dspace cover page - ETH E
Werbung

Research Collection
Doctoral Thesis
Beitrag zur Kenntnis der Biologie und Verbreitung einiger
Stechmückenarten in der Schweiz
Author(s):
Büttiker, Willi
Publication Date:
1948
Permanent Link:
https://doi.org/10.3929/ethz-a-000096535
Rights / License:
In Copyright - Non-Commercial Use Permitted
This page was generated automatically upon download from the ETH Zurich Research Collection. For more
information please consult the Terms of use.
ETH Library
ZUR
BEITRAG
DER
KENNTNIS
VERBREITUNG
UND
BIOLOGIE
EINIGER STECHMÜCKENARTEN
IN
DER SCHWEIZ
VON DER
EIDGENÖSSISCHEN TECHNISCHEN HOCHSCHULE
IN ZÜRICH
ZUR
DER
WÜRDE
EINES
ERLANGUNG
DOKTORS
DER
NATURWISSENSCHAFTEN
GENEHMIGTE
PROMOTIONS ARBEIT
VORGELEGT VON
Wilü BÜTTIKER
VON
Referent
:
ÖLTEN
Herr Prof. Dr. 0. Schneider-Orelli
Korreferent
:
Herr Prof. Dr.
IMPRIMERIE
J. Seiler
LA CONCORDE
LAUSANNE
1948
Meinen lieben Eltern.
Abdtuck
aus
der Schweizerischen
den Mitteilungen
Entomologischen Gesellschaft
Band XXI. Heft 1. 1948.
Aus dem Entomologischen Institut der Eidgenössischen Technischen
Hochschule in Zürich
Beitrag
und
zur
Kenntnis der
Biologie
Verbreitung einiger Stechmückenarten
in der Schweiz
von
Willi Büttiker
Rheinfelden
Inhaltsverzeichnis
Seite
Vorwort
I. Einleitung
2
:
1. Sanitäre
4
2.
5
Bedeutung der Stechmücken
Systematik und Synonymie
3. Allgemeines über die untersuchten Arten
II. Materialbeschaffung
1.
und
Materialbeschaffung
2. Zuchttechnik
Zuchttechnik
9
:
12
13
III. KÖRPERBAU:
1. Das Ei
23
2. Die Larven
27
3. Die
32
4. Die
34
Puppen
Imagines
5. Das Darmsystem
34
2
WILLI
IV. Abiotische Aussenfaktoren
:
1. Einfluss der
BÜTTIKER
Seite
41
Temperatur
A. Temperatur und
41
Entwicklungsdauer
B. ThermopraeferendumCVorzugstemperatur)
C. Temperatur und Aktivität
"...
78
Luftfeuchtigkeit
2. Der Einfluss der
79
85
A. Hygrotaxis, Hygropraeferendum
B. Luftfeuchtigkeit und Lebensdauer
93
3. Der Einfluss der Strahlung
A. Strahlung und
B. Phototaxis
C. Strahlung und
.
60
68
94
104
109
Stoffwechselphysiologie
.
Eiablage
112
4. Der Einfluss des Luftdruckes
A. Luftdruck und Entwicklungsdauer
B. Luftdruck und Eiablage
C. Weitere Beobachtungen über den Einfluss des Luftdruckes auf den
113
116
_
Eiablagetermin
V. Die Rassen von Anopheles
der Schweiz
".
maculipennis
und
ihre
Verbreitung
in
Rassenproblems
Gesichtspunkte des Rassenproblems
3. Die Verbreitung der maculipennis-Rassen in der Schweiz
4. Grösse und Zeichnungsmuster von Eiern schweizerischer Herkunft
118
1. Die historische Entwicklung des
122
2. Neue
VI. Biologische Beobachtungen
an
Culiciden
117
124
.
130
:
Entwicklungsverlauf einzelner Eigelege
2. Das Schwimmvermögen der Culiciden
1.
132
135
VII. Zusammenfassung
142
VIII. Literatur-Verzeichnis
144
VORWORT
Bei der Durchsicht der reichhaltigen Stechmückenliteratur zeig¬
sich einige Punkte, welche nicht oder kaum der wissenschaft¬
lichen Untersuchung unterzogen wurden. So fehlen teilweise genaue
Angaben über die Entwicklungszeit und den Einfluss abiotischer
Faktoren (Temperatur, Luftfeuchtigkeit, Licht usw.) auf die ver¬
schiedenen Entwicklungsstadien der Culiciden. In der vorliegenden
Arbeit wurden eine Reihe dieser Faktoren einzeln geprüft. Die
ten
BIOLOGIE
UND
VERBREITUNG
EINIGER
STECHMÜCKENARTEN
3
erhaltenen Resultate gestatten in biologischer und oekologischer
Hinsicht ein besseres Urteil.
Der Abschnitt über Morphologie bestätigt in der Hauptsache
die Befunde früherer Autoren.
Den systematischen Fragen der Formengruppe von Anopheles
maculipennis wurde besondere Beachtung geschenkt. Es zeigte sich
dabei deutlich, dass es sich hier um eine biologisch und oekologisch
recht unterschiedliche Gruppe handelt.
Frühere Untersuchungen
über die Verbreitung der Anophelen in der Schweiz (z. B. GalliValerio, 23) gehen auf eine Zeit zurück, wo man innerhalb der
maculipennis-Gruppe keine Rassenunterschiede machte. Die in dieser
Arbeit erschienenen diesbezüglichen kartographischen Aufnahmen
sind neu.
—
Speziellen biologischen Beobachtungen, besonders im Hinblick
mit Culiciden, wurde ein weiterer Abschnitt
gewidmet.
Meine Untersuchungen über die Frage einer obligaten Symbiose
von Larven oder Imagines mit pflanzlichen
Mikroorganismen fielen
negativ aus.
Die Abbildungen beziehen sich meist auf die vier hauptsächlich
untersuchten Arten ; die Zeichnungen wurden mit Hilfe des Abbé*
sehen Zeichenapparates ausgeführt.
Mit den praktischen Arbeiten begann ich im Verlaufe des Früh¬
jahres 1946 im Entomologischen Institut der Eidgenössischen Tech¬
auf Zuchtversuche
nischen Hochschule in Zürich. Den experimentellen Teil konnte
ich im Sommer I947abschliessen. Um stets genügend Untersuchungs¬
material zur Verfügung zu haben, war es vorerst nötig, eine Anzahl
Culicidenzuchten in Betrieb zu setzen.
Meinem verehrten Lehrer, Herrn Prof. Dr. O. Schneider-Orelli,
Vorstand des Entomologischen Institutes der Eidgenössischen Tech¬
nischen Hochschule, danke ich herzlich für seine wohlwollende
Unterstützung und für die mir erteilten wertvollen Ratschläge.
Herrn Prof. Dr. N. H. SwELLENGREBEL, Instituut voor tropische
Hygiene, Amsterdam, bin ich zu bestem Dank verpflichtet für die
Ermöglichung der Bibliothekbenützung im dortigen Indisch Insti¬
tuut, sowie für die Überlassung von Material von Anopheles macu¬
lipennis
var.
atroparvus.
Herrn Dr. R. Wiesmann in Basel verdanke ich wertvolle Anre¬
gungen im Hinblick auf die vorliegende Arbeit.
I.
1. Sanitäre
EINLEITUNG
Bedeutung der Stechmücken
Die Stechmücken (Culicidae, Diptera) sind die hauptsächlich¬
Urheber der weitverbreiteten und gefürchteten Mücken-,
Schnaken- oder Gelsenplage. Die Gabelmücken (Anopheles-Arten)
im besondern sind die hauptsächlichsten Verbreiter des Wechsel¬
fiebers des Menschen (Malaria), das nicht nur in verschiedenen
Teilen Europas verkommt, sondern ganz besonders tropische und
subtropische Gebiete schwer heimsucht. In Mitteleuropa waren bis
vor kurzer Zeit noch Malariaherde bekannt1.
Die schweizerischen Fiebergebiete lagen vor allem an den Mittel¬
landseen und den sogenannten Föhntälern. Zwischen 1870 und 1890
erlosch, ähnlich wie in Schweden und Finnland, ein Malariaherd
nach dem andern, trotzdem die malariaübertragenden Mücken
zahlenmässig kaum stark zurückgingen (SwELLENGREBEL, 81). Die
eigentlichen Gründe, welche zum Erlöschen der Malaria in Mittel¬
europa führten, sind allerdings nicht bekannt (Geigy, 26).
Die Vertreter der Gattung Anopheles sind für die Übertragung
der Menschenmalaria verantwortlich, während Culex, i4edes und
Theobaidia die Überträger der Vogelmalarien sind, z. B. Proteosoma
praecox durch C. pipiens, Aedes communis, A. aegypti und A. mariae
(Reichenow, 68), bei zahlreichen Vogelarten ferner Proteosoma circumflexum durch Theobaidia annulata.
Aedes aegypti (syn. Stegomyia fasciata) überträgt die Viren der
Gelbfieber, von denen das Stadt-Gelbfieber und das Denguefieber
die bekanntesten sind. In den Tropen und Subtropen werden die
Erreger der Elephantiasis, Microfilaria bancrofti (Nematelminthes,
Vermes) und Filarien des Hundes, Filaria immitis und F. repens
durch Stechmücken, hauptsächlich der Gattungen Culex und Aedes,
sten
übertragen.
Unsere einheimische Singschnake (Culex pipiens) ist an der
Übertragung von Viren der Geflügelpocken und Geflügeldiphterie
beteiligt (Martini, 52). Gewisse Vogeltrypanosomen werden eben¬
falls durch Stechmücken übertragen ; im allgemeinen gelten diese
Erreger als harmlos, da sie nach kurzer Massenvermehrung in eine
chronische, den Vogel nicht weiter schädigende Infektion übergehen
(Martini, 52 ; Steinhaus, 80).
1
Schweiz(Galli-Valerio, 24): 1870-1890; Baden(Schwietert, 77): 1905-1915
noch 169 Fälle; Württemberg (Schwietert, 77): 1895, 15 Fälle.
BIOLOGIE UND
VERBREITUNG EINIGER
STECHMÜCKENARTEN
5
BRAUN und Caspari (6) gelang im Experiment die Übertragung
des Paratyphus-Bazillus durch Culex pipiens. Die Bazillen persistier¬
ten während 3—4 Wochen im Körper von C. pipiens; durch den
Kot der Stechmücken mussten sie wieder ausgeschieden werden.
Beim Zerdrücken infizierter Mücken oder vermittelst infizierter
Nahrungsmittel können diese Bazillen verbreitet werden ; die Über¬
tragung erfolgt aber offenbar selten.
Die Geflügelspirochaete (Spirochaete gallinarum) ist der haupt¬
sächlichste Vertreter der Rückfallfiebergruppe. Nach Zuelzer
cit. lit. Braun, 6) spielt C. pipiens als Überträger eine grosse Rolle.
In Nord- und Südamerika werden weitere Viren durch Culiciden
übertragen. So wird die in St. Louis (U.S.A.) 1933 erstmals auf¬
getretene Menschen-Encephalitis durch Culex tarsalis übertragen
(Matheson, 53) ; im Experiment sind sechs weitere Culicidenarten
für die Übertragung nachgewiesen worden.
Die Pferde-Encephalitis (Gehirn- und Rückenmarksentzündung)
wurde in den U. S. A. schon vor 1900 untersucht ; das erregende
Virus konnte aber erst 1931 isoliert werden. Als Überträger ist
Culex tarsalis nachgewiesen (MATHESON, 53). Nebst den gewöhn¬
lichen Eiterinfektionen können Culiciden bei empfindlichen Per¬
sonen starke quaddel-förmige Hautreaktionen als allergische Erschei¬
nungen hervorrufen.
Schliesslich kommt den Stechmücken als Ruhestörer, beson¬
ders während der Nacht, eine nicht zu unterschätzende praktische
Bedeutung
zu.
2.
Die
Systematik und Synonymie
Stellung der in der vorliegenden Arbeit
systematische
berücksichtigten Arten
ist nach MARSHALL
(46) folgende
:
Insecta
Ordnung
Unterordnung
Familien
Unterfamilie
Tribus
Diptera
Nematocera
Culicidae
Culicinae
Genus
Anophelini
Anopheles
Subgenus
Anopheles
Art>en
A.
A.
Culiiictm
une
Cultex
Culex
bifurcatus (syn. claviger)
maculipennis
Aëdtes
Aëdimorphus Finlaya
C. pipiens
Aides vexans
Aedes geniculatus
6
WILLI
A.
Rassen
BUTTIKER
maculipennis
var.
messeae
var.
melanoon
var.
typicus
Die Nematoceren enthalten eine
grosse
Anzahl
von
Familien
(z. B. Tipulidae, Psychodidae, Culicidae, Chironomidae, usw.), wobei
die Familie der Culicidae wieder in drei Unterfamilien zerfällt,
nämlich in
Culicinae
/-./ "ïf
•
Chaobormae
eigentliche Stechmücken
\
)
nicht stechende Mücken
Die ungefähr 1400 beschriebenen Culicidenarten werden dem
Tribus der Anophelini einerseits und der Culicini anderseits zuge¬
teilt und im gesamten in dreissig Genera getrennt.
Sämtliche Vertreter der Anophelini gehören den folgenden drei
Gattungen
an :
£>•
11
I
m
EuroPa nicht
vertreten
Anopheles
Die Gattung Anopheles erfährt eine weitere Aufspaltung in vier
Subgenera :
subg.
subg.
subg.
subg.
Anopheles
1
Stethomyia
Nyssorhynchus >
)
Myzomyia
in
Europa nicht
vertreten
Sämtliche Vertreter der Culicini, ungefähr 1250 an der Zahl,
werden in 27 verschiedene Genera eingeteilt, wovon fünf Gattungen
in Mitteleuropa vorhanden sind, nämlich : Culex, Aedes, Theobaidia,
Orthopodomyia, Taeniorhynchus.
Die mitteleuropäischen Vertreter
den Untergattungen
subg. Culex
subg. Neoculex
an.
der
Gattung Aedes gehören
BIOLOGIE UND VERBREITUNG
Die hauptsächlichsten
Unterschiede zwischen Anophelini und Culicini
Culicini
Anophelini
Stadium
Eier
EINIGER STECHMUCKENARTEN
1. Mit
Schwimmkammern
ausgerüstet.
2. Werden einzeln auf die
Wasseroberfläche abge¬
legt.
mit Schwimm¬
1. Nicht
kammern ausgerüstet.
2. Werden separat abgelegt
oder
zu
Schiffchen
sammengeklebt
trockenem
Überflutung
dem
Boden
;
oder
zu¬
auf
zur
gelangen¬
oder
auf
Wasseroberfläche.
Larven
1. Atmen durch ein Paar
Siphonen, welche sich
dorsal am 8. Segment
befinden.
2. Liegen horizontal an der
Wasseroberfläche.
3. Nahrung wird von der
Wasseroberfläche einge¬
nommen. Der Kopf wird
um 180° gedreht.
Puppen
1. Mit mehr oder weniger
kegelförmigen Atmungs¬
röhren.
2. Mit Borste an jedem
apicalen Ende der Abdo¬
minalsegmente (III bis
VII).
3. Mit einer zusätzlichen
der
Borste
ventralen
1. Atmen durch ein Paar
Stigmen, welche tubus¬
artig verlängert sind (Si¬
phon).
2. Liegen kopfabwärts an
der Wasseroberfläche.
3. Schwebende Partikel bil¬
den die Nahrung oder
die Larven bewegen mit
den Mundbürsten tote,
pflanzliche Substanz.
1. Mit mehr oder weniger
zylinderförmigen Atem¬
röhren.
2. An jedem apicalen Ende
der Abdominalsegmente
ein meist gezacktes Haar.
3. Ohne zusätzliche Borste.
Schwimmpaddeln.
Imagines
1. Haben keine Schuppen
am Abdomen.
haben
Weibchen
2. Die
lange und schmale Pal¬
pen,
ungefähr gleich lang
wie die Proboscis.
3. Die Männchen haben
lange, am Ende verbrei¬
tete Kiefertaster.
4. Körperachse
abgewin¬
kelt zur Unterlage.
1. Mit dichtbeschupptem
Abdomen.
Weibchen haben
2. Die
kurze Kiefertaster, 1/5
bis 1/i so lang wie der
Rüssel.
3. Die Männchen haben
lange, federförmige Kie¬
fertaster.
4. Körperachse parallel zur
Unterlage.
8
WILLI
BÜTTIKER
Die hauptsächlichsten Unterschiede zwischen den Gattungen
Culex und Aedes
Cule
1. Werden
zu
Aedes
Schiffchen
abgelegt.
2. Mit Carolla(Abb. 4, b).
Eizahn meisselförmig
spitzt (Abb. 4, m).
zuge¬
1. Werden
einzeln
legt.
abge¬
2. Ohne CaroIIa.
Eizahn
Druck-Knopf
als
(Abb. 4, h und i).
Keine generellen Unterschiede.
(Für die Bestimmung vergl. Marshall, 46.)
Das Abdominalhaar A/VIII
hat die Ansatzstelle vom
Seitenrande entfernt.
Das Abdominalhaar A/VIII
ist am Seitenrande inseriert.
(Abb. 12, b)
Männchen
:
Die letzten 2 Abschnitte
der Kiefertaster sind dünn
und aufwärts gebogen (bis
90°).
Männchen
:
Subg. Aedes
Vs
:
Die Kiefer¬
lang wie die
Proboscis. Subg. Finlaya u.
Subg. Aëdimorphus gleich
lang wie Proboscis.
taster
so
allen
Bei
mitteleuropäi¬
schen Arten länger.
Weibchen
:
stumpf
(Abb. 17, c).
Die Cerci sind
Weibchen
Die
:
sind lang
(Abb. 17, b).
Cerci
gezogen
aus¬
In der vorliegenden Arbeit wurden folgende Arten eingehend
berücksichtigt :
1.
Culex pipiens LlNNÉ (1758)
C. molestus FoRSKAL (1775)
syn. C. domesticus Germar (1817)
syn. C. haematophagus FlCALBI (1893)
syn.
2. Aedes
Meigen (1830)
Culex vexans MeigEN (1820)
Culex sylvestris THEOBALD (1901)
Aedes enochorus Howard (1917)
vexans
syn.
syn.
syn.
BIOLOGIE UND
VERBREITUNG
3. Aedes geniculates OLIVIER
9
STECHMÜCKENARTEN
EINIGER
(1791)
A. Anopheles maculipennis Meigen (1818)
syn. A. lewisi Ludlow (1921)
syn. A selengensis Ludlow (1921)
5. Anopheles bifmeatus MEIGEN (1818)
syn. Anopheles claviger Meigen (1804)
syn. A. griscescens Stephens (1828)
syn. A. Villosus ROBINEAU, Desvoidy (1827).
Die wissenschaftlichen Bezeichnungen stammen einesteils
dem Griechischen, andernteils aus dem Lateinischen :
Aedes (gr)
Ochlerotatus
(gr.)
Anopheles (gr.)
Culex (lat.)
=
=
aus
unangenehm
lästig
=
nutzlos, beschwerlich
=
Bezeichnung
von
Mücke nach PuNIUS
Die Vulgärnamen sind mannigfaltig ; im deutschen Sprachgebiet
werden die stechenden Culiciden als Mücken, Gnitzen, Gelsen,
Schnaken und Schnogge, im Holländischen muggen, moeskieten
und im Englischen als Biting gnats und mosquitoes benannt. Die
französischen Ausdrücke sind : moustique, cousin ; die italienische
Bezeichnung
3.
:
zanzzara.
Allgemeines über die untersuchten Arten
Culex pipiens. Gemeine Singschnake.
Charakteristische Brutgebiete : Gartenfässer, Uberlaufgruben,
Jaucheansammlungen, eutrophe Wassertümpel. Larven und Puppen
nur in stehendem Wasser. Imago dringt in menschliche Behausungen
ein. Sie ist regelmässiger Blutsauger an Hausgeflügel und wildleben¬
den Vögeln. Vom Frühjahr bis im Spätsommer nicht so lästig für
den Menschen wie vor der Überwinterung, wo sie als Hausstech¬
mücke die grösste Bedeutung aller mitteleuropäischen Arten erlangt.
Die geographische Verbreitung erstreckt sich über die palaearktische Region, das östliche Nordamerika, Canada, Pazifik-Küste,
südliches Südamerika. Ausserdem ist diese Art in Nord- und SüdAfrika, Madagaskar, China und Japan beheimatet. In der eigentlichen
tropischen Zone fehlt sie jedoch.
Die Weibchen überwintern, nachdem sie eine Blutmahlzeit
eingenommen haben, von September bis April. Larven, Puppen und
Adulte können von August bis September (ausnahmsweise Oktober)
gefunden werden. Es findet .nur eine Diapause der überwinternden
Weibchen statt, da die Männchen im Herbst absterben.
10
WILLI
Aedes
BUTTIKER
vexans.
Charakteristische Brutgebiete sind temporäre Wasseransamm¬
lungen, überschwemmte Wiesen, Randgebiete flacher Seen und
langsam fliessender Flüsse (Altwasser). Die Imago dringt nicht in
menschliche Siedlungen ein, sticht aber den Menschen im Freien
häufig. Auch während des Tages kann A. vexans sehr zudringlich
werden, so dass diese Art in verschiedenen Gebieten (Nordamerika)
Sie ist regelmässiger Blut¬
zu den lästigsten Stechmücken gehört.
sauger an Menschen, Vögeln und Kaltblütern, und ihre geogra¬
phische Verbreitung erstreckt sich über die palaearktische, nearktische und die orientalische Region.
Die Larven können von April bis September gefunden werden,
die Imagines von Mai bis September. Innerhalb der Eihülle macht
die noch nicht geschlüpfte Junglarve die Latenzzeit durch. Euro¬
päische Beobachtungen stimmen darin überein, dass die Hauptent¬
wicklungszeit auf die Frühjahrs-Hochwasserperioden entfällt, und
dass nur eine Generation im Jahr wegen der Diapause erhalten wird.
Beobachtungen aus U. S. A. dagegen (New York : Matheson, 53)
stellen deutlich zwei Brutperioden fest, nämlich eine erste im Früh¬
jahr und eine zweite Ende August/anfangs September.
Aedes geniculates.
Typischer Vertreter der Baumhöhlenfauna und deshalb mit Ano¬
pheles plumbeus öfters vergesellschaftet. Die Larven von Aedes geni¬
culates können während des ganzen Jahres in hohlen Baumstämmen,
Asthöhlen und Baumstöcken angetroffen werden, sofern die Wasser¬
verhältnisse es erlauben. Imagines entwickeln sich in den Monaten
April bis September. A. geniculates dringt nicht in menschliche Sied¬
lungen ein, sticht aber den Menschen im Walde sehr häufig. Sie ist
in Europa bis Kleinasien beheimatet. Die Diapause wird im Larven¬
stadium durchgemacht.
Anopheles
oder
maculipennis
(Formengruppe)
:
Gabelmücke,
Malaria¬
Wechselfiebermücke.
Randgebiete seichter
Wassertümpel, Altwasserläufe und vegetationsreiche Ent¬
wässerungsgräben. Die Imagines der Rassen labranchiae, sicaulti und
atroparvus dringen ganz besonders häufig zur Blutaufnahme während
des Sommers in menschliche Siedlungen ein. Es sind dies die anthropophilen Rassen, im Gegensatz zu den zoophilen (Tabelle Nr. 53).
Sie sind für die Übertragung der Malaria in Europa verantwortlich.
Die unterschiedliche Bevorzugung der Wirte ist der Grund der
malariafreien und malariaverseuchten Anopheles-Gebiete. Die Bio¬
logie der einzelnen macu/ïpennrs-Rassen unterscheidet sich zum
Die charakteristischen Brutorte sind die
Seen,
BIOLOGIE UND
Teil sehr stark.
vorhanden.
VERBREITUNG
EINIGER
STECHMUCKENARTEN
Morphologische Unterschiede sind
nur
11
geringe
ist über ganz Europa, Nordafrika und
Aus der Schweiz sind drei Rassen
verbreitet.
Asien
in
Teil
zum
nachgewiesen, nämlich : messeae, melanoon, typicus.
(Im Gebiete des Rheins, des Ticino, der Rhône und des Mittel¬
landes sind diese maculipennis-Rassen häufiger, als allgemein ange¬
nommen wird. In gewissen Seegegenden ist diese Art Gegenstand
der hauptsächlichsten Mückenplage [Neuenburger-, Bieler- und
Anopheles maculipennis
Murtensee].)
Die Überwinterung der in der Schweiz vorkommenden Rassen
ist allein bei messeae ganz abgeklärt, wo die Weibchen in Kellern,
feuchten und kalten Ställen überwintern. Es scheint, dass die Weib¬
chen der andern beiden Rassen eher ausserhalb menschlicher Siede¬
lungen die kalte Jahreszeit überdauern (z. B. Höhlen, hohle Baum¬
strünke). Die Männchen sterben im Verlaufe des Herbstes ab, so
dass die befruchteten Weibchen eine Latenzzeit von Oktober bis
April durchmachen. Larven, Puppen und Imagines finden sich von
April bis Oktober.
A. bifurcatus.
Als charakteristische Brutorte gelten Waldtümpel, moorige Ent¬
wässerungsgräben, grössere und kleinere Wasseransammlungen hin¬
der Litoralzone der Seen ; ausnahmsweise können Larven auch in
Gartenfässern angetroffen werden. A. bifurcatus dringt nicht in
menschliche Behausungen ein, sticht aber den Menschen ausserhalb
ter
Siedlungen häufig, bevorzugt in der Regel Säugetiere. In Paläs¬
hauptsächlichste Überträger der Malaria. Die
Verbreitung erstreckt sich vor allem über Europa, Kleinasien und
Nordafrika, wobei neuerdings mehrere morphologisch kaum unter¬
scheidbare Rassen abgetrennt werden.
Die Larven lassen sich während des ganzen Jahres finden,' ins¬
besondere vom Herbst bis Frühjahr. Die Imagines weisen in Mittel¬
europa die grösste Häufigkeit im Frühjahr und Herbst auf. Die
Diapause wird in den Larvenstadien durchgemacht.
der
tina ist diese Art der
II.
MATERIALBESCHAFFUNG
UND
1.
ZUCHTTECHNIK
Materialbeschaffung
Das für die Versuche benötigte Material stammt z. T. direkt
dem Freiland, z. T. aber auch aus den eigenen Culicidenzuch~
ten. Wo kein kontinuierliches Züchten möglich war {Aedes vexans, A.
geniculatus, Anopheles maculipennis var. messeae, var. melanoon und
var. typicus), war ich auf geeignete Fundplätze angewiesen. Meist
konnten jeweils die genügenden Exemplare beschafft werden.
aus
Culex pipiens
Zürich-Altstetten
Zürich-Oerlikon
Zürich-Witikon
Zürich-Oerlikon
Aedes
überwinternde Ç
Eigelege, 3 Generationen
22.2.46
26.5.46
weitergezüchtet
überwinternde $
Eischiffchen
9.4.47
16.4.47
vexans
Rapperswil
Rapperswil
Rapperswil
Eier
Eier
Eier
an
Moos
1.4.47
15.4.47
25.4.47
Aedes geniculatus
Zürich-Affoltern
Larven
Larven
Larven
Larven
Larven
10.5.47
1.6.47
5.6.47
18.7.47
8.47
Flaach
Larven
Möhlin
Larven
1.6.47
10.9.47
Zürich-Rehalp
Freudwil b/Üster
Zürich, Rehalp
Zürich, Rehalp
Anopheles maculipennis
var.
var.
La
typicus
messeae
Sauge
La Sauge
überwinternde $
überwinternde ?
7.3.46
10.11.46
BIOLOGIE
La
UND
Sauge
La Sauge
Zûrich-Afîoltern
Zürich-Afloltern
var.
VERBREITUNG
EINIGER
13
STECHMÜCKENARTEN
überwinternde $
gravide $
gravide ?
gravide $
9.3.47
26.5.47
9.6.47
15.7.47
melanoon
La Sauge
La Sauge
Zûrich-Afîoltern
gravide $
gravide ?
Eier
29.3.46
26.5.47
30.5.47
Anopheles bifurcatus
Möhlin (Aargau)
Möhlin
Aarau-Rohr
Rapperswil
Larven
Larven
Larven
Larven
31.3.46
2.2.47
24.11.46
15.4.47
2. Zuchttechnik
Um die nötigen Larven, Puppen und Imagines für die Unter¬
suchungen zur Verfügung zu haben, war es vorerst notwendig,
geeignete Culicidenzuchten in Betrieb zu nehmen.
Nicht alle Arten konnten im Labor zur Fortpflanzung gebracht
werden, da besonders die Raumfrage für die Paarung von grösster
Wichtigkeit ist. Man unterscheidet eurygame von den stenogamen
Arten ; erstere kopulieren nur in sehr geräumigen Käfigen, letztere
stellen kleinere Ansprüche an die Umgebung, da die Männchen
keinen « Paarungstanz » durchführen. Verschiedene Autoren (RouBAUD, 73 ; Hackett u. Missiroli, 58) weisen auf diese Eigenschaf¬
ten hin. Ausserdem spielen Faktoren eine grosse Rolle, die im Wei¬
teren zur Diskussion kommen werden. Bei C. pipiens gibt es nach
neueren
Untersuchungen von RoUBAUD (73) nebeneinander eury¬
game und stenogame Rassen. Stenogamie ist eines der Hauptmerk¬
male von C. molestus (frühere Bezeichnung für C. pipiens autogenicus).
Für Laborzuchten ist also diese Art am besten geeignet, zumal sie
noch homodynam ist, d. h. eine unbegrenzte Anzahl von Genera¬
tionen ohne Diapause durchlaufen kann. Im Gegensatz dazu steht
C pipiens, die nach 4—5 Generationen zur Diapause übergeht.
Bei den durchgeführten C. piprens-Zuchten erreichte ich in kei¬
nem der zur Verfügung stehenden Käfige Kopulationen von C.
pipiens.
Dagegen gelang ein Weiterzüchten im Versuchshaus, dessen gesamter
Raum von 6 X 3 X 2,5 m als Käfig diente. Einschränkend ist zu
bemerken, dass dieses Pflanzenhaus ungünstige Temperatur- und
Feuchtigkeitsverhältnisse aufwies, so dass besonders die Sterblich¬
keit der Imagines während der Sommermonate sehr hoch
war.
Es
14
WILLI
BUTTIKER
konnten aber vom Mai bis August 1946 2 bis 3 Generationen erhalten
werden, da den Weibchen stets Eiablageplätze und Hühner als
Blutspender zur Verfügung standen.
Gute Zuchtresultate ergaben sich mit C. apicalis, dessen Weibchen
im Versuchshaus leicht in grosser Anzahl zur Eiablage schritten. Es
standen hiezu Aquariengläser und Heuaufguss zur Verfügung. Als
Blutspender wirkten drei Feuerunken (Bombinator pachypus), die
in einem Glasgefäss den Mücken leicht zugänglich waren. Die Eientwicklungsstadien dieser Culicidenart fanden aber in den vorliegenden
Untersuchungen keine Verwendung.
Über die Verhältnisse bei A. maculipennis gibt die Tabelle Nr. 53
(letzte Kolonne) Aufschluss. Demnach sind die drei schweizerischen
Rassen eurygam, können also nur in gross-dimensionierten Räumen
zur Fortpflanzung gebracht werden. Es wurde in den Jahren 1946
und 1947 wiederholt versucht, das gesamte Versuchshaus (6 X 3 X
2,5 m) als Zuchtkäfig für diese Art zu benützen ; ich erhielt aber
in keinem Falle Eigelege, obgleich geeignete Eiablageplätze in Form
von Glasschalen und Meerschweinchen als Blutspender zur Verfü¬
gung standen. Die Gründe hiezu sind ebenfalls in den ungünstigen
mikroklimatischen Verhältnissen, vor allem Temperatur und Luft¬
feuchtigkeit, zu suchen. Durch tägliches Bespritzen der Wände und
des Bodens konnte die Feuchtigkeit im Sommer bestenfalls auf 70 %
gebracht werden. Die Larven von var. messeae, melanoon und typicus
erhielt ich jeweils von eingefangenen Weibchen, die zur Eiablage
im Thermostaten gebracht werden konnten oder durch Sammeln
im Freiland.
Die in dieser Arbeit berücksichtigten Aëdes-Arten wurden nicht
auf die Züchtungsmöglichkeit geprüft.
Die Blutspender
Eine sehr wichtige Rolle spielen bei der Zucht die speziellen
Blutspender, die je nach Art zur Verfügung stehen müssen. Literatur¬
angaben sind aus letzter Zeit reichlich vorhanden (Bos, 9 ; LANGERON,
42). Es ist bekannt, dass eine Culex pipiens-Rasse Vögel bevorzugt,
eine andere hauptsächlich den Menschen sticht. Aus den eigenen
Laborbeobachtungen zu schliessen, nimmt C. apicalis sehr gern
Lurchenblut auf. Aedes Vexans sucht zur Blutaufnahme vornehmlich
Lurche auf, während Anopheles maculipennis je nach Varietät anthropophil oder zoophil ist. Messeae, melanoon und typicus gelten als
zoophil, wobei wohl ausschliesslich Säuger als Blutspender in Betracht
kommen. Diesen Tatsachen wurde Rechnung getragen, indem wäh¬
rend der ganzen Versuchszeit Hühner und Meerschweinchen, z. T.
auch Lurche (Bombinator pachypus) verfügbar waren. Die Wirtsspezifität konnte, aus den Resultaten zu schliessen, bestätigt werden.
BIOLOGIE
UND
VERBREITUNG
EINIGER
15
STECHMÜCKENARTEN
C. pipiens.
Folgende Liste gibt
aufnahme
eine Übersicht über die kontrollierte Blut¬
an :
Haushuhn (1—4 Monate alt)
Kanarienvogel
Meerschweinchen
Weisse Maus
Feuerunke (Bombinator pachypus)
über
200 Fälle
»
2
»
0
»
0
»
0
Bos und NlESCHULZ (8) geben Resultate über die Fütterung von
C. pipiens an Kanarienvögeln. Bei diesen Experimenten wurden spe¬
zielle Käfige verwendet, wobei 84 % der Mücken zur Blutaufnahme
schritten. Die Fütterung an Kanarienvögeln ist meines Erachtens
zu umständlich und zu
teuer, da stets mit Verlusten an Vögeln zu
rechnen ist (Verletzungen, zu starke Blutaufnahme).
Bos und NlESCHULZ (9) veröffentlichen weitere Resultate über
Fütterungsversuche an Haushühnern. Es zeigte sich, dass die Stech¬
freudigkeit jeweils am Ende der Überwinterung am grössten ist.
Weiter geht daraus hervor, dass in der ersten Nacht 19 %, nach
3 Tagen und Nächten 45 % und nach 6 Tagen und Nächten 71 %
aller verwendeten C. pipiens-Weibchen Blut sogen.
Für die Fütterung an Haushühnern wurde folgende Anordnung
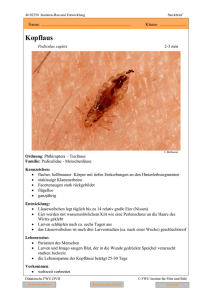
getroffen : In einem Draht-Gazekäfig, welcher die befruchteten
Abb. 1.
—
Zuchtkäfige : Im Vordergrund Drahtkäfig für Kücken
Hintergrund Gazekafig fur Mücken.
;
im
16
WILLI
BUTTIKER
Weibchen enthielt, wurde über Nacht ein Drahtgeffechtkäfig kleineren
Formats mit den darin eingeschlossenen Kücken eingesetzt. Die
Dimensionen betrugen für den Gazekäfig 58 X 43 X 43 cm, für das
Drahtgeflecht, je nach Grösse der Kücken :
14
19
40
x
X
X
10 X4,5
14 X 6,0
24 X 14
cm
(vergl. Abb. 1).
(für ausgewachsene Hühner).
cm
cm
darauf geachtet werden, dass die Maschenweite der Käfige
ist (18 X 18 mm im Minimum), damit ein freies Passieren
der Mücken zu den Blutspendern gewährt wird. Durch Zukleben
des Schnabels wurde es den Hühnern verunmöglicht, die im Käfig
herumfliegenden Culiciden zu erhaschen.
Es
muss
gross genug
Aedes
üexans.
Bei dieser Art wurde nur ein Versuch
Lurchen zeigte.
an Vögeln und
durchgeführt, der das
Saugen
Anopheles maculipennis
var.
messeae.
Versuchsweise wurden neben Meerschweinchen auch Hühner,
weisse Mäuse und Feuerunken abwechselnd in die Versuchskäfige
eingesetzt, wobei sich nachfolgende Resultate ergaben :
Meerschweinchen
100
% (April 1947) 1
iT»L
no/
Huhner
U
i
reuerunken
no/
n
Weisse Mäuse
U
%
%
l
J
)
„
aller
j
den
saugen•?
i
Weibchen,
w
12 Stück (1946)
Unmittelbar nach Beendigung der Winterruhe sind die Weib¬
chen am stechfreudigsten. Die Meerschweinchen, welche am Rücken
ungefähr auf eine Fläche von 6 auf 8 cm geschoren wurden, lockten
in den Abendstunden die Mücken nach kurzer Zeit an.
Als Aufenthaltsräume dienten wiederum die Gazekäfige mit den
Ausmassen 21 X 21 X 34 cm. über Nacht wurde in jeden Käfig
ein Meerschweinchen eingeschlossen, z. T. in einem kleinen Draht¬
käfig, z. T. ohne, diesen sodass die Mücken ungehindert an die
geschorenen Stellen Zutritt hatten.
Weisse Mäuse bewährten sich nicht als Blutspender, da sie
durch ihr unruhiges Verhalten die Mücken am ungestörten Saugen
hinderten. Trotzdem die weissen Mäuse auf einem Spannbrettchen
fixiert wurden, konnten sie sich doch so gut bewegen, dass die Anopheles-Weibchen meist vom Stechen Umgang nahmen.
biologie und
verbreitung
Die Ernährung
mit
einiger
stechmückenarten
der Imagines mit Fruchtsäften
Blut-Zuckersaft-Gemisch
17
und
Werden die Imagines der Stechmücken mit Pflanzensäften gefüt¬
wird ihre Lebensdauer dadurch beträchtlich verlängert.
so
Meist kommt es bei den von mir untersuchten Arten oder Rassen
zur
Fettspeicherung, jedoch nie zur Ei-Reifung. Die Männchen
nehmen überhaupt nur Fruchtsäfte oder Wasser zu sich, da ihre
Mundwerkzeuge nicht zum Stechen geeignet sind.
Die zuckerhaltigen Pflanzensäfte gelangen vor allem in den
paarigen und unpaarigen Saugmagen (Oesophagus-Ausstülpungen ;
vergl. Abb. 20). Hier werden sie bis zum Verbrauch gespeichert.
Vielfach werden Aufblähungen der Divertikel beobachtet, was auf
Gärungsvorgänge zurückzuführen ist. ScHAUDINN (cit. lit. 31) ver¬
mutete Symbionten, die einen alkoholischen Abbau der aufgenomme¬
nen Zuckerstoffe bewirken (COä-Bildung). Eysell (17) glaubte, dass
es sich bei den in dem Saugmagen befindlichen Gasblasen nur um
geschluckte Luft handelt. HECHT (31) dagegen weist auf die Möglich¬
keit einer häufigen, aber nicht obligaten Hefepilzinfektion hin.
Meine eigenen Beobachtungen bestätigen die Ansicht, dass keine
obligatorischen Symbionten in den Oesophagusdivertikeln vorhanden
sind (über mehr oder weniger regelmässig anzutreffende Mikro¬
organismen bei Culiciden siehe Steinhaus, 80), dass aber mit der
Nahrung eingedrungene Hefepilze den Zucker zur Gärung bringen.
Das Kohlendioxyd bewirkt eine Blähung der Saugmagen und des
gesamten Abdomens, so dass der Hinterleib fast vollständig durch¬
sichtig erscheint.
In der folgenden Zusammenstellung sind die Beobachtungsresultate über die Fütterung (überwinterte Weibchen) von C. pipiens
mit verschiedenen Fruchtsäften und Blut-Fruchtsaftmischungen
enthalten :
tert,
Fütterung
mit
Fruchtsäften.
Orangensaft
Orangensaft
als Nahrung verschmäht.
wird auch nach Tagen nicht angenommen.
und Traubenzucker, gemischt im Verhältnis 1:1, wird
Traubenzuckerlösung
unter 15 % ergab Blähungen des Darmsystems.
Vor allem füllten sich die Oesophagusdivertikel mit Blasen. Frisch
untersuchte, aus dem Winterquartier eingefangene Weibchen
52 Stück) mehr oder weniger grosse
zeigten bis zu 100 % (n
Luftblasen. Eine Traubenzuckerlösung von ca. 40 % hat keine
Schädigungen zur Folge. Die Verdauung ist normal, wobei eine
deutliche Fettspeicherung, nicht aber eineEireifung festzustellen ist.
Apfelstücke, welche noch Fruchtsaft abgeben können, werden als
=
Nahrungsquelle
2
auch benützt.
18
BÜTTIKER
WILLI
Mit etwas Wasser vermengtes Dattelfleisch bietet eine vorzügliche
Nahrung. Frisch eingefangene, überwinterte Weibchen und
Sommermücken beiden Geschlechts saugen gierig
Futter. Aus den bisherigen Erfahrungen
tenen
vom
zu
dargebo¬
schliessen,
bildet die Dattel das vorzüglichste Futter für Stechmücken. Die
bei Zimmertemperatur und
Lebensdauer gefütterter Culiciden
dreimal
ist
länger.
täglicher Wasserzugabe
—
—
Fütterung
mit
Blut-Fruchtsaft-Mischun§en.
Eine Mischung von Dattelfleisch mit frischem, nicht geronnenem
Schweineblut wird zum Teil von den Mücken aufgenommen.
