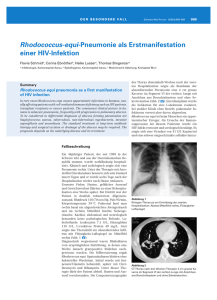
Prophylaxe der Rhodococcus-equi-Pneumonie bei Fohlen durch
Werbung

Aus der Klinik für Pferde der Tierärztlichen Hochschule Hannover Prophylaxe der Rhodococcus equi-Pneumonie bei Fohlen durch Vakzination mit Rhodococcus equi-Impfstoff und Adjuvans CpG XXXX INAUGURAL-DISSERTATION zur Erlangung des Grades eines Doktors der Veterinärmedizin (Dr. med. vet.) durch die Tierärztliche Hochschule Hannover Vorgelegt von Axel-Günther Hullmann aus Leer Hannover 2006 Wissenschaftliche Betreuung: Univ.- Prof. Dr. E. Klug 1. Gutachter: Univ.- Prof. Dr. E. Klug 2. Gutachter: Prof. Dr. M. Hoedemaker Tag der mündlichen Prüfung: 29.05.2006 Meinen Eltern Johann-Gerhard und Christa Hullmann gewidmet. Inhaltsverzeichnis 1 2 Einleitung ......................................................................................................... 11 Literaturübersicht............................................................................................ 13 2.1 Rhodococcus equi...................................................................................... 13 2.1.1 Taxonomie .......................................................................................... 13 2.1.2 Morphologie und Kultur ....................................................................... 14 2.1.3 Epidemiologie und Pathogenese......................................................... 14 2.1.4 Virulenzfaktoren .................................................................................. 16 2.1.5 Klinische Symptome............................................................................ 18 2.1.6 Pathologie ........................................................................................... 19 2.1.6.1 Makroskopisch ............................................................................ 19 2.1.6.2 Histologisch................................................................................. 20 2.1.7 Labordiagnostik................................................................................... 21 2.1.7.1 Hämatologie ................................................................................ 21 2.1.7.2 Mikrobiologie ............................................................................... 22 2.1.7.3 PCR ............................................................................................ 23 2.1.7.4 Nachweis von Antikörpern .......................................................... 24 2.1.8 Bildgebende Verfahren ....................................................................... 26 2.1.9 Therapie .............................................................................................. 27 2.1.10 Prophylaxe .......................................................................................... 28 2.1.10.1 Mutterschutzimpfungen............................................................... 29 2.1.10.2 Impfung der Fohlen ..................................................................... 29 2.1.10.3 Impfstudien ................................................................................. 30 2.1.10.4 Hyperimmunplasma .................................................................... 31 2.1.10.5 Sonstige Prophylaxemaßnahmen ............................................... 33 2.2 Das Immunsystem beim Fohlen im Zusammenhang mit R. equi ............... 33 2.2.1 Die humorale Abwehr.......................................................................... 34 2.2.2 Die zelluläre Immunität........................................................................ 36 2.2.2.1 Toll-like-Rezeptoren (TLRs) ........................................................ 37 2.2.2.2 Die zelluläre Abwehr im Zusammenhang mit R. equi.................. 37 2.3 Adjuvantien ................................................................................................ 40 2.3.1 CpG-Motive ......................................................................................... 41 2.3.1.1 CpG-Motive als Impfstoff-Adjuvantien......................................... 42 2.3.1.2 CpG-Motive in der Therapie ........................................................ 44 2.4 Die intranasale Applikation von Impfstoffen ............................................... 45 3 Material und Methode...................................................................................... 47 3.1 Probandengut............................................................................................. 47 3.1.1 Haltung................................................................................................ 47 3.1.2 Impfungen und Entwurmungen ........................................................... 48 3.1.3 Erstversorgung der Fohlen.................................................................. 48 3.1.4 Bedingungen für die Aufnahme in die Studie ...................................... 49 3.1.5 Einteilung der Probanden in Gruppen ................................................. 50 3.2 Rhodococcos equi-Impfstoff....................................................................... 50 3.3 Serumprobe und Impfung........................................................................... 51 3.4 Untersuchung der Fohlen........................................................................... 53 4 5 6 7 8 9 3.4.1 Klinische Untersuchung des Respirationstraktes ................................ 53 3.4.2 Labordiagnostische Untersuchung...................................................... 54 3.4.3 Weiterführende Untersuchung des Respirationstraktes ...................... 54 3.5 Der Rhodococcus equi-ELISA.................................................................... 57 3.5.1 Das Antigen......................................................................................... 57 3.5.2 Chargenprüfung und Antigenauswertung............................................ 57 3.5.3 Herstellung der Rhodococcus equi-ELISA-Testplatten ....................... 57 3.5.4 Durchführung des ELISA .................................................................... 58 3.5.5 Auswertung des ELISA ....................................................................... 59 3.6 Statistische Auswertung ............................................................................. 61 Ergebnisse ....................................................................................................... 63 4.1 Das Auftreten von Lungenabszessen......................................................... 63 4.1.1 Morbidität ............................................................................................ 63 4.1.2 Erkrankungsalter ................................................................................. 64 4.2 Klinische Symptome und Leukozytenkonzentration im Blut ....................... 68 4.3 Ultraschallbefunde...................................................................................... 70 4.4 Ergebnisse der ELISA-Untersuchungen..................................................... 72 4.4.1 Vergleich des Anti-R. equi-IgG-Gehalts aller Fohlen........................... 72 4.4.2 Vergleich zwischen erkrankten und gesunden Fohlen ........................ 74 4.4.3 Einfluss der maternalen Antikörper ..................................................... 76 Diskussion ....................................................................................................... 78 5.1 Diskussion von Material und Methode........................................................ 78 5.1.1 Probanden .......................................................................................... 78 5.1.2 Impfprotokoll........................................................................................ 79 5.1.3 Untersuchungen der Fohlen................................................................ 81 5.2 Ergebnisse ................................................................................................. 83 5.2.1 Morbidität und Befunde ....................................................................... 83 5.2.2 Untersuchung des Anti-R. equi-IgG-Gehalts im Fohlenserum ............ 87 5.3 Schlussfolgerungen.................................................................................... 91 Zusammenfassung.......................................................................................... 93 Summary .......................................................................................................... 95 Literaturverzeichnis ........................................................................................ 97 Anhang ........................................................................................................... 115 Abkürzungsverzeichnis Abb. Abbildung AGP Agargel-Präzipitation BALF bronchoalveoläre Lavage-Flüssigkeit BHV bovines Herpesvirus bzw. beziehungsweise °C Grad Celsius ca. Circa CAMPPhänomen nach den Entdeckern Christie-Atkins-Munch-Petersen Cluster of Differentiation (System zur Bezeichnung von CD Differenzierungsantigenen) cm Zentimeter CpG Cytosin-Phosphat-Guanosin CTL zytotoxische T-Zelle DNA Desoxyribonukleinsäure dl Deziliter EHV Equines Herpesvirus ELISA enzyme linked immunosorbent assay EU Elisa Units g Erdbeschleunigung (g = 9,81 m/s2) HIV humane immunodeficiency virus H-Ketten schwere Ketten i.e. id est Ig Immunglobulin IL Interleukin IFN Interferon kDa Kilodalton kg Kilogramm KGW Körpergewicht kV Kilovolt L-Ketten leichte Ketten LPS Lipopolysaccharid MALT mucosa-associated lymphoid tissue mAs Milliampère-Sekunde mg Milligram µg Mikrogramm MHC major histocompatibility complex MHz Megahertz ml Milliliter µl Mikroliter mm Millimeter µm Mikrometer MTP Mikrotiterplatte NK-Zellen natürliche Kilerzellen OD Optische Dichte O.I.E. Office International des Epizooties PAMP pathogen-associated molecular patterns PCR polymerase chain reaction p.o. per os PRR pattern recognition receptors R. equi Rhodococcus equi RNA Ribonucleinsäure SD Standardabweichung SDS-Page Sodiumdodecylsulfat-Polyacrylamidgel-Elektrophorese s.o. siehe oben ssp. Subspezies TBS Tracheobronchialsekret TBST-Puffer Tris buffered saline Tween®20-Puffer Th T-Helferzelle TLR Toll-like-Rezeptor TNF Tumornekrosefaktor Vap virulenz-assoziierte Proteine VLP Virus-like particels z.B. zum Beispiel Die chemischen Elemente entsprechend abgekürzt. werden dem internationalen Periodensystem Kapitel 1: Einleitung 1 Einleitung Eine Vielzahl von Krankheitserregern, die adulte Säugetiere ohne das Auftreten von klinischen Symptomen erfolgreich eliminieren können, lösen in juvenilen Individuen schwere Erkrankungen aus. Die Gründe für dieses Phänomen liegen nicht nur in den Fähigkeiten der Erreger dem Immunsystem zu entgehen, sondern auch in der unzureichenden Immunantwort der Jungtiere. In der Humanmedizin gilt als erwiesen, dass Neugeborene Immunsystem jedoch ebenso von immunkompetent einer Polarisierung sind in wie Richtung Erwachsene, der ihr humoralen Immunantwort bestimmt wird (KOVARIK u. SIEGRIST 1998). Rhodococcus equi ist ein Krankheitserreger, der intrazellulär überleben kann und bei Fohlen in den ersten vier bis sechs Lebensmonaten lebensbedrohliche pyogranulomatöse Lungenentzündungen hervorruft, während klinische Fälle bei adulten Pferden die Ausnahme sind. Da die Erkrankungen durch Rhodococcus equi eine kosten- und zeitintensive Therapie erfordern, die darüber hinaus erhebliche Nebenwirkungen hervorrufen kann, ist der Bedarf an einer Prophylaxe sehr groß. Bisher konnten jedoch weder die Vakzinierung der Mutterstuten noch eine aktive oder passive Immunisierung der Fohlen einen reproduzierbaren Schutz der Fohlen bewirken. Sowohl erwachsene als auch juvenile Pferde, die sich erfolgreich mit Rhodococcus equi auseinandergesetzt haben, weisen in der bronchoalveolären Lavage-Flüssigkeit einen signifikant höheren Gehalt an IFN-γ sezernierenden T-Lymphozyten auf als erkrankte Fohlen (HINES et al. 2003). Durch den Nachweis dieses Zytokins wird die Dominanz der zellulären Abwehr bei den nicht erkrankten Tieren und ihre entscheidende Rolle in der Bekämpfung von Rhodococcus equi deutlich. In der Human- und Veterinärmedizin liegen Erkenntnisse aus Untersuchungen in vivo und in vitro vor, dass Bestandteile bakterieller DNA, die Oligodeoxynukleotide, das Immunsystem so beeinflussen können, dass die zelluläre Komponente der Antwort gesteigert wird. Entscheidende Strukturelemente der Oligodeoxynukleotide sind Cytosin-Guanosin-Sequenzen, so genannte CpG-Motive (KRIEG 2002). Beim Pferd 11 Kapitel 1: Einleitung deuten bisher vor allem Studien an isolierten Leukozyten die Wirkung verschiedener CpG-Motive an (RANKIN et al. 2001, WATTRANG et al. 2005). In der Auseinandersetzung mit Erregern, die vor allem über die Schleimhäute aufgenommen werden, wie im Fall von Rhodococcus equi, verfügen Wirbeltiere mit dem schleimhautassoziierten Immunsystem über einen Schutzmechanismus, der zum Teil unabhängig vom restlichen Immunsystem arbeitet. Unter anderem werden bei wiederholter Auseinandersetzung mit einem Agens Gedächtniszellen gezielt an die betroffene Schleimhautregion gelenkt (JANEWAY u. TRAVERS 2002). Die intranasale Applikation eines Rhodococcus equi -Impfstoffs mit einem CpG-Motiv als Adjuvans wurde beim Fohlen in der Literatur bisher nicht beschrieben. In der vorliegenden Untersuchung sollen die Mechanismen des mukosalen Immunsystems durch eine intranasale Impfung genutzt und mit Hilfe des Adjuvans CpG XXXX besonders die zelluläre Komponente stimuliert werden, um eine schützende Immunität gegen Rhodococcus equi aufzubauen. 12 Kapitel 2: Literaturübersicht 2 Literaturübersicht 2.1 Rhodococcus equi 2.1.1 Taxonomie Rhodococcus equi (R. equi) wurde erstmals 1923 in Schweden von MAGNUSSON beschrieben. Da es sich um ein grampositives, unbewegliches pleomorphes nicht sporenbildendes Bakterium handelte, das sowohl zur Gelatineverflüssigung als auch zur Indolbildung führte, ordnete MAGNUSSON (1923) den Erreger der eitrigen Pneumonie des Pferdes der Gattung Corynebakterien zu und bezeichnete ihn als Corynebacterium equi. Darauf folgten weitere Beschreibungen des Keims durch MIESSNER und WETZEL (1923), BULL (1924) sowie von RAJAGOPALAN (1936). Über die Taxonomie des Erregers wurde anfänglich kontrovers diskutiert, wobei sowohl von JENSEN (1934) als auch von KRASIL`NIKOV (1966) aufgrund der Ähnlichkeit mit Mykobakterien die Bezeichnung Mycobacterium equi vorgeschlagen wurde. HOLTH und AMUNDSEN (1936) gelang es, das Bakterium aus tuberkulös veränderten Lymphknoten des Schweins als „spezifisches, säurefestes kokkobazilläres Bakterium“ nachzuweisen, so dass 1940 von PLUM die Bezeichnung Corynebacterium Magnusson-Holth vorgezogen wurde. GORDON (1966) befürwortete die Benennung Mycobakterium rhodococcus. Jedoch nach der Feststellung, dass das Bakterium N-glykolierte Muraminsäurereste enthält, was für Nokardien, Mykobakterien und Rhodokokken spezifisch ist, schlugen GOODFELLOW und ALDERSON (1977) vor, den Erreger den Rhodokokken zuzuordnen. Weitere Resultate von DNA- und RNA-Analysen unterstrichen den Unterschied zu Mykobakterien und Corynebakterien (SUZUKI et al. 1981), so dass die Bezeichnung Rhodococcus equi etabliert wurde. Der Name Rhodococcus leitet sich von den morphologischen Veränderungen während der Wachstumsphase ab, einem Zyklus aus filamentartigen und kokkoiden Formen „rod to coccus“ (PRESCOTT 1991). Aufgrund verschiedener Antigen-Strukturen auf der Polysaccharidkapsel werden 27 Serotypen unterschieden, die regional getrennt voneinander auftreten, aber nicht mit der Einteilung in virulente und avirulente Stämme übereinstimmen (NAKAZAWA et al. 1987). Schließlich wurde R. equi zusammen mit Mycobacterium ssp., Corynebacterium ssp. und weiteren Gattungen 13 Kapitel 2: Literaturübersicht in die phylogenetische Gruppe der nokardioformen Actinomyceten eingeordnet (PRESCOTT 1991). 2.1.2 Morphologie und Kultur Nach 24- bis 48-stündiger, aerober Bebrütung von R. equi auf festem, nicht selektivem Nährboden z.B. Blutagar zeigen sich unregelmäßige ein bis vier Millimeter große, runde Kolonien mit glattem Rand (PRESCOTT 1991). Die Kolonien sind feucht, glatt, glänzend, schleimig mit konfluierender Tendenz, lachsfarbener Pigmentierung und weisen einen erdigen Geruch auf (MAGNUSSON 1923, PRESCOTT 1991). Das Temperaturoptimum liegt bei etwa 30 °C (TAKAI et al. 1987). Charakteristisch für R. equi kann auf Schafsblutagar ein CAMP-ähnliches Phänomen (nach Staphylococcus den aureus Entdeckern beobachtet Christie-Atkins-Munch-Petersen) werden. Dabei kommt es durch mit das Zusammenwirken des Equi-Faktors von R. equi und dem Hämolysin von Staphylococcus aureus zu einer halbmondförmigen Zone vollständiger Hämolyse (SELBITZ 2002). Beim Equi-Faktor handelt es sich um ein Exoenzym mit membranolytischen Eigenschaften, das unter anderem aus der Cholesteroloxidase und der Phospholipase C besteht (PRESCOTT 1991). Um den kulturellen Nachweis von R. equi zu verbessern, hat sich der Einsatz von Selektivmedien, die sich neben den idealen Wachstumsbedingungen für R. equi vor allem durch die Hemmung unerwünschter Begleitflora auszeichnen, bewährt (WOOLCOCK et al. 1979, BARTON u. HUGHES 1981, MARTENS et al. 1982, TAKAI u. TSUBAKI 1985, VON GRAEVENITZ u. PÜNTER-STREIT 1995). Unter dem Mikroskop zeigt sich R. equi als grampositiver, pleomorpher, unbeweglicher Keim (SELBITZ 2002). 2.1.3 Epidemiologie und Pathogenese R. equi tritt weltweit auf und stellt vor allem für Fohlen im Alter von zwei Wochen bis sechs Monaten einen der wichtigsten Infektionserreger dar (BARTON u. HUGHES 1980, GIGUÈRE u. PRESCOTT 1997). In diesem Altersabschnitt werden auf betroffenen Gestüten eine Morbidität von 40 bis 80 % und die Mortalität von 5 bis 14 Kapitel 2: Literaturübersicht 17 % beobachtet (MARTENS et al. 1982). Fohlen im Alter unter zwei Wochen zeigen sich für eine experimentelle Infektion besonders anfällig (MARTENS et al. 1989). Sowohl die induzierte (MARTENS et al. 1982) als auch die natürliche Infektion führen zu Lungenläsionen vorwiegend im cranioventralen Bereich (MARTENS et al. 1982, BARTON u. EMBURY 1987). Die Manifestation in den genannten Lungenarealen und das gehäufte Auftreten bei staubiger Umgebung in den Sommermonaten (CHAFFIN et al. 2003) geben Hinweise dafür, dass die klassische R. equiPneumonie durch die aerogene Aufnahme des Erregers verursacht wird (BARTON u. EMBURY 1987, AINSWORTH 1999). Zu vernachlässigen ist der orale, intrauterine oder omphalogene Infektionsweg ebenso wie das Eindringen des Erregers durch Hautwunden oder durch wandernde Parasitenlarven (BARTON u. HUGHES 1980). Die orale Infektion bzw. das Abhusten und anschließende Herunterschlucken des Erregers führen in einigen Untersuchungen bei etwa 50 % der Fohlen, bei denen in der Sektion eine R. equi-Pneumonie diagnostiziert wurde, zu einer intestinalen Manifestation (ZINK et al. 1986, TAKAI et al. 1995). Für das Auftreten von makroskopisch sichtbaren Darmläsionen ist die Aufnahme von großen Keimmengen über längere Zeit Voraussetzung (ZINK et al. 1986). Bei der Beurteilung der Rennleistung eines Fohlenjahrgangs fiel auf, dass nur 54 % der nach einer R. equi-Pneumonie wieder genesenen Fohlen mindestens ein Rennen laufen, was sich im Vergleich zum Durchschnitt der gesamten Fohlenpopulation, von der pro Jahr 65 % ins Rennen gehen, signifikant unterscheidet (AINSWORTH et al. 1998). Die Fohlen jedoch, die ins Rennen gehen, zeigen die gleiche Leistung wie Fohlen, die nicht erkrankt waren. Im Gegensatz zu der hohen Prävalenz bei Fohlen werden Erkrankungen durch R. equi bei adulten Pferden nur in Einzelfällen beschrieben (PRESCOTT 1991). VENGUST et al. (2002) identifizierten R. equi in einer Trachealsekretprobe als Ursache bei einem an Pneumonie erkrankten Wallach und ZINK et al. (1986) beschreiben, dass eine Infektion mit R. equi nur sehr selten ursächlich für die Infertilität von Stuten verantwortlich ist. 15 Kapitel 2: Literaturübersicht Rhodococcus equi besitzt eine hohe Tenazität in der Umwelt: Er überlebt bei direkter Sonneneinstrahlung im Boden über 12 Monate (MAGNUSSON 1938) und ist unempfindlich gegenüber einigen Desinfektionsmitteln, wie z.B. Hypochlorsäure (MAGNUSSON 1938, BARTON u. HUGHES 1980). GOODFELLOW und MINNIKEN (1977) zählen Rhodokokken zu den obligat aeroben Saprophyten, die sich besonders gut in trockenen, sandigen Böden vermehren (BARTON u. HUGHES 1984). Im frisch rektal entnommenen Kot von Pferden endemisch betroffener Betriebe kann R. equi stets isoliert werden. Jedoch liegen die Keimzahl und die Isolierungsrate deutlich unter denen in am gleichen Ort genommenen Bodenproben (TAKAI u. TSUBAKI 1985). Daraus wird geschlossen, dass der Gastrointestinaltrakt nicht das natürliche Habitat von R. equi ist, sondern er in einem Zyklus zwischen den Tieren und dem Boden ihrer Umgebung lebt (TAKAI u. TSUBAKI 1985). Das Vorkommen von R. equi im Boden auch unabhängig von der Pferdehaltung unterstützt diese These (WOOLCOCK et al. 1980, BARTON u. HUGHES 1984). Der Keim kann im Kot von allen Herbivoren mit Weidegang nachgewiesen werden, wohingegen er im Hunde- und Katzenkot nur selten vorkommt (CARMAN u. HODGES 1987). Allerdings führt er bei anderen Tieren als Pferden nur vereinzelt zu den charakteristischen Erkrankungsformen an Lunge und Lymphknoten (CARMAN u. HODGES 1987). In der Humanmedizin tritt eine R. equi-Erkrankung ausschließlich bei immunsupprimierten Personen wie HIV-positiven Patienten oder während einer Glykokortikoid-Behandlung auf (TAKAI et al. 1994, ARLOTTI et al. 1996). Das typische Krankheitsbild wird wie in der Veterinärmedizin durch eine eitrige, abszedierende Pneumonie und selten durch intracerebrale, paraspinale und subcutane Abszesse geprägt. In der Anamnese wird häufig der Kontakt der erkrankten Person mit Pferden genannt (TAKAI et al. 1994). 2.1.4 Virulenzfaktoren Die unterschiedliche Virulenz der R. equi-Stämme beruht auf dem Vorhandensein so genannter Virulenzfaktoren, die von den pathogenen Stämmen getragen werden (TAKAI 1997). Zu ihnen zählen die Kapselpolysaccharide, die die phagozytierenden 16 Kapitel 2: Literaturübersicht Leukozyten hemmen, das Exoenzym Cholesteroloxidase, das Einfluss auf die Lysosomenmembran ausübt und Zellwandbestandteile, die im Zusammenhang mit der Granulombildung gesehen werden. (HONDALUS 1997). Auf der Bakterienoberfläche präsentierte Proteine, deren Genom extrachromosomal auf Plasmiden codiert wird, stellen weitere Virulenzfaktoren dar (HONDALUS 1997). R. equi-Stämme, die nicht aus erkrankten Tieren, sondern aus Bodenproben isoliert werden, enthalten selten diese Plasmide (SELLON et al. 2001). Auf einem 85 bis 90 kDa großen Plasmid liegen die Gene für eine Familie von sieben so genannten virulenz-assoziierten Proteinen (Vap): Vap A und C bis H (TAKAI et al. 2000). Die genaue Reaktionsweise der meisten 15-17 kDa großen Proteine ist noch nicht bekannt, aber einigen konnte durch Deletion der Gene eine Wirkung zugeordnet werden. Wird das VapA aus virulenten Bakterien entfernt, so verlieren sie ihre Fähigkeit, innerhalb von Makrophagen zu überleben oder Pneumonien in Fohlen auszulösen (HONDALUS u. MOSSER 1994, WADA et al. 1997, JAIN et al. 2003). VapA expremierende R. equi-Stämme zeigen in vitro eine bis zu 70 % stärkere zytotoxische Wirkung an Mäusemakrophagen als die Stämme, denen dieses Protein fehlt (LUEHRMANN et al. 2004). Mit Hilfe von Elektronenmikroskopie, Western Blotting und anderen Verfahren wird der nekrotische Charakter des Zelltods erkannt. Das Gen für VapB liegt auf einem weiteren Plasmid und wurde bei an R. equi erkrankten HIV-positiven Menschen isoliert. Dagegen wurde es bisher beim Fohlen kaum entdeckt und zeigt im Mäusemodell eine deutlich geringere Virulenz als VapA (LUEHRMANN et al. 2004). Die Expression der Virulenzproteine zeigte in vitro eine Abhängigkeit von den Inkubationsbedingungen: Nur bei Temperaturen zwischen 34 und 41 °C und nur in nährstoffarmen Medien können die Virulenzproteine mit Hilfe von SDS-Page ermittelt werden (TAKAI et al. 1991). Da die Bildung von VapA vor allem bei einem sauren pH-Wert und einer Temperatur von 38 °C nachzuweisen ist, vermuten TAKAI et al. (1995), dass die Virulenzfaktoren insbesondere in den Phagolysosomen exprimiert werden, in denen solche Bedingungen vorherrschen. 17 Kapitel 2: Literaturübersicht 2.1.5 Klinische Symptome Die klassische R. equi-Pneumonie der Fohlen verursacht die klinischen Symptome einer chronischen eitrigen, abszedierenden Bronchopneumonie (MARTENS et al. 1982). Da Fohlen in der Lage sind, den Verlust an funktionellem Lungengewebe sehr lange zu kompensieren (GIGUÈRE u. PRESCOTT 1997), treten zunächst unspezifische Symptome, wie erhöhte Körperinnentemperatur, Niedergeschlagenheit oder geringgradige Erhöhung der Atemfrequenz auf (MARTENS et al. 1982, FALCON et al. 1985, ALTHAUS 2004). Eine Tachypnoe kann solange unentdeckt bleiben bis die Fohlen durch Umgebungstemperatur, Zwangsmaßnahmen oder Bewegung belastet werden (AINSWORTH 1999). Das spätere Krankheitsbild wird durch deutlichere Anzeichen wie hohes Fieber bis 41,5 °C, hochgradige Tachypnoe mit dadurch bedingter verstärkter Bauchatmung (MARTENS et al. 1982), mittel- bis hochgradigen, produktiven Husten oder bilateralen, mukopurulenten Nasenausfluss geprägt (FALCON et al. 1985). Der progradiente Krankheitsverlauf führt zur Hyperventilation und zyanotischen Maulschleimhäuten (MARTENS et al. 1982, FALCON et al. 1985), wobei der Tod schließlich aufgrund von Asphyxie und kardialer Schwäche eintritt (HARAKAWA u. MORITA 1949). Das plötzliche Eintreten des Todes wird nur selten beschrieben, während das Sterben, begleitet von Husten, mukopurulentem Nasenausfluss und stark erhöhter Atemfrequenz, innerhalb weniger Tage trotz intensiver Betreuung und Therapie die Regel ist (MARTENS et al. 1982, BEECH u. SWEENEY 1991). Neben der chronischen R. equi-Pneumonie kann selten auch eine akute bis subakute Verlaufsform auftreten, die sich durch akute Atemnot und eine Körperinnentemperatur von über 40 °C und ein schnell eintretender Tod ohne vorangegangene respiratorische Symptome darstellt (MARTENS et al. 1982). Sowohl bei der chronischen als auch bei der akuten Verlaufsform sind die Befunde der Lungenauskultation nicht eindeutig: Sind bei gering- bis mittelgradigen Erkrankungen über den betroffenen Lungenarealen ein in- und expiratorisches Giemen und Knistern zu hören, werden in schweren Fällen nur tracheobronchiale Geräusche wahrgenommen, da die peripheren Lungenbereiche bereits verdichtet 18 Kapitel 2: Literaturübersicht sind (GIGUÈRE u. PRESCOTT 1997). Darüber hinaus führen hochgradige entzündliche Veränderungen der Lunge, insbesondere ausgedehnte periphere Abszesse, sogar zu einer vollständigen Dämpfung der Lungengeräusche (GIGUÈRE u. PRESCOTT 1997). Da die Befunde nicht unbedingt mit dem Schweregrad der Pneumonie korrelieren, bietet sich die Auskultation alleine für den Untersucher nicht als zuverlässiges Diagnostikum an, sondern muss mit anderen klinischen Befunden und weiterführenden Untersuchungen betrachtet werden (FALCON et al. 1985). Neben den pulmonalen Erscheinungsformen tritt vor allem Durchfall als Folge von ulcerativen Darmläsionen auf (BARTON u. HUGHES 1980, AINSWORTH 1999). An zweiter Stelle stehen durch Osteomyelitiden sowie durch septische und aseptische Arthritiden bedingte Bewegungsstörungen (AINSWORTH 1999). Aseptische Arthritiden werden durch Immunkomplexablagerungen verursacht, die auch zur Entwicklung von Uveitis, Hypopyon und Anämie führen können (KENNEY 1994, AINSWORTH 1999). Selten kommt es zur Bildung von Hautabszessen (AINSWORTH 1999) oder Serositiden wie Peritonitis, Pleuritis und/oder Perikarditis (ZINK et al. 1986). 2.1.6 Pathologie 2.1.6.1 Makroskopisch Das typische Sektionsbild einer mit R. equi infizierten Lunge wird geprägt von einer subakuten bis chronischen eitrigen Bronchopneumonie mit Abszessbildung und assoziierter suppurativer Lymphadenitis (ELLENBERGER u. GENETZKY 1986, AINSWORTH 1999). Multifokale Abszesse miliarer Größe bis hin zu sieben Zentimeter Durchmesser befinden sich vor allem im ventralen Bereich (MARTENS et al. 1982, FALCON et al. 1985, AINSWORTH 1999), während sie in caudalen Lungenlappen nur vereinzelt anzutreffen sind (ZINK et al. 1986). In etwa 20 % der Fälle sind die Abszesse generalisiert über die Lunge verstreut. Zu einem gleichen Anteil wird der pathologische Befund von einzelnen Abszessen bestimmt (ZINK et al. 1986). Der Abszessinhalt weist eine zähflüssige bis käsige Konsistenz, eine graurote bis gelbliche Farbe und einen unangenehmen Geruch auf (AINSWORTH 1999). Das die Abszesse umgebende Lungengewebe ist verdichtet oder eingeschmolzen 19 Kapitel 2: Literaturübersicht (BARTON u. HUGHES 1980), während die Pleura nur selten in das Entzündungsgeschehen einbezogen wird (ELLENBERGER u. GENETZKY 1986). An der Schnittfläche der betroffenen Lungenbereiche tritt mukopurulentes Sekret aus den Bronchien aus (BARTON u. HUGHES 1980). Im fortgeschrittenen Verlauf der Infektion findet sich häufig eine Lymphadenitis im Hals- und Thoraxbereich, wobei die Lymphknoten durch eine bis zu achtfache Größenzunahme und eine ödematöse oder sogar nekrotische Schnittfläche auffallen (SMITH u. JANG 1980, ZINK et al. 1986). Bei experimenteller Infektion wurde festgestellt, dass die Lungenschäden mit der Infektionsdosis korrelieren (WADA et al. 1997). In etwa der Hälfte der Sektionsfälle werden multifokale ulzerative Enteritiden in absteigender Häufigkeit im Colon, Caecum und Dünndarm beobachtet. In dreiviertel der Fälle sind die zugehörigen Mesenterial- und Darmlymphknoten vergrößert und zeigen in Einzelfällen eine abszedierendene Entzündung (ZINK et al. 1986). Etwa ein Drittel der Sektionstiere zeigen darüber hinaus entzündliche Veränderungen der Gelenke. Bei einer seltenen systemischen Verbreitung von R. equi können Abszesse in Leber, Nieren und Haut (MARTENS et al. 1982, AINSWORTH 1999) oder Serositiden gefunden werden (ZINK et al. 1986). 