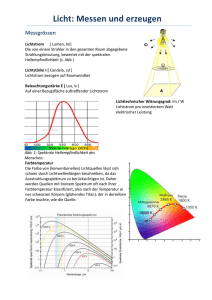
Morphologie und synaptische Interaktion von Neuronen einer
Werbung

September 2004 X. Jahrgang D 13882 F ISSN 0947-0875 3.04 Morphologie und synaptische Interaktion von Neuronen einer kortikalen Kolumne Funktionelle Magnetresonanz-Tomografie des menschlichen Gehirns Thrombin-verwandte Proteasen als Signalmoleküle im Gehirn Kortikale Kontrolle zielgerichteter Bewegungen Neuroforum 3/04 217 INHALT For all lab animals ● 3-dimension precision manipulator for left- & right-hand use ● Optional fine adjustment in the 3rd axis ● Choice of animal adapters, ear bars & accessories ● Versatile attention testing system for rats & mice ● 5-choice serial reaction task ● Pellet feeder or liquid dispenser configuration ● Assess incorrect, correct & premature responses ● Analyze acoustic, tactile & fear-potentiated startle ● Control 4 units with one PC ● User-defined trial sequences ● Complex pre-pulse designs ● Outputs response latency & amplitude ● Study open field behavior or home-cage activity ● Variable box sizes and infrared sensor densities ● Vertical movement detection ● Detailed spatial & temporal analysis of locomotion DE SI GN ● NE W ● Fully computerized custom systems for rats and mice ● Includes ready-to-use trials such as FR, VR, PR, FI, VI, DRH and DRL ● Create your own schedules with the unique program composer! 5-Hole-Box The complete solution for drug research Startle Response ● ● For all arenas including open field, water maze, elevated plus maze, radial maze... ● Outputs distance travelled, time spent, latencies, entries, speed, rotation ● With key-board event recorder Motility Systems VideoMot 2 - Video Activity System Operant Behavior Systems for Life Sciences and Laboratories Stereotaxic Systems Sophisticated Research Instrumentation Contact us for other products and details. TSE Technical & Scientific 218 Equipment GmbH Siemensstr. 21 61352 Bad Homburg/Germany Phone: +49 (0 ) 61 72 -7 89-0 Fax: +49 (0 ) 61 72 -7 89-50 0 Neuro forum 3/04 E-Mail: info@TSE-Systems .de Internet: http://www.TSE-Systems.de INHALT INHALT 179 HAUPTARTIKEL Zum Titelbild: Eine reife und eine unreife Körnerzelle (grün) im hippocampalen Hirnschnitt der Ratte sind in ein Netzwerk von GFAP-positiven Gliazellen (rot) eingebettet (Siehe Artikel des Quartals S. 244). n ....... Joachim Lübke und Dirk Feldmeyer Morphologie und synaptische Interaktion von Neuronen einer kortikalen Kolumne 220 Peter Dechent und Jens Frahm Funktionelle Magnetresonanz-Tomografie des menschlichen Gehirns 229 Tanuja Rohatgi und Georg Reiser Thrombin-verwandte Proteasen als Signalmoleküle im Gehirn: Protease-aktivierte Rezeptoren bei neuronaler Schädigung und Neuroprotektion 237 Neurowissenschaftliche Gesellschaft Vorstand der Amtsperiode 2003/2005 Präsident: Prof. Dr. Herbert Zimmermann, Frankfurt/M. Vizepräsident: Prof. Dr. Klaus-Peter Hoffmann, Bochum Schatzmeister: Prof. Dr. Andreas Draguhn, Heidelberg ARTIKEL DES QUARTALS Christoph Schmidt-Hieber, Peter Jonas, Josef Bischofberger Enhanced synaptic plasticity in newly generated granule cells of the adult hippocampus 244 INSTITUTSVORSTELLUNG Ralf Gold und Mathias Bähr Interdisziplinäres Institut für MS-Forschung in Göttingen Bereich Humanmedizin der Universität Göttingen und Gemeinnützige Hertie-Stiftung 247 Generalsekretär: Prof. Dr. Helmut Kettenmann, Berlin Sektionssprecher Computational Neuroscience: Prof. Dr. Klaus Pawelzik, Bremen Entwicklungsneurobiologie/Neurogenetik: Prof. Dr. Sigrun Korsching, Köln INTERNETVORSTELLUNG Jürgen Konrad Mai und ThomasVoß „ATLAS of the HUMAN BRAIN“ 249 Klinische Neurowissenschaften: Prof. Dr. Mathias Bähr, Göttingen NACHRICHTEN AUS DER NEUROWISSENSCHAFTLICHEN GESELLSCHAFT Kognitive Neurowissenschaften und Verhalten: Prof. Dr. Niels Birbaumer, Tübingen NWG Preis für den Bundeswettbewerbes Jugend forscht 2004 Protokoll der Mitgliederversammlung am 12. Juli 2004 während des FENS Forum 2004 in Lissabon 246 251 Molekulare Neurobiologie: Prof. Dr. Hans Werner Müller, Düsseldorf STELLENMARKT 252 Neuropharmakologie und -toxikologie: Prof. Dr. Werner J. Schmidt, Tübingen HEIMKEHRERBÖRSE 253 Systemneurobiologie: Prof. Dr. Hermann Wagner, Aachen BÜCHER Zelluläre Neurobiologie: Prof. Dr. Tobias Bonhoeffer, Martinsried Neuroforum 3/04 Gehirn und Verhalten 253 AUSBLICK/IMPRESSUM 254 219 MORPHOLOGIE UND SYNAPTISCHE INTERAKTION VON NEURONEN EINER KORTIKALEN KOLUMNE Morphologie und synaptische Interaktion von Neuronen einer kortikalen Kolumne Joachim Lübke und Dirk Feldmeyer Zusammenfassung Ein grundlegendes Prinzip des Neokortex ist seine Organisation in funktionelle Kolumnen und ein weitreichendes horizontales System neuronaler Verbindungen. Diese Kolumnen mit ihren Netzwerken von Neuronen, die für alle sensorischen Hirnregionen nachgewiesen wurden, sind maßgeblich an der Generierung unterschiedlichster Wahrnehmungsprozesse unseres Gehirns beteiligt. Bisher ist jedoch relativ wenig darüber bekannt, wie komplexe Sinnesleistungen auf zellulärer Ebene verarbeitet werden. In den letzten zehn Jahren wurden zahlreiche neuronale Verbindungen innerhalb und zwischen kortikalen Schichten funktionell und strukturell untersucht. Eine der wichtigsten Techniken ist dabei die simultane Patch-clamp Ableitung synaptisch gekoppelter Neurone bei gleichzeitiger intrazellulärer Biocytin-Füllung, die eine korrelierte Struktur-Funktionsanalyse erlaubt. Neben Gemeinsamkeiten zeigten sich jedoch große Unterschiede hinsichtlich der funktionellen und morphologischen Eigenschaften der synaptischen Transmission. Daraus folgt die Notwendigkeit, jede einzelne neuronale Verbindung separat zu beschreiben. Ziel dieser Untersuchungen ist letztlich, die Interaktion der einzelnen Verbindungen innerhalb des neuronalen Netzwerkes einer kortikalen Kolumne zu verstehen. Abstract Morphology and synaptic interaction between neurons in a cortical column A basic feature of the neocortex is its organisation into functional columns and a longrange horizontal system of neuronal connections. These columns together with their network of neurones, present in all sensory cortices, are the cellular substrate for sensory perception in the brain. Currently, relatively little is known how these complex functions are realised at the level of individual neurones or networks. In the last decade numerous neuronal connections within or between cortical layers have been studied both at a functional and structural level. Paired patch-clamp recordings from synaptically coupled neurones combined with simultaneous biocytinfillings is one of the most important techniques allowing a correlated structure-function analysis. Besides similarities, clear differences with respect to both physiology and morphology of synaptic transmission were found. Therefore it is essential to investigate each neuronal connection individually. The aim of these studies is to understand the interaction of individual neuronal connections within the framework of a cortical column. Key words: cortical column, synaptic transmission, cortical signal flow, neuronal connectivity, synaptic efficacy Einleitung Ein generelles Organisationsprinzip sensorischer Areale des Neokortex ist ihre Gliederung in funktionelle vertikal orientierte Kolumnen. Das Konzept der kolumnären Organisation der Hirnrinde geht in erster Linie auf grundlegende Untersuchungen zur Repräsentation der Mechanorezeption im so 220 genannten „Fässchen“- oder „Barrel“-Feld des somatosensorischen Kortex zurück (Mountcastle 1957; Mountcastle und Powell 1959). Ein zentraler Befund dieser Arbeiten war, dass alle Neurone einer vertikalen, durch alle kortikalen Schichten verlaufenden Kolumne nur durch eine der im somatosensorischen Kortex repräsentierten Sinnesmodalitäten (in diesem Fall: Schnurrhaarbe- wegung, Druckwahrnehmung der Haut und Tiefensensibilität) erregt werden. Diese klare Trennung einzelner sensorischer Modalitäten in alternierenden Kolumnen, in denen Neurone ähnliche Eigenschaften besitzen, führte zum Konzept der funktionellen kolumnären Organisation sensorischer Kortizes. Mit der Entdeckung von Orientierungs-, Richtungs- und okulären Dominanzkolumnen im visuellen Kortex durch Hubel und Wiesel (1962, 1963) wurde dieses Organisationsprinzip bestätigt und erweitert. Neben den funktionellen vertikalen Kolumnen existiert ein diskontinuierliches, weitreichendes System von intra- bzw. translaminären neuronalen Verbindungen, welche zum einen verschiedene kortikale Areale, zum anderen die kortikalen Schichten miteinander verbinden. Dieses weitreichende System ist jedoch im Wesentlichen auf exzitatorische Neurone beschränkt. Diese Verbindungen scheinen bei der Synchronisation von räumlich nicht benachbarten, weit auseinander liegenden Ensembles von Pyramidenneuronen kortikaler Kolumnen mit ähnlicher Funktion, z.B. okulärer Dominanz und/oder ähnlicher Richtungs- bzw. Orientierungsspezifität, eine wichtige Rolle zu spielen. Ein besonders attraktives System zum Studium neuronaler Netzwerke innerhalb einer kortikalen Kolumne ist der so genannte „Barrel“-Kortex von Nagetieren. Diese „Barrels“ stellen die topographisch exakte Repräsentation des Schnurrhaarsystems von Nagern im Neokortex dar. Dabei ist jedem Schnurrhaar ein definiertes „Barrel“ zugeordnet (Woolsey und van der Loos 1970). Im Vergleich zu anderen sensorischen Kortizes ist hier die Definition einer kortikalen Kolumne anhand struktureller Merkmale viel klarer, da die „Barrels“ schon im ungefärbten Präparat sichtbar sind (Abbildung 1C). Zudem ist die Zuordnung zum peripheren Rezeptor und zur sensorischen Modalität eindeutig, so dass eine klare Struktur-Funktionsbeziehung möglich ist. Ein Weg, derartige Struktur-Funktionsbeziehungen zu untersuchen ist das akute Gehirnschnittpräparat. Ein grundlegender Vorteil gegenüber in vivo Experimenten im Ganztier ist hierbei der relativ einfache Zugang zu den neuronalen Strukturen, so dass gezielte Messungen von identifizierten Neuronen unter optischer Kontrolle möglich sind (Abbildung 1A, B). Erste simultane Ableitungen von synaptisch gekoppelten Neuronen des Neokortex wurden von Thomson und Mitarbeitern (1994) publiziert. In dieser Arbeit wurden synaptische Verbindungen zwischen Pyramidenzellen der kortikalen Neuroforum 3/04 Abb. 1: Synaptisch gekoppelte, exzitatorische Neurone in Schicht 4 und Schicht 2/3 des „Barrel“-Kortex. Infrarot-Differenzialinterferenzkontrast (IR-DIC)-Aufnahmen einer postsynaptischen Pyramidenzelle in Schicht 2/3 (A) und einer präsynaptischen Sternzelle in Schicht 4 (B) bei hoher Mikroskopvergrößerung. Die beiden Elektroden in (C) zeigen die relative Lage des prä- und postsynaptischen Neurons zueinander. Im ungefärbten Gehirnschnittpräparat sind drei benachbarte „Barrels“ gut zu erkennen, die durch Septa eindeutig voneinander getrennt sind. Die beiden synaptisch gekoppelten Neurone liegen direkt in bzw. oberhalb eines kortikalen „Barrels“, d.h. beide Neurone sind in derselben kortikalen Kolumne lokalisiert. Schichten 5 und 6 abgeleitet und anschließend morphologisch dargestellt. Dadurch konnte eine Beziehung zwischen der Amplitude des exzitatorischen postsynaptischen Potentials und der Anzahl der synaptischen Kontakte hergestellt werden; d.h. hier wurden erstmals funktionelle Eigenschaften synaptischer Transmission mit morphologischen Parametern korreliert. Leider waren von über 1000 gemessenen Neuronenpaaren nur 35 tatsächlich miteinander verbunden, und nur bei drei Verbindungen war ein qualitativ ausreichender Strukturerhalt gegeben, so dass viele wichtige Aspekte synaptischer Transmission weiterhin ungeklärt blieben. Ein Durchbruch waren gezielte simultane Patch-clamp Ableitungen von identifizierten Pyramidenzellen der Schicht 5 (Markram et al. 1997). Dieser experimentelle Ansatz ermöglicht, gezielte Fragen zu funktionellen und strukturellen Parametern synaptischer Transmission zu stellen: Welche Rolle spielen funktionelle Faktoren wie die Größe und der Zeitverlauf der postsynaptischen Leitfähigkeitsänderung bei der Effizienz der synaptischen Übertragung und wie beeinflussen morphologische Faktoren wie die dendritische Konfiguration, axonale Projektion sowie die Anzahl und Verteilung synaptischer Kontakte zusammen mit den aktiven und passiven elektrophysiologischen Eigenschaften des prä- und postsynaptischen Neurons die Effizienz synaptischer Übertragung? Individuelle synaptische Verbindungen wurden dann systematisch durch Verwendung des „Barrel“-Kortex-Gehirnschnittpräparats in Bezug auf das neuronale Netzwerk einer kortikalen Kolumne untersucht (Feldmeyer et al. 1999). Hier wurde der Versuch unternommen, die Konnektivität oder Verbindungsrate am Beispiel von exzitatorischen Neuronen der Schicht 4 abzuschätzen und einen Bezug zur kolumnären Struktur herzustellen (Lübke et al. 2000; s.a. Abbildung 1, 3). Dadurch wurde erstmals ein Brückenschlag zwischen der zellulären Beschreibung neuronaler Verbindungen und der Systemebene erreicht. In den folgenden Kapiteln wird der heutige Kenntnisstand zur Morphologie und Physiologie synaptischer Transmission im Neokortex anhand der wichtigsten Parameter zusammengefasst und basierend auf diesen Erkenntnissen versucht, den Signalfluss in einer kortikalen Kolumne darzustellen. Analyze this! Innovative tools for behavioral research Scientists studying animal behavior have an increasing need for accurate Noldus Information Technology bv quantitative data. As a behavioral neuro- Wageningen, The Netherlands scientist, you need sensitive observational Phone: +31-317-497677 research tools with a maximum degree of E-mail: [email protected] automation. Our integrated solutions for data collection, analysis, management and visualization are today’s premier Noldus Information Technology GmbH Freiburg, Germany Phone: +49-761-4701600 E-mail: [email protected] tools for the study of behavior, locomotion and acoustics. EthoVision - Video tracking system for automation of behavioral experiments Noldus Information Technology Inc. Leesburg, VA, U.S.A. The Observer - System for collection and analysis of observational data, live or from video Phone: +1-703-771-0440 UltraVox - System for automatic Konnektivität in einer kortikalen Kolumne Toll-free: 1-800-355-9541 E-mail: monitoring of ultrasonic vocalizations [email protected] Lange Zeit wurde angenommen, dass die Konnektivität im Neokortex eher zufällig ist (Braitenberg und Schüz 1998). Dagegen sprechen neuere Untersuchungen, die eindeutig belegen, dass es mehre- www.noldus.com Neuroforum 3/04 221 MORPHOLOGIE UND SYNAPTISCHE INTERAKTION VON NEURONEN EINER KORTIKALEN KOLUMNE Abb. 2: Uni- und bidirektionale (reziproke) synaptische Verbindung. (A) Ein Aktionspotential in Zelle 1 löst kein exzitatorisches postsynaptisches Potential (EPSP) in Zelle 2 aus; umgekehrt löst ein Aktionspotential in Zelle 2 ein EPSP in Zelle 1 aus; die Verbindung ist demnach uni-direktional. (B) Aktionspotentiale in Zelle 1 als auch in Zelle 2 lösen EPSPs in Zelle 2 bzw. Zelle 1 aus; d.h. die Verbindung ist bidirektional; in derartigen Verbindungen kommt es zu rekurrenter Erregung. re Grundprinzipien der kortikalen Organisation und neuronalen Verschaltung gibt. Qualitativ kann zwischen lokalen, intralaminären Verbindungen (innerhalb einer kortikalen Schicht) und translaminären (zwischen einzelnen kortikalen Schichten) bzw. transkolumnären (zwischen kortikalen Kolumnen) unterschieden werden. Darüber hinaus ist die synaptische Verschaltung entweder uni-direktional oder reziprok (bidirektional; Abbildung 2). Während die uni-direktionale Verschaltung einen einseitig gerichteten Signalfluss bedingt, ermöglicht die bidirektionale Kopplung eine rekurrente Erregung bzw. eine Rückkopplungsinhibition (Feed-back inhibition). Die überwiegende Zahl der bisher untersuchten synaptischen Verbindungen des Neokortex ist lokal (d.h. innerhalb einer Distanz von <150 µm) und intralaminar. Dies gilt für exzitatorische, aber mehr noch für inhibitorische synaptische Verbindungen. Ein hoher Prozentsatz der intralaminären Verbindungen ist reziproker Natur. Nach heutigem Kenntnisstand sind translaminäre Verbindungen dagegen uni-direktional (Thomson 1997; Reyes und Sakmann 1999; Tarczy-Hornoch et al. 1999; Feldmeyer et al. 2002). Die synaptische Signaltransduktion ist ebenfalls gerichtet und zwar im Wesentlichen von den sensorischen Relay-Kernen des Thalamus nach Schicht 4 des Neokortex, der ersten Station kortikaler Signalverarbeitung (Abbildung 3, 4). Von dort wird die synaptische Erregung über exzitatorische Neurone der kortikalen Schicht 4 zunächst auf Pyramidenzellen der Schicht 2/3 und dann auf solche der Schicht 5 übertragen, d.h. der Informationsfluss ist strikt uni-direktional. In Übereinstimmung hiermit finden sich synaptische Verbindungen 222 zwischen Schicht 4 und Schicht 2/3 (Feldmeyer et al. 2002) bzw. Schicht 2/3 und Schicht 5 häufig (Thomson 1997; Reyes und Sakmann 1999). Synaptische Verbindungen von Schicht 5 nach Schicht 2/3 sind dagegen bis heute nicht nachgewiesen worden. Auf zellulärer Ebene spielt die Zielspezifität der Axone zur Beschreibung der Konnektivität individueller neokortikaler Verbindungen eine wesentliche Rolle. Dabei kommt sowohl zelluläre (d.h. neuronenklassenspezifische) wie auch subzelluläre Zielspezifität vor, d.h. synaptische Kontakte terminieren an einem bestimmten neuronalen Kompartiment, entweder am Soma, den Dendriten oder am Axonhügel. Für die verschiedenen Klassen GABAerger Interneurone ist eine derartige Zielspezifität seit langem bekannt; Untersuchungen an exzitatorischen synaptischen Verbindungen haben jedoch gezeigt, dass auch hier eine gerichtete Termination axonaler Boutons (Verdickungen des Axons im Bereich eines synaptischen Kontakts) vorliegt. Kartierungen zur räumlichen Verteilung synaptischer Verbindungen von Pyramidenneuronen der kortikalen Schicht 5 mittels Kombination von Patch-clamp Ableitungen und durch Photolyse freigesetztes Glutamat aus einer licht-sensitiven inaktiven Vorstufe (engl. caged-compounds) haben gezeigt, dass offensichtlich Subpopulationen von Pyramidenneuronen in der Schicht 5 existieren. Diese weisen in Bezug auf ihre synaptischen Eingänge schichtenspezifische Unterschiede auf (Schubert et al. 2001). Leider kann mit diesem Versuchsansatz der Ursprung dieser Inputs (d.h. die Projektionsneurone), welche zur synaptischen Antwort beitragen, nur unzureichend zurückverfolgt werden. Quantitativ wird die Konnektivität eines gegebenen Netzwerks durch die Häufigkeit der synaptischen Kopplung zwischen verschiedenen Neuronentypen bestimmt. Dabei sind allerdings folgende Punkte kritisch zu betrachten und bei der Berechnung von Konnektivitätsraten zu berücksichtigen: (1) Die experimentelle Wahrscheinlichkeit bzw. der Bias (Erfahrung, Selektion häufig vorkommender Neuronentypen usw.), im akuten Gehirnschnitt eine Verbindung zu finden; (2) Die Wahrscheinlichkeit synaptisch gekoppelter Neurone in akuten Gehirnschnitten stimmt nicht mit dem tatsächlichen Grad der Konnektivität in vivo überein; (3) Synaptisch gekoppelte Neurone zeigen ein weites Spektrum unterschiedlicher Morphologien bezüglich ihres axonalen Projektionsmusters, der Dichte und Verteilung axonaler Boutons und ihrer Anordnung (z.B. in Form neuronaler Cluster) innerhalb einer Hirnregion oder in einer kortikalen Kolumne. Der Grad der Konnektivität einer Verbindung wurde zuerst an Hand der experimentellen Erfolgsrate, eine synaptische Verbindung zu finden, bestimmt. Dies ist jedoch kein allzu verlässlicher Parameter, da je nach experimentellen Bedingungen die Konnektivitätsrate starken Schwankungen unterliegt, wie das Beispiel exzitatorischer Schicht 5-Verbindungen verdeutlicht (1.4%, Thomson et al. 1993; 15%, Markram et al. 1997; 10%, Sjöström et al. 2001). Die experimentelle Erfolgsrate kann daher nur als ein grobes Maß zur Beschreibung der Konnektivität einer Verbindung herangezogen werden. Um zu realistischen Angaben zu kommen, müssen also Parameter wie die räumliche Verteilung und Dichte synaptischer Boutons, die mittlere Anzahl synaptischer Kontakte pro Verbindung und die Anzahl von Neuronen, die innerhalb des rezeptiven Feldes des präsynaptischen Axons liegen, für die Berechnung der Konnektivität mit einbezogen werden. Bis heute liegen solche Analysen nur für intralaminäre Verbindungen zwischen Schicht 5-Pyramidenzellen (15-25%, Markram 1997) und exzitatorischen Neuronen der Schicht 4 vor (20%, Feldmeyer et al. 1999). Aber auch hier bleibt unberücksichtigt, dass die Konnektivitätsrate mit steigender Distanz der prä- und postsynaptischen Neurone (intra- vs. transkolumnäre Verbindungen) deutlich abnimmt und stark von der unterschiedlichen Dichte und Verteilung synaptisch gekoppelter Neurone in den einzelnen kortikalen Schichten (Feldmeyer et al. 1999, 2000) und von der Zielspezifität einzelner Neurone einer gegebenen Verbindung beeinflusst wird (Lübke et al. 2000, 2003). Diese Abhängigkeit wird besonders deutlich, weil z.B. der Anteil reziproker Neuroforum 3/04 Verbindungen (~30% für intralaminäre Verbindungen) die berechnete Konnektivitätsrate deutlich übersteigt. Reziproke kortikale Verbindungen werden als Feed-back -Verstärker synaptischer Signale angesehen, obwohl der Nettoeffekt der Verstärkung kritisch von der synaptischen Effizienz, der Feed-forward-Hemmmung und dem Vorkommen bzw. der Dichte solcher Verbindungen abhängt (Douglas et al. 1995; Chance et al. 1999). Ein weiterer wichtiger Parameter zur Beschreibung der Konnektivität ist die Anzahl und Verteilung synaptischer Kontakte am postsynaptischen Neuron. Die Zielspezifität synaptischer Kontakte GABAerger Interneurone auf Prinzipalneurone oder andere GABAerge Interneurone ist im Gegensatz zu rein exzitatorischen Verbindungen relativ gut untersucht (Kawaguchi und Kubota 1997, 1998; Tamás et al. 1997, 1998; Finnerty et al. 1999; Galarreta und Hestrin 1999, 2001; Gupta et al. 2000; Krimer und Goldmann-Rakic 2001; Kaiser et al. 2004). Dabei etablieren die Axone GABAerger Interneurone en passant Boutons an spezifischen Kompartimenten des Zielneurons, d.h. entweder am Soma, an den Dendriten oder direkt am Axoninitialsegment, welches den unterschiedlichen Einfluss der Hemmung auf die postsynaptische Partnerzelle (selektive Unterdrückung synaptischer Inputs an unterschiedlichen Kompartimenten des Zielneurons, Blockade der Initiierung von Na+ und Ca2+ Spikes usw.) und damit auch innerhalb eines Netzwerks erklärt. Neuere Studien zu synaptischen Verbindungen zwischen exzitatorischen Neuronen des Neokortex zeigen eindeutig, dass sich solche Verbindungen auch durch eine hohe Zielspezifität auszeichnen. In allen bisher untersuchten Verbindungen sind synaptische Kontakte exklusiv nur am Dendritenschaft bzw. an dendritischen Dornenfortsätzen (engl. spines) zu finden, und dort hauptsächlich an sekundären und tertiären basalen Dendriten (63-85%) lokalisiert (Makram et al. 1997a; Feldmeyer et al. 1999, 2002; Lübke et al. 2000, 2003; Silver et al. 2003). Die große Mehrzahl synaptischer Kontakte befindet sich zudem relativ nah am Soma (~100 µm) und trägt so auf Grund der geringen elektrotonischen Distanz zur synaptischen Effizienz bei, da die elektrische Filterung des postsynaptischen Potentials nicht sehr stark ist. Interessanterweise scheint es Ausnahmen zur Verteilung synaptischer Kontakte einer exzitatorischen Verbindung zu geben. Die bisher beschriebenen Verbindungen sind alle durch eine synaptische Innervationsdomäne gekennzeichnet, die sich hauptsächlich im Bereich der basalen Dendriten des postsynaptischen Neurons befindet. Für die Verbindungen zwischen Pyramidenzellen der Schicht 2/3 und der Schicht 5 sind synaptische Kontakte nicht nur an basalen Dendriten lokalisiert, sondern auch in der Nähe der Endverzweigung des apikalen Dendriten. Dies ist die Region, an der dendritische Kalzium-Aktionspotentiale ausgelöst werden, so dass hier EPSPs zu deren Auslösung beitragen können. Fine surgical instruments and accessories for research • Spring scissors • Forceps • Scalpels • Sutures • Retractors • Clamps • And much more Fine Science Tools GmbH Fahrtgasse 7 - 13 D-69117 Heidelberg Germany Tel.: +49 (0) 62 21 / 90 50 50 Fax: +49 (0) 62 21 / 60 00 01 E-Mail: [email protected] Web: www.finescience.com Signalfluss in einer kortikalen Kolumne (Kanonischer Signalfluss) Der Signalfluss bzw. die Signalverarbeitung in einer kortikalen Kolumne ist für die exzitatorische Transmission mittlerweile recht gut untersucht. Neben funktionellen Eigenschaften wie