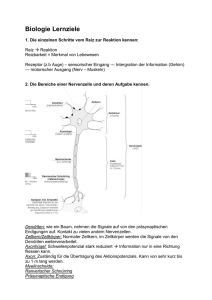
Volltext
Werbung

Aus dem Veterinär-Anatomischen Institut der Veterinärmedizinischen Fakultät der Universität Leipzig und dem Anatomischen Institut der Medizinischen Fakultät der Albert-Ludwigs-Universität Freiburg Einfluss des clostridialen C3 Toxins auf die Dendritenmorphologie und Spinebildung von CA1 Pyramidenzellen in Hippocampus-Schnittkulturen der Maus - eine quantitative lichtmikroskopische Untersuchung Inaugural-Dissertation zur Erlangung des Grades eines Doctor medicinae veterinariae (Dr. med. vet.) durch die Veterinärmedizinische Fakultät der Universität Leipzig eingereicht von Thorsten Hintze aus Freiburg im Breisgau Leipzig, 2010 Mit Genehmigung der Veterinärmedizinischen Fakultät der Universität Leipzig Dekan: Prof. Dr. Uwe Truyen Betreuer: Prof. Dr. Johannes Seeger Prof. Dr. Bernd Heimrich Gutachter: Prof. Dr. Johannes Seeger, Veterinär-Anatomisches Institut, Universität Leipzig Prof. Dr. Bernd Heimrich, Institut für Anatomie und Zellbiologie, AlbertLudwigs-Universität Freiburg Prof. Dr. Michael Stoffel, Abteilung Veterinäranatomie, Vetsuisse Fakultät, Universität Bern, Schweiz Tag der Verteidigung: 5. Oktober 2010 II Inhaltsverzeichnis 1 Einleitung und Fragestellung ............................................................................................... 1 2 Literaturübersicht................................................................................................................. 3 2.1 Einführung...................................................................................................................... 3 2.2 Rho GTPasen .................................................................................................................. 4 2.3 Initialisierung und Auswachsen von Neuriten............................................................. 6 2.4 Einwirkung der Rho Aktivität auf die Dendriten- und Spinebildung....................... 7 3 Tiere, Material und Methoden........................................................................................... 10 3.1 Versuchsplanung .......................................................................................................... 10 3.2 Schnittkulturen............................................................................................................. 10 3.2.1 Tiere ........................................................................................................................ 10 3.2.2 Herstellung der Schnittkulturen .............................................................................. 11 3.2.3 Inkubation................................................................................................................ 11 3.3 Biocytinfüllung der Pyramidenzellen ......................................................................... 12 3.4 Fixierung und Visualisierung der Pyramidenzellen ................................................. 12 3.5 Resultierende Behandlungsgruppen........................................................................... 13 3.6 Lösungsansätze ............................................................................................................. 16 3.7 Quantitative Auswertung der dendritischen Parameter .......................................... 17 3.7.1 Rekonstruktion der CA1 Pyramidenzellen.............................................................. 17 3.7.2 Analyse der Daten ................................................................................................... 18 3.8 Statistische Tests........................................................................................................... 18 4 Ergebnisse ............................................................................................................................ 20 4.1 Allgemeine Befunde zu den CA1 Pyramidenzellen................................................... 20 4.2 Morphologie der hippocampalen CA1 Pyramidenzellen ......................................... 20 4.2.1 Serumhaltige Kontrollgruppe .................................................................................. 21 4.2.2 Serumfreie Kontrollgruppe ..................................................................................... 23 4.2.3 C3 bot Toxinbehandelte Experimentalgruppe......................................................... 24 III 4.3 Quantitative Analyse.................................................................................................... 27 4.3.1 Dendritenlänge ........................................................................................................ 28 4.3.1.1 Gesamtlänge der Basaldendriten ...................................................................... 28 4.3.1.2 Gesamtlänge der Apikaldendriten .................................................................... 29 4.3.2 Verzweigungspunkte............................................................................................... 30 4.3.2.1 Anzahl der Verzweigungspunkte der Basaldendriten ...................................... 30 4.3.2.1 Anzahl der Verzweigungspunkte der Apikaldendriten .................................... 30 4.3.3 Anzahl der Spines.................................................................................................... 31 4.3.3.1 Anzahl der Spines der Basaldendriten ............................................................. 31 4.3.3.2 Anzahl der Spines der Apikaldendriten ........................................................... 32 4.3.3.3 Gesamtzahl der Spines ..................................................................................... 33 4.3.4 Spinedichte ............................................................................................................. 34 4.3.4.1 Spinedichte der Basaldendriten ........................................................................ 34 4.3.4.2 Spinedichte der Apikaldendriten ...................................................................... 35 4.3.4.3 Spinedichte der gesamten Zelle........................................................................ 36 4.4 Zusammenfassung der Ergebnisse ............................................................................. 37 5 Diskussion ............................................................................................................................ 39 5.1 Einführung in die Problemstellung ............................................................................ 39 5.2 Bewertung des experimentellen Ansatzes .................................................................. 40 5.3 Evaluierung der Ergebnisse ........................................................................................ 41 6 Zusammenfassung............................................................................................................... 44 7 Summary .............................................................................................................................. 46 8 Literaturverzeichnis............................................................................................................ 48 Anhang .................................................................................................................................... 57 Danksagung............................................................................................................................. 60 IV Verwendete Abkürzungen ABC Avidin-Biotin-Peroxidase-Komplex ACSF artificial cerebrospinal fluid aqua bidest. aqua bidestillata BME basal medium Eagle CA1 Cornu ammonis Region 1 CA3 Cornu ammonis Region 3 C3 bot C3 Exoenzym von Clostridium botulinum DAB Diaminobenzidintetrahydrochlorid DG Gyrus dentatus (dentate gyrus) DIV days in vitro GAP GTPase activating proteins GDI guanine nucleotide dissociation inhibitors GEF guanine nucleotide exchange factors MEM minimum essential medium MW Mittelwert P0 erster postnataler Tag PB Phosphatpuffer PFA Paraformaldehyd SD Standardabweichung V 1 Einleitung und Fragestellung Lokale Pyramidenzellen bilden die Hauptneurone des Hippocampus. Sie werden entsprechend ihrer Position und Morphologie ihrer Dendriten in CA1 und CA3 Pyramidenzellen differenziert. Ihre Dendriten besitzen postsynaptische Vorwölbungen, so genannte Spines, welche in einem spezifischen Verteilungsmuster angeordnet sind. Das Wachstum und die Entwicklung von Nervenzellen werden durch Rho-GTPasen entscheidend beeinflusst. Um die Charakteristik dieser Einflussnahme näher untersuchen zu können, wurde für diese Studie ein Agens gesucht, mit dessen Hilfe die Aktivität der kleinen GTPasen manipuliert werden kann. Eine etablierte Methode zur Inaktivierung von Rho stellt die Anwendung des C3 Toxins von Clostridium botulinum dar. Dieses Exoenzym wird von C- und D-Stämmen des Bakteriums gebildet und inaktiviert Rho-Proteine durch ADPRibosylierung (AKTORIES u. FREVERT 1987, CHARDIN et al. 1989). Die resultierende Eliminierung der Rho-Wirkung führt zur Dominanz seiner beiden Gegenspieler Rac und Cdc42 und damit zur Entfaltung von deren Wirkung auf die Morphologie der Nervenzelle. Diese Studie verfolgt das Ziel der qualitativen und quantitativen Analyse der morphologischen Veränderungen von intrazellulär mit Biocytin gefüllten und dadurch identifizierbaren CA1 Pyramidenzellen. Dabei wurden an organotypischen HippocampusSchnittkulturen der Maus, in denen sich bekanntermaßen Pyramidenzellen ähnlich entwickeln wie in vivo (FROTSCHER et al. 1990), die Effekte der C3bot Toxin-Applikation auf die Verzweigung der Dendriten sowie Anzahl und Dichte der dendritischen Spines untersucht. Methodisch wurden mehrere Versuchsgruppen gebildet, um mögliche Effekte durch die toxininduzierte Inaktivierung von Rho vergleichend nachweisen zu können. Um eine Beeinflussung des C3 Toxins durch möglicherweise im Serum enthaltene Proteasen ausschließen zu können, wurde die toxinbehandelte Zellgruppe serumfrei inkubiert. Bei diesem Versuchsaufbau war im Vorfeld ungewiss, ob die verwendeten Schnittkulturen eine serumfreie Inkubation unbeschadet überstehen und sich physiologisch entwickeln können. Neben einer weiteren serumfrei inkubierten Gruppe, deren Funktion darin bestand, auszuschließen, dass in der Toxingruppe möglicherweise zu beobachtende Effekte tatsächlich auf das Toxin und nicht etwa auf Effekte durch den Serumentzug zurückführen sind, bildeten normal serumhaltig inkubierte Schnittkulturen die Kontrollgruppe. Die Inkubation dauerte 14 Tage, während die Dauer der Toxinexposition zwischen vier und sechs Stunden betrug. Mit Hilfe eines Lichtmikroskops und eines Computers wurden 1 zweidimensionale Rekonstruktionen der biocytin-markierten CA1 Pyramidenzellen erstellt, und die Gesamtlänge der Dendriten, die Anzahl der dendritischen Verzweigungspunkte sowie die Gesamtzahl und Dichte der dendritischen Spines gemessen und statistisch zwischen den Gruppen verglichen. In vorangehenden Studien haben sich die Autoren mit der Untersuchung der Toxinwirkung auf dissoziierte Zellen befasst, deren Typ teilweise nicht bestimmt wurde (AHNERTHILGER et al. 2004, PILPEL u. SEGAL 2004). Bisher nicht geprüft worden ist, ob das C3 Exoenzym auf Neurone, die sich im Gewebeverband befinden, die gleichen in der Literatur beschriebenen Wirkungen ausübt oder ob es zu abweichenden Beobachtungen kommt. Folgende Fragen sollten beantwortet werden: 1. Überleben die hippocampalen Neurone einen kurzzeitigen Serumentzug, der vor Applikation des C3 bot Toxins behandlungstechnisch unumgänglich ist? 2. Führt die Behandlung mit dem C3 Toxin zu strukturellen Veränderungen der Dendritenarchitektur? 3. Kommt es nach der C3 Toxin-Behandlung zu Veränderungen in der Spinezahl bzw. Spinedichte? 4. Sind strukturelle Veränderungen an allen Dendritenabschnitten erkennbar oder kommt es zu selektiven Veränderungen bestimmter Dendritenabschnitte? 