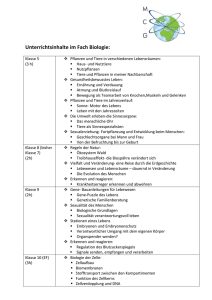
4_Evolution des Lebens
Werbung

J. Herlet; 2012-2016 Arbeit in 2 Teilen: I) Organisation des Lebendigen; II) Evolution des Lebens 1. Die kosmische Evolution bis zur Entstehung des Lebensraums Erde Wie wir heute wissen, entstand das Universum vor etwa 14 Milliarden Jahren mit einem „Urknall“ und dehnt sich seither aus. Damals war die gesamte Materie und Energie des heutigen Universums in einer kosmischen Keimzelle auf unvorstellbar kleinem Raum verdichtet. Der Urknall ist für uns nicht vorstellbar, er bezeichnet auch keine „Explosion“ in einem bestehenden Raum, sondern die gemeinsame Entstehung von Materie, Raum und Zeit. Aus diesem Urknall ist ein Universum hervorgegangen, in dem sich zunächst nur gasförmige Wolken aus Wasserstoff (75%) und Helium (25%) bildeten, den beiden leichtesten Elementen. Durch die Wirkung der Schwerkraft verdichteten sich die Wasserstoff- und Helium-Gase des frühen Universums zu ersten Sternen und Galaxien. Das Innere der zu Sternen kollabierenden Gaswolken erreichte durch den Gravitationsdruck so hohe Temperaturen und Dichten, dass atomare Verschmelzungsprozesse zündeten, bei denen Schritt für Schritt alle schwereren Elemente, wie z.B. Kohlenstoff, Stickstoff, Sauerstoff bis hin zu Eisen entstanden. Massereiche Sterne verbrennen schnell, Sterne von 10-facher Sonnenmasse haben ihren Brennstoffvorrat schon nach 100 Millionen Jahren aufgebraucht und enden dann in einer gewaltigen Supernova-Explosion. Bei diesen Supernovae wird der größte Teil des Sternes mit allen darin enthaltenen Elementen als Gas und Staub wieder an den Weltraum abgegeben, wo er als Baumaterial für die Bildung nachfolgender Sterngenerationen und Planeten verwendet werden kann. Alle Atome der lebenden Materie, die schwerer als Wasserstoff sind, wurden daher im Inneren längst erloschener Sterne gebrannt. Der Wasserstoff, der in Form von Wasser und anderen molekularen Verbindungen in uns steckt, entstand jedoch bereits in den ersten Minuten nach dem Urknall. So entstand auch das Sonnensystem mit seinen Planeten vor etwa 5 Milliarden Jahren aus einem interstellarem Gas- und Staubnebel, der durch seine eigene Gravitation kollabierte. Die Entstehung unseres Planetensystems ist noch nicht genau verstanden, grob muss sich folgendes abgespielt haben: der in sich zusammenstürzende Gasnebel geriet analog einer Pirouette in immer schneller Rotation, wobei sich die Materie im Zusammenspiel mit der Schwerkraft in einer flachen, mit rotierenden Scheibe um das Zentrum zu konzentrierte. Diese hat sich nach Geburt der Sonne unter Einfluss lokaler Gravitationszentren und Kollisionen nach und nach weiter zu Protoplaneten und schließlich zu den Planeten verdichtet. Dies erklärt auch, warum alle Planeten in Drehrichtung der Sonne um diese kreisen, und ihre Umlaufbahnen annähernd in der Äquatorebene der Sonne liegen.So entstand vor ca. 4.600 Millionen Jahren auch die Erde als eine weißglühende Kugel aus zähflüssigem Magma. Mit der Sonne entstand ein Stern vom Typ G, d.h. von einer Größe und Zusammensetzung, die eine lange Lebensdauer von ca. 10 Milliarden Jahren gleichbleibender Ausdehnung und gleichmäßiger Strahlungsleistung ermöglicht. Mit der Erde entstand ein Planet, dessen Bahn genau in der begrenzten Sphäre der Sonne verläuft, in der die Oberflächen-Temperatur eines Planeten im Rahmen der Toleranzgrenzen für Leben bleibt (auf dem es flüssiges Wasser geben kann; die Venus ist dafür zu heiß, der Mars vermutlich zu kalt) Das erste Äon der Erdgeschichte von vor etwa 4.600 bis etwa 4.000 Millionen Jahren wird Hadaikum (auch „vorgeologische Ära“) genannt. In dieser Phase wurde die Erde zunächst noch massiv von weiteren Materiebrocken und Kleinplaneten bombardiert, die durch das junge Sonnensystem schwirrten. Relativ gesichert ist die Theorie der zufolge auch der Mond vor 4,5 Milliarden Jahren durch Einschlag eines etwa Mars-großen Protoplaneten entstand, der das Wegschleudern einer großen Magmamenge bewirkte. Der Mond hat daher eine identische geochemische Zusammensetzung wie die Erdkruste. Das ständige Bombardement führte zu einem Anwachsen und einer Aufheizung der Erdmasse. Das glutflüssige Gestein differenzierte sich allmählich, die schwereren Elemente wie Eisen und Nickel sanken ab und bildeten den Erdkern, die leichten, wie Silizium und in Mineralien gebundener Sauerstoff formten allmählich Erdmantel und Erdkruste. Aus dieser Formationsphase ging die Erde vor etwa 4,4 Milliarden Jahren als eine Kugel mit einer verfestigten Rinde aus nacktem Granitund Basaltfels sowie einem heißen, glutflüssigem Inneren, und - daraus resultierend - einer heftigen vulkanischen Aktivität hervor. Das älteste Gestein der Erde aus dem Norden Kanadas (Kanadisches Schild) wurde auf ein Alter von 4,3 Milliarden Jahren datiert. Die Erde drehte sich damals noch so schnell, dass der Tag nur 10 Stunden hatte, der Mond befand sich fast 7 mal näher an der Erde, die Sonneneinstrahlung war wesentlich schwächer. (Die Abbremsung der Erdrotation seither durch die Gezeitenkräfte des Mondes resultierte in einer entsprechenden Zunahme des Bahndrehimpulses und damit des Abstands des Mondes.) Für die spätere Entstehung des Lebens auf dem Planeten Erde waren neben dem passenden Sonnenabstand vor allem dessen Größe und chemische Zusammensetzung wichtig. Auf Grund der richtigen Größe der Erde und des damit verbundenen Gravitationsdrucks konnte sich in ihrem Inneren ein glutflüssiger äußerer Eisenkern bilden. (Der innere Eisenkern ist wegen des zu hohen Gravitationsdruckes trotz einer Temperatur von ca. 6000 Grad Celsius fest). Durch Anwachsen und weitere Verdichtung des inneren Kerns sowie durch den Zerfall radioaktiver Elemente (wie Uran, Thorium oder Radium) in den Erdschalen wird das Erdinnere bis heute und weit in Zukunft in einem Maß weiter aufgeheizt, dass es flüssig bleibt und ständigen Wärmezirkulationen unterliegt. Ohne diese Konvektionsströmungen im Erdinneren gäbe es kein Magnetfeld, das uns von dem energiereichen Sonnenwind abschirmt, keinen Vulkanismus, der die Erdatmosphäre und die Erdozeane geschaffen hat, und keine Wanderbewegungen der Erdplatten (Plattentektonik), welche die Gebirgsbildung in Gang hält und das Abtragen der Kontinente und ihr Absinken in die Ozeane verhindert. Die Ur-Atmosphäre der Erde und die Ur-Ozeane entstanden in einer relativ kurzen, nur einige Jahrzehntausende dauernden Übergangsepoche als Folge dieser vulkanischen Aktivität. Dabei wurden im Erdinneren eingeschlossener Wasserdampf und Gase in ungeheuren Mengen herausgeschleudert. Wasserdampf kondensierte zu Regen und prasselte auf die noch weit über 100 Grad heiße Erdoberfläche nieder um sofort wieder zu verdampfen. Während dieser Abkühlungsphase besaß die Erde eine unglaublich dichte Atmosphäre aus kochendem Wasserdampf und Gasen, der kein Schimmer Sonnenlicht durchdringen ließ, dazu jahrtausendelang anhaltende gewaltige Wolkenbrüche und pausenlos tobende Gewitter. Als die Erdoberfläche hinreichend abgekühlt war sammelte sich das Regenwasser in den großen Senken der Erdoberfläche und bildete die Ur-Ozeane. Die Atmosphäre aus Gasen wie Kohlendioxyd, Methan Ammoniak, Schwefelwasserstoff, Stickstoff, und wurde klar und durchsichtig, enthielt aber keinen freien Sauerstoff, was an den Verwitterungsspuren von Gesteinen der damaligen Erdepoche erkennbar ist. Entstammt alles irdische Oberflächenwasser vulkanischer Aktivität? Der über Jahrmilliarden anhaltende Vulkanismus hat jedenfalls weiter enorme Mengen von Gestein und Wasser an die Erdoberfläche gebracht. Auch heute stoßen die Vulkane der Erde pro Jahr noch ca. 3 km3 Gestein und große Mengen Wasserdampf aus. Einer anderen Theorie zu Folge entstammt aber auch ein erheblicher Anteil des irdischen Wassers aus Einschlägen wasserreicher Asteroiden in der Frühzeit der Erde. Weitgehend ungeklärt ist, warum auf den anderen erdähnlichen Planeten wie Venus und Mars kaum oder kaum mehr Wasser zu finden ist. 2. Die Entstehung des Lebens auf der Erde durch chemische Evolution Die Einheit des Lebens. Aus den Erkenntnissen der modernen Biochemie wissen wir heute, dass alles Leben stammesgeschichtlich miteinander verwandt ist, also aus einer Urzelle stammt, von deren Überleben einst alles abhing. Dies ergibt sich eindeutig aus der Universalität zentraler Lebensstrukturen und Prozesse. Alle Lebewesen bauen DNA und RNA aus den gleichen 5 Nukleotid-Bausteinen auf, alle nutzen die gleichen 20 genetisch programmierten Aminosäuren für den Aufbau ihrer Proteine, alle steuern die ProteinBiosynthese über den universell gültigen genetischen Code, allen dient ATP als zentraler Energiezwischenspeicher für Zellarbeit. Auch der Grad der Unterschiede bei den Proteinen gleicher Funktion ist sehr gering. Leben aus dem Kosmos? Wir wissen heute, dass im Weltraum organische Grundstoffe vorkommen, dass es ferner in den obersten Schichten der Atmosphäre mikroskopische Organismen gibt, die auch einen ungeschützten Aufenthalt im Weltraum überleben würden, und dass es für diese kleinsten Organismen prinzipiell auch möglich ist, mittels des Strahlendrucks der Sonne in Weltraum hinaus zu wandern. Also könnte auf diese Weise auch Leben auf die Erde gelangt sein. Dies ist jedoch sehr unwahrscheinlich und würde das Problem der Lebensentstehung nur an einen anderen Ort im Kosmos verlagern. Der biochemische Aufbau des Lebens aus Proteinen und Nukleinsäuren spricht auch eindeutig dafür, dass das Leben unter den spezifischen Bedingungen der frühen Erde entstanden ist. Leben als Ergebnis einer chemischen Evolution: Man nimmt daher heute an, dass das Leben auf der Erde in einem etwa 500 bis 700 Millionen Jahre dauernden Prozess einer chemischen (abiotischen) Evolution entstand, der nach Formation des Planeten Erde, seiner Uratmosphäre und der Urozeane vor ca. 4,4 Milliarden Jahren begann. Da alle heutigen und auch alle fossil erhaltenen Lebewesen einer Abstammung sind, ist diese abiotische Entstehung von Leben entweder ein sehr unwahrscheinlicher und daher einmaliger Akt gewesen, oder Leben konnte sich deshalb nur einmal bilden, weil die Entstehung neuen Lebens unter dem Konkurrenzdruck existierenden Lebens unmöglich ist; spätere Ansätze der Natur in diese Richtung wurden einfach aufgefressen, bevor sie sie sich etablieren konnten. Die ältesten bisher entdeckten fossilen Lebensformen sind 3,4 -3,5 Milliarden Jahre alte Einschlüsse algenartiger Einzeller ähnlich heutigen Cyanobakterien sowie fossile Algenriffe, sogenannte Stromatolithen, die aus Ablagerungen von Stoffwechselprodukten frühzeitlicher Mikroorganismen in Gewässern entstanden sind. (Mögliche Lebensspuren aus der Isua-Formation in Grönland, deren Alter auf 3,8 Mrd. Jahre datiert wurde, sind umstritten. Bei diesen 3 µm großen Objekten deutet der C- Isotopenanteil auf photosynthetisch aktive Mikroorganismen hin.) Bei aller Primitivität stellen diese Organismen bereits ziemlich komplizierte Lebensformen dar. Klar ist auch, dass die ersten Lebensformen viel einfacher gewesen sein müssen als das einfachste heutige Bakterium. Auch im einfachsten heute bekannten Bakterium laufen gleichzeitig Hunderte chemischer Reaktionen ab. Dieses komplexe Zusammenwirken hat sich sicher über lange Zeiträume aus ursprünglicheren Lebensformen Schritt für Schritt entwickelt. Aber auch die Urzelle muss bereits die Grundeigenschaften des Lebens besessen haben, einen Stoffwechsel zur Energieversorgung und die Fähigkeit zur Vermehrung. Für die Entstehung von Leben waren zwei entscheidende Voraussetzungen gegeben, das Vorhandensein von Wasser und das Fehlen von freiem Sauerstoff in der Uratmosphäre. Wasser ist der stoffliche Mittler für alle biochemischen Reaktionen und hat eine Reihe lebensstützender Eigenschaften (s. Teil 1, Kapitel 4). Das Leben ist im Wasser entstanden und hat dieses auch nach seiner Entstehung für ca. 3 Milliarden Jahre nicht verlassen. Alle bekannten Lebensvorgänge finden in Anwesenheit von Wasser statt. Auch unsere Zellen bestehen zum größten Teil aus Wasser. Freier Sauerstoff in der Uratmosphäre wäre mit dem Wasserkreislauf auch in die Meere gelangt und hätte wegen seiner hohen chemischen Aktivität die Bildung und hinreichende Beständigkeit komplexer organischer Moleküle gar nicht zugelassen. Er entstand erst später durch das Leben selbst, und dieses musste dann lernen, die zersetzende Wirkung des Sauerstoffs im Zellstoffwechsel auszuschalten und diesen in der Zellatmung zu nutzen. Der Ablauf der chemischen Evolution bis zur Zündung des Lebens kann heute nur ansatzweise skizziert werden, von einem wirklichen Verständnis ist man noch sehr weit entfernt. Unterscheiden kann man dabei (wenigstens) 4 spezifische Phasen: 1. Die Entstehung organischer Grundbausteine (Nukleotide, Aminosäuren etc.) in Uratmosphäre und Ur-Ozeanen und deren Anreicherung in den Ur-Ozeanen. 