5 Exemplare in einer Nacht.) Die Nahrung wird jedoch
(16 %
verdaut, ohne dass es dabei zur Ausreifung der Eier kommt. Die
Fettkörper vergrössem sich deutlich.
Serumflüssigkeit von frischem Schweineblut und Dattelfleisch,
Mischung 1:1. Diese Mischung wird ebenfalls von einer Anzahl
6 Exemplare in
der Weibchen als Nahrung verwendet (25 %
einer Nacht). Die Tiere zeigen eine Anschwellung des Fett¬
körpers, eine Eientwicklung findet aber nicht statt.
Ähnliche Resultate erhielten Täte und VlNCENT (13).
=
=
Die Dauer
des
Stechens
Zu diesen Beobachtungen eigneten sich überwinterte Weibchen
A. maculipennis var. messeae, die an meiner Hand zum Saugen
von
ansetzten
(März 1946).
Gemessene Zeiten des Stechens
Min.
Min.
Min.
Min.
Min.
1 Min.
1
1
1
1
1
(A. maculipennis
10 Sek.
Sek.
Sek.
Sek.
Sek.
45 Sek.
:
35
35
35
35
Mittelwert
:
1
1
2
2
3
/
Min. 50 Sek.
Min. 55 Sek
Min. 10 Sek,
Min. 15 Sek
Min. 10 Sek.
Min. 52 Sek
atroparvus [Amsterdam, Holland]
Stechzeit : 1 Min. 45 Sek.)
var.
Die Eireifung
:
(Praeovipositionszeit)
Wie in Abschnitt IV a) darauf hingewiesen wird, ist die Praeovipo¬
sitionszeit ebenfalls von der Temperatur abhängig. Im Verlaufe der
es sich, dass für eine normale Eireifungszeit
C schädlich sind, was sich im Ablegen
über
27°—28°
Temperaturen
unreifer, heller Eier und im Abgeben des im Darm gespeicherten
Blutes bemerkbar macht.
Untersuchungen zeigte
BIOLOGIE
UND VERBREITUNG
EINIGER
19
STECHMÜCKENARTEN
Die günstigsten Wassertemperaturen für die Eireifung liegen
für Culex pipiens, A. maculipennis var. messeae, var. melanoon, var.
typicus, um 18—23° C.
Die
Luftfeuchtigkeit wurde auf
ca.
90—100% gehalten. Es
scheint aber, dass dieser Faktor keine grosse Rolle spielt, da das
vollgesogene Weibchen genügend Flüssigkeitsmengen enthält und
somit von einer Wasseraufnahme unabhängig ist.
Freilandbeobachtungen gehen dahin, dass gravide Weibchen
einerseits in feuchten und andererseits in sehr trockenen Räumen
die Praeovipositionszeit verbringen. Der Temperatur kommt meines
Erachtens grössere Wichtigkeit zu.
Die Eiablage
Culex pipiens.
Die Grösse der Gefässe spielt bei Culex pipiens für die Eiablage
eine weitaus grössere Rolle als bei A. maculipennis. In den kleinen
Präparategläsern von 10 cm Höhe und 2 cm Durchmesser konnten
in keinem Falle Eier erhalten werden. Die Drahtgazekäfige von
43 X 43 X 58 cm eigneten sich besser dazu. Es wurden den Weib¬
chen Glasschalen mit jauchigem Wasser (Kaninchen- oder Hühner¬
kot) dargeboten, auf dessen Oberfläche die Eischiffchen in der Regel
über Nacht abgelegt wurden. Der Vorzugstemperatur bei der Eiablage
wurde Rechnung getragen, indem zwischen Käfigboden und Wasser¬
schale ein Heizkissen gelegt wurde. Die Wassertemperatur betrug
meist 21—24° C (vergl. Hecht, 32).
Anopheles maculipennis (var.
messeae,
melanoon, typicus).
Versuchsresultate über die Vorzugstemperatur einiger Anophelen
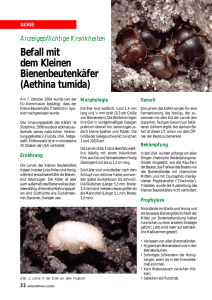
bei der Eiablage veröffentlichten Weyer und HüNDERTMARK (39.)
Bis kurze Zeit vor der Eiablage blieben die Weib¬
chen in der Temperaturstufe von 22° und wurden
dann in die Stufe von 27° übersiedelt. Stand¬
gläschen von 11 cm und 3 cm Durchmesser dienten
als Gefässe zur Eiablage, in welche in den untersten
Teil Brunnenwasser eingefüllt und ein Streifen
Fliesspapier eingelegt wurde. Als Abschluss nach
oben (siehe Abb. 2) diente ein Wattebausch.
—
Die Pflege
der
Eier
Nach der Eiablage gelangten die Eier in die
Aquariengläser (12 Liter Inhalt)
oder in Einmachgläser von 1 bis 2 Liter Inhalt.
entsprechenden
Abb. 2.
für die
—
Gefäss
Eiablage
(A. maculipennis).
20
WILLI
Die Pflege
BÜTTIKER
der
Larven
Culex pipiens.
WEYER (93) zeigte, dass
C. pi'piens-Larven sich schneller mit
Kalbsleber
als
Blut entwickelten ; die Larven¬
mit
pulverisierter
sterblichkeit war aber sehr hoch, nahm jedoch bei Zugabe von Heu¬
aufguss stark ab. In einer spätem Arbeit (94) wies derselbe Autor
darauf hin, dass er bei seinen Larvenzuchten Heuaufguss allein
erfolgreich verwendete. Roubaud (74) benützte Weizen und erhielt
mit gereinigtem Kasein zusammen die besten Resultate. Für C.
pipiens und C. aegypti war auch Sojabohnenmehl sehr günstig. Die
Zuchttemperatur betrug ca. 26° C. Mit Vitamin B versehenes Futter
ergab schlechtere Resultate, vielleicht im Zusammenhang mit der
starken Bakterienentwicklung.
Eigene Versuche mit Heuaufguss, Meerschweinchenkot und
getrockneter Lebersubstanz ergaben die besten Resultate. Es wurde
darauf geachtet, keinen üppigen Pilzüberzug auf der Wasserober¬
fläche zu erhalten, der den Larven die Luftaufnahme verunmöglicht.
Die Lebersubstanz * wurde ungefähr alle 3 Tage in kleinen Quan¬
titäten dargeboten, so dass kein grosser Futterüberfluss vorhanden
war. Der Meerschweinchenkot gelangte in zerriebenem Zustand in die
Kulturen ; diese zusätzliche Nahrung wurde gierig von den Larven,
welche sich auf dem Grunde aufhielten, aufgenommen. Man muss
darauf bedacht sein, einerseits keine Hungerkulturen, andererseits
keine « Uberschusskulturen » (Intoxikationszuchten) zu bekommen.
Ulmann (89) geht in seiner Publikation eingehend auf die Bevöl¬
kerungsprobleme von Normal-, Hunger- und Intoxikationszuchten
bei C. molestus ein. Bei Nahrungsmangel und Nahrungsüberfluss
ergeben sich ähnliche Resultate, die hauptsächlich in der Verschie¬
bung des Geschlechtsverhältnisses zugunsten der Männchen und
in der Reduktion der Eizahl bei graviden Weibchen bestehen.
Es ist nötig, das Nährmedium täglich von abgestorbenen Larven
und Puppen und von grösseren Nahrungsresten zu reinigen. Alle
3 bis 5 Tage erfolgte ein Wasserwechsel, wobei die Entwicklungs¬
stadien in neue Kulturgefässe übertragen wurden. Die Fütterung
führte ich alle 2 bis 3 Tage mit Hilfe eines kleinen Löffels (Inhalt
5 mg) durch. Die Larven nahmen das fein zerriebene Futter
ca.
(getrocknetes Leberpulver und Hundebiscuits) schon von der Ober¬
fläche weg.
Anopheles maculipennis (verschiedene Varietäten).
Nachdem die Junglarven in den Präparategläsern (Abb. 2) aus¬
geschlüpft waren, kamen sie entweder in Einmachgläser von 1 —2 Liter
Geliefert
von
Fa.
«
Opopharma » G.
m.
b. H., Zürich.
BIOLOGIE UND
VERBREITUNG
EINIGER
STECHMÜCKENARTEN
21
Inhalt oder in Aquariengläser von 12 Liter Inhalt, welche sich am
besten bewährten.
Unter die Zuchtgläser kam ein elektrisches Heizkissen zu liegen,
das nach Wahl Temperaturen von 18—27° C lieferte. Nach den
anfänglichen Misserfolgen in der Aufzucht der Larven, die durch
den Lichtmangel verursacht wurden, verwendete ich elektrische
Glühbirnen (60 u. 75 Watt). Wie später auf Abb. 38 hingewiesen
wird, trat durch die Lichteinwirkung eine Entwicklung mit nur
geringen Larvenverlusten ein.
Das Futter bestand aus einer Mischung von Hundebiscuit und
1
getrockneten Daphnien (Copepodae, Crustacea) im Verhältnis 1:1.
Zur Vermeidung der Verpilzung wurde nur in kleinen Mengen"
gefüttert, sodass die dargebotene Nahrungsmenge für ca. 12 Stunden
ausreichte. Hundebiscuit allein ergab auch befriedigende Resul¬
tate.
Weyer (94) berichtet über Fütterungsversuche an A. maculipennis mit Piscidin, Hefe, Leber und Blut, sowie einer Vorstufe
des Hypophysen-Vorderlappen-Präparates « Preloban » (I. G. Far¬
ben). Das letztere schien einen fördernden Einfluss auf die Larven¬
entwicklung bewirkt zu haben. Eigene Versuche mit dem handels¬
üblichen Präparat « Preloban » blieben bei Unterbelichtung der
Kulturen ohne Erfolg ; eine Verbesserung der Zuchtresultate konnte
bei richtiger Lichtdosierung nicht beobachtet werden. Ohne mir
ein endgültiges Urteil zu bilden, scheint es aber sehr unwahrschein¬
lich, dass dieses Präparat, welches bei Menschen hormonale Man¬
gelerscheinungen (Endokrine Magersucht, Amenorrhoe) beseitigt,
geeignet ist, Wachstumsstörungen bei Culiciden-Larven zu elimi¬
nieren.
BoRRANI (5) u. a. vermuten, dass Lemna-Arten eine hemmende
Wirkung auf Anopheles ausüben. Ein Laborversuch zeigte, dass in
einem Gefäss 50 Junglarven von A. maculipennis var. messeae schon
innerhalb 2—3 Tagen abstarben. Die Wasseroberfläche war so
kompakt mit den Blättern dieser Pflanze überdeckt, dass dieser
Abschluss den Larven die Atmung verunmöglichte.
Aedes
vexans.
Die Aufzucht von Aedes vexans bot keine Schwierigkeiten. In der
Regel können durch Begiessen der Moospflanzen, wo die Weibchen
ihre Eier abgelegt haben, die Eilarven zum Schlüpfen gebracht
werden. Diese schlüpfen, sofern sie die Diapause schon durch¬
gemacht haben, innerhalb 15—20 Minuten. Als Zuchtgefässe bewäh¬
ren sich vor allem Aquariengläser von ca. 12 Liter Inhalt; getrock¬
netes Riedgras und Moosstengel mit ihrem Überzug an Mikroben
1
Früher öfters als Fischfutter
unter
dem Namen
«
Piscidin
»
im Handel.
22
WILLI
BUTTIKER
bilden ein gutes Futter für alle Larvenstadien. Aber auch Einzel¬
zuchten in 10—12 cm hohen und 3—4 cm dicken Glaszylindern
ergeben gute Erfolge. Das Auswechseln des Wassers scheint nicht
nötig zu sein, ebenso ist das Zugeben von Hundebiscuit normaler¬
weise nicht nötig.
Aedes geniculates.
RoUBAUD (74) veröffentlicht Resultate über die Notwendigkeit
des Wechsels zwischen Feuchtigkeit und Trockenheit, Wärme und
Kälte, damit die Eier von A. geniculates überhaupt schlüpfen kön¬
nen ; für die Latenzzeit der Eier führt er eine Dauer von 8—13 Mona¬
ten
an.
Meine eigenen Erfahrungen gehen dahin, dass die Larvenkul¬
turen vielfach unter Verpilzung leiden. Faulende Holzbestandteile,
wie Stengel und Laub, wurden jeweils mitsamt ca. 500 cm3 Stock¬
wasser ins Labor mitgenommen, um für die Larven möglichst natür¬
liche Zuchtbedingungen zu haben. Die zusätzliche Nahrung bestand
aus fein zerriebenem Hundebiscuit, das täglich in kleinen Mengen
den Kulturen zugegeben wurde. Bei Uberdosierung litten sie jedoch
stets unter Verpilzung, besonders in den oberen Temperaturstufen.
Aus normalen Larvenzuchten ergaben sich mühelos Puppen, die
sich auch gut zu Imagines entwickelten.
Culex apicalis.
Im Verlauf der Monate Mai, Juni und Juli erhielt ich im Pflanzen¬
haus ca. 100 Eigelege von C. apicalis (syn. C. pyrenaïcus). Die schlüp¬
fenden Larven gediehen bei Zuchtbedingungen, wie sie für C. pipiens
vorlagen, gut, besonders in den mit Grünalgen versehenen Zucht¬
behältern. Die ausschlüpfenden Imagines hatten die Möglichkeit,
Feuerunken Blut aufzunehmen, wodurch es ihnen ermöglicht
an
wurde, Eier auszureifen. Offenbar fand die Kopulation im geschlos¬
senen Räume des Versuchshauses statt.
Die Pflege
der
Puppen
Das Puppenstadium nimmt keine Nahrung
dere Pflege ist daher nicht anzuwenden.
Die Pflege
der
zu
sich. Eine beson¬
Imagines
Die Imagines erhielten täglich morgens und abends Wasser, das
fein zerstäubt in die Draht-und Baumwoll-Gazekäfige gespritzt wurde.
Den Imagines wurde Dattelfleisch, das sich am besten als Nahrung
eignet, oben auf dem Käfig dargeboten, wobei das Futter durch die
Maschen des Käfigs hindurch erreicht werden konnte. Leider war
BIOLOGIE UND
VERBREITUNG
EINIGER
STECHMÜCKENARTEN
23
der
Feuchtigkeitsgehalt und die Temperatur des Versuchshauses
grossen Schwankungen ausgesetzt. Während des Sommers betrugen
die Temperatur-Minima und -Maxima ca. 20° und 38° C, die Min¬
dest-, und Höchstwerte der Luftfeuchtigkeit ca. 30 %, resp. 80 %
F. Die optimalen Werte halten sich ungefähr in folgendem Rahmen :
r.
Temperatur
Luftfeuchtigkeit
22 bis 28° C
.
(normale Aktivität)
80-100%.
Aus der Bestimmung des Thermohygrogrammes geht hervor, dass
bei niederen Temperaturwerten und hoher Luftfeuchtigkeit die
längste Lebensdauer erreicht wird.
III.
KÖRPERBAU
Die Culiciden sind Insekten mit holometaboler oder vollstän¬
; sie durchlaufen nach dem Ei vier Larvenstadien,
verpuppen sich und entwickeln sich dann zur geschlechtsreifen Imago.
Im folgenden Kapitel kommen ausschliesslich die Entwicklungs¬
stadien der in dieser Arbeit berücksichtigten Arten zur Besprechung.
diger Verwandlung
1. Das Ei
Die Eihülle besteht aus drei verschiedenen
Schichten. Die innerste, welche den Eidotter
und den Keim umschliesst, wird als Membrana
vitellina bezeichnet. Die mittlere und die äusserste
Schicht werden als Chorion zusammengefasst.
Das Endochorion (Mittelschicht) ist opak und
hart, während das Exochorion (äusserte Schicht)
biegsam und durchsichtig ist.
Beim frischgelegten Ei ist das Endochorion
ebenfalls durchsichtig, aber unter natürlichen
Umständen wird es nach kurzer Zeit dunkler.
Culex pipiens.
Die Eier werden auf die Wasseroberfläche
abgelegt und zu sogenannten « Eischiffchen »
(Abb. 3) von 180 bis 350 Stück zusammengeklebt.
Abb. 3.
Culex pipiens, Eigelege
von
oben. Vergr.,
7-fach. Orig.
•
24
BÜTTIKER
WILLI
Selten steigt die Eizahl über 350 Stück ; Gelege von weniger
als 100 Eiern sind bei C. pipiens sehr selten. Ein einzelnes aufgebro¬
chenes Ei gelangt in Abb. 4, a zur Darstellung. Bei fortgeschrittener
Entwicklung ist der Embryo durch die Eihülle hindurch sichtbar1.
Das Auszählen von Eigelegen aus der Umgebung von Zürich
ergab folgende Zahlen (Tab. 1)
:
Tabelle Nr. I
Datum
Fundort
Anzahl
d. Eier
Datum
Fundort
Anzahl
d. Eier
15.4.47
Zürich ETH
Zürich ETH
170
205
4.5.47
12.5.47
Zürich ETH
Zürich ETH
290
316
24.4.47
Zürich ETH
Zürich ETH
320
360
1.6.47
18.6.47
Zürich ETH
Zürich ETH
283
336
Zch-Örlikon
280
300
305
212
325
294
18.6.47
18.6.47
18.6.47
Zürich ETH
Zürich ETH
Zürich ETH
308
284
310
25.4.47
»
»
»
»
»
Frisch gelegte Eier sind weiss, werden dann gelblichgrün und
innert 1—2 Stunden schwarz. Die Grösse der Eier schweizerischer
Herkunft beträgt im Durchschnitt :
Länge 0,720
mm
Breite
mm
0,131
~«
n
~~
^
Mit Hilfe des Eizahnes (Abb. 4, m), der bei Culex pipiens scharf
zugespitzt ist, öffnet die Junglarve die Eihülle. Kopfabwärts gelangt
die schlüpfende Larve ins Wasser.
Aedes
vexans.
Die Eier werden von den Weibchen einzeln in feuchtes Gras,
in Moos usw. gelegt. Bevorzugt sind Orte, die periodisch über¬
schwemmt werden. Das Ei von A. vexans wird in Abb. 4, d dar¬
gestellt. Mit Hilfe des Eizahnes (Abb. 4, h, i) gelangt die Eilarve
nach aussen, indem der Körperinnendruck einen Chitinknopf am
Kopf (Druckknopf-System) an die Eihülle drückt und diese zum
Reissen bringt.
nur
1
Die Eier der andern mitteleuropäischen Culex-Arten lassen sich nicht oder
schwer von den p/prens-Eiern unterscheiden.
BIOLOGIE UND VERBREITUNG
Abb. 4.
60-fach.
25
EINIGER STECHMUCKENARTEN
C. pipiens, aufgebrochenes Ei, 50-fach.
b, C. pipiens, Ei mit Embryo,
C. pipiens, abnormes Ei, 50-fach.
d, A. vexans, Ei, 50-fach.
e, A.
maculipennis var. messeae, aufgebrochenes Ei, 50-fach. —/, A. macalipennis var. typicas,
50-fach.
h, A. vexans,
g, A. maadipenms var. messeae, von der Seite gesehen.
Eizahn von der Seite, 300-fach.
i, A. vexans, Eizahn von oben, 300-fach.
k, A.
maculipennis var. messeae, Eizahn v. oben, 300-fach.
I, A. maculipennis var. messeae,
Mikropyle des Eies, ca. 250-fach.
m, C. pipiens, Eizahn von oben, 300-fach.
a,
—
—
—
c,
—
—
—
—
—
—
—
—
Anopheles maculipennis.
Die
einzeln
aufgelegten
Eier
besitzen
beidseitig luftgefüllte
Schwimmkammern. Der Umriss der Eier ist spindelförmig. Die
Körnelung des Exochorions bedingt das Zeichenmuster, welches für
die einzelnen Varietäten charakteristisch und deshalb
tischer Bedeutung ist (Abb. 4, e ; f).
von
systema¬
26
WILLI
BÜTTIKER
Der Eizahn beruht ebenfalls auf dem Druckknopf-System ; die
drückt beim Schlüpfen einen Chitinknopf an die
Eihülle, wobei die letztere aufreisst (Abb. 4, k). Die Mikropyle liegt
am vordem Ende des Eies und ermöglicht während der Eiablage das
Eindringen der Spermien aus den Spermatheken (Abb. 4,1).
Die Eizahl eines legenden Weibchens ist starken Schwankungen
unterworfen und beträgt nach eigenen Beobachtungen ca. 150 Stück
Körperflüssigkeit
(Tab. 2).
Tabelle Nr. 2
Gesogen
12.3.46
12.3.46
12.3.46
20.3.46
20.3.46
20.3.46
26.5.47
20.3.46
26.5.47
26.5.47
26.5.47
Varietät
Abgelegt
15.3.46
15.3.46
Herkunft
messeae
messeae
16./17.3.46
messeae
23.3.46
23.3.46
23.3.46
messeae
messeae
messeae
29./30.5.47
messeae
22.6.47
23.3.46
messeae
melanoon
29./30.5.47
29./30.5.47
29./30.5.47
melanoon
melanoon
melanoon
melanoon
2.8.47
2.8.47
typicus
La Sauge
La Sauge
La Sauge
La Sauge
La Sauge
La Sauge
La Sauge
Zürich
La Sauge
La Sauge
La Sauge
La Sauge
Zürich
Zürich
Eizahl
ca.
ca.
ca.
ca.
145
190
120
125
120
120
142
183
115
155
240
253
143
155
Junge Weibchen legen mehr Eier als alte ; nach bisherigen Beo¬
bachtungen kann ein Weibchen gegen 10 Gelege legen.
Das Ausmessen schweizerischer macu/i'penm's-Eier ergab im Mittel
folgende Werte (Tab. 3) :
n
=
20
Tabelle Nr. 3
Var.
Länge
Schwimmkammerlänge
messeae
691
266
Var. melanoon
Var. typicus
710
301
670
265
BIOLOGIE
UND VERBREITUNG
EINIGER
STECHMÜCKENARTEN
27
2. Die Larven
Die Larven (IV. Stadium) aller drei in dieser Arbeit berücksich¬
tigten Culiciden-Gattungen sind als Ubersichtsbilder dargestellt
(Abb. 5 und 6). Anopheles lässt sich leicht von den Vertretern der
Culicini unterscheiden, da die Larven der ersteren waagrecht an
der Wasseroberfläche liegen und dorsal am VIII. Segment nicht
erweiterte Tracheenöffnungen haben, während die Culicini kopfabwärts an der Wasseroberfläche haften und ein deutliches Siphonalrohr besitzen. Für die Systematik ist die Körperbehaarung (Chaetotaxis) sehr wichtig (MARSHALL, 46).
Culex pipiens.
Die Zeichnung (Abb. 7) gibt die Anordnung der Mundwerk¬
in schematischer Weise wider. Die paarigen, fächerförmigen
Mundbürsten der Oberlippe (Abb. 8) sind beweglich und führen
durch ihren rhythmischen Schlag der Larve im Wasser Infusorien,
Algen, Bakterien und weitere Kleinlebewesen zu. Die paarigen Oberund Unterkiefer tragen ebenfalls Strudelhaare, die durch ihre Bewe¬
gung an der Zuführung der Nahrung beteiligt sind. Die Taster der
Unterkiefer sind rudimentär. Die Fühler tragen nebst büscheligen
Haaren an ihrer Spitze eine Anzahl Sinnesorgane. Die Unterlippe
zeichnet sich infolge stärkerer Chitinisierung durch dunklere Farbe
aus. Der Epi- und Hypopharynx kleiden die Mundhöhle nach oben
und unten aus, und beide gehen an ihrem Ende in den Pharynx
über. Der Hypopharynx im speziellen trägt eine grosse Anzahl
Geschmacksorgane, die am vorderen Ende als Zäpfchen und Papillenreihen gut erkenntlich sind.
Von den andern mitteleuropäischen Cü/e*-Arten lässt sich
C. pipiens im Larvenstadium am besten an der Form der Abdominalsiphonen und der Siphonalindices (Basisbreite zu Gesamtlänge)
zeuge
unterscheiden.
C. pipiens
C. apicalis
C. hortensis
Die
Siphonalindex
»
»
4
7
5,6
Abb. 10, A
Abb. 10, B
Abb. 10, C
Bedornung der
Siphonen weist ebenfalls charakteristische
als Unterscheidungsmerkmale verwendet
werden können. C pipiens und C. apicalis weisen normalerweise
4 Borstengruppen auf, im Gegensatz zu C. hortensis, welcher dorsal
Eigenschaften auf,
bis 8
die
Borstengruppen trägt. Weiter
lassen sich die IV. Larven
von
Abb. 5.
Anopheles maculipennis; IV. Larvenstadium. Vergr.
—
ca.
I5-fach Orig.
Abb. 6.
A, IV.-Larve von
Aedes vexans.
B, IV.-Larve
von Culex pipiens.
—
—
Abb. 7.
Mundwerkzeuge
einer Culicidenlarve, schema¬
tisiert.
kt,
f, Fühler.
Kiefertaster.
ok, Ober¬
kiefer.
ol, Oberlippe.
ul, Unter¬
uk, Unterkiefer.
—
—
—
—
—
—
—
lippe.
Abb. 8.
a,
—
Culex pipiens. Viertes Larvenstadium, Mundwerkzeuge, Orig. 75-fach.
d, Unterlippe.
e, Fühler,
b, Oberkiefer.
c, Unterkiefer.
Oberlippe.
—
—
—
f, Epipharynx.
—
g,
Hypopharynx.
—
30
WILLI
BÜTTIKER
C pipiens und C. apicalis an der Anordnung der Haare unterschei¬
den, abgesehen vom Siphonalindex, der bei der ersten 4, bei der
letzteren 7 beträgt.
Die Gewichtszunahme während der Larvenentwicklung wurde
bei jeder Häutung verfolgt, wobei folgende Zahlen resultierten
(Tab. 4)
:
C. pipiens.
Gewichtszunahme,
n
Stadium
=
10
*
pro
Zuwachs¬
faktor
Stück
ms
TABELLE Nr. 4
pro
Stadium
Stück
mg
Frisch geschl. Eilarve
0.1
IV. Larve,
III. Larve
0.2
0.8
2,3
Entwicklung
Puppe, anf. d. Entw.
Imago, v. d. Schlüpfen
Imago, mit Blut ca.
am
Ende d.
....
2
4
3
Zuwachs¬
faktor
.
4,1
4,1
4,0
7,0
2
2
Auf Abb. 9 sind die obigen Verhältnisse bildlich dargestellt.
Von besonderem Interesse ist eine Erscheinung, welche man
Mantiden und Dixippus kennt, dass eine Eiablage resp. Ge¬
von
schlechtsreife der weiblichen Tiere erst nach einer weiteren Ver¬
doppelung des Gewichts eintritt. Die Praeovipositionsperiode ent¬
spricht also physiologisch
(BODENHEIMER, 4).
einem
weiteren
Entwicklungsstadium
Anopheles maculipennis.
Die Kopfzeichnung der IV.-Larven kann in den meisten Fällen
als Bestimmungsmerkmal für die beiden ähnlich aussehenden. A. macu¬
lipennis und A. hifurcatus verwendet werden. Grenzfälle, wo eine
sichere Bestimmung anhand der dunklen Farben nicht mehr möglich
ist, konnten bei schweizerischen Larven festgestellt werden. Die
flächenhafte Ausdehnung und Intensität dieser dunklen Flecken
variiert sehr stark, sowohl an Larven verschiedener wie gleicher
Herkunft (Abb. 11, A und B). Mit Hilfe der Chaetotaxis kann die
Artzugehörigkeit bestimmt werden.
Die Rückenzeichnung der Larven variiert ebenfalls stark und
wird durch die verschieden grosse Ausdehnung der Zellgruppen von
Oenocyten bestimmt.
Die Gewichtszunahme im Laufe der Larvenentwicklung verlief
folgendermassen (Tab. 5)
:
BIOLOGIE
UND
VERBREITUNG EINIGER
Culex pipiens.
Abb. 9.
Gewichtszunahme während
31
STECHMÜCKENARTEN
—
der
Larvenentwicklung. I.
IV. Larvenstadium.
bis
•y
Ti/
yPuppê
*
-
z-
"y
/
n^i
i
6
Taye
8
Abb. 10.
Abdominalsiphon
verschiedener Cu/ejr-arten ;
IV. Larvenstadium.
A,
Culex pipiens, Siphonalindex
4.
B, C. apicalis, Sipho¬
nalindex
7.
C, C. hortensis, Siphonalindex
5,6.
—
—
=
—
=
—
=
Abb. 11.
Kopfkapseln
der IV. Larven von A,
—
Anopheles maculipennis ;
Zürich-AfToItern.
B,
—
Anopheles
bifurcatus
Möhlin.
;
32
WILLI
BUTTIKER
Anopheles maculipennis
Gewichtszunahme
n
var.
messeae
TABELLE Nr. 5
10
=
IV. Larve
Frisch geschlüpfte
Eilarve (I.)
0.1
0.2
0.9
2.7
mg/St.
Stadium
mg/St.
Stadium
am
Ende der
6.4
6.4
6.2
Puppe, frisch geschl..
Imago, vor d. Schlüpfen
.
Die Gewichtszunahme von der frischgeschlüpften Eilarve bis zur
geschlüpften Imago beträgt demnach bei Anopheles maculipennis
ca.
das 60—fache und bei Culex pipiens das 40—fache.
3. Die
Puppen
Das IV. Larvenstadium geht durch eine Häutung in die Puppe
(Abb. 12, a) über, die sehr beweglich ist. Der Körper besteht aus
einem kräftig entwickelten Céphalothorax und einem gut geglie¬
derten Abdomen, welches sich aus acht Segmenten zusammensetzt.
Mit Hilfe der Ruderfinne (« Paddel ») kann die Puppe sich lebhaft
hin und her bewegen. Am Céphalothorax befinden sich zwei trompetenförmige Atemröhren, die mit ihren Öffnungen an die Wasser¬
oberfläche ragen.
Die morphologischen Unterschiede der Puppen mitteleuropäi¬
scher Culiciden-Arten sind gering. Culex pipiens unterscheidet sich
von Aedes vexans und Anopheles maculipennis durch das vom Seiten¬
rande abgesetzte Haar A/VIII (VIII. Segment, vergl. Abb. 12, b;
c ; d) ; i4ê'(/es-Arten unterscheiden sich von Anopheles im Puppen¬
stadium an den zusätzlichen Paddelborsten, die bei Aedes fehlen
(Marshall, 46).
Bei Puppen von A. maculipennis ergaben
folgende Werte (Tab. 6) :
n=
die
Längenmessungen
Tabelle Nr. 6
10
Freiland
Körperlänge, ausgestreckt
Körperende).
(Kopf
—
Abb. 13 stellt einen
.
.
5,8
mm
Labor
4,9
mm
Querschnitt durch die Puppe von C. pipiens dar.
BIOLOGIE
UND
VERBREITUNG
EINIGER
STECHMÜCKENARTEN
33
Abb. 12.
b-d, Schwanzfinnen (Paddel) von Culiciden.
a, Puppe von Culex pipiens.
b, Culex pipiens.
d, Aedes vexans.
c. Anopheles maculipennis.
e, Labiumf, Spitze der Oberlippe.
spitze von A. maculipennis.
g, Spitze des Hypopharynx.
h, Spitze des Oberkiefers.
i, Spitze des Unterkiefers.
k, Schematischer Quer¬
schnitt durch die Mundwerkzeuge. Punktiert : Oberlippe ; gekreuzt schraffiert :
Oberkiefer ; weiss : Unterkiefer ; schwarz : Hypopharynx ; einfach schraffiert : Unter¬
1, Sechstes Tergit von Culex apicalis.
lippe.
m, Sechstes Tergit von Culex pipiens.
n. Sechstes Tergit von Culex horlensis.
—
—
—
—
—
—
—
—
—
3
—
—
—
—
—
34
WILLI
4. Die
BÜTTIKER
Imagines
(Abb. 14)
Am
Kopf sind Scheitel, Hinterhaupt und Schläfen mit Haaren
und Schuppen bedeckt. Die Mundwerkzeuge sind stark verlängert
und beim Weibchen noch in gleicher Zahl wie bei beissenden Mund¬
teilen. Als Beispiel gelangen die Mundteile eines Männchens und
eines Weibchens von Anopheles maculipennis vergleichsweise zur Dar¬
stellung (Abb. 15 und Abb. 16) : Die unpaare Oberlippe (ol) umfasst
den Nahrungsgang ; ihr angelagert sind die paarigen Ober- und
Unterkiefer (ok resp. uk), die beim Männchen eine schwächere Aus¬
bildung zeigen. Der Hypopharynx als unpaarer Teil enthält den
Speichelgang. Die Unterlippe (ul) umfasst sämtliche Mundwerk¬
zeuge, wie dies aus dem schematischen Rüsselquerschnitt (Abb. 12, k)
hervorgeht.
Die Oberlippe ist im Prinzip eine Hohlnadel ; der darin vor¬
handene Nahrungsgang geht an der Basis in den Pharynx über.
Die Oberkiefer (ok) sind bei den Weibchen dünne, sehr schmale
Blätter, die am Spitzenrand fein gesägt sind. Die Unterkiefer (uk)
sind ähnlich wie die Oberkiefer, tragen aber stärkere Zähne. Am
Grunde der Unterkiefer befinden sich die viergliedrigen Kiefer¬
taster (die reduziert sein können). Der Hypopharynx (hy) ist ein
langes, schwertförmiges Blatt, in dessen mittleren Verdickungsleiste der Ausführungsgang der Speicheldrüse bis zur Spitze geht.
An das lange; ziemlich gleichdicke Kinn schliessen die beiden Labellen
an, welche die ungebildeten Lippentaster darstellen (Abb. 12, e).
Diese Labiumspitze übernimmt beim Stechakt die Führung des
Stechborstenbündels. Die Unterlippe bleibt also ausserhalb der
Wunde. Die Fühler können als « fadenförmig, quirlig behaart »
bezeichnet werden.
Der Thorax trägt nebst den feingegliederten Beinen die Flügel
mit ursprünglicher Aderung (Abb. 17, d). Das Abdomen ist bei
Anopheles behaart, bei den Vertretern der Culicini reichlich beschuppt.
Morphologisch lassen sich die adulten Vertreter mitteleuropäi¬
scher CaleX'Arten am besten an der Tergitzeichnung unterscheiden
(Abb. 12,1,
m,
n)
;
(MARSHALL, 46).
S. Das
Darmsystem
Nach der Ernährung unterscheiden sich Larven und Imagines
der Culiciden auffallend stark. Das Puppenstadium nimmt keine Nah¬
von der Atmung abgesehen
rung auf, sondern begnügt sich
mit den gespeicherten Reservestoffen. Die beweglichen Mund—
—
Abb. 14.
messeae,
Abb.
13.
—
—
A. maculipennis
Weibchen. 6-fach
var.
Orig.
C. pipiens; Querschnitt durch
Puppe, kurz vor dem Schlüpfen der Imago.
I, Auge.
2, Oberes
Vergr. 20-fach Orig.
3, Unteres CerebralCerebral-Ganglion.
5, Fühler.
4, Beinanlagen.
Ganglion.
—
—
—
—
—
6, Riechnerv.
—
7, Thoracalmuskulatur.
8,
9, Mitteldarm.
Oesophagus-Ausstülpungen.
10, Magen.
II, Fettkörper.
12, Bauch¬
muskulatur.
13, Tracheen._— 14, Malpighi15, Abdominalpaddel.
gefässe.
—
—
—
—
—
—
—
—
Abb. 15.
A. maculipennis var. messeae,
Männchen. Kopf mit Mundwerkzeu¬
f, Fühler.,
hy
gen, 25-fach Orig.
kt, Kiefertaster.
Hypopharynx.
ol, Oberlippe.
ok, Oberkiefer.
uk, Unterkiefer.
ul, Unterlippe.
—
—
—
—
—
—
—
—
A. maculipennis var. messeae, Weibchen.
Abb. 16.
Kopf mit Mundwerkzeugen, 25-fach. Orig.
f, Fühler.
hy, Hypopharynx^— kt, Kiefertaster.
uk, Unter¬
ok, Oberkiefer.
ol, Oberlippe.
kiefer.
ul, Unterlippe.
—
—
—
—
—
—
—
Abdomen von CuliAbb 17.
ciden-Weibchen. —a, Anopheles.
d,
b, Aedes.
c, Culex.
co,
Flügel einer Culicide.
Costalader.
sc, Subcostalader.
Radialadern.
m
r 1—5,
ca.
I—2,
I—1, Medialadern.
Cubitaladern.
an, Analader.
ca.F, CubitoaF, Analfalte.
Analfalte.
—
—
—
—
—
—
—
—
—
—
—
—
C. pipiens. Larvendarm; IV. Larven¬
2, Pharynx.
stadium, Orig.
I, Mundöffnung.
4, Ausstülpungen des 8-teiI.
3, Oesophagus.
Mitteldarms.
6, Fintritt der 5
5, Magen.
Abb. 18.
—
—
—
—
—
—
—
Malpighi'schen Gefässe in den Enddarm (Pylorus).
8, Rektalblase mit Rektal¬
7, Malpighigefässe.
—
—
drüsen.
—
9, After.
BIOLOGIE UND VERBREITUNG EINIGER STECH MÜCKEN ARTEN
37
bürsten der Larve ermöglichen die Aufnahme fester Bestandteile
während die Imagines mit ihren stechenden Mundwerkzeugen nur
befähigt sind, flüssige Nahrung
zu
saugen
(Blut, Fruchtsäfte).
Larven (Abb. 18, Darm, und Abb. 8, Mundwerkzeuge).
Es können am Verdauungskanal bei allen Arten drei Haupt¬
abschnitte unterschieden werden, nämlich Vorder-, Mittel- und
Enddarm. Der Vorderdarm besteht zur Hauptsache aus Pharynx
und Oesophagus, der Mitteldarm aus den acht Blindsäcken und
dem Magen, der Hinterdarm aus Pylorus und Enddarmblase. Beim
Pylorus treten die fünf Malpighi'schen Gefässe in den Darm ein
(Abb. 18).
Die Mitteldarm-Ausstülpungen (Drüsendivertikel) produzieren
Verdauungsfermente. Beim lebenden Tier scheinen sie ockerfarbig
bis dunkelbraun durch den Körper hindurch und zeigen rhythmische
Bewegungen. Der Vorderdarm ist, ähnlich wie bei den ausgewachse¬
nen Tieren, in den Mitteldarm eingestülpt (Valvula cardiaca. Abb.
19, a). An dieser Übergangsstelle befindet sich der sogenannte Imaginalring (ektodermal). Ein einfaches Zylinderepithel am Anfang des
Mitteldarmes bildet die hyaline, aus Proteinstoffen bestehende,
peritrophische Membran, die den Verdauungssekreten und den
Abbauprodukten Durchtritt gewährt und ausserdem geeignet ist,
das Darmepithel vor Verletzungen zu schützen. Sie wird in dem
Verhältnis erneuert, wie Verluste nach Übertritt in den Hinterdarm
eintreten.
Die Drüsendivertikel (Coeca) sind einfache, sackförmige Aus¬
buchtungen des Darms, welche ballenförmige Sekrete in ihr Lumen
abgeben. Bei den Dipteren ist die Sekretion in Form kleiner, mit
feiner Haut umhüllten Blasen die Regel (merocrine Sekretion).
An der Grenze des Mittel-und Hinterdarmes befindet sich der
Pylorus, wo die Malpighi'schen Gefässe in den Enddarm einmün¬
den. Vor dem After erweitert sich der Enddarm zur Rektalblase,
auf welcher Rektalpapillen vorhanden sind. Die Malpighigefässe sind
Hinterdarmdivertikel. Es fällt ihnen
anatomisch betrachtet
durch ihre Sonderstellung die Funktion als Exkretionsorgan zu.
Bei den Culiciden und Psychodiden sind fünf Malpighigefässe,
während bei den meisten Dipteren pur deren vier vorkommen. Das
Epithel, das das Lumen der Röhren umgibt, ist einsichtig ; seine
Zellen von bedeutender Grösse bilden ein regelmässiges Mosaik
mehr oder
und enthalten
je nach physiologischem Zustand
weniger Ausscheidungsstoffe (Abb. 19, B). Die Kerne sind rundlich
und das Plasma enthält die Ausscheidungsstoffe in Form von Körn¬
chen, die sich im zentralen Teil der Zelle häufen. Aus den bisherigen
eigenen Beobachtungen zu schliessen, ist bei unbelichteten oder
schlecht belichteten Larven die Körneranreicherung besonders gross.
—
—
—
—
38
WILLI
I
«„„„„
BÜTTIKER
B.
,."•
1
*-Vi"l'ffl
A. Culex pipiens; IV. Larvenstadium. Ubergangszone von Vorder- zu
Abb. 19.
Mitteldarm [Valvula cardiaca]. Orig.
BS, Ballenförmige Sekrete.
I, ImaginaiV, Valvula cardiaca: 1, Ring¬
MD, Mitteldarm.
Oe, Oesophagus.
ring.
muskulatur des Oesophagus.
3, Mitteldarm-Ausstülpungen.
2, Cuticula.
5, Nahrungspartikel.
4, Peritonialgewebe mit Urat-Ausscheidungen.
6, Peri7, Mitteldarmgewebe.
trophische Membran.
B. A. maculipennis var. messeae; IV. Larve. Endabschnitt eines Tubus der Malpighi'schen Gefässe ; Zellen mit Uratkristallen.
—
—
—
—
—
—
—
—
—
—
—
—
Bei den Puppen scheinen die Exkretstofie nicht mehr in so grosser
Menge vorhanden zu sein. Neben den Ca-, Na-, NH3- Urat kommen
in zweiter Linie Leucin und Calciumoxalat, Na- und Ca-Carbonat
in Betracht (WlGGLESWORTH, 97).
Imagines (Abb. 20).
