Document
Werbung

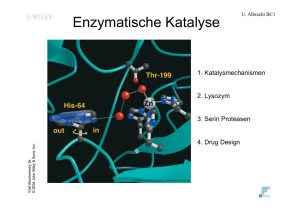
U. Albrecht BC1 Voet Biochemistry 3e © 2004 John Wiley & Sons, Inc. 1. Überblick 2. Stoffwechselquellen für Acetyl-Coenzym A 3. Enzyme des CitronensäureCyclus 4. Regulation des Citronensäure-Cyclus 5. Amphibole Natur des Citronensäure-Cyclus U. Albrecht BC1 1. Überblick Der Citronensäure-Cyclus kommt in Eukaryonten und Prokaryonten vor. Wird auch Tricarbonsäure-Cyclus (TCA-Cyclus) oder Krebs-Cyclus genannt. Voet Biochemistry 3e © 2004 John Wiley & Sons, Inc. Die Acetylgruppe eines Acetyl-CoA-Moleküls wird zu 2 Molekülen CO2 oxidiert und die Energie wird gespeichert um später ATP zu Erzeugen. Reaktionen 8 Enzyme -> es entstehen: 2 CO2 3 NADH -> in oxidative Phosph. 1 FADH2-> „ „ „ 1 GTP U. Albrecht BC1 Tertiärer Alkohol -> schwer oxidierbar Voet Biochemistry 3e Page 766 © 2004 John Wiley & Sons, Inc. Oxalacetat wird regeneriert -> unendliche Zahl von Acetylgruppen Kann mit einem Oxalacetat Oxidiert werden. Sekundärer Alkohol -> leichter oxidierb. U. Albrecht BC1 2. Stoffwechselquellen für Acetyl-Coenzym A Thioester KohlenhydratFettsäurenAminosäurenabbau Energiereiche Bindung Amidbindung Voet Biochemistry 3e Page 768 © 2004 John Wiley & Sons, Inc. Vitamin Chemische Struktur von Acetyl-Coenzym A A. Pyruvat-Dehydrogenase-Multienzymkomplex U. Albrecht BC1 Pyruvat Vorstufe von Acetyl-CoA. Multienzymkomplex Pyruvate decarboxylase multienzym Complex (PDC) katalysiert. Multienzymkomplex= nicht kovalent assoziierte Enzyme die 2 or mehr sequentielle Reaktionen katalysieren. Komplex in E. coli beschrieben. PDC Enzyme: Voet Biochemistry 3e Page 769 © 2004 John Wiley & Sons, Inc. Pyruvate dehydrogenase (E1) Dihydrolipoyl transacetylase (E2) Dihydrolipoyl dehydrogenase (E3) intact complex “core” complex U. Albrecht BC1 Structural organization of the E. coli PDC. Voet Biochemistry 3e Page 769 © 2004 John Wiley & Sons, Inc. E2 core 24 subunits E1 E3 Combined 60 subunit complex U. Albrecht BC1 Voet Biochemistry 3e Page 769 © 2004 John Wiley & Sons, Inc. Comparison of the X-ray structures of the dihydrolipoyl transacetylase (E2) cores of PDCs The cubic Azobacter vinelandii E2 core. The dodecahedral B. stearothermophilus E2 core. Gram-negative Bakterien Gram-positive Bakterien und Eukaryontische Mitochondrien (20 E2 trimere, 30 E1 tetramere, 12 E3 dimere Mitochondrien von Gram-Positiven ? Mit. Haben aber doppelmembran Wie gram-negative.-> Sequenzvergleiche Ricketsia prowazekii (typhus) -> gram neg. aber dodecahedral U. Albrecht BC1 Multienzymkomlexe sind katalytisch effizient 1. Enzymatische Reaktionsraten sind limitiert durch die Frequenz mit der das Substrat mit dem Enzym kollidiert. In einem Multienzymkomplex wird die Diffusion zwischen den aktiven Zentren minimiert -> Ratenverbesserung Voet Biochemistry 3e © 2004 John Wiley & Sons, Inc. 2. Kanalisierung metabolischer Zwischenprodukte wird möglich -> Verringerung von Nebenreaktionen. 3. Reaktionen können koordiniert kontrolliert werden. U. Albrecht BC1 Acetyl-CoA Bildung geschieht in 5 Schritten Voet Biochemistry 3e Page 770 © 2004 John Wiley & Sons, Inc. Pyruvate + CoA + NAD+ -> acetyl-CoA + CO2 + NADH The five reactions of the PDC. U. Albrecht BC1 Voet Biochemistry 3e Page 771 © 2004 John Wiley & Sons, Inc. The Coenzymes and Prosthetic Groups of Pyruvate Dehydrogenase. Reaktion 1 Reaktion 2 Reaktion 4 Reaktion 3 Voet Biochemistry 3e © 2004 John Wiley & Sons, Inc. wrong goes to reaction 2 Reaktion 5 Voet Biochemistry 3e Page 771 © 2004 John Wiley & Sons, Inc. U. Albrecht BC1 Struktur von Lipoamid Der