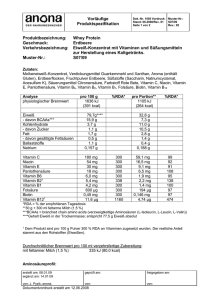
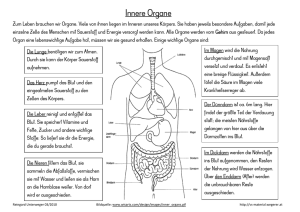
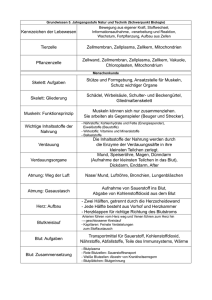
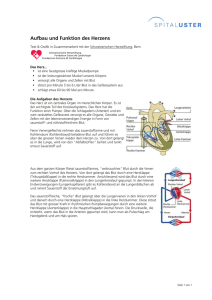
Inhaltsverzeichnis
Werbung

Aus der Medizinischen Tierklinik der Veterinärmedizinischen Fakultät der Universität Leipzig Untersuchungen zum antioxidativen Status bei verschiedenen Formen der Dislocatio abomasi des Rindes im Blut der V. jugularis und der V. epigastrica Inaugural-Dissertation zur Erlangung des Grades eines Doctor medicinae veterinariae (Dr. med. vet.) durch die Veterinärmedizinische Fakultät der Universität Leipzig eingereicht von Gregor Dinges aus Darmstadt Leipzig, 2004 Mit Genehmigung der Veterinärmedizinischen Fakultät der Universität Leipzig Dekan: Herr Prof. Dr. Gotthold Gäbel Betreuer: PD Dr. habil. Manfred Fürll, Medizinische Tierklinik der Veterinärmedizinischen Fakultät Leipzig Gutachter: PD Dr. habil. Manfred Fürll, Medizinische Tierklinik der Veterinärmedizinischen Fakultät Leipzig Prof. Dr. Ute Schnurrbusch, Ambulatorische und Geburtshilfliche Tierklinik der Veterinärmedizinischen Fakultät Leipzig Univ.-Prof. Dr. M. Stangassinger, Institut für Physiologie, Physiologische Chemie und Tierernährung, Lehrstuhl für Physiologie Tierärztliche Fakultät der Ludwig-Maximilian Universität, München Tag der Verteidigung: 10. August 2004 Meinen Eltern Inhaltsverzeichnis Seite 1. Einleitung und Zielstellung 1 2. Literaturübersicht 3 2.1 Die Labmagenverlagerung (Dislocatio abomasi, DA) des Rindes vor dem Hintergrund oxidativer Pathomechanismen 3 2.1.1 Dislocatio abomasi bei Milchkühen 3 2.1.2 Ischämie und Reperfusion 9 2.1.2.1 Entstehung von Radikalen durch Abbau von ATP während lokaler Ischämie/Reperfusion 11 2.1.2.2 Auswirkungen von Ischämie/Reperfusion auf Membranstrukturen 12 2.1.2.3 Aktivierung der körpereigenen Abwehr 13 2.2 Radikale 15 2.2.1 Definition und oxidativer Stress 15 2.2.2 Einteilung der Radikale 16 2.2.2.1 Aktivierte Sauerstoffspezies (ASS) 16 2.2.2.2 Stickoxidradikale (NO.) 17 2.2.2.3 Lipidperoxidation (LPO) 18 2.3 Antioxidantien 20 2.3.1 Definition und Vorkommen 20 2.3.2 Nichtenzymatische Antioxidantien 21 2.3.2.1 Metallbindende Proteine 21 2.3.2.2 Nieder- und mittelmolekulare Antioxidantien 22 2.3.2.2.1 Fettlösliche Antioxidantien 22 2.3.2.2.2 Wasserlösliche Antioxidantien 25 2.3.2.3 TEAC-Konzentration 27 2.3.3 Enzymatische Antioxidantien 28 2.3.3.1 Enzyme zur Glutathionregeneration 28 2.3.3.2 Superoxiddismutasen (SOD) 34 2.3.3.3 Katalasen 38 2.4 Kreatinkinase 38 3. Eigene Untersuchungen: Tiere, Materialien und Methoden 41 3.1 Tiere 41 3.2 Versuchsanordnung, Gruppeneinteilung und Versuchsdurchführung 41 3.3 Material 42 3.3.1 Blutentnahme aus der V. jugularis 42 3.3.2 Blutentnahme am Labmagen 43 3.4 Probenaufbereitung 43 3.4.1 Gewinnung von Serum 43 3.4.2 Gewinnung von Plasma 43 3.4.3 Herstellung von Erythrozytenpellets 44 3.5 Probenbestimmung 44 3.5.1 Klinisch-chemische Untersuchungsmethoden 44 3.5.2 Superoxiddismutase (SOD) 45 3.5.3 Glutathionperoxidase (GPX) 47 3.5.4 Trolox equivalent antioxidative capacity (TEAC) 49 3.5.5 Hämoglobin (Hb) 50 3.6 Statistische Auswertung 50 4. Ergebnisse 52 4.1 Klinisch-chemische Parameter des Fett-, Leber-, Eiweiß- und Mineralstoffwechsels 54 4.2 Parameter im peripheren und abomasalen Blut 55 4.2.1 Vergleich der Chloridkonzentrationen im peripheren und abomasalen Blut 56 4.2.2 Lactatkonzentration 56 4.2.2.1 Lactatkonzentrationen im peripheren Blut 56 4.2.2.2 Lactatkonzentrationen im abomasalen Blut 57 4.2.2.3 Vergleich der Lactatkonzentrationen im peripheren und abomasalen Blut 58 4.2.3 58 Creatinkinase-Aktivität 4.2.3.1 Creatinkinase-Aktivitäten im peripheren Blut (CK(p)) 58 4.2.3.2 Creatinkinase-Aktivitäten im abomasalen Blut (CK(lm)) 59 4.2.3.3 Vergleich der Creatinkinase-Aktivitäten im peripheren und abomasalen Blut 60 4.2.4 60 Superoxiddismutase-Aktivität 4.2.4.1 Superoxiddismutase-Aktivitäten im peripheren Blut (SOD(p)) 60 4.2.4.2 Superoxiddismutase-Aktivitäten im abomasalen Blut (SOD(lm)) 61 4.2.4.3 Vergleich der Superoxiddismutase-Aktivitäten im peripheren und abomasalen Blut 62 Glutathionperoxidase-Aktivität 64 4.2.5 4.2.5.1 Glutathionperoxidase-Aktivitäten im peripheren Blut (GPX(p)) 64 4.2.5.2 Glutathionperoxidase-Aktivitäten im abomasalen Blut (GPX(lm)) 64 4.2.5.3 Vergleich der Glutathionperoxidase-Aktivitäten im peripheren und 4.2.6 abomasalen Blut 65 TEAC-Konzentration 66 4.2.6.1 TEAC-Konzentrationen im peripheren Blut (TEAC(p)) 66 4.2.6.2 TEAC-Konzentrationen im abomasalen Blut (TEAC(lm)) 67 4.2.6.3 Vergleich der TEAC-Konzentrationen im peripheren und abomasalen Blut 68 4.3 TEAC-Konzentrations-Verlauf post operationem 69 4.3.1 TEAC-Konzentrations-Verlauf bei Tieren mit LDA 70 4.3.1.1 Zusammenfassende Darstellung der TEAC-Konzentrationen bei Kühen mit LDA 3 72 4.3.2 TEAC-Konzentrations-Verlauf bei Tieren mit RDA 73 5. Diskussion 78 5.1 Gruppeneinteilung der Tiere 78 5.2 Methode der Blutentnahme am Labmagen 78 5.3 Ausgewählte klinische Befunde bei Kühen mit DA 79 5.4 Messergebnisse und Vergleich 79 5.5 Vergleich der Ergebnisse nach der Herkunft des Blutes 89 5.6 TEAC-Konzentrationsverlauf post operationem 92 6. Zusammenfassung 97 7. Summary 99 8. Literaturverzeichnis Anhang 101 Abkürzungsverzeichnis α alpha Abb. Abbildung ADP Adenosindiphosphat AMP Adenosinmonophosphat Ǻ Ǻngström ASS Aktivierte Sauerstoff Spezies ATP Adenosintriphosphat BHB Beta-Hydroxy-Butyrat Ca2+ Kalziumionen °C Grad Celsius CK Kreatinkinase DA Dislocatio abomasi DGKC Deutsche Gesellschaft für Klinische Chemie g Gramm -GT -Glutamyl-Transpeptidase GPX Glutathionperoxidase Hb Hämoglobin H2O2 Wasserstoffperoxid Ig A Immunglobulin A Ig G Immunglobulin G K+ Kaliumionen l Liter LDA Left Dislocatio Abomasi LMV Labmagenverlagerung μmol Mikromol mmol Milimol Na+ Natriumionen NAD+ / NADH +H+ Nicotinamidadenindinukleotid NADPH Nicotinamidadenindinukleotidphosphat ORAC Oxygen Radical Absorbance Capacity PG Prostaglandin RDA Right Dislocatio Abomasi RDA cT Right Dislocatio Abomasi cum Torsio RDA sT Right Dislocatio Abomasi sine Torsio ROS Reactive Oxygen Species SOD Superoxiddismutase Tab. Tabelle TEAC Trolox Equivalent Antioxidative Capacity TRAP Total Radical-trapping Antioxidant Parameter TROLOX 6-hydroxy-2,5,7,8-tetramethylchroman-2carboxylic acid U Unit V. Vena Einleitung und Zielstellung __________________________________________________________________________________________ 1. Einleitung und Zielstellung Nach wie vor stellt die Dislocatio abomasi (DA) eine der häufigsten nichtinfektiösen Erkrankungen des Rindes in der Frühlaktation dar, die zu großen wirtschaftlichen Verlusten führt (POIKE u. FÜRLL 2000, PEHRSON u. STENGÄRDE 2000). Bisherige Beobachtungen haben gezeigt, dass neben Störungen des Fett- und Energiestoffwechsels auch Perfusionsstörungen am Labmagen und oxidativer Stress das klinische Bild der DA prägen und für die weitere Prognose mit verantwortlich sind (FÜRLL et al. 1999). Untersuchungen gesunder Kühe post partum zum antioxidativen System zeigten: im peripartalen Zeitraum (zwei Wochen a. p. bis zwei Wochen p. p.) befinden sich die TEAC-Konzentrationen bei gesunden Kühen auf dem niedrigsten Niveau und steigen nachfolgend kontinuierlich an (WILKEN u. FÜRLL 2002) zwei Wochen p. p. hat die SOD bei gesunden Kühen ebenfalls die niedrigste Aktivität, vier bis sechs Wochen p. p. steigt sie signifikant an (SATTLER 2001) die CK-Aktivität (Median) steigt von 60 U/l a. p. auf 170 U/l am ersten Tag p. p. und sinkt in den folgenden vier Tagen wieder auf 90 U/l (NAURUSCHAT u. FÜRLL 2002) die GPX-Aktivität bei gesunden Kühen im peripartalen Zeitraum ist abhängig von der Selenversorgung, bei Aktivitäten von >130 U/g Hb kann von einer ausreichenden Versorgung ausgegangen werden (WOLF 1998) Zum Zeitpunkt der Diagnose der Labmagenverlagerung stellt sich das antioxidative System wie folgt dar: die Aktivität der SOD entspricht weitgehend der von gesunden Kühen, sie liegt bei fast 50 % der Kühe unter 7000 U/l Erythrozytenlysat, die Aktivität der GPX beträgt bei 75 % der Kühe 250 – 500 U/mg Hb (SATTLER u. FÜRLL 2002) Kühe mit linksseitiger DA (LDA) weisen niedrigere Aktivitäten der SOD als Kühe mit rechtsseitiger DA (RDA) auf (SATTLER u. FÜRLL 2002) bei Kühen mit Labmagenverlagerung ist die TEAC-Konzentration im Vergleich zu gesunden Kühen niedriger (GOERRES u. FÜRLL 2002) Zum antioxidativen Status nach operativer Rückverlagerung liegen folgende Ergebnisse vor: die Aktivität der SOD im Serum sinkt eine und drei Stunden p. op. gegen die Nachweisgrenze ab (FÜRLL et al. 2004) -1- Einleitung und Zielstellung __________________________________________________________________________________________ bei den LDA steigen die Aktivitäten der SOD im Serum nach 24 Stunden wieder auf den präoperativen Bereich an, bei den RDA dagegen bleiben sie erniedrigt (FÜRLL et al. 2004) Zur Situation am Labmagen selbst wies GEISHAUSER (1995) in in-vitro Untersuchungen während und nach Verlagerung auf Schädigungen der Labmagenmuskulatur hin, die auf die Bildung von Radikalen zurückzuführen sind. Klinisch konnten diese Ergebnisse nur anhand des makroskopischen Bildes intra operationem und anhand peripheren Blutes, das vor der Entnahme bereits die Leber passiert hatte, nachvollzogen werden. Wittek et al. (2004) dagegen konnten in venösem Blut der V. epigastrica dextra bei Kühen mit RDA eine geringere Sauerstoffsättigung und höhere Lactatkonzentrationen als bei Kühen mit LDA nachweisen, die ebenfalls auf Ischämie/Reperfusion hinweisen. Zielstellung der vorliegenden Arbeit ist es, die Auswirkungen der verschiedenen Formen der DA auf den antioxidativen Status der Kühe nicht nur durch Analysen im Blut der V. jugularis, sondern auch im Blut der unmittelbar vom Labmagen wegfließenden V. epigastrica zu beleuchten, um die Auswirkungen der DA am Labmagen selbst besser bewerten zu können. Die abomasale Blutgewinnung erfolgt intra operationem direkt nach Rückverlagerung des Labmagens aus der V. gastroepiploica dextra im Bereich des Pylorus. Zunächst wird über ausgewählte klinisch-chemische Parameter des Fett-, Leber-, Eiweiß- und Mineralstoffwechsels ein Gesamtbild der Kühe mit DA ermittelt. Nachfolgend werden Chlorid- und Lactatkonzentration sowie die Aktivität der Creatinkinase im peripheren mit dem abomasalen Blut verglichen. Zur Einschätzung der oxidativen Stresssituation werden SOD- und GPX-Aktivität sowie die TEAC-Konzentration herangezogen, auch hier erfolgt der Vergleich von peripherem zu abomasalem Blut. Im Anschluss an die operative Rückverlagerung des Labmagens wird der antioxidative Status in Form der TEAC im peripheren Blut über einen Zeitraum von 12 bzw. 16 Stunden postoperativ verfolgt, um den Ablauf der Regeneration von Seiten der nichtenzymatischen Antioxidantien in den ersten Stunden nach der Operation nachzuvollziehen. . -2- Einleitung und Zielstellung __________________________________________________________________________________________ 2. 