Das Darmsystem kann ebenfalls in drei Abschnitte gegliedert
werden : Vorder-, Mittel- und Hinterdarm. Die ektodermalen
Vorder- und Hinterdarm
sind chitinisiert. Der
Abschnitte
Mitteldarm weist eine epitheliale Struktur auf, zur Erzeugung der
Verdauungsfermente und zur Nahrungsresorption. Der Vorderdarm
besteht aus Pharynx und Oesophagus, mit Ausstülpungen.
—
—
VERBREITUNG EINIGER
BIOLOGIE UND
C. pipiens.
Abb. 20.
Darm. ca. 10-fach. Orig.
—
Imaginaler
I, Oeso¬
2, Paariger Saugmagen.
3, Unpaariger Saugmagen, mit
Luftblasen.
4, Magen [Fundus].
5, Eintritt der 5 Malpighi'schen
Gefässe.
7,
6, Enddarm.
phagus.
—
—
—
—
—
—
—
Enddarmblase
Malpighi-gefässe.
mit Rektalpapillen.
—
C. pipiens, Weibchen.
Abb. 21.
Frontalschnitt durch den Kopf auf
der Höhe des Pharynx. Vergr. 90fach. Orig.
1, Vordere Pharynx2, Pharynx, Vorderab¬
pumpe.
schnitt.
3, Radialer Zugmuskel.
5, Unter¬
4, Chitinspange.
—
—
—
—
—
—
6, Oesophagus.
schlundganglion.
C, Cerebralganglion.
A, Augen.
—
—
—
—
T, Thorax.
—
S.
Speicheldrüsen.
Abb. 22.
—
Culex pipiens, Weibchen.
Querschnitt durch die vordere Pharynxpumpe. Orig.— 1, Längsmuskel.
2, Zugmuskel.
3, Zugmuskel.
4, Chitingerüst.
5, Ansatzstelle
—
—
—
—
der Muskulatur.
6, Variabler
Hohlraum des Pharynx.
7, Zug¬
muskel [Dilatator].
—
—
STECHMÜCKENARTEN
39
40
WILLI
Den grössten
Teil des
BUTTIKER
Pharynx nimmt die hintere Pharynx-
Pumpe (Abb. 21) ein, die aus drei konkaven Chitinplatten besteht,
welche an ihren Ecken durch biegsame Häute verbunden sind.
Kräftige Muskeln, die an den Platten befestigt sind, ermöglichen ein
sehr schnelles Erweitern und Schliessen des Hohlraumes. Beim
Erweitern fliesst infolge Unterdrucks die Flüssigkeit in den Oeso¬
phagus. Die vordere Pharynx-Pumpe (Abb. 22), welche kleiner und
schwächer ist, schliesst den Pharynx gegen die Mundwerkzeuge ab.
Im paarigen und unpaarigen Saugmagen wird die Nahrung gespei¬
chert. Sie gelangt nach und nach durch den Proventriculus in den
Mitteldarm. Der paarige Saugmagen liegt im Thorax dorsal, während
der viel voluminösere, unpaarige bis weit ins Abdomen (5. Abdo¬
minalsegment) reicht. Normalerweise werden Fruchtsäfte in die
Divertikel geleitet, während Blut direkt dem Mitteldarm zugeführt
wird.
In der Gegend des Oberschlundganglions ist der Pharynx nur
ein dünner Kanal. Die hintere Pharynx-Pumpe ist beim Männchen
viel schwächer entwickelt als beim Weibchen. Der Hypopharynx
steht mit den dreilappigen Speicheldrüsen in Verbindung, die bei
Männchen kleiner sind als bei Weibchen. Die Ausfuhrgänge beider
Drüsenhälften nähern sich im Hals und vereinigen sich im Kopfe
zum gemeinsamen Speichelgang des Hypopharynx.
Der Mitteldarm beginnt in einem langen, dünnen Schlauch
(Magenschlauch oder Cardia), der in den kurzen, dicken Magen¬
sack (Fundus) übergeht. Die peritrophische Membran wird im
Ubergangsgebiet von Vorder-zu Mitteldarm gebildet, die in Form
einer glasartig dursichtigen Membran die eingeschlossene Speise¬
masse nach hinten schiebt.
Unmittelbar beim Übergang vom Mittel-zum Hinterdarm münden
die fünf Malpighigefässe ein. Nebst der kräftigen Muskulatur hat
der Enddarm auch eine chitinige Auskleidung. Der Mastdarm, als
hinterer Enddarmabschnitt, trägt sechs kleine Rektaldrüsen und geht
durch einen faltenreichen Gang in den After über.
Die Malpighi'schen Gefässe sind gleichartige, lange, unver¬
zweigte Schläuche mit engem Sammelkanal.
ABIOTISCHE
IV.
AUSSENFAKTOREN
1. Einfluss der
A.
Die
Temperatur
und
Ausbreitung der Wärme
Temperatur
Entwicklungsdauer
von
einer Stelle des Raumes
zur
andern kann durch Wärmeleitung, Wärmemitführung (Konvektion)
und Wärmestrahlung geschehen: Die Wärmeübertragung durch Lei¬
tung und Mitführung ist auf Erwärmung der Zwischenschicht ange¬
wiesen, nicht aber die Übertragung durch Strahlung.
In diesem Kapitel werden nur die Beziehungen zwischen der
übertragenen Wärme und der abhängigen Entwicklungszeit aller
Entwicklungsstadien untersucht. Die Wärmestrahlung wird im Kapitel
« Einfluss der
Strahlung » behandelt.
Für die bei der Entwicklung der Insekten sich abspielenden
Lebensprozesse sind eine Reihe von Aussenfaktoren, wie Luftfeuch¬
tigkeit, Licht und Wärme, von grosser Wichtigkeit. Aber kein anderer
Faktor hat in der Ökologie der Tiere eine solche Bedeutung wie
die Temperatur "erlangt. Der tägliche und jährliche Wechsel sowie
die regionale Verschiedenheit haben mannigfaltige Auswirkungen zur
Folge. Innerhalb eines bestimmten Temperaturbereiches ist Leben
und Entwicklung möglich, ausserhalb davon tritt der Tod ein.
Die Grenzen des vitalen Bereiches sind weitgehend von Art zu
Art spezifisch und können bei den verschiedenen Entwicklungs¬
stadien der gleichen Art schwanken. Desgleichen sind oft auch
Unterschiede im Verhalten der Männchen und Weibchen der Tem¬
gegenüber festzustellen.
Eine Temperaturerhöhung wirkt sich bis einem gewissen Grade
im Sinne einer Entwicklungs-Beschleunigung aus ; oberhalb der kür¬
zesten Entwicklungszeit tritt wieder eine Wachstumshemmung ein,
die bis zur Entwicklungsgrenze zunimmt und oberhalb davon den
Wärmetod zur Folge hat. Desgleichen findet unterhalb eines gewissen
als Entwicklungsnullpunkt (k, « developmental zero »)
Punktes
bezeichnet
keine Entwicklung mehr statt. Die Temperatur ist
also in erster Linie ausschlaggebend für die Entwicklungsdauer, und
ihre Beziehung zur Entwicklungszeit kann in der Formel
peratur
—
—
42
WILLI
c
=
t
(T—k)
BUTTIKER
c
=
t
=
T
k
=
=
const.
Tage
Temperatur °C
Entwicklungsnullpunkt
{Hyperbelkurve nach BoDENHElMER) ausgedrückt werden.
Mit diesem mathematischen Ausdruck ist
gegebenen Punkten die
es
möglich,
von
zwei
Entwicklungskurve zu bestimmen.
bemerken, dass sich beobachtete und
gesamte
Einschränkend ist aber zu
errechnete Werte nur in einem beschränkten Bereich decken. Ver¬
schiedene Autoren weisen auf die Entwicklungsstörungen im Gebiete
des Entwicklungsnullpunktes und im Bereich der hohen Tempe¬
raturen hin, Parker (65) an Eiern von Melanoplus m. mexicanus, VAN
HEERDT (36) an Forficula aurkularia. Martini (48) unterzieht in
seiner Arbeit die Hyperbelformel einer kritischen mathematischen
Betrachtung.
Der Entwicklungsnullpunkt eines Insektes gibt über seine ther¬
mischen Ansprüche an das Milieu guten Aufschluss und kann des¬
halb für die ökologische Beurteilung der Insekten von grosser Bedeu¬
tung sein.
Über maximale und minimale Temperaturen, welche von Insek¬
ohne Schaden überdauert werden können, gibt Chapman(IO)
einige Angaben (Buprestiden-Larven unter Holzrinde bis zu 52° C,
Callosamia promethea [Saturniidae, Lep.] von —35° C).
Eine gute Zusammenfassung der bis jetzt durchgeführten Expe¬
rimente über den Einfluss der Temperatur auf die Entwicklungszeit
bei einer Anzahl von Insekten gibt VAN Heerdt (36), so dass auf
diese Literaturübersicht verwiesen werden kann.
Untersuchungen über die Entwicklungsdauer wurden von ver¬
schiedenen Autoren beschrieben. Ihre Resultate sind jedoch recht
unterschiedlich, was zum Teil auf die ungleichen Zuchtbedingungen
zurückgeführt werden kann. Ältere Larven können während einer
ganzen Anzahl von Tagen bei nicht zusagender Nahrung auskom¬
men, stellen aber in dieser Zeit das Wachstum ein. Die Beobachtungs¬
resultate können deshalb zu ganz fehlerhaften Schlüssen führen.
Angaben über die Entwicklungszeit mitteleuropäischer Culiciden veröffentlichen Eckstein (14) und Galli-Valerio (22), wobei
der Temperaturfaktor meist ausser Betracht gelassen wird. Genauere
Angaben macht Martini (52), hauptsächlich über den Temperatureinfluss bei A. maculipennis und C. pipiens. In seiner Publikation über
die Bionomie einiger Stechmücken (47) werden A. bifurcatus u. a.
einer näheren Untersuchung unterworfen. CoRADETTI (12) befasst
sich zur Hauptsache mit A. maculipennis-R&ssen, dessen Unter¬
ten
suchungen verschiedene Entwicklungszeiten ergeben.
Neu in der vorliegenden Arbeit sind die genauen Untersuchun¬
über die Entwicklungszeit bei verschiedener Temperatur,
gen
BIOLOGIE
UND
VERBREITUNG
EINIGER
STECHMÜCKENARTEN
43
besonders bei Aedes
pennis
var.
vexans, A. geniculatus und Anopheles maculiDie Experimente an Culex pipiens bestätigen
früheren Resultate oder ergaben entsprechende
messeae.
Teil die
Korrekturen.
Ebenfalls neu sind alle detaillierten Untersuchungen über die
vier Larvenstadien, sowie des Puppenstadiums. Bei Culex pipiens
und Anopheles maculipennis var. messeae kamen dazu noch die Eiér,
deren Entwicklungszeiten in Abhängigkeit der Temperatur ermittelt
werden konnten.
zum
Methode
Für die Entwicklungsversuche stand ein Reihenthermostat
(WlESMANN, Jenny, 96) zur Verfügung, bei dem die beiden Tempe¬
raturextreme variiert werden konnten ; die Wärme zwischen 35 und
50° C, die Kälte im Bereich zwischen —20 und +I0°C. Gesamt¬
haft sind sieben Einzelabteile vorhanden, die ein stufenweises Anstei¬
gen der Temperatur ergeben. Die Entwicklungszeiten der unter¬
suchten Culiciden wurden in den Stufen von +8°, 16°, 22°, 27°,
32° und 34° C beobachtet. Die Genauigkeit des Thermostaten war
im Durchschnitt zufriedenstellend ; bei starker Sonneneinstrahlung
ergaben sich aber Temperaturerhöhungen bis zu ±0,5° C, beson¬
ders in den mittleren Stufen (Tabelle Nr. 7).
Im Verlaufe der Untersuchungen stellte es sich heraus, dass für
die Entwicklung der Anopheles maculipennis-Rassen Strahlungs¬
energie auf die Larven einwirken muss. Es wurden deshalb in sämt¬
liche Abteile des Thermostaten elektrische Birnen von 25 Watt ein¬
gebaut und täglich während 12 Stunden in Betrieb genommen
(Abschnitt « Einfluss der Strahlung »). Die Larvensterblichkeit konnte
auf diese Weise von ca. 95 % auf 20 % und weniger reduziert wer¬
den (Abb. 38).
Durch die unterschiedlich schnelle Entwicklung von Männchen
und Weibchen ergibt sich in allen Stadien eine Streuung, die bis
zum Entwicklungsende je nach Temperatur
1—5 Tage ausmachen
kann. Die Beobachtungsresultate stellen also Mittelwerte dar; diese
dienen zur Berechnung der Bodenheimer'schen Kurve.
Resultate
Culex pipiens.
Während ECKSTEIN (14) über C. pipiens nur wenige Daten gibt
(Entwicklung des Eies 5 Tage, von Ei bis Imago 3 Wochen), stellt
Martini (52) eine Tabelle auf, welche Daten verschiedener Her¬
kunft berücksichtigt (Tabelle 8) :
NlESCHULZ und Bos (59) konnten einmal bei 25° C Versuchs¬
temperatur schon nach 12
Tagen, in einem andern Fall
aber
erst
44
WILLI
BÜTTIKER
nach 18 Tagen, bei gleicher Temperatur Imagines erhalten. Diesel¬
ben Autoren geben für 29° C eine Entwicklungszeit von 16—17 Tagen
an, mit der Bemerkung, dass von einer Entwicklungshemmung, wie
sie RoUBAUD beobachtet hatte, keine Anzeichen vorhanden waren.
Martini (47) führt in seiner Arbeit über die Bionomie einiger euro¬
päischer Stechmücken, speziell über C. pipiens, keine Daten an.
MARSHALL (46) gibt für die Eientwicklung die Zeit von 2 Tagen an.
Meine beobachteten und die daraus errechneten Werte für die
Entwicklungszeit von C. pipiens sind in Tabelle Nr. 9 und Abb. 23
zusammengestellt.
Von einer Entwicklungshemmung der Larven bei 25—28° C,
wie sie RoUBAUD (74) beschreibt, konnte in den durchgeführten
Versuchen nichts festgestellt werden. Vermutlich handelt es sich bei
den beschriebenen Versuchen um Tiere von Hunger- oder Intoxi¬
kationszuchten, denn er bemerkt, dass nach einer schnellen Ent¬
wicklung eine Hemmung in der Entwicklung eintritt. Nur ein gerin¬
ger Teil der Larven verpuppte sich nach etwa 15 Tagen, die andern
Exemplare blieben im IV. Stadium und starben nach etwa 2 Monaten
ab. NlESCHULZ und Bos (59) experimentierten wahrscheinlich mit
Normalzuchten und konnten deshalb keine Entwicklungshemmung
feststellen. Ulmann (89) beschreibt eingehend seine Untersuchun¬
gen an C. molestus (frühere Bezeichnung von C pipiens autogenicus)
in Hunger- und Intoxikationszuchten und zeigt, dass in beiden
Fällen eine sehr grosse Larvensterblichkeit eintritt. Ausser einer
Verschiebung des Geschlechtsverhältnisses ergibt sich auch eine
Verlängerung der Entwicklungszeit.
Eine Entwicklungshemmung, durch zu hohe Temperatur verur¬
sacht, ergab sich bei Puppen von C. pipiens im Bereich von 35—36° C.
Wie bei den durchgeführten Versuchen an den Larvenstadien,
konnte eine hohe Sterblichkeit beobachtet werden, so dass die oberste
unschädliche Entwicklungstemperatur ca. 34—35° C betragen mag.
Die beobachteten durchschnittlichen Entwicklungszeiten sämt¬
licher Entwicklungsstadien (Larven I bis IV und Puppen, exklusive
Eier) betragen bei Normalzuchten und einer Wassertemperatur von
16°
22°
27°
32°
Aedes
15,5 bis 17,0 Tage
»
10,0 » 11,0
»
7,5 »
8,0
»
6,7
C
C
C
C
vexans.
über die Entwicklungszeit von Aedes vexans konnte nur eine
Angabe gefunden werden, nämlich bei ECKSTEIN (14) : « Die Zeit des
Wachstums der vexans-Larven hängt ausserordentlich ab von der
Temperatur des Wassers und der vorhandenen Nahrung. Ist die
Wassertemperatur hoch,
so
vermögen sich die Larven sehr schnell,
BIOLOGIE UND VERBREITUNG
EINIGER
STECHMÜCKENARTEN
45
I. Larve.
•
o
Beobachtung
Rechnung
Tage
15
30
•
o
Beobachtung
Rechnunn
20
10
<e
Tagt
Abb. 23.
—
10
15
10
15
Culex pipiens. Die Entwicklung der einzelnen Stadien, in Abhängigkeit
von der Temperatur.
0°
tbrits
tbrits
+
2.5
0.5
masgnal
5°
8,0
rhes
52,0
0.1-
grnuutkanraewphmceST
der
12
60
0,5
10°
egaT
egaT
°C
+
°C
+
III
15°
45
5-6
0.5
°C
+
+
IV
°C
5.51
°C
4
31
0,5
°81-71
0,22
II
3
26
22»
3
10
25»
25°
8
7
0.5
Nr,
+
0.5
+
+
0.5
°C
Nr.
VII
°C
VI
°C
0,72
I
0.23
netatsomrehtnehieR tiekgiuaneG
des
ntehgcaerlthrcesv
rebü
ellebaT
0.43
ellebaT
IV
44
44
5,73
5,23
0,22
III
4
6
5
12
—
ca.
5,65
5
5
7,3
0,71-5,51
71-61
31-11
5,73
II
4,0
3.0
2,5
2.5
3,5
3,5
3,0
2,0
3,5
4,0
5,61
0,11
5,21
7,41
0,11
41-21
I
0.11-0,01
3,5
1.5
2.2
1.8
1,3
2.2
2,6
2,5
4,01
5,3-0,3
4-5,3
5,2-2
5,21
5
0,8-5,7
1.0
1,5
2,0
0,2-5,1
1.5
5,1>
5.2-0,2
0.2-5.1
5.2-0.2
22°
27°
tethcaboeb tenhcerre tethcaboeb tenhcerre tethcaboeb tenhcerre .hcaboeb
16°
1,7
6,8
1,5
6.7
7.7
32°
—
1,5
1.5
1,2
1.2
1,5
1,5
1.5
0,8
1.5
1.6
1.7
1,5
1,5
1.5
2.0
1,2
1,0
1.5
1.0
1.5
1.7
.hcerre
8°
.boeb
k
.hcerTC
etatluseR-sgnulkciwtnE
C
.boeb
C.
1,5
1,6
1,5
53.6
1,5
1,2
57,0
hcerre
.°43
sneipip
von
aus
Nr.
9
hc,illltehtcsirsarDe
.thcaboeB
hparg
eretieW
ne g n u k r e m e B
ellebaT
eppuP
evraL
evraL
evraL
evraL
reiE
48
WILLI
BUTTIKER
schon in
etwa 6-8 Tagen soweit zu entwickeln, dass sie sich verpuppen
können. Das Puppenstadium dauert dann etwa 1 bis 2 Tage, sodass
die ganze Entwicklung von vexans unter Umständen etwa nur 8—10
Tage in Anspruch zu nehmen braucht. »
Die Entwicklungsdauer von Aedes vexans -ist, verglichen mit
andern Culiciden, tatsächlich sehr kurz. Als Bewohner temporärer
Wasseransammlungen, die besonders in den Monaten Juni, Juli und
August hohe Temperaturen aufweisen, ist diese Insektenart auf eine
schnelle Entwicklung zur geschlechtsreifen Imago angewiesen.
Die beobachteten und daraus errechneten Entwicklungszeiten für
A. vexans sind aus Tabelle Nr. 11 und Abb. 24 ersichtlich. Die beo¬
bachteten durchschnittlichen Entwicklungszeiten sämtlicher Ent¬
wicklungsstadien (Larven I bis IV, Puppen, exkl. Eier) bei einer
16° C betrugen 16,5 bis 18,5 Tage
Wassertemperatur von
22° C
10,5
»
27° C
7,5 bis 8,0
»
30° C
6,3 »
7,0
»
34° C
5,5 »
6,0
Entwicklungsstörungen bei langer Einwirkung treten nach Labor¬
versuchen erst zwischen 36° und 37° C auf, wobei auch eine Hemmung
der Entwicklung zu beobachten ist. Parallel dazu geht ebenfalls
eine höhere Larvensterblichkeit (Tabelle Nr. 11).
Die minimale Entwicklungstemperatur beträgt 7,0 bis 7,6° C,
die maximale dagegen für kurze Zeit über 41° C, für längere Zeit
nicht über 36° C. Die I. Larven und Puppen konnten sich entwickeln,
starben aber unmittelbar vor der Verpuppung ab. Die Temperatur¬
stufe von 36° C stellt also den Grenzwert der Entwicklungstempe¬
dar. Freilandmessungen ergaben folgendes Zahlenmaterial
ratur
(Tab. 10).
Vergleicht
diese Entwicklungsresultate mit den ökologischen
ist eine enge Bindung zwischen Mücke und Biotop
unverkennbar. Die ausserordentlich rasche Entwicklungszeit dieser
Culicide bietet Gewähr, dass in diesen temporären Wasseransamm¬
lungen sämtliche Stadien innerhalb nützlicher Frist durchlaufen wer¬
den können. Des weitern können hohe Temperaturen, wie sie in
diesen seichten, moorigen oder steinigen Gewässertypen vorkommen,
ohne Schaden überdauert werden. Marshall (46) vermutet, dass
es sich bei A. vexans um eine ursprünglich südliche (vielleicht zentral¬
afrikanische) Culicidenart handle, führt aber keine weiteren Gründe
zu dieser Annahme an. Die vorliegenden Temperaturversuche sowie
die Experimente über die Aktivität verstärken doch Marshall's
Vermutung, wenn es auch an der genügenden Zahl von ökologischen
und physiologischen Untersuchungen fehlt. Die Aktivitätskurve und
die einzelnen Aktivitätsstufen sprechen aber gegen eine solche
Annahme, da die Kältestarre zum Beispiel nur wenig über derjenigen
von Anopheles maculipennis var. messeae liegt und nicht subtropische
man
Verhältnissen,
so
BIOLOGIE
UND
VERBREITUNG
EINIGER
STECHMÜCKENARTEN
49
II, Larve.
•
Beobachhma
o
Rechnung
20-
Tage
5
10
•
Beobachtung
o
Rechnung
5
S
4
Tage
10
1y
10
Aedes vexons. Die Entwicklung
Abb. 24.
der einzelnen Stadien, in Abhängigkeit von
der Temperatur.
—
,
Tage
50
WILLI
BÖTTIKER
Temperaturverlauf am Brutplatz von Aedes vexans.
Ort: Rapperswil; Datum: 20.7.1947.
Tabelle Nr. 10
Zeit
H20
Luft/Sonne
Bemerkungen
0500
0600
0800
1000
1200
1400
1500
1600
1800
2000
2100
2200
16,2
16,3
17,1
21,3
27,0
28,1
29,1
28,8
26,3
22,5
21,6
20,5
16,1
16,2
16,8
24,3
31,5
36,8
Wetterlage : nach nächt¬
lichem Gewitter Aufhel¬
lung und sonniges,
heisses Wetter.
36,2
32,1
26,2
25.1
24,2
Monges, seichtes Litoralgewässer des Zürichsees, mit Moos und
Schilf bewachsen. Während den Überschwemmungen bevorzugter Brut¬
ort von Aedes vexans, die hier in ungeheuren Mengen ihre Entwicklung
durchlaufen.
oder tropische Merkmale aufweist wie zum Beispiel Aedes sollicitons
(vergl. Abschnitt « Temperatur und Aktivität ») mit einem Grenzwert
der Kältestarre
von
+17° C.
Aedes geniculates.
Über die Entwicklungsdauer von Aedes-geniculatus bei verschie¬
denen Temperaturstufen konnten keine Angaben früherer Autoren
gefunden werden.
Durch Einzelzuchten, wie sie mit C. pipiens, Aedes vexans und
Anopheles maculipennis
sich
folgende Resultate
var.
messeae
durchgeführt wurden, ergaben
:
Siehe Tabelle Nr. 13 und Abb. 25.
Sehr auffällig ist die hohe Temperaturempfindlichkeit im Bereich
über 32° C. Kurze Zeit kann allerdings diese Wärmestufe ohne Scha¬
den ertragen werden, wie dies aus den Aktivitätsversuchen (vergl.
BIOLOGIE
UND
VERBREITUNG
STECHMÜCKENARTEN
EINIGER
°C
/. Larve.
//. Larve.
30-
20-
10
k-
0
5
Tage
10
'C
30-
IV. Larve.
20-
10-
e
Tay
10
Abb. 25.
Aedes genicalatus. Die Entwick¬
lung der einzelnen Stadien, in Abhängigkeit
von der Temperatur.
—
51
1
IV
Ei
vom
Mai
0,52
5.91
6.33
5,04
9.92
III
7,1
7,6
7,5
7,3
bis
rehcilt inhcshcruD
lirpA
ca.
32
k:
40
33
84
49
36
242
für
des
treW
latoT
tethcaboeb
7,0
.rre
II
4,5
tethcaboeb
eppuP
evraL
evraL
evraL
evraL
muidatS
I
.3,7
nedneglofhcan
5,2-2
5,4-4
5,3-3
5,3-3
5,81-5,61
serhaJ
in
1.7
3,3
3
2
2,5
1.3
4,5
3.3
4.0
2,3
4,71
en.
22°
.boeb
Eix
5,01
.esuapaiD
2,7
2,0
2,3
1.3
1,7
0,01
.rre
16°
7,5
1,3
1,5
1,7
1,3
1.3
2,0
1,5
1,0
1,0
1.7
2,0
27°
tethcaboeb
8°
0,8-5,7
2-5,1
.rre
k
0,7-3,6
2,1-1
0,1<
2-5.1
30°
tethcaboeb
)k—T(
C
6,5
1,1
ca.
1
ca.
1
3/4
1,5
1,3
1,5
1.5
1,7
0,9
*rif
.rre
.tsnok
=
6-5,5
34°
tethcaboeb
ebagnA(
C= t
5.5
0,9
1,3
0,7
1.1
1,5
.rre
von
ca.
—
)t(0.1>
)f(2
Nr.
36°
tethcaboeb
in
—
0,8
~
1.4
11
.rre
ellebaT
etatluseR-sgnulkciwtnE
)negaT
snaxev sedeA
BIOLOGIE
UND
Temperaturverlauf
VERBREITUNG
EINIGER
STECHMÜCKENARTEN
53
in einem
Brutplatz von Aedes geniculates
(Fichten-, Lärchen-, Buchenmischwald)
Zürich-Rehalp
Zeit
31:5-/1.6.1947
H20
Tabelle Nr. 12
Luft
Luft
H20
(Sonne)
20,8
19,7
23,3
20,7
Bemerkungen
315
2000
2100
19,1
18,3
y2 Std. später 17,4°
1
Über,
1
regen.
Nacht
Gewitter-
1.6
0600
0700
0800
0900
1000
16,0
16,3
163
17,0
17,6
1100
1200
1300
1400
1500
1600
1700
1800
1900
2000
2100
20,0
19,6
20,2
24,8
21,7
20,9
20,6
20,3
19,9
19,4
19,0
.
.
15,8
16,5
17,0
17,5
18,6
16,2
17,0
17,8
20,1
25,4
20,4
20,4
21,0
22,7
21,8
22,8
22,0
22,2
21,5
21,0
20,5
27,0
30,4
33,8
35,7
35,6
35,0
34,0
32,5
29,5
25,2
24,9
Regen.
Beginn der direkten
Sonneneinstrahlung.
i
i
'
Ende der direkten
Sonneneinstrahlung.
Tagesschwankung 9° C.
Messungen
:
H2O
Luft H2O
:
Luft/Sonne
Unmittelbar unter der Wasseroberfläche:
2 cm über der Wasseroberfläche im Schatten.
Direkte Sonnenbestrahlung des Thermometers.
:
ca.
:
Abschnitt « Temperatur und Aktivität ») hervorgeht. So starben
jeweils die Larven I. Stad. und Puppen bei 34° C ab, ohne sich noch
gehäutet zu haben. Im II. und III. Larvenstadium konnte eine Ent¬
wicklungszeit von 2,5 bezw. 7 Tagen beobachtet werden. Futter
war stets in genügender Menge vorhanden, so dass diese Erscheinung
wohl kaum dem Nahrungsmangel ; zugeschrieben werden kann.
IV
III
)k—T(
muidatS
evraL
evraL
evraL
evraL
eppuP
ttinhcshcruD
II
6.75
5,27
5,94
50
0,83
ca.
7,3
7.8
7.8
7.2
7
8,0
ca.
—
latoT
tethcaboeb
.tsnoc
I
—
6
y2
5,23-0.13
7-2y
0,9<
5.5
5.5
0.5-5.4
tethcaboeb
MT.
4.0
5.0
5,0
8.5
4.0
3.3
22°
3.3
2.7
3.5
5.5
2,5
5.5
7.0
52.13
57.4
.rre
16°
91-0.81
)5.4(
5.3-3
tethcaboeb
8°
3.81
.rre
k
2.5
2.0
2.5
3.0
0.41-5.31
0,4-5.3
27°
tethcaboeb
etatluseR-shcusreV
C
—
2.5
2.4
3.0
3.0
3.7
32o
2.0
1.6
2,0
2.0
2.5
t
2.5
3.0
7.31
.rre
von
.hcaboeb
der
0,11
.rre
gnulkciwtnE
=
n.
—
Tg.
tot
2
)!(0,4
n.
2
9.9
2,2
1.7
1.5
1.8
2.7
Nr.
34°
Tg
tot
0(5.3
tethcaboeb
sedeA
C= t
13
.rre
ellebaT
setalucineg
BIOLOGIE UND VERBREITUNG
EINIGER
55
STECHMÜCKENARTEN
Nach dem Temperaturverlauf eines Brutortes von A. geniculates
(Tabelle Nr. 12) zu schliessen, ist nicht anzunehmen, dass seine
Wassertemperatur über 28° C steigt.
Die beobachteten mittleren Entwicklungszeiten der Larvenstadien
I bis IV und Puppen betrugen bei einer Wassertemperatur von
31.0 bis 32,5 Tage
»
19,0
18,0
»
13,5 » 14,0
16° C:
22° C:
27° C:
»
Anopheles maculipennis.
ECKSTEIN (14) gibt für die Ei-Entwicklung ohne besondere Tem¬
peraturangabe 7 Tage, für das Larven- und Puppenstadium zusam¬
men 4 Wochen an, während Martini (47) für das Ei 3 Tage und
« manchmal wesentlich länger »,
für die Larven gut 2—3 Wochen
und für die Puppen ca. 3 Tage anführt. Im später erschienenen
« Lehrbuch der medizinischen Entomologie » (52) sind durch Martini
folgende Entwicklungszeiten angegeben (z. T. nach CoRADETTl).
Tabelle Nr. 14
°c
24-27
20-22
16-19
CoRADETTl
Larve
II
Larve
I
EÎ
2
11/4
2
21/4
fast 3
2
4
6
5
(12) veröffentlicht
Larve
III
Larve
IV
21/2
21/2
41/2
41/4
4%
6 1/2
Puppe
gesamt
2%
141/2
3
18
41/2
30%
in seiner Arbeit Resultate über die
Entwicklungsdauer der verschiedenen Rassen
x
aus der folgende Angaben entnommen sind
:
von
A. maculipennis,
Versuchstemperatur 23° C
September 1933
Tabelle Nr. 15
Durchschn. Entwicklungsdauer
von Ei bis Puppe
Rasse
labranchiae
atroparüus
messeae
.
.
.
.
.
.
....
19 Tage
19 Tage
20 Tage
22 Tage
22 Tage
1
Ausserdem enthält diese_ Arbeit Angaben über Versuche mit_ verschiedenen
gefärbten Zuchtgefässen, wobei die durchschn. Entwicklungszeit zwischen «weiss»
und «dunkel»
um
1
Tag
variiert.
56
WILLI
BUTTIKER
Die durchnittlichen Unterschiede in der Entwicklungszeit der
einzelnen maculipennis-Rassen betragen demnach 0 bis 3 Tage. Es
wäre wünschenswert gewesen, ähnliche Entwicklungsversuche mit
schweizerischem Material durchzuführen ; wegen zu grossen Tem¬
peraturdifferenzen im Reihenthermostat musste aber davon abgesehen
werden. Die Streuung zwischen minimaler und maximaler Ent¬
wicklungszeit ist bei 22° C mit ca. 2—3 Tagen so gross, dass nur
sehr genaue Mittelwerte verglichen werden können. Der Einfluss
der Strahlung muss ebenfalls berücksichtigt und auf alle Fälle bei
allen Versuchen konstant gehalten werden (vergl. Abschnitt « Einfluss
der Strahlung »).
Zur eingehenden Untersuchung gelangte nur Anopheles maculipennis var. messeae, wobei folgende Entwicklungsresultate erhalten
werden konnten (Tabelle Nr. 17, Abb. 26). Daraus geht deutlich
hervor, dass die beobachteten mittleren Entwicklungsstufen der
Larvenstadien I bis IV und Puppen folgende Zeitwerte betrugen :
(16° C)
22° C
27° C
34° C
(27,5 bis 29,0 Tage)
16,0
17,5
»
12,0
9,3
»
»
13,0
»
»
Die Entwicklungshemmungen, hervorgerufen durch zu hohe Tem¬
peratur, traten im Bereich von 35-37° C auf, wobei Verlängerungs¬
zeiten bis zu 12 Stunden beobachtet werden konnten.
Versuche im tieferen Temperaturbereich von 8° C ergaben deut¬
lich, dass alle Entwicklungsstadien auf Kälte sehr empfindlich sind.
Nach einiger Zeit starben die Versuchstiere ab, nachdem der grösste
Teil des gespeicherten Fettes aufgezehrt war. Bei dieser Temperatur¬
stufe wurde von den Larven kaum noch Nahrung aufgenommen. Es
wurde versucht, mit frisch gehäuteten Larven aus dem Freiland durch
Überführen in 8° C eine Weiterentwicklung zu erreichen ; alle Expe¬
rimente in dieser Beziehung blieben aber erfolglos, da sämtliche Tiere
vor der nächsten Häutung schon abstarben.
BIOLOGIE
UND
VERBREITUNG
57
EINIGER STECHMUCKENARTEN
I. Larve.
m
o
Beobachtung
Rechnung
•
o
Beobachtung
Rechnung
20
10
10
Tage
m
5
15
II. Larve.
20
10
10
5
Tage
10
10
15
m. Larve.
30-
20
0
Tage
5
Tage
10
15
'C
IV. Larve.
30-
Puppe.
30»
Beobachtung
Rechnuno
20
10
20
10
-
S
Abb. 26.
—
Tage
10
15
Tage
10
15
Anopheles maculipennis var. messeae. Die Entwicklung der einzelnen Stadien
in Abhängigkeit von der Temperatur.
ttinhcshcruD
IV
muidatS
evraL
evraL
evraL
evraL
eppuP
latoT
III
k=
7.0
7,0
7,2
7,1
7,0
5.44
2,48
0,05
0.03
4.04
5.44
II
6.8
n.
Tg.
tot
11
tot
.gT7.n
tethcaboeb
I
—
—
.rre
—
—
3,5
5
3.0
3.3
5.7
1.5
2.5
2,0
3.5
2,0
2,0
3,0
2,7
5.6
5,0
4,4
3,3
9.5
5.0
8,72
5.5-5
6.5-5
5,4-0,4
tethcaboeb
7.1
5,71-0,61
.rre
Ei
7.61
3-5,2
6-5.5
3-5,2
3-5,2
tethcaboeb
k
0,31-0,21
5,2-2
5,4-4
.rre
C
8°
16°
22°
27°
2.2
0,21
2,2
2,0
1,5
2.5
52.4
5.2-2
tethcaboeb
=
—
3,5
>1
2,0
32°
1.8
1,6
1,2
2.0
3,4
1,8
0,01
0.2<
2-5,1
.rre
.tsnok
)k—T(
sin epilucam selehponA etatluseR-sgnulkciwtnE
von
9.3
ca.
Nr.
34°
1,7
1.5
1
<2
3.0
8,1<
.rre
tethcaboeb tethcaboeb
.rav
C= t
9,3
1,7
3.1
1.1
7.9
1.5
1.7
17
.rre
ellebaT
eaessem
BIOLOGIE
UND
VERBREITUNG
EINIGER
Temperaturen eines Brutplatzes
var.
La
Sauge
17.
von
Anopheles maculipennis
messeae
Tabelle Nr. 16
Aug. 1947
Zeit
H20
Luft/Sonne
0500
0600
0800
1000
1200
1400
1500
1600
1800
2000
2100
18.2
18,2
18,5
20,6
26,2
27,9
28,1
27,8
25,2
21,4
20,5
16,3
16,4
19.1
25,4
31,2
38,4
38,6
37,1
31,0
25,8
22.3
Temperatur- und Feuchtigkeitsverlauf
Anopheles maculipennis
örtlichkeit
Datum: 17.
:
in einem Aufenthaltsort
var.
0530
0630
August 1947
Tabelle Nr. 18
92%
92%
Bemerkungen
Dämmerung. Starker Einflug
? durch die Öffnungen.
1
18,2
19.5
20.8
25.2
26.1
25,8
22,2
17,3
17.1
Innerhalb
Umgerechnet
12
X
40
X
15.
von_ 5 Minuten
auf eine Stunde,
85%
80%
72%
50%
45%
45%
65%
90%
90%
von
vor
den Ein¬
Nur noch vereinzelter
Einflug1.
Männchen
0830
0930
1030
1230
1430
1630
1830
2030
2130
La Sauge
unbenutzter militärischer Bunker,
16.2
16,3
von
messeae
Lufttemperatur Luftfeuchtigkeit
°C
% rel.
Zeit
59
STECHMÜCKENARTEN
fluglöchern.
tanzen
Abenddämmerung. Starker Aus¬
flug von 6* + ?.<? tanzen vor den
Ausflugsöffnungen.
flogen durch eine Bunkeröffnung ca. 40 Mücken ein.
während dieser der Einflug stattfindet, ergeben sich
(Einflugöffnungen)
=
7200 Mücken.
60
WILLI
Temperatur
BUTTIKER
und
Praeovipositionszeit
Die minimalen Praeovipositionszeiten von Culex pipiens und
Anopheles maculipennis var. messeae unterliegen der Wärmesummen¬
regel nach BoDENHEIMER. Die in der nachfolgenden Tabelle ange¬
führten Zahlen sind beobachtete Mindestwert (Tab. 19).
Tabelle Nr. 19
Temp. °C
Culex pipiens
5
3
18
22
25
Tage
Tage
2,5 Tage
Anopheles maculipennis
var.
messeae
3,5 Tage
2,5 Tage
2,0 Tage
r
Praeovipositionszeit ist
gleich wie die einzelnen Ent¬
wicklungsstadien der Insekten
temperaturabhängig. Dazu können
barometrische Schwankungen und andere Umgebungseinflüsse eine
Verschiebung der Eiablage um 3—10 Tage zur Folge haben (vergl.
Abschnitt 4, « Einfluss des Luftdruckes »).
Die
—
—
B. Thermopraeferendum
(Vorzugstemperatur)
Wirken richtungsgebende Reize auf das Insekt ein, so erfolgt
die Orientierung durch Ortsbewegungsreaktionen oder sogenannte
Taxien. Je nach den auslösenden Reizen kann man von Thermo¬
taxis, Phototaxis, Chemotaxis, Geotaxis usw. sprechen.
Die Phobotaxis ist eine Unterscheidungsreaktion, die auf einer
Unterschiedsempfindlichkeit beruht. Sie besteht darin, dass das
Tier nach einem Reizoptimum strebt, das als Indifferenzzone keiner¬
lei Reaktionen auslöst, vor höhern und niedern Reizquantitäten
aber zurückschreckt (Flucht- und Schreckreaktion). Am bekann¬
testen sind die bei den Insekten beobachteten Bewegungsreaktionen
dieser Art auf Grund von Wärmereizen (Thermophobotaxis). In
einem Reizgefälle (Temperaturgefälle), das man im Experiment
durch eine sogenannte Temperaturorgel herstellen kann, sammeln
sich die Versuchstiere allmählich auf Grund phobischer Reaktionen
in einer mehr oder weniger breiten Zone, dem Temperaturoptimum
(Vorzugstemperatur oder Thermopraeferendum) an. Diese Zone
kann übrigens ganz nach dem physiologischen Zustande, dem Ent¬
wicklungsstadium, auch bei ein und derselben Art beträchtliche
Unterschiede aufweisen.
BIOLOGIE UND
VERBREITUNG
EINIGER
STECHMÜCKENARTEN
61
In der Literatur wird diese Zone, welche von den Insekten durch
Bewegungen aufgesucht wird, auch als thermotaktisches Opti¬
aktive
mum,
Wahltemperatur,
Lieblingswärme
und
Behaglichkeitstem¬
peratur bezeichnet. BoDENHElMER und SCHENKIN (4) benützen in
ihren Arbeiten Vorzugstemperatur und Thermopraeferendurn, wäh¬
rend Herter (cit. lit. 36) thermotaktisches Optimum verwendet.
UvAROV (90) und WlGGLESWORTH (97) verwenden « praeferendum »
und « temperature preference ».
Seit einigen Jahren wurden eine ganze Anzahl Insekten auf das
Thermopraeferendurn geprüft. Als Erster hat Graber 1887 mit
Periplaneta orientalis Untersuchungen bezüglich der Vorzugstempe¬
ratur durchgeführt. Nach VAN HëERDT (36) haben HERTER 1924 und
Bodenheimer 1928—1931 mit Coleopteren, Weber 1924 mit Haematopinus suis, Martini 1929 mit Pediculus vestimenti, Andersen 1931
mit Sitona lineata, Krijgsma und Windred 1930 mit Lyperosia,
Kelin 1933 mit Periplaneta americana, NlESCHULZ 1933—1935 mit
verschiedenen Dipteren experimentiert.