Konnektivität, Effizienz und Verlässlichkeit synaptischer Verbindungen bestimmen strukturelle Charakteristika wie die dendritische Konfiguration, axonale Projektion sowie die Anzahl und Verteilung morphologisch identifizierter synaptischer Kontakte den Signalfluss bzw. Signalverarbeitung einer kortikalen Kolumne. Sensorische Signale aus der Peripherie erreichen über die spezifischen Relay-Kerne des Thalamus die Eingangsstation des Neokortex, Neuroforum 3/04 223 MORPHOLOGIE UND SYNAPTISCHE INTERAKTION VON NEURONEN EINER KORTIKALEN KOLUMNE Abb. 3: Synaptische Verbindungen im Signalfluss einer kortikalen Kolumne. NEUROLUCIDA Rekonstruktionen synaptisch verbundener Paare exzitatorischer Neurone in der Signalkette einer kortikalen Kolumne. Die Dendriten der präsynaptischen Neurone sind rot dargestellt und ihre Axone in blau. Die postsynaptischen Neurone sind in weiß wiedergegeben und ihre Axone in grün. Der Signalfluss ist so dargestellt wie in Abbildung 4 detailliert beschrieben. die Schicht 4 (Abbildung 3, 4). Die thalamokortikalen Afferenzen terminieren zum großen Teil an exzitatorischen Sternzellen, d.h. diese Zellen repräsentieren die erste Station kortikaler Signalverarbeitung. Für den visuellen und den somatosensorischen Kortex konnte gezeigt werden, dass synaptische Verbindungen dieser Neurone durch eine hohe Effizienz gekennzeichnet sind, welche unter anderem durch die Höhe der mittleren EPSP-Amplitude (1.0-1.6 mV) und der geringen Fehlerrate (<5%) bzw. dem niedrigen Koeffizienten der Variation (CV < 0.4; Quotient aus der Standardabweichung und dem Mittelwert der EPSP-Amplitude; er gibt die relative Varianz des mittleren Messwertes an) ausgedrückt wird (Stratford et al. 1996; Tarczy-Hornoch et al. 1999; Feldmeyer et al. 2002). Einige dieser Schicht 4-Verbindungen sind so stark, dass es zur Auslösung postsynaptischer Aktionspotentiale kommt, die wahrscheinlich auf die vergleichsweise hohe Beteiligung von NMDA-Rezeptoren (~ 40%) an der Gesamtamplitude zurückzuführen sind (Feldmeyer et al. 1999). Auf Grund der hohen Konnektivität zwischen exzitatorischen Sternzellen dieser Schicht (Feldmeyer et al. 1999; Lübke et al. 2003) kommt diesen Neuronen nicht nur eine Schlüsselstellung bei der Signalübertragung, sondern auch eine „Feed-back-Verstärker“Funktion selbst schwacher sensorischer Signale aus der Peripherie zu (Douglas et al. 1995). Morphologisch sind exzitatorische Sternzellen durch die vertikale, auf die Kolumne beschränkte, Projektion ihres Axons charakterisiert (Abbildung 3, 4). Diese Axonkollateralen ziehen dabei durch alle kortikalen Schichten mit einer präferentiellen Projektion in Schicht 2/3, wo die en passant Bou224 tons zielspezifisch an Pyramidenneuronen zu finden sind. Der Signalfluss von Schicht 4 nach Schicht 2/3 (translaminar) ist uni-direktional, d.h. nur in Richtung der Schicht 2/3 und nicht umgekehrt und bleibt auf die Kolumne beschränkt (Feldmeyer et al. 2002; Lübke et al. 2003; s. a. Abbildung 3, 4). Obwohl die Distanz zwischen prä- und postsynaptischem Neuron nicht unerheblich ist, ist diese synaptische Verbindung trotz ihrer relativ niedrigen Effizienz (mittlere EPSP-Amplitude 0.7 mV) sehr zuverlässig. Für die Signalübertragung von Schicht 4 nach Schicht 2/3 scheint daher eine wesentlich höhere Schwelle der Aktivierung bzw. die gleichzeitige Rekrutierung vieler Sternzellen notwendig zu sein. Pyramidenneuronen der Schicht 2/3 kommt folglich eher eine Filterfunktion für die laterale Ausbreitung der Erregung innerhalb dieser Schicht zu. Im Gegensatz zur kolumnären Organisation der Axone der exzitatorischen Schicht 4-Neurone verfügen Schicht 2/3-Pyramidenneurone über zwei axonale Domänen: eine vertikale, hauptsächlich kolumnär organisierte Domäne und ein weitreichendes diskontinuierliches System horizontaler Verbindungen präferentiell in den Schichten 2/3 und 5, mit einer Projektion individueller Axonkollateralen von 2-4 mm über die Kortexoberfläche (Abbildung 3). An dieser Stelle des Neokortex kann der Signalfluss zwei Richtungen nehmen, entweder innerhalb der Kolumne (intrakolumnär) oder über Kolumnen hinweg (transkolumnär). Die Zielstruktur von Schicht 2/3Pyramidenzellaxonen sind hauptsächlich andere Schicht 2/3- oder Schicht 5-Pyramidenneurone. Für synaptische Verbindungen (Paarableitungen) zwischen Pyramidenneuronen der Schicht 2/3 liegen bis heute haupt- sächlich Befunde zur lokalen (Distanz zwischen prä- und postsynaptischen Neuron ~ 50 µm), intrakolumnären (Reyes und Sakmann 1999; Atzori et al. 2001; Feldmeyer et al. 2003; Thomson und Bannister 1998, 2003) und nur eine Arbeit zur transkolumnären Konnektivität (Yoshimura et al. 2000) vor. Der Signalfluss innerhalb von Schicht 2/3 ist sowohl uni- (Atzori et al. 2001) als auch bidirektional und wird über die vertikale und horizontale axonale Domäne vermittelt. Obwohl die Konnektivität von Pyramidenneuronen der Schicht 2/3 lokal (Radius von ~ 100 µm) sehr hoch ist, ist die Effizienz dieser Verbindung vergleichsweise niedrig (mittlere EPSP Amplitude 0.3-1.0 mV). Auch die Wahrscheinlichkeit der Neurotransmitter-Freisetzung (probability of release, kurz Pr) und die synaptischen Antworten nach repetitiver Stimulation sind bei diesen Verbindungen in verschiedenen sensorischen Kortizes sehr unterschiedlich. Während im auditorischen Kortex zwei Subpopulationen synaptischer Schicht 2/3-Pyramidenzell-Verbindungen, eine mit niedrigem Pr (0.13) und eine zweite mit hohem Pr (0.68) vorkommen, existieren im Barrel-Kortex nur solche mit hohem Pr (0.93). Synaptische Verbindungen zwischen Neuronen gleichen Typs aber unterschiedlichen physiologischen Eigenschaften sind auch für den motorischen, somatosensorischen und visuellen Kortex nachgewiesen (Thomson 1997). Diese Befunde implizieren, dass sowohl innerhalb eines sensorischen Areals als auch zwischen sensorischen Kortizes sehr heterogene Verbindungstypen zwischen Pyramidenzellen der kortikalen Schicht 2/3 existieren. Die Signaltransduktion über diese Verbindungen ist daher in verschiedenen sensorischen Arealen sehr unterschiedlich und wahrscheinlich dem jeweiligen sensorischen Input angepasst. Zudem repräsentieren Schicht 2/3-Pyramidenzellen eine sehr heterogene Population innerhalb der Klasse der Pyramidenneurone hinsichtlich ihrer dendritischen Geometrie und axonalen Projektion, die möglicherweise das morphologische Substrat für die Unterschiede in funktionellen Eigenschaften darstellen. Intralaminäre Schicht 2/3-Pyramidenzellverbindungen sind integrative Elemente der Signalverarbeitung innerhalb einer kortikalen Kolumne. Über ihr diskontinuierliches System weitreichender horizontaler Axone findet die transkolumnäre Signaltransduktion statt. Eine mögliche Funktion könnte daher in der Synchronisation elektrischer Aktivität neuronaler Ensembles mit ähnlichen funktionellen Eigenschaften über einen großen Bereich Neuroforum 3/04 JOACHIM LÜBKE UND DIRK FELDMEYER der Kortexoberfläche, d.h. über mehrere kortikale Kolumnen hinweg, liegen. Die andere Zielstruktur von Schicht 2/3Pyramidenzellaxonen sind Pyramidenzellen der Schicht 5, die in der Regel eine große terminale dendritische Endaufzweigung in Schicht 1 ausbilden. Die Pyramidenzellen der Schicht 5 stellen neben denen der Schicht 6 das „Output“-System des Neokortex dar, z.B. in verschiedene subkortikale Gehirnregionen (z.B. Thalamus, Pons) bis hin zur sensorischen Peripherie (Abbildung 3, 4). Translaminäre Verbindungen zwischen Pyramidenneuronen der Schicht 2/3 und Schicht 5 sind wahrscheinlich uni-direktional, wie die zwischen Schicht 4 nach Schicht 2/3 (Thomson 1997; Thomson und Bannister 2003; Reyes und Sakmann 1999). Die mittlere EPSP Amplitude variiert stark (0.1-0.8 mV), zudem scheint diese Verbindung eine gemessen an anderen Verbindungen relativ niedrige synaptische Effizienz zu haben. Ihre funktionelle Bedeutung ist relativ unklar, es ist jedoch vorstellbar, dass sie eine intrakolumnäre Relay-Station zur Integration kortikaler Signale von intra- und transkolumnären neuronalen Ensembles darstellt. Neben Pyramidenzellen der Schicht 6 repräsentieren die der Schicht 5 den kortikalen Output zu subkortikalen Gehirnregionen bzw. zur sensorischen Peripherie und sind daher strukturell und funktionell am besten charakterisiert (Thomson et al. 1993; Deuchars et al. 1994; Markram und Tsodyks 1996; Markram et al. 1997a, b; Tsodyks und Neuroforum 3/04 Abb. 4: Signalfluss in einer kortikalen Kolumne. Die Abbildung zeigt schematisch den „kanonischen“ Signalfluss in einer kortikalen Kolumne eines sensorischen Areals des Großhirns. Als Beispiel wurde der somatosensorische „Fässchen“- oder „Barrel“-Kortex der Nager gewählt, weil hier in der kortikalen Schicht 4 die morphologischen Korrelate der kortikalen Kolumne, die „Barrels“ zu sehen sind (s. Text). Über thalamische Relay-Kerne erhält der Kortex sensorische Eingänge aus der Peripherie, die hauptsächlich in Schicht 4 terminieren. Exzitatorische Neurone der Schicht 4 verstärken das Signal und leiten es präferentiell an Pyramidenzellen der Schicht 2/3 weiter. Von dort wird es auf andere Pyramidenzellen in Schicht 2/3 oder auch auf solche in Schicht 5 übertragen. Pyramidenzellen in Schicht 5 sind ebenfalls stark reziprok untereinander verbunden und sind die Ausgangsstation des sensorischen Kortex; von hier gelangt das sensorische Signal in andere Kortexareale und subkortikale Bereiche. Markram 1997). Die Verbindung von Pyramidenzellen der kortikalen Schicht 5 zeichnet sich durch eine relativ hohe Effizienz der synaptischen Transmisson aus, was sich in einer hohen mittleren EPSP Amplitude (1.3 mV), einem niedrigen CV von 0.52 und einer geringen Fehlerrate (14%) ausdrückt. Ensembles von synaptisch gekoppelten Schicht 5-Pyramidenneuronen scheinen sowohl intra- als auch transkolumnär bei der Generierung, Synchronisation und Aufrechterhaltung elektrischer Aktivität ein wichtiges Element der Signaltransduktion bzw. Signalverarbeitung darzustellen. Im visuellen Kortex kommt eine zusätzliche Verbindung zwischen exzitatorischen Schicht 4-Neuronen und Schicht 6-Pyramidenneuronen vor (Tarczy-Hornoch et al. 1999). Interessanterweise weicht diese Verbindung in vielerlei Hinsicht von den oben 225 MORPHOLOGIE UND SYNAPTISCHE INTERAKTION VON NEURONEN EINER KORTIKALEN KOLUMNE beschriebenen exzitatorischen Verbindungen ab. Sie zeichnet sich durch eine vergleichsweise sehr niedrige synaptische Effizienz aus, hat einen niedrigen Pr (0.13) und zeigt eine im Gegensatz zu anderen exzitatorischen Verbindungen frequenzabhängige Faszilitierung der EPSP Amplitude. Obwohl bis heute die Vielzahl möglicher synaptischer Verbindungen des kolumnären Netzwerks des Neokortex noch nicht ausreichend bzw. gar nicht beschrieben ist, kann zusammenfassend festgestellt werden, dass intralaminäre Verbindungen in der Regel durch eine hohe synaptische Effizienz, eine geringe Fehlerrate und einen hohen Anteil reziproker Verbindungen charakterisiert sind. Im Gegensatz dazu sind translaminäre Verbindungen ein weitgehend kolumnäres, uni-direktionales System, welches den gerichteten Informationsfluss über die einzelnen kortikalen Schichten gewährleistet (Abbildung 3, 4). Ein zentrales Element kortikaler Signalverarbeitung stellen die exzitatorischen Neurone der Schicht 4 dar, über deren axonale Projektion die Information selektiv in obere Kortexschichten und innerhalb der kortikalen Kolumne verteilt wird. Über das weitreichende System horizontaler Axonkollateralen der Pyramidenneurone der Schichten 2/3 und 5 wird die Ausbreitung der sensorischen Information über kortikale Kolumnen hinweg und über ihre callosale Projektion zur Gegenseite ein Abgleich beider kortikaler Hemisphären sichergestellt. Inhibitorische Interneurone In dem oben beschriebenen Modell zum Signalfluss in einer kortikalen Kolumne sind GABAerge Interneurone bewusst ausgeklammert worden und werden hier gesondert betrachtet. Obwohl diese Neurone nur ca. 10-15% der gesamten Neuronenpopulation des Neokortex repräsentieren und verschiedene Typen (Klassen) dieser Neurone schon sehr lange bekannt sind, ist ihre Funktion und damit ihre Rolle innerhalb des kolumnären Netzwerks im Gegensatz zum Hippokampus bis heute relativ unklar. Die hohe Zielspezifität ihrer Axone an verschiedenen Kompartimenten des Zielneurons führt zu einer deutlich unterschiedlichen selektiven Art der Inhibition innerhalb des kortikalen Netzwerks, die weit über den Mechanismus der Feed-forward- und Feed-back-Inhibition hinausgeht (Kawaguchi und Kubota 1997, 1998; Buhl et al. 1997; Tamás et al. 1997, 1998; Galarreta und Hestrin 1999, 2001; Markram et al. 1998; Reyes et al. 1998; Gibson et al. 1999; Zilberter et al. 1999; 226 Gupta et al. 2000; Krimer und GoldmanRakic 2001; Kaiser et al. 2004; Karube et al. 2004). Es gilt heute als gesichert, dass GABAerge Interneurone via chemischer und elektrischer Synapsen untereinander neuronale Ensembles bilden, die für die Synchronisation und Aufrechterhaltung oszillatorischer Aktivität verantwortlich sind (Galarreta und Hestrin 1999, 2001; Gibson et al. 1999; Támas et al. 2000). Synaptische Verbindungen zwischen exzitatorischen und inhibitorischen Neuronen sind rekurrent und scheinen eine wichtige Rolle bei der Stabilisierung des kortikalen Netzwerks zu spielen, indem sie das Auftreten von „run away“ Exzitation und paroxysmaler neuronaler Aktivität unterdrücken, wie sie in unkontrollierten exzitatorischen Netzwerken vorkommen kann. Darüber hinaus unterliegen exzitatorische Synapsen einer viel stärkeren Depression als inhibitorische Synapsen, selbst während Perioden anhaltender neuronaler Aktivität. Sie besitzen daher auch einen intrinsischen Inhibitionsmechanismus, der zur Stabilität neuronaler Netzwerke beiträgt (Galarreta und Hestrin 1998). Im Gegensatz dazu wird durch die verschiedenen Typen GABAerger Interneurone ein hoher Grad der Flexibilität bei der Regulation neuronaler Aktivität erreicht. Synaptische Effizienz und Verlässlichkeit synaptischer Übertragung Für ein genaues Verständnis neuronaler Verrechnungsprozesse in einer kortikalen Kolumne ist aber nicht nur die Konnektivität zwischen Neuronen sondern auch die Stärke oder Effizienz der verschiedenen synaptischen Verbindungen von entscheidender Bedeutung. Sie wird durch drei voneinander unabhängige Parameter bestimmt: (1) der Anzahl der Freisetzungsstellen des Neurotransmitters n, (2) der Wahrscheinlichkeit der Transmitterfreisetzung aus synaptischen Vesikeln Pr pro Verbindung, und (3) der quantalen Amplitude q, d.h. der Signalamplitude pro synaptischen Kontakt. Synaptische Effizienz ist somit das Produkt dieser drei Parameter E = n · q · Pr und wird durch die Amplitude postsynaptischer Potentiale (EPSP oder IPSP, je nachdem, ob die Verbindung exzitatorisch oder inhibitorisch ist) oder Ströme (EPSC oder IPSC) ausgedrückt. Individuelle synaptische Verbindungen des Neokortex zeigen große Unterschiede bezüglich ihrer synaptischen Effizienz (Feldmeyer und Sakmann 2000; zusammengefasst durch Thomson und Bannister 2003). Synaptische Verbindungen zwischen exzitato- rischen Sternzellen der kortikalen Schicht 4 zeichnen sich durch eine hohe EPSP Amplitude (1.6 mV) bei niederfrequenter Stimulation aus (Feldmeyer et al. 1999), während andere kortikale Verbindungen, z.B. solche zwischen Pyramidenneuronen der Schicht 6 und Sternzellen der Schicht 4, eine 16-fach niedrigere EPSP Amplitude (0.1 mV) aufweisen (Stratford et al. 1996). Die große Mehrzahl exzitatorischer Verbindungen des Neokortex ist jedoch durch eine relativ hohe Wahrscheinlichkeit der Transmitterfreisetzung charakterisiert. Translaminäre synaptische Verbindungen zwischen Sternzellen und Schicht 2/3-Pyramidenneuronen im Barrel-Kortex haben hohe Pr Werte (0.8). Ähnlich verhält es sich bei intralaminären Verbindungen zwischen Pyramidenneuronen der Schicht 2/3 und zwischen Sternzellen der Schicht 4 (Feldmeyer et al. 1999). Demzufolge besitzen eine Reihe von intrakortikalen synaptischen Verbindungen eine Pr, die fast so hoch ist wie diejenige der sehr verlässlichen thalamokortikalen Verbindungen (Gil et al. 1999). Daneben existieren jedoch auch Verbindungen mit eher niedrigen Pr, z.B. Verbindungen zwischen Schicht 6-Pyramidenneuronen und Schicht 4-Neuronen des visuellen Kortex (0.37-0.53; Stratford et al. 1996) und einer Subpopulation synaptisch gekoppelter Schicht 2/3-Pyramidenneuronen im auditorischen Kortex (0.13, Atzori et al. 2001). Diese Befunde sprechen für eine hohe Effizienz und Verlässlichkeit der exzitatorischen synaptischen Transmission neokortikaler (intrinsischer) Verbindungen und wurden bis dato für intrinsische kortikale Verbindungen kontrovers diskutiert (Stevens 1994). Die Wahrscheinlichkeit der Transmitterfreisetzung und damit die Amplitude der EPSPs bzw. EPSCs hängt entscheidend von der Dynamik, d.h. dem „Funktionszustand“ des präsynaptischen Neurons ab. Die repetitive Stimulation des präsynaptischen Neurons einer exzitatorischen Verbindung des Neokortex führt im nachgeschalteten postsynaptischen Neuron in der Regel zu synaptischer Depression (engl. Paired Pulse Depression), während inhibitorische Verbindungen oft eine synaptische Faszilitierung (engl. Paired Pulse Fascilition) zeigen. Das frequenzabhängige Verhalten einer Synapse, ihre Kurzzeit-Plastizität, ist oft zielneuronspezifisch (Stratford et al. 1996; Reyes und Sakmann 1999; Reyes et al. 1998; Galaretta und Hestrin 1998; Makram et al. 1998; Thomson und Bannister 1998; Varela et al. 1999; Gupta et al. 2000; Krimer und Goldman-Rakic 2001; Feldmeyer et al. 2002; Kaiser et al. 2004); regionenspezifisch (AtzNeuroforum 3/04 JOACHIM LÜBKE UND DIRK FELDMEYER ori et al. 2001), wird über die postnatale Entwicklung reguliert (Reyes und Sakmann 1999) und zeigt einen hohen Grad der Variabilität bezüglich der Spezifität der Verbindung (Tsodyks und Markram 1997). Sensorische Deprivation, z.B. das Trimmen der Schnurrhaare, scheint nach neueren Untersuchungen nicht nur einen strukturellen Effekt auf die axonale und dendritische Morphologie intra- bzw. transkolumnärer Verbindungen zu besitzen, sondern auch einen signifikanten funktionellen Einfluss auszuüben, z.B. auf die Kurzzeitdynamik der synaptischen Transmission (Finnerty et al. 1999; Keller und Carlson 1999; Bender et al. 2003; Maravall et al. 2004, Petersen et al. 2004). Die Effizienz der Signalübertragung sensorischer Information ist maximal für eine bestimmte Frequenz der Aktivierung an einer Synapse. Neokortikale Synapsen, die neuronale Aktivität herunterfahren, tragen wesentlich zur räumlichen Verarbeitung von Signalen während niederfrequenter Stimulation bei, z.B. während spontaner Entladung. Synapsen mit faszilitierenden Eigenschaften scheinen dagegen erst bei hochfrequenter Stimulation (9-70 Hz) optimal zu operieren und könnten daher eher bei evozierter Aktivität kortikaler Neurone rekrutiert werden (Fuhrmann et al. 2002). Es ist jedoch weiterhin unklar, ob spontane Aktivität in vivo (im wachen Tier) und evozierte Aktivität durch sensorische Stimulation als gleichwertig angesehen werden können (Moore und Nelson 1998, Zhu und Connors 1999; Brecht und Sakmann 2003). stellen nahezu übereinstimmt (Korn et al. 1981; Redman 1990; Gulyas et al. 1993; Korn et al. 1994). Auf der anderen Seite liegen immer mehr ultrastrukturelle Daten vor, die belegen, dass die Zahl sogenannter „docked vesicles“ an zentralen Synapsen unterschiedlich hoch ist, aber definitiv mehr als nur ein Vesikel an die präsynaptische Membran gedockt ist, welches für eine multivesikuläre Freisetzung spricht (Harris und Sultan 1995; Schikorski und Stevens 1997; Xu-Friedmann et al. 2001; Millar et al. 2002). Diese morphologischen Befunde werden durch physiologische Studien unterstützt, die ebenfalls eine multivesikuläre Transmitterfreisetzung an zentralen Synapsen postulieren (Wadiche und Jahr 2001; Oertner et al. 2002). Der Vergleich beider Hypothesen lässt den Schluss zu, dass a priori wahrscheinlich beide Mechanismen an zentralen Synapsen vorkommen. Für die Verbindung zwischen exzitatorischen Sternzellen und Schicht 2/3-Pyramidenzellen haben wir zeigen können, dass pro Aktivierung eines synaptischen Kontakts dieser Verbindung nur ein Transmitter-Vesikel freigesetzt wird (Silver et al. 2003). Demnach folgt ein synaptischer Kontakt dem Alles-oder-Nichts-Prinzip: Entweder wird Transmitter freigesetzt, daraus resultiert ein postsynaptisches Signal, oder es kommt zu keiner Antwort. Die Fluktuation der Signalamplitude (EPSP) ist also allein auf die unterschiedliche Zahl aktiver Kontakte pro Verbindung zurückzuführen. Ob dies jedoch ein allgemein gültiges Prinzip für andere zentrale oder kortikale Synapsen ist, muss in Folgearbeiten noch eindeutiger geklärt werden. Quantale Eigenschaften der synaptischen Übertragung Ein synaptischer Kontakt besteht aus morphologischer Sicht aus einer präsynaptischen Neurotransmitter-Freisetzungsstelle, an der ein „Pool“ synaptischer Vesikel lokalisiert ist, einer postsynaptischen Dichte, an der je nach Synapsentyp verschiedene NeurotransmitterRezeptoren zu finden sind und einem synaptischen Spalt zwischen prä- und postsynaptischer Membran. Es ist bis heute jedoch unklar, wie synaptische Übertragung in einer definierten Verbindung abläuft. Ein bestimmender Faktor ist hier die Anzahl an Neurotransmitter-Freisetzungsstellen. Es ist aber nicht eindeutig geklärt, ob alle morphologisch identifizierten synaptischen Kontakte einer Verbindung auch funktionell sind (Phänomen der silent synapses), und ob es pro Kontakt mehr als nur eine Neurotransmitter-Freisetzungsstelle gibt. So gesehen bestimmt die Anzahl an Freisetzungsstellen die Eigenschaften synaptischer Übertragung, z.B. ob uni- bzw. multivesikulärer und/oder uni- bzw. multiquantale Freisetzung des Neurotransmitters aus synaptischen Vesikeln vorliegt. Diese Problematik wurde bisher nur in wenigen Studien thematisiert (Korn et al. 1981; Gulyas et al. 1993; Silver et al. 2003). Trotz großer Unterschiede in Bezug auf die Anzahl identifizierter synaptischer Kontakte der bisher untersuchten synaptischen Verbindungen (zwischen InterneuronMauthnerzelle im Goldfisch, hippokampaler CA3 PyramidenzelleInterneuron und der kortikalen Sternzelle-Schicht 2/3-Pyramidenzelle der Ratte) zeigte sich, dass die Anzahl funktionell bestimmter Transmitter-Freisetzungsstellen der Anzahl elektronenmikroskopisch identifizierter synaptischer Kontakte entsprach. Diese Ergebnisse sprechen eindeutig für funktionelle Kontakte mit nur einer Freisetzungsstelle. Bis heute wird kontrovers diskutiert, ob an zentralen Synapsen uni- oder multivesikuläre Freisetzung an aktiven Zonen stattfindet, d.h. ob nur ein oder mehrere Vesikel an der Freisetzung des Neurotransmitters pro Aktivierung beteiligt sind. Die Hypothese one site one vesicle resultiert aus klassischen Studien, in denen die quantale Freisetzung, bestimmt durch statistische Analyse synaptischer Potentiale, mit der Anzahl morphologisch identifizierter FreisetzungsNeuroforum 3/04 227 MORPHOLOGIE UND SYNAPTISCHE INTERAKTION VON NEURONEN EINER KORTIKALEN KOLUMNE Literatur Atzori, M., Lei, S., Evans, D.I.P., Kanold, P.O., Phillips-Tansey, E., McIntyre, O. und McBain, C.F. (2001): Differential synaptic processing separates stationary from transient inputs to the auditory cortex. Nature Neuroscience 4: 12301237. Braitenberg, V. und Schüz, A. (1998): Anatomy of the Cortex: Statistics and geometry. Berlin: Springer Verlag. Ed. 2. Buhl, E.H., Tamás, G., Szilágyi, T., Stricker, C., Paulsen, O. und Somogyi, P. (1997): Effect, number and location of synapses made by single pyramidal cells onto aspiny interneurones of cat visual cortex. Journal of Physiology 500: 689-713. Deuchars, J., West, D.C. und Thomson, A.M. (1994): Relationships between morphology and physiology of pyramid-pyramid single axon connections in rat neocortex in vitro. Journal of Physiology 478: 423-435. Feldmeyer, D., Egger, V., Lübke, J. und Sakmann, B. (1999): Reliable synaptic connections between pairs of excitatory layer 4 neurones within a single ‘barrel’ of developing rat somatosensory cortex. Journal of Physiology 521: 169-190. Feldmeyer, D. und Sakmann, B. (2000): Synaptic efficacy and reliability of excitatory connections between the principal neurones of the input (layer 4) and output layer (layer 5) of the neocortex. Journal of Physiology 525: 31-39. Feldmeyer, D., Lübke, J., Silver, R.A. und Sakmann, B. (2002): Synaptic connections between layer 4 spiny neurones and layer 2/3 pyramidal cells in rat barrel cortex: physiology and anatomy of interlaminar signaling within a cortical column. Journal of Physiology 538: 803-822. Galarreta, M. und Hestrin, S. (1999): A network of fast-spiking cells in the neocortex connected by electrical synapses. Nature 402: 72-75. Gulyás, A.I., Miles, R., Sik, A., Tóth, K., Tamamaki, N. und Freund T.F. (1993): Hippocampal pyramidal cells excite inhibitory neurons through a single release site. Nature 366: 683-687. Hubel, D.H. und Wiesel, T.N. (1962): Receptive fields, binocular interaction and functional architecture in the cat’s visual cortex. Journal of Physiology 160: 106-154. Kawaguchi, Y. und Kubota, Y. (1997): GABAergic cell subtypes and their synaptic connections in rat frontal cortex. Cerebral Cortex 7: 476-486. Korn, H., Friller, A., Mallet, A., Legrende, P. und Faber, D.S. (1981): Fluctuating response at a central synapse: n of binomial fit predicts number of presynaptic boutons. Science 213: 898901. Korn, H., Sur, C., Stephane, C., Legrende, P. und Faber, D.S. (1994): The one-vesicle hypothesis and multivesicular release. In: Stärnje, L., Greengard, P., Grillne, S., Hökfeldt, T. und Ottoson, D. (Hrsg.) Molecular and cellular mechanisms of neurotransmitter release. New York: Raven Press. Lübke, J., Egger, V., Sakmann, B. und Feldmeyer, D. (2000): Columnar organization of dendrites and axons of single and synaptically coupled neurons in layer 4 of the rat barrel cortex. Journal of Neuroscience 20: 5300-5311. 