2 2 Literaturübersicht 2.1 Einführung Das Gehirn stellt das Steuerungszentrum aller kognitiven, motorischen und emotionalen Funktionen dar. Es in der Lage, Informationen über unsere Umgebung aufzunehmen, zu verarbeiten, weiterzuleiten und zu speichern. Zentrales strukturelles Element sind die Nervenzellen oder Neurone. Schätzungen zufolge besitzt das menschliche Gehirn etwa 50 bis 100 x 109 Neurone. Diese sind über so genannte Synapsen auf komplexe Art und Weise miteinander verschaltet. Trotz der Komplexität zeigen diese Verschaltungen oftmals ein hohes Maß an Ordnung, was sich in der Ausbildung ganz bestimmter neuronaler Schaltkreise widerspiegelt. Das gewährleistet die zielgerichtete Weiterleitung von Signalen und ermöglicht die neuronalen Informationen korrekt zu verarbeiten und angemessen auf neuronale Stimuli zu antworten. Die physiologische Reifung von Nervenzellen ist damit essentiell für die normale Funktion des Zentralnervensystems. Entsprechend können Störungen der neuronalen Differenzierung beziehungsweise der synaptischen Konnektivität die Signalübertragung beeinflussen und sich in kognitiven Defiziten ebenso wie in motorischen oder emotionalen Fehlfunktionen niederschlagen. Ganz allgemein durchlaufen Neuronen nach ihrer Geburt und Wanderung zum Zielort morphologische Veränderungen: Erstens die Ausbildung eines Dendritenbaumes, mit dem die Zellen in der Lage sind, Informationen von anderen Neuronen aufzunehmen, und zweitens das Auswachsen eines efferenten Axons. Über spezifische Kontaktstrukturen, die Synapsen, findet die Übermittlung der Informationen statt. Im Bereich der Dendriten erfolgt durch zwei verschiedene Lokalisationen der Synapsen eine Regulation des Informationstransfers. Nervenfasern sind imstande, mit dem nachgeschalteten Dendriten entweder so genannte Schaftsynapsen einzugehen oder präsynaptische Boutons an postsynaptischen Dornen, den Spines, auszubilden. Allgemein gilt, dass Schaftsynapsen inhibitorische Synapsen darstellen, wohingegen Spinesynapsen exzitatorische Verknüpfungen repräsentieren. Eine Reihe von Krankheiten geht mit morphologischer Fehlentwicklung der Neurone einher. So ist bekannt, dass bei gesicherter Diagnose bestimmter neurologischer oder psychiatrischer Erkrankungen wie zum Beispiel Demenz, stärkerer Depression, Schizophrenie oder Epilepsie eine geringere Dichte oder Missbildungen der dendritischen Spines zu beobachten sind. Diese Veränderungen treten auch auf bei Zustand nach Traumata, chronischem Medikamenten3 missbrauch, sowie in Folge normalen Alterns (BLANPIED u. EHLERS 2004, FIALA et al. 2002, NIMCHINSKY et al. 2002). Bestimmte Erkrankungen führen zu einer Abnahme der Spinedichte: Schizophrene Patienten zeigen eine Verarmung an Spines im Bereich des präfrontalen Cortex (GLANTZ u. LEWIS 2001), während Patienten mit Epilepsie unter einem Spinemangel im Hippocampus leiden (ISOKAWA 2000, SWANN et al. 2000, JIANG et al. 1998). Bei chronisch neurodegenerativen Erkrankungen wie Morbus Alzheimer oder Morbus Parkinson liegen im Frühstadium der Erkrankung oft Spineverlust oder andere dendritische Veränderungen vor (SELKOE 2002). Im Gegensatz dazu führt chronischer Abusus von Kokain und Amphetaminen zu einer erhöhten Spinedichte in der Hirnregion, die mit Suchterkrankungen assoziiert ist, dem Nucleus accumbens (NORRHOLM et al. 2003, ROBINSON u. KOLB 1999). Außerdem bestehen Zusammenhänge zwischen einigen erblichen Formen der mentalen Retardierung mit Defekten in der Proteinexpression, die die Spineentwicklung und deren Aufrechterhaltung reguliert (DÖLEN u. BEAR 2005). Verschiedene pathologische Veränderungen der Spines führen zu unterschiedlicher klinischer Symptomatik der einzelnen Erkrankungen, weil die Spinemorphologie in engem Zusammenhang mit der synaptischen Funktion steht (CALABRESE et al. 2006). Erkrankungen des Zentralnervensystems sind bislang vergleichsweise unbefriedigend therapierbar. Grundlagenforschung in diesem Bereich kann zu weiteren Erkenntnissen führen und letztendlich neue und aussichtsreiche Therapieformen auf den Weg bringen. 2.2 Rho GTPasen Seit einigen Jahren gibt es eine Reihe von Hinweisen, wonach Proteine der Rho-Familie der GTPasen sowie verwandte Moleküle an neuronalen Differenzierungsprozessen maßgeblich beteiligt sind. So kontrollieren Rho-GTPasen sowohl Dendritenwachstum und –differenzierung als auch das Axonwachstum, steuern die axonale Wegfindung und sind für Bildung und Aufrechterhaltung der dendritischen Spines verantwortlich (LEEMHUIS et al. 2007). Aufgrund ihrer großen Bedeutung für die genannten Vorgänge ist es keineswegs verwunderlich, dass Mutationen der die Regulatoren und Effektoren der Rho-GTPasen codierenden Gene mit neurologischen Leiden in enger Verbindung stehen (GOVEK et al. 2005). 4 Die Rho-Familie der kleinen GTPasen stellt eine Untergruppe der übergeordneten Familie der Ras-GTPasen dar (VAN AELST u. D'SOUZA-SCHOREY 1997, HALL 1998). Neben Rho (RhoA, RhoB, RhoC) gehören Rac (Rac1, Rac2, RacE), Cdc42 und weitere GTPasen zur Rho-Familie (VAN AELST u. D'SOUZA-SCHOREY 1997, BURRIDGE u. WENNERBERG 2004). RhoA, Rac und Cdc42 sind an der Spinebildung und der Bestimmung ihrer Gestalt maßgeblich beteiligt (NEWEY et al. 2005). Rho GTPasen sind niedrig-molekulare Guanin-Nukleotid-bindende Proteine, die als binäre Schalter fungieren. Durch das Umschalten einer einzigen GTPase können mehrere verschiedene Signalübertragungswege miteinander koordiniert aktiviert werden. Um diese komplexen zellulären Prozesse zu steuern, nutzen die GTPasen einfache biochemische Mechanismen. Sie wechseln zwischen zwei konformativen Zuständen hin und her: Im aktiven Status an GTP und im inaktiven Status an GDP gebunden. Durch Hydrolyse wird GTP in GDP umgewandelt. Im „eingeschalteten“ Status (GTPgebunden) interagieren die Rho GTPasen mit einem von etwa 60 Zielproteinen und generieren solange ein Signal, bis der Schalter durch Hydrolyse wieder auf „aus“ gestellt wird (MACKAY u. HALL 1998). Dieses Prinzip hat sich im Laufe der Evolution fortentwickelt, so dass Säugerzellen über mehrere hundert solcher GTPasen verfügen. Das Genom des Menschen verfügt über die Information zur Bildung von über 60 GTPaseAktivatoren (guanine nucleotide exchange factors, GEFs) und über 70 -Inaktivatoren (GTPase activating proteins, GAPs), was auf eine sehr sorgfältige Kontrolle des „Rho-Schalters“ hindeutet. Die GEFs aktivieren die GTPasen, indem sie den Austausch von gebundenem GDP zugunsten von GTP fördern (SCHMIDT u. HALL 2002). Im Gegenzug arbeiten die GAPs als negative Regulatoren der GTPasen, indem sie die intrinsische Rate der GTP-Hydrolyse einer GTPase erhöhen (BERNARDS 2003, BERNARDS u. SETTLEMAN 2004). Als weitere Negativ-Regulatoren fungieren die so genannten guanine nucleotide dissociation inhibitors (GDIs), die den Austausch von GDP gegen GTP verhindern und außerdem die intrinsische GTPase-Aktivität von GTP-gebundenen GTPasen hemmen (ZALCMAN et al. 1999, HOFFMAN et al. 2000). Die Erkenntnis, dass konstitutiv-aktive und dominant-negative Mutanten derselben Rho-GTPase abnormale Phänotypen verursachen können, unterstreicht die Bedeutung einer strikten Regulation der Rho-GTPase-Aktivität für die normale zelluläre Funktion. Ferner kommt der Fähigkeit, zwischen der aktiven GTP-gebundenen und inaktiven GDP-gebundenen Form zu wechseln, eine besondere Bedeutung zu (LUO et al. 1994). Aktivierte, GTP-gebundene Rho-GTPasen interagieren mit spezifischen Effektormolekülen, um Signale zu übermitteln (BUCHSBAUM et al. 2002, BUCHSBAUM et al. 2003). 5 Am intensivsten aus der Rho-Familie der kleinen GTPasen sind RhoA, Rac1 und Cdc42 erforscht worden. Charakteristisch für sie ist ihr bedeutender Einfluss auf das Aktinzytoskelett. Untersuchungen an kultivierten Fibroblasten haben gezeigt, dass die Aktivierung von RhoA, Rac1 und Cdc42 einen Umbau des Aktinzytoskeletts im Sinne einer Bildung von Stressfasern, fokalen Adhäsionen, Lamellipodien beziehungsweise filopodischen Mikrospikes zur Folge hat. Es gibt außerdem Hinweise, dass sie neben der vorstehend beschriebenen Funktion der Regulation des Aktinzytoskeletts einen bedeutsamen Einfluss auf die Zellpolarität, Membrantransportbahnen und die Aktivierung von Transkriptionsfaktoren haben. Darüber hinaus wirken sie regulatorisch auf Zellproliferation und neuronale Differenzierung (BAGRODIA et al. 1995, COSO et al. 1995, MINDEN et al. 1995, ZHANG et al. 1995, BROWN et al. 1996, VAN AELST u. D'SOUZA-SCHOREY 1997, HALL 1998). Für die Neuritenbildung sind die Rho-GTPasen unabdingbar. Die Untersuchung, in welcher Art und Weise die Beeinflussung der Rho-GTPasen zu einer Veränderung der Neuritenbildung führt, ist ein wesentlicher Bestandteil der vorliegenden Arbeit. 2.3 Initialisierung und Auswachsen von Neuriten Durch die Regulation der zytoskelettalen Reorganisation und der mikrotubulären Orientierung üben Rho-GTPasen einen tief greifenden Einfluss auf die Neuritogenese aus. Membranrezeptoren werden durch extrazelluläre Signalmoleküle aktiviert. Im Verlauf der daraufhin einsetzenden Neuritogenese wachsen zylindrische Fortsätze aus dem Zellkörper der Neuronen, aus denen sich später die Axone beziehungsweise Dendriten herausbilden. Entsprechend ihrer genetischen Codierung entwickelt jede Neuronenpopulation ihre Neuriten individuell verschieden. In jedem Fall verliert die Zelle ihre ursprünglich runde Gestalt durch die Ausbildung einer oder mehrerer Wachstumsknospen, aus denen sich jeweils ein Neurit entwickeln wird. Für den Prozess der Entstehung und der weiteren Entwicklung der Neuriten sind koordinierte Wechselwirkungen notwendig zwischen dem Aktinzytoskelett einerseits, das die Energie für das Zellwachstum bereitstellt, und dem mikrotubulären Netzwerk, das der Stabilisierung und dem Erhalt der Neuriten dient, andererseits (DA SILVA u. DOTTI 2002). Die Rho GTPasen üben zum Teil gegensätzliche Effekte auf das Nervenzellwachstum aus: Während Rho die Initialisierung der Neuriten verhindert und darüber hinaus deren Retraktion bewirkt, fördern Rac und Cdc42 die Herausbildung von Lamellipodien und Filopodien und sind damit für die Schaffung von Nervengewebe essentiell (TIGYI et al. 1996, KOZMA et al. 6 1997, KRANENBURG et al. 1997, SEBÖK et al. 1999, BROUNS et al. 2001, FUJITA et al. 2001). Dieses Prinzip der antagonistischen Wirkung mag in der Modulation von neuronalen Fortsätzen, die für die Wegfindung der Neuriten und die synaptische Plastizität wesentlich sind, eine gewichtige Rolle spielen. Beispielsweise steigert konstitutiv-aktives Rac1 beziehungsweise reduziert dominant-negatives Rac1 die Neuritenausdehnung in dissoziierten Hippocampusneuronen der Ratte (SCHWAMBORN u. PÜSCHEL 2004). Konstitutiv-aktives Rho hingegen blockiert das Auswachsen von Neuriten von kultivierten hippocampalen Neuronen (DA SILVA et al. 2003, SCHWAMBORN u. PÜSCHEL 2004). Durch die Ausschaltung von Rho durch C3 Exoenzym wird sowohl in Hippocampusneuronen der Ratte als auch in Hinterwurzelganglien des Huhnes die Neuritenentfaltung begünstigt (JIN u. STRITTMATTER 1997, SARNER et al. 2000, DA SILVA et al. 2003, FOURNIER et al. 2003). Als wichtigste Information erschließt sich aus vielen Studien, dass die Rho GTPasen für die Neuritenbildung von Primärneuronen von substantieller Bedeutung sind und eine präzise Regulation der Prozesse für die physiologische Reifung von Nervenzellen erforderlich ist. 2.4 Einwirkung der Rho Aktivität auf die Dendriten- und Spinebildung Die Dendriten bilden eine komplexe baumähnliche Struktur aus. Damit sind sie in der Lage, Informationen von anderen Neuronen zu empfangen und zu verarbeiten. JAN u. JAN (2001) haben beschrieben, dass der Grad der dendritischen Verzweigung annähernd der Größenordnung des Informationsflusses an das Neuron entspricht. Während des Prozesses der Dendritenausbreitung bilden sich stetig winzige fadenähnliche Gebilde, die Filopodien, aus und ziehen sich wieder zurück. Sie bleiben bestehen, wenn eine Stabilisierung in dem Sinne eintritt, dass das Maß der Neubildung dasjenige des Rückzugs übertrifft. Aus dieser Zunahme an Substanz resultiert eine räumliche Ausdehnung des Neurons (CLINE 2001, VAN AELST u. CLINE 2004). Untersuchungen haben gezeigt, dass konstitutiv-aktives Rho drastisch das Wachstum des Dendritenbaums von Pyramidenzellen in organotypischen Schnitten des Hippocampus der Ratte (NAKAYAMA et al. 2000) und kultivierten Hippocampusneuronen von Ratte und Maus reduziert (AHNERT-HILGER et al. 2004, PILPEL u. SEGAL 2004). Hingegen erhöht die dominant-negative Rho-Mutante signifikant die Wachstumsrate des Dendritenbaums und damit seine Ausdehnung in kultivierten Hippocampusneuronen der Maus (AHNERTHILGER et al. 2004). Dieser negative Einfluss von Rho spiegelt sich in zahlreichen 7 Modellsystemen wider, und zwar nicht nur im Bereich der Wirbeltiere sondern auch der Wirbellosen (RUCHHOEFT et al. 1999). Ein frühes Entwicklungsstadium der Spines stellen die Filopodien dar. Sie sind winzige Vorwölbungen der Dendritenmembran, lang und dünn in ihrer Gestalt, mit weniger F-Aktin als die Spines. Filopodien befinden sich auch an den Wachstumsknospen der Dendriten, wo sie unabhängig von neuronaler Aktivität für Wachstum und Verzweigung der Dendriten sorgen. Die als frühes Stadium der Spines am Dendritenschaft sitzenden Filopodien sind hingegen für ihre Weiterentwicklung von neuronaler Aktivität abhängig (PORTERACAILLIAU et al. 2003). Sie befinden sich im steten Wechsel zwischen Wachstum und Rückzug, um die Begegnungschance mit wachsenden Zielaxonen zu erhöhen. Sie haben eine durchschnittliche Lebensdauer von nur etwa zehn Minuten (ZIV u. SMITH et al. 1996). Sobald der Kontakt zu einem Axon hergestellt ist, bildet sich entweder physikalisch in Form von Zell-Zell-Adhäsion oder chemisch über lokal freigesetzte Signale eine synaptische Verbindung aus. Reifungsschritte schließen sich an. Während der gesamten Synaptogenese ist eine Kommunikation zwischen den wachsenden prä- und postsynaptischen Teilen der Synapse erforderlich (GODA u. DAVIS 2003). Die Filopodien werden schließlich durch Spines ersetzt (MATUS 2005), die im Vergleich zu den Filopodien eine gut abgegrenzte Kopf- und Halsstruktur aufweisen (HARRIS 1999, HERING u. SHENG 2001). Das Zytoskelett sowohl der Spines als auch der Filopodien basiert vorwiegend auf Aktin. Untersuchungen von BONHOEFFER u. YUSTE (2002) und LISMAN (2003) legen nahe, dass die Rho GTPasen eine entscheidende Rolle bei Bildung, Fortbestand und physiologischer Funktion der Spines spielen. Der Austausch der vorhandenen Rho-GTPasen der Neuronen durch Mutanten ergibt charakteristische Spine-Phänotypen. Dabei vermindert konstitutiv-aktives