Van Heerdt (36) stellt fest, dass bis jetzt ungefähr an 70 Insekten¬
arten das Thermopraeferendurn untersucht worden sind. UVAROV (90)
schliesst aus seinen Untersuchungen, dass das Thermopraeferen¬
durn in der Oekologie der Insekten von grosser Bedeutung ist. Die
Experimente
von
Hecht (32)
an
Weibchen
von
Culex pipiens
erge¬
ben ebenfalls die Tatsache, dass die bei der Eiablage bevorzugte Brut¬
ortstemperatur zur Besiedlung eines bestimmten Biotopes von ent¬
scheidender Bedeutung ist.
Die grosse Zahl der Untersuchungsmethoden, welche von den
verschiedenen Autoren angewandt wurden, brachte es mit sich, dass
die Resultate nicht einheitlich sind u. daher nicht ohne weiteres
verglichen werden dürfen. So ergaben alte und neue Untersuchungen
von HERTER an Liogryllus domesticus in der Vorzugstemperatur eine
Differenz von 10° C. Van Heerdt (36) gibt eine genaue Zusammen¬
stellung der bis jetzt angewandten Versuchseinrichtungen wieder.
Diese Apparaturen wurden von den Autoren als Temperaturorgel
oder Temperaturwahlapparat bezeichnet, wobei sich der erstere
Ausdruck am besten durchgesetzt hat.
Meines Wissens existieren ausser den Veröffentlichungen VON
Hecht (34) und Hundertmark (39) über den Wärm,esinn bei Culiciden keine weiteren Arbeiten. Hecht (34) beschreibt eingehend die
Vorzugstemperaturen der Weibchen von Culex pipiens und Anopheles
maculipennis (verschiedene Rassen) bei der Eiablage, während HüNDERTMARK (39) zum Schlüsse kommt, dass es unmöglich sei, ein
Reagieren auf Temperatur bei Winter- und Sommergenerationen
von Anopheles maculipennis festzustellen.
'
62
BÜTTIKER
WILLI
Methode
als wasserbewohnende Stadien
Für die Larven und Puppen
wurde eine Temperaturorgel aus Zinkblech nach folgenden Grössen¬
massen konstruiert :
—
Länge
Breite
Höhe
Höhe der Scheidewand
.
.
—
50
9
8
7
cm
cm
vergl. Abb. 27
cm
cm
Die Temperaturen zwischen Gefässboden und Wasseroberfläche
variierten sehr stark. Die Temperaturwerte wurden jeweils an der
Wasseroberfläche abgelesen. Da die ganze Versuchseinrichtung ziem¬
lich starken Temperaturschwankungen unterworfen war, mussten ca.
®©@
®
©
ILLLLLL
«
Abb. 27.
B, Kammer, mit
A, Heizplatte.
Temperaturorgel », schematisch.
Wasser gefüllt.
C, Wasseroberfläche..— D, Scheidewand.
E, Thermometer.
—
—
—
—
—
—
F, Eis-Kochsalz-Mischung.
alle Halbstunden Heizplatte und Kältemischung kontrolliert werden.
Für die Imagines wurde dieselbe Apparatur benützt, wie sie bei
den Versuchen des Hygropraeferendums zur Verwendung kam. Die
Glasschalen mit den Reguliersalzen jedoch wurden daraus entfernt.
An einem Ende dagegen kamen zwei Heizkissen als Wärmespender,
andern wurde eine Glasschale mit einer Eis-Kochsalzlösung
am
aufgestellt. In jedem Abteil zeigte ein Thermometer die Oberflächen¬
temperatur an.
Der Einfluss der Luftfeuchtigkeit auf den Versuchsverlauf konnte
leider nicht verhindert werden. Somit war bei den tiefen Tempera¬
turstufen eine höhere Luftfeuchtigkeit vorhanden.
evruK
0081
0061
0041
0021
0001
0080
tieZ
.tbA
.leR
1
9
8
9
10
0
5
1
2
3
1
n
12
11
12
12
11
12
°C
36
II
I
0
0
n
2
°c
1
0
2
1
2
1
n
III
13
14
14
14
14
14
°C
n
II
0
1
4
2
1
2
n
16
17
17
17
17
17
°C
°c
IV
n
3
2
1
1
1
1
n
34°
32°
35°
0
0
30°
29°
30°
1
1
,sutacrufib seerleeihtsphocnuAsreV
let iM-.tarepmeT
n igebshcusreV
64.21.5
5431
0021
0011
tieZ
.mudnerefearpomrehmTutadshcusreV
2
1
1
1
9
10
35
5
°C
n
I
I.
27°
26°
27°
°c
20
20
19
21
20
30
°C
III
V
1
1
2
3
1
2
n
IV
23
22
23
23
22
23
°C
16
13
n
VI
evraL
001—58
.mudnerefearpomrehTtiekgithcueftfuL
%.
1
3
2
1
2
1
n
23°
23°
23°
°C
-
25
25
24
25
25
24
°C
VII
rhoR
ogamI
=
1
4
1
1
2
1
n
=
10
8
n
V
50
20°
20°
20°
°C
,uaraA
n
27
26
26
27
26
26
°C
St.
3
6
7
n
VI
2
3
3
4
4
5
n
28%
28
28
28
29
28
°C
IX
18°
18°
17°
°C
°C
20
35
36
5
5
n
VII
21
16°
16°
15°
°C
Nr.
31
30
30
31
33
30
31
4
<30
X
Nr.
30
3
n
ellebaT
,64.11.42
IIIV
sneipip xeluC
von
ellebaT
,net etstlA
64
WILLI
BÜTTIKER
Resultate
'
Culex pipiens.
Larven : Obschon im Gebiet von 21—25° C eine Anhäufung der
Larven beobachtet wurde, sind die Resultate nicht deutlich. Die
Stufen von über 30° C enthielten nach kurzer Zeit keine Larven
mehr. Im Bereich der tiefen Tem100 7
wurden die Körper¬
peraturstufen
% \
\
so dass die
gehemmt,
bewegungen
80
Vorzugstemperatur nur undeutlich
\
\
zum Ausdruck kam. Sie liegt un¬
SO
\
\
gefähr zwischen 21° und 25° C.
\
70
Imagines, Sommertiere : Unter¬
sucht wurden die Weibchen ; bei
--\
m
einer Luftfeuchtigkeit von ca. 85
/
\
—100 %. Sämtliche 50 Tiere wur¬
i /
so
den im Abteil I eingesetzt. Die
\ /
W
detaillierten Resultate gehen aus der
40
V
Tabelle Nr. 20 und Abb. 28 her¬
A
Um die Übersichtlichkeit zu
vor.
30
/i
wahren, kamen nur 4 Kurven zur
/ \
/ \9-W°
20
Abbildung, nämlich die Tempera¬
/ i
turstufen von 9°C, 14° C, 28° C
10
28°
und 30° C. Die Dauer des Versu¬
U°_
ches betrug 12 Stunden, wobei je¬
0
10
1Z
S
8
weils die Imagines nach 2 Stunden
Stunden
wieder kontrolliert wurden. Die
Kurve des Thermopraeferendums
Culex pipiens. ThermopraeAbb. 28.
ferendum, überwinternde Weibchen.
(30—31°) ergab die Prozentzahlen
nach der Beobachtungszeit von
_
-sß^—
-
—
1
2 Stunden
4
,6
8
10
12
6% sämtlicher Tiere
60%
»
»
62%
»
66%
»
»
70%
»
72%
»
»
»
»
»
»
»
»
»
Die Temperaturstufe von 28° C weist maximal nur 10 % aller
Imagines auf. Längere Einwirkung von Temperaturen über 36° C
schädigen die Tiere. Im Thermostat gehaltene Tiere sterben bald in
dieser Stufe ab, sofern sie nicht Gelegenheit haben, eine tiefere
Temperatur aufzusuchen. Es kann also angenommen werden, dass
die Vorzugstemperatur von C. pipiens um 30—32° C liegt. Genauere
Temperaturangaben,
wie sie
von
andern Autoren mit
Hilfe
von
0022
0002
tieZ
lairetaM
0061
0081
0341
tieZ
In
64.11.42 lairetaM
:
nevraL
74.4.51
in
%
nennogeb hcusreV
4%
I
°C
8°
n
2
.mudnerefearpomrehT
und
VII
0
0
n
31°
32°
30°
°c
8%
4
n
2
0
n
II
II
11°
°C
28°
29°
27°
°c
10
III
20%
50
n
5
4
n
III
17°
°C
°C
25°
27°
26°
IV
34%
17
19
21
n
IV
IV.
n
ereitshcusreV mutadshcusreV
,
s
u
t
a
c
r
u
f
i
b
seleh7p4o.n4A.91
nevraL
let iM-.tarepmeT
tztesegnie
ereitshcusreV mutadshcusreV
,
s
u
t
a
c
r
u
f
i
b
sele6h4p.o2n1A.6
IV.V
.mudnerefearpomrehT
I
20°
°C
°C
23°
25°
24°
,nevraL
,nevraL
54
V
26%
13
n
18
14
n
V
23°
°C
22°
23°
21°
°C
-
6%
3
n
9
9
n
VI
VI
,liwsrep aR
uaraA
II.
27°
°C
20°
19°
20°
°C
2%
1
n
5
2
n
34°
°C
23
16°
16°
15°
22
°C
Nr.
VII
Nr.
VII
)tot(
ellebaT
ellebaT
,rhoR
1
0
0
0
10
n
1.4
0
0
0
7°
%
3.41
0012
0391
0081
nnigeB
0361
0051
74.6.7
tieZ
0051
mudnerefearpomrmeuhtTadshcusreV
kcütS
ca.
3
2
2
2
10
n
ca.
2,9
2,9
2,9
4,3
%
13°
7
7
7
6
10
n
ca.
8,6
0,01
0.01
0.01
3,41
nevraL
70
%
16°
35
43
42
10
26
n
ca.
3,16
0,06
1,73
0,05
3.41
sin epilucam
A.
%
21°
10
30
22
16
17
n
ca.
4.32
8,22
5,13
9,24
3,41
.rav
von
%
26°
3
4
2
3
10
n
ca.
:
4,3
2,9
5,7
4,3
%
3,41
3,41
tfnukreH sucipyt
der
31°
1
0
0
0
10
n
1.4
0
0
0
24
%
37°
Nr.
ca.
3.41
ELLEBAT
.hcaalF
BIOLOGIE
UND
VERBREITUNG
EINIGER
67
STECHMÜCKENARTEN
publiziert werden, finde ich nicht zweckmässig. THOMSON
(87) schlägt eine besondere Darstellungsweise vor, die den indivi¬
duellen Unterschieden der Versuchstiere besser Rechnung trägt,
Dezimalen
indem
die Tiere
einer
protokolliert
Versuchsreihe getrennt
und
graphisch dargestellt werden.
Verglichen mit andern Dipteren (NlESCHULZ, 60, 61 und 62),
ergeben sich folgende Resultate :
Culex pipiens ?, Sommertiere (eig. Beob.) 30 bis
Culex pipiens $, Wintertiere
(eig. Beob.) ca. 2
Musca domestica <$
Musca domestica $
Stomoxys calcitrans $
Fannia canicularis $
31° C
6° C
-
34,2°
33,1°
27,9°
23,9°
C
C
C
C
Anopheles bifurcatus.
Larven
:
Es kamen die Stadien I bis IV
folgende Resultate erzielt wurden
Larven
I.
II.
III.
IV.
Thermopraeferendum
»
»
»
»
»
»
zur
Untersuchung, wobei
:
:
bei
bei
bei
bei
Abb. 29
23° C
23-25° C
ca.
(ca. 23°)
20-23° C
(Tabellen Nr. 21 bis Nr. 23.)
Es geht daraus hervor, dass sich das Thermopraeferendum im
Verlaufe der Larvenentwicklung nicht ändert. Charakteristisch ist
IPC
«.
vznL---Tr-_-L7'i/.3T»
1,5
vu
m
Abb. 30.
der Larven
3
Stunden
—
Anopheles bifurcatus. Thermopraeferendum der I. und II. Larven, überwinternd.
Abb. 29.
—
4,5
S
Thermopraeferendum
von
A. maculipennis
typicus.
var.
68
WILLI
BUTTIKER
Ansteigen der Häufigkeitskurve im Gebiet der hohen
Temperaturen bis zum Thermopraeferendum, während unterhalb
der Vorzugstemperatur die Kurve langsamer ausklingt. Dies liegt
darin begründet, dass die hohen Temperaturen einen höheren Akti¬
vitätsgrad der Larven zur Folge haben. Die Tiere dieser Gebiete
können deshalb rascher die Praeferendumszone aufsuchen, als jene
des kalten Bereiches mit entsprechend niedriger Aktivität.
Das Praeferendum kann als Bereich von 23-25° C, resp. 20-23° C
angegeben werden. Es scheint nicht zweckmässig, bei Culiciden
genauere Temperaturangaben zu machen, da die Oberflächentem¬
peratur durch Wasserströmung kontinuierlich verändert wird. Die
Genauigkeit entspricht somit den Angaben mittelst Dezimalen nicht.
das steile
Anopheles maculipennis
var.
typicus.
IV. Larven : Die maximale Anzahl der Larven befindet sich im
Bereich von 21° C, wie dies aus der detaillierten Zusammenstellung
(Tabelle Nr. 24) und Abb. 30 hervorgeht.
Das Thermopraeferendum von A. bifurcatus unterscheidet sich
also nicht grundsätzlich von demjenigen der messeae-Larven.
C. Temperatur
und
Aktivität
Die Aktivität ist eine der auffallendsten und wichtigsten Aus¬
drucksformen der Lebenstätigkeit. Sie drückt sich äusserlich durch
die Muskeltätigkeit, besonders der Fortbewegungsorgane, die ihrer¬
seits vom Nervensystem ausgelöst wird, aus.
Die Abhängigkeit der Aktivität von der Temperatur ist sehr
deutlich und erlaubt es, verschiedene Aktivitätsstufen zu unter¬
scheiden. Eine Abgrenzung der einzelnen Aktivitätsstufen bietet
zweifelsohne Schwierigkeiten, da meist keine scharfen Grenzen
bestehen, sodass eine subjective Beurteilung nicht auszuschliessen
ist. Aus praktischen Gründen kann man von den einzelnen Lebens¬
äusserungen nur die Bewegungsunterschiede als Einteilungsprinzip
verwenden. Ein allgemein gültiges Schema zur Charakterisierung
der einzelnen Stufen und Grenzpunkte lässt sich nicht geben, da
die Bewegungsäusserungen verschiedener Arten nicht gleich sind.
Man kann daher von dieser Methode nur Annäherungswerte ver¬
langen, die aber für praktisch-ökologische Zwecke durchaus verwert¬
bar sind.
Eine präzisere Untersuchungsmethode kann mit Hilfe der Zeit¬
messung für einen bestimmten zurückgelegten Weg verwendet wer¬
den (vergl. Spezieller Teil), eine wesentliche Einschränkung der
Variation für die Fixpunktwerte wird sich aber kaum dar aus ergeben,
da mit individuellen Unterschieden der Tiere zu rechnen ist.
BIOLOGIE
UND
VERBREITUNG
EINIGER
STECHMÜCKENARTEN
69
In den Untersuchungen über den Einfluss der Temperatur auf
die Aktivität soll versucht werden, einige Punkte eingehender zu
bearbeiten. Eine genaue Kenntnis der Lebensweise der Culiciden
ist von grosser Bedeutung und kann für die Praxis wertvolle Angaben
liefern.
Untersuchungen über den Aktivitätsgrad bei Insekten wurden
schon von verschiedenen Autoren besrhrieben, so von Arnim über
Lymantria monacha (1), VAN Heerdt über Forficula auricularia (36),
Nieschulz an Stomoxys calcitrans (60), und anderen. Jvanova (41)
veröffentlicht Beobachtungen an A. maculipennis var. messeae und
atroparvus, wobei auch der Einfluss des Lichtes berücksichtigt wird.
Dieser Autor kommt zum Schluss, dass die erwähnten maculipennisRassen zwei Aktivitätsmaxima (das erste bei 17° C, das zweite bei
30—32° C) haben und dass der Aktivitätsbereich zwischen 6° und
40° C liegt. Eine weitere Angabe über Temperatur und Aktivität
enthält die Publikation v. Rudolfs (75) über Aèdes sollicitons, eine
Culicide hauptsächlich des subtropischen Nordamerikas (Atlantik¬
küste von Texas, Oklahoma, New Mexico). Vergleichsweise wurden
seine Resultate bei den entsprechenden Untersuchungen mittel¬
europäischer Culiciden benützt (s. Tabelle Nr. 28).
In der vorliegenden Arbeit gelangen die einzelnen Entwicklungs¬
stadien zur Untersuchung. Neu in diesem Zusammenhange ist die
Prüfung sämtlicher vier Larvenstadien von A. bifurcatus, die Unter¬
suchung der angeführten Puppen und Imagines. Ausgenommen
dagegen ist A. maculipennis var. messeae. Im Vergleich zu IVANOVA
(41) ergaben sich aber bei meinen Larvenuntersuchungen andere
Resultate.
Methode
Die Abgrenzung der Aktivitätsstufen und
nach BODENHEIMER (1929, 4) durchgeführt :
1. Beginn der Kältestarre
2. Leichte Bewegung von Gliedmassen
3. Tiere kriechen mit Unterbrechung
4. Normale Aktivität
5. Starke Aktivität
6. Höchste Erregung
7. Beginn der Wärmeparalyse
8. Wärmetod.
Grenzpunkte wurde
Zwei Proben zu je 10 Untersuchungs-Tieren (Larven, Puppen oder
Imagines) kamen aus dem Zuchtgefäss bei Zimmertemperatur in
je einen Glastubus von ca. 10 cm Höhe und 2,5 cm 0. Die erste Probe
wurde für den Versuch oberhalb von 4° C verwendet.
Für die Versuche eignete sich die Apparatur, wie sie in Abb. 31
dargestellt ist. NlESCHULZ (60) empfiehlt für die Aktivitätsversuche
70
WILLI
BUTTIKER
ein Bassin, welches zur Sterilisation medizinischer Instrumente ver¬
wendet wird.
Zur Kühlung wurde jeweils eine Eiskochsalzmischung
hergestellt, mit welcher Temperaturen bis zu —8° C erreicht wurden.
—
Die Erwärmung der
Heizplatte, wobei die
Versuchsapparatur geschah durch eine elektr.
Temperaturhöhe von 46° C die obere Grenze
bildete.
Die relative Luftfeuchtigkeit war in den Versuchen mit Imagines
nicht genau zu kontrollieren und kaum konstant zu halten. Um den
Fehler möglichst klein zu machen,
wurden jeder Probe einige Tropfen
Wasser zugegeben. Die relative
Luftfeuchtigkeit betrug deshalb an¬
nähernd 100 %, so dass die Ver¬
suchsresultate miteinander vergli¬
chen werden können.
Bei den Larven von Culex pipiens,
Aedes geniculates, Anopheles maculipennis und A. bifurcatus wurde die
Schwimmgeschwindigkeit
Abb
31.
-
Versuchsapparatur
zur
Bestimmune der Aktivitatsstuten, tur
,
Larven, Puppen und Imagines. Sehematisch.
B, Ther¬
Â, Becherglas.
—
—
C, Gefäss mit Culiciden.
D, Becherglas mit Eis-KochsalzE, Isoliermaterial aus
Mischung.
F, Eis- und Kochsalz.
Sägemehl.
G, Heizplatte.
mometer.
—
gemessen
und graphisch dargestellt.
Die Aktivitätsuntersuchungen er¬
streckten sich meistens über eine
Zeitspanne von je ungefähr 1 %
Stunden. Man muss sich vergegen¬
wärtigen, dass die Körpertemperatur
der Insekten bei Temperaturverän¬
derungen erst mit einer gewissen zeitHchen Verschiebung den Wärmegrad
des
.
sie
.
i
umgebenden
n «•
i
•
Mediums
an¬
(Chapman, 10).
Untersuchungsresultate der
genommen
hat
Die
einzelnen Arten wurden entspre¬
chend miteinander verglichen und
darnach ausgewertet. Bei A. bifur¬
catus diente die Untersuchung aller
4 Larvenstadien dazu, einen all¬
unterschiedlichen Verlauf der Aktivitätskurve während der
Larvenentwicklung feststellen zu können.
—
—
—
—
fälligen
ganzen
Resultate
Larvenstadium I. bis IV. bei der
gleichen Art (A. bifurcatus).
Am 11. Dezember 1946 standen für die
genügend I. und II. Larven
an
den III. und IV. Larven
zur
erst
Aktivitätsuntersuchungen
Verfügung, während die Versuche
am 18. April 1947 zur Ausführung
BIOLOGIE
UND
VERBREITUNG
EINIGER
STECHMÜCKENARTEN
71
kamen. Es zeigte sich aber, dass die Charakteristik der Kurve bei
allen Stadien dieselbe ist, und dass das Versuchsdatum keinen Einfluss auf das Resultat hatte. Die Aktivitätskurven der einzelnen Ent¬
wicklungsstadien überdecken sich praktisch. Individuelle Unter¬
schiede und kleine Unregelmässigkeiten im Experimentierungsverlauf
ergaben nur unbedeutende Abweichungen. In der Tabelle Nr. 25
sind die Resultate zusammengefasst.
Larvenstadium IV. verschiedener Arten.
Das Einfrieren im Wasser wird von keiner der untersuchten
Arten ertragen. Selbst A. bifurcatus als Überwinterer in der Larven¬
form ist demnach ziemlich kälteempfindlich. Eigenbewegungen der
Larven ermöglichen das Fernbleiben von der eigentlichen Gefrier¬
zone des Wassers, was auch aus einigen Freilandbeobachtungen her¬
vorgeht (Vergl. Blatt « Feldbeobachtungen » hiezu, Seite 78).
Bei 0° C verhalten sich 4 Arten gleich, nämlich C. pipiens,
A. vexons, A. geniculates und A. maculipennis var. messeae. Eine
Ausnahme davon macht A. bifurcatus, welcher schon bei dieser
Temperatur schwache, allerdings sehr träge Schwimmbewegungen
ausführt. Das Ansteigen der Aktivitätskurve nimmt bei allen
Arten ungefähr denselben Verlauf. Die Wärmestarre liegt bei den
ersten vier Arten um 37—40° C, während sie bei A. bifurcatus
bei ca. 34—35° C liegt. Die detaillierten
als Kaltwasserbewohner
Versuchsergebnisse sind in Tabelle Nr. 26 zusammengestellt. Die
von Ivanova (41) an Anopheles maculipennis-Larven durchgeführten
Aktivitätsversuche konnten nur zum Teil bestätigt werden ; so
konnte das in jener Arbeit erwähnte erste Maximum nicht beobachtet
werden. Nach IVANOVA liegt das erste Maximum bei 17°C, das von
einem Minimum bei 23—28° C abgelöst wird ; ein zweites Anwachsen
der Aktivität beginnt bei 30—32? C und erreicht sein Maximum
bei 37—39° C. Verglichen mit schweizerischem Material stimmen
aber obere und untere Aktivitätsgrenze gut überein.
—
—
Puppenstadium.
Ein ähnliches Bild ergibt sich aus den Experimenten mit Puppen.
Das Einfrieren in Wasser erträgt keine dieser Arten. Bei ungefähr
3—4° C beginnen die ersten deutlichen Schwimmbewegungen, mit
A. bifurcatus als Ausnahme. Es sind dort schon oberhalb +0° C deut¬
liche Schwimmbewegungen wahrzunehmen. Der Anstieg der Akti¬
vitätskurven ist bei allen Arten in grossen Zügen gleich. Bei A. bifur¬
catus liegen die Stufen um einige Grade tiefer, so dass der Wärme¬
tod hier bei 35° C eintritt, bei den andern um 40—41° C. In der
nachstehenden Tabelle Nr. 27 sind die Resultate enthalten.
72
WILLI
BUTTIKER
Imaginalstadium.
Sommergenerationen : Culex pipiens und Anopheles maculipennis
messeae zeigen grosse Ähnlichkeit ; bei C. pipiens liegen die
Aktivitätsstufen ungefähr 1 ° C tiefer. Einen grösseren Unterschied
var.
vexans, bei welcher der Kältetod um 0° C liegt. Die
Kältestarre endet erst bei +5° C. Die ersten deutlichen Aktivitätsan¬
zeichen treten meist bei 6—7° C auf. Die Wärmestarre, sowie der
Wärmetod, befindet sich ebenfalls 1°C höher als bei A. maculi¬
pennis var messeae. Interessant in diesem Zusammenhange sind die
Untersuchungen V. Rudolfs (75) an Aèdes sollicitons, einer südlichen
Culicidae aus Nordamerika, bei der die Kältestarre bei ungefähr
17° C liegt. Sämtliche Aktivitätsstufen liegen aus diesem Grunde
innerhalb eines engeren Bereiches, nämlich zwischen 17° C und
44° C (Differenz 27°), während sie bei A. maculipennis var. messeae
zwischen 4° C und 40° C (Differenz 36° C) liegen. Die detaillierten
Untersuchungsresultate sind in der nachfolgenden Tabelle 28 und
der Abb. 32 zusammengefasst.
zeigt Aedes
Abb. 32.
Aktivitätsstufen und Aktivitätskurven der Imagines verschiedener Culiciden.
I, Culex pipiens.
2, Aedes Vexans.
3, Aedes sollicitais.
4, Anoph. maculipennis
—
—
—
—
var.
überwinternde
—
messae.
Weibchen : Die Aktivität erfährt während der
Diapause eine sehr deutliche Reduktion. Nachdem die Weibchen
ihren Uberwinterungsort aufgesucht haben, verbleiben sie
mit
in Ruhestellung. Bei den
Ausnahme bei der Wasseraufnahme
vorliegenden Versuchsexemplaren handelt es sich um Winterweib¬
chen von Culex pipiens und Anopheles maculipennis var. messeae, die
2. November 1946, resp. 13.11.46, untersucht wurden. Mit
am
diesen verglichen wurden die Sommermücken vom 30. Juli 1947,
woraus deutlich hervorgeht, dass bei entsprechenden Temperaturen
im Winter immer ein tieferer Aktivitätsgrad erreicht wird (Tabelle
Nr. 29).
In einer gesonderten Untersuchung versuchte ich einen allfälli¬
gen Unterschied im Verhalten von Männchen und Weibchen fest¬
zustellen. Bei allen Aktivitätsstufen zeigten aber C. pipiens und
A. maculipennis var. messeae gleiches Verhalten.
'
—
—
37
36
35
35
eznerG
33
des
30
an
d.
4
ca.
-1
0
+1
16
2
32
mumixaM
nn.itgäteiBv
.negnug
hcil
.tiekgihäf
fl.
ben
ab.
der
der
12.
zu.
¬eweB
nedrew
emhanuZ
driW
ressaW
enho
-4
¬itkA
elatämtriosnnetdnnIrehännA
negnugeweB
46
in
Be¬
¬mmiwhcS
¬tued
nemhen
e,nteet.ghrnceuhigcrieelgwtneuu.ebdnmeginwuhgceSw
gnugi.dnäeghacrStre ,ethcieL
e
n
e
r
o
r
f
e
g
n
i
E
nevraL
11.
37
36
35
34
35
32
33
30
16
4
2
+1
0
-1
ca.
-4
°C
eznerG
der
des
d.
mumixaM
.tätiv
nnigeB
an
.negnug
elamron dnrehännA
hcil
den
emhanuZ
.negnug
in
fl.
ben
ab.
driW
¬rets
°C
der
¬itkA
zu.
¬eweB
¬tued
nemhen
negntuätgiesnwneeetbngIm.ntuiigekweghwihecäBSf
etethc.irreehgcniult,ueetdhcieL
¬mmiwhcS
11.
12.
46
¬rew
¬eweB
enho
ressaW
g.nnueggairdtärhecS
e
n
e
r
o
r
f
e
g
n
i
E
nevraL
II.
37
36
35
35
34
32
33
30
16
4
2
+1
0
—1
ca.
-4
°c
eznerG
des
d.
mumixaM
.tätiv
nnigeB
an
den
ben
ab.
in
.negnug
driW
¬rets
muidatsnevraL
I.
von
der
zu.
4. 47
A.
15.
fl.
der
nemhen
¬eweB
krats
ressaW
nevraL
emuidhactussnreevvrsatLätivitkA
an
Be¬
37
36
35
35
32
33
34
30
4
an
2
°C
-4
ca.
-1
0
+1
16
eznerG
der
des
d.
mumixaM
.tätiv
nnigeB
elamroN
nedrew
elamrontätdinsrneethnäInnA
n
e
g
n
u
g
e
w
e
B
.
t
i
e
k
g
i
h
ä
f
.rehciletmuhedanuZ
en.tenegetgnhacurigtrreeegwnegu.bnnumeg,egiitndwhäuchhgicceeSSwL
e
n
e
r
o
r
f
e
g
n
i
E
¬mmiwhcS
¬rew
enho
¬rets
.III
nevraL .VI—.I
34
.sleshcewf otS .dotemräW .sleshcewf otS .dotemräW .sleshcewf otS .dotemräW .sleshcewf otS .dotemräW
ben
IV.
ab.
15.
zu.
nemhen
¬itkA
.erratsemräW
netregietseg .err.attästeivmirtäkWA netregnieettrseeggiet.seergr.attästeivmirtäkWA netregnieettrseeggiet.seergr.attästeivmirtäkWA netregnieettrseeggietseg .tätivitkA
n
e
t
r
e
g
i
e
t
s
e
g
¬itkA
n.neegg.nnrueuhggceeiwlwtnteäeueBtbeigdsmnnueitgnweIhwceSB
nedrew
driW
hcon
.negnug
enho
,negart
gissülf
das
4. 47
nrefos
.tsi
er¬
Nr.
nevraL
ressaW
¬eweB
krats
enegte.nrtuehghcceiirwleetgubneumd,eitwhhccieSL
gnugidähcSenerorfegniE
25
¬rets
muidatsnevraL
ellebaT
muidatsnevraL
sutacrufib
40
39
38
37
35
33
32
31
16
30
8
3
4
der
ednE
+2
d.
ekratS
etsrE
-1
der
d.
der
der
des
nedrew
nur
im
Eis
41
40
39
37
33
32
18
12
5
+2
+4
-1
—2
ednE
°c
.tätiv
der
13.
der
der
der
5. 47
d.
der
nnigeB
.errats
nnak
nereirfniE
—2
.sl.esdhocteewfmroätWS
erebO
.ndro
n.neirrgaetBs
.droegnu .täntniivigtekBA
..ttäättiivviittkkAA eeemllhaaammnrruooZNN
-egnu
¬emräW
eznerG
.e.rrdaottseemmrrääWW .negnugeweB
mgunmugiexwaeMB
.
n
e
.errat
ge-
ge-
¬sem.rtäätWivitkAnn.iggieetBs
.gnugemwueBmi.xgiaetMs
.etmähtiavnitukZA
netsre
¬itkA
.tätivitkA
.elamroN
.negenmuhgaenwueZB .neegnmuhganenew.ugerZneBhucgieltwueeBd
.er atsetläK .er atset.leärK atsetläK
¬etläK
eginie
.nedrew negartre
.keS
8. 46
40
41
39
38
36
31
17
30
7
+4
-2
—I
nnigeB
der
.negnug
netendroeg
.dotemräW
..eerrrraattsseemmrrääWW
d.
der
der
der
°C
un¬
¬eweB
netendro
mumixaM
.geweB
¬egnu
.täntniviigtekBA
.tätivitkA
elamroN
emhanuZ
.ddootteettllääKK
sneipip xeluC
30.
3. 6. 47
41
40
-1
+5
7
9
17
32
35
37
38
39
gen
der
d.
der
—2
A.
d.
3. 6. 47
°C
.dotemräW
..eerrrraattsseemmrrääWW
ednE
ednE
.er atsetläK
.
.ddootteettllääKK
snaxev sedeA
°c
.tseg
der
in
un-
¬nugewebm iwhcS
.treg
.täntniivigtekBA
.tätivmiutkmAixaM
¬ietseg nnigeB
.tätivitkA
.tätiveiltkaAmroN
emhanuZ ekratS
.hciltued
.gnuge,weerBats.elttluäeKd
nerredarotetsieerttfllnääiKKE
mieb
.siE
sin epilucam
sutalucineg
e
h
c
u
s
r
e
v
s
t
ä
t
i
v
i
t
k
A
nevraL
A.
37
36
34
35
35
32
33
30
16
4
+2
0
—1
—4
°C
.
men
gen
zu.
des
der
krats
tätis
«
ven
A.
der
an
negnugeweB
nedrew
driW
gnug
ethcieL
enho
rereg
15.
».
ge¬
ab.
bei
Nr.
4. 47
nebrets
eaessem
.VI(
an
.sl.esdhocteewfmroätWS
eznerG
.erratsemräW
netreg
nnigeB
.tät i v i t k A n e t r e g i e t s
.tätivimtkuAmixaM
-ietseg
-nugeweB elamroN
¬idähcS
¬hen
¬netnI
e.¬thecnitulhgtcueie.rwdenegebngumnuigwehwceSB
,neegrarrattrseetläK
.kriwniE
enerorfegniE
¬raL
26
¬näl
sutacrufib
ellebaT
)muidats
41
39
40
36
31
28
bis
16
.dotemräW
erebO
erebO
.tätiv
d.
mumixaM
nnigeB
elamroN
5
der
d.
¬itkA
.tätiv
nnigeB
+3
d.
41
40
39
38
36
15
18
34
10
5
3-4
-2
tsres
¬itkA
.negnnuegsehwceaBwnA sekratS
.
e
r
.daottseettllääKK
sneipip xeluC
—2
-1
nnigeB
der
.dotemräW
der
der
der
3. 6. 47
Ak¬
39
40
40
38
36
30
35
18
bis
8
5
3-4
—2
der
.dotemräW
nenriegbeOB
erebO
.tätivit
mumixaM
nnigeB
elamroN
.masgnal
ella hcoN
ednE
etsrE
°c
°C
der
der
der
3. 6. 47
35
34
33
31
16
11
3
2
1
+0
-0
—1
-2
°C
an
rhes
mumixaM
elamroN
¬suä
.neegrnautgseetwleaBK .dotetläK
sedeA
3. 8. 46
A.
d.
d.
d.
18.
zu.
4. 47
reba
ekratS
an
.dotemräW
nenriegbeOB
mumixaM
elamroN
nem.htednnaweg
hcilmeiz
.masgnal
Nr.
remmi
emhanuZ
etsrE
sin epilucam selehponA
ednE
.ltued
.rav
ehcusrevstätsuitvailutckiAneg
°C
eznergleshcewf otS
krats
27
hcon
nnigeB
.negnug
eaessem
¬ebmmiwhcS .negnugew
.
e
r
a
t
s
e
t
l
ä
K
.dotetläK
¬itkA
.ezn.eerzgnleersghscteäwtfivoittSkA
netr.eegrireat.tstesägetimvriä.tWnkAegnugeweB .eznergleshcewf otS
.
e
r
r
a
t
s
e
m
r
ä
W
.
e
z
n
e
r
g
s
t
ä
t
i
v
i
t
k
A
t
ä
t
i
s
n
e
t
n
I
.
e
r
r
a
t
s
e
m
r
ä
W
n
e
g
n
u
g
e
w
e
B
.tätivitkA
nneeggnnuuggeewweebbmm,hciiiwwlhhtucceSSd netregiet.steägtivitkA .tätivitkA netrengeitertesgeigetseg .tätivitkA netregnieett.rsteeäggtiievtistekgA
.
n
e
g
n
u
r
e
s
u
ä
s
t
ä
i
v
.tätiv.ittäktAivietkdAnemhenuZ
.
n
e
'
g
e
w
e
b
m
i
w
h
c
S
¬ewe.bnmemginuwghecwSeB
..eerr..daalottttssueeeetttdlllnäääUKKK
sutacrufib
ellebaT
neppuP
41
39
40
23
35
um
18
12
8
9
7
den
.neg
4
a.
elamroN
negeilF
emasgnaL
d.
.ltued etsrE
xeluC
+3
der
der
43
41
42
39
40
19
37
12
10
6
7
5
4
+3
—I
°C
nnigeB
.xaM
nohcs
.tsiD ezruk
.tug
.legülF
enieB
.negnugew.ereBhnc.eieglrtnuauetgdseetwleäBK
¬rew
sneipip
-5
.erratsemräW.dotemräW .e.rnreagtnseumgreäwWeb.heer.nrGdeaohtetcsgeieimemtrrzsäänaWWHA
d.
.tätivitkA .gietseg
der
d.
.tätivit
nnigeB
elamroN
.ltueD
etsrE
.tdnaweg
.netre.gtiäneteitvsgientgukgAewebgulF .tänteigvint.ukgegAögemnwrueegbveggwuulelFFB
n
¬
n
u
g
e
w
e
b
t
i
e
r
h
c
S
Ak¬
.negnugewebheGugeweB
.
n
e
g
n
e
t
e
t
h
c
i
r
e
g
n
u
...eeerrr.daaaottttssseeeettttllllääääKKKK
snaxeV sedeA
ehcusrevstätivitkA
°C
an
45
44
44-
42
40
39-
20
39
17
17
°C
.tnU
A.
erebO
erebO
.dotemräW
..ezenzenregrsgtsätäitvivtiktAkA
.eznerg'.wefrortaStsemräW
.t.ättäitviivtiktAkAetetrreedgniiemtrseeVG
.
e
r
a
t
s
e
t
l
ä
K
32)
snatàillos
senigamI
»
42
40
41
39
36
31
16
23
10
6
4
+3
-5
°C
A.
.er.rdaottseemmrrääWW
nnigeB
ten
der
der
zu
zu.
un¬
¬e.nedrrroaetgsne.umnreägWnugmeuwmeiBxaM
ne.nnneidgreebw negtneundgreoweegB
.tätivitkA teältaimvirtokNA
sin epilucam
etsrE
.neg
.er atsetläK
tsaf
.krem tmmin
nengenguö.grmeetrhwäecteviiblvghtiuutelekGFdA
nohcs
.lamron
.t u g
nedrew
.rav
Nr.
28
¬nugeweB ehciltued
eaessem
ellebaT
.bbA(
41
egitsah
35
Die
23
12
Bei
bei
Die
ellA
.ret
18
.hcsit
ereiT
trofos
39
40
.er.rdaottseemmrrääWW
d.
negeilf
ereiT
in
ereiT
8-9
thcin
hcua
der
;
;
bei
.nie
¬dnawneegmhen
des
die
reba
7
rhem
negnures uästä iv tkA
dnis
nur
-salG
5
41
nedrew
egitsaH
bei
39
40
36
35
23
18
9
12
6
7
8
4
+3
—5
Nur
auf
elamroN
°c
d.
ezruk
.nie
tiv
nemhen ereiT .tdnaw
.tug
,rehc
etsrE
4
eniek
dnis
,legülF n.engeegwneuBg
dnis
.ltued
reba
nereiT nella
.drew
hcon
thcin
¬iltued
46
30.
ge¬
7. 47
42
41
40
39
36
31
16
10
6
b.4
+3
—5
°C
ten
den
zu.
men
neres
etsrE
etsrE
11.
der
mumixaM
in
reba
¬aler nohcs
.ltued
2.
d.
der
tmmin
;
dlab
hcon
nedrew
_.negnugeweB n.eegrnautgseetwleäBK
dnretniwrebü
sneipip xeluC
+3
nnigeB
;
negnures uästä iv tkA
egitsah
¬rew
zum
enho
ereiT
rabkrem
.nie
¬hen
¬suä
krats
¬tued
.negnugeweB n.eegrnautg.sreeetwhlceäiBKl
ereitremmoS
ehcusrevstätivitkA
-5
42
nnigeB
Bei
gen
d.
u.
13.
A.
11.
46
;
40
41
39
36
31
16
10
6
4
+3
-5
°C
var
ten
der
mumixaM
zu.
der
zu
tendroeg
eniek
lieT
eginie
an
tmmin
tsaf
thcer
rehmu
negeilf
reba
etsrE
dnretniwrebü
enieB
¬nu.gleegwüelBF .ltu.eedrnuatsetläK
sin epilucam
senigamI
°c
.nedrew
un¬
,lamron
rabkrem
.¬seurärsattästeivmirtäkWA etne.ennhdecrgioneeuzgrnneAus
.erratsemr.änWeg.neur.grdeaotwtseeeBmmrrääWW
.er.rdaottseemmrrääWW negnugewegbnhureGt ühcsrE
.er.rdaottseemmrrääWW ¬endroegnutend.rnoeeggnnuugeweB .enrergantusgemerwäeWbgul.nFterhhücfieegzsnuAa ¬siretka.grgnanuhulcleettssffaallhhccSS
.z.nnaetsginDugewebheG
¬e.nedrrroaetgsn.eunmreägWnugeweB
n
e
g
n
u
g
e
w
e
b
g
u
l
F
.
t
ä
t
i
v
i
t
k
A
g
n
u
l
e
t
s
f
a
l
h
c
S
t
g
n
u
r
e
t
ü
h
c
g
n
u
l
e
t
s
f
a
l
h
c
S
eähtcisviittsikrA
nennigeb negnugeweB
.ssersEsäfeg
..gnul entgsenfguaöllmhtercätSetsvifvagiluhtckFSA ¬etk.agrnualhcetnnseefaggslnnshuuuclgSgfeen.wwiteEedbbnggauuwlleFFg .negnungeegwnneeubgggöuemlwrFeebvgguullFF gnur.egtnuülhcestrsEehuR
¬
w
b
i
e
r
h
c
S
emasgnaL
.
n
e
g
n
u
g
e
w
e
b
h
e
G
n.negeggnennuraruafegletpebmwüeehxBcEsrE
.
t
l
e
t
s
e
g
t
s
e
f
emasgnaL
nnee.gtgndnunug.agrewewehwegcebibglthuuelFGd
enieK
nohcs
30.
Nr.