2.1.6.2 Histologisch Histologisch weisen die von R. equi ausgelösten Pneumonien primär einen pyogranulomatösen Charakter auf (ELLENBERGER u. GENETZKY 1986). Das Zentrum der Abszesse enthält nekrotisches Zellmaterial und ist umgeben von zugrunde gehenden neutrophilen Granulozyten, während die Peripherie massenhaft von Makrophagen infiltriert ist. Selten sind einzelnen Riesenzellen und Lymphozyten zu finden. Das Abszess umgebende Lungenparenchym ist hyperämisch und ödematös. Sowohl im nekrotischen Abszessinhalt, in den Alveolen als auch im Zytoplasma der Makrophagen und neutrophilen Granulozyten sind unterschiedlich viele grampositive Stäbchenbakterien zu erkennen (HILLIDGE 1986). Im Verlauf der Erkrankung prägen zunehmend nekrotische Makrophagen das histologische Bild (LUEHRMANN et al. 2004). Ferner 20 werden histologisch neben der Kapitel 2: Literaturübersicht pyogranulomatösen Pneumonie eine akute purulente Bronchitis und Peribronchitis diagnostiziert: In den Alveolen sind eingewanderte Makrophagen und neutrophile Granulozyten und in den Bronchien Fibrin- und selten Eiterablagerungen zu erkennen (ELLENBERGER u. GENETZKY 1986). 2.1.7 Labordiagnostik Der Einsatz von Antibiotika erfordert immer eine exakte Diagnose basierend auf einer klinischen und erforderlichenfalls weiterführenden labordiagnostischen Untersuchungen (UNGEMACH et al. 2000). Im Fall von R. equi ist eine unterstützende Labordiagnostik besonders wichtig, weil die klinischen Symptome erst in Erscheinung treten, wenn die Schädigung der Lunge weit fortgeschritten ist (AINSWORTH 1999). An jedes Testsystem zum Nachweis einer Infektion mit R. equi wird die Forderung sowohl einer hohen Sensitivität (i.e. der Anteil der Falschnegativen ist möglichst gering) als auch einer hohen Spezifität (i.e. der Anteil der Falschpositiven ist möglichst gering) gestellt, um entweder durch rechtzeitiges Erkennen die Prognose zu verbessern oder zu hohe Kosten und unnötige Nebenwirkungen der Therapie zu vermeiden (COHEN et al. 2000, MARTENS et al. 2002). 2.1.7.1 Die Hämatologie Bestimmung der Blutleukozytenkonzentration als unspezifischer Entzündungsparameter ist auf Gestüten mit endemischer Rhodokokkose im Rahmen eines Überwachungsprogrammes bei den jungen Fohlen hilfreich. Da zu Beginn einer Infektion mit R. equi nahezu alle Fohlen eine Leukozytose entwickeln (GIGUÈRE et al. 2003b, CHAFFIN et al. 2004), sollten ab einer Konzentration von 13.000 Leukozyten/μl Blut weitergehende spezifische Untersuchungen wie Sonographie oder Radiologie durchgeführt werden (GIGUÈRE et al. 2003b). Zu erwähnen ist jedoch, dass in einer Studie, die 66 an abszedierender Pneumonie erkrankte Fohlen umfasste, bei 75 % der Tiere, die bei der Ultraschalluntersuchung Lungenabszesse aufwiesen, die Konzentration der Leukozyten unter 13.000/μl Blut lag (ALTHAUS 2004). 21 Kapitel 2: Literaturübersicht In der Früherkennung einer R. equi-Pneumonie an 165 Fohlen war die Bestimmung der Leukozytenzahl gegenüber einer Messung der Fibrinogenkonzentration von signifikant höherem diagnostischen Nutzen, wobei als Referenz der positive kulturelle Nachweis von R. equi aus der trachealen Lavage diente (GIGUÈRE et al. 2003b). Die Bestimmung der Leukozytenzahl wies bei einem Grenzwert von 13.000 Zellen pro μl Blut eine Sensitivität von 95,2 % und eine Spezifität von 61,2 %, bei einem Grenzwert von 15.000 Zellen pro μl Blut eine Sensitivität von 78,6 % und eine Spezifität von 90,8 % auf. Demgegenüber zeigte die Messung der Fibrinogenkonzentration bei einem Grenzwert von 400 mg/dl eine Sensitivität von 91 % und eine Spezifität von 51 %, bei einem Grenzwert von 500 mg/dl eine Sensitivität von 63 % und eine Spezifität von 68 % im Vergleich zum kulturellen Nachweis von R. equi. 2.1.7.2 Mikrobiologie Das für R. equi spezifische kulturelle Wachstumsverhalten lässt eine sichere Identifizierung des Erregers zu. Am lebenden Tier kann der Erreger aus dem Nasentupfer oder dem Tracheobronchialsekret (TBS) isoliert werden. Auf einem Gestüt mit endemischer Rhodokokkose gelang nur bei 52 von 217 Fohlen (24 %), die bei der Ultraschalluntersuchung Lungenabszesse oder Pneumonien aufwiesen, die Isolierung des Erregers aus dem Nasentupfer (MEYER-HAMME 2004). Dagegen konnte R. equi aus 120 derselben 217 Proben (54 %) des TBS nachgewiesen werden. Diese Quote konnte auf demselben Gestüt ein Jahr später bestätigt werden (HEYERS 2005). MARTENS et al. (1982) schätzten ebenfalls den Erregernachweis aus dem TBS aufgrund nicht konstant positiver Ergebnisse bei erkrankten Fohlen als unsicher ein. Kulturell negative Befunde werden von SELLON et al. (2001) kritisch diskutiert, da obligat intrazelluläre Keime schwer anzuzüchten sind und das Wachstum von R. equi außerdem durch eine schneller wachsende Begleitflora gehemmt werden kann. In einer weiteren Studie allerdings konnte der Erreger neun Tage nach experimenteller Infektion durch Inhalation von R. equi aus dem TBS aller Fohlen kulturell nachgewiesen werden (ANZAI et al. 1997). GIGUÈRE und PRESCOTT (1997) berichten über die Möglichkeiten einer falschen Diagnose, wenn R. equi über kontaminierten Staub eingeatmet und im TBS nachgewiesen wird, aber 22 Kapitel 2: Literaturübersicht keine Manifestation in der Lunge stattfindet. Ein Nachweis des Erregers ohne Zusammenhang mit einer Erkrankung wurde auch von HEYERS (2005) beschrieben. R. equi wurde bei 30 % der 40 Fohlen aus dem TBS isoliert, bei denen weder durch die klinische noch die sonographische Untersuchung pathologische respiratorische Befunde erhoben wurden. Zu bedenken bleibt allerdings, dass aufgrund der physikalischen Gegebenheiten die sonographische Untersuchung Abszesse, die medial von belüfteten Lungenbereichen liegen, nicht darstellen kann (COHEN et al. 2000). Zuverlässig ist dagegen der kulturelle Nachweis von R. equi aus Sektionsmaterial wie Lymphknoten oder Lungenabszessen (MARTENS et al. 1982). 2.1.7.3 PCR Die Polymerase-Kettenreaktion (englisch Polymerase Chain Reaction, PCR) ist gegenüber der Kultur schneller und ermöglicht auch den Nachweis von abgetöteten Erregern oder ihren Bestandteilen (PERSING et al. 1993). Für den Nachweis von R. equi sind verschiedene PCR-Methoden entwickelt worden. BELL et al. (1996) konzentrierten sich auf eine in allen R. equi-Stämmen vorkommende Gensequenz, die keine Differenzierung zwischen virulenten und avirulenten Stämmen ermöglichte. Um die Stämme unterscheiden zu können, wurde eine PCR-Methode entwickelt, mit der das Gen amplifiziert wird, das für das Virulenzassoziierte Protein A (VapA) codiert (TAKAI et al. 1998). Mit diesem Verfahren gelang es OLDFIELD et al. (2004) in allen von erkrankten Pferden stammenden R. equi-Proben, das für VapA codierende Genom zu isolieren. Dagegen wurde mit der VapA-PCR bei experimentell infizierten Fohlen in nur 51 % der Proben aus dem Tracheobronchialsekret (TBS) R. equi nachgewiesen, während dieselben Proben in der Kultur zu 100 % einen positiven Erregerbefund ergaben (ANZAI et al. 1997). In einer Studie von HEYERS (2005) wies die Kultur mit 52 % ebenfalls eine höhere Sensitivität im Vergleich zu zwei PCR-Varianten (AceA500- und IdeR500-PCR) mit 40 % bzw. 33 % auf. Hinsichtlich der Spezifität zeigten sich beide PCR-Varianten mit je 83 % der Kultur mit 70 % überlegen. Als Erklärung für das insgesamt zu niedrige Niveau der Sensitivität vermutet der Autor das Vorhandensein von Inhibitoren im TBS und die zum Teil sehr hohe Viskosität des Sekrets, welche die Präparation von 23 Kapitel 2: Literaturübersicht bakterieller DNA beeinträchtigen kann. Weitere Entwicklungen im Bereich der PCR beruhen auf der Identifizierung der Gene anderer Virulenzfaktoren wie zum Beispiel die PCR für das Gen der Cholesteroloxidase (LADRON et al. 2003). 2.1.7.4 Nachweis von Antikörpern Der indirekte Erregernachweis beruht auf der Bindung von Antikörpern an spezifischem Antigen, so dass es möglich ist mit einem bekannten Reaktionspartner das Vorhandensein des jeweils anderen in einer Probe nachzuweisen. Durch die Etablierung von quantitativen Testverfahren und die Erstellung von Referenzwerten können Aussagen sowohl über die Immunkompetenz als auch über eine aktuelle Krankheitssituation eines Patienten getroffen werden. Die für die Diagnose von R. equi entwickelten Antikörpernachweise beruhen auf der Agargel-Präzipitation (AGP), der Hämolysehemmungsreaktion und vor allem auf dem enzyme linked immunosorbent assay (ELISA). Bei der AGP werden Antigen und Antikörper in löslicher Form auf eine Agar-Gel-Platte gegeben. Im Kontaktbereich entstehen durch die Antigen-Antikörper-Komplexe so genannte Präzipitationslinien. Anhand dieser Linien kann eine semiquantitative Aussage über das Vorliegen von Antikörpern gemacht werden. Der Antikörpernachweis durch die Hämolysehemmungsreaktion beruht auf der Hemmung der Exoenzyme des Equi-Faktors durch die gesuchten Antikörper. Auf der Grundlage der ELISA-Technik fanden die meisten Entwicklungen statt, dem folgendes Prinzip zu Grunde liegt: R. equi-spezifisches Antigen wird auf der Oberfläche einer Mikrotiterplatte (MTP) fixiert und dient als Fänger-Antigen. Sind im Serum entsprechende Antikörper vorhanden, kommt es zur Bildung von AntigenAntikörper-Komplexen. Nach einer Inkubationszeit werden nicht gebundene Antikörper von der Platte gewaschen und enzymmarkierte Sekundärantikörper, die gegen equines IgG gerichtet sind, aufgetragen. Wiederum werden nicht gebundene Antikörper von der MTP gespült. Das an die Sekundärantikörper gekoppelte Enzym setzt ein hinzugefügtes, farbloses Substrat in ein farbiges Produkt um. Die Intensität der entstandenen Färbung kann anschließend photometrisch als Extinktion gemessen werden. Durch den Vergleich mit einem Kontrollserum erfolgt die Berechnung der Konzentration in der zu untersuchenden Probe. Die verschiedenen 24 Kapitel 2: Literaturübersicht ELISA-Methoden, die im Zusammenhang mit R. equi etabliert wurden, unterscheiden sich vor allem in der Art und in der Präparation des Fängerantigens. Für den ersten veröffentlichten ELISA wurde das Rhodococcus equi-Antigen aus einem erkrankten Fohlen isoliert (ELLENBERGER et al. 1984). Zunächst wurde die Probe mit einer Pufferlösung verdünnt, anschließend autoklaviert und zentrifugiert, so dass das Antigen aus dem Überstand gewonnen werden konnte. Ein Jahr später folgten der so genannte „California-ELISA“ von HIETALA et al. (1985) und der ATCC-6939-ELISA von TAKAI et al. (1985), die beide auf einer vergleichbaren Antigenpräparation basieren. HIETALA et al. (1985) erhielten das von ihnen verwendete Antigen aus R. equi-Kulturen, die auf Mueller-Hinton-Agar angezüchtet wurden. Mit Phosphatpuffer wurden die Kulturen vom Nährboden gelöst, und das Antigen nach Zentrifugation aus dem Überstand gewonnen. Das Antigen für den ATCC-6939-ELISA wurde durch Tween 20® aus dem apathogenen Stamm ATCC 6939 entwickelt, dem er seinen Namen verdankt (TAKAI et al. 1985). Dieser Stamm weist eine hohe Kreuzreaktivität mit anderen R. equi-Stämmen und damit eine hohe Sensitivität auf. Für den Einsatz des ELISAs in der Diagnostik geben TAKAI et al. (1985) an, dass der Nachweis positiv ist, wenn die optische Dichte (OD) den Wert 0,3 überschreitet. In Kombination mit einer klinischen Überwachung beurteilten HIGUCHI et al. (1997a) den ATCC-6939-ELISA als geeignet für die Früherkennung einer R. equi-Infektion. Sie stellen darüber hinaus zur Diskussion, auf Gestüten mit endemischer Rhodokokkose bei allen Fohlen im Alter von 35 bis 65 Tagen generell den Antikörpergehalt zu untersuchen, da einige Fohlen mit einer OD über 0,9 in der Sektion eine weit fortgeschrittene Erkrankung mit R. equi in der Lunge und im Intestinaltrakt zeigten. In einer weiteren Studie, in der bei klinisch auffälligen Fohlen sowohl ein kultureller Erregernachweis aus dem Tracheobronchialsekret als auch eine Antikörpermessung im Serum durchgeführt wurde, bestätigten HIGUCHI et al. (1997b) die hohe Sensitivität des ELISAs, da von den 21 Fohlen mit positivem kulturellem Erregernachweis nur eins mit einer OD unter 0,3 auffiel. Allerdings wies der ATCC-6939-ELISA eine mäßige Spezifität auf, da unter den zehn kulturell negativen Tieren drei falsch positive mit einer OD über 0,3 waren. TAKAI et al. (1996) etablierten ein ELISA-Verfahren, das in der Präparation des Antigens mit dem 25 Kapitel 2: Literaturübersicht ATCC-6939-ELISA übereinstimmt, aber das Antigen eines virulenten Stammes enthält und nach diesem ATCC 33701-ELISA genannt wird. Um den Antikörpernachweis weiter zu verfeinern, präparierten PRESCOTT et al. (1996) das in allen virulenten Stämmen enthaltene virulenzassoziierte Protein A (VapA) als Fängerantigen. Die Möglichkeiten der ELISA-Techniken in der Diagnostik werden in der Literatur kritisch beurteilt. Schon bei der klinischen Erprobung der ersten ELISA-Methoden wurde festgestellt, dass es keine Korrelation zwischen dem Antikörpergehalt und dem Schweregrad der Erkrankung gibt (ELLENBERGER et al. 1984, HIETALA et al. 1985). Ähnlich kritisch fällt das Ergebnis einer Studie aus, in der ein Vergleich des ATCC-6939-ELISAs nach TAKAI et al. (1985), des ATCC-33701-ELISAs nach TAKAI et al. (1996), des VapA-ELISA nach PRESCOTT et al. (1996), der AgargelPräzipitation und der Hämolysehemmungsreaktion durchgeführt wurde (MARTENS et al. 2002). Die Fragestellungen waren, ob es möglich sei aus dem Ergebnis einer einzigen Serumprobe bereits vor dem Auftreten von klinischen Erscheinungen eine Erkrankung mit R. equi zu diagnostizieren. Weiter sollte ermittelt werden, ob die Genauigkeit der ersten Frage durch eine gepaarte Serumprobe zu verbessern sei, und schließlich, ob es serologisch möglich sei, infizierte von gesunden Fohlen zu unterscheiden. Die Ergebnisse zeigten, dass kein Verfahren in der Lage war, auch nur eine der Anforderungen befriedigend zu erfüllen. In einem weiteren Vergleich der etablierten ELISA-Techniken und einer Agargel-Präzipitation lieferte der „CaliforniaELISA“ bei einem Grenzwert von 40 % die besten Ergebnisse hinsichtlich Sensitivität (59 %) und Spezifität (88 %) (GIGUÈRE et al. 2003a). Die Schlussfolgerungen fielen jedoch ebenfalls negativ aus, da keine serologische Methode in der Lage war, gesunde von tatsächlich erkrankten Fohlen zu unterscheiden und daher ein Einsatz als alleiniges Diagnostikum ausgeschlossen wird. 2.1.8 Bildgebende Verfahren Um die Diagnose einer R. equi-Pneumonie weiter abzusichern, stehen als bildgebende Verfahren die Röntgen- und die sonographische Untersuchung zur Verfügung. Beim Fohlen sind für die transkutane sonographische Untersuchung der 26 Kapitel 2: Literaturübersicht Lunge sowohl Sektor- als auch Linearscanner mit einer Frequenz von 5 bis 7,5 MHz geeignet (O´BRIEN u. BILLER 1997). Besonders Veränderungen im peripheren Bereich der Lunge können mittels Sonographie dargestellt werden (COHEN et al. 2000, ALTHAUS 2004), wobei der Sonographie eine höhere Sensitivität als der röntgenologischen Untersuchung in diesen Bereichen zugesprochen wird (COHEN et al. 2000). Jedoch ermöglicht die Sonographie weder eine Darstellung der Befunde, die medial von belüftetem Lungengewebe liegen, noch lässt sie eine exakte Aussage über die Ausdehnung von Lungenveränderungen in die Tiefe zu (GIGUÈRE u. PRESCOTT 1997). Um solche pathologischen Veränderungen bildlich darzustellen, ist die radiologische Untersuchung der Lunge erforderlich, für deren Durchführung bei Fohlen im Alter von einem Monat eine Einstellung von 70 bis 74 kV und 0,14 mAs bei einer Film/Folien-Kombination mit einer Empfindlichkeit von 400 empfohlen wird (COHEN et al. 2000). Bei Fohlen älter als drei Monate können im Röntgenbild darstellbare Lungenabszesse nicht nur von R. equi hervorgerufen werden, sondern unter anderem auch von Streptococcus equi ssp. zooepidemicus (LAVOIE et al. 1994, RAMIREZ et al. 2004). Dagegen sind solche Befunde bei sehr jungen Fohlen pathognomonisch für eine Infektion mit R. equi (HILLIDGE 1986). Darüber hinaus sehen FALCON et al. (1985) im Röntgenbild darstellbare schwerwiegende noduläre und kavernöse Läsionen als Kennzeichen für ein weit fortgeschrittenes Stadium der Erkrankung, da sie mit einer letalen Entwicklung häufiger einhergehen als mit einer Genesung der Fohlen. Ebenso erkannten AINSWORTH et al. (1998), ausgehend von einem selbst erstellten Score von eins bis vier, dass Fohlen ab einem Röntgen-Score von drei oder höher signifikant geringere Überlebensraten aufweisen. In einer retrospektiven Studie über die Untersuchung von 17 an R. equi-Pneumonie erkrankten Fohlen bezeichnen RAMIREZ et al. (2004) nach Abwägung der Vor- und Nachteile die Sonographie als eine angemessene Alternative zur Röntgenuntersuchung. 2.1.9 Therapie Das Hauptaugenmerk in der Therapie einer Erkrankung mit R. equi liegt, wie bei jedem bakteriellen Erreger, auf der Bekämpfung der Ursache durch eine geeignete Antibiotikagabe. Die Eigenschaften von R. equii, sowohl innerhalb der Makrophagen 27 Kapitel 2: Literaturübersicht zu überleben als auch granulomatöse Veränderungen mit verkäsendem Inhalt im Lungengewebe hervorzurufen, schränken die Auswahl der Chemotherapeutika erheblich ein (HONDALUS u. MOSSER 1994). Als wirkungsvollste Therapie gegen R. equi wurde die Kombination von dem aus der Ansamycingruppe stammenden Rifampicin mit dem zu den Makrolidantibiotika gehörenden Erythromycin entdeckt (SWEENEY et al. 1987). Mit einer Dosierung von 25 mg/kg KGW p.o. Erythromycin dreimal täglich und 5 mg/kg KGW p.o. Rifampicin zweimal täglich konnte ein Therapieerfolg von 88 % erreicht werden (HILLIDGE 1987). Die Nachteile dieses Therapieprotokolls liegen in den hohen Kosten und den Nebenwirkungen, die bei den Fohlen als Diarrhöe und Hyperthermie und bei den Stuten als schwere Kolitis auftreten können (GIGUÈRE u. PRESCOTT 1997). Mit Azithromycin wurde ein weiteres Antibiotikum für die Rhodokokkentherapie entdeckt (JACKS et al. 2001), das sich alleine eingesetzt als ebenso wirksam wie die Kombination aus Erythromycin und Rifampicin zeigte (PILTZ 2004). Der Einsatz von Tulathromycin, ein weiteres Makrolidantibiotikum, befindet sich in der Erprobungsphase (KERTH 2005, HÖHENSTEIGER 2005). Über die Antibiotikagabe hinaus verbessert die Verwendung von nichtsteroidalen Antiphlogistika das Allgemeinbefinden eines erkrankten Fohlens (AINSWORTH 1999). Zur Verringerung des Atemwiderstandes und zur Erhöhung der mukozilliären Clearence können Bronchodilatatoren eingesetzt werden (COHEN 2000). Sollte sich der Allgemeinzustand drastisch verschlechtern oder sich eine Atemnot einstellen, wird eine Sauerstoffinsufflation notwendig (5 l/min) (COHEN 2000). Um geeignete Umgebungsverhältnisse zu schaffen, sollten die Fohlen in einen gut belüfteten und temperierten Stall gebracht werden. Daneben sollte sowohl auf Pflege als auch auf ausreichende Nahrungs- und Flüssigkeitsaufnahme geachtet werden (GIGUÈRE u. PRESCOTT 1997). 2.1.10 Prophylaxe Da R. equi ubiquitär vorkommt und die von ihm ausgelösten Erkrankungen eine zeitund kostenintensive Therapie erfordern, die Nebenwirkungen induziert, ist die Entwicklung einer Prophylaxe von großem Interesse. Es werden insbesondere Strategien verfolgt, um eine Immunität in den Fohlen aufzubauen, die entweder auf 28 Kapitel 2: Literaturübersicht der Übertragung schützender Antikörper an die Fohlen oder auf einer aktiven Immunisierung der Fohlen selbst beruhen. 2.1.10.1 Mutterschutzimpfungen Die Vakzinierung der Mutterstuten, um so die Fohlen vor einer Erkrankung durch R. equi zu schützen, führte zu unterschiedlichen Studienergebnissen. Ein R. equiTotimpfstoff erhöhte den spezifischen Antikörpergehalt im Kolostrum der geimpften Stuten und im postkolostralen Serum der zugehörigen Fohlen, blieb aber für die Fohlen ohne Schutzwirkung gegen die R. equi-Pneumonie (MADIGAN et al. 1991). In einer weiteren Studie zeigten die Fohlen von mit Lebendimpfstoff vakzinierten Stuten ebenfalls eine signifikant erhöhte Antikörperkonzentration gegenüber der Kontrollgruppe, unterschieden sich jedoch nach experimenteller Infektion weder hinsichtlich der Erkrankungs- noch der Überlebensrate von den Fohlen ungeimpfter Tiere (MARTENS et al. 1992). In einer mehrjährigen Studie hingegen konnte durch Immunisierung von fast 800 Mutterstuten mit einem R. equi-Totimpfstoff in der Zeit von 1992 bis 1995 die Mortalität von 3 auf 1,2 % deutlich reduziert werden (BECÚ et al. 1997). CAUCHARD et al. (2004) verglichen die Mutterschutzimpfung durch eine VapA-Antigen-haltige Vakzine, der ein wasserlösliches Nanopartikel-Adjuvans (Montanide IMS 3012) zugesetzt wurde, mit der Verabreichung eines formalininaktivierten Ganzzell-Impfstoffs. Die gemessenen Antikörper der VapA-Gruppe zeigten sowohl hinsichtlich der Konzentration im Fohlenserum am 30. und am 45. Lebenstag als auch in der Opsonisierungsaktivität im Fohlen- und im Stutenserum gegenüber den Antikörpern der anderen Impfgruppe bzw. der Kontrollgruppe ein signifikant besseres Ergebnis. Hier schienen allerdings die Antikörper beider Impfgruppen eine Schutzwirkung zu übertragen, da nur in der Kontrollgruppe vier der 15 Fohlen eine Pneumonie entwickelten. 2.1.10.2 Impfung der Fohlen Ebenso wie bei der Mutterschutzvakzinierung führt die Verwendung eines R. equiTotimpfstoffs bei der aktiven Immunisierung der Fohlen nicht zu einheitlichen Ergebnissen. Manche Autoren beschreiben den Einsatz eines Totimpfstoffs als wirkungslos (BECÚ et al. 1997, PRESCOTT et al. 1997b). In einer Studie wird von 29 Kapitel 2: Literaturübersicht der Reduzierung der Morbidität und der Letalität durch einen formalin-inaktivierten R. equi-Impfstoff sowohl alleine als auch in Kombination mit einer EHV-2-Vakzine in Bezug auf eine Feldinfektion berichtet (VARGA et al. 1997). Ein oral verabreichter R. equi-Lebendimpfstoff führte zu einer Immunität, die bei experimenteller Infektion sowohl eine Erkrankung verhinderte als auch eine schnelle Beseitigung des Erregers aus der Lunge bewirkte (CHIRINU-TREJO et al. 1987). Jedoch gilt der Einsatz eines Lebendimpfstoffs auf endemisch mit R. equi betroffenen Gestüten aufgrund der möglichen Vermehrung des Erregers im Fohlendarm als zu gefährlich. Der Aufbau einer schützenden Immunität durch einen Lebendimpfstoff konnte später durch HOOPER-McGREVY et al. (2005) bestätigt werden, die vier Fohlen im Alter von 2 Tagen, einer Woche und drei Wochen oral mit dem virulenten Stamm ATCC 33701 immunisierten und am Tag der letzten Impfung intratracheal infizierten. Weder in der klinischen Beobachtung noch in der zwei Wochen nach Infektion durchgeführten Sektion wurden in der Impfgruppe Hinweise auf eine Pneumonie gefunden. Darüber hinaus war im Lungengewebe der immunisierten Tiere der Erregergehalt signifikant geringer als bei der Kontrollgruppe, in der alle Fohlen eine Pneumonie mit deutlichen klinischen Erscheinungen und Sektionsbefunden entwickelten. Damit wurde gezeigt, dass der orale Weg für eine Immunisierung mit virulenten R. equi-Stämmen nicht immunsuppressiv ist und dass Fohlen im Alter von drei Wochen eine schützende Immunantwort aufbauen können (HOOPER-McGREVY et al. 2005). Es wird jedoch, um die Risiken im Einsatz eines Lebendimpfstoffs zu minimieren, die Erforschung schwach virulenter Mutanten, die dennoch einen ausreichenden Schutz bieten können, für notwendig gehalten. 2.1.10.3 Impfstudien Im Mittelpunkt der Entwicklung neuer Impfstoffe stand in den letzten Jahren das virulenz-assoziierte Protein A (VapA), das im Mäusemodell eine auf einer verstärkten Th1-Antwort beruhende Immunität hervorruft (PRESCOTT et al. 1997a). Darüber hinaus induziert VapA in Pferden die Bildung schützender VapA-Antikörper, die in experimentell mit R. equi infizierten Mäusen zu einer dosisabhängigen Eliminierung des Erregers aus allen Geweben führen (FERNANDEZ et al. 1997). Eine DNAVakzine auf der Grundlage des Genoms, das für VapA codiert, induzierte nach 30 Kapitel 2: Literaturübersicht gleichzeitiger intradermaler und intranasaler Verabreichung in erwachsenen Ponies eine Erhöhung der Antikörper gegen VapA sowohl im Serum als auch in der bronchoalveolären-Lavage-Flüssigkeit (BALF) (LOPEZ et al. 2003). Des Weiteren zeigten aus der BALF entnommene Zellen in vitro eine signifikant höhere Proliferationsrate auf eine erneute Konfrontation mit dem VapA als auf ein unspezifisches R. equi-Antigen. Bei Fohlen steigerte dieser Impfstoff zwar nicht die zelluläre Aktivität, aber sowohl im Serum als auch in der BALF zeigten zwei von fünf Tieren erhöhte spezifische Antikörpergehalte gegen VapA. Sowohl eine DNAVakzine auf der Basis des VapA als auch das VapA selbst wurden im Mäusemodell getestet (VANNIASINKAM et al. 2005). Beide Reagenzien induzierten eine deutliche Th1-Antwort, die im Fall der DNA-Vakzine durch Zugabe von Interleukin 12 (IL-12) noch verstärkt werden konnte. IL-12 spielt eine Schlüsselrolle bei der Differenzierung von omnipotenten Lymphozyten in Th1-Zellen und unterstützt somit die zelluläre Immunantwort (PARK u. SCOTT 2001). Im Gegensatz zu einem gleichzeitig im Mäusemodell getesteten Lebendimpfstoff bewirkte weder die DNA-Vakzine noch das VapA einen ausreichenden Schutz gegen eine experimentelle Infektion. Eine protektive Immunität wurde jedoch später mit dem gleichen Versuchsaufbau beobachtet (HAGHIGHI et al. 2005). Mit einer DNA-Vakzine auf der Basis von VapA intramuskulär oder intraperitoneal immunisierte Mäuse zeigten im Vergleich zu Kontrolltieren eine signifikant höhere Eliminierung des Erregers vier Tage nach intravenöser Applikation einer R. equi-haltigen Lösung. Darüber hinaus erhöhte die Zugabe der DNA-Sequenz, die für IL-12 codiert, die Eliminierungsrate signifikant, reduzierte jedoch im gleichen Maße den spezifischen Antikörpergehalt, ein Phänomen, das bereits bei DNA-Impfungen gegen Mycobacterium tuberculosis und Mycobacterium avium beobachtet wurde. 2.1.10.4 Hyperimmunplasma Das Serum von Tieren, die eine Infektion mit R. equi erfolgreich überstanden haben, an junge Fohlen zu verabreichen ist eine Prophylaxemaßnahme, über die schon seit langem berichtet wird (SCHMIEDHOFFER 1922). In den neunziger Jahren des letzten Jahrhunderts wurde über den erfolgreichen Einsatz von Hyperimmunplasma berichtet, das von Spendertieren gewonnen wurde, die mit einem R. equi- 31 Kapitel 2: Literaturübersicht Lebendimpfstoff oder einem inaktivierten R. equi-Stamm immunisiert wurden (MARTENS et al. 1989, 1992, MADIGAN et al. 1991). Die Beobachtung, dass das infundierte Plasma einen besseren Schutz bietet als die passive Immunisierung durch Kolostrum wird auf im Kolostrum nur in geringen Konzentrationen vorhandene Faktoren der humoralen Abwehr wie Komplement, Interferone, Zytokine und Fibronektin zurückgeführt (MARTENS et al. 1989, 1992, MADIGAN et al. 1991). Unterstützt wird diese Ansicht von der Erkenntnis, dass Serum von immunkompetenten Pferden, ohne gesteigerten R.-equi-Antikörpergehalt, ebenso wirksam ist wie Hyperimmunplasma (PERKINS et al. 2001). An 28 Fohlen wurde ein handelsübliches Hyperimmunplasma mit einem Plasma verglichen, das nur aufgereinigte Immunglobuline gegen die Virulenzfaktoren VapA und VapC enthielt (HOOPER-McGREVY et al. 2001). Dass beide Präparate im Vergleich zur Kontrollgruppe die klinischen Symptome und die Schäden in der Lunge reduzierten, wird mit der Schutzwirkung der Antikörper gegen VapA und VapC erklärt. Es wird aber auch nicht ausgeschlossen, dass trotz der Aufreinigung der Immunglobuline bisher noch nicht identifizierte Proteine in der Infusionslösung vorhanden waren (HOOPER-McGREVY et al. 2001). Neben einer unterschiedlichen Präparation des Hyperimmunplasmas werden unterschiedliche Zeitpunkte und Häufigkeiten der Infusionen angegeben. Solange sie vor der ersten Exposition mit dem Erreger stattfindet, wird eine einmalige Infusion bis zum 60. Lebenstag als ausreichend erachtet (MADIGAN et al. 1991). Für Gestüte mit hoher Prävalenz von R. equiErkrankungen und hoher Mortalität wird eine Infusion von einem Liter Hyperimmunplasma während der ersten Lebenstage und eine zweite von ebenfalls einem Liter in der dritten Lebenswoche vorgeschlagen (COHEN et al. 2000). Um die unterschiedlichen Präparationen (Lebend- oder Totimpfstoff) und Applikationszeiten bzw. –häufigkeiten verschiedenen vergleichbar zu Hyperimmunplasmen machen, auf wurde einem die Gestüt Wirksamkeit mit von endemischer Rhodokokkose überprüft (GIGUÈRE et al. 2002). Die Erkrankungsrate der 68 infundierten Fohlen fiel mit 19,1 % niedriger aus als die der 80 unbehandelten mit 30 %, wies damit aber keine signifikante Verbesserung auf. Trotz der identischen Präparation und nahezu gleichen Antikörpergehalten im Serum der Spendertiere wie MARTENS et al. (1992) konnten HURLEY und BEGG (1995) bei infundierten Fohlen 32 Kapitel 2: Literaturübersicht weder einen ausreichenden Schutz noch eine Abschwächung der Symptome beobachten. Der Einsatz von Hyperimmunplasma, das sieben Tage nach experimenteller Infektion zu therapeutischen Zwecken verabreicht wurde, zeigte keinen Einfluss auf die klinischen Erscheinungen oder den Krankheitsverlauf (CHAFFIN et al. 1991). 