Lipolysyl-Arm befördert die Zwischenprodukte von einer Untereinheit des Enzyms zur nächsten Wie werden Zwischenprodukte von einem Enzym zum nächsten weitergegeben? Liposylyl-Arm wie eine Liane -> schwingt von E1 nach E3 Voet Biochemistry 3e © 2004 John Wiley & Sons, Inc. E2 Lianen schwingen auch gegeneinander -> gegenseitige Acetylierung -> Jede E1 UE kann nehrere E2 UE acetylieren und jede E3 UE kann mehrere E2 reoxidieren. U. Albrecht BC1 Voet Biochemistry 3e Page 773 © 2004 John Wiley & Sons, Inc. Die Struktur von E2 der PDC Domain structure of the dihydrolipoyl transacetylase (E2) subunit of the PDC. U. Albrecht BC1 Coenzym A Voet Biochemistry 3e Page 773 © 2004 John Wiley & Sons, Inc. lipoamid X-Ray structure of a trimer of A. vinelandii dihydrolipoyl transacetylase (E2) catalytic domains. U. Albrecht BC1 Voet Biochemistry 3e Page 773 © 2004 John Wiley & Sons, Inc. Lys Seitenkette zu welcher die Lipoylgruppe gebunden wird ist in einem type 1 β turn. The NMR structure of the A. vinelandii dihydrolipoyl transacetylase (E2) lipoyl domain. U. Albrecht BC1 Voet Biochemistry 3e Page 773 © 2004 John Wiley & Sons, Inc. Bindungstellen für E1 und E3 The NMR structure of the peripheral subunit-binding domain from B. stearothermophilus E2. U. Albrecht BC1 E1 E2 Voet Biochemistry 3e Page 774 © 2004 John Wiley & Sons, Inc. E2 catalytic domain - E1 binding d. E3 Electron microscopy–based images of the bovine kidney pyruvate dehydrogenase complex at ~35 Å resolution. Arsenverbindungen sind giftig weil sie Lipoamid maskieren U. Albrecht BC1 Arsenverbindungen sind toxisch da sie kovalente Verbindungen mit Sulfhydrylgruppen eingehen können. -> Inaktivierung lipoamidhaltiger Enzyme Pyruvat-Dehydrogenase α-Ketoglutarat-Dehydrogenase Voet Biochemistry 3e © 2004 John Wiley & Sons, Inc. -> Unterbrechung der Atmung Organische Arsenvergiftungen toxischer für Mikroorganismen als für den Menschen -> zu Beginn des 20. Jahrhunderts Syphilis-Behandlung und TrypanosomenBehandlung -> = erste Antibiotika, aber schwere Nebenwirkungen. Anektote Napoleon: Arsen in Haar -> Vergiftet? - grüne Tapeten mit Pigment KupferArsenat (CuHAsO4) .-> in feuchtheissem Klima -> Pilze konvertieren dies zu Trimethyl Arsin ((CH3)3As) welches flüchtig und sehr giftig ist. U. Albrecht BC1 Die Struktur von E1 Neben PDC noch andere sehr ähnliche Multienzymkomplexe: α-Ketoglutarat dehydrogenase Komplex (Reaktion 4 im Citrat Zyklus) Verzweigte Ketten α-ketosäure dehydrogenase Komplex (degradation von Ile, Leu und Val) Alle in Familie von 2-Ketosäure Dehydrogenase mit gleichem E3 enzym und spezifischen E1 und E2 Struktur jedoch bei allen ähnlich. α-Untereinheiten β-Untereineheiten Voet Biochemistry 3e Page 776 © 2004 John Wiley & Sons, Inc. TPP cofaktoren an Bindungsstelle von Untereinheiten. X-Ray structure of E1 from P. putida branched-chain α-keto acid dehydrogenase. (a) The α2β2 heterotetrameric protein. Voet Biochemistry 3e Page 776 © 2004 John Wiley & Sons, Inc. U. Albrecht BC1 X-Ray structure of E1 from P. putida branched-chain α-keto acid dehydrogenase. (b) A surface diagram of the active site region. U. Albrecht BC1 B. Mechanismus der Dihydrolipoyl Dehydrogenaase (E3) 2 electron acceptor sites: Voet Biochemistry 3e © 2004 John Wiley & Sons, Inc. Fad and disulfid bond U. Albrecht BC1 Röntgenstruktur von Dihydrolipoamid Dehydrogenase 2 Subunits Interface Domäne Zentral Domäne NAD+ Voet Biochemistry 3e Page 777 © 2004 John Wiley & Sons, Inc. FAD NAD+ Bindungsdomäne FAD Bindungsdomäne X-Ray structure of dihydrolipoamide dehydrogenase (E3) from P. putida