2.1 Literaturübersicht Die Labmagenverlagerung (Dislocatio abomasi, DA) des Rindes vor dem Hintergrund oxidativer Pathomechanismen 2.1.1 Dislocatio abomasi bei Milchkühen Vorkommen Die Dislocatio abomasi (DA) gilt als eine der häufigsten Erkrankungen von Hochleistungsrindern (GEISHAUSER 1995, REHAGE et al. 1996). Durch Rückgang der Milchleistung, geringere Fruchtbarkeit, hohe Tierarztkosten und Totalverluste kommt es jährlich zu immensen wirtschaftlichen Schäden (FÜRLL et al. 1998). Bei der DA handelt es sich um eine vorwiegend peripartal auftretende Erkrankung von Hochleistungsrindern (FÜRLL u. KRÜGER 1999, WOLF et al. 2001, ZADNIK et al. 2001). In vielen Staaten wird in den letzten Jahren ein vermehrtes Auftreten der DA verzeichnet, unter anderem in den USA und Schweden (PEHRSON u. STENGÄRDE 1998), in Griechenland (KARATZIAS 1998) und der Schweiz (EICHER et al. 1998). Zunächst galten vor allem Tiere zwischen dem vierten und siebten Lebensjahr als gefährdet, mittlerweile sind auch jüngere Tiere vermehrt betroffen (DIRKSEN 1987, KARATZIAS 1998). Im jahreszeitlichen Verlauf treten besonders im Winter und Frühjahr die meisten DA auf (FÜRLL et al. 1997). Ätiologie und Pathogenese Hauptfaktoren für die Entstehung der Labmagenverlagerung sind eine gestörte Labmagenmotorik und eine Gasansammlung im Labmagen (DIRKSEN 1995). Diese „zentralen Auslöser“ der Verlagerung werden selbst aber durch eine Vielzahl von Faktoren beeinflusst bzw. hervorgerufen, so dass man von einem „multifaktoriellen Komplex“ spricht (MARTENS 1998, Tab. 1). Mit zu den Auslösern gehören Störungen des Fettstoffwechsels, die bei Kühen um die Geburt herum schon lange bekannt sind (REID 1980). Durch die verringerte Futteraufnahme in der Endgravidität (etwa zwei Wochen a.p.) und den gesteigerten Energiebedarf in der Frühlaktation steigt die Lipolyse im Fettgewebe, die Leber wird mit einem Anstrom an freien Fettsäuren (FFS) überschwemmt, wobei eine Konzentration an FFS im Plasma >0,65 μmol/l als Zeichen verstärkter Fettmobilisation angesehen wird (BARTLEY 1989). Die FFS können -3- Literatur __________________________________________________________________________________________ sich in den Leberzellen wieder zu Triglyceriden (TG) zusammenlagern, so dass es zur Verfettung der Leberzellen kommt. Dies wird als Steatosis hepatis bezeichnet, sobald mehr als 30 mg TG pro Gramm Leber (Nassgewicht) enthalten sind (OIKAWA et al. 1997). Bei Tieren mit DA wird regelmäßig auch das Lipomobilisationssyndrom festgestellt, wobei Ursache und Wirkung in Bezug auf auslösende Faktoren nicht genau zu trennen sind (FÜRLL et al. 1997). So wiesen Kühe mit DA bei Untersuchungen von REHAGE et al. (1996) zu 32 % eine hochgradige, zu 40 % eine mittelgradige und zu 28 % eine geringgradige oder gar keine Leberverfettung auf. Als Begleiterscheinung ist bei Kühen mit DA und Lipomobilisation eine verringerte Plasmatocopherolkonzentration (Vitamin E) zu finden, die als vermindertes Antioxidans die Lipid-peroxidation im Lebergewebe fördern und so zum Zelltod führen kann (MUDRON et al. 1997). Auch wenn Ursache/Wirkung des Lipomobilisationssyndroms bei der DA nicht genau geklärt werden können, so stellt das Lipomobilisationssyndrom einen der limitierenden Faktoren bei der Regeneration der Tiere dar (REHAGE et al. 1996). Hochleistungskühe im peripartalen Zeitraum gelten aufgrund der hohen Milchproduktion grundsätzlich als gefährdet, in hypocalcämische Zustände zu geraten (Hypocalcämie: < 1,7 mmol/l Gesamtcalcium). Da diese grenzwertigen Calciumkonzentrationen nicht nur Auswirkungen auf die Skelettmuskulatur haben, sondern auch auf die Motilität des Magendarmkanals, gelten sie per se schon als Risikofaktor für die DA (KÜMPER 1995 a, GEISHAUSER et al. 1996 b, ZADNIK et al. 2001). Während derselben liegen z.T. hypocalcämische Zustände vor (WARD et al. 1994). Tab. 1: Mögliche Ursachen der Dislocatio abomasi (Dirksen 1995) Gestörte Labmagenmotorik Hypotonie Hohe Dilatation Histaminfreisetzung Gasansammlung Transportbehinderung Konzentration Adrenalin- -“- durch gestörte abomasaler FFS Duodenale Säuerung Labmagenmotorik Hypocalcämie Prostaglandin E2 Hyperketonämie Vergrößerter Gestörte Entleerung via Abomasus-Kanal Hyperinsulinämie Labmagen im Alter Erhöhte Gasproduktion Hypergastrinämie und Alkalämie Futtermitteln bei Endotoxämie besonderen Gestörte Absorption bzw. Diffusion im Labmagen -4- Literatur __________________________________________________________________________________________ In Untersuchungen zur Motorik und dem Entleerungsmechanismus des Labmagens über myogen und nerval vermittelte mechanische Aktivitäten der Labmagenmuskulatur sowie den Plexus myentericus konnte GEISHAUSER (1995) zeigen, dass bei Kühen mit DA eine Fehlfunktion im Antrum pylori vorliegt, die zur gestörten Labmagenaktivität führt. Als mögliche Ursachen nennt er hierfür eine gestörte Produktion und Freisetzung von Neurotransmittern (Noradrenalin) sowie eine vermehrte Stickstoffmonoxidbildung, die die Muskelkontraktion hemmen. Die Rolle des Stickoxids wird auch in weiterführenden in-vitro Untersuchungen von REICHE et al. (1998) deutlich, da es diesen gelang, durch Hemmung der Stickoxid-Synthase (NOS) eine höhere cholinerge kontraktile Aktivität der Labmagenmuskulatur von Kühen mit DA hervorzurufen. Zusätzlich lag in-vitro in den Muskelzellen eine verringerte Sensibilität gegenüber Acetylcholin (ACH) vor, die dazu führt, dass erst bei erhöhten ACH-Konzentrationen die Aktivierung erfolgt. Zusätzliche Erkrankungen können die Entstehung der DA begünstigen. Hierzu zählen Erkrankungen des Genitale, wie sie nach Geburten auftreten können, Erkrankungen des Euters, der Klauen und Gelenke sowie Pneumonien und parasitäre Erkrankungen (FÜRLL et al. 1998). Nachteilig können sich zudem die Fütterung und die Änderung der Haltungsbedingungen post partum mit dem Einstieg in die Milchproduktion auswirken (MARTENS 1998). Klinisches Bild der Labmagenverlagerung Der Labmagen liegt bei Rindern physiologisch ventral mit dem Fundus auf der Bauchwand und erstreckt sich mit dem Antrum nach dorsal, etwa bis in Höhe der rechten Hungergrube. Aus dieser Position heraus kann er sich nach links (LDA) und nach rechts (RDA) verlagern. Die klinische Diagnose der Labmagenverlagerung erfolgt hauptsächlich durch die Perkussionsauskultation, bei der aufgrund der Gasansammlung das typische „Klingeln“ erkannt werden kann. Eine Feldstudie an Holstein-Friesian-Kühen zeigte, dass 75 % aller Labmagenverlagerungen nach links (LDA) und 25 % nach rechts (RDA) erfolgen (MARTIG 1996, WOLF et al. 2001). Die linksseitige Verlagerung wird präoperativ in drei Grade eingeteilt (LDA 1-3), wobei der dritte Grad der größten Ausdehnung im Perkussionsfeld entspricht. Sie kann über längere Zeit und intermittierend („Pendellabmagen“) auftreten, da noch Ingesta den Labmagen passieren können. Die Tiere magern mit der Zeit ab und zeigen einen deutlichen Leistungsabfall. Bei der rechtsseitigen Verlagerung handelt es sich um ein akutes und lebensbedrohliches Geschehen, da zusätzlich zur Aufgasung (RDA sT) noch eine Drehung (RDA cT) erfolgen kann, die den Zu- und Abfluss des Blutes massiv einschränkt. -5- Literatur __________________________________________________________________________________________ Eine Drehung erfolgt in 99 % aller Fälle nach links und unterbindet die Passage der Ingesta ab einer Drehung um mehr als 180° vollständig (KÜMPER 1995 b). Die Tiere zeigen in der Regel am ersten Tag Inappetenz und Leistungsabfall, am zweiten Tag Apathie und blutigen Kot (Melaena). Spätestens am dritten Tag liegen sie fest und können nachfolgend verenden (FÜRLL et al. 1999). Der blutige Kot entsteht dabei durch Ulzerationen des Labmagens, die dort aufgrund der verringerten Motorik durch die Enzyme des Labmagens selbst verursacht werden (GEISHAUSER et al. 1996 b). Die verringerte Motorik des Labmagens kann bis zur Atonie des Labmagens führen, als Folge werden die Ingesta nicht weiterbefördert, es kommt zur Labmagenobstipation (REICHE et al. 1998). Stoffwechselveränderungen bei Kühen mit DA Da die DA häufig im peripartalen Zeitraum auftritt, muss bei veränderten Stoffwechselparametern darauf geachtet werden, dass diese nicht nur durch die DA verändert werden, sondern auch Nachwirkungen der vorangegangenen Geburt sein können. KLEISER u. FÜRLL (1998) untersuchten das Blut von Kühen mit DA am dritten Tag post partum. Dabei zeigte sich, dass nur bei den Parametern β-Hydroxy-Butyrat (BHB), Cholesterol, freie Fettsäuen (FFS), Bilirubin, Aspartat-Amino-Transferase (ASAT), Creatinkinase (CK) sowie den Leukozyten gesicherte Unterschiede zur Kontrollgruppe auftraten (Tab. 2). Tab. 2: Stoffwechselparameter mit gesichertem Unterschied zur Kontrollgruppe bei Kühen mit DA am dritten Tag p. p. (modifiziert nach KLEISER u. FÜRLL 1998) Parameter Einheit Referenzbereich BHB mmol/l < 1,0 Cholesterol mmol/l > 2,0 