228 Lübke, J., Roth, A., Feldmeyer, D. und Sakmann, B. (2003): Morphometric analysis of the columnar innervation domain of neurons connecting layer 4 and layer 2/3 of juvenile rat barrel cortex. Cerebral Cortex 13: 1051-1063. Markram H., Lübke J., Frotscher M., Roth A. und Sakmann B. (1997a): Physiology and anatomy of synaptic connections between thick tufted pyramidal neurones in the developing rat neocortex. Journal of Physiology 500: 409-440. Mountcastle, V.B. (1957): Modality and topographic properties of single neurons of cat’s somatic sensory cortex. Journal of Neurophysiology 20: 408-434. Petersen, C.C., Brecht, M., Hahn, T.T. und Sakmann, B. (2004): Synaptic changes in layer 2/3 underlying map plasticity of developing barrel cortex. Science 304: 739-742. Schikorski, T. und Stevens, C.F. (2001): Morphological correlates of functionally defined synaptic vesicle populations. Nature Neuroscience 4: 391-395. Silver, R.A., Lübke, J., Sakmann, B. und Feldmeyer, D. (2003): Uniquantal transmission at excitatory synaptic contacts in barrel cortex. Science 302: 1981-1984. Támas, G., Somogyi, P. und Buhl, E.H. (1998): Differentially interconnected networks of GABAergic interneurons in the visual cortex of the cat. Journal of Neuroscience 18: 4255-4270. Thomson, A.M., Deuchars, J. und West , D.C. (1993): Single axon excitatory postsynaptic potentials in neocortical interneurons exhibit pronounced paired pulse facilitation. Neuroscience 54: 347-360. Thomson, A.M., West, D.C., Wang, Y. und Bannister, A.P. (2002): Synaptic connections and small circuits involving excitatory and inhibitory neurons in layers 2-5 of adult rat and cat neocortex: triple intracellular recordings and biocytin labelling in vitro. Cerebral Cortex 12: 936-953. Wadiche, J.I. und Jahr, C.E. (2001): Multivesicular release at climbing fiber-Purkinje cell synapses. Neuron 32: 301-313. Eine ausführliche Literaturliste kann bei den Autoren angefordert werden. Studium der Biologie in Göttingen. 19871991: Diplom- und Doktorarbeit am MaxPlanck-Institut für biophysikalische Chemie, Abt. Neurobiologie (Prof. Dr. O.- D. Creutzfeldt). 1991-1993: Postdoctoral Fellow (Stipendiat der Royal Society of Science) am Department of Human Anatomy (Prof. Dr. Ray Guillery) der University of Oxford. 19931995: „Von Helmholtz“-Stipendiat des BMBF am Institut für Anatomie der Universität Freiburg (Prof. Dr. Michael Frotscher). 1997: Wolfgang-Bargmann-Preis der Anatomischen Gesellschaft. 1999: Habilitation im Fach Anatomie an der Universität Freiburg. 2000: Ernennung zum Hochschuldozenten C2. 2003: Berufung auf die Abteilungsleiterstelle Zelluläre Neurobiologie am Institut für Medizin, Forschungszentrum Jülich GmbH. Dirk Feldmeyer: geboren 1960; 1979-1983: Studium der Biologie, Germanistik und Pädagogik an der Ruhr-Universität Bochum. 19841987: Doktorarbeit am Institut für Zellphysiologie (Prof. Dr. H. C. Lüttgau). 1990-1995: Postdoctoral Fellow (DFG, Welcome Trust) am Department of Pharmacology, UCL London (Prof. S.G. Cull-Candy).1994: Forschungsaufenthalt (Stipendiat des British Council und des Japanese Ministry of Education) am Brain Research Institute in Tokyo (Prof. T. Takahashi). 1995: Berufung auf eine Arbeitsgruppenleiterstelle an das Max-Planck-Institut für medizinische Forschung, Abt. Zellphysiologie in Heidelberg (Direktor: Prof. Dr. med. B. Sakmann). 2000: Habilitation und Ernennung zum Hochschuldozenten für das Fach Physiologie an der Universität Heidelberg. 2001: Berufung zum Editor des Journals of Physiology (London). Ab August 2004 Berufung als Abteilungsleiter Zelluläre Neurobiologie am Institut für Medizin, Forschungszentrum Jülich GmbH. Korrespondenzadressen Danksagung Wir möchten an dieser Stelle der Deutschen Forschungsgemeinschaft für die kontinuierliche finanzielle Unterstützung in Form von Einzel bzw. SFB-Anträgen danken, die wesentlich zum Erfolg unserer Forschungsarbeiten beigetragen haben. Unser besonderer Dank gilt Prof. Dr. Bert Sakmann und Prof. Dr. Michael Frotscher für die stetige wissenschaftliche Unterstützung unserer Forschungsvorhaben. Kurzbiographien Joachim Lübke: geboren 1956; 1974-1977: Ausbildung zum biologisch-technischen Assistenten. 1981-1982: Begabten-Abitur am Abendgymnasium in Göttingen. 1982-1987: Joachim Lübke Institut für Medizin Abteilung Zelluläre Neurobiologie Forschungszentrum Jülich GmbH D-52425 Jülich Tel.: ++ 49 (0) 24 61 61 22 88 Fax: ++ 49 (0) 24 61 61 28 20 e-mail: [email protected] Dirk Feldmeyer Institut für Medizin Abteilung Zelluläre Neurobiologie Forschungszentrum Jülich GmbH D-52425 Jülich Tel.: ++49 (0) 24 61 61 59 14 Fax: ++49 (0) 24 61 61 28 20 e-mail: [email protected] Neuroforum 3/04 PETER DECHENT UND JENS FRAHM Funktionelle MagnetresonanzTomografie des menschlichen Gehirns Peter Dechent und Jens Frahm Technik liegt dagegen in ihrer hohen Sensitivität und der Möglichkeit, mit Hilfe radioaktiv markierter Substanzen eine molekular oder zellulär spezifische Bildgebung zu erreichen. Parallel zur PET-Technik hat sich in den vergangenen 20 Jahren die MagnetresonanzTomografie (MRT) zum wichtigsten Verfah- Zusammenfassung Die bildgebenden Verfahren der Magnetresonanz-Tomografie (MRT) sind nicht nur unverzichtbar für die medizinische Diagnostik, sie haben sich zudem zu wichtigen nichtinvasiven Werkzeugen in der biomedizinischen Forschung entwickelt. Dies gilt insbesondere für die grundlegenden und klinischen Neurowissenschaften. So ermöglicht die funktionelle MRT des menschlichen Gehirns eine detaillierte Charakterisierung kortikaler Systeme und Netzwerke von der primären Sensorik bis zur kognitiven Informationsverarbeitung. Aufgrund der wachsenden Verfügbarkeit geeigneter MRT-Geräte werden entsprechende Ansätze der Hirnbildgebung für Wissenschaftler unterschiedlicher Fachrichtungen zugänglich. Ziel dieses Beitrages ist es daher, die wesentlichen Elemente eines funktionellen MRT-Experimentes zu erläutern – von der Übersetzung der eigentlichen Fragestellung in ein geeignetes Stimulationsparadigma über die Aufnahme hämodynamisch vermittelter MRT-Signale als Korrelate neuronaler Aktivitätsänderungen bis hin zu Aspekten der Datenauswertung und Visualisierung. Letztendlich verspricht das Verfahren die reduktionistische Vorgehensweise in der Neurobiologie durch ein integratives Konzept zu ergänzen und genetische ebenso wie erfahrungsabhängige Einflüsse mit der Funktion des Gehirns zu verbinden. Abstract Functional magnetic resonance imaging of the human brain Apart from its leading role in diagnostic imaging of the central nervous system, magnetic resonance imaging (MRI) has evolved into an important research tool in both basic and clinical neuroscience. Functional MRI of the human brain offers a detailed characterization of cortical networks ranging from primary sensory processing to cognition. In view of the increasing availability of suitable MRI systems for neuroscientists across multiple disciplines, the purpose of this contribution is to discuss the key elements of a typical functional MRI experiment – the design of a paradigm that properly translates the actual question into an MRI-compatible stimulus protocol, the recording of hemodynamically mediated MRI signals that correlate with changes in neural activity, and specific challenges of data acquisition and analysis. The approach promises to complement molecular and cellular neurobiology by an integrative concept that links genetic and experience-dependent information to function of the intact brain. Key words: MRI; human brain; brain function; functional mapping Bildgebende Verfahren in der Hirnforschung Bis zur Mitte des 20. Jahrhunderts wurden Einblicke in die Funktionsweise des menschlichen Gehirns fast ausschließlich aus Läsionsstudien sowie mit Hilfe intrakortikaler Ableitungen elektrischer Signale und ebensolcher Stimulationen gewonnen. Systematische wissenschaftliche Studien waren daher durch die Verfügbarkeit geeigneter Patienten einerseits und die Invasivität der verwendeten Methoden andererseits erheblich eingeschränkt. Erst mit der Entwicklung nicht-invasiver und insbesondere bildgebender Verfahren konnte die Erforschung des Neuroforum 3/04 intakten menschlichen Gehirns in den vergangenen zwei Jahrzehnten entscheidend vorangetrieben werden. Erste eindrucksvolle Ergebnisse wurden seit Beginn der 80er Jahre mit der Positronen-Emissions-Tomografie (PET) erzielt, obwohl das Verfahren nur über eine begrenzte räumliche (lineare Bildelemente von etwa 5-8 mm) und zeitliche Auflösung (etwa eine Minute) verfügt. Zudem führt die Notwendigkeit, ein kurzlebiges radioaktives Präparat zu verabreichen, zu einem erheblichen radiochemischen Aufwand und einer Strahlenbelastung, die einen uneingeschränkten Einsatz bei gesunden Versuchspersonen verhindert. Die besondere Chance der PET- Abb. 1: Schema eines fMRT-Experimentes. ren der diagnostischen Bildgebung entwikkelt. Komplementär zu Röntgentechniken stellt die MRT nicht die Knochen, sondern die Verteilung des Wassers in den Körpergeweben dar und bietet einen exzellenten Weichgewebekontrast. Zu Beginn der 90er Jahre hat man zudem entdeckt, dass sich bestimmte Messvarianten nutzen lassen, um Hirnfunktionen bildlich darzustellen: genauer gesagt, die mit ihnen verbundenen regionalen Änderungen des Blutflusses und der Sauerstoffversorgung im Gewebe. Die folgenden Abschnitte beschreiben zunächst die wesentlichen Grundlagen des Standardver229 FUNKTIONELLE MAGNETRESONANZ-TOMOGRAFIE DES MENSCHLICHEN GEHIRNS A B C D E F Abb. 2: Die Elemente eines fMRT-Experimentes (hier: visuelle Stimulation mit einem Schachbrettmuster im Vergleich zu einem grauen Bild) betreffen: (A) die Wahl eines Volumens (hier: eine Schicht durch den okzipitalen Kortex), (B) die MRTDarstellung der Anatomie, (C) die Aufnahme einer dynamischen Zeitserie von MRTBildern während der Kontrollbedingung und (D) während der Stimulationsbedingung (Pfeile: Bereiche mit erhöhter Signalintensität im okzipitalen Kortex), (E) eine Grauwert-Karte der berechneten Korrelationskoeffizienten (hell: hoch korreliert, grau: nicht korreliert, dunkel: entgegengesetzt korreliert) und (F) die Überlagerung einer farbkodierten Karte der statistisch signifikanten Aktivierungen mit einem geeigneten Referenzbild. Komplex aus Sauerstoff und Hämoglobin ist Desoxyhämoglobin ein paramagnetisches Molekül, das die MRT-Eigenschaften der benachbarten Wassermoleküle beeinflusst und wie ein endogenes Kontrastmittel wirkt. Beispielsweise führen erhöhte Desoxyhämoglobin-Konzentrationen – etwa nach einer kurzen Anoxie oder einem Anhalten des Atems – zu einer MRT-Signalreduktion in venösen Gefäßen und unmittelbar benachbartem Hirngewebe. Ein durch eine veränderte Hirnfunktion erzeugter positiver BOLD-Kontrast, insbesondere ein MRT-Signalanstieg bei visueller Stimulation, wurde erstmals 1992 von mehreren Arbeitsgruppen beschrieben (Bandettini et al. 1992; Blamire et al. 1992; Frahm et al. 1992; Kwong et al. 1992). Seitdem hat sich dieser Ansatz aufgrund seiner einfachen Anwendung und hohen Sensitivität zusammen mit der EchoPlanar-Tomografie als schneller Bildgebungssequenz für die Mehrzahl der fMRTAnwendungen durchgesetzt. Ebenso wie die elektromagnetischen Signaländerungen einer Elektroenzephalogramm (EEG)-Aufzeichnung vermitteln die hämodynamischen Antworten der fMRT nur ein indirektes Korrelat veränderter Hirnaktivität. Das Schema in der Abbildung 1 verdeutlicht, dass zunächst die wissenschaftliche Fragestellung in ein geeignetes Stimulationsparadigma übersetzt werden muss, welches auf neuronaler Ebene zu einer re- gionalen Änderung der synaptischen Aktivität führen sollte. Dieser Wechsel zwischen zwei Hirnfunktionszuständen erzeugt mittels neurovaskulärer Kopplungsmechanismen einen Stimulus-bedingten BOLD-Kontrast, der sich bezüglich der zugrundeliegenden Desoxyhämoglobin-Konzentration als das Ergebnis metabolischer (erhöhter Sauerstoffverbrauch = mehr Desoxyhämoglobin) und physiologischer Prozesse (stark erhöhter Blutfluss = viel weniger Desoxyhämoglobin) ergibt. In der Tat führt eine erhöhte Hirnaktivität in der Regel zu einer Netto-Reduktion der Desoxyhämoglobin-Konzentration und damit zu einem fMRT-Signalanstieg entsprechend einer positiven BOLD-Antwort. Schließlich erfordert die dynamische Aufzeichnung von Signaländerungen in einer fMRT-Bildserie, die fortlaufend während mehrfacher Stimulus-Wechsel gemessen wird, eine Nachverarbeitung der eigentlichen Bilddaten. Sie beinhaltet in der Regel eine zeitliche Korrelation des Paradigmas mit den dynamischen BOLD-Signalen sowie eine statistische Bewertung und Visualisierung der Ergebnisse als zwei- oder dreidimensionale Karte der Hirnaktivitätsänderungen. Die Abbildung 2 veranschaulicht den praktischen Ablauf eines einfachen fMRTExperimentes. Zunächst wird das interessierende Hirnvolumen, in diesem Beispiel eine einzelne Schicht durch den okzipital gelegenen visuellen Kortex ausgewählt (2A) und fahrens der funktionellen MRT (fMRT), das auf dem sogenannten BOLD-Kontrast beruht (BOLD = blood oxygenation level dependent). Anschließend werden einige Bedingungen und Probleme diskutiert, die bei der praktischen Durchführung einer fMRT-Untersuchung unbedingt Beachtung verdienen. Die physikalischen Grundlagen der Magnetresonanz-Verfahren werden in einem Exkurs dargestellt. Funktionelle MagnetresonanzTomografie Der erstmalig von Ogawa et al. (1990) am Gehirn der Ratte beschriebene BOLD-Kontrast bezieht sich auf MRT-Bilder, die mit einem Gradientenechosignal aufgenommen werden, und spiegelt Veränderungen in der intravaskulären Konzentration des Desoxyhämoglobins wider. Im Gegensatz zum 230 Abb. 3: Das Ergebnis eines fMRT-Experimentes zur funktionellen Charakterisierung des visuellen Systems (20/22 Schichten, räumliche Auflösung 2x2x2 mm3, Messzeit 4.5 Minuten) zeigt Aktivierungen (rot) und Deaktivierungen (blau). Neuroforum 3/04 PETER DECHENT UND JENS FRAHM mit konventionellen anatomischen MRTAufnahmen dokumentiert (2B). Dieses Volumen wird dann mit einer Repetitionszeit von etwa 2-3 s für üblicherweise einige Minuten aufgenommen, wodurch man einen dynamischen Rohdatensatz von MRT-Bildern erhält. Zeitgleich werden der Testperson visuelle Stimuli für jeweils einige Sekunden (entsprechend einiger sequentieller MRT-Bilder) präsentiert. Da die BOLDfMRT lediglich Änderungen der Hirnaktivität identifiziert, müssen mindestens zwei verschiedene Stimuli miteinander verglichen werden: hier sind es ein schwarz-weiß-invertierendes Schachbrettmuster (Stimulationsbedingung) und ein grauer Bildschirm (Kontrolle). Eine absolute Quantifizierung von Hirnaktivität ist mit der fMRT nicht möglich. Bei einem Vergleich eines gemittelten Bildes aus der Kontroll- (2C) und Stimulationsphase (2D) sind bereits Signalerhöhungen in den mit Pfeilen markierten Bereichen des visuellen Kortex zu erkennen. Für die genaue Kartierung entsprechender Areale bietet sich aufgrund der zeitlichen Charakteristik der MRT-Signaländerungen eine (Kreuz-) Korrelationsanalyse an, die das Stimulationsparadigma als Rechteckfunktion mit dem MRT-Intensitätsverlauf in jedem Bildpunkt vergleicht und eine GrauwertKarte der Korrelationskoeffizienten erzeugt (2E). Nach einer statistischen Bewertung (Baudewig et al. 2003) werden die Bildpunkte mit signifikanten Signaländerungen farbkodiert und vorzugsweise dem Originalschnittbild überlagert (2F). Um der hämodynamisch verzögerten BOLD-Antwort Rechnung zu tragen (siehe unten), wird die Referenzfunktion bei der Berechnung der Korrelationskarte in der Regel um 2-4 s verschoben. Als Erweiterung dieses einfachen Ansatzes kann die Rechteckfunktion darüber hinaus mit einer geeigneten hämodynamischen Antwortfunktion gefaltet werden. Dies führt insbesondere bei ereigniskorrelierten Paradigmen, die aus einer Folge sehr kurzer und schnell aufeinanderfolgender Stimuli bestehen, zu einer verbesserten Sensitivität. Mit dem beschriebenen Grundkonzept ist es möglich, nicht nur eine einzelne Schicht, sondern gleichzeitig weite Teile des Gehirns funktionell zu untersuchen. Beispielsweise zeigen die Abbildungen 3 und 4 eine umfassende Beteiligung des okzipitalen und parietalen Kortex an der visuellen Informationsverarbeitung. Obwohl die Darstellung der Ergebnisse als zweidimensionale Aktivierungskarte die häufigste Form der Visualisierung ist, erschwert sie in vielen Fällen, beispielsweise bei gekippter SchichtfühNeuroforum 3/04 schließender Extrahierung der Kortexoberfläche (6B) einen weiteren Verarbeitungsschritt dar. Um allerdings die Projektion der funktionellen Daten auf die komplexe Faltung der Hirnoberfläche zu vermeiden, kann die Kortexoberfläche mathematisch „entfaltet“ und in eine flache Karte der Hirnwindungen und -furchen mit überlagerten Aktivierungen überführt werden (6C). Als weitere Option zur Auswertung der funktionellen Ergebnisse bietet sich eine Gruppenanalyse an. Dabei sind jedoch interindividuelle Unterschiede sowohl in der Hirnanatomie als auch bei der Ausführung des Paradigmas – vorzugsweise bei höheren kognitiven Aufgaben – zu berücksichtigen. Auch die Transformation der individuellen Hirnanatomie in ein standardisiertes Koordinatensystem bedingt bereits einen Informationsverlust durch räumliche Ungenauigkeiten und Filterungen. Mechanismen Abb. 4: Ausgewählte Schichten aus Abb. 3 zeigen außer striatären und extrastriatären visuellen Arealen die lateralen Kniekörper, in denen die visuelle Information von den optischen Trakten auf die Sehstrahlungen übertragen wird, die frontalen Augenfelder, die die motorische Steuerung von Augenbewegungen vermitteln und den Intraparietalkortex, der an Prozessen wie der räumlichen Wahrnehmung, Navigation und mentalen Rotation beteiligt ist. rung, die genaue anatomische Zuordnung eines aktivierten Areals. In solchen Fällen können die funktionellen Daten, wie in Abbildung 5 illustriert, mit einem dreidimensionalen anatomischen Datensatz koregistriert und aus allen drei Raumrichtungen betrachtet werden. Die Abbildung 6 stellt mit der Segmentierung des Gehirns und an- Trotz vielfältiger Anwendungen der fMRT sind die exakten Mechanismen der Übertragung neuronaler Aktivitätsänderungen in eine hämodynamische Reaktion noch nicht vollkommen verstanden. Dies trifft auch auf die zentrale Frage nach dem Ursprung der von der fMRT beobachteten BOLD-Antwort zu: Ist eine Membrandepolarisation am Axon oder eine synaptische Übertragung der eigentliche Auslöser? Erste wichtige Hinweise lieferten Messungen des Blutflusses in Kombination mit der Aufzeichnung von single-unit Aktivitäten (Aktionspotentiale) und lokalen Feldpotentialen (synaptische Aktivität) am zerebellären Kortex der Ratte (Mathiesen et al. 1998). Darauf aufbauend untersuchten Logothetis et al. (2001) gleichzeitig multi-unit Aktivitäten, lokale Feldpotentiale und Abb. 5: Koregistrierung der zweidimensional gemessenen Aktivierungen (hier: das extrastriatäre bewegungssensitive visuelle Areal V5) mit einer dreidimensionalen MRTAufnahme der Anatomie. 231 FUNKTIONELLE MAGNETRESONANZ-TOMOGRAFIE DES MENSCHLICHEN GEHIRNS Abb. 6: Erweiterte Darstellungen der (A) koregistrierten funktionellen und anatomischen Daten (hier: der Intraparietalkortex) durch eine (B) Segmentierung und (C) anschließende Entfaltung des Gehirns, die nur noch der Oberfläche der grauen Hirnsubstanz entspricht (hellblau: Hirnwindungen, dunkelblau: Hirnfurchen). BOLD fMRT-Signale an anästhesierten Affen während visueller Stimulation. Sie konnten zeigen, dass die zeitliche Entwicklung der hämodynamischen Reaktion besser mit derjenigen der lokalen Feldpotentiale und damit synaptischer Übertragung korreliert, als mit der Charakteristik von multi-unit Aktivitäten. Somit korrespondiert die in der fMRT üblicherweise dargestellte Hirnaktivität eher mit der Verarbeitung eingehender Informationen als der Übermittlung ausgehender Signale – unabhängig davon, dass in den meisten Fällen beide Aspekte eng miteinander gekoppelt sind. Dieses Konzept wird durch Untersuchungen am Menschen mit der transkraniellen Magnetstimulation (TMS) und gleichzeitiger fMRT unterstützt (Baudewig et al. 2001a; Bestmann et al. 2003). Die TMS moduliert in Abhängigkeit von der Intensität die elektrische Hirnaktivität im Kortex – beispielsweise im primär motorischen Areal (M1) – durch die externe Applikation sehr kurzer (etwa 200 µs), aber intensiver Magnetfeldpulse (etwa 2 Tesla). In Abbildung 7 werden Aktivierungskarten bei überschwelliger repetitiver TMS (rTMS) von M1 mit Auslösung einer Fingerbewegung mit den Ergebnissen bei unterschwelliger rTMS ohne periphere Muskelaktivität verglichen. Während überschwellige rTMS die primären motorischen und somatosensorischen Areale (M1/ S1) sowie das supplementäre motorische Areal (SMA) aktiviert, führt die unterschwellige Stimulation zu reduzierter Aktivierung im SMA und zu keiner Aktivierung in M1/S1. Diese Beobachtung deutet darauf hin, dass nicht die in M1/S1 erzeugten Ak232 tionspotentiale oder ihre Modulation durch unterschwellige rTMS eine hämodynamische Reaktion auslösen, sondern nur die im SMA über kortiko-kortikale Faserverbindungen eingehenden Informationen, die eine synaptische Übertragung erfordern. In physiologischer Hinsicht entspricht eine positive BOLD-Antwort, d.h. ein fMRT-Signalanstieg in Reaktion auf eine neuronale Aktivitätsänderung, einer Verringerung der absoluten Konzentration an Desoxyhämoglobin in den lokalen Mikrogefäßen. Die zugrunde liegenden Mechanismen, ebenso wie ihre jeweiligen zeitlichen Charakteristika, sind weiterhin Gegenstand vor allem tierexperimenteller Untersuchungen und theoretischer Modellierungen. Übereinstimmung besteht darin, dass eine funktionell bedingte Änderung der Hirnaktivität regionale Änderungen des Sauerstoffverbrauchs, des (arteriellen) Blutflusses und des (venösen) Blutvolumens verursacht. Offenkundig kommt es zunächst zu einem erhöhten Sauerstoffverbrauch, also einer verstärkten Produktion von Desoxyhämoglobin, dem mit kurzer Latenz eine überproportionale Erhöhung des Blutflusses folgt. Dieser Effekt bedingt ein Überangebot an oxygeniertem Hämoglobin und wäscht somit einen Teil des Desoxyhämoglobins aus dem venösen Schenkel der Mikrozirkulation aus. Zeitlich verzögert folgt dem erhöhten Blutfluss eine Zunahme des Blutvolumens, was wiederum eine Erhöhung der Desoxyhämoglobin-Konzentration in betroffenen Bildelementen zur Folge hat. In der Summe überwiegt im gesunden Gehirn der Einfluss des Blutflusses und die damit verbundene Abnahme der Desoxyhämoglobin-Konzentration. Die zu erreichende zeitliche und räumliche Auflösung der BOLD-fMRT hängt daher maßgeblich von der zeitlichen Entwicklung der einzelnen Mechanismen ab. Zeitliche Auflösung Abb. 7: Die Kombination der fMRT mit der transkraniellen Magnetstimulation (TMS) über dem primären motorischen Handareal führt bei überschwelliger Stimulation (links: 110% RMT) zu Aktivierungen der primären motorischen und sensorischen Areale (M1/S1) sowie des supplementären motorischen Areals (SMA). Im Gegensatz dazu aktiviert eine unterschwellige Stimulation (rechts: 