in complex with FAD and NAD+. (a) The homodimeric enzyme. Voet Biochemistry 3e Page 777 © 2004 John Wiley & Sons, Inc. U. Albrecht BC1 X-Ray structure of dihydrolipoamide dehydrogenase (E3) from P. putida in complex with FAD and NAD+. (b) The enzyme’s active site region. Katalytischer Reaktionszyklus der Dihydrolipoyl Dehydrogenase Voet Biochemistry 3e Page 778 © 2004 John Wiley & Sons, Inc. 2 substrates U. Albrecht BC1 Charge Transfer complex U. Albrecht BC1 Charge Transfer Complex FAD S Voet Biochemistry 3e Page 780 © 2004 John Wiley & Sons, Inc. NAD+ The complex of the flavin ring, the nicotinamide ring, and the side chain of Cys 63 observed in the X-ray structure of human erythrocyte glutathione reductase (very similar to Dihydrolipoyl dehydrogenase). The reaction transferring an electron pair from dihydrolipoyl dehydrogenase’s redox-active disulfide in its reduced form to the enzyme’s bound flavin ring. U. Albrecht BC1 C. Kontrolle der Pyruvat-Dehydrogenase A) Kontrolle durch Produktinhibition Voet Biochemistry 3e Page 781 © 2004 John Wiley & Sons, Inc. PDC reguliert den Eintritt von aus Kohlenhydraten stammenden Acetyleinheiten In Citronesäure Cyclus. E1 irreversibler Schritt -> genaue Regulation da sonst keine anderen Wege zur Synthese von Acetyl-CoA in Säugern. Produkt Inhibition: NADH und Acetyl-CoA treiben die reversiblen Reaktionen E2 und E3. U. Albrecht BC1 B) Kontrolle durch Phosphorylierung und Dephosphorylierung Voet Biochemistry 3e Page 781 © 2004 John Wiley & Sons, Inc. Phospohatase und Kinase Phospohorylierung eines Ser Restes in E1 3. Enzyme des Citronensäure-Cyclus U. Albrecht BC1 A. Citrat-Synthase Kondensation von Acetyl-CoA mit Oxalacetat. -> Induced Fit Enzym, sequenzieller Mechanism. Oxalacetat bindet zuerst -> Konformationsänderung -> Aectyl-CoA bindet. Voet Biochemistry 3e Page 782 © 2004 John Wiley & Sons, Inc. Oxalacetat Bindungsstelle Conformational changes in citrate synthase. (a) Space-filling drawing showing citrate synthase in the open conformation. Conformational changes in citrate synthase. (b) Space-filling drawing showing citrate synthase in the closed, substrate-binding conformation. Mechanismus und Stereochemie der Citrat-Synthase Reaktion U. Albrecht BC1 Voet Biochemistry 3e Page 783 © 2004 John Wiley & Sons, Inc. Aldol-Claisen-Esterkondensation in 3 Schritten U. Albrecht BC1 B. Aconitase Voet Biochemistry 3e Page 784 © 2004 John Wiley & Sons, Inc. Aconitase katalysiert die reversible Isomerisierung von Citrat und Isocitrat mit cis-Aconitat als Zwischenprodukt. U. Albrecht BC1 Fluorocitrat hemmt Aconitase Fluoroacetat sehr giftig (LD50 bei Ratten= 0.2 mg/kg Körpergewicht) Kommt in Blättern bestimmter Pflanzen Aus Afrika, Australien und Südamerika vor. Selbst keine toxische Wirkkung wird aber In den Zellen umgewandelt zu Fluorocitrat. Voet Biochemistry 3e © 2004 John Wiley & Sons, Inc. Dies hemmt Aconitase spezifisch und Inhibiert den Citrat transport durch die Mitochondrienmembran. U. Albrecht BC1 C. NAD+ -abhängige Isocitrat-Dehydrogenase Isocitrat-Dehydrogenase katalysiert die oxidative Decarboxylierung von Isocitrat zu α-Ketoglutarat, Wobei erstmalig im Citronensäure-Cyclus CO2 und NADH gebildet wird. Voet Biochemistry 3e Page 785 © 2004 John Wiley & Sons, Inc. In Säugern 2 verschiedene Formen: 1) Citronensäure-Cyclus Enzym in Mitocheondrien, NAD+ als Coenzym. 