2.1.10.5 Sonstige Prophylaxemaßnahmen Verbesserungen in Haltung und Management, um Fohlen vor einer Infektion mit R. equi zu schützen, beziehen sich auf die epidemiologischen Eigenschaften des Erregers und insbesondere auf seine Verbreitungswege. Besonders Gestüte mit einer großen Zahl an Stuten bzw. Fohlen und mit viel Durchgangsverkehr zeigen ein erhöhtes Risiko, Rhodokokkose als Bestandproblem zu entwickeln (CHAFFIN et al. 2003). In die Überlegungen der Betriebsführung sollten Maßnahmen, die den Keimdruck senken, wie das Auskoffern von kontaminierten Paddocks, die Vermeidung von Staub durch Beregnung von Treibwegen, die Pflege der Grasnarbe, die Reduzierung der Herdengröße oder ein Rotationssystem auf den Weiden aufgenommen werden (COHEN et al. 2000, CHAFFIN et al. 2003). Um die Fohlen vor weiteren Agenzien zu schützen, die ihre Gesundheit beeinflussen können, wird ein konsequentes Impf- und Entwurmungsprogramm empfohlen (FALCON et al. 1985). Besonders auf Gestüten mit endemischen R. equi-Pneumonien wird der Aufbau eines Früherkennungsystems geraten, das je nach möglichem bzw. erwünschtem Umfang aus Temperaturkontrolle, Lungenauskultation, Messung der Leukozytenzahl sowie Röntgen- und Ultraschalluntersuchung bestehen kann (COHEN et al. 2000, GIGUÈRE et al. 2003b). 2.2 Das Immunsystem beim Fohlen im Zusammenhang mit R. equi Neben uneinheitlichen Angaben zum Erfolg der Prophylaxe sind Erkenntnisse zur Reaktion des Immunsystems der Fohlen auf eine Infektion mit R. equi noch unvollständig und der Grund, warum Fohlen in dem Altersabschnitt bis sechs Monate eine besondere Anfälligkeit für diesen Erreger aufweisen, noch nicht geklärt. Erste Hypothesen für diese Prädisposition der Fohlen beriefen sich auf das zeitliche 33 Kapitel 2: Literaturübersicht Zusammentreffen der noch insuffizienten humoralen und zellulären Immunität im Alter von acht bis sechzehn Wochen (YAGER 1987, TAKAI et al. 1995) mit dem Schwinden maternaler Antikörper (AINSWORTH 1999). 2.2.1 Die humorale Abwehr Mitte der neunziger Jahre des letzten Jahrhunderts wurde die Nomenklatur des equinen Immunglobulinsystems überarbeitet. Die Analyse der Organisation der Gensegmente für die konstanten Regionen der schweren Ketten (cH-Gene) der equinen Immunglobuline (WAGNER et al. 1998) ergab, dass das Pferd neben je einem IgM, IgE und IgA die genetische Information für mindestens sechs IgG Subtypen besitzt. Zur Vereinfachung wurde auf dem Immunglobulin und Fc-Rezeptor Workshop in Uppsala, Schweden (2001) eine neue Nomenklatur festgelegt, die der Tabelle 1 zu entnehmen ist. Tab. 1: Nomenklatur des equinen Immunglobulinsystems Ig-Isotypen alte Nomenklatur Ig-Isotypen IGH Gene(b) GenBank neue Nomenklatur(a) accession number IgM IgGa IgM IgG1 IgG2 IgG3 IgG4 IgG5 IgG6 IgE IgA IgG(T) IgGb IgG(T) IgGc IgE IgA IGHM IGHG1 IGHG2 IGHG3 IGHG4 IGHG5 IGHG6 IGHE IGHA L49414 AJ302055 AJ302056 AJ312379 AJ302057 AJ312380 AJ312381 AJ305046 nt (a) Immunoglobulin und Fc-receptor Workshop in Uppsala, Schweden (2001) (http://medicine.uiowa.edu/CigW). (b) Die offizielle Immunglobulin Heavy Chain Gene Loci (IGH)-Nomenklatur, festgelegt auf der internationalen ImMunoGeneTics database (IMGT) (http://imgt.cines.fr). Etwa fünf bis zehn Wochen post natum erreichen die R. equi-Antikörper, die das Fohlen mit dem Kolostrum von der Stute erhält, ihre geringste Konzentration im „California-ELISA“, während ein Anstieg der endogen gebildeten R. equi-Antikörper ab der sechsten bis neunten Woche in diesem Testsystem gemessen wird (HIETALA et al. 1985, PAUL 2005). PRESCOTT et al. (1996) beobachten im VapA-ELISA die 34 Kapitel 2: Literaturübersicht erste Serokonversion durchschnittlich im Alter von acht bis zehn Wochen. Bei der kontroversen Diskussion über die prophylaktischen Effekte von Kolostrum und Hyperimmunplasma konnte die Frage nach der Bedeutung der übertragenen Antikörper in vivo bisher nicht beantwortet werden (MADIGAN et al. 1991, MARTENS et al. 1992, HOOPER-McGREVY et al. 2001, PERKINS et al. 2001). In vitro verbessert eine erhöhte Konzentration spezifischer R. equi-Antikörper sowohl die Opsonisierung der Erreger als auch die Verschmelzung der Phagolysosomen in den Makrophagen, die von mit R. equi konfrontierten juvenilen oder adulten Pferden stammen (ARDANS et al. 1986, HIETALA u. ARDANS 1987). Jedoch variiert die Opsonisierungsaktivität zwischen Seren verschiedener Pferde, denn es wurden unterschiedliche Phagozytoseraten von Hefen durch neutrophile Granulozyten trotz einer nahezu gleichen Konzentration von IgG, das gegen ein Antigen aus der Zellwand von Hefen gerichtet war, beobachtet (GRÖNDAHL et al. 1997). Neben der Menge eines Immunglobulins wird die Opsonisierungsaktivität von der Antigenspezifität und dem Isotyp bestimmt (JANEWAY u. TRAVERS 2002). Auf einem Gestüt mit endemischer Rhodokokkose konnte kein Zusammenhang zwischen dem Auftreten einer Pneumonie und dem zu diesem Zeitpunkt im Serum gemessenen Antikörpergehalt festgestellt werden, da im Serum von 94 erkrankten und 21 nicht erkrankten Fohlen keine Unterschiede im „California-ELISA“ von HIETALA et al. (1985) nachgewiesen wurden (TRISKATIS 2004). Immunkompetente adulte Pferde wiesen 14 Tage nach experimenteller Infektion mit R. equi einen signifikant höheren Gehalt an R. equi- und VapA-spezifischen IgGa (IgG1)- und IgGb (IgG4)-Antikörper als am Tag der Infektion im Serum auf (LOPEZ et al. 2002). HOOPER-McGREVY et al. (2003) stellten fest, dass sich zwischen juvenilen und adulten Pferden, die sich erfolgreich mit virulenten Feldstämmen von R. equi auseinandergesetzt hatten, und erkrankten Fohlen eine unterschiedliche Aufteilung in der IgG-Fraktion zeigte, die Antigen-Spezifitäten für die sieben virulenzassoziierten Proteine (Vap) A und C bis H besaßen. Bei den gesund gebliebenen Tieren lagen im Verhältnis mehr Antikörper des Isotyps IgGa (IgG1) vor und weniger IgGb (IgG4) oder IgGT (IgG3/IgG5), während bei den Fohlen, die eine Pneumonie entwickelten, der Gehalt der Isotypen IgGb (IgG4) und IgGT (IgG3/IgG5) 35 Kapitel 2: Literaturübersicht erhöht war. Da die erfolgreiche Eliminierung von R. equi mit einer Th1-Antwort einhergeht, spiegelt IgGa (IgG1) möglicherweise den Th1-Charakter einer Immunantwort wider und IgGb (IgG4) und IgGT (IgG3/IgG5) die Polarisierung in Richtung einer Th2-Antwort. In einer späteren Studie konnte diese Verteilung der Isotypen allerdings nicht mehr nachvollzogen werden (HOOPER-McGREVY et al. 2005). Die mit einer Lebendvakzine immunisierten und nach experimenteller Infektion klinisch gesund gebliebenen Fohlen zeigten vor allem einen signifikant erhöhten Gehalt an IgGT (IgG3/IgG5). Die Autoren schlossen aus dieser Beobachtung, dass die isotypenspezifische Antikörper-Antwort nicht als allgemeingültiger Indikator für einen Schutz angesehen werden kann, sondern sie abhängig sei sowohl vom Infektionsweg als auch davon, ob es sich um eine experimentelle Infektion handelt oder um eine natürliche. 2.2.2 Die zelluläre Immunität Da für die Lymphozyten weder die Antigenspezifität ihrer Rezeptoren noch die Immunantwort, die in ihnen ausgelöst wird, falls ein Antigen an die Rezeptoren bindet, im Voraus festgelegt ist, sind diese Zellen auf weitere Aktivierungssignale angewiesen. Dadurch wird gewährleistet, dass eine Immunantwort nur dann ausgelöst wird, wenn das erkannte Antigen seinem Ursprung nach weder körpereigen noch von einem harmlosen Umweltkeim stammt (MEDZHITOV u. JANEWAY 1997). Eine Aktivierung von T-Lymphozyten bedarf der so genannten Costimulierung durch die Expression von CD 80-Molekülen, die gleichzeitig von den Zellen durchgeführt wird, die auch das Antigen präsentieren. Für die Unterscheidung, ob das gebundene Antigen pathogenen oder apathogenen Ursprungs ist, haben die antigenpräsentierenden Zellen des angeborenen Immunsystems bestimmte Zielstrukturen. Sie nutzen Molekülmuster der Mikroorganismen, die essentiell für ihr Überleben und deshalb kaum Veränderungen wie Mutationen unterworfen sind. Dazu zählt zum Beispiel die von allen gram-negativen Bakterien in ihrer Zellmembran getragenen Lipopolysaccharide (LPS). Für die Erkennung dieser Strukturen, die in der Literatur pathogen-associated molecular patterns (PAMPs) genannt werden, besitzt das angeborene Immunsystem eine Reihe von Rezeptoren, die so genannten pattern recognition receptors (PRRs). Neben frei im Plasma zirkulierenden PRRs gibt 36 Kapitel 2: Literaturübersicht es auch zellgebundene Rezeptoren, zu denen die Toll-like receptors (TLR) zählen, die unter anderem auf antigenpräsentierenden Zellen wie den Dendritischen Zellen oder auf Makrophagen zu finden sind (MEDZHITOV u. JANEWAY 1997). 2.2.2.1 Toll-like-Rezeptoren (TLRs) Der Name Toll-like-Rezeptoren stammt von der bei der Fruchtfliege Drosophila als erstes entdeckten Rezeptorfamilie „Toll“ ab, wo sie nach experimenteller Infektion die Bildung bakterizider und fungizider Proteine induziert (LEMAITRE et al. 1996). In Säugetieren löst die Bindung von spezifischen Liganden an TLRs die Expression von Zytokinen und costimulatorischen Molekülen aus, die für eine Aktivierung des erworbenen Immunsystems notwendig sind (UNDERHILL u. OZINSKY 2002). Nach Bindung der spezifischen Liganden finden verschiedene Enzymaktivierungen statt, die zur Bildung von Zytokinen wie TNFα, IFN-γ, IL-1β, IL-6, IL-8, IL-10 und IL-12 oder der oberflächlichen Expression von Molekülen wie CD 80 führen (KRUTZIK et al. 2001). 2.2.2.2 Das Die zelluläre Abwehr im Zusammenhang mit R. equi Wissen über die Mechanismen der zellulären Immunität bei der Auseinandersetzung mit R. equi ist seit der Mitte der neunziger Jahre des letzten Jahrhunderts deutlich gewachsen. Die Anfälligkeit von Fohlen bis zum Alter von sechs Monaten wurde bis dahin mit der kurz nach der Geburt unzureichenden und erst mit zunehmendem Alter steigenden Phagozytoseaktivität der Alveolarmakrophagen begründet (ZINK et al. 1985). Mit der Erkenntnis, dass R. equi in der Lage ist, innerhalb dieses Zelltyps zu überleben (HIETALA u. ARDANS 1987), rückten andere Zellen ins Interesse der Forschung. Neutrophile Granulozyten sind in vitro in der Lage 99 % aller opsonisierten R. equi innerhalb von 15 Minuten zu phagozytieren (YAGER 1987). Fohlen, die im Untersuchungszeitraum bis zum sechsten Lebensmonat an einer R. equi-Pneumonie erkranken, weisen in der zweiten und vierten Lebenswoche signifikant weniger neutrophile Granulozyten in ihrem Serum auf als Fohlen, die gesund bleiben (CHAFFIN et al. 2004). Im Mäusemodell wurde darüber hinaus gezeigt, dass eine Inaktivierung dieses Zelltyps durch monoklonale Antikörper zu signifikant höheren Erregermengen in Leber, Milz 37 Kapitel 2: Literaturübersicht und Lunge gegenüber Kontrolltieren führt, wenn die Tiere einer experimentellen Infektion ausgesetzt werden (MARTENS et al. 2005). Eine weitere Erklärung auf die Frage der Prädisposition für R. equi liefern Unterschiede der Leukozytenpopulationen in der Bronchoalveolären LavageFlüssigkeit (BALF) und im Blut adulter und juveniler Pferde. Bei Fohlen liegt in der Lunge eine deutlich geringere Konzentration der T-Lymphozyten vor, wobei sich insbesondere die Anzahl der CD4+- und CD8+T-Zellen auf niedrigerem Niveau bewegt, auch wenn die Menge der CD4+T-Zellen schneller das Niveau erwachsener Pferde erreicht (BALSON et al. 1997). Die Gesamtzahl der Lymphozyten im Blut steigt bis zum dritten Lebensmonat an und kann im Alter von ein bis sechs Monaten sogar die Werte der Adulten übertreffen, wobei der Anteil der B-Lymphozyten bei Fohlen deutlich erhöht ist (FLAMINIO et al. 1999). Ebenso weisen mit R. equi infizierte Fohlen ein in Richtung der B-Lymphozyten verschobenes Verhältnis der Lymphozytenfraktion im Vergleich zu gesunden Gleichaltrigen auf, woraus FLAMINIO et al. (1999) den Schluss ziehen, dass ein Mangel an zytotoxischen TZellen der Grund für die Anfälligkeit gegenüber R. equi in diesem Alter ist. Im Mäusemodell bewirkte die Übertragung von CD4+-T-Zellen, die aus mit R. equi infizierten adulten Pferden gewonnen wurden und Interferon (IFN)-γ sezernieren, sowohl eine vollständige Beseitigung des Erregers aus dem Körper als auch eine Vorbeugung von Lungenläsionen (KANALY et al. 1996). Virulente Rhodokokkenstämme greifen in erkrankten Fohlen auf die Zytokinexpression ein und unterdrücken die IFN-γ-Produktion von CD4+-T- Lymphozyten (GIGUÈRE et al. 1999). Die aus der BALF adulter Pferde, die 14 Tage zuvor durch Inhalation mit R. equi infiziert wurden, isolierten und anschließend mit R. equi-Antigen oder mit VapA inkubierten Lymphozyten zeigten eine signifikant höhere IFN-γ-Produktion als Zellen, die vor der Auseinandersetzung mit dem Erreger entnommen wurden (LOPEZ et al. 2002). HINES et al. (2003) wiesen mit einer Durchflusszytometrie in der BALF immunkompetenter adulter Pferde als Antwort auf die Konfrontation mit virulenten R. equi-Stämmen eine signifikant höhere Zahl von CD4+ und CD8+T-Zellen nach, die IFN-γ sezernieren. In einer in vitro Studie reagierten T-Lymphozyten, die aus der BALF von eine Woche zuvor durch Inhalation mit R. equi infizierten Pferden 38 Kapitel 2: Literaturübersicht entnommen wurden, auf die Inkubation mit VapC, VapD und VapE mit der Expression von IFN-γ (KOHLER et al. 2003). PATTON et al. (2004) zeigten, dass R. equi-spezifische CD8+ zytotoxische T-Lymphozyten (CTL) die Zellen zerstören, die mit dem intrazellulär persistierenden Erreger infiziert sind, während nicht infizierte unversehrt blieben. Dabei wurde festgestellt, dass die Antigenpräsentation unabhängig vom üblichen MHC-1-Weg geschieht, der bei der Reaktion intrazellulärer Erreger wie Viren abläuft, sondern dass CD1+-Zellen für die Antigenpräsentation und damit die Stimulierung der CTLs verantwortlich sind (PATTON et al. 2004). Bei einer erneuten Auseinandersetzung mit R. equi beruht die Eliminierung des Keims bei immunkompetenten adulten Pferden auf einem starken lokalen Übertritt von antigenspezifischen T-Gedächtniszellen aus den betroffenen Lymphknoten in die Lunge (PATTON et al. 2005). Während die CTLs der Fohlen, die kurz nach der Geburt und im Alter von drei Wochen aus dem Blut gewonnen wurden, nicht in der Lage waren, infizierte Zellen zu zerstören, zeigt sich nach etwa acht Wochen eine CTL-Aktivität, die mit der adulter Tiere vergleichbar ist. Auf Gestüten mit endemischer Rhodokokkose zeigten im Untersuchungszeitraum an einer R. equi-Pneumonie erkrankte Fohlen sowohl mit zwei als auch mit vier Wochen im Verhältnis signifikant weniger CD4+- und mehr CD8+T-Lymphozyten als Gleichaltrige, die gesund blieben (CHAFFIN et al. 2004). Die Erkenntnisse von FLAMINIO et al. (1999) in Bezug auf die B-Lymphozyten konnten in der Studie von CHAFFIN et al. (2004) nicht nachvollzogen werden, da keine Differenzen zwischen kranken und gesunden Fohlen festgestellt wurden. Und schließlich stellten CHAFFIN et al. (2004) fest, dass nicht nur einzelne Zelltypen, sondern auch die Gesamtzahl der Leukozyten im Blut von Fohlen, die später im Untersuchungszeitraum eine Pneumonie entwickelten, im Alter von zwei und vier Wochen signifikant geringer war als bei denen, die gesund blieben. Die Autoren betonen, nicht der Beobachtung von GIGUÈRE et al. (2003b) zu widersprechen, die den Anstieg der Leukozytenzahl als gute Methode zur Früherkennung identifizierten, da in beiden Fällen zum Zeitpunkt der Diagnose einer Erkrankung mit R. equi eine Leukozytose vorliegt. Werden Makrophagen durch das von T-Zellen sezernierte IFN-γ und durch den von Makrophagen selbst produzierten Tumornekrosefaktor-α aktiviert, sind sie in vitro in 39 Kapitel 2: Literaturübersicht der Lage R. equi durch freigesetzte Sauerstoff- und Stickstoffradikale abzutöten (DARRAH et al. 2000). Das VapA von R. equi wird über Toll-like Rezeptor-2 der Makrophagen erkannt, was zur Bildung von Th1-Zytokinen wie Interleukin (IL)-12 oder Tumornekrosefaktor-α führt (DARRAH et al. 2004). Die Autoren vermuten, dass die Anfälligkeit der Fohlen gegenüber R. equi dadurch verstärkt wird, dass die Stimulierung des Immunsystems über TLR-2 unzureichend ist. Für den neonaten Menschen ist bekannt, dass er nicht in der Lage ist, eine gänzlich Th1 polarisierte Immunantwort zu erzeugen, und aus der Nabelschnur entnommene humane Leukozyten zeigen eine zu vernachlässigende IL-12-Antwort nach Aktivierung von TLRs im Vergleich zu Adulten. 2.3 Adjuvantien Gereinigte Antigene sind allein selten immunogen und erfordern den Zusatz von Adjuvantien, die als Substanzen definiert sind, welche die Immunogenität von Antigenen erhöhen (JANEWAY u. TRAVERS 2002). Adjuvantien können die Immunogenität der Proteine auf zwei Weisen verstärken: Erstens können sie lösliche Proteinantigene in partikuläres Material umwandeln, das die antigenpräsentierenden Zellen schneller aufnehmen. Dies geschieht etwa durch Anlagern der Antigene an Aluminiumpartikel oder durch Emulsion in mineralischem Öl. Die Umwandlung löslicher Proteine in unlösliche Partikel erhöht die Immunogenität in einem gewissen Grad, Adjuvantien wirken jedoch verhältnismäßig schwach, wenn sie nicht Bakterien oder deren Produkte enthalten, die als zweiter Faktor eines Adjuvans zur Erhöhung der Immunogenität beitragen. Das vollständige Freundsche Adjuvans ist eine Öl- und Wasseremulsion, Adjuvantien sind die abgetötete abgetötete Mykobakterien Bordetella enthält. Andere pertussis-Bakterien, bakterielle bakterielle Polysaccharide, bakterielle Hitzeschockproteine und bakterielle DNA. Die Wirkungen zeichnen sich durch eine verzögerte Antigenfreisetzung, verstärkte Aufnahme durch Makrophagen, Induktion von Costimulatoren in den Makrophagen (JANEWAY u. TRAVERS 2002) aus. Bakterielle Zellwandbestandteile oder abgetötete Bakterien weisen den Nachteil auf, dass sie selbst als Antigene erkannt werden und damit zu ungewollten Immunantworten führen können. Moderne Adjuvantien, wie CpG-Motive, 40 Kapitel 2: Literaturübersicht rufen solche unerwünschten Reaktionen aufgrund der geringen Molekülgröße und der Erkennung durch spezifische Rezeptoren nicht hervor. 2.3.1 CpG-Motive Die Fähigkeit bakterieller DNA, das Immunsystem von Säugetieren zu stimulieren, ist seit den Achtziger Jahren des letzten Jahrhunderts bekannt. Die tumorunterdrückende Wirkung einzelsträngiger DNA wird durch die erhöhte Produktion der Interferone (IFN) α, β, γ und die gesteigerte Vermehrung der natürlichen Killerzellen (NK)-Zellen erklärt (TOKUNAGA et al. 1988). Als entscheidende Strukturelemente für die immunstimulierende Wirkung dieser Oligodeoxynukleotide wurde ein zentraler Cytosin-Guanosin-Kern erkannt (KRIEG et al. 1995). Die so genannten CpG-Motive sind einzelsträngige DNA-Sequenzen, die in 5’-3’-Richtung ein Cytosin-Phosphat-Guanosin-Dinukleotid (CpG-Dinukleotid) besitzen, wobei am C5-Atom des Cytosins keine Methylgruppe gebunden ist. Die Inversion zu Guanosin-Cytosin oder die Methylierung der Oligodeoxynukleotide hebt die immunstimulierende Wirkung auf (HAECKER et al. 2002). Letzteres ist der Grund, warum die DNA von Wirbeltieren diese Wirkung nicht besitzt, da sie in der Regel methyliert vorliegt. Aus Bakterien, Viren oder Parasiten gewonnene natürliche DNA enthält ein Phosphodiester (PD)-Grundgerüst, während synthetisierte entweder ein PD-Rückgrat haben, das dem natürlichen nachempfunden ist, oder ein Phosphothioat (PT)-Rückgrat. Letzteres hemmt den schnellen Abbau durch ubiquitär vorkommende Nukleasen und bewirkt eine schnellere Aufnahme in Makrophagen (SESTER et al. 2000). Es bestehen Speziesunterschiede bezüglich der optimalen Sequenz der zwei flankierenden Nukleinsäuren sowohl in 3’-Richtung als auch in 5’Richtung und sie haben entscheidenden Einfluss auf die Immunantwort. So wurde bislang für folgende Spezies die entsprechende „beste“ Sequenz bestimmt (HARTMANN et al. 2000, BAUER et al. 2001, KAMSTRUP et al. 2001, RANKIN et al. 2001): Maus: GACGTT Mensch/Rind/Katze/Hund/Pferd: GTCGTT Schwein: ATCGAT. 41 Kapitel 2: Literaturübersicht Daneben bewirkt auch eine zunehmende Länge der Sequenz, d.h. die Anzahl der CG-Dinukleotide, eine Verstärkung der Immunantwort, so dass die aktuell eingesetzten synthetischen Sequenzen zum Teil aus über zwanzig Nukleotiden bestehen (HAECKER et al. 2002). Außerdem wurde festgestellt, dass die flankierenden Basen, aufgrund ihrer Wirkung in immun-stimulatorische und immunneutralisierende Sequenzen eingeteilt werden können. Im Vertebratengenom überwiegen die immun-neutralisierenden Sequenzen. Dies erklärt zusätzlich, warum vor allem bakterielle DNA Einfluss auf das Immunsystem hat (KRIEG et al. 1998a). Zu den in der Humanmedizin beobachteten Effekten synthetischer CpG-Motive zählen in vitro sowohl die B-Zell-Proliferation, die Aktivierung dendritischer Zellen, Makrophagen und Monozyten als auch die verstärkte Aktivität der natürlichen Killerzellen (KRIEG 2002). In vivo verstärken CpG-Motive die CD4+T-Zell-Antwort, die in Th1-Richtung abläuft (KRIEG 2002). In einer in vitro Studie konnte gezeigt werden, dass equine Leukozyten auf die Stimulierung durch synthetische CpG-Motive mit der Sezernierung von sowohl IFN-α und IFN-β als auch IL-6 und IL-12 reagieren (WATTRANG et al. 2005). Die Qualität der Zytokinantwort variierte je nach Sequenz der Oligodeoxynukleotide, insbesondere in Bezug auf die Anzahl der vorhandenen CpG-Dinukleotide. Des Weiteren unterschied sich die Antwort equiner Leukozyten auf die eingesetzten Oligodeoxynukleotide von der humaner, muriner oder porciner Zellen aus vorherigen Studien. 2.3.1.1 CpG-Motive als Impfstoff-Adjuvantien Die Verwendung von CpG-Motive als Adjuvantien zeigt im Mäusemodell bereits nach der ersten Immunisierung eine 66-fache Steigerung der antigen-spezifischen Immunantwort im Vergleich zu Aluminiumverbindungen (HARTMANN et al. 2000). VLEUGELS et al. (2002) berichten bei der Vakzinierung von Hühnern über eine lang anhaltende Ig und IgG-Antwort durch den Zusatz von CpG-Motiven. Die Höhe des Gesamtimmunglobulingehalts nach einmaliger Anwendung entsprach dabei dem, der sonst nach einer Boosterimpfung gemessen wird. Porcine Blutmonozyten reagieren in vitro auf die Zugabe von CpG-Motiven mit Proliferation und Sekretion von IL-6, IL- 42 Kapitel 2: Literaturübersicht 12 und TNF-α (KAMSTRUP et al. 2001). Darüber hinaus erzeugten sowohl CpG- als auch GpC-Motive, die einem Schweineimpfstoff zugesetzt wurden, nach einmaliger intramuskulärer Applikation neben einer gesteigerten systemischen Immunantwort auch einen signifikant erhöhten IgA-Spiegel (VAN DER STEDE et al. 2002). Über die Wirkung auf canine und feline Zellen wurde bisher aus in vitro Studien berichtet, dass verschiedene CpG-Motive in diesen Tierarten ebenfalls eine Aktivierung der Lymphozyten induzieren (WERNETTE et al. 2002). RANKIN et al. (2001) wiesen in einer übergreifenden Studie die Proliferation induzierende Potenz verschiedener CpG-Motive bei allen untersuchten Spezies (Huhn, Hund, Katze, Kaninchen, Maus, Pferd, Schaf, Schwein und Ziege) nach. Die Vorteile im Einsatz von CpG-Motiven liegen nicht nur in der insgesamt stärkeren Immunantwort, sondern auch in der gesteigerten zellulären Abwehrreaktion. Im Rahmen einer Impfung gegen das bovine Herpesvirus-1 (BHV-1) bewirkte der Einsatz von CpG-Motiven die Änderung einer Th2-polarisierten in eine ausgeglichene Th2:Th1-Antwort (RANKIN et al. 2002). Auch im Vergleich zu anderen TLR-Liganden erzeugen CpG-Motive einen besseren Schutz, der unter anderem auf der signifikant gesteigerten Aktivität der spezifischen zytotoxischen T-Zellen beruht, die im Zusammenhang mit intrazellulären Erregern von entscheidender Bedeutung sind (SCHWARZ et al. 2003). In adulten Mäusen wurden durch eine Booster-Impfung mit CpG-Motiven die antigenpräsentierenden Zellen aktiviert, so dass sowohl Th1Zytokine als auch IgG2a und IgM vermehrt gebildet wurden (KOVARIK et al. 1999). Dagegen reagierten die antigenpräsentierenden Zellen juveniler Mäuse zwar in vitro auf CpG-Motive mit der Sekretion von IL-12, in vivo aber konnte der bei den Adulten beobachtete Effekt nicht im gleichen Maße wiederholt werden, da die Immunantwort zwar von Th1-Zytokinen geprägt war, jedoch keine eindeutige Polarisierung erreichte. Im Gegensatz zu Aluminiumverbindungen besitzen CpG-Motive den Vorteil, dass sie wenig bis gar keine Gewebsirritationen erzeugen (RANKIN et al. 2002, VLEUGELS et al. 2002). Darüber hinaus konnten HORNER et al. (2000) zeigen, dass CpG-Motive nach mukosaler Gabe sowohl eine Steigerung der lokalen als auch der systemischen Immunantwort induzieren. 43 Kapitel 2: Literaturübersicht Der alleinige Einsatz von CpG-Motiven als Adjuvantien wird von manchen Autoren wegen nicht ausreichender Wirkung kritisch diskutiert. Die Verwendung von Aluminiumverbindungen führte im Rahmen einer intramuskulären Impfung gegen das bovine Herpesvirus-1 (BHV-1) zu einer stärkeren Immunantwort, aber auch dosisabhängig zu lokalen Gewebsreizungen (RANKIN et al. 2002). Ebenso zeigt beim Schaf die subcutane Verabreichung der Kombination von CpG-Motiven mit Adjuvantien auf Basis von Mineralöl eine gute Depotwirkung, so dass im Vergleich zum alleinigen Zusatz von MineralölAdjuvantien die Konzentration und damit die Nebenwirkungen deutlich reduziert werden können (IOANNOU et al. 2002). Im Mäusemodell zeigen CpG-Motive in hohen Dosierungen von 50 bis 100 μg pro Tier Nebenwirkungen wie Splenomegalie oder Lymphadenopathien (JOSEPH et al. 2002, STORNI et al. 2004). Eine weitere kritische Beobachtung ist die Zersetzung der CpG-Motive durch DNAsen des Körpers, womit die von manchen Autoren beobachtete kurze Wirkungsdauer erklärt werden kann. Um diesen Nachteilen aus dem Wege zu gehen, wurde eine einfache und schnell durchzuführende Möglichkeit getestet, um CpG-Motive zusammen mit dem Antigen in Vesikel zu verpacken (JOSEPH et al. 2002). Diese Formulierung löste in Mäusen mit niedriger Dosierung schon nach einmaliger sowohl intramuskulärer als auch intranasaler Applikation die gleiche Th1-Antwort aus wie die 50-fache Menge des löslichen CpG-Motivs. Zu ähnlichen Ergebnissen führt die Applikation von CpG-Motiven, die in so genannten virus-like particles (VLPs) aus dem Bakteriophagen Qβ untergebracht wurden (STORNI et al. 2004). Die gleichzeitige Verpackung von CpG-Motiven und dem Antigen, gegen das immunisiert werden soll, in ein solches Vesikel, wies im Mäusemodell keinerlei Nebenwirkungen auf und konnte sogar eine T-Zell-Antwort erzeugen, die mit der eines Lebendimpfstoffs vergleichbar ist. 2.3.1.2 CpG-Motive in der Therapie Neben dem Anwendungsgebiet als Impfstoffzusatz wird auch der Einsatz von CpGMotiven in der Therapie in der Literatur beschrieben. Adulte Mäuse, die experimentell mit dem intrazellulären Erreger Listeria monozytogenes infiziert waren, konnten sich erst nach Gabe von CpG-Motiven erfolgreich von ihm befreien (KRIEG et al. 1998b). 44 Kapitel 2: Literaturübersicht Der positive Einfluss von CpG-Motiven auf die Eliminierung von Leishmania major wird im Wesentlichen auf drei Mechanismen zurückgeführt (DITTMER u. OLBRICH (2003). Zum einen wird die Bildung von MHC-1 und MHC-2 in spezifischen antigenpräsentierenden Zellen wie den dendritischen Zellen gesteigert, was entscheidend ist, damit das Antigen den T-Zellen präsentiert wird. In den dendritischen Zellen wird darüber hinaus die Bildung von IL-12 gesteigert, was die IFN-γ-Produktion und damit die T-Zell-Antwort steigert. Als drittes induzieren CpGMotive die Bildung von IL-6 über TLR-9, was den hemmenden Effekt von regulierenden T-Zellen blockt, also eine Hemmung der T-Zellen aufhebt. Im Zusammenhang mit der Tumortherapie zeigte sich die Wirkung von CpG-Motiven im Mäusemodell in der vermehrten Aktivierung der dendritischen Zellen und der zytotoxischen T-Zellen, wodurch der Tumor erfolgreich bekämpft wurde (DAVILA u. CELIS 2000). 2.4 Die intranasale Applikation von Impfstoffen Die Schleimhautoberflächen des Körpers sind wie die äußere Haut direkt mit der Umgebung verbunden und daher einer großen Zahl von Krankheitserregern ausgesetzt. Neben mechanischen (tight junctions der Epithelzellen), chemischen (Enzyme, niedriger pH-Wert) und mikrobiologischen Barrieren (kommensale Mikroflora) befindet sich in der Nähe der Oberflächen das Immunsystem der Schleimhaut, das als MALT (mucosa-associated lymphoid tissue) bezeichnet wird (JANEWAY u. TRAVERS 2002). Dieses Element der adaptiven Immunantwort zeichnet sich durch seine Unabhängigkeit von den lymphatischen Geweben des restlichen Körpers aus (TIZARD 2000). Durch so genannte homing-Rezeptoren wandern die Lymphozyten aus den regionalen Lymphknoten gezielt zu der betroffenen Schleimhautoberfläche, wodurch das Geschehen lokal begrenzt bleibt (JANEWAY u. TRAVERS 2002). Ein besonderes Kennzeichen der Schleimhautoberflächen ist das vermehrte Aufkommen von IgA, das durch Neutralisation das Interagieren von eingedrungenen Krankheitserregern mit den Epithelzellen verhindert. Die präzise lokale Immunantwort gilt auch für die Gedächtniszellen, so dass eine den Infektionsweg über die Schleimhaut nachahmende Impfung sinnvoll erscheint. Allerdings sind 45 Kapitel 2: Literaturübersicht inaktivierte Antigene alleine wirkungslos, da sie ähnlich wie Proteine der Nahrung oder viele Bestandteile der Umgebungsluft von spezifischen T-Zellen erkannt werden, aber ohne eine Costimulierung keine Antwort des adaptiven Immunsystems auslösen. Dagegen zeigen intranasal applizierte Lebendimpfstoffe wie gegen die bovine oder feline Rhinotracheitis (TIZARD 2000) und Pseudomonas aeruginosa (PRIEBE et al. 2002) die Ausbildung einer schützenden Immunität. Weitere Berichte über intranasal verabreichte inaktivierte Vakzine zeigten nur in Kombination mit Adjuvantien wie dem Choleratoxin (WU et al. 1997), dem Protoxin von Bacillus thuringiensis (MORENO-FIERROS et al. 2002) oder CpG-Motiven (McCLUSKIE et al. 2002, JIANG et al. 2003) einen Impferfolg in Form von signifikant erhöhten IgASpiegeln oder verbesserten Proliferationsraten der antigenspezifischen Lymphozyten. Im Zusammenhang mit einer Impfung gegen R. equi wird die intranasale Applikation einer DNA-Vakzine beschrieben, um Einflüsse der maternalen Antikörper zu umgehen, wobei das Impfprotokoll in der serologischen Untersuchung nur bei einigen Probanden zu einem Unterschied im Vergleich zur Kontrollgruppe führte (LOPEZ et al. 2003). Ziel der vorliegenden Arbeit war es, die beschriebenen Mechanismen des mukosalen Immunsystems durch eine intranasale Impfung zu nutzen und mit Hilfe von CpG XXXX die Antwort auf die Vakzinierung in die Th1-Richtung zu polarisieren. Die intranasale Applikation eines R. equi-Impfstoffs mit einem CpG-Motiv als Adjuvans wurde bisher in der Literatur beim Fohlen nicht beschrieben. R. equi stellt als intrazellulär persistierender Erreger ein geeignetes Modell dar, um zu eruieren, ob diese Impfstoffverabreichung und -formulierung geeignet ist, beim Fohlen eine Immunität aufzubauen, die beim Menschen und im Mäusemodell auf der Aktivierung der zellulären Immunantwort beruht. 