90% AMT) nicht M1/ S1, sondern nur das axonal verbundene SMA. Verändert aus Bestmann et al. (2003). Die Abbildung 8 verdeutlicht die zeitliche Entwicklung der hämodynamischen Antwortfunktion am Beispiel einer einfachen visuellen Stimulation. Der initial erhöhte Sauerstoffverbrauch ist mit den üblicherweise verwendeten fMRT-Verfahren nicht zu detektieren. Ob sich seine Existenz durch einen kurzen Signalabfall – also eine negative BOLD-Antwort – tatsächlich nachweisen lässt, wird in der Literatur zudem kontrovers diskutiert. Stattdessen zeigt sich in der Regel als Antwort auf einen 10 s Stimulus ein um etwa 2 s verzögerter, durch die lokale Blutflusserhöhung verursachter Signalanstieg, der sein Maximum nach etwa 10 bis 15 s erreicht (8A). Nach Beendigung der Stimulation fällt das BOLD-Signal wiederum mit einer erheblichen Latenz zunächst bis unter die ursprüngliche Basisintensität ab bevor es nach etwa 60 s auf den Ausgangswert zurückkehrt (Fransson et al. 1998). Der Neuroforum 3/04 PETER DECHENT UND JENS FRAHM das Signal-Rausch-Verhältnis, den funktionellen Kontrast und die durch Suszeptibilitäten verursachten Signalauslöschungen und Verzerrungen. So kann ein großes Bildelement, d.h. eine schlechte räumliche Auflösung, zu falsch-positiven Aktivierungen aufgrund von Partialvolumeneffekten mit nicht-aktivierter Hirnsubstanz führen oder in der Nähe von Gewebe-Luft-Übergängen erhebliche Signalauslöschungen verursachen. Auch die Verringerung der Auflösung durch räumliches Filtern der Daten im Rahmen der Nachverarbeitung sowie durch eine Transformation in standardisierte Koordinatensysteme führt zu einer weiteren Verschlechterung der Spezifität der beobachteten Aktivierungen und ihrer Kongruenz mit der Hirnanatomie. Im Allgemeinen werden fMRT-Untersuchungen mit einer räumlichen Auflösung von 2-4 mm Kantenlänge in der Bildebene und 2-6 mm Schichtdicke verwendet. Damit ist bei Einzelmesszeiten eines fMRT- Abb. 8: Die zeitliche Charakteristik der hämodynamischen Antwortfunktion für eine visuelle Stimulation (schraffierte Boxen) mit unterschiedlich langen Stimulations- und Kontrollbedingungen: (A) 10 s Stimulation mit 90 s bzw. (B) 20 s Kontrolle, (C) 0.2 s Stimulation mit 90 s bzw. (D) 9.8 s Kontrolle. Verändert aus Fransson et al. (1998) und (1999). post-Stimulus undershoot-Effekt wird aufgrund von Kontrastmittelstudien am Nagergehirn zumeist als ein anhaltend erhöhtes Blutvolumen bei bereits normalisiertem Blutfluss verstanden. Allerdings können Beiträge durch einen ebenfalls nachhaltig erhöhten Sauerstoffverbrauch nicht ausgeschlossen werden. Ein qualitativ gleiches Bild der BOLDAntwort ergibt sich auch bei erheblich verkürzter Stimulusdauer (8C) (Fransson et al. 1999). Vor allem die relativ lang andauernden negativen Signalkomponenten müssen bei der Planung eines fMRT-Experimentes sowie beim Design eines geeigneten Stimulationsparadigmas beachtet werden. So verdeutlichen die Abbildungen 8B und 8D, dass die für ein ausreichend gutes KontrastRausch-Verhältnis notwendigen Wiederholungen der Stimulations- und Kontrollperioden einen gewissen zeitlichen Abstand erfordern, um eine physiologische Trennung der beteiligten Hirnfunktionszustände zu gewährleisten. Daraus ergibt sich, dass viele neuropsychologische Standardtests nicht ohne weiteres in ein fMRT-taugliches Stimulationsparadigma übersetzt werden können. Die nur begrenzte zeitliche Auflösung der fMRT wird zum Teil durch ihre Fähigkeit kompensiert, auch BOLD-Reaktionen auf Neuroforum 3/04 sehr kurze und/oder einmalige Reize zu detektieren. Diese Empfindlichkeit kann insbesondere durch die Ableitung eines EEGSignals während der fMRT-Messung für eine räumlich hochaufgelöste Darstellung einzelner kortikaler (pathologischer) Ereignisse genutzt werden. Die Abbildung 9 zeigt mögliche Anwendungen der kombinierten EEGfMRT am Beispiel eines Epilepsie-Patienten, der auch zwischen den generalisierten Anfällen Epilepsie-typische Muster im EEG aufweist. Derartige interiktale EEG-Veränderungen wurden während einer kontinuierlichen fMRT-Messung (im Ruhezustand ohne Stimulation) aufgezeichnet und nach der Untersuchung für eine Korrelationsanalyse mit den fMRT-Daten verwendet. Auf diese Weise konnte ein eng umschriebenes Gebiet in der Inselregion des Patienten mit den EEG-synchronen Aktivitätsänderungen identifiziert werden. Inwieweit dieses Areal die eigentliche Quelle der epileptischen Aktivität oder nur ein Projektionsgebiet darstellt, verbleibt eine zu klärende Frage (Baudewig et al. 2001b). Räumliche Auflösung Die Wahl der räumlichen Auflösung für ein fMRT-Experiment ist oft von entscheidender Bedeutung. Sie hat direkten Einfluss auf Abb. 9: Die Kombination der fMRT mit EEGMessungen an einem Patienten mit generalisierter Epilepsie ergibt (oben) ein Areal kortikaler Aktivität in der Inselregion, das (unten) durch eine Korrelation mit den im EEG identifizierten Epilepsietypischen Mustern (schraffierte Boxen) berechnet wurde. Verändert aus Baudewig et al. (2001b). 233 FUNKTIONELLE MAGNETRESONANZ-TOMOGRAFIE DES MENSCHLICHEN GEHIRNS Abb. 10: Der Einfluss der räumlichen Auflösung auf die Spezifität der Aktivierungskarten: (A) 3x3 mm2, (B) 2x2 mm2, (C) 1x1 mm2 und (D) 0.5x0.5 mm2 bei jeweils gleicher Schichtdicke (4 mm). Bildes von unter 100 ms eine Volumenabdeckung großer Teile des Gehirns mit einer zeitlichen Auflösung von 2-3 s möglich (vgl. Abbildungen 3, 7 und 9). Die Vorteile einer höheren räumlichen Auflösung für spezifische Fragestellungen werden in Abbildung 10 deutlich. Die visuellen Repräsentationen in den primären Arealen der Sehrinde leiden bei einer häufig gewählten Auflösung von nur 3x3 mm2 (10A) stark unter Partialvolumeneffekten. Die Erhöhung der Auflösung über 2x2 mm2 (10B) zu 1x1 mm2 (10C) führt zwar zu einer deutlichen Verringerung dieser Problematik, jedoch erst bei 0.5x0.5 mm2 (10D), d.h. bei einer Reduktion des Bildelementvolumens um den Faktor 36 im Vergleich zu Abb. 10A, beschränkt sich die Aktivierung weitgehend auf die graue Hirnsubstanz. Der Preis sehr hoher räumlicher Auflösung ist eine Verlängerung der Messzeit für ein einzelnes fMRT-Bild, so dass entsprechende Untersuchungen im Grenzfall auf eine einzelne Schicht begrenzt werden müssen, um eine Zeitauflösung von 2-3 s aufrechtzuerhalten. Neurowissenschaftliche Anwendungen beziehen sich beispielsweise auf Fragen nach der Kodierung einzelner Fingerrepräsentationen im primären motorischen Kortex (siehe unten) oder auf die Untersuchung der augenspezifischen Okulardominanzbanden im primären visuellen Kortex (Dechent und Frahm 2000). Probleme und Herausforderungen Die praktische Ausführung eines fMRT-Experimentes ist durch vielfältige technische und physiologische Probleme gefährdet. Insbesondere ist eine intakte neurovaskuläre Kopplung die Voraussetzung dafür, 234 dass neuronale Aktivitätsänderungen tatsächlich ihre hämodynamische Entsprechung finden. Pharmakologische Modulationen, auch im Zusammenhang mit vasoaktiven Medikamenten bei der Untersuchung von Patienten, die Gabe von Anästhetika etwa bei der Untersuchung von Kindern, oder pathologische Veränderungen bei zerebrovaskulären Erkrankungen sind nur einige Beispiele, bei denen diese Grundlage der BOLD-fMRT in Frage gestellt ist. In technischer Hinsicht stellen vor allem Stimulus-korrelierte (Kopf-) Bewegungen ein ernstzunehmendes Problem dar, da sie an starken Kontrastgrenzen, etwa zwischen der grauen Hirnsubstanz und der zerebrospinalen Flüssigkeit, falsch-positive Aktivierungen vortäuschen können. Suszeptibilitätsartefakte und geometrische Verzerrungen an den Übergängen zwischen Hirngewebe und luftgefüllten Räumen, beispielsweise in den basalen Anteilen des Gehirns, betreffen die vielleicht schwerwiegendste Herausforderung. Sie wird in Abbildung 11 am Beispiel der Amygdala demonstriert Abb. 11: Suszeptibilitätsartefakte und geometrische Verzerrungen. (A) Während die bilateral lokalisierte Amygdala (Pfeile) in strukturellen MRT-Aufnahmen gut zu erkennen ist, führen (B) Suszeptibilitätsartefakte in dieser Region bei einer in der Literatur verwendeten Auflösung von 3.75x3.75 mm2 zu einer völligen Auslöschung in den funktionellen MRT-Bildern. Verändert aus Merboldt et al. (2001). (C) Geometrische Verzerrungen der fMRTBilder verschieben die Aktivierungen (hier: Fingerbewegungen im primären motorischen Handareal) bei einer Überlagerung der Aktivierung mit einem anatomischen MRT-Bild fälschlich auf die postzentrale Hirnwindung. (D) Dieses Problem kann nur vermieden werden, wenn die Aktivierungskarte einem gleichermaßen verzerrten Originalbild überlagert wird, so dass die motorische Repräsentation erwartungsgemäß entlang der Zentralfurche erscheint. (Merboldt et al. 2001), die aufgrund ihrer Beteiligung an der Emotionsverarbeitung immer häufiger im Fokus von fMRT-Studien steht. Da die verantwortlichen makroskopischen Suszeptibilitätseffekte entscheidend vom Volumen der einzelnen Bildelemente abhängen, können Signalauslöschungen wie in der Amygdala-Region (11B) bis zu einem gewissen Grad durch eine Erhöhung der räumlichen Auflösung und/oder eine Verringerung der Schichtdicke vermieden werden. Auf jeden Fall ist es erforderlich, dass entsprechende fMRT-Untersuchungen die Qualität ihrer Ergebnisse durch eine Dokumentation der Originalbilder belegen. Ein direkt verwandtes Problem sind geometrische Verzerrungen und die sich daraus ergebende räumliche Inkongruenz zwischen fMRT-Bildern (einschließlich der Aktivierungskarten) und hochaufgelösten anatomischen MRT-Bildern. Eine unkritische Überlagerung wie in Abbildung 11C führt dann zu qualitativ falschen Aussagen: hier wurde die motorische Repräsentation der Hand auf die postzentrale Hirnwindung, also in den somatosensorischen Kortex, positioniert. Diese künstliche Verschiebung verschwindet erst, wenn die Aktivierungskarte einem fMRT-Originalbild aus der gleichen Messung überlagert wird (11D). Für das genaue Ergebnis eines fMRT-Experimentes ist nicht zuletzt das exakte Design des Stimulationsparadigmas von entscheidender Bedeutung. Wie eine scheinbar subtile Änderung zu einem völlig anderen Ergebnis führen kann, ist in Abbildung 12 am Beispiel der Fingersomatotopie im M1 Handareal verdeutlicht. Nach Entfaltung des Kortex im Bereich der Zentralfurche weisen die kortikalen Repräsentationen einzelner Fingerbewegungen bei einer Kontrolle gegen Ruhe große Überlappungen auf (Abbildung 12, obere Reihe). Verwendet man dagegen einen differentiellen Ansatz (Abbildung 12, untere Reihe), in dem einzelne Fingerbewegungen gegen die Bewegung eines anderen Fingers verglichen werden, so erhält man eine somatotope Anordnung funktionell dominanter Repräsentationen, die vom Daumen (D1) im anterio-inferio-lateralen Bereich bis zum kleinen Finger (D5) im posterio-superiomedialen Anteil von M1 reichen (Dechent und Frahm 2003). Dieses scheinbar widersprüchliche Ergebnis wird verständlich bei Betrachtung der unterschiedlichen Aufgabenstellungen für Fingerbewegungen: einerseits muss bei Greifbewegungen der Hand die Koordination mehrerer Finger gewährleistet sein, andererseits müssen auch Neuroforum 3/04 PETER DECHENT UND JENS FRAHM völlig isolierte Einzelfingerbewegungen wie etwa beim Spielen eines Musikinstrumentes möglich sein. Exkurs Ausblick Exkurs: Physikalische Grundlagen Die Erforschung der Funktionsweise des menschlichen Gehirns mit Hilfe der fMRT wird sich von der bisherigen Konzentration auf primäre sensorische und motorische Systeme auf die Informationsverarbeitung höherer kognitiver und emotionaler Leistungen und ihren Störungen erweitern. Bereits jetzt zeigen die vielfältigen Untersuchungsmöglichkeiten zunehmend auch in einfachen sensorischen Systemen eine starke Modulation durch Aufmerksamkeitseffekte oder sogenannte top-down-Prozesse. Herausforderungen entstehen vor allem durch die größere Komplexität der zu untersuchenden Prozesse, ihre im Gegensatz zu exekutiven Motorfunktionen nur begrenzt überprüfbare Ausführung und den großen Einfluss selbst kleinster Unterschiede in der Wahl des Stimulationsparadigmas. Darüber hinaus ist zu erwarten, dass wachsende interindividuelle Unterschiede in der strategischen Bearbeitung einer kognitiven Aufgabe die Interpretation der Ergebnisse erschweren werden. In methodischer Hinsicht wird die durch die Echo-Planar-Messtechnik bedingte Empfindlichkeit gegenüber Suszeptibilitätssprüngen als grundsätzliches Problem weitestgehend bestehen bleiben und vor allem Untersuchungen des ventralen frontalen Kortex, des vorderen Hippocampus und der Amygdala belasten. Verbesserungen sind Bei den Verfahren der nuklearmagnetischen Resonanz (NMR) wird der Untersuchungsgegenstand oder die Versuchsperson in ein statisches Magnetfeld eingebracht (hier: 3 Tesla = etwa 60.000-faches Erdmagnetfeld). Dadurch richten sich die magnetischen Dipolmomente geeigneter Atomkerne (hier: Wasserstoff) parallel und antiparallel zum statischen Magnetfeld aus. Aufgrund des Besetzungsunterschiedes zwischen den energetisch ungleichen Orientierungen stellt sich eine Gleichgewichtsmagnetisierung in Richtung des statischen Magnetfeldes ein. Dieser Gleichgewichtszustand kann durch einen kurzen Hochfrequenz-Impuls im UKW-Bereich (hier: 123 MHz) gestört werden. Anschließend kehrt die Magnetisierung in den Gleichgewichtszustand zurück, wobei die Atomkerne ihrerseits ein Radiofrequenzsignal abgeben, das als NMR-Signal aufgezeichnet und weiterverarbeitet werden kann. Für die Magnetresonanz-Tomografie (MRT) werden räumlich unterscheidbare NMR-Signale durch magnetische Zusatzfelder (Magnetfeldgradienten) erzeugt, die weniger durch immer höhere Magnetfeldstärken zu erwarten, die diese Probleme linear verstärken, sondern vielmehr durch parallele Bildgebungstechniken wie sie zur Zeit allgemein auf modernen MRT-Syste- sowohl während der Hochfrequenz-Anregung als auch während der Datenaufnahme geschaltet werden. Da die Frequenz des NMR-Signals direkt proportional zur Magnetfeldstärke ist, entspricht das durch Fourier-Transformation erhaltene konventionelle NMR-Spektrum bei örtlich variabler Magnetfeldstärke einer eindimensionalen Projektion der Intensität des Untersuchungsobjektes in Richtung des zugeschalteten Gradienten. Aus einer Vielzahl von Einzelmessungen mit veränderter Ortskodierung lassen sich zwei- oder dreidimensionale Bilder des Objektes rekonstruieren. Die strukturelle Bildgebung stellt die derzeit wichtigste Anwendung der MRT mit etwa 60 Millionen Untersuchungen jährlich dar. Sie profitiert vor allem von den exzellenten Weichgewebekontrasten des Verfahrens sowie einer hohen Sensitivität bei pathologischen Veränderungen. Darüber hinaus weist die MRT eine Vielzahl spezieller Techniken auf, von denen für neurowissenschaftliche Fragen vor allem spektroskopische Untersuchungen des Hirnstoffwechsels, diffusionsbasierende Darstellungen der axonalen Konnektivitäten und individuellen Nervenfaserverläufe sowie funktionelle Kartierungen kortikaler Systeme von Interesse sind. men eingeführt werden. Mittelfristig werden zudem fMRT-Untersuchungen mit simultaner EEG-Aufzeichnung an Bedeutung gewinnen: Sie verbinden zwei unabhängige physiologische Korrelate neuronaler Abb. 12: Fingerrepräsentationen im primären motorischen Handareal (M1) nach Segmentierung und Entfaltung des Gehirns (vgl. Abb. 6). (Oben) Während eine Bewegung gegen Ruhe zu starken Überlappungen führt, ergibt (unten) ein differentieller Vergleich unterschiedlicher Fingerbewegungen eine geordnete somatotope Reihung von D1: Daumen, D2: Zeigefinger, D3: Mittelfinger, D4: Ringfinger und D5: kleiner Finger. Verändert aus Dechent und Frahm (2003). Neuroforum 3/04 235 FUNKTIONELLE MAGNETRESONANZ-TOMOGRAFIE DES MENSCHLICHEN GEHIRNS Aktivität sowie komplementäre technische Stärken und bieten damit hervorragende Voraussetzungen für neue Einsichten in die raumzeitliche Organisation wesentlicher Hirnfunktionen und ihrer Störungen. Vorhersehbare Anwendungen reichen von der Charakterisierung epileptogener Prozesse bis zur Verfolgung synaptischer Plastizität im Zusammenhang mit Reorganisationsprozessen bei Lernvorgängen oder bei der Rehabilitation nach Hirnverletzungen. Klinische Anwendungen der fMRT versprechen ein großes Potential für die Diagnostik neurologischer und psychiatrischer Erkrankungen. Allerdings muss vor allzu schnellen und unkritischen Einsätzen gewarnt werden. Zur Zeit wird die fMRT vielfach in der prächirurgischen Funktionsdiagnostik erprobt, um intaktes Gewebe von der zu entfernenden zerebralen Raumforderung abzugrenzen. Voraussetzung dabei ist, dass für die spezifische Hirnregion ein geeignetes Paradigma vorliegt. Während dies für viele primäre sensomotorische Areale gegeben ist, gestaltet sich die praktische Umsetzung für Gebiete mit multimodaler kognitiver oder gar unbekannter Kodierung als sehr schwierig bis unmöglich. Psychiatrische Erkrankungen stellen daher für die fMRT ein ebenso lohnendes wie schwieriges Feld dar. Beispielsweise gibt es bisher nur wenige neuropsychologische Testverfahren, die in ein robustes – hämodynamisch kompatibles – fMRT-Stimulationsparadigma übertragen werden konnten oder gar bezüglich der Vergleichbarkeit der Testergebnisse an gesunden Versuchspersonen ausreichend überprüft wurden. Zusammenfassend ist festzustellen, dass sich die wachsende Bedeutung der fMRT für die Hirnforschung vor allem aus der Nicht-Invasivität, dem raumzeitlichen Auflösungsvermögen, der Empfindlichkeit und der großen experimentellen Flexibilität ergibt. Damit begegnet das Verfahren der Erkenntnis, dass sich spezifische kognitive und emotionale Leistungen des Gehirns nicht allein aus den zugrundeliegenden molekularbiologischen Komponenten oder synaptischen Übertragungsmechanismen ableiten lassen, sondern Untersuchungen des intakten Gesamtsystems erfordern. Die reduktionistische Vorgehensweise in der neurobiologischen Grundlagenforschung wird auf diese Weise durch ein integratives Konzept ergänzt, das die genetische Information – über den Schritt geeigneter Tiermodelle – mit der Funktion des Gehirns verbindet. 236 Literatur Bandettini, P.A., Wong, E.C., Hinks, R.S., Tikofsky, R.S. und Hyde, J.S. (1992): Time course EPI of human brain function during task activation. Magn. Reson. Med. 25: 390-397. Baudewig, J., Siebner, H.R., Bestmann, S., Tergau, F., Tings, T., Paulus, W. und Frahm, J. (2001a): Functional MRI of cortical activations induced by transcranial magnetic stimulation (TMS). Neuroreport 12: 3543-3548. Baudewig, J., Bittermann, H.J., Paulus, W. und Frahm, J. (2001b): Simultaneous EEG and functional MRI of epileptic activity: a case report. Clin. Neurophysiol. 112: 1196-1200. Baudewig, J., Dechent, P., Merboldt, K.D. und Frahm, J. (2003): Thresholding in correlation analyses of magnetic resonance functional neuroimaging. Magn. Reson. Imaging 21: 11211130. Bestmann, S., Baudewig, J., Siebner, H.R., Rothwell, J.C. und Frahm, J. (2003): Subthreshold high-frequency TMS of human primary motor cortex modulates interconnected frontal motor areas as detected by interleaved fMRI-TMS. Neuroimage 20: 1685-1696. Blamire, A.M., Ogawa, S., Ugurbil, K., Rothman, D., McCarthy, G., Ellermann, J.M., Hyder, F., Rattner, Z. und Shulman, R.G. (1992): Dynamic mapping of the human visual cortex by high-speed magnetic resonance imaging. Proc. Natl. Acad. Sci. U.S.A. 89: 11069-11073. Dechent, P. und Frahm, J. (2000): Direct mapping of ocular dominance columns in human primary visual cortex. Neuroreport 11: 32473249. Dechent, P. und Frahm, J. (2003): Functional somatotopy of finger representations in human primary motor cortex. Hum. Brain Mapp. 18: 272-283. Frahm, J., Bruhn, H., Merboldt, K.D. und Hänikke, W. (1992): Dynamic MRI of human brain oxygenation during rest and photic stimulation. J. Magn. Reson. Imaging 2: 501-505. Fransson, P., Krüger, G., Merboldt, K.D. und Frahm, J. (1998): Temporal characteristics of oxygenation-sensitive MRI responses to visual activation in humans. Magn. Reson. Med. 39: 912-919. Fransson, P., Krüger, G., Merboldt, K.D. und Frahm, J. (1999): Temporal and spatial MRI responses to subsecond visual activation. Magn. Reson. Imaging 17: 1-7. Kwong, K.K., Belliveau, J.W., Chesler, D.A., Goldberg, I.E., Weisskoff, R.M., Poncelet, B.P., Kennedy, D.N., Hoppel, B.E., Cohen, M.S., Turner, R., Cheng, H.M., Brady, T.J. und Rosen, B.R. (1992): Dynamic magnetic resonance imaging of human brain activity during primary sensory stimulation. Proc. Natl. Acad. Sci. U.S.A. 89: 5675-5679. Logothetis, N.K., Pauls, J., Augath, M., Trinath, T. und Oeltermann, A. (2001): Neurophysiological investigation of the basis of the fMRI signal. Nature 412: 150-157. Mathiesen, C., Caesar, K., Akgoren, N. und Lauritzen, M. (1998): Modification of activity-dependent increases of cerebral blood flow by excitatory synaptic activity and spikes in rat cerebellar cortex. J. Physiol. 512: 555-566. Merboldt, K.D., Fransson, P., Bruhn, H. und Frahm, J. (2001): Functional MRI of the human amygdala? Neuroimage 14: 253-257. Ogawa, S., Lee, T.M., Kay, A.R. und Tank, D.W. (1990): Brain magnetic resonance imaging with contrast depending on blood oxygenation. Proc. Natl. Acad. Sci. U.S.A. 87: 98689872. Kurzbiografien Peter Dechent: Studium der Biologie in Mainz, Manchester, Stockholm und Göttingen. 2001 Promotion am Max-Planck-Institut für biophysikalische Chemie in Göttingen. Danach Postdoc in der Biomedizinischen NMR Forschungs GmbH. Seit 2004 Leiter der Forschergruppe „MR-Forschung in der Neurologie und Psychiatrie“, ein Gemeinschaftsprojekt des Bereichs Humanmedizin der Georg-August-Universität Göttingen und des Max-Planck-Instituts für biophysikalische Chemie in Göttingen, mit Unterstützung des Landes Niedersachsen. Jens Frahm: Studium der Physik in Göttingen. 1977 Promotion am Max-Planck-Institut für biophysikalische Chemie in Göttingen. 