7. 47
nedrew
etsrE
.neg
.er atsetläK
29
¬nugeweB .ltuednu
eaessem
ereitremmoS
ellebaT
78
WILLI
Freilandbeobachtungen
an
Larven
BÜTTIKER
von
A.
bifurcatus.
Aus einem Randtümpel des Katzensees bei Zürich wurden Eis¬
stücke entnommen, im Laboratorium aufgetaut und auf Insekten
(speziell Culicidae) untersucht.
Probe I. 25. Januar 1947 : Ungefähr aus der Mitte des Tümpels
wurde ein Eisstück von 35 X 35 cm herausgeschlagen ; seine Dicke
betrug 8 cm. Unmittelbar unter der Eisdecke hielten sich Larven von
Theobaidia annulata und A. bifurcatus auf, von denen durch Herauspipettieren 3 Larven von Theobaidia annulata und 4 von A. bifurca¬
tus (II. und III. Stadien) erhalten werden konnten.
Beim Auftauen der Eismasse kamen 16 Larven von Dixa spec.
(lebend), eine Larve von Chironomus spec, (tot), ein Exemplar von
Hydrometra (tot) und 1 Wassermilbe zum Vorschein.
Probe II. 2. Februar 1947. An derselben Stelle wurde ein gleich
grosses Eisstück dem Tümpel entnommen. Nebst 8 eingefrorenen
Larven von Dixa spec, (lebend) enthielt die Probe 12 verlassene
Köcher von Trichopteren und eine Larve von Chironomus (tot).
2. Der Einfluss der
Luftfeuchtigkeit
Es ist für die Tiere von grösster Wichtigkeit, ein bestimmtes
Gleichgewicht zwischen dem Wassergehalt des Körpers und dem
des umgebenden Mediums zu halten. Der Betrag des Wassers variiert
bei den verschiedenen Arten sehr stark ; so unterscheidet sich der
Gehalt bei einigen Wasserinsekten von der Umgebung nur um
1 Prozent (CHAPMAN, 10). Nebst den amphibischen Insekten, welche
grösste Feuchtigkeitsunterschiede überdauern können, kennt man
« xerophile
», die in sehr trockenem Milieu leben können. Die Feuch¬
tigkeitsdifferenzen halten sich aber bei ihnen in engen Grenzen.
Als besonderer Faktor wirkt die Feuchtigkeit in tiergeogra¬
phischer Hinsicht in der Begrenzung des Verbreitungsgebietes.
Feuchtigkeit kann in Form von flüssigem und gasförmigem
Wasser vorliegen ; der feste Zustand des Wassers wird nicht in
diese Bezeichnung einbezogen. Es ist möglich, den Feuchtigkeits¬
gehalt der Luft in verschiedener Weise auszudrücken, nämlich als
relative Feuchtigkeit in Prozenten der bei einer bestehenden
Temperatur möglichen Sättigung ;
Sättigungsdefizit in mm Hg ;
Dampfspannung in mm Hg ;
absolute Feuchtigkeit in g H2O pro dm3.
Das Psychrometer, welches auf dem Prinzip des Feuchtigkeits¬
und Trockenheits- Thermometers basiert, ist das gebräuchlichste
BIOLOGIE
UND
VERBREITUNG
EINIGER
STECHMÜCKENARTEN
79
Instrument, das auf einfache Art die relative Feuchtigkeit bestimmen
lässt. In kleinen Räumen ist es aber wegen der durchzuführenden
Schwingbewegungen nicht geeignet, so dass in diesen Fällen ein
geeichtes Haar hygrometer verwendet werden muss. In den beschrie¬
benen Laboruntersuchungen kamen Haar hygrometer zur Anwen¬
dung, während Freilandmessungen mit dem Psychrometer durch¬
geführt wurden.
Besteht ein Feuchtigkeitsgefälle, so kann bei vielen Insekten, die
bis jetzt eingehend auf den Einfluss der abiotischen Faktoren unter¬
sucht wurden, eine deutliche Vorliebe für einen bestimmten Bereich
beobachtet werden. Analog dem Thermopraeferendum kann dieser
als Hygropraeferendum bezeichnet werden ; die benützte Laborein¬
richtung, welche das Herstellen stufenförmiger Feuchtigkeitswerte
ermöglichte, als Feuchtigkeitsorgel ».
Die Abhängigkeit der Lebensdauer von der Luftfeuchtigkeit bei
konstanter und verschiedener Temperatur wird in einem speziellen
Abschnitt des vorliegenden Kapitels besprochen.
Beim Thermopraeferendum übt die Feuchtigkeit eine reiz¬
physiologische Wirkung aus, bei der Beziehung von Feuchtigkeit
zur Lebensdauer dagegen sind es zur Hauptsache Einwirkungen auf
«
die
Stoffwechselphysiologie.
A. Hygrotaxis, Hygropraeferendum (Vorzugsfeuchtigkeit)
Van Heerdt (36) gibt in seiner Arbeit über Forficula avicularia
eine gute
Literaturübersicht, auf die an dieser Stelle hingewiesen
werden soll. Im folgenden werden einige der wichtigsten neueren
Arbeiten erwähnt :
Krijgsman und WlNDRED (1930) konnten bei ihren Versuchs¬
tieren (Stomoxys calcitrans und Lyperosia exigua) nur eine hygropositive Reaktion erhalten, nachdem die Tiere lange Zeit gehungert
hatten.
Kenneday (1937) berichtet über das Verhalten von Locusta
migratoria, die eine ausgesprochene Vorliebe für trockene Luft auf¬
wies.
GuNN (1937) fand bei Blattiden nur eine positive Reaktion,
nachdem sie einige Zeit ohne Wasseraufnahme waren.
WlGGLESWORTH (1937) beschreibt ausführlich das Reagieren von
Pediculus humanus corporis auf Luftfeuchtigkeit. Frisch geschlüpfte
Eilarven vermeiden Feuchtigkeitswerte von 95 %. Tiere, die an eine
hohe Luftfeuchtigkeit gewöhnt sind (95 %), vermeiden anfänglich
eine tiefere, doch erfährt das Verhalten eine Änderung, bis zu ca.
50 %.
PlELAN (1940) schliesst aus seineTi Versuchen, dass das Wahr¬
nehmen der Feuchtigkeit bei Pediculus in den Antennen lokalisiert
ist.
80
WILLI
BUTTIKER
Teubner (84) schreibt, dass die Sommermücken schneller auf
mikroklimatische Unterschiede reagieren als Wintermücken und
dass A. maculipennis übersättigter Luft ausweicht.
De Meillon (55) stellt fest, dass A. gambiae und A. funestus stets
die hohe Luftfeuchtigkeit der niederen vorzogen. THOMPSON R. C. M.
(cit. lit. 84) beobachtet, dass C. Jatigans Feuchtigkeiten oberhalb
95 % meidet. Hungertiere nehmen angeblich noch eine Differenz
von 1 % wahr. Hundertmark (38) stellt bei seinen eigenen Unter¬
suchungen fest, dass bei A. maculipennis var. superpictus eine Ver¬
schiebung des Hygropraeferendums eintritt, nämlich von 81 %
(I.Tag) auf 97% (2. Tag).
Methode
Das
bestand
Gefäss, in dem die Imagines
einem 130 cm langen, 10
aus
« Feuchtigkeitsorgel ».
Abb. 33.
Quer¬
schnitt durch eine Versuchskammer.
2 Glasplatte.
J, Wand des Holzgehäuses.
4, Gaze
3, Trennungswand aus Glas.
als Abschluss gegen Petrischale.
5,
7,
6, Salz.
Übersättigte Salzlösung.
Petrischale.
—
—
—
—
—
—
—
—
zur
Beobachtung gelangten,
hohen und 13 cm tiefen
Holzbehälter, der oben mit
einem Glasdeckel versehen
war (Abb. 33). Es befanden
sich darin 9 Scheidewände
(Glas 8 cm hoch), die den In¬
nenraum in 10 gleiche Kam¬
mern einteilten. Das Feuch¬
tigkeitsgefälle wurde nach der
Zwölfer'schen Methode mit¬
telst übersättigten Salzlösun¬
gen erreicht (vergl. Zwölfer,
99), wobei für diese Stoffe
cm
folgende theoretische Werte
angegeben werden (Tab. 30).
Die
Versuchsbedingun¬
gen
änderten
Versuches
tant.
Die Dauer eines Versuches
von
Unter¬
Untersuchung.
Sie blieben aber jeweils wäh¬
rend eines durchgeführten
suchung
zu
annähernd kons¬
lag zwischen ein bis zwei Tagen.
Resultate
C. pipiens*
Überwinternde Weibchen. Die Versuchstiere kamen im Verlaufe
3 Tagen zur Beobachtung, deren Resultate in Tab. Nr. 31
zusammengestellt sind. Im Verlaufe der Beobachtungszeit sammelten
sich immer mehr Tiere im Gebiet hoher Luftfeuchtigkeit an, sodass
von
BIOLOGIE
UND
VERBREITUNG
EINIGER
STECHMÜCKENARTEN
81
der Prozentsatz der Imagines in der wassergesättigten Kammer von
26 auf 67 anstieg. Sämtliche Imagines, welche sich ursprünglich in
den Kammern mit niedriger Luftfeuchtigkeit befanden, begaben sich
in das Gebiet höchster Luftfeuchtigkeit. Die Aktivität der Winter¬
mücken, besonders bei C. pipiens, ist selbst bei relativ hohen Tem¬
peraturen gering, sodass eine weitere Verschiebung zugunsten der
flohen Kammern (neun und zehn) zu erwarten gewesen wäre. Nach
3 Tagen wurde der Versuch abgebrochen, der deutlich genug zeigte,
dass C. pipiens an Orten mit wässerig-feuchter Unterlage über¬
wintert. Es besteht offenbar das Bedürfnis, öfters flüssiges Wasser
aufzunehmen. Exemplare, die längere Zeit an trockenen Stellen ver¬
weilen, wechseln auch im Latenzzustande den Standort, um an
geeigneter Stelle Wasser aufzunehmen. Die Abb. 34 enthält drei
Kurven :
Kurve n (Anfang) gibt die Prozentwerte zu Beginn des Ver¬
suches wider, Kurve n (Ende) dieselben am Schlüsse. Die Feuchtig¬
keitswerte kommen in der Kurve der rel. Feuchtigkeit zum Aus¬
druck.
Abtohl
Abb. 34.
—
Z345S789
Hygropraeferendum
von
10
C. pipiens, überwinternde Weibchen.
A. maculipennis (verschiedene Rassen).
schreibt eingehend über das ÜnterscheiLuftfeuchtigkeit der drei in Deutschland vor¬
«
:
Anophelen sind wahlfähige Insekten, die
Hundertmark (37)
dungsvermögen
der
Rassen
kommenden
optimale mikroklimatische Gebiete aufsuchen können. Im Wahl¬
versuch wird von den Mücken stets die Kammer mit der jeweilig
höheren Luftfeuchtigkeit vor der mit der niederen Luftfeuchtigkeit
stark bevorzugt ». Die Varietäten unterscheiden sich nach dem glei¬
chen Autor im Verhalten gegenüber der Luftfeuchtigkeit :
Hygropraeferendum
:
atroparvus
messeae
typicus
«
...
100 %
97 %
95 %
r.
r.
r.
F.
F.
F.
o
m
kl
OOOOOOOOOO
OOOOOOOOOO
o
100%
O
et
2
os
Z
W
0
e5v
M
1
!
1
1
1
1
1
1
iriuMnuMrimiPiuMnm
O^ ejs c3n ej\ 0^ e^s 0"n o\ c^n 0s
%
26
=
u
_)
j
H
B
*t>st^aOvOr>.t30tv.<»2
os
vp
tNe>itv)T}-u^-^rr>.i-».rvoo
e»aoe»ooooooooaoaOoO
B
cnencnen-*ï-TcN-<i--*j-'*i-
u
n
o
en
o
0
u
:3
4-»
U.
os
vP
oN
u
00
vp
ejv
^
4
II
O
m
sP
o
z
w
vO
3=20% 4=26%
80
%
ca.
20
=
OO
E
B
II
3
V
-B
IV.
rv
NaC
ca.
vp
eî-
11
vO
E
tu
B
os
vO
O D"
"
Z
-cs'^
CV.CM-
U.
01
VO
70
b
OS
U
vp
nO ^O nO nÔ n© ^D vO \Ô nû >0
1^
11
B
_,—
vO
in
U
=3
SC
60
os
m
ca.
vp
m
m
O
D"
U
es
^^
00
c
B
»*
ipt
C
«.
-4-t
X
os
•>r
a
u g"
n
^5
e>v
5J
<
en
3
o^
os
tT°
O
en
vp
eu
e--)
&
V*
OS
o
e»
g"
c?v
0
60
>>
X
_
b.
a
0
i/Mri.—e3NCOtvmt>qe>jcn
*r-'l--<rcncnenenenenen
o
u
et
eîv
—
B
eu
co
ca.
vP
B
ä
B
ut;
46
b.
OS
en
E
\Ou",ii/\,i/\,ir\.ir-\.u'\ir\,ifMrN.
.—-«
60
g,0
"vP
B
4-»
u
et
•*
—
—
^—
u
—
1-
vP
cyv
B
20
%
ca.
20
=
flC\!f\——
3
cu
O.
E
eu
CA
9,"
eu
»
^-5
0^
0
U
3
OOOOOOOOOO
OOOOOOOOOO
0,Ne>|ONt30<NC2\C3001tJNCO
O.—
—
O
—
—
O
—
—
O
r>,
rv
rv
rv
"*.
"^
"^
"^
en
i"
<N
•*
o
oj
rvl
e>|
Anfang Schlus
lettiM
etnezorP/n
:lairetaM
5101
5431
64.11.51
5461
0080
0021
64.11.61
lettiM
F.
58%
eguaS
r.
17
17
15
39
40
42
40
13
10
40
10
14
40
11
%
RF
2
5
3
2
4
16%
.voN
n
n
RF
6491
im
15
5,71
5,71
%
mudnerefearporgyH
2
)24(
I
5%
1
2
I
1
n
3
45
44
45
2%
n
45 •2
45
45
%
RF
4
RF
)05(
/o
3%
2
1
n
5
60
59
60
60
60
60
%
RF
sin e:pmiluutcaadmssheclueshrpeoVnA
°C
3%
1
1
1
n
6
/o
RF
1
1
n
7
2%
.voN
15.
.6491
.rav
10.
)56(
eaessem
La
70
70
61
70
70
71
%
RF
0%
n
8
%
RF
)57(
20
3%
•
2
1
n
9
80
80
80
81
80
80
%
RF
9%
1
2
2
2
2
n
90
001-09
001-09
17°
Nr.
10
ca.
C.
32
%RF
001-09
001—09
001-09
ellebaT
:rutarepmetshcusreV
e
r
e
i
t
s
h
c
u
s
r
e
V
84
WILLI
BUTTIKER
Bei den Untersuchungen von Hundertmark (37) handelt es sich
Sommer mucken. Ich verwandte für meine Experimente über¬
winterte Weibchen (messeae). Während des Versuches wurde dif¬
fuses Licht verwendet, so dass eine störende Wirkung vermieden
werden konnte. Die Raumtemperatur betrug ca. 17° C.
Aus der Tabelle Nr. 32 und Abb. 35 gehen die detaillierten
Resultate hervor.
um
r.E
-ifOO
_:
%
*~
-
80
~60
^sJssä^s^
-40
-20
Abteil:
Abb. 35.
—
0
8
1
Anopheles maculipennis
var. messeae,
Weibchen.
9
10
Hygropraeferendum
;
überwinternde
Demnach wurden die niedrigen Feuchtigkeitsstufen bevorzugt
Der mittlere Bereich enthält nur wenig Tiere,
r. F.).
während die Kammer der höchsten Feuchtigkeit von 90—100 % r. F.
wieder einen höhern Prozentsatz aufwies. Die Imagines nahmen
jeweils an Kondenswassertropfen Flüssigkeit auf ; nach einiger Zeit
flogen aber die Mücken aus der feuchten Zone in die trockene.
Verglichen mit C. pipiens ergibt sich also ein prinzipieller Unter¬
schied. Dieser besteht darin, dass A. maculipennis var. messeae nur
zur Wasseraufnahme feuchte Stellen bevorzugt, als Uberwinterungsort aber trockenes Milieu aktiv aufsucht. C. pipiens dagegen hält sich
vornehmlich an dauernd feuchten Stellen auf, und zwar während
und nach der Wasseraufnahme. Diese Tatsachen stimmen mit den
Beobachtungen an den Uberwinterungsplätzen sehr gut überein, wo
regelmässig C. pipiens an nassen Orten anzutreffen ist (Wasser¬
leitungen, nasse Mauern), während A. maculipennis var. messeae, scharf
von diesen getrennt, an trockenen Orten überwintert.
Feuchtigkeitsmessungen an Uberwinterungsorten ergaben für
A. maculipennis var. messeae und C. pipiens folgende Werte (gleiches
(bis 25 %
Lokal)
:
Rübenkeller, La Sauge.
7. März 1946
11. Nov. 1946
März 1947
C. pipiens
C. pipiens
C. pipiens
100
% ; A.
100%;
100%;
mac.
mess.
»
»
»
»
70 % 5° C.
75-80% 8° C.
60—70% 9° C.
BIOLOGIE
UND
VERBREITUNG
EINIGER
B. Luftfeuchtigkeit
und
STECHMÜCKENARTEN
85
Lebensdauer
über die Lebensdauer von Insekten bei verschiedener Feuchtig¬
keit liegen verschiedene Literaturangaben vor. Eine Verlängerung
der Entwicklungsdauer, hervorgerufen durch niedere Luftfeuchtig¬
keit, beschrieben erstmals HUNTER und PlERCE (40) bei Anthonomus
grandis.
Bacot und Martin (2) berichten, dass Xenopsylla cheopis als
Imago bei 89 % r. F. ohne Nahrung 152 Stunden am Leben bleibt,
dagegen nur 27 Stunden bei 27 % r. F. (32° C). Es handelte sich
dabei um Tiere, die vor Versuchsbeginn gefüttert wurden ; im
Gegensatz zu Leeson (44), der mit frisch geschlüpften Imagines
experimentierte. Es ergaben sich dabei andere Resultate.
Angaben finden sich bei WlGGLESWORTH (97) über Anopheles
spec, welche 24 Stunden bei 0 % r. F. und 32° C und 24 Stunden bei
100 % r. F. und 37° C ertragen konnte. Hundertmark (37) publi¬
ziert Ergebnisse über die Lebensdauer der drei Anopheles maculipennis-Rassen (messeae, atroparvus und typicus) ; er stellt fest, dass
die Lebenszeit bei den Männchen geringer ist als bei den Weibchen,
und dass atroparvus viel länger lebt als messeae ; typicus nimmt eine
Mittelstellung ein.
Über die Abhängigkeit der Lebensdauer der Culiciden von der
umgebenden Luftfeuchtigkeit veröffentlicht auch MARTINI (52) Beo¬
bachtungsresultate, die übereinstimmend eine Verlängerung der
Lebensdauer mit steigender Luftfeuchtigkeit nachweisen. MARTINI
stellt fest, dass die Lebensdauer der Culiciden zu verschiedenen
Jahreszeiten nicht gleich ist. Nebst Temperatur und Luftfeuchtig¬
keit spielt die aufgenommene Nahrungs- und Wassermenge eine
entscheidende Rolle. Anopheles punctipennis konnte in einem Versuch
bis zu 231 Tagen mit Dattel- und Wassernahrung am Leben erhalten
werden, während die voll Blut gesogenen Weibchen bis zu 205 Tagen
lebten. Die durchschnittliche Lebensdauer betrug aber nur 90,4 Tage.
Frisch aus den Puppen geschlüpfte Imagines, die nicht gefüttert
werden, zehren von den gespeicherten Stoffen (Fett, Kohlehydrate) ;
sie sind daher physiologisch anders zu beurteilen als mit Pflanzen¬
säften und Blut versehene Exemplare. Nicht ausser Acht zu lassen
sind die Unterschiede zwischen Sommertieren und überwinterten
Weibchen ; letztere weisen in der Diapause auch bei hohen Tem¬
peraturstufen eine geringe Aktivität auf. Durch die weniger häufigen
und langsameren Bewegungen der Wintermücken, die also in die¬
sem Falle nicht nur von der Temperatur abhängig sind, reduziert
sich dementsprechend der Nahrungsverbrauch beträchtlich, so dass
die gespeicherten Reserven für eine längere Lebensdauer ausreichen.
Bei den vorliegenden Untersuchungen handelt es sich um die
beiden praktisch wichtigsten mitteleuropäischen Culiciden (Culex
11
ooooooooooooooooo
ooooooooooooooooo
33
O
ta
1
^9
1
1
1
1
1
1
1
1
1
1
1 1
1
1
1
1
\0 nO \0 nO ^O vO nO vO nû vO \0 nO vO nO vO nO v©
X
3
OOOOOOOOOOOOOOOOO
11
33
P
Z
7Î
^9
3
OOOOOOOOOOOOOOnOnOnOnOnOOOOO
OOOOOOOOOOOOOOOOOOOOOOOOnOnOnOnO
OOOOO
3
1
*»
NO
—N)UJ^U,on
|
O
_.
NO
11
33
O^
oooooooooooooooooooooooo
O
7<
3
11
33
o
Z
^9
MUiNJOOvûSSoo |
^jOoOonOnOnOOOOOO
^l-J^J^I^^JOOOOOOOOOO
Ni
3
o NO 00 *J On -U. t-O
1
—
—
—
33
go
o
z
n
^9
3
33
P
0**00000
—NiNiKi' O
MSIOOCDnOvCOOO
"
3
33
NO
00
|
UJ
UJVJiON
|
—NJ O
UiUiONOOO
nP
OQ
2
O
O^
ta
O
1
o
OOOOOOOOOOOOnOnOOO—'NJ-U
UiVJiUiVJiVJ1nJ1\Ji\JiOnOnOnOnOn
o
71
O—
0O -4
3
|
|
—ON o
«J On
33
O
3
N
Ui On V^i VJi \Jl
"nP
3
—NJUi^J O
11
P
Substanze
33
^J"^100
3
ONJV/i
n
09
Tage
bu>blnbuibu'buibuibu>bu<
o
——
opovavjoNONUtyijUjUOJWNjjv}
BIOLOGIE
UND
VERBREITUNG
EINIGER
STECHMÜCKENARTEN
87
pipiens und Anopheles maculipennis var. messeae), bei denen die
Lebensdauer als Funktion der Luftfeuchtigkeit bestimmt wurde.
Der kombinierte Einfluss Temperatur und Luftfeuchtigkeit kommt
im Abschnitt b), « Kombination von Luftfeuchtigkeit und Tempe¬
ratur bezüglich der Lebensdauer » zur weiteren Besprechung.
Konstante
Temperatur
Methode
Verschiedene Feuchtigkeitsgrade wurden bei einer konstant
gehaltenen Temperatur mit übersättigten Salzlösungen in gut verschliessbaren Rezipienten hergestellt. Um die gewünschten Abstu¬
fungen zu erreichen, kamen die übersättigten Salzlösungen nach
Zwölfer (99) zur Anwendung, die folgende theoretische Werte
ergeben (Tab. 33) :
Die erhaltene Luftfeuchtigkeit variierte jeweils im Verlaufe des
Versuches ; ihre Veränderungen sind aus den entsprechenden Tabel¬
len ersichtlich. Die Auszählung der gestorbenen Tiere wurde täglich
zweimal durchgeführt, nämlich morgens 0800 und abends 1900 oder
2000 Uhr.
Resultate
Culex pipiens.
Lebensdauer überwinterter Weibchen, am Ende der Diapause
(Tab. Nr. 34) : Alle Exemplare in sehr niedriger Luftfeuchtigkeit
(P2O5, 7—8 % r. F.) starben schon nach sehr kurzer Zeit ab. Die
Mortalität betrug innerhalb 1,5 Tagen 100 %. In den höheren Feuch¬
tigkeitsstufen verlängerte sich dementsprechend die Lebensdauer.
Besonders auffallend gross war der Unterschied zwischen der Stufe
von 84 % r. F. und 100 % r. F., wie dies in den Kurven VIII resp. IX
zum Ausdruck kommt. In der Stufe von 88 % r. F. betrug nach
8 Tagen die Mortalität 100 %, während sie sich bei wassergesättigter
Luft in derselben Zeit auf 0 % belief. Die Weibchen der letzteren
Kategorie starben erst im Verlaufe weiterer 6 bis 8 Wochen (April,
Mai) ab, da die Nahrungsreserven bis zu diesem Zeitpunkt voll¬
ständig aufgebraucht waren. Das Maximum der Lebensdauer über¬
winterter Weibchen beträgt demnach unter normalen Verhältnissen
schätzungsweise 8—8 % Monate (September bis April, Mai).
Die Mortalitätskurven sind meist deutliche Sigmoïden, welche
das Integral der Gauss'schen Glockenkurven darstellen. PratoLUNGO (101) erhält in seiner Arbeit über die Prüfung von Insektiziden
ähnliche Resultate. Die Sterblichkeitskurven stellen ebenfalls Sig¬
moïden dar.
88
WILLI
BUTTIKER
Lebensdauer frisch geschlüpfter Männchen und Weibchen : Auch bei
diesen Versuchstieren kam ganz deutlich das starke Feuchtigkeits¬
bedürfnis zum Ausdruck. Die durchschnittliche Lebensdauer frisch
geschlüpfter Imagines ohne Nahrungsaufnahme beider Geschlechter
beträgt bei 22° C
:
rel. F.
Männchen
Weibchen
8%
20%
34%
56%
76%
93%
100%
0,2 Tage
»
0,2
0,25 Tage
»
0,3
»
0,6
»
1,0
»
2,0
»
2,4
»
2,5
0,4
0,7
1,4
1,6
)>
»
»
»
Der Unterschied in der Lebensdauer zwischen Männchen und Weib¬
chen kann im Zahlenverhältnis 2 : 3 bis 3 : 4 angegeben werden.
Die Lebensdauer mit Pflanzensaft gefütterter Weibchen : Als Unter¬
suchungsobjekte dienten frisch geschlüpfte Weibchen, welche mit
Dattelfleisch gefüttert wurden. Es stand ihnen dreimal täglich Wasser
Verfügung, das mit Hilfe eines Zerstäubers auf den Aufenthalts¬
käfig gespritzt wurde. Verursacht durch die Sonneneinstrahlung,
schwankte die Raumtemperatur im Versuchshause zwischen 18° und
zur
30° C. Die beobachteten Lebenszeiten sind
wegen
diesen starken
Temperaturschwankungen nur Annäherungswerte.
Die längste beobachtete Lebensdauer betrug im
70 Tage ; ungefähre Mittelwerte ca. 20—30 Tage.
Anopheles maculipennis
Lebensdauer
liegt bei A.
Im Herbst
bilden aber
vor.
Maximum
messeae.
überwinternder Weibchen
Der Überwinterungsmodus
ähnlich wie bei Culex pipiens.
nehmen die Weibchen eine Blutmahlzeit zu sich ; sie
Fett anstatt Eier und zehren im Verlaufe der Latenzzeit
an kühlen, dunklen Stellen von den gespeicherten Stoffen. Im Früh¬
jahr erfahren die Weibchen mit eintretender Temperaturerhöhung
eine deutliche Aktivitätssteigerung. Durch eine erneute Blutmahlzeit
wird es den Weibchen ermöglicht, die Eier auszureifen und sie nach
der Praeovipositionszeit von ca. 3 Tagen abzulegen.
Die in dieser Untersuchung verwendeten messeae-Weibchen
15 Stück) wurden am 9. März 1947 dem Uberwinterungsort
(n
(Raumtemperatur : +4° C) entnommen und in 10 Präparategläschen
=
maculipennis
var.
messeae
:
BIOLOGIE
UND
VERBREITUNG
EINIGER
STECHMÜCKENARTEN
89
in die Thermostatstufen von —2 bis —3° C und 100 % r. F. über¬
führt. Alle Exemplare starben aber erst zwischen Juli und Mitte
September ab. Es ergibt sich daraus eine Lebensdauer von 10—12
Monaten, nämlich von September bis Juli-September des folgenden
Jahres. Nach der Abb. 37 (Mortalitätsdiagramm) liegt das für die
Lebensdauer optimale Gebiet bei 100 % r. F. und 0° C.
Lebensdauer frisch geschlüpfter Männchen und Weibchen ohne
Fütterung : Es ergeben sich bei gleichen Versuchsbedingungen
ähnliche Resultate wie bei Culex pipiens. Die maximalen Werte liegen
bei 100 % r. F., die minimalen bei geringster Luftfeuchtigkeit. Die
Versuchstemperatur betrug wiederum 22° C.
Durchwegs haben die Männchen eine kürzere Lebensdauer als
die Weibchen, wie dies aus folgender Zusammenstellung deutlich
hervorgeht
:
rel. F.
Männchen
Weibchen
8%
20%
0,6 Tage
»
0,7
»
1.5
2,1
2,4
2,4
»
2,5
0,8 Tage
»
1,0
34%
56%
76%
93%
»
»
»
100%
2,0
2,8
3,4
3,5
3.5
»
»
»
»
»
Kombination
von Luftfeuchtigkeit und Temperatur
bezüglich der Lebensdauer
(Mortalitätsdiagramm)
Die Bedeutung der Luftfeuchtigkeit für die Insekten tritt speziell
im Zusammenhange mit dem Stoffwechsel, der Aktivität und Mor¬
talität in Erscheinung. Der Einfluss Iässt sich hauptsächlich bei
Hungertieren und Entwicklungsstadien, denen eine Wasserzufuhr
aussen her unmöglich ist, nachweisen ; er steigert sich mit der
Temperatur, und zwar nicht nur deshalb, weil Temperaturerhöhungen
die Verdunstung an sich erhöhen, sondern auch wegen der dadurch
bedingten Steigerung der Atmungsintensität, welche Gelegenheit zu
Wasserverlusten gibt.
Die kombinierte Einwirkung von Temperatur und Feuchtigkeit
auf die Sterblichkeit, die von Stadium zu Stadium, von Art zu Art
von
verschieden ist, kann man im sogenannten Mortalitätsdiagramm dar¬
stellen. Man erhält so, indem man die Punkte mit gleichen Sterblich¬
keitswerten (oder der gleichen Lebensdauer) verbindet, Kurven, die
90
WILLI
BUTTIKER
sich um ein Zentrum scharen, welches man als «vitales Optimum»
bezeichnen kann.
Im Thermohygrogramm sind also drei Grössen vorhanden, näm¬
lich Temperatur, relative Feuchtigkeit und Lebensdauer. Auf dem
Wege des physiologischen Experimentes hat man versucht, diese
wichtigsten Faktoren zu analysieren. Die Methodik derartiger Ver¬
suche besteht darin, die einzelnen Stadien bei den verschiedensten
Kombinationen der Faktoren und ihre Wirkung auf die Lebensdauer zu
prüfen. Man bedient sich dazu der sogenannten Thermohygrostaten,
die es gestatten, die gewünschte Kombination herzustellen und
konstant zu halten. Von manchen der wirtschaftlich wichtigen Insek¬
ten existieren die Mortalitätsdiagramme, so z. B. von Carpocapsa
pomonella (Shelford, 79), von der Nonne, Panolis flammea (ZwöLFER,
98), von Schistocerca gregaria (BoDENHElMER, 4). Ähnliche Unter¬
suchungen an mitteleuropäischen Culicidenarten sind mir nicht
bekannt.
Methode
:
Zur erreichung der gewünschten Luftfeuchtigkeit kamen sieben
überstättigte Salzlösungen nach ZwöLFER (99) zur Anwendung. Aus¬
serdem stand der Reihenthermostat mit 7 Stufen zur Verfügung,
wodurch 49 Kombinationen erziehlt wurden. Es kamen folgende
Feuchtigkeits- und Temperaturwerte zur Anwendung :
1
Chem. Verbindung
.
.
.
Feuchtigkeit theoretisch %
Praktisch erreicht
%
2
3
4
5
6
P20B ZnCU MgCl2 Ca(N03)2 NaCl KN03
7
H20
0
10
34
56
76
93
100
8
20
34
56
76
93
95-100
-2°
+2°
10°
]7o
22°
27°
350
Resultate
Culex pipiens.
Die ungefütterten und frischgeschlüpften Weibchen erreichten
eine maximale Lebensdauer von 4—5 Tagen bei —2° bis +2° und
100 % r. F. Im normalen Temperaturbereich von 15 bis 27° kann
bei 100 % r. F. eine Lebensdauer von ca. 2—3 Tagen erreicht werden.
BIOLOGIE
UND
VERBREITUNG
EINIGER
91
STECHMÜCKENARTEN
Im Bereich über 30° C nimmt die Lebensdauer sehr schnell ab,
wobei sie bei 35° C und 8 % r. F. nur noch 0,1 Tag (2 x/2 Std.)
beträgt.
Das Gebiet der grössten Lebensdauer liegt zwischen —2° und
+2° C und bei 100 % r. F., das Pessimum dagegen bei 35° und 8 %
relativer Luftfeuchtigkeit. An Stelle der Isomortalen sind in der
Abb. 36 Kurven gleicher Lebensdauer eingezeichnet.
Die Männchen wiesen eine merklich kürzere Lebenszeit auf als
die Weibchen ; sie betrug je nach Temperaturstufe 2I$ bis % des
entsprechenden Weibchenalters. In einer Vergleichsreihe kamen die
männlichen Imagines bei 100 % r. F. zur Untersuchung. Es ergaben
sich
folgende Resultate (Tab. 35)
Lebensdauer
von
:
frischgeschlüpfter
Culex pipiens bei
Männchen und Weibchen
100 % rel.
Feuchtigkeit
(ohne Nahrungsaufnahme)
Durchschnittliche Werte
°c
Männchen
—2
2
+2
2,5
10
17
22
27
35
Tage
TABELLE Nr. 35
Weibchen
Differenz
3.5 Tage
1,5 Tage
1.5 »>
1.5
1.0 Tag
4
»
3.5
2
2
»
3
»
2.5
»
2.0
0,66 Tag
—
1.5 Tag
»
0,5
Anopheles maculipennis
Dar.
»
—
0,5
0,16
»
»
ungefähres
Verhältnis
>
23
7a
7s
—
74
74
messeae.
Die Verhältnisse bei dieser Culicidenart liegen ähnlich wie bei
Culex pipiens. Die Männchen weisen ebenfalls eine kürzere Lebens¬
dauer auf als die Weibchen. Die Resultate der Versuche mit frisch¬
geschlüpften und ungefütterten Weibchen sind in der Abb. 37 ein¬
liegt das Gebiet mit der längsten Lebensdauer
bei niedriger Temperatur (—2 bis +2° C) und höchster Luftfeuch¬
tigkeit (100 % r. F.). Die Lebensdauer bei Weibchen beträgt im
Maximum 13—14 Tage, bei Männchen um 10 Tage. Das Pessimum
befindet sich im Diagramm dargestellt diagonal gegenüber der Stelle,
hohe Temperaturen und geringe Luftfeuchtigkeit eingetragen
wo
sind. Bei 35° C und 8 % r. F. ergibt sich demnach eine Lebensdauer
von nur 0,2 Tagen (= ca. 4,8 Std.).
getragen. Wiederum
92
WILLI
BUTTIKER
Tage
0.5
03
0.2
0,1
0
10
Abb. 36.
o/oKl
12
1H
12
—
10
20
C. pipiens, Weibchen. Mortalitäts-Diagramm.
8
2
6
100
90
/
/ /
V
°C
30
,
Tage
j,
i
/ /
80
\
1.0
70
60
*
s..
50
W
\
^
30
20
10
\
A
V.
s
Abb. 37.
—
20°
10°
A.
maadipennis
//
^ s
'
\-
0°
OS
var.
messeae,
30
/
°C
Weibchen. Mortalitäts-Diagramm.
Q2
BIOLOGIE
UND
VERBREITUNG
EINIGER
93
STECHMÜCKENARTEN
Aus der Tabelle Nr. 36 können die vergleichenden Ergebnisse
Experimente über die Lebensdauer von Männchen und Weibchen
entnommen werden. Das Leben der Männchen ist durchschnittlich
Va bis 1U kürzer als dasjenige der Weibchen.
der
Lebensdauer
von
frischgeschlüpfter
Männchen und Weibchen
Anopheles maculipennis var. messeae bei 100 %
(ohne Nahrungsaufnahme)
Durchschnittliche Werte
°c
Männchen
-2
6,5 Tage
»
8,0
4,5
»
3,0
2,5
2,2
1,0 Tag
+2
10
17
22
27
35
»
»
»
r.
F.
TABELLE Nr. 36
Weibchen
10
12
Differenz
Tage
6—7
4
3,5
3,5 Tage
4,0
»
2,5
»
»
1
Tag
1
»
»
3,0
1,5 Tag
0,8
0,5
>>
»
ungefähres
Verhältnis
V'
%
3U
3U
3U
3u
%
In diesem Zusammenhang wurde die Lebensdauer von mit
Pflanzensaft und Blut genährten Weibchen verglichen, wobei sehr
grosse Unterschiede festzustellen waren.
bei 5°C und 100% r. F. :
frisch geschlüpft, ungefüttert :
frisch geschlüpft, mit Pflanzensaft
gefüttert :
gefüttert (Wintermücken)
mit Blut
Lebensdauer
»
:
3. Der Einfluss der
»
10 Tage
40 Tage
5—6 Monate
Strahlung
Bei der Grosszahl aller Lebewesen, speziell auch der Insekten,
spielt die Strahlung eine lebenswichtige Rolle ; sie ist eine der grössten
Energiequellen der Lebewesen überhaupt. Bei der Betrachtung der
Eigenschaften verschiedener Wellenlängen hat sich gezeigt, dass sich
allem die kurzen Wellen durch ihre chemisch-physikalische
vor
Wirksamkeit auszeichnen. Es ist aber nötig, dass das Licht absor¬
biert wird, um überhaupt einen Effekt zu erzielen.
94
WILLI
BUTTIKER
Die Bedeutung der Strahlung für einen Organismus muss vor¬
sichtig beurteilt werden, da verschiedene Lichtintensitäten auch ver¬
schiedene Effekte zur Folge haben (TrÖNDLE, 88). Die Untersu¬
chungen von LAURENS und Hooke (43) zeigen, dass die Strahlungen
verschiedener Wellenlänge verschiedene Werte des Körper¬
von
wachstums haben.
Die Experimente von Lutz (45) deuten darauf hin, dass Insek¬
ten auf kurze Wellen sehr empfindlich sind und Uberdosierung zu
Schädigungen führt.
Belaard (3) untersuchte 1858 bei Musca den Einfluss verschie¬
dener Lichtfarben und kam zum Resultat, dass das Licht die Körpergrösse beeinflusst. Die grössten Exemplare entstanden unter violett,
die kleinsten unter grün. Das Grössenverhältnis betrug 3:1.
Gal (21) machte Versuche mit Exemplaren von Bombyx mori,
die ein unterschiedliches Wachstum und eine unterschiedliche
Eiproduktion bei verschiedenem Licht ergaben.
Davey (13) studierte den Einfluss von x-Strahlen (Röntgen¬
strahlen) auf Tribolium ; er konnte eine Verlängerung der Lebens¬
dauer bei täglicher Bestrahlung feststellen.
Packard (64) stellte eine Beziehung zwischen der Dosis von
Röntgenstrahlen und der Sterblichkeit der Eier von Drosophila fest.
Mavor (54) wies bei Drosophila deutliche Verschiedenheit in der
Empfindlichkeit auf x-Strahlen während der Entwicklung nach.
Nach der bisherigen Literatur kann darauf geschlossen werden,
dass die Wirkung von Art zu Art unterschiedlich ist und bei den
Entwicklungsstadien stark variiert.
Parker (65) züchtete Heuschrecken unter künstlichem Licht. Es
konnte aber kein Unterschied zu den Kontrolltieren des Tages¬
lichtes festgestellt werden.
Der Einfluss des langwelligen Lichtes wurde nicht so eingehend
untersucht wie derjenige des kurzen. Allgemein wurde angenommen
(Chapman, 10), dass das ultraviolette Licht für die Metamorphose
des Insektes nötig sei.
Halberstaedter, Goldhaber und Hecht (35) untersuchten die
Wirkung von Röntgen-Strahlen (X-Rays) auf die Entwicklung von
Calliphora erythrocephala und Culex piptens, wobei eine unterschied¬
liche Empfindlichkeit festgestellt wurde.
A. Strahlung
und
Stoffwechselphysiologie (Photosynthese)
Bei den Larvenzuchten von Anopheles macttlipennis var. messeae
und var. melanoon zeigte sich ein auffallend starkes Lichtbedürfnis
während der Entwicklung. Ungenügende oder fehlende Bestrahlung
führte zum Absterben der Larven. Um Anhaltspunkte über den
Wirkungsgrad des Lichtes zu bekommen, wurde folgende Versuche
durchgeführt
:
BIOLOGIE UND VERBREITUNG
Versuch
a :
EINIGER
STECHMÜCKENARTEN
95
Larvensterblichkeit bei völliger Dunkelheit.
Bei normalen Fütterungsbedingungen kamen je 50 Larven in
fast völliger Dunkelheit (Thermostat), resp. unter Lichteinfluss einer
elektrischen Glühlampe von 25 Watt zur Aufzucht. Die Wassertem¬
peratur betrug in beiden Gefässen ca. 22° C. Der Verlauf der Larven¬
sterblichkeit kommt in der Abb. 38 zum Ausdruck : Die Larven¬
zucht unter Lichteinfluss weist eine Larvensterblichkeit von 23 %
auf (Kurve I), diejenige im Thermostat-Abteil 94 %. In andern
Fällen, wo vor die Thermostatfenster noch schwarze, metallene
Schutzplatten montiert wurden, stieg die Larvensterblichkeit sogar
auf 100 %. Aus den vorliegenden Ergebnissen kann geschlossen
werden, dass die Strahlung für das Gedeihen der Larven von Bedeu¬
tung ist. Diese Tatsache bildete den Ausgangspunkt einiger weiterer
Experimente über Strahlungseinwirkung bei Culiciden.
