett. In: KNUSSMANN, R. (Hrsg.): Vergleichende Biologie des Menschen. Lehrbuch der Anthropologie und Humangenetik Stuttgart 1988, 421–443. THOMPSON, J. L./B. ILLERHAUS (1998): A new reconstruction of the Le Moustier 1 skull and investigation of internal structures using 3-D-µCT data. Jour. Hum. Evol. 35, 1998, 647–665. ZOLLIKOFER, C. P. E./PONCE DE LEÓN, M. S./MARTIN, R. D./ P. STUCKI (1995): Neanderthal computer skulls. Nature 375, 1995, 283–285. PD. Dr. Jörg Orschiedt Historisches Centrum Hagen Eilper Straße 71-75 58091 Hagen [email protected] Flora Gröning M.A. PALAEO Biology (S-Block) University of York PO Box 373 York YO10 5YW fl[email protected] Prof. Dr. Thorsten M. Buzug Institut für Medizintechnik Universität zu Lübeck Ratzeburger Allee 160 23538 Lübeck [email protected] 41 aktuell Körperhöhenschätzung an ausgewählten neolithischen und bronzezeitlichen Skeletten Katrin Schmidt, Ronny Bindl und Horst Bruchhaus Zusammenfassung – In dieser Studie wurde versucht, erste Hinweise zur Verwendbarkeit verschiedener Körperhöhenschätzmethoden ausgehend von einer kleinen Serie (n = 10) vollständig erhaltener spätneolithischer und frühbronzezeitlicher Skelette zu finden. Als Bezugsgröße diente die Körperhöhe nach FULLY und PINEAU (1960). Bei Schätzung der Körperhöhe aus den Gesamtknochenlängen eigneten sich die Formeln von PEARSON (1899) und TELKKÄ (1950) am besten. Die höchsten Abweichungen lieferten die Regressionsformeln von ROTHER ET AL. (1971, 1973, 1978) DUPERTUIS und HADDEN (1951) sowie BREITINGER (1937) und BACH (1965). Mittels Körperhöhenschätzung aus Langknochenfragmenten konnten ebenso brauchbare Ergebnisse erzielt werden, vorausgesetzt die Teilstrecken waren ausreichend lang und die Messpunkte sicher und leicht zu identifizieren. Hierbei wurden zwei Möglichkeiten zur Rekonstruktion der Körperhöhe aus Langknochenfragmenten angewendet. Bei der indirekten Methode (u. a. nach STEELE/MCKERN 1969; JACOBS 1992; WRIGHT ET AL. 2003), wurde aus den Abschnitten zunächst die Knochenlänge bestimmt, bevor aus dieser Knochenlänge die Körperhöhe mittels gut bekannter Regressionsformeln rekonstruieret werden konnte. Für die hier untersuchten Langknochen Humerus, Femur und Tibia lieferten die Regressionsgleichungen von WRIGHT ET AL. (2003) die geringsten Abweichung von der gemessenen Knochenlänge. Bei anschließender Rekonstruktion der Körperhöhe aus den berechneten Knochenlängen zeigten sich ähnliche genaue Ergebnisse wie bei Schätzung der Körperhöhe aus gemessenen Langknochenlängen. Die am Beispiel des Femur durchgeführte direkte Schätzung der Körperhöhe aus Fragmentmaßen (SIMMONS ET AL. 1990) schien weniger geeignet als die indirekte Methode. Schlüsselwörter – Körperhöhenschätzung, Langknochenfragmente, Neolithikum, Bronzezeit Abstract – In this study, first indications regarding the applicability of several stature estimation methods were acquired using a small sample of well-preserved skeletons (n = 10) from the later Neolithic and early Bronze-age. The anatomical body height based on the method of FULLY and PINEAU (1960) was therefor used as reference value. After calculating the stature from measured complete long bone lenghts, it could be shown that the equations of PEARSON (1899) and TELKKÄ (1950) yield to the best results when compared with the anatomical body height. The highest deviations were derived using the equations of ROTHER ET AL. (1971, 1973, 1978), DUPERTUIS and HADDEN (1951) as well as BREITINGER (1937) and BACH (1965). To estimate the body height from long bone fragments, a direct (SIMMONS ET AL. 1990) and an indirect (i.a. STEELE/MCKERN 1969; JACOBS 1992; WRIGHT ET AL. 2003) approach were tested. When applying the indirect method, the long bone lenght was initially estimated from the lenght of the fragments, before stature could be calculated from this estimated long bone length via well-known regression equations. For the here investigated long bones humerus, femur and tibia, the equations of WRIGHT ET AL. (2003) provide the lowest deviations from the measured long bone lengths. The following reconstruction of stature from the calculated long bone lenghts lead to similar accurate results as the estimation of body height from the measured complete long bone lenghts. On the example of the femur, the direct approach of stature estimation from long bone fragments was performed and it could be shown, that this method seems less suitable than the indirect approach. Keywords – stature estimation, fragmentary long bones, neolithic, Bronze-age Einleitung und Zielstellung In der Paläanthropologie spielt die Interpretation von Skelettmaterial hinsichtlich Größe und Gestalt eines Individuums eine wichtige Rolle, da die mittels Körperhöhenschätzung gewonnenen Individualdaten einen Beitrag zu den Lebensumständen eines Individuums liefern können In der Literatur werden zwei Grundprinzipien zur Körperhöhenschätzung beschrieben: Zum einen ist dies die so genannte „anatomische Methode“, welche die Körperhöhe aus der Summe der Maße verschiedener Skelettelemente rekonstruiert und sich in erster Linie auf die Untersuchungen von FULLY (1956) sowie FULLY und PINEAU (1960) stützt. Die Autoren entwickelten für die Berechnung der Skeletthöhe Gleichungen, denen die Summe dieser Skelettelemente zugrunde liegt und geben zusätzlich einen entsprechenden Korrekturfaktor für die Weichteilkomponente (Zwischenwirbelscheiben etc.) an. Somit ist es wichtig, dass bei Schätzung der Körperhöhe nach der „anatomischen Methode“ alle die Skelettelemente vorhanden und möglichst intakt sind, welche einen Beitrag zur Körperhöhe des Individuums leisten. Dies ist in der Regel nur bei wenigen Skeletten der Fall. Der Vorteil dieser Methode liegt darin, dass mit ihrer Hilfe die Körperhöhe sehr exakt geschätzt werden kann, da die Formeln relativ unabhängig von den Proportionsverhältnissen im Skelett sind und damit weder durch das Geschlecht noch durch das Al- Archäologische Informationen 30/1, 2007, 51-69 Bulletin de la Société Suisse51d‘Anthropologie 13 (1), 2007 Bulletin der Schweizerischen Gesellschaft für Anthropologie 13 (1), 2007 Katrin Schmidt, Ronny Bindl und Horst Bruchhaus ter oder die Verschiedenheit zwischen Populationen beeinflusst werden können. Infolgedessen kann die „anatomische Körperhöhe“ vor allem bei (prä)historischen Skeletten als tatsächliche Körperhöhe angenommen werden und damit als Basis für verschiedene Berechnungen und Vergleiche dienen. Das zweite Prinzip zur Schätzung der Körperhöhe beruht auf der linearen Proportionalität von Knochenlänge und Körperhöhe. Hierbei wird versucht, mittels