FFS μmol/l < 2,0 Bilirubin μmol/l < 6,0 ASAT U/l < 100 CK U/l < 200 Leukozyten G/l > 4,0 Veränderung bei DA Die DA wird in der Regel von einer Ketose begleitet. Die erhöhte Konzentration an Ketonkörpern wie dem β-Hydroxy-Butyrat ist Folge des Energiedefizits, das durch die Frühlak-tation hervorgerufen wird und durch eine erhöhte Lipomobilisation gekennzeichnet -6- Literatur __________________________________________________________________________________________ ist (STÖBER u. DIRKSEN 1982). Weitere Belastungen des Fettstoffwechsels machen die Parameter FFS und Cholesterol deutlich, die Beeinträchtigung der Leberfunktion zeigt sich in erhöhten Enzymaktivitäten und erhöhter Bilirubinkonzentration. Bei Tieren mit RDA konnte MEYLAN (1999) zudem gesichert verringerte Chloridkonzentrationen im Blut im Vergleich zu LDA nachweisen. Aus ihrer Sicht machen Herzfrequenzen > 90/min, ein Serum-Harnstoff 10 mmol/l, eine Kaliumkonzentration 3,3 mmol/l und eine Chloridkonzentration 85 mmol/l eine weitere wirtschaftliche Nutzung der Kühe auch nach überstandener Verlagerung unwahrscheinlich, was aber vielfach bezweifelt wird. Zusätzlich postuliert sie eine präoperative Unterscheidung zwischen rechtsseitiger Verlagerung ohne und mit Torsion über eine im Falle der Verlagerung mit Torsion deutlich höhere γ-GT. In ihren Untersuchungen überlebten 81,2 % der Tiere eine rechtsseitige Verlagerung ohne Torsion und 67,3 % der Tiere eine Verlagerung mit Torsion. Auch der Säure-Base-Haushalt zeigt Veränderungen während der DA. Tiere mit DA weisen im Blut eine Alkalose auf, wenn diese bei den Tieren mit RDA auch nur im Anfangsstadium besteht. Damit einher geht der erhöhte Basenüberschuss (> 2 mmol/l) und ein zu niedriger pH-Wert im Harn (< 7) (FÜRLL et al. 1999). GEISHAUSER et al. (2000) weisen in Herden mit DA auf die Erhöhung des MilchfettMilchprotein-Quotienten (FEQ) hin. Stoffwechseldiagnostik im Labmagensaft Untersuchungen an Milchkühen zum pH-Wert im Labmagensaft, der physiologischerweise einen Wert von 2 besitzt, zeigen in den ersten drei Wochen post partum einen höheren Wert als in der Mitte der Laktation. Der pH-Wert erreicht seinen Höhepunkt etwa zwei Wochen post partum und fällt nachfolgend wieder ab. Diese Erhöhung des pH-Wertes ermöglicht ein abnormes Bakterienwachstum im Labmagen, was zu gesteigerter Gasbildung führen und somit Aufgasung und Verlagerung des Labmagens fördern kann (VAN WINDEN et al. 2002). Um die Vorgänge während einer DA am Labmagen direkt beleuchten zu können, haben GEISHAUSER et al. (1996 a, b) den Labmagensaft präoperativ sowie einen und drei Tage postoperativ untersucht. Im Labmagen steigt bei einer DA die Chloridkonzentration, während die Kalium-, Magnesium-, Calcium- und Phosphatkonzentrationen absinken. Präoperativ sinkt der pH-Wert bei linksseitigen Verlagerungen, während er bei den rechtsseitigen DA mit Torsion unverändert bleibt. Im Labmagen sind die Natrium- und Kaliumkonzentration bei linksseitigen DA vor der Operation geringer als die der rechtsseitigen. Die Konzentrationen von Magnesium, Calcium und Phosphat verhalten sich umgekehrt. Nach der Operation sinkt -7- Literatur __________________________________________________________________________________________ die Natriumkonzentration, während die Kalium-, Magnesium-, Calcium- und Phosphatkonzentrationen wieder ansteigen. Die intraabomasale Chloridkonzentration sinkt bei den linksseitigen DA nach der Operation, während sie bei den rechtsseitigen zunächst weiter ansteigt. Am dritten Tag nach der Operation sind die Konzentrationen an Kalium, Magnesium, Calcium und Phosphat noch etwas erniedrigt, ebenso der pH-Wert, während die Chloridkonzentration nach wie vor erhöht ist. Diese Veränderungen sind über die geringere Futteraufnahme der Tiere mit Labmagenverlagerung zu erklären. Das Futter bleibt länger im Pansen bzw. Labmagen und kann erst nach der Operation in den Labmagen bzw. Dünndarm gelangen. Bei den rechtsseitigen DA mit Torsion ist zusätzlich noch ein im Vergleich zu den linksseitigen Verlagerungen erhöhter Reflux aus dem Labmagen zu beachten, der für eine spätere Normalisierung der Zustände sorgt. Dazu kommt eine gesteigerte Sekretion der Labmagendrüsen vor der Operation. Da die Veränderungen bis zum dritten Tag nach der Operation andauern, ist bis dahin auf eine gestörte Sekretion und Motilität des Labmagens zu schließen (GEISHAUSER et al. 1996 b, DELGADO-LECAROZ et al. 2000). Veränderungen der Labmagenwand Die verringerte Motorik und Gasansammlung im Labmagen ist die Voraussetzung für die DA (CONSTABLE et al. 1992). Dies gab Anlass, die glatte Muskulatur des Labmagens genauer zu untersuchen, da diese für die Motorik des Labmagens verantwortlich ist. Dabei konnten hinsichtlich der Spontanaktivität bei gesunden Tieren und deren Zuordnung zu bestimmten Rassen keine Rückschlüsse auf spätere DA gewonnen werden, da die Spontanaktivität keinem gleichbleibenden Muster folgt (ZULAUF et al. 2002). REICHE et al. (1998) untersuchten die Motilität der glatten Muskulatur des Labmagens in Nähe des Pylorus in vitro bei gesunden und bei Kühen mit DA. Dabei stellten sie bei Kühen mit DA verminderte Aktivitäten der Muskulatur fest, die durch Hemmung der Stickoxidsynthase erhöht werden konnten. Daraus schlossen sie auf Beteiligung von Stickoxidradikalen. Erhöhte Konzentrationen von Stickoxidradikalen konnten auch schon bei anderen Motilitätsstörungen im Darm, wie dem Ileus nachgewiesen werden (DE WINTER et al. 1997). Die Quelle der Stickoxidradikale ist bislang noch ungeklärt, als mögliche Bildungsstätten werden Nervenzellen (Neurotransmitter) (AL GOUHL et al. 1995), Zellen des lokalen Immunsystems (SALZMANN 1995) sowie die Muskelzellen selbst angesehen (MURTHY et al. 1993). Um die Konzentration der Stickoxidradikale bei Kühen zu minimieren, wird versucht, deren mögliche Beseitigung durch das antioxidative System näher zu erforschen (FÜRLL et al. 1999). -8- Literatur __________________________________________________________________________________________ Therapie Bei der Therapie der Labmagenverlagerung steht die Rückverlagerung im Vordergrund. Diese kann konservativ über Wälzen und invasiv über einen sog. „blind stitch“ (TITHOF u. REBHUN 1986), über eine Laparatomie von links („Utrechter Methode“), über eine Laparotomie von rechts nach DIRKSEN (BÜCKNER 1993, STEINER 1996, BECVAR et al. 2001) sowie endoskopisch (JANOWITZ 1998) erfolgen. Postoperativ gilt es, den Kreislauf mittels Infusionen (0,9%ige NaCl-Lösung + Glucose) zu stabilisieren, weiteren Entzündungserscheinungen vorzubeugen (Antibiose + Antiphlogistika), die Verdauung zu fördern (Pansenstimulans) sowie Imbalancen des Elektrolyt-, des SäureBasen-Haushaltes und des Fettstoffwechsels auszugleichen (FÜRLL et al. 1999). 2.1.2 Ischämie und Reperfusion Unter Ischämie (griech.: „ischein“-zurückhalten; „haima“-Blut) versteht man eine verminderte Durchblutung in einem Organ oder einem Körperteil, die durch einen Gefäßverschluss hervorgerufen wird. Während der Ischämie kommt es zur Unterversorgung mit Sauerstoff (Hypoxie) und Nährstoffen im betroffenen Gebiet, bei vollständiger Unterbindung sogar zur Anoxie (TILL 1999). Der Hauptenergielieferant des Gewebes, ATP, kann nicht mehr regeneriert werden. Kann die Atmungskette aufgrund der Hypoxie nicht mehr ausreichend ablaufen, bleiben nur die anaerobe Glykolyse und die Verschiebung des Creatinkinase- und des Adenylatkinase-Gleichgewichtes (siehe Abb. 1) mit einer weitaus geringeren Regeneration von ATP. So kommt es in den betroffenen Arealen auf längere Sicht zum Energiemangel mit Ansammlung von Lactat und AMP. Das Lactat kann diagnostisch mit als Hinweis für das Vorliegen einer Ischämie bei Kühen mit RDA cT genutzt werden (WITTEK et al. 2004). Der Anstieg von AMP und Lactat wird in der letzten Zeit als der Beginn der nachfolgenden Schädigungen im Ischämie-/Reperfusionssyndrom angesehen, da beim Abbau desselben Radikale und weitere zellschädigende Vorgänge in Gang gesetzt (ÖHLENSCHLÄGER 1993 a). Creatinkinase (1) Creatinphosphat + ADP + H + ATP + Creatin Adenylatkinase (2) 2 ADP ATP + AMP -9- werden Literatur __________________________________________________________________________________________ Abb. 1: ATP-Regeneration unter sauerstoffgesättigten (1) und hypoxischen (2) Bedingungen (modifiziert nach ÖHLENSCHLÄGER 1993 a) Vom Energiemangel sind alle energieverbrauchenden Prozesse betroffen. Auf zellulärer Ebene sind besonders die Ionenpumpen (z.B. Na+-K+-Pumpe, Ca2+-Pumpe und Na+-Ca2+Austauscher) zu nennen, die zu Imbalancen des osmotischen Gleichgewichtes von Intra- und Extrazellulärraum führen (CHAUDRY 1983). Intrazellulär kann daraus ein pH-Wert-Abfall um fast eine Einheit folgen (LAIPPLE 2000), insgesamt wird das Membranpotential verringert (KEELER et al. 1966, LEAF 1973). So werden auch Struktur und Funktion der semipermeablen Zellmembran gestört, was zum Austritt von negativ geladenen Proteinen aus dem Zellinneren führen kann. Letztendliche Folge der Ischämie ist der Zelltod durch Versagen der einzelnen Organellen, Strukturverlust der Membranen und Zellödem durch Wassereinstrom, was als hydropische Degeneration bezeichnet wird (LEAF 1973, LAMBOTTE 1977, LEHMANN et al. 1996). Daraus können Störungen der Mikrozirkulation, Organversagen aufgrund zellulären Unterganges und Gesamttod des Organismus resultieren. Die Reperfusion ischämischer Bereiche oder Organe kann zu weiteren Schäden führen. Die ersten Sekunden nach Beseitigung des Ischämieverursachers (z.B. Volvulus von Darmabschnitten) erfolgt eine vollständige Durchblutung wie vor der Ischämie. Diese Durchblutung („intestinal blood flow, IBF“) verringert sich nachfolgend zunehmend („noreflow phenomenon“) (REES et al. 1994, AKGÜR et al. 1996), denn der positiven Reoxygenierung stehen mehrere negative Effekte gegenüber. So erhalten die hyperosmotischen Zellen Wasser, was, durch die gestörte Membranfunktion noch