2. Die Entstehung komplexer Biomoleküle als elementare Grundbausteine des Lebens durch Verkettung von Aminosäuren zu Proteinen und von Nukleotiden zu Nukleinsäuren. 3. Die Entstehung präbiotischer Zellstrukturen, mit einem primitiven Stoffwechsel, der diesen Strukturen aber eine gewisse Stabilität verlieh. 4. Die Entstehung einer ersten replikationsfähigen Urzelle Die Entstehung organischer Grundbausteine aus anorganischen Verbindungen der Atmosphäre und des Meeres wird durch das Miller-Urey-Experiment gestützt. Dabei setzt man ein Gemisch aus Wasserdampf und Gasen der Uratmosphäre elektrischen Entladungen aus, welche Gewitterblitze nachbilden sollen. Bei diesem und ähnlichen Experimenten entstanden nach gewisser Zeit viele organische Grundbausteine heutiger Lebewesen (z.B. Aminosäuren, Nukleotid-Basen wie Adenin, einfache Fettsäuren). Durch den Wasserkreislauf haben sich diese Bausteine im Lauf der Zeit auch in den Ur-Ozeanen angereichert. Als Energiequelle könnten auch vulkanische Wärme oder UV-Strahlen gedient haben. Die Entstehung komplexer Biomoleküle: Die Entstehung von Nukleotiden oder langkettigen Membran-bildenden Lipiden sowie die Verkettung (Polymerisation) von Aminosäuren zu Proteinen bzw. von Nukleotiden zu Nukleinsäuren ist jedoch durch diese Experimente kaum zu erklären. Die Entstehung und Anreicherung dieser Biomoleküle in einer „Ursuppe“ von Lebensbausteinen hat sehr wahrscheinlich eine kontinuierliche Energieversorgung auf chemischer Basis erfordert, die beständig Energie zur Synthese dieser Biomoleküle geliefert hat. Geeignete Urerde-Szenarien sind noch Gegenstand der Forschung. Die oberen Schichten der Ur-Ozeane wurden von dem damals viel näher um die Erde kreisenden Mond derart aufgewühlt, dass sich darin kaum die notwendige Konzentration an organischen Stoffen hätte ansammeln können. Heute werden eher warme Flachgewässer oder am Boden des Urmeeres gelegene Hydrothermal Quellen (schwarze Raucher) als die Orte vermutet, in denen das Leben entstand. In Flachgewässern konnten sich komplexe Biomoleküle trotz der damals sehr intensiven UVStrahlung anreichern, ohne gleich wieder zerstört zu werden. Dies wird durch den nach seinem Entdecker benannte Urey-Effekt gestützt. Danach hat das energiereiche UV-Licht der Sonne, das die Atmosphäre wegen des Fehlens von Sauerstoff zunächst ungehindert durchdringen konnte, die Wassermoleküle an der Oberfläche der Seen und Ozeane solange in entweichenden Wasserstoff und sich in der Atmosphäre anreichenden Sauerstoff gespalten, bis eine Sauerstoffkonzentration von 0,1 Prozent der heutigen erreicht war. Damit wurde die Filterwirkung gegen das UV-Licht so stark, dass die Spaltung von Wassermolekülen zum Abbruch kam. Sauerstoff absorbiert das UV-Licht der Sonne durch Bildung und Zerfall von Ozon (3O2-> 2O3->3O2). Die sich so einpendelnde geringe Sauerstoffkonzentration bildete mit den sonstigen Eigenschaften der damaligen Atmosphäre daher einen UV-Filter, der genau in dem Strahlungsfenster optimalen Schutz bot, in dem Proteine und Nukleinsäuren, die zentralen Lebensbausteine am strahlenempfindlichsten sind. Eine geeignete Energiequelle für die Bildung komplexer Biomoleküle ist allerdings für Flachgewässer schwieriger zu erklären. Als besonders interessanter Ansatz gilt heute die von Günter Wächtershäuser entwickelte Theorie der Eisen-Schwefel-Welt, nach der das Leben auf der Erde an der Oberfläche von Eisen-Schwefel-Mineralen entstanden ist, die sich heute noch durch geologische Prozesse an Tiefsee-Vulkanen bilden und zur Frühzeit der Erde noch wesentlich häufiger auftraten. Das Konzept bietet mehrere Vorteile: die Grundbausteine (Methan, Ammoniak, H2 etc.) der Biosynthese kommen in vulkanischen Gasen vor, eine verlässliche Energiequelle für die Entstehung komplexer Bio-Moleküle könnte durch Bildung von Pyrit (FeS2) und Wasserstoff aus Schwefelwasserstoff und Eisenmonosulfid (H2S + FeS -> FeS2+ H2 + Energie) bestanden haben. Organismen, die sich heute chemosynthetisch von Schwefel-Verbindungen ernähren (oder fotosynthetisch auf Basis H2S und fotochemischer Eigenschaften von Pyritkristallen), stehen dem Ursprung des Lebens möglicherweise sehr nahe. Zusätzlich bieten Pyrit und andere Eisen-Schwefel-Minerale positiv geladene Oberflächen, an denen sich die überwiegend negativ geladenen Biomoleküle konzentrieren und miteinander reagieren können. Auch laufen chemische Umsetzungen bei erhöhter Temperatur viel schneller ab. Die Bildung präbiotischer Strukturen (Zellvorläufer) Wahrscheinlich war der Urozean über eine lange Phase der Entstehung des Lebens mit einer großen Zahl von organischen Verbindungen angefüllt, die sich ständig in unterschiedlichen Zusammensetzungen verklumpten und wieder zerfielen. Dabei entstanden irgendwann auch Verklumpungen, die sich von ihrer Umgebung durch eine membranartige Grenzschicht abgrenzten und durch diese zusammengehalten wurden, und die über einen einfachen Stoffwechsel- und Energie-Austausch auch eine gewisse Stabilität erreichten. Durch verschiedene Experimente konnte gezeigt werden, dass sich unter geeigneten Bedingungen Proteinaggregate bilden können, die sich von ihrer Umwelt membranartig abgrenzen und mit dieser Umwelt Stoffe austauschen (Koazervate, Mikrosphären). Die Entstehung einer ersten replikationsfähigen Urzelle: Aus einer solchen zellähnlichen Struktur muss dann irgendwann ein vermehrungsfähiges System entstanden sein. Nach der sogenannten „RNA-Welt-Hypothese“ könnten einfache RNA-Moleküle, die ihre eigene Replikation katalysierten und sich daher bevorzugt vermehrten, Vorläufer der ersten Organismen gewesen sein. Einige von diesen erlangten die Eigenschaft, die Synthese gewisser Proteine zu katalysieren, die selbst wieder die Synthese der RNA katalysierten. Dieses sich gegenseitig bedingende Verhältnis zwischen Nukleinsäuren (hier RNA) und Proteinen ist allem Anschein nach die elementare Wurzel dessen, was wir Leben nennen. Immerhin hatte die Natur auch für diesen letzten Schritt der chemischen Evolution von einer Ursuppe aus anorganischen Verbindungen zur ersten vermehrungsfähigen Zelle hunderte von Millionen Jahren des Probierens. Diese Theorie wird insbesondere durch die Entdeckung gestützt, dass RNA nicht nur (wie die DNA) als Speicher der Erbinformation dient, sondern neben Proteinen an der katalytischen Aktivität der Ribosomen zur Proteinsynthese entscheidend beteiligt ist. Die Ribosomen finden sich bei allen Lebewesen und gelten daher als sehr ursprüngliche Zellbausteine. RNA wird für älter gehalten als DNA, auch deshalb, weil die heutige DNA-Replikation mit der Bildung eines RNA-Startmoleküls beginnt. Die Synthese von Proteinen ist ohne DNA, aber nicht ohne RNA möglich. 3. Die Evolutionstheorie Die biologische Evolutionstheorie erklärt und beschreibt die Entstehung der Arten als das Ergebnis von Evolution. Sie wurde 1858 von Charles Darwin (zeitgleich und unabhängig von Alfred Russel Wallace) begründet. In seinem berühmten 1859 erschienenen Buch On the Origin of Species formulierte Darwin 2 revolutionäre Thesen: (1) Alle Arten stammen durch Evolution von einem/wenigen gemeinsamen Vorfahren ab. (2) Hauptursache der Evolution der Arten ist die natürliche Selektion der zufällig besser angepassten Varianten. Bis Mitte des 19. Jahrhunderts war die Schöpfungstheorie weit verbreitet, nach der Gott die Welt und alle Lebensformen in einem Schöpfungsakt erschaffen hat, der einige Tausend oder höchstens Millionen Jahre zurück liegen sollte. Erste Vorstellungen, dass sich die heutigen Lebensformen aus Urformen über lange Zeiten entwickelt haben, finden sich aber schon bei Empedokles (ca. 450 v. Chr.). Auch bei Kant finden sich Vermutungen zur Verwandtschaft aller Lebensformen und zur Entstehung neuer Arten aus Vorläuferformen. Darwins und Russels Theorie stützte sich auf zwei wesentliche Erkenntnisse: 1. Kampf ums Dasein: alle Lebewesen müssen sich in ihrem jeweiligen Lebensraum einem ständigen Kampf um die zu knappen Überlebensressourcen stellen. 2. Selektion: Es überleben vor allem die Arten, die am besten an den jeweiligen Lebensraum angepasst sind („survival of the fittest“), die also Eigenschaften besitzen, die Ihnen die besten Überlebens- und Fortpflanzungschancen ermöglichen. Die ständige Selektion der genetisch am besten an die jeweiligen Lebensräume angepassten Lebensformen ist der Motor für die allmähliche Entstehung neuer Arten. Entscheidend für diese sogenannte natürliche Selektion ist die Wahrscheinlichkeit der Fortpflanzung, die nicht unbedingt auf physischer Stärke beruht. Eine wichtige Rolle spielt z.B. auch die Bevorzugung von Sexualpartnern mit bestimmten Merkmalen (sexuelle Selektion), möglicherweise der Grund für Ausbreitung der Blauäugigkeit seit ihrer Entstehung vor 8000 bis 10.000 Jahren. Die Selektion setzt dabei an der genetischen Variabilität einer Population an. Die Ursachen dieser Variabilität wurden aber erst in den Folgejahrzehnten im Rahmen der „synthetischen Evolutionstheorie“ (August Weismann, Julian Huxley u.a.) begründet. Dabei werden alle Prozesse („Evolutionsfaktoren“) betrachtet, durch die der Genpool einer Population, das ist die Gesamtheit aller Erbanlagen in dieser Population, verändert wird. Der wichtigste Evolutionsfaktor neben Selektion sind Mutationen, aber auch Rekombination, Gendrift, Genfluss, geografische Isolation und vernetzte Evolution spielen eine Rolle. Mutationen sind spontane (zufällige) Veränderungen der Erbinformation, also der BasenSequenzen der Zell-DNA. Durch Mutationen entstehen immer wieder Individuen mit veränderten Eigenschaften. In manchen Fällen ermöglichen die neuen Eigenschaften eine bessere Überlebenschance in dem betreffenden Lebensraum oder ein Ausweichen in einen anderen Lebensraum. Die so begünstigten Individuen pflanzen sich zahlreicher fort. Mutationen sind also die Voraussetzung dafür, dass immer wieder neue Selektionen stattfinden können. Für die Evolution von Bedeutung sind nur Mutationen, die in der DNA von Zellen stattfinden, aus denen Keimzellen hervorgehen. Nur dann wird die veränderte Erbanlage über die befruchtete Eizelle auf die Nachkommen übertragen. Mutationen sind meistens rezessiv (z.B. viele Erbkrankheiten). An einem rezessiven Merkmal kann die Selektion nur dann ansetzen, wenn dieses von beiden Eltern übertragen wird. Außerdem können die Zellen auf Grund der Redundanz der Erbinformation in der Doppelstrang- DNA gewisse Gen-Mutationen auch reparieren. Rekombination bezeichnet die Kombination männlicher und weiblicher Erbanlagen bei sexueller Vermehrung. Dadurch wird der Genpool einer Population laufend durchmischt, es entstehen immer wieder Individuen mit neuen Erbanlagen und Eigenschaften. Unter Gendrift versteht man eine zufallsbedingte Änderung des Genpools, z.B. durch