bevölkerungs- und geschlechtsspezifischer Regressionsgleichungen von der Gesamtlänge einzelner Langknochen auf die Körperhöhe zu schließen. Demgemäß wird diese Vorgehensweise auch als „mathematische Methode“ bezeichnet (u. a. PEARSON 1899; TELKKÄ 1950; BREITINGER 1935 und BACH 1967; DUPERTUIS/HADDEN 1951; TROTTER/GLESER 1952, 1958, DE MENDONCA 2000). Liegen die Langknochen fragmentiert vor, so lässt sich auch aus Knochenfragmenten die Körperhöhe auf mathematischem Weg rekonstruieren. Hierzu gibt es zwei unterschiedliche Ansatzweisen. So kann die Körperhöhe, ähnlich wie bei Vorhandensein intakter Langknochen, direkt aus Abschnittsmaßen rekonstruiert werden, nur das die Berechnung der Körperhöhe nicht von der Gesamtlänge des Knochens sondern von dem entsprechenden Teilmaß ausgeht. Diese Methode wird als direkte Methode der Körperhöhenschätzung aus Langknochenfragmenten bezeichnet, allerdings finden sich in der Literatur hierzu nur wenige Regressionsgleichungen. (SIMMONS ET AL. 1990, HOLLAND 1992, CHIBBA/BIDMOS 2006). Der zweite Weg, nämlich die indirekte Körperhöhenschätzung aus Extremitätenknochenfragmenten, erweist sich als sehr komplex und birgt einen hohen Fehler wegen des kumulativen Effektes zweier Standardfehler, da zunächst die Langknochenlänge aus der Fragmentlänge und anschließend die Körperhöhe aus der rekonstruierten Langknochenlänge geschätzt werden muss. Ferner ergeben sich oft Schwierigkeiten bei der genauen Definition der Messpunkte. Studien zur Schätzung der Knochenlänge und/oder Körperhöhe aus Langknochenfragmenten existieren u. a. für Maße an Radius, Ulna (MÜLLER 1935; GRALLA/FUDALI 1973) und Fibula (WRIGHT ET AL. 2003), die meisten Untersuchungen konzentrieren sich allerdings auf Abschnittsmaße an Humerus, Femur und Tibia (STEELE/MCKERN 1969; SONDER/KNUSSMANN 1985; SIMMONS ET AL. 1990; JACOBS 1992; WRIGHT ET AL. 2003). Die untersuchten Längenteilstrecken der verschiedenen Autoren sind durch markante Struk- aktuell turen am Knochen abgegrenzt und beinhalten verschiedene Punkte entlang der Epiphyse und Diaphyse. Bei diesen Strukturen kann es sich beispielsweise um Muskelansatzstellen oder um durch Gelenkarchitektur bedingte Segmentgrenzen handeln. Andere Autoren verwenden zur Schätzung der Knochenlänge „Standardmaße“ (MARTIN 1928; MARTIN/SALLER 1957 bzw. MARTIN/BREUER 1988), welche kurze Teilstrecken und Breiten- bzw. Umfangsmaße umfassen (SIMMONS ET AL. 1990; FUNKE 2006) oder sowohl „Standardmaße“ als auch Längenteilstrecken (SONDER/KNUSSMANN 1985) beinhalten. In dieser Arbeit soll der Schwerpunkt auf die drei wichtigsten zur Körperhöhenschätzung geeigneten Langknochen, d. h. Humerus Femur und Tibia, gelegt werden. Hierbei gilt es, die Eignung verschiedener Fragmentmaße zur Körperhöhenschätzung zu überprüfen. Zunächst wurde bei einer kleinen Serie von Skeletten unterschiedlicher Zeitstellung (Glokkenbecherkultur, Linienbandkeramik, Schnurkeramik, Aunjetitzer Kultur) die Körperhöhe nach FULLY und PINEAU (1960) rekonstruiert. Bei allen untersuchten Skeletten lagen die Skelettelemente, welche zur Körperhöhe beitragen, vollständig vor. Damit waren alle Voraussetzungen für den Vergleich der „anatomischen Körperhöhe“ mit den nach anderen Methoden ermittelten Körperhöhen vorhanden (siehe Teil 1). Anschließend wurden die Gesamtlängen nach MARTIN (1928); MARTIN/SALLER (1957) bzw. MARTIN/BREUER (1988) gemessen (Femur M1/M2; Humerus M1; Tibia: M1a). Aus diesen Messwerten wurde mittels linearer Regressionsgleichungen die Körperhöhe nach verschiedenen Methoden bestimmt (u. a. nach PEARSON 1899; TROTTER/GLESER 1952; BREITINGER 1937 und BACH 1965) (siehe Teil 2). Später erfolgte eine Messung verschiedener Abschnittsmaße, wobei es sich zum einen um Standardmaße und zum anderen um durch andere Autoren definierte Längenteilstrecken handelte. Die Höhe der Abweichungen der mittels Abschnittsmaßen berechneten Knochenlängen von den in Teil 2 gemessenen Knochenlängen geben Hinweise auf die Verwendbarkeit der zur Längenmaßrekonstruktion verwendeten Abschnittsmaße (siehe Teil 3). Mittels dieser unterschiedlichen Methoden, welchen Skelettmaterial verschiedener Zeitstellungen und Populationen zugrunde liegt, wird bei den von uns untersuchten Skeletten die Gesamt- 52 Körperhöhenschätzung an ausgewählten neolithischen und bronzezeitlichen Skeletten länge der Langknochen rekonstruiert. Diese berechneten Langknochenlängen dienen schließlich als Basis für die Berechnung der Körperhöhe. Hierbei kommen wiederum verschiedene bekannte Regressionsformeln (u. a. PEARSON 1899; TELKKÄ 1950; TROTTER/GLESER 1952) zum Einsatz (siehe Teil 4). Als Beispiel für die oben genannte direkte Methode sollen die Regressionsformeln für das Femur nach SIMMONS ET AL. (1990) untersucht und ein Vergleich der nach dieser Methode rekonstruierten Körperhöhe mit der nach der indirekten Methode berechneten Körperhöhe durchgeführt werden, da die Autoren in ihrer Arbeit gleichermaßen Regressionsformeln für die direkte als auch für die indirekte Schätzung der Körperhöhe aus Abschnittsmaßen angeben (siehe Teil 5). Der Vergleich der Körperhöhen, welche wir aus den gemessenen und berechneten Knochenlängen ermitteln konnten, mit den nach FULLY und PINEAU (1960) rekonstruierten Körperhöhen, sind ein anschauliches Beispiel für die Problematik des unkritischen Einsatzes von Methoden (siehe auch REICHELT ET AL. 2003). Tabelle 3 zeigt alle Messpunkte für die Messung der verwendeten Abschnittsmaße an Femora, Tibiae und Humeri. In den Abbildungen 1, 2 und 3 sind die einzelnen Messstrecken an Humerus, Femur und Tibia eingetragen: Grün sind die Martinmaße dargestellt (gemessen bei Sonder und Knussmann 1988 (SK); SIMMONS ET AL. 1990 (SI); FUNKE 2006 (FK)). Rot sind die Abschnitte nach WRIGHT ET AL. 2003 (W) und blau die Abschnitte nach STEELE und MCKERN 1969 (SMK) eingezeichnet. Die Maße von JACOBS 1992 (J) entsprechen den Maßen von STEELE und MCKERN (1969) und wurden daher nicht extra eingetragen. Die Angaben hinter den Abkürzungen SMK/J oder W (z. B. SMK1…SMK5, W0-5, W1-6 usw.) geben die Bezeichnungen der Autoren für die entsprechenden Messstrecken an. Angaben wie SMK 1-2, J 2-3-4 etc. bedeuten, dass es sich um kombinierte Regressionsgleichungen handelt, d. h., dass die Länge verschiedener Teilstrecken für die Berechnung der Knochenlänge verwendet wurde. Bei jedem der 10 Skelette wurden die berechneten Langknochenlängen von den gemessenen Längen subtrahiert und die entsprechenden Abweichungen grafisch dargestellt. Ähnliches gilt für die aus den berechneten Knochenlängen bestimmten Körperhöhen sowie den direkt aus den Abschnittsmaßen