zusätzlich begünstigt, zum Platzen der Zellen führen kann (LEAF 1973). Zellen und Organellen, die auf anaerobe Glykolyse eingestellt sind, erhalten wieder Sauerstoff. Der pH-Wert steigt binnen kurzer Zeit wieder auf das Ausgangsniveau. Wird der pH-Wert dagegen nur langsam nach oben korrigiert, liegt die Überlebensrate der Zellen weitaus höher. Die während der Ischämie entstandenen Metaboliten von AMP und dabei freigewordene Radikale werden im Organ bzw. Organismus gestreut und rufen Reaktionen des gesamten Körpers hervor (COGHLAN et al. 1991, SCHOENBERG u. BEGER 1993). Bezeichnenderweise folgt auf die Phase der Ischämie eine vom Darm selbst ausgehende Minderdurchblutung, durch die der Körper selbst Reperfusionsschäden vermindert (WILKINS et al. 1994). - 10 - Literatur __________________________________________________________________________________________ In vitro Untersuchungen am Mäusedarm (BIELEFELDT u. CONKLIN 1997) zur Motilität vor, während und nach Ischämie/Reperfusion zeigten, dass die Motilität während der Hypoxie abnimmt, nach 15-minütiger Ischämie noch Reize beantwortet werden und nach einer Stunde keine Reaktion mehr erfolgt. In der frühen Phase der Reperfusion werden Reize wieder beantwortet, mit zunehmender Dauer der Reperfusion allerdings mit abnehmender Tendenz. Gingen der Reperfusion 15 Minuten Ischämie voraus, erreichte die Motilität wieder 100 %, nach einstündiger Ischämie allerdings nur noch 47 % der Ausgangsmotilität. Bei Rindern mit DA war bislang der Grad einer eventuellen Ischämie nur intra operationem anhand des makroskopischen Bildes des Labmagens abzuschätzen. Als Hilfsmittel dienten dabei die Stauung der Blutgefäße, die zyanotische Färbung sowie die Spannung der Labmagenwand durch die Gasansammlung. Wurden Kühe später seziert, konnten anhand der Histologie genaue Aussagen über Ischämie- bzw. Reperfusionsschäden getroffen werden (AUPPERLE et al. 1998). WITTEK u. FÜRLL (2002) gelang es erstmals, anhand des abomasalen Gasdrucks und -volumens sowie der Sauerstoffsättigung am verlagerten Labmagen im Vergleich zur Sauerstoffsättigung am dorsalen Blindsack des Pansens Aussagen über die Per-fusion des verlagerten Labmagens am lebenden Tier zu treffen. Bei allen Kühen mit DA waren die Gasmenge sowie der Gasdruck positiv miteinander korreliert. Bei den Kühen mit LDA konnten keine Korrelationen zwischen Gasdruck oder -volumen und der Sauerstoffsättigung am Labmagen festgestellt werden. Die Sauerstoffsättigung am Labmagen war geringer als am dorsalen Pansenblindsack. Dies bedeutet, dass im Falle einer LDA nur geringe Perfusionsstörungen vorliegen. Bei den Tieren mit RDA konnten negative Korrelationen zwischen Gasdruck oder –volumen und Sauerstoffsättigung am Labmagen festgestellt werden. Die Sauerstoffsättigung am Labmagen unterschied sich signifikant von der des Pansenblindsackes. Dies lässt auf deutliche Perfusionsstörungen bei Tieren mit RDA schließen, die nach der Rückverlagerung zu Reperfusionsschäden und Labmagenatonie führen können. 2.1.2.1 Entstehung von Radikalen durch Abbau von ATP während lokaler Ischämie/Reperfusion Adenosinmono- (AMP) und Adenosindiphosphat (ADP) werden, wenn sie nicht zu ATP regeneriert werden können, in den ischämischen Bezirken abgebaut. Dabei entsteht aus den Purinanteilen Hypoxanthin. Unter sauerstoffgesättigten Bedingungen erfolgt der weitere Abbau durch die NAD+-abhängige Xanthindehydrogenase, die „überschüssige“ Elektronen auf NAD+ überträgt. Unter hypoxischen Bedingen wird das Enzym unter Ca2+-Einfluss zur - 11 - Literatur __________________________________________________________________________________________ Xanthinoxidase transformiert, wobei Sulfhydryl-Gruppen reversibel oxidiert werden und/oder irreversible proteolytische Spaltungen vorgenommen werden. Dies hat zur Folge, dass „überschüssige“ Elektronen nicht mehr auf NAD+ übertragen werden, sondern auf Sauerstoff, der in der Phase der Reperfusion wieder ausreichend vorhanden ist. Dabei entsteht das sehr reaktive Superoxid (GRANGER et al. 1981, BULKLEY 1987) (Abb. 2, Abb. 8). Ort dieser Transformation der Xanthindehydrogenase stellen vor allem Endothelzellen dar, da das Enzym hier durch Glycosaminoglycane gebunden wird. Demzufolge sind auch an den Gefäßwänden die größten Schäden zu erwarten (NAKAZAWA et al. 1996). Welchen Einfluss Hypoxanthin und die Xanthinoxidase auf die nachfolgend gesteigerte Permeabilität haben, geht aus Untersuchungen von GRANGER et al. (1986) hervor. Es wurde gezeigt, dass Infusionen von Hypoxanthin und Xanthinoxidase die gleiche Permeabilitätserhöhung im nicht-ischämischen Darm hervorrufen wie das komplette Ischämie-/Reperfusionsgeschehen. Pathomechanismen des Allein diese Infusionen reichen aus, die Ischämie-/Reperfusionssyndroms in Gang zu setzen (JOHNSON et al. 1981, GROGAARD et al. 1982). MURPHY et al. (1997) zeigten in vivo am Rattendarm, dass eine Ischämiezeit über sechzig Minuten zu einer Steigerung der Aktivität der Xanthinoxidase und zu einer Erhöhung der Gefäß- und Schleimhautpermeabilität führten. Die für die Schadwirkung verantwortliche Umwandlung der Xanthinoxidase findet allerdings nur in den Endothelzellen des Darmes statt, die Mukosazellen spielen nur eine untergeordnete Rolle (SCHILLER et al. 1993). Dies wird durch die Tatsache gestützt, dass die gastrointestinale Schleimhaut von einer Mucinschicht überzogen wird, die sie einerseits vor Oxidantien aus dem Lumen schützt, andererseits aber auch entstehende Radikale aus den oberen Schleimhautschichten beseitigen kann (PARKS 1989). Bei Pferden mit Kolonverlagerung und Volvulus konnte festgestellt werden, dass auftretende Ischämie-/Reperfusionsschäden nicht durch die Xanthinoxidase hervorgerufen werden, da kein Anstieg der Aktivität dieses Enzyms erfolgte. Dementsprechend blieb auch die Behandlung mit Allopurinol erfolglos. In Bezug auf die Gabe von Vitamin E, GPX und Selen bestand zwischen Versuchs- und Kontrolltieren kein Unterschied. In Untersuchungen an Hasen mit Ischämie-/Reperfusionssyndrom am Darm mit nachfolgender Behandlung mit Vitamin C, Vitamin E, Prednisolon und Mannitol zeigte sich dagegen, dass Mannitol und Vitamin C im Gegensatz zu Prednisolon und Vitamin E die Schleimhautschäden verringern, dass Vitamin C und Mannitol zu geringeren Konzentrationen an MDA führen und dass Vitamin E, Vitamin C und Mannitol eine höhere Konzentration an GSH hervorrufen (GÜNEL et al. 1998). - 12 - Literatur __________________________________________________________________________________________ 2.1.2.2. Auswirkungen von Ischämie/Reperfusion auf Membranstrukuren Während der Ischämie und der frühen Reperfusion kommt es zu Veränderungen der Membranstrukturen. Die semipermeablen Membranen verlieren die Asymmetrie und werden undicht. Dadurch wird der ungehinderte Austausch zwischen dem Extrazellulärraum und dem Zytoplasma möglich (LAMBOTTE 1977). Mögliche Ursachen hierfür sind die gestörte Neusynthese von Membranlipiden oder deren Umverteilung aufgrund der verminderten Aktivität der ATP-abhängigen Translokase. Schließlich verliert das Zytoskelett unter anderem durch den Wassereinstrom als physikalische Kraft seine Verbindung zu den Membranen. Dies führt besonders bei kontraktilen Zellen während der Kontraktion zu weiteren Schädigungen (CORBUCCI et al. 1985). Die betroffenen Zellen tragen auch über die Aktivierung des Prostaglandinsystems selbst zu ihrer Beseitigung bei. Durch endogene Phospholipasen werden bei Membranschäden Vorstufen der Prostaglandine aus Membranlipiden und deren Bestandteilen wie der Arachidonsäure freigesetzt. Die Arachidonsäure ist dabei eines der wichtigsten Ausgangssubstrate für die Prostaglandin- (PG), Thromboxan- und Leukotriensynthese (LT). Über Cyclooxigenase-katalysierte Reaktionen werden PG E2, PG F2, Thromboxan A2 und PG I2 gebildet, die die Hauptfaktoren der weiteren Entzündungsreaktion sind. So gelten PG F und Thromboxan als Zeichen für eine Leukozyteninfiltration im Gewebe (MURPHY et al. 1997). PAPPARELLA et al. (1997) stellten beim Schwein einen Anstieg der Prostaglandinkonzentration während der Reperfusion des Darmes fest. Bemerkenswert ist, dass ab einer bestimmten Ischämiedauer keine Prostaglandinkonzentration in Bezug auf die Dauer der quantitative Zunahme der Ischämie erfolgt (BOROS et al. 1991). Besonders betroffen sind Endothelzellen im Gebiet der Ischämie/Reperfusion. Alle weiteren Schädigungen von Geweben und Organen nehmen hier ihren Anfang (LAIPPLE 2000), können zum Zelluntergang sowie zu ausgedehnten Nekrosen großer Organgebiete führen (LUTHER et al. 1996), von denen auch der Labmagen bei einer Verlagerung betroffen sein kann. Die für den Magen-Darm-Kanal wichtigsten Folgen einer Ischämie mit nachfolgender Reperfusion bestehen in der Freisetzung von lysosomalen Hydrolasen, die zu einer gesteigerten Gefäßpermeabilität (SCHILLER et al. 1993) und zu Schleimhautschäden („gastric stress ulcers“) führen (YOSHIDA et al. 1998). Diese können auch bei Rindern mit Labmagenverlagerung auftreten und klinisch über die veränderten Faeces und einen Anstieg von Gastrin im Plasma diagnostiziert werden (OK et al. 2001). - 13 - Literatur __________________________________________________________________________________________ Weitere Schädigungen der Schleimhaut im Magen-Darm-Kanal erfolgen durch eine erhöhte Sekretion von Albumin ins Lumen, die mit der Dauer der Ischämie positiv korreliert ist (GROGAARD et al. 1982). 