46 Kapitel 3: Material und Methode 3 Material und Methode In der vorliegenden Arbeit wurde ein R. equi-Impfstoff mit und ohne Adjuvans bei Fohlen in einer kontrollierten und randomisierten Blindstudie auf seine klinische Wirksamkeit und seinen Einfluss auf den Antikörperverlauf untersucht. 3.1 Probandengut Auf einem Warmblutgestüt in Norddeutschland, auf dem endemisch durch R. equi verursachte Pneumonien auftreten, wurden 108 Fohlen in der Zeit von Juni bis Juli 2005 geimpft und standen bis zum Alter von vier Monaten (125. Lebenstag) zur Verfügung. Es handelte sich bei den Probanden um 46 Stutfohlen und um 62 Hengstfohlen verschiedener Warmblutrassen. 3.1.1 Haltung Die Mutterstuten wurden während der Wintermonate in mit Stroh eingestreuten Laufställen und über den Sommer Tag und Nacht auf der Weide gehalten. Der Abfohlbereich war von den übrigen Stallungen räumlich getrennt und der Personenverkehr eingeschränkt. Ante partum wurden die Stuten in 3 x 4 Meter große Abfohlboxen verbracht, die vor dem Aufstallen sowohl mit Wasser als auch mit einem Universalreiniger gereinigt und anschließend mit einem Desinfektionsmittel behandelt wurden. Die Stuten wurden in den Abfohlboxen entweder auf Stroh oder auf Späne gehalten. Um sowohl die Geburtsüberwachung als auch die Geburtshilfe zu erleichtern und zur Reduktion des Keimgehaltes während der Geburt, wurde der Schweif der Stuten mit einer elastischen Einmalbinde einbandagiert. Weiterhin wurde den Stuten zwecks Keimreduktion die Innenseite der Hinterbeine und das Euter mit einer jodhaltigen Seife (Braunol®, Braun, Melsungen) gewaschen. Das Waschen der Hinterbeine und des Euter unterblieb, wenn die Stuten zu starke Abwehrbewegungen zeigten. Nach etwa acht bis zehn Tagen wurden die Stuten mit Fohlen aus dem Abfohlbereich in Laufställe verbracht. Diese Laufställe bestanden aus einem mit Stroh eingestreuten überdachten Bereich und einem Paddock, dessen Boden mit 47 Kapitel 3: Material und Methode Beton befestigt war. Die Gruppengrößen schwankten je nach Stallgebäude zwischen acht und zwölf Mutterstuten mit den dazugehörigen Fohlen. Mitte Mai begann die Weidehaltung für die Fohlen, die zu diesem Zeitpunkt älter als 5 Wochen waren. Dabei wurden die Herden in Größen von etwa 30 Mutterstuten zusammengestellt. Wurde ein Fohlen im Rahmen der regelmäßigen Untersuchung als beobachtungsoder therapiewürdig beurteilt, so brachte man Stute und Fohlen zurück in die Laufställe. 3.1.2 Impfungen und Entwurmungen Die Mutterstuten wurden zwei Mal jährlich sowohl gegen das equine Herpesvirus 1 und 4 als auch gegen Influenza immunisiert und alle zwei Jahre wurde eine Tetanusimpfung durchgeführt. Die Entwurmung der Stuten erfolgte alle drei Monate. Die Fohlen wurden das erste Mal am zehnten Lebenstag und anschließend alle vier Wochen entwurmt. 3.1.3 Erstversorgung der Fohlen Nach der Geburt wurden die Vitalfunktionen des Fohlens überprüft, ein Klistier (Practoclyss® Friedeberg-Fresenius Kabi, Bad Homburg) verabreicht und der Nabel mit einer ethanolhaltigen Jodlösung desinfiziert. Anschließend wurde das Stutenkolostrum mittels Refraktometrie auf seine IgG-Konzentration untersucht (MARKUS 2005). Der Brechungsindex wurde mit einem Zuckerrefraktometer (Digital Handheld ´Pocket´ Refractometer PAL-1, Fa. ATAGO CO, LTD, Tokyo, Japan) gemäß der Bedienungsanleitung des Herstellers bestimmt. Ein Brechungsindex über 25 % kennzeichnet ein Kolostrum mit einem IgG-Gehalt von über 50 g/l also guter Qualität. Nahmen die Fohlen in den ersten zwei Lebensstunden weniger als 250 ml Kolostrum auf, wurde ihnen abgemolkenes Kolostrum der Mutterstute mit einer Flasche eingegeben. War das Abmelken der Stute nicht möglich oder das Kolostrum von zu schlechter Qualität, stand tiefgefrorenes, gestütseigenes Kolostrum zur Verfügung. Acht bis zehn Stunden nach der Geburt wurde der IgG–Gehalt im EDTABlut der Fohlen mittels Snap Foal IgG Test (IDEXX, Blue Ridge Pharmaceuticals, Westbrook, Marine, USA) bestimmt. Bei einem IgG-Gehalt unter 800 mg/dl, wurde den Fohlen je nach Qualität 250 bis 500 ml Kolostrum aus der betriebseigenen 48 Kapitel 3: Material und Methode Kolostrumbank eingegeben. War der IgG-Gehalt nach weiteren vier Stunden nicht auf mindestens 800 mg/dl angestiegen, wurde ein Liter eines Plasmas infundiert, das von adulten Pferden des Betriebes stammte. Im Zusammenhang mit der Blutentnahme wurde den Fohlen 20 ml eines Immunplasmas gegen Rota- und Coronaviren (Eurovet, Smöaunn, Dänemark) oral verabreicht und der Nabel erneut desinfiziert. Die Kontrolle und Desinfektion des Nabels erfolgte bis zum etwa zehnten Lebenstag zweimal täglich. Wegen schwerer Durchfallerkrankungen im März 2005, die sowohl durch Rota- und Coronaviren als auch durch Escherichia coli ausgelöst wurden, erhielten die Fohlen der Abfohlsaison 2005 seitdem in den ersten zehn Lebenstagen eine orale Antibiotikatherapie mit den Wirkstoffen Colistin, Neomycin und Sulfadimidin. 3.1.4 Bedingungen für die Aufnahme in die Studie Alle in der Studie vorkommenden Fohlen wurden auf dem gleichen Gestüt geboren und erhielten über den Untersuchungszeitraum die gleichen Haltungs- und Fütterungsbedingungen. Fohlen wurden nur in die Studie aufgenommen, wenn das Allgemeinbefinden der Mutterstute keine Störungen aufwies, die Trächtigkeitsdauer in den physiologischen Grenzen von 315 bis 355 Tagen lag, die Fohlen durch eine ungestörte Geburt zur Welt kamen und ihr Allgemeinbefinden ungestört war. Ferner musste der IgG-Gehalt acht bis zehn Stunden post natum einen Wert von mindestens 800 mg/dl betragen, um von einer ausreichenden Kolostrumaufnahme ausgehen zu können. Von besonderer Bedeutung war die uneingeschränkte Funktionsfähigkeit der Lunge. Unmittelbar vor der hier zu besprechenden Studie hat auf dem gleichen Gestüt eine Untersuchung der Lunge von 138 Fohlen innerhalb der ersten 24 Stunden post natum stattgefunden. Dabei wurden bei den Fohlen weder durch die klinische noch die sonographische Untersuchung pathologische respiratorische Befunde erhoben (BAUMANN 2006, persönliche Mitteilung). Somit wurde in der vorliegenden Studie beim neonaten Fohlen die auskultatorische Untersuchung der Lunge als hinreichend akzeptiert. Stellte sich im Verlauf der Studie bei einem Probanden eine schwere Störung des Allgemeinbefindens ein (z.B. Kolik, Fraktur, Polyarthritis), deren Genese unabhängig von R. equi war, führte dies zum Ausschluss aus der Studie. 49 Kapitel 3: Material und Methode 3.1.5 Einteilung der Probanden in Gruppen Der vorliegende Versuch wurde als Blindstudie durchgeführt und erst nach Beendigung der Versuchsreihe und Analyse der Ergebnisse klärte der Hersteller des Impfstoffs, die Wirtschaftsgenossenschaft deutscher Tierärzte eG (WdT, Garbsen), Serumwerk Memsen, den Inhalt der Flaschen auf. Die Fohlen der Studie wurden in folgende drei Gruppen aufgeteilt: Gruppe A = Impfstoff A (n=36) Gruppe B = Impfstoff B (n=36) Gruppe C = Impfstoff C (n=36) Die Zuteilung der Fohlen in die Gruppen erfolgte randomisiert durch Auswürfeln. Gruppe A war die Placebogruppe, der eine 0,9 % Kochsalzlösung verabreicht wurde. Die Fohlen der Gruppe B erhielten den Totimpfstoff mit Zusatz des Adjuvans CpG XXXX. Den Fohlen der Gruppe C wurde der Totimpfstoff ohne den Zusatz eines Adjuvans appliziert. 3.2 Rhodococcos equi-Impfstoff Bei den für die Herstellung des Impfstoffs verwendeten R. equi-Stämmen handelt es sich um drei virulente Stämme, die im August 2003 am Institut für Mikrobiologie der Stiftung Tierärztliche Hochschule Hannover aus Tracheobronchialsekretproben von erkrankten Fohlen des gleichen Gestüts isoliert wurden. Die Stämme wurden bei der Wirtschaftsgenossenschaft deutscher Tierärzte eG (WdT, Garbsen), Serumwerk Memsen, lyophilisiert aufbewahrt und wurden vor der Herstellung erneut auf Reinheit und Identität überprüft. Nach Kultivierung in einer Nährbouillon wurden die Stämme mit Formalin inaktiviert. Anschließend erfolgte das Einstellen der Bakterienkonzentration auf 109 pro ml Impfstoff sowie die Zugabe von 0,5 % Phenol als Konservierungsmittel. Die so hergestellte Lösung wurde in zwei Portionen geteilt, von denen einer 500 μg/ml CpG XXXX als Adjuvans zugefügt wurde, während die 50 Kapitel 3: Material und Methode andere ohne Adjuvans blieb. Anschließend folgte die Abfüllung in Injektionsflaschen zu je 4 ml und die Prüfung auf Sterilität gemäß der Europäischen Pharmakopoe. Die Sequenz des eingesetzten CpG-Motivs XXXX wird auf Wunsch der Wirtschaftsgenossenschaft deutscher Tierärzte eG (WdT, Garbsen) an dieser Stelle nicht veröffentlicht. Um den Einfluss von DNAsen im formalininaktivierten Impfstoff auf das Adjuvans auszuschließen, wurde qualitativ die Enzymaktivität im Institut für Physiologische Chemie der Stiftung Tierärztliche Hochschule Hannover bestimmt. Eine zu diesem Zweck dem Impfstoff hinzugefügte bekannte Menge DNA blieb nahezu vollständig erhalten, so dass ebenfalls nicht von einem Abbau des CpG-Motivs durch DNAsen des Impfstoffs auszugehen ist. 3.3 Serumprobe und Impfung Bei den in die Studie aufgenommenen Probanden wurde acht bis zehn Stunden nach der Geburt neben einer EDTA-Blutprobe auch eine erste Serumprobe entnommen. Hierfür wurde die Vena jugularis mit einer 1,2 x 40 mm großen Kanüle (Sterican®, B. Braun, Melsungen) punktiert und etwa 4 ml Blut in einer Serummonovette (Sarstedt®, Nürnbrecht) aufgefangen. Die gewonnenen Proben wurden für höchstens zwei Tage in einem Kühlschrank bei unter 4 °C gelagert, damit sich die festen Bestandteile des Blutes absetzen konnten. Zur besseren Trennung wurden die Proben für sechs Minuten bei 3000 x g zentrifugiert, der Überstand in Kryoröhrchen abpipettiert und bis zur Untersuchung bei -20 °C eingefroren. Die Serumprobennahme erfolgte nach folgendem Schema: Die erste Serumprobe wurde acht bis zehn Stunden post natum, die Zweite am sechsten bis achten Lebenstag, die dritte am 13. bis 15. und die vierte am 20. bis 22. Lebenstag gewonnen. Die fünfte Serumprobe stand für den 27. bis 29. Lebenstag an und war die letzte, die so genau terminiert wurde, da die weiteren Proben im Rahmen der alle zwei Wochen stattfindenden klinischen Untersuchung genommen wurden. 51 Kapitel 3: Material und Methode Die erste Impfung wurde zeitgleich mit der ersten Serumprobe durchgeführt. Aus einem dem jeweiligen Fohlen zugeordneten Behältnis wurden in zwei 2,0 ml Spritzen (Braun, Melsungen) jeweils 1,0 ml der Lösung und etwa 1,0 ml Luft aufgezogen, bevor zur Zerstäubung im Nasenraum ein 50 mm langer Intranasal-Applikator aus Kunststoff mit einem Durchmesser von 4 mm (Pfizer, Karlsruhe) über einen Lueraufsatz auf die Spritzen gesteckt wurde. Zur Applikation wurde der Außenflügel der jeweiligen Nüster nach dorsal gehoben und die Spritze mit nach unten gerichtetem Intranasal-Applikator in die Nase eingeführt ohne die Schleimhaut zu berühren (siehe Abb. 1). Auf diese Weise wurde der Impfstoff in beide Nüstern sowohl aus der Spritze als auch mit der Luft aus dem Applikator gedrückt. Die zweite Impfung wurde auf den 20. bis 22. Lebenstag festgesetzt und fand im Anschluss an die vierte Serumprobe statt. Um beim Auftreten unerwünschter Reaktionen auf die Impfstoffgabe rechtzeitig reagieren zu können, wurden die Fohlen nach Applikation noch etwa 15 Minuten lang beobachtet. Abb. 1: Intranasale Applikation beim Fohlen. Der Außenflügel der Nüster wird nach dorsal gehoben und die Spritze mit nach unten gerichtetem Intranasal-Applikator in die Nase eingeführt, ohne dabei die Schleimhaut zu berühren. 52 Kapitel 3: Material und Methode 3.4 Untersuchung der Fohlen Zur Überprüfung des Gesundheitsstatus wurden die Probanden zu Beginn der Studie im Zwei-Wochen-Rhythmus klinisch allgemein untersucht. Wegen des vermehrten Auftretens von interstitiellen Pneumonien wurden ab September 2005 alle Fohlen, die jünger als drei Monate waren, wöchentlich einer Untersuchung unterzogen. Dabei wurden die Haltung, das Verhalten, insbesondere das Saugverhalten (inkl. Euterfüllung der Mutterstute) beobachtet sowie die rektal ermittelte Körperinnentemperatur gemessen. Darüber hinaus wurden die Kotkonsistenz, der Nabel und die Füllung der Gelenke beurteilt. Die klinische Allgemein- und die Lungenuntersuchung erfolgten für die Probanden von Geburt an bis zu einem Alter von mindestens 17 Wochen. 3.4.1 Klinische Untersuchung des Respirationstraktes Bei der klinischen Untersuchung des Respirationstraktes wurde der Nasenausfluss und die Größe der Mandibularlymphknoten beurteilt. Eine vermehrt abdominale Atmung, Nüsternblähen, das Einsinken der Interkostalräume oder spontaner sowie auslösbarer Husten wurde als ein schwerwiegendes Anzeichen einer Erkrankung des Respirationstraktes beurteilt. Auskultatorisch wurden die Trachea und die Lunge beidseits auf Giemen, Rasseln oder Knistern über mindestens zwei vollständige Atemzüge untersucht. Die auskultatorischen Befunde der Lunge wurden an drei Lokalisationen, cranio-ventral, Mitte und caudo-dorsal erhoben. Zur Beurteilung des Schweregrades einer Lungenerkrankung wurden die Punkte jedes klinischen Symptoms zum „klinischen Score“ addiert (nach OHNESORGE et al. 1998) (siehe Tab. 2). Die Zahlenwerte können zwischen 0 und 13 liegen, wobei Fohlen mit Werten von 0 – 1 als gesund, mit 2 – 3 als geringgradig, mit 4 – 6 als mittelgradig und Scorewerte von 7 oder mehr als hochgradig erkrank bezeichnet wurden. 53 Kapitel 3: Material und Methode Tab. 2: Klinischer Score zur Beurteilung des Schweregrads der klinischen Symptome (nach OHNESORGE et al. 1998) Merkmal Atemfrequenz Nasenausfluss Hustenauslösung Lnn. mandibulares Ruhedyspnoe Lungenauskultation Tracheaauskultation Befund Scorewert < 80 > 80 Nein serös, seromukös Purulent nicht auslösbar Mehrfach Spontan o.b.B. Vergrößert Nein Einsinken der Interkostalräume, Nüsternblähen vesikulär, vesikulär verschärft Rasseln, Knistern, Giemen o.b.B Rasseln 0 1 0 1 2 0 1 2 0 1 0 Gesamtscore 3 0 2 0 2 0 – 13 3.4.2 Labordiagnostische Untersuchung Weiterhin wurde der Gesundheitszustand durch die Bestimmung des Leukozytengehalts im Blut überwacht. Zu Ermittlung der Leukozytenzahl wurde Blut aus der Vena jugularis externa in ein mit Kalium-EDTA Probenröhrchen gefüllt. Die Leukozytenzahl wurde mit Hilfe eines Hämatologie-Analysator (Sysmex KX-21N, Sysmex, Kobe, Japan) nach dem elektrischen Widerstandsprinzip ermittelt. 3.4.3 Weiterführende Untersuchung des Respirationstraktes Bei den Probanden dieser Studie wurde bei dem Auftreten folgender Symptome eine sonographische Untersuchung der Lunge durchgeführt. • Leukozytose von über 13 x 109 Leukozyten/l im Blut • hochgradig ausgeprägte Atemnebengeräusche (Rasseln, Giemen) bei der Auskultation des Respirationstraktes • Tachypnoe in Verbindung mit Nüsternblähen und abdominaler Atmung • Rektal ermittelte Körperinnentemperatur von über 40 °C 54 Kapitel 3: Material und Methode • Husten • Kümmern. Die sonographische Untersuchung der Lunge erfolgte an stehenden von zwei Hilfspersonen fixierten Fohlen mit dem Ultraschallgerät „Sonovet 2000“ der Firma Kretztechnik AG, Tiefenbach, Österreich. Es handelt sich dabei um ein tragbares, akkubetriebenes Ultraschallgerät mit einem 7,5 MHz Linearscanner. Der Schallkopf wies eine Länge von 6,5 cm und eine Breite von 1,7 cm auf. Zur sonoraphischen Untersuchung der Lunge wurde ein von dorsal durch den M. longissimus dorsi, nach ventral durch das Zwerchfell und nach cranial durch das Schulterblatt begrenztes Untersuchungsfeld geschoren, mit 25 %-iger Propanollösung (Biesterfeld, Hamburg) entfettet und mit einem Ultraschallgel (BLR Sonic Ultraschallgel, Diagonal, Waldeck, Münster) versehen. Die Befunde wurden in einem vorgefertigtem Befundbogen eingetragen (siehe Abb. 2). Der Befundbogen sieht eine Einteilung der zwölf Interkostalräume in drei horizontal verlaufende Felder vor, so dass die ultrasonographisch erhobenen Befunde insgesamt 27 Bereichen einer Lungenseite zugeordnet wurden. Ultraschallbefunde, wie Abszesse und pneumonisch veränderte Bereiche wurden im Ultraschallgerät gespeichert und am selben Tag deren Abmessungen bestimmt. Um das Ausmaß der Lungenveränderungen zu quantifizieren, wurde ein Abzsess-Score errechnet, indem die Durchmesser aller Abszesse an einem Untersuchungstag beim untersuchten Probanden addiert wurden. Zusätzlich wurden alle Abszesse gezählt, wobei ihre Anzahl und ihre Tiefe den Schweregrad der Lungenveränderungen bestimmten. 55 Kapitel 3: Material und Methode Abb. 2: Befundbogen für die sonographische Untersuchung der Lunge 56 Kapitel 3: Material und Methode 3.5 Der Rhodococcus equi-ELISA Zur Bestimmung des Gehaltes an Antikörpern im Serum der Fohlen, die gegen R. equi gerichtet waren, wurde der von TRISKATIS (2004) modifizierte ELISA von HIETALA et al. (1985) verwendet. Die Untersuchung der Proben wurde in der Gesellschaft für Innovative Veterinärdiagnostik (IVD GmbH), An-Institut der Stiftung Tierärztliche Hochschule Hannover von Oktober bis Dezember 2005 durchgeführt. 3.5.1 Das Antigen Der in dieser Studie für die Antigenpräparation eingesetzte R. equi-Stamm wurde auf dem gleichen Gestüt 2003 im Rahmen der Studie von TRISKATIS (2004) aus dem Tracheobronchialsekret eines erkrankten Fohlens isoliert. Durch die 16-S-PCR nach SELLON et al. (2001) war der Erreger als R. equi Stamm identifiziert und mit Hilfe der PCR-Untersuchung nach MARKAI et al. (2002) der Virulenzfaktor VapA nachgewiesen worden. Das Antigen wurde nach der Methode von HIETALA et al. (1985) präpariert, wobei die Modifikationen von HIGUCHI et al. (1997a) berücksichtigt wurden. 3.5.2 Chargenprüfung und Antigenauswertung Vor der Durchführung des ELISA wurden eine Antigenauswertung und eine Chargenprüfung vorgenommen. Dazu wurden die Platten wie auch in den Hauptversuchen nach der Anleitung 3.5.3 vorbereitet und die Versuche, wie in 3.5.4 angegeben, durchgeführt. Ein definiertes Panel an Seren, das im Frühjahr 2004 von adulten Stuten des gleichen Gestüts gewonnen wurde, wurde sowohl auf den neu gecoateten als auch auf im Rahmen der Dissertation von MARKUS (2005) verwendeten Platten eingesetzt. Diese Vorversuche für den Ganzzell-ELISA zeigten, dass für das Fängerantigen eine Beschichtungskonzentration von 3,125 μg/ml geeignet war. 3.5.3 Herstellung der Rhodococcus equi-ELISA-Testplatten Das zur Verfügung stehende Antigen wurde mit Coatingpuffer (Rezept im Anhang) in einem Verhältnis von 1:320 verdünnt, um eine Konzentration von 3,125 μg/ml zu erzielen. 100 μl dieser Lösung wurden in jede Vertiefung einer zu beschichtenden 57 Kapitel 3: Material und Methode Polysorp®-Mikrotiterplatte (Firma Nunc, Wiesbaden) gegeben und die Plattencharge über Nacht bei 4°C inkubiert. Nach dreimaligem Waschen der Platte mit jeweils 200 μl TBST-Waschpuffer pro Vertiefung (ELISA-Washer PW 96, Tecan, Crailsheim) wurde die Restfeuchte ausgeschlagen und die Platten für eine Stunde mit der Öffnung nach unten zeigend bei Raumtemperatur (ca. 20 °C) getrocknet. Anschließend folgte die Lagerung der Mikrotiterplatten bei -20 °C. 3.5.4 Durchführung des ELISA Für die Untersuchungen mit dem R. equi-ELISA in der vorliegenden Arbeit wurden ausschließlich Mikrotiterplatten der Charge verwendet, die am 11.10.2005 gecoatet wurde. Alle im Zusammenhang mit dem ELISA stehenden Reagenzien waren zum Zeitpunkt ihres Einsatzes auf Raumtemperatur erwärmt. Als erstes wurden die Positivkontrolle, die Negativkontrolle und die zu untersuchenden Seren mit Tris buffered saline-Tween® 20 (TBST)-Waschpuffer (Rezept siehe Anhang) in einem Verhältnis von Serum zu Puffer von 1:50 vorverdünnt. Anschließend wurden alle Wells der ELISA-Platte mit 100 µl TBST befüllt. Im nächsten Schritt wurden von der Positivkontrolle, der Negativkontrolle und den zu untersuchenden Seren je 100 µl der Vorverdünnung in die zweite Zeile der ELISA-Platte aufgebracht; die erste Zeile diente zur Bestimmung des Leerwertes. Die Positivkontrolle wurde in das erste Well, die Negativkontrolle in das zweite und die Proben in ihre jeweiligen Vertiefungen der zweiten Zeile gegeben. Von Zeile B bis Zeile H wurde mit dem vorgelegten TBST eine geometrische 1:2 Verdünnungsreihe angelegt (siehe Tab. 3). Die so präparierte Platte inkubierte für 60 min bei Raumtemperatur auf einem Schüttelrotator IKA®Schüttler MTS 4 (Fa. IKA-Labortechnik, Staufen). Anschließend wurde die Platte mit Hilfe des ELISA-Washers PW 96 (Fa. Tecan, Crailsheim) einem dreimaligen Waschvorgang mit je 200µl TBST-Waschpuffer unterzogen und die Restfeuchte durch Ausklopfen aus der Platte geschlagen. Das anschließend zugesetzte Konjugat, Peroxidase-conjugated Affine Pure Goat Anti-Horse IgG (H+L) (Ziege-anti-Pferd-IgGPO) (Fa. Dianova, Hamburg) wurde mit TBST-Waschpuffer im Verhältnis 1:15.000 verdünnt und jede Kavität mit je 100 µl befüllt. Die Platte inkubierte danach für weitere 30 Minuten bei Raumtemperatur auf dem Schüttelrotator, bevor sie wiederum dreimal mit dem ELISA-Washer gewaschen und ausgeschlagen wurde. Befanden 58 Kapitel 3: Material und Methode sich im Serum spezifische Antikörper gegen das R. equi-Antigen, entstanden Immunkomplexe, an die sich das Konjugat anlagerte. Im dritten Schritt wurden in jedes Well je 100 µl des chromogenen Substrats Tetramethylbenzidin-Substratpuffer (TMB, Fa. Viro-Immun Labor-Diagnostika GmbH, Oberursel) gegeben, das mit der Peroxidase-Komponente des Konjugats reagierte. Nachdem die Platte für zehn Minuten bei Raumtemperatur inkubierte, wurde der dabei zu beobachtende Farbumschlag durch 100 µl einer 0,5 molaren Schwefelsäurelösung gestoppt. Die photometrische Messung als optische Dichte (OD) bei 450 nm erfolgte mit Hilfe des ELISA-Reader SLT-Spectra (Fa. SLT-Labinstruments Deutschland GmbH, Crailsheim). Tab. 3: Mikrotiterplattenbelegung des Rhodococcus equi-ELISA 1 2 3 4 Leer- Leer- Leer- Wert Wert Wert B + - C ↓ ↓ D Titration A 5 6 7 8 9 10 11 12 Leer- Leer- Leer- Wert Wert Wert Probe 1 Probe 2 Probe 9 Probe 10 ↓ ↓ ↓ ↓ Titration Titration E F G H + Positivkontrolle, - Negativkontrolle 3.5.5 Auswertung des ELISA Mit Hilfe der Software Magellan® (Fa. Tecan, Crailsheim) erfolgte sowohl die Bearbeitung der Rohdaten als auch die Prüfung der Validitätskriterien. Die ELISAAktivität der Proben wurde auf Grundlage der gemessenen optischen Dichte (leerwertkorrigierte Einzelwerte der Titrationsreihe) nach der Referenz- Standardmethode (FRANZ et al. 1989) in ELISA-Units (EU) ermittelt. Dafür wurden 59 Kapitel 3: Material und Methode die Extinktionswerte der Titrationsreihe des Positivserums um den Leerwert korrigiert, die Ergebnisse dekadisch logarithmiert und zur Berechnung einer Regressionsgeraden herangezogen. Die erste Verdünnungsstufe des Positivserums wurde dabei gleich 100 EU gesetzt. Anhand der für die Proben ermittelten EU-Werte konnten die entsprechenden R. equi-IgG-Konzentrationen abgeleitet werden. Die ELISA-Ergebnisse wurden als valide eingestuft, wenn die niedrigste Verdünnung der Positivkontrolle OD-Werte zwischen 1,0 und 2,5 aufwies und die jeweilige Regressionsgerade eine Steigung von tanα zwischen 0,7 und 1,1 besaß (FRANZ et al. 1989). Kriterien für die Auswertbarkeit der Serumproben waren ein Variationskoeffizient von unter 20 %, nach O.I.E.-Richtlinien (Office International des Epizooties 2004), und die Linearität und Parallelität der logarithmierten OD-Werte im Vergleich zur Positivkontrolle (siehe Abb. 3). Für die Regressionsanalyse zur Durchführung der Konzentrationsberechnung sind mindestens drei Werte notwendig. Um Fehler in der Durchführung auszuschließen und die Auswertbarkeit zu prüfen, wurden alle Serumproben, deren Ergebnisse nicht auswertbar waren, da in der Titration ein Variationskoeffizient von über 20 % vorlag, erneut im R. equi-ELISA untersucht. Darüber hinaus wurde der Gesamt-IgG-Gehalt dieser Seren im von MARKUS (2005) modifizierten IgG-ELISA nach LUFT (2000) ermittelt. 60 Kapitel 3: Material und Methode log Leerwert korrigierte OD-Werte 3,5 Positivkontrolle Fohlenserum 3,0 2,5 2,0 1,5 1,0 0,0 1,0 2,0 3,0 4,0 5,0 6,0 7,0 8,0 log Verdünnung Abb. 3: Linearität und Parallelität der logarithmierten OD-Werte in den jeweiligen Verdünnungsstufen des Positivserums und eines Fohlenserums (Kontrollgruppe). 3.6 Statistische Auswertung Gegenstand der statistischen Auswertung waren folgende Punkte: 1. Der Vergleich der Morbidität in den drei Fohlengruppen 2. Der Vergleich von Alter, klinischem Score, Konzentration der Leukozyten, Anzahl der Abszesse und Summe der Abszess-Durchmesser in den drei Fohlengruppen zum Zeitpunkt der Diagnose 3. Vergleich des Gehalts an Anti-R. equi-Immunglobulin G zu bestimmten Zeitpunkten im Untersuchungszeitraum zwischen den drei Fohlengruppen 4. Zusammenhang zwischen dem Gehalt an Anti-R. equi-Immunglobulin G acht bis zehn Stunden post natum mit dem Auftreten einer R. equi-Pneumonie und dem Erkrankungsalter. 61 Kapitel 3: Material und Methode Im Bezug auf die Morbidität wurden jeweils zwei Gruppen mit dem Fisher-Test verglichen. Die Entwicklung der Neuerkrankungen in den drei Fohlengruppen in der Darstellung nach Kaplan-Meier wurde im Wilcoxon-Test auf Unterschiede überprüft. Zum Zeitpunkt der Diagnose wurden die Gruppen hinsichtlich des Alters der Fohlen, des klinischen Scores, der Zahl der Leukozyten, der Anzahl der Abszesse, der Summe der Abszess-Durchmesser und des Anti-R. equi-Immunglobulin G-Gehalts im Kruskal-Wallis-H-Test miteinander verglichen. Die Vergleiche bezüglich des Gehalts an Anti-R. equi-IgG zu bestimmten Zeitpunkten und hinsichtlich des Parameters AUC (Area Under Curve) wurden mit dem Kruskal-Wallis-H-Test überprüft. Der Vergleich des Gehalts an Anti-R. equi-IgG acht bis zehn Stunden post natum zwischen den erkrankten und den gesunden Fohlen wurde mit dem MannWhitney U-Test untersucht. Der Zusammenhang zwischen dem Gehalt an Anti-R. equi-IgG acht bis zehn Stunden post natum und dem Erkrankungsalter wurde mit der Spearman Rangkorrelation eruiert. Die statistischen Berechnungen wurden mit dem Programmpaket STATISTICA der Firma StatSoft, Inc., Tulsa, USA von Herrn Dr. W. REIMERS, Adendorf, durchgeführt. Die Bewertung der statistischen Tests erfolgte anhand Irrtumswahrscheinlichkeit p < 0,05 (siehe Tab. 4). Tab. 4: Irrtumswahrscheinlichkeiten (p-Werte) zur Darstellung der Signifikanzen p-Wert Signifikanz p > 0,05 nicht signifikant p < 0,05 schwach signifikant * p < 0,01 signifikant ** p < 0,001 hoch signifikant *** 62 Symbol der Kapitel 4: Ergebnisse 4 Ergebnisse Ziel der vorliegenden Arbeit war zu überprüfen, ob durch die eingesetzten Impfstoffformulierungen und den intranasalen Applikationsweg ein Schutz vor einer R. equi-Pneumonie bei Fohlen aufgebaut werden kann und wie sich während des Untersuchungszeitraums im Serum der Probanden der Gehalt an Anti-R. equiImmunglobulin G (IgG) im ELISA nach HIETALA et al. (1985) verhält. Die in die Untersuchung involvierten Probanden erfüllten alle die unter 3.1.4. genannten Bedingungen zur Aufnahme in die Studie. Zwei Fohlen der Kontrollgruppe (Nr. 80 und Nr. 103) und ein Fohlen der Impfgruppe ohne Adjuvans (Nr. 4) verstarben vor Ablauf des Untersuchungszeitraums an Erkrankungen, die nicht den Respirationstrakt betrafen. Bei den im Institut für Pathologie der Stiftung Tierärztliche Hochschule Hannover an den Tieren Nr. 4 und Nr. 103 durchgeführten Sektionen konnte R. equi aus Lungenabszessen isoliert werden. Die intranasale Vakzination konnte im Zusammenhang mit anderen Maßnahmen wie der klinischen Untersuchung oder einer Punktion der Vena jugularis ohne stärkere Abwehrreaktionen bei allen Probanden durchgeführt werden. Im direkten Beobachtungszeitraum 15 Minuten nach Applikation der Lösungen und auch bei den nachfolgenden Untersuchungen der Studie wurden bei keinem Tier systemische oder lokale unerwünschte Impfreaktionen festgestellt. 4.1 Das Auftreten von Lungenabszessen 4.1.1 Morbidität Der Verdacht einer R. equi-Pneumonie wurde gestellt, wenn klinische Befunde und/oder eine Leukozytose vorlagen und mindestens ein Lungenabszess sonographisch darstellbar war. Um den Einfluss der intranasal verabreichten Lösungen zu vergleichen, wurde eruiert, wie viele Probanden der jeweiligen Gruppen solche Befunde bei der Ultraschalluntersuchung zeigten und wann sie festgestellt wurden. In der Kontrollgruppe zeigten 61,1 % (22 von 36) der Fohlen mindestens einen Lungenabszess im Ultraschall, in der Impfgruppe mit Adjuvans CpG XXXX 36,1 % 63 Kapitel 4: Ergebnisse (13 von 36) und in der Impfgruppe ohne Adjuvans 58,3 % (21 von 36). Diese Unterschiede wurden mit dem Fisher-Test miteinander verglichen und erwiesen sich nicht als statistisch signifikant (siehe Tab. 17 im Anhang). In Abbildung 4 wird präsentiert, bei welchem relativen Anteil der drei Fohlengruppen während des Untersuchungszeitraums mindestens ein sonographisch darstellbarer Lungenabszess festgestellt wurde. Morbidität [%] 70 60 50 40 30 20 10 0 Kontr.gr. R.e.m.A. R.e.o.A. Kontr.gr = Kontrollgruppe; R.e.m.A. = R. equi-Impfstoff mit Adjuvans CpG XXXX; R.e.o.A. = R. equi-Impfstoff ohne Adjuvans CpG XXXX. Abb. 4: Anteil der erkrankten Fohlen an der Gesamtpopulation der Gruppen angegeben in Prozent [%]. Kontrollgruppe n=36; Impfgruppe mit Adjuvans CpG XXXX n=36; Impfgruppe ohne Adjuvans n=36. 