rekonstruierten Körperhöhen und deren Abweichungen von der Körperhöhe nach FULLY und PINEAU (1960). Hier sollen aus Platzgründen mittels Boxplots nur die gemittelten Abweichungen für die männlichen und weiblichen Individuen dargestellt werden. Die Boxplots geben Auskunft über Mittelwert, Standardabweichung, den minimalen und maximalen Messwert (Spannweite), zusätzlich werden auch Extremwerte angezeigt. Negative Werte innerhalb der Diagramme indizieren eine Überschätzung der gemessenen Knochenlänge bzw. der Körperhöhe, positive Werte eine Unterschätzung. Die Erstellung der Diagramme erfolgte mit dem Programm SPSS 14.0 for Windows. Material und Methoden Für die Untersuchung standen 10 vollständig erhalten Skelette (4 männliche und 6 weibliche Individuen) aus den in Tabelle 1 aufgeführten spätneolithischen und frühbronzezeitlichen Kulturen zur Verfügung (Altersklassen: frühadult – matur). Zunächst erfolgte bei diesen Skeletten eine Rekonstruktion der Körperhöhe nach der Methode von FULLY und PINEAU (1960). Hierzu werden die Messwerte für Basion-Bregma-Höhe des Craniums (M17), Höhe des Axis (M1a), Wirbelkörperhöhen von Cervikalwirbel 2 bis Sakralwirbel 1 (M1), physiologische Femurlänge (M2), Tibialänge (M1) sowie Höhe von Talus und Calcaneus nach anatomischer Reposition addiert und in die von FULLY und PINEAU (1960) erstellte Gleichung eingesetzt. Anschließend wurden die Gesamtlängen der Langknochen (Femur M1/M2; Humerus M1, Tibia: M1a) sowie die Teilmaße an Humerus, Femur und Tibia gemessen. Die Messung erfolgte für die rechte und linke Körperseite getrennt, für die weiteren Berechnungen wurde allerdings der Mittelwert aus beiden Körperseiten verwendet. War der Knochen einer Körperseite vorhanden, so wird der entsprechende Messwert dieser Seite für die Berechnungen genutzt. 53 aktuell Katrin Schmidt, Ronny Bindl und Horst Bruchhaus Abb. 3 Tibia – Messstrecken. Abb. 1 Humerus – Messstrecken. Teil 2: Schätzung der Körperhöhe aus intakten Langknochen Humerus Bei Anwendung verschiedener Regressionsformeln zur Schätzung der Körperhöhe aus der Länge des Humerus (M1 oder M2) zeigt sich vor allem für die Gleichungen von PEARSON (1899), und DE MENDONCA (2000) eine Unterschätzung der Körperhöhe um bis zu 10 cm, wobei die Formeln von DE MENDONCA (2000) bei Frauen besser geeignet scheinen (geringere Spannbreite der Abweichungen). Die stärkste Unterschätzung der Körperhöhe findet sich bei beiden Geschlechtern bei Verwendung der Formeln von ROTHER ET AL. (1971, 1973, 1978), hier liegen die Abweichungen für Männer bei 5 bis 17 cm, für Frauen 2 bis 13 cm unterhalb der tatsächlichen Körperhöhe. Eine Überschätzung der Körperhöhe um bis zu 7 cm lässt sich bei Anwendung der Formeln für weiße Abb. 2 Femur – Messstrecken. Ergebnisse Teil 1: Schätzung der „anatomischen Körperhöhe“ nach Fully und Pineau (1960) Die entsprechenden „anatomischen Körperhöhen“ für die einzelnen Skelette sind in Tabelle 2 dargestellt, sie werden in dieser Untersuchung wie die tatsächliche Körperhöhe der Individuen gehandhabt und stellen die Grundlage für den Vergleich mit den nach anderen Methoden berechneten Körperhöhen dar. aktuell 54 Körperhöhenschätzung an ausgewählten neolithischen und bronzezeitlichen Skeletten Individuen von DUPERTUIS und HADDEN (1951) finden. Bei männlichen Individuen scheinen die Formeln von TELKKÄ (1950) und OLIVIER ET AL. (1976) am besten geeignet, bei Frauen nur die Formeln von OLIVIER ET AL. (1967), da ihre mittleren Abweichungen die geringsten Unterschiede von der Körperhöhe nach FULLY und PINEAU (1960) aufweisen. Alle anderen Formeln führen zu einer leichten Überschätzung der Körperhöhe. Die Ergebnisse sind in Abb. 4 und 5 grafisch dargestellt. sere Schätzergebnisse als die Formeln für weiße Individuen. Die Formeln von ROTHER ET AL. (1971, 1973, 1978) führen zu einer Unterschätzung der Körperhöhe bei beiden Geschlechtern. Eine hohe Spannbreite der Abweichungen findet sich bei Verwendung der Formeln von BREITINGER (1937) und BACH (1965). Für die männlichen Individuen erweisen sich die Regressionsformeln von PEARSON (1899), TROTTER und GLESER „Blacks“ (1952) und OLIVIER ET AL. (1976) am besten geeignet. Bei den weiblichen Individuen liefert die Formel von PEARSON (1899) die besten Ergebnisse. Die anderen Regressionsformeln führen im Schnitt zu einer Überschätzung der Körperhöhe. Die Ergebnisse sind in Abb. 8 und 9 grafisch dargestellt. Femur Für das Femur (M1 oder M2) zeigen bei beiden Geschlechtern die Gleichungen von BREITINGER (1937) und BACH (1965) eine hohe Spannbreite der Abweichungen. Auch hier führen die Formeln von ROTHER ET AL. (1971, 1973, 1978) zu einer starken Unterschätzung der Körperhöhe, bei Männern um bis zu 7cm, bei Frauen um bis zu 12 cm. Eine hohe Spannbreite der Abweichungen findet sich auch bei Anwendung der Formeln von HAUSER ET AL. (2005) auf weibliche Femora. Recht gute Schätzergebnisse liefern die Formeln von TELKKÄ (1950), hier liegt bei beiden Geschlechtern der mittlere Schätzwert nah an der Körperhöhe nach FULLY und PINEAU (1960). Auch die Formeln von OLIVIER ET AL. (1967) und DE MENDONCA (2000) liefern akzeptable Schätzergebnisse, dies gilt vor allem für männliche Individuen. Im Schnitt erfolgt bei Schätzung der Körperhöhe aus Femurlängen eine leichte Unterschätzung der Körperhöhe, allerdings weniger stark als bei Humerus und Tibia. Die Spannbreite der Abweichungen ist bei weiblichen Individuen höher als bei männlichen Individuen. Die Ergebnisse sind in Abb. 6 und 7 grafisch dargestellt. Teil 3: Schätzung der Knochenlänge aus Abschnittsmaßen Humerus Betrachtet man die Abweichungen der geschätzten Humeruslänge (Abb. 10 und 11) von der größten Länge des Humerus (M1), so zeigt sich für die einzelnen Geschlechter ein unterschiedliches Bild. Bei Anwendung der Regressionsgleichungen von STEELE und MCKERN (1969) zeigen die Frauen nur geringe Abweichungen von der gemessenen Knochenlänge, während hingegen für die Männer hohe Abweichungen mit einer hohen Spannbreite zu beobachten sind. Die stärksten Abweichungen bei den männlichen Individuen kommen bei Regression der Strecken Caput humeri – distalster Punkt der Aufwulstung des Caputrandes (SMK 1) sowie proximaler Rand Fossa olecrani – distaler Rand Fossa olecrani (SMK 4) auf die Knochenlänge M1 zustande. Die Regressionsformeln von WRIGHT ET AL. (2003) führen bei den männlichen Individuen zu einem sehr exakten Schätzergebnis für M1, die Spannbreite der Abweichungen ist gering. Daneben finden sich bei den weiblichen Individuen wesentlich höhere Einzelabweichungen, wobei auch hier die Spannbreite eher gering ist. Frauen zeigen die höchste