RhoA Spinedichte und Spinelänge von Pyramidenzellen in Hippocampusschnitten von Ratte und Maus und kultivierten Hippocampusneuronen der Ratte (NAKAYAMA et al. 2000, TASHIRO et al. 2000, PILPEL u. SEGAL 2004). Die Hemmung von Rho führt zu unterschiedlichen Effekten. Durch die Inhibition von Rho mittels C3 Exoenzym kommt es zu einer Verlängerung des Spinehalses, einer unerwarteten Verringerung der Spinedichte und einer erhöhten Filopodiendichte in kultivierten Hippocampusneuronen der Ratte (PILPEL u. SEGAL 2004). Darüber hinaus werden dadurch Dichte und Länge von Spines einiger Kortikal- und Hippocampus-Pyramidenzellen der Maus in organotypischen Schnitten erhöht (TASHIRO et al. 2000), obwohl dominant-negatives RhoA die Spinedichte von Pyramidenzellen aus 8 Hippocampusschnitten der Ratte nicht verändert. Diese widersprüchlichen Forschungsergebnisse mögen darin begründet sein, dass lokale Unterschiede der RhoAAktivität auftreten. Bisherige Forschungsarbeiten befassen sich mit der Untersuchung der Toxinwirkung auf dissoziierte Zellen, deren Typ teilweise nicht bestimmt wurde (AHNERT-HILGER et al. 2004, PILPEL u. SEGAL 2004). Nicht untersucht worden ist, ob das C3 Exoenzym auf Neurone, die sich im Gewebeverband befinden, die gleichen in der Literatur beschriebenen Wirkungen ausübte oder ob es zu abweichenden Beobachtungen käme. Zusammenfassend belegen die Untersuchungen, dass die Rho GTPasen eine herausragende Rolle bei Bildung und Funktion von unterschiedlichsten Strukturelementen dieser hochdifferenzierten Zellen spielen, vor allem durch die Regulation des zellulären Zytoskeletts. Außerdem kommt ihnen eine wichtige Funktion in der dendritischen und synaptischen Plastizität zu, was als essentiell für Lernen und Gedächtnis angesehen wird. Ihre große Bedeutung wird untermauert durch das Auftreten von zahlreichen Dysfunktionen des Nervensystems in Verbindung mit Störungen der Rho GTPase-Signalgebung. Als Fernziel besteht die Möglichkeit einer therapeutischen Intervention mit Ziel einer Beeinflussung der Rho GTPase-Aktivität. 9 3 Tiere, Material und Methoden 3.1 Versuchsplanung Im Rahmen der vorliegenden Dissertation sollte an identifizierten CA1 Pyramidenzellen in kultivierten Hippocampusschnitten der Maus die Wirkung von clostridialem C3 Toxin auf die Dendritendifferenzierung und die Spinebildung analysiert werden. Zur Darstellung der CA1 Pyramidenzellen wurden diese intrazellulär mit dem Tracer Biocytin gefüllt, um sie anschließend bezüglich der Parameter Dendritenlänge, Dendritenverzweigung, Gesamtzahl an Spines und Spinedichte zu vergleichen. Es wurden drei unterschiedlich behandelte Untersuchungsgruppen gebildet. Gruppe 1 bestand aus Hippocampuskulturen, die mit normalem serumhaltigem Kulturmedium inkubiert worden waren. Da das Serum möglicherweise aktive Proteasen enthielt, die die Wirkung des Toxins hätten beeinflussen oder neutralisieren können, bestand die zweite Gruppe aus Kulturen, die zwölf Stunden vor der intrazellulären Füllung auf serumfreies Medium gewechselt worden waren. Diese Gruppe diente als serumfreie Kontrolle. Die dritte Gruppe stellte die Experimentalgruppe dar, welche unmittelbar vor der Tracerinjektion für vier bis sechs Stunden mit 120 nM C3 bot Toxin inkubiert worden war. Auch bei dieser Gruppe wurde am Abend vor dem Experiment das normale Medium durch serumfreies Medium ersetzt und vier bis sechs Stunden vor dem Experiment das Toxin zugesetzt. Die Bereitstellung der intrazellulär mit Biocytin gefüllten Zellen erfolgte durch Herrn PD Dr. Rudi Deisz, Zentrum für Anatomie, Charité Universitätsmedizin, Berlin. Das C3 bot Toxin wurde von Frau Prof. Dr. Gudrun Ahnert-Hilger, Zentrum für Anatomie, Charité Universitätsmedizin, Berlin, zur Verfügung gestellt. 3.2 Schnittkulturen 3.2.1 Tiere Zur Herstellung der Schnittkulturen wurden neugeborene C57 BL/6 Mäuse (P0-P1) der Tierhaltung des Zentrums für Anatomie, Institut für Zellbiologie und Neurobiologie, Charité Universitätsklinik, Berlin verwendet. Sie wurden entsprechend den Europäischen Richtlinien für Tierschutz bei einem 12-Stunden-Tag-Nacht-Rhythmus bei konstanter Raumtemperatur und –feuchtigkeit gehalten. Futter und Wasser standen ihnen ad libitum zur Verfügung. Genehmigungsbescheid: Aktenzeichen T 0340/00 10 Genehmigende Behörde: Landesamt für Arbeitsschutz, Gesundheitsschutz und technische Sicherheit Berlin Sitz und Datum: Berlin, 12.02.2001 3.2.2 Herstellung der Schnittkulturen Die hippocampalen Schnittkulturen wurden nach der in unserer Arbeitsgruppe etablierten Methode hergestellt (SAVASKAN et al. 2002). Die Mäuse wurden am ersten postnatalen Tag (P0) dekapitiert, die Kopfschwarte und die Schnittfläche mit 100-prozentigem Isopropanol desinfiziert. Alle weiteren präparativen Arbeitsschritte erfolgten unter sterilen Bedingungen. Unter einer Laminar Flow Bench wurden zunächst Kopfhaut, Schädeldach und Hirnhäute jeweils mit frischem Präparierbesteck entfernt. Anschließend wurde das Gehirn mit Hilfe eines Spatels aus der Schädelhöhle heraus gehoben und in 4°C kaltes Präparationsmedium (Lösungsansätze, Nr. 1) überführt. Nachdem die rostralen Gehirnanteile auf Höhe des Hypophysenstiels und die kaudalen Anteile auf Höhe des Mittelhirns mit einem Skalpell abgetrennt worden waren, wurde ein Mediansagittalschnitt gelegt und unter einer Stereolupe die Hippocampi beider Hemisphären herauspräpariert. Im Anschluss wurde der mid-septotemporale Anteil des Hippocampus mit einem McIllwainTissue-Chopper in 400 µm dicke transversale Scheiben geschnitten. Mit zwei Spateln wurden diese Schnitte vorsichtig von der Schneideunterlage abgehoben und in Präparationsmedium überführt. Pro Hippocampus erhielt man durchschnittlich fünf Schnitte. Unter visueller Kontrolle wurden diejenigen Hippocampusschnitte zur Kultivierung ausgewählt, die keine sichtbaren Verletzungen aufwiesen. Die Schnitte wurden dann in einem Tropfen Präparationsmedium auf einen Spatel aufgenommen und auf angefeuchtete Millicell CMMembran-Einsätze (Millipore 0,2 µm) übertragen. Die Einsätze wurden schließlich in 6-well Platten (Becton-Dickinson) gelegt, die mit jeweils 1200µl Inkubationsmedium (Lösungsansätze, Nr. 2) gefüllt waren. 3.2.3 Inkubation Die Schnittkulturen wurden bei 37°C, 5% CO2 und 100% Luftfeuchtigkeit in einem Zellkulturschrank 14 Tage lang inkubiert. Das Inkubationsmedium wurde dreimal pro Woche gewechselt. Kulturen, die als serumfreie Kontrolle beziehungsweise als toxinbehandelte Experimentalgruppe dienen sollten, wurden ca. 12 Stunden vor der intrazellulären Füllung über Nacht auf serumfreies Inkubationsmedium (Lösungsansätze, Nr. 3) gesetzt. Dem 11 Medium der Kulturen der Toxin-Behandlungsgruppe wurden am Experimentaltag 120 nM C3 bot zugegeben und es wurde für mindestens vier bis sechs Stunden inkubiert. 3.3 Biocytinfüllung der Pyramidenzellen Zur Darstellung der CA1 Pyramidenzellen in ihrer Gesamtheit mit Dendritenbaum und Axon wurden die Zellen am Inkubationstag (DIV) 14 mit dem anterograden Tracer Biocytin (1%ige Lösung) intrazellulär gefüllt. Bei der Biocytinfüllung kam das Patch Clamp Verfahren zur Anwendung. Eine Mikropipette aus Glas, die durch ein besonderes Herstellungsverfahren unter vorsichtiger Erhitzung über einer Flamme und kontinuierlichem maschinellen Zug auf einen Durchmesser ihrer Spitze von bis zu weniger als einem Mikrometer ausgezogen worden war, wurde dabei vorsichtig auf die Zellkörpermembran der zu füllenden Zelle gesetzt. Die Zelle befand sich dazu in einer Begasungskammer mit ACSF (Lösungsansätze, Nr. 4). Durch Erzeugen eines leichten Unterdrucks wurde eine dichte Verbindung zwischen Glaswand und Membran geschaffen, eine sogenannte „On-Cell“- Konfiguration. Verstärkte man nun den Unterdruck oder legte man einen kurzen Spannungspuls an, wurde die Zellmembran durchbrochen und die Glaspipette bildete in der „Whole-Cell“-Konfiguration eine Einheit mit der ganzen Zelle. Zugleich wurde über die Pipette das gelöste Biocytin in die Zelle appliziert. Biocytin, ein Komplex aus Biotin und Lysin, wurde dann über den aktiven axonalen Transport anterograd bis in die Axonendigungen transportiert. Darüber hinaus verteilte es sich im gesamten Dendritenbaum. Anschließend wurden die aufgrund ihrer Lokalisation im Pyramidenzellband mit Biocytin intrazellulär gefüllten CA1 Neurone für eine Stunde im Brutschrank reinkubiert, um den axonalen Transport des Biocytins und seine Diffusion in die peripheren Dendritenabschnitte zu ermöglichen. 3.4 Fixierung und Visualisierung der Pyramidenzellen Der Reinkubation folgte die Fixation der Schnittkulturen. Dazu wurden die Zellkultureinsätze in eine Petrischale mit 1%igem Paraformaldehyd (PFA) (Lösungsansätze, Nr. 7) und 1%igem Glutaraldehyd (Lösungsansätze, Nr. 8) in 0.1 M Phosphatpuffer (PB) (Lösungsansätze, Nr. 5) bei einer Temperatur von 4°C überführt und über Nacht bei Raumtemperatur stehen gelassen. Am Folgetag wurde dreimal für jeweils 10 min mit 0,1 M PB gespült. Für eine Zeitdauer von 10 min wurden die Schnitte in 3% H2O2 in 0,1 M PB eingelegt und damit die endogene Peroxidase inaktiviert. Es folgten 6 bis 8 Spülvorgänge in 0,1 M PB für jeweils 10 min. 12 Zur Detektion der biocytinmarkierten Zellen und ihrer Fortsätze kam die ABC-Methode (HSU et al. 1981) zur Anwendung. Dazu wurden die Schnitte über Nacht mit dem Avidingekoppelten Meerrettichperoxidasekomplex ABC-Elite (Vector-Laboratories, Burlingame, California, USA) (1:100 in 0,1 M PB) bei 4°C im Dunkeln auf dem Schüttler inkubiert. Am folgenden Tag wurde wieder 6 bis 8x für jeweils 10 min in 0,1 M PB gespült. Die Visualisierung der Zellen basiert auf einer Nickel-Kobalt-intensivierten DAB-Reaktion (ADAMS 1981); die Schnittkulturen wurden jeweils zunächst in einer Lösung aus 0,07%igem 3,3’-Diaminobenzidintetrahydrochlorid (DAB) (Sigma), 15 µl Cobaltchlorid (CoCl2) und 20 µl Nickelammoniumsulfat (NiNH4SO4) in 5 ml DAB für 25 min vorinkubiert. Durch Zugabe von 16,5 µl H2O2 (0.001%) wurde die Peroxidasereaktion in Gang gesetzt. Durch die Peroxidasereaktion wird das farblose DAB zu einem schwarzblauen Reaktionsprodukt umgewandelt. Der Ablauf dieser Färbereaktion wurde für 3 bis 5 min unter einem Lichtmikroskop visuell kontrolliert und schließlich durch 3x Spülen für jeweils 10 min in 0,1 M kaltem PB gestoppt. Im Anschluss wurden die Schnitte in einem Tropfen Aqua bidest. auf einen gelatinierten Objektträger (Lösungsansätze, Nr. 6) aufgezogen und bei Raumtemperatur über Nacht getrocknet. Daraufhin erfolgte eine stufenweise Dehydrierung in einer aufsteigenden Alkoholreihe (30, 50, 60, 70, 80, 96, 100%, je 3 min), wobei die Kulturen auf dem Objektträger mit Alkohol überschichtet wurden und dieser vorsichtig wieder abgesaugt wurde. Zum Schluss wurden die Präparate mit Entellan überschichtet und mit einem Deckglas eingedeckelt. 3.5 Resultierende Behandlungsgruppen Aufgrund der experimentellen Anordnung ergaben sich drei Experimentalgruppen: • Serumhaltige Kontrollgruppe • Serumfreie Kontrollgruppe • C3 bot Toxin-behandelte Experimentalgruppe 13 Verworfen wurden diejenigen Zellen, bei denen die intrazelluläre Füllung zu sichtbaren Schäden des Somas oder der Dendriten geführt hat. Außerdem wurden jene Zellen nicht ausgewertet, bei denen es infolge einer Leckage zu unspezifischen DAB-Ablagerungen kam und eine Auswertung nicht machbar war. Auf einer Schnittkultur befanden sich ein bis maximal drei markierte CA1 Pyramidenzellen. Zur Übersichtlichkeit sind die einzelnen Arbeitsschritte der Zellbehandlung beginnend von der Inkubation der Kulturen bis zur Analyse der Biocytin-gefüllten Neurone in Abbildung 1 schematisch zusammengefasst. Abbildung 1: Schematische Darstellung des experimentellen Protokolls Dem Schema ist zu entnehmen, dass sich die 3 Experimentalgruppen (1, 2 und 3) erst gegen Ende der Inkubationszeit unterscheiden: An DIV 13 wird bei den Kulturen der serumfreien Kontrollgruppe sowie der toxinbehandelten Experimentalgruppe das normale Medium durch serumfreies Medium ersetzt. An DIV 14 wird den Kulturen der Experimentalgruppe (3) 120 nM zugesetzt. Die weiteren Abläufe sind in den 3 Behandlungsgruppen identisch. 14 C3 bot dem Medium Zur Illustration der so gewonnenen CA1 Pyramidenzellen in hippocampalen Schnittkulturen dient die Abbildung 2. Abbildung 2: Beispiel einer hippocampalen Schnittkultur mit intrazellulär Biocytin-gefüllter CA1 Pyramidenzelle an DIV 14 Man erkennt neben dem Zellkörper (schwarzer Pfeil) deutlich den Apikaldendriten (weißer Pfeil) mit seinen Verzweigungen. Am basalen Pol des Somas ist ein Basaldendrit zu erkennen. Die Markierung (Punkte) umreißt das Pyramidenzellband (CA1, CA3) und den Gyrus dentatus (DG). 15 3.6 Lösungsansätze 1.) Präparationsmedium für Schnittkulturen: 50 ml Minimum Essential Medium (MEM) (Gibco) in zweifacher Konzentration, sterilfiltriert, mit 1 ml Glutamin und 49 ml sterilem Aqua ad injectione. Mittels 1 M NaOH auf einen pH-Wert von 7,35 eingestellt. 2.) Inkubationsmedium: 25 ml MEM (2x), sterilfiltriert, 25 ml Basal Medium Eagle (BME) ohne Glutamin, 25 ml hitzeinaktiviertes Horse Serum (Gibco), 1 ml Glutamin und 3,125 ml 20% sterile Glukoselösung (0,625%) mit sterilem Aqua ad injectione auf 100 ml aufgefüllt. Mittels steriler 1 M NaOH bzw. HCl wird der pH-Wert auf 7,2 bis 7,3 eingestellt. 