1993 Habilitation im Fach Physikalische Chemie und seit 1997 außerplanmäßiger Professor an der Georg-AugustUniversität Göttingen. Seit 1992 wissenschaftlicher Leiter und Geschäftsführer der Biomedizinischen NMR Forschungs GmbH am Max-Planck-Institut für biophysikalische Chemie in Göttingen. Er hat zahlreiche Preise und Auszeichnungen für grundlegende Arbeiten zur Magnetresonanz-Tomografie erhalten und ist z.Z. stellvertretender Sprecher des Zentrums für Neurobiologie des Verhaltens sowie Mitglied des DFG Forschungszentrums Molecular Physiology of the Brain und des Instituts für Multiple Sklerose-Forschung. Korrespondenzadresse Dr. Peter Dechent MR-Forschung in der Neurologie und Psychiatrie Bereich Humanmedizin Georg-August-Universität Göttingen D-37099 Göttingen Tel.: ++ 49 (0) 551 3913 140 Fax: ++ 49 (0) 551 3913 243 e-mail: [email protected] Neuroforum 3/04 TANUJA ROHATGI UND GEORG REISER Thrombin-verwandte Proteasen als Signalmoleküle im Gehirn: Proteaseaktivierte Rezeptoren bei neuronaler Schädigung und bei Neuroprotektion Tanuja Rohatgi und Georg Reiser Zusammenfassung Thrombin, eine Serinprotease mit zentraler Bedeutung für die Blutgerinnung, kann Rezeptoren auf der Plasmamembran von Zellen aktivieren. Diese Protease-aktivierten Rezeptoren (PARs) wurden kürzlich auch im Gehirn gefunden. Dort sind diese Rezeptoren insbesondere bei Neuroprotektion oder Schädigung im Zusammenhang mit pathologischen Zuständen von Bedeutung. Man unterscheidet 4 Subtypen, die als PAR-1 bis PAR-4 bezeichneten Rezeptoren, die alle in Mensch, Ratte und Maus exprimiert sind und große Homologie zwischen diesen Spezies aufweisen. PARs besitzen einen einzigartigen Aktivierungsmechanismus, weil sie durch die Spaltung des Rezeptorproteins irreversibel in den aktiven Zustand versetzt werden. Das durch die Proteolyse neu entstandene extrazelluläre N-terminale Ende des Rezeptor-Polypeptids wird als ein vorher verborgenes N-terminales Ende offen gelegt und wirkt nun als an das Protein gekoppelter, aktivierender Peptidligand. PARs sind in vielen Bereichen im zentralen und im peripheren Nervensystem exprimiert. PARs scheinen bei der Pathologie neurodegenerativer Erkrankungen, wie Alzheimer oder HIV-Encephalitis eine Rolle zu spielen. Thrombin löst im Gehirn über PARs die Proliferation von Astrozyten aus, was für die reaktive Astrogliose von Bedeutung ist. Andererseits wirkt Thrombin janus-ähnlich, da es bei hohen Konzentrationen Nervengewebe schädigt, dagegen in sehr niedrigen Konzentrationen, welche im leicht geschädigten Nervengewebe erreicht werden, eine protektive Rolle hat. Abstract Thrombin-type proteases as signaling molecules in the brain: the role of protease activated receptors in neuronal damage and in neuroprotection Thrombin, mainly known as a serine protease with central significance in blood coagulation, is able to activate receptors on the plasma membrane of cells. These proteaseactivated receptors (PARs) were recently found also in brain. There the receptors are significant for neuronal development, as well as for neuroprotection or damage during pathological situations. Molecular biology research has identified four subtypes of PARs, G protein-coupled receptors. All four subtypes, which are named PAR-1 to PAR-4 were found to be expressed in human, rat and mouse. PARs have a unique activation mechanism, because proteolytic cleavage leads to irreversible conversion of the protein into its active state. In this case, receptor activation after proteolysis results in a new extracellular N-terminal domain. This unmasks the tethered ligand, a sequence of about 6 amino acids, which binds specifically on an extracellular region of the receptor. This feature can be used experimentally for activating the receptor without proteolysis, by utilizing synthetic peptides of those 5-6 amino acid residues. PARs are expressed in many regions of the central and peripheral nervous system. PARs seem to play a role in the pathology of neurodegenerative disorders, like Alzheimer’s disease or HIV-encephalitis. Thrombin induces proliferation of brain astrocytes mediated by PARs, which is of significance for reactive astrogliosis. On the other side thrombin has a janus-like action: at high concentration it causes damage of nervous tissue, at very low concentrations which are reached in slightly damaged tissue it plays a specific protective role. Key words: Protease-activated receptors; thrombin; neurodegeneration; neuroprotection Neuroforum 3/04 Einleitung: Thrombin als eine Protease der Blutgerinnung - und Thrombin im Gehirn? Übermäßiger Blutverlust als Folge von Wunden oder Gewebeverletzungen wird durch den Anstoß einer Blutgerinnungskaskade verhindert, die zur Bildung eines Blutpfropfens am Verletzungsort führt. Unser Blutkreislaufsystem besitzt zahlreiche Proteine, die in dieser Kettenreaktion mitwirken, darunter die Serinprotease Thrombin, deren proteolytische Aktivität Fibrin als eine unauflösliche Verklumpung bildet. Das 39 kDa-Protein Thrombin entsteht aus dem Vorläufer Prothrombin, einem 71.6 kDaProtein. Dies geschieht als Folge einer proteolytischen Spaltaktivität durch den Faktor Xa, ein anderes an der Gerinnungskaskade beteiligtes Protein (Abbildung 1). Bis vor kurzem hatte man angenommen, dass Prothrombin und folglich Thrombin nur im Blut vorkommen und dort spezifisch die Aufgabe der Gerinnung erfüllen. Dann stellte sich heraus, dass Thrombin auch die Blutplättchenaggregation auslöst. Inzwischen ist klar, dass Thrombin verschiedene Zelltypen, wie Makrophagen, Neutrophile, Monozyten und Endothelzellen, aktiviert und darüber hinaus die Proliferation glatter Gefäßmuskelzellen anstoßen kann. Man entdeckte dann, dass Thrombin sogar in primären neuronalen Zellkulturen morphologische Veränderungen herbeiführt. Thrombin bewirkt nämlich ein Einziehen von Neuriten, wohingegen die Zugabe der Thrombinantagonisten ProteaseNexin-1 (PN-1) und Hirudin diesen Thrombin-Effekt hemmte. Bei der Untersuchung menschlicher Fibroblasten wurde der spezifische Thrombinhemmer PN-1 identifiziert. Hirudin ist das Agens, mit dem der Blutegel beim Blutsaugen die Gerinnung hemmt. Ein völlig neues Konzept zur Thrombinwirkung ergab sich aus der Erkenntnis, dass Thrombin, wenn es an Zelloberflächenrezeptoren bindet, mit mehreren Übertragungswegen verknüpft ist. Allerdings wurden bis 1991 weder der Rezeptor noch die thrombinabhängigen Übertragungswege identifiziert. Erst dann wurde der Thrombin-Rezeptor, der auch Protease-aktivierter Rezeptor-1 (PAR-1) genannt wurde, aus humanem Gewebe kloniert. Nachfolgend wurden noch drei weitere Mitglieder der PAR-Familie (PAR-2, PAR-3 und PAR-4) gefunden. Im Zusammenhang mit PAR-2 zeigte sich, dass nicht nur Thrombin, sondern auch andere Serinproteasen, wie Trypsin und Granzym A, PARs aktivieren können. Untersuchun237 DIE ROLLE VON PROTEASE-AKTIVIERTEN REZEPTOREN BEI NEURONALER SCHÄDIGUNG Abb. 1: Erzeugen von Thrombin aus Prothrombin und die direkt durch proteolytische Effekte des Thrombin beeinflussten Wege. Proteolytische Spaltung von Prothrombin durch Faktor Xa erzeugt aktives und funktionelles Thrombin. Thrombin seinerseits wirkt proteolytisch und erleichtert dadurch die Aggregation der Blutplättchen, ermöglicht den endgültigen katalytisch aktivierenden Schritt der Blutgerinnungskaskade. Nach der Bindung von Thrombin an Rezeptoren auf Zelloberflächen aktiviert es diese durch Spaltung. gen in unserem Labor konnten erstmals das Vorhandensein von funktionell aktivem PAR-1 und PAR-2 in primären AstrozytenKulturen aus Rattenhirn nachweisen. Großes Interesse auf diesem Gebiet fand die Entdeckung eines gehirnspezifischen Serinprotease-Hemmers, des Glia-Derived Nexin (GDN), das aus Gliazellen in Kultur gewonnen wurde. Später stellte sich heraus, dass GDN mit PN-1 identisch ist, weshalb man beide nunmehr als PN-1 bezeichnet. Prothrombin und PN-1 sind zugleich im Rattenhirn vorhanden. Darüber hinaus haben Expressionsstudien gezeigt, dass PAR-1 sowohl in embryonalen als auch in postnatalen und adulten Hirnen präsent ist. Wir konnten sogar alle vier PAR-Subtypen im adulten Rattenhirn nachweisen. Bei traumatischen Hirnverletzungen oder Schlaganfall kann ein Durchbrechen der Blut-HirnSchranke zusätzlich den exogenen Serinproteasen des Serums den Zugang vom Blut zum Hirn öffnen und damit Hirnschädigungen in Form von Vernarbungen und Ödemen verstärken. Diese Schädigungen können Gehirnkrämpfe oder Zelltod verursachen. Im Folgenden werden nach einer Einführung in das Gebiet der Proteasen zuerst die Beson238 derheiten der PARs und daraufhin die Bedeutung von PARs im Gehirn dargestellt. Proteasen als Enzyme und als Liganden der Protease-Aktivierten Rezeptoren (PARs) Proteasen stellen die größte Enzymgruppe dar. Sie wirken hochspezifisch, indem sie Peptidbindungen durch Hydrolyse irreversibel spalten. Unsere Sicht auf Proteasen wurde durch die Forschung der vergangenen Jahre dramatisch verändert. So spricht man von ihnen inzwischen auch als „Signalscheren“. Die katalytische Aktivität der Proteasen kann eine Vielzahl verschiedener Zellfunktionen einleiten und regeln. Dazu gehören Proliferation, Differenzierung, Migration, Apoptose, Wundheilung und Angiogenese (Gefäßbildung). Ihrem katalytischen Wirkungsmechanismus folgend werden die Proteasen in Gruppen eingeteilt, die entsprechend jener Aminosäure benannt werden, die bei der katalytischen Wirkung wesentlich ist: Das sind als eine Gruppe die Serinproteasen, wie Thrombin, Trypsin und Chymotrypsin, welche im Brennpunkt dieses Übersichtsartikels stehen. Die Proteasen werden meistens zunächst als harmlose inaktive Proenzyme gebildet und verharren so gewissermaßen in einem Standby-Modus, bis ihre Aktivität erforderlich ist. Proteasen spalten sehr spezifisch und begrenzt verschiedene Substratproteine. Es ist äußerst wichtig, die jeweiligen Substrate zu identifizieren, weil diese schließlich die potenziellen Angriffspunkte für eine therapeutische Behandlung pathologischer Zustände sind. Der vorliegende Artikel wird sich auf ein bestimmtes Substrat für die Serinproteasen, die Protease-aktivierten Rezeptoren (PARs), konzentrieren. Für die Existenz, Versorgung und Aufrechterhaltung einer Homöostase insbesondere im Gehirn ist die Kommunikation zwischen den Zellen durch ein Netzwerk von Signalen unabdingbar. Auch die Serinproteasen Thrombin und Trypsin wirken als Signal gebende Moleküle, indem sie an Rezeptoren an der Zelloberfläche, nämlich die PARs, binden, diese aktivieren und so diverse Zellfunktionen regulieren. Abbildung 2 zeigt Struktur und Aktivierungsmechanismus der PARs. PARs sind an G-Proteine gekoppelte Rezeptoren und gehören zu einer großen Familie von Transmembranrezeptoren. PARs zeigen die dafür typischen sieben transmembranen Helices (TMD1 bis TMD7 in Abbildung 2): Bei diesen besteht eine Verbindung von drei extrazellulären und drei intrazellulären Schleifen mit einem extrazellulären Nterminalen und einem intrazellulären C-terminalen Bereich. PARs bedienen sich eines einzigartigen Aktivierungsmechanismus, indem sie den extrazellulären proteolytischen Spaltungsvorgang in ein transmembranäres Signal umwandeln (Coughlin 2000). Um den Rezeptor zu aktivieren, bedarf es dieser Spaltung des Rezeptors an einer spezifischen Stelle im extrazellulären N-terminalen Bereich durch die wirksame Liganden-Protease. Die Spaltung lässt ein neues N-terminales Ende als Liganden aufscheinen, der jedoch an die Polypeptidkette des Rezeptors gekoppelt bleibt. Lediglich etwa sechs Aminosäure-Reste innerhalb dieses Bereichs interagieren als neu hervorgetretener, fest verkoppelter Ligand mit der extrazellulären Schleife 2 und den Transmembranbereichen des gespaltenen Rezeptors und lösen folglich transmembranäre Signale aus. So tragen die PARs ihren eigenen selbstaktivierenden Liganden, der jedoch den Rezeptor nur dann aktiviert und damit ein innerzelluläres Signal initiieren kann, wenn er zuvor der Proteaseaktivität ausgesetzt worden ist. Allerdings kann bei Abwesenheit der aktiven Protease der intakte Rezeptor auch durch die Neuroforum 3/04 TANUJA ROHATGI UND GEORG REISER Bindung des „gekoppelten Liganden“ als synthetisch hergestelltes Peptid aktiviert werden. Das Peptid kann so den Rezeptor ohne die Notwendigkeit einer proteolytischen Spaltung aktivieren. Es wurde gefunden, dass die für PAR-1 und -2 bekannten aktivierenden Peptide ähnliche physiologische Rezeptor-Reaktionen hervorrufen wie die Agonisten-Proteasen. Letztlich interagiert der aktivierte Rezeptor über die intrazellulären Schleifen 2 und 3 mit verschiedenen G-Proteintypen und übermittelt derart das Signal (Coughlin 2000). Die verschiedenen PAR-Gene und die pharmakologische Klassifizierung der PARs Für die vier Subtypen der PAR, PAR-1 bis PAR-4, zeigt Tabelle 1 die Größe der Exons und Introns im Genom, die Aktivierungsund Inaktivierungsproteasen, die bei Aktivierung entstandenen Spaltstellen des Proteins beim Menschen und die Peptidsequenzen der gekoppelten Liganden der einzelnen PARs bei Mensch, Maus und Ratte. Die vier Subtypen zeigen eine identische Genstruktur: zwei Exons (kodierende Region), die im Falle von PAR-1 bis PAR-3 durch ein einziges, sehr langes Intron (nicht-kodierende, unterbrechende Region) von 4000 bis 22000 Basen getrennt sind. PAR-4 unterscheidet sich insofern, als hier das Intron sehr kurz ist, nämlich nur 274 Basen. In allen vier Fällen enthält das erste kurze Exon die 5’-nichttranslatierte Sequenz, das Start-Codon und ein Signal-Peptid, wohingegen das zweite, lange Exon für das Rezeptorprotein und die 3’-nichttranslatierte Sequenz kodiert. Die Gene von PAR-1, PAR-2 und PAR-3 befinden sich auf demselben menschlichen Chromosom 5, nämlich auf der Bande Nr. 13 des langen Arms (q) (die Bezeichnung des Genortes ist also 5q13); dagegen ist PAR-4 auf der Bande 12 des kurzen Arms (p) von Chromosom 19 (19p12) lokalisiert. Alle vier Subtypen haben große Ähnlichkeit untereinander und sind quer durch die verschiedenen Spezies Mensch, Maus und Ratte in hohem Maße konserviert. Die pharmakologische Klassifizierung der PARs erfolgt durch Zuordnung der aktivierenden Liganden-Proteasen. Der hauptsächliche Agonist von PAR-1, PAR-3 und PAR4 ist die Serinprotease Thrombin, die sowohl im Blutplasma als auch im zentralen Nervensystem (ZNS) vorkommt. Eine weitere Protease, die als PAR-Agonist vor allem auf PAR-2, aber auch auf PAR-1 wirkt, ist Trypsin, ein Verdauungsenzym, das im gastrointestinalen Trakt als inaktives Zymogen geNeuroforum 3/04 Abb. 2: Aktivierungsmechanismus der PARs. Proteolytische Aktivierung durch Serinproteasen am N-Terminus ergibt einen „gekoppelten“ Liganden, dessen Sequenz durch das Trapez gezeigt ist, welches dann an die Stelle des blauen Trapezes, auf der extrazellulären Schleife 2 des Rezeptors bindet und ihn somit aktiviert. Synthetische aktivierende Peptide, welche der Sequenz des gekoppelten Liganden des Rezeptors entsprechen, können an dieselbe Aktivierungsstelle binden und so den Rezeptor ohne proteolytische Spaltung aktivieren. Die intrazellulären Signale werden sowohl C-terminal als auch durch die zweite intrazelluläre Schleife über das G-Protein weitergegeben. ECL extrazelluläre Schleife, nummeriert 1 bis 3; ICL intrazelluläre Schleife, nummeriert 1 bis 3; TMD Transmembrandomäne, nummeriert 1 bis 7. bildet wird. Da Trypsin nicht an Expressionsorten von PAR-2 wie im Hirn vorhanden ist, suchte man nach alternativen Agonisten für PAR-2 und fand dabei die Tryptase, eine tetramere Serinprotease, die nach einer allergischen Reaktion von Mastzellen freigesetzt wird und ihrerseits PAR-2 aktiviert. Interessanterweise setzen Rattenhirnschnitte eine trypsinartige Serinprotease (P22) frei, die in der Lage ist, extrazelluläre Matrix zu verdauen und PAR-2 zu stimulieren. Neben Thrombin und Trypsin gibt es noch weitere Serinproteasen, die ebenfalls PARs aktivieren können. Granzym A gehört zu diesen. Es wird in zytotoxischen T-Lymphozyten gebildet und kann PAR-1 in Neuronen und Astrozyten aktivieren. Die proteolytische Wirkung von Granzym A hat sowohl die Retraktion von Neuriten als auch die Umkehrung der Astrozyten-Fortsatzbildung zur Folge. Allerdings können andere Serinproteasen, die in Tabelle 1 bei den jeweiligen PARs angegeben sind, PARs inaktivieren. Dazu gehören Cathepsin G, Elastase, Proteinase 3 und Plasmin. Der Inaktivierungsmechanismus beruht in allen Fällen auf einer proteolytischen Spaltung mitten im gekoppelten Liganden, wodurch der Rezeptor unempfänglich für eine Aktivierungsprotease, wie Thrombin, wird. Verteilung der PARs im Gehirn Studien in unserem Labor konnten das Vorhandensein aller vier PAR-Subtypen im Gehirn nachweisen. Abbildung 3A-D zeigt jeweils Beispiele für deren Verteilung in verschiedenen Arealen des Rattenhirns (Striggow et al. 2001). Eine detaillierte immunhistochemische Analyse ergab, dass das PAR1-Protein im Hippocampus reichlich in der Pyramidenzellschicht vorhanden ist, verglichen mit einem eher niedrigen Expressionsniveau in den Neuronen im Kortex, Thalamus, Hypothalamus, Striatum und Amygdala. PAR-1-Expression, sowohl als mRNA als auch als Protein, wurde in embryonalem und postnatalem Hirngewebe festgestellt. PAR-2 und PAR-3-Proteine finden sich in hohem Maße in Hippocampus, Kortex, Amygdala, Thalamus, Hypothalamus und Striatum. PAR-2-Protein wird ferner wäh239 DIE ROLLE VON PROTEASE-AKTIVIERTEN REZEPTOREN BEI NEURONALER SCHÄDIGUNG Abb. 3: Immunzytochemischer Nachweis der PARs im Gehirn. Koronarschnitte vom adulten Rattenhirn mit verschiedenen Arealen nach Anfärbung mit spezifischen Antikörpern für PAR-1 bis -4. (A) PAR-1 Immunreaktivität in der rechten Hemisphäre des Gehirns; (B) PAR2 Immunreaktivität in der Hippocampusformation. PAR-2 positive Zellen in der Pyramidalschicht, Körnerzellen des Gyrus dentatus und Moosfasern. (C) PAR-3 Anfärbung im Hippocampus. PAR-3 Immunreaktivität in Pyramidenzellen, Körnerzellen des Gyrus dentatus and des Subiculum. (D) PAR-4 Immunreaktivität in der Moosfaserschicht der Hippocampusformation. Maßstab jeweils, 2 mm (A), 500 µm (B und C) und 100 µm (D). Abkürzungen: AN, amygdaloid nucleus; CO, cortex; CPu, caudate putamen; GrDG, granular cell layer of dentate gyrus; HF, hippocampal formation; HTH, hypothalamus; LMol, stratum lacunosum moleculare; mf, mossy fibres; Mol, stratum moleculare of the dentate gyrus; Py, pyramidal cell layer. (Ausschnitte übernommen von Striggow et al. 2001) rend der Embryogenese im ZNS exprimiert. Dagegen zeigt sich das PAR-4-Protein in neuronalen Somata, in den Axonen und Dendriten des Hippocampus sowie allen Schichten des Kortex, von Thalamus, Hypothalamus und Amygdala. Aus unserem Labor wurden des Weiteren auf mRNA- wie auf Protein-Ebene funktionelle Koexpressionen von PAR-1 bis -4 in Ratten-Astrozyten in Kultur berichtet (Wang et al. 2002a). Unter Einsatz von RT-PCR und Immunomarkierung konnten wir kürzlich die Expression von PAR-1 und PAR-3 in Ratten-Oligodendrozyten in Kultur sowie auch in einer Oligodendroglia-Zelllinie belegen. PAR-3 zeigt dort jedoch keine offenkundige funktionelle Aktivität. Eine weitere Beobachtung weckt sicherlich unser Interesse. PARs sind auch in vielen anderen Regionen des Nervensystems in 240 hohem Maße exprimiert. Im Rückenmark zum Beispiel ist PAR-1 mRNA reichlich in Motoneuronen und in den Hinterwurzelganglien zu finden, ferner in den präganglionären Neuronen des autonomen Nervensystems. PAR-2-Expression auf Proteinebene lässt sich während der Embryogenese im peripheren Nervensystem (PNS) entdecken. PAR-2 wird ferner von primären Rückenmarksafferenzneuronen exprimiert. Vor kurzem wurde mit Hilfe der Immunocytochemie die Expression von PAR-4 in peripheren Nerven gefunden. Mechanismen der Signalübertragung durch PARs Die Spaltung des N-terminalen extrazellulären PAR-Abschnitts durch die AgonistProtease ist von der Bindung des aufgedeck- ten gekoppelten Liganden mit Aktivierung des Rezeptors gefolgt (Abbildung 2). Damit wird eine Veränderung der Rezeptorkonformation bewirkt, was in der Folge den Rezeptor mit heterotrimeren G-Proteinen interagieren lässt. Diese Proteine binden die Guaninnukleotide GDP und GTP und werden deshalb so genannt. Die PAR-Rezeptor-GProtein-Interaktion stößt nun folgenden Prozess an: GDP wird auf der großen Untereinheit des G-Proteins durch GTP ersetzt und ergibt den dadurch aktivierten G-ProteinGTP-Komplex. Dieser kann verschiedene Enzyme oder Ionenkanäle als Effektoren aktivieren. Die Signalübertragung durch GProteine endet, wenn GTP zu GDP hydrolysiert ist und nachfolgend die G-Proteine dadurch wieder in ihren inaktiven Zustand zurückgeführt sind. PAR-1 ist innerhalb der PAR-Familie hinsichtlich seines Signal-Mechanismus bisher am besten verstanden. Eine Aktivierung von PAR-1 durch einen Agonisten resultiert meistens in einer Stimulierung der Phospholipase C (PLC) über das Gq-Protein. Aktivierte PLC hydrolysiert in der Plasmamembran ein Phospholipid zu Inositol(1,4,5)-trisphosphat (IP3) und Diacylglycerol (DAG). Die Bildung des IP3 führt zur Mobilisierung von intrazellulärem Ca2+, DAG seinerseits hat an der Aktivierung von Proteinkinase C (PKC) Anteil. Eine weitere Enzymfamilie, die in der PAR-Signalübertragung involviert ist, sind die mitogen-aktivierten Proteinkinasen (MAPK). Sie spielen eine zentrale Rolle bei Wachstum, Proliferation, Entwicklung und Überleben aller eukaryotischen Organismen. Die Signalübertragung von PARs in Astrozyten des Gehirns wurde intensiv untersucht. Astrozyten bilden eine Hauptgruppe der Gliazellen und sind äußerst wichtig im Gehirn, da sie auch die Neuronenaktivität beeinflussen, vielleicht sogar steuern. Nach Verletzungen proliferieren Astrozyten und können in der Folge zu Synapsen wandern, die durch die Verletzung nicht geschädigt wurden. Die Unterstützung funktioneller Synapsen und die Entstehung von Progenitor-Stammzellen für die Schaffung aller Typen reifer Hirnzellen, einschließlich Neuronen im adulten Hirn, gehören zu den Charakteristika, die im Zusammenhang mit Astrozyten diskutiert werden. Es ist deshalb entscheidend, den Mechanismus, welcher der Proliferation der Astrozyten im ZNS zugrunde liegt, zu verstehen. Studien in unserem Labor konnten nachweisen, dass Thrombin eine erhebliche Stimulation der Proliferation von Rattenastrozyten in Kultur auslöst (Wang et al. 2002a). Dieser Prozess wird auf dem MAPK-Signalweg ermögNeuroforum 3/04 TANUJA ROHATGI UND GEORG REISER licht (Wang et al. 2002b). Der stimulierende Effekt von Thrombin oder PAR-1 aktivierendem Peptid wird durch die Aktivierung von PAR-1 angestoßen und läuft über zwei parallele intrazelluläre Schienen. Eine dieser beiden Reaktionen involviert die Aktivierung des Pertussis-Toxin-sensitiven GiProteins. Pertussis-Toxin, welches für die Entstehung der Keuchhustenerkrankung verantwortlich ist, bewirkt auf molekularer Ebene eine Hemmung des Gi-Proteins. Vom GiProtein wandert das Signal zur Phosphatidylinositol-3-Kinase und bewirkt schließlich eine Phosphorylierung der die Genexpression steuernden Proteine ERK 1/2. Der zweite Transduktionsweg verbindet das Gq-Protein mit der PLC, deren Aktivierung zur Entstehung von IP3 und DAG führt und in der Konsequenz die Freisetzung von intrazellulärem Kalzium und die Aktivierung der PKC zur Folge hat. Die beiden Signalübertragungswege stehen unter einander in Verbindung (Wang et al. 2002b; Wang und Reiser 2003). Kürzlich konnten wir an einer Oligodendrozyten-Zelllinie darlegen, dass auch in diesen Zellen der durch PAR-1-Aktivierung induzierte intrazelluläre Kalziumanstieg vor allem das Ergebnis der Ca2+-Freisetzung aus intrazellulären Speichern ist. Hinsichtlich der intrazellulären Signalmechanismen des PAR-2 ist bisher deutlich weniger bekannt. Physiologische und pathophysiologische Funktionen von Thrombin und PARs im Gehirn Die Vielzahl der in den letzten Jahren erschienenen Publikationen zum Thema PARs beleuchtet das wachsende Interesse an deren Bedeutung vor allem auch für zerebrale Vorgänge. Dennoch stehen wir trotz rascher Fortschritte immer noch am Anfang des Verständnisses der von ihnen kontrollierten zellulären Funktionen. Die jüngsten Studien haben gezeigt, dass diese Rezeptorfamilie an einer erheblichen Anzahl neurologischer Prozesse beteiligt ist, indem sie eine kritische Rolle in der Aufrechterhaltung der Balance zwischen Neuroprotektion und Neurodegeneration, bei Entzündung, Verletzung und verschiedenen Krankheitszuständen spielt. Abbildung 4 stellt einige der wesentlichen funktionellen Konsequenzen einer PAR-Aktivierung im Gehirn an den Zelltypen Neuronen, Astrozyten und Mikroglia und bezogen auf neurodegenerative Erkrankungen dar. Thrombin kann im Nervensystem Wachstum, Aufrechterhalten der neuronalen Funktion und morphologische Veränderungen inNeuroforum 3/04 Abb. 4: Funktionelle Bedeutung der PAR Aktivierung im Gehirn. PAR Aktivierung durch Thrombin bewirkt diverse zelluläre und morphologische Veränderungen in den Hauptzelltypen des Gehirns, Neuronen, Astrozyten and Mikroglia. Deren Wechselwirkungen sind wesentlich für Entwicklung und Folgen bei einigen neurodegenerativen Erkrankungen. duzieren. Im Gehirn beeinflusst es alle drei Hauptzelltypen. Thrombin löst in Neuronen eine Veränderung der Zellmorphologie aus. Die proteolytische Aktivität von Thrombin resultiert über PAR-1 in einem Einziehen neuritischer Fortsätze, ein nach Entzug von Thrombin aus dem umgebenden Medium reversibler Vorgang. Auch auf Astrozyten wurde die Wirkung von Thrombin detailliert untersucht. In Kultur verursacht Thrombin eine Umkehr der sternförmigen Morphologie der Astrozyten und führt sie in eine flache epitheliale Form über. Der oben besprochene mitogene Effekt, welcher in einer Proliferation der Astrozyten resultiert, wird ausgelöst. Allerdings schließen sich die beiden Thrombin-Effekte an Astrozyten bezüglich der erforderlichen Konzentration von Thrombin gegenseitig aus. Bei der niedrigsten Konzentration von Thrombin wurde kein mitogener Effekt beobachtet, wohingegen die Umkehr der sternförmigen Morphologie ausgelöst wurde. Unsere Untersuchungen haben ergeben, dass die Zugabe von Thrombin und das Aktivieren von PAR-1 bis PAR-3 durch die entsprechenden Peptidliganden zur Astrozytenproliferation führt (Wang et al. 2002a). Weiterhin stimuliert Thrombin die Freisetzung des potenten vasokonstriktorischen Peptids Endothelin-1 aus Astrozyten. Thrombin induziert auch die Arachidonsäurebildung in Astrozyten, wobei die Arachidonsäure ih- rerseits die Thrombin-induzierten Kalziumantworten in den Astrozyten unterdrückt. Nicht nur bei Astrozyten sondern auch bei der Mikroglia löst Thrombin Proliferation über Aktivierung des PAR-1 aus. Mikroglia sind residente Makrophagen im ZNS und spielen bei verschiedenen ZNS-Erkrankungen eine wesentliche Rolle, indem sie Entzündung und neuronalen Zelltod kontrollieren. PAR-1-Aktivierung reguliert CD40 hoch, welches ein transmembranes Glykoprotein ist, das in Immunzellen, wie B-Lymphozyten, aktivierten T-Zellen und Monozyten, exprimiert wird. Aktivieren der PARs über Thrombin hat indirekt auch einen Effekt auf die Expression eines anderen Typs der G-Protein-gekoppelten Rezeptoren, nämlich des metabotropen Glutamatrezeptors, welcher bei synaptischer Plastizität