4.1.2 Erkrankungsalter Als Zeitpunkt der Erkrankung wurde der Tag festgelegt, an dem das erste Mal mindestens ein Lungenabszess sonographisch darstellbar war. Aus den einzelnen Daten wurde das mittlere Alter berechnet und zwischen den Gruppen verglichen (siehe Abb. 5). In der Kontrollgruppe traten Lungenabszesse durchschnittlich mit 63,8 ± 20,5 Tagen, in der Impfgruppe mit Adjuvans CpG XXXX mit 77,6 ± 27,7 Tagen 64 Kapitel 4: Ergebnisse und in der Impfgruppe ohne Adjuvans mit 91,1 ± 32,6 Tagen auf. Der paarweise Vergleich der drei Fohlengruppen zeigt, dass zwischen der Kontrollgruppe und der Impfgruppe ohne Adjuvans mit p = 0,0065 ein statistisch signifikanter Unterschied nachzuweisen ist (siehe Tab. 18 im Anhang). Alter [Tage] 140 ** 120 100 80 60 40 20 0 Kontr.gr. R.e.m.A. R.e.o.A. Kontr.gr = Kontrollgruppe; R.e.m.A. = R. equi-Impfstoff mit Adjuvans CpG XXXX; R.e.o.A. = R. equi-Impfstoff ohne Adjuvans CpG XXXX. Abb.5: Erkrankungsalter in den einzelnen Gruppen angegeben in Tagen. Kontrollgruppe n=36; Impfgruppe mit Adjuvans CpG XXXX n=36; Impfgruppe ohne Adjuvans n=36. Dargestellt sind Mittelwerte und dazugehörige Standardabweichungen. Um nicht nur das durchschnittliche Erkrankungsalter der drei Fohlengruppen, sondern auch die genaue Entwicklung über den Untersuchungszeitraum zu vergleichen, wurde die Anzahl neuerkrankter Fohlen in bestimmten Altersabschnitten betrachtet (siehe Abb. 6). Nach der vierten Lebenswoche war in der Impfgruppe ohne Adjuvans ein Fohlen erkrankt. In der fünften und sechsten Woche wiesen aus der Kontrollgruppe vier Fohlen, aus der Impfgruppe mit Adjuvans CpG XXXX zwei und aus der Impfgruppe ohne Adjuvans ein Fohlen Lungenabszesse auf. In der siebten und achten 65 Kapitel 4: Ergebnisse Lebenswoche erkrankten aus der Kontrollgruppe acht, aus der Impfgruppe mit Adjuvans CpG XXXX ein und aus der Impfgruppe ohne Adjuvans drei Tiere neu. Nach der zehnten Woche kamen als weitere Erkrankte in der Kontrollgruppe drei, in den beiden Impfgruppen jeweils zwei Fohlen hinzu. Nach der zwölften Lebenswoche wiesen vier weitere Probanden der Kontrollgruppe, drei der Impfgruppe mit Adjuvans und zwei der Impfgruppe ohne Adjuvans sonographisch darstellbare Lungenabszesse auf. In der dreizehnten und vierzehnten Lebenswoche entwickelten in den drei Gruppen jeweils zwei weitere Fohlen Abszesse in der Lunge. In der sechzehnten Lebenswoche erkrankte in der Kontrollgruppe ein und in den beiden Impfgruppen jeweils drei Probanden neu. In der siebzehnten und achtzehnten Lebenswoche kamen nur in der Impfgruppe ohne Adjuvans sieben weitere erkrankte Fohlen hinzu. Anzahl neuerkrankter Fohlen 9 8 Kontr.gr. R.e.m.A. 7 R.e.o.A. 6 5 4 3 2 1 0 4. 6. 8. 10. 12. 14. 16. 18. Alter der Fohlen in Wochen Kontr.gr = Kontrollgruppe; R.e.m.A. = R. equi-Impfstoff mit Adjuvans CpG XXXX; R.e.o.A. = R. equi-Impfstoff ohne Adjuvans CpG XXXX. Abb.6: Anzahl neuerkrankter Fohlen der drei Gruppen in bestimmten Altersabschnitten. Kontrollgruppe n=36; Impfgruppe mit Adjuvans CpG XXXX n=36; Impfgruppe ohne Adjuvans n=36. 66 Kapitel 4: Ergebnisse Um zu zeigen, wie sich diese Erkrankungsrate im Vergleich zur Gesamtpopulation der jeweiligen Gruppe verhält, wurde in Abb. 7 die Darstellung nach Kaplan-Meier gewählt. Die Kurve der Kontrollgruppe liegt deutlich unter denen der beiden Impfgruppen. Die Tiere dieser Gruppe wiesen also insgesamt eher und häufiger Abszesse auf als die Tiere der Impfgruppen. Der Unterschied war im Wilcoxon-Test mit p = 0,031 schwach signifikant. Die Kurven der beiden Impfgruppen sind bis zum 112. Tag nahezu gleich und trennen sich erst ab diesem Zeitpunkt deutlich voneinander. gesunde Fohlen [%] 110 Kontr.gr. R.e.m.A R.e.o.A. 100 90 80 70 60 50 40 30 0 14 28 42 56 70 84 98 112 126 140 Alter der Fohlen [d] Kontr.gr = Kontrollgruppe; R.e.m.A. = R. equi-Impfstoff mit Adjuvans CpG XXXX; R.e.o.A. = R. equi-Impfstoff ohne Adjuvans CpG XXXX. Abb.7: Verlauf des Anteils gesunder Fohlen in den Gruppen in Abhängigkeit vom Lebensalter nach Kaplan-Meier. Kontrollgruppe n=36; Impfgruppe mit Adjuvans CpG XXXX n=36; Impfgruppe ohne Adjuvans n=36. 67 Kapitel 4: Ergebnisse Aus der Kontrollgruppe zeigten drei Fohlen sonographisch darstellbare Lungenabszesse nach dem Untersuchungszeitraum im Alter von 134, 135 und 138 Tagen und zwei weitere Probanden erkrankten nach bereits abgeschlossener Therapie erneut. In der Impfgruppe mit Adjuvans CpG XXXX wurden bei zwei Fohlen nach Beendigung der Studie im Alter von 134 und 135 Tagen Lungenabszesse im Ultraschall erkannt. Ferner erkrankten aus der Impfgruppe ohne Adjuvans drei Fohlen nach Abschluss der Arbeit im Alter von 138, 140 und 142 Tagen und ein Fohlen nach Therapie wiederholt. 4.2 Klinische Symptome und Leukozytenkonzentration im Blut In der vorliegenden Studie wurde die sonographische Diagnostik durchgeführt, wenn klinische Befunde oder eine Leukozytose den Verdacht einer Lungenerkrankung ergaben (siehe 3.4.3). Der klinische Score nach OHNESORGE et al. (1998) quantifiziert die Ergebnisse der klinischen Untersuchung wie sie unter 3.4.1 beschrieben ist. Somit können Unterschiede hinsichtlich des durchschnittlichen Schweregrades der klinischen Symptome zum Zeitpunkt der Befunderhebung ermittelt werden. Die Probanden der drei Gruppen wiesen zu dem Zeitpunkt, an dem zum ersten Mal mindestens ein Lungenabszess im Ultraschall sichtbar geworden war, einen durchschnittlichen klinischen Score auf, der sie als geringgradig erkrankt einstufte. In der Kontrollgruppe lag er bei 2,4 mit einer Standardabweichung von 2,1, in der Impfgruppe mit Adjuvans CpG XXXX bei 3,4 mit einer Standardabweichung von 2,4 und in der Impfgruppe ohne Adjuvans bei 3,5 mit einer Standardabweichung von 2,2. Ein statistisch signifikanter Unterschied ist nicht nachzuweisen (siehe Tab. 19 im Anhang). Die Schwankungsbreite lag zwischen sieben als höchstem Wert in allen drei Gruppen und Null als niedrigstem Wert, der ebenfalls in allen Fohlengruppen vorkam. Bei der klinischen Untersuchung waren nicht bei allen erkrankten Fohlen respiratorischen die relativ Erkrankung wie unspezifischen verschärfte klinischen Symptome Atemgeräusche, Husten einer oder Nasenausfluss festzustellen. So ergab die Untersuchung im klinischen Score nach OHNESORGE et al. (1998) bei neun Fohlen aus der Kontrollgruppe, bei zwei Tieren aus der Impfgruppe mit Adjuvans CpG XXXX und bei vier Probanden aus der 68 Kapitel 4: Ergebnisse Impfgruppe ohne Adjuvans einen Wert, der sie als gesund einstuft. Abbildung 8 zeigt die mittleren Werte im klinischen Score in den drei Fohlengruppen. Klinischer Score 7 6 5 4 3 2 1 0 Kontr.gr. R.e.m.A R.e.o.A. Kontr.gr = Kontrollgruppe; R.e.m.A. = R. equi-Impfstoff mit Adjuvans CpG XXXX; R.e.o.A. = R. equi-Impfstoff ohne Adjuvans CpG XXXX. Abb.8: Klinischer Scores nach OHNESORGE et al. (1998) zum Diagnosezeitpunkt. Kontrollgruppe n=36; Impfgruppe mit Adjuvans CpG XXXX n=36; Impfgruppe ohne Adjuvans n=36. Dargestellt sind Mittelwerte und dazugehörige Standardabweichungen. Im Rahmen der klinischen Untersuchung wurde auch die Konzentration der Blutleukozyten gemessen, deren Auswertung in Abbildung 9 dargestellt ist. Die Fohlen wiesen zum Diagnosezeitpunkt Leukozytenkonzentrationen im Blut auf, die im Durchschnitt in der Kontrollgruppe bei 16,9 x 109 pro Liter Blut mit einer Standardabweichung von 3,4, in der Impfgruppe mit Adjuvans CpG XXXX bei 14,0 x 109 pro Liter Blut mit einer Standardabweichung von 3,9 und in der Impfgruppe ohne Adjuvans bei 14,7 x 109 pro Liter Blut mit einer Standardabweichung von 3,6 lagen. Es ist im paarweisen Vergleich der Probandengruppen kein statistisch signifikanter Unterschied nachzuweisen (siehe Tab. 20 im Anhang). Betrachtet man die erkrankten Probanden aus allen Gruppen, so weisen 64,3 % (36 von 56) der Fohlen eine Leukozytenkonzentration von über 13.0 x 109 pro Liter Blut auf (siehe Abb. 9). 69 Kapitel 4: Ergebnisse Leukozyten [x 109/l] 25 20 15 10 5 0 Kontr.gr. R.e.m.A R.e.o.A Kontr.gr = Kontrollgruppe; R.e.m.A. = R. equi-Impfstoff mit Adjuvans CpG XXXX; R.e.o.A. = R. equi-Impfstoff ohne Adjuvans CpG XXXX. Abb.9: Leukozytenanzahl pro Liter Blut zum Diagnosezeitpunkt. Kontrollgruppe n=36; Impfgruppe mit Adjuvans CpG XXXX n=36; Impfgruppe ohne Adjuvans n=36. Angegeben sind Dargestellt sind Mittelwerte und dazugehörige Standardabweichungen. 4.3 Ultraschallbefunde Um den Schweregrad der Ultraschallbefunde zu vergleichen, werden sowohl die Anzahl der Abszesse als auch die Summe der Abszessdurchmesser gegenübergestellt. Zum Diagnosezeitpunkt wiesen die Fohlen der Kontrollgruppe im Durchschnitt 2,2 Abszesse mit einer Standardabweichung von 1,2, in der Impfgruppe mit Adjuvans CpG XXXX 1,5 Abszesse mit einer Standardabweichung von 1,5 und in der Impfgruppe ohne Adjuvans 3,9 Abszesse mit einer Standardabweichung von 4,0 auf. Im Vergleich ist zwischen den Impfgruppen ein statistisch schwach signifikanter Unterschied nachweisbar (siehe Tab. 21 im Anhang). Die Daten streuten in der Kontrollgruppe zwischen 1 und 5, in der Impfgruppe mit Adjuvans CpG XXXX zwischen 1 und 6 und in der Impfgruppe ohne Adjuvans von 1 bis 17, wobei in der letzten Gruppe drei Fohlen besonders auffielen: Nr. 18 mit 11, Nr. 22 mit 17 und Nr. 70 Kapitel 4: Ergebnisse 64 mit 9 Abszessen. Abbildung 10 zeigt die mittlere Anzahl der Lungenabszesse in den Gruppen, die zum Zeitpunkt der Diagnose dargestellt wurden. Anzahl der Abszesse 8 7 6 * 5 4 3 2 1 0 Kontr.gr. R.e.m.A. R.e.o.A. Kontr.gr = Kontrollgruppe; R.e.m.A. = R. equi-Impfstoff mit Adjuvans CpG XXXX; R.e.o.A. = R. equi-Impfstoff ohne Adjuvans CpG XXXX. Abb.10: Abszessanzahl zum Diagnosezeitpunkt. Kontrollgruppe n=36; Impfgruppe mit Adjuvans CpG XXXX n=36; Impfgruppe ohne Adjuvans n=36. Dargestellt sind Mittelwerte und dazugehörige Standardabweichungen. Neben der Anzahl der Abszesse wird die Summe der Abszessdurchmesser betrachtet, damit Fohlen, bei denen ein einzelner Abszess mit einem großen Durchmesser dargestellt wurde, besser mit Fohlen mit mehreren kleinen Abszessen und umgekehrt zu vergleichen sind. Dieser Parameter weist bei Fohlen in der Kontrollgruppe im Durchschnitt 37 mm mit einer Standardabweichung von 30, in der Impfgruppe mit Adjuvans CpG XXXX 26 mm mit einer Standardabweichung von 25 und in der Impfgruppe ohne Adjuvans 60 mm mit einer Standardabweichung von 71 auf. Ein statistisch signifikanter Unterschied ist im Vergleich der Probandengruppen 71 Kapitel 4: Ergebnisse nicht nachzuweisen (siehe Tab. 22 im Anhang). In Abbildung 11 sind die mittleren Summen der Abszessdurchmesser dargestellt. Summe der Abszessdurchmesser [mm] 140 120 100 80 60 40 20 0 Kontr.gr. R.e.m.A. R.e.o.A. Kontr.gr = Kontrollgruppe; R.e.m.A. = R. equi-Impfstoff mit Adjuvans CpG XXXX; R.e.o.A. = R. equi-Impfstoff ohne Adjuvans CpG XXXX. Abb.11: Summe der Abszessdurchmesser zum Diagnosezeitpunkt angegeben in Millimeter [mm]. Kontrollgruppe n=36; Impfgruppe mit Adjuvans CpG XXXX n=36; Impfgruppe ohne Adjuvans n=36. Dargestellt sind Mittelwerte und dazugehörige Standardabweichungen. 4.4 Ergebnisse der ELISA-Untersuchungen Die im „California-ELISA“ nach HIETALA et al. (1985) untersuchten Serumproben wurden zu den in 3.3 genannten Zeitpunkten gewonnen und wie in 3.5 beschrieben analysiert. 4.4.1 Vergleich des Anti-R. equi-IgG-Gehalts aller Fohlen Werden die Mittelwerte im Gehalt von Anti- R. equi-IgG aller Fohlen der drei Gruppen während des Untersuchungszeitraums betrachtet, zeigt sich ein nahezu identischer Verlauf vom ersten Lebenstag bis zur siebten bzw. achten Lebenswoche auf. Bis 72 Kapitel 4: Ergebnisse einschließlich zu diesem Zeitpunkt ist kein signifikanter Unterschied zwischen den Gruppen nachzuweisen. In der neunten bzw. zehnten Lebenswoche erfährt die Kurve der Kontrollgruppe einen deutlichen Anstieg, während die Kurve der Impfgruppe mit Adjuvans CpG XXXX nur eine leichte Erhöhung zeigt und die der Impfgruppe ohne Adjuvans zu diesem Zeitpunkt abfällt. Dieser Unterschied konnte im Vergleich der drei Gruppen als statistisch schwach signifikant nachgewiesen werden (p = 0,044), jedoch nicht im paarweisen Vergleich (siehe Tab. 23 im Anhang). Von der elften bzw. zwölften Woche an fallen die Gehalte an Anti- R. equiIgG in allen drei Gruppen ab. Die Unterschiede in diesem Zeitraum sind im Vergleich der drei Gruppen ab der 15./16. Woche statistisch signifikant (p = 0,008) und bei den Beprobungen der 17./18. Woche schwach signifikant (p = 0,26). Im paarweisen Vergleich der Gruppen sind die Unterschiede zwischen der Kontrollgruppe und der Impfgruppe ohne Adjuvans statistisch signifikant (15./16. Woche) bzw. schwach signifikant (17./18. Woche) (siehe Tab. 24 und Tab. 25 im Anhang). In Serumproben, die im Zeitraum der Diagnose entnommen wurden, konnte kein signifikanter Unterschied zwischen den Gruppen nachgewiesen werden. Um nicht nur einzelne Beprobungen, sondern auch den gesamten Verlauf des mittleren Gehalts an Anti-R. equi-Immunglogulin G in den Gruppen miteinander vergleichen zu können, ist ein wichtiges Kriterium der Parameter AUC (Area Under Curve = Fläche unter der Kurve). Er ist definiert als die von der Abszisse des Koordinatensystems und der individuellen Verlaufskurve des Anti-R. equi-IgG jedes Tieres über die Zeit eingeschlossenen Fläche, angegeben in ELISA UNITS x Wochen (EU x Wochen). AUC kann für die Fälle berechnet werden, in denen zu allen zwölf Beprobungszeitpunkten eine Messung des IgG vorlag. Dies sind insgesamt 89 Fälle (25 der Kontrollgruppe; 33 der Impfgruppe mit Adjuvans; 31 der Impfgruppe ohne Adjuvans). Im Kruskal-Wallis-Test wurde kein statistisch signifikanter Unterschied zwischen den Gruppen hinsichtlich der mittleren Fläche unter der Kurve nachgewiesen (siehe Tab 26 im Anhang). In Abbildung 12 ist der durchschnittliche Gehalt an Anti-R- equi-IgG aller Fohlen der drei Gruppen während des Untersuchungszeitraums dargestellt. 73 Kapitel 4: Ergebnisse Anti-R. equi-IgG [EU] 80 Kontr.gr. 70 R.e.m.A R.e.o.A 60 50 40 30 20 10 0 0 1 2 3 4 5/6 7/8 9/10 11/12 13/14 15/16 17/18 Alter der Fohlen in Wochen Kontr.gr = Kontrollgruppe; R.e.m.A. = R. equi-Impfstoff mit Adjuvans CpG XXXX; R.e.o.A. = R. equi-Impfstoff ohne Adjuvans CpG XXXX. Abb.12: Anti- R. equi-Immunglobulin G (IgG) im Serum aller Fohlen der drei Gruppen während des Untersuchungszeitraums angegeben in ELISA UNITS [EU]. Kontrollgruppe n=36; Impfgruppe mit Adjuvans CpG XXXX n=36; Impfgruppe ohne Adjuvans n=36. Dargestellt sind die Mittelwerte. 4.4.2 Vergleich zwischen erkrankten und gesunden Fohlen Neben einem Vergleich des mittleren Gehalts an Anti-R. equi-IgG der gesamten Gruppenpopulation soll eruiert werden, ob ein Unterschied in der Immunantwort zwischen den gesunden und den erkrankten Fohlen festzustellen ist. Im Vergleich der gesunden Fohlen der drei Gruppen konnte zu keinem Beprobungstermin ein statistisch signifikanter Unterschied nachgewiesen werden. Abbildung 13 zeigt den 74 Kapitel 4: Ergebnisse durchschnittlichen Gehalt an Anti-R. equi-IgG der während des Untersuchungszeitraums gesunden Fohlen der drei Gruppen. Anti-R. equi-IgG [EU] 80 Kontr.gr. R.e.m.A R.e.o.A 70 60 50 40 30 20 10 0 0 1 2 3 4 5/6 7/8 9/10 11/12 13/14 15/16 17/18 Alter der Fohlen in Wochen Kontr.gr = Kontrollgruppe; R.e.m.A. = R. equi-Impfstoff mit Adjuvans CpG XXXX; R.e.o.A. = R. equi-Impfstoff ohne Adjuvans CpG XXXX. Abb.13: Anti- R. equi-Immunglobulin G (IgG) im Serum der gesunden Fohlen der drei Gruppen während des Untersuchungszeitraums angegeben in ELISA UNITS [EU]. Kontrollgruppe n=36; Impfgruppe mit Adjuvans CpG XXXX n=36; Impfgruppe ohne Adjuvans n=36. Dargestellt sind die Mittelwerte. Beim Vergleich der erkrankten Probanden aus den drei Fohlengruppen ist ein statistisch signifikanter Unterschied nur in der 15./16. Lebenswoche nachzuweisen (siehe Tab. 27 im Anhang). In Abbildung 14 ist der durchschnittliche Gehalt an Anti-R. equi-IgG der im Untersuchungszeitraum erkrankten Fohlen der drei Gruppen dargestellt. 75 Kapitel 4: Ergebnisse Anti-R. equi-IgG [EU] 80 Kontr.gr. 70 R.e.m.A. R.e.o.A. 60 50 40 30 20 10 0 0 1 2 3 4 5/6 7/8 9/10 11/12 13/14 15/16 17/18 Alter der Fohlen in Wochen Kontr.gr = Kontrollgruppe; R.e.m.A. = R. equi-Impfstoff mit Adjuvans CpG XXXX; R.e.o.A. = R. equi-Impfstoff ohne Adjuvans CpG XXXX. Abb.14: Anti- R. equi-Immunglobulin G (IgG) im Serum der erkrankten Fohlen der drei Gruppen während des Untersuchungszeitraums angegeben in ELISA UNITS [EU]. Kontrollgruppe n=36; Impfgruppe mit Adjuvans CpG XXXX n=36; Impfgruppe ohne Adjuvans n=36. Dargestellt sind die Mittelwerte. Im Vergleich innerhalb der Fohlengruppen zwischen den gesunden und den erkrankten Probanden wurde zu keinem Zeitpunkt und in keiner Gruppe ein statistisch signifikanter Unterschied nachgewiesen. 4.4.3 Einfluss der maternalen Antikörper Die erkrankten Fohlen sowohl der Kontrollgruppe als auch der Impfgruppe mit Adjuvans CpG XXXX wiesen einen niedrigeren durchschnittlichen Gehalt an Anti-R. 76 Kapitel 4: Ergebnisse equi-IgG acht bis zehn Stunden post natum auf als die Fohlen, die im Untersuchungszeitraum nicht erkrankten (siehe Tab. 5). Allerdings konnte in keiner Fohlengruppe ein statistisch signifikanter Unterschied nachgewiesen werden (siehe Tab. 28 im Anhang). In der Kontrollgruppe war dieser Unterschied mit p = 0,095 nur knapp nicht statistisch signifikant. Tab.5: Anti-R equi-Immunglobulin G (IgG) in ELISA UNITS [EU] acht bis zehn Stunden post natum differenziert nach den Gruppen insgesamt und unterteilt in gesunde und erkranke Fohlen. Kontrollgruppe n=36; Impfstoff mit Adjuvans CpG XXXX n=36; Impfstoff ohne Adjuvans n=36. Dargestellt sind Mittelwerte ( X ) und dazugehörige Standardabweichungen (SD). Kontr.gr. krank gesund Anzahl der Tiere (n) 22 14 X [EU] 18,9 28,6 SD 25,2 25,4 R.e.m.A krank gesund 13 22 12,9 24,0 9,1 28,9 R.e.o.A. krank gesund 21 15 22,7 19,3 32,1 21,1 Kontr.gr = Kontrollgruppe; R.e.m.A. = R. equi-Impfstoff mit Adjuvans CpG XXXX; R.e.o.A. = R. equi-Impfstoff ohne Adjuvans CpG XXXX. Bei der Kontrollgruppe ist ein statistisch signifikanter Zusammenhang zwischen dem Gehalt an Anti-R. equi-IgG acht bis zehn Stunden post natum und dem Erkrankungsalter festzustellen, der in den beiden Impfgruppen nicht nachzuweisen war (siehe Tab. 29 im Anhang). In keiner Fohlengruppe ist ein statistisch signifikanter Einfluss des Gehalts Anti-R. equi-IgG acht bis zehn Stunden post natum auf den Parameter AUC festzustellen (siehe Tab. 30 im Anhang). Es fielen 28 Serumproben auf, die auch nach Wiederholung nicht ausgewertet werden konnten, weil die gemessene optische Dichte (OD) nicht linear mit der Konzentration geringer wurde bzw. ihre Kurve nicht parallel zur Positivkontrolle verlief. Die Ursache für dieses Phänomen konnte nicht abschließend geklärt werden, da die Ergebnisse im Immunglobulin G -ELISA nach LUFT (2000) keine Hinweise ergaben. 77 Kapitel 5: Diskussion 5 Diskussion In der vorliegenden Studie wurde auf einem endemisch mit Rhodococcus equi betroffenen Warmblutgestüt untersucht, ob durch die intranasale Applikation eines formalininaktivierten Impfstoffs, dem das Adjuvans CpG XXXX zugefügt wurde, ein Schutz gegen Erkrankungen durch R. equi aufgebaut werden kann. Zu diesem Zweck erhielten 108 Fohlen nach Randomisierung entweder den aus drei virulenten R. equi-Stämmen bestehenden Impfstoff alleine, den Impfstoff mit dem Adjuvans CpG XXXX oder eine Placebolösung. 5.1 Diskussion von Material und Methode 5.1.1 Probanden Um in dieser Arbeit die Vergleichbarkeit des Patientenmaterials und die Aussagekraft der Untersuchungen zu gewährleisten, wurden Gruppengrößen von je 36 Fohlen gewählt. Da alle Tiere auf demselben Betrieb in der Zeit von Juni bis Juli 2005 geboren und zusammen mit ihren Mutterstuten unter den gleichen Bedingungen gehalten und gefüttert wurden, ist eine Beeinflussung der Ergebnisse durch diese Faktoren auszuschließen. Die Geschlechterverteilung von 46 Stut- und 62 Hengstfohlen kann als repräsentativ für die Gesamtpopulation angesehen werden. Damit sich jahreszeitliche Bedingungen nicht auf die Ergebnisse der Studie auswirken konnten, erfolgte eine randomisierte Einteilung der Gruppen. Der Beobachtungszeitraum wurde bis zur 17. bzw. 18. Lebenswoche festgelegt, da in dieser Zeit die meisten Fohlen klinische Symptome äußern (GIGUÈRE u. PRESCOTT 1997). Es lagen somit homogene Bedingungen vor, wie sie bisher in anderen Impfstudien im Zusammenhang mit R. equi mit dieser Probandenzahl und über diesen Untersuchungszeitraum nicht beschrieben wurden. CHIRINO-TREJO et al. (1987) untersuchten sechs Fohlen, die bei der ersten Impfung zwischen einer und drei Wochen alt waren, und drei gleichaltrige Kontrolltiere. Die Untersuchungen endeten sechs Wochen nach der ersten Immunisierung. VARGA et al. (1997) untersuchten insgesamt 74 gleichaltrige Fohlen, die auf drei endemisch mit R. equi betroffenen Gestüten gehalten wurden, wobei sie nur auf zwei Betrieben für den Vergleich der Wirksamkeit mit den immunisierten 78 Kapitel 5: Diskussion Tieren ungeimpfte Fohlen als Kontrolltiere ließen. Die Wirksamkeit des verabreichten Totimpfstoffs wurde bei allen Tieren in Bezug auf eine Feldinfektion in Verbindung mit der Impfung der Mutterstuten untersucht und nach neun Wochen beendet. PRESCOTT et al. (1997b) impften ebenfalls Stuten und Fohlen und wählten acht Impf- und sechs Kontrollpaare. Die Fohlen wurden im Alter von drei und fünf Wochen geimpft und über den Untersuchungszeitraum von 16 Wochen in Hinblick auf einen Schutz gegen eine Feldinfektion beobachtet. LOPEZ et al. (2003) standen fünf Fohlen, die bei der ersten Impfung acht bis fünfzehn Tage alt waren, und fünf Kontrolltiere im gleichen Alter zur Verfügung. Die Impfwirkung wurde bis zum 45. Lebenstag nicht hinsichtlich eines erzeugten Schutzes gegenüber einer Infektion mit R. equi, sondern in Bezug auf die Antikörper- und Zytokinantwort untersucht. HOOPER-McGREVY et al. (2005) wählten für ihre Untersuchung gleichaltrige Fohlen, von denen vier den Impfstoff erhielten und drei als Kontrolltiere blieben und die alle im Alter von fünf Wochen euthanasiert wurden, um anschließend eine Sektion durchzuführen. 5.1.2 Impfprotokoll In der vorliegenden Studie wurde die erste Impfung acht bis zehn Stunden post natum durchgeführt, nachdem durch Messung des Gehalts an IgG im Blut und eine klinische Untersuchung eine Störung des Allgemeinbefindens auszuschließen war. Somit wurde ein früherer Termin gewählt als in einer Studie mit einer für das virulenz-assoziierte Protein A (VapA) codierenden DNA-Vakzine, in der die Fohlen das erste Mal im Alter von acht bis fünfzehn Tagen immunisiert wurden (LOPEZ et al. 2003). Jedoch wurde die gleiche Absicht verfolgt, indem möglichst vor dem ersten Erregerkontakt eine Immunität aufgebaut und durch die intranasale Applikation eventuelle Einflüsse von maternalen Antikörpern umgangen werden sollten. Letzteres konnten LOPEZ et al. (2003) bei einigen Fohlen durch den Nachweis von VapA-spezifischen Antikörpern in der BALF nachweisen. Aus den gleichen Gründen wurde in einer nach Beginn der vorliegenden Studie veröffentlichten Untersuchung der zweite Lebenstag für eine orale Immunisierung mit einem attenuierten Lebendimpfstoff gewählt, wodurch in den Fohlen eine Immunität entwickelt wurde, die einer experimentellen Infektion standhielt (HOOPER-McGREVY et al. 2005). Für 79 Kapitel 5: Diskussion die zweite Impfung wurde in der vorliegenden Studie der 20. bis 22. Tag gewählt, da Erkrankungen regelmäßig bis zum Alter von drei Wochen auftreten (ZINK et al. 1986). Hinsichtlich der Häufigkeit und der Zeitpunkte der Wiederholungsimpfungen sind in der Literatur sehr unterschiedliche Angaben zu finden. LOPEZ et al. (2003) applizierten die DNA-Vakzine ein zeites Mal intranasal und intradermal im Alter von 22 bis 29 Tagen und anschließend das Protein VapA über die gleichen Applikationswege 30 Tage nach der ersten Impfung. HOOPER-McGREVY et al. (2005) wiederholten die oralen Impfungen im Alter von einer und drei Wochen. CHIRINO-TREJO et al. (1987) wählten für die erste orale Applikation das Alter von ein bis drei Wochen, wiederholten die Impfung zwei, drei und vier Wochen später und boten damit den Fohlen einen Schutz, der eine experimentelle Infektion abwehrte. VARGA et al. (1997) impften intramuskulär auf dem ersten Gestüt 24 Fohlen gegen R. equi und EHV-2 im Alter von drei, fünf und sieben Wochen, auf dem zweiten Gestüt 14 Fohlen gegen R. equi und EHV-2 im Alter von drei und fünf Wochen und auf dem dritten Gestüt 15 Fohlen nur gegen R. equi mit fünf und sieben Wochen. PRESCOTT et al. (1997b) impften die Probanden ebenfalls intramuskulär und im Alter von drei und fünf Wochen. Bei der vorliegenden Studie handelt es sich um die erste klinische Erprobung, bei der ein CpG-Motiv als Impfstoff-Adjuvans beim Fohlen eingesetzt wird. Das CpG-Motiv XXXX wurde ausgewählt, da es im Vergleich zu anderen auf equine Leukozyten in vitro eine stärkere stimulierende Wirkung zeigt, was sich in einer erhöhten Proliferationsrate dieser Zellen widerspiegelt (RANKIN et al. 2001). Da beim Pferd bisher keine Erkenntnisse hinsichtlich einer geeigneten Dosierung als ImpfstoffAdjuvans vorliegen, wurden 500 μg des CpG XXXX mit 109 R. equi pro ml als vielversprechende Impfstoffformulierung bei der Verabreichung von 1 ml pro Nüster gewählt. Die Entscheidung für diese Dosierung berücksichtigt Ergebnisse, die mit Dosen von 500 μg an Primaten bei intradermaler (VERTHELYI et al. 2003) und mit 100 μg im Mäusemodell bei subkutaner Verabreichung (DAVILA u. CELIS 2000) erzielt wurden. Da die intramuskuläre Verabreichung von bis zu 10 mg eines CpGMotivs beim Kalb (RANKIN et al. 2002) beschrieben wurden ohne unerwünschte Nebenwirkungen zu beobachten, waren in dieser Studie unerwünschte Reaktionen 80 Kapitel 5: Diskussion als unwahrscheinlich zu betrachten. Wie in der vorliegenden Studie setzten auch VARGA et al. (1997) einen formalininaktivierten R.equi-Impfstoff ein, fügten als Adjuvans jedoch eine Aluminiumverbindung hinzu. PRESCOTT et al. (1997b) setzten das virulenz-assoziierte Protein A (VapA) zur Immunisierung ein, konnten jedoch im Vergleich zur Kontrollgruppe keinen Schutz aufbauen, sondern es erkrankten von den geimpften Probanden im Verhältnis mehr als von den Kontrolltieren. CHIRINOTREJO et al. (1987) und HOOPER-McGREVY et al. (2005) verwendeten attenuierten R. equi-Lebendimpfstoffe und konnten damit erfolgreich eine schützende Immunität aufbauen. Die intradermale und intranasale Verabreichung einer DNAVakzine und des aufgereinigten Proteins VapA durch LOPEZ et al. (2003) brachte Erkenntnisse hinsichtlich des Applikationsweges, der Impfstoff wurde jedoch nicht im Hinblick auf eine Infektion überprüft. Die Verwendung von CpG-Motiven als Adjuvantien bei intranasaler Applikation wird vielfach in der Literatur am Mäusemodell beschrieben, um die notwendige Aktivierung des Immunsystems für die Bildung von Gedächtniszellen zu gewährleisten (McCLUSKIE et al. 2002, JIANG et al. 2003). 5.1.3 Untersuchungen der Fohlen Um die Wirksamkeit der eingesetzten Impfstoffformulierungen und den Schweregrad der Erkrankung zu beurteilen, wurden die klinischen Symptome, die Leukozytenzahl im Blut sowie die Anzahl und der Durchmesser der im Ultraschall darstellbaren Abszesse verglichen. Diese Parameter wurden im Rahmen eines Überwachungsprogramms zur Früherkennung einer R. equi-Pneumonie erfasst, das an Vorschläge von COHEN et al. (2000) für endemisch betroffene Betriebe angelehnt ist. In den bisher veröffentlichten R. equi-Impfstudien spielt die klinische Untersuchung eine zentrale Rolle. Für die endgültige Diagnose wurde vor allem der kulturelle Erregernachweis durchgeführt, zum Teil im Rahmen einer Sektion. CHIRINO-TREJO et al. (1987) wählten als Parameter für die Auswertung die klinische Untersuchung und führten drei Wochen nach der letzten Immunisierung über den Zeitraum einer Woche eine viermalige experimentelle Infektion durch Inhalation und zehn bzw. 14 Tage nach der ersten Exposition die Sektion aller in die Studie involvierten Fohlen mit histologischer und mikrobiologischer Untersuchung. 81 Kapitel 5: Diskussion VARGA et al. (1997) wählten für die Kontrolle des Impfschutzes gegenüber einer Feldinfektion die klinische Beobachtung, die serologische Untersuchung in Form der indirekten Hämagglutination und die kulturelle Untersuchung von Nasentupfern der erkrankten bzw. von Proben aus der Sektion der verendeten Probanden. PRESCOTT et al. (1997b) kombinierten die klinische Beobachtung mit der kulturellen Untersuchung klinisch auffälliger Fohlen. LOPEZ et al. (2003) richteten ihre Aufmerksamkeit auf den Nachweis von VapA-spezifischen IgG-Isotypen sowohl in der BALF als auch im Serum. Darüber hinaus wurde in der BALF mit Hilfe einer PCR ein erhöhtes Vorkommen von IFN γ- oder IL-4-sezernierenden Zellen überprüft. In der Studie von HOOPER-McGREVY et al. (2005) wurden neben den Parametern der klinischen Untersuchung, die Konzentrationen von Leukozyten, Fibrinogen und VapA-spezifischen Isotypen im Blut bzw. Serum bestimmt. Kurz nach der letzten Impfung im Alter von drei Wochen wurden alle Fohlen intratracheal infiziert und 14 Tage später euthanasiert, um eine Sektion einschließlich mikrobiologischer Untersuchung durchzuführen. Da die sonographische Untersuchung in der Literatur für die Diagnose einer R. equiPneumonie für ebenso geeignet beschrieben wird wie die radiologische Untersuchung (GIGUÈRE u. PRESCOTT 1997, RAMIREZ et al. 2004), keine Strahlenbelastung verursacht und sich außerdem auf dem Gestüt in der Früherkennung der Rhodokokkose als zuverlässig gezeigt hat (ALTHAUS 2004), wurde in der vorliegenden Studie die Sonographie als Diagnostikum einer R. equiPneumonie verwendet. Obwohl Lungenabszesse ebenfalls durch andere Erreger wie Streptococcus equi ssp. zooepidemicus hervorgerufen werden können (LAVOIE et al. 1994, RAMIREZ et al. 2004), sprechen nachfolgende Punkte für eine Beteiligung von R. equi als Ursache der sonographischen Befunde. Auf dem Gestüt wurde in den letzten drei Jahren im Tracheobronchialsekret von Fohlen mit sonographisch darstellbaren Lungenabszessen wiederholt R. equi nachgewiesen, zuletzt am 10.10.2005 (ROTHHAAR 2006, persönliche Mitteilung). Zusätzlich zeigte eine im Jahr 2003 auf dem Gestüt durchgeführte Studie sowohl in den Stallungen als auch auf den Weiden eine ubiquitäre Verbreitung von R. equi, wobei aus 80 % der Bodenproben von den Weiden der Erreger kulturell isoliert werden konnte (MEYER- 82 Kapitel 5: Diskussion HAMME 2004). Schließlich liefert die vorliegende Studie einen indirekten Erregernachweis, da alle involvierten Fohlen im Serum einen Anstieg des Antikörpergehalts im ELISA nach HIETALA et al. (1985) zeigten. Ein direkter Erregernachweis wurde in der vorliegenden Arbeit nicht durchgeführt, da sowohl die Kultur als auch die PCR nur über eine geringe Sensitivität und Spezifität verfügen (MEYER-HAMME 2004, HEYERS 2005). 