eine Naturkatastrophe oder Seuche. Auch Genfluss, also die Durchmischung von 2 Populationen der gleichen Art, oder eine durch veränderte Umweltbedingungen ausgelöste geografische Isolation kann eine evolutionäre Weiterentwicklung begünstigen. Vernetzte Evolution ist ein weiteres Muster in der Evolution vieler biologischer Arten. Dabei entstehen biologische Verwandtschaftsverhältnisse nicht durch Abstammung sondern z.B. durch horizontalen Gentransfer zwischen Bakterienarten (dies fördert die Verbreitung , der Antibiotika-Resistenz), durch virale Transduktion (Gen-Übertragung durch Viren ist auch im menschlichen Genom nachgewiesen) oder durch Kreuzung (Hybridbildung) zwischen hinreichend kompatiblen Arten. Höherentwicklung - die Entwicklung des Lebens vom Einfachen zum Komplexen: Hatten sich im Verlauf der Entwicklung vorteilhafte Strukturen herausgebildet, wurden sie künftig nicht mehr aufgegeben. Dadurch addieren sich in den Organismen vorteilhafte Eigenschaften. Das führt zu jener Tendenz, die mit „Höherentwicklung“ bezeichnet wird. Gewisse vorteilhafte Konstruktionsprinzipien wurden dabei von der Natur auch mehrfach unabhängig erfunden oder zumindest von elementaren Ansätzen unabhängig zu ausgereiften Konstruktionen weiter entwickelt, so z.B. Mehrzelligkeit, sexuelle Vermehrung, Augen, Zähne, Warmblütigkeit (bei Vögeln und Säugetieren). Selbstorganisation, Emergenz und Seitensprünge der Evolution: Emergenz und Selbstorganisation sind Begriffe, die hin und wieder im Zusammenhang mit der Evolution eines Systems genannt werden. Selbstorganisation bezeichnet dabei die Tendenz zu einer Strukturbildung auf Grund einer sich selbst verstärkenden Wirkung (z.B. durch positive Rückkopplung). Mit Emergenz ist das überraschende Auftauchen einer neuen Systemeigenschaft gemeint, die sich z.B. durch Zusammenwirkung unabhängig entwickelter Merkmale oder auf Grund neuer externer Bedingungen herausbildet. Was den Zufall genetischer Variationen so erfolgreich macht ist die Auslese. Gelegentlich beobachten wir einen Seitensprung der Evolution: eine Struktur, die im Hinblick auf eine bestimmte Funktion selektiert wurde, wird sekundär für eine neue Funktion brauchbar, so wie das Federkleid der Vögel zunächst wohl der Wärmeregulierung und erst danach auch dem Fliegen diente. Stammesentwicklung (Phylogenese) und biologische Systematik: Die evolutionäre Entwicklung der Arten (Stammesentwicklung, Phylogenese) lässt sich in Form eines „Abstammungsbaumes“ darstellen. Eine Art (Species) bezeichnet dabei eine homogene Gruppe von Lebewesen, die – bei sexueller Vermehrung – miteinander fruchtbare Nachkommen erzeugen können, oder - bei ungeschlechtlicher Vermehrung - in allen wesentlichen biologischen Merkmalen übereinstimmen. Eine monophyletische Gruppe enthält genau alle Nachfahren einer Stammart sowie die Stammart selbst. Die jüngste Art, die gemeinsame Stammart zweiter Nachfolge-Arten ist, wird auch als „Link“ bezeichnet, bzw. als „missing link“, wenn diese Art noch nicht bekannt oder fossil erfasst ist. Weiter lassen sich die Arten nach dem Grad ihrer stammesgeschichtlich begründeten gemeinsamen Merkmale klassifizieren. Diese Einteilung der Arten wird auch als biologische Systematik (Taxonomie) bezeichnet. Eine Hierarchie solcher Klassen (Taxa) bilden hierbei: Art, Gattung, Familie, Ordnung, Klasse, Stamm. Der Mensch (Art „homo sapiens“) z.B. gehört zur Gattung Homo, zur Familie der Hominiden (Menschenaffen), zur Ordnung der Primaten, zur Klasse der Säugetiere und zum Stamm der Chordatiere (Unterstamm: Wirbeltiere). Das Tempo der Evolution: Gibt es Zeiten und Umstände, in denen das Leben qualitative Sprünge machte, in denen besonders viele neue Lebensformen entstanden? Als „Makroevolution“ wird ein Evolutionsvorgang bezeichnet, der über Artgrenzen hinaus führt, bei dem also eine neue Gattung, Familie, Ordnung, Klasse oder gar Stamm entsteht, oder bei dem neue Anpassungszonen besiedelt werden oder ganz neue morphologische Strukturen entstehen. Solche Evolutionssprünge sind nach Meinung der Mehrzahl der Evolutionsbiologen meist jedoch nur eine Summierung von sehr vielen kleineren sogenannten mikro- evolutionären Veränderungen über lange Zeiträume hinweg. Die Dynamik und Richtung des Evolutionsprozesses wird aber sicher durch äußere Umweltbedingungen stark beeinflusst. Beispiele hierfür sind: 1. Populationsgröße: Je kleiner eine Population ist, umso schneller können sich vorteilhafte Mutationen durchsetzen. Ein über viele Fortpflanzungszyklen anhaltender, in eine bestimmte Richtung wirkender Selektionsdruck auf eine kleine (z.B. in geografischer Isolation lebende) Population kann daher den Genpool rasch in Richtung auf die Herausbildung und Vervollkommnung des selektierten Merkmals verändern. 2. Massenaussterben: Von den Milliarden Tier- und Pflanzenarten, die einmal auf der Erde gelebt haben, hat kaum eine die gesamte Erdgeschichte hindurch überlebt. 99,9 Prozent aller jemals entwickelten Arten sind ausgestorben, viele eher sang und klanglos, andere in katastrophalen Sterbewellen, die nicht nur eine Art, sondern gleich Tausende auslöschten. Im Verlauf der Erdgeschichte kam es zu mehreren Massenaussterben, die den Verlauf der Evolution entscheidend beeinflussten. Nach solchen Ereignissen wurden große potentielle Lebensräume zur Neueroberung durch Leben frei. Es folgte für gewöhnlich eine Phase der Expansion der überlebenden Organismen, oft durch Adaption an nicht besetzte ökologische Nischen. Als Ursachen solcher Aussterben werden das Verschwinden von Lebensraum durch abrupten Klimawandel, insbesondere durch Eiszeiten, oder als Folge von starkem Vulkanismus oder von Meteoriteneinschlag angenommen. Beim katastrophalsten Massenaussterben der Erdgeschichte vor circa 250 Millionen Jahren starben 75-90% aller Arten aus. Die Dinosaurier, die danach entstanden, wurden nach einem weiteren Massenaussterben vor etwa 200 Millionen Jahren zur dominierenden Landwirbelgruppe und starben selbst beim großen Massenaussterben vor 66 Millionen Jahren aus, zusammen mit fast allen anderen damals lebenden Saurierarten. Dies wiederum begünstigte den Aufschwung und die Entfaltung der Säugetiere. 3. Das Erdmagnetfeld: Ein „Anheizen der Evolution“ ergab sich vermutlich in gewissen Phasen der Erdgeschichte z.B. dadurch, dass sich das Magnetfeld der Erde etwa alle 300.000 Jahre umpolte (zuletzt vor 780.000 Jahren). Während solcher Phasen ist die Erde dem Sonnenwind stärker ausgesetzt, was sicher zu höheren DNA-Mutationsraten durch diese hochenergetische Strahlung führte, und die Evolution möglicherweise anheizte. 4. Die Kambrische Explosion: so wird die umfassende Umformung der Tierwelt zu Beginn des Kambriums vor etwa 550 Millionen Jahren bezeichnet. In einem geologisch kurzen Zeitraum von bis zu 5 bis 10 Millionen Jahren entstanden viele neue Arten, und es entwickelten sich die grundlegenden Baupläne vieler mehrzelliger Tierstämme, die seitdem die Erde bevölkern. Zu dieser Artenvervielfältigung hat sicher der im Zeitraum seit 700 MYA (Million Years Ago) stark ansteigende Sauerstoffgehalt der Atmosphäre und die damit verbundene Möglichkeit einer höheren Energieausbeute aus der Nahrung entscheidend beigetragen, was auch die Entwicklung Energie-verbrauchenden Fähigkeiten und die Entwicklung spezieller Organe zur Sauerstoffaufnahme (Tracheen, Kiemen, Lunge) begünstigte. Nach jüngsten Forschungsergebnissen könnte aber auch eine geologisch bedingte massive Zunahme der Kalziumkonzentration in den Urmeeren die Entstehung von Hartschalen und Stützskeletten sowie das Zusammenlagern von Zellen in dieser Epoche entscheidend gefördert haben. Die anscheinende Beschleunigung des Evolutionstempos seit der kambrischen Explosion ist vermutlich aber auch darauf zurückzuführen, dass alle fundamentalen Prinzipien, die höherem Leben zugrunde liegen, in der unvorstellbar langen Zeit davor bereits als Keime oder Ansätze entwickelt waren, so dass es bei der Ausbildung „höherer Lebensfunktionen nur noch um die Perfektion von ansatzweise bereits vorhandenen Funktionen ging. (z.B. Augenflecken bereits beim Einzeller). Ferner nahmen durch die Zunahme der Merkmale auch die Ansatzpunkte für Mutationen immer weiter zu, was sicher auch zu einer Beschleunigung der Artenvielfalt und zu einer kürzeren durchschnittlichen Lebensdauer einzelner Arten führte. Aus Fossilienfunden kann man ableiten, dass die durchschnittliche Lebensdauer einer Art seit dem Kambrium immer weiter abnahm von 2,7 Millionen Jahren bis einigen hunderttausend Jahren im Jura. 5. Methoden der Erforschung der Entwicklungsgeschichte des Lebens 1) Molekulargenetische Analysen Dass alle heutigen Lebensformen einen gemeinsamen Ursprung haben, zeigt sich u.a. am prinzipiell gleichen Aufbau aller Proteine aus 20 Aminosäuren und an der universell gültigen Bauvorschrift zur Synthese von Proteinen (genetischer Code). Anhand der Unterschiede der DNA oder funktionsspezifischer Proteine kann aber auch auf den Grad der Verwandtschaft und auf den Zeitabschnitt beginnender Auseinanderentwicklung einzelner Arten geschlossen werden. Das Atmungsenzym Citochrom c aus den Mitochondrien der Eukaryoten besteht aus über 100 Aminosäuren. Es unterscheidet sich vom Menschen zum Rhesusaffen nur um 1 Aminosäure, die Unterschiede von den Säugern zu den Vögeln sind 10-15, zu den Fischen etwa 20 zur einzelligen Hefe 40-50 Aminosäuren. 2) Vergleichende Individualentwicklung Ernst Haeckel formulierte die Hypothese, dass die Embryonalphase eines Individuums die Evolution seiner Art rekapituliert (Biogenetische Grundregel). Diese Regel wird heute jedoch als zu starke Vereinfachung betrachtet. Wiederholt werden nicht die Adultstadien, sondern die frühen Embryonalstadien der jeweiligen Vorformen einer Art. Beispiele einer solchen Wiederholung sind das Auftreten von Kiemenspalten bei allen Wirbeltierembryonen. Dies belegt, dass alle heutigen Wirbeltiere von Fischen abstammen. 3) Vergleichende Morphologie, d.h. die Auswertung von strukturellen (morphologischen) und anatomischen Merkmalen von Fossilien und heutigen (rezenten) Lebewesen. Schon Darwin sah in Ähnlichkeiten morphologischer Strukturen wichtige Hinweise für eine entwicklungsgeschichtliche Verwandtschaft. Z.B. besitzen alle Säugetiere 7 Halswirbel, die vorderen Extremitäten der Säugetiere, die Flügel der Vögel und die Flossen von Delphinen haben die gleiche Skelettstruktur (1 langer Knochen - 2 Parallelknochen - 5 Endglieder). Morphologische Stufenreihen ermöglichen den zeitlichen Ablauf der Evolution zu erkennen. ( z.B. die Stufenreihe von Herz und Blutkreislauf der Wirbeltiere: Fische: ein Vorhof, eine Kammer, nur ein Blutkreislauf; Amphibien: zwei Vorhöfe, eine Kammer, starke Vermischung von arteriellem (sauerstoffreichem) und venösem Blut; Reptilien: unvollständige Trennung der Herzkammer; Vögel, Säugetiere: vollständige Trennung auch der Herzkammer, damit keinerlei Vermischung von sauerstoffreichem und sauerstoffarmen Blut im Herzen.) Von den homologen Strukturen und Merkmalen, die auf einen gemeinsamen evolutionären Ursprung zurück zuführen sind, sind analoge Merkmale zu unterscheiden, die durch Anpassung an eine ähnliche Funktion und ähnliche Umweltbedingungen entstanden sind. Beispiele für eine solche biologische Konvergenz sind die Flossenbildung bei Fischen und Walen oder die Flügel von Vögeln, Fledertieren und Flugsauriern. 