mitwirkt. Nach PAR-1-Aktivierung durch Thrombin ist die Expression eines Typs des metabotropen Glutamatrezeptors, mGluR5, auf Astrozyten reduziert. Astroglia haben eine bedeutende Aufgabe bei der Aufrechterhaltung der Glutamattransmission. Kenntnis der Regulation der astrozytären Funktionen während pathologischer Zustände, welche durch erhöhte Thrombinspiegel charakterisiert sind, kann von großer Bedeutung für das Verständnis zellulärer Wechselwirkungen bei Krankheitszuständen im ZNS sein. Die Aktivierung von PAR-1 über Throm241 DIE ROLLE VON PROTEASE-AKTIVIERTEN REZEPTOREN BEI NEURONALER SCHÄDIGUNG Tab. 1: Charakteristika der PAR-Gene und der proteinspaltenden Aktivierungen der vier PAR-Subtypen, PAR-1 bis -4. Exons (Anzahl der kodierten Aminosäuren (AS)) und Introns (Anzahl Basen) der humanen PAR-Gene. PARs können meist durch mehrere Proteasen aktiviert bzw. inaktiviert werden, wie hier zusammengefasst. Die durch Agonist entstehende Spaltstelle am humanen PAR und die Sequenz des aktivierenden Peptids (gekoppelter Ligand) der PARs bei Mensch (h), Maus (m) und Ratte (r). TF, tissue factor; Faktoren VIIa und Xa, Faktoren der Gerinnungskaskade. PAR-1 PAR-2 PAR-3 PAR-4 Exon 1 (human) AS 1-29 AS 1-27 AS 1-22 AS 1-37 Exon 2 (human) AS 30-425 AS 28-397 AS 23-374 AS 38-385 Intron (human) 22,000 Basen 14,000 Basen 4,000 Basen 274 Basen Thrombin Trypsin Factor Xa Granzym A Trypsin Tryptase Factor Xa TF/ Factor VIIa Thrombin Thrombin Trypsin Cathepsin G Arg 41-Ser 42 Arg 34-Ser 35 Lys 38-Thr 39 Arg 47-Gly 48 Gekoppelter Ligand SFLLRN (h) SFFLRN (m, r) (Peptidsequenz) SLIGKV (h) SLIGRL (m, r) TFRGAP (h) SFNGCP (m) GYPGQV (h) GYPGKF(m) Aktivierende Proteasen Spaltstelle (human) Inaktivierende Proteasen Plasmin Cathepsin G Elastase Proteinase 3 Chymase Chymase Elastase bin in hippocampalen Neuronen hemmt die durch NMDA-Rezeptoren vermittelte Schmerzübertragung auf einem Weg, welcher den Endothelin-A-Rezeptor betrifft. Letzterer ist ein weiterer G-Protein-gekoppelter Rezeptor. Endothelin-Rezeptoren, die wichtig bei der Regulation des kardiovaskulären Systems sind, wurden auch auf den Gliazellen im ZNS gefunden. Ein Gebiet, auf dem die Erforschung der PARs besondere Aufmerksamkeit erregt hat, ist deren mögliche Mitwirkung bei der Kontrolle von Entzündung und bei Verletzungen. PAR-1 zum Beispiel wurde nach einer Gesichtsnervverletzung herunterreguliert. Eine jüngst durchgeführte Studie ergab, dass eine milde Quetschung des Sehnervs in der Ratte, welche ein Modell für ZNSTrauma darstellt, zu einer vorübergehenden Hochregulation aller PARs führt (Rohatgi et al. 2003). Bei der Nervquetschung entsteht aus Prothrombin an der Stelle der Gewebeläsion durch eine verletzungsbedingte Aktivierung Thrombin, welches dann einen Sekundärschaden verursacht. Thrombin kann in neuralen Zellen über die Rezeptor-vermittelte proteolytische Aktivierung überraschenderweise gegensätzliche Wirkungen auslösen, entweder apoptotischen Zelltod oder aber Neuroprotektion. Wie ist das zu verstehen? Beides wurde in Kultur sowohl bei Astrozyten als auch bei Neuronen beobachtet. Diese an isolierten Zellen gewonnene Einsicht löste diverse Studien aus, die nach einer eventuell kon242 Elastase zentrationsabhängigen Wirkung des Thrombin in vivo forschten. Dies wurde insbesondere bei Gewebeverletzung und Wundheilung untersucht, wie im Fall verschiedener Gehirnverletzungsmodelle. Diese Modelle beinhalten Sauerstoff-Glukose-Entzug an hippocampalen Schnittkulturen (Striggow et al. 2001), Quetschung des Nervus opticus sowie hypoglykämische und oxidative Stressbedingungen. Eine Studie unseres Labors stellte fest, dass Thrombin bei einer niedrigen Konzentration (50 nM oder weniger) neuroprotektiv wirken kann, wohingegen bei höherer Konzentration ein neurodegenerativer Effekt ausgelöst wird (Striggow et al. 2000). Beides wurde an dem Modell der organotypischen hippocampalen Schnittkulturen in vitro untersucht. Die parallel durchgeführte in vivo Studie ergab, dass nach einer Hirnischämie der Ratte Thrombin eine konzentrationsabhängige Wirkung bezüglich Absterben oder Überleben der Neurone hat (Striggow et al. 2000). Eine andere Untersuchung zeigte, dass die Thrombin-vermittelte PAR-1-Aktivierung Neuronen und Astrozyten gegen Belastungen schützt, zu denen oxidativer Stress oder Hypoglykämie gehören, wohingegen hohe Konzentrationen von Thrombin sowohl für Neuronen als auch Astrozyten, die unter Normalbedingungen kultiviert wurden, toxisch waren. Dabei offenbarte sich ferner, dass all diese Effekte durch den endogenen Serinprotease-Inhibitor des Gehirns, PN-1, gehemmt werden. Dies belegt die Notwendigkeit der proteolytischen Aktivität von Thrombin über PARs. Die meisten Wirkungen von Thrombin sind durch den Rezeptor PAR-1 vermittelt, wobei jüngst auch die Rolle des PAR-4 bei proinflammatorischen Prozessen herausgestrichen wurde. An Hirnmikroglia sind proinflammatorische Effekte des Thrombin nicht nur durch PAR-1, sondern auch durch PAR4 vermittelt. PAR-2 Agonisten sind in neurogener Inflammation beteiligt und sind toxisch für hippocampale Neurone. Daraus folgt, dass PAR-2-Aktivierung zur Neurodegeneration beitragen kann. Schlaganfall, nämlich Zerebralinfarkt, bewirkt ein plötzliches neurologisches Versagen, welches zum einen durch Verschluss eines zerebralen Blutgefäßes verursacht ist, wodurch ischämische Nekrose des Gehirns ausgelöst wird. Andererseits kann auch der Riss eines Blutgefäßes verantwortlich sein, wodurch im Gehirn oder im Subarachnoidalraum eine intracraniale Blutung entsteht. Unsere Studien zur PAR-Expression in verschiedenen Tiermodellen der zerebralen Ischämie haben gezeigt, dass PARs bei der Pathophysiologie der zerebralen Ischämie beteiligt sind: die mRNA-Expression der PARs wird nach Ischämie transient verändert. Hingegen offenbarte sich in Knockout-Mäusen, welche den PAR-1 nicht mehr hatten, eine deutliche Reduktion des Infarktvolumens nach einer transienten zerebralen Ischämie (Junge et al. 2003). Dies weist auf eine neurodegenerative Rolle des PAR-1 während des Schlaganfalls und der Ischämie hin, sowie auf das Auslösen von Neuroprotektion in Folge eines Mangels an PAR-1. Das Fehlen auffälliger Unterschiede zwischen Wildtyp und PAR-1 Knock-out-Mäusen nach Durchlässigmachen der Bluthirnschranke bei Ischämie beweist, dass PAR1-Signale mit neuronalem oder glialem Zellüberleben zusammenhängen. Ein weiterer Aspekt, welcher die Wirkung von Thrombin als einer möglichen neuroprotektiven Substanz unterstützt, ist seine Beteiligung beim ischämischen Präkonditionieren (Striggow et al. 2001; Xi et al. 2003). Beim Präkonditionieren entwickelt das Gehirn eine Toleranz gegenüber einem den Zelltod auslösenden Insult, wenn das Gehirn mit einem physiologisch stresshaften, jedoch weitaus weniger gravierenden Insult vorbehandelt wird. So bietet Präkonditionieren eine mögliche Therapie insbesondere für Schlaganfallgefährdung. Wir haben gezeigt, dass endogenes Thrombin beim ischämischen Präkonditionieren beteiligt ist, womit der Schaden nach einer folgenden schweren Ischämie vermindert wird und sich eine Neuroforum 3/04 TANUJA ROHATGI UND GEORG REISER Neuroprotektion einstellt (Striggow et al. 2000). Präkonditionieren durch Anwenden von Thrombin verringert auch die durch zerebrale Blutung ausgelöste Gehirnödembildung. PARs werden ferner mit verschiedenen neurodegenerativen Erkrankungen in Zusammenhang gebracht. Auf dem Gebiet der Alzheimerschen Erkrankung ist es essenziell, die biologischen Wirkungen des β-Amyloid-Proteins, ein ursächliches Agens der Alzheimer-Erkrankung, zu erforschen. Thrombin schwächt die durch β-AmyloidProtein ausgelöste neuronale Degeneration ab. Thrombin-induzierte Effekte können dabei durch das PAR-1 aktivierende Peptid vermittelt werden, was darauf hinweist, dass PAR-1 bei Alzheimer-Erkrankungen beteiligt ist. Eine Studie aus jüngster Zeit hat ergeben, dass bereits nanomolare Konzentrationen von Thrombin in der Lage sind, eine schnelle Hyperphosphorylierung des TauProteins in hippocampalen Neuronen der Maus durch PAR herbeizuführen, wodurch apoptotischer neuronaler Zelltod ausgelöst wird. Das Tau-Protein, welches eines der Mikrotubuli-assoziierten Proteine ist, kommt als axonales Phosphoprotein auch im normalen Gehirn vor; bei Alzheimer-Erkrankung ist Tau jedoch hyperphosphoryliert und ein wesentlicher Bestandteil der neurofibrillären Geflechte. Deren Anzahl scheint proportional zum Grad der Demenz bei Alzheimer-Patienten zu sein. Daher unterstreicht die Aktivität des Thrombin über PAR-1 und PAR-4 bei der Auslösung der Tau-Hyperphosphorylierung und nachfolgender Bildung der Geflechte die besondere Bedeutung von Thrombin und PARs bei der Pathogenese der Alzheimer-Erkrankung. Eine weitere neurodegenerative Erkrankung, bei der Thrombin und PARs eine Rolle zu spielen scheinen, ist die Parkinsonkrankheit. Bei der Parkinsonkrankheit findet sich eine Abnahme der Anzahl der Neuronen, welche den Neurotransmitter Dopamin freisetzen, wodurch sich eine verminderte Koordination der Bewegung sowie eine Steifheit der Extremitäten ergibt. Mehrere Berichte haben gezeigt, dass Thrombin-vermittelte Aktivierung der Mikroglia bei neuropathologischen Prozessen des dopaminergen Zelltods in der substantia nigra der Ratte beteiligt ist. Auch bei Humanimmundefizienz (HIV)-bedingter Neurodegeneration wurde ein Bezug zu Thrombin und PAR-1 gefunden. Die HIV-Encephalitis ist ein Hauptgrund der Infektion im Gehirn bei HIV-Patienten. Encephalitis ist eine Entzündung viralen oder mikrobiellen Ursprungs. Neuroforum 3/04 Aktivieren und Hochregulieren von PAR-1 scheinen bei Gehirnentzündung zum neuronalen Schaden der HIV-Infektion beizutragen. Abschließende Bemerkungen Die Funktionen der PARs als Rezeptor-Familie und die des Thrombin und anderer proteolytischer Rezeptoraktivatoren sind wesentlich im normalen wie im kranken Zustand des Nervengewebes wirksam. Welche Chancen können sich dadurch eröffnen? Strategien, um die PAR-Aktivierung durch PAR-Agonisten oder PAR-Antagonisten zu beeinflussen, stellen ein neuartiges therapeutisches Potenzial dar. Dies kann helfen, Prozesse in Gang zu setzen, um Langzeitschäden bei neurodegenerativen Erkrankungen zu reduzieren (Ossovskaya und Bunnett 2004). Eine wichtige und offensichtliche Frage ist jedoch bis jetzt trotz intensiver Forschung auf diesem Gebiet noch nicht beantwortet, nämlich, welche Typen von Proteasen während normaler oder pathologischer Bedingungen im Gehirn die PAR-Aktivierung auslösen und ob auch Peptide als aktivierende Agenzien dies übernehmen können. Literatur Coughlin, S.R. (2000): Thrombin signalling and protease-activated receptors. Nature 407: 258264. Junge, C.E., Sugawara, T., Mannaioni, G., Alagarsamy, S., Conn, P.J., Brat, D.J., Chan, P.H. und Traynelis, S.F. (2003): The contribution of protease-activated receptor 1 to neuronal damage caused by transient focal cerebral ischemia. Proc Natl Acad Sci USA 100: 1301913024. Ossovskaya, V.S. und Bunnett, N.W. (2004): Protease-activated receptors: contribution to physiology and disease. Physiol Rev 84: 579-621. Rohatgi, T., Sedehizade, F., Sabel, B.A. und Reiser, G. (2003): Protease-activated receptor subtype expression in developing eye and adult retina of the rat after optic nerve crush. J Neurosci Res 73(2): 246-254. Striggow, F., Riek, M., Breder, J., Henrich-Noack, P., Reymann, K.G. und Reiser, G. (2000): The protease thrombin is an endogenous mediator of hippocampal neuroprotection against ischemia at low concentrations but causes degeneration at high concentrations. Proc Natl Acad Sci USA 97: 2264-2269. Striggow, F., Riek-Burchardt, M., Kiesel, A., Schmidt, W., Henrich-Noack, P., Breder, J., Krug, M., Reymann, K.G. und Reiser, G. (2001): Four different types of protease-activated receptors are widely expressed in the brain and are up-regulated in hippocampus by severe ischemia. Eur J Neurosci 14: 595-608. Wang, H. und Reiser, G. (2003): The role of the Ca2+-sensitive tyrosine kinase Pyk2 and Src in thrombin signalling in rat astrocytes. J Neurochem 84: 1349-1357. Wang, H., Ubl, J.J. und Reiser, G. (2002a): Four subtypes of protease-activated receptors, co-expressed in rat astrocytes, evoke different physiological signaling. Glia 37: 53-63. Wang, H., Ubl, J.J., Stricker, R. und Reiser, G. (2002b): Thrombin (PAR-1)-induced proliferation in astrocytes via MAPK involves multiple signaling pathways. Am J Physiol Cell Physiol 283: C1351-1364. Xi, G., Reiser, G. und Keep, R.F. (2003): The role of thrombin and thrombin receptors in ischemic, hemorrhagic and traumatic brain injury: deleterious or protective? J Neurochem 84: 3-9. Eine ausführliche Literaturliste kann von den Autoren angefordert werden. Danksagung Die Untersuchungen wurden von der DFG und dem BMBF unterstützt. Wir danken Gisela Reiser, M.A. für die gründliche Durchsicht des Textes. Kurzbiographien Prof. Dr. Georg Reiser: 1967-1973 Studium Physik und Chemie, Ludwig-Maximilians-Universität München und Universität Lausanne; Dissertation am Max-Planck-Institut für Biochemie, Martinsried und Promotion 1977; 1979 -1982 Wissenschaftlicher Assistent, Physiologisch-chemisches Institut der Julius-Maximilians-Universität Würzburg; 1982-1984 Forschungsstipendium der Deutschen Forschungsgemeinschaft, Honorary Research Assistant am University College, Dept. of Biophysics, London; 19841994 Wissenschaftlicher Angestellter und Akad. Rat, Physiologisch-chemisches Institut der Eberhard-Karls-Universität Tübingen; 1987 Habilitation für das Fach Physiologie an der Medizinischen Fakultät (Theoretische Medizin) der Universität Tübingen; ab 1991 Leiter einer eigenständigen Arbeitsgruppe: „Molekulare Neurophysiologie“; 1992 Habilitation für das Fach Physiologische Chemie und Biochemie an der Fakultät für Chemie und Pharmazie der Universität Tübingen; 1994 Professur für Biochemie / Neurobiochemie an der Otto-von-GuerickeUniversität Magdeburg und Direktor des Instituts für Neurobiochemie. Seit 1996 Sprecher des DFG-Graduiertenkollegs: „Biologische Grundlagen von Erkrankungen des Nervensystems“ an der Medizinischen Fakultät der Universität in Magdeburg. Forschungsarbeiten zu (i) molekularen Mechanismen der Neurotransmitter-vermittelten Signaltransduktion in Gliazellen, einschließlich physiologischer Funktionen von ATP 243 ARTIKEL DES QUARTALS und Thrombin, sowie deren Rezeptoren, (ii) Ca2+-Regulation und Energiestoffwechsel bei Neuroprotektion und Neurodegeneration bei Ischämie; (iii) Beteiligung der Mitochondrien bei Regulation astrocytärer Funktionen und Glia-Neuron-Wechselwirkung. Dr. Tanuja Rohatgi: 1991-1996 Studium Zoologie und Molekular-Biologie, Universität Delhi, Indien. 1996-1999: Research fellow, Centre for Biochemical Technology, Delhi, Indien. 2000-2004: Promotionsstudium an der Otto-von-Guericke-Universität, Magdeburg. 2004 Promotion in Neurobiochemie, Universität Magdeburg. Liste der Abkürzungen Korrespondenzadresse DAG Diacylglycerol ERK Extracellular signal regulated kinase GDP Guanosin Diphosphat GTP Guanosin Triphosphat HIV Human Immunodefizienz Virus Inositol 1,4,5-trisphosphat IP3 MAPK Mitogen-aktivierte Proteinkinase PNS Peripheres Nervensystem PARs Protease Aktivierte Rezeptoren PN-1 Protease Nexin 1 PLC Phospholipase C PKC Proteinkinase C ZNS Zentrales Nervensystem Prof. Dr. G. Reiser Institut für Neurobiochemie Otto-von-Guericke-Universität Magdeburg Medizinische Fakultät Leipziger Straße 44 D-39120 Magdeburg Tel.: ++ 49 (0) 391 671 3088 Fax: ++ 49 (0) 391 671 3097 e-mail:[email protected] ARTIKEL DES QUARTALS Vorgestellt von Michael Frotscher Institut für Anatomie und Zellbiologie der Albert-Ludwigs-Universität Freiburg, Postfach 111, 79001 Freiburg Enhanced synaptic plasticity in newly generated granule cells of the adult hippocampus Christoph Schmidt-Hieber, Peter Jonas, Josef Bischofberger Erschienen in Nature, 13 May 2004; 429, 184-187 (2004). Die meisten von uns haben während ihres Studiums noch gelernt, dass die Bildung von Nervenzellen etwa zum Zeitpunkt der Geburt abklingt. Später wurde diese Aussage dahingehend revidiert, dass in einzelnen Hirnregionen die Neurogenese noch anhalten kann, insbesondere wenn diese Hirnregionen spät gebildet werden. Hierzu gehört der Gyrus dentatus der Hippocampusformation. Wir wissen heute, dass in dieser Hirnregion lebenslang neue Nervenzellen gebildet werden. Es kam als eine ziemliche Überraschung, dass diese Neubildung von Neuronen durch physische Aktivität kontrolliert wird (van Praag et al. 1999). Mancher Läufer unter uns hat sich an dieser Stelle wohl ganz zufrieden zurückgelehnt. Warum aber werden diese neuen Nervenzellen gebildet? Wir alle wissen, dass der Hippocampus eine wichtige Rolle bei Lern- und Gedächtnisprozessen spielt. Hat die lebenslange Neubildung von Nervenzellen etwas mit Lernen und Gedächtnis zu tun? Wir wissen, dass neugebildete Nervenzellen nicht am Ort ihrer Bildung verharren, sondern zu ihrem Be244 stimmungsort hinwandern. Wie muss man sich diesen Prozess im ausdifferenzierten Gehirn vorstellen? Wir wissen aus den Arbeiten von Pasko Rakic, dass neugebildete Nervenzellen entlang von Radialgliafasern migrieren. Ist denn ein solches Radialgliagerüst im adulten Gyrus dentatus vorhanden? Wir haben auch gelernt, dass Radialgliazellen Vorläuferzellen von Neuronen sind. Wie kann man sich Nervenzellneubildung und neuronale Migration im Gyrus dentatus des ausdifferenzierten Gehirns vorstellen? Vor allem aber wollen wir wissen, was denn die Funktion dieser neugebildeten Neurone ist und wie sie sich mit ihren auswachsenden Fortsätzen in das präexistente Netzwerk des Hippocampus integrieren. Zu diesem Fragenkatalog haben SchmidtHieber et al. mit ihren Untersuchungen einen wesentlichen Beitrag geleistet. Die Autoren fanden, dass reife und junge Körnerzellen des Gyrus dentatus unterschiedliche funktionelle Eigenschaften aufweisen. Neugebildete unreife Körnerzellen sind viel leichter erregbar als die benachbarten ‚alten’ Nervenzellen. Christoph Schmidt-Hieber, Peter Jonas und Josef Bischofberger (von links) haben die Rolle von jungen Körnerzellen im adulten Hippocampus untersucht. Mehr noch, auch die Induktion von assoziativer synaptischer Plastizität ist deutlich erleichtert. Während bei reifen Körnerzellen zur Induktion synaptischer Verstärkung (LTP) neben der Afferenzstimulation auch “Burst”Entladungen der postsynaptischen Körnerzelle erforderlich waren, konnte bei unreifen Körnerzellen LTP bereits durch eine Kombination von afferenter Stimulation mit einzelnen postsynaptischen Spikes induziert werden. Damit ist die Schwelle für die LTP-Induktion bei den jungen Nervenzellen signifikant niedriger. Bevor die Autoren jedoch diese Schlussfolgerung ziehen konnten, mussten sie die beiden unterschiedlichen Zellen, junge und alte Körnerzellen, eindeutig identifizieren. Dabei kam ihnen zunächst zu Hilfe, dass junge Körnerzellen immer am unteren Rand des Körnerzellbandes gelegen sind, im Gyrus dentatus also eine outside-in-Schichtung (im Gegensatz zur inside-out-Schichtung des Neokortex) vorliegt. Weiterhin haben sie die abgeleiteten Zellen mittels intrazellulärer Biocytinfüllung markiert und mit Antikörpern gegen PSA-NCAM angefärbt. Bekanntlich wird diese embryonale Form von NCAM im adulten Hippocampus nur noch in neugebildeten Körnerzellen exprimiert. Schließlich Neuroforum 3/04 ARTIKEL DES QUARTALS wissen wir aus Untersuchungen zur postnatalen Entwicklung der Körnerzellen sehr genau, wie junge und wie reife Körnerzelldendritenbäume aussehen (Abb. 1). Die Autoren waren damit in der Lage, die funktionellen Eigenschaften ihrer abgeleiteten Zellen direkt solchen unreifen oder differenzierten Körnerzellen zuzuordnen. Welche Bedeutung haben nun die Ergebnisse? Ich habe bereits die Frage aufgeworfen, dass es zu verstehen gilt, wie die neugebildeten Körnerzellen in das Netzwerk der Hippocampusformation integriert werden. Werden sie überhaupt in das Netzwerk integriert? Untersuchungen anderer Autoren hatten Hinweise dafür geliefert, dass ein beträchtlicher Teil der neugeborenen Zellen abstirbt. Andere Untersuchungen haben aufgezeigt, dass eine abwechslungsreiche Umgebung oder räumliches Lernen die Wahrscheinlichkeit für das Überleben der Neurone deutlich erhöht (Gage 2000). So haben auch SchmidtHieber et al. ihre Labortiere nicht in „normalen“ Käfigen gehalten, sondern besonders große Käfige mit Kletterwänden, Laufrädern und Kletterröhren benutzt. Eine höhere synaptische Plastizität während der räumlichen Exploration könnte für das Überleben der neugebildeten Zellen nützlich sein. Die Eigenschaften der neugebildeten Neurone wurden von Schmidt-Hieber et al. im isolierten Hirnschnittpräparat untersucht. Sind die ermittelten Befunde auch auf die in-vivo Situation übertragbar, so ergeben sich weitere interessante Zusammenhänge zwischen erhöhter Plastizität und räumlichem Lernen. Im Zentrum eines definierten Ortes („place field“) feuern die betreffenden Körnerzellen mit „Burst“-Entladungen. „Burst“-Entladungen kombiniert mit afferenter Stimulation induzierten bei den ausdifferenzierten Körnerzellen LTP, während LTP bei jungen Körnerzellen bereits dann induziert werden konnte, wenn afferente Stimulation mit Einzelentladungen kombiniert war. Dies kann bedeuten, dass junge Körnerzellen gleichsam ein größeres Einzugsgebiet beim räumlichen Lernen besitzen. Mit zunehmendem Alter würde bei dieser Annahme dann eine höhere Ortsspezifität für das räumliche Lernen entstehen. Wenngleich auch diese Annahmen noch überprüft werden müssen, zeigen die vorliegenden Untersuchungen jedoch in eindrucksvoller Weise auf, dass morphologisch eindeutig als unreif identifizierte Körnerzellen eine deutlich höhere synaptische Plastizität als ausdifferenzierte Körnerzellen besitzen. Literatur van Praag, H., Christie, B.R., Sejnowski, T.J. und Gage, F.H. (1999): Running enhances neuro- Neuroforum 3/04 Abb. 1: Neugebildete Körnerzellen befinden sich am inneren Rand der Körnerzellschicht. A.) Das konfokale Fluoreszenzbild zeigt eine reife und eine unreife Körnerzelle im hippocampalen Hirnschnitt der Ratte. Während der elekrophysiologischen Ableitung wurden die Zellen mit Biocytin gefüllt und anschließend fluoreszenzmarkiert (grün). Diese Neurone sind in ein Netzwerk von GFAP-positiven Gliazellen (rot) eingebettet. B.) Vergrößerte Darstellung der unreifen Zelle. Maßstab in Abb. A.) und B.) 100 µm bzw. 20µm. genesis, learning, and long-term potentiation in mice. Proc Natl Acad Sci USA 96: 1342713431. Gage, F.H. (2000): Mammalian neural stem cells. Science 287: 1433-1438. Fragen an die Autoren Frage: Wie sind Sie auf die im Artikel beschriebenen Befunde gestoßen? Welche Arbeiten haben Sie zu Ihrer Fragestellung inspiriert? Josef Bischofberger: Bereits vor einigen Jahren ist mir aufgefallen, dass sich die Körnerzellen des Hippocampus am inneren Rand der Körnerzellschicht erstaunlich inhomogen verhalten. Dies war damals sehr störend. Als sich dann herausstellte, dass es sich hier um morphologisch und immunhistochemisch distinkte Teilpopulationen (eben reife und unreife Neurone) handelt, konnten wir der Ursache endlich auf den Grund gehen. Wichtige Arbeiten, die uns bestärkt haben, die Funktion dieser Zellen zu studieren, kamen aus den Gruppen von Fred Gage und Elizabeth Gould, die unter anderem zeigen konnten, dass adulte Neurogenese auch im humanen Hippocampus vorkommt (Erikson et al. 1998, Nat Med 4:1313-1317) und dass die neugebildeten Nervenzellen das Lernen erleichtern (Shors et al. 2001, Nature 410:372-376). Peter Jonas: Meine Abteilung beschäftigt sich mit den Mechanismen der synaptischen Übertragung und der funktionellen Plastizität im Hippocampus. Dabei haben wir uns insbesondere mit der Funktion präsynaptischer Elemente und der Struktur und Funktion inhibitorischer Interneurone befasst. Es schien, als ob wir die wesentlichen zellulären Komponenten des hippocampalen Netz- werkes verstanden hätten. Als mehrere Gruppen vor einigen Jahren Evidenzen für adulte Neurogenese veröffentlichten, waren wir sehr überrascht und wollten auch dieser neuen strukturellen Plastizitätsform auf den Grund gehen. Frage: Wann haben Sie begonnen, sich für die Neurowissenschaften zu interessieren? Christoph Schmidt-Hieber: Am Ende meiner Schulzeit interessierte ich mich sehr für künstliche Intelligenz, unter anderem wegen der damals aktuellen Diskussion über das Schachduell zwischen Deep Blue und Kasparow. Die Beschäftigung mit dem Thema weckte letztendlich auch mein Interesse an der „Natürlichen Intelligenz“, insbesondere an den Mechanismen von Lernen und Gedächtnis. Josef Bischofberger: Während der ersten Semester meines Physikstudiums kam mir ein Buch des Psychiaters und Neurologen Hoimar von Ditfurth in die Hände: „Der Geist fiel nicht vom Himmel – Die Evolution unseres Bewusstseins“. Von da an hat mich das Thema nicht mehr losgelassen. Frage: Warum sind Sie Wissenschaftler geworden? Christoph Schmidt-Hieber: Bin ich (noch) nicht. Josef Bischofberger: Weil ich gerne wissen würde, wie das Gehirn funktioniert. Frage: Wer oder was hat Sie wissenschaftlich besonders geprägt? Christoph Schmidt-Hieber: Im Rahmen meines Studiums vor allem das PhysiologieSeminar bei Professor Jonas und damit der erste Kontakt mit der Welt der Wissenschaft. 