15
Tage
Abb. 38.
Larvensterblichkeit bei Anopheles maculipennis var. messeae, im Verlaufe
der Larvenentwicklung.
(I), Aufzucht unter Einwirkung der Sonnenstrahlung.
II, Aufzucht der Larven im Thermostat ohne Bestrahlung.
—
—
—
In der Folge wurde ein Versuch durchgeführt, der den Einfluss
der verschiedenen Strahlungsstärken der verwendeten Glühlampen
abklären sollte.
Versuch b
:
Larvensterblichkeit bei verschiedener Strahlungs-Intensität.
Stufenweise wurden elektrische Glühbirnen von steigender
Stärke angeordnet, zu denen je ein Zuchtgefäss mit 50 Eilarven
gehörte. Die Lichtstärken der verwendeten Lampen betrugen * :
1
Der
Daten sehr
Vertretung
zu
der Firma
Dank verpflichtet.
Philips
in Zürich bin ich für
Mitteilung
dieser
96
WILLI
16,5 Watt
25
40
60
75
100
200
BUTTIKER
140 Lumen
»
240
»
440
»
740
»
990
»
1420
»
2750
(Lichtstrom)
Der prozentuale Anteil der einzelnen Spektralgebiete an der
Strahlung einer normalen Wolframwendel-Glühlampe Philips » ist
folgender :
«
Ultraviolett
Sichtbarer Bereich (4000 —7000
Infrarot
À).
praktisch
0
ca.
9
ca.
91
%
%
%
Glühlampenlicht weist dem Tageslicht gegenüber ein Manko
blauen und grünen, aber einen Überfluss an gelben und roten
Strahlen auf. Die Larven wurden bei den vorliegenden Versuchen
somit einer überdosierten Gelb- und Rotstrahlung ausgesetzt.
Die Bestrahlung der Larven erfolgte täglich während 12 Stun¬
den ; die restliche Zeit des Tages verbrachten die Larven in völliger
Dunkelheit.
Die Versuchsresultate sind in Tabelle 37 ersichtlich. Es geht
daraus hervor, dass Larven, die vollständig unbestrahlt in der Dun¬
kelheit gehalten wurden, innerhalb kurzer Zeit abstarben. Es ergab
sich in diesem Falle eine Sterblichkeit von 100 %. Die weiteren
Zahlenwerte waren :
Das
an
Lampenstärke
Kontrolle (voll. Dunkelheit)
16,5 Watt
25
»
40
»
60
75
100
200
»
»
»
»
Sterblichkeit
%
70%
20%
14%
100
4%1
6%)
Minimum d. Sterblichkeit
10%
%
100
Demnach liegt das Optimum im Gebiet zwischen den Lampen¬
stärken von 60 und 75 Watt, die eine Larvensterblichkeit von nur
A %, resp. 6 % ergaben. Die grösste Larvensterblichkeit liegt bei
200 Watt. Sie lässt sich durch Schädigung infolge zu hoher Strah¬
lungsintensität (Verbrennung !) erklären.
Aus dem vorliegenden Versuch geht aber nicht hervor, welche Ge¬
biete des Spektrums hauptsächlich am Strahlungseffekt beteiligt sind.
BIOLOGIE UND
Versuch
Die
VERBREITUNG
EINIGER
STECHMÜCKENARTEN
97
Ultraviolett- und Infrarot-Strahlung.
c : Der Einfluss der
folgenden Teiluntersuchungen bezweckten den Effekt der
ultravioletten und infraroten Strahlung zu ermitteln. Es wurde den
Fettkörpern, den Pigmentstoffen, den Oenocyten und den Uratausscheidungen besonders Beachtung geschenkt.
Leider konnte eine genaue qualitative Untersuchung der Wellen¬
längen bezüglich des photosynthetischen Effektes nicht durchge¬
führt werden, da ein Mikroskop-Comparimeter für die Prüfung
nicht zur Verfügung stand. Anhand des Absorptionsdiagrammes
bätte auf die anwesenden Pigmentstoffe und auf ihre Absorptions¬
fähigkeit geschlossen werden können.
Das Vergleichen der Fettkörper von Larven, welche bei ver¬
schiedener Intensität bestrahlt wurden, zeigte merkliche Unter¬
schiede. Die Menge des gespeicherten Fettes, als auch die Aachenmassige Ausdehnung der Fettkörper und die Menge der darin gelös¬
ten Pigmentstoffe, zeigten deutliche Zusammenhänge mit der Bestrahlungsintensität.
Dem Fettkörper fällt die Aufgabe der Fettspeicherung zu ; dieses
Speicherorgan kann allerdings auch weitere Stoffe, wie Proteine und
Glycogen, enthalten. Fettropfen und Proteünkörner treten gewöhn¬
lich bald auf, nachdem die Nahrungsaufnahme begonnen hat. Am
Ende der Larvenentwicklung sind normalerweise die Fettzellen mit
Fett, Protein und Glycogen angereichert (WlGGLESWORTH, 97).
Ausserdem sind im gespeicherten Fett Pigmentstoffe vorhanden, die
bei den Stoffwechselvorgängen eine entscheidende Rolle spielen.
Vermutlich sind chemische Stoffe der Pterine bei der Entwicklung
von Bedeutung. Carotinoide (Lipochrome) werden oft aus Pflanzen
absorbiert und im Blut und in den Geweben angereichert (Fett¬
körper von Pyrrhocoris). Wie weit Carotinoide in der Insekten¬
physiologie eine Rolle spielen, ist nicht bekannt (WlGGLESWORTH,
97, S. 335).
Die Farbe der Oenocyten im frischen Zustand ist oft hellgelb bis
ockerfarbig ; bei einigen Insekten sind sie farblos. Vielfach sind sie
mit den Fettkörpern eng verbunden. Sie sind oft gelappt und ziehen
sich über die Körperoberfläche hin. Über ihre Funktionen herrscht
Unklarheit ; aber die meisten Autoren stimmen darin überein, dass
die Oenocyten Organe sind, die ihre Sekrete dem Blut übergeben.
Vermutlich haben die Oenocyten Anteil an der Pigmentspeicherung,
Sekretion von Stoffwechselprodukten aus dem Blut, Produktion von
Cytolysefermenten und Häutungshormonen und schliesslich an der
Produktion
Versuch d
von
Hormonen für die Gonadenentwicklung.
Ultraviolett-Bestrahlung.
für die nachfolgenden Teil-Versuche eine Ultra¬
violettlampe (U. V.-Filter und Original Hanau-Lampe [S 300]) mit
:
Es standen
7
dunkel
Kontrole
w
100
Bem rk.
0
—
0
CS
— — —
CS
\o
ooo
CS CS-TT
•f
Tin
— —
fCS — — COCOT
CO CO CO CO CO CO coco
—
Pup ePup en
Bem rk.
2
r^oooovooo
CSCOCOCSCSCSCOCO
CS
—
Bem rk.
45
1
»n
CO
CS
tot
Tf
f
CO
CO
co-^coco
co
T
mm
Pup enImagines weiter Imagine schlüpfen geschi.
47
3 3
—
CO
OCN
cocs
Ovt>
CSCS
CO
CO
geschi.
Pup e
o
48
1
U
r^ oooo oo oo oo ov es
CSCSCSCSCSCSCSCS
tot
O
CO
I
O
tot
oooo
00
cscs
cscs
CS
1
^irAininininvûrs
t**
CSCSCSCSCSCSCSCS
es
vom
cscs
43
mm
cscs
mvo
cscs
CSCOvOt-s
—
r*>
geschi.
Puope Pup enImago weiter Imagines chlüpfen
Bem rk.
1
a
O
tot
7
40
1
cs^-ct-co-'l-^rpmvo
CSCSCSCSCSCSCSCS
vO
Ol T >« TT UO vO vO
CS
m,f
cscs
cscs
es es es
vo
r»r-s
r>.oo
OvOvO
Pup enweiter Imagines chlüpfen
Bem rk.
2
O
— es es co
co ** m vo
CSCSCSCSCSCSCSCS
tot total
Datum Begin Bei.der
geschi.
Imagines chlüpfen
Pup eImagoWeiter
U
CT<0O
CS
—
Bem rk.
16.5
3 3
1
U
1
W
geschi.
P.
O
SS
CSCS
Pup e Pup enheschl. weiter schl. tot
Im.
Bem rk.
tot
£s
geschi.
coco
•«r
CSCSCSCSCSCSCSCS
tot
O
ES
CS CS CO
es
vO
uo->icscs
cscs
CS CS CS
es
cscs
cscs
COCO CO
—
csco
•tj-u-i
\or-<»
co-t-m
—
•
•
—
CS
— —
CS CS CS
vO vO v© vO vO vO vO vO
fA^iAvOrsoOONO
CSCSCSCSCSCSCSCO
geschi.
15
23
24
24
13
14
15
16
1
24
12
I.
1
23
24
10
11
1
8
9
•c
24
25
23
24
24
23
24
24
24
7
6
egaT
epmaL
.pmeT
1
2
3
4
5
24
latoT
20
.tfpülhcseg
senigamI
)III(
)VI(
neppuP .tiew
.eppuP
tftpfüplühlchscesgeg senigamI
treirapärp
treirapärp ..llppmmeexxEE
.lgät
15"
1
10
tot
evraL
etoT
tlezsgnulhartseB
uanaH
gnukriW
300
29
29
29
29
29
30
29
29
29
30
29
29
30
29
•c
: 30
15"
.pmeT
dnatsbA
net eloivartlu
S
1
1
1
I.
latoT
22
.tfpülhcseg
senigamI
cm
)VI(
)III(
.lgät
.eppuP
;
X=
von
bis
mit
tretifrpaüplährcpseg senigamI
treirapärp ..llppmmeexxEE
tiezsgnulhartseB
sethciL
: U. V.
4
25
25
26
25
26
26
27
27
10
"C
24
24
25
26
26
A.
À
.vraL
eloT
nessikzieH
4662
nevraL
auf
=
26
50
30"
gen
n
.III
.pmeT
0024
sin epilucam
des
.tfpülhcseg
latoT
senigamI
.
e
d
e
i
h
c
s
r
e
t
n
u
u.
in
dnis
.xatot
.vraL
d.
z.
2 T.
;
T.
sie
evitisop ekrats
gitfärk
.nneevprpauLP
-nessörG ekratS .tdnaweg
.geweB
rhes
-ohP
Nr.
¬iez
.lgät
.rav
Die
10
38
eto T
gnulhartseB
ELLEBAT
.eaessem
nehcsimehc-hcsinagrO sin epileutcrahmüfeghcrud dirolhcirtnomitnA
nnak
mit
enie
tteF
enagrO
¬tteF
reprök
-oneO
netyc
nrreH
Die
eniek
.gni
,ssiew
ebraF
.hcier
rhes
nleguk
s.
auf
P.
39,
und
A)
a.
da
d.
nur
-tarU
*
von
die
i. d.
.hrov
ellat
b.
am
nefpor
llarp
¬lhaz
¬tteF
«
thcin
.mehe
esylanA .lpid
.ehcälfreborepröK
von
.rav
gut
von
d.
B)
den
39,
mit
in
,tsöleg
ginteflälreKz
.bbA(
dnalierF
ich
hcan
nohcs
eaessem
nelleZ
¬tteF
¬tteF
nevraL
IV.
auf
bis
«
.ned
,hcil
»
;
in
,llarp
gineslslaeMz
torarf.nbIbA(
mit
der
auf
der
an
den
»
39,
C)
und
nur
in
¬räps
neggiengeawd
.bbA(
.rav
der
Aus
Der
len
bei
Die
dem
Im
die
den
rhes
thcin
¬tteF
tteF
.nednah ¬nahrov
:
:
P.
der
mit
gissam hcielg
nelleZ neznag
ressorg
egneM
39,
;
Nr.
D)
thcin
tteF
eaessem
*
hcan
.nedrew
)muidatsnevraL
.VI(
bis
,nev
ebraF
.ssiew
,gnudlhicbislunAuärb elamroN ,gnudlhiscbriseludnAnuoärsbebelamroN
eirehpireP
bis
sawte
gnun
ngnunberdoiäioFtnkiaetehorcrbsariCapFyt tiehn,ersaebwthbcAis nednaB hcodejhcodejekndnenadudrfrueewBv
tnetsissA s,erleenhsphocnOA
.dnawhcsertegviezneeddniuonkietSornaeCginie ecirnPe-srroalChcseg
.nerupS
gtnhuctiireeLB( hcir.üeZditoäntiitsorreavCinU tutitsnI
murtkeps noitprosbA
.forP
tetual
.)muidatsnevraL
nevitagen neretztel
, ) r erraK
.II(
¬leZ
un¬
wie
¬raL
ssorg
el atsirktarU
ebraF
.ehcälfrebo
regnireg
netllhhaartzsneAb ¬¬trpeuparhöKeretztel ,shsciielwhcäs
hcielg rhäfeg hcilnuärb
¬hedsuA elamron negieZ
nelleZ nenleznie
.tllüfeg nellat .)hecbirlahFcier tsöleg
¬sirK
in
die
dnis
39 a
;
¬rov
tlhef
.bbA(
ELLEBAT
torarfnI
ssorg
nefporetfteeonFtürsgtlnleemhg(iP n-.sülirhrgckältslaterphUuah.g,ihbcrialfrräepksco ,.hncüirlgrlälpesh ,.hrecelilhlaphticsriieerprk
.tgfipdelnmäuatrtshslcisroekvetgfaroUntesltlneezmtgtieFP ,tedlmirboefgnsueafporT enfleogtusk,tttnteeedmlFgibiePgsu.,athlciülfhecigenra ef ,ohntcesiltllnheeczimtegtriePF .tllüfeg
t eloivartlU
tiehleknuD
,sin eptileulcoaimvarstellUehponAgnulhartseB etatluseR
thcilnennoS
BIOLOGIE
der grössten
Werten
men
UND
VERBREITUNG
EINIGER
101
STECHMÜCKENARTEN
Strahlungsintensität bei 2664 bis 2536 Â und den
von 2378 und 4200 Â zur Verfügung 1.
extre¬
Völlig in Dunkelheit aufbewahrte Eilarven (je 50 Stück) wurden
bestrahlt.
ersten Schlüpftag an täglich während 15", resp. 30
Eine weitere Zucht wurde täglich 15" bestrahlt, ausserdem aber
vom
noch mit einem Heizkissen auf ca. 29° C erwärmt.
Aus dem Versuchs-Protokoll (Tabelle Nr. 38) ist
dass eine Larvensterblichkeit von
zu
entnehmen,
% bei \5" tägl. Bestrahlung
% bei 15" tägl. Bestrahlung und Heizkisesn
48 % bei 30" tägl. Bestrahlung
60
56
resultierte. Es kann daraus geschlossen werden, dass die doppelte
Bestrahlung von je 15" die Larvensterblichkeit von 60 % auf 48 %
reduzierte. Die mit dem Heizkissen versehene Zucht nahm eine
Mittelstellung ein, wobei die schnellere Entwicklungszeit wohl auf
die Wärme (eventuell Infrarot-Bestrahlung) zurückzuführen ist.
Anopheles maculipennis var. messeae
Bestrahlung täglich 12 Std. mit infrarotem Licht
n
Tabelle Nr. 39 b
50
=
Tage
°C
1
2
3
4
5
6
7
8
9
10
11
12
24
24
23
22
21
22
23
23
23
24
24
24
24
23
24
24
13
14
15
16
Tote Larven
Bemerkungen
8
10
12
IV. Larve
13
1. Puppe
Zwei Imagines
16
geschlüpft
18
Total 32 Imagines geschlüpft
1
Für die
Benützung der U.-V.-Lampe spreche
ich
Herrn Prof.
Pharmazeutisches Institut der ETH meinen herzlichsten Dank
aus.
Dr. Büchi,
102
WILLI
BÜTTIKER
Durch eine Vergleichsreihe mit steigenden Bestrahlungszeiten hätte
das Bestrahlungs-Optimum ermittelt werden können ; diese Unter¬
suchungen mussten aber aus Zeitmangel auf später verschoben
werden.
Tabelle Nr. 39 a enthält die Beschreibung der Fettkörper,
Oenocyten und Pigmentstoffe, die mit den Larven der InfrarotBestrahlung und der Kontrollzucht verglichen wurden.
Aus dem vorliegenden Versuche geht hervor, dass eine richtig
dosierte U. V.-Bestrahlung durchaus fähig ist, die Photosynthese
aufrecht zu erhalten. Verglichen mit den Resultaten der InfrarotBestrahlung, ergibt sich eine intensivere physikalisch-chemische
Wirkung der U. V.-Strahlen.
Versuch
e ;
Infrarot-Bestrahlung.
Für die Infrarot-Untersuchungen kamen wiederum 50 Eilarven
A. maculipennis var messeae zur Anwendung. Die Versuchs¬
anordnung bestand aus einem zylindrischen Blechgefäss von 10 cm
Durchmesser und 15 cm Höhe. Der Infrafilter (Agfa Nr. 85, schwarz,
Absorption bis 755 uu) wurde in den Blechdeckel eingebaut, so
dass die Larven nur während der Belichtungszeit von 12 Stunden
unter dem Einfiuss des infraroten Lichtes standen ; die gesamte
übrige Strahlung konnte auf diese Weise ausgeschaltet werden.
Aus der Tabelle Nr. 39 b sind die Resultate im einzelnen zu
entnehmen. Durchschnittlich herrschte eine Temperatur von 23° C ;
die Larvensterblichkeit betrug 36 %, wobei zu bemerken ist, dass die
Zuchtbedingungen nicht ideal waren. Das Zuchtgefäss war für
50 Larven (besonders des III. und IV. Stadiums) zu klein, so dass
die Wirkungen des « Crowdings » ebenfalls zu berücksichtigen sind
von
(Ulmann, 89).
Es ergeben sich folgende Resultate :
Eine Bestrahlung von infraroten Wellen von täglich 12 Stunden
genügte bei den beschriebenen Verhältnissen, dass sich die Larven
normal entwickeln konnten. Verglichen mit den Versuchen der
Ultraviolett-Bestrahlung, ergibt sich eine viel kleinere Wirksam¬
keit des langwelligen Lichtes (wie dies auch bei andern Insekten
durch verschiedene Autoren festgestellt wurde), da eine U. V.¬
Bestrahlung von 30" der Infrarotzeit von 12 Stunden täglich gegen¬
überstand und in grossen Zügen den gleichen Effekt zeigte.
Larvensterblichkeit bei U. V.
»
»
bei Infrarot
Versuch f
:
Aufzucht
Wiederum
von
wurden
=
=
48
36
%/30 Sek. 1 Bestrahlung
%/12 Std. |
täglich
Larven bei völliger Dunkelheit.
50
frischgeschlüpfte
völlig unbelichteten Gefäss aufgezogen. Bis
am
Eilarven in einem
Ende des II. Larven-
BIOLOGIE UND VERBREITUNG EINIGER
STECHMÜCKENARTEN
103
stadiums blieben einige Larven am Leben, die andern Starben
schon vorher ab. Die Lebenduntersuchung zeigte grosse Unter¬
schiede
zu den bestrahlten Exemplaren. In den Zellen des Fett¬
körpers fehlte vor allem das Fett vollständig. Somit fehlten auch
die Pigmentstoffe, die normalerweise darin gelöst sind. Dafür waren
Uratkristalle in ausserordentlicher Menge vorhanden. Die Fettzellen
des fehlenden Fettes stark eingeschrumpft. Die Larven¬
sterblichkeit betrug nach ca. 11 Tagen 100 %. Die Resultate werden
in der Tabelle Nr. 37 mit den Ergebnissen der bestrahlten Larven
waren wegen
verglichen (Abb. 39, D).
Abb. 39.
Anopheles maculipennis var. messeae, IV. Larven, ca. 800 fach Orig. Die
Wirkung verschiedener Wellenlängen auf den Fettkörper der Larven.
A, normale
Sonnenbeleuchtung, Freiland.
B, Infrarote Strahlen.
C, Ultraviolette Strahlen.
D, Kontrollversuch ; in völliger Dunkelheit aufgezogene Larve.
I, Fettropfen.
2, Oenocyten.
3, Uratkristall.
4, Zellgrenze.
5, Zellkern der Fettzelle.
—
—
—
—
—
—
—
—
—
—
104
WILLI
BÜTTIKER
B. Phototaxis
Um das Verhalten der Tiere beurteilen zu können, ist es nötig,
zwischen Organismen zu unterscheiden, die nur Helligkeitsempfin¬
dungen haben, und andere, die bildhaft sehen. Die ersten Ent¬
wicklungsstadien unterscheiden in der Regel nur zwischen hell und
dunkel ; die Imagines dagegen haben
mit wenigen Ausnahmen
ein bildliches Sehvermögen. Es wurde deshalb angenommen, dassalle diese Organismen mechanisch auf Helligkeitswerte reagieren,
entweder positiv oder negativ. Nach neueren Untersuchungen dagegen
scheint es wahrscheinlicher, dass verschiedene Umgebungseinflüsse
—
—
einer Modifikation der Lichtkreation führen.
Die Untersuchungen von Lutz (45) deuten darauf hin, dassInsekten auf kurze Wellen besonders empfindlich sind und im ultra¬
violetten Bereich noch sehen können.
Äussere physikalische Reizwirkungen werden u. a. durch die
sogenannten Taxien ausgedrückt. Die Topotaxis führt zur Orientie¬
rung in eine bestimmte Reizrichtung. Dabei dreht sich das Insekt
durch Wendereaktionen in einem Reizfeld in eine bestimmte Rich¬
tung zur Reizquelle. Bei der Fortbewegung entsteht auf diese Weise
meist eine gerichtete Bewegung, die entweder zur Reizquelle hin
(positive Topotaxis), oder von ihr weg (negative Topotaxis) führt.
Dabei werden Störungen in der Bewegungsrichtung infolge von Ver¬
schiebungen oder durch Kompensationsbewegungen ausgeglichen.
Die Einstellungsziele können verschiedenster Natur sein, je nach der
Reizmodalität, welche sie anwenden (Farben, Formen, Düfte usw.).
Ihr Reizwert als Ziel kann angeboren sein oder aber durch Erfahrung;
erhalten werden.
Man kann im Folgenden vier Kategorien unterscheiden (ElDMANN,
zu
a)
b)
c)
d)
Tropotaxis
Telotaxis
Menotaxis
Mnemotaxis
(Symmetrieeinstellung)
(Zieleinstellung,
gradlinig
Lichtquelle, als Phototelotaxis
Insekten)
(Kompensationsbewegungen)
zur
bei
(Gedächtnisorientierung).
Literatur : Eine Zusammenfassung der wichtigsten Arbeiten über
den Einfluss des Lichtes auf Insekten, speziell auch über die Photo¬
taxis, ist bei CHAPMAN (10) zu finden, sodass sich hier eine Auf¬
zählung erübrigt. Angaben über Culiciden sind darin jedoch keine
vorhanden.
Im nachfolgenden Abschnitt soll besonders die Phototaxis einer
näheren Prüfung unterzogen werden. Zur Untersuchung gelangten
Larven, Puppen und Imagines von Culex pipiens und Anopheles
maculipennis
var.
messeae.
BIOLOGIE
UND VERBREITUNG
EINIGER
STECHMÜCKENARTEN
105
Methode
Larven und Puppen : Ein Wasserbehälter von 1,40 m Länge,
20 cm Breite und 10 cm Tiefe wurde mit Wasser gefüllt, worin sich
die zu untersuchenden Larven und Puppen frei bewegen konnten.
Im senkrechten Abstand von 25 cm wurden sechs elektrische Glüh¬
birnen mit zunehmender Stärke stufenweise montiert.
Auf diese Weise konnte ein Gefälle der Lichtintensität erreicht
werden. Zur Anwendung kamen Glühbirnen von 16,5, 25, 40, 60,
75 und 100 Watt. Zur Kontrolle wurde das eine Behälterende von
der direkten Lichteinwirkung abgeschirmt. Die Versuchstiere gelang¬
nach 24 Stunden zur Auszählung.
Imagines. Für die flugfähigen Imagines kam ein Holzgefäss von
1.40 m Länge, 10 cm Breite und 8 cm Höhe zur Anwendung. Als
oberer Abschluss diente eine Glasplatte, welche das ungehinderte
Beobachten ermöglichte. Als Lichtquelle diente eine elektrische Glüh¬
birne von 40 Watt, im Abstand von 20 cm ; ihr Standort war an
einem Ende des Holzgefässes, so dass eine gerichtete Lichtstrahlung
ten
entstand.
Resultate
Culex pipiens.
Larven (IV. Stadium); (Tabelle Nr. 40). Das Minimum der Larven¬
zahl liegt deutlich bei der niedrigsten Lichtintensität, d. h. beim
abgeschirmten Bereich der « Lichtorgel ». Die Anzahl der Versuchs¬
tiere nimmt ungefähr linear zu, um am Schluss durch leichtes Anstei¬
gen ein Maximum von 34 % aller Larven zu erreichen (vergl. Tabelle
Nr. 40). Durch andere, ähnliche Versuche konnte die positive Photo¬
taxis der Larven bestätigt werden.
Phototaxis der Larven
n
=
von
Culex pipiens
Tabelle Nr. 40
70
Datum
Dunkelheit
16,5 W
25 W
40 W
60W
75 W
100W
1.9.47
2.9.47
10
2
10
5
10
7
10
9
10
11
10
14
10
22
Puppen (Tabelle Nr.41). Auf dieselbe Weise wurden die Puppen
auf ihr phototaktisches Verhalten geprüft. Aus den beobachteten
Zahlen ergibt sich ein vermehrtes Ansteigen der Puppenzahl gegen
die höhere Lichtintensität. Die positive Phototaxis der Puppen ist
0081
74.9.42
0081
74.9.32
mutaD
,74.9.22 :nnigeB
BB
gnuthcirthciL
74.2.81
74.2.71
mutaD
evruK
ereitshcusreV
AA
18
19
34
3
72%
6,2
%
.
41%
h
tiehleknuD
00.81
n
n
1
4
5.61
n
2
7
=
2
4,2
%
4%
9%
W
4.2
%
8,4
3
der
n
4
2
50
sixatotohP
1
:
1
5
25
n
3
2
nevraL
$
4%
11%
W
von
4.2
%
6.2
4
A.
1
2
n
1
3
5
40
%
4.2
2.1
%
4%
6.5
W
n
3
1
6
%
2
3
6.2
2,1
sin epilucam
0080
48
%
8%
W
6.5
60
n
2
0
.rav
7,07
0091
tieZ
=
0
%
4,2
7
1
8
0
2
n
:
75
0
4%
W
17%
:
%
4,2
'8
17°
0
3
n
C
9
1
4
%
r.
6.2
0
%
100
4%
44
2.1
9%
W
Nr.
1
42
%
10
Nr.
n
24
F.
ellebaT
eaessem
rutatrieepkmgeitthschuceufstrfeuVL
001—09
6,41
74.2.81/71
sixatotohP mutadshcusreV
von
0,05
ellebaT
ineiptp xeluC
BIOLOGIE
UND
VERBREITUNG EINIGER
Phototaxis der Puppen
n
=
von
STECHMÜCKENARTEN
107
Culex pipiens
50 Stück
Tabelle Nr. 41
Datum
Dunkelheit
16,5 W
25 W
40 W
60 W
75 W
100W
1.9.47
2.9.47
7
0
7
7
1
0
7
1
7
4
7
14
8
28
mit dem Maximum von 57 % noch ausgeprägter als bei den Larven
mit maximal 33 %.
Imagines. Überwinterte Weibchen (Tabelle Nr. 42). Sämtliche
Versuchstiere wurden im Abteil 10 zu Beginn des Versuches ein¬
gesetzt. Schon nach wenigen Minuten verteilten sie sich in die ein¬
zelnen Abteile. Nach 13 Stunden befanden sich ca. 71 % am licht¬
nahen Ende der Versuchsapparatur.
Imagines. Einwinternde Weibchen : Beobachtungen an der Uberwinterungsplätzen zeigten eine deutliche negative Phototaxis, indem
die sich einwinternden Weibchen stets in dunkle (und feuchte)
Räume sich begaben. Dies geschah hauptsächlich in den Monaten
September-Oktober.
Folgender Laborversuch erläutert die Verhältnisse in dieser
Beziehung. Die Versuchsbedingungen waren mit 90—100 % r. F.
und 19° C ungefähr konstant.
n
=
50
Versuchsdatum: 17.11.47.
Tabelle Nr. 43
Zeit
1
2
3
4
5
6
7
8
9
to
0800
1000
1200
6
2
2
3
1
0
5
3
I
6
2
0
5
2
6
1
4
8
2
7
7
4
5
9
15
7
11
25
Lichtrichtung
1
0
_>.
Sommermücken. Nüchterne Weibchen: Als eigentliche Dämmerungs¬
und Nachttiere zeigten die Weibchen während den Abendstunden
ihre grösste Aktivität. Bei hohen Lichtintensitäten (am Tage) sind
sie negativ, bei niedern (in der Dämmerung) aber positiv photo¬
taktisch. Freilandbeobachtungen zeigten während des Sommers regel¬
mässig diese Eigenschaft.
evruK
AA
BB
gnuthcirthciL
28
74.3.21
74.3.11
0080
0302
mutaD
mutaD
0081
74.9.52
ereitshcusreV
tieZ
=
16
17
n
?
12
0,75
1
40%
%
6,06
tiehleknuD
30
3
1
2
n
3,6
%
3
5,61
1
2
n
W
3,6
%
7,2
3
10%
sixatotohP
=
:
3
0
0
n
25
4
0
0
%
W
10%
neppuP
n
7,01
2
1
n
74.3.21/11 mutadshcusreV
retretniwrebü sixatotohP
A.
7,2
%
3,6
5
4
40
nehcbieW
1
2
n
von
3,6
%
7,2
6
A.
13%
W
sin epilucam
von
3
I
0
n
60
%
0
3,6
7
W
10%
sin epilucam
.rav
der
0
1
n
2
0
%
3,6
8
75
1
0
n
7%
W
3,6
0
%
9
3
3
4
%
10
46
45
10%
w
Nr.
100
Nr.
n
7,01
2,41
ELLEBAT
ellebaT
eaessem
BIOLOGIE
UND
VERBREITUNG
STECHMÜCKENARTEN
EINIGER
Sommermücken. Blutgesogene Weibchen
109
Aus Freiland- und Labor¬
dass sie während der
in dunklen Räumen verbleiben.
:
beobachtungen geht übereinstimmend hervor,
Praeovipositionszeit
stets
Anopheles maculipennis
var.
messeae.
Larven (IV. Stadium) ; (Tabelle Nr. 44). Die wiederholt durch¬
geführten Experimente an Larven von messeae zeigten jeweils dasselbe
Bild der Lichtflucht. Tabelle Nr. 44 veranschaulicht die vorgefun¬
denen Verhältnisse. Zu Beginn des Versuchs wurden 46 Larven
auf das Gefäss gleichmässig verteilt. Bei der ersten Kontrolle nach
ca. 6 Stunden fanden sich am abgeschirmten Ende 19 Larven (=41 %)
ein, während der lichtexponierte Teil nur geringe Prozentwerte
aufwies.
Nach 24 Stunden ergab die Kontrolle im Prinzip ein gleiches
Bild. Von den 25 noch nicht zur Verpuppung übergegangenen
Larven befanden sich 18 Exemplare (= 72 %) an der unbelichteten
Stelle, während sich der Rest der Tiere ziemlich gleichmässig auf
den
übrigen Teil der Versuchseinrichtung verteilte.
Das Resultat ist schwierig zu deuten, da Anopheles maculipennis
var. messeae nach andern durchgeführten Versuchen grössere An¬
sprüche an die Belichtung als zum Beispiel Culex pipiens stellte. Zu
erwarten wäre*also eine noch ausgeprägtere positive Phototaxis. Für
das gegenteilige Verhalten könnte möglicherweise die Zusammen¬
setzung des verwendeten Lampen-Lichtes verantwortlich gemacht
werden. Es wäre interessant, diese Verhältnisse noch einer ein¬
gehenderen Prüfung zu unterziehen.
Puppen (Tabelle Nr. 45). Die negative Phototaxis kommt auch
Entwicklungsstadium zum Ausdruck. 40 % der frisch
geschlüpften Puppen befanden sich im unbelichteten Bereich des
in diesem
Wasserbehälters.
Imagines, überwinterte Weibchen : Aus den Beobachtungen geht
deutlich hervor, dass überwinterte Weibchen stark positiv photo¬
taktisch sind. Die von ungefähr gleichen Versuchsbedingungen
erhaltenen Werte sind in der Tabelle Nr. 46 detailliert zusammen¬
gestellt.
C. Strahlung
und
Eiablage
Die Mehrzahl der Culiciden zeigt die grösste Aktivität in der
Morgen- und Abenddämmerung und während der Nacht. Viele der
wichtigsten Lebensfunktionen gelangen in dieser Zeit zur Ausfüh¬
rung, wie Blutaufnahme, Kopulation, Eiablage und Ortsveränderung.
Manche weitere Tätigkeit ist ebenfalls mehr oder weniger vom
Helligkeitsgrad der Umgebung abhängig.
110
WILLI
BUTTIKER
BRUES (100) veröffentlicht einige Beobachtungen an Aedes aegypti,
wonach eine Eireifung nur bei intermittierender Belichtung der
Weibchen
möglich
sei. Andere
Literaturangaben sind mir über dieses
Problem nicht bekannt geworden.
Methode
Die vorliegenden Versuche an A. maculipennis var. messeae wurden
speziell bezüglich dem Einfluss des sichtbaren Lichtes auf die lege¬
reifen Weibchen durchgeführt.
Bei der Anordnung der Versuche wurden 3 Gruppen zu je
20 Weibchen gebildet, die verschiedener Belichtung ausgesetzt
waren. Das Material war von gleicher Herkunft und die Ovarien
waren bei allen Weibchen im gleichen Reifezustand. Für die Belich¬
von 25 Watt im Abstand von
10 cm benützt. Sämtliche Experimente wurden im Thermostat bei
22° C durchgeführt. Die täglichen Belichtungszeiten waren bei den
einzelnen Gruppen wie folgt :
tung wurde eine elektrische Glühbirne
a)
b)
c)
24 Stunden pro Tag (Tabelle Nr. 47, Kol. 1)
12 Stunden pro Tag (Tabelle Nr. 47, Kol. 2)
0 Stunden pro Tag (Tabelle Nr. 47, Kol. 3)
Die Weibchen wurden in Präparategläschen von 10
2 cm Durchmesser während der Beobachtungszeit
die Luftfeuchtigkeit annähernd 100 % betrug.
cm Länge und
gehalten, wobei
Resultate
Die detaillierten Versuchsresultate der Belichtungsversuche sind
in der Tabelle Nr. 47 aufgeführt. Kolonne 1 enthält Angaben über
die Weibchen, welche während täglich 24 Std. belichtet wurden. Es
erfolgte vom 29. Juli bis zum 13. Aug. 1947 keine Eiablage; die
Weibchen starben bis zu diesem Datum, also innerhalb von 16 Tagen,
ab. In der Kolonne 2 sind die Angaben über Weibchen, die je 12 Stun¬
den abwechselnd dem Licht und der Dunkelheit ausgesetzt wurden.
Nach 4 Tagen wurden die ersten sechs Eigelege vorgefunden. Bei
der nächstfolgenden Dunkelperiode kamen weitere 10 Gelege zur
Ablage, und die letzten 2 .Weibchen legten ihre Eier erst nach
6 Tagen 1. Insgesamt starben 2 Weibchen. Kolonne 3 enthält die
Beobachtungsresultate der in völliger Dunkelheit gehaltenen Weib¬
chen. Nach 4 Tagen wurde die überwiegende Mehrzahl der Gelege
nur
1
Wie weit der Barometerdruck
schwer entscheiden.
an
dieser
Verzögerung mitbeteiligt ist,
lässt sich
ellA
8
Im
zu
2
;
.tot
.tot
egalba
remmI
neznag
den
2
die
,nebah
enho
.egalba
tulB
nehc
tats
remmI
in
17
eniek
?
Ei¬
Ei¬
¬bieW
hcon
reiE
dnis
hcon
nohcs
eniek
tgeleg
nohcs
.nebrotseg nehcbieW
.nebrotseg
,nehcbieW eretiew
.
n
e
b
r
o
t
s
e
g
nehcbieW
.thcarbrev
¬omrehT .etnemirepxE
.)sdnenbnai(gneeBgos,esgdnebA
der
8.5/.4
8.4/.3
8.3/.2
7.92
7.92
negnukremeB
nehcbieW
ennoloK
tethcileb gidnäts
1
$
10
6
tulB
mutaD
smnepilucam
?
.tnnak
.tot
.negelegiE
.tot
hcoN
2
1
1
thcin dnurG
be¬
mit
nehcnbeihecWbieW
nehcbieW .tgelegba
in
der
ge¬
egeleG elamron nebah
nehcbieW eretiew
.tedlib
egelnegeihEcbeileaWmron
nebah
2
negoseg
nehc
tats
.t.hectanrebmrievrepxE
¬omrehT
der
¬bieW
der
3.8
in
auf
$
8.3/.2
7.92
7.92
ennoloK
.eaessem
.rav
.)sdnenbnai(geB ,sdnebA
negnukremeB
gnuthcileB ednereit imretni
ssulfniE
A.
1
19
den
der
.egeleG
.ned
.etnem
tulB
mutaD
gnuthcileB
von
die
3
Nr.
in
¬rev
¬nufegrov egeleG
tatsomreh.Tthcarb
¬irenpexEhcbieW
.)sdneba( negosengnigeB
negnukremeB
tiehleknuD regillöv
ennoloK
ellebaT
egalbaiE
8.31
8.31/.21
8.11/.01
8.01/.9
8.9/.8
7.92
7.92
mutaD
47
112
WILLI
BÜTTIKER
den Weibchen abgegeben (19 Stück) ; kurze Zeit darauf erfolgte
Ablage des letzten Geleges. Die Sterblichkeit betrug 0 %.
Zusammenfassend kann gesagt werden, dass bei völliger Dunkel¬
heit die Eiablage am besten ist, während bei andauernder Einwir¬
kung des Lichtes die Legetätigkeit verhindert wird. Die abwechselnde
Belichtung hat eine kleine Verzögerung des Hauptablagetermins zur
Folge.
von
die
4. Der Einfluss des Luftdruckes
Die
Kraft, welche wir als Druck des umgebenden Mediums
ist das Gewicht der Atmosphäre, des Wassers oder des
Bodens auf das Tier. Der atmosphärische Druck wechselt mit den
allgemeinen atmosphärischen Verhältnissen an einem bestimmten
Ort. Die grössten Druckunterschiede werden aber durch Höhen¬
differenzen hervorgerufen. Der Luftdiuck kann mit Hilfe des Queck¬
silberbarometers gemessen werden, doch ist dabei die jeweilige
Umgebungstemperatur zu berücksichtigen ; Barometer mit luftleerer
messen,
Dose sind davon weitgehend unabhängig.
Es ist schwierig zu unterscheiden zwischen dem direkten Ein¬
fluss des Druckes und der Wirkung anderer mitbeteiligter Faktoren.
Durch die Druckverminderung wird z. B. auch der Gasdruck des
Atmungssystems verändert ; die meisten Untersuchungen zeigen
die Beeinflussbarkeit der Entwicklung durch Druckveränderungen
der
Atmungsluft (CHAPMAN, 10).
Greenwood (28) experimentierte mit Larven von Cossus cossus,
indem er sie dem atmosphärischen Druck von +25 und —30 mm
während 20 Minuten im ersten und 2 Stunden 40 Minuten im zwei¬
ten Falle aussetzte. Diese Larven verpuppten sich normal, nachdem
sie in die früheren Verhältnisse zurückgeführt wurden. Der Autor
schliesst, dass die Fähigkeit, einen grossen Druckwechsel zu über¬
dauern, mit der Tracheenatmung zusammenhängt.
Versuche mit Fliegen, Schmetterlingen, Käfern u. a. ergaben,
dass selbst ein Luftdruck von Vio ooo mm während 90 Sekunden
ertragen werden kann. Beobachtungen an Corethra, Chironomus u. a.
gehen dahin, dass mehrere Atmosphären (at) ertragen werden kön¬
nen, und dass auch ein rascher Druckwechsel auf das Schlüpfen
keinen Einfluss hat.
Es gibt einige Anhaltspunkte, dass sich die Lebensäusserungen
vieler Insekten unter barometrischen Depressionszuständen steigern.
Verschiedene Autoren weisen auf die besonders grosse Aktivität
blutsaugender Dipteren vor Gewitterregen hin (WELLINGTON, 91).
Genaue Beobachtungen und Experimente wurden über den
Einfluss der Luftdruckabnahme auf die Länge des Puppenstadiums
und auf das Schlüpfen der Puppen von Gross- und Kleinschmetter¬
lingen durchgeführt. Es geht daraus hervor, dass ein gleichmässiger,
BIOLOGIE
UND
VERBREITUNG
EINIGER
STECHMÜCKENARTEN
113
normaler Luftdruck, der während des ganzen Puppenstadiums wirkt,
dieses verlängert, während ein anhaltend verminderter Luftdruck
dasselbe beträchlich verkürzt (UvAROV, 90). So dauert bei Pieris
rapae das Puppenstadium bei einem Druck von 710—728 mm nur
9 Tage, während es normalerweise 15 Tage beträgt (760 mm). Die
Verkürzung der Entwicklungszeit beträgt demnach 40 %.
Das Schlüpfen der Imagines bei Lepidopteren findet in den
meisten Fällen nur bei fallendem Barometerdruck statt. Eine Druck¬
verminderung von 1 mm soll schon ausreichen, das Auskommen eines
schlüpfreifen Falters zu erzielen. Die Wirkung des Luftdruckes soll
nach PlCTET (67) eine rein mechanische sein : innerhalb der Puppen¬
hülle ist das fertig ausgebildete Insekt solange unfähig diese zu zer¬
sprengen, bis eine Druckdifferenz einen relativ höhern Druck der
Körperflüssigkeit schafft.