4) Fossilienfunde Da die Geschichte des Lebens zu einem guten Teil aus Fossilienfunden rekonstruiert wurde, sollten wir uns zunächst klar machen, dass wir nur einen sehr kleinen Teil kennen. Der Grund: „Ein Fossil zu werden, ist nicht einfach“ (Bill Bryson), nur einer von einer Milliarde Knochen wird vermutlich zu einem Fossil. Von allen heute in Deutschland lebenden Menschen werden also (bei 82 Millionen Menschen mit je 206 Knochen) nur ca. 17 Knochen zum Fossil werden, nicht einmal ein Zehntel eines vollständigen Skeletts. Und das muss dann auf der Fläche von gut 350.000 km² erst einmal gefunden werden. Von den geschätzt bis 50 Milliarden Arten, die die Erde in ihrer Geschichte hervorgebracht hat, sind ca. 250.000 als Fossilien bekannt: eine unter 200.000 Arten. Außerdem ist das vermittelte Bild nicht neutral: Tiere mit Gehäusen werden viel leichter als Weichtiere zu Fossilien, und große Organismen sind leichter zu finden als mikroskopisch kleine. Fossilien können entstehen, wenn der Zerfallsprozess der Materie aufgehalten wird, z.B. durch Sauerstoffabschluss bei Einlagerung in Sedimente, allmähliche Anreicherung mit Mineralkörnern und druckabhängige Verfestigung (Versteinerung). Die Altersbestimmung von Gesteinen und Fossilien erfolgt in erster Linie mittels Methoden der radiometrische Datierung. Dabei wird für bestimmte, natürlich vorkommende radioaktive Isotope das Mengenverhältnis Mutter- zu Tochter-Isotop bestimmt. Da man oft recht genau begründen kann, wie dieses Verhältnis zu Lebzeiten des Fossils oder bei Entstehung des Gesteins gewesen sein muss, kann man aus den beiden Verhältnissen - heute zu damals - und der bekannten Zerfallsrate des Isotops das Alter des Fossils bzw. Gesteins bestimmen. Es gibt eine ganze Reihe solcher Datierungsverfahren. Die Uran-Blei-Datierung beruht auf dem Zerfall von Uran235 / Uran238 in Blei und eignet sich z.B. zur Altersbestimmung der Erde (Halbwertszeiten 704 MJ / 4500 MJ). Die Radion-KarbonMethode beruht auf dem Zerfall von C14 in N14 (Halbwertszeit 5.730 Jahre) und eignet sich z.B. zur Altersbestimmung abgestorbener organischer Materialien bis zu einem Alter von etwa 60.000 Jahren. Generell hängt die Genauigkeit dieser Verfahren davon ab, wie zutreffend die Annahmen über die ursprünglichen Isotopenverhältnisse sind, und ob weitere Effekte als nur die Zerfallsraten für das heutige Isotopenverhältnis zu berücksichtigen sind. Sogenannte „Brückenfossilien“ zeigen Verzweigungsstellen im Baum der Evolution, sind also Übergangsformen zwischen zwei Taxa (Stämmen, Ordnungen etc.). Beispiele sind der Urvogel Archäopteryx (ca. 150 MYA/Million Years Ago), der Merkmale von Reptilien und Vögeln aufweist, oder der Fischlurch Ichthyostega (360 MYA), der Fisch- und AmphibienMerkmale aufweist. Als lebende Fossilien werden gemeinhin Arten bezeichnet, die in morphologisch gleicher oder sehr ähnlicher Form schon vor Hunderten von Jahrmillionen gelebt haben. Oft haben sie einmal weite Bereiche der (damaligen) Erde besiedelt, kommen jedoch heute nur noch in wenigen Regionen vor (Reliktvorkommen) und galten teilweise bereits als ausgestorben. Geografische Isolation, passende Rückzugsgebiete und ein nicht zu große Spezialisierung können zur Langlebigkeit dieser Arten beigetragen haben. Beispiele aus dem Pflanzenreich sind Baumfarne und der Ginko-Baum, Beispiele aus dem Tierreich sind Quastenflosser (Fossilien-Datierungen 400-70 MYA, Wiederentdeckung 1938) und Lungenfische (Fossilien-Datierungen seit 400 MYA), beides die engsten Verwandten der ausgestorbenen Stammart aller Landwirbeltiere. 6. Die biologische Evolution im Präkambrium Fassen wir zusammen: vor etwa 4,4 Milliarden Jahren war die Abkühlung der Erdkruste soweit fortgeschritten, dass sich Uratmosphäre und Ur-Ozeane bilden konnten, und in ihnen die chemische Evolution einsetzen konnte. Vor etwa 3,7 bis 4 Milliarden Jahren dürften die ersten kernlosen Zellen entstanden sein, dann hat sich die Natur nochmals 3 Milliarden Jahre Zeit gelassen bis zum Beginn der Evolution höherer mehrzelliger Lebewesen vor rund 700 Millionen Jahren. Erst dann wagte das Leben auch den Sprung auf das Land, das in der langen Zeit davor überwiegend aus Gestein, Geröll und Sand bestand und bar jeden Lebens war. Kontinente entstanden und zerbrachen wieder, Gebirge formten sich und wurden abgetragen. Die Tage wurden allmählich länger, denn die Drehung der Erde um ihre Achse verlangsamt sich durch die Reibung der Gezeiten. Vor 700 Millionen Jahren betrug der Tag nur etwa 20 h. Die etwa 4,6 Milliarden Jahre seit der Entstehung der Erde werden aus geologischer Sicht in vier Äonen unterteilt. Als Hadaikum, auch „vorgeologische Ära“, wird das erste Äon der Erdgeschichte von vor etwa 4.600 bis etwa 4.000 Millionen Jahren bezeichnet. Im Hadaikum verfestigte sich die Erdkruste, die Ur-Ozeane entstanden, und einige hundert Millionen Jahre chemischer Evolution liefen ab. Das daran anschließende Archaikum erstreckte sich von vor ca. 4.000 bis ca. 2.500 Millionen Jahren. Im Archaikum entstand das Leben auf der Erde und es wurden grundlegende Lebensprozesse wie Proteinbiosynthese, DNA-Replikation, Glykolyse und Photosynthese erfunden (biochemische Grundlagen: siehe Teil-1, Kapitel 5 und 6). Die biologische Evolution begann vor 3,7 bis 4 Milliarden Jahren mit der Entstehung der ersten Zellen. Die oxygene Photosynthese wurde vor ca. 3,3 bis 3,5 Milliarden Jahren, erfunden. Der dabei entstehende Sauerstoff wurde jedoch gleich wieder durch die noch in großen Mengen vorhandenen leicht oxidierbaren Stoffe (z.B. Eisenerze) chemisch gebunden. Erst nachdem diese Verbindungen vor etwa 2,5 Milliarden Jahren fast komplett oxidiert waren, stieg der Sauerstoffgehalt in der Atmosphäre und in den Ozeanen bis auf 2-3%, also rund zehn Prozent des heutigen Werts von 21%. Das Proterozoikum (gr. „frühe Lebewesen“) ist das dritte Äon der Erdgeschichte. Es reicht vom Ende des Archaikums vor ca. 2.500 Millionen Jahren bis zum Beginn des Kambriums vor ca. 550 Millionen Jahren, es umfasst also fast 2 Milliarden Jahre, oder fast die Hälfte der erdgeschichtlichen Entwicklung. Kennzeichen des Proterozoikums ist die Entstehung einer sauerstoffhaltigen Atmosphäre und damit einhergehend die Entwicklung aeroben, Sauerstoffnutzenden Lebens. • Erste Bakterien mit aerober Zellatmung entwickelten sich als Reaktion auf die allmähliche Anreicherung von freiem Sauerstoff in den Ur-Ozeanen vor 2,3 bis 2,5 Milliarden Jahren. • Die ersten einfachen Mehrzeller (Zellkolonien) traten vor etwa 2,1 Milliarden Jahren auf. • Vor 2,1 bis 1,5 Milliarden Jahren, entwickelten sich die ersten einzelligen Eukaryoten mit Zellkern und spezialisierten Organellen, Mitochondrien zur Zellatmung und Plastiden zur Photosynthese. • Seit 1000 MYA (Million Years Ago) stieg der Sauerstoffgehalt der Erdatmosphäre von 2%-3% zunächst allmählich, dann ab etwa 700 MYA stark bis auf etwa 12% zu Beginn des Kambriums (550 MYA) an. Es bildete sich außerdem eine Ozon-Schicht und damit eine Abschirmung der energiereichen UV-Strahlen.In Folge entwickelten sich vielzellige Tiere, die sich fossil ab ca. 650 MYA nachweisen lassen. Das Leben begann sich aus dem Wasser auch auf das Land auszubreiten. Mit Abschluss des Proterozoikums an der Wende zum Kambrium scheinen auch die meisten Stämme des Tierreichs als Ausgang für die folgende sogenannte kambrische Explosion bereits vorhanden gewesen zu sein. Der blaue Planet: Durch die Zunahme des Sauerstoffs in der Atmosphäre gegen Ende des Proterozoikums wurde das blau-wellige Licht stärker gestreut, was den Himmel und die UrOzeane allmählich in blauem Licht erscheinen ließ. (In der Frühzeit der Erde könnte der Himmel – bedingt durch Methan in der Atmosphäre - für unser Auge eher orange-rötlich gewirkt haben. Farben kamen allerdings erst mit der Entwicklung unseres Sehapparates in die Welt. Das schmale Frequenzband sichtbaren Lichts, das die Erdatmosphäre durchlässt, wird von unserem Sehapparat (Augen + Gehirn) in mit Farben unterlegte Teilfrequenzbänder differenziert.) Schneeball Erde: Fachwissenschaftlich umstritten ist eine Theorie, nach der die Erde vor etwa 700 Millionen Jahren (und eventuell auch in früheren Phasen) komplett mit Eis bedeckt und auch die Ozeane größtenteils zugefroren waren. Unstrittig ist, dass es im Lauf der Erdgeschichte immer wieder weit verbreitete Eiszeiten gab. Das Phanerozoikum (gr. „sichtbares Leben“) ist das jüngste Äon der Erdgeschichte, es umfasst die jüngsten 550 Millionen Jahre bis heute und gliedert sich in drei Zeitalter, und diese wiederum in mehrere Perioden: • Paläozoikum (Erdaltertum): Kambrium bis Perm; Beginn vor ca. 550 Millionen Jahren. • Mesozoikum (Erdmittelalter): Trias bis Kreide; Beginn vor ca. 250 Millionen Jahren. • Känozoikum (Erdneuzeitalter): Beginn vor 65 Millionen Jahren bis heute Das Erdaltertum ist geprägt von der Entfaltung höheren tierischen und pflanzlichen Lebens und dem Auszug des Lebens aus dem Wasser, das Erdmittelalter ist das Zeitalter der Saurier, die Erdneuzeit ist geprägt von der Entwicklung der Säugetiere bis hin zum Menschen. Die erste Periode des Erdaltertums ist das Kambrium. Die Äonen vor dem Phanerozoikum fasst man auch unter dem Begriff Präkambrium zusammen. Die geochronologische Systematik der Perioden des Phanerozoikums wird durch das Auftreten charakteristischer Leitfossilien in aufeinander aufbauenden Gesteins- und Sedimentschichten der Erdkruste begründet und definiert. Die Bestimmung von Alter und Dauer dieser Perioden erfolgt mittels radiometrischer Datierung. Meilensteine der Evolution im Präkambrium: (1) Erfindung der Photosynthese Vermutlich gewannen die ersten Lebewesen ihre Energie aus dem Abbau von abiotisch entstandenen organischen Nährstoffen oder chemosynthetisch aus einfachen anorganischen Verbindungen, wie z.B. heutige Schwefelbakterien. Die Photosynthese, also der Aufbau von energiereichen Kohlehydraten aus anorganischen Stoffen (CO2 und H2O oder H2S) mit Hilfe von Lichtenergie, wurde sehr früh in der Geschichte des Lebens erfunden. Möglicherweise geschah sie unter dem evolutionären Druck der ersten Ernährungskrise des Lebens, verursacht durch eine rapide Abnahme abiotisch entstandener, organischer Nährstoffe in den Ozeanen. Als gesichert gilt, dass zunächst, vermutlich bereits vor etwa 3,5-3,7 Milliarden Jahren, eine einfachere, anoxygene Photosynthese (z.B. auf Basis Schwefelwasserstoff H2S) erfunden wurde, die noch heute bei einigen Bakterien vorkommt. Zellen mit lichtabsorbierenden Porphyrin-Molekülen bildeten dabei möglicherweise den Ausgangspunkt. Die oxygene Photosynthese, bei der Wasser (H2O) statt H2S gespalten und Sauerstoff frei gesetzt wird, wurde wahrscheinlich bereits vor 3,3 bis 3,5 Milliarden Jahren durch Vorgänger der heutigen Cyanobakterien (Blaualgen) genutzt. Sie wird heute von allen Pflanzen, Algen und vielen Bakterienarten praktiziert und erzeugt den gesamten in der Erdatmosphäre vorkommenden Sauerstoff. (biochemische Grundlagen: s. Teil-1, Kapitel 6). (2) Erfindung der Zellatmung Die Zellatmung, also die energetische Auswertung von Nährstoffen mittels Sauerstoff, wurde vor ca. 2,3 bis 2,5 Milliarden Jahren unter dem evolutionären Druck der Zunahme freien Sauerstoffs in der Atmosphäre und den Ur-Ozeanen erfunden. Sauerstoff ist sehr reaktionsfreudig, er reagiert leicht mit einigen empfindlichen organischen Biomolekülen, auch mit freien organischen Substanzen, und zerstört diese. Für die anaeroben frühen Lebewesen war die vor 2,5 Milliarden Jahren beginnende Freisetzung von Sauerstoff in die Atmosphäre und im Wasser eine Katastrophe. Nur wenige dieser Organismen überlebten, konnten sich in sauerstofffreie Lebensräume zurückziehen oder entwickelten Enzyme, die toxische Sauerstoffverbindungen unschädlich machten. Schließlich entwickelten sich in einigen Mikroorganismen Enzyme und Stoffwechselprozesse, die es erlaubten den Sauerstoff zur kontrollierten chemischen Verbrennung (Oxydation) der Nährstoffe ausnutzen, um Energie für das Wachstum der eigenen Zellen zu erzeuge. Damit war die Zellatmung geboren. (biochemische Grundlagen: s. Teil-1, Kapitel 6). (3) Evolution der Eukaryoten Die ältesten Mikrofossilien, welche weitgehend unumstritten als Überbleibsel von Eukaryoten