245 ARTIKEL DES QUARTALS Die Zusammenhänge zwischen den grundlegenden Funktionen des menschlichen Körpers und deren Störungen bei Krankheiten haben mich sehr fasziniert. Josef Bischofberger: Was die Neurophysiologie betrifft, so erinnere ich mich an ein Seminar in der Arbeitsgruppe von Erwin Neher in Göttingen über Struktur und Funktion von Ionenkanälen. Das fand ich damals sehr aufregend! Sehr beeindruckend war für mich aber auch die Tübinger Zeit und der damals dort arbeitende Mathematiker Simon Ruijsenaars. Er hat uns in den ersten Semestern beigebracht, dass der wichtigste Schritt bei der Bearbeitung eines Problems darin besteht, sich sehr genau zu überlegen, ob prinzipiell eine Lösung existiert. Der Rest ergibt sich dann fast von alleine. Frage: Welche menschlichen Eigenschaften sind Ihres Erachtens für eine erfolgreiche wissenschaftliche Karriere eine wichtige Voraussetzung? Christoph Schmidt-Hieber: Die Beantwortung dieser Frage überlasse ich lieber anderen. Josef Bischofberger: Ich denke insbesondere Neugierde und Begeisterungsfähigkeit. Frage: Wie schätzen Sie die gegenwärtige Situation an den deutschen Universitäten ein? Christoph Schmidt-Hieber: Meiner Erfahrung nach hängen Erfolg und Misserfolg im Studium sehr viel mehr von der eigenen Initiative als von den Rahmenbedingungen ab. Insofern möchte ich mich nicht über knappe Kassen und geringe personelle Mittel beklagen. Josef Bischofberger: Die Situation ist meiner Ansicht nach an vielen Hochschulen im Moment nicht so schlecht. Viele der neu eingeführten Regelungen (12 Jahresbeschränkung, Juniorprofessoren, Stellenabbau) werden allerdings zu großen Problemen führen, wenn hier keine gesetzlichen Nachbesserungen vorgenommen werden. Peter Jonas: Als ich vor fast zehn Jahren an die Universität Freiburg berufen wurde, hatte ich die Vision, ein Zentrum mit optimalen Rahmenbedingungen für neurophysiologische Spitzenforschung zu schaffen. Ich habe dieses Ziel nicht ganz erreicht, bin ihm aber zumindest näher gekommen. Leider ist die Arbeit in den letzen Jahren zunehmend schwieriger geworden. Die Grundprobleme sind: Viel zu hohe Lehrbelastung, fehlende „tenure track“-Stellen für den wissenschaftlichen Nachwuchs, zu wenig Mittel für die Grundlagenforschung, zu geringe Leistungsorien246 tierung bei der Ressourcen-Verteilung und mangelhafte Klarheit in den politischen Rahmenbedingungen. Frage: Was raten Sie begabten Studenten, die sich für eine wissenschaftliche Laufbahn interessieren? Christoph Schmidt-Hieber: Ich kann hier nur für meine Doktorarbeit sprechen. An dem erfolgreichen Verlauf des Projektes waren vor allem zwei Faktoren beteiligt: Zum einen die hervorragende Betreuung, zum anderen die sehr weit entwickelte Hirnschnitt-Technik am Institut von Professor Jonas. Josef Bischofberger: Man sollte möglichst früh den persönlichen Kontakt zu Wissenschaftlern in verschiedenen Labors suchen. Laborpraktika und Hiwi-Jobs bieten hier eine effektive Möglichkeit, Arbeitsgruppen kennen zu lernen und seinen Platz zu finden. Frage: Wie würden Sie die Sonnen- und Schattenseiten Ihres Wissenschaftlerlebens beschreiben? Christoph Schmidt-Hieber: Die Sonnenseiten: Den Dingen auf vielfältige Weise auf den Grund gehen zu können; recht große Freiheiten, was beispielsweise Zeitplanung und Themenauswahl betrifft. Die Schattenseiten: Hoher Arbeitseinsatz, der unter Umständen auch umsonst gewesen sein kann. Josef Bischofberger: Im Allgemeinen schlägt man sich oft mit kurzen Zeitverträgen herum. Diese unsichere Zukunft ist sicher eine der größten Schwierigkeiten unseres Berufs, insbesondere wenn man Familie mit Kindern hat. Zu den Sonnenseiten gehören die kleinen Glücksgefühle nach einer erfolgreichen Publikation, die Möglichkeit, Dinge zu sehen, die noch niemand zuvor gesehen hat und die Kommunikation mit ganz unterschiedlichen und oft sehr interessanten Menschen quer über den Globus. Frage: Womit beschäftigen Sie sich, wenn Sie nicht forschen oder lehren? Christoph Schmidt-Hieber: Derzeit vor allem mit der Examensvorbereitung, ansonsten Reisen, Sport und Fotografie. Peter Jonas: In meiner minimalen Freizeit mache ich Bergläufe auf Schauinsland, Kandel und andere Freiburger Hausberge, gerne auch gemeinsam mit dem Rezensenten unserer Arbeit. Manchmal nehme ich Manuskripte und ein Diktiergerät mit. Ob dies die Neurogenese fördert, ist für mich dabei nicht von primärem Interesse. Josef Bischofberger: Wenn Zeit bleibt, mache ich gerne Jazz mit Posaune oder Bass. Ich jogge nicht so viel wie manch anderer, fahre aber dafür mit dem Fahrrad durch den Schwarzwald. NWG Preis für den Bundeswettbewerbes Jugend forscht 2004 Die Neurowissenschaftliche Gesellschaft sieht ihre Aufgabe nicht nur darin, etablierte Wissenschaftler, die bereits ihren Weg in die Neurowissenschaft gefunden haben, zu vertreten. Sie bemüht sich vielmehr intensiv auch um den wissenschaftlichen Nachwuchs und setzt dabei bereits in der Schule an. Seit Jahren schon werden bundesweit Fortbildungsveranstaltungen für Biologie-Lehrer der gymnasialen Oberstufe veranstaltet, um die neuesten Erkenntnisse neurowissenschaftlicher Forschung in die Schulen zu tragen. In diesem Jahr stiftet die Gesellschaft zum ersten Mal einen Sonderpreis für den Bundeswettbewerb Jugend forscht. Dieser Sonderpreis wurde für die beste Arbeit auf dem Gebiet der Neurowissenschaft vergeben und ist mit 500,– D dotiert. Außerdem wurde einer der Preisträger zu der im kommenden Februar stattfindenden Jahrestagung der Neurowissenschaftlichen Gesellschaft nach Göttingen eingeladen, wobei die Gesellschaft die Reise- und Hotelkosten übernimmt. Zusätzlich erhält er kostenlos für ein Jahr Neuroforum. Preisträger sind drei Schüler aus Berlin und Potsdam (Niko Hübner-Kosney (20 Jahre), Katharina Hoffmann (20 Jahre) und Arvid Heise (20 Jahre). Sie erhalten den Preis für Ihre Arbeit „Entspannung auf Knopfdruck Wirkungsweise und Wirkungsnachweis einer selbst gebauten Mind Machine“. Wenn der Radiowecker anspringt, behält Niko Hübner-Kosney das erste Lied nach dem Aufwachen noch lange im Ohr. Warum das so ist, hat er gemeinsam mit Katharina Hoffmann und Arvid Heise herausgefunden. Das Gehirn ist morgens im Alpha-Zustand - entspannt und in hohem Maße aufnahme- und konzentrationsfähig. Die drei wollten zeigen, dass es möglich ist, die Gehirnwellen und damit den Bewusstseinszustand gezielt zu verändern. Testpersonen erhielten über Kopfhörer rechts und links Töne mit verschiedener Hertz-Zahl, außerdem über eine mit LCDs bestückte Sonnenbrille gleichzeitig unterschiedliche Blinksignale. Mit Hilfe eines EEGs wiesen die Jungforscher nach, dass das Gehirn als Reaktion auf die Reize veränderte Gehirnwellen produziert. Nach 20 bis 30 Minuten an der Mind Machine fühlten sich die Testpersonen entspannt und konzentriert, ihr Puls ging ruhig und gleichmäßig. Eine einfache Methode, Stresssituationen zu bewältigen und die Konzentration zu steigern. Neuroforum 3/04 Preisgünstige Studienausgabe: Lexikon der Neurowissenschaft INSTITUTSVORSTELLUNG Interdisziplinäres Institut für MS-Forschung in Göttingen Bereich Humanmedizin der Universität Göttingen und Gemeinnützige Hertie-Stiftung ,– Früher g 596 jetzt g 99,-!! Ralf Gold und Mathias Bähr Die multiple Sklerose ist eine der häufigsten Erkrankungen des Nervensystems, an der etwa 120.000 Menschen in Deutschland leiden. Obschon die Krankheit seit mehr als einhundert Jahren bekannt ist und erforscht wird, ist die eigentliche Ursache noch immer unklar und es fehlen sicher wirksame Maßnahmen zur Eindämmung und Heilung (s.a. Übersichtsarbeit Aktas und Zipp im NeuroForum 2.04). In den letzten Jahren wird dazu in den nächsten zehn Jahren bis zu 4,6 Mio. a zur Verfügung stellen. Göttingen hatte sich im Jahr 2001 im öffentlichen Wettbewerb „Molekulare Neuroimmunologie der Multiplen Sklerose“ der GHS durchgesetzt und damit den Zuschlag für die Fördermittel der Stiftung erhalten, so dass das erste interdisziplinäre Institut für Multiple-Sklerose- Forschung in der Bundesrepublik am 31.03.2004 in Göttingen Das vierbändige Lexikon der Neurowissenschaft vermittelt einen umfassenden und im Bereich deutschsprachiger Nachschlagewerke einzigartigen Überblick zu allen neurowissenschaftlichen Fachgebieten. Das gesamte derzeit verfügbare Wissen wird von kompetenten und renommierten Vertretern der einzelnen Fachdisziplinen in rund 14.500 Stichwörtern und 31 ausführlichen Essaybeiträgen verständlich vermittelt und über ein weitreichendes Verweissystem erschlossen. Mit der preiswerten Studienausgabe sparen Sie u 497,– gegenüber der Originalausgabe (mit der sog. Kombi-Ausgabe sogar u 745,–) !! „Das ‚Lexikon der Neurowissenschaft‘ ist ein außerordentlich gelungenes Nachschlagewerk, das sowohl Neurobiologen wie auch all jene begeistern wird, die sich für dieses zukunftsweisende Fachgebiet interessieren. (…)“ Gehirn und Geist 2001, 4 Bde., ca. 450 S. pro Bd., geb. – insgesamt: ca. 14.500 Stichwörter, rund 1.200 Abb. und Tab., 31 Essays ■ Gesamtausgabe Buch: u 99,-, ISBN 3-8274-0451-7 ■ Gesamtausgabe CD-ROM: u 99,-, ISBN 3-8274-0456-8 ■ Gesamtausgabe Buch + CD-ROM: u 149,–, ISBN 3-8274-0455-X ■ haben sich jedoch die Hinweise darauf verstärkt, dass die eigentliche Ursache der multiplen Sklerose in einer Störung des körpereigenen Immunsystems liegt. Die Gemeinnützige Hertie-Stiftung (GHS) will die wissenschaftlichen Erkenntnisse auf diesem Gebiet mit einem herausragenden, erfolgversprechenden und langfristigen Forschungsvorhaben gezielt vorantreiben und Neuroforum 3/04 offiziell eröffnet werden konnte. Das Forschungsinstitut hat seinen Sitz in der 2. Etage im Waldweg 33 in Göttingen (s. Bild). Die Räumlichkeiten dort wurden mit Unterstützung des Landes Niedersachsen ausund umgebaut. Im Rahmen des Eröffnungssymposiums wurde auch der ehemalige Direktor der Abteilung Neurologie, Prof. Dr. Helmut Bauer, anlässlich seines 90. Ge- ! Ausführliche Informationen unter www.elsevier.de 247 Bestellen können Sie ■ telefonisch 07071/935369 ■ per Fax: 07071/935393 ■ per Mail: [email protected] INSTITUTSVORSTELLUNG burtstages geehrt. Prof. Bauer hatte wesentlichen Anteil an der MS-Forschung und Therapie nach dem 2. Weltkrieg und entscheidend dazu beigetragen, dass die deutsche MS-Forschung wieder eine internationale Reputation erhielt. Mit einer aufsehenerregenden Publikation von Trapp (NEJM 1998; 338: 278) wurde das Interesse der MS-Forschung an den neurodegenerativen Aspekten dieser chronisch-entzündlichen Erkrankung des Zentralnervensystems neu geweckt. Daraus ergab sich auch, dass innovative, interdisziplinäre Ansätze erforderlich sind. In einem international begutachteten Wettbewerb der GHS zum Thema „Molekulare Neuroimmunologie“ wurde ein unter Leitung von Professor Dr. Mathias Bähr (Direktor der Abt. Neurologie – Bereich Humanmedizin) und Privatdozent Dr. Harald Neumann (Forschergruppe Neuroimmunologie des European Neuroscience Institute der Universität Göttingen) gestellter Antrag mit dem Titel „Molecular mechanisms and repair strategies of demyelination and axonal damage in Multiple Sclerosis“ zur Förderung ausgewählt. Die am Institut tätigen Arbeitsgruppen wollen die neuroimmunologischen Mechanismen erforschen, die dem für die multiple Sklerose so charakteristischen Untergang der Nervenfaserscheiden und Nervenfasern zugrunde liegen. Ziel des Instituts ist es darüber hinaus, Strategien zur Reparatur dieser Schäden zu entwickeln. Schwerpunkte der geplanten Forschung sind die Analyse der molekularen Mechanismen der Myelin- und axonalen Schädigung, der zytotoxische Interaktionen zwischen Immunzellen und Neuronen sowie die Entwicklung neuer Strategien der Gen- und Zellersatztherapie in experimentellen MSModellen. An dem neuen Institut sind sechs wissenschaftliche Einrichtungen aus Göttingen vertreten: Die Abteilung Neurologie mit ihrem Schwerpunkt Neuroimmunologie des Bereichs Humanmedizin (Prof. Bähr), die Abteilung für Neuropathologie (Prof. Brück), das Max-Planck-Institut für experimentelle Medizin (Prof. Nave), die Forschergruppe Neuroimmunologie des European Neuroscience Instituts (ENI; PD Dr. Neumann), die Biomedizinische NMR For248 schungs GmbH am Max-Planck-Institut für Biophysikalische Chemie Göttingen (Prof. Frahm) und das Deutsche Primatenzentrum (Prof. Fuchs). Diese werden ergänzt durch Arbeitsgruppen des Instituts für Rekonstruktive Neurobiologie der Universität Bonn (Prof. Brüstle) und das Institut für Neuroloimmunologie der Charité Berlin (Frau Prof. Zipp). Die Leiter dieser Abteilungen sind im Vorstand vertreten und wählen aus Ihrer Mitte einen geschäftsführenden Vorsitzenden für jeweils drei Jahre. Dieser koordiniert die wissenschaftlichen Aktivitäten des IMSF und vertritt das Institut gegenüber dem Drittmittelgeber. Nach Beginn der Förderung im 2. Halbjahr 2002 wurden von den Antragstellern bereits neun Einzelprojekte und sechs aufwendige interdisziplinäre experimentelle Fragestellungen erfolgreich bearbeitet. Besonderer Wert wurde auf die Verknüpfung molekularbiologischer, elektrophysiologischer, feingeweblicher und kernspintomographischer Methodik gelegt. Parallel zum Start der Förderung erfolgte der Ausbau und die Einrichtung einer kompletten Forschungsetage im Gebäude Waldweg 33. Der damit verbundene Stiftungslehrstuhl „Experimentelle Neuroimmunologie“ wird von der GHS finanziert. Auf den Lehrstuhl wurde Prof. Dr. Ralf Gold berufen. Im Januar 2004 konnte seine Abteilung Experimentelle Neuroimmunologie ihre wissenschaftliche Arbeit beginnen, und durch die räumliche Verflechtung mit den Antragstellern die Synergieeffekte noch verstärken. Die GHS arbeitet in den Schwerpunkten Neurowissenschaften/multiple Sklerose, Europäische Integration und Erziehung zur Demokratie. Dem Willen des Stifters, Georg Karg, entsprechend wurde der medizinischen Forschung seit Gründung der Stiftung im Jahre 1974 eine hohe Priorität eingeräumt, wobei die Erforschung der multiplen Sklerose im Vordergrund stand. Die GHS ist heute die größte private Förderinstitution der Hirnforschung in Deutschland; allein in den letzten drei Jahren wurden die Neurowissenschaften mit durchschnittlich mehr als zehn Millionen C pro Jahr gefördert. Mit dem Hertie-Institut für Klinische Hirnforschung in Tübingen hat die Stiftung im Jahr 2000 das größte und modernste Zentrum für Neurologie in Deutschland gegründet, das im Juni 2004 offiziell eröffnet wurde. Auf dem Gebiet der multiplen Sklerose ist die GHS – sowohl in der Forschung als auch der sozialmedizinischen Nachsorge – die aktivste deutsche Stiftung und hat bislang insgesamt mehr als 30 Millionen C an Fördermitteln aufgewendet. Originalarbeiten aus der bisherigen IMSF Förderung Diem, R., Hobom, M., Maier, K., Weissert, R., Storch, M.K., Meyer, R. und Bähr, M. (2003): Methylprednisolone increases neuronal apoptosis during autoimmune CNS inflammation by inhibition of an endogenous neuroprotective pathway. J Neurosci 23 (18): 6993-7000. Kempermann, G. and Neumann, H. (2003): Microglia: The enemy within. Science 302: 1689-1890, invited perspective. Hobom, M., Storch, M.K., Weissert, R., Maier, K., Radhakrishnan, A., Kramer, B., Bähr, M. und Diem, R. (2004): Mechanisms and Time Course of Neuronal Degeneration in Experimental Autoimmune Encephalomyelitis. Brain Pathol 14: 148-157. Iliev, A., Stringaris, A.K., Nau, R. and Neumann, H. (2004): Neuronal injury mediated via stimulation of microglial toll-like receptor-9 (TLR9). FASEB J. 18: 412-414. Nitsch, R., Pohl, E.E., Smorodchenko, A., InfanteDuarte, C., Aktas, O. und Zipp, F. (2004): Direct impact of T cells on neurons revealed by two-photon microscopy in living brain tissue. J Neurosci. 24: 2458-2464. Michailov, G.V., Sereda, M.W., Brinkmann, B.G., Fischer, T.M., Haug, B., Birchmeier, C., Role, L., Lai, C., Schwab, M.H. und Nave, K.A. (2004): Axonal neuregulin-1 regulates myelin sheath thickness. Science 304: 700-703. Korrespondenzadresse Prof. Dr. Ralf Gold Universität Göttingen und Gemeinnützige Hertie-Stiftung, Institut für MS-Forschung Waldweg 33, D-37073 Göttingen Tel.: ++ 49 (0) 551 391 3331 e-mail: [email protected] Fehlende Mitgliederadressen Von folgenden Mitgliedern fehlt uns die korrekte Anschrift: Brand, Antje (vormals Martinsried) Czernilofsky, Dr. Armin (vormals Wien, Österreich) Dammermann, Dr. Björn (vormals Hamburg) Diesmann, Dr. Markus (vormals Göttingen) Dudel, Prof. Dr. J. (vormals München) Franken, Gilbert (vormals Magdeburg) Haase, Annely (vormals Hannover) Hartmann, Dr. Jana (vormals München) Horstmann, Sonja (vormals München) Morgenstern, Dr. Eve (vormals Berlin) Nedvetsky, Pavel (vormals Giessen) Olcese, Prof. Dr. James (vormals Hamburg) Riess, Prof. Dr. med. Olaf (vormals Tübingen) Rybak, Dr. Jürgen (vormals Würzburg) Schwarz, Stephan (vormals Bonn) Stover, Dr. John F. (vormals Philadelphia, USA) Für Hinweise sind wir dankbar. Neuroforum 3/04 INTERNETVORSTELLUNG INTERNETVORSTELLUNG „ATLAS of the HUMAN BRAIN“ Jürgen Konrad Mai und ThomasVoß Bildgebende Verfahren, insbesondere die Magnetresonanztomographie, haben die Beurteilungsmöglichkeiten von Hirnstrukturen revolutionär erweitert. Mit verbesserter Auflösung und zunehmender Darstellungsgenauigkeit wächst auch der Bedarf an In- terpretationshilfen für die gewonnenen Bilddaten des menschlichen Gehirns. Die nötigen Informationen können durch einen Referenz-Atlas vermittelt werden, wenn dieser konsistent, detailliert, funktions- und klinisch orientiert ist. Das Anforderungsprofil der Abb. 1: Konzept des elektronischen Atlas. Die verschiedenen Einzelkomponenten (Module) des Atlas (beispielhaft in der ersten Bildreihe dargestellt) können miteinander zu so genannten Navigatoren kombiniert werden. Dreidimensional korrespondierende Schichten werden dabei automatisch eingelesen, so dass beim Verfolgen einer Struktur in einem Schnitt die jeweils korrelierten Schnitte in den anderen beiden Raumebenen angezeigt werden und die betroffene Struktur in das Zentrum des Koordinatenkreuzes platziert wird. Im MR- und Brain-Navigator werden aus einer Serienschnittsequenz die orthogonal hierzu gelegenen Schnitte berechnet und das gemeinsame Volumen gekennzeichnet. Im HeadNavigator werden drei zueinander senkrecht stehende anatomische 1 cm-Scheiben von unterschiedlichen anatomischen Präparaten dargestellt. Der Anschluss an eine Datenbank mit faktischem und lexikalischem Wissen wird derzeit vorbereitet. Neuroforum 3/04 Nutzer ist allerdings sehr unterschiedlich. Es reicht von Studierenden, die diese Informationen als Lern-Programm zur Anatomie von Kopf und Gehirn nutzen wollen, über medizinisches Fachpersonal und Ärzte, die Interpretationshilfen für bildgebende Verfahren benötigen, bis zu Wissenschaftlern, die sehr detaillierte Informationen und Spezialwissen nebst wissensbasierter Datenbank abrufen wollen. Im Folgenden soll ein den unterschiedlichen Anforderungen entsprechender Atlas vorgestellt werden, in dem Daten makro- und mikroskopischer Auflösung repräsentiert sind. Dieser “Atlas of the Human Brain” (Elsevier/Academic Press, 2004), der in gedruckter und elektronischer Version vorliegt, kann als Unterrichtsmedium, Trainingstool für die Weiterbildung und als Forschungsinstrument eingesetzt werden. Der gedruckte Atlas kombiniert einen makroskopischen Atlas, der das Gehirn in seiner natürlichen Umgebung darstellt, mit einem mikroskopischen Atlas, der detailliert die Histologie des Gehirns vermittelt. Der “makroskopische Atlas” zeigt drei Schnittführungen, jeweils repräsentiert durch ein anatomisches Präparat, detailgetreue Umzeichnung, korrespondierende Röntgenabbildungen und Magnetresonanztomogramme. Jede Schnittsequenz zeigt sowohl die Ober- als auch die Unterseite der Schnittflächen der anatomischen Präparate. Der Nutzer kann diese wie Buchseiten durchblättern und analysieren, die Einzelstrukturen folglich von Schnitt zu Schnitt verfolgen. Der „mikroskopische Atlas“ umfasst ausschließlich das Gehirn. Er beruht auf einer histologischen Frontalschnittserie, die über Jahre von mehreren Neurowissenschaftlern untersucht und beschrieben wurde. Dem Publikationsumfang nach dürfte es das bestuntersuchte menschliche Gehirn sein. Diese Studien validieren die Abgrenzungen in diesem Atlas. Die Schnittsequenz ist im standardisierten Hirnraum repräsentiert; daher ist die Lage jedes Einzelschnittes, als Fotografie repräsentiert, eindeutig definiert. Jede fotografische Einzelabbildung wird durch eine Umzeichnung ergänzt. Die CD-ROM enthält zusätzlich zum Inhalt des gedruckten Buches Farbabbildungen, Originalliteratur zum Atlasgehirn, weitere Magnetresonanztomogramme in korrespondierenden Schnittebenen und 3-DNavigationsmöglichkeiten. Sie werden dem Nutzer über eine einfache webbasierte Oberfläche zugänglich gemacht. Der mikroskopische Atlasteil (Hirnteil) wird auf der CDROM um durchsuchbare Volltexte (im pdfFormat) der Originalarbeiten zu diesem Atlasgehirn ergänzt. Darunter finden sich 249 INTERNETVORSTELLUNG Abb. 2: Das Java-Programm „BrainNavigator“ basiert auf ca. 200 farbigen Schemazeichnungen des im mikroskopischen Atlas präsentierten Gehirns. Es berechnet frei wählbare orthogonale Ebenen, deren gemeinsamer Schnittpunkt mittels eines Fadenkreuzes angezeigt wird. X,Y,Z-Koordinaten lassen sich manuell eingeben und deren Position auf den Schnitten anzeigen. Der jeweils zugehörige Koronarschnitt wird im linken Bildfenster wahlweise entweder als vergrößerte Umzeichnung oder als Originalschnitt abgebildet. In ausgewählten Bezirken können Zellbilder bei höherer Vergrößerung eingeblendet werden. schwer zugängliche deskriptive Arbeiten von H. Brockhaus und R. Hassler sowie morphometrische Untersuchungen, die an den meisten wichtigen Hirnstrukturen vorgenommen wurden. Da es sich hierbei häufig um Vergleichswerte - auch zu pathologisch veränderten Gehirnen - handelt, steht hiermit ein einzigartiges Vergleichsmaterial für zukünftige Untersuchungen zur Verfügung. Die CD-ROM enthält außerdem sog. Navigatoren (Abbildung 1). Hier sind die MR-Bilddatensätze (MR-Navigator), sowie die makroskopischen (HeadNavigator) und mikroskopischen (BrainNavigator) Schnittserien in jeweils drei orthogonalen Schnittebenen angeordnet. Die dreidimensional korrespondierenden Schichten werden dabei automatisch eingelesen, so dass beim Verfolgen einer Struktur in einem Schnitt die jeweils korrelierten Schnitte in den anderen beiden Raumebenen angezeigt werden und die betroffene Struktur in das Zentrum des Koordinatenkreuzes platziert wird. Auf diese Weise kann sich der Nutzer jeden Bildpunkt auf den drei orthogonalen Bilddatensätzen anzeigen lassen und die topographischen Nachbarschaftsverhältnisse interessierender Strukturen studieren. Im Java-Programm „BrainNavigator“ kann die gesamte Hemisphäre berücksichtigt werden oder allein der Subkortex, wodurch eine Darstellung in höherer Auflösung ermöglicht wird. Der Koronarschnittebene ist der zugehörige Myelinschnitt unterlegt; in ihn lassen sich die Areale einblenden, von denen Fotografien von der Zytomorphologie 250 der gekennzeichneten Region verfügbar sind. Das Programm erlaubt den Import von Fremdkoordinaten, beispielsweise aus der funktionellen Bildgebung, so dass Aktivierungsorte im entsprechenden histologischen Umfeld dargestellt werden können. Umfangreiche Informationen zum Inhalt aller Atlaskomponenten bietet die Website www.thehumanbrain.info, auf der auch Updates abrufbar sind. Ein Demo-Applet des BrainNavigator kann auf der Atlaswebsite evaluiert werden. Die genannte Website enthält gegenwärtig zwei 3-D Navigationsprogramme, die unabhängig von Atlas oder CD-ROM nutzbar sind: MR-Navigator und BrainNavigator, unter dem Menüpunkt „Navigators“ aufrufbar, entsprechen in abgespeckter Form den bereits beschriebenen Programmen zur Navigation im MR- bzw. histologischen Bilddatensatz. Der Zugriff auf das BrainNavigator Applet (www.thehumanbrain.net/navigator/demo.html) setzt allerdings eine schnelle Webanbindung und eine aktuelle Java-Version voraus. Der BrainNavigator erlaubt das Browsen durch die Farbdiagramme des Atlashirns und errechnet neue virtuelle Schnittebenen. Der Farbcode hilft, wichtige Hirnstrukturen dreidimensional zu verfolgen; welche dies sind, erfährt der Benutzer, indem er den Mauspfeil darauf platziert. Die manuelle Koordinatenangabe setzt das Fadenkreuz direkt auf die gewünschte Position. Die Website ist Teil eines webbasierten Atlaskonzepts, das neben den geschilderten Strukturinformationen auch Informatio- nen aus Datenbanken enthält und in standardisierter Weise dem Nutzer zur Verfügung stellt. Die im vorgestellten „Atlas of the Human Brain“ enthaltenen Komponenten sind komplex und, wie Module für unterschiedliche Anwendungsschwerpunkte, vielfältig kombinierbar. Es findet deshalb im Rahmen der 2. Internationalen Santa Lucia Konferenz: „The Human Brain – Modelling and Remodelling“ am 6. Oktober 2004 in Rom (www.thehumanbrain.org) ein Workshop statt, in dem die verschiedenen Komponenten und Anwendungsbeispiele (z.B. für die funktionelle Bildgebung und Neurochirurgie) vorgestellt werden. Korrespondenzadresse Prof. Dr. Jürgen Konrad Mai Abteilung Anatomie H.