3.) Serumfreies Inkubationsmedium: 50 ml MEM (2x), sterilfiltriert, 25 ml BME, 1 ml Glutamin (200 mM), 3.125 ml 20% Glucose, mit 20,875 ml Aqua ad injectione auf 100 ml aufgefüllt. Mit sterilem 1 M NaOH wird der pH-Wert auf 7,2 bis 7,3 eingestellt. 4.) Medium der Begasungskammer: ACSF (artificial cerebrospinal fluid) [in mM: 124 NaCl; 3 KCl; 1,3 MgSO4; 1,24 NaH2PO4; 2,4 CaCl2; 26 NaHCO3; 10 d-Glukose; pH 7,4]. Das Medium ähnelt in seiner Zusammensetzung dem Liquor cerebrospinalis und simuliert eine natürliche Zellumgebung. 5.) 0,1 M Phosphatpuffer (PB): 28,4 g wasserfreies Na2HPO4 werden in 1000 ml Aqua bidest. und 8,3 g NaH2PO4-H2O in 300 ml Aqua bidest. gelöst. Letztere Lösung wird zu ersterer Lösung hinzu titriert, bis ein pH-Wert von 7,35 erreicht ist. 6.) Gelatinisierte Objektträger: Die Objektträger werden nach dem Spülen und vollständigen Trocknen kurz in eine filtrierte Gelatinelösung eingetaucht und im Anschluss über Nacht im Wärmeschrank bei 37°C getrocknet. Gelatinelösung: 5 g Gelatine (Bloom 300) werden in 50 ml Aqua bidest. unter Erwärmung gelöst, mit Aqua bidest. auf 1000 ml aufgefüllt und 0,5 g Chromalaun (CrK(SO4)2 x 12 H2O) sowie 10 mg Thymol zugegeben. 7.) Fixativ: 99 ml PB werden mit 1 ml Paraformaldehyd (PFA) versetzt. 8.) Glutaraldehyd 1%ig: 4 ml 25%iges Glutaraldehyd und 1 ml PFA in 95 ml PB 16 3.7 Quantitative Auswertung der dendritischen Parameter 3.7.1 Rekonstruktion der CA1 Pyramidenzellen Die Schnittkulturen mit biocytingefüllten CA1 Pyramidenzellen wurden randomisiert nummernkodiert, so dass die Rekonstruktion in einer Blindstudie durchgeführt wurde und dem Experimentator nicht bekannt war, welche Schnittkultur welcher Experimentalgruppe zuzuordnen war. Zur Rekonstruktion der Biocytin-markierten CA1 Pyramidenzellen wurden diese mittels eines Zeiss Axiophot Lichtmikroskops mit motorisiertem Kreuztisch bei 63facher Vergrößerung des Objektivs mit Ölimmersionstechnik betrachtet und mit dem Programm Neurolucida (Firma MicroBrightField) in ihrer x-, y- und z-Ausdehnung rekonstruiert. Neurolucida ist eine spezielle Software zum Erstellen zwei- und dreidimensionaler anatomischer Abbildungen und zur Rekonstruktion von Nervenzellen geeignet. Mit Hilfe eines Joysticks werden unter visueller Kontrolle präzise Bewegungen des motorisierten Tisches gesteuert und über eine Mikrometerschraube eine exakte Fokussierung des Objektivs ermöglicht. Per Digitalkamera wird das mikroskopische Bild der zu rekonstruierenden Struktur auf einen Computerbildschirm übertragen. Im Folgenden werden die wesentlichen Arbeitsschritte skizziert: Mit der Rekonstruktion der Zelle wird am Zellkörper begonnen. Bei entsprechender Einstellung wird die Kontur des Somas mit der Computermaus exakt auf dem Bildschirm umfahren und diese Daten gespeichert. Im zweiten Schritt erfolgt die Erfassung der Zellfortsätze. Die Software ist so konfiguriert, dass zwischen Axon, Basal- und Apikaldendrit unterschieden werden kann. Analog zur Vorgehensweise bei der Rekonstruktion des Zellkörpers kann jeder Fortsatz in seinem Verlauf bis zum Ende verfolgt werden. Dabei kann der Durchmesser jeden Fortsatzes parallel mit erfasst werden (diese Daten wurden nicht in die Auswertung einbezogen). Während der Rekonstruktion der Zelle wird stets die Einstellung der Z-Achse mit der Mikrometerschraube reguliert, wodurch die Zellausläufer in ihrer dreidimensionalen Ausdehnung durch die gesamte Tiefe des Schnittes verfolgt werden können. Als Arbeitsvorgabe wurden nur Zellausläufer verfolgt, die mit dem Soma in Verbindung standen, da eine Zuordnung eines isolierten Dendritensegments zu der zu rekonstruierenden Zelle nicht zu treffen war, wenn mehrere Zellen pro Kultur Biocytin-gefüllt waren. 17 Nach jedem Verzweigungspunkt wechselt das Programm automatisch die Farbe des rekonstruierten Abschnitts, so dass jeder Verzweigungsordnung konstant eine bestimmte Farbe zugeordnet wird. Als weitere Option des Systems können an beliebiger Stelle so genannte markers gesetzt werden. Hier wurden sie dazu verwendet, um die Spines einzuzeichnen. 3.7.2 Analyse der Daten Um sämtliche mit Neurolucida erarbeitete Daten zu analysieren, steht dem System ein weiteres Programm desselben Herstellers, der Neuroexplorer, zur Verfügung. Er ist in der Lage, jeden beliebigen Parameter einer rekonstruierten Zelle zu errechnen und tabellarisch darzustellen. Aus der großen Fülle der Information wurden die für die Arbeit relevanten Datentabellen ausgewählt und in Excel-Dateien exportiert. 3.8 Statistische Tests Die statistische Auswertung wurde mit Hilfe des Statistikprogramms SAS 8.2 in der Abteilung Biomathematik der Universität Freiburg (mit Unterstützung durch Herrn Prof. Dr. Schulte Mönting) durchgeführt. Zur Abklärung, ob die Behandlung mit C3 bot beziehungsweise der Serumentzug zu quantitativen Effekten bei der Dendritenmorphologie beziehungsweise Spinezahl oder -dichte führt, wurden folgende statistischen Tests durchgeführt. Da für die Anwendung der Varianzanalyse normalverteilte Grundgesamtheiten mit gleichen Varianzen Voraussetzung sind, wurde in dieser Analyse der nichtparametrische KruskalWallis Test (MONTGOMERY 2001) angewandt. Dieser Test eignet sich zum Vergleich zwischen drei oder mehr unabhängigen Gruppen von Daten. Im Gegensatz zur parametrischen Varianzanalyse verlangt er keine besonderen Voraussetzungen bezüglich der Datenverteilung (z.B. Normalverteilung). Seine einzige Forderung ist das Vorliegen von ordinalskalierten Variablen. Ähnlich wie andere nicht-parametrische Tests verwendet der Kruskal-Wallis Test zur Berechnung der Statistik die Rangplätze der Daten anstatt ihrer Rohwerte. Da er keine Vorgaben zur Verteilung der Daten macht, ist er allerdings nicht so stark wie die Varianzanalyse. 18 Neben dem Kruskal-Wallis Test kam zur quantitativen Datenanalyse auch die Varianzanalyse (HARTUNG u. ELPELT 1989), die eine Normalverteilung der Grundgesamtheiten vorschreibt, zur Anwendung. Ermöglicht wurde ihre Anwendung durch logarithmische Transformation der Daten. Dadurch wird eine definierte Verteilung der Daten erzielt und die Voraussetzung der Normalverteilung ist erfüllt. Wie auch der Kruskal-Wallis Test wurde die Varianzanalyse dazu verwendet, statistische Hypothesen über signifikante Unterschiede zwischen den Mittelwerten zu überprüfen. Sowohl der Kruskal-Wallis Test als auch die Varianzanalyse zeigen an, ob zwischen drei oder mehr unabhängigen Gruppen signifikante Unterschiede der Mittelwerte vorliegen. Allerdings machen sie keine Aussage darüber, zwischen welchen beiden Mittelwerten ein signifikanter Unterschied auftritt. Das zu bestimmen, ist die Aufgabe eines post hoc Tests. Der Tukey Test (LINTON u. HARDER 2007), der für diese Analyse herangezogen wurde, stellt einen solchen post hoc test dar, der einen paarweisen Vergleich der Mittelwerte durchführt, um die sich signifikant unterscheidenden Gruppen anzuzeigen. In den Graphiken werden signifikante Unterschiede zwischen den Mittelwerten der Gruppen wie folgt gekennzeichnet: p < 0,05 (Irrtumswahrscheinlichkeit α < 5%) *** p < 0,01 (Irrtumswahrscheinlichkeit α < 1%) * 19 4 Ergebnisse 4.1 Allgemeine Befunde zu den CA1 Pyramidenzellen Zur Rekonstruktion wurden nur Kulturen herangezogen, die keine Anzeichen von Degeneration erkennen ließen. Insgesamt sind für diese Arbeit 120 Slicekulturen angelegt worden. Bei der visuellen Kontrolle unter dem Mikroskop wiesen 52 dieser Kulturen eine Biozytinmarkierung auf. Einige Zellen waren aufgrund einer Leckage nicht vollständig gefüllt, sondern zeigten stattdessen durch ausgelaufenes Biozytin einen dunklen Hof um den Zellkörper herum. Diese unspezifischen Biocytinablagerungen machten eine Rekonstruktion unmöglich. Unter allen rekonstruierten Zellen wurden aus jeder der drei Behandlungsgruppen randomisiert 6 Zellen ausgewählt und deren Daten quantitativ analysiert. 4.2 Morphologie der hippocampalen CA1 Pyramidenzellen Die CA1 Pyramidenzellen aller drei Behandlungsgruppen zeigten unter dem Mikroskop folgende für diese Zellart typische Merkmale: einen dreieckigen bis ovoiden Zellkörper, einen Apikaldendriten und mehrere Basaldendriten. Alle Dendriten waren mehrfach verzweigt und dicht mit postsynaptischen Dornen besetzt. Vom Soma abgehende Biocytin-gefüllte Axone waren bei einem Großteil der Zellen sichtbar. Diese wurden nicht weiter analysiert. Nachfolgend werden aus jeder der drei Behandlungsgruppen beispielhaft zwei CA1 Pyramidenzellen dargestellt. Die Pyramidenzellen wurden bei unterschiedlicher Vergrößerung (10fache, 20fache und 63fache Vergrößerung des Objektivs) digital photographiert und erfasst. Zu den ausgewählten Beispielen von CA1 Pyramidenzellen wird auch die NeurolucidaRekonstruktion als Farbausdruck abgebildet. Da jeder Dendritenordnung eine Farbkodierung zugrunde liegt, lässt sich somit der Grad der Differenzierung leicht erkennen. 20 4.2.1 Serumhaltige Kontrollgruppe In der serumhaltigen Kontrollgruppe zeigten alle intrazellulär gefüllten und analysierten CA1 Pyramidenzellen ihre typische Erscheinungsform. Deutlich erkennbar war ein Apikaldendrit, der sich distal vom Soma aufzweigte. Von ihm gingen mehrere Seitendendriten ab. Am basalen Pol der Zellen entsprangen zahlreiche Basaldendriten. Alle Dendriten zweigten sich in Dendriten weiterer Ordnungen auf. Sowohl Apikal- als auch Basaldendriten waren dicht mit postsynaptischen Spines besetzt (siehe Abbildung 3B und C). Abbildung 3: Biocytin-gefüllte CA1 Pyramidenzelle der serumhaltigen Kontrollkultur am DIV 14. (A) Die Übersichtsaufnahme zeigt den intensiv angefärbten Zellkörper (schwarzer Pfeil) mit dem Apikaldendriten (weißer Pfeil). (B) zeigt den in (A) eingerahmten Dendritenabschnitt des Apikaldendriten bei hoher Vergrößerung. Deutlich sind einige postsynaptische Spines mit ausgeprägtem Spinekopf zu erkennen (Pfeile). (C) Software-unterstützte Rekonstruktion derselben CA1 Pyramidenzelle. Das Axon ist mit einem Pfeil gekennzeichnet. Die verschiedenen Ordnungen der Dendritenverzweigung sind verschiedenfarbig abgebildet (Stammdendriten blau, Dendriten 2. Ordnung gelb usw.). Die dendritischen Spines sind als kleine rote Punkte gekennzeichnet. 21 Abbildung 4: Zweites Beispiel einer CA1 Pyramidenzelle der serumhaltigen Kontrollgruppe (A) Die in der Übersichtsvergrößerung dargestellte biocytingefüllte CA1 Pyramidenzelle zeigt im Vergleich zu Abbildung 3 eine etwas unterschiedliche Morphologie. Der Apikaldendrit ist stärker verzweigt, ebenso die beiden Basaldendriten, die an der linken Seite des Zellkörpers entspringen. Der schwarze Pfeil zeigt auf das vom Soma abgehende Axon. Es ist eine zweite Zelle des Pyramidenzellbandes zu sehen (weißer Pfeil), die jedoch eine unvollständige Biocytinfüllung aufweist. Bei der Anwendung des Patch Clamp Verfahrens tritt gelegentlich über ein Leck in der Zellmembran Biocytin aus, was zur perizellulären dunklen Färbung durch das Reaktionsprodukt führt. Zellen wie diese wurden nicht in die Untersuchung mit einbezogen. (B) zeigt den in (A) eingerahmten Abschnitt des Apikaldendriten bei stärkerer Vergrößerung. Ein dichter Spinebesatz des Dendritenabschnitts ist erkennbar (Pfeile). (C) Rekonstruktion derselben CA1 Pyramidenzelle. Der Pfeil deutet auf das Axon. 22 4.2.2 Serumfreie Kontrollgruppe Der zeitlich begrenzte Serumentzug führte nicht zum Absterben der hippocampalen Schnittkulturen. Die intrazellulär mit Biocytin gefüllten CA1 Pyramidenzellen behielten unter diesen in vitro Bedingungen ihre typischen morphologischen Merkmale. Ein Apikaldendrit und mehrere Basaldendriten waren auch in dieser Behandlungsgruppe erkennbar. Generell zeigte der Vergleich der Mittelwerte für Dendritengesamtlänge (siehe Kapitel 4.3.1.1 und 4.3.1.2) und Spineanzahl (siehe Kapitel 4.3.3.1, 4.3.3.2 und 4.3.3.3) zwischen dieser und der serumhaltigen Kontrollgruppe eine leichte Abnahme der Werte. Da diese Parameter aber nicht signifikant abwichen, kann man folgern, dass der ca. zwölfstündige Serumentzug keinen messbar schädlichen Einfluss auf die CA1 Pyramidenzellen ausgeübt hat, welcher die Durchführbarkeit der Experimente in Frage gestellt hätte. Abbildung 5: CA1 Pyramidenzelle der serumfreien Kontrollgruppe (A) Übersichtsaufnahme: Der Zellkörper dieser CA1 Pyramidenzelle (schwarzer Pfeil) ist länglich ovoid. Deutlich sichtbar ist wiederum die polare Ausrichtung des Dendritenbaums. Apikal entspringt ein erst unverzweigter Stammdendrit (weißer Pfeil). Basaldendriten entspringen am gegenüber liegenden Zellpol. Zusätzlich hat diese Zelle einen seitlich am Soma abgehenden Dendriten. Alle Dendriten sind dicht mit Spines besetzt (B, C und D). 23 Abbildung 6: CA1 Pyramidenzellen der serumfreien Kontrollgruppe (A) Zwei biocytingefüllte Zellen der CA1 Region. Die Apikaldendriten sind deutlich sichtbar, während die Basaldendriten außerhalb der Fokusebene liegen (siehe schwarzen Pfeil und vergleiche C). Der weiße Pfeil zeigt auf unspezifische Biocytin-Präzipitate. (B) In der Ausschnittsvergrößerung (Rechteck in A) sieht man spinebesetzte Dendriten (Pfeil). (C) Rekonstruktion beider Nervenzellen aus (A) in rekonstruierter Form. Aus Gründen der besseren Übersichtlichkeit sind die beiden Zellen nicht in ihrem originalen Abstand zueinander, sondern separat abgebildet. Man sieht deutlich, dass die Pyramidenzellen in der Morphologie ihrer Dendriten variieren. 