Die MENDELschen Regeln
Werbung

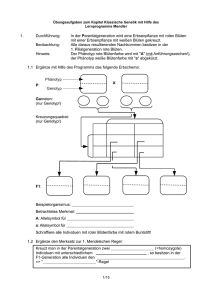
Klassische Genetik Pollen F1 P P 21.1 Die Ergebnisse der Mendelschen Experimente 1 Die Mendelschen Regeln P O RT R ÄT Seitdem die Menschen sich mit der Züchtung von Tieren und Pflanzen beschäftigten, musste ihnen klar gewesen sein, dass bestimmte Merkmale von Generation zu Generation vererbt werden. Ab dem 18. Jahrhundert sind Experimente zum Beispiel von KÖLREUTER und GÄRTNER belegt, die das Wie und Warum der Vererbung aufhellen sollten. Trotz vieler tausend Versuche war den Forschern kein Erfolg vergönnt. Diesen sollte erst der Mönch Johann Gregor MENDEL haben. Er vermied drei entscheidende Fehler seiner Vorgänger: Erstens kreuzte er nur verschiedene Rassen einer Art miteinander und überschritt bei seinen Experimenten die Artgrenze nicht, zweitens beschränkte er sich bei den Erbgängen auf wenige zu beobachtende Merkmale und drittens arbeitete er mit jeweils reinerbigen Elternpflanzen. Diese geben ein bestimmtes Merkmal, wie die Samenfarbe, immer nur in einer Ausprägung, zum Beispiel gelb, weiter. MENDEL überprüfte verschiedene Erbsenrassen über zwei Jahre auf die erforderliche Reinerbigkeit, indem er sie im Gewächshaus sechs Generationen lang züchtete. In einem Experiment beschäftigte MENDEL sich mit der Farbe der Erbsensamen. Für die Johann Gregor Mendel (1822–1884) 20 Klassische Genetik Johann MENDEL wurde im mährischen Heinzendorf (in Tschechien) geboren. Aus wirtschaftlicher Not trat er 1843 dem Augustinerstift in Brünn bei. Er erhielt den zusätzlichen Namen Gregor. Da er für die Seelsorge ungeeignet war, durfte er sich Lehre und Forschung widmen. Bei Kreuzungsexperimenten mit Saaterbsen entdeckte er Regelmäßigkeiten bei der Vererbung, die 1866 veröffentlicht wurden. Erbanlage oder das Gen dieses Merkmals gibt es zwei verschiedene Zustandsformen, die heute Allele genannt werden und die gelbe oder grüne Samenfarbe festlegen. MENDEL entfernte in einem Experiment zuerst die Staubgefäße der Blüten gelbsamiger Pflanzen. Dann bestäubte er mit einem Pinsel die Narben dieser Blüten mit Pollen einer grünsamigen Erbse. Anschließend schützte er die Blüten durch kleine Stoffsäckchen vor Fremdbestäubung. Die Ausgangspflanzen einer solchen Kreuzung nennt man Eltern- oder Parentalgeneration, abgekürzt „P“. Aus diesem Ansatz erhielt MENDEL nur gelbsamige Pflanzen, die 1. Tochter- oder Filialgeneration, abgekürzt „F1“. MENDEL wollte nun wissen, ob die Erbinformation für die grüne Samenschale ganz verschwunden war und überließ deshalb die F1 sich selbst. Erbsen sind Selbstbestäuber. In der sich nun entwickelnden 2. Filialgeneration (F2) erhielt MENDEL wieder gelbe und grüne Erbsensamen. Er schloss daraus, dass das Allel für die grüne Samenfarbe in der F1 auch noch vorhanden war, aber von dem Allel für gelbe Erbsen in der Ausprägung überlagert wurde. Er nannte das Allel für gelbe Erbsen deshalb dominant (lat. dominare, beherrschen) über das für grüne und kürzte es mit einem großen „G“ ab. Das Allel für grüne Erbsen nannte er entsprechend rezessiv (lat. recedere, sich zurückziehen) gegenüber dem Allel für gelbe und kürzte es mit einem kleinen „g“ ab. In den Zygoten, aus denen die F1 entstanden ist, mussten jeweils zwei verschiedene Allele zu dem Merkmal Samenfarbe vorliegen. Dieser Zustand wird heterozygot (gr. heteros, verschieden) genannt. Analog benutzt man für reinerbige Lebewesen den Begriff homozygot (gr. homos, gleich). Aus dem obigen Versuchsergebnis schloss MENDEL weiterhin, dass der Erscheinungstyp eines Lebewesens, der Phänotyp, nicht der Summe der enthaltenen Gene, dem Vererbungs- oder Genotyp, entsprechen muss. MENDELs Genialität lag nun darin, dass er die alle F1 Selbstbestäubung F2 F1 F 6022 2001 1 F2 Pollen Selbstbestäubung F1 P P F2 F 315 101 32 108 1 F2 