Abweichung bei der Strecke proximalster Punkt Tuberculum minus – distalster Punkt Trochlea humeri (W0-5). Mäßige bis hohe Abweichungen von M1 sind bei Verwendung des Martinmaßes M3 (obere Epiphysenbreite) bei beiden Geschlechtern zu beobachten. Tibia Bei Schätzung der Körperhöhe aus der Länge der Tibia (M1 oder M1a) zeigt sich bei männlichen Individuen für die Formeln von TELKKÄ (1950) eine Überschätzung der Körperhöhe um 8 bis 14 cm. Beide Geschlechter zeigen hohe Abweichungen bei Anwendung der Gleichungen von DUPERTUIS und HADDEN (1951), sowie TROTTER (1970). Generell zeigen die Formeln für schwarze Individuen bes- 55 aktuell Katrin Schmidt, Ronny Bindl und Horst Bruchhaus Die Ergebnisse für das Femur zeigen, dass insbesondere die von STEELE und MCKERN (1969) definierten Strecken Caput femoris – Trochanter minor (SMK 1), Trochanter minor – proximalste Ausladung Facies poplitea (SMK 2) sowie proximalste Ausladung Facies poplitea – proximaler Rand Fossa intercondylaris (SMK 3) und proximalster Rand Fossa intercondylaris – distalster Punkt Condylus medialis (SMK 4) zu einer hohen Abweichung bei der Berechnung der maximalen Länge des Femur (M1) führen. Vor allem auch die kombinierten Regressionsgleichungen, welche die Messpunkte Trochanter minor und Facies poplitea beinhalten, führen zu einer hohen Abweichung von der Abb. 4 KHS - Humerus (Männer). Abb. 5 KHS - Humerus (Frauen). Abb. 6 KHS - Femur (Männer). Abb. 7 KHS - Femur (Frauen). Femur aktuell 56 Körperhöhenschätzung an ausgewählten neolithischen und bronzezeitlichen Skeletten Abb. 9 KHS - Tibia (Frauen). Abb. 8 KHS - Tibia (Männer). Abb. 10 Abweichungen von der gemessenen Humeruslänge (M1). Regression der Segmentlängen auf die Knochenlängen nach den Regressionsformeln verschiedener Autoren. Männliche Individuen. 57 aktuell Katrin Schmidt, Ronny Bindl und Horst Bruchhaus sionsgleichungen finden, so zeigen sich zum Teil bei beiden Geschlechtern noch höhere Abweichungen von M1 als bei Verwendung der Längenteilstrecken (Abb. 14). Einzig die Maße M3a (Trochanter-Condylus lateralis-Länge) und M5 (Diaphysenlänge) und M22 (Dicke des Condylus lateralis) bei SONDER und KNUSSMANN (1985) führen zu einer geringen Abweichung von M1. Die Berechnung von M1 mittels der Maße M6 und M8 führt im Schnitt zu einer Überschätzung von M1; M7, M13, M18 und M25 unterschätzen die gemessene Femurlänge im Mittel. gemessenen Femurlänge M1. Insgesamt gilt, dass die Spannbreite der Abweichungen bei Frauen im Schnitt höher ist als bei Männern, bei den männlichen Individuen jedoch höhere Einzelabweichungen von der gemessenen Knochenlänge zu finden sind. Bei Anwendung der Regressionsformeln von JACOBS (1992) zeigt sich ein ähnliches Bild, da dieser die gleichen Messpunkte wie STEELE und MCKERN (1969) verwendet und der Berechnung der Regressionsgleichungen nur anderes Skelettmaterial (meso- und neolithische Skelette) zugrunde legt. Auch hier sind die Abweichungen von M1 für die oben aufgeführten Punkte beträchtlich, allerdings ist die Spannbreite der Abweichungen nicht ganz so hoch wie bei STEELE und MCKERN (1969). Die geringsten Abweichungen von der gemessenen Femurlänge M1 können mittels der Regressionsformeln von WRIGHT ET AL. (2003) erreicht werden. Dies gilt gleichermaßen für beide Geschlechter. Betrachtet man die untersuchten Maße nach MARTIN (1928), welche bei FUNKE (2006) sowie SIMMONS ET AL. (1990) und SONDER und KNUSSMANN (1985) (ausschließlich Männer) Eingang in die Regres- Tibia Für die Tibia gilt zu sagen, dass bei den weiblichen Individuen generell geringere Abweichungen von der gemessenen ganzen Länge der Tibia (M1) zu finden sind als bei männlichen (Abb. 15 und 16). Bei den weiblichen Individuen zeigt sich ein Trend zur Unterschätzung der Tibialänge, während sich Über- und Unterschätzung bei männlichen Individuen die Waage halten. Einzige Aus- Abb. 11 Abweichungen von der gemessenen Humeruslänge (M1). Regression der Segmentlängen auf die Knochenlängen nach den Regressionsformeln verschiedener Autoren. Weibliche Individuen. aktuell 58 Körperhöhenschätzung an ausgewählten neolithischen und bronzezeitlichen Skeletten nahme bildet der Abschnitt proximalster Punkt Tuberositas tibiae – Punkt Linienzusammenfluss ausgehend vom Unterende der Tuberositas (SMK 2) bei den Frauen. Hier zeigt sich eine starke Überschätzung der Tibialänge M1 um zum Teil mehr als 7 cm. Die Anwendung der Formeln von STEELE und MCKERN (1969) führen bei weiblichen Individuen zu geringeren Abweichungen von der gemessenen Knochenlänge als bei Männern. Hierbei führen kombinierte Formeln aus drei oder vier Segmenten zu recht guten Ergebnissen (SMK 10 – 14). Grundsätzlich scheinen sich die Regressionsgleichungen von STEELE und MCKERN (1969) etwas besser zur Schätzung der Knochenlänge M1 zu eigenen als die Formeln von JACOBS (1992). Ähnlich wie bei Femur und Humerus sind die geringsten Abweichungen von der gemessenen Tibialänge (sowohl für Männer als auch für Frauen) bei Berechnung von M1 mittels der Formeln von WRIGHT ET AL. (2003) zu finden. Allgemein lässt sich erkennen, dass die Schätzung der Knochenlänge aus Tibiafragmenten mit die höchsten Abweichungen von der gemessenen Knochenlänge produzieren (im Schnitt bis zu 3 bis 4 cm, im Extremfall sogar über 7 cm). Im Gegensatz dazu scheint der Humerus am besten zur Knochenlängenschätzung geeignet, hier finden sich die geringsten absoluten Abweichungen sowie die geringste Spannweite der Abweichungen (maximale Abweichungen im Bereich von 2,5 bis 3 cm). Teil 4: Schätzung der Körperhöhe aus berechneten Knochenlängen Zur Berechnung der Körperhöhe konnten nur die Formeln der Autoren verwendet werden, welche die Körperhöhe aus dem Maß M1 (maximale Länge des Femur, größte Länge des Humerus, ganze Länge der Tibia) Langknochen bestimmen, da nur dieses Maß aus den einzelnen Abschnitten rekonstruiert wurde, so dass nicht ganz so viele Regres- Abb. 12 Abweichungen von der gemessenen Femurlänge (M1). Regression der Segmentlängen auf die Knochenlängen nach den Regressionsformeln verschiedener Autoren. Männliche Individuen. 