4.2.3 C3 bot Toxinbehandelte Experimentalgruppe Diese Behandlungsgruppe wurde wie die serumfreie Kontrollgruppe 12 Stunden vor Experimentbeginn (intrazelluläre Füllung mit Biocytin) unter Serumentzug inkubiert. Zusätzlich wurden dem Medium 4 bis 6 Stunden vor Beginn 120 nM C3 bot zugegeben. Alle Biocytin-gefüllten CA1 Pyramidenzellen dieser Gruppe zeigten eine polare Ausrichtung mit Apikaldendrit und Basaldendriten. Die Dendriten waren mit Spines besetzt. Insgesamt ähnelten die Zellen in ihrer Morphologie denen der serumfreien Kontrollgruppe. Dennoch hatte die Zugabe des clostridialen Toxins Einfluss auf die Dendritenmorphologie und den 24 Spinebesatz. Die quantitative Analyse der jeweiligen Parameter ist im Ergebnisteil unter Nummer 4.3 enthalten. Abbildung 7: CA1 Pyramidenzelle der C3 bot-toxinbehandelten Experimentalgruppe (A) In der Übersichtsabbildung ist eine der quantitativ analysierten Biocytin-gefüllten Pyramidenzellen der hippocampalen CA1 Region dargestellt. In der Abbildungsebene ist das Soma des Neurons nicht zu erkennen. Der schwarze Pfeil deutet auf den schwach verzweigten Apikaldendriten, der weiße Pfeil auf einen Basaldendriten. Ein Axon ist nicht sichtbar. (B) Detailvergrößerung des in (A, oberer Rahmen) eingefassten Dendritenabschnitts. Der dargestellte Ausschnitt beinhaltet sowohl apikale als auch basale Dendritenabschnitte. Alle Dendritensegmente haben Spinebesatz (siehe Pfeil). (C) Hohe Vergrößerung des in (A, unterer Rahmen) umfassten Abschnitts eines Basaldendriten mit postsynaptischen Dornen (siehe Pfeil). (D) Neurolucida-Rekonstruktion dieser CA1 Pyramidenzelle. Im Vergleich zur serumhaltigen Kontrollgruppe fällt ein wenig verzweigter Apikaldendrit auf. 25 Abbildung 8: 2. Beispiel einer CA1 Pyramidenzelle der C3 bot-toxinbehandelten Gruppe (A) In der Übersicht sieht man deutlich den ovoiden Zellkörper, der am Apikalpol einen schwach verzweigten Apikaldendriten entlässt (weißer Pfeil). Am gegenüberliegenden Pol des Somas entspringen mehrere ebenfalls gering verzweigte Basaldendriten. Das Axon ist Biocytin-gefüllt und schwach sichtbar (schwarzer Pfeil). (B) Abbildung des in (A) eingerahmten Abschnitts des Apikaldendriten in der Detailvergrößerung. Vereinzelt sind postsynaptische Spines zu erkennen (Pfeil). (C) Neurolucida-Rekonstruktion dieses Neurons 26 4.3 Quantitative Analyse Mit Hilfe der Neuroexplorer-Software wurden für jede der in die quantitative Analyse einbezogenen Zellen folgende Parameter berechnet: • Länge der Basaldendriten (gesamt) • Länge des Apikaldendriten (gesamt) • Anzahl der Verzweigungspunkte der Basaldendriten • Anzahl der Verzweigungspunkte des Apikaldendriten • Gesamtzahl der Spines • Anzahl der Spines an den Basaldendriten • Anzahl der Spines am Apikaldendriten • Spinedichte der gesamten Zelle • Spinedichte der Basaldendriten • Spinedichte des Apikaldendriten Anhand der ausgewählten statistischen Tests (Kruskal-Wallis-Test, Varianzanalyse und Tukey-Test) wurden Experimentalgruppen die Daten (serumhaltige auf signifikante Kontrollgruppe, Unterschiede serumfreie zwischen den Kontrollgruppe und toxinbehandelte Gruppe) untersucht. Genaue Angaben zu den statistischen Prüfverfahren sind dem Material- und Methodenteil, Kapitel 3.8, zu entnehmen. 27 4.3.1 Dendritenlänge 4.3.1.1 Gesamtlänge der Basaldendriten Die Pyramidenzellen der serumhaltigen Kontrollgruppe wiesen die längsten Dendritenbäume auf basaler Seite auf (1838,38 µm ± 1043,89 µm, MW ± SD). Die serumfreie Kontrolle hatte kürzere Basaldendriten (1651,55 µm ± 919,42 µm). Die C3 Toxin-Behandlungsgruppe zeigte die kleinsten Werte für die Länge der Basaldendriten (1020,32 µm ± 357,61µm). Die statistische Auswertung dieser Daten zeigte keine signifikanten Unterschiede zwischen den einzelnen Gruppen an: Der Kruskal-Wallis-Test ergab einen p-Wert von 0,14 und die Varianzanalyse zeigte nach logarithmischer Tranformation der Daten einen p-Wert von 0,23 an. Unter Toxineinfluss waren die Basaldendriten zwar im Mittel kürzer, aber nicht signifikant abweichend. Abbildung 9: Graphische Darstellung der Gesamtlänge der Basaldendriten. Jedes Symbol steht für den Wert eines der untersuchten Neurone. Die Querbalken entsprechen den Mittelwerten der jeweiligen Behandlungsgruppe. Den Mittelwerten sind nach oben und unten die Werte für ihre Standardabweichung zugeordnet. 28 4.3.1.2 Gesamtlänge der Apikaldendriten Ähnlich sieht es bei der Betrachtung der Länge der apikalen Dendritenbäume aus. Bezüglich dieses Merkmals unterschied sich die Serumkontrolle (2628,35 µm ± 1247,10 µm) sogar noch eindrücklicher sowohl von der serumfrei kultivierten Kontrollgruppe (1218,51 µm ± 624,34 µm) als auch der C3 bot-toxinbehandelten Gruppe (1075,90 µm ± 597,25 µm). Hier ergab die statistische Analyse der Daten ebenfalls keine signifikanten Unterschiede zwischen den Gruppen. In Bezug auf die Länge der Apikaldendriten bestand zumindest jedoch eine starke Tendenz zur Dendritenverkürzung unter serumfreien Kulturbedingungen, die unter Toxineinfluss nochmals deutlicher wurde. Eine etwaige toxinbedingte Wechselwirkung zwischen Apikal- und Basaldendriten (also eine Verlängerung der einen Dendritenart bei gleichzeitiger Verkürzung der anderen) konnte durch eine weitere Varianzanalyse ausgeschlossen werden. Abbildung 10: Graphische Darstellung der Apikaldendriten-Gesamtlänge. Jedes Symbol steht für die Länge eines Apikaldendriten. Der jeweilige Mittelwert der Gruppen ist als Querbalken, die entsprechende Standardabweichung senkrecht dazu dargestellt. 29 4.3.2 Verzweigungspunkte 4.3.2.1 Anzahl der Verzweigungspunkte der Basaldendriten Am stärksten verzweigt waren die Basaldendriten der serumhaltig kultivierten CA1 Pyramidenzellen (23.83 ± 10,34 Verzweigungspunkte), während der Grad der Verzweigung über die serumfrei kultivierte Kontrollgruppe (17,33 ± 10,23 Verzweigungspunkte) bis hin zur toxinbehandelten Gruppe (10,50 ± 3,62 Verzweigungspunkte) stark abnahm. Im Rahmen der statistischen Analyse zeigte der Kruskal-Wallis Test einen p-Wert von 0,07, die Varianzanalyse von 0,08 an. In Bezug auf die Anzahl der basalen Verzweigungspunkte traten also keine signifikanten Unterschiede zwischen den Gruppen auf. Abbildung 11: Graphische Darstellung der Verzweigungspunkte auf basaler Seite. Anhand der Symbole ist die Anzahl der basalen Verzweigungspunkte aller untersuchten Zellen abzulesen. Die Querbalken stehen für die Mittelwerte der einzelnen Behandlungsgruppen. Die Werte ihrer Standardabweichungen sind senkrecht dazu eingezeichnet. 4.3.2.1 Anzahl der Verzweigungspunkte der Apikaldendriten Die Serum-Kontrollgruppe wies stark verzweigte Apikaldendriten auf (21,83 ± 8,28 Verzweigungspunkte). Der Grad der Verzweigung nahm bei der serumfrei kultivierten Gruppe ab (13,83 ± 3,43 Verzweigungspunkte). Die Abnahme war noch ausgeprägter nach Toxinbehandlung (9,67 ± 5,89 Verzweigungspunkte). Beide statistischen Tests zeigten eine 30 signifikante Reduktion der apikalen Verzweigungspunkte in der toxinbehandelten Gruppe im Vergleich zur serumhaltigen Kontrolle. Der Kruskal-Wallis Test ergab einen p-Wert von 0,037, die Varianzanalyse von 0,036. Die serumfreie Kontrollgruppe nahm laut Tukey-Test eine indifferente Zwischenstellung ein und war somit von keiner der anderen beiden Gruppen signifikant verschieden. Auch hier traten keine Wechselwirkungen zwischen apikaler und basaler Seite auf, was durch eine weitere Varianzanalyse gezeigt werden konnte. Abbildung 12 Graphische Darstellung der Verzweigungspunkte der Apikaldendriten. Die Mittelwerte der drei Gruppen sind als Querbalken, ihre Standardabweichungen senkrecht dazu eingezeichnet. * p ≤ 0,05 4.3.3 Anzahl der Spines 4.3.3.1 Anzahl der Spines der Basaldendriten Bei der Analyse der Basaldendriten fiel der Unterschied zwischen den Gruppen bezogen auf die Anzahl der Spines nicht so markant aus. Trotzdem besaßen auch hier die serumhaltig inkubierten Pyramidenzellen die meisten Spines (350,67 ± 266,87). Die Gesamtzahl der Spines an den Basaldendriten in der serumfrei inkubierten Kontrollgruppe betrug 339,00 ± 271,67. Am geringsten mit Spines besetzt waren die Basaldendriten der toxinbehandelten CA1 Neurone (105,67 ± 91,85). Hier ergab die statistische Auswertung keine signifikanten 31 Unterschiede zwischen den verschiedenen Behandlungsgruppen. Der Kruskal-Wallis Test gab einen p-Wert von 0,12, die Varianzanalyse von 0,09 an. Abbildung 13: Die Anzahl der Spines der Basaldendriten graphisch dargestellt. Die Querbalken kennzeichnen die Mittelwerte der jeweiligen Behandlungsgruppe, die senkrechten Balken ihre Standardabweichungen. 4.3.3.2 Anzahl der Spines der Apikaldendriten Bei diesem Parameter fielen bei Betrachtung der Werte deutliche Unterschiede zwischen den drei Gruppen auf. Am dichtesten mit Spines besetzt waren die Apikaldendriten der serumhaltigen Kontrollgruppe (561,00 ± 358,91). Der Mittelwert für die Anzahl der Spines war um ca. 58% verringert (235,83 ± 127,29) für die serumfrei inkubierten Pyramidenzellen. Eine weitere Abnahme der Spines an den Apikalendriten war bei der toxinbehandelten Gruppe zu beobachten (109,50 ± 83,48). Die statistische Auswertung erbrachte signifikante Unterschiede bezüglich des Parameters Spinezahl an Apikaldendriten. Unter Toxineinfluss kam es zu einer signifikanten Abnahme im Vergleich zur serumhaltigen Kontrollgruppe mit einer Irrtumswahrscheinlichkeit p von 0,032 (Kruskal-Wallis Test) beziehungsweise 0,030 (Varianzanalyse). Laut Tukey-Test wich die serumfreie Kontrollgruppe nicht signifikant von einer der beiden anderen Gruppen ab. 32 Abbildung 14: Graphische Darstellung der Anzahl der Spines im Bereich der Apikaldendriten. Die Querbalken kennzeichnen die Mittelwerte der jeweiligen Behandlungsgruppe. Ihnen sind nach oben und unten die Werte ihrer Standardabweichung zugeordnet. * p ≤ 0,05 4.3.3.3 Gesamtzahl der Spines Die meisten postsynaptischen Dornen besaßen die Neurone der serumhaltigen Kontrollgruppe (911,67 ≤ 560,39). Wie auch bei den vorherigen Parametern war eine Verminderung der Spineanzahl über die serumfreie Kontrollgruppe (574,83 ≤ 378,81) bis hin zur toxinbehandelten Gruppe (215,17 ≤ 161,72) zu beobachten. Auch in Bezug auf die Gesamtzahl der Spines zeigte die toxinbehandelte Gruppe im Vergleich zur serumhaltigen Kontrollgruppe eine signifikante Abnahme. Der Kruskal-Wallis Test ergab einen p-Wert von 0,020, die Varianzanalyse von 0,018. Die serumfreie Kontrolle nahm laut Tukey-Test wiederum eine indifferente Zwischenstellung ein. 33 Abbildung 15: Grafik der Gesamtzahl der Spines aller ausgewerteter CA1 Pyramidenzellen. Mittelwerte und Standardabweichungen der Gruppen sind als Quer- bzw. vertikale Balken eingezeichnet. * p ≤ 0,05 4.3.4 Spinedichte 4.3.4.1 Spinedichte der Basaldendriten Die Neurone der serumhaltig inkubierten Kontrollgruppe waren mit 0,18 ± 0,08 Spines/µm am dichtesten Spine-besetzt. Die Besatzdichte nahm von der serumhaltigen zur serumfrei inkubierten Kontrollgruppe mit 0,17 ± 0,08 Spines/µm leicht, zur toxinbehandelten Gruppe dagegen stark ab (0,11 ± 0,08 Spines/µm). Die Mittelwerte der Behandlungsgruppen waren nicht signifikant voneinander verschieden. 34 Abbildung 16: Graphische Darstellung der Spinedichte der Basaldendriten jeder der 18 untersuchten Pyramidenzellen. Die Querbalken zeigen die Mittelwerte der jeweiligen Behandlungsgruppe an. Ihnen sind nach oben und unten die Werte ihrer Standardabweichung zugeordnet. 4.3.4.2 Spinedichte der Apikaldendriten In der serumhaltigen Kontrollgruppe hatten die Neurone eine mittlere Spinedichte von 0,19 ± 0,07 Spines/µm. Die serumfreie Kontrolle zeigte mit einem Mittelwert von 0,20 ± 0,08 Spines/µm eine leicht höhere Besatzdichte an. Mit 0,09 ± 0,05 Spines/µm waren die Neurone der C3 bot Toxin-behandelten Gruppe am wenigsten dicht mit Spines besetzt. Statistisch betrachtet war die Spinedichte der Apikaldendriten nicht signifikant unterschiedlich zwischen den einzelnen Behandlungsgruppen. Für eine Ausdünnung der Spines durch die Toxinbehandlung gab der Kruskal-Wallis Test eine Irrtumswahrscheinlichkeit von 0,051, die Varianzanalyse von 0,047 an. Obwohl der durch die Varianzanalyse ermittelte p-Wert das Signifikanzniveau knapp unterschritt, zeigte der post hoc durchgeführte Tukey-Test keine signifikanten Unterschiede zwischen den drei Gruppen an. Auch in Bezug auf die Spinedichte traten keine statistisch nachweisbaren Wechselwirkungen zwischen basaler und apikaler Seite auf. 35 Abbildung 17: Schaubild der apikalen Spine-Besatzdichte aller untersuchten Nervenzellen. Die Querbalken zeigen die Mittelwerte der jeweiligen Behandlungsgruppe, die senkrechten Balken die Werte ihrer Standardabweichung an. 4.3.4.3 Spinedichte der gesamten Zelle Nicht nur absolut hatten die Neurone der serumhaltig inkubierten Untersuchungsgruppe eine signifikant höhere Gesamtzahl an Spines als die Nervenzellen der toxinbehandelten Gruppe (vergleiche 3.3.3.3). Auch für den Parameter Spinedichte ergab die Analyse signifikante Unterschiede zwischen der Serumkontrolle mit 0,19 ± 0,04 Spines/µm und der Toxingruppe mit 0,10 ± 0,06 Spines/µm. Die Zellen der serumfreien Kontrolle (0,18 ± 0,06 Spines/µm) waren ähnlich dicht mit Spines besetzt wie die der serumhaltigen Kontrollgruppe. Der durchschnittliche Abstand der Spines zueinander nahm unter Toxineinfluss signifikant zu gegenüber der serumhaltigen Kontrolle. Der Kruskal-Wallis Test gab dafür eine Irrtumswahrscheinlichkeit von 0,049 an, die Varianzanalyse von 0,029. Laut Tukey-Test verhielt sich die serumfreie Kontrollgruppe ein weiteres Mal indifferent. 