erhaltenen Samen der F2 zahlenmäßig auswertete: Es entstanden 6022 gelbe sowie 2001 grüne Erbsensamen, was etwa einem ganzzahligen Verhältnis von 3 : 1 entspricht. Erbgänge, bei denen man jeweils ein Merkmal betrachtet, werden monohybrid (gr. monos, einzig; hybrida, Mischling) genannt. MENDEL kreuzte in einem dihybriden (gr. di- = zwei) Erbgang auch Erbsenrassen, die sich in zwei Merkmalen unterschieden, zum Beispiel Pflanzen mit gelb-runden Samen mit Pflanzen mit grün-runzeligen Samen. In der 1. Filialgeneration beobachtete er nur gelbrunde Samen. In der 2. Filialgeneration gab es die neuen Merkmalskombinationen grün-runde sowie gelb-runzelige Samen. Wieder zählte MENDEL die Samen aus und kam auf 315 mit dem Phänotyp gelb-rund und 108 mit grün- rund. Außerdem erhielt er 101 Samen des Phänotyps gelb-runzelig und 32 grün-runzelige Erbsen. MENDEL war von der Gültigkeit seiner Ergebnisse überzeugt und verschickte zusätzlich zu der Veröffentlichung Sonderdrucke seiner Arbeit. Zu seiner großen Enttäuschung blieben die Versuche unbeachtet. So wandte er sich nach seiner Wahl zum Abt des Klosters im Jahre 1868 anderen Aufgaben zu. Erst um 1900 wurden MENDELs Versuche wieder aufgegriffen. TSCHERMAK, CORRENS und DE VRIES veröffentlichten Arbeiten mit eigenen Kreuzungsergebnissen und bezogen sich auf die Schlussfolgerungen des Mönchs aus Brünn. CORRENS war es auch, der die Ergebnisse des Mönchs in prägnante Sätze fasste und sie „MENDELsche Regeln“ nannte. Klassische Genetik 21 GG gg G G g g Parentalgeneration P Gameten Gg 1. Filialgeneration F1 Uniformität Gg Gg Gg G g G g G g G GG Gg g Gg gg Gameten 2. Filialgeneration F2 Spaltung Aus solch einem Kreuzungsansatz leitet sich die 2. MENDELsche Regel ab, die auch Spaltungsregel heißt: Kreuzt man die Individuen der F1 untereinander, so ist die F2 nicht uniform, sondern die Merkmale spalten in bestimmten Zahlenverhältnissen auf, und zwar beim dominant-rezessiven Erbgang im Phänotyp im Verhältnis 3 : 1 und im Genotyp im Verhältnis 1 : 2 : 1. 22.1 Kombinationsquadrat eines monohybriden Erbganges GGRR ggrr GR GR gr gr Gg Rr Gg Rr Gg Rr Gg Rr GR Gr gR gr GR Gr gR gr P Gameten Die 1. MENDELsche Regel wird auch Uniformitäts- (lat. uniformis, eine Gestalt) und Reziprozitätsregel (lat. reciprocus, auf derselben Bahn zurückkehrend) genannt. Sie lautet: Kreuzt man zwei Individuen einer Art, die sich in einem Merkmal unterscheiden, für das sie reinerbig sind, so sind ihre Nachkommen in der F1 in bezug auf dieses Merkmal untereinander uniform. Dabei ist es gleichgültig, welche der beiden Rassen Mutter oder Vater stellt. F1 Uniformität Gameten GR Gr gR gr GR GGRR GGRr GgRR GgRr Gr GGRr GGrr GgRr Ggrr gR GgRR GgRr ggRR ggRr gr GgRr Ggrr ggRr ggrr F2 Spaltung und Neukombination 9 : 3 : 3 : 1 22.2 Kombinationsquadrat eines dihybriden Erbganges P-Generation F1-Generation RR rot WW weiß RW rosa R-Allel R-Allel R-Allel viel Enzym pro Zeiteinheit W-Allel W-Allel weniger Enzym pro Zeiteinheit rot viel Farbstoff rosa weniger Farbstoff weiß kein Farbstoff kein Enzym 22.3 Erbgang mit unvollständiger Dominanz der Allele für rote und weiße Blütenfarbe W-Allel 22 Klassische Genetik Bei einem dihybriden Erbgang kommt die 3. MENDELsche Regel zum Tragen, die Rekombinantenregel (lat. recombinare, wieder verknüpfen). Kreuzt man Individuen derselben Art, die sich in mehreren Merkmalen unterscheiden, für die sie reinerbig sind, so sind ihre Nachkommen in der F1 in bezug auf dieses Merkmal untereinander uniform. In der F2 treten neben den Merkmalskombinationen der Eltern auch neue Zusammenstellungen auf. Bei einem dihybriden Erbgang spaltet der Phänotyp in der F2 im Verhältnis 9 : 3 : 3 : 1 auf. CORRENS entdeckte auch neue Zusammenhänge, nämlich Erbgänge mit unvollständiger Dominanz. Ein Beispiel dafür ist die Vererbung der