59 aktuell Katrin Schmidt, Ronny Bindl und Horst Bruchhaus sionsformeln wie bei Schätzung der Körperhöhe aus intakten Langknochen zum Einsatz kamen. Betrachtet man die Ergebnisse, die man bei der Schätzung der Körperhöhe aus den berechneten Knochenlängen erhält (Abb. 17-22), so zeigt sich Abb. 13 Abweichungen von der gemessenen Femurlänge (M1). Regression der Segmentlängen auf die Knochenlängen nach den Regressionsformeln verschiedener Autoren. Weibliche Individuen. Abb. 14 Abweichungen von der gemessenen Femurlänge (M1). Regression von Martinmaßen auf die Knochenlängen basierend auf den Regressionsformeln verschiedener Autoren. aktuell 60 Körperhöhenschätzung an ausgewählten neolithischen und bronzezeitlichen Skeletten kein grundlegender Unterschied zwischen den einzelnen Knochen, alle scheinen gleich gut zur Schätzung der Körperhöhe geeignet. Die Abweichungen von der Körperhöhe nach FULLY und PINEAU (1960) liegen im Schnitt bei maximal +/- 10 cm. Abb. 15 Abweichungen von der gemessenen Tibialänge (M1a). Regression der Segmentlängen auf die Knochenlängen nach den Regressionsformeln verschiedener Autoren. Männliche Individuen. Abb. 16 Abweichungen von der gemessenen Tibialänge (M1a). Regression der Segmentlängen auf die Knochenlängen nach den Regressionsformeln verschiedener Autoren. Weibliche Individuen. 61 aktuell Katrin Schmidt, Ronny Bindl und Horst Bruchhaus Geschlechtern gleichermaßen zu einer starken Unterschätzung der Körperhöhe von ca. 5 bis 12 cm. Die Formeln von BREITINGER (1937) und BACH (1965) führen zu einer Überschätzung der Körperhöhe bei den Individuen der von uns untersuchten Stichprobe. Bei Schätzung der Körperhöhe aus berechneten Femurlängen erweisen sich die Regressions- Abb. 17 Schätzung der Körperhöhe aus berechneten Humeruslängen (M1) basierend auf den Regressionsgleichungen verschiedener Autoren. Männliche Skelette. Abb. 18 Schätzung der Körperhöhe aus berechneten Humeruslängen (M1) basierend auf den Regressionsgleichungen verschiedener Autoren. Weibliche Skelette. Abb. 19 Schätzung der Körperhöhe aus berechneten Femurlängen (M1) basierend auf den Regressionsgleichungen verschiedener Autoren. Männliche Skelette. Abb. 20 Schätzung der Körperhöhe aus berechneten Femurlängen (M1) basierend auf den Regressionsgleichungen verschiedener Autoren. Weibliche Skelette. So führen die Regressionsformeln von ROTHER ET AL. (1971, 1973, 1978) bei allen Knochen und beiden aktuell 62 Körperhöhenschätzung an ausgewählten neolithischen und bronzezeitlichen Skeletten Abb. 21 Schätzung der Körperhöhe aus berechneten Tibialängen (M1a) basierend auf den Regressionsgleichungen verschiedener Autoren. Männliche Skelette. Abb. 22 Schätzung der Körperhöhe aus berechneten Tibialängen (M1a) basierend auf den Regressionsgleichungen verschiedener Autoren. Weibliche Skelette. formeln von PEARSON (1899), TROTTER und GLESER „Blacks“ (1952) sowie DE MENDONCA (2000) als gut geeignet, hier liegen die geschätzten mittleren Körperhöhen nahe an der Körperhöhe nach FULLY und PINEAU (1960). Für die Tibia liefern ebenfalls die Formeln von PEARSON (1899), aber auch die Regressionsgleichungen von GENOVÉS (1967), welche hier zusätzlich verwendet wurden, gute Ergebnisse in Hinblick auf die Schätzung der Körperhöhe. Die Regressionsformeln von TROTTER und GLESER (1952) sowie TROTTER (1970) führen bei Schätzung der Körperhöhe aus berechneten Humeruslängen zu den besten Ergebnissen (mittlere Abweichung von etwa 1 bis 3 cm). Die Ergebnisse für die direkte Schätzung der Körperhöhe aus den Maßen M13, M18 und M25 (nach SIMMONS ET AL. 1990) lassen kaum Unterschiede zu den Ergebnissen bei indirekter Körperhöhenberechnung erkennen. Die mittlere Abweichung von der Körperhöhe nach FULLY und PINEAU (1960) liegt bei direkter Schätzung der Körperhöhe aus den Abschnittsmaßen etwas niedriger als bei indirekter Körperhöhenschätzung, allerdings lässt sich bei direkter Schätzung eine höhere Spannbreite der Abweichungen erkennen (siehe Abb. 23). Diskussion Teil 5: Direkte Schätzung der Körperhöhe aus Abschnittsmaßen am Beispiel des Femur und Vergleich mit den Ergebnissen bei indirekter Körperhöhenschätzung aus diesen Maßen Generell kann man feststellen, dass es schwierig ist, geeignete Regressionsformeln für (prä-) historisches Skelettmaterial zu finden. Die meisten der aktuellen Regressionsformeln basieren auf Skelettserien, welche eine mehr oder weniger starke Beeinflussung durch die säkulare Akzeleration zeigen. Es ist unklar, inwieweit Akzelerationsprozesse auch in prähistorischen oder historischen Zeiten gewirkt haben können. Es steht außer Frage, dass die rezenten Individuen eine wesentlich höhere durchschnittliche Körperhöhe aufweisen als die Individuen im Neolithikum und der Bronzezeit. Möglicherweise bestehen SIMMONS et al. (1990) erstellten Regressionsformeln am Femur, um zum einen mittels der Maße M13, M18 und M25 die Körperhöhe direkt zu bestimmen, geben aber zum anderen auch Regressionsgleichungen an, mit denen aus diesen Maßen zunächst die maximale Länge des Femur (M1) bestimmt werden kann, um darauf folgend aus dieser berechneten Knochenlänge die Körperhöhe zu berechnen. 63 aktuell Katrin Schmidt, Ronny Bindl und Horst Bruchhaus Abb. 23 Vergleich der Abweichungen bei direkter Schätzung der Körperhöhe und indirekter Schätzung der Körperhöhe aus den Maßen M13, M18 und M25 nach den Regressionsgleichungen von SIMMONS ET AL. (1990). zwischen (prä)historischem und rezentem Material auch Proportionsunterschiede innerhalb der einzelnen Körperabschnitte (z. B. unterschiedliches Verhältnis von Rumpflänge zu Extremitätenlänge) und Unterschiede in dem prozentualen Anteil, welchen die einzelnen Langknochen zur Gesamtkörperhöhe beitragen. Diese Einflüsse erschweren die Körperhöhenschätzung bei (prä)historischem Skelettmaterial und erfordern eine kritische Betrachtung der Ergebnisse. rückführen, dass er keinen Beitrag zur Körperhöhe leistet und von Veränderungen der Körperhöhe im Laufe der Zeit am wenigsten beeinflusst bleibt. Dennoch kann er zur Körperhöhenschätzung verwendet werden, allerdings sollte die Schätzung der Körperhöhe aus Femur- und Tibialängen der Schätzung der Körperhöhe aus Humeruslängen vorgezogen werden. Femur und Tibia scheinen gleich gut zur Körperhöhenschätzung geeignet. Generell ist eine Tendenz zur Überschätzung der Körperhöhe zu erkennen, am geringsten ist diese Tendenz bei Rekonstruktion der Körperhöhe aus Femurlängen ausgeprägt. Begründen lässt sich die durchschnittliche Überschätzung der Körperhöhe möglicherweise damit, dass die zur Berechnung der Körperhöhe aus intakten Langknochen verwendeten Regressionsformeln weitestgehend an rezentem Material erstellt wurden. Dieses zeigt in den meisten Fällen bereits eine Beeinflussung durch die säkulare Akzeleration und besitzt somit eine höhere durchschnittliche Körperhöhe als das von uns untersuchte (prä)historische Skelettmaterial. Vor allem bei der für die Körperhöhenrekonstuktion aus Tibialängen erzielen die Gleichungen von PEARSON (1899) gute Schätzergebnisse. Das von PEARSON (1899) untersuchte Material ist von säkularen Veränderungen noch wenig beeinflusst, und da die vowiegend