36 Abbildung 18: Graphik der Gesamt-Spinedichte der 18 analysierten Pyramidenzellen. Die Querbalken kennzeichnen die Mittelwerte der jeweiligen Behandlungsgruppe, die senkrechten Balken geben ihre Standardabweichung an. * p ≤ 0,05 4.4 Zusammenfassung der Ergebnisse Als wesentliche Ergebnisse konnten mehrere signifikante Unterschiede zwischen der toxinbehandelten Experimentalgruppe und der serumhaltigen Kontrollgruppe festgestellt werden. Die Behandlung mit dem clostridialen C3 Toxin führte zu einer signifikanten Reduktion der dendritischen Verzweigung im Bereich der Apikaldendriten. Außerdem konnte nach der Behandlung eine signifikant verminderte Anzahl der Spines sowohl an den Apikaldendriten als auch an den Dendriten insgesamt (basal + apikal) beobachtet werden. Auch im Hinblick auf die Spinedichte ergab die Toxinbehandlung im Vergleich zur SerumKontrollgruppe signifikant verminderte Werte bezogen auf das gesamte Neuron (basal + apikal). An den Basaldendriten war ebenfalls eine Verminderung der genannten Parameter zu verzeichnen. Hier bewegten sich die Werte allerdings nicht im Bereich der statistischen Signifikanz. In Bezug auf die Dendritenlänge führte Toxineinfluss ebenfalls zu einer leichten, jedoch nicht signifikanten Abnahme. Die serumfreie Kontrollgruppe nahm stets eine indifferente Zwischenstellung ein. 37 Tabelle 1 vermittelt einen Überblick über die wesentlichen Ergebnisse der quantitativen Analyse. Merkmal Gesamtlänge Submerkmal Statistik Basaldendrit nicht signifikant p = 0.14 nicht signifikant p = 0.07 Apikaldendrit Beschreibung Verzweigungspunkte Basaldendrit nicht signifikant p = 0.07 Apikaldendrit signifikant p = 0.04 Basaldendrit nicht signifikant p = 0.12 Apikaldendrit signifikant p = 0.03 C3 Toxingruppe ist – verglichen mit der serumhaltigen Kontrolle lediglich mit einem Fünftel der Spines besetzt (*). Im Vergleich zur serumfreien Kontrolle weisen die toxinbehandelten Zellen im Mittel immer noch weniger als die Hälfte der Spines auf (n. s.) Gesamt signifikant p = 0.02 C3 Toxin-Gruppe weist die geringste Spineanzahl auf, statistisch signifikant gegenüber der serumhaltigen Kontrollgruppe Basaldendrit nicht signifikant p = 0.17 Apikaldendrit nicht signifikant p = 0.05 Gesamt signifikant p = 0.049 C3 Toxingruppe signifikant schwächer verzweigt als serumhaltige Kontrollgruppe Anzahl der Spines Spinedichte Toxinbehandlung bewirkt signifikante Reduktion der Spinedichte im Vergleich zur Serum-Kontrolle. Auch gegenüber der serumfreien Kontrolle Tendenz zu weniger dicht spinebesetzten Dendriten Tabelle 1: Überblick der Ergebnisse der quantitativen Analyse, getrennt nach untersuchten Parametern sowie Dendritentyp 38 5 Diskussion 5.1 Einführung in die Problemstellung Dendriten stellen ein wichtiges strukturelles Element von Nervenzellen dar. Sie fungieren als Zielstrukturen von Nervenfasern, die von anderen benachbarten beziehungsweise weiter entfernten Neuronen stammen. So können Nervenfasern einerseits am Dendritenschaft funktionelle Kontakte ausbilden. Diese so genannten Schaftsynapsen werden als symmetrische inhibitorische Kontakte angesehen. Neben den Schaftsynapsen gibt es Kontakte an speziellen Ausstülpungen des Dendritenbaums. Diese Ausstülpungen werden als Protrusionen oder Spines bezeichnet. Zahlreiche Literaturbefunde belegen, dass diese Spines postsynaptische Strukturen für exzitatorische Nervenfasern darstellen (MATUS 2005, CALABRESE et al. 2006). Spines sind sehr dynamische Strukturen, die gebildet und eingezogen werden können und unter dem Einfluss synaptischer Aktivität stabilisiert werden. Es gibt Literaturhinweise, dass es nach Blockade synaptischer Aktivität zu Veränderungen der Form dieser Spines kommt (DRAKEW et al. 1998). Andererseits führt gesteigerte neuronale Aktivität wie Langzeitpotenzierung ebenfalls zu Spinebildung an Dendriten (ENGERT u. BONHOEFFER 1999, BONHOEFFER u. YUSTE 2002). Das heißt, Modulation der Aktivität hat Einfluss auf den Spinebesatz der Dendriten und damit die Verschaltung zwischen den Neuronen. Stabilisierung und Verdrahtung von Nervenzellen untereinander sind wichtige Voraussetzungen für die normale Funktion des Zentralnervensystems. In der vorliegenden Arbeit wollten wir nun der Frage nachgehen, inwieweit Neurotoxine in einer subletalen Dosis von Nervenzellen toleriert werden beziehungsweise zu strukturellen Veränderungen führen. Voraussetzung für eine detaillierte Analyse der Wirkung von clostridialem C3 Toxin war die Identifikation von Neuronen, die standardisierte Exposition gegenüber dem C3 Toxin und die Zugänglichkeit behandelter Neurone für eine quantitative Analyse. Die Besonderheit dieser Arbeit besteht darin, dass die Untersuchungen an identifizierten Neuronen durchgeführt worden sind. Bisherige Literaturbefunde zur Wirkung von C3 bot beziehen sich auf die Arbeit an dissoziierten Neuronen und die Wirkung auf die Axone der kultivierten Zellen (OLENIK et al. 1997, AHNERT-HILGER et al. 2004). Dies hat den Nachteil, dass keine zellspezifischen Effekte beobachtet werden können. 39 5.2 Bewertung des experimentellen Ansatzes Die hier vorgestellten Untersuchungen wurden an einem durch zahlreiche Literaturbefunde gut etablierten Forschungsmodell der organotypischen Hirnschnittkulturen durchgeführt (HEIMRICH u. FROTSCHER 1991, HEIMRICH u. FROTSCHER 1993a, HEIMRICH u. FROTSCHER 1993b, ZAFIROV et al. 1994). In Untersuchungen von ZIMMER u. GÄHWILER (1984) und FROTSCHER et al. (2000) wurden in den letzten Jahren zahlreiche quantitative Analysen zu Spine- und Dendritenentwicklung durchgeführt und im normalen Tier verglichen. In diesen Untersuchungen konnte gezeigt werden, wie in vivo die Spineentwicklung stattfindet und wie die CA1 Pyramidenzellen ihre morphologischen Merkmale beibehalten und als solche aufgrund des Dendritenbaums und Spinebesatzes identifiziert werden können. Ausgehend von diesen Befunden gab es zusätzlich zahlreiche Untersuchungen, in denen die in-vitro-Bedingungen experimentell modifiziert wurden, was dann zu Veränderungen der neuronalen Morphologie beziehungsweise Konnektivität führte (BRINKS et al. 2004, MAYER et al. 2005). Die Möglichkeit, Hirnschnittkulturen unter standardisierten Bedingungen über einen längeren Zeitraum kultivieren und Zellen aufgrund ihrer Morphologie und Lokalisation im Hirngewebe identifizieren zu können, war eine wesentliche Grundlage dafür, strukturelle Veränderungen nach C3 Toxin-Behandlung sowohl qualitativ als auch quantitativ zu erfassen. Um den Einfluss der verschiedenen Behandlungsarten auf die Dendritendifferenzierung und Spinebildung der hippocampalen CA1 Pyramidenzellen zu untersuchen, wurden drei Untersuchungsgruppen gebildet. Die erste Gruppe umfasste Hippocampuskulturen, die in serumhaltigem Kulturmedium inkubiert worden waren. Die zweite Experimentalgruppe bestand aus Kulturen, deren Kulturmedium einige Stunden vor intrazellulärer Applikation des Tracers Biocytin gegen serumfreies Medium ausgetauscht worden war, um eine mögliche Beeinträchtigung des Toxins durch im Serum enthaltene Proteasen auszuschließen. Toxinbehandelte Kulturen, die kurz vor der Toxininjektion ebenfalls serumfrei inkubiert worden waren, bildeten die dritte Gruppe. Bei einigen intrazellulären Füllungen mit Biocytin kommt es zum Auslaufen von Biocytin aus der penetrierten Zelle. Dies führt nach DAB-Entwicklung zu einem hohen Hintergrund durch unspezifischen Niederschlag um die mit Biocytin gefüllte Zelle. Damit sind sowohl eine Rekonstruktion als auch die quantitative Erfassung der Spines nicht zuverlässig möglich. Die betroffenen Zellen sind trotz Füllung nicht auswertbar und wurden nicht in die Analyse mit 40 einbezogen. Aus allen drei Gruppen konnten biocytingefüllte CA1 Pyramidenzellen für Rekonstruktion und quantitative Analyse der dendritischen Parameter herangezogen werden. Dies bestätigt, dass unter den gewählten in vitro-Bedingungen hippocampale Neurone ihre morphologische Charakteristik beibehalten, wie dies in früheren Untersuchungen an Schnittkulturen aus dieser Arbeitsgruppe schon gezeigt werden konnte (HEIMRICH u. FROTSCHER 1991, FROTSCHER et al. 1995). Die jeweiligen Gruppen zeigten entsprechend ihrer Behandlung unterschiedliche Effekte auf Dendritenorganisation und Spinebesatz. Dies deutet auf einen Effekt nach Serumentzug beziehungsweise C3 Toxin-Exposition hin. Hierbei kommen mehrere beeinflussende Faktoren in Betracht: Der Serumentzug könnte die intrazelluläre Füllbarkeit der Zelle mit Biocytin beeinträchtigen. Dieser Faktor ist insofern zu vernachlässigen, da wir die Dendriten sowohl in den serumfreien sowie serumfreien und zusätzlich toxinbehandelten Gruppen bis in ihre distalen Abschnitte mikroskopisch verfolgen konnten. Ganz generell ist die intrazelluläre Biocytinfüllung eine viel genutzte und allgemein anerkannte Methode zur Darstellung von Neuronen (IRINTCHEV et al. 2005). Zum anderen könnte Serumentzug einen Mangel an Nährstoffen nach sich ziehen, was letztendlich morphologisch sichtbare Veränderungen hervorrufen könnte. Da jedoch keine signifikanten Unterschiede bezüglich Dendritenlänge, Anzahl der Verzweigungspunkte, Anzahl der Spines und Spinedichte zu den Pyramidenzellen aus unbehandelten Kontrollkulturen messbar waren, sind eventuelle Effekte zu vernachlässigen. Die Zugabe von C3 Toxin führt zu signifikanten morphologischen Veränderungen beim Vergleich von serumhaltig inkubierter Kontrollgruppe und C3 Toxin Behandlungsgruppe. Dies deutet darauf hin, dass die Applikation von C3 bot maßgeblich Dendritenmorphologie und Spinebesatz beeinflußt. 5.3 Evaluierung der Ergebnisse Im folgenden Abschnitt werden noch einmal die wichtigsten Befunde aus den Untersuchungen der einzelnen Behandlungsgruppen Serumhaltige Kontrollgruppe, Serumfreie Kontrollgruppe sowie C3 bot Toxingruppe zusammengefasst und diskutiert. Unabhängig von der jeweiligen Behandlung haben die CA1 Pyramidenzellen aller drei Gruppen überlebt und wir konnten in allen Kulturen die Neurone durch intrazelluläre Füllung mit Biocytin als CA1 Pyramidenzellen identifizieren. Tendenziell war festzustellen, dass alle erfassten Werte der einzelnen Parameter in der serumfreien Kontrollgruppe verglichen mit der serumhaltigen Kontrollgruppe abnahmen. Die 41 geringen Veränderungen bezüglich Dendritenlänge, Anzahl der Verzweigungspunkte, Anzahl der Spines und Spinedichte zwischen den beiden serumhaltig bzw. serumfrei inkubierten Kontrollgruppen könnten Effekte nach geänderter Nährstoffversorgung sein. Im Vergleich zwischen der serumhaltigen Kontrollgruppe und der mit C3 bot Toxin behandelten Gruppe kam es gegenüber dem Vergleich zwischen den beiden Kontrollgruppen zu einer noch stärkeren Verminderung von Dendritenlänge, Anzahl der Verzweigungspunkte, Anzahl der Spines sowie Spinedichte. Über alle analysierten morphologischen Parameter traten nur für einzelne signifikante Unterschiede auf. Keine signifikanten Abweichungen waren hinsichtlich Länge der Apikaldendriten, Länge der Basaldendriten, Verzweigungspunkte der Basaldendriten, Spineanzahl der Basaldendriten, Spinedichte der Apikal- und Spinedichte der Basaldendriten zu beobachten. Demgegenüber waren die Anzahl der Verzweigungspunkte und die Anzahl der Spines an den Apikaldendriten signifikant verringert. Die ebenfalls signifikante Abnahme der Gesamtzahl der Spines und der Gesamtspinedichte bei der C3 bot Toxin-Behandlungsgruppe im Vergleich zur serumhaltigen Kontrollgruppe beruht vor allem auf den stark ausgeprägten strukturellen Unterschieden an den Apikaldendriten. Dies lässt auf eine selektive Wirkung des C3 bot Toxins auf das neuronale Kompartiment, den apikalen Dendritenbaum mit seinen postsynaptischen Spines, schließen. Diese selektive Wirkung des Toxins deutet auf rezeptorvermittelte Effekte hin und könnte durch unterschiedliche Rho-Rezeptoren bzw. Unterschiede in deren Verteilung beruhen. Aus der Literatur gibt es Hinweise, dass das Toxin nicht ubiquitär, sondern an bestimmten Zellkompartimenten wirkt. AHNERT-HILGER et al. (2004) untersuchten den Einfluss von RhoA auf die dendritische beziehungsweise axonale Differenzierung in dissoziierten Zellen. Sie konnten zeigen, dass Aktivierung von RhoA zu einer verstärkten axonalen und gleichzeitig verminderten dendritischen Entwicklung führt. Umgekehrt hemmt Inaktivierung des RhoA durch C3 Isoformen axonales Wachstum und fördert Dendritenwachstum. Diese Befunde stehen in Widerspruch zu den vorliegenden Untersuchungsergebnissen. Die Inaktivierung von RhoA durch das C3 Toxin hat zu einer zwar nicht signifikanten jedoch tendenziellen Verkürzung der Dendriten geführt. Eine mögliche Erklärung hierfür könnten die angewandten unterschiedlichen in vitro-Systeme sein. In dissoziierten Kulturen werden unterschiedliche Neuronentypen aus embryonalem Hirngewebe verwendet. Diese Neurone zeigen eine sehr starke morphologische Variabilität, die eine Identifizierung des Neuronentyps nach morphologischen Kriterien weitgehend unmöglich macht. Es könnte durchaus möglich sein, dass unterschiedliche Neuronenpopulationen sehr unterschiedlich auf C3 bot Toxin-Behandlung reagieren. Demgegenüber sind in dieser Untersuchung identifizierte CA1 Pyramidenzellen in 42 Hippocampusschnittkulturen analysiert und verglichen worden. Mit in Betracht gezogen werden muss auch eine Wirkung des Toxins auf die in den Hippocampusschnittkulturen vorhandene intrahippocampale axonale Projektion der Schafferkollateralen, welche die CA3 Pyramidenzellen mit den CA1 Neuronen verbinden und an den Dendriten der CA1 Pyramidenzellen endigen. Die Unterschiede, die wir an den Neuronen beobachtet haben, könnten also auch Sekundäreffekte darstellen, die in Zusammenhang mit einer Toxinwirkung stehen. In neueren Untersuchungen konnten solche Effekte von C3 bot auf Axonwachstum und axonale Kollateralisation nachgewiesen werden (AHNERT-HILGER et al. 2004, HÖLTJE et al. 2008). Zusammenfassend konnte in dieser Studie gezeigt werden, dass die Behandlung von identifizierten hippocampalen CA1 Pyramidenzellen mit C3 bot Verzweigungsmuster und Spinezahl selektiv am Apikaldendrit dieses Zelltyps verändert, jedoch keinerlei messbare Wirkung an den Basaldendriten entfaltet. Dies deutet auf einen rezeptorvermittelten Effekt des Toxins hin. Weiterführende Untersuchungen mit einem biologisch inaktiven C3 Toxin von Clostridium botulinum sollen helfen zu klären, inwieweit das biologisch aktive Toxin in direktem Zusammenhang mit diesen Veränderungen steht. Außerdem sollen weitere in vitro Untersuchungen mit C3 bot Behandlung dazu dienen, den Signalweg, der zu diesen morphologischen Veränderungen führt, näher zu charakterisieren. 