2.1.2.3 Aktivierung der körpereigenen Abwehr Die Aktivierung der körpereigenen Abwehr im Rahmen von Ischämie und Reperfusion erfolgt unter anderem über das Komplementsystem. Durch die Veränderungen der Zellen im betroffenen Areal wird direkt der Faktor C 3b aktiviert. Daraufhin läuft die gesamte Komplementkaskade ab. Besondere Bedeutung kommt hierbei den Faktoren C 3 und C 5a zu. Durch sie werden „vasoaktive Mediatoren“ zur Erweiterung der Blutgefäße freigesetzt. Dabei werden sie durch Histamin aus gewebsgebundenen Mastzellen und Leukozyten unterstützt. BOROS et al. (1989) zeigten, dass in der Reperfusionsphase ein Anstieg des Histamins erfolgt, den sie auf die zunehmende Freisetzung von Sauerstoffradikalen zurückführen. Der Komplementfaktor C 5a erhöht zudem die Radikalbildung durch neutrophile Granulozyten und Gewebsmastzellen, wenn diese durch Antikörper wie Ig G und Ig A stimuliert werden (WARREN et al. 1987). Die Komplementfaktoren wirken weiterhin chemotaktisch auf polymorphkernige Granulozyten (PMN) und Makrophagen, die lytische Enzyme enthalten. Viele dieser Enzyme besitzen eine hohe Aktivität zur Bildung von H2O2, was in der Phase der Reperfusion mittels Sauerstoff und der HABER-WEISS-Reaktion (Abb. 4) zur Bildung von hochreaktiven Radikalen führt (WARREN et al. 1987, ALLISON et al. 1988). Die Enzyme und mit ihnen die Radikale werden freigesetzt, wenn die Granulozyten an den einverleibten Zellteilen zugrunde gehen („frustrane Phagozytose“) (JOHNSTON u. LEHMEYER 1977, JOHNSON et al. 1981) (siehe Abb. 2). Die Granulozyten sezernieren auch das Interleukin 1 (IL-1) und das Leukotrien B4 (LT-B4). Das Interleukin 1 führt neben der Stimulierung der Produktion von „Akute-Phase-Proteinen“ zu einer Steigerung der B- und T-Zellproliferation. Die B-Zellen sind für die Produktion von Immungloulinen verantwortlich. Dabei stimulieren Ig A und Ig G Makrophagen zur Bildung und Freisetzung von O2- und H2O2. Studien haben belegt, dass Ig G sowohl Neutrophile als auch Gewebsmakrophagen zur Radikalfreisetzung stimuliert, Ig A jedoch nur die Gewebsmakrophagen (WARREN et al. 1987). Das Leukotrien B4, das durch Aktivierung der Phospholipase A2 freigesetzt wird (ZIMMERMANN et al. 1990), verursacht Adhäsion und Chemotaxis von Leukozyten und fördert deren Aggregation, Enzymfreisetzung und die Radikalproduktion (PARKS et al. 1983, ZIMMERMANN et al. 1990, LEHR et al. 1991). Die Leukozyten „rollen“ dabei nach Ausbildung von besonderen Adhäsionsmolekülen am Endothel entlang, werden fest gebunden und emigrieren ins Gewebe - 14 - Literatur __________________________________________________________________________________________ hinein (DEL MAESTRO 1982, GRANGER et al. 1986). Heften sich die Leukozyten am Endothel im Endstromgebiet fest, so verhindern sie nachfolgend die Passage von Erythrozyten und damit die Reoxygenierung (LAIPPLE 2000). Dies ist als Teil des „noreflow“-Phänomens zu sehen (MENGER et al. 1992). Mastzellen und ihre Degranulation spielen im Ischämie-/Reperfusionssyndrom nur eine untergeordnete Rolle (SZABO et al. 1997). I S ATP C ADP H AMP Ä Adenosin M Inosin Xanthindehydrogenase Komplementkaskade (C5a) Oxidation von SH-Gruppen und/oder proteolytische Spaltung PMN Chemotaxis O2- LT B4 Xanthinoxidase I O2- + H2O2 + Xanthin + Harnsäure Hypoxanthin E R E P E R F U S I O N Abb. 2: Ischämie-/Reperfusionssyndrom in Hinblick auf ATP-Abbau und Aktivierung der körpereigenen Abwehr (modifiziert nach ENGELHART 1999) 2.2 Radikale 2.2.1 Definition und oxidativer Stress Elektronen befinden sich auf Orbitalen um den Atomkern herum. Ein Orbital kann maximal zwei Elektronen, ein Elektronenpaar, beinhalten. Befindet sich nur ein Elektron auf einer solchen Orbitalbahn, spricht man von einem ungepaarten Elektron, bezogen auf ein gesamtes Atom oder Molekül, von einem Radikal. Dabei können in einem solchen auch mehrere ungepaarte Elektronen vorliegen (HALLIWELL et al. 1992, SCHILLER et al. 1993). - 15 - Literatur __________________________________________________________________________________________ Im gesunden Körper und der Zelle besteht ein Gleichgewicht zwischen gebildeten Radikalen und deren Bindung oder Abbau durch enzymatische und nichtenzymatische Antioxidantien. Ist dieses Gleichgewicht durch erhöhte Bildung von Radikalen und/oder durch verminderte antioxidative Kapazität gestört, spricht man von „oxidativem Stress“ (ÖHLENSCHLÄGER 1993 a). Durch Radikale hervorgerufene Schäden können sich auf einzelne Zellstrukturen (z.B. DNA/RNA), Zellorganellen (z.B. Mitochondrien, Membransysteme), auf ganze Zellen (z.B. Tumorzellentstehung/Zelltod), Organe (z.B. Lunge, Haut) oder auf den kompletten Organismus (z.B. Alterung, Immunschwäche) beziehen (DORMANDY 1983). Der Nachweis von Radikalen kann direkt oder indirekt erfolgen. Die direkte Bestimmung wird aber durch die Reaktionsfreudigkeit und die Kurzlebigkeit erschwert (SCHELLER 1999). Daher wird die indirekte Bestimmung über die Messung von Reaktionsprodukten (z.B. Malondialdehyd) oder die Bestimmung über den Verbrauch von Antioxidantien bevorzugt. 2.2.2 Einteilung der Radikale 2.2.2.1 Aktivierte Sauerstoffspezies (ASS) Die ASS, auch „reactive oxygen species“ (ROS) genannt, entstehen aus Sauerstoff und können ineinander umgewandelt werden. Ihr Abbau erfolgt letztendlich zu Wasser. Tab. 3: Reaktive Sauerstoffspezies, ihre Bildung, von ihnen und durch sie hervorgerufene Schädigungen und ihr Abbau Wasserstoffperoxid (H2O2) Superoxidanion (O2-.) ASS Bildung Schädigung - aus molekularem Sauerstoff durch Flavienzyme, Mono-, Di- und Lipoxygenasen - an Übergangsmetallen und Semichinon (FONG et al. 1973) - von Struktur- und Enzymproteinen - von Zytochrom C - von Trägern von SH-Gruppen - durch Bildung von Peroxiden, Singulettsauerstoff und Hydroxylradikalen - durch hohe Reaktivität, kurze Lebensdauer - durch eigene Toxizität, aber besonders durch Bildung von Hydroxylradikalen (HALLIWELL u. GUTTERIDGE 1984; Abb. 4) - durch Flavienzyme, Superoxiddismutasen - in Mitochondrien, bes. der Leber - 16 - Abbau - Superoxiddismutasen (SCHILLER et al. 1993) - Katalasen und Oxidasen (z.B. Xanthinoxidase, ÖHLENSCHLÄ GER 1993 a) Singulett-Sauerstoff (1O2 / O2 (1g) / O2 ) Hydroxylradikal (HO. ) Literatur __________________________________________________________________________________________ - durch Reduktion von (H2O2) - durch sehr kurze Halbwertszeit, überall möglich hohe Reaktivität - von DNA/RNA (HARRIS 1992 b) - von Trägern von -SH-Gruppen - energiearme und- reiche Form - während vieler Stoffwechselprozesse, Abbau von Zytochrom p-450, über Myeloperoxidase und Reduktion von Prostaglandinen (TILL 1999) - von Doppelbindungen (in ungesättigten Fettsäuren von Membranen, in Aminosäuren und Nukleinsäuren) (ÖHLENSCHLÄGER 1993 a) - Mannit, Harnsäure, Vitamin E, Ethanol, GSH, LCystein, Melatonin (JENTZSCH 1997) - ß-Carotin Das gefährlichste Radikal stellt das Hydroxylradikal dar, da es aus Wasserstoffperoxid überall gebildet werden kann (Abb. 3). (1) HABER-WEISS-Reaktion Fe oder Cu HO. + HO- + O2 H2O2. + O2. (2) FENTON-Reaktion (GUTTERIDGE et al. 1990) H2O2 + Fe2+ Fe3+ + HO. + HO- (3) Radiolyse des Wassers über -Strahlen (HARRIS 1992) H + HO + e-aqu. H2O + hv γ (4) Reduktion durch Semichinone QH + H2O2 Abb. 3: Q + H2O + HO Reaktionstypen des Wasserstoffperoxids unter Bildung weiterer Radikale (modifiziert nach HALLIWELL u. GUTTERIDGE 1984) 2.2.2.2 Stickoxidradikale (NO ) - 17 - Literatur __________________________________________________________________________________________ Stickoxidradikale bedrohen den Körper sowohl in der Gas- als auch in gelöster Form. Dabei befindet sich das ungepaarte Elektron in der Mitte des Moleküls. (1) NO + O2- ONOO- (2) ONOO- + H+ ONNOH (3) ONOOH OH + NO2 H+ + NO3- Abb. 4: Entstehung von Stickoxidradikalen (modifiziert nach HALLIWELL et al. 1992) Stickoxidradikale selbst sind nicht sehr reaktiv. Sie werden dies aber, sobald sie sich in Verbindungen wie dem -S-Nitrothiol (S-NO), dem Nitrosyl-iron (Fe-NO), dem Nitrogendioxid (NO2 ), dem Peroxynitrit (ONOO-), dem Nitrat (NO2-) oder dem Nitrit (NO3-) befinden (NAKAZAWA et al. 1996). Diese Verbindungen treten besonders bei der Entzündungsreaktion und deren Abwehr durch die Phagozytose auf. Am häufigsten ist hierbei das Peroxynitrit zu finden, das auf SH-Gruppen als Oxidans wirkt (HALLIWELL et al. 1992). In jüngerer Zeit wird diskutiert, ob in Bezug auf das Ischämie-/Reperfusionssyndrom nicht die Peroxynitrite im Gegensatz zu den Superoxid- und Hydroxylradikalen den schädigenderen Einfluss ausüben (NAKAZAWA et al. 1996). Hieraus würden auch für die Therapie von Ischämie-/Reperfusionsschäden wichtige Änderungen resultieren, da die SOD wenig Einfluss auf den Abbau der Peroxynitrite hat. Bei der Regulation des Gefäßtonus nimmt das Stickoxid am Endothel eine positive Rolle in der Regulation ein. In Phasen der Hypoxie/Ischämie werden die Gefäße im betroffenen Gebiet durch den EDCF („endothelium-derived-constricting-factor“) eng gestellt. Folgt der Ischämie die Reperfusion mit einem Anstieg der Sauerstoffkonzentration, so wird vom Endothel der EDRF („endothelium-derived-relaxing-factor“), der dem Stickoxid entspricht, freigesetzt, der zur Weitstellung der Gefäße führt. Diese Weitstellung verringert die Adhäsion des Endothels und damit die Plättchen- und PMN-Aggregation an diesem. Ist die Reperfusion allerdings mit einer großen Freisetzung von Sauerstoffradikalen verbunden, so inaktivieren diese den EDRF, so dass die Gefäße sich wieder verengen und oben genannter Verschluss eintreten kann „noreflow phenomenon“) (AKGÜR et al. 1996). In der Labmagenwandmuskulatur spielt Stickoxid als Neurotransmitter eine bedeutende Rolle. Es wird im Antrum pylori durch Neurone synthetisiert. Untersuchungen von GEISHAUSER (1995) zeigen, dass bei Kühen mit Labmagenverlagerung vermehrt Stickoxid - 18 - Literatur __________________________________________________________________________________________ gebildet wird, was zur Hemmung der Muskelkontraktion in der Labmagenwand und damit zur Reduktion der Labmagenmotorik führt. Die veringerte Motorik hat bei der Entstehung, dem Verlauf und der Regeneration der Labmagenverlagerung große Bedeutung. In der Entstehung begünstigt eine verringerte Motorik die Gasansammlung. Damit wird der Schweregrad der Verlagerung (LDA 1-3, RDA s/c T) maßgeblich beeinflusst. Kommt die Motorik durch Verringerung der Stickoxidkonzentration wieder in Gang, werden Gas und Ingesta weiterbefördert, der Labmagen kann sich bei den LDA spontan rückverlagern („Pendellabmagen“). Nach erfolgter operativer Reposition spielt das Wiedereinsetzen der Motorik für die weitere Prognose der Kühe eine entscheidende Rolle. 