säkularen Skelettveränderungen laut Angaben einiger Autoren (...) an den distalen Knochen der unteren Extremität vonstatten gehen, scheinen die Formeln von Pearson für unser Material im Hinblick auf die Tibia als besonders geeignet. Auch die Formeln von zu Teil 1: Mittels der Methode nach FULLY und PINEAU (1960) kann die tatsächliche Körperhöhe mit einer Genauigkeit von ± 2,05 cm bestimmt werden. Die berechnete Körperhöhe stellt somit nur Annäherung an die wirkliche Körperhöhe des Individuums dar, allerdings kann mit Hilfe der „anatomischen Methode“ die Körperhöhe exakter geschätzt werden als mit mathematischen Methoden. Vor allem für prähistorisches und historisches Material stellt die „anatomische Methode“ die einzige Möglichkeit dar, mit welcher eine Körperhöhe berechnet werden kann, die als Bezugsgröße für Vergleiche mit anderen Körperhöhenschätzmethoden zu gebrauchen ist. zu Teil 2: Bei Schätzung der Körperhöhe aus intakten Langknochen erweist sich der Humerus als der Knochen mit den höchsten Einzelabweichungen von der Körperhöhe. Die weniger gute Eignung des Humerus lässt sich unter Umständen darauf zu- aktuell 64 Körperhöhenschätzung an ausgewählten neolithischen und bronzezeitlichen Skeletten TELKKÄ (1950) liegen mit ihren Schätzergebnissen im Bereich der Körperhöhe nach FULLY und PINEAU (1960), die einzige Ausnahme lässt sich bei den männlichen Tibiae erkennen. Bei allen drei Langknochen führen die Regressionsgleichungen von ROTHER ET AL. (1971, 1973, 1978) zu einer starken Unterschätzung der Körperhöhe, was bereits von RÖSING (1988) kritisiert wurde. Skelett Zeitstellung Auch BREITINGER (1937) und BACH (1965) erweisen sich in unserer Untersuchung als ungeeignet, was möglicherweise damit zusammenhängt, dass die von ihnen untersuchte Stichprobe sich zu stark von unserer unterscheidet. zu Teil 3: Die Regressionsgleichungen nach SONDER und KNUSSMANN (1988) wurden in der Auswertung der Geschlecht Körperhöhe FULLY & PINEAU (1960) [cm] Femur Tibia (M1) (M1a) (M1) 01 152,8 415,5 346,0 298,5 Sk-Nr. Humerus 01 Schnurkeramik weiblich 02 Schnurkeramik weiblich 02 160,9 415,0 334,0 305,5 03 Schnurkeramik weiblich 03 160,7 436,0 371,5 317,0 04 179,5 500,0 423,0 359,0 05 159,9 446,5 349,0 305,0 04 Schnurkeramik männlich 05 Glockenbecherkultur weiblich 06 164,1 439,0 364,0 296,0 männlich 07 165,6 449,0 358,5 320,0 08 162,2 437,0 350,0 308,5 09 158,8 434,0 361,0 309,5 10 152,3 410,0 331,0 296,0 06 Glockenbecherkultur 07 Bandkeramik männlich 08 Bandkeramik männlich 09 Aunjetitzerkultur weiblich 10 Aunjetitzerkultur weiblich Tab. 2 Anatomische Körperhöhen und gemessene Knochenlängen. Tab. 1 Übersicht Skelettmaterial. Humerus Autor Maß/ Abschnittsbezeichnung Männer mittlere Abschnittslänge [mm] Tab. 3 Mittlere Abschnittslängen Humerus. Frauen sd mittlere Abschnittslänge sd [mm] [mm] [mm] MARTIN & SALLER (1957) M3 49,5 2,9 41,0 2,8 STEELE & SMK/J1 43,4 2,9 41,0 2,8 MC KERN (1969)/ SMK/J2 54,0 11,6 58,6 4,1 JACOBS (1992) SMK/J3 185,3 19,6 166,9 16,6 6,2 WRIGHT et al. (2003) SMK/J4 17,6 2,2 SMK/J5 20,9 2,8 21,3 3,8 1,6 W 0-5 282,8 32,9 267,4 8,5 W 0-6 300,3 33,4 285,2 8,0 W 1-5 277,0 32,6 262,3 7,3 W 1-6 296,0 32,9 280,4 7,3 W 1-7 313,3 33,4 298,2 7,6 W 2-5 269,2 30,1 257,0 8,2 W 2-6 287,2 30,7 275,6 8,1 W 2-7 305,8 29,9 290,2 7,1 W 3-5 237,7 29,6 227,1 10,3 W 3-6 254,7 32,5 244,5 8,6 W3-7 273,7 32,4 258,2 13,1 sd = Standardabweichung 65 aktuell Katrin Schmidt, Ronny Bindl und Horst Bruchhaus Ergebnisse nach reiflicher Überlegung weggelassen, da sie nur auf männliche Individuen anwendbar sind, in dieser Studie aber sowohl männliche als auch weibliche Individuen untersucht wurden. Für das Humerus sind die Strecken SMK 1 und SMK 4 zur Schätzung der Knochenlänge ungeeignet. Bei SMK 1 bestand das Problem der Identifizierbarkeit des distalsten Punktes der Aufwulstung des Caputrandes, daneben ist diese Messstrecke recht kurz und nicht schaftübergreifend. Die Messpunkte der Strecke SMK 4 lassen sich zwar leicht identifizieren (proximaler und distaler Rand der Fossa olecrani), stellen aber nur ein extrem kurzes Segement des Humerus dar und sind daher zur Knochenlängenschätzung ungeeignet. Die hohen Abweichungen für den Punkt SMK 2 bei den Frauen kommen durch die schwere Identifizierbarkeit des Punktes des Linienzusammenflusses ausgehend vom Unterende der Tuberositas tibiae zustande. Dieser Messpunkt charakterisiert den Übergang der Linien in die Margo anterior der Tibia. Aufgrund der geringeren Robustizität der weiblichen Knochen sind diese Linien schwieriger zu identifizieren. Auch bei den männlichen Individuen zeigt sich bei der Schätzung der Knochenlänge aus diesem Segment eine erhöhte Abweichung, diese ist allerdings nicht ganz so hoch wie bei den weiblichen Individuen. Tab. 4 Mittlere Abschnittslängen Femur. Betrachtet man die einzelnen Teilstrecken am Femur, so zeigt sich, dass die Stelle der proximalsten Ausladung der Facies poplitea, welche durch den Zusammenfluss der Linea supracondylaris medialis und lateralis zur Linea aspera (Parallelwerden beider Linien) definiert wird, nur sehr schwer zu identifizieren ist. Je nach Grad der Muskularisierung ist die Linea aspera am Knochen mehr oder weniger stark ausgeprägt, so dass bei einigen weniger robusten Knochen dieser Punkt schwer abzugrenzen war (siehe auch WRIGHT ET AL. 2003 und JACOBS 1992). Die Stelle, an welcher Linea supracondylaris medialis und lateralis parallel werden, hängt sehr stark von subjektiven Empfinden ab und variiert je nach Betrachter. Auch Regressionsgleichungen, welche den Mittelpunkt des Trochanter minor als Segmentgrenze beinhalten, führen zu hohen Abweichungen von der tatsächlichen Femurlänge, obwohl dieser Punkt am Knochen weniger schwierig zu identifizieren ist und auch die Messung ohne Probleme durchgeführt werden kann. Die geringeren Spannbreiten der Abweichungen bei JACOBS (1992), welcher die gleichen Messstrecken verwendet wie STEELE und MCKERN (1969), lassen sich möglicherweise dadurch begründen, dass den Kalkulationen von JACOBS (1992) meso- und neolithisches Skelettmaterial zugrunde liegt und dieses im Gegensatz zu den Femur Autor Männer Maß/ Abschnittsbezeichnung MARTIN & SALLER (1957) aktuell Frauen mittlere Abschnittslänge sd mittlere Abschnittslänge sd [mm] [mm] [mm] [mm] M3a 415,5 6,7 407,3 15,7 M6 29,4 2,3 26,4 2,5 M8 87,6 5,5 82,1 4,9 M5 351,0 2,5 338,3 13,6 M7 26,6 1,9 26,3 1,2 M13 94,5 6,3 88,1 6,1 M18 45,3 0,9 43,0 1,9 M21 76,3 3,4 -- -- M22 59,2 1,8 56,9 1,8 M25 35,9 2,0 34,8 1,7 STEELE und SMK/J1 75,1 3,3 71,8 5,1 MC KERN (1969)/ SMK/J2 179,2 12,6 182,9 16,0 JACOBS (1992) SMK/J3 144,8 13,8 135,8 11,4 