43 6 Zusammenfassung Thorsten Hintze Einfluss des clostridialen C3 Toxins auf die Dendritenmorphologie und Spinebildung von CA1 Pyramidenzellen in Hippocampus-Schnittkulturen der Maus - eine quantitative lichtmikroskopische Untersuchung Veterinär-Anatomisches Institut Veterinärmedizinische Fakultät, Universität Leipzig und Anatomisches Institut Medizinische Fakultät, Albert-Ludwigs-Universität Freiburg im Breisgau Eingereicht im Februar 2010 47 Seiten, 18 Abbildungen, 1 Tabelle, 83 Literaturangaben, Anhang Schlüsselwörter: Hippocampus, CA1 Pyramidenzellen, Rho Proteine, C3 Toxin, Hirnschnittkulturen Lokale Pyramidenzellen sind die Hauptneurone des Hippocampus und können durch ihre Position und die Morphologie ihrer Dendriten als CA1 und CA3 Pyramidenzellen identifiziert werden. Die Dendriten der exzitatorischen Pyramidenzellen sind mit postsynaptischen Vorwölbungen, den so genannten Spines, bedeckt, welche in einem spezifischen Verteilungsmuster angeordnet sind. Neurotoxine wie das C3 Toxin von Clostridium botulinum sind funktionelle Substanzen, die die neuronale Morphologie verändern und die neuronale Funktion beeinflussen können. In dieser Studie wurden die morphologischen Veränderungen von intrazellulär mit Biocytin gefüllten CA1 Pyramidenzellen qualitativ und quantitativ analysiert. Die hippocampalen Schnittkulturen, in denen sich bekanntermaßen Pyramidenzellen ähnlich entwickeln wie in vivo, wurden dazu herangezogen, die Effekte der C3bot Toxin-Applikation auf die Verzweigung der Dendriten sowie Anzahl und Dichte der dendritischen Spines zu untersuchen. Drei Gruppen von Zellen wurden verglichen: Erstens Neurone, die in serumhaltigem Medium inkubiert worden waren, zweitens Nervenzellen, die 44 in einem Medium ohne Serum inkubiert worden waren und drittens Zellen, die unter Serumentzug dem C3bot Toxin ausgesetzt worden waren. Die Inkubation dauerte 14 Tage, während die Dauer der Toxinexposition zwischen vier und sechs Stunden betrug. Mit Hilfe eines Computers wurden zweidimensionale Nachbildungen der biocytin-markierten CA1 Pyramidenzellen erstellt, und die Gesamtlänge der Dendriten, die Anzahl der dendritischen Verzweigungspunkte und die Gesamtzahl und Dichte der dendritischen Spines gemessen und statistisch ausgewertet. Signifikante Unterschiede wurden zwischen der mit C3 Toxin behandelten Gruppe und der serumhaltig inkubierten Kontrollgruppe beobachtet. Diese signifikanten morphologischen Veränderungen traten selektiv an den Apikaldendriten der toxinbehandelten CA1 Pyramidenzellen auf. Aus der Behandlung resultierte eine Reduktion der Anzahl apikaler Verzweigungspunkte, der Anzahl der apikalen Spines, der Gesamtzahl (basal und apikal addiert) der Spines sowie der Gesamtspinedichte. Im Gegensatz dazu ergaben sich keine signifikanten Unterschiede zwischen der toxinbehandelten Gruppe und der ohne Serum inkubierten Kontrollgruppe, obwohl der Serumentzug im Vergleich zur serumhaltig inkubierten Kontrollgruppe die Entwicklung der Zellen beeinflusste. Auf Grundlage der beobachteten Veränderungen können wir schließen, dass die Behandlung mit C3 bot einen starken Einfluss selektiv auf die Morphologie der Apikaldendriten ausübt. Der Mechanismus, der dieser selektiven Empfindlichkeit der Apikaldendriten gegenüber dem C3 bot Toxin zugrunde liegt, wird Gegenstand weiterer Untersuchungen sein. 45 7 Summary Thorsten Hintze Influence of Clostridium botulinum C3 toxin on dendrite morphology and spine formation of CA1 pyramidal cells in hippocampal slice cultures of the mouse – a quantitative light microscopic investigation Institute of Anatomy, Histology and Embryology Faculty of Veterinary Medicine, University of Leipzig and Institute of Anatomy Faculty of Medicine, University of Freiburg Submitted in February 2010 47 pages, 18 figures, 1 table, 83 references, appendix Keywords: Hippocampus, CA1 Pyramidal Cells, Rho Proteins, C3 Toxin, brain slice cultures Local Pyramidal cells are the principal neurons of the hippocampus and can be identified by their position and their dendritic morphology as CA1 and CA3 pyramidal cells. The dendrites of excitatory pyramidals are covered with postsynaptic protrusions the so-called spines, which are arranged in a specific distribution pattern. Neurotoxins like the C3 toxin of clostridium botulinum are functional substances which can alter the neuronal morphology and influence neuronal function. In this study the morphologic changes of intracellular biocytin-filled CA1 pyramidal cells were qualitatively and quantitatively analysed. The hippocampal slice cultures, wherein the pyramidal cells are known to develop similarly as in vivo, were used to study the effects of C3 toxin application on dendritic branching and the number and density of dendritic spines. Three groups of cells were compared, namely cells incubated in medium containing serum, cells incubated in medium without serum and cells exposed to C3bot toxin under serum deprivation. The duration of incubation accounted for 14 days, while the exposition of the toxin was between 4 and 6 hours. Computer-assisted two dimensional reconstructions of biocytin-labelled CA1 pyramidal neurons were performed and the total 46 length of dendrites, number of dendritic nodes, total number and density of dendritic spines were measured and statistically evaluated. Significant differences were observed between the group treated with the C3 toxin and the control group incubated with serum containing medium. These significant morphological changes appeared selectively on apical dendrites of treated CA1 pyramidal neurons. The treatment resulted in a decrease of the number of the apical nodes, the number of the apical spines, the total (basilar and apical) number of spines and the total spine density. In contrast there are no significant differences between the toxin treated group and the control group without serum, although the deprivation of serum affected the development of the cells in comparison to the control cells incubated with serum. Based upon the observed changes we can conclude that C3bot treatment exerts a strong impact selectively on apical dendrite morphology. The mechanism forming the basis of this selective susceptibility of apical dendrites to C3bot requires further investigations. 47 8 Literaturverzeichnis Adams JC. Heavy metal intensification of DAB-based HRP reaction product. J Histochem Cytochem. 1981; 29: 775. Ahnert-Hilger G, Höltje M, Grosse G, Pickert G, Mucke C, Nixdorf-Bergweiler B, Boquet P, Hofmann F, Just I. Differential effects of Rho GTPases on axonal and dendritic development in hippocampal neurones. J Neurochem. 2004; 90: 9-18. Aktories K, Frevert J. ADP-ribosylation of a 21-24 kDa eukaryotic protein(s) by C3, a novel botulinum ADP-ribosyltransferase, is regulated by guanine nucleotide. Biochem J. 1987; 247: 363-8. Bagrodia S, Derijard B, Davis RJ, Cerione RA. Cdc42 and PAK-mediated signaling leads to Jun kinase and p38 mitogen-activated protein kinase activation. J Biol Chem. 1995; 270: 27995-8. Bernards A. GAPs galore! A survey of putative Ras superfamily GTPase activating proteins in man and Drosophila. Biochim Biophys Acta. 2003; 1603: 47-82. Bernards A, Settleman J. GAP control: regulating the regulators of small GTPases. Trends Cell Biol. 2004; 14: 377-85. Blanpied TA, Ehlers MD. Microanatomy of dendritic spines: emerging principles of synaptic pathology in psychiatric and neurological disease. Biol Psychiatry. 2004; 55: 1121-7. Bonhoeffer T, Yuste R. Spine motility. Phenomenology, mechanisms, and function. Neuron. 2002; 35: 1019-27. Brinks H, Conrad S, Vogt J, Oldekamp J, Sierra A, Deitinghoff L, Bechmann I, AlvarezBolado G, Heimrich B, Monnier PP, Müller BK, Skutella T. The repulsive guidance molecule RGMa is involved in the formation of afferent connections in the dentate gyrus. J Neurosci. 2004; 24: 3862-9. 48 Brouns MR, Matheson SF, Settleman J. p190 RhoGAP is the principal Src substrate in brain and regulates axon outgrowth, guidance and fasciculation. Nat Cell Biol. 2001; 3: 361-7. Brown JL, Stowers L, Baer M, Trejo J, Coughlin S, Chant J. Human Ste20 homologue hPAK1 links GTPases to the JNK MAP kinase pathway. Curr Biol. 1996; 6: 598-605. Buchsbaum RJ, Connolly BA, Feig LA. Interaction of Rac exchange factors Tiam1 and RasGRF1 with a scaffold for the p38 mitogen-activated protein kinase cascade. Mol Cell Biol. 2002; 22: 4073-85. Buchsbaum RJ, Connolly BA, Feig LA. Regulation of p70 S6 kinase by complex formation between the Rac guanine nucleotide exchange factor (Rac-GEF) Tiam1 and the scaffold spinophilin. J Biol Chem. 2003; 278: 18833-41. Burridge K, Wennerberg K. Rho and Rac take center stage. Cell. 2004; 116: 167-79. Calabrese B, Wilson MS, Halpain S. Development and regulation of dendritic spine synapses. Physiology (Bethesda). 2006; 21: 38-47. Chardin P, Boquet P, Madaule P, Popoff MR, Rubin EJ, Gill DM. The mammalian G protein rhoC is ADP-ribosylated by Clostridium botulinum exoenzyme C3 and affects actin microfilaments in Vero cells. Embo J. 1989; 8: 1087-92. Cline HT. Dendritic arbor development and synaptogenesis. Curr Opin Neurobiol. 2001; 11: 118-26. Coso OA, Chiariello M, Yu JC, Teramoto H, Crespo P, Xu N, Miki T, Gutkind JS. The small GTP-binding proteins Rac1 and Cdc42 regulate the activity of the JNK/SAPK signaling pathway. Cell. 1995; 81: 1137-46. Da Silva JS, Dotti CG. Breaking the neuronal sphere: regulation of the actin cytoskeleton in neuritogenesis. Nat Rev Neurosci. 2002; 3: 694-704. 49 Da Silva JS, Medina M, Zuliani C, Di Nardo A, Witke W, Dotti CG. RhoA/ROCK regulation of neuritogenesis via profilin IIa-mediated control of actin stability. J Cell Biol. 2003; 162: 1267-79. Dölen G, Bear MF. Courting a cure for fragile X. Neuron. 2005; 45: 642-4. Drakew A, Frotscher M, Deller T, Ogawa M, Heimrich B. Developmental distribution of a reeler gene-related antigen in the rat hippocampal formation visualized by CR-50 immunocytochemistry. Neuroscience. 1998; 82: 1079-86. Engert F, Bonhoeffer T. Dendritic spine changes associated with hippocampal long-term synaptic plasticity. Nature. 1999; 399: 66-70. Fiala JC, Spacek J, Harris KM. Dendritic spine pathology: cause or consequence of neurological disorders? Brain Res Brain Res Rev. 2002; 39: 29-54. Fournier AE, Takizawa BT, Strittmatter SM. Rho kinase inhibition enhances axonal regeneration in the injured CNS. J Neurosci. 2003; 23: 1416-23. Frotscher M, Heimrich B, Schwegler H. Plasticity of identified neurons in slice cultures of hippocampus: a combined Golgi/electron microscopic and immunocytochemical study. Prog Brain Res. 1990; 83: 323-39. Frotscher M, Zafirov S, Heimrich B. Development of identified neuronal types and of specific synaptic connections in slice cultures of rat hippocampus. Prog Neurobiol. 1995; 45: viixxviii. Frotscher M, Drakew A, Heimrich B. Role of afferent innervation and neuronal activity in dendritic development and spine maturation of fascia dentata granule cells. Cereb Cortex. 2000; 10: 946-51. Fujita A, Hattori Y, Takeuchi T, Kamata Y, Hata F. NGF induces neurite outgrowth via a decrease in phosphorylation of myosin light chain in PC12 cells. Neuroreport. 2001; 12: 3599-602. 50 Glantz LA, Lewis DA. Dendritic spine density in schizophrenia and depression. Arch Gen Psychiatry. 2001; 58: 203. Goda Y, Davis GW. Mechanisms of synapse assembly and disassembly. Neuron. 2003; 40: 243-64. Govek EE, Newey SE, Van Aelst L. The role of the Rho GTPases in neuronal development. Genes Dev. 2005; 19: 1-49. Hall A. Rho GTPases and the actin cytoskeleton. Science. 