Über den Einfluss des Barometerdruckes auf die Eiablage bei
Dipteren, spec, bei Culiciden, liegen ebenfalls Beobachtungen vor.
(Marshall, 46). Übereinstimmend geht daraus hervor, dass bei
Depressionen die Eiablage besonders zahlreich erfolgt.
Parmann (66) berichtet, dass verschiedene Fliegenarten in einen
Zustand von Nervosität geraten, sobald der Barometer druck zu
fallen beginnt. Er zitiert das Beispiel von Stomoxys calcitrans und
Musca domestica, welche vor einem Sturm in grosser Menge vor¬
handen waren, während nachher keine mehr beobachtet werden konn¬
ten. Allerdings ist damit nicht bewiesen, dass der Luftdruck die
alleinige Ursache dieses Verhaltens war.
A. Luftdruck
und
Entwicklungsdauer
Bei den durchgeführten Experimenten sollte vor allem der Ein¬
fluss des Luftdruckwechsels auf die Entwicklung der Eier, Larven
und Puppen festgestellt werden. Meines Wissens existieren keine
Untersuchungen über den Einfluss des Luftdruckes auf die Ent¬
wicklungsgeschwindigkeit der Jugendstadien bei Culiciden.
Im vorliegenden Kapitel werden die einzelnen Stadien von
Culex pipiens und z. T. von Anopheles maculipennis var. messeae
«iner näheren Prüfung unterzogen.
Methode
Die Versuchsapparatur bestand aus zwei Glas-Rezipienten,
«inem Barographen und einer Wasserstrahlpumpe, welche eine
Depression bis zu 40—50 mm Hg. resp. 680 mm erzeugen konnte.
Die Versuchsanordnung für Imagines ist aus Abb. 40 ersichtlich.
Für die Experimente an den Larven und Puppen wurde die
Anordnung etwas modifiziert, indem an das T-Rohr (Abb. 40, 3)
ein Verbindungsschlauch zu einer Flasche geführt wurde. Dieser
s
114
WILLI
BUTTIKER
Q)
®
Abb. 40.
1, Gefäss für Imagines.
Versuchsapparatur für Luftdruckexperimente.
4, Zuleitung
3, Zweigstück des T-Rohres.
2, Gefäss für Larven und Puppen.
zur Wasserstrabipumpe.
5, Rezipient I.
6, Barograph.
7, Rezipient II.
—
—
—
—
—
—
—
—
Glasbehälter befand sich im Thermostaten (22°) und enthielt die
Larven und Puppen.
Die Druckschwankungen im Verlaufe des Versuches konnten
dem Papierstreifen der Barographentrommel entnommen werden.
Resultate
Eier
von
A. maculipennis
var. messeae.
Zwei Eigelege zu je 150 Eiern kamen zur Untersuchung. Das
eine wurde einem Unterdruck von 25—30 mm ausgesetzt, während
das andere als Kontrolle bei normalem, lokalen Barometerdruck
(Zürich ca. 730 mm) zur Beobachtung kam. Es resultierten in beiden
Fällen die gleichen Entwicklungszeiten, wie dies aus nachfolgender
Tabelle Nr. 48 hervorgeht, sodass von einem messbaren Einfluss auf
die Entwicklungszeit nicht gesprochen werden kann. Die Entwick¬
lungsdauer von 2,5 bis 3 Tagen entspricht dem normalen Wert bei
einer Temperatur von 22° C (vergl. Abschnitt « Temperatur und
Entwicklung »).
Tabelle Nr. 48
Eier bei 25—30
(700—795
mm
mm
Depression
Hg)
2,5-3 Tage
Kontrolle
(ca 730 mm)
2,5-3 Tage
BIOLOGIE UND
Larvenstadien
von
Die bei 30
gegenüber den
VERBREITUNG
EINIGER
115
STECHMÜCKENARTEN
Culex pipiens.
Depression durchgeführten Versuche zeigten
mm
Kontrolltieren annähernd gleiche Resultate. Die
Schlüpfvorgänge waren bei allen Tieren normal. Kleinere Unter¬
schiede in der Entwicklungszeit können auch auf Differenzen in der
Ernährung zurückgeführt werden. Wie an anderer Stelle gezeigt
wird, spielt die Nahrungsquantität für die Entwicklungsdauer eine
Rolle.
Das Mass der zeitlichen Streuung in der Entwicklungszeit sämt¬
licher Larvenstadien ist zum Teil gross, so dass eigentlich nur Mit¬
telwerte verglichen werden können.
grosse
Tabelle Nr. 49
Larven bei 30
mm
(700—705
Depression
Hg)
Kontrolle (ca 730
mm
8—8,5 Tage
Puppen
von
Hg)
mm
8—8,5 Tage
Anopheles maculipennis
var.
messeae.
mm (Hg-Säule : 695—700 mm). 40 frisch¬
die nicht über 8 Std. alt waren, wurden in zwei
Gruppen zu je 20 Stück getrennt. Die erste Gruppe kam in das
Gefäss mit einem Unterdruck von ca. 25 mm, während die zweite
dem normalen Barometerstand ausgesetzt wurde. Die Versuchstem¬
peratur betrug jeweils 22° C, wobei folgende Werte resultierten
Druckdifferenz
:
30^-35
geschlüpfte Puppen,
(Tab. 50)
:
Tabelle Nr. 50
Beginn
des Versuches
:
Puppen bei 695—700
13.8.47
«?
0800
2,5 Tage
mm
?
3
Tage
Kontrolle (ca. 730 mm)
?
S
2,5 Tage
3
Tage
Die verschiedenen Entwicklungszeiten für Männchen und Weib¬
chen entsprechen den Werten der Entwicklungsversuche im Zusam¬
menhange mit der Temperatur. Die Entwicklungszeit der Männchen
ist bei den Culiciden kürzer. Der Schlüpfvorgang war in allen Fällen
normal.
116
WILLI
BUTTIKER
Verglichen mit den eingangs erwähnten Experimenten an Schmet¬
terlingen (Uvarov, 90) ergibt sich ein grosser Unterschied, da die
Culicidenpuppen im Gegensatz zum entsprechenden Stadium bei
Lepidopteren nicht
werden können.
zur
Verkürzung der Entwicklungszeit veranlasst
Druckdifferenz : 680 mm (Hg-Säule : 40—50 mm). Die Versuche
wurden mit einer modifizierten Apparatur wiederholt, indem an
Stelle des Barographen ein Quecksilbermanometer verwendet wurde.
Der Luftdruck von 40—50 mm Hg-Säule wirkte auf 20 Puppen ein ;
bei gleichen Temperaturverhältnissen, aber Normalbarometerdruck
(730 mm), standen vergleichsweise ebenfalls 20 Puppen zur Ver¬
fügung.
Es zeigten sich bei den Puppen schon nach einigen Stunden
ungewöhnliche Schwimmbewegungen im Depressionsgefäss. Nach
1,5 Tagen wiesen einige Exemplare typische Ermattungsanzeichen
auf. Während die Kontrolltiere sich normal weiter entwickelten,
starben die ersten Puppen des Unterdruckgefässes nach ca. 1 % Tagen,
die letzten bis zum Ende des dritten Tages. Sie lagen mit ausge¬
streckten Abdomen unmittelbar unter der Wasseroberfläche. Es
geht daraus hervor, dass ein Druckunterschied von ca. 680 mm eine
100-prozentige Mortalität bewirkt.
B. Luftdruck
Culex pipiens
:
und
Eiablage
Weibchen.
Druckdifferenz : 25—35 mm. Die Versuche mit C. pipiens blieben
erfolglos, da die Weibchen im Rezipienten (Abb. 40, 5) in keinem
Falle zur Eiablage schritten. Die Anzahl der befruchteten und mit
Blut vollgesogenen Weibchen betrug 25. Sie starben jeweils nach
durchschnittlich 8—9 Tagen ab, ohne die Eier vorher abgelegt zu
haben. Die Gründe zu diesem Verhalten sind mir nicht bekannt,
doch vermute ich, dass die Luftfeuchtigkeit infolge Wasserver¬
dunstung in der Brutschale zu hoch und dass der Raum des Rezipien¬
ten als künstlicher Brutort zu klein war.
Anopheles maculipennis
var. messeae ;
Weihchen.
Druckdifferenz : 25—35 mm. Die Weibchen dieser Art verhielten
gleichen Versuchsbedingungen ganz anders. 21 befruchtete
Weibchen, welche frisch Blut aufgenommen hatten, wurden in den
Rezipienten eingeschlossen und einer Depression von 25—35 mm
ausgesetzt. 12 Tiere fanden als Kontrollexemplare Verwendung.
Eine deutliche Vorverschiebung des Eiablagetermins konnte bei
den Weibchen des Unterdruckgefässes festgestellt werden (Tab. 51) :
sich bei
BIOLOGIE UND
VERBREITUNG
Blutaufnahme
am
EINIGER
117
STECHMÜCKENARTEN
30.7.47 abends oder nachts
Tabelle Nr. 51
Eier
abgelegt
Druckdifferenz 25—30
am
Im
Gegensatz
zu
C. pipiens lässt sich also A. maculipennis
den beschriebenen
unter
mm
0
12 ?
15 ?
6 S
1./2.8.47
2./3.8.47
messeae
Kontrolle 730
mm
Versuchsbedingungen
zur
var.
Eiablage
bringen.
C. Weitere Beobachtungen über
auf
Culex pipiens
:
den
den
Einfluss
des
Luftdruckes
Eiablagetermin
Weibchen.
Frühjahr 1946 fiel mir die Tatsache
um einige Tage
verspätet hatten. Die näheren Verhältnisse ergeben sich aus der
Bei den Laborzuchten im
auf, dass sich die Culex-Weibchen mit der Eiablage
Tabelle Nr. 52.
Tabelle Nr. 52
Anzahl
Weibchen
3
2
I
2
2
2
Blut gesogen
am
Eiablage
Diff. in
am
I./2.4.1946
2./3.4.1946
3./4.4.1946
4./5.4.1946
8./9.4.46
(10 Gelege)
5./6.4.1946
9./10.4.I946
15.9.1946
Tagen
7
6
5
4
3
5
Eizahl
ca.
120
200
180
200
200
250
Wirksamer Barometer¬
druck, maximal
8./9.4.1946
mit 4,9
mm
In der Nacht vom 8. auf den 9. April 1946 wurden also 10 Eigelege auf die Wasseroberfläche abgelegt. Die Differenz variiert zwi¬
schen 7 und 3 Tagen bei gleichen Temperaturverhältnissen. Die
wie dies auch aus andern Beobachtun¬
Dauer von 3 Tagen kann
als minimale Praeovipositionszeit angesehen wer¬
gen hervorgeht
den. Für A. maculipennis führt Marshall (46) in seinem CulicidenWerk eine Praeovipositionszeit von 4—10 Tagen an, welche jeweils
von den atmosphärischen Druckverhältnissen abhängig ist.
—
—
V.
UND
DIE
RASSEN
IHRE
VON
A.
VERBREITUNG
MACULIPENNIS
IN
DER
SCHWEIZ
1. Die historische Entwicklung des Rassenproblems
Sergent (78) vermutete schon 1903 gewisse Unterschiede in
Grösse und Gewohnheiten zwischen algerischen und französischen
maca/ipenms-I ndividuen. Roubaud (69) als erster hatte die Idee,
dass bei dieser Art verschiedene Rassen vorkommen. Er trennte sie
in eine zoophile und eine anthropophile Rasse und wollte morpho¬
logische Zusammenhänge (Zahl der Maxillenzähne) feststellen. Diese
Vermutungen wurden aber später widerlegt.
Praktisch zur selben Zeit folgerte Wesenberg-Lund (92), dass der
Wechsel vom weide- zum stallgefütterten Vieh in Dänemark eine
Modifikation der Gewohnheiten der Anophelen mit sich brachte.
Im selben Jahr vermutete der gleiche Autor, dass man annehmen
könne, eine Rasse vor sich zu haben, die den Menschen nicht sticht.
Falleroni (20) beobachtete 1922 an Anopheles maculipennis, dass die
Eier von verschiedenem Aussehen waren (grau und dunkel), die
Weibchen dagegen waren morphologisch nicht zu unterscheiden.
Falleroni erklärte, dass die Unterschiede bei den Eiern konstant
wären und dass sie eine Trennung der beiden Rassen ermöglichen
würden. Dies führte zur Bezeichnung A. maculipennis var. messae
Fall, (graue Eier) und A. maculipennis var. labranchiae Fall, (dunkle
Eier).
Van Thiel (86) beobachtete in Holland, dass Anopheles maculi¬
pennis Verschiedenheiten in der Grösse zeigten, je nachdem sie
verseuchten oder malariafreien Gebieten kamen.
SwELLENGREBEL (82) und seine Schüler zeigten, dass die körper¬
lichen Unterschiede von A. maculipennis holländischer Herkunft
nicht auf äussere Einflüsse zurückzuführen sind. Als Folge weiterer
Untersuchungen nannte VAN Thiel (85) 1926 die kurzgeflügelte
« holländische »
Rasse Anopheles maculipennis var. atroparvus VAN
Thiel.
Später schlug Falleroni vor, diese Rasse als basilei zu bezeich¬
nen ; aber dieser Name wurde nicht beibehalten.
aus
BIOLOGIE
UND
VERBREITUNG EINIGER
STECHMÜCKENARTEN
119
Hackett und MiSSIROLl (56) wiesen auf die Zusammenhänge
zwischen macu/ipenms-Rassen in malariafreien und malariaverseuch¬
ten Gebieten hin, was zur Abtrennung weiterer biologischer Rassen
führte, nämlich
A. maculipennis
A. maculipennis
var.
var.
Fall. 1926
labranchiae Fall. 1926.
messeae
HACKETT und Martini wiesen auf die Notwendigkeit hin, weitere
Bestimmungsmerkmale als nur die Unterschiede am männlichen
Kopulationsapparat zur Verfügung zu haben.
La Face (18) führte morphologische Untersuchungen an Larven
der von Falleroni beschriebenen Rassen durch und zeigte die Möglich¬
keit, gewisse Merkmale an den Larven für die Rassengliederung zu
verwenden.
RoUBAUD und seine Mitarbeiter legten dar, dass in der Natur
auch Bastarde vorkommen können (typicus und atroparvus).
De Buck und SwELLENGREBEL nahmen an, dass die von Fallebeschriebenen Eier aus Italien mit den holländischen morpho¬
logisch identisch seien. Die Rassen typicus und messeae bilden zusam¬
eine Gruppe der gestreiften Eier, während labranchiae und
men
atroparvus der Gruppe mit gefleckten Eiern angehören.
Falleroni beobachtete unter « messeae »-Eiern vollständig dunkel¬
gefärbte, wobei die gefleckten Partien fast ganz verschwanden. Diese
Varietät wurde von Hackett (29) als « melanoon » bezeichnet, deren
schwarze Eier sich von den schwärzlichen A. bifmeatus-Eiern durch
die netzartige Oberfläche unterscheiden.
Eine Lokalrasse von A. maculipennis wurde 1934 von RoUBAUD
(69 b) aus der Normandie beschrieben, die sich biologisch ähnlich
wie atroparvus verhält und als maculipennis var. fallax bezeichnet
wurde. RoUBAUD (70) beschrieb ebenfalls eine neue Rasse von
RONI
Marokko, A. maculipennis sicaulti, welche dem Eitypus entsprechend
in die Nähe von labranchiae gehört. MiSSIROLl (57) fand im selben
Jahr in Sizilien ebenfalls eine neue Rasse, welche er als « pergusae »
bezeichnete und Ähnlichkeit mit labranchiae aufweist.
Schliesslich stellten RoUBAUD und Treillard (71) eine neue
maculipennis-Rasse in Portugal fest, die mit atroparvus verglichen
werden kann, sich aber durch die Schwimmkammer und den
Schwimmkammerindex unterscheidet.
In der Tabelle Nr. 53 sind die Merkmale der verschiedenen
Rassen nach HACKETT und MiSSIROLl (30) zusammengestellt.
um
d.
dex
ca.
17.
:
als
.tlek
I
vom
tnemgeS nelhartS .53,0
raaH
.53,0
¬mmiwhcS nnreelmhamratkS
,nez
:xed
:
ca.
d.
bis
mit
.004,0
dex
62,0
n¬nrierdenämblaaksmreviswnhacrST
hcon
bei
elknud
als
;
tsben
den
.nekcelF
u
a
r
g
l
e
k
n
u
D
nremmak
xedninerlehmamratkS
der
20.
2
nella
7291
6291
6291
8191
inorellaF
asseM
eaihcnarbaL
suvraportA
inorellaF
sucipyT
leihT
negieM
Van
II.
-kiwtne
gidnätslroavanhuramlaP rem¬nmiarkemmmiawkhmcSiwhcS
,
t
k
c
e
l
f
e
g
hcilnuärB
¬nirem akm iwhcS
raah
14.
,dneh
iewZ
.netsroB
redo
;
und
ererhem
ezruk
tes
ein
I
I
als
vom
vom
II.
II.
wie
etsroB
raaH
raaH
.raaH
hcilnhä tnemgeS
von
raaH
rhaJ
rotuA
der
esiew
,mag
,magonets ,lihpooZ
.manydoreteh
1
vom
II.
und
und
des
des
des
¬liet
.noitanrebihimeS
-onets
fpmuts
eressuÄ
.legülF egnaL .gnal
etsroB
reba
,lihpoporhtnA ¬.eltesgnüiletFlzltaipssengeuhzectrdlelesiitshsuceÄSg -ramlaP.tlekciwttnneemgeS ¬.rnaekwcheclsF negi,kucaerglleH
snehcdlihcS ¬redeifeg .lnhöweg
.magyrue ,lihpooZ
nevraL
reiE
essaR
sin epilucam.A elamkremnes aR nenedeihcsrev gnus afnem asuZ
elknud
-.whcS
.nredna
tnemgeS r¬emsmsöirwghcS ,u,arregdlnläeBH
snehcdlihcS gidnätsl ovnu.tlekciwtne
egnal ,zruk
hcan
eressuÄ
TTEKCAH
etludA
etsroB
fpmuts
.manydoreteh .legülF
ehcsigoloiB
,magyrue ,lihpooZ
Nr.
53
elamkreM
ellebaT
,)03(
.tzrükeg lLORlSSlM
den
:
neloP
ettiM
5391
6391
DUABUOR
UORISSlM
DUABUOR
ttekcaH
allaF
itluaciS
easugreP
icanruobmaC
bplahuS
siweL
5391
DUABUOR
noonaleM
6391
4391
ttekcaH
drallierT
und
dex
mit
Der
eaihc
:
reba
zu
.nih
den
.ttalg
der
der
von
¬ieb
.04,0
4391
und
ginew
.shucvirlanphoräta,ttkäcteelifreaGV
.nekcelF
uarg
,kcid
.3,0
¬-nar.nbtaiflireertmtsäetgaekinrmaaVribwmhecMS
,hciltuednu rneepkrcöekliEF
dex
ehan
,gid
raaH
raaH
nehciltued,hcilnhä
1
1
des
des
:
II.
gimröfnedaf.tletsär ¬ni.r9e2m.0a-k42m,0iwhcS
II
thcin
des
II.
ve¬
¬ieb
¬nätsl,lgoivtnruarasmeltanpemgeS
¬lap
thcin
I
ezruk
eressuÄ
-lap
se.tgintermagreaSm dnemhdenneudzrew,uarreglnkensuidE
se.tgintermagreaSm ¬¬smämmilweghecrSnxue,dtnkncaierlrfebemgmmeaMk
.n,ezgroazwrhebcüS
raaH
sig
des
,mag
.legülF
etsroB
am
nressuä
eressuÄ
des
zu¬
.snehcdlihcS netztipseg
lieT
etsroB
ehcälfrmeebgOikcel,ezirvawhcS
zteN
mit
gnerts
.eaihc
der
.manyd
,tläitehiproapVorhtnA ,tztipseguz snehcdlihcS
rheS
.manydomo,hlihpooZ
¬on e t s
-na r b a l
hcilnhä
¬omoh
.manydomoh
,magonets ,lihpooZ
.lihpooZ
122
WILLI
2. Neue
BÜTTIKER
Gesichtspunkte des Rassenproblems
In der neueren Culiciden-Literatur ist diese Unterscheidung
übernommen worden ; allerdings variiert darin die Anzahl der
genannten Rassen. MARTINI (52) und ElCHLER (15) beispielsweise
möchten systematisch diese Rassen z. T. als selbständige Arten ein¬
reihen. ElCHLER definiert, dass die « Art » eine Gruppe sehr ähnlicher
Individuen ist, die sich in der Regel in der Lebensweise ähnlich ver¬
halten, sich von andern in derselben Gegend lebenden verwandten
Arten abtrennen und sich nicht zu bastardieren pflegen. Im Gegen¬
satz dazu bezeichnet man als Unterarten oder Rassen solche nächst¬
verwandte Formen, die zu einer Art gehören, aber durch geogra¬
phische Trennung isoliert sind und dies auch an äusserlich erkenn¬
baren Merkmalen zeigen.
Neue Art- und Rasse-Einteilung, in
1.
Anlehnung
an
Anopheles typicus (Meigen) (= maculipennis
ElCHLER (15).
sens,
strict.), Gebirgs-
form.
Frühere Bezeichnung A. maculipennis typicus (Meigen) oder
A. maculipennis maculipennis (Meigen).
Eine Aufteilung in verschiedene Rassen scheint nach neueren
Untersuchungen (Weyer, 95) möglich.
2.
Anopheles
messeae
(Falleroni).
Rassegliederung
A.
:
(Falleroni) (Mittel-, Nord- und OstEuropa).
A. messeae subalpinus (Lewis und Hackett) (Balkanländer).
A. messeae melanoon (Hackett).
Weitere, noch nicht unterschiedene Rassen vermutlich in
Osteuropa.
Rassebastarde möglich.
3.
messeae
messeae
Anopheles labranchiae (FALLERONI) Küstenform,
Rassegliederung
in Brackwasser.
:
A. labranchiae atroparvus (van Thiel) (Mittel- und Nordeu¬
ropa).
A. labranchiae fallax (RoUBAUü) (Normandie). In die Nähe
von atroparvus gehörend.
A. labranchiae cambournaci (RoUBAUD und TreillARD) (Mit¬
telmeergegend).
A. labranchiae labranchiae
Spanien).
(Falleroni)
(Süd-Italien,
Süd-
BIOLOGIE
UND
VERBREITUNG
EINIGER STECHMÜCKENARTEN
A. labranchiae sicaulti (Roubaud) (Marokko).
A. labranchiae pergusae (MlSSIROLl) (Sizilien).
von
4.
123
In die Nähe
sicaulti gehörend.
Anopheles elutus (Evans) Brackige Sumpfgebiete,
gebiet, Griechenland.
Mittelmeer¬
(elutus) steht
den andern Arten
und ist auch äusserlich
gut zu unterscheiden (wenigstens für den, der sie kennt). In Deutsch¬
land kommt sie nicht vor ; in den Mittelmeerländern gehört sie zu
den gefürchtetsten Malariaüberträgern, besonders in den brackigen
Küstensümpfen Griechenlands, deren Salzwasser ihre Larven gut
ertragen. Ob elutus wirklich identisch ist mit sacharovi, wie einige
Autoren annehmen, bedarf wohl noch der Bestätigung. Der Name
sacharovi ist älter und hätte daher das Vorrecht. Da beide aber aus
ganz verschiedenen Gegenden beschrieben werden, sind es vielleicht
verschiedene Rassen derselben Art. »
Die Mücken der verschiedenen Anopheles-Rassen selbst sind
nicht unterscheidbar, da die morphologischen Unterschiede zu
minim sind, ihre Eier gehören aber deutlich verschiedenen Typen
an. Am besten unterscheiden sich die Rassen aber durch die Lebens¬
gewohnheiten, wie Uberwinterungsmodus, Art der Eiablage, Stech¬
gewohnheiten, Ansprüche an den Biotop usw. Aus diesen Gründen
könnte es sich um verschiedene echte Arten handeln. Eine Klarstel¬
lung' wird auch notwendig, weil z. B. die messeae aus verschiedenen
Ländern nicht gleich aussehen, sondern entsprechend den einzelnen
geographischen Arealen verschiedenen Untertypen entsprechen
(Weyer, 95). Diesen Untertypen würden also die wirklichen geo¬
graphischen Rassen entsprechen. Von einer endgültigen Aufspaltung
der maca/«/>enms-Gruppe ist deshalb meines Erachtens noch Umgang
zu
nehmen, da noch weitere morphologische Untersuchungen
besonders der Eier aus verschiedenen Gegenden des gesamten
durchzuführen sind.
Verbreitungsgebietes
Nach den Untersuchungen von UngurEANU und Shute (102)
ist es neuerdings möglich, die Imagines des Formenkreises maculipennis anhand der Flügelschuppen zu Unterscheiden. Vergleiche an
eigenem Material bestätigten die Resultate der beiden Autoren.
Untersuchungen über den Chromosomensatz bei den verschie¬
denen Rassen könnten möglicherweise weitere Resultate liefern, die
eine bessere Unterscheidung der Rassen ermöglichen. Die Detail¬
untersuchungen an den einzelnen macu/i/Senm's-Rassen verschiedener
Herkunft sind noch zu wenig zahlreich, um ein endgültiges Urteil
über sie zu bilden. Ich möchte deshalb in dieser Arbeit an der alten
systematischen Einteilung festhalten.
ElCHLER
der
(15) schreibt
: «
Diese Art
macu/ipenms-Gruppe verhältnismässig fern
—
—
124
WILLI
3. Die
BÜTTIKER
Verbreitung der maculipennis-Rassen
in der Schweiz
Über die Verbreitung der Anophelen in der Schweiz veröffent¬
lichte Galli-Valerio eine Reihe von Beobachtungen. Besonders in
einer Arbeit (23) berücksichtigte er das Verbreitungsgebiet von
A. macalipennis, A. bifurcatus und A. nigripes. Dem wissenschaft¬
lichen Stand der damaligen macu/r'penrHs-Untersuchungen (1918)
zufolge war es ihm unmöglich, Rassenunterschiede festzustellen.
Diese Fundangaben dienten Gaschen (26) zur Errichtung einer
Verbreitungskarte, welche durch seine eigenen Funde erweitert
wurde.
Von den ungefähr 11 europäischen maculipennis-Rassen sind
aus der Schweiz deren drei bekannt, nämlich : messeae, melanoon,
und typicus.
Abb. 41.
—
a,
Die
Verbreitung
von
A. macalipennis
in
Europa (nach
melanoon in
Europa (nach
messeae
var.
Christophers; II).
Abb. 41.
—
b, Die Verbreitung
von
A.
maculipennis,
var.
Christophers; II).
Abb. 41.
—
c,
von A. maculipennis
Christophers; II).
Die Verbreitung
var.
typicus in Europa (nach
BIOLOGIE UND VERBREITUNG
Fundorte
von
A.
EINIGER
maculipennis
var.
125
STECHMÜCKENARTEN
messeae
in der Schweiz
Tabelle Nr. 54
Nr.
1
Nähere
Fundort
Affoltern bei Zürich
Imagines
Bezeichnung des Fundortes
aus
Viehstall im Labor Eier ab¬
Fund¬
datum
17.4.47
gelegt. Eier und Larven in den umliegenden
2
3
4
5
Arlesheim/BL
Brestenberg/AG
Buchs/SG
Büren a. d. A./BE
6
7
Bürliacker/AG
Dietikon/ZH
8
Egelsee/ZH
9
10
11
12
13
14
Effretikon/ZH
Gattikon/ZH
Genf
Katzensee/ZH
Klingnau/AG
Mauensee/LU
15
Rheineck/SG
16
Rheineck/SG
17
18
Rheinfelden/AG
Oberkirch/LU
19
20
Sitten/VS
La Sauge/VD
21
Spreitenbach/AG
22
23
Villeneuve/VD
Wohlen/AG
24
Zürich
Sümpfen häufig.
Waldweiher.
Hinter der Litoralzone des Hallwylersees
In den Altwassern des Rheins.
In den Altwassern der Aare, nur geringe
Anzahl.
Kleiner Teich in der Talsohle, sehr häufig
Altwasser der Limmat, gegen Geroldswil,
11.5.47
10.5.47
30.5.47
18.5.47
veraigten Stellen,
25.4.47
sehr häufig.
In der Randzone
an
spärlich.
Weiher mit sehr starker Entwicklung
Lemna mimor.
Weiher oberhalb des Dorfes.
Botanischer Garten.
von
Randpartien, nicht häufig
Altwasser des Rheins gegen Koblenz.
in der Seegegend,
verbreitet.
Hinter Litoralzone des Bodensees, in Ent¬
wässerungsgraben, sehr häufig.
Im Altwasser des Rheins, in der Nähe eines
Bauernhofes, Larven sehr häufig.
Auweiher.
Unmittelbar am Sempachersee in Entwäs¬
serungsgraben, verbreitet.
Nebenfluss der Rhône.
In den Ställen des Oekonomie-Gebäudes ;
als überwinternde $. Sommermücken in
ehemaligem Militärbunker, sehr häufig ;
als Larven in Entwässerungsgraben, sehr
Entwässerungsgraben
häufig.
In
den umliegenden Kiesgruben, recht
häufig.
Entwässerungsgraben, Eier und Larven.
Entwässerungsgraben an der Bahnlinie
Wohlen-Bremgarten.
Gartenanlage der Eidg. Techn. Hochschule
Stadtzentrum.
10.5.47
24.4.46
21.6.47
15.6.45
28.4.47
10.5.47
9.6.46
2.5.47
2.5.47
15.8.47
9.6.46
7.47
7.3.46
20.5.46
7.4.47
II. 1946
30.7.47
126
WILLI
Fundorte
von
BÜTTIKER
A. maculipennis
var.
melanoon in der Schweiz
Tabelle Nr. 55
Nr.
Fundort
1
2
Aarau/AG
Affoltern b/Zürich
3
4
Ariesheim/BL
Aeschi/SO
5
6
7
Baldegg/LU
Büren a. d. A./BE
Dietikon/ZH
Nähere
Bezeichnung
des Fundortes
Altwasser der Aare, Schachen.
Randtümpel des Katzensees, in den Ställen
eines Bauernhauses, sehr häufig.
Waldweiher oberhalb des Dorfes.
Feuerweiher des Dorfes, mit typicus zusam¬
Fund¬
datum
28.4.46
17.4.47
11.5.47
18.5.47
men.
Entwäss.-Graben am Baldeggersee, häufig.
Im Altwasser der Aare, spärlich.
In den Altwassern der Limmat gegen
Geroldswil.
Vereinzelt in den Altwassern der Aare.
In den Schachen der alten Aare.
An den Ufern des Hasli-Sees, stellenw.
8
9
10
Döttingen/AG
Dozingen/BE
Hasli/ZH
11
12
Klingnau/AG
Murgenthal/BE
13
14
Oristal
15
Oerlikon-Zürich
Gartenfässer.
16
17
18
19
Reazzino/TI
20
La
Altwasser des Ticino-FIusses, verbreitet.
Altwasser des Rheins.
In den Altwassern der Reuss.
Hinter der Utoralzone des Sempacher-Sees
in Entwässerungsgräben.
In den umliegenden Entwässerungsgräben
Larven sehr häufig, Imagines im Sommer in
Ställen u. Bunkern, im Winter in Kellern.
In kleinen Weihern an der Strasse nach
Eiken.
In Tümpeln und Gräben häufig.
9.6.46
18.5.47
24.4.46
10.5.47
18.5.47
22.4.46
häufig.
Moos-See/BE
(Liestal)/BL
Rheineck/SG
Rottenschwil/AG
Sempach/Lu
Sauge/VD
21
Stein/AG
22
Verzasca-Delta/TI
In den Aare-Schachen, vereinzelt.
An den bewachsenen Ufern des kleinen
Sees, recht häufig.
An den Randzonen des Moos-Sees, häufig.
In den Weihern der Fischzuchanstalt, stel¬
lenweise häufig.
10.5.47
18.5.47
8.6.46
11.7.47
Sommer
1946 und
1947
25.8.47
2.5.47
10.5.47
9.6.46
7.3.46
u.
a.
•
6.7.47
25.8.47
BIOLOGIE
Fundorte
UND VERBREITUNG
von
A. maculipennis
EINIGER
var.
127
STECHMÜCKENARTEN
typicus in der Schweiz
Tabelle Nr. 56
Nr.
Fundort
1
Affoltern-Zürich
2
Amlikon/TG
3
4
5
Aeschi/SO
Balchensthal/ZH
6
Basel
7
Bern
8
9
Boniswil/AG
Cadenazzo/TI
Nähere Bezeichnung des Fundortes
Fund¬
datum
Aus einem Viehstall, Nähe Katzensee,
4 Gelege erhalten.
Aus Altwasser der Thur, Grundwasser¬
Sommer
1947
2.5.47
tümpel.
Baldegg/LU
10
Flaach/ZH
11
FuIIy/VS
12
13
14
15
Gettnau/Zug
16
Mollis/GL
17
18
Magadino/TI
Stein-Säckingen/AG
19
20
Villeneuve/VD
Wermetswil/ZH
Gontensch wil /AG
Hettlingen/ZH
Möhlin/AG
Im Feuerweiher, mit melanoon zusammen.
Im Quellenweiher, häufig.
Entwässerungsgraben hinter der Litoralzone des Baldegger-Sees.
Riehen, Weiher des Vogelschutz-Reser¬
18.5.47
29.6.47
9.6.47
vates.
11.5.47
20.6.46
Im Teich des Gartens der Eidg. Versuchs¬
anstalt Liebefeld.
Aus dem Entwässerungsgraben beim « Risi ».
Beim « Parco Selvaggio » in den Grund¬
wasserkanälen, regelmässig und häufig.
In kleinen Grundwasser-und Altwasser¬
tümpeln der Thur, sehr häufig.
In den verkrauteten Meliorationsgräben der
Talsohle.
Aus Tümpeln der Kiesgruben, recht häufig.
Aus einem Brunnentrog, mit Culex pipiens.
Vereinzelt in einigen Feldtümpeln.
In einem Gänseweiher des « Ziegelhofes » in
sehr grosser Zahl. Eutrophierung sehr
9.6.46
25.8.47
1.6.47
11.7.47
8.6.46
7.8.47
1.6.47
22.8.47
gross !
Fundorte
1
2
3
4
5
von
3.5.47
25.8.47
6.7.47
7.4.47
29.6.46
A. maculipennis in der Schweiz, ohne Rassenangabe
Airolo/TI
Bonfol/BE
Tramelan/BE
Rheinfelden/AG
Schaffhausen
Aus einem grossen Wassergraben in statt¬
licher Anzahl.
Aus einem Seitenarm des Tessin-FIusses.
An der Randzone der beiden kleinen Seen
an der Strasse Stein-Eiken.
Im Altwasser der Rhône, Eier u. Eilarven.
Im Feuerweiher des Dorfes in grosser
Anzahl.
Gegen Ambri-Piotta, in Tümpeln
Waldweiher.
Etang de la Gruyère, Randzone.
Männchen und Weibchen eingefangen.
Aus Tümpel, in Stadtnähe.
4.6.45
12.6.45
12.6.45
Juli 47
29.5.45
128
WILLI
BUTTIKER
Aus den Verbreitungskarten (Abb. 41, a, b und c) der Rassen
(CHRISTOPHERS, 11) ist die Verbreitung von messeae, melanoon und
typicus in Europa zu entnehmen. Messeae erstreckt sich über den
ganzen Kontinent, ebenso die Varietät typicus. Eine mehr südliche
Form scheint melanoon zu sein, die
aus den bisherigen Resultaten
ihre Hauptverbreitung in Italien hat.
zu schliessen
Als erste gehen Gaschen (25) und BoRRANl (5) auf die Rassen
von maculipennis des schweizerischen Verbreitungsgebietes ein. Der
—
—
meldet Fundorte aus den Kantonen Waadt, Uri, Graubünden
und Wallis, BoRRANl aus dem Kanton Tessin.
Meine eigenen Untersuchungen erstrecken sich auf das gesamte
•Gebiet der Schweiz. In der Verbreitungskarte sind die drei bis
jetzt gefundenen Rassen von A. maculipennis gesondert aufgeführt
erstere
(Abb. 42).
Gesetzmässigkeiten der Verbreitung
Messeae.
Die schweizerischen Fundorte sind in der Tabelle Nr. 54
zusam¬
mengestellt. Übereinstimmend geht aus den Funden hervor, dass
diese Rassen hauptsächlich in grösseren Tümpeln, an Uferpartien
der Seen, in den Entwässerungsgräben mit reichlicher Vegetation,
in seichten, warmen Wasserlachen ihre Brutorte haben. Mit Vor¬
liebe werden Wasseransammlungen gewählt, die eine starke Algen¬
vegetation (z. B. Spirogyra) aufweisen. Der Eutrophierungsgrad ist
erfahrungsgemäss höher als bei typicus, aber ungefähr gleich wie
bei den Brutorten von melanoon. Dementsprechend werden Grundund Quellwasseransammlungen gemieden. Stark saure Gewässer wer¬
den verschmäht, so dass nur ausnahmsweise messeae in MoorgeAvässern anzutreffen ist. Vielfach ist messeae mit melanoon vergesell¬
schaftet.
Melanoon.
Die schweizerischen Fundorte sind in Tabelle Nr. 55
zusammen¬
gestellt. Vegetationsreiche Entwässerungsgräben, seichte Wasser¬
ansammlungen mit starker Algenvegetation (z. B. Spirogyra), Alt¬
wasser mit starken Temperaturschwankungen sind die beliebtesten
Brutorte. Der Eutrophierungsgrad der Gewässer ist in der Regel
ähnlich wie bei messeae.
Typicus.
Die schweizerischen Fundorte sind in Tabelle Nr. 56
zusammen¬
gestellt. Das Weibchen legt die Eier vorzugweise auf die Oberfläche
kleiner Wasseransammlungen, wie Quellteiche, kleine Wassertümpel,
welche mit dem Grundwasser in Verbindung stehen ; kleine Tümpel
BIOLOGIE UND VERBREITUNG EINIGER
STECHMÜCKENARTEN
y
Abb. 42.
—
Die Fundorte
von
5
Anopheles macalipennis in der Schweiz nach Rassen
getrennt.
9
129
130
WILLI
BUTTIKER
und Weiher mit konstanten und relativ tiefen Temperaturen werden
bevorzugt ; eutrophierte Gewässer dagegen werden gemieden.
Das günstigste Verbreitungsgebiet für alle drei Rassen ist das
schweizerische Mitteland mit seinen Süsswasserseen. Die tief in die
Alpen eingeschnittenen Täler der Rhone, des Rheins und Tessins
bieten ebenfalls güngstige Brutorte 1. Bis zu Ende des 19. Jahrhun¬
derts waren Teile dieser Gebiete autochthone Malariaherde (vergl.
Galli-Valerio, 23).
4. Grösse und
von
Zeichnungsmuster
Eiern schweizerischer Herkunft
HACKETT, Martini und MjSSiROLl (58) verwendeten für ihre
sog. Schwimmkammer-Index, den man wie
Untersuchungen den
folgt ausdrücken kann
:
Schwimmkammer-Index
=
Länge
der Schwimmkammer
Länge des Eies
GASCHEN (25) veröffentlicht
die mit eigenen Resultaten
a
b
Messungen an schweizerischem Material,
verglichen werden.
Tabelle Nr. 57
Schwimmkammer¬ Schwimmkammer¬
index (Gaschen)
index (Büttiker)
Rasse
A. maculipennis
A. maculipennis
A. maculipennis
Rasse
Messeae
Melanoon
Typicus
var.
messeae
var.
melanoon
var.
typicus
Eilänge (Gaschen)
0,724
0,685
0,712
mm
mm
mm
0,447
0,527
0,435
Eilänge (Büttiker)
n
0,691
0,710
0,670
Eier
.
=
20
mm
mm
von
0,385
0,424
0,396
Herkunft
Zürich-Aff.
Zürich-Aff.
Zürich-Aff.
Weibchen in Gefangen¬
schaft abgelegt.
1
Die Fundorte anderer Autoren (Gaschen, 25) und (BoRRANl, 5) sind dabei
nicht berücksichtigt. Anopheles bifurcatus und A. nigripes Hess ich ebenfalls ausser
Betracht ; eine Veröffentlichung der gesamten von mir in der Schweiz gesammelten
Culiciden wwd später erscheinen.
BIOLOGIE
UND
VERBREITUNG
EINIGER
131
STECHMÜCKENARTEN
Die grossen Unterschiede lassen sich vielleicht auf individuelle
Messfehler zurückführen. Es ist kaum anzunehmen, dass das Material
innerhalb eines kleinen Areals so starken Schwankungen unterworfen
ist. Diese Untersuchungen mögen auf die Schwierigkeit des statis¬
Vergleichens von Untersuchungsmaterial verschiedener Au¬
aufmerksam machen.
Aus verschiedenen Landesgegenden der Schweiz wurden Eier
der mikroskopischen Untersuchung unterzogen. Eine Auswahl der
Zeichenmuster kommt in Abb. 43 zur Darstellung. Die beiden
Querbänder sind bei allen abgebildeten Eiern deutlich zu erkennen.
Die helle Felderung ist verschieden ausgeprägt, und die dunkle
Grundtönung variiert oft beim selben Eigelege sehr stark. Auch hier
treten Querbänder auf, die allerdings schwach angedeutet sind. Die
Grundtönung des Eies ist schwarzbraun, sie kann aber je nach Alter
in der Intensität variieren. Am deutlichsten sind die beiden Quer¬
bänder bei typicus vorhanden. Die Eipole weisen ebenfalls eine
Dunkelfärbung auf. Die Grundfärbung ist silbrig-grau, so dass sich
die Querbänder besonders kontrastreich abheben.
tischen
toren
Abb. 43.