2.2.2.3 Lipidperoxidation (LPO) Besondere Angriffspunkte für Radikale während des oxidativen Stresses bieten ungesättigte Fettsäuren und Lipoproteine in Membranen. Dabei entstehen resonanzstabilisierte Lipidradikale (Pentadienylradikale), die für die verselbstständigte Kettenreaktion der Lipidperoxidation verantwortlich sind (Abb. 5). -H+ R1 R1 R2 R2 R1 R2 Abb. 5: Initialisierung der Lipidperoxidation durch Bildung eines Lipidradikals aus Lipiden mit mehrfachen Doppelbindungen (modifiziert nach ENGELHART 1999) Treffen Lipidradikale auf molekularen Sauerstoff, wie er in der Phase der Reperfusion vorhanden ist, kommt es zur Bildung von Lipidperoxylradikalen, die wiederum selbst Lipidradikale aus anderen Lipiden bilden können. Folge der Lipidperoxidation ist der Verlust der strukturellen und funktionellen Einheit von Zellmembranen (ENGELHART 1999). Eine Beendigung der Kettenreaktion ist nur möglich, wenn zwei Lipidperoxylradikale unter Abgabe von Sauerstoff einen Alkohol und ein Aldehyd bilden, die stabil bleiben (Abb. 6, (1)), - 19 - Literatur __________________________________________________________________________________________ oder wenn ein Lipidperoxylradikal mit einem Lipidradikal (Abb. 6, (2)) eine stabile Verbindung eingeht. (1) LOO + LOO (2) LOO + L LO + LO + O2 L-OH + L=O LOOL Abb. 6: Beendigung der Lipidperoxidation (modifiziert nach ENGELHART 1999) Eine weitere, auch therapeutisch nutzbare Möglichkeit zur Beendigung stellt die Reaktion mit Vitamin E dar, das nachfolgend durch Vitamin wieder regeneriert werden kann (s. 2.3.2.2.1, 2.3.2.2.2). Desweiteren eignen sich auch das Vitamin-E-Analogon TROLOX („6-hydroxy2,5,7,8-tetramethyl-chroman-2-carbonic acid“), allrac -tocopherol und BHT („2,6-ditertbutyl-p-cresole“) zur therapeutischen Verminderung der Lipidperoxidation (BALTHAZARY et al. 1999). Ein Produkt der Lipidperoxidation ist das Malondialdehyd (MDA), das nur noch zu einem geringen Prozentsatz (1-5 %) aus Peroxiden von mehrfach ungesättigten Fettsäuren besteht. Es kann zur Messung des Ausmaßes der Lipidperoxidation herangezogen werden. Hierbei wird häufig die TBA-Methode („thiobarbituric acid“) genutzt, die MDA unter den mit TBA reagierenden Substanzen misst (BALTHAZARY et al. 1999). Die Bildung der Lipidradikale beginnt während der Ischämie, besonders betroffen sind die Endothelzellen im ischämischen Bereich (LAIPPLE 2000). Ausreichend ist hierbei schon eine Ischämiedauer von 7 1 Minuten. Die Lipidperoxidation führt so relativ früh zu nachweislichen Folgeschäden, wohingegen andere Pathomechanismen erst nach einer Ischämiezeit von über 30 Minuten nachweisbare Folgeschäden hervorrufen. Problematisch im Bezug auf die Labmagenverlagerung ist das Fettmobilisationssyndrom, das durch Geburt und verlagerten Labmagen forciert wird und zur Verfettung der Leber führen kann. Aufgrund des Syndroms fallen im Körper erhöhte Mengen an Fettsäuren an, die bei gleichzeitig erhöhter Konzentration an Radikalen („oxidativer Stress“) günstige Bedingungen für die Lipidperoxidation darstellen (FÜRLL et al. 2001). So beeinflussen die Störungen des Fettstoffwechsels auch den postoperativen Verlauf der Labmagenverlagerung negativ (MUDRON et al. 1994, REHAGE et al. 1996). Begünstigt wird die Lipidperoxidation durch eine Ketose, wie sie bei Rindern mit Labmagenverlagerung häufig besteht. Beim Menschen wurde durch JAIN et al. (1998) nachgewiesen, dass Ketonkörper die Lipidperoxidation - 20 - Literatur __________________________________________________________________________________________ fördern, die Radikalfreisetzung erhöhen, die Aktivität der SOD herabsetzen und Gefäßendothelien schädigen. GEISHAUSER et al. (2000) sehen in der Vermeidung einer subklinischen Ketose bei Rindern sogar die Möglichkeit, eine Labmagenverlagerung zu verhindern, nachdem sie in früheren Untersuchungen feststellten, dass Tiere mit erhöhten Konzentrationen an -Hydroxybuttersäure häufiger an einer Labmagenverlagerung erkrankten als Kühe mit physiologischen Konzentrationen (GEISHAUSER et al. 1998). 2.3 Antioxidantien 2.3.1 Definition und Vorkommen Unter Antioxidantien verstand man ursprünglich nur Stoffe mit kettenbrechenden Eigenschaften. Heute bezeichnet man jede Substanz als Antioxidans, die, wenn sie in Vergleich zu dem zu oxidierenden Substrat nur in geringer Menge vorhanden ist, deutlich die Oxidation dieses Stoffes verringert oder ganz verhindert (HALLIWELL 1989). Die Stoffgruppen, aus denen Antioxidantien bestehen, sind vielfältig und reichen von Zuckern über Proteine mit und ohne Metallbestandteil, bis hin zu Vitaminen und Enzymen. Sie sind zum Teil als natürliche Abwehrsysteme im Körper vorhanden, zum Teil aber auch über die Nahrung aufzunehmende. So steigt die antioxidative Kapazität im Serum nach der Aufnahme von Erdbeeren, Spinat, Rotwein und Vitamin C innerhalb von vier Stunden signifikant an (CAO et al. 1998). Während des Wegfangens von Radikalen verändern sich die Antioxidantien strukturell. Je nachdem, ob die Strukturänderung reversibel oder irreversibel ist, können sie regeneriert werden, ansonsten werden sie ausgeschieden. 2.3.2 Nichtenzymatische Antioxidantien 2.3.2.1 Metallbindende Proteine Tab. 4: Metallbindende Proteine als Teil nichtenzymatischer Antioxidantien Name Molekulargewicht / Aufbau Bildungsort Aufgabe - 21 - Besonderheiten Literatur __________________________________________________________________________________________ Transferrin Lactoferrin Coeruloplasmin 75-80000 75-88000 160000 Leber, RES Eisenbindung und –transport vom Darm zur Zielzelle Neutrophile Granulozyten Leber -“- - „latente Eisenbindungskapazität“ (nur 1/3 Eisenspeicher belegt) (DORMANDY 1978) - Stop der Fe-abhängigen Lipidperoxidation (NAKAZAWA et al. 1996) - vor allem in Sekreten (Milch) (NAKAZAWA et al. 1996) - Wertigkeitswechsel von Fe2+ zu Fe3+; - Wertigkeitswechsel auch bei „ferroxidase-like activity“ Kupfer, nur geringere (DORMANDY 1978) Affinität - akute Phase Protein Bilirubin - aus Erythrozytenabbau - Scavenger von Superoxid- und Hydroxylradikalen Ferritin + Hämosiderin intrazellulär -Bindung von Fe3+ zur Speicherung Bildungsort Aufgabe Name Molekulargewicht / Aufbau - Marker für Produktion an Singulettsauerstoff (ÖHLENSCHLÄGER 1993 b) Besonderheiten - Häm alleine starkes Peroxidans Hämoglobin Fe2+ im Zentrum; Globinanteil mit vier Peptidketten mit je einem Hämmolekül - Sauerstofftransport; Schutz vor dessen unkontrollierter Oxidation - Häm + H2O2: Oxo-ironspecies; fördern Lipidperoxidation; setzt Fe2+ frei - daher Plasmabindung: Met-/ Hämoglobin an Haptoglobin, Häm an Hämopexin (HALLIWELL u. GUTTERIDGE 1990) - Aufrechterhaltung kolloidosmotischer Druck Albumin 68000 Leber - Transport von FFS, Bilirubin, Mg2+, Cu2+, Fe2+ - Scavenger von HOCl 2.3.2.2 Nieder- und Mittelmolekulare Antioxidantien 2.3.2.2.1 Fettlösliche Antioxidantien -Tocopherol (Vitamin E) - 22 - - Sulfhydrylgruppen neutralisieren Radikale Literatur __________________________________________________________________________________________ Vitamin E ist ein fettlösliches, ausschließlich von Pflanzen synthetisiertes Vitamin. Es besteht aus einem Chromanring mit unterschiedlichen Substituenten und einer isoprenoiden Seitenkette. Die Aufnahme erfolgt über die Nahrung, die Resorptionsrate ist aber nur sehr gering, so dass eine Supplementierung nur parenteral erfolgen sollte. In den Zellen findet sich Vitamin E im Plasmalemm und den Mitochondrien (KOLB 1989). Zu den Aufgaben von Vitamin E gehört u.a. das „Quenching“ (siehe Vitamin A) von Singulett-Sauerstoff. Hierbei übernimmt Vitamin E ohne Elektronenübertragung Energie vom Singulett-Sauerstoff, der dadurch in seine energieärmere Form, den Triplett-Sauerstoff übergeht. Die überschüssige Energie gibt Vitamin E als Wärme ab (ENGELHART 1999). Eine Hauptfunktion von Vitamin E ist die Unterbrechung der Lipidperoxidation (Abb. 7; HALLIWELL et al. 1992). Lipid-O2 + Tocopherol-OH Lipid-O2H + Tocopherol-O- Abb. 7: Beendigung der Lipidperoxidation durch Vitamin E Diese Unterbrechung erfolgt noch bevor das Lipidradikal weiter reagieren kann. Das Vitamin E-Radikal bleibt stabil und reagiert nicht im Sinne der Peroxidation weiter, da es über Vitamin C oder das Coenzym Q (Abb. 8) wieder regeneriert wird (HALLIWELL et al. 1992). (1) Tocopherol-O + Ascorbat Semidehydroascorbat + Tocopherol-OH (2) Tocopherol-O + CoQH2 Tocopherol-OH + CoQH Abb. 8: Regenerationsmöglichkeiten von Tocopherolradikalen Das Ubichinon wird wiederum über den Elektronentransfer in der Mitochondrienmembran vom oxidierten Q10 zum reduzierten Q10H2 regeneriert. Es wirkt zudem antioxidativ auf mitochondriale Lipide (CORBUCCI et al. 1986). Darüber hinaus wird die 5-Lipoxygenase in ihrer Aktivität gehemmt, so dass es zu einer Verringerung der Leukotriensynthese in Entzündungsgebieten kommt. Unter anderem wird damit auch die Chemotaxis von neutrophilen Granulozyten verhindert, die zu einer erhöhten Radikalproduktion im Entzündungsgebiet beitragen (ENGELHART 1999). In der Behandlung und Prävention eingesetztes -Tocopherol in Verbindung mit Taurin und Selen bewirkt nach Ischämie und Reperfusion im Splanchnikusgebiet der Ratte einen gesteigerten Blutdruck und eine bessere Durchblutung des betroffenen Gebietes (YOSHIDA - 23 - Literatur __________________________________________________________________________________________ et al. 1998). Ebenso stellten STOHRER et al. (1998) fest, dass bei Ratten mit der höchsten Supplementierung an -Tocopherol nach einer experimentellen fokalen cerebralen Ischämie der geringste Reperfusionsschaden folgte. Bei Rindern, die unter Leberverfettung mit nachfolgendem Leberschaden leiden, ist in Plasma und Leber