2) in Mitochondrien und Cytosol, NADP+ als Coenzym. = sek. Alkohol = Keton Konnte noch nicht bewiesen werden U. Albrecht BC1 D. α-Ketoglutarat-Dehydrogenase α-Ketoglutarat-Dehydrogenase katalysiert die oxidative Decarboxylierung einer α-Ketocarbonsäure, wodurch das zweite CO2 und das zweite NADH des CitronenSäure-Cyclus grei werden. Reaktion ähnlich der von Pyruvat-Dehydrogenase-Multienzymkomplex. 3 Untereinheiten: Voet Biochemistry 3e © 2004 John Wiley & Sons, Inc. α-Ketoglutarat-Dehydrogenase (E1) Dihydrolipoyl-Transsuccinylase (E2) Dihydrolipoyl-Dehydrogenase (E3) E3 ist identisch mit E3 von PDC. U. Albrecht BC1 E. Succinyl-CoA-Synthase Hydrolysiert die energiereiche Verbindung Succinyl-CoA unter Kopplung an die Synthese eines Energiereichen Nucleosidtriphospphates. In Säugern -> GTP, in Pflanzen ATP. Beachte: Enzymnahmen können auch die Rückreaktionen benennen (wie hier). Voet Biochemistry 3e Page 787 © 2004 John Wiley & Sons, Inc. Durch Succinyl-CoA-Synthase katalysierte Reaktionen: Bildung eines succinyl phosphates, ein “high-energy” mixed anhydride. Voet Biochemistry 3e Page 787 © 2004 John Wiley & Sons, Inc. U. Albrecht BC1 Formation of phosphoryl–His, a “high-energy” intermediate. U. Albrecht BC1 Voet Biochemistry 3e Page 787 © 2004 John Wiley & Sons, Inc. Transfer of the phosphoryl group to GDP, forming GTP. GTP Bildung äquivalent zu ATP Bildung da ineinander umwandelbar. GTP + ADP GDP + ATP freie Enthalpie = 0 U. Albrecht BC1 F. Succinat-Dehydrogenase Dehydrierung von Succinat zu Fumarat Voet Biochemistry 3e © 2004 John Wiley & Sons, Inc. Enzym gehemmt durch Malonat (ein C atom weniger als Succinat). U. Albrecht BC1 FAD ist kovalent an die Succinat-Dehydrogenase gebunden Normalerweise FAD nicht kovalent in Enzymen Gebunden Aber in Succinat dehydrogenase schon -> Problem:Wie wird FADH2 reoxidiert damit Enzym wieder funktionieren kann? Voet Biochemistry 3e Page 787 © 2004 John Wiley & Sons, Inc. Das Enzym wird über die Elektronentransportkette reoxidiert. Einziges and innere Mitochondrienmembran gebundenes Enzym des Citronensäure-Cyclus -> Auch als Complex II der Oxidationskette bekannt. Reoxidiert durch Coenzym Q U. Albrecht BC1 G. Fumarase Voet Biochemistry 3e Page 788 © 2004 John Wiley & Sons, Inc. Es ist nicht klar ob der Mechanismus über ein Carbanion oder einen Carbokation ÜbergangsZustand geht. U. Albrecht BC1 Voet Biochemistry 3e © 2004 John Wiley & Sons, Inc. H. Malat-Dehydrogenase Regeneration von Oxalacetat U. Albrecht BC1 4. Regulation des Citronensäure-Cyclus Diese Enzyme arbeiten weit weg vom Gleichgewichtszustand unter natürlichen Bedingungen -> Voet Biochemistry 3e © 2004 John Wiley & Sons, Inc. Diese Enzyme bestimmen Geschw. des Citratzyklus -> Kontrollpunkte U. Albrecht BC1 Regulation durch: 1) Verfügbarkeit der Substrate 2) Produkthemmung 3) Hemmung durch Zwischenprodukte Da Sauerstoffverbrauch, NADH Reoxidation und ATP-Produktion eng miteinander gekopplet sind, muss der Citronensäure-Cyclus über Rückkopplungsmechanismen, die seine NADH-Produktion auf den EnergiverBrauch abstimmen, reguliert werden. Voet Biochemistry 3e Page 791 © 2004 John Wiley & Sons, Inc. Wichtigste Regulatoren: -) Acetyl-CoA, Oxalacetat, NADH +) ADP, Ca2+ Keine komplizierte Regulation über Hormone (vrgl. Glykogenabbau), da Keine Speicherstoffe gebildet werden. Produkte kurzfristig direkt an Atmung gekoppelt. 5. Amphibole Natur des Citronensäure-Cyclus U. Albrecht BC1 1. Glucose-Biosynthese 2. Lipid-Biosynthese 3. Aminosäure-Biosynthese Voet Biochemistry 3e Page 793 © 2004 John Wiley & Sons, Inc. 4. Porphyrin-Biosynthese 5. Oxidation von Aminosäuren
![Figure 14-1 Plot of ln[A] versus time for a first](http://s1.studylibde.com/store/data/011570233_1-88128775b0e44d7bbbef6452196ac987-300x300.png)