—
Eier
von
A. maculipennis verschied. Rassen, a—d,
melanoon.
f—k, var. typicus.
—
var. messeae.
—
e,
var.
VI.
BIOLOGISCHE
AN
BEOBACHTUNGEN
CULICIDEN
1. Entwicklungsverlauf einzelner Eigelege
Bei vielen Insektenarten ist festzustellen, dass sich die Männchen
durchschnittlich rascher entwickeln als die Weibchen und auch früher
schlüpfen. Diese Erscheinung wird als Protandrie bezeichnet. Sie
ist besonders unter den Fliegen verbreitet und verhindert in gewissem
Grade die Geschwister-Ehe.
über die Verhältnisse bei Culiciden schreibt Martini (52), dass
die Männchen von A. maculipennis früher schlüpfen als die Weibchen.
Es ist dies die einzige mir bekannte Literaturangabe über Protandrie
bei Stechmücken.
Bei den vorliegenden Versuchen kommen zwei Arten zur Beob¬
achtung, nämlich C. pipiens und A. maculipennis var. messeae.
Methode
Je ein Eigelege wurde im Labor aufgezogen, und die Larven wur¬
den mit der üblichen Nahrung gefüttert. Die Wassertemperatur betrug
ca. 18° C. Die geschlüpften Imagines wurden bei C. pipiens dreimal
täglich ausgezählt, nämlich um 0800, 1200 und 1800, bei A. maculi¬
pennis um 0800 und 1900 Uhr.
Resultate
Culex pipiens.
Die Beobachtungen ergaben (Tab. Nr. 58) eine deutliche Pro¬
tandrie bei C. pipiens. Das Schlüpfen der Weibchen beginnt 1 bis
1 Yz Tage nach dem ersten Auftreten von Männchen. Dagegen
erschienen noch während zwei Tagen Weibchen, nachdem alle
Männchen geschlüpft
waren.
gesonderten Kolonne werden die im Stundendurchschnitt
schlüpfenden Tiere aufgeführt. Es geht daraus hervor, dass in der
Regel die Quote am Morgen grösser ist als nachmittags.
In einer
82
80
1
2
1
1
5
162
5
1
2
1
1
8
16
16
8
3
1
3
5
2
6
10
13
23
12
8
30
5
4
8
18
12
1
4
4
1
1
6
2
6
8
4
17
3
5
2
6
9
2
7
6
.lhnceshegcbliheaWznA .lhnceshegcnlnhäaMznA
—
.U?(
emmuS
1
der
?
61.0
52,0
52.0
41.0
52.1
33.1
41.1
52.1
33,1
41.2
00,3
06,1
03,2
61,2
72,0
34.1
00,1
00,4
66,1
573,0
—
t _i-nnhecdsuhtcSrud
xeluC
0081
0021
0081
0080
0021
0080
0081
0021
0081
0080
0021
0080
0022
0071
0011
0042
0071
0311
0011
0080
0000
)ednutS(
tieZ
C°81
ca.
sgat
sgat
sthcan
sgat
sgat
—
sgat
-segaT
mumixaM
ednefpülhcS sneipip
:
4
22
43
43
24
26
Tag
(1)
pro
Nr.
58
lhaztmaseG
ellebaT
senigamI
latoT
64.8.92
64.8.82
64.8.72
64.8.62
64.8.52
64.8.42
mutaD
ruta6r4e.p8m.e4t2/s3h2cusreV
S
.lhcseg
lhaZ
30
~~
—
—
—
—
3
1
2
—.
5
—
6
1
4
2
2
?
2
3
4
3
4
4
1
1
—
—
1
—
2
—
4
—.
29
lhaZ
—
—
—
.lhcseg
1
2
1
59
1
2
1
0
4
3
2
8
1
9
0
4
3
7
4
4
4
I
1
0
+
emmuS
cî
von
9
Vu
=
Vu =
Vu
=
Vu =
Vu =
Vn =
=
=
Vu =
Vi3
Vir
Ve =
Vu =
Vb =
Vu =
=
0=
Vu =
Via
80.0
—
03
0.2
0.2
0.7
0.2
0.5
0.4
0.2
0.6
0.3
0.4
0.3
90.0
0.2
80.0
=
Vu =
Vis
80.0
t inhcshcrud
¬nednutS
9
—
—
sthcan
sgat
sgat
—
sgat
—
sgat
—.
sgat
bis
—
—
sthcan
.grom
sthcan
.dba
mumixaM
segelegiE
sisnenepiiglaumcIamedsneelfehppüolnhAcS
senie
.rav
latoT
0091
74.6.72
74.6.62
74.6.52
74.6.42
74.6.32
74.6.22
74.6.12
74.6.02
74.6.91
74.6.81
0080
0091
0080
0022
0080
0091
0070
0042
0080
0091
0080
0091
0080
0091
0080
0091
0080
0080
0091
.pmeT
mutaD
°22—81
tieZ
C
1
5
—
8
—
10
—
4
—
10
—
10
—
7
—
1
—
Tag
3
—
pro
Nr.
59
lhaztmaseG
ellebaT
eaessem
BIOLOGIE UND
VERBREITUNG EINIGER
STECHMÜCKENARTEN
135
Es sind zwei Schlüpf maxima vorhanden; das erste Maximum
rührt von den Männchen, das zweite von den Weibchen her (Abb.44).
Das Geschlechtsverhältnis beträgt 1: 1 für Normalfälle ; bei Hungerund Intoxikations-Zuchten sind die Männchen in der Überzahl
(Ulmann, 89).
21.8.1946
Abb. 44.
—
Culex pipiens.
25.
Schlüpfen
Anopheles maculipennis
27.
28.
var.
von
28.
29.8.1948
Männchen und Weibchen eines
Eigeleges.
messeae.
Ähnliche Verhältnisse ergaben sich bei A. maculipennis var.
messeae. Ein Heines Eigelege wurde bei 18—20° Wassertemperatur
aufgezogen.
Auch bei dieser Culicidenart schlüpften die Männchen zuerst,
die Weibchen folgten im Abstände von 2 Tagen. Das Schlüpf¬
maximum der Männchen fiel auf den vierten Tag, dasjenige der
Weibchen auf den siebenten und achten Tag, auf den Schlüpfbeginn
bezogen. Die Schlüpfmaxima beider Geschlechter fallen auf die
hellen Tagestunden, da der Stundendurchschnitt von 0800 bis
1900 höher liegt als von 1900 bis 0800 des folgenden Tages (vergl.
Tabelle Nr. 59).
Das Geschlechtsverhältnis lautet auch bei dieser Art 1 : 1 (30 et :
29$).
Zusammenfassend kann gesagt werden, dass den untersuchten
Culiciden eine deutliche Protandrie eigen ist. Der Schlüpf Vorgang
entfällt bei beiden Arten vorwiegend auf die Tagesstunden, zum
kleineren Teil auf die Nacht.
•
2. Das Schwimmvermögen der Culiciden
Untersuchungen über
das Schwimmvermögen der Culiciden hat
A. maculipennis durchgeführt. Aus seinen
Beobachtungen geht hervor, dass eine Schwimmtiefe von 10 cm nur
bei 37—40° C erreicht wird. Angaben weiterer Autoren konnten
L. V. IVANOVA (41)
an
136
WILLI
BÜTTIKER
nicht erhalten werden ; es scheint aber, dass Untersuchungen über
das Schwimmvermögen sehr spärlich durchgeführt wurden.
In diesem Kapitel werden die Tauchzeiten und Schwimmtiefen
einer näheren Prüfung unterzogen. Zur Untersuchung gelangten
Larven und Puppen von C. pipiens, A. vexans, A. geniculates und
A. maculipennis.
Versuch Nr. 1
:
Maximale Schwimmtiefen im Verlaufe
der
Larvenentwicklung
Methode
von C. pipiens, A. vexans und A. maculipennis var.
wurden zu drei Versuchsreihen in ein Pyrexglasrohr von
150 cm Höhe und 12 cm Durchmesser überführt. Zweimal täglich
wurden die maximalen Schwimmtiefen gemessen, wobei die Larven
durch Erschüttern der Wasseroberfläche zum Tauchen veranlasst
wurden.
Um eine Durchschnittstemperatur von 18° C zu erhalten, wurde
ein gleichmässig erwärmter Raum für die Versuche benützt. Die
Larven wurden täglich zweimal mit der üblichen Nahrung (Piscidin„
Je 20 Eier
messeae
Hundebiscuit, Heuaufguss) gefüttert.
Resultate
C. pipiens.
C. pipiens erfahren eine merkliche Zunahme der
ersten Lebenstagen. Bis vor dem Errei¬
chen des Puppenstadiums wird eine maximale Schwimmtiefe von
150 cm erreicht (Tabelle Nr. 60).
Larven
von
Schwimmfähigkeit in den
A.
vexans.
Die ausserordentlich rasche Zunahme der Schwimmfähigkeit bei
A. vexans ist aus der Abb. 45 ersichtlich. Nach vier Tagen wird- von
den II. Larven schon eine maximale Tiefe von 150 cm erreicht. Die
Versuche mussten mangels eines grösseren Zylinderrohres abge¬
brochen werden, da die gesamte Tiefe des Versuchsglases schon
nach vier Tagen erreicht wurde (Tabelle Nr. 61).
A.
maculipennis
var.
messeae.
Im Gegensatz zu den bereits beschriebenen Arten nimmt hier die
Schwimmfähigkeit im Verlaufe der Larventwicklung nur sehr lang¬
sam zu. Bei den vorliegenden Versuchsverhältnissen erreichte die
verpuppungsreife Larve nach 21,5 Tagen eine maximale Tiefe von
45 cm (Tabelle Nr. 62).
BIOLOGIE
150
II
cm
UND
137
VERBREITUNG EINIGER STECHMUCKENARTEN
/ A. vexans
100-
20
Abb. 45.
—
Das Schwimmvermögen verschiedener Culiciden, in
Tage
Abhängigkeit ihres
Entwicklungsalters.
Zusammenfassend kann gesagt werden, dass die erreichten
Schwimmtiefen bei den untersuchten Arten sehr unterschiedlich
sind. Es lassen sich oekologische und bewegungs-physiologische
Zusammenhänge ableiten, da z. B. A. maculipennis var. messeae als
Brüter seichter Wasseransammlungen nur in geringe Tiefen schwimmt,
während C. pipiens als Brüter tieferer Gewässer auch ein besseres
Schwimmvermögen aufweist.
Ganz besonders interessant liegt der Fall bei Aedes vexans, dessen
II. Larve zwei stark erweiterte Hauptäste der Tracheen aufweist
(Abb. 46, C). Verglichen werden mit den Tracheenstämmen von
A. vexans diejenigen von A. maculipennis var. messeae und C. pipiens.
.
Maximale Schwimmtiefe
der
von
Aedes
vexans
im Verlaufe
Larvenentwicklung
Tabelle Nr. 6I
Datum
Stunde
I6.4.47
1500
2130
0800
1800
0800
1800
0800
1800
0800
I7.4.47
I8.4.47
19.4.47
20.4.47
1800
Schwimmtiefe in cm
Bemerkungen
Larven geschlüpft
2-3.
10
32
—
80
—
135
150
—
ca.
17
26
28
34
40
45
55
12
15
16
20
3.0-2,0
8.5
II.
ca.
.tS-.L
C°81
.pmetressaW
tfpnüelvhrcasleigE
in
cm
2
64.9.3
64.9.2
64.9.1
64.8.13
64.8.03
64.8.92
64.8.82
von
mutaD
der
0081
0080
0081
0080
0081
0080
0081
0081
0080
0081
0080
0080
0081
0080
ednutS
gnulkciwtnenevraLnegnukremeB efeitmmiwhcS
efeitmmiwefhucaSlreelVamixaM
im
150
140
145
—
121
114
65
75
85
95
105
in
IV.
ca.
L.
St.
gn u p p u p r e V
.tS-.L
.III
57
cm
Nr.
60
negnukremeB efeitmmiwhcS
ellebaT
sneipip xeluC
0081
0080
0081
0081
0080
0080
0081
0081
0080
0080
0081
0080
0021
0080
ednutS
64.8.72
64.8.62
64.8.52
64.8.42
64.8.32
64.8.22
64.8.12
64.8.02
mutaD
3
3
4
4
5
4
5
5
5
6
5
6
5
6
7
7
2-3
2
cm
negnukremeB efeitmmiwhcS
.rav
in
der
mutaD
dnerhäw eaessem
0081
0080
0080
0081
0022
0080
0080
0081
0080
0081
0080
0081
0080
0081
0081
0081
0080
0080
0080
0081
0061
0080
ednutS
'
0,5
—
nenfepvürlahlciSE
ca.
74.5.01
74.5.9
74.5.8
74.5.7
74.5.6
74.5.5
74.5.4
74.5.3
74.5.2
74.5.1
74.4.03
d.
0081
0080
0080
0081
0081
0080
0080
0081
0080
0081
0080
0081
0081
0080
0081
0080
0080
0081
0081
0080
0081
0080
ednutS
von
45
30
40
30
30
25
24
24
24
19
—
—
19
20
15
17
18
13
10
in
8
9
cm
Nr.
62
negnukremeB efeitmmiwhcS
ellebaT
sin genpiullukccaimwtsneelneehvproanAL efeîtm îwhcS elamixaM
74.4.92
74.4.82
74.4.72
74.4.62
74.4.52
74.4.42
74.4.32
74.4.22
74.4.12
74.4.02
74.4.91
mutaD
140
WILLI
BÜTTIKER
Diese anatomische Besonderheit erlaubt natürlich eine Speicherung
grösserer Luftmengen, was eine längere Schwimmdauer zur Folge
hat. Dadurch kann Aèdes vexons schon im II. Larvenstadium eine
beträchtliche Schwimmtiefe erreichen.
Versuch Nr. 2
:
Mittlere Schwimmtiefen
Methode
Je 50 Larven (IV. Stadium) eines Eigeleges von Culex pipiens,
A. geniculates und A. maculipennis var. messeae wurden in einen Stand¬
zylinder von 50 cm Höhe übertragen. Bei der Erschütterung der
Wasseroberfläche flüchtete sich die Grosszahl der Larven nach unten,
wobei jeweils nur eine Larve nach freier Wahl zur Messung der
Schwimmtiefe benützt wurde (Wassertemperatur ca. 18° C).
Resultate
Durch die Wiederholung der Versuche ergaben sich folgende
Mittelwerte :
a) Culex pipiens erreichte eine mittlere Schwimmtiefe von 22,4 cm
b) Aedes geniculatus eine solche von 19,4 cm ;
c) Anopheles maculipennis var. messeae nur 6,1 cm.
;
Wieder ergaben sich Zusammenhänge zwischen oekologischen
und bewegungs-physiologischen Gesichtspunkten ; A. maculipennis
als Brüter untiefer Wasseransammlungen erreicht nur eine geringe
Schwimmtiefe im Vergleich zu den andern beiden untersuchten
Arten, während A. geniculatus nahe an den Wert von C. pipiens
herankommt. Diese mittleren Zahlenwerte ergeben ein noch deut¬
licheres Bild als die Höchstwerte des Versuches 1 über die erreichten
Schwimmtiefen.
Versuch Nr. 3
:
Bestimmung der mittleren Tauchzeiten
Puppen
der Larven und
Methode
-
Ähnlich wie bei Versuch 2 wurden je 50 Larven (IV. Stadium)
desselben Eigeleges von C. pipiens, A. geniculatus und A. maculi¬
pennis var. messeae in einem Standzylinder von 50 cm Höhe geprüft.
Ebenso gelangten je 30 Puppen von Culex pipiens, Aedes vexans und
Anopheles maculipennis var. messeae zur Prüfung. Durch Erschüt¬
terung der Wasseroberfläche flüchtete sich jeweils die Grosszahl der
BIOLOGIE UND
141
VERBREITUNG EINIGER STECHMUCKENARTEN
Larven nach unten. Auf diese Weise konnten pro Teilversuch die
Tauchzeiten von durchschnittlich 5—6 Larven, resp. Puppen mit
Hilfe der Stoppuhr gemessen werden.
Resultate
Aus der Wiederholung der
mittlere Tauchzeiten (Tab. 63) :
ergaben sich folgende
Versuche
Tabelle Nr. 63
Culiciden
—
Art
IV. Larven
Puppen
104"
23.7
55.8
43.2
4.5"
>300
"
"
"
"
A. maculipennis
var. messeae
.
11
"
Zusammenfassend kann gesagt werden, dass die Puppen eine
viel kürzere Tauchzeit aufweisen als die Larven. Bei den unter¬
suchten /4ëc?es-Arten kann die ausserordentliche Tauchfähigkeit mit
der gespeicherten Luft in den voluminösen Tracheenstämmen in
Beziehung gebracht werden. Entsprechend der geringen Tauchtiefe
von A. maculipennis var. messeae sind auch die Tauchzeiten nur kurz.
Abb. 46.
—
Tracheen-Haupt¬
Culicidenlarven.
Verschieden stark vergrössert.
A. Culex pipiens, IV.
Orig.
Larve.
B, Aedes vexans, I.
Larve.
C, Aedes vexans, II.
Larve.
D, Anopheles maculi¬
pennis var. messeae, IV. Larve.
stämme
von
—
—
—
—
VII.
ZUSAMMENFASSUNG
1. In der vorliegenden Arbeit werden die Ergebnisse von Metho¬
den zur Aufzucht von Culiciden gegeben. Als Larvenfutter hat sich
im Grossen und Ganzen Hundekuchen, als Imaginalfutter Dattel¬
fleisch als Zusatznahrung gut bewährt.
2. Die minimale Praeovipositionszeit von C. pipiens und A. maculipennis messeae beträgt 4 Tage, resp. 3,5 Tage (18° C).
3. Der Temperatureinfluss auf die Entwicklungszeiten der ver¬
schiedenen Culicidenstadien wird bestimmt ; die Entwicklungszeit
von I. Larve bis Imago beträgt :
C. pipiens
A. vexans
A. geniculates
A. maculipennis
....
10,4
10,0
18,3
16,7
22° C
4. Das Thermopraeferendum der Larven, Puppen und Ima¬
gines ist verschieden. So liegt es bei Culex pipiens Larven bei 21
25° C, und bei A. bijurcatus Larven bei 20—25° C, bei den Imagines
(?) der selben Art aber bei 30—31 °C im Sommer, im Winter bei
2—6° C. Eine Verschiebung des Thermopraeferendums im Verlauf
der Larvenentwicklung tritt nicht ein.
—
5. Die Aktivitätskurven und Aktivitätsstufen der einzelnen
Arten stimmen durchschnittlich miteinander gut überein. Anopheles
bijurcatus als Kaltwasserform zeigt die grössten Abweichungen, da
eine Aktivität auch bei +0° noch zu beobachten ist. Eine starke
Zunahme der Aktivität erfolgt im Gebiet von 4—8° C.
6. Imagines von C. pipiens und A. maculipennis var. messeae zeigten
bei den durchgeführten Untersuchungen über die Vorzugsfeuchtig¬
keit gegenteiliges Verhalten. Erstere bevorzugte hohe Luftfeuch¬
tigkeit (bis 100 %), während die letztere die trockensten Stellen der
Feuchtigkeitsorgel aufsuchten (bis 25 %).
7. Der Bereich der längsten Lebensdauer liegt bei C. pipiens und
A. maculipennis ($ ungefüttert) bei —2° C bis +2° C und 100 % Vor¬
zugsfeuchtigkeit. Die maximale Lebensdauer beträgt bei C. pipiens
BIOLOGIE
UND
VERBREITUNG
EINIGER
STECHMÜCKENARTEN
143
($ ungefüttert) ca. 4 Tage, bei A. maculipennis var. messeae ($ unge¬
füttert) ungefähr 12 Tage. Das Verhältnis der Lebensdauer bei
Männchen und Weibchen beträgt bei beiden Arten 2 : 3 bis 3 : 4.
8. Die Larvensterblichkeit bei künstlicher Aufzucht nimmt durch
Einwirkung elektromagnetischer Strahlung stark ab. Die Strahlung
übt einen starken Einfluss auf die Fett- und Pigmentbildung aus.
8. Die Phototaxis ist je nach Entwicklungsstadium und dem
physiologischen Zustand der Tiere verschieden. Larven und Puppen
von C. pipiens sind deutlich positiv phototaktisch, während Imagines
vor der Diapause stehend negativ phototaktisch sind. Imagines nach
der Diapause sind am Morgen positiv phototaktisch und am Abend
negativ phototaktisch. Bei A. maculipennis var. messeae sind Larven,
und Puppen bei Kunstlicht im Gegensatz zu C. pipiens negativ
phototaktisch.
10. Einen messbaren Einfluss des Luftdruckes auf die Entwi¬
cklungsdauer lässt sich bei Eiern, Larven und Puppen von C. pipiens
und A. maculipennis var. messeae nicht nachweisen.
11. Es werden die Zusammenhänge zwischen Druckverminderung
und Eiablage bei A. maculipennis var. messeae gezeigt. Die kürzeste
Praeovipositionszeit beträgt 3,5 und die längste ca. 10 Tage bei
18° C.
12. Die in der Schweiz beobachteten A. maculipennis-Rassen
sind messeae, melanoon und typicus.
13. Eine Neueinteilung der maculipennis-Rassen, welche den neu
zu erstellenden Arten Rechnung trägt, wird eingeführt.
14. Es werden einige Ei-Zeichenmuster der drei in der Schweiz
vorkommenden Rassen von A. maculipennis abgebildet.
15. Der für die systematischen Fragen wichtige Schwimmkam¬
mer-Index beträgt bei
Messeae
Melanoon
Typicus
0,385
0,424
0,396
Zu andern schweizerischen Messungen ergaben sich aber beacht¬
liche Unterschiede.
16. C. pipiens und A. maculipennis weisen eine ganz deutliche
Protandrie auf.
17. Das Ausschlüpfen der Imagines aus den Eiern erfolgt haupt¬
sächlich während den Tagesstunden.
VIII.
LITERATUR-VERZEICHNIS
(1) Arnim, Frh. von, 1936. Zur Kenntnis der Temperaturabhängigkeit der Nonne
(Lymantria monacha L.). Ztschr. f. angew. Entomologie, Bd. 22.
(2) Bacot A. W., Martin C. J., 1924. The perspective Influence of Temperature
and Moisture upon the Survival of the Rat-Flea (Xenopsylla cheopis) away
from its host. Journal of Hygiene 23.
(3) Belaard J., 1858. Note relative à l'influence de la lumière sur les animaux.
Compt. rend. acad. sei. 46.
(4) Bodenheimer, F.-S., 1929. Studien zur Epidemiologie, Oekologie und Physiologie
der afrikan. Wanderheuschrecke (Schistocerca gregaria Fàrsk.). Z. f. angew.
Ent. 15.
(5) BoRRANI, E., 1937. Osservazioni
e
ricerche sulla distribuzione dei culicidi. Diss.
Losanna.
(6) BRAUN H., Caspari, E., 1939. Kann Culex pipiens bakterielle Infektionserreger
verbreiten? Schweiz. Zeitschr. f. allg. Path, und Bakt. Basel, Vol. II, Fase. 3.
(7) De Bück, A., SwelleNGREBEL, N.-H., 1931. Das Vorkommen von zwei ver¬
schiedenen Rassen des A. maculipennis als Erklärung des Anophelismus sine
Malaria. Verhandl. d. deutsch. Zoolog. Gesellschaft XXXIV.
(8) Bos, A., Nieschulz, 0., 1939. über die Fütterung von Mücken an Kanarien¬
vögeln. Zbl. Bakt. 144, Jena.
1940. Einige Weitere Versuche mit überwinternden
(9)
Exemplaren von Culex
pipiens. Zool. Anz., 129, Leipzig.
(10) CHAPMAN, R. A., 1931. Animal Ecology. McGraw-Hill, New-York—London.
(11) CHRISTOPHERS, R., 1934. A brief Guid to the Varieties of «Anopheles maculi¬
pennis ». Quart. Bull, of the Health Organisation of the League of Nations,
—
Vol. Ill, No. 4.
(12) Corradetti, A., 1934. Ricerche sulla biologia delle diverse razze di Anopheles
maculipennis. Riv. d. Malariol., Roma.
(13) Davey, W. P., 1919. Prolongation of the Life of Tribolium confusum apparently
due to small Doses of X-Rays. Journ. Exptl. Zool. 28.
(14) Eckstein, F., 1920. Die einheimischen Stechmücken. München.
(15) Eichler, W. Rassenfragen bei Malariamücken. Erscheint demnächst.
(16) ElDMANN, H., 1941. Lehrbuch der Entomologie. Paul Parey, Berlin.
(17) Eysell, Ad., 1924. Die Krankheitsüberträger und Krankheitserreger unter den
Arthropoden. Menses Hdb. d. Tropenkrankheiten, Bd. I, 3. Aufl. Leipzig.
(18) La Face, L. SuU'esistenza di razze diverse di A. maculipennis. Riv. di MalarioIogia, Vol. X, N° 6.
(19)
—
Ricerche sulla biologia del Culex pipiens. L'alimentazione
di Malariol. 5.
e
Vibemamento. Riv.
(20) Falleroni, D., 1924. Studio sugli Anopheles maculipennis nelle Paludi Pontine.
Tipographia Economica (Recanati).
(21) Gal, J., 1898. L'influence des lumières colorées sur le développement des vers à soie.
BIOLOGIE UND
VERBREITUNG
EINIGER STECHMÜCKENARTEN
145
(22) Galli-Valerio, B., 1916 Culiciden-Beobachtungen. Centrbl. f. Balrt., Bd. 78.
1917. La Distribution géographique des Anophélines en Suisse. Bull. d. Schweiz.
(23)
—
Ges.-Amtes, Nr. 39.
(24)
—
1918. Sechzehn Jahre Untersuchungen über Culiciden und Malaria. Arch. f.
Schiffs- und Tropenhygiene, 22.
1940. Contribution à l'étude des anopheles de Suisse. Mem. Soc.
vaud. Sei. nat., Vol. 6, N° 6.
Geigy, R., 1945. Malaria in der Schweiz. (Mit Verbreitungskarte) Acta tropica
Basel, Vol. 2, Nr. 1.
(25) Gaschen, H.,
(26)
(27) GrASSI, B., 1921. Razza biologica di anofeli che non punge l'uomo. Un singolarissimo caso di anofelismo senza malaria. R. C. Acad. Naz. d. Lincei. Ser. 5,
2 sem., Vol XXX (2), p. 2.
(28) GREENWOOD, M., 1906. The Effects of Rapid Decompression on Larvae. Journ.
Physiol., 35.
(29) Hackett, L. W., 1934. The present status of our knowledge of the subspecies of
«A. maculipennis». Trans. Roy. Soc. Med. and Hyg. XXVIII (2).
(30) HACKETT, L. W„ MissiroLI, A., 1935. The varieties of Anopheles maculipennis
and their relation to the distribution of malaria in Europe. Riv. di Malariologia,
Vol. XIV, No. 1,
p.
45.
über die Sprosspilze der Ösophagus-Ausstülpungen und über die
Giftwirkung der Speicheldrüsen von Stechmücken. Arch. f. Schiffs- und Tropenhyg., Bd. 32.
1930. Ober den Wärmesinn der Stechmücken bei der Eiablage. Riv. d. Malario¬
logia, IX. N° 6.
1932. Experimentelle Beiträge zur Biologie der Stechmücken. II. Zeitschr. f.
angew. En t., Bd. XIX.
1934. Experimentelle Beiträge zur Biologie der Stechmücken. V. (Wärmesinn der
A. maculipennis-Rassen bei der Eiablage). Arch. f. Schiffs- und Tropenhygiens, Bd. 38.
(31) Hecht, 0., 1928.
(32)
—
(33)
—
(34)
—
(35) Hecht, O., Goldhaber, Halberstaedter, 1943. The Effect of X-Rays on the
Developement of Insects. I. Irradiation in Larval Stage. Growth. 7.
(36) VAN Heerdt, P.-F., 1946. Einige physiologische en oecologische Problemen bij
Forficula auricularia. Diss. Utrecht.
(37) Hundertmark, A., 1938. über das Luftfeuchtigkeits-Unterscheidungsvermögen
und die Lebensdauer der drei in Deutschland vorkommenden Rassen von A. mac.
bei verschied. Luftfeuchtigkeiten. Ztschr. f. angew. Ent., Bd. 25.
(38) HUNDERTMARK, J., 1941. Versuche und Beobachtungen über das Verhalten von
A. maculipennis bei verschiedener Luftfeuchtigkeit und Temperatur. Ztschr. f.
angew.
Entom., Bd. 27, Heft 4.
(39) Hundertmark, J., Weyer, F., 1941.
Versuche über die Vorzugstemperatur einiger
Anophelen bei der Eiablage. Riv. di Malariologia, N° 4.
<40) Hunter, W. D., Pierce, W. D., 1932. Mexican Cotton Holl Weevil,
2. Session
sen.
62.
Docum. No. 305.
Kongress,
(41) Ivanova, L. V., 1940. The Influence of Temperature and the Behaviour of A.
maculipennis-Larvae. Med. Parasit. 9, 58—70, Moskau.
(42) Langeron, M., 1942. Précis de Microscopic Masson, Paris.
(43) Laurens, H., Hooke, H. D., 1920. Studies on the Relative Physiological Value
of Spectral Lights. Journ. Exptl. Zool. 30.
(44) Leeson, H. S., 1932. The Effect of Temperature and Humidity
of certain unfed Rat-Fleas. Parasitology 24.
(45) Lutz, F.-E., 1924. A Study of Ultraviolet in Relation
of Insects. Anm. N. Y. Acad. Sei., 29.
10
to the
upon the
Survival
Flowervisiting Habits
146
WILLI
BÜTTIKER
(46) Marshall, J., 1938. British Mosquitoes. British Museum of Natural History.
London.
(47) MARTINI, E., 1921. Zur Bionomie
48)
49)
(50)
—
—
—
Stechmücken. Arch. f. Schiffs- und
unserer
Tropenhygiene, Bd. 25.
1925. über die Wärmesummenregel. Ztschr.
angew. Entomologie, 4.
1928. über die Kettenlinie und die Exponentialkurve überhaupt als Bilder für
die Abhängigkeit der Entwicklungsdauer von der Wärme. Ztschr. f. angewandte
Entomologie, 14.
1931. Die Rassenf rage bei Anopheles maculipennis. Arch. f. Schiffs- und Tropen¬
hygiene. Vol. 35.
(51) Martini, E., Teubner. Ober das Verhalten von Stechmücken bei verschiedenen
Temperaturen und Luftfeuchtigkeiten. Arch. f. Schiffs- und Tropenhyg.
Band 37, Beiheft 1.
(52) Martini, E., 1946. Lehrbuch der Medizinischen Entomologie, 3. Aufl., Fischer,
Jena.
(53) Matheson, R., 1944. Handbook, of the Mosquitoes of North America. Comstock,
Ithaca-New-York.
(54) Mavor, J. W., 1947. A Comparison of the Susceptibility to X-Rays of Drosophila
melanogaster at Various Stages of the Life-cycle. Journ. Exptl. Zool., 47.
(55) DE Meillon, B., 1935. Key to the full grown larvae and adults of South African
anopheline mosquitoes. S. A. lust. Med. Res. Nr. 28.
(56) Missiroli, A., Hackett, L. W., 1927. La regressione spontanea delta malaria
in alcune regioni d'ltalia. Riv. di Malariologia, Vol. VI, N° 2.
(57) Missiroli, A., 1935. Une nouvelle variété d'Anophèles maculipennis, trouvée à
2000 pieds : A. maculipennis var. pergusiae. Ann. d'Igiene, Vol. 45, p. 333.
(58) Missiroli, A., Hackett, L., Martini, E., 1933. Le razze di Anopheles maculi¬
pennis
e
la loro importanza nella distribuzione délia malaria in alcune regioni
d'Europa. Riv. di Malariologia, vol. 12.
(59) Nieschulz, 0., Bos, A., 1931. über die Larvenentwicklung von Culex pipiens
bei höheren Temperaturen. Ztschr. f. Parasitenkd., 3. Band, 4. Heft.
(60) NlESCHULZ, 0., 1933. über die Temperaturbegrenzung der Aktivitätsstufen von
Stemoxys calcitrans. Ztschr. f. Parasitenkd., 6. Bd., 2. Heft.
1933. über die Bestimmung der Vorzugstemperatur von Insekten. Zool. Anzeiger
(61)
—
(62)
—
21.
1934. Über die Vorzugstemperatur
Entomologie, Bd. 21, Heft 2.
von
Stemoxys calcitrans. Ztschr. f.
angew.
(63) Nieschulz, 0., Toit, R. DU, 1935. über die Temperaturabhängigkeit der Aktivität
und die Vorzugstemperatur von Musca domestica und Fannia canicular is.
Zoolog. Anzeiger, 110.
(64) Packard, C, 1926. The Measurement of Quantitative Biological Effects of X-Rays.
Journ. Cancer Research, 10.
(65) Parker, J. R., 1930. Some Effects of Temperature and Moisture
mexicanus. Univ. of Minn.
(66) P ARM AN, D. C,
Ag. Expt. Sta. Bull. 223.
1920. Observations
Activity. Journ. Econ. Ent., 13.
on
the
upon
Melanoplus
Effect of Storm Phenomena
(67) PlCTET, A.,_ 1918. Les Eclosions de Papillons
Soc. lepidopt. Genève, Vol. IV., Fasc. 1.
et
la Pression
on
Insect
barométrique. Bull.
(68) Reichenow, E., 1932. Die Entwicklung von Proteosoma circumflexum in Theobaldia annulata, nebst Beobachtungen über das Verhalten anderer Vogelplasmodien in Mücken. Jen. Ztschr. f. Naturw., 67. Bd., Festschrift L. Plate.
(69a) Roubaud, E., 1921. La différenciation des races zootropiques d'anophèles et la
régression spontanée du paludisme. Bull. Soc. Path. Exot. I, 14.
BIOLOGIE UND VERBREITUNG EINIGER
(69b)
(70)
STECHMÜCKENARTEN
147
RoUBAUD E., 1934. Un type racial nouveau de I'Anopheles maculipennis. Bull.
Soc. Path. Exot. T. XXVII, n° 8, p. 737.
1935. Variété nouvelle de VAnopheles maculipennis au Maroc: A. maculipennis
var. sicaulti (nov. var.). Bull. Soc. Path. Exot. T. XXVIII, n° 2, p. 107.
—
(71) RoUBAUD, E., TREILLARD, M., 1936. Sur
une
variété portugaise de
l'Anopheles
maculipennis (groupe atroparvus). Bull. Soc. Path. Exot. T. XXIX, p. 726.
(72) Roubaud, E., Colas-Belcour, J., Treillard, M., 1937. Hybridation naturelle
de deux biotypes considérés comme amixiques de l'Anopheles maculipennis (var.
typicus et atroparvus). Bull. Soc. Path. Exot. T. XXX. n° 7.
(73) Roubaud. E., Meyer, J., 1942. La nourriture larvaire n'influe pas le développe¬
ment de l'autogenese chez les races de Culex pipiens spécifiquement anautogènes.
Ann. Parasit, hum. comp. 12.
(74) Roubaud, E., Greiner, P., 1942. Quelques observations sur l'aliment des larves
de culicides (facteurs B et substances protéiques). Bull. Soc. Path. Exot. 35,
Paris.
(75) Rudolfs, W., Chemotropism of Mosquitoes. New Jersey Agricultural Experiment
Stations, March 1922, Bulletin 367.
(76) Schaudinn, F., 1904. Generations- und Wirtswechsel bei Trypanosomen und
Spirochaeten. Arb. a. d. Kaiserl. Gesundheitsamte, Bd. 20.
(77) Schwietert, K., 1935. Hat die Malaria noch heute für Deutschland eine Bedeu¬
tung? Diss. Hamburg.
(78) Sergent, E., 1903. C. R. Soc. Biol.. LV.
(79) Shelford, V. E., 1927. An Experimental Investigation of the Relations of the
Coddling Moth to Weather and Climate. III. Nat. Hist. Survey Bull., Vol. XVI.
(80) Steinhaus, E., 1946. Insect Microbiology. Comstock, Ithaca-New York.
(81) Swellencrebel, A.-H. Eine Vergleichung der Änderungen, welche die jährl.
Zahlen der Malariakranken und Anophelen in den letzten Jahren in der Umge¬
bung Amsterdams aufwiesen. Abh. a.d. Geb. d. Auslandkunde, Hamburg,
Bd. 26 (Festschrift Nocht).
(82) Swellencrebel, A. H., de Buck, A., Schoute, E. On Anophelism without
malaria around Amsterdam. Proc Kon. Akad.
XXX (1), p. 61.
v.
Wetensch.
te
Amsterdam,
(83) Tate, P., Vincent, H., 1936. The Biology of autogenous and anautogenous Races
of Culex pipiens. Parasitology, 1936.
(84) Teubner, E, Martini, E., 1933. über das Verhalten von Stechmücken, besonders
von Anopheles maculipennis, bei verschiedenen Temperaturen und Luftfeuchtig¬
keiten. Beiheft, Arch. f. Schiffs- u. Tropenhygiene, Bd. 37.
(85) VAN Thiel, P.-H., 1927. Sur l'origine des variations de taille de Anopheles macu¬
lipennis. Bull. Soc. Path. Exot. XX, n° 4.
1926. Maxillenzahnzahl und Flügellänge bei Anopheles maculipennis. Arch.
f. Schiffs- u. Tropenhygiene XXX, (1).
(87) Thomsen, E., Thomsen, M., 1937. über das Thermopraeferendum der Larven
einiger Fliegenarten. Ztschr. f. vergl. Physiologie, 24. Bd., 3. Heft, Berlin.
(88) TrÖnDLE, A., 1910. Der Einfluss des Lichtes auf die Plasmahaut. Jahrb. wiss.
Bot., 48.
(86)
—
(89) Ulmann, E., 1941.
Die regulatorische
Bedeutung der Bevölkerungsdichte für das
natürliche Gleichgewicht einer Art. Experimentalversuche mit
autogenicus Roub. Ztschr. f. angew. Entomologie, 28, Berlin.
Culex pipiens
(90) Uvarov B.-P., 1931. Wetter und Klima in ihren Beziehungen zu den Insekten.
Ztschr. f. angew. Entomologie, Bd. 17.
(91) Wellington, W. S., 1945. Conditions governing the Distribution of Insects in
the free Atmosphere. The Canadian Entomologist, Nr. 1, Jan.
148
WILLI
BÜTTIKER
(92) Wesenberg-Lund, C, 1920—1921. Contribution to the Biology of the Danish
Culiddae. Mem. Acad. Roy. Sc. et Lettr. Danemark, VII.
(93) Weyer, F., 1934. Der Einfluss der Larvenernährung auf die Fortpflanzungs-Phy¬
siologie verschiedener Stechmücken. Arch. f. Schiffs- und Tropenhygiene,
Nr. 9, Leipzig.
1936. Einige Erfahrungen bd der Aufzucht von Stechmückenlarven. Zbl.
(94)
—
Bakt., 136.
(95)
(96)
(97)
(98)
(99)
1942. Bestimmungsschlüssel für die Anopheles-Wdbchen und -Larven in Europa,
Nordafrika und Westasien. Merkbl. Med. wichtiger Insekten, Tropeninstitut
Hamburg, 12.
Wiesmann, R., Jenny, J., 1938. Vollautomatischer Elektro-Brückenthermostat
« rVädenswih. Landw.
Jahrbuch d. Schweiz.
WlGGLESWORTH, V. B., 1942. The Principles of Insect Physiology, II. Edition
Methuen & Co., London.
ZwÖLFER, W., 1931. Studien zur Oekologie und Epidemiologie der Insekten. 1.
Kieferneule, Panolis flammea. Zeitschrift für angewandte Entomologie, Bd. 17.
1932. Methoden zur Regulierung von Temperatur und Luftfeuchtigkeit. Ztschr.
—
—
f. angew. Entom., Bd. XIX.
Nachtrag
(100) Brues, CT., 1946. Insect Dietary. Harvard.
(101) Pratolongo, U., 1947. Anticrittogamid ed insettiddi. Trattati di Agricultura,
Roma.
(102) Ungureanu E., Shute P.-J., 1947. The Value of the Wing Scales as an Aid
to the Taxonomy of adult Anopheles maculipennis. The Roy. Ent. Soc.,
London, Vol. 16, Nr. 7-8.
Imprimerie
La
Concorde, Lausanne (Suisse).
CURRICULUM
Am 5. August 1921
VITAE
wurde ich, Willi Büttiker,
von
Ölten, in
Ölten geboren. Ich bin Sohn des Paul Büttiker, Bankdirektor, und
seiner Gattin Elsa
geb.
von
Arx, wohnhaft in Rheinfelden.
In Rheinfelden besuchte ich 1928-1933 die Gemeindeschule und
1933-1937 die Bezirksschule.
der
1937
Kantonsschule in Aarau ein.
prüfung (Typus C)
trat
ich in die Oberrealschule
Nach bestandener
im Herbst 1940 Hess ich mich
an
Maturitäts¬
der Eidgenös¬
sischen Technischen Hochschule, Abteilung für Naturwissenschaften,
immatrikulieren.
Frühjahr 1942 bestand ich das erste Vordiplom in Mathe¬
matik, Geologie und Chemie und 1943 das zweite Vordiplom in
Physik, Geographie une Pétrographie.
Im
Im Sommer 1945 schloss ich
—
durch Militärdienst verzögert
das naturwissenschaftliche Studium ab. Die Promotionsarbeit
ich
1945
begann
Herrn
Prof.
Leitung von
Dr. Schneider-Orelli im Entomologischen Institut der E. T. H., wo
ich seit diesem Zeitpunkt bis zum übertritt am 1. Januar 1948 in
die Firma J; R. Geigy A. G., Basel, als Assistent tätig war.
im
November
—
unter
Zürich, Ende Dezember 1947.
der