anerkannt sind, fanden sich in etwa 1,5 Milliarden Jahre altem Gestein in Nordaustralien. Bei den etwa 2,1 Milliarden Jahre alten Mehrzeller-Fossilien Gabonionta und Grypania (siehe nächster Abschnitt) handelt es sich vermutlich um organisierte Zellkolonien von Prokaryoten. Die bis zu 1,8 Milliarden Jahre alten Acritarcha-Mikrofossilien weisen zumindest eine für Eukaryoten typische Zellgrößen auf. Als gesichert gilt, dass alle heutigen Eukaryoten einen gemeinsamen Ursprung haben. Dieser Stammvater der Eukaryoten entstand vermutlich vor 1,5 -2,1 Milliarden Jahren. Entscheidender Vorteil dieser neuen Lebensarchitektur ist die durch voneinander abgegrenzte Zellbereiche und spezialisierte Organelle (Zellkern, Mitochondrien, Plastiden usw.) mögliche Leistungssteigerung bzgl. Anzahl und Komplexität der Stoffwechselprozesse sowie die dadurch mögliche höhere genetische Informationsvielfalt. Die Eukaryoten besitzen bis 1.000 Mal mehr Erbmaterial DNA als die kernlosen Prokaryoten. Sie sollten die sexuelle Fortpflanzung „erfinden“, und aus ihnen sollten sich die großen Reiche der Pflanzen, Pilze und Tiere entwickeln, die heute das Bild der Erde bestimmen. Protisten, d.h. ein- bis wenigzellige Eukaryoten, die keinem dieser 3 Reiche zugeordnet werden können, sind aber auch heute noch sehr vielfältig vorhanden (z.B. die Gruppe der Amöben oder die Augentierchen). Bis heute unverstanden ist die Entstehung des Zellkerns. Die meisten Theorien gehen von einer Fusion eines Archaeon mit einem Bakterium aus, da das Genom der Eukaryoten sowohl Übereinstimmungen mit dem von Archaeen als auch mit dem von Bakterien aufweist. Möglicherweise war der Wirt der Fusion auch ein damals noch existierender Vertreter der RNA-Welt, was die RNA-basierte Proteinsynthese außerhalb des Zellkerns erklären könnte. Auch horizontaler Gentransfer könnte bei der Entstehung und Entwicklung der Eukaryoten eine Rolle gespielt haben. Mit Loki (Lokiarchaeota) wurde in Atlantiksedimenten kürzlich ein Archaeon entdeckt, dessen Erbgut viele spezifisch eukaryotische Gene enthält. Weitgehend akzeptiert ist die Endosymbionten-Theorie, nach der zunächst die Mitochondrien durch symbiotischen Einverleibung aerober Bakterien in frühen Eukaryoten entstanden sind, sich dann bei einigen dieser Organismen, den Vorläufern der Grünalgen und Pflanzen, auch die Chloroplasten (Plastiden) durch eine entsprechende Endosymbiose mit Blaualgen-ähnlichen Prokaryoten entwickelt haben. Die dabei von der Wirtszelle eingefangenen Prokaryoten wurden nicht sofort verdaut, sondern versorgten ihren Jäger mit Energie bzw. stellten Glukose-Moleküle durch Photosynthese her. Die eingefangenen Symbionten wurden dann im Laufe der Zeit zu Zellorganellen. Diese Theorie wird dadurch gestützt, dass Mitochondrien und Plastiden von einer doppelten Zellmembran umgeben sind, ein eigenes Genom (DNA) enthalten und über eine eigene Proteinbiosynthese verfügen; sie können sich durch Teilung selbst vermehren. (4) Erfindung der Mehrzelligkeit Die Mehrzelligkeit ist im Laufe der Evolution mehrmals unabhängig entstanden. Echte Mehrzeller bilden sich nicht durch Zusammenschluss selbständiger Zellen (obwohl heute auch solche einfache Lebensformen bekannt sind), sondern aus einem Teilungsprozess ohne Verselbständigung der Tochterzellen und durch eine zunehmende Arbeitsteilung und Spezialisierung einzelner Zellgruppen. Der evolutionäre Vorteil liegt wohl zum einen in der Körpergröße (schützt vor gefressen werden), dann in Effizienzsteigerung durch Arbeitsteilung und Spezialisierung (z.B. schnellere Beweglichkeit, bessere Nährstoffspeicherung). Dazu mussten die vielzelligen Organismen erst einen Mechanismus entwickeln, sich zu spezialisierte Teilzellpopulationen und Zelltypen zu differenzieren. Vermutlich wird dies durch zeitlich abgestimmte Aktivierungen und Blockierungen relevanter Gene gesteuert. Ferner mussten Mechanismen des Zusammenhaltens (Adhäsion) von Zellen und der Informationsübertragung zwischen Zellen entwickelt werden. (Das Zusammenhalten der Zellen übernehmen “biologische Klebstoffe”. Neben diesem gibt es außen an der Zellmembran spezifische “Nieten”, die dafür sorgen, dass sich nur Zellen einer bestimmten Art verbinden, und “Rezeptoren”, die bestimmte Moleküle binden können auf diese Art übertragen Zellen Informationen.) Der individuelle Alterstod: Vielzeller sind nicht mehr wie Einzeller potenziell unsterblich, sondern ihr Körper stirbt nach einer gewissen Zeit aus Altersschwäche ab. Einzeller sterben dagegen nur durch äußere Einwirkung, zum Beispiel wenn sie gefressen werden, austrocknen oder verhungern, aber niemals aus inneren Ursachen von selbst. Die ältesten fossil bekannten vielzelligen Lebewesen traten möglicherweise bereits vor etwa 2,1 Milliarden Jahren im Proterozoikum auf. Bei den etwa 1 cm langen spiraligen Fossilien Grypania spiralis handelte es sich vermutlich um kettenförmige Zellkolonien aus Bakterien. Die 2010 im afrikanischen Gabun entdeckten Gabonionta sind bis zwölf Zentimeter große, morphologisch relativ komplex strukturierte Fossilien, möglicherweise aber auch noch keine echten Mehrzeller mit differenzierten Zelltypen, sondern eine Kolonie gleichartiger Zellen, die eine organisierte Morphologie verbunden mit einer Aufgabenteilung bildeten. Vielzellige Tiere (Metazoa) lassen sich fossil erstmals ab einem Alter von etwa 650 Millionen nachweisen (z.B. Ediacara-Fauna, Schwämme). Möglicherweise konnte sich komplexes vielzelliges Leben erst dann entwickeln, als genug Sauerstoff in der Luft war, um durch Zellatmung ausreichend Energie für das aufwändigere, da zu koordinierende vielzellige Leben zu gewinnen. (5) Erfindung der Sexualität Die bloße Zellteilung als Methode der Vermehrung hat den großen Nachteil, dass unabhängig voneinander entstandene Mutationen nicht in einem Individuum zusammen treffen können, was die Variabilität des Genmaterials einer Art und damit die schnelle Anpassungsfähigkeit an die Umwelt einschränkt. Selbst einfache Bakterien verfügen daher oft über einen Konjugation genannten Mechanismus zum Austausch von Erbanlagen. Dieser horizontale Gentransfer findet teilweise sogar zwischen verschiedenen Bakterienarten statt und trägt unter anderem zur Ausbreitung von Antibiotika-Resistenzen bei. Mit der Entwicklung der Eukaryoten und ihrer komplexeren DNS kam die Methode des Genaustausches aber offenbar an Grenzen. Die Erfindung der sexuellen Fortpflanzung beruht zunächst auf einer Verdopplung der genetischen Information (Chromosomen) bei den eukaryotischen Einzellern. Das war auch deswegen vorteilhaft, weil ein „Fehler“ auf einem Chromosom durch das andere Chromosom ausgeglichen werden konnte. Bei sexueller Fortpflanzung werden aus solcher Zellen mit doppeltem (diploiden) Chromosomensatz nun durch Halbierung (Reduktionsteilung) spezielle Fortpflanzungszellen (Keimzellen) mit wieder einfachem Chromosomensatz gebildet; bei Verschmelzung zweier solcher Keimzellen im Akt der Befruchtung entsteht dann der Kern (die Zygote) eines neuen Lebewesens. In diesem sind die Chromosomensätze wieder diploid und rekombiniert, d.h. zusammengesetzt aus 2 verschiedenen Keimzellen, vorhanden. Da die sexuelle Fortpflanzung also spezialisierte Keimzellen braucht, ist sie nur bei echten Vielzellern möglich. Die Spezialisierung der Keimzellen führte weiter, es entstanden große, nährstoffreiche Eizellen und kleine, schwimmfähige Spermien-ähnliche Zellen – womit die Unterscheidung zwischen männlich und weiblich erfunden war. Die sexuelle Vermehrung bedeutete zunächst einen gravierenden Nachteil in Gestalt der Männchen: nur etwa die Hälfte der Nachkommen kann wieder gebären und benötigt dafür außerdem einen weiblichen Partner. Bei Zwittern wie dem Regenwurm kann ein Tier sowohl männliche als auch weibliche Keimzellen bilden, was zwar die Variationsvielfalt einschränkt, aber auch vom Zwang befreit, zur Fortpflanzung immer einen geeigneten Partner finden zu müssen. Bei Pantoffeltierchen, einzelligen Protisten, finden bereits sexuelle Vorgänge mit einem vollständigen Austausch der genetischen Information statt. Dabei lagern sich – wie bei der Konjugation von Bakterien – zwei Zellen aneinander an. Dann tauschen sie über einen Verbindungskanal jeweils einen zuvor gebildeten haploiden Zellkern aus, der mit der verbliebenen Zellkernhälfte wieder zu einem diploiden Kern verschmilzt. Anschließend entstehen durch ungeschlechtliche Fortpflanzung (Zweiteilungen) Tochterzellen mit einer neuen Eigenschaftskombination. 7. Die biologische Evolution seit dem Kambrium Das vierte Äon der Erdgeschichte, das Phanerozoikum (Zeitalter des „sichtbaren Lebens“) umfasst die jüngsten 550 Millionen Jahre der Erdgeschichte bis heute und gliedert sich in Paläozoikum (Erdaltertum), Mesozoikum (Erdmittelalter) und Känozoikum (Erdneuzeit). Das Erdalterum ist geprägt von der Entfaltung höheren tierischen und pflanzlichen Lebens und dem Auszug des Lebens aus dem Wasser, das Erdmittelalter ist das Zeitalter der Reptilien, die Erdneuzeit ist geprägt von der Entwicklung der Säugetiere bis hin zum Menschen. Das Kambrium ist die erste geologische Epoche (Periode) des Erdaltertums. In den letzten 150 Millionen Jahren vor Beginn des Kambriums stieg der Sauerstoffanteil der Atmosphäre von zuvor etwa 3% auf 12% stark an. Dies ermöglichte eine bessere energetische Verwertung von Nährstoffen und begünstigte in der Folge einen enormen Evolutionsschub der Mehrzeller, die sogenannte kambrische Explosion. Ferner kam es zur Bildung von Ozon (O3) in höheren Schichten der Atmosphäre und damit einer Abschirmung der Erdoberfläche von UV-Strahlen, was für die Entwicklung des Lebens auf dem Land eine wichtige Rolle spielte. Ursache dieser Sauerstoffzunahme könnten starke Algenblüten gewesen sein, die durch eine geologisch ausgelöste, vermehrte Düngung der Urzeit-Ozeane durch Phosphor angefacht wurden. Darauf deuten marine Ablagerungen von Phosphatgestein im Erdaltertum hin, die immer dann auftraten, wenn Kontinentalverschiebung und Gletscher besonders starke Erosion verursachten. Aber auch ein explosives Ergrünen der Landflächen durch Algen zu Ende des Präkambriums könnte den Anstieg der Sauerstoffkonzentration bewirkt haben. In einem geologisch kurzen Zeitraum entstanden viele neue Arten, und es entwickelten sich die grundlegenden Baupläne vieler mehrzelliger Tierstämme mit echtem Zellgewebe, die seitdem die Erde bevölkern, darunter die ersten Zweiseitentiere (Bilateria) mit einer zentralen Körperachse, achsensymmetrischem Körperbau und einem Zentralnervenstrang vom Kopf zum Rumpfende, von denen u.a. alle Gliederfüßer (u.a. Insekten, Spinnentiere, Trilobiten) und Wirbeltiere abstammen. Ein entscheidender Schritt für die Evolution des Lebens zu dieser Zeit stellt die Entwicklung einer schützenden Hartschale in Form eines Außenskeletts dar. Die damit verbundene längere Erhaltung und bessere Fossilbildung der Hartteile hatte auch einen plötzlichen Anstieg der bis in unsere Tage erhaltenen Fossilien zur Folge. Manche Forscher interpretieren die kambrische Explosion deshalb auch als Explosion des Fossil-Aufkommens. Landgang: Der Auszug aus dem Wasser und die Besiedlung von Landflächen durch lebende Organismen begann bereits im Präkambrium. Der Landgang von Grünalgen, Cyanobakterien und Pilzflechten wird anhand von Proteinsequenz-Analysen auf einen Zeitraum von vor 700 bis 1000 Millionen Jahren geschätzt. Im Erdaltertum nahmen dann auch Pflanzen und Tiere nach und nach vom Land Besitz. Dies ist durch fossile Belege relativ gut dokumentiert. Dem Landgang der Pflanzen (480-460 MYA/ Million Years Ago) folgte der der Gliederfüßer (ca. 450 MYA). Der Landgang der Wirbeltiere (400-360 MYA) vollzog sich durch einen evolutionären Übergang von Fischen zu Vorformen der heutigen Amphibien. Durch den Landgang hat sich