SMK/J4 38,4 2,5 35,9 1,2 66 Körperhöhenschätzung an ausgewählten neolithischen und bronzezeitlichen Skeletten Tibia Autor Maß/ Abschnittsbezeichnung Tab. 5 Mittlere Abschnittslängen Tibia. Männer Frauen mittlere Abschnittslänge sd mittlere Abschnittslänge sd [mm] [mm] [mm] [mm] STEELE und SMK/J1 21,5 5,7 19,3 1,8 MC KERN (1969)/ SMK/J2 65,6 17,1 63,1 5,1 JACOBS (1992) SMK/J3 165,3 20,9 153,5 17,5 SMK/J4 103,9 14,5 94,5 5,3 WRIGHT et al. (2003) SMK/J5 17,1 1,8 15,8 1,3 W 0-6 353,6 30,6 328,3 15,4 W 1-6 336,3 30,6 312,9 15,5 W 1-7 357,0 33,5 331,1 15,2 sd = Standardabweichung Es lässt sich feststellen, dass es an der Tibia die größten Schwierigkeiten bei der Identifikation der Segmente gab. Sowohl der Punkt des Linienzusammenflusses ausgehend vom Unterende der Tuberositas tibiae als auch der Überkreuzungspunkt der Margo anterior über die zentrale Tibiaachse lassen sich nur schwer klar abgrenzen, womit sich auch die hohen Abweichungen bei Anwendung dieser Formeln begründen lassen. Die geringsten Probleme bei der Identifikation der Messpunkte gab es am Humerus, daher führen diese Regressionsgleichungen auch zu den geringsten Abweichungen von der gemessenen Knochenlänge. rezenten Skeletten bei STEELE und MCKERN (1969) eine geeignetere Referenzstichprobe für das von uns untersuchte Skelettmaterial darstellt. WRIGHT ET AL. (2003) erstellten ihre Regressionsgleichungen anhand von Maya-Skeletten, welche eine geringe durchschnittliche Körperhöhe aufweisen. Da auch das von uns untersuchte Skelettmaterial eine ähnlich geringe Körperhöhe und ähnliche Proportionen besitzt, erweisen sich diese Regressionsgleichungen aus genanntem Grund möglicherweise als besonders geeignet. Betrachtet man die Schätzergebnisse für die Körperhöhe bei Verwendung der Maße nach MARTIN (1928), so zeigen nur zwei Messstrecken eine gute Korrelation mit der Knochenlänge, was mit hoher Wahrscheinlichkeit auf die Diaphysenübergreifende Länge dieser Messstrecken zurückzuführen ist. Von den horizontalen Martinmaßen eignet sich nur Maß 22 zur Bestimmung der Körperhöhe. Die anderen Horizontalmaße spiegeln in erster Linie nur Robustizitätsverhältnisse (Durchmesser/Umfang des Knochens) wider und stehen nicht im direkten Zusammenhang mit der Knochenlänge, auch wenn Individuen mit robusten Knochen oftmals auch längerer Knochen und eine größere Gesamtkörperhöhe besitzen. Viele Autoren geben an, dass Maß M13 für die Schätzung der Körperhöhe gut geeignet ist, was in unserer Untersuchung nicht bestätigt werden konnte. Bedingt durch die hohe Variabilität des Corpus-CollumWinkels variiert auch die Länge dieser Messstrecke sehr stark und scheint zur Körperhöhenschätzung nur wenig geeignet. Der einzige Vorteil, welchen die Martinmaße gegenüber den Längenteilstrekken anderer Autoren besitzen, ist ihre problemlose Identifizierbarkeit und die Reproduzierbarkeit der Messungen am Knochen. zu Teil 4: Bei Schätzung der Körperhöhe aus den berechneten Knochenlängen spielt es anscheinend nur eine untergeordnete Rolle, ob die Abweichung von der gemessenen Knochenlänge hoch oder niedrig ist. Es spiegelt sich bei allen Knochen ein ähnlicher Trend wider, der auch schon bei Schätzung der Körperhöhe aus den gemessenen Knochenlängen (Teil 1) gezeigt werden konnte. Bestimmte Regressionsgleichungen scheinen für das von uns untersuchte Material ungeeignet, weitestgehend unabhängig davon, mit welchem der drei Knochen die Regression auf die Körperhöhe durchgeführt wird. Hierbei liegen die Abweichungen, welche man bei Berechnung der Körperhöhe aus intakten Langknochenlängen erhält, nur geringfügig unter den Abweichungen, die bei Schätzung der Körperhöhe aus berechneten Langknochenlängen erzielt wurden. Wie schon von anderen Autoren bestätigt (z. B. RÖSING 1988), eigenen sich die Regressionsformeln von ROTHER ET AL. (1971, 1973, 1978) nicht zur Schätzung der Körperhöhe, da sie genau wie 67 aktuell Katrin Schmidt, Ronny Bindl und Horst Bruchhaus bei Schätzung der Körperhöhe aus den gemessenen Knochenlängen einen starken Trend zur Unterschätzung dieser aufweisen. Auch BREITINGER (1937) und BACH (1965) erweisen sich in unserer Untersuchung als ungeeignet, wie auch bereits bei Schätzung aus den gemessenen Längen gezeigt wurde. Gut geeignet scheinen die Formeln von PEARSON (1899), TELKKÄ (1950) sowie TROTTER und GLESER (1952, 1958). Schlussfolgerungen Letzten Endes lässt sich sagen, dass die von uns erzielten Ergebnisse einen deutlichen Trend bezüglich der Eignung bestimmter Methoden erkennen lässt. Man sollte sich jedoch immer vor Augen halten, dass die Ergebnisse bei Anwendung der Methoden auf Skelettmaterial anderer Zeithorizonte oder zwischen Populationen gleicher Zeitstellung variieren können. Es kann die Aussage getroffen werden, dass auch aus Knochenfragmenten eine halbwegs annehmbare Schätzung der Körperhöhe möglich ist. Allerdings sollte man darauf achten, die Knochenlänge zunächst nur mittels Segmenten zu bestimmen, welche ausreichend lang und gut zu identifizieren sind und welche gut mit der Gesamtlänge des entsprechenden Knochens korrelieren. Diese Segmente sollte man anschließend verwenden, um die Knochenlänge mit der für die untersuchte Stichprobe am besten geeigneten Regressionsformeln (in unserem Fall die von WRIGHT ET AL. 2003) zu rekonstruieren. Zur Schätzung der Körperhöhe sollten die Regressionsformeln verwendet werden, welche für die Stichprobe die besten Ergebnisse liefern. Da sich die Formeln nach ROTHER ET AL. (1971, 1973, 1978) sowie BREITINGER (1937) und BACH (1965) für die hier untersuchten Skelette als ungeeignet erwiesen, sollte man vor ihrer Anwendung überprüfen, ob sie für das zu untersuchende Material brauchbare Schätzergebnisse liefern oder ihnen die Anwendung anderer Formeln vorzuziehen ist. Für weitere Untersuchungen wäre es sinnvoll, eine größere und folglich statistisch repräsentative Skelettserie einer bestimmten Population zusammenzustellen, für die eine Schätzung der Körperhöhe nach der Methode von FULLY und PINEAU (1960) vorgenommen werden kann. Alle zu untersuchenden Skelette dieser Population sollten möglichst geschlechtsbestimmt sein, weiterhin sollte eine Einteilung entsprechend der anthropologischen Altersklassen bekannt sein. Diese Skelettserie sollte genügend Individuen von verschiedenen Körperhöhengruppierungen (groß–klein) sowie von verschiedenen Altersklassen enthalten, um eine differenziertere Untersuchung der einzelnen Methoden bezogen auf unterschiedlich große/kleine sowie unterschiedlich alte Individuen durchführen zu können. zu Teil 5: Vergleicht man die Ergebnisse, welche man bei Schätzung der Körperhöhe