5.2 Ergebnisse 5.2.1 Morbidität und Befunde In der vorliegenden Studie wurde durch den Einsatz des CpG-Motivs XXXX als Impfstoff-Adjuvans keine signifikant niedrigere Erkrankungssrate im Vergleich zur Kontrollgruppe oder zur Impfgruppe ohne Adjuvans erzeugt. Der Unterschied im Vergleich zur Kontrollgruppe war mit einer Irrtumswahrscheinlichkeit von 0,065 allerdings nur knapp nicht statistisch signifikant und darf somit als eine Tendenz interpretiert werden. Die Fohlen der Kontrollgruppe erkrankten frühzeitig in größerer Zahl, so dass sich der Morbiditätsverlauf in Abhängigkeit vom Alter statistisch schwach signifikant von denen der beiden Impfgruppen unterscheidet (siehe Abb. 7). Im Durchschnitt erkrankte die Kontrollgruppe signifikant früher als die Impfstoffgruppe ohne Adjuvans. Aufgrund des anzunehmenden ständigen Kontakts mit dem Erreger kann also für beide Vakzinegruppen eine gewisse Wirkung der Impfstoffe diskutiert werden. Jedoch hielt diese Immunität nur bei der Adjuvansgruppe auch einem erhöhten Infektionsdruck stand, wie er vermutlich zum Zeitpunkt des Aufstallens in der letzten Oktober- bzw. ersten Novemberwoche herrschte. Die Stallungen, in welche die Stuten und ihre Fohlen gebracht wurden und in denen zuvor auch kranke Fohlen untergebracht waren, wurden weder gesäubert noch desinfiziert. Da von diesem Ereignis Probanden aus allen drei Gruppen tangiert waren, blieb die Vergleichbarkeit erhalten. Aus der Impfgruppe mit Adjuvans erkrankte unter diesen Umständen nur ein Fohlen, aber aus der Impfgruppe ohne Adjuvans neun Fohlen. Dass aus der Kontrollgruppe nach dem Aufstallen kein weiteres Fohlen erkrankte, ist mit der Tatsache zu erklären, dass zu diesem Zeitpunkt bereits 61,1 % der Probanden dieser Gruppe erkrankt waren und sich somit bereits mit dem Erreger auseinandergesetzt hatten. 83 Kapitel 5: Diskussion Im Vergleich der Gruppen hinsichtlich der klinischen Symptome und der Blutleukozytenkonzentration am Tag der Diagnose „Lungenabszess“ konnte kein statistisch signifikanter Unterschied nachgewiesen werden. Die Impfgruppe ohne Adjuvans weist zum Zeitpunkt der Diagnose eine statistisch schwach signifikant höhere Abszesszahl auf als die Adjuvansgruppe. Bei differenzierter Betrachtung dieses Phänomens fällt jedoch auf, dass besonders die nach dem Aufstallen erkrankten Probanden der Gruppe C eine sehr hohe Abszesszahl aufwiesen, was wiederum durch den erhöhten Infektionsdruck zu diesem Zeitpunkt erklärt werden kann. Die Korrelation zwischen Keimmenge und Schweregrad der Befunde wird in der Studie von WADA et al. (1997) aus Sektionen von an R. equi erkrankten Fohlen bestätigt. Das Ereignis des Aufstallens berücksichtigend hat die Verabreichung eines R. equi-Totimpfstoffs mit oder ohne Adjuvans somit keinen Einfluss auf den Schweregrad der Erkrankung. Dass bei 15 der 56 erkrankten Fohlen aus allen drei Gruppen Lungenabzesse bei der Ultraschalluntersuchung erkannt wurden, obwohl sie im klinischen Score nach OHNESORGE et al. (1998) einen physiologischen Wert aufwiesen, stimmt mit den Befunden bei der Etablierung des Systems auf dem Gestüt überein (ALTHAUS 2004) und mit der Beobachtung von FALCON et al. (1985), dass die Auskultation der Lunge alleine keine zuverlässigen Hinweise auf eine R. equi-Pneumonie liefert. Die Kombination der klinischen Untersuchung mit der Messung der Leukozyten im Blut führte in dieser Studie zur Erkennung von symptomlos erkrankten Fohlen, da 14 der 15 klinisch unauffälligen Ultraschalluntersuchung eine Fohlen mit Leukozytose Lungenabszessen aufwiesen. Dieser bei Nutzen der der Leukozytenmessung in der Diagnose einer R. equi-Pneumonie wird durch die Beobachtungen von CHAFFIN et al. (2004) und GIGUÈRE et al. (2003b) bestätigt. Die bisher im Zusammenhang mit R. equi an Fohlen durchgeführten Impfstudien weisen unterschiedliche Ergebnisse auf. So konnten die Untersuchungen mit attenuiertem Lebendimpfstoff von CHIRINO-TREJO et al. (1987) und HOOPERMcGREVY et al. (2005) einen Impfschutz aufbauen, der eine experimentelle Infektion abwehrte. In beiden Studien erkrankten sämtliche Probanden der Kontrollgruppen 84 Kapitel 5: Diskussion mit signifikant stärkeren klinischen Symptomen und mussten vor dem ursprünglich geplanten Termin euthanasiert werden, während die geimpften Fohlen die Auseinandersetzung mit dem Erreger mit nur leichten bzw. ohne pathologische Befunde überstanden. In den Sektionen unterschieden sich die Befunde sehr deutlich und in der Studie von HOOPER-McGREVY et al. (2005) waren die Lungen der geimpften Fohlen sogar frei von pathologischen Veränderungen. Gleiches gilt für die aus festgelegten Lungenarealen entnommenen Proben für die Keimzahlbestimmung, wo in der Studie von HOOPER-McGREVY et al. (2005) die Kontrolltiere eine Konzentration bis zu 1011 pro Gramm aufwiesen und die geimpften Tiere nur 102. In der Studie von VARGA et al (1997) erkrankten durch eine Feldinfektion weder Fohlen aus der Gruppe, die Impfstoff gegen R. equi und EHV-2 erhielt, noch aus der R. equi-Impfgruppe. Bei den drei verstorbenen der insgesamt 21 Fohlen der Kontrollgruppen wurde in allen drei Fällen in der Sektion R. equi aus Lungenproben isoliert. Weitere fünf Fohlen wiesen klinische Symptome wie Fieber, Husten und Dyspnoe auf, R. equi wurde bei keinem Fohlen der Kontrollgruppen aus den Nasentupfern nachgewiesen. Die von PRESCOTT et al. (1997b) beschriebene Immunisierung mit dem Protein VapA konnte nicht vor einer Erkrankung mit R. equi schützen, da vier der acht geimpften Probanden erkrankten, aber nur eins der sechs Kontrolltiere. Alle erkrankten Krankheitssymptome und Fohlen einen wiesen positiven deutliche respiratorische Erregernachweis aus den Trachealspülproben auf. Die Abhängigkeit von einer Polarisierung der Immunantwort in Th1-Richtung bei der Abwehr von R. equi wird in zahlreichen Studien beschrieben. So transferierten KANALY et al. (1996) IFN-γ-sezernierende CD4+-Zellen von adulten Pferden in Mäuse, denen solche Zellen fehlen und konnten eine experimentelle Infektion abwehren. DARRAH et al. (2000) wiesen in vitro nach, dass Makrophagen, die durch das Th1-Zytokin IFN-γ aktivert wurden, den intrazellulär persistierenden Erreger vollständig eliminierten. HINES et al. (2003) zeigten einen signifikant höheren Gehalt von IFN-γ-sezernierenden CD4+- und CD8+-T-Lymphozyten bei adulten und juvenilen Pferden, die sich erfolgreich mit R. equi auseinandersetzten. Und schließlich lieferten PATTON et al. (2005) eine mögliche Erklärung für die Altersprädisposition, indem sie 85 Kapitel 5: Diskussion zeigten, dass bei neugeborenen und drei Wochen alten Fohlen eine geringere in vitro Aktivität zytotoxischer T-Zellen vorliegt als im Alter von acht Wochen (PATTON et al. 2005). Die erfolgreichen Einsätze von attenuierten Lebendimpfstoffen beim Fohlen (CHIRINO-TREJO et al. 1987, HOOPER-McGREVY et al. 2005) und die Ansätze im Zusammenhang mit einer DNA-Vakzine beim Fohlen (LOPEZ et al. 2003) und bei der Maus (HAGHIGHI et al. 2005) zeigen Möglichkeiten auf, um die zelluläre Komponente des Immunsystems für die Prophylaxe zu nutzen. Auf diesem Wege kann die von KOVARIK et al. (1999) bei Neugeborenen von Mensch und Maus nachgewiesene und daher bei allen anderen Säugetieren zu vermutende Th2Polarisierung überwunden werden. Ferner wird durch die mukosale Impfstoffapplikation der Einfluss von maternalen Antikörpern umgangen. Der Einsatz eines Lebendimpfstoffs birgt jedoch das hohe Risiko, dass sich in einem endemisch betroffenen Bestand geimpfte Tiere zu einer zusätzlichen Infektionsquelle entwickeln. CpG-Motive sind eine mögliche Alternative, denn KOVARIK et al. (1999) zeigten in vitro, dass es möglich ist durch CpG-Motive Leukozyten von juvenilen Mäusen so zu stimulieren, dass sie Th1-Zytokine sezernieren. In der Literatur wird der positive Effekt von CpG-Motiven als Impfstoff-Adjuvans bei Schweinen, Rindern und Geflügel beschrieben (KAMPSTRUP et al. 2001, RANKIN et al. 2002, VLEUGELS et al. 2002). Dass CpG-Motive stimulierend auf das equine Immunsystem wirken, wurde ebenfalls in in vitro Studien gezeigt (RANKIN et al. 2001, WATTRANG et al. 2005). In der vorliegenden Feldstudie wurde kein statistisch signifikanter Unterschied im Vergleich zu den anderen Gruppen nachgewiesen, so dass in nachfolgenden Studien zu klären ist, ob CpG-Motive ein wirkungsvolles Adjuvans beim Fohlen darstellen. Dass in dieser Studie kein vollständiger Impfschutz entwickelt wurde, kann sowohl durch die nicht ausreichende Menge des Adjuvans bedingt sein, die von den antigenpräsentierenden Zellen aufgenommen wurde, als auch durch eine Beteiligung weiterer Erreger neben R. equi an der Entstehung der Lungenabszesse. Mechanische Verluste bei der Applikation oder der Abbau durch DNAsen können zu einer Reduzierung der Ursprungsmenge des CpG-Motivs geführt haben (JOSEPH et al. 2002). Ferner kann die Menge des Adjuvans ausreichend gewesen sein, aber 86 Kapitel 5: Diskussion nicht genügend Zellen gleichzeitig das Antigen und das Adjuvans aufgenommen haben. Um sowohl den Abbau durch DNAsen als auch die gleichzeitige Aufnahme von Adjuvans und Antigen zu gewährleisten, bleibt zu klären, ob durch die Verpackung des Antigens zusammen mit einem CpG-Motiv in Liposomen oder in so genannten Virus-like Particels (JOSEPH et al. 2002, STORNI et al. 2004) bessere Erfolge zu erzielen sind bzw. dieses Verfahren auch für die intranasale Applikation zu verwenden ist. Des Weiteren ist in nachfolgenden Studien zu eruieren, ob bisher noch nicht untersuchte CpG-Motive beim Pferd eine stärker stimulierende Wirkung auf das Immunsystem ausüben. Nicht nur das Adjuvans, sondern auch das verwendete Antigen kann für die Unvollständigkeit des Schutzes verantwortlich sein. Denn trotz der bereits geschilderten hohen Wahrscheinlichkeit, dass die Lungenabszesse durch R. equi verursacht wurden, kann eine Beteiligung anderer Erreger nicht ausgeschlossen werden. Darüber hinaus ist zu überprüfen, ob die hier vorgenommene zweifache Impfung durch eine mehrfache Vakzinierung zu ersetzen ist, wie sie in anderen Impfstudien beschrieben wurde. Es ist nicht zu erwarten, dass zusätzliche intranasale Applikationen des Impfstoffs Komplikationen hervorrufen werden, da in dieser Studie die intranasale Verabreichung der Lösungen weder mit vermehrten Abwehrreaktionen der Fohlen noch mit unerwünschten lokalen oder systemischen Reaktionen verbunden war. Diese Beobachtungen stimmen mit den Beschreibungen von GROGAN u. McDONELL (2005) überein. Neben der Akzeptanz der intranasalen Applikation zeigten die 22 Pferde und Ponys in ihrer Studie weder unerwünschte Nebenwirkungen noch vermehrte Abwehrbewegungen gegen Manipulationen am Kopf. 5.2.2 Untersuchung des Anti-R. equi-IgG-Gehalts im Fohlenserum Ein weiteres Ziel dieser Studie war es, die Wirkung der eingesetzten Impfstoffformulierungen auf die humorale Immunantwort zu überprüfen. Da es sich bei dem „California“-ELISA nach HIETALA et al. (1985) um ein bereits etablitiertes Verfahren handelt (TRISKATIS 2004, PAUL 2005), das im Vergleich zu anderen serologischen Nachweisverfahren eine hohe Sensitivität (88 %) und Spezifität (59 %) aufweist (GIGUÈRE et al. 2003a), stellte seine Verwendung eine gute Ergänzung für 87 Kapitel 5: Diskussion diese Studie dar. Die Proben wurden bei der Durchführung des ELISAs titriert, um durch die Erstellung von hohen Verdünnungsstufen sicher zu sein, dass die gemessenen Extinktionen durch gebundenes IgG verursacht wurden. Beim von TRISKATIS (2004) und PAUL (2005) verwendeten Doppelansatz ohne Titration kann dies nicht abschließend eruiert werden. Die Ergebnisse der serologischen weisen ebenso wie die dargestellten Beobachtungen aus der klinischen und sonographischen Untersuchung nur vereinzelt statistisch signifikante Unterschiede zwischen den drei Fohlengruppen auf. Der beobachtete Verlauf des Gehalts an Anti-R. equi-IgG in der Kontrollgruppe entspricht den Erkenntnissen Untersuchungsmethoden unterscheiden. Ebenso und wie aus die in anderen Studien, Probentermine sich der vorliegenden wenn von wurden auch dieser die die Studie Fohlen der Kontrollgruppen in den Arbeiten von PRESCOTT et al. (1996) und PAUL (2005) mit kolostralen Antikörpern versorgt und waren wahrscheinlich vom ersten Tag an in Kontakt mit R. equi. PRESCOTT et al. (1996) stellten einen Abfall in der Menge der maternalen Antikörper bis zur vierten bis achten Woche und Paul (2005) bis zur vierten Woche fest. Mit Beginn der endogenen Antikörperbildung beschreiben die Studien einen vergleichbaren Anstieg, der auch hier beobachtet wurde mit einem Höhepunkt in der neunten bis zwölften Woche (PRESCOTT et al. 1996) bzw. zwölften Woche (PAUL 2005). VARGA et al. (1997) beobachteten bei den Kontrolltieren ebenfalls einen Anstieg zur neunten Woche hin, wo diese Studie allerdings endete. Da in den genannten und der vorliegenden Arbeit die Auseinandersetzung der Fohlen mit R. equi nur als sehr wahrscheinlich zu betrachten und ihr exakter Zeitpunkt nicht zu benennen ist, kann dieser Anstieg des Antikörpergehalts Hinweise über den Charakter der Immunantwort geben aber keine Beweise. Dieser Punkt ist ebenso zu berücksichtigen wie die Bewertung von GIGUÈRE et al. (2003a), dass zurzeit keine serologische Methode zur Verfügung stehen, die in der Diagnostik von R. equi eingesetzt werden kann. Auch PAUL (2005) wies im „California“-ELISA nach HIETALA et al. (1985) keinen signifikanten Unterschied im Gehalt an Anti-R. equi-IgG zwischen erkrankten und gesunden Probanden zum Zeitpunkt der Diagnose nach. 88 Kapitel 5: Diskussion Dass sich die Kurven der beiden Impfgruppen während des Zeitraums, in dem Reaktionen sowohl auf die erste als auch auf die zweite Impfung zu erwarten waren, nicht von der Kontrollgruppe unterscheiden, kann als Bestätigung des nicht vorhandenen Impfschutzes interpretiert werden. Es bleibt zu klären, ob bei der Adjuvansgruppe das Nichtansteigen des IgG-Gehalts für eine Polarisierung der Immunantwort in Th1-Richtung spricht. HAGHIGHI et al. (2005) berichten bei der Induktion einer Th1-Antwort gegen R. equi mit Hilfe einer DNA-Vakzine von einem Rückgang der spezifischen Antikörperkonzentration im Mäusemodell. Da hinsichtlich des Parameters AUC (Area Under Curve) kein statistisch signifikanter Unterschied nachzuweisen war, ist auch keine Aussage zu treffen, ob in einer Gruppe eine Polarisierung der Immunantwort in Th1- oder Th2-Richtung vorlag. Der signifikant höhere Gehalt an Anti-R. equi-IgG, den die Kontrollgruppe in der neunten bzw. zehnten Woche aufweist kann auf die große Zahl neuerkrankter Fohlen in der achten bzw. neunten Woche zurückgeführt werden. Ob ein aufgrund der hohen Zahl neuerkrankter Fohlen der Impfgruppe ohne Adjuvans in der 17. bzw. 18. Lebenswoche ebenfalls zu erwartender Anstieg im IgG-Gehalt hätte beobachtet werden können, war wegen des festgelegten Untersuchungszeitraums nicht zu eruieren. Im Vergleich der erkrankten Fohlen der drei Fohlengruppen untereinander war ebenso wenig zu irgendeinem Beprobungstermin ein Unterschied im Gehalt an AntiR. equi-IgG zu erkennen wie im Vergleich der kranken und gesunden Probanden innerhalb der Gruppen. Auch zum Zeitpunkt der Diagnose lag kein statistisch signifikanter Unterschied zwischen den Gruppen im Gehalt an Anti-R. equi-IgG vor. Die Intention war es, Hinweise zu bekommen, die denen von HINES et al. (2003) entsprechen, die anhand des signifikant höheren Gehalt von IFN-γ-sezernierenden CD4+- und CD8+-T-Lymphozyten bei adulten und juvenilen Pferden, die sich erfolgreich mit R. equi auseinandersetzten eine Th1-Polarisierung in der Immunantwort nachwiesen. Durch eine Impfung der Mutterstuten gegen R. equi kann der Antikörpergehalt im Kolostrum und dadurch auch im Fohlenserum gesteigert, jedoch kein Schutz gegen eine Erkrankung entwickelt werden, was in mehreren Studien bestätigt wurde (MADIGAN et al. 1991, MARTENS et al. 1992, TRISKATIS 2004). Somit wird die 89 Kapitel 5: Diskussion Beobachtung dieser Studie unterstützt, dass die Entstehung einer R. equiPneumonie in keinem statistisch signifikanten Zusammenhang steht zur Höhe des Gehaltes an Anti-R. equi-IgG im Serum der Fohlen acht bis zehn Stunden post natum. Da in der Kontrollgruppe dieser Zusammenhang jedoch nur knapp nicht statistisch signifikant ist und eine signifikante Korrelation zwischen einem höheren Gehalt an Anti-R. equi-IgG und einem späteren Erkranken der Probanden besteht, kann ein positiver Einfluss der kolostralen Antikörper für ungeimpfte Fohlen in der Auseinandersetzung mit dem Erreger nicht völlig verneint werden. Außerdem gelang es BECÙ et al. (1997) durch die Immunisierung der Stuten die Mortalität auf Gestüten mit einer repräsentativen Anzahl von Fohlen zu senken. Auch CAUCHARD et al. (2004) konnten durch Impfung der Mutterstuten die Fohlen vor einer Feldinfektion schützen. Wenn allerdings kein vollständiger Schutz gegen R. equi vermittelt werden kann, muss die Verlagerung in ein höheres Alter kritisch betrachtet werden. Auf den in erster Linie endemisch betroffenen Gestüten bedeutet dies aufgrund des erhöhten Gewichts der Fohlen eine Steigerung der Medikamentenmenge und damit der Kosten. Anhand der stark streuenden Gehalte an Anti-R. equi-IgG im Serum acht bis zehn Stunden post natum wird ersichtlich, dass die Fohlen sehr unterschiedlich mit maternalen Antikörpern ausgestattet waren. Um einen Einfluss dieser auf eine Impfstudie ausschließen zu können, ist die Verabreichung von gepooltem Kolostrum notwendig. Dies stößt jedoch bei Untersuchungen mit Probandenzahlen wie in der vorliegenden Studie an finanzielle und personelle Grenzen. Die in anderen R. equi-Impfstudien beschriebenen serologischen Untersuchungen sind aufgrund der dort verwendeten Methoden oder der anderen Termine der Immunisierungen nur bedingt mit der vorliegenden zu vergleichen. VARGA et al. (1997) setzten eine indirekte Hämagglutination ein und nahmen die Serumproben am zweiten und vierten Lebenstag und im Alter von drei, fünf, sieben und neun Wochen. Sie zogen aus dem bis zur fünften Lebenswoche in allen Fohlen abfallenden und von da an in den geimpften Fohlen signifikant höherem Antikörpergehalt im Vergleich zu den Kontrolltieren den Schluß, dass dieser Anstieg auf der endogenen Antikörperbildung beruht und auch für den Schutz verantwortlich ist. LOPEZ et al. (2003) wiesen in adulten Pferden als Reaktion auf die DNA-Vakzine einen Tag vor 90 Kapitel 5: Diskussion und 24 bzw. 45 Tage nach der ersten Impfung sowohl in der BALF als auch im Serum eine signifikante Erhöhung der VapA-spezifischen Antikörper auf, während nur zwei der fünf geimpften Fohlen auf diese Weise auf die Impfung reagierten. Diese beiden Fohlen zeigten erhöhte Gehalte an den Isotypen IgGa und IgGb, wobei es sich dabei um die veraltete Nomenklatur handelt. HOOPER-McGREVY (2005) untersuchten in den Serumproben, die nach der experimentellen Infektion über zwei Wochen jeden zweiten Tag genommen wurden nicht nur die IgG-Isotypen, sondern auch verschiedene Antigenspezifitäten hinsichtlich der virulenz-assoziierten Proteine. Da sich die Ergebnisse aber völlig von denen ihrer eigenen Studie von 2003 unterschieden, kamen sie zu dem Schluss, dass die Fraktionierung der Isotypen von zu vielen anderen Faktoren wie Infektionsweg bzw. Impfung oder Infektion abhängig ist, als dass daraus ein Zusammenhang mit der Polarisierung der Immunantwort gezogen werden kann. 5.3 Schlussfolgerungen Die vorliegende Untersuchung stellt die erste Studie dar, in der die Wirkung eines CpG-Motivs als Impfstoff-Adjuvans beim Pferd in vivo überprüft wurde. Die Ergebnisse zeigen keinen statistisch signifikanten Effekt, aber eine Tendenz, dass durch die intranasale Applikation eines formalin-inaktivierten R. equi-Totimpfstoffs zusammen mit einem CpG-Motiv bereits in den ersten drei Lebenswochen eine schützende Immunität gegen die Entstehung einer R. equi-Pneumonie aufgebaut werden kann. Die intranasale Applikation wurde von den Fohlen gut geduldet und es waren weder lokale noch systemische unerwünschte Impfreaktionen zu beobachten. Die erkrankten Fohlen aller Gruppen wiesen eine ähnliche Ausprägung der klinischen und der sonographischen Befunde auf. Trotz der unterschiedlichen Reaktionen im ELISA nach HIETALA et al. (1985) wurden statistisch signifikante Unterschiede hinsichtlich des Charakters der Immunantworten weder zwischen den Fohlengruppen noch zwischen den erkrankten und den gesunden Probanden nachgewiesen. In nachfolgenden Arbeiten bleibt zu klären, welche Dosierung eines CpG-Motivs beim Pferd eine vollständig schützende Aktivierung der zellulären Immunität bewirkt, ob es weitere oder sogar stärker immunstimulierende CpG-Motive für Pferde gibt und 91 Kapitel 5: Diskussion ob eine häufigere Applikation dieser Impfstoffformulierung einen vollständigen Schutz gegen R. equi bietet. 92 Kapitel 6: Zusammenfassung 6 Zusammenfassung Hullmann, Axel-Günther: Prophylaxe der Rhodococcus equi-Pneumonie bei Fohlen durch Vakzination mit Rhodococcus equi-Impfstoff und Adjuvans CpG XXXX In der vorliegenden Studie wurde auf einem endemisch von Rhodococcus equi betroffenen Warmblutgestüt erstmals untersucht, ob durch einen formalininaktivierten R. equi-Impfstoff in Kombination mit dem CpG-Motiv XXXX bei intranasaler Applikation ein Schutz gegen Erkrankungen durch R. equi aufgebaut werden kann. Zu diesem Zweck erhielten 108 Fohlen nach Randomisierung aus für den Untersucher verblendeten Gefäßen entweder den aus drei R. equi-Stämmen bestehenden Impfstoff alleine, den Impfstoff mit der Adjuvans CpG XXXX oder eine Placebolösung. Für die erste intranasale Applikation wurde der Zeitpunkt acht bis zehn Stunden post natum und für eine zweite der 20. bis 22. Lebenstag gewählt. Die Beobachtung der Impfwirkung erfolgte bis mindestens zur 17. bzw. 18. Lebenswoche im Rahmen eines auf dem Gestüt etablierten Überwachungsprogramms für R. equi, das neben der klinischen Lungenuntersuchung die Messung der Blutleukozyten und eine transthorakale Ultraschalluntersuchung der Lunge beinhaltet. Bei allen Probanden wurde darüber hinaus die Entwicklung des Gehalts an Anti-R. equiImmunglobulin G im von TRISKATIS (2004) modifizierten ELISA nach HIETALA et al. (1985) über die ersten 17 Lebenswochen überprüft. Im Rahmen dieser Studie wurde kein statistisch signifikanter Unterschied in der Morbidität zwischen den Fohlengruppen nachgewiesen. In der Kontrollgruppe erkrankten 61,1 % (22 von 36) der Fohlen, in der Impfgruppe mit Adjuvans CpG XXXX 36,1 % (13 von 36) und in der Impfgruppe ohne Adjuvans 58,3 % (21 von 36). Diese Ergebnisse zeigen jedoch eine deutliche Tendenz, dass durch die Verwendung eines CpG-Motivs als ImpfstoffAdjuvans ein Schutz gegen Rhodococcus equi bei Fohlen innerhalb der ersten drei Lebenswochen aufgebaut werden kann. Die für die Immunisierung gewählte intranasale Applikation wurde von 93 Kapitel 6: Zusammenfassung den Fohlen ohne weitere Abwehrreaktionen geduldet und erzeugte weder lokal noch systemisch unerwünschte Impfreaktionen. Die erkrankten Fohlen aller Gruppen zeigten eine ähnliche Ausprägung der klinischen und der sonographischen Befunde. Trotz der unterschiedlichen Reaktionen im ELISA nach HIETALA et al. (1985) wurden statistisch signifikante Unterschiede hinsichtlich des Charakters der Immunantworten weder zwischen den Fohlengruppen noch zwischen den erkrankten und den gesunden Probanden nachgewiesen. 94 Kapitel 7: Summary 7 Summary Hullmann, Axel-Günther: Prophylaxis of Rhodococcus equi-pneumonia in foals by vaccination with an Rhodococcus equi vaccine and the adjuvans CpG XXXX In the present study it was investigated for the first time if an inactivated Rhodococcus equi vaccine in combination with the adjuvans CpG XXXX was able to establish a protection against R. equi pneumonia. To achieve this 108 foals were randomisely administered either the vaccine containing three R. equi-strains, the vaccine associated with the adjuvance CpG XXXX or a placebo intranasally on a farm for warmblooded horses endemically affected by R. equi. The vials containing one of the formulations mentioned above were coded. The first intranasal application was given eight to ten hours after birth and the second on the 20th till 22nd day of life. In order to evaluate the efficiency of the vaccines the foals were monitored including clinical examination of the lungs, counting blood leukocytes and transthoracal ultrasonography of the lungs till the 17th or 18th week of life. Furthermore, the Anti-R. equi-IgG in blood of any foal was observed with the ELISA of HIETALA et al. (1985) modified by TRISKATIS (2004) for the first 17 weeks of life. No statistical significant difference concerning the morbidity was noticed between the three foal groups. In the control group there were 61,1 % (22 of 36) sick foals, in the group administrated the vaccine associated with the adjuvans CpG XXXX 36,1 % (13 of 36) and in the group without any adjuvans 58,3 % (21 of 36). Because of these results there is a clear tendency to etablish a protection against R. equi in foals in the first three weeks of life by using a CpG-motif as an adjuvans. The intranasal administration was tolerated by the foals without any resistance and no local or systemic leasions were seen. Taken together the sick foals of all groups showed similar clinical and ultrasonographic signs. Although different reaction patterns could be seen in the ELISA of HIETALA et al. (1985) no statistical significant differences were shown 95 Kapitel 7: Summary regarding the induction of adverse immune reaction between the three foal groups or sick and healthy foals. 96 Kapitel 8: Literaturverzeichnis 8 Literaturverzeichnis AINSWORTH, D.M., S.W. EICKER, A.E. YEAGER, C.R. SWEENEY, L. VIEL, D. TESAROWSKI, J.-P. LAVOLIE, A. HOFFMAN, M.R. PARADIS, S.M. REED; H.N. ERB, E. DAVIDOW u. M. NALEVANKO (1998): Associations between physical examination, laboratory, and radiographic findings and outcome and subsequent racing performance of foals with Rhodococcus equi infection: 115 cases (1984-1992). J. Am. Vet. Med. Assoc. 213, 510-515 AINSWORTH, D. (1999): Rhodococcus equi infections in foals. Equine Vet. Educ. 11, 191-198 ALTHAUS, O.P. (2004): Sonographie der Lunge: Eine Hilfe zur Früherkennung der Rhodococcus equiPneumonie beim Fohlen. Hannover, Tierärztliche Hochschule, Diss. ANZAI, T., R. WADA, A. NAKANISHI, M. KAMADA, S. TAKAI, Y. SHINDO u. S. TSUBAKI (1997): Comparison of tracheal aspiration with other tests for diagnosis of Rhodococcus equi pneumonia in foals Vet. Microbiol. 56, 335-345 ARDANS, A.A., S.K. HIETALA, M.S. SPENSLEY u. A. SANSOME (1986): Studies of naturally occuring and experimental Rhodococcus equi (Corynebacterium equi) pneumonia in foals. In: Proceedings, 32 Ann. Meet Am. Ass. Equine Pract., 129-144 ARLOTTI, M., G. TOBOLI, G.L. MOSCETELLI, G. MAGNANI, R. MASERETI, V. BORGHI, M. ANDREONI, M. LIBANORE, L. BONAZZI, A. PISCINA u. R. CIAMMARUGHI (1996): Rhodococcus equi infection in HIV-positive subjects: a retrospective analysis of 24 cases. Scand. J. Infect. Dis. 28, 463-467 BALSON, G.A., G.D. SMITH u. J.A. YAGER (1997): Immunophenotypic analysis of foal bronchoalveolar lavage lymphocytes. Vet. Microbiol. 56, 237-246 BARTON, M.D. u. K.L. HUGHES (1980): Corynebacterium equi: A review. Vet. Bull. 50, 65-80 97 Kapitel 8: Literaturverzeichnis BARTON, M.D. u. K.L. HUGHES (1981): Comparison of three techniqes for isolation of Rhodococcus (Corynebacterium) equi from contaminated sources. J. Clin. Microbiol. 13, 219-221 BARTON, M.D. u. K.L. HUGHES (1984): Ecology of Rhodococcus equi. Vet. Microbiol. 9, 65-76 BARTON, M.D. u. D.H. EMBURY (1987): Studies of the pathogenesis in Rhodococcus equi infections in foals. Aust. Vet. J. 64, 332-339 BAUER, S., G.J. KIRSCHNUNG, H. HACKER, V. REDECKE, S. HAUSMANN, S. AKIRA, H. WAGNER u. G.B. LIPFORD (2001): Human TLR9 confers responsiveness to bacterial DNA via species-specific CpG motif recognition. Immunol. 98, 9237-9242 BECÚ, T., G. POLLEDO u. J.M. GASKIN (1997): Immunprophylaxis of Rhodococcus equi pneumonia in foals. Vet. Microbiol. 56, 193-204 BELL, K.S., J.C. PHILP u. N. CHRISTOFI (1996): Identification of Rhodococcus equi using the polymerase chain reaction. Lett. Appl. Microbiol. 23, 72-74 BULL, L.B. (1924): Corynebacterial pyaemia of foals. J. Comp. Pathol. 