das Leben ein neues Ökosystem erschlossen, musste auch inhärente Nachteile (höherer Energieverbrauch durch Tragen des eigenen Körpergewichts, Ausgleich von großen Temperaturunterschieden, Wassermangel und damit das Problem, Ballast und Giftstoffe effizient zu entsorgen) durch neue Formen der Adaption überwinden. Begünstigt wurde der Landgang durch die Abschirmung der harten UV-Strahlung durch den sich ausbildenden Ozonfilter. Eine größere Unabhängigkeit vom Wasser entwickelten ca. 300 MYA die Amniontiere (Vorfahren der Reptilien, Vögel und Säugetiere), bei denen sich der Embryo in einer Fruchtwasser-gefüllten Blase – bei Vögeln und den meisten Reptilien zusätzlich durch eine hartschalige Eihülle aus Kalk geschützt – entwickelt. Perioden des Erdaltertums (Paläozoikum): Kambrium (542- 485 MYA): Am Anfang steht die Entwicklung fast aller wirbellosen Tierstämme in einem Zeitraum von 5 bis 10 Millionen Jahren (Kambrische Explosion); erstmalig Tiere mit äußerem Skelett (Hartschalen), darunter viele muschel- und krebs- artige Formen, wie die als Leitfossilien geltenden Trilobiten (ausgestorben 250 MYA) und erste aquatische Räuber. Gegen Ende des Kambriums ereignete sich ein großes Massenaussterben, etwa 80% aller Tierarten starben aus. Das Klima war gemäßigt mit zunehmender Erwärmung und Anstieg des atmosphärischen CO2-Anteils bis auf das 12-fache (!) des heutigen Niveaus (Sauerstoffgehalt nur ca. 12,5%); der große Südkontinent Gondwana umfasste die damals zusammenhängenden Kontinente Südamerika, Afrika, Antarktika, Australien und Indien. Ordovizium (485- 444 MYA): Landgang der Moose; erste marine Riffgemeinschaften, erste noch kieferlose Fische (z.B Schleimaale); gegen Ende globale Abkühlung mit Vereisung der Südhalbkugel, möglicherweise ausgelöst durch eine CO2-Reduktion in der Atmosphäre als Folge der Ausbreitung der Landpflanzen; damit einhergehend großes Massenaussterben. Silur (444- 419 MYA): Ausbreitung erster Gefäßpflanzen mit Wurzeln an den Ufern der Meere und Flüsse; Landgang der Gliederfüßer wie Skorpione, Krebse; erste Wirbeltiere (Kiefermäuler) und deren Weiterentwicklung zu ersten Knorpel- und Knochenfischen. Global warmes Klima mit hohem atmosphärischen CO2 -Gehalt; Vereinigung kleinerer Urkontinente zu Laurussia nördlich des Äquators; Kaledonische Gebirgsbildung (u.a. Appalachen und Rumpfgebirge Skandinaviens). Devon (419- 359 MYA): Zeitalter der Fische (dominierend: bis 10 m lange Panzerfische); aus den Knochenfischen entwickeln sich Quastenflosser und Lungenfische, in einer Schwesterlinie die Amphibien als erste Landwirbeltiere; erste geflügelte Insekten. Massive Ausbreitung der Landpflanzen und damit Anstieg des atmosphärischen Sauerstoffgehalts und Rückgang des CO2 -Gehalts; erste Wälder aus Bärlappgewächsen, Urfarnen (Biomasse der ältesten Kohlevorkommen); Klima warm; gegen Ende großes Massenaussterben (u.a. alle Panzerfisch-Arten). Karbon (359- 299 MYA): Zeitalter der Farne und Bärlapp-Pflanzen mit bis zu 40 m hohen baumartigen Formen; erste Samenpflanzen (Nacktsamer); Diversifikation der Insekten und Amphibien; Entwicklung erster Amniontiere als Schwestergruppe der Amphibien mit einer Fruchtwasserblase (Amnion) für die Embryo-Entwicklung, die sich damit unabhängig vom Wasser fortpflanzen können. Spätere Aufspaltung dieser Amniontiere in Synapsiden (Vorfahren der Säugetiere) und Sauropsiden (Vorfahren der Reptilien und Vögel); beide Gruppen sind zunächst Reptilien-artig, aber mit unterschiedlichem Schädelbau. Entstehung des letzten Superkontinents Pangäa durch Vereinigung der Urkontinente Gondwana und Laurussia; dabei Bildung vieler heutiger Faltgebirge (u.a. Ural, Pamir, europäische Mittelgebirge, Antiatlas); feucht-warmes Klima im Wechsel mit Eiszeiten; Atmosphäre bis ca. 32% Sauerstoff, CO2 ca. 2-faches heutiges Niveau. Perm (299-252 MYA): Die Synapsiden stellten die häufigsten und gleichzeitig größten Landwirbeltiere; aus ihnen entwickeln sich „säugerähnliche Reptilien“ (Therapsiden) mit einigen morphologischen Säuger-Merkmalen, welche die Stammart der Säugetiere enthalten. Parallel entfalteten sich die Reptilien (Sauropsiden). Gegen Ende des Perms ereignete sich das größte Massenaussterben der Erdgeschichte: in einem Zeitraum von ca. 200.000 Jahren starben etwa 95% aller meeresbewohnenden Arten, etwa 2/3 aller Reptilien- und Amphibien-Arten sowie 1/3 aller Insektenarten aus. Als primäre Ursache werden gewaltige Vulkanausbrüche im urzeitlichen Sibirien vermutet, welche große Mengen CO2 freisetzten und damit eine starke Klimaerwärmung und eine Versauerung der Ozeane verursachten. Perioden des Erdmittelalters (Mesozoikum): Trias (252-201 MYA): Der bereits im Perm erfolgte Umschwung der Pflanzenwelt von Farnen zu Nacktsamern setzte sich in der Trias weiter fort. Die Reptilien nahmen einen ungeheuren Aufschwung. Es entstanden viele neue Ordnungen, u.a. die Dinosaurier (als zunächst kleine Reptilien), Flugsaurier, Fischsaurier (Ichtyosaurier), Krokodile und Echsen. Aber auch Übergangsformen zu den Säugetieren sind möglicherweise im späten Trias entstanden. Der Superkontinent Pangäa, der fast die gesamte Landmasse enthielt und sich vom Nordpol bis zum Südpol spannte, ermöglichte die uneingeschränkte Verbreitung der Landtiere. Am Ende der Trias ereignete sich ein Massenaussterben, dem fast alle Landwirbeltiere zum Opfer fielen, möglicherweise ausgelöst durch gewaltigen MagmaFreisetzungen des beginnenden Auseinanderbrechens von Pangäa. Das Klima der Trias war warm bis heiß und trocken (atmosphärischer O2-Anteil ca. 16%). Jura (201-145 MYA): Die in der Trias erschienenen Dinosaurier entfalteten sich und wurden zur dominierenden Lebensform. Dabei entstanden auch die größten Landtiere aller Zeiten, pflanzenfressende Sauropoden mit bis 40 m Länge und 70 Tonnen Gewicht. Aus dem frühen Jura von China stammt der vermutet älteste Fund eines Säugetier-Fossils (Hadrocodium), der „Urvogel“ Archäopteryx wurde in Gesteinsschichten des späten Jura gefunden (Solnhofen, Eichstätt). Die Flora wurde von den Nacktsamern dominiert (darunter Nadelholzgewächse wie Mammutbäume, aber auch Ginkgos und Palmfarne). Während des frühen Jura zerfiel Pangäa in die Bruchstücke Nordamerika, Eurasien und den Südkontinent Gondwana. Das Klima war warm (atmosphärischer O2-Anteil 26%). Kreide (145- 66 MYA): Reptilien, speziell auch Saurier entwickelten neue Arten und breiteten sich aus; es entstanden Riesenformen von Flugsaurier und Dinosaurier (u.a. Tyrannosaurus Rex in Nordamerika). Die bereits in der Jura begonnene Aufspaltung der Ursäuger und Vögel in verschiedene Entwicklungslinien setzte sich fort. Während der Kreide entwickelten sich die ersten strauchigen Blütenpflanzen, darunter Laubbäume wie Ahorn, Eiche und Walnuss. Dies begünstigte eine neue ökologische Nahrungskette Insekten-Vögelkleine Raubsäuger. Gräser breiteten sich auf dem Festland aus. Der Zerfall von Gondwana, der bereits im Jura begonnen hatte, setzt sich in der Kreide fort. Es kommt zur Trennung des noch zusammenhängenden Australien/Antarktika und des zu Beginn der Kreide ebenfalls noch zusammenhängenden Afrika/Südamerika, auch Indien spaltet sich ab. Beginn der alpidischen Gebirgsbildung. Das Klima in der Kreide war allgemein warm und ausgeglichen. Die Pole waren eisfrei, entsprechend war der Meeresspiegel sehr hoch (atmosphärischer O2-Anteil 30 %, auch hoher CO2 -Gehalt ). Am Ende der Kreidezeit kam es zu einem weltweiten Massenaussterben, dem u.a. fast alle Saurierarten zum Opfer fielen. Als Ursache wird der Einschlag eines Meteoriten nahe der Halbinsel Yucatan angesehen, aber auch großflächige Vulkanausbrüche in Vorderindien können zu einer Klimakatastrophe beigetragen haben. Ungeklärt ist, wieso nur bestimmte Tiergruppen aussterben und andere nicht. So gingen zwei ganze Ordnungen mit allen ihren Familien, Gattungen und Arten zugrunde - ohne Ausnahme, ohne Überlebende, darunter alle Nicht-Vogel-Dinosaurier, Flugsaurier, Plesiosaurier und Fischsaurier, während alle anderen Wirbeltiergruppen (Fische, Amphibien, Reptilien, Vögel und Säuger) überlebten. Nimmt man eine Verdunkelung des Planeten und die fast vollständige Einstellung der Photosynthese an, sollten alle Tierarten gleich betroffen sein. Auch das Überleben Blüten-bestäubender Insekten kann nur schwer erklärt werden. Perioden der Erdneuzeit (Känozoikum): Paläogen (65 - 23 MYA): Nach dem Aussterben der großen Dinosaurier kam es zur Entwicklung der Vögel und der höheren Säugetiere (Plazenta-Tiere). Letztere differenzierten sich schnell und wurden zu den beherrschenden Landtieren. Im Paläogen nahmen die Kontinente in etwa die heutigen Positionen ein. Die Indische Platte begann auf die Eurasische zu treffen, es bildete sich der Himalaya und Karakorum. Durch plattentektonische Vorgänge bildeten sich auch die amerikanischen Kordilleren und die Faltgebirge des alpidischen Gürtels (u.a. Alpen, Karpaten, Kaukasus). Die einzelnen Kontinente waren anfangs isoliert, über eine Landbrücke zwischen Afrika und Eurasien, die sich vor 27 Millionen Jahren bildete, konnten sich die Tiere jedoch wieder weit verbreiten. Nur Australien nahm eine andere Entwicklung. Neogen (23 MYA – 2,6 MYA): Das Neogen ist gekennzeichnet durch die Entfaltung der Säugetiere und der Vögel. Zwischen den Kontinenten Nord- und Südamerika bildete sich eine Landbrücke. Nachdem die Antarktis schon seit etwa 30 Millionen Jahren vergletschert war, kam es im Zuge einer klimatischen Abkühlung vor 2,6 Millionen auch zu einer starken Vergletscherung der Nordhemisphäre. Quartär (2,6 MYA – heute): Das Quartär ist gekennzeichnet durch die Entwicklung der Gattung Mensch (Homo) bis zum heutigen Homo sapiens. Es beginnt mit dem jüngsten Eiszeitalters der Erdgeschichte (dem sogenannten Pleistozän), welches mit einem Wechsel von Kaltzeiten und Warmzeiten bis zum Ende der letzten Eiszeit vor 11.700 Jahren andauerte. Zu Ende des Pleistozäns, im Zeitraum von 50.000 bis 10.000 Jahren vor unserer Zeit, kam es weltweit zu einem Massenaussterben von großen Säugetieren. Mit Ausnahme Afrikas und des südlichen Asiens starben alle Arten mit über 1000 Kilogramm Gewicht und 80% aller Arten mit 100 bis 1000 Kilogramm Gewicht aus. Inwieweit klimatische Veränderungen oder die Bejagung durch den Menschen (Overkill-Hypothese) oder beides die Schuld dafür tragen, ist immer noch umstritten. Heute ist sicher der Mensch für das sich beschleunigende Aussterben vieler Arten verantwortlich. Nach neuesten Schätzungen liegt die derzeitige Aussterberate von 10 bis 150 Arten pro Tag um den Faktor 100 bis 1000 über der normalen Aussterberate stabiler Ökosysteme. Dies und der Klimawandel sind wohl die gravierendsten Merkmale einer neuen Epoche der Erdgeschichte, des Anthropozän. Die Entwicklung höherer Arten: Wirbeltiere: Die Wirbeltiere (Vertebrata) umfassen nahezu 58.000 rezente Arten, u.a. alle Knorpel-und Knochenfische, die Amphibien, Reptilien, Vögel und Säugetiere. Dies sind nach Schätzungen etwa ein Prozent aller jemals existierenden Arten von Wirbeltieren. Weltweit sind bisher mehrere zehntausend fossile Arten entdeckt worden /(Wikipedia). Ein wesentlicher Vorteil der Wirbeltiere ist, dass die innere Skelettstütze ein im Vergleich zu Lebensformen ohne Skelett oder zu Schalen-Tieren enormes Größenwachstum ermöglicht. Ferner werden Gehirn und Sinnesorgane der Wirbeltiere durch einen knöchernen Schädel geschützt. Bei den Insekten setzt der Außenpanzer neben dem Atmungssystem der Tracheen, einem Netzwerk kleiner Röhrchen und Kanäle mit punktuellen Öffnungen nach außen, dem Größenwachstum dagegen sehr früh eine Begrenzung. Diese Sackgasse der Evolution wird bei manchen Insektenarten durch eine Arbeitsteilung und Spezialisierung auf Ebene der Gesamtpopulation kompensiert. Die frühe Evolution der Wirbeltiere wird im obigen Abriss des Erdaltertums