Blütenfarbe bei Nelken. Die rote Blütenfarbe wird durch ein Allel „R“ verursacht, das viel roten Farbstoff produziert. Entsprechend ist der Phänotyp der Blüte rot. Das Allel für die weiße Blütenfarbe „W“ codiert ein funktionsunfähiges Enzym, das keinen Farbstoff bilden kann, sodass die Blüte weiß bleibt. Bei Mischerbigkeit kommt das Allel „R“ nur einmal vor, deshalb wird pro Zeiteinheit nur eine geringere Menge des Enzyms und damit eine entsprechend geringere Menge des Farbstoffs gebildet. Der Phänotyp dieser Blüte ist rosa, daher nennt man diese Form der Merkmalsausbidung intermediär (lat. intermedius, dazwischen). Bei der unvollständigen Dominanz kann der Phänotyp der Heterozygoten aber auch mehr dem einen oder anderen Elterntyp zuneigen. GG gg G G g g Parentalgeneration P Gameten Gg 1. Filialgeneration F1 Uniformität Gg Gg Gg G g G g G g G GG Gg g Gg gg Gameten 2. Filialgeneration F2 Spaltung Aus solch einem Kreuzungsansatz leitet sich die 2. MENDELsche Regel ab, die auch Spaltungsregel heißt: Kreuzt man die Individuen der F1 untereinander, so ist die F2 nicht uniform, sondern die Merkmale spalten in bestimmten Zahlenverhältnissen auf, und zwar beim dominant-rezessiven Erbgang im Phänotyp im Verhältnis 3 : 1 und im Genotyp im Verhältnis 1 : 2 : 1. 22.1 Kombinationsquadrat eines monohybriden Erbganges GGRR ggrr GR GR gr gr Gg Rr Gg Rr Gg Rr Gg Rr GR Gr gR gr GR Gr gR gr P Gameten Die 1. MENDELsche Regel wird auch Uniformitäts- (lat. uniformis, eine Gestalt) und Reziprozitätsregel (lat. reciprocus, auf derselben Bahn zurückkehrend) genannt. Sie lautet: Kreuzt man zwei Individuen einer Art, die sich in einem Merkmal unterscheiden, für das sie reinerbig sind, so sind ihre Nachkommen in der F1 in bezug auf dieses Merkmal untereinander uniform. Dabei ist es gleichgültig, welche der beiden Rassen Mutter oder Vater stellt. F1 Uniformität Gameten GR Gr gR gr GR GGRR GGRr GgRR GgRr Gr GGRr GGrr GgRr Ggrr gR GgRR GgRr ggRR ggRr gr GgRr Ggrr ggRr ggrr F2 Spaltung und Neukombination 9 : 3 : 3 : 1 22.2 Kombinationsquadrat eines dihybriden Erbganges P-Generation F1-Generation RR rot WW weiß RW rosa R-Allel R-Allel R-Allel viel Enzym pro Zeiteinheit W-Allel W-Allel weniger Enzym pro Zeiteinheit rot viel Farbstoff rosa weniger Farbstoff weiß kein Farbstoff kein Enzym 22.3 Erbgang mit unvollständiger Dominanz der Allele für rote und weiße Blütenfarbe W-Allel 22 Klassische Genetik Bei einem dihybriden Erbgang kommt die 3. MENDELsche Regel zum Tragen, die Rekombinantenregel (lat. recombinare, wieder verknüpfen). Kreuzt man Individuen derselben Art, die sich in mehreren Merkmalen unterscheiden, für die sie reinerbig sind, so sind ihre Nachkommen in der F1 in bezug auf dieses Merkmal untereinander uniform. In der F2 treten neben den Merkmalskombinationen der Eltern auch neue Zusammenstellungen auf. Bei einem dihybriden Erbgang spaltet der Phänotyp in der F2 im Verhältnis 9 : 3 : 3 : 1 auf. CORRENS entdeckte auch neue Zusammenhänge, nämlich Erbgänge mit unvollständiger Dominanz. Ein Beispiel dafür ist die Vererbung der Blütenfarbe bei Nelken. Die rote Blütenfarbe wird durch ein Allel „R“ verursacht, das viel roten Farbstoff produziert. Entsprechend ist der Phänotyp der Blüte rot. Das Allel für die weiße Blütenfarbe „W“ codiert ein funktionsunfähiges Enzym, das keinen Farbstoff bilden kann, sodass die Blüte weiß bleibt. Bei Mischerbigkeit kommt das Allel „R“ nur einmal vor, deshalb wird pro Zeiteinheit nur eine geringere Menge des Enzyms und damit eine entsprechend geringere Menge des Farbstoffs gebildet. Der Phänotyp dieser Blüte ist rosa, daher nennt man diese Form der Merkmalsausbidung intermediär (lat. intermedius, dazwischen). Bei der unvollständigen Dominanz kann der Phänotyp der Heterozygoten aber auch mehr dem einen oder anderen Elterntyp zuneigen.