37, 295-298 CARMAN, M.G. u. R.T. HODGES (1987): Distribution of Rhodococcus equi in animals, birds and from the environment. New Z Vet. J. 35, 114-115 CAUCHARD, J., C. SEVIN, J.J. BALLET u. S. TAOUJI (2004): Foal IgG and opsonizing anti-Rhodococcus equi antibodies after immunization of pregnant mares with a protective VapA candidate vaccine. Vet. Microbiol. 104, 73-81 CHAFFIN, M.K., R.J. MARTENS, J.G. MARTENS u. R.A. FISKE (1991): Therapeutic effects of immune plasma in foals with Rhodococcus equi pneumonia. Equine Vet. J. 12, Suppl, 23-29 CHAFFIN, M.K., N.D. COHEN u. R.J. MARTENS (2003): Evaluation of equine breeding farm management and preventative health practices as risk factors for development of Rhodococcus equi pneumonia in foals. J. Am. Vet. Med. Assoc. 222, 476-485 98 Kapitel 8: Literaturverzeichnis CHAFFIN, M.K., N.D. COHEN, R.J. MARTENS, R.F. EDWARDS, M. NEVILL u. R. SMITH (2004): Hematologic and immunophenotypic factors associated with development of Rhodococcus equi pneumonia of foals at equine breeding farms with endemic infection. Vet. Immunol. Immunpathol. 100, 33-48 CHIRINO-TREJO, J.M., J.F. PRESCOTT und J.A. YAGER (1987): Protection of foals against experimental Rhodococcus equi pneumonia by oral immunization . Can. J. Vet. Res. 51, 444-447 COHEN, N.D., M.K. CHAFFIN u. R.J. MARTENS (2000): Control and prevention of Rhodococcus equi pneumonia in foals. Compend. Contin. Educ. Pract. Vet. 22, 1062-1070 DARRAH, P.A., M.K. HONDALUS, Q. CHEN, H. ISCHIROPOULOS u. D.M. MOSSER (2000): Cooperation between reaktive oxygen and nitrogen intermediates in killing of Rhodococcus equi by activated macrophages. Infect. Immun. 68, 3587-3593 DARRAH, P.A., M.C. MONACO, S. JAIN, M.K. HONDALUS, D.T. GOLENBOCK u. D.M. MOSSER (2004): Innate immune responses to Rhodococcus equi. J. Immunol. 173, 1914-1924 DAVILA , E. u. E. CELIS (2000): Repeated administration of cytosine-phosphorothiolated guanine-containing oligonukleotides together with peptide/protein immunization results in enhanced CTL responses with anti-tumor activity. J. Immunol. 165, 539-547 DITTMER, U. u. A.R.M. OLBRICH (2003): Treatment of infectious diseases with immunstimulatory oligodeoxynucleotides containing CpG motifs. Curr. Opp. Microbiol. 6, 472-477 ELLENBERGER, M.A., M.L. KAEBERLE u. J.A. ROTH (1984): Equine humoral immune response to Rhodococcus (Corynebacterium) equi. Am. J. Vet. Res. 45, 2428-2430 ELLENBERGER, M.A. u. R.M. GENETZKY (1986): Rhodococcus equi infections: literature review. Compend. Contin. Educ. Practic. Vet. 8, 414-423 99 Kapitel 8: Literaturverzeichnis FALCON, J., B.P. SMITH, R. O´BRIEN, G.P. CARLSON u. BIBERSTEIN (1985): Clinical and radiographic findings in Corynebacterium equi pneumonia of foals. J. Am. Vet. Med. Assoc. 186, 593-599 FERNANDEZ, A.S., J.F. PRESCOTT u. V.M. NICHOLSON (1997): Protective effect against Rhodococcus equi infection in mice of IgG purified from horses vaccinated with virulence associated protein (VapA)-enriched antigens. Vet. Microbiol. 56, 187-192 FLAMINIO, M.J.B.F., B.R. RUSH u. W. SHUMAN (1999): Peripheral blood lymphocyte subpopulations and immunoglobulin concentrations in healthy foals and foals with Rhodococcus equi pneumonia. J. Vet. Intern. Med. 13, 206-212 FRANZ, B., B. WARLICH u. A. OLTROGGE (1989): ELISA for detection of porcine antibodies against Erysipelothrix rhusiopathiae. Immunobiol., Suppl. 4, 134 GIGUÈRE, S. u. J.F. PRESCOTT (1997): Clinical manifestations, diagnosis, treatment and prevention of Rhodococcus equi infections in foals. Vet. Microbiol. 56, 313-334 GIGUÈRE, S., B.N. WILKIE u. J.F. PRESCOTT (1999): Modulation of cytokine response of pneumonic foals by virulent Rhodococcus equi. Infect. Immun. 67, 5041-5047 GIGUÈRE, S., J. GASKIN, C. MILLER u. J.L. BOWMANN (2002): Evaluation of commercially available hyperimmune plasma product for prevention of naturally acquired pneumonia caused by Rhodococcus equi in foals. J. Am. Vet. Med. Assoc. 220, 59-63 GIGUÈRE, S., J. HERNANDEZ, J. GASKIN, J.F. PRESCOTT, S. TAKAI u. C. MILLER (2003a): Performance of five serological assays for diagnosis of Rhodococcus equi pneumonia in foals. Clin. Diag. Lab. Immunol. 10, 241-245 GIGUÈRE, S., J. HERNANDEZ, J. GASKIN, C. MILLER u. J.L. BOWMANN (2003b): Evaluation of white blood cell concentration, plasma fibrinogen concentration and an agar gel immundiffusion test for early identification of foals with Rhodococcus equi pneumonia. J. Am. Vet. Med. Assoc. 222, 775-781 GOODFELLOW, M. u. G. ALDERSON (1977): The actinomycete-gemus: Rhodococcus: A home for the rhodococcus complex. J. Gen. Microbiol. 100, 99-122 100 Kapitel 8: Literaturverzeichnis GOODFELLOW, M. u. D.E. MINNIKIN (1977): Norcadioform bacteria. Ann. Rev. Microbiol. 31, 159-180 GORDON, R.E. (1966): Some strains in search of a genus: Corynebacterium, Mycobacterium, Nocardia or what? J. Gen. Microbiol. 43, 329-343 GRAEVENITZ, VON, A., u. V. PÜNTER-STREIT (1995): Development of a new selective plating medium for Rhodococcus equi. Microbiol. Immunol. 39, 283-284 GRÖNDAHL, G., A. JOHANNISSON u. W. JENSEN (1997): Opsonic effect of equine plasma from different donors. Vet. Microbiol. 56, 313-334 GROGAN, E.H. u. S.M. McDONELL (2005): Behavioral responses to two intranasal vaccine applicators in horses and ponies. J. Am. Vet. Med. Assoc. 226, 1689-1693 HAECKER, G., V. REDECKE u. H. HAECKER (2002): Activation of the immune system by bacterial CpG-DNA. Immunol. 105, 245-251 HAGHIGHI H.R. u. J.F. PRESCOTT (2005): Assessment in mice of VapA-DNA vaccination against Rhodococcus equi infection. Vet. Immunol. Immunopathol. 104, 215-225 HARAKAWA, T. u. S. MORITA (1949): Observation on Corynebacterium equi isolated from abscesses occurring in the lungs of foals. Jpn. J. Vet. Sci. 11, 63-74 HARTMANN, G., R.D. WEERATNA, Z.K. BALLAS, P. PAYETTE, S. BLACKWELL, I. SUPARTO, W. L. RASMUSSEN, M. WALDSCHMIDT, D. SAJUTHI, R.H. PURCELL, H.L. DAVIS u. A.M. KRIEG (2000): Delineation of a CpG phosphorothioate oligodeoxynucletide for activating primate immune responses in vitro and in vivo. J. Immunol. 164, 1617-1624 HIETALA, S.K., A.A. ARDANS u. A. SANSOME (1985): Detection of Corynebacterium equi specific antibody in horse by enzyme-linked immunosorbent assay. Am. J. Vet. Res. 4, 13-15 101 Kapitel 8: Literaturverzeichnis HIETALA, S.K. u. A.A. ARDANS (1987): Interaction of Rhodococcus equi with phagozytic cells from Rhodococcus equi exposed and non-exposed foals. Vet. Microbiol. 14, 307-320 HIGUCHI, T., S. HASHIKURA, C. GOJO, T. INUI, S. SATOH, M. YOSHIDA, T. ISHIYAMA, H. YAMADA u. S. TAKAI (1997a): Clinical evaluation of the serodiagnostic value of enzyme-linked immunosorbent assay for Rhodococcus equi infection in foals. Equine Vet. J. 29, 274-278 HIGUCHI, T., S. HASHIKURA, S. HAGIWARA, C. GOJO, T. INUI, S. SATOH, M. YOSHIDA, M. FUJII, D. HIDAKA, S. TSUBAKI u. S. TAKAI (1997b): Isolation of virulent Rhodococcus equi from transtracheal aspirates from foals serodiagnosed by enzyme-linked immunosorbent assay. J. Vet. Med. Sci. 59, 1097-1101 HILLIDGE, C.J. (1986): Review of Corynebacterium (Rhodococcus) equi lung abscesses in foals: Pathogenesis, diagnosis and treatment. Vet. Rec. 119, 261-264 HILLIDGE, C.J. (1987): Use of Erythromycin-Rifampicin combination in treatment of Rhodococcus equi pneumonia. Vet. Microbiol. 14, 337-342 HINES, S.A., D.M. STONE, M.T. HINES, D.C. ALPERIN, D.P. KNOWLES, L.C. NORTON, M.J. HAMILTON, W.C. DAVIS u. T.C. McGUIRE (2003): Clearance of virulent but not avirulent Rhodococcus equi from the lungs of adult horses is associated with intracytoplasmatic interferon production by CD4+ and CD8+ T-Lymphocytes Clin. Diagn. Lab. Immunol. 10, 208-215 HÖHENSTEIGER, NINA (2005): Nachweis der Konzentration von Tulathromycin im Plasma und in der bronchoalveolären Lavageflüssigkeit beim Fohlen mittels Hochdruckflüssigkeitschromatographie mit zweifacher Massenspektroskopie nach i.m. Applikation, mit und ohne Kombination vom Rifampicin. Tierärztliche Hochschule Hannover, Diss. HOLTH, H. u. H. AMUNDSEN (1936): Forsatte undersökelserover baciltypene ved tuverkulose hos svinet på Ostlandet. Nor. Veterinaertidskr. 48, 2-17 HONDALUS, M.K. u. D.M. MOSSER (1994): Survival and replication of Rhodococcus equi in macrophages. Infect. Immun. 62, 4167-4175 102 Kapitel 8: Literaturverzeichnis HONDALUS, M.K. (1997): Pathogenesis and virulence of Rhodococcus equi. Vet. Microbiol. 56, 257-268 HOOPER-McGREVY, K.E., S. GIGUÈRE, B.N. WILKIE u. J.F. PRESCOTT (2001): Evaluation of equine immunoglobulin specific for Rhodococcus equi virulenceassociated proteins A and C for use in protecting foals against Rhodococcus equiinduced pneumonia. A.J.V.R. 62, 1307-1313 HOOPER-McGREVY, K.E., B.N. WILKIE u. J.F. PRESCOTT (2003): Immunglobulin G subisotype responses of pneumonic and healthy, exposed foals and adult horses to Rhodococcus equi virulence-associated proteins. Clin. Diagn. Lab. Immunol. 10, 245-351 HOOPER-McGREVY, K.E., B.N. WILKIE u. J.F. PRESCOTT (2005): Virulence-associated protein-specific serum immunoglobulin G-isotype expression in young foals protected against Rhodococcus equi pneumonia by oral immunization with virulent R. equi. Vaccine 23, 5760-5767 HORNER, A.A., N. CLINMAN, A. RONAGHY u. E. RAZ (2000): Mucosal adjuvanticity of immunostimulatory DNA sequences. Semin. Immunolpathol. 22, 133-146 HURLEY, J.R. u. A.P. BEGG (1995): Failure of hyperimmune plasma to prevent pneumonia caused by Rhodococcus equi in foals. Aust. Vet. J. 72, 418-420 IOANNOU, X.P., S.M. GOMIS, B. KARVONEN, R. HECKER, L.A. BABIUK u. S. VAN DRUNEN LITTLE-VAN DEN HURK (2002): CpG-containing oligodeoxynucleotides, in combination with conventional adjuvants, enhance the magnitude and schange the bias of the immune responses to a herpesvirus glycoprotein. Vaccine 21, 127-137 JACKS, S., S. GIGUÈRE, R.R. GRONWALL, M.P. BROWN, A. KELLY u. B.S. MERRITT (2001): Pharmacokinetics of azithromycin and concentration in body fluids and bronchoalveolar cells in foals. Am. J. Vet. Res. 62, 1870-1875 JAIN S., B.R. BLOOM u. M.K. HONDALUS (2003): Deletion of VapA encoding Virulence Associated Protein A attenuates the intracellular actinomycete Rhodococcus equi. Mol. Microbiol. 50, 115-128 103 Kapitel 8: Literaturverzeichnis JANEWAY, C. A., P. TRAVERS, M. WALPORT u. M.J. SHLOMCHIK (2002): Immunologie. Verlag Spektrum Akademischer Verlag Heidelberg, Berlin, Oxford, 5. Auflage JENSEN, H.L. (1934): Studies on saprophytic mycobacteria and corynebacteria. PROC. Linn. Soc. N.S.W. 59, 19-61 JIANG, W., H.J. BAKER u. B.F. SMITH (2003): Mucosal immunization with Helicobacter, CpG DNA and Cholera Toxin is protective. Infect. Immun. 71, 40-46 JOSEPH, A., I. LOURIA-HAYON, A. PLIS-FINAROV, E. ZEIRA, Z. ZAKAY-RONES, E. RAZ, T. HAYASHI, K. TAKABAYASHI, Y. BARENHOLZ u. E. KEDAR (2002): Liposomal immunostimulatory DNA sequences (ISS-ODN): An efficient parenteral and mucosal adjuvant for influenza and hepatitis B vaccines. Vaccine 20, 3342-3354 KAMSTRUP, S., D. VERTHELYI u. D.M. KLINMAN (2001): Response of porcine peripheral blood mononuclear cells to CpG-containing oligodeoxynucleotides. Vet. Microbiol. 78, 353-362 KANALY, S.T., S.A. HINES u. G.H. PALMER (1996): Transfer of a CD4+ Th1 cell line to nude mice effects clearance of Rhodococcus equi from the lung. Infect. Immun. 64, 1126-1132 KENNEY, D.G., S.C. ROBBINS, J.F. PRESCOTT, A. KAUSHIK u. J.D. BAIRD (1994): Development of reactive arthritis and resistance to erythromycin and rifampicin in a foal during treatment for Rhodococcus equi pneumonia. Equine Vet. J. 26, 246-248 KERTH, R. (2005): Untersuchung der Wirksamkeit von Tulathromycin bei der Behandlung von Lungenabszessen bei Fohlen. Tierärztliche Hochschule Hannover, Diss. KOHLER, A.K., D.M. STONE, M.T. HINES, B.A. BYRNE, D.C. ALPERIN, L.K. NORTON u. S.A. HINES (2003): Rhodococcus equi secreted antigens are immunogenic and stimulate a type 1 recall response in the lung of horses immune to Rhodococcus equi infection. Infect. Immun. 71, 6329-6337 104 Kapitel 8: Literaturverzeichnis KOVARIK, J. u. C.A. SIEGRIST (1998): Immunity in early life. Immunol. Today 19, 150-152 KOVARIK, J., P. BOZOTTI, L. LOVE-HOMAN, M. PIHLGREN, J.L. DAVIS, P.-H. LAMBERT, A.M. KRIEG u. C.-A. SIEGRIST (1999): CpG oligodeoxynucleotides can circumvent the Th2 polarization of neonatal responses to vaccines but may fail to fully redirect Th2 responses established by neonatal priming. J. Immunol. 162, 1611-1617 KRASIL`NIKOV, N.A. (1966): Keys to the Actinomycetales. Isr. Programme Sci. Trans., Jerusalem KRIEG, A.M., A.-K. YI, S. MATSON, T.J. WALDSCHMIDT, G.A. BISHOP, R. TEASDALE, G. KORETZKY u. D. KLINMAN (1995): CpG motifs in bacterial DNA trigger direct B-cell activation. Nature 374, 546-549 KRIEG, A.M., T. WU, R. WEERATNA, S.M. EFLER, L. LOVE-HOMAN, L. YANG, A.K. YI, D. SHORT u. H.L. DAVIS (1998a): Sequence motifs in adenoviral DNA block immune activation by stimulatory CpG motifs. Proc. Natl. Acad. Sci. USA 95, 12631-12636 KRIEG, A.M., T. WU, R. WEERATNA, S.M. EFLER, L. LOVE-HOMAN, L. YANG, A.K. YI, D. SHORT u. H.L. DAVIS (1998b): CpG DNA induces sustained IL-12 expression invivo and resistance to Listeria monocytogenes challenge. J. Immunol. 161, 2428-2434 KRIEG, A.M. (2002): CpG motifs in bacterial DNA and their immune effects. Annu. Rev. Immunol. 20, 709-760 KRUTZIK, S. R., P. A. SIELING u. R. L. MODLIN (2001): The role of Toll-like receptors in host defense against microbial infection. Curr. Opin. Immunol. 13, 104-108 LADRON, N., M. FERNANDEZ, J. AGUERO, B. GONZALEZ ZORN, J.A. VAZQUEZBOLAND u. J. NAVAS (2003): Rapid identification of Rhodococcus equi by a PCR assay targeting the choE gene. J. Clin. Microbiol. 41, 3241-3245 LAVOIE, J.P., I. FISET u. S. LAVERTY (1994): Review of 40 cases of lung abscesses in foals and adult horses. Equine Vet. J. 26, 348-352 105 Kapitel 8: Literaturverzeichnis LEMAITRE, B., E. NICOLAS, L. MICHAUT, J.M. REICHHART u. J.A. HOFFMANN (1996): The dorsoventral regulatory gene cassette spatzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell 86, 973–983. LOPEZ, A.M., M.T. HINES, G.H. PALMER, D.C. ALPERIN u. S.A. HINES (2002): Identification of pulmonary T-Lymphocyte and serum antibody isotype responses associated with protection against Rhodococcus equi. Clin. Diagn. Lab. Immunol. 9, 1270-1276 LOPEZ, A.M., M.T. HINES, G.H. PALMER, D.P. KNOWLES, D.C. ALPERIN u. S.A. HINES (2003): Analysis of anamnestic immune responses in adult horses and priming in neonates induced by a DNA vaccine expressing the VapA gene of Rhodococcus equi. Vaccine 21, 3815-3825 LUFT, C.S. (2000): Untersuchungen zur systemischen Verfügbarkeit von Immunglobulin G beim neugeborenen Fohlen. Diss. Uni-München LUEHRMANN, A., N. MAUDER, T. SYDOR, E. FERNANDEZ-MORA, J. SCHULZELUEHRMANN, S. TAKAI u. A. HAAS (2004): Necrotic death of Rhodococcus equi-infected macrophages is regulated by virulenceassociated plasmids. Infect. Immun. 72, 853-862 MADIGAN, J.E., S.K. HIETALA u. N.S. MULLER (1991): Protection against naturally acquired Rhodococcus equi pneumonia in foals by administration of hyperimmune plasma. J. Reprod. Fert. 44, Suppl, 571-578 MAGNUSSON, H. (1923): Spezifische infektiöse Pneumonie beim Fohlen. Ein neuer Eitererreger beim Pferd. Arch. Wiss. Prakt. Tierheilkd. 50, 22-38 MAGNUSSON, H. (1938): Pyaemia in foals caused by Corynebacterium equi. Vet. Rec. 50, 1359-1468 MARKAI, L., S. TAKAI, M. TAMURA, A. TSUKAMOTO, R. SEKIMOTO, Y. SASAKI, T. KAKUDA, S. TSUBAKI, J. VARGA, L. FODOR, N. SOLYMOSI u. A. MAJOR (2002): Characterization of plasmid types in Rhodococcus equi isolates from foals, pigs, humans and soil in Hungary. Vet. Microbiol. 88, 377-384 106 Kapitel 8: Literaturverzeichnis MARKUS, G. (2005): Bestimmung der Immunglobulin G-Konzentration mittels Refraktometrie, Kolostrometrie und ELISA-Technik sowie Untersuchung weiterer Inhaltsstoffe im Stutenkolostrum. Tierärztliche Hochschule Hannover, Diss. MARTENS, R.J., R.A. FISKE u. H.W. RENSHAW (1982): Experimental subacute foal pneumonia induced by aerosol administration of Corynebacterium equi. Equine Vet. J. 14, 111-116 MARTENS, R.J., J.G. MARTENS, R.A. FISKE u. S.K. HIETALA (1989): Rhodococcus equi foal pneumonia: Protective effects of immune plasma in experimentally infected foals. Equine Vet. J. 21, 249-255 MARTENS, R.J., J.G. MARTENS u. R.A. FISKE (1992): Failure of passive immunisation by colostrum from immunised mares to protect foals against Rhodococcus equi pneumonia. Equine Vet. J. 12, Suppl., 19-22 MARTENS, R.J., N.D. COHEN, M.K. CHAFFIN, S. TAKAI, C.L. DOHERTY, A.B. ANGULO u. R.F. EDWARDS (2002): Evaluation of 5 serologic assays to detect Rhodococcus equi pneumonia in foals. J. Am. Vet. Med. Assoc. 221, 825-833 MARTENS, R.J., N.D. COHEN, S.L. JONES, T.A. MOORE u. J.F. EDWARDS (2005): Protective role of neutrophils in mice experimentally infected with Rhodococcus equi. Infect. Immun. 73, 7040-7042 McCLUSKIE, M.J., R.D. WEERATNA, P.J. PAYETTE u. H.L. DAVIS (2002): Parenteral and mucosal prime-boost immunization strategies in mice with hepatitis B surface antigen and CpG DNA. FEMS Immunol. Med. Microbiol. 32, 179-185 MEDZHITOV, R. u. C.A. JANEWAY (1997): Innate immunity: the virtues of nonclonal system of recognition. Cell. 91, 295-298 MEYER-HAMME, M.B. (2004): Rhodococcus equi und Streptococcus equi ssp. Zooepidermicus aus Nasentupfern und Tracheobronchialsekret bei lungenkranken Fohlen. Tierärztliche Hochschule Hannover, Diss. 107 Kapitel 8: Literaturverzeichnis MIESSNER, K.K. u. R, WETZEL (1923): Corynebacterium pyogenes (equi) als Erreger einer infektiösen, abszedierenden Pneumonie der Fohlen. Dtsch. Tierärtl. Wochenschr. 31, 449-454 MORENO-FIERROS, L., E.J. RUIZ-MEDINA, R. ESQUIVEL, R. LÓPEZ-REVILLA u. S. PINA-CRUZ (2003): Intranasal Cry1Ac Protoxin is an effective mucosal and systemic carrier and adjuvant of Streptococcus pneumoniae polysaccharides in mice. Scan. J. Immunol. 57, 45-55 NAKAZAWA, M., Y. ISAYAMA u. M. KASHIWAZAKI (1987): Diagnosis of Rhodococcus equi infection in foals by the agar gel diffusion test with protein antigen. Vet. Microbiol. 15, 105-119 O´BRIEN, R.T. u. D.S. BILLER (1997): Field imaging of the respiratory tract. Vet. Clin. N. Am. Equine Pract. 13, 487-499 OHNESORGE, B., C. TRÖTSCHEL u. E. DEEGEN (1998): Bestimmung von Totraum und expiratorischem Mischluftvolumenverhältnis zur Diagnostik chronischer Lungenerkrankung Pferdeheilkd. 14, 450-455 OLDFIELD, C., H. BONELLA, L. RENWICK, H.I. DODSON, G. ALDERSON u. M. GOODFELLOW (2004): Rapid determination of VapA/VapB genotype in Rhodococcus equi using a differential polymerase chain reaction methode. Antonie Van Leeuwenhoek. 85, 317-326 PARK, A.Y. u. P. SCOTT (2001): Il-12: keeping cell-mediated immunity alive. Scand. J. Immunol. 53, 529-532 PATTON, K.M., T.C. McGUIRE, D.G. FRASER u. S.A. HINES (2004): Rhodococcus equi-infected macrophages are recognized and killed by CD8+T Lymphocytes in a major histocompatibility complex class I-unrestricted fashion. Infect. Immun. 72, 7073-7083 PATTON, K.M., T.C. McGUIRE, M.T. HINES, R.H. MEALEY u. S.A. HINES (2005): Rhodococcus equi-specific cytotoxic T lymphocytes in immune horses and development in asymptomatic foals. Infect. Immun. 73, 2083-2093 108 Kapitel 8: Literaturverzeichnis PAUL, M. (2005): Verlauf des Rhodococcus equi-Antikörpertiters beim Fohlen: Vergleich von Fohlen mit und ohne Anti-Rhodococcus equi-Hyperimmunserumgabe Tierärztliche Hochschule Hannover, Diss. PERKINS, G.A., A. YEAGER, H.N. ERB, D.V. NYDAM, T.J. DIVERS u. J.L. BOWMAN (2002): Survival of foals with experimentally induced Rhodococcus equi infection given either hyperimmune plasma containung Rhodococcus equi antibody or normal equine plasma. Vet. Ther. 2, 334-346 PERSING D.H. (1993): Diagnostic molecular microbiology. Current challenges and future directions. Diagn. Microbiol. Infect. Dis. 16, 159-163 PILTZ, K. (2004): Vergleichende Behandlung von Rhodococcus equi-Pneumonien bei Fohlen mit Azithromycin und Rifampicin in Kombination mit Erythromycin bzw, Trimethoprim/Sulfadiazin. Tierärztliche Hochschule Hannover, Diss. PLUM, N. (1940): On corynebacterial infections in swine. Preliminary report, Cornell Vet. 30, 14-20 PRESCOTT, J.F. (1991): Rhodococcus equi: an animal and human pathogen. Clin. Microbiol. Rev. 4, 20-34 PRESCOTT, J.F., A.S FERNANDEZ, V.M. NICHOLSON, M.C. PATTERSON, J.A. YAGER, L. VIEL u. G. PERKINS (1996): Use of a virulence-associated protein based enzyme-linked immunosorbent assay for Rhodococcus equi serology in horses. Equine Vet. J. 28, 344-349 PRESCOTT, J.F., M.C. PATTERSON, V.M. NICHOLSON, B. MOREIN u. J.A. YAGER (1997a): Assessment of the immunogenic potential of Rhodococcus equi virulence associated protein (VapA) in mice. Vet. Microbiol. 56, 213-225 PRESCOTT, J.F., V.M. NICHOLSON, M.C. PATTERSON, M.C. ZANDONA MELEIRO, A. CATERINO DE ARAUJO, J.A. YAGER u. M.A. HOLMES (1997b): Use of Rhodococcus equi virulence-associated protein for immunization of foals against R. equi pneumonia. Am. J. Vet. Res. 58, 356-359 109 Kapitel 8: Literaturverzeichnis PRIEBE, G.P., M.M. BRINIG, K. HATANO, M. GROUT, F.T. COLEMAN, G.B. PIER u. J.B. GOLDBERG (2002): Construction and characterization of a live, attenuated aroA deletion mutant of Pseudomonas aeruginosa as acandidate intranasal vaccine. Infect. Immun. 70, 1507-1517 RAJAGOPALAN, V.R. (1936): Pneumonia in foals due to Corynebacterium equi. Indian Vet. J. Sci. Anim. Husb. 7, 38-53 RAMIREZ, S., G.D. LESTER u. G.R. ROBERTS (2004): Diagnostic contribution of thoracic ultrasonography in 17 foals with Rhodococcus equi pneumonia. Vet. Radiol. Ultrasound 45, 172-176 RANKIN R., R. PONTAROLLO, X. IOANNOU, A.M. KRIEG, R. HECKER, L.A. BABIUK u. S. VAN DRUNEN LITTLE-VAN DEN HURCK (2001): CpG motiv identification for veterinary and laboratory species demonstrates that sequence recognition is highly conserved. Antisense Nucleic Acid Drug Dev. 11, 333-340 RANKIN R., R. PONTAROLLO, S. GOMIS, B. KARVONEN, P. WILLSON, B.I. LOEHR, D.L. GODSON, L.A. BABIUK, R. HECKER u. S. VAN DRUNEN LITTLEVAN DEN HURCK (2002): CpG-containung oligodeoxynucleotides augment and switch the immune responses of cattle to bovine herpesvirus-1 glycoprotein D. Vaccine 20, 3014-3022 SCHMIEDHOFFER, S. (1922): Über eine infektiöse eitrige Lungenentzündung bei Saugfohlen. D. Tierärztl. Wochenschrift, S. 613 SCHWARZ, K., T. STORNI, V. MANOLOVA, A. DIDIERLAURENT, J.C. SIRARD, P. RÖTHLISBERGER u. M.F. BACHMANN (2003): Role of Toll-like receptors in costimulating cytotxic T cell responses. Eur. J. Immunol. 33, 1465-1470 SELBITZ, H.J. (2002): Infektionen mit Rhodococcus equi. In: Rolle, M., MAYR, A. (Hrsg.): Medizinische Mikrobiologie, Infektions- und Seuchenlehre. Ferdinand Enke Verlag, Stuttgart, 7. Auflage, 553-554 SELLON, D.C., T.E. BESSER, S.L. VIVRETTE u. R.S. McCONNICO (2001): Comparison of Nucleic Acid Amplification, Serology and Microbiologic Culture for Diagnosis of Rhodococcus equi Pneumonia in Foals. J. Clin. Microbiol. 39, 1289-1293 110 Kapitel 8: Literaturverzeichnis SESTER, D.P., S. NAIK, S.J. BEASLEY, D.A. HUME u. K.J. STACEY (2000): Phosphorothioate backbone modification modulates macrophage activation by CpG DNA. J. Immunol. 165, 4165-4173 SMITH, J.E. u. S.S. JANG (1980): Isolation of Corynebacterium equi from a foal with an ulcerated leg wound and a pectoral abscess. J. Am. Vet. Assoc. 177, 623-624 STEDE, Y. VAN DER, F. VERDONCK, S. VANCAENEGHEM, E. COX u. B.M. GODDEERIS (2002): CpG-oligodinucleotides as an effective adjuvant in pigs for intramuscular immunizations. Vet. Immunol. Immunopathol. 86, 31-41 STORNI, T., C. RUEDL, K. SCHWARZ, R.A. SCHWENDENER, W.A. RENNER u. M.F. BACHMANN (2004): Nonmethylated CpG motifs packaged into virus-like particles induce protective cytotoxic T cell responses in the absence of systemic side effects. J. Immunol. 172, 1777-1785 SUZUKI, K., K. KANEKO u. K. KOMAGATA (1981): Desoxyribonucleic acid homologies among coryneform bacteria. Int. J. Syst. Bacteriol. 31, 131-138 SWEENEY, C.R., R.W. SWEENEY u. T.J. DIVERS (1987): Rhodococcus equi Pneumonia in 48 Foals: Response to Antimicrobial Therapy. Vet. Microbiol. 14, 329-336 TAKAI, S., S. KAWAZU u. S. TSUBAKI (1985): Enzyme-linked immunosorbent assay for (Rhodococcus) equi infection. Am. J. Vet. Res. 46, 2166-2170 diagnosis of Corynebacterium TAKAI, S. u. S. TSUBAKI (1985): The incidence of Rhodococcus equi in domestic animal and soil. Jpn. J. Vet. Sci. 47, 493-496 TAKAI, S., T. FUJIMORI, K. KATSUZAKI u. S. TSUBAKI (1987): Ecology of Rhodococcus equi in horses and their environment on horse breeding farms. Vet. Microbiol. 14, 233-239 TAKAI, S., K. KOIKE, S. OHBUSHI, C. IZUMI u. S. TSUBAKI (1991): Identification of 15- to 17 kilodalton antigens associated with virulent Rhodococcus equi. J. Clin. Microbiol. 29, 439-443 111 Kapitel 8: Literaturverzeichnis TAKAI, S., Y. SASAKI, T. IKEDA, Y. UCHIDA, S. TSUBAKI u. T. SEKIZAKI (1994): Virulence of Rhodococcus equi isolates from patients with and without AIDS. J. Clin. Microbiol. 94, 457-460 TAKAI, S., Y. SASAKI u. S. TSUBAKI (1995): Rhodococcus equi infections in foal. Current concepts and implication for future research. J. Equine Vet. Sci. 6, 105-119 TAKAI, S., D. HIDAKA, F. MASUNOBU et al. (1996): Serum antibody responses of foals to virulence-associated 15- to 17-kilodalton antigens of Rhodococcus equi. Vet. Microbiol. 52, 63-71 TAKAI, S. (1997): Epidemiology of Rhodococcus equi infections: A review. Vet. Microbiol. 56, 167-176 TAKAI, S., G. VIGO, H. IKUSHIMA. T. HIGUCHI, S.T. HAGIWARA and Y. SASAKI, S. HASHIKURA, S. TSUBAKI, T. ANZAI, u. M. KAMADA (1998): Detection of virulent Rhodococcus equi in tracheal aspirate samples by polymerase chain reaction for rapid diagnosis of Rhodococcus equi pneumonia in foals. Vet. Microbiol. 61, 59-69 TAKAI, S., S.A. HINES, T. SEKIZAKI, V.M. NICHOLSON, D.A. ALPERIN, M. OSAKI, D. TAKAMATSU, M. NAKAMURA, K. SUZUKI, N. OGINO, T. KAKUDA, H. DAN u. J.F. PRESCOTT (2000): DNA sequences and comparison of virulence plasmids from Rhodococcus equi ATCC 33701 and 103. Infect. Immun. 68, 6840-6847 TIZARD, I. R. (2000): Veterinary Immunology: An Introduction. 6th ed. TOKUNAGA, T., S. YAMAMOTO u. K. NAMBA (1988): A synthetic single-stranded DNA, poly(dG,dC), induces interferon-α/β and –γ, augments natural killer activity, and suppresses tumor growth. Jpn. J. Cancer Res. 79, 682-686 TRISKATIS, A.-L. (2004): Semiquantitative Bestimmung von Antikörpern gegen Rhodococcus equi in Serum und Kolostrum bei Stuten und Fohlen mittels ELISA und der Vergleich mit Befunden der Lungenuntersuchung. Hannover, Stiftung Tierärztliche Hochschule, Diss. 112 Kapitel 8: Literaturverzeichnis UNDERHILL, D.M. u. A. OZINSKY (2002): Toll-like receptors: key mediators of microbe detection. Curr. Opin. Immunol. 14, 103-110 UNGEMACH, F.R., H. BODE, A. BÖTTNER, H. BOTTERMANN, L. GOSSENS, M. KIETZMANN, R. KROKER, S. SCHÜLLER, K. SIMON, L.H. WIELER u. G. WITTKOWSKI (2000): Leitlinien für den sorgfältigen Umgang mit antimikrobiell wirksamen Tierarzneimitteln. Dt. Tierärzteblatt Nov. 2000 VANNIASINKAM, T., M.D. BARTON. u. M.W. JEUZENROEDER (2005): Immune response to vaccines based upon the VapA protein of the horse pathogen, Rhodococcus equi, in a murine model. Int. J. Med. Microbiol. 294, 437-445 VARGA, J., L. FODOR, M. RUSIVA, I. SOÓS u. L. MAKRAI (1997): Prevention of Rhodococcus equi pneumonia of foals using two different inactivated vaccines. Vet. Microbiol. 56, 205-212 VENGUST, M., H. STAEMPFLI, J.F. PRESCOTT (2002): Rhodococcus equi pleuropneumonia in an adult horse. Can. Vet. J. 43, 706-708 VERTHELYI, D., M. GURSEL, R.T. KENNEY, J.D. LIFSON, S. LIU, J. MICAN u. D.M. KLINMAN (2003): CpG Oligodeoxynucleotides protect normal and SIV-infected Macaques from Leishmania infection. J. Immunol. 170, 4717-4723 VLEUGELS, B., C. VERVERKEN u. B.M. GODDEERIS (2002): Stimulatory effect of CpG sequences on humoral response in chicken. Poult. Sci. 81, 1317-1321 WADA, R., M. KAMADA, T. ANZAI, A. NAKANISHI, T. KANEMARU, S. TAKAI u. S. TSUBAKI (1997): Pathogenicity and virulence of Rhodococcus equi in foals following intratracheal challenge. Vet. Microbiol. 56, 301-312 WAGNER, B., G. OVERESCH, A.S. SHEORAN, M.A. HOLMES, C. RICHARDS, W. LEIBOLD u. A. RADBRUCH (1998): Organization of the equine immunoglobulin constant heavy chain genes. III. Alignment of c mu, c gamma, g epsilon and c alpha genes. Immunobiol. 199; 105-118 113 Kapitel 8: Literaturverzeichnis WATTRANG, E., M. BERG u. M. MAGNUSSON (2005): Immunostimulatory DNA activates produktion of type I interferons and interleukin-6 in equine peripheral blood mononuclear cells in vitro. Vet. Immunol. Immunopathol. 107, 265-279 WERNETTE, C.M., B.F. SMITH, Z.L. BARKSDALE, R. HECKER u. H.J. BAKER (2002): CpG oligodeoxynucleotides stimulate canine and feline immune cell proliferation. Vet. Immunol. Immunopathol. 84, 223-236 WOOLCOCK, J.B., A.- M.T. FARMER u. M.D. MUTIMER (1979): Selective medium for Corynebactrium equi Isolation. J. Clin. Microbiol. 21, 640-642 WOOLCOCK, J.B., A.M. FARMER u. M.D. MURTIMER (1980): Epidemiology of Corynebacterium equi in horses Res. Vet. Sci. 28, 87-90 WU, H.Y., E.B. NIKOLOVA, K.W. BEAGLEY, J. H. ELDRIDGE u. M.W. RUSSELL (1997): Development of antibody-secreting cells and antigen-specific T cells in cervical lymph nodes after intranasal immunization. Infect. Immun. 65, 227-235 YAGER, J.A. (1987): The pathogenesis of Rhodococcus equi pneumonia in foals. Vet. Microbiol. 