aus den berechneten Femurlängen erhält (indirekte Methode), von den Ergebnissen, die man bei direkter Schätzung der Körperhöhe aus den gleichen Femurabschnitten erhält (beides nach der Methode von SIMMONS ET AL. 1990), so erweist sich die indirekte Methode aufgrund der geringeren Spannbreite der Abweichungen in Bezug auf das untersuchte Material als besser geeignet. Begründen lässt sich dies unter Umständen damit, dass die untersuchten Strecken sehr kurz sind und ausschließlich Horizontalmaße darstellen. Diese leisten keinen wesentlichen Beitrag zur Körperhöhe und korrelieren folglich nur sehr wenig mit ihr. Die Korrelation der Horizontalmaße mit der Knochenlänge ist dagegen um einiges höher, so dass es angebracht erscheint, zunächst die Knochenlänge aus diesen Maßen zu berechnen und anschließend aus der berechneten Knochenlänge die Körperhöhe zu schätzen. Möglicherweise lässt sich bei direkter Schätzung der Körperhöhe aus Längenteilstrecken, welche Diaphysen übergreifend sind, ein besseres Ergebnis erzielen als es für die indirekte Schätzmethode der Fall ist. Allerdings wurde dies hier nicht untersucht. aktuell 68 Körperhöhenschätzung an ausgewählten neolithischen und bronzezeitlichen Skeletten Literatur REICHELT, E. (2003): Dissertation 2003. BACH, H. (1965): Zur Berechnung der Körperhöhe aus den langen Gliedmaßenknochen weiblicher Skelette. Anthrop Anz. 20, 1965, 12-21. RÖSING, F. W. (1988): Körperhöhenrekonstruktion aus Skelettmaßen. In: R. KNUSSMANN (Hrsg.): Anthropologie. Handbuch der vergleichenden Biologie des Menschen. Bd.1: Wesen und Methoden der Anthropologie. Gustav Fischer Verlag, Stuttgart, New York 1988. BREITINGER, E. (1937): Zur Berechnung der Körperhöhe aus den langen Gliedmaßenknochen. Anthropol Anz. 14, 1937, 14: 249-247. ROTHER, B. (1971): Möglichkeiten und Grenzen der Körperhöhenrekonstruktion aus den Maßen langer Röhrenknochen. Med. Diss. Leipzig 1971. DUPERTIUS, C. W./J. A. HADDEN (1951): On the reconstruction of stature from long bones. Am J Phys Anthropol. 9, 1951, 15-54. SIMMONS, T./JANTZ, R. L./M. W. BASS (1990): Stature estimation from fragmentary femora: A revision of the Steele method. J Forensic Sci. 35(3), 1990, 628-636. FULLY, G./H. PINEAU (1960): Determination de la stature au moyen du squelette. Ann Med Leg. 40, 1960, 145-154. SONDER, E./R. KNUSSMANN (1985): Zur Körperhöhenbestimmung männlicher Individuen aus Femur-, Tibiaund Humerusfragmenten. Z Morph Anthrop. 75(2), 1985, 131-153. FUNKE, C. (2006): Dissertation 2006. GENOVÉS, S. (1967): Proportionality of the long bones and their relation to stature among Mesoamericans. Am J Phys Anthropol. 26, 1967, 67-78. STEELE, G. D./T. W. MCKERN (1969): A Method for Assessment of Maximum Long Bone Length and Living Stature from Fragmentary Long Bones. Am J Phys Anthropol. 31, 1969, 215-228. HAUSER, R./SMOLINSKI, J./T. GOS (2005): The estimation of stature on the basis of measurements of the femur. Forensic Sci Int. 147, 2005, 185-190. TELKKÄ, A. (1950): On the prediction of human stature from the long bones. Acta Anat. 9, 1950, 103-117. JACOBS, K. (1992): Estimating femur and tibia length from fragmentary bones: an evaluation of Steele´s (1970) method using a prehistoric European sample. Am J Phys Anthropol. 89(3), 1992, 333-345. TROTTER, M./G. C. GLESER (1952): Estimation of stature from long bones of American Whites and Negroes. Am J Phys Anthropol. 10, 1952, 10, 463-514. – (1952): Corrigenda to „Estimation of Stature from Long Limb Bones of American Whites and Negroes“. Am J Phys Anthropol. 47, 1952, 355-356. MARTIN, R./R. KNUSSMANN (1988): Anthropologie, Band I/1 und I/2, Wesen und Methoden der Anthropologie Gustav-Fischer-Verlag Stuttgart/New York, 1. Auflage 1988. – (1958): A re- evaluation of stature based on measurements taken during life and of long bones after death. Am J Phys Anthropol. 16, 1958, 79-124. MARTIN, R./K. SALLER (1957): Lehrbuch der Anthropologie, Band I und II, Gustav-Fischer-Verlag Stuttgart, 3. Auflage 1957. WRIGHT, L. E./M. A. VÁSQUEZ (2003): Estimating the Length of Incomplete Long Bones: Forensic Standards From Guatemala. Am J Phys Anthropol. 120, 2003, 233-251. LORKE, D./MÜNZER, H./E. WALTER (1953): Zur Rekonstruktion der Körpergröße eines Menschen aus den langen Gliedmaßenknochen. Zeitschrift gerichtl. Medizin. 42, 1953, 189-202. Katrin Schmidt Ronny Bindl Dr. Horst Bruchhaus Institut für Humangenetik und Anthropologie Friedrich-Schiller-Universität Jena Kollegiengasse 10 07740 Jena [email protected] [email protected] [email protected] OLIVIER, G. (1963): La détermination de la stature d´après les os longs des membres. Bull Mém Soc d´Anthrop Paris 11e série 4, 1963, 443-449. OLIVIER, G./H. TISSIER (1975): Estimation de la stature feminine d´après les os longs des membres. Bull Mém Soc d´Anthrop Paris 2e série 13, 1975, 297-306. PEARSON, K. (1899): On the reconstruction of stature of prehistoric races. Mathematic contributions to the theory of evolution. Transa Roy Soc A. 192, 1899, 169-244. 69 aktuell Inhaltsverzeichnis / Table of contents WINFRIED HENKE Paläoanthropologie - Standortbestimmung einer innovativen Disziplin ..............................1 RONNY BINDL, RAINER NITSCHE und HORST BRUCHHAUS Altersschätzung von Skelettmaterial - Untersuchungen zur Obliteration der Nähte des Schädels ......................................................................................25 JÖRG ORSCHIEDT Virtuelle Rekonstruktion und stereolithographisches Modell eines jungneolithischen Schädelfundes aus der Blätterhöhle in Hagen, NRW .............................35 CLAUDIA GERLING und MICHAEL FRANCKEN Das linearbandkeramische Gräberfeld von Schwetzingen ............................................43 KATRIN SCHMIDT, RONNY BINDL und HORST BRUCHHAUS Körperhöhenschätzung an ausgewählten neolithischen und bronzezeitlichen Skeletten ...........51 BIRGIT GROSSKOPF und ALEXANDER GRAMSCH Leichenbrand erzählt vom Umgang mit den Toten - Die interdisziplinäre Rekonstruktion ritueller Handlungen am Beispiel eines Urnengräberfelds der Lausitzer Kultur ....................71 FABIAN LINK Gesichtsrekonstruktionen von drei alamannischen Individuen aus dem Gräberfeld Basel-Gotterbarmweg (5./6. Jh. n. Chr.) .................................................................81 SIMONE KAHLOW Die Pest als Interpretationsproblem mittelalterlicher und frühneuzeitlicher Massengräber ............................................................................................97 Absender/Expéditeur: Historische Anthropologie Institut für Medizingeschichte Universität Bern Fabrikstrasse 29d CH – 3012 Bern Switzerland