skizziert: Die ältesten Kiefermäuler, von denen fast alle Fische und alle Landwirbeltiere abstammen entwickelten sich vor ca. 450 Millionen Jahren. Der Auszug der Wirbeltiere aus dem Wasser begann vor ca. 400-360 Millionen Jahren. Neueren Erkenntnissen nach ist die Stammart aller Landwirbeltiere (Tetrapoden) ein ausgestorbener naher Verwandter der heutigen Lungenfische und Quastenflosser, bei der sich aus je zwei Paar Brust- und Bauchflossen bereits Knochen-verstärkte Bein-ähnlichen Gliedmaßen entwickelt hatten, mit denen sie sich auf dem Sumpfboden von flachen Süßgewässern bewegten. Ein Fossil mit solchen Merkmalen ist Tiktaalik (Fundort Ellesmere-Island, 380 MYA, Klasse: Fleischflosser). Aus den Landwirbeltieren entwickelten sich die heutigen Amphibien und – seit dem Karbon 360-300 MYA - parallel dazu die Gruppe der Amnioten, deren Embryonen sich in einem mit Fruchtwasser gefüllten Amnion entwickeln. Das Amnion schützt die Embryos vor dem Austrocknen und unterstützt den embryonalen Stoffwechsel. Bei rezenten eierlegenden Amnioten – alle Vögel, Krokodile, Schildkröten sowie die meisten anderen Reptilien - wird das Amnion von der innersten Membran der Eihaut gebildet. Die Ernährung des Embryos erfolgt durch den Eidotter. Die kalkige Eischale bietet zusätzlichen Schutz, auch gegen Austrocknung. Alle Säugetiere bis auf die Kloakentiere sind dagegen lebendgebärend, bei ihnen ist die Fruchtwasserblase das Amnion und der Embryo wird über den Stoffwechsel der Mutter versorgt. Es gibt auch einige lebendgebärenden Echsen- und Schlangenarten, sowie Zwischenformen, bei denen das Ei bis zum Schlüpfen der Jungen im Mutterleib verbleibt (z.B. Kreuzotter, viele Seeschlangen). Auch Amphibien, Fische und Insekten sind überwiegend eierlegend, den Eiern fehlt jedoch das Amnion, die Eiablage erfolgt in der Regel im Wasser. Auch hier gibt es lebendgebährende Arten und Zwischenformen. Die Amnioten spalteten sich etwa 300 MYA in zwei Evolutionslinien auf: 1) Synapsiden: aus ihnen entwickelten sich vor etwa 270 Millionen (im Perm) zunächst „säugerähnliche Reptilien“ (Therapsiden), aus letzteren dann über einen Zeitraum von ca. 80 Millionen Jahren allmählich die ersten Säugetiere. Alle Reptilien-artigen Vorfahren der Säugetiere sind ausgestorben. 2) Sauropsiden: aus diesen entwickelten alle heutigen Reptilien und Vögel. Dabei führen getrennte Entwicklungslinien zu Schildkröten, Echsen und Schlangen, ausgestorbenen Saurierarten (wie die Fischsaurier) und zu den sogenannten Archosauriern. Aus letzteren spalteten sich ab ca. 230 MYA die Evolutionslinien der Krokodile, der Flugsaurier und der Dinosaurier ab. Aus den Dinosaurier entwickelten sich ab einem Zeitraum vor circa 150- 200 Millionen Jahren die Vögel. Warmblüter: Warmblütige Organismen können ihre Körpertemperatur selbst regulieren. Bei warmblütigen Organismen wie Vögeln und Säugetieren wird die gleichmäßige Körpertemperatur durch die Stoffwechselaktivität der Tiere erzeugt. Dies äußert sich insbesondere in einem deutlich höheren Energieverbrauch (pro Körpergewicht) als bei sogenannten wechselwarmen Tieren. Warmblütige Tiere benötigen in der Regel daher eine besondere Wärmeisolierung in Form eines Fell- oder Federkleides. Warmblüter können auch bei Kälte schnell fliehen bzw. jagen. Heute lebende Reptilien sind wechselwarm (kaltblütig). Je nach Umgebungstemperatur erwärmt sich ihr Körper oder kühlt sich ab, was sich auf die Geschwindigkeit ihres Stoffwechsels und ihre Bewegungsfähigkeit auswirkt. Dasselbe wurde lange Zeit auch für die Echsen des Erdmittelalters angenommen. Einige Hinweise sprechen dafür, dass auch die Dinosaurier, flink wie sie waren, und als nahe Verwandte der warmblütigen Vögel, eine gleichmäßige, wenn auch vergleichsweise niedrige Körpertemperatur aufrecht erhalten konnten; das Gleiche wird für die Fischsaurier und die reptilienartigen Vorfahren der Säuger angenommen. Die Warmblütigkeit bei Vögeln und Säugern hat sich daher vermutlich aus einer einfachen Urform seit 300 Millionen getrennt weiter entwickelt. „Der Schlaf der Landtiere ist möglicherweise eine Erinnerung unserer Gene an 300 Millionen Jahre nächtlicher Kältestarre.“ /(HvD) Fische schlafen nicht, Wale, Delphine und auch bestimmte Vogelarten können abwechselnd mit jeweils einer Gehirnhälfte schlafen. Säugetiere: Zu ihren kennzeichnenden Merkmalen gehören das Säugen des Nachwuchses mit Milch, die in den Milchdrüsen der Weibchen produziert wird, sowie Warmblütigkeit und ein Fell aus Haaren. Die Säugetiere sind bis auf die Unterklasse der eierlegenden Ursäuger (Kloakentiere, z.B. Schnabeltier, Ameisenbär) lebend gebärend. Die beiden anderen Unterklassen sind die Beutelsäuger (z.B. Koala, Känguru) und die höheren Säugetiere (Plazenta-Tiere), die etwa 94% aller Säugetier-Arten umfassen. Von den Säuger-ähnlichen Reptilien des Perms überlebte die Gruppe der Cynodontia das große Massensterben am Ende des Perms. Diese meist Fleisch-fressenden Räuber mit Körperlängen bis 1 m waren vermutlich bereits warmblütig und behaart. Die Entwicklung zu ersten Säugern war ein graduell verlaufender Prozess, der vermutlich im späten Trias zu ersten Brückenformen, im frühen Jura dann zu ersten echten Säugern führte. Fossilien, die als mögliches „ältestes Säugetier“ diskutiert werden, sind bis zu 190 Millionen Jahre alt. Die meisten Forscher gehen von einem gemeinsamen Urahn aller Säugetiere aus. Juramaia sinensis (Spitzmaus-groß, 160 MYA, China) gilt als möglicher erster Vertreter der höheren Säugetiere; zweifelsfrei mit heutigen Arten verwandte Säugetiere traten erst nach Aussterben der Saurier zu Beginn der Erdneuzeit auf. Die rezenten „höheren Säuger“ lassen sich verhältnismäßig eindeutig in rund 20 Ordnungen einteilen; deren Verwandtschaftsgrad und Entwicklung über ausgestorbene Zwischenformen ist teilweise noch umstritten. Die Ordnung der Primaten hat sich vermutlich bereits vor 80-90 Millionen Jahren herausgebildet. Die ältesten zweifelsfrei den Primaten zuzuordnenden Fossilfunde (Archicebus achilles) sind etwa 55 Millionen Jahre alt und zeigen bereits die Aufspaltung in die Unterordnungen Feuchtnasenaffen und Trockennasenaffen; aus letzteren gingen vor18 Millionen Jahren die Familie der Menschenaffen (Hominidae) hervor. Diese Familie spaltete sich später auf in die Entwicklungslinien der Orang-Utans etwa 15 MYA, der Gorillas etwa 10 MYA, der Schimpansen und der Menschen, jeweils etwa 7 MYA. Vögel: Die Evolution der Vögel beginnt im Jura, vor 200-150 Millionen Jahren. Die Vorfahren der Vögel waren nach Ansicht der Mehrzahl der Forscher kleine Raubdinosaurier. Archäopteryx („Urvogel“) gilt als "Brückenform" zwischen den Reptilien und den Vögeln und als Vorfahre der heutigen Vögel. Er besaß Federn und beherrschte den Gleitflug. Die Länge des SchwanzSkeletts und der mit Zähnen besetzte Kiefer sind dagegen typische Reptilienmerkmale. Die heutigen Vogelgruppen mit ihren zahnlosen Kiefern haben sich seit Beginn der Erdneuzeit herausgebildet, erste Vertreter (nach Fossilienfunden in China) aber schon in der Kreidezeit 130 MYA. Nach dem großen Massenaussterben am Ende der Kreidezeit vor 66 Millionen Jahren stellen die Vögel zusammen mit den Krokodilen die einzigen überlebenden Archosaurier dar. Obwohl sich die Entwicklungslinien der heutigen Vögel und Krokodile bereits vor etwa 250 Millionen Jahren trennten, lässt sich die Verwandtschaft beider Gruppen aufgrund einer Reihe von Merkmalen (z.B. vollständige Trennung der Herzkammern) begründen. Die Stoffwechselrate der Vögel liegt noch höher als die der Säugetiere. Generell ist der Sauerstoffbedarf zur Nährstoffverbrennung bei kleineren Tieren wegen des relativ größeren Wärmeverlustes (größeres Verhältnis Oberfläche zu Volumen) relativ größer als bei großen. Dadurch dass Vögel einem starken Selektionsdruck zu Leichtigkeit ausgesetzt waren, entwickelten sich besonders effiziente Lungen. Die Luft fließt bei ihnen nicht nur bei der Einatmung in die Lunge, sondern sie fließt beim Ein- und Ausatmen durch die Lunge. Dadurch kann in der Lunge ein Gegenstrom von Luft und Blut realisiert werden, was die O2Aufnahme weit effektiver macht. „Ein Spatz ist in der Lage, den Himalaya zu überfliegen, während eine Maus dort oben ohnmächtig zusammen brechen würde.“ 8. Die Geschichte des Lebens im Zeitraffer Unser intuitives Zeitgefühl ist abgestimmt auf unsere Lebensspanne, die sich in Jahrzehnten bemisst. Auch zivilisationsgeschichtliche Zeiträume scheinen wir noch ganz gut zu überblicken, wir gliedern sie in Perioden, vor der Neuzeit das Mittelalter, das römische Reich, die Antike usw. Aber machen wir uns einmal klar, was allein die letzten 2000 Jahre vor unserer Zeit bedeuten? Etwa 120 Generationen liegen zwischen damals und uns, das sind viele tausende verschiedene Ahnen in unserer direkten Ahnenkette; Menschen, die mit uns verbunden sind, ohne die wir nicht da wären, wenn sie nicht gelebt und sich fortgepflanzt hätten. Was wissen wir von diesen Personen, ihren Schicksalen? Carl Sagan: „Der Widerstand gegen Darwin und Wallace geht zum teil auf unser Unvermögen zurück, uns das Verstreichen von Jahrtausenden vorzustellen, ganz zu schweigen von Äonen. Was bedeuten 70 Millionen Jahre für Wesen, deren Lebenszeit höchstens ein Millionstel davon beträgt? Wir sind wie Schmetterlinge, die einen Tag lang umher flattern und denken, es sei die Ewigkeit“ Wenn man die 13, 7 Milliarden Jahren der Geschichte des Universums auf 1 Jahr projiziert, entsprechen 433 Jahre Echtzeit einer Sekunde. Das Lebensalter eines Menschen entspricht ca. 0,2 Sekunden, nur ein Wimpernschlag, aber immerhin noch eine messbare Größe. Nach dieser verkürzten Zeitskala erfolgt der Urknall am 1 Januar 0.0h, das Universum wird durchsichtig als das Jahr gerade mal 14 Minuten alt ist. Irgendwann zwischen dem 5. und dem 13. Januar entstehen die ersten Masse-reichen Megasterne. In den Sternen entstehen durch Kernfusionen alle schweren Elemente bis zum Eisen, und aus den bereits nach einigen Stunden Brennzeit erfolgenden Supernova-Explosionen, die durch Gravitationskollaps dieser Mega-Sterne ausgelöst werden, noch schwereren Elemente. In den Tagen des 20. bis 23. Januar haben die ältesten heute bekannten Galaxien und Quasare ihr Licht ausgesandt. Dann geschieht lange Monate nichts weiter, als dass, angeheizt durch Zusammenstöße von Galaxien und Supernova-Explosionen, immer neue Generationen von Sternen entstehen, und sich auch immer mehr schwerere Elemente in den Gas- und Staubwolken der Galaxien ansammeln. Anfang September entsteht auf diese Weise die Sonne und mit ihr unser Planetensystem. Das erste Leben auf der Erde ist vermutlich am 19. September entstanden. In den folgenden 2 Monaten entwickelte sich das Leben in den Ur-Ozeanen zu allmählich immer komplexeren Mikroorganismen, darin die Urformen der Pflanzen, Pilze und Tiere. Am am 16. Dezember ereignet sich in die sogenannte kambrische Explosion, die ersten Pflanzen besiedeln das Land. Am 21. Dezember kriechen erste Amphibien an Land. Ungefähr zu Weihnachten, beim Übergang vom Perm zum Trias gab es auf der Erde das schlimmste von insgesamt 5 bekannten Massensterben, zwischen 70 und 90% aller Lebewesen starben zu dieser Zeit aus. Nun treten die Saurier ihren Siegeszug an, bis zu ihrem plötzlichen Aussterben am 28. Dezember beim Übergang von der Kreidezeit in die Erdneuzeit als Folge eines gewaltigen Meteoriteneinschlags. Die Dinosaurier machten Platz für eine rasche Evolution der Säugetiere. Durch Zusammenstoß von Kontinentalplatten falten sich nun auch die großen Gebirge der heutigen Erdoberfläche auf, der Himalaya und die Alpen. Am 31. Dezember, knapp 4 Stunden vor Mitternacht spaltet sich die Linie der Hominiden vom Entwicklungsast der übrigen Primaten ab. Der moderne Mensch, Homo sapiens, entwickelt sich erst etwa 6 Minuten vor Mitternacht, die großen Weltreligionen entstehen in den letzten 15 Sekunden des Kalenderjahres, die modernen Naturwissenschaften in der letzten Sekunde. 