14, 225-232 ZINK, M.C., J.A. YAGER, J.F. PRESCOTT u. B.N. WILKIE (1985): In vitro phagozytosis and killing of Corynebacterium equi by alveolar macrophages of foals. Am. J. Vet. Res. 46, 2171-2174 ZINK, M.C., J.A. YAGER u. N.L. SMART (1986): Corynebacterium equi infections in horses, 1958-1984: a review of 131 cases. Can. Vet. J. 27, 213-217 114 Kapitel 9: Anhang 9 Anhang Rezepte Coatingpuffer (150 mmol Carbonat-Bicarbonat-Puffer) Rezeptur: 5,088 g Natriumcarbonat (Na2CO3) 8,568 g Natriumbicarbonat (NaHCO3) ad 1000,0 ml Aqua dest. pH-Wert kontrollieren (zwischen 9,4-9,8) Lagerung bei 20°C ± 3°C, Haltbarkeit 1 Woche 10fach Tris buffered saline Rezeptur: 80,0 g Natriumchlorid (NaCl) 2,0 g Kaliumdihydrogenphosphat (KH2PO4) 11,4 g Dinatriumhydrogenphosphat (Na2HPO4) 2,0 g Kaliumchlorid (KCl) ad 1000,0 ml Aqua dest. Lagerung bei 20°C ± 3°C, Haltbarkeit 1 Woche Waschpuffer (TBST) Rezeptur: 100 ml 10fach Tris buffered saline 0,5 ml Tween® 20 (AppliChem, Darmstadt) ad 1000,0 ml Aqua dest. Lagerung bei 20°C ± 3°C, Haltbarkeit 1 Woche 115 Kapitel 9: Anhang Tab.6: Tag der Diagnose, Ergebnisse der klinischen, sonographischen Untersuchung und die gemessene Leukozytenkonzentration am Tag der Diagnose und der Gehalt im Serum an Anti-Rhodococcus equi-Immunglobulin G im entsprechenden Altersabschnitt angegeben in ELISA UNITS [EU] der Fohlen 1 bis 18. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). n.a. = nicht auswertbar Anzahl der Abszesse Summe der Abszesstie fen [mm] Anti-R. equi-IgG [EU] Fohlennr. Gruppe Datum Alter [d] klin. Score Leukozyte n x109 pro l Blut 1 B 27.09.2005 76 2 9,1 1 10 36,1 2 A 20.09.2005 89 1 13,7 1 10 21,0 3 B 19.09.2005 79 0 16,3 1 10 19,1 4 C 26.08.2005 63 6 21,3 6 90 6,2 5 B 16.08.2005 37 5 23,6 1 20 1,8 6 A 7 A 8 C 9 B 10 B 11 A 12 B 13 B 12.09.2005 63 3 14,7 1 25 n.a. 14 A 15 A 15.08.2005 52 3 20,8 5 110 n.a. 16 A 06.10.2005 97 3 16,7 1 10 2,8 17 B 18 C 07.11.2005 130 5 7,4 11 130 3,5 116 Kapitel 9: Anhang Tab.7: Tag der Diagnose, Ergebnisse der klinischen, sonographischen Untersuchung und die gemessene Leukozytenkonzentration am Tag der Diagnose und der Gehalt im Serum an Anti-Rhodococcus equi-Immunglobulin G im entsprechenden Altersabschnitt angegeben in ELISA UNITS [EU] der Fohlen 19 bis 36. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). Leukozyten 9 Anzahl der Abszesse Summe der Abszesstie fen [mm] Anti-R. equi-IgG [EU] Datum Alter [d] klin. Score x10 pro l Blut C 07.11.2005 127 1 13,4 4 70 3,8 21 C 29.08.2005 44 5 21,0 2 45 7,4 22 C 07.11.2005 114 5 12,8 17 330 4,0 23 A 30.9.2005 82 2 17,0 1 10 21,8 24 C 02.08.2005 51 2 15,9 1 20 2,5 25 B 26 B 27 C 31.10.2005 117 2 9,1 5 85 2,4 28 C 29 A 30 C 19.08.2005 66 0 15,4 1 10 9,2 31 A 06.10.2005 109 5 12,7 3 38 34,8 32 B 02.08.2005 38 5 9,8 3 95 1,5 33 B 34 C 05.09.2005 87 3 16,0 1 20 7,4 35 C 11.10.2005 98 3 14,0 3 50 3,5 36 B Fohlennr. Gruppe 19 B 20 117 Kapitel 9: Anhang Tab.8: Tag der Diagnose, Ergebnisse der klinischen, sonographischen Untersuchung und die gemessene Leukozytenkonzentration am Tag der Diagnose und der Gehalt im Serum an Anti-Rhodococcus equi-Immunglobulin G im entsprechenden Altersabschnitt angegeben in ELISA UNITS [EU] der Fohlen 37 bis 54. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). Leukozyten 9 Anzahl der Abszesse Summe der Abszesstie fen [mm] Anti-R. equi-IgG [EU] Datum Alter [d] klin. Score x10 pro l Blut C 15.08.2005 56 2 18,9 2 30 93,0 42 B 04.10.2005 102 7 13,3 1 12 15,7 43 C 44 C 02.08.2005 22 1 19,7 2 50 0,3 45 C 04.10.2005 81 3 14,3 1 15 3,2 46 A 47 A 23.08.2005 38 2 13,9 2 20 1,3 48 A 49 A 50 C 11.10.2005 102 7 15,1 3 30 10,2 51 C 31.10.2005 117 3 14,2 1 10 10,8 52 C 53 A 15.08.2005 43 0 22,6 1 15 9,4 54 C 06.11.2005 125 2 15,7 3 30 5,7 Fohlennr. Gruppe 37 B 38 C 39 C 40 B 41 118 Kapitel 9: Anhang Tab.9: Tag der Diagnose, Ergebnisse der klinischen, sonographischen Untersuchung und die gemessene Leukozytenkonzentration am Tag der Diagnose und der Gehalt im Serum an Anti-Rhodococcus equi-Immunglobulin G im entsprechenden Altersabschnitt angegeben in ELISA UNITS [EU] der Fohlen 55 bis 72. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). n.a. = nicht auswertbar Leukozyten Summe der Abszesstie fen [mm] Anti-R. equi-IgG [EU] Fohlennr. Gruppe Datum Alter [d] klin. Score x109 pro l Blut Anzahl der Abszesse 55 C 28.09.2005 108 7 10,9 3 40 3,2 56 A 17.10.2005 94 6 11,2 1 12 14,3 57 A 06.09.2005 52 2 19,2 4 75 8,0 58 B 59 B 05.09.2005 55 7 14,9 6 60 3,4 60 A 15.08.2005 62 1 21,8 1 20 5,9 61 B 62 B 63 A 15.08.2005 54 4 21,0 3 100 n.a. 64 C 07.11.2005 120 5 14,6 9 130 9,3 65 B 04.11.2005 116 6 13,3 1 10 9,4 66 B 67 B 15.08.2005 64 0 13,9 1 30 0,7 68 B 69 A 05.10.2005 80 2 15,7 2 30 n.a. 70 C 71 A 03.08.2005 53 1 15,7 2 20 11,5 72 A 15.08.2005 55 0 15,0 3 75 13,6 119 Kapitel 9: Anhang Tab.10: Tag der Diagnose, Ergebnisse der klinischen, sonographischen Untersuchung und die gemessene Leukozytenkonzentration am Tag der Diagnose und der Gehalt im Serum an Anti-Rhodococcus equi-Immunglobulin G im entsprechenden Altersabschnitt angegeben in ELISA UNITS [EU] der Fohlen 74 bis 92. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). Datum Alter [d] klin. Score x109 pro l Blut Anzahl der Abszesse Summe der Abszesstie fen [mm] Anti-R. equi-IgG [EU] 31.10.2005 115 7 13,1 2 25 9,7 29.08.2005 68 1 12,8 4 45 3,8 A 19.09.2005 64 0 17,9 3 60 68,1 84 C 15.08.2005 53 1 15,7 1 10 116,0 85 A 09.08.2005 36 1 16,7 1 20 24,1 86 C 87 B 20.09.2005 94 3 8,1 1 10 6,3 88 B 89 A 90 B 91 C 07.11.2005 118 3 9,9 3 50 8,9 92 A Leukozyten Fohlennr. Gruppe 74 C 75 C 76 A 77 C 78 C 80 A 81 C 82 C 83 120 Kapitel 9: Anhang Tab.11: Tag der Diagnose, Ergebnisse der klinischen, sonographischen Untersuchung und die gemessene Leukozytenkonzentration am Tag der Diagnose und der Gehalt im Serum an Anti-Rhodococcus equi-Immunglobulin G im entsprechenden Altersabschnitt angegeben in ELISA UNITS [EU] der Fohlen 93 bis 110. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). n.a. = nicht auswertbar Leukozyten Anzahl der Abszesse Summe der Abszesstie fen [mm] Anti-R. equi-IgG [EU] Datum Alter [d] klin. Score x109 pro l Blut A 23.08.2005 48 2 17,4 1 10 13,7 95 A 26.08.2005 46 7 16,0 1 10 10,3 96 A 97 B 98 B 99 C 100 A 02.08.2005 46 0 15,2 2 40 10,1 101 A 102 B 103 A 15.08.2005 60 5 23,7 3 40 3,4 104 C 105 A 106 B 29.08.2005 60 2 15,2 1 30 n.a. 107 A 26.09.2005 76 5 14,7 3 40 7,6 108 B 12.10.2005 110 2 13,7 1 10 14,1 109 B 24.10.2005 115 2 16,4 1 12 5,2 110 B Fohlennr. Gruppe 93 B 94 121 Kapitel 9: Anhang Tab.12: Anti-Rhodococcus equi-Immunglobulin G-Gehalt angegeben in ELISA UNITS im Serum der Fohlen Nr. 1 bis 24 zu jedem Untersuchungszeitpunkt. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). n.a. = nicht auswertbar Alter der Fohlen in Wochen Fohlennr. Gruppe 1. LT 1 2 3 4 5/6 7/8 9/10 11/12 13/14 15/16 17/18 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 B A B C B A A C B B A B B A A A B C B C C C A C 25,2 71,4 4,7 109,3 24,1 37,4 29,3 30,6 4,3 13,3 9,0 6,1 2,0 94,9 15,5 40,0 86,0 13,1 78,4 3,5 3,2 8,1 17,2 1,6 12,6 30,3 0,9 24,6 4,8 8,8 7,1 14,7 1,6 3,2 5,4 3,1 0,4 24,1 5,7 12,5 59,8 3,4 29,6 1,8 1,1 2,6 5,9 2,3 7,2 28,1 1,1 20,5 5,2 5,5 7,9 10,5 1,3 3,4 5,2 3,3 3,0 24,0 7,3 14,0 49,2 3,0 24,5 1,2 0,7 2,4 4,1 3,5 6,3 20,3 1,3 25,1 5,7 4,2 6,1 8,1 1,0 3,5 5,1 2,3 1,8 25,1 9,2 14,2 57,7 3,2 21,7 0,9 1,6 2,9 3,0 2,1 6,2 21,8 1,2 14,7 3,8 1,9 4,9 5,3 0,8 3,3 6,8 1,8 1,2 9,2 3,4 2,5 19,4 0,4 8,3 0,5 0,9 3,7 2,2 1,2 4,5 15,9 0,6 8,6 1,8 1,3 2,5 2,7 15,9 2,1 7,2 2,1 2,1 11,8 3,8 5,6 33,6 6,1 10,9 22,5 7,4 10,1 3,8 3,9 3,5 28,9 12,4 6,2 1,6 6,8 37,0 1,2 39,7 3,5 5,8 12,0 113,0 6,5 n.a. 4,3 12,6 30,8 6,4 11,8 32,6 17,2 6,3 2,5 26,3 20,6 16,0 tot 5,7 9,7 29,3 3,4 53,5 8,5 12,6 23,2 n.a. 11,3 n.a. 4,2 12,0 30,8 9,5 13,3 20,4 9,8 229,2 7,5 36,1 21,0 19,1 tot 5,4 15,3 21,0 4,2 30,9 7,5 11,7 13,4 n.a. 14,8 n.a. 3,8 17,6 35,1 4,3 13,3 13,1 7,7 21,8 1,2 17,5 8,6 11,2 tot 5,3 12,7 19,1 4,1 13,2 4,9 12,3 9,8 36,6 8,2 n.a. 2,8 6,8 5,6 4,4 6,6 6,4 7,2 15,6 7,2 23,6 16,2 8,1 tot 3,1 10,6 11,0 2,9 9,4 7,4 11,1 15,5 51,3 10,2 n.a. 2,7 9,8 8,9 6,7 7,5 8,9 4,0 9,0 3,2 46,5 4,4 5,8 tot 2,1 6,6 8,2 1,5 5,3 5,3 7,1 9,1 27,6 5,1 n.a. 1,9 6,1 3,5 3,8 3,8 3,3 9,8 9,7 2,2 122 Kapitel 9: Anhang Tab.13: Anti-Rhodococcus equi-Immunglobulin G-Gehalt angegeben in ELISA UNITS im Serum der Fohlen Nr. 25 bis 48 zu jedem Untersuchungszeitpunkt. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). n.a. = nicht auswertbar Alter der Fohlen in Wochen Fohlennr. Gruppe 1. LT 1 2 3 4 5/6 7/8 9/10 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 B B C C A C A B B C C B B C C B C B C C C A A A 8,0 10,5 6,1 46,7 62,4 32,7 3,7 2,6 30,0 114,4 14,2 72,0 2,6 3,2 70,6 94,9 5,5 5,5 36,8 3,0 45,5 36,4 2,7 12,7 1,3 4,8 0,9 10,6 14,8 7,7 13,0 1,5 3,9 24,6 2,7 25,1 0,8 0,4 17,0 18,5 4,3 6,7 18,6 0,2 17,1 21,1 0,1 4,0 1,0 3,7 1,3 9,3 10,2 4,9 9,9 1,1 2,9 24,6 2,3 18,6 2,0 2,6 16,7 17,5 4,6 6,2 14,3 0,4 16,6 19,5 0,5 3,9 2,8 2,5 0,7 4,7 5,3 2,3 6,7 0,9 1,9 14,0 1,2 15,4 0,6 0,6 9,5 9,7 2,3 1,9 8,2 0,3 10,9 11,6 0,1 1,7 0,8 5,0 1,4 4,0 3,7 1,0 4,8 1,8 1,1 8,6 1,0 9,9 1,1 0,3 6,4 9,8 0,1 1,9 5,8 0,3 6,8 5,3 0,4 1,9 5,4 3,8 2,1 n.a. 2,8 2,4 6,3 1,5 0,8 5,7 0,9 30,1 2,0 0,4 5,4 8,9 3,8 11,8 3,7 0,7 6,7 4,8 1,3 2,3 5,6 3,3 2,9 n.a. n.a. 3,8 31,8 9,0 10,1 5,9 11,9 25,3 2,7 1,1 50,1 n.a. 93,0 109,0 13,3 0,5 6,4 39,0 0,8 3,9 6,4 4,6 4,9 79,0 41,0 9,2 68,2 5,9 3,5 6,0 9,8 11,5 12,1 4,1 18,7 130,0 27,2 40,0 38,1 0,5 2,2 32,8 0,7 3,5 123 11/12 13/14 15/16 17/18 5,2 3,5 3,9 32,0 26,0 6,3 50,0 4,0 3,8 7,4 11,2 12,5 29,7 5,1 11,7 43,1 10,9 28,6 24,6 2,4 3,2 46,1 1,4 3,4 4,9 5,7 3,2 17,9 18,4 6,9 fehlt 7,1 4,1 3,5 3,5 4,6 12,4 2,3 4,7 17,6 4,7 15,7 32,2 11,4 3,0 77,0 3,6 7,0 1,7 1,8 2,1 8,5 9,5 3,2 34,8 3,5 4,0 4,8 3,2 4,8 8,7 2,9 6,0 15,7 6,3 16,3 26,0 50,0 4,3 65,0 3,9 8,9 2,2 19,3 2,4 4,8 6,6 3,3 20,1 3,7 7,2 6,0 4,2 5,8 9,0 3,4 5,8 9,8 2,7 11,1 15,9 42,6 2,7 35,4 2,8 4,4 Kapitel 9: Anhang Tab.14: Anti-Rhodococcus equi-Immunglobulin G-Gehalt angegeben in ELISA UNITS im Serum der Fohlen Nr. 49 bis 72 zu jedem Untersuchungszeitpunkt. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). n.a. = nicht auswertbar Alter der Fohlen in Wochen Fohlennr. Gruppe 1. LT 1 2 3 4 5/6 7/8 9/10 11/12 13/14 15/16 17/18 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 A C C C A C C A A B B A B B A C B B B B A C A A 10,0 35,2 21,3 5,7 2,7 0,8 23,4 10,0 0,6 15,5 26,4 63,6 17,2 1,9 1,4 19,6 12,0 14,4 19,3 21,5 4,4 2,0 13,6 89,3 2,9 15,9 14,2 22,4 3,8 0,6 16,1 6,4 1,3 11,5 9,6 29,6 9,2 0,8 0,5 5,4 5,4 6,4 4,9 11,8 1,0 1,2 13,8 30,3 4,6 16,6 10,9 16,5 1,7 1,2 23,0 4,0 0,4 6,6 7,8 23,5 5,2 1,0 0,7 3,8 3,8 3,3 3,3 6,6 1,0 1,3 6,3 34,0 1,5 9,7 5,3 9,2 2,6 0,4 10,8 0,7 0,2 3,5 4,1 10,4 6,5 3,3 0,6 2,2 2,2 1,2 2,3 5,9 1,2 1,6 6,7 21,9 1,4 7,0 5,0 6,2 0,9 1,0 5,7 0,6 1,0 1,9 2,7 7,1 6,8 3,0 0,6 2,5 2,2 1,0 1,5 4,9 3,0 1,4 3,0 17,2 8,3 26,8 30,9 5,8 9,4 123,0 4,7 4,4 1,7 1,2 2,5 5,8 4,0 24,4 1,7 1,6 10,7 5,5 0,6 2,0 1,3 0,8 2,3 8,6 14,6 17,5 23,4 13,6 9,5 13,5 14,0 8,7 8,0 5,1 9,7 16,3 3,5 31,4 n.a. 7,2 14,1 16,9 0,6 7,5 1,2 1,3 11,5 13,6 11,4 7,2 20,0 5,2 6,3 36,7 4,1 19,0 n.a. 3,3 2,6 5,9 17,3 24,6 69,8 100,0 8,8 12,2 0,7 17,3 168,0 1,2 56,8 25,4 7,1 6,9 27,8 5,0 7,6 23,2 3,6 15,8 231,0 4,6 3,4 21,0 7,8 35,3 64,9 38,9 16,5 13,4 72,9 11,2 n.a. 8,9 51,9 20,9 124 13,0 10,2 11,0 4,4 5,8 13,9 2,5 14,3 n.a. 3,0 2,9 66,3 6,1 39,1 40,0 24,6 10,8 15,8 31,8 8,7 52,6 34,1 43,3 13,1 12,4 12,3 10,8 2,9 5,2 10,5 3,2 24,0 96,0 3,9 2,5 63,0 8,8 44,9 34,2 14,7 9,4 24,2 20,0 10,7 49,7 37,4 62,0 11,2 5,8 8,0 4,4 2,5 3,7 5,7 2,0 9,5 95,0 3,5 1,3 31,7 3,6 42,5 32,5 9,3 8,0 12,7 9,6 3,8 17,3 16,3 28,1 12,4 Kapitel 9: Anhang Tab.15: Anti-Rhodococcus equi-Immunglobulin G-Gehalt angegeben in ELISA UNITS im Serum der Fohlen Nr. 74 bis 98 zu jedem Untersuchungszeitpunkt. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). n.a. = nicht auswertbar Alter der Fohlen in Wochen Fohlennr. Gruppe 1. LT 1 2 3 4 5/6 7/8 9/10 11/12 13/14 15/16 17/18 74 75 76 77 78 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 C C A C C A C C A C A C B B A B C A B A A A B B 42,7 11,5 18,7 2,4 10,2 31,4 12,0 6,2 11,0 9,1 4,8 6,7 11,1 10,6 37,9 2,0 5,4 16,1 1,8 2,2 1,3 1,7 19,8 4,2 15,2 2,5 3,4 1,8 2,2 4,8 3,4 3,5 5,4 3,3 2,5 1,4 5,3 3,6 28,2 0,7 5,5 12,2 1,1 1,0 0,9 1,1 21,1 8,6 7,6 1,3 2,6 0,5 1,4 2,5 3,2 6,1 3,2 2,5 1,8 1,1 2,6 1,9 21,7 0,3 0,8 3,9 0,2 0,2 0,3 0,4 18,1 2,1 5,2 2,3 2,5 1,0 1,9 1,2 2,7 1,2 2,1 2,0 3,1 1,9 3,1 3,4 27,7 4,6 0,5 3,3 0,5 1,1 0,5 1,6 15,4 6,0 1,7 0,8 1,0 0,5 1,0 1,0 1,4 2,2 1,2 0,9 0,7 1,2 1,1 0,5 7,0 0,3 1,5 3,6 1,2 0,6 0,6 0,5 6,4 1,4 2,0 3,1 0,6 1,0 0,6 0,8 4,0 4,2 1,3 1,5 24,1 1,4 17,7 1,5 8,6 2,5 1,0 3,3 7,0 18,8 3,6 1,0 16,4 2,7 5,2 5,7 6,8 6,3 2,1 n.a. 8,4 22,2 3,8 116,0 105,0 5,2 26,8 5,1 11,3 4,1 2,7 13,0 8,4 13,7 10,3 2,0 10,1 123,0 6,3 5,2 n.a. 9,8 2,5 3,8 6,9 17,9 68,1 26,3 52,0 20,1 12,2 5,4 6,1 7,4 7,8 18,7 9,1 38,3 fehlt 4,0 5,1 123,0 9,0 6,7 43,6 17,6 3,5 tot 10,1 23,7 23,6 n.a. 27,8 26,0 9,7 8,0 4,8 6,6 5,9 14,5 10,6 98,0 6,6 13,0 10,1 56,9 6,5 4,3 27,8 10,3 1,1 tot 10,1 45,8 13,2 80,0 25,3 11,4 6,3 23,8 4,4 27,3 7,7 6,8 16,4 70,0 5,4 3,7 4,4 55,0 6,5 7,6 21,7 9,2 3,3 tot 9,6 n.a. 10,4 n.a. 23,2 9,9 5,0 19,2 5,0 29,9 4,8 4,7 12,1 35,7 4,9 5,1 4,8 23,0 7,3 8,5 20,7 9,7 3,4 tot 7,7 n.a. 14,1 63,6 22,1 8,8 7,6 25,9 14,2 41,7 8,9 7,5 15,9 46,6 fehlt 5,4 8,0 38,9 125 Kapitel 9: Anhang Tab.16: Anti-Rhodococcus equi-Immunglobulin G-Gehalt angegeben in ELISA UNITS im Serum der Fohlen Nr. 99 bis 110 zu jedem Untersuchungszeitpunkt. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). n.a. = nicht auswertbar Alter der Fohlen in Wochen Fohlennr. Gruppe 1. LT 1 2 3 4 5/6 7/8 9/10 11/12 13/14 15/16 17/18 99 100 101 102 103 104 105 106 107 108 109 110 C A A B A C A B A B B B 2,3 1,2 2,5 8,8 21,9 1,6 31,6 4,0 6,9 10,1 21,0 27,5 3,1 5,3 2,3 4,1 13,1 6,0 14,6 1,6 3,1 7,5 6,3 21,4 0,5 2,0 1,6 3,0 10,6 3,9 11,2 1,0 1,1 3,9 2,0 17,2 3,7 5,4 1,1 2,1 4,5 1,9 5,1 0,7 0,8 3,0 1,0 10,0 1,0 1,4 1,1 2,2 4,1 2,0 5,6 5,3 0,4 2,1 3,8 9,3 3,2 10,1 85,0 4,0 6,9 13,9 10,5 n.a. 1,5 3,2 2,6 20,4 53,8 69,8 82,0 5,3 3,4 33,9 6,6 n.a. 3,4 6,3 2,9 7,5 n.a. 327,0 76,0 9,0 203,0 34,6 11,9 n.a. 12,9 79,0 10,1 10,1 34,0 n.a. 28,8 9,9 tot 21,2 6,0 n.a. 7,6 44,0 9,6 7,5 23,8 54,0 14,3 5,0 tot 14,3 7,5 96,0 5,7 28,5 3,6 7,5 19,0 36,8 10,7 7,3 tot 11,7 7,3 87,7 9,2 14,1 5,2 3,9 17,1 33,8 14,2 10,9 tot 13,2 6,4 77,7 4,7 8,3 3,5 2,6 126 Kapitel 9: Anhang Tab.17: Irrtumswahrscheinlichkeiten (p) zum Unterschied der Morbidität der Fohlengruppen im Fisher-Test. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). Gruppe A Gruppe A Gruppe B 0,059 Gruppe C 1,000 Gruppe B Gruppe C 0,059 1,000 0,098 0,098 Tab.18: Irrtumswahrscheinlichkeiten (p) zum Unterschied des mittleren Erkrankungsalters der Fohlengruppen im Kruskal-Wallis-Test. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). Gruppe A Gruppe A Gruppe B 0,4901 Gruppe C 0,0065 Gruppe B Gruppe C 0,4901 0,0065 0,6130 0,6130 Tab.19: Irrtumswahrscheinlichkeiten (p) zum Unterschied des mittleren klinischen Scores nach OHNESORGE et al. (1998) der Fohlengruppen im Kruskal-WallisTest. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). Gruppe A Gruppe A Gruppe B 0,568 Gruppe C 0,268 Gruppe B Gruppe C 0,568 0,268 1,000 1,000 Tab.20: Irrtumswahrscheinlichkeiten (p) zum Unterschied der mittleren Leukozytenkonzentration im Blut zum Zeitpunkt der Diagnose in den Fohlengruppen im Kruskal-Wallis-Test. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). Gruppe A Gruppe A Gruppe B 0,066 Gruppe C 0,165 Gruppe B Gruppe C 0,066 0,165 1,000 1,000 127 Kapitel 9: Anhang Tab.21: Irrtumswahrscheinlichkeiten (p) zum Unterschied der mittleren Anzahl der Abszesse in den Fohlengruppen zum Diagnosezeitpunkt im Kruskal-WallisTest. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). Gruppe A Gruppe A Gruppe B 0,222 Gruppe C 0,785 Gruppe B Gruppe C 0,222 0,785 0,018 0,018 Tab.22: Irrtumswahrscheinlichkeiten (p) zum Unterschied der durchschnittlichen Summe der Abszesstiefen zum Zeitpunkt der Diagnose in den Fohlengruppen im Kruskal-Wallis-Test. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). Gruppe A Gruppe A Gruppe B 0,637 Gruppe C 0,674 Gruppe B Gruppe C 0,637 0,674 0,067 0,067 Tab.23: Irrtumswahrscheinlichkeiten (p) zum Unterschied des mittleren Gehaltes an Anti-R. equi-IgG in ELISA UNITS in der 9./10. Woche der Fohlengruppen im Kruskal-Wallis-Test. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). Gruppe A Gruppe A Gruppe B 0,143 Gruppe C 0,061 Gruppe B Gruppe C 0,143 0,061 1,000 1,000 Tab.24: Irrtumswahrscheinlichkeiten (p) zum Unterschied des mittleren Gehaltes an Anti-R. equi-IgG in ELISA UNITS in der 15./16. Woche in den Fohlengruppen im Kruskal-Wallis-Test. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). Gruppe A Gruppe A Gruppe B 0,218 Gruppe C 0,006 Gruppe B Gruppe C 0,218 0,006 0,514 0,514 128 Kapitel 9: Anhang Tab.25: Irrtumswahrscheinlichkeiten (p) zum Unterschied des mittleren Gehaltes an Anti-R. equi-IgG in ELISA UNITS in der 17./18. Woche in den Fohlengruppen im Kruskal-Wallis-Test. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). Gruppe A Gruppe A Gruppe B 0,783 Gruppe C 0,022 Gruppe B Gruppe C 0,783 0,022 0,318 0,318 Tab.26: Irrtumswahrscheinlichkeiten (p) zum Unterschied der mittleren Fläche unter der Kurve (AUC) in den Fohlengruppen im Kruskal-Wallis-Test. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). Gruppe A Gruppe A Gruppe B 0,576 Gruppe C 0,141 Gruppe B Gruppe C 0,576 0,141 1,000 1,000 Tab.27: Irrtumswahrscheinlichkeiten (p) zum Unterschied des mittleren Gehaltes an Anti-R. equi-IgG in ELISA UNITS in der 15./16. Woche der erkrankten Fohlen in den Gruppen im Kruskal-Wallis-Test. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). Gruppe A Gruppe A Gruppe B 0,550 Gruppe C 0,0088 Gruppe B Gruppe C 0,550 0,0088 0,5312 0,5312 Tab. 28: Irrtumswahrscheinlichkeiten (p) zum Unterschied des mittleren Gehalts an Anti-R equi-Immunglobulin G (IgG) acht bis zehn Stunden post natum zwischen den erkrankten und gesunden Fohlen jeder Gruppe im MannWhitney-U-Test. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). Irrtumswahrscheinlichkeit (p) Gruppe A 0,095 Gruppe B 0,649 Gruppe C 0,975 129 Kapitel 9: Anhang Tab.29: Korrelationskoeffizienten zur Berechnung des Zusammenhanges zwischen Anti-R. equi-IgG acht bis zehn Stunden post natum und dem Erkrankungsalter. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). Spearman R p-Niveau Gruppe A 0,520 0,013 Gruppe B 0,038 0,901 Gruppe C -0,040 0,862 Tab.30: Korrelatinskoeffizienten und Irrtumswahrscheinlichkeiten (p) bei der Berechnung des Einflusses des Anti-R. equi-IgG-Gehaltes 8 – 10 Stunden post natum auf den Parameter AUC in den Gruppen in Spearmans Rangkorrelation. Gruppe A: Kontrollgruppe (n=36); Gruppe B: Impfstoff mit Adjuvans CpG XXXX (n=36); Gruppe C: Impfstoff ohne Adjuvans (n=36). Spearman R p-Niveau Gruppe A 0,247 0,235 Gruppe B -0,045 0,804 Gruppe C 0,147 0,431 130 Kapitel 9: Anhang Verzeichnis der Tabellen Tab. 1: Nomenklatur des equinen Immunglobulinsystems ...................................... 34 Tab. 2: Klinischer Score zur Beurteilung des Schweregrads der klinischen Symptome (nach OHNESORGE et al. 1998) .............................................. 54 Tab. 3: Mikrotiterplattenbelegung des Rhodococcus equi-ELISA ........................... 59 Tab. 4: Irrtumswahrscheinlichkeiten (p-Werte) zur Darstellung der Signifikanzen .. 62 Tab.5: Anti-R equi-Immunglobulin G (IgG) in ELISA UNITS [EU] acht bis zehn Stunden post natum differenziert nach den Gruppen insgesamt und unterteilt in gesunde und erkranke Fohlen ........................................... 77 Tab.6: Tag der Diagnose, Ergebnisse der klinischen, sonographischen Untersuchung und die gemessene Leukozytenkonzentration am Tag der Diagnose und der Gehalt im Serum an Anti-Rhodococcus equiImmunglobulin G im entsprechenden Altersabschnitt angegeben in ELISA UNITS [EU] der Fohlen 1 bis 18..................................................... 116 Tab.7: Tag der Diagnose, Ergebnisse der klinischen, sonographischen Untersuchung und die gemessene Leukozytenkonzentration am Tag der Diagnose und der Gehalt im Serum an Anti-Rhodococcus equiImmunglobulin G im entsprechenden Altersabschnitt angegeben in ELISA UNITS [EU] der Fohlen 19 bis 36................................................... 117 Tab.8: Tag der Diagnose, Ergebnisse der klinischen, sonographischen Untersuchung und die gemessene Leukozytenkonzentration am Tag der Diagnose und der Gehalt im Serum an Anti-Rhodococcus equiImmunglobulin G im entsprechenden Altersabschnitt angegeben in ELISA UNITS [EU] der Fohlen 37 bis 54................................................... 118 Tab.9: Tag der Diagnose, Ergebnisse der klinischen, sonographischen Untersuchung und die gemessene Leukozytenkonzentration am Tag der Diagnose und der Gehalt im Serum an Anti-Rhodococcus equiImmunglobulin G im entsprechenden Altersabschnitt angegeben in ELISA UNITS [EU] der Fohlen 55 bis 72................................................... 119 Tab.10: Tag der Diagnose, Ergebnisse der klinischen, sonographischen Untersuchung und die gemessene Leukozytenkonzentration am Tag der Diagnose und der Gehalt im Serum an Anti-Rhodococcus equiImmunglobulin G im entsprechenden Altersabschnitt angegeben in ELISA UNITS [EU] der Fohlen 74 bis 92................................................... 120 131 Kapitel 9: Anhang Tab.11: Tag der Diagnose, Ergebnisse der klinischen, sonographischen Untersuchung und die gemessene Leukozytenkonzentration am Tag der Diagnose und der Gehalt im Serum an Anti-Rhodococcus equiImmunglobulin G im entsprechenden Altersabschnitt angegeben in ELISA UNITS [EU] der Fohlen 93 bis 110................................................. 121 Tab.12: Anti-Rhodococcus equi-Immunglobulin G-Gehalt angegeben in ELISA UNITS im Serum der Fohlen Nr. 1 bis 24 zu jedem Untersuchungszeitpunkt............................................................................ 122 Tab.13: Anti-Rhodococcus equi-Immunglobulin G-Gehalt angegeben in ELISA UNITS im Serum der Fohlen Nr. 25 bis 48 zu jedem Untersuchungszeitpunkt............................................................................ 123 Tab.14: Anti-Rhodococcus equi-Immunglobulin G-Gehalt angegeben in ELISA UNITS im Serum der Fohlen Nr. 49 bis 72 zu jedem Untersuchungszeitpunkt............................................................................ 124 Tab.15: Anti-Rhodococcus equi-Immunglobulin G-Gehalt angegeben in ELISA UNITS im Serum der Fohlen Nr. 74 bis 98 zu jedem Untersuchungszeitpunkt............................................................................ 125 Tab.16: Anti-Rhodococcus equi-Immunglobulin G-Gehalt angegeben in ELISA UNITS im Serum der Fohlen Nr. 99 bis 110 zu jedem Untersuchungszeitpunkt............................................................................ 126 Tab.17: Irrtumswahrscheinlichkeiten (p) zum Unterschied der Morbidität der Fohlengruppen im Fisher-Test .................................................................. 127 Tab.18: Irrtumswahrscheinlichkeiten (p) zum Unterschied des mittleren Erkrankungsalters der Fohlengruppen im Kruskal-Wallis-Test.................. 127 Tab.19: Irrtumswahrscheinlichkeiten (p) zum Unterschied des mittleren klinischen Scores nach OHNESORGE et al. (1998) der Fohlengruppen im Kruskal-Wallis-Test .............................................................................. 127 Tab.20: Irrtumswahrscheinlichkeiten (p) zum Unterschied der mittleren Leukozytenkonzentration im Blut zum Zeitpunkt der Diagnose in den Fohlengruppen im Kruskal-Wallis-Test ..................................................... 127 Tab.21: Irrtumswahrscheinlichkeiten (p) zum Unterschied der mittleren Anzahl der Abszesse in den Fohlengruppen zum Diagnosezeitpunkt im Kruskal-Wallis-Test ................................................................................... 128 132 Kapitel 9: Anhang Tab.22: Irrtumswahrscheinlichkeiten (p) zum Unterschied der durchschnittlichen Summe der Abszesstiefen zum Zeitpunkt der Diagnose in den Fohlengruppen im Kruskal-Wallis-Test ..................................................... 128 Tab.23: Irrtumswahrscheinlichkeiten (p) zum Unterschied des mittleren Gehaltes an Anti-R. equi-IgG in ELISA UNITS in der 9./10. Woche der Fohlengruppen im Kruskal-Wallis-Test ..................................................... 128 Tab.24: Irrtumswahrscheinlichkeiten (p) zum Unterschied des mittleren Gehaltes an Anti-R. equi-IgG in ELISA UNITS in der 15./16. Woche in den Fohlengruppen im Kruskal-Wallis-Test ..................................................... 128 Tab.25: Irrtumswahrscheinlichkeiten (p) zum Unterschied des mittleren Gehaltes an Anti-R. equi-IgG in ELISA UNITS in der 17./18. Woche in den Fohlengruppen im Kruskal-Wallis-Test ..................................................... 129 Tab.26: Irrtumswahrscheinlichkeiten (p) zum Unterschied der mittleren Fläche unter der Kurve (AUC) in den Fohlengruppen im Kruskal-Wallis-Test ...... 129 Tab.27: Irrtumswahrscheinlichkeiten (p) zum Unterschied des mittleren Gehaltes an Anti-R. equi-IgG in ELISA UNITS in der 15./16. Woche der erkrankten Fohlen in den Gruppen im Kruskal-Wallis-Test. ...................... 129 Tab.28: Irrtumswahrscheinlichkeiten (p) zum Unterschied des mittleren Gehalts an Anti-R equi-Immunglobulin G (IgG) acht bis zehn Stunden post natum zwischen den erkrankten und gesunden Fohlen jeder Gruppe im Mann-Whitney-U-Test ............................................................................... 129 Tab.29: Korrelationskoeffizienten zur Berechnung des Zusammenhanges zwischen Anti-R. equi-IgG acht bis zehn Stunden post natum und dem Erkrankungsalter ............................................................................... 130 Tab.30: Korrelatinskoeffizienten und Irrtumswahrscheinlichkeiten (p) bei der Berechnung des Einflusses des Anti-R. equi-IgG-Gehaltes 8 – 10 Stunden post natum auf den Parameter AUC in den Gruppen in Spearmans Rangkorrelation...................................................................... 130 133 Kapitel 9: Anhang Verzeichnis der Abbildungen Abb. 1: Intranasale Applikation beim Fohlen. .......................................................... 52 Abb. 2: Befundbogen für die sonographische Untersuchung der Lunge..……...…...56 Abb. 3: Linearität und Parallelität der logarithmierten OD-Werte in den jeweiligen Verdünnungsstufen des Positivserums und eines Fohlenserums (Kontrollgruppe)................................................................... 61 Abb. 4: Anteil der erkrankten Fohlen an der Gesamtpopulation der Gruppen angegeben in Prozent [%] ........................................................................... 64 Abb. 5: Erkrankungsalter in den einzelnen Gruppen angegeben in Tagen ............. 65 Abb. 6: Anzahl neuerkrankter Fohlen der drei Gruppen in bestimmten Altersabschnitten......................................................................................... 66 Abb. 7: Verlauf des Anteils gesunder Fohlen in den Gruppen in Abhängigkeit vom Lebensalter nach Kaplan-Meier........................................................... 67 Abb. 8: Klinischer Scores nach OHNESORGE et al. (1998) zum Diagnosezeitpunkt....................................................................................... 69 Abb. 9: Leukozytenanzahl pro Liter Blut zum Diagnosezeitpunkt ............................ 70 Abb.10: Abszessanzahl zum Diagnosezeitpunkt ...................................................... 71 Abb.11: Summe der Abszesstiefen zum Diagnosezeitpunkt angegeben in Millimeter [mm]............................................................................................ 72 Abb.12: Anti- R. equi-Immunglobulin G (IgG) im Serum aller Fohlen der drei Gruppen während des Untersuchungszeitraums angegeben in ELISA UNITS [EU] ...................................................................................... 74 Abb.13: Anti- R. equi-Immunglobulin G (IgG) im Serum der gesunden Fohlen der drei Gruppen während des Untersuchungszeitraums angegeben in ELISA UNITS [EU] ...................................................................................... 75 Abb.14: Anti- R. equi-Immunglobulin G (IgG) im Serum der erkrankten Fohlen der drei Gruppen während des Untersuchungszeitraums angegeben in ELISA UNITS [EU] ...................................................................................... 76 134 Danksagung Herrn Prof. Dr. E. Klug danke ich für die freundliche Überlassung des Dissertationsthemas. Ich danke Frau Dr. M. Venner für die intensive Betreuung und die gute Zusammenarbeit, wodurch die zügige Durchführung dieser Arbeit ermöglicht wurde. Herrn P. Schockemöhle danke ich herzlich für die großzügige finanzielle Unterstützung und die Bereitstellung der Pferde für die Studie. Ich danke der Wirtschaftsgenossenschaft deutscher Tierärzte eG (WdT, Garbsen), namentlich Herrn Dr. Schaufuß und Herrn Dr. Steller, für die gute Zusammenarbeit. Frau Dr. K. Strutzberg-Minder danke ich sehr herzlich für die kompetente Beratung und die stets zügige Korrektur meines Manuskripts. Bei den Mitarbeitern der IVD GmbH bedanke ich mich für die freundliche Aufnahme und Zusammenarbeit im Labor. Herrn Dr. Jens Rohwer, dem immunologischen Vater dieser Arbeit, gilt mein besonderer Dank für die scheinbar selbstverständlichen Hilfestellungen von der Planung des Versuchaufbaus bis zur Korrektur des Manuskripts. Den Mitarbeitern des Gestüts Lewitz, vor allem Herrn P. Baumgart und Herrn F. Pieper, gilt mein Dank für die Hilfe bei der Durchführung der Arbeit. Meinen Mitstreiterinnen Ina Baumann, Inke Gravert, Nina Höhensteiger, Birte Reinhold, Saskia Walther und Bianca Weimar danke ich für die gute Zusammenarbeit und für die Hilfe bei der Probennahme. Ganz besonders möchte ich meinen Eltern danken, die mir das Studium und die Dissertation überhaupt erst ermöglicht und immer an mich geglaubt haben. Danke für die Gewissheit, dass Ihr stolz auf mich seid. Mein größter Dank gilt meiner Freundin Silke! Nur durch Deine uneingeschränkte Unterstützung war es überhaupt möglich, diese Arbeit anzufertigen! Danke, dass es Dich für mich gibt!