1998; 279: 509-14. Harris KM. Structure, development, and plasticity of dendritic spines. Curr Opin Neurobiol. 1999; 9: 343-8. Hartung J, Elpelt B. Multivariate Statistik. Lehr- und Handbuch der angewandten Statistik. 3. Aufl. München/Wien; 1989. Heimrich B, Frotscher M. Differentiation of dentate granule cells in slice cultures of rat hippocampus: a Golgi/electron microscopic study. Brain Res. 1991; 538: 263-8. Heimrich B, Frotscher M. Formation of the septohippocampal projection in vitro: an electron microscopic immunocytochemical study of cholinergic synapses. Neuroscience. 1993; 52: 815-27. Heimrich B, Frotscher M. Slice cultures as a model to study entorhinal-hippocampal interaction. Hippocampus. 1993; 3 Spec No: 11-7. Hering H, Sheng M. Dendritic spines: structure, dynamics and regulation. Nat Rev Neurosci. 2001; 2: 880-8. Höltje M, Hofmann F, Lux R, Veh RW, Just I, Ahnert-Hilger G. Glutamate uptake and release by astrocytes are enhanced by Clostridium botulinum C3 protein. J Biol Chem. 2008; 283: 9289-99. 51 Hoffman GR, Nassar N, Cerione RA. Structure of the Rho family GTP-binding protein Cdc42 in complex with the multifunctional regulator RhoGDI. Cell. 2000; 100: 345-56. Hsu SM, Raine L, Fanger H. Use of avidin-biotin-peroxidase complex (ABC) in immunoperoxidase techniques: a comparison between ABC and unlabeled antibody (PAP) procedures. J Histochem Cytochem. 1981; 29: 577-80. Irintchev A, Rollenhagen A, Troncoso E, Kiss JZ, Schachner M. Structural and functional aberrations in the cerebral cortex of tenascin-C deficient mice. Cereb Cortex. 2005; 15: 95062. Isokawa M. Remodeling dendritic spines of dentate granule cells in temporal lobe epilepsy patients and the rat pilocarpine model. Epilepsia. 2000; 41 Suppl 6: S14-7. Jan YN, Jan LY. Dendrites. Genes Dev. 2001; 15: 2627-41. Jiang M, Lee CL, Smith KL, Swann JW. Spine loss and other persistent alterations of hippocampal pyramidal cell dendrites in a model of early-onset epilepsy. J Neurosci. 1998; 18: 8356-68. Jin Z, Strittmatter SM. Rac1 mediates collapsin-1-induced growth cone collapse. J Neurosci. 1997; 17: 6256-63.6 Kozma R, Sarner S, Ahmed S, Lim L. Rho family GTPases and neuronal growth cone remodelling: relationship between increased complexity induced by Cdc42Hs, Rac1, and acetylcholine and collapse induced by RhoA and lysophosphatidic acid. Mol Cell Biol. 1997; 17: 1201-11. Kranenburg O, Poland M, Gebbink M, Oomen L, Moolenaar WH. Dissociation of LPAinduced cytoskeletal contraction from stress fiber formation by differential localization of RhoA. J Cell Sci. 1997; 110 (Pt 19): 2417-27. Leemhuis J, Henle F, Meyer DK. VIP induces the elongation of dendrites and axons in cultured hippocampal neurons: role of microtubules. Peptides. 2007; 28: 1700-5. 52 Linton LR, Harder LD. Biology 315 - Quantitative Biology Lecture Notes. University of Calgary, Calgary; 2007. Lisman J. Actin's actions in LTP-induced synapse growth. Neuron. 2003; 38: 361-2. Luo L, Liao YJ, Jan LY, Jan YN. Distinct morphogenetic functions of similar small GTPases: Drosophila Drac1 is involved in axonal outgrowth and myoblast fusion. Genes Dev. 1994; 8: 1787-802. Mackay DJ, Hall A. Rho GTPases. J Biol Chem. 1998; 273: 20685-8. Matus A. Growth of dendritic spines: a continuing story. Curr Opin Neurobiol. 2005; 15: 6772. Mayer D, Fischer H, Schneider U, Heimrich B, Schwemmle M. Borna disease virus replication in organotypic hippocampal slice cultures from rats results in selective damage of dentate granule cells. J Virol. 2005; 79: 11716-23. Minden A, Lin A, Claret FX, Abo A, Karin M. Selective activation of the JNK signaling cascade and c-Jun transcriptional activity by the small GTPases Rac and Cdc42Hs. Cell. 1995; 81: 1147-57. Montgomery DC. Design and Analysis of Experiments. 5. Aufl. Danvers; 2001. Nakayama AY, Harms MB, Luo L. Small GTPases Rac and Rho in the maintenance of dendritic spines and branches in hippocampal pyramidal neurons. J Neurosci. 2000; 20: 532938. Newey SE, Velamoor V, Govek EE, Van Aelst L. Rho GTPases, dendritic structure, and mental retardation. J Neurobiol. 2005; 64: 58-74. Nimchinsky EA, Sabatini BL, Svoboda K. Structure and function of dendritic spines. Annu Rev Physiol. 2002; 64: 313-53. 53 Norrholm SD, Bibb JA, Nestler EJ, Ouimet CC, Taylor JR, Greengard P. Cocaine-induced proliferation of dendritic spines in nucleus accumbens is dependent on the activity of cyclindependent kinase-5. Neuroscience. 2003; 116: 19-22. Olenik C, Barth H, Just I, Aktories K, Meyer DK. Gene expression of the small GTP-binding proteins RhoA, RhoB, Rac1, and Cdc42 in adult rat brain. Brain Res Mol Brain Res. 1997; 52: 263-9. Pilpel Y, Segal M. Activation of PKC induces rapid morphological plasticity in dendrites of hippocampal neurons via Rac and Rho-dependent mechanisms. Eur J Neurosci. 2004; 19: 3151-64. Portera-Cailliau C, Pan DT, Yuste R. Activity-regulated dynamic behavior of early dendritic protrusions: evidence for different types of dendritic filopodia. J Neurosci. 2003; 23: 7129-42. Robinson TE, Kolb B. Alterations in the morphology of dendrites and dendritic spines in the nucleus accumbens and prefrontal cortex following repeated treatment with amphetamine or cocaine. Eur J Neurosci. 1999; 11: 1598-604. Ruchhoeft ML, Ohnuma S, Mcneill L, Holt CE, Harris WA. The neuronal architecture of Xenopus retinal ganglion cells is sculpted by rho-family GTPases in vivo. J Neurosci. 1999; 19: 8454-63. Sarner S, Kozma R, Ahmed S, Lim L. Phosphatidylinositol 3-kinase, Cdc42, and Rac1 act downstream of Ras in integrin-dependent neurite outgrowth in N1E-115 neuroblastoma cells. Mol Cell Biol. 2000; 20: 158-72. Savaskan NE, Alvarez-Bolado G, Glumm R, Nitsch R, Skutella T, Heimrich B. Impaired postnatal development of hippocampal neurons and axon projections in the Emx2-/- mutants. J Neurochem. 2002; 83: 1196-207. Schmidt A, Hall A. Guanine nucleotide exchange factors for Rho GTPases: turning on the switch. Genes Dev. 2002; 16: 1587-609. 54 Schwamborn JC, Püschel AW. The sequential activity of the GTPases Rap1B and Cdc42 determines neuronal polarity. Nat Neurosci. 2004; 7: 923-9. Sebök A, Nusser N, Debreceni B, Guo Z, Santos MF, Szeberenyi J, Tigyi G. Different roles for RhoA during neurite initiation, elongation, and regeneration in PC12 cells. J Neurochem. 1999; 73: 949-60. Selkoe DJ. Alzheimer's disease is a synaptic failure. Science. 2002; 298: 789-91. Swann JW, Al-Noori S, Jiang M, Lee CL. Spine loss and other dendritic abnormalities in epilepsy. Hippocampus. 2000; 10: 617-25. Tashiro A, Minden A, Yuste R. Regulation of dendritic spine morphology by the rho family of small GTPases: antagonistic roles of Rac and Rho. Cereb Cortex. 2000; 10: 927-38. Tigyi G, Fischer DJ, Sebök A, Yang C, Dyer DL, Miledi R. Lysophosphatidic acid-induced neurite retraction in PC12 cells: control by phosphoinositide-Ca2+ signaling and Rho. J Neurochem. 1996; 66: 537-48. Van Aelst L, D'Souza-Schorey C. Rho GTPases and signaling networks. Genes Dev. 1997; 11: 2295-322. Van Aelst L, Cline HT. Rho GTPases and activity-dependent dendrite development. Curr Opin Neurobiol. 2004; 14: 297-304. Zafirov S, Heimrich B, Frotscher M. Dendritic development of dentate granule cells in the absence of their specific extrinsic afferents. J Comp Neurol. 1994; 345: 472-80. Zalcman G, Dorseuil O, Garcia-Ranea JA, Gacon G, Camonis J. RhoGAPs and RhoGDIs, (His)stories of two families. Prog Mol Subcell Biol. 1999; 22: 85-113. Zhang S, Han J, Sells MA, Chernoff J, Knaus UG, Ulevitch RJ, Bokoch GM. Rho family GTPases regulate p38 mitogen-activated protein kinase through the downstream mediator Pak1. J Biol Chem. 1995; 270: 23934-6. 55 Zimmer J, Gähwiler BH. Cellular and connective organization of slice cultures of the rat hippocampus and fascia dentata. J Comp Neurol. 1984; 228: 432-46. Ziv NE, Smith SJ. Evidence for a role of dendritic filopodia in synaptogenesis and spine formation. Neuron. 1996; 17: 91-102. 56 Anhang Hier sind die Rohdaten aller in die Untersuchung aufgenommener Parameter der Nervenzellen in jeweils aufsteigender Reihenfolge aufgeführt. Mittelwert und Standardabweichung der einzelnen Gruppen sind außerdem abzulesen. Mittelwert Standardabweichung Serumhaltige Kontrolle 926,6 µm 1151,8 µm 1235,8 µm 1711,1 µm 2275,8 µm 3729,2 µm 1838,38 µm 1043,89 µm Serumfreie Kontrolle 633,3 µm 720,3 µm 1145,1 µm 2338,9 µm 2414 µm 2657,7 µm 1651,55 µm 919,42 µm C3 Toxin 680,7 µm 852,4 µm 887,3 µm 903,2 µm 1101,3 µm 1697 µm 1020,32 µm 357,61 µm Tabelle A1: Gesamtlänge der Basaldendriten der 18 untersuchten Zellen Mittelwert Standardabweichung Serumhaltige Kontrolle 617 µm 1690,4 µm 2818,5 µm 3163,6 µm 3573,7 µm 3906,9 µm 2628,35 µm 1247,10 µm Serumfreie Kontrolle 374,7 µm 781,3 µm 1029,3 µm 1326,2 µm 1712,5 µm 2087,1 µm 1218,51 µm 624,34 µm C3 Toxin 534,8 µm 534,8 µm 985,6 µm 992,7 µm 1259,6 µm 2147,9 µm 1075,9 µm 597,25 µm Serumfreie Kontrolle 3 10 14 22 24 31 17,33 10,23 C3 Toxin 5 8 10 12 13 15 10,5 3,62 Tabelle A2: Länge der Apikaldendriten Mittelwert Standardabweichung Serumhaltige Kontrolle 10 16 21 25 35 36 23,83 10,34 Tabelle A3: Anzahl der basalen Verzweigungspunkte 57 Mittelwert Standardabweichung Serumhaltige Kontrolle 6 22 23 23 28 29 21,83 8,28 Serumfreie Kontrolle 9 10 15 16 16 17 13,83 3,43 C3 Toxin 2 4 10 11 13 18 9,67 5,89 Tabelle A4: Anzahl der Verzweigungspunkte auf apikaler Seite Mittelwert Standardabweichung Serumhaltige Kontrolle 112 114 253 254 682 689 350,67 266,87 Serumfreie Kontrolle 52 59 206 441 612 664 339 271,67 C3 Toxin 36 40 62 71 153 272 105,67 91,85 Tabelle A5: Anzahl der postsynaptischen Spines im Bereich der Basaldendriten Mittelwert Standardabweichung Serumhaltige Kontrolle 54 260 534 701 777 1040 561 358,91 Serumfreie Kontrolle 55 99 269 303 331 358 235,83 127,29 C3 Toxin 12 47 89 89 196 224 109,5 83,48 Tabelle A6: Anzahl der postsynaptischen Spines der Apikaldendriten Mittelwert Standardabweichung Serumhaltige Kontrolle 308 513 648 813 1459 1729 911,67 560,39 Tabelle A7: Gesamtzahl der postsynaptischen Spines 58 Serumfreie Kontrolle 107 158 509 799 881 995 574,83 378,81 C3 Toxin 48 87 151 242 267 496 215,17 161,72 Mittelwert Standardabweichung Serumhaltige Kontrolle 0,12/ µm 0,22/ µm 0,30/ µm 0,18/ µm 0,09/ µm 0,15/ µm 0,18/ µm 0,08/ µm Serumfreie Kontrolle 0,25/ µm 0,19/ µm 0,25/ µm 0,18/ µm 0,08/ µm 0,08/ µm 0,17/ µm 0,08/ µm C3 Toxin 0,25/ µm 0,06/ µm 0,08/ µm 0,02/ µm 0,076/ µm 0,17/ µm 0,11/ µm 0,08/ µm Serumfreie Kontrolle 0,13/ µm 0,13/ µm 0,15/ µm 0,18/ µm 0,27/ µm 0,32/ µm 0,20/ µm 0,08/ µm C3 Toxin 0,02/ µm 0,09/ µm 0,09/ µm 0,09/ µm 0,09/ µm 0,18/ µm 0,09/ µm 0,05/ µm Serumfreie Kontrolle 0,27/ µm 0,22/ µm 0,20/ µm 0,18/ µm 0,11/ µm 0,11/ µm 0,18/ µm 0,06/ µm C3 Toxin 0,21/ µm 0,07/ µm 0,09/ µm 0,02/ µm 0,08/ µm 0,13/ µm 0,10/ µm 0,06/ µm Tabelle A8: Spinedichte der Basaldendriten Mittelwert Standardabweichung Serumhaltige Kontrolle 0,09/ µm 0,15/ µm 0,19/ µm 0,20/ µm 0,22/ µm 0,29/ µm 0,19/ µm 0,07/ µm Tabelle A9: Spinedichte der Apikaldendriten Mittelwert Standardabweichung Serumhaltige Kontrolle 0,17/ µm 0,18/ µm 0,24/ µm 0,24/ µm 0,18/ µm 0,13/ µm 0,19/ µm 0,04/ µm Tabelle A10: Gesamt-Spinedichte 59 Danksagung An dieser Stelle möchte ich all jenen meinen Dank aussprechen, die mir in verschiedenster Hinsicht geholfen und zum Gelingen dieser Arbeit beigetragen haben. Als erstes möchte ich Herrn Prof. Dr. Johannes Seeger herzlich dafür danken, dass er gerne als Doktorvater die Betreuung meiner Arbeit übernommen hat. Mein besonderer Dank gilt Herrn Prof. Dr. Bernd Heimrich für die Überlassung des Themas, die Herstellung der Schnittkulturen und die nette Unterstützung in allen Arbeitsschritten der Dissertation. Sehr herzlich möchte ich mich dafür bedanken, dass er stets für mich erreichbar war und sich viel Zeit genommen hat. Eine bessere Betreuung hätte ich mir nicht wünschen können. Herrn Prof. Dr. Jürgen Schulte Mönting spreche ich meinen besten Dank für die arbeitsintensive statistische Auswertung meiner Daten aus. Frau Dr. Astrid Rollenhagen hat mich geduldig in die Handhabung der Neurolucida eingearbeitet und mir einige Tricks und Kniffe gezeigt, die meine Arbeit sehr erleichtert haben. Dafür vielen lieben Dank. Herr Dr. Imre Vida stand mir immer dann sehr hilfsbereit zur Seite, wenn die Neurolucida nicht so funktionierte, wie ich mir das von ihr wünschte. Vielen herzlichen Dank für diese Unterstützung. Bei Herrn Dr. Rudi Deisz bedanke ich mich sehr für die intrazelluläre Biocytinfüllung der CA1-Pyramidenzellen. Mein herzlicher Dank gilt auch Frau Prof. Dr. Gudrun Ahnert-Hilger, die freundlicherweise das C3 bot Toxin zur Verfügung gestellt hat. Ganz herzlich möchte ich mich bei meinem guten Freund Dr. Ralf Schmälzle bedanken, der mir mit wertvollen Tipps zum Layout der Arbeit und Korrekturlesen sehr behilflich war. Meinen Eltern möchte ich besonders herzlich dafür danken, dass sie mich während all der Jahre des Studiums und der Promotion in jeder Hinsicht unterstützt haben. Darüber hinaus war mir mein Vater bei der sprachlichen Durchsicht dieser Arbeit eine große Hilfe. Zu guter Letzt möchte ich meiner Frau Virna für ihre Liebe danken, die mir für die Arbeit an meiner Dissertation viel Kraft gab. 60