9. Leben - Zufall oder Notwendigkeit Alle Lebensformen auf der Erde sind miteinander verwandt, stammen von der gleichen Urzelle ab, die vor ca. 4 Milliarden Jahren auf der Erde entstand. Die Einheit und Durchgängigkeit des Lebens zeigt sich in den biochemischen Grundprozessen der Zellen (z.B. dem genetischen Code), welche allen Lebewesen gemeinsam sind. Wie ist es aber zu erklären, dass alles Leben auf der Erde von einer ersten Urzelle abstammt und dem Überleben dieser Urzelle abhing? Ist die Entstehung des Lebens ein so einmaliger sich nie wiederholender Vorgang? Ist es ferner allein durch Mutation und Selektion erklärbar, dass das Leben scheinbar zielgerichtet immer wieder so enorme qualitative Sprünge machte, wie mit der Entstehung der Urzelle selbst, der Erfindung von Photosynthese und Zellatmung, der Entwicklung höherer Lebewesen bis hin zur Entstehung von Intelligenz und Bewusstsein? Auf diese Fragen gibt es 3 grundsätzlich mögliche Antworten (Hoimar von Ditfurth): 1. Leben hat sich zwangsläufig in der heutigen Form entwickelt, weil es eine dem Leben innewohnende, naturgesetzlich nicht zu erklärende „schöpferische Kraft“ gibt, welche die Entstehung und Entwicklung des Lebens treibt und lenkt. 2. Leben ist gebunden an die Grundbausteine und Formen nach denen es auf der Erde entstanden ist. Jeder einzelne Entwicklungsschritt ist das Ergebnis des rein zufälligen Aufeinandertreffens der diesen Schritt jeweils ermöglichenden Faktoren. In diesem Fall wäre die Entstehung des Lebens so astronomisch unwahrscheinlich, dass die Erde mit nahezu absoluter Sicherheit der einzige belebte Himmelskörper wäre und der Mensch allein und einzigartig in dieser Welt (anthropozentrische Naturphilosophie) 3. Leben ist in vielen Formen denkbar (auch wenn uns andere Formen unbekannt und nicht vorstellbar sind). Auf der Erde entwickelte sich Leben auf Basis von Proteinen und Nukleinsäuren allein deshalb, weil dies die beiden einzigen Molekülarten von ausreichender Kompliziertheit und Wandlungsfähigkeit waren, die in genügender Menge zur Verfügung standen. Vitalismus versus Mechanismus: Die erste dieser möglichen Antworten postuliert eine naturgesetzlich nicht zu erklärende, Lebenskraft als eigenständiges Prinzip und damit einen Wesensunterschied zwischen organischer und anorganischer Materie. Dieses schöpferische Lebensprinzip, das man auch „Gott“ nennen kann, soll Grundlage alles Lebendigen sein. Diese Auffassung nennt man Vitalismus (lateinisch: vita = Leben). Dagegen steht der Mechanismus, der die Entstehung und Entwicklung des Lebens ausschließlich auf naturgesetzliche, chemische und physikalische Grundprinzipien, also auf erkennbare und messbare Eigenschaften der Materie zurückführt. Die Antworten 2 und 3 oben entsprechen einer mechanistischen Naturphilosophie. Sie unterscheiden sich nur in der gegensätzlichen Bewertung, ob die Entstehung von Leben auf Basis der geltenden Naturgesetze als astronomischer Zufall anzusehen ist, oder angesichts vieler Möglichkeiten und astronomisch vieler Versuche der Natur über unvorstellbar lange Zeiträume hinweg eher zwangsläufig erfolgte. Die Wurzeln dieser naturwissenschaftlich wie weltanschaulich bedeutsame Frage reichen bis in die griechische Philosophie zurück. Als ein Vorläufer des Vitalismus kann Aristoteles gelten, der ein eigenständiges Lebensprinzip postulierte, das er Entelechie nannte. Nach Platon dagegen ist die unzerstörbare Form (die Naturgesetze) das Primäre, der Stoff (die Materie) folgt der Form. Dies entspricht dem Standpunkt, dass alle Erscheinungsformen der Materie, also auch das Lebendige, aus den Naturgesetzen herleitbar sind. Die Frage der Urzeugung: Bis zum 19. Jahrhundert glaubte man allgemein auf Grund der Beobachtung, dass Maden oder Schimmelpilze scheinbar spontan auftraten, wenn organische Stoffe sich selbst überlassen werden, dass die spontane Entstehung von Leben aus toter Materie ein alltäglicher Prozess sei. Dies wurde im 19. Jahrhundert durch Louis Pasteur widerlegt, der zeigte, dass auch diese Organismen aus biologischen Vorläuferorganismen entstanden sind. Man hat auch bis heute nie beobachtet, dass Leben neu entstanden ist. Mit der Erkenntnis, dass alles Leben aus dem Leben kommt (lat: Omne vivum ex vivo), stellte sich gleichzeitig auch die Frage nach der Urzeugung des Lebens. Charles Darwin vermutete, dass das Leben in einem „warmen, kleinen Teich, in dem alle Arten von Ammoniak und Phosphor-Salzen, Licht, Hitze, Elektrizität usw. vorhanden waren“ begonnen haben könnte und dass „heutzutage so etwas sofort verschlungen oder absorbiert würde, was nicht der Fall war, bevor die lebenden Kreaturen entstanden.“ (Brief an Joseph Dalton Hooker, 1871) Heute herrscht in den Naturwissenschaften die Auffassung vor, dass erste Leben auf der Erde nicht spontan entstanden ist, sondern sich als Ergebnis einer Jahrmillionen Jahre andauernden chemischen, also naturgesetzlichen Evolution herausgebildet hat. Diese mechanistische Auffassung wird auch durch in-vitro Experimente gestützt, die zeigen, dass Teilprozesse von Lebensprozessen auch außerhalb des Organismus unter geeigneten Laborbedingungen ablaufen. 2002 gelang es erstmals, ein Poliovirus durch Reproduktion des genetischen Bauplans im Labor künstlich herzustellen. Kürzlich (SZ 21.5.2010) ist es erstmals gelungen ein Lebewesen „in vitro“ herzustellen, in dem man in eine Bakterienhülle einen künstlich synthetisierten DNA-Strang einbrachte. Das anthropische Prinzip: Das anthropische Prinzip (von griechisch anthropos» Mensch«), besagt, dass das Universum, das wir beobachten, für die Entwicklung intelligenten Lebens geeignet sein muss, da wir andernfalls nicht hier wären, es beobachten und physikalisch beschreiben könnten. An diese an sich triviale Feststellung kann man jedoch eine Reihe naturwissenschaftlicher und philosophischer Fragestellungen knüpfen: 1. Welche grundlegenden Eigenschaften des Universums und der Materie sind notwendige Voraussetzungen für die Entwicklung intelligenten Lebens (wie wir es kennen)? 2. Welche anderen Formen von Leben sind im Universum denkbar? 3. Wie wahrscheinlich ist die Entwicklung von intelligentem Leben im Universum? Diese Fragestellungen zielen über die Suche nach einer Erklärung für die Urzeugung des Lebens auf der Erde hinaus. Nach heutigem Stand der Wissenschaften erfordern adaptive und informationsverarbeitende Lebensprozesse die kontrollierte und doch schnelle Interaktion hochkomplexer Moleküle und damit zumindest eine hinreichende Stabilität des Lebensraums bei gemäßigten Temperaturen, möglicherweise auch Kohlenstoff und flüssiges Wasser. Augenscheinlich mussten schon für das Sonnensystem und den Planeten Erde eine Menge Dinge zusammen kommen, um die Entstehung von Leben auf der Erde zu ermöglichen, u.a. • Eine ausreichend lange stabile Brenndauer des Sternes; massereichere Sterne haben viel kürzere Lebensdauer, bei 10-facher Masse z.B. nur 10 Millionen Jahre. • Eine habitable Zone der Galaxis (z.B. ausreichender Abstand zu Supernovae-Explosionen) • Ein Planetensystem mit stabilen Planetenbahnen (daher auch kein Doppelstern-System). • Der richtige Abstand zur Sonne: er ermöglicht gemäßigte Temperaturen, die Entstehung der Ozeane, beständige komplexe Moleküle (5% näher oder 15% weiter entfernt wäre außerhalb Ökosphäre der Sonne) • Der richtiger Planet: die richtigen Elemente in den richtigen Mengen (z.B. Kohlenstoff, Stickstoff, Wasser); ein schützendes Magnetfeld (durch flüssigen Eisenkern); die Schaffung der Atmosphäre, von Meeren und Festlandgebieten durch Vulkanismus und Plattentektonik; eine Uratmosphäre ohne freien Sauerstoff usw. • Ein Verlauf der Evolution mit ausreichend langen Zeiten der Stabilität, unterbrochen von Phasen mit starkem Anpassungsdruck, sowie das Ausbleiben vernichtender Katastrophen. Ansätze, die Wahrscheinlichkeit weiterer (intelligenter) Lebensformen außerhalb der Erde zu berechnen, sind sehr umstritten um kommen je nach Annahmen zu sehr unterschiedlichen Ergebnissen von astronomisch unwahrscheinlich (rare-earth Hypothese) bis zu einigen 100 oder gar 1000 intelligenten Zivilisationen allein in unserer Galaxis (nach Modellen der DrakeGleichung). Zwar gibt es mindestens 100 Milliarden Sterne im Milchstraßensystem, und nach neueren Beobachtungen und Hochrechnungen bis zu 20 Milliarden Planeten, die von ihrer Masse und Umlaufdauer her eventuell als Lebenswelten in Frage kommen könnten, aber wie viele davon erfüllen die vielen Voraussetzungen, die zur Entstehung von Leben auf der Erde erfüllt sein mussten? Und wie viele extraterrestrische Lebensformen haben eine höhere, zu technischer Kommunikation befähigten Intelligenz entwickelt? Von den Milliarden von Arten, welche die Erde hervorgebracht hat, hat dies bisher nur der Mensch geschafft, und dies auch nur in den letzten 150 Jahren einer 4 Milliarden Jahre umfassenden Evolution. Feinabstimmung der Naturkonstanten: Schließlich bleibt die Frage nach den Naturgesetzen selbst. Offensichtlich sind sie so beschaffen, dass die kosmische Evolution ein Universum mit komplexen Formen der Materie in Form der vielfältigen Elemente und Moleküle sowie Sternen und Planeten und Leben hervorbringen konnte. Das ist nicht selbstverständlich. So werden die atomaren und molekularen Prozesse wesentlich durch die Werte von 4 Naturkonstanten bestimmt; dies sind die Massen von Proton und Elektron sowie die Stärken von elektromagnetischer und starker Kraft. Hätten diese Naturkonstanten nur geringfügig andere Werte, gäbe es keine stabilen molekularen Systeme oder keine Sterne im Universum. Man spricht in diesem Zusammenhang auch von der Feinabstimmung der Naturkonstanten. Andere Beispiele dafür sind die Asymmetrie der Entstehung von Materie und Anti-Materie beim Urknall ohne die keine Materie überlebt hätte, oder die Entstehung von Kohlenstoff (12C) als Basis für Leben durch Fusion von 3 Helium-Atomen (Drei-AlphaProzess) im Inneren von Sternen, welche nur durch eine erstaunliche naturgesetzliche Resonanz der Kernenergie-Niveaus der beteiligten Atomkerne zustande kommt. Verbirgt sich hinter dieser „Feinabstimmung“ ein noch unbekannter naturgesetzlicher Zusammenhang, gibt es – wie manche Wissenschaftler meinen - unendlich viele ParallelUniversen und darunter dann zwangsläufig auch solche, in denen sich komplexe Formen der Materie bis hin zu beobachtendem Leben bilden können, oder gibt es doch hinter allen Naturphänomenen eine innere Ziel- und Zweckgerichtetheit (teleologische Auffassung)? Die Unwahrscheinlichkeit des Seins: Jeder von uns ist mit der Urzelle durch eine ununterbrochene Kette von Lebensprozessen verbunden. Alle Lebewesen sind miteinander verwandt, wir selbst und jeder Grashalm, jeder Schmetterling haben irgendwo in der langen Lebenskette einen gemeinsamen Vorfahren. Die Anzahl unserer Vorfahren geht in die Milliarden. Jeder Mensch, aber auch jede biologische Art verdankt seine/ ihre Existenz dem unwahrscheinlichen Glücksfall, dass die Kette der zu ihm/ zu ihr führenden Einzelleben nie abriss. Unsere individuelle Existenz oder auch die spezielle Ausprägung unserer Art verdanken wir daher wohl einem astronomischen Zufall, der Fortgang des Lebens und die zunehmende Höherentwicklung und Diversifikation der Arten erfolgte dagegen wohl zwangsläufig nach den Gesetzen der Evolution. Der Begriff „Zufall“ an dieser Stelle soll bedeuten, dass für jedes zufällige Ereignis nur eine bestimmte Wahrscheinlichkeit des Eintretens gegeben ist. Es bleibt natürlich eine offene, philosophische Frage, ob uns dies nur subjektiv so erscheint, objektiv dagegen jedes Ereignis (und damit z.B. auch jede „Willensentscheidung“) kausal determiniert ist.