-Heine University Duesseldorf Universitaetsstr. 1 D-40225 Duesseldorf Tel./Fax: 0211 / 81-12763 (-12367) Tel.: 0211 / 3119005 eMail: [email protected] Neuroforum 3/04 NWG / NACHRICHTEN n ....... Neurowissenschaftliche Gesellschaft Neurowissenschaftliche Gesellschaft e.V. Protokoll der Mitgliederversammlung am 12. Juli 2004 während des FENS Forum 2004 in Lissabon, 12.00-13.00 Uhr Versammlungsleiter ist der Präsident der Neurowissenschaftlichen Gesellschaft, Prof. Dr. Herbert Zimmermann Protokollführer sind der Generalsekretär der Neurowissenschaftlichen Gesellschaft, Prof. Dr. Helmut Kettenmann und Meino Gibson. Die Zahl der erschienenen Mitglieder beträgt 30. Die Versammlung wurde satzungsgemäß einberufen, die Tagesordnung war den Mitgliedern bei der Einberufung mitgeteilt worden, eine Ergänzung der Tagesordnung wurde nicht gewünscht. sam die finanzielle Verantwortung. Das Local Organizing Committee (LOC) wurde gleichermaßen mit deutschen wie mit österreichischen Mitgliedern besetzt. Hotelzimmer wurden bereits reserviert, da in dieser Zeit keine Saison ist, sind die Preise moderat. Die Nutzung der Wiener Öffentlichen Verkehrsmittel wird kostenlos sein. Die Ausstellungsfläche im Kongresszentrum ist größer als in Lissabon, Poster und Industrie werden gemischt sein. Die Firma Herlitz, die auch die Ausstellung beim SfN Meeting betreut, wird sich um die Industrie kümmern. Für junge Wissenschaftler wird es ein spezielles Programm geben. 3. Bericht des Schatzmeisters Beginn: Ende: 12.00 Uhr 13.00 Uhr Tagesordnung: 1. 2. 3. 4. 5. 6. 7. 8. Begrüßung durch den Präsidenten FENS Forum 2006 in Wien Bericht des Schatzmeisters Mitgliederstand Göttinger Tagung 2005 Schilling-Preis Aktivitäten der Gesellschaft Verschiedenes 1. Begrüßung durch den Präsidenten H. Zimmermann begrüßt die Anwesenden und eröffnet die Sitzung. Das Protokoll der letzten Mitgliederversammlung vom 14. Juni 2003 wird mit 30 Ja-Stimmen und 0 Nein-Stimmen und 0 Enthaltungen angenommen. 2. FENS Forum 2006 in Wien (8. – 12. Juli 2006) Alois Saria berichtet über den Stand der Vorbereitungen für das FENS Forum 2006 im Austria Center in Wien. NWG und ANA (Austrian Neuroscience Association) sind gleichberechtigte Partner und tragen gemeinNeuroforum 3/04 A. Draguhn gibt den Kassenbericht für das Jahr 2003: die Einnahmen betrugen 235.438,50 C, die Ausgaben 224.153,22 C. Der Kassenbestand zum 31.12.03 betrug auf dem Girokonto 14.546,14 C, dazu kamen Sparanlagen in Höhe von 62.299,93 C. Damit beläuft sich das Vermögen der Gesellschaft zum 31.12.03 auf 76.846,07 C. Die Kasse wurde von den Kassenprüfern Professor U. Dirnagl und Professor A. Elepfandt geprüft. Beide Kassenprüfer bestätigen die ordentliche Kassenführung und empfehlen der Mitgliederversammlung, dem Ergebnis der Kassenprüfung zuzustimmen. Die Mitgliederversammlung entlastet den Schatzmeister auf der Grundlage des Berichts der Kassenprüfer mit 29 Ja-Stimmen, 1 Enthaltung und 0 Nein-Stimmen. 4. Mitgliederstand Die Zahl der Mitglieder steigt weiterhin. Der Mitgliederstand am 30. Juni 2004 beträgt 1.658 Mitglieder, davon 1.302 ordentliche Mitglieder und 356 studentische Mitglieder. Die Verteilung der Mitglieder in den Sektionen ist weiterhin wenig verändert, wobei die Tab. 1: Entwicklung der Mitgliederzahlen Mitglieder 1995 1996 1997 1998 1999 2000 2001 2002 2003 2004 792 920 1.038 1.229 1.343 1.508 1.538 1.562 1.616 1.658 Sektion Computational Neuroscience immer noch mit Abstand die kleinste Sektion ist. Die großen Sektionen mit über 10 % sind Zelluläre Neurowissenschaften, Molekulare Neurobiologie, Systemneurobiologie, Klinische Neurowissenschaften und Kognitive Neurowissenschaften und Verhalten. 5. Göttinger Tagung 2005 H. Zimmermann berichtet über die Vorbereitungen zur nächsten Göttinger Jahrestagung der Gesellschaft, die ausnahmsweise Tab. 2: Sektionszugehörigkeit (Stand 30. Juli 2004) Zelluläre Neurowissenschaften Molekulare Neurobiologie Systemneurobiologie Klinische Neurowissenschaften Kognitive Neurowissenschaften u. Verhalten Neuropharmakologie und -toxikologie Entwicklungsneurobiologie und Neurogenetik Computational Neuroscience 2001 2001 2001 2001 24,5% 12,8% 13,2% 6,1% 18,5% 5,2% 7,5% 1,2% 24,3% 17,7% 15,0% 13,8% 12,0% 8,4% 7,5% 1,3% 24,3% 17,5% 14,9% 14,1% 11,8% 8,5% 7,6% 1,5% 23,8% 17,4% 14,6% 13,9% 12,6% 8,5% 7,5% 1,7% 251 NWG / NACHRICHTEN nicht im Sommer, sondern im Februar (17. – 20. Februar 2005) stattfinden wird. Grund dafür ist vor allem, dass es ab 2005 keine Pfingstferien, in denen das Hörsaalgebäude bisher verfügbar war, mehr geben wird, zum anderen aber auch, dass die Tagung nun in den Händen einer neuen lokalen Organisatorin – Frau Professor Dr. Kerstin Krieglstein – liegt, die im Sommer kommenden Jahres nicht verfügbar ist. In den weiteren Jahren kann die Tagung aber wieder im Sommer stattfinden. Der NWG Vorstand favorisiert für 2007 die dritte Juliwoche. Die bei der Vorbereitung der Göttinger Tagung anfallenden Aufgaben wurden zwischen der lokalen Organisatorin und der Geschäftsstelle der NWG aufgeteilt. Ein lokales Organisationsgremium unterstützt Frau Krieglstein. H. Zimmermann hat an die DFG einen Antrag zur Unterstützung der Jahrestagung gestellt. 6. Schilling-Preis Nachdem es auf der letzten Göttinger Tagung zwei Preisverleihungen gab – Novartis Preis und TILL Photonics Preis, – hat die Firma Novartis für 2005 ihre Zusage für einen Preis leider nicht wiederholt. Stattdessen konnte auf Initiative von H. Zimmermann und mit Unterstützung von Prof. Heinz, Wässle beim Stifterverband für die Deutsche Wissenschaft der mit 20.000 Euro dotierte Schilling-Preis der Neurowissenschaftlichen Gesellschaft eingeworben werden. Er ist als Nachwuchspreis für Bewerber bis zu einem Alter von max. 35 Jahren als STELLENMARKT Forschungspreis ausgeschrieben. Bewerbungsschluss ist der 1. September 2004. Die Verleihung wird auf der Göttinger Jahrestagung im Februar stattfinden. H. Zimmermann dankt Herrn Wässle für seinen Einsatz. 7. Aktivitäten der Gesellschaft H. Zimmermann berichtet kurz über die Aktivitäten der Gesellschaft. Die Lehrerfortbildung ist nach wie vor sehr erfolgreich und hat sich vieler Orts zu einer festen Einrichtung etabliert. Das von der Hertie-Stiftung zur Verfügung stehende Geld wird noch für knapp zwei weitere Jahre reichen. Die Methodenkurse finden ebenfalls in bewährter Form statt. H. Zimmermann dankt Herrn Reifenberger für sein Engagement. Wie auch in den vergangenen Jahren wurden 100 Abstract Slots für das SfN Meeting in San Diego verteilt. Auch können NWG Mitglieder sich wieder zur reduzierten Gebühr für Mitglieder registrieren. Neuroforum läuft nach wie vor problemlos. Die Geschäftsstelle ist im Augenblick dabei, alle Neuroforum-Autoren um die Erlaubnis zu bitten, die Abbildungen auf die Homepage der NWG stellen zu dürfen. Diese Bildquellen sind besonders als Service für die Lehre gedacht. Ebenfalls auf die Homepage gesetzt wurde die DFG-Broschüre zu Tierversuchen. Erstmals hat die NWG in diesem Jahr auf Anregung von Herrn Pflüger einen Sonderpreis in Höhe von 500 Euro im Rahmen des Bundeswettbewerbs „Jugend forscht“ ausgeschrieben. Preisträger sind drei Schüler aus Potsdam und Berlin mit Ihrer „Mind Machine“. Die Urania, ein traditionsreicher Berliner Volksbildungsverein, hat die NWG zur Unterstützung eines Programms „Halbzeit in der Dekade des Gehirns“ aufgefordert. Die NWG wird sich ideell an dem Programm beteiligen. 8. Verschiedenes J. Deitmer fragt an, ob es möglich ist, auf die DFG bezüglich der gegenwärtig sehr langen Bearbeitungszeiten Einfluss zu nehmen. C. Becker hält dem entgegen, dass sich die Bearbeitungszeit auf Grund der Umstellung auf Fachkollegien verlängert hat und man hier etwas Geduld haben solle. Versammlungsleiter: Prof. Dr. Herbert Zimmermann (Präsident) Protokollführer Prof. Dr. Helmut Kettenmann (Generalsekretär) 252 Neuroforum 3/04 BÜCHER HEIMKEHRER BÖRSE Moritz Hertel, New York, USA Ausbildung – Studium der Biologie in Basel und Würzburg – Diplomarbeit: MPI für Neurobiologie, Abteilung Yves-Alain Barde „Purification and Biological Characterization of the BDNF Precursor Protein“ – Doktorarbeit: ETH Zürich Prof. Sabine Werner in Zusammenarbeit mit Prof. Christian Alzheimer (LMU München, jetzt Kiel) „Novel Players in Neuroprotection and Tissue Reorganisation after Hippocampal Lesions“ – Seit 2002 Postdoc an der Rockefeller Universität in New York, im Labor von Prof. Fernando Nottebohm „Screen for Genes involved in Adult Neurogenesis in the Brain of Songbirds“ Forschungsarbeit Nach meiner Doktorarbeit, während der ich mich mit Läsionen des murinen Hippocampus beschäftigt hatte, richtete sich mein Interesse zunehmend auf die adulte Neurogenese. Die besonders hohe Rate von neu entstehenden Nervenzellen im Gehirn von ausgewachsenen Singvögeln ließ mich darüber nachdenken ob sich dieses System für die gezielte Suche nach Genen eignet, die hierbei eine Rolle spielen. In dieser Hinsicht war bisher wenig publiziert und das Labor von Fernando Nottebohm versprach die besten Vorraussetzungen für eine solche Arbeit. In den letzten 2 Jahren habe ich erfolgreich Screeningverfahren verwendet, um Fragen zum Themenkomplex adulte Neurogenese und neuronaler Lebenszyklus zu beantworten. Dabei kamen Methoden wie: Differential Display, Laser Micro Dissection, Subtrakive Hybridisierung und cDNA Arrays zur Anwendung. Für meine weitere Karriere in Deutschland oder dem benachbarten Ausland möchte ich einige dieser Ergebnisse weiterverfolgen. Dabei aber sicher auch zur Maus oder einem anderen Organismus, an dem mehr Genetik möglich ist, zurückkehren. Weitere Experimente am Vogel sind sicher Erfolg versprechend, auch im Hinblick auf die gerade verfügbar gewordenen Werkzeuge wie einen cDNA Chip vom Zebrafinken, die Publikation des Hühnergenoms etc. Referenzen – Professor Sabine Werner; ETH Zürich – Professor Christian Alzheimer; Universität Kiel – Professor Fernando Nottebohm; The Rockefeller University NY Kontaktadresse Moritz Hertel The Rockefeller University, Box 137 1230 York Avenue New York, NY 10021 Tel: 001 212 327 8381 (6h back in time) Fax: 001 212 327 8312 [email protected] Gehirn und Verhalten Besprochen von Katharina Braun, Otto-von-Guericke Universität, Institut für Biologie, Brenneckestr. 6, 39118 Magdeburg Gehirn und Verhalten ist ein Lehrbuch, in dem auf didaktisch klare und effiziente Weise die vielschichtigen Bezüge zwischen Hirnfunktionen und Verhalten aufgezeigt werden. Es deckt sowohl die Bereiche der Biopsychologie und Verhaltens-/Neurobiologie, als auch die medizinisch-klinischen Bereiche der Neurologie und Psychiatrie und auch die Neuropharmakologie in präNeuroforum 3/04 gnant formulierter, übersichtlicher Weise ab. Hervorzuheben ist die Betonung auf die systemisch-funktionellen Aspekte, die in vielen Lehrbüchern zugunsten der genetisch-molekularen Mechanismen oft etwas zu kurz kommen, und mit dem das Buch eine „Marktlücke“ schließen kann. Als nützliche Orientierungshilfe ist jedem Kapitel ein kurzer Abschnitt voraus- Neueintritte Folgende Kolleginnen und Kollegen dürfen wir als Mitglieder der Neurowissenschaftlichen Gesellschaft begrüßen: Albrecht, Jan Dominik (Magdeburg) Albrecht, Juliane (Mainz) Baeuerle, Peter (Frankfurt/Main) Bikbaev, Dr. Artur F. (Bochum) Conrad, Verena (Stralsund) Doebler, Stefanie (Frankfurt/Main) Fink, Prof. Dr. Heidrun (Berlin) Gastens, Dr. Alexandra (Hannover) Haeusler, Dr. Udo (Königsbach-Stein) Hertel, Dr. Moritz (New York, USA) Jaeger, Philipp (Berlin) Joshi, Illah (Bochum) Leuchtenberger, Dr. Stefanie (Mainz) Moisel, Dr. Marion (Berlin) Neumann, Dr. Nicola (Tübingen) Poell, Florian (Magdeburg) Prange-Kiel, Dr. Janine (Hamburg) Rose, Dr. Michael (Hamburg) Schira, Mark M. (San Francisco, USA) Schmidt, Manuela (Göttingen) Steiner, Michel (München) Tziridis, Konstantin (Tübingen) Utz, Kathrin (Trier) Wesarg, Dr.-Ing. Thomas (Freiburg) Der Mitgliedsstand zum 15. Juli 2004 beträgt 1.662 Mitglieder. gestellt, in dem stichwortartig ein Überblick über die Inhalte des Kapitels gegeben wird. Hiermit, sowie mit den als „Exkurs“ bezeichneten eingeschobenen thematischen Blöcken zur Vertiefung bestimmter Aspekte der im Text ausgeführten Themen, wird jedes Kapitel zu einer Einheit, die auch einzeln gelesen und verstanden werden kann. Schön kompakt und übersichtlich ist das Kapitel, in dem das neurowissenschaftliche Methodenspektrum einschließlich der Datenanalyse sehr übersichtlich dargestellt wird, dies beleuchtet vor allem für die Studenten die technologische Vielfalt in den Neurowissenschaften. Die Kapitel zu den 253 IMPRESSUM / BÜCHER Ausblick Folgende Beiträge werden für die nächsten Ausgaben von Neuroforum vorbereitet: Migrationsstörungen und Epilepsie Carola A. Haas Axogliale Interaktion und Myelinisierung- oder wie ein erster Kuss in Umhüllung resultiert Eva-Maria Krämer und Jacqueline Trotter Experimentell Therapiestrategien akuter Rückenmarksverletzungen – eine intrgrative Perspektive Jan M. Schwab, Klaus Brechtel und Christian-Andreas Müller Impressum Geschäftsführerin: Angelika Lex Herausgeber: Neurowissenschaftliche Gesellschaft e.V. Bankverbindung: Berliner Bank AG, BLZ 100 200 00, Kto.-Nr. 810 505 1800 Homepage: http://nwg.glia.mdc-berlin.de Anzeigen: top-ad Bernd Beutel Hammelbächerstr. 30 69469 Weinheim Tel.: 06201/29092-0, Fax: 06201/29092-20 e-mail: [email protected] Redaktion: Helmut Kettenmann (v.i.S.d.P.) Meino Alexandra Gibson Redaktionsanschrift: Max-Delbrück-Centrum für Molekulare Medizin (MDC) Robert-Rössle-Str. 10, 13092 Berlin Tel./Fax: 030 9406 3133/3819 e-mail: [email protected] Redaktionsgremium: Matthias Bähr, Göttingen Cord-Michael Becker, Erlangen Niels Birbaumer, Tübingen Tobias Bonhoeffer, Martinsried Andreas Draguhn, Heidelberg Ulf Eysel, Bochum Karl Friedrich Fischbach, Freiburg Michael Frotscher, Freiburg Klaus-Peter Hoffmann, Bochum Sigismund Huck, Wien Sigrun Korsching, Köln Georg W. Kreutzberg, Martinsried Hans Werner Müller, Düsseldorf Wolfgang H. Oertel, Marburg Klaus Pawelzik, Bremen Hans-Joachim Pflüger, Berlin Werner J. Schmidt, Tübingen Petra Störig, Düsseldorf Hermann Wagner, Aachen Herbert Zimmermann, Frankfurt/Main Verlag: Elsevier GmbH Spektrum Akademischer Verlag GmbH Slevogtstr. 3-5, 69126 Heidelberg Tel.: 06221/9126-300 Fax: 06221/9126-370 http://www.spektrum-verlag.com 254 Satz: polycom Media Service Brunnenstr. 128, 13355 Berlin Tel.: 030/26484087, Fax: 030/26484088 Druck, Auslieferung, Vertrieb, Abo-Service: Druckhaus Beltz, Herr Herzog Tilsiter Str. 17 69502 Hemsbach Tel.: 06201/703-134, Fax: 06201/703-100 e-mail: [email protected] Titelgestaltung: Eta Friedrich, Berlin Erscheinungsweise viermal im Jahr. Neuroforum ist das Publikationsorgan der Neurowissenschaftlichen Gesellschaft. Bezugspreise (jeweils zzgl. Versandkosten): Einzelheft EUR 25,-; Jahresabonnement Inland Einzelperson EUR 45,-; Jahresabonnement Inland Firmen, Bibliotheken EUR 89,Studentenabonnement EUR 15,- bei Vorlage der Immatrikulationsbescheinigung o.ä. Eine Abonnement-Bestellung kann innerhalb von zwei Wochen schriftlich bei Druckhaus Beltz widerrufen werden. Für das Ausland gelten besondere Tarife. Das Abonnement gilt zunächst für ein Jahr und verlängert sich jeweils um ein weiteres Jahr, falls es nicht spätestens sechs Wochen vor Ablauf gekündigt wird. Bei Nichtlieferung aus Gründen, die nicht vom Verlag zu vertreten sind, besteht kein Anspruch auf Nachlieferung o. Erstattung vorausbezahlter Bezugsgelder. Gerichtsstand, Erfüllungs- u. Zahlungsort ist Heidelberg. Sinnessystemen und den motorischen Systemen sind kompakt und übersichtlich aufgebaut, es fehlt zwar die Vertiefung in die molekularen und physiologischen Details, dafür wird aber im Gegensatz zu vielen anderen Lehrbüchern immer wieder auf die Bezüge zu den Verhaltensleistungen hingewiesen, so daß ein ganzheitliches Verständnis der Hirnfunktionen vermittelt wird. Diese ganzheitliche Darstellung setzt sich auch in den folgenden Kapiteln zu den Themen Gedächtnis, Emotion, Sprache und Pharmakologie etc. fort. Das Kapitel über die Interaktion von Gehirn, Hormonund Immunsystemen würde man sich nicht zuletzt auch aufgrund seiner Aktualität noch umfassender und detaillierter wünschen, denn in den meisten Lehrbüchern wird dieses komplexe Thema doch eher vernachlässigt. Ebenfalls um ein Vielfaches umfangreicher würde man sich den Bereich der Entwicklungsneurobiologie wünschen, und zwar weniger die zellulären, genetisch-molekularen Aspekte betreffend, die in den meisten anderen Lehrbüchern bereits relativ extensiv behandelt werden, sondern der systemisch-funktionelle Aspekt der Entwicklung von Gehirn und Verhalten sollte hier ausführlicher dargestellt werden, denn auch diese Betrachtungsebene kommt in den meisten Lehrbüchern zu kurz. Die vielen, gut gelungenen und klaren Abbildungen erleichtern und vertiefen das Verständnis der im Text erklärten Inhalte ganz erheblich, umso bedauerlicher ist es, dass es sich nur um schwarz-weiß Darstellungen handelt. Im Zuge einer sicherlich irgendwann einmal angestrebten zweiten Auflage sollte unbedingt mit dem Verlag eine Ausstattung mit farbigen Abbildungen ausgehandelt werden, denn auch beim Lernen für Klausuren und Prüfungen dürften die Studenten dem Spruch „das Auge liest bzw. lernt mit“ folgen! Insgesamt ist das Buch klar und übersichtlich gegliedert und repräsentiert eine systemisch-funktionelle Betrachtungsweise von Gehirn und Verhalten, die den Studenten der Psychologie Medizin und Biologie ein ganzheitliches Verständnis der Hirnfunktionen vermittelt. Monika Pritzel, Matthias Brand, Hans J. Markowitsch Gehirn und Verhalten Ein Grundkurs der physiologischen Psychologie Spektrum Akademischer Verlag 1. Aufl. 2003, 496 S., 16 Abb., geb., J 49,95, ISBN 3-8274-0248-4 Neuroforum 3/04 n NWG-NACHRICHTEN Neurowissenschaftliche Gesellschaft ....... Neurowissenschaftliche Gesellschaft e.V. Beitrittserklärung: Ich optiere für folgende 2 Sektionen: Hiermit erkläre ich meinen Beitritt zur Neurowissenschaftlichen Gesellschaft e.V. (bitte ankreuzen) Eintrag in das Mitgliederverzeichnis: Name Vorname Verhaltensneurobiologie Zelluläre Neurobiologie Entwicklungsneurobiologie und Neurogenetik Neuropharmakologie und -toxikologie Systemneurobiologie Molekulare Neurobiologie Klinische Neurowissenschaften Computational Neuroscience Titel Dienstadresse Universität/Institut/Firma Ich bin Student ja nein (Bescheinigung anbei) Jahresbeitrag: (bitte ankreuzen) Straße 50,– D/Jahr ordentliches Mitglied 25,– D/Jahr Studenten, Mitglieder im Ruhestand, Arbeitslose PLZ, Ort Tel./Fax/eMail Privatadresse Straße Überweisung: Bankverbindung: Berliner Bank AG, Blz: 100 200 00, Kto.-Nr.: 810 505 1800 Einzug über VISA-Kreditkarte: Einzug über EUROcard: Kartennummer Exp.Date PLZ, Ort Betrag Tel./Fax Name Unterschrift Datum/Unterschrift des neuen Mitglieds BANKEINZUGSERMÄCHTIGUNG Ich unterstütze den Antrag auf Beitritt zur Neurowissenschaftlichen Gesellschaft e.V.: Ich ermächtige die Neurowissenschaftliche Gesellschaft e.V. von meinem Datum/Unterschrift Konto Nr. Ich unterstütze den Antrag auf Beitritt zur Neurowissenschaftlichen Gesellschaft e.V.: bei der Bank BLZ Datum/Unterschrift einmal jährlich den Mitgliedsbeitrag in Höhe von C einzuziehen Ort, Datum Unterschrift Neurowissenschaftliche Gesellschaft e.V. Meino Alexandra Gibson Max-Delbrück-Centrum für Molekulare Medizin Zelluläre Neurowissenschaften Robert-Rössle-Straße 10 Neuro forum Berlin 3/04 D-13092 Kontoinhaber Anschrift 255 Gö tti ng en 17 -2 0, 20 05 | GE of N the NE UR Ge OB rm IO an LO GY Ne CO uro NF sci ER en EN ce CE So cie ty TT IN Main Speakers Symposia 왘 Fe bru ary GÖ Threshold currents: modulators of neuronal excitability 왘 Amyloid and neurodegeneration 왘 Ion channels and transporters in the cochlea: 왘 Barry Dickson, Vienna, Austria from current to molecule to pathology 왘 Pushing toward the limits of what insects can know: Axon guidance at the Drosophila midline Case studies for comparative cognition 왘 Signals in early neural development 왘 Christopher Miller, Waltham, USA 왘 Brain plasticity and cognition: cellular mechanisms Proteins that move ions across and clinical perspectives 왘 Extracellular matrix molecules in regeneration membranes: our evolving picture and synaptic plasticity (Roger Eckert Lecture) 왘 Efference copies and corollary discharge mechanisms in sensory and mental processing 왘 Hannah Monyer, Heidelberg, Germany 왘 Real time processing vs. variability of neural responses 왘 Plasticity and task-dependence of auditory processing Molecular determinants for 왘 The integrated role of glial cells in the CNS: new synchronous oscillatory network activity methodological approaches 왘 Cellular and molecular control of vertebrate neurogenesis 왘 Bill Newsome, Stanford, USA 왘 Use of two-photon fluorescence microscopy to Parietal cortex and the neural study neuronal calcium in brain slices and in representation of ’value’ the intact brain (Otto Creutzfeld Lecture) 왘 Neuronal injury and infection 왘 Nitric oxide/cyclic nucleotide signalling as regulator of developmental processes and cell motility 왘 Miguel Nicolelis, Durham, USA in the nervous system Computing with neural ensembles 왘 New vistas on insect vision 왘 Genomic and proteomic expression profiling in neural repair 왘 Martin Schwab, Zürich, Schweiz 왘 Brain-computer-interfaces (BCI): neuroprotheses Axonal repair in the adult mammalian for the paralysed central nervous system 왘 Neural mechanisms of visual perception and learning in man and monkey 왘 Amyotrophic Lateral Sclerosis (ALS) and motoneuron Chaired by Prof. Dr. Kerstin Krieglstein disease: From basic molecular and cellular mechanisms to novel clinical applications and Prof. Dr. Herbert Zimmermann 왘 What the nose tells the brain – News and views in olfactory coding 왘 Function of the glial cell line derived neurotrophic factor family in development and disease 왘 Possible mechanisms contributing to memory consolidation during sleep 왘 Comparative insights into genetic and activitydependent mechanisms of CNS development bil du ng r. :D W ern er Zu sc hr att 6 th Ab Me eti ng 30 th 왘 Leonardo Cohen, Bethesda, USA Mechanisms of cortical reorganization underlying recovery of motor function after stroke (Ernst Florey Lecture) er, M ag de bu rg Homepage: http://www.neuro.uni-goettingen.de Organization Stipends http://www.neuro.uni-goettingen.de Neurowissenschaftliche Gesellschaft e. V. Geschäftsstelle Meino Alexandra Gibson Max Delbrück Center for Molecular Medicine Robert Roessle Str. 10 D-13125 Berlin Phone: +49 30 9406 3336 Fax: +49 30 9406 3819 eMail: [email protected] Prof. Dr. Kerstin Krieglstein Georg-August Universität Abt. Anatomie/Neuroanatomie Kreuzbergring 36 D-37075 Göttingen Phone: +49 551 39 7051 Fax: +49 551 39 14016 eMail: [email protected] 256 The German Neuroscience Society provides stipends for young qualified investigators. The deadline for application is August 31, 2004. Please send the application including 왘 short CV copy of the abstract list of publications letter of recommendation from a senior scientist 왘 왘 왘 to the Geschäftsstelle of the Neurowissenschaftliche Gesellschaft e. V. Neuroforum 3/04 Gestaltung: Eta Friedrich, Berlin http://nwg.glia.mdc-berlin.de