eine verminderte Vitamin E-Konzentration zu finden. Durch die Fettleber steigen MDA und Triglyceride an, insgesamt resultiert somit aus der gesteigerten Lipidperoxidation ein verminderter antioxidativer Status (MUDRON et al. 1999). Genauer aufgegliedert haben Kühe mit dem höchsten Triglyceridanteil in der Leber die höchste Vitamin E-Konzentration in der Leber und die niedrigste im Plasma. Es zeigt sich also eine positive Korrelation zwischen Triglyceridkonzentration der Leber und MDA und eine negative Korrelation zur Vitamin E-Konzentration des Plasmas (MUDRON et al. 1997). Milchkühe in Stresssituationen zeigen im Serum erhöhte Aktivitäten der Creatinkinase (CK) und erhöhte Konzentrationen an Cortisol und Harnstoff. Wurden die Tiere jedoch über einen Monat mit Vitamin E supplementiert, fiel der Anstieg der CK geringer aus. Plasma, Erythrozyten und Leber zeigen nach Supplementation erhöhte Konzentrationen an Vitamin E. Daraus folgt, dass Vitamin E nur dann zur Stressbekämpfung eingesetzt wird, wenn es im Übermaß vorhanden ist (NOCKELS et al. 1996). Provitamin A Die Provitamine des Retinols (Vitamin A) sind die Carotinoide -, - und -Carotin. Sie enthalten mindestens eine -Iononringstruktur und werden ausschließlich in Pflanzen synthetisiert und müssen über die Nahrung aufgenommen werden. Die Umwandlung zu Vitamin A erfolgt in Darm und Leber. Im Gegensatz zu Vitamin E ist eine Speicherung in Form der Carotinoide in der Leber möglich (KOLB 1989). Aus antioxidativer Sicht kommt dem Vitamin A die Rolle der Ausbildung und Stabilisierung biologischer Membranen/Epithelien zu („Epithelschutzvitamin“). In den Membranen liegt es in einem Komplex mit Proteinen vor und ist für das „Quenchen“ von Singulett-Sauerstoff verantwortlich. Die dabei aufgenommene, überschüssige Energie kann -Carotin wieder in Form von Wärme abgeben (HARRIS 1992 b). Teilweise werden dem Provitamin A auch kettenbrechende Eigenschaften zugeschrieben (ENGELHART 1999). Unter Vitamin A-Mangel stehende Mäuse weisen im Magen-Darm-Kanal eine erhöhte Konzentration an GSH, reduziertem Glutathion auf, während die Aktivität der Se-abhängigen Glutathionperoxidase deutlich abfällt. Für den Magen-Darm-Trakt bedeuten diese Vitamin A- 24 - Literatur __________________________________________________________________________________________ Mangel-Zustände eine erhöhte Anfälligkeit gegenüber Tumoren und toxischen Stoffen (DE WAZIER u. ALBRECHT 1987). Bei Rindern führt Vitamin A-Mangel zur Hyperkeratose der Labmagenschleimhaut, woraus katarrhalische Entzündungen mit Verdickung der Mukosa und Atonie des Labmagens resultieren können. Dies wiederum gilt als wichtiger Faktor bei der Entstehung der Labmagenverlagerung. Werden die defizitären Kühe wieder ausreichend mit Vitamin A versorgt, können die Inzidenz herabgesetzt und die klinischen Symptome der Labmagenverlagerung z. T. deutlich gemildert werden (MARKUSFELD 1989). Wird Vitamin A in zu großen Dosen verabreicht, so wirkt es prooxidativ. Dies wurde unter anderem bei Rauchern festgestellt, die nach einer Supplementierung mit 20 mg/d Betacarotin eine erhöhte Lungenkrebsinzidenz aufwiesen (WOODFORD u. WHITEHEAD 1998). Neben dem Provitamin A existieren noch eine große Reihe weiterer pflanzlicher Carotinoide wie Zeaxanthin, Astaxanthin und Canthaxanthin. Auch diese sind in der Lage, unter Bildung von Carotinoidradikalen (Car. bzw. (R-Car).) Radikale zu neutralisieren. Ihre Effektivität als Antioxidans wird mehr durch die Art des zu eliminierenden Radikals als durch das jeweilige Carotinoid bedingt (MORTENSEN et al. 1997). Ubichinon (Coenzym Q) Ubichinon bildet im Rahmen der Atmungskette ein Hilfssubstrat, das Wasserstoffionen sammelt, die zum Teil von NADH+H+, aus der Succinat-Dehydrogenase oder aus der Oxidation stammen (CORBUCCI et al. 1986). Es kann in oxidierter (Q10) und in reduzierter (Q10H2) Form vorliegen. Seine Wasserstoffionen können dazu verwendet werden, um Ketten zu brechen und somit Radikale wie R-OO und R-O zu neutralisieren. Ubichinon ist lipophil und daher in die Lipidphase der Mitochondrienmembran eingelagert, allerdings in weitaus geringeren Konzentrationen als Vitamin E. Es ist auch sehr viel reaktionsträger in Bezug auf seine kettenbrechenden Eigenschaften als Vitamin E (HALLIWELL et al. 1992). 2.3.2.2.2 Wasserlösliche Antioxidantien Ascorbinsäure (Vitamin C) Die Ascorbinsäure ist das En-Diol-Lacton der Keto-L-Gulonsäure. Alle Tierarten außer Primaten, Meerschwein und Regenbogenforelle können sie selbst synthetisieren. Vitamin C kann nur in geringem Umfang gespeichert werden, überschüssiges wird über die Niere ausgeschieden. Als wasserlösliches Vitamin tritt es im Zytosol sowie im Plasma auf (KOLB 1989). - 25 - Literatur __________________________________________________________________________________________ Die wichtigste Eigenschaft des Vitamin C ist seine reduzierende Wirkung, so dass es bei vielen Reaktionen als Redoxpartner beteiligt ist. Es hat die Fähigkeit, O2-, HO2-, wasserlösliche Peroxylradikale (RO2.), Thiyl- und Sulfenylradikale abzufangen. Karzinogene Nitrosamine werden zu unschädlichen Produkten reduziert, die Lipidperoxidation durch Hämoglobin- oder Myoglobin-Wasserstoffperoxid-Gemische wird verringert (LEHMANN et al. 1996). Damit wird der Freisetzung von Eisen aus dem Häm vorgesorgt, indem FerrylProteine gebildet werden. Vitamin C ist außerdem ein wirksamer Scavenger von HOCl, Singulett-Sauerstoff und Hydroxylradikalen, sobald diese in wässriger Lösung auftreten. Die reduzierenden Eigenschaften werden bei der Regeneration von -Tocopheryl-Radikalen zu Vitamin E und bei der Reduktion von Stickoxid-Radikalen, die bei Angriff von Superoxiden und Hydroxylradikalen auf Desferrioxamine entstehen, deutlich. Schließlich schützt es Plasmalipide gegen Peroxidation durch neutrophile Granulozyten (NAKAMURA et al. 1997) und bewahrt vor den in Zigarettenrauch enthaltenen Oxidantien (HALLIWELL u. GUTTERIDGE 1990). Nach erfolgter Reduktion wird das oxidierte Vitamin C über Enzyme unter Beihilfe von NADH+H+ und reduziertem Glutathion zerlegt, wobei Oxalate, Threonate und andere Stoffe entstehen. Eine eher seltene Möglichkeit ist aber auch die Regeneration über GSH (ÖHLENSCHLÄGER 1993 b). Trotz der antioxidativen Eigenschaften kann Vitamin C auch oxidationsfördernd wirken, so dass unter Anwesenheit von Kupfer- und Eisen-Salzen die Lipidperoxidation gefördert wird (NAKAMURA et al. 1997). Da die Übergangsmetalle aber nur unter pathologischen Bedingungen auftreten, kann Vitamin C auch nur unter pathologischen Bedingungen prooxidativ wirken. Aus diesem Grund ist der therapeutische Effekt von Vitamin C-Gaben bei Krankheit fragwürdig. So ergab eine Studie bei Rauchern mit Supplementierung von Vitamin C eine Erhöhung der Inzidenz an Lungenkrebs (WOODFORD u. WHITEHEAD 1998). Glutathion Glutathion ist ein aus den Aminosäuren Glutaminsäure, Glycin und Cystein bestehendes Tripeptid, das extraribosomal in der Leber synthetisiert wird. Über Blut und Galle wird es im Körper verteilt (NAKAMURA et al. 1997). Als wasserlösliches Peptid tritt es im Cytosol und in den Mitochondrien auf. Seine herausragende Eigenschaft ist die Fähigkeit, sich leicht zu einer Disulfidform oxidieren zu lassen. So können Superoxidradikale und Hydroxylradikale abgefangen werden. Vor allem in den Erythrozyten, in denen Glutathion in größeren Mengen auftritt, ist dies von Bedeutung, da diese sehr empfindlich gegen Oxidation sind. Ist zu wenig - 26 - Literatur __________________________________________________________________________________________ Glutathion vorhanden oder verbraucht, werden die Erythrozytenenzyme und die Membranen zerstört, es kommt zur Hämolyse. Um dies zu vermeiden, können Zellen bei einer Belastung mit Radikalen und oxidiertem GSSG den Hexose-Mono-Phosphat-Weg aktivieren, um Elektronen zu erhalten, die dem oxidierten Glutathion als Reduktionsmittel dienen (COCHRANE 1991). Harnsäure (Urat) Die Harnsäure, die in der Leber und zu geringen Teilen auch in Dünndarm und Niere synthetisiert wird, ist ein Endprodukt des Purinbasenabbaus (KOLB 1989). Aus antioxidativer Sicht gilt Harnsäure im Plasma als guter Scavenger von Singulett-Sauerstoff, HOCl und Peroxyl-Radikalen. Zudem bindet sie Eisen- und Kupferionen, so dass diese nicht zur Radikalbildung zur Verfügung stehen. So kann auch die Lipidperoxidation verhindert werden (HALLIWELL u. GUTTERIDGE 1990). Reagiert Harnsäure allerdings mit Oxidantien, können Harnsäureradikale daraus entstehen, die selbst schädigende Wirkung haben, aber wiederum durch Vitamin C reduziert und damit unschädlich gemacht werden können (HALLIWELL u. GUTTERIDGE 1990). Ein weiterer Nebeneffekt in Geweben mit Hypoxie besteht darin, dass beim Abbau von Xanthin bzw. der Synthese von Harnsäure das dazu notwendige Enzym sich in seine Oxidaseform umwandelt und es nunmehr bei der Reaktion zur Bildung von hyperreaktivem Superoxidanion kommt (BERGHAUS 1997) (Abb. 9). Physiologische Bedingungen: Xanthin- Xanthin + H2O + NAD + Harnsäure + NADH + H+ dehydrogenase Hypoxische Bedingungen: Xanthin- Harnsäure + 2 O2-. + 2 H+ Xanthin + H2O + 2 O2 Oxidase Abb. 9: Umwandlung der Xanthindehydrogenase zur Xanthinoxidase unter hypoxischen Bedingungen (modifiziert nach BERGHAUS 1997) 2.3.2.3 TEAC-Konzentration Die Messung der nichtenzymatischen Antioxidantien in einem Substrat erfolgt zumeist komplex. Dabei kommen verschiedene Verfahren zur Anwendung. Zum einen kann der antioxidative Status zu einer bestimmten Zeit gemessen werden, zum anderen der Grad einer antioxidativen Schutzfunktion einer Probe gegenüber einem Oxidans. Trolox, Vitamin C und Vitamin E bieten beispielsweise einen kompletten Schutz vor definierten Oxidantien. Kleine - 27 - Literatur __________________________________________________________________________________________ Moleküle, wie Melatonin und GSH, aber auch Albumin, bieten nur zu einem gewissen Teil Schutz vor Oxidantien (CAO et al. 1995). Um beide Komponenten in einem zu vereinigen, geht man in letzter Zeit dazu über, die antioxidative Kapazität als „area under curve“ zu bestimmen, d.h. man misst das Ausmaß des Schutzes vor Oxidantien über eine bestimmte Zeit. Dieses Verfahren wird als TRAP („total radical-trapping antioxidant parameter“) (GHISELLI et al. 1995, TUBARO et al. 1998) bezeichnet, wenn es für alle Arten von Radikalen anwendbar ist, und als ORAC („oxygen radical absorbance capacity“), wenn es für aktivierte Sauerstoffstufen (ASS) angewendet wird (CAO et al. 1993, CAO et al. 1995, NINFALI u. ALUIGI 1998). Zumeist wird dabei die antioxidative Fähigkeit der Probe mit einem definierten Antioxidans verglichen. Bei der TEAC-Konzentration stellt TROLOX, ein Vitamin E-Analogon, dieses definierte Antioxidans dar (MILLER et al. 1996, ROMAY et al. 1996, LANDS et al. 2000). Des weiteren kommen Methoden zum Einsatz, bei denen die Chemilumineszenz von bestimmten Stoffen in Anwesenheit von Antioxidantien verändert wird (BAADER et al. 2000, HERMANN 2000). Diese neuen Methoden bieten neben der Bestimmung der totalen antioxidativen Kapazität auch gleichzeitig die Möglichkeit zur Bestimmung einzelner Antioxidantien wie Vitamin C und Harnsäure (POPOV u. LEWIN 2000). Der Nachteil vieler Methoden besteht darin, dass sie für alle Arten von Proben anwendbar sein sollen, so dass Differenzen zwischen der Summe der einzelnen Antioxidantien zur Gesamtbestimmung auftreten (WOODFORD u. WHITEHEAD 1998). So wurden für die TEAC-Konzentrations-Messung auf die Probenart abgestimmte Testkits entwickelt, so dass sie für die Messung der wasserlöslichen Antioxidantien im Plasma (siehe 2.3.2.2) sehr zuverlässig sind. Wir haben uns für dieses Verfahren entschieden, da es durch gute Reproduzierbarkeit und relativ leichte Anwendbarkeit bei großen Probenmengen gute Vergleichsmöglichkeiten bietet. Bei gesunden Kühen liegt die TEAC-Konzentration zwischen 200 und 300 μmol/l (WILKEN u. FÜRLL 2002). Im zeitlichen Abstand zur Geburt zeigt sich ein steigender Trend der TEAC-Konzentration. Signifikante Unterschiede zwischen den einzelnen Leistungsgruppen der Milchproduktion bestehen nicht. Im Laktationsverlauf nimmt die TEAC-Konzentration zu. Mutterkühe haben im Vergleich zu Hochleistungskühen signifikant geringere TEACKonzentrationen (WILKEN u. FÜRLL 2002). Bei kranken Kühen (Klauenerkrankungen, Mastitiden, Metritiden, DA etc.) sind die TEAC-Konzentrationen niedriger als die der Kontrollgruppe, wobei der Abstand zum Partus vergleichend miteinbezogen wurde (GOERRES u. FÜRLL 2002). - 28 - Literatur __________________________________________________________________________________________ 2.3.3 Enzymatische Antioxidantien 2.3.3.1 Enzyme zur Glutathionregeneration Glutathionperoxidasen (GSHPx; GPX) Die Glutathionperoxidase ist eine Wasserstoffperoxid-Oxidoreduktase (E.C 1.11.1.9). Sie bildet aus Glutathion ein Disulfid, um aus Wasserstoffperoxid und organischen Hydroperoxiden Wasser zu gewinnen. (PARKS 1989) (Abb. 10). (1) H2O2 + 2 GSH GSSG + 2 H2O (2) R-OOH + 2 GSH GSSG + R-OH + H2O Abb. 10: Entstehung von Glutathiondisulfiden (modifiziert nach PARKS 1989) Die Glutathionperoxidase dient dem Schutz der Zellmembranen, im besonderen der Mitochondrienmembranen (KÜHN 1997). Von MILLS et al. wurde 1957 das erste Mal von einer Glutathionperoxidase in Rindererythrozyten berichtet (KÜHN 1997). Die Struktur der GPX besteht aus einem Tetramer mit vier Untereinheiten, die eine Peptidkette mit einem Massengewicht von 22000, ungefähr 180 Aminosäuren und jeweils ein Selenatom besitzen. Die einzelnen Monomere sind dabei fast kugelförmig mit einem Radius von 19 Å (WENDEL u. CIKRYT 1980). Die Angaben sind nur ungefähr zu treffen, da jede Spezies ihre eigenen Arten von GPX hat, die durch unterschiedliche spezifische Gewichte bei fast gleicher Selenkonzentration gekennzeichnet sind (BAUMGARTNER 1983, Tab. 5). Tab. 5: Vergleich der relativen Dichte und der Selenkonzentration der GPX verschiedener Tierarten (modifiziert nach BAUMGARTNER 1983) Autoren Jahr Tierart/Gewebe Relative Dichte g Selen/Molekül FLOHÉ et al. 1973 Rinder- 83000 4,4 22000 3,8 erythrozyten OH et al. 1974 Schaferythrozyten BELL u. COWEY 1982 Regenbogen- 100000 Forelle NAKAMURA et al. 1997 Rattenleber - 29 - 75-76000 4,24 Literatur __________________________________________________________________________________________ In Bezug auf die Einteilung der Enzyme innerhalb der Glutathionperoxidasen gibt es mehrere Varianten. ÖHLENSCHLÄGER (1993 b) nennt zunächst drei Grundtypen der GPX: I. GPX mit Selenocystein als aktives Zentrum II. GPX als sulfhydrylhaltige Enzyme III. Selen-unabhängige Enzyme Hierbei kommen I.+ II. im Zytosol und den Mitochondrien von Pilzen, Algen und Tieren vor, sind aber bei höheren Pflanzen und Bakterien nicht zu finden. Die Gruppe der selenocysteinhaltigen GPX ist dabei stabiler gegenüber Schäden, die durch Peroxide verursacht werden als die Gruppe der sulfhydrylhaltigen Enzyme. Die Selen-unabhängigen Enzyme sind bei Tieren nicht zu finden und können kein Wasserstoffperoxid, sondern nur andere Peroxide umsetzen. SCHOLZ (1991) dagegen beschreibt, dass bei Tieren in Milz, Herzmuskel, Erythrozyten, Gehirn, Thymus, Fettgewebe und quergestreifter Muskulatur ausschließlich Selen-abhängige GPX zu finden ist. In Leber, Lunge, Nebenniere, Hoden und Niere treten jedoch Selen-ab- und -unabhängige Arten der GPX auf. In der Leber ist insgesamt der höchste Gehalt an Selen-unabhängiger GPX zu finden. Die Glutathionperoxidasen lassen sich auch nach ihrem Vorkommen und dem Zeitpunkt ihrer Entdeckung einteilen: I. zytosolische Glutathionperoxidase (cGPX): FLOHÉ et al. (1973) II. Phospholipidhydroperoxid-Glutathionperoxidase (PHGPX): URSINI et al. (1985) III. glykosilierte extrazelluläre Glutathionperoxidase (pGPX): BRIGELIUS-FLOHE et al. (1999) IV. gastrointestinale Glutathionperoxidase (GIGPX): CHU et al. (1993) Als gemeinsames Kennzeichen gilt, dass alle GPX Hydroxide mit Hilfe von Thiolen reduzieren. Sie besitzen alle eine Triade aus Selenocystein, Tryptophan und Glutamin als aktives Zentrum. Unterscheiden lassen sich die vier Gruppen anhand ihrer Substrate. So reduzieren die cGPX und die GIGPX nur Wasserstoffperoxid und lösliche organische Peroxide, während die PHGPX und die pGPX auch komplexe Hydroperoxide wie Phospholipidhydroperoxide umsetzen. Die PHGPX kann als einzige auch Cholesterinhydroperoxide reduzieren. Dies ist vor allem beim Schutz von Membranen vor Lipidperoxidation von Bedeutung (URSINI et al. 1985). - 30 - Literatur __________________________________________________________________________________________ Einzeln betrachtet stellt sich die zytosolische GPX als Homotetramer dar, mit einem 1Seleno-Cystein-Rest, der jeweils von einem von vier Argininresten gebunden wird. Diese Argininreste wiederum umgeben das katalytische Zentrum. Die cGPX findet sich in höheren Konzentrationen in Erythrozyten, Phagozyten, Leber und Niere (FLOHÉ 1997). Bei der Entgiftung von Peroxiden arbeitet sie effektiver als die Katalase. Fehlt die cGPX dem Körper, so wird dieser sehr anfällig gegenüber Schäden, die aus dem Ischämie-/Reperfusionssyndrom hervorgehen. Dies wurde anhand von „cGPX-knockout-Mäusen“ nach Myokardischämie untersucht (YOSHIDA et al. 1998). Darüber hinaus lässt sich die cGPX als Marker für den Selen-Status eines Organismus heranziehen, da sie als Selenspeicher mit regulativer Funktion fungiert. Die Phospholipid-Hydroperoxid-GPX tritt als Monomer auf. Als besondere Eigenschaft ist bei ihr zu nennen, dass sie zusammen mit -Tocopherol der Peroxidation von Membranlipiden Einhalt gebietet (MAIORINO et al. 1989). Sie kommt in hohen Konzentrationen in den Hoden vor (ROVERI et al. 1992, ROVERI et al. 2001). Bei der extrazellulären GPX handelt es sich um ein Tetramer, das vor allen im Plasma (TAKAHASHI et al. 1987), in der Milch und anderen extrazellulären Flüssigkeiten vorkommt. Die pGPX wird in Leber und Niere synthetisiert. Durch ihre sehr niedrige Plasmakonzentration wird ihre Bedeutung als Plasmaperoxidase in Frage gestellt. Trotz der großen Ähnlichkeit der gastointestinalen GPX und der zytosolischen GPX in Struktur und Funktion hat die GIGPX doch eine weitaus größere Affinität zu organischen Peroxiden als die cGPX, was für ihre Funktion als Reduktionsmittel für Peroxide, die aus der Nahrung stammen, von großer Bedeutung ist. Dieser Zusammenhang wird durch die negative Korrelation des GIGPX-Status mit den aus dem Darmlumen abtransportierten Hydroperoxiden untermauert. Eine hohe Aktivität der GIGPX ist eventuell auch für eine höhere Resistenz gegenüber Kolonkrebs beim Menschen verantwortlich (CHU et al. 1993). Allgemein gilt für alle Arten der intrazellulären GPX, dass sie ihre Aktivitäten zu 75 % frei im Cytosol und zu 25 % locker an die Mitochondrienmembran gebunden, entfalten. Sie treten nicht in den Peroxisomen auf. Eine Messung der GPX-Aktivität kann spektrophotometrisch über die Extinktionsabnahme des Substrates NADPH bei 334 nm und bei konstanter Temperatur und GSH-Konzentration erfolgen. Dazu bieten sich im Blut die Erythrozyten an, wobei ein signifikanter Unterschied zwischen der Aktivität der GPX im Hämolysat und der Aktivität in einer Suspension aus gewaschenen Erythrozyten von Schafen besteht (SHEPPARD u. MILLAR 1981). Die Aktivität im Plasma beträgt insgesamt weniger als 1 % - 31 - Literatur __________________________________________________________________________________________ der Gesamtaktivität im Blut. Nach der Definition von PAGLIA und VALENTINE (1938) repräsentiert 1 mU Enzymaktivität die Oxidation von 1 mmol NADPH zu NADP+ pro Minute bei 25 °C (BACKALL u. SCHOLZ 1979). Die Bildung der Erythrozyten-GPX erfolgt im Knochenmark in den Erythroblasten und im weiteren Verlauf durch Aufnahme aus dem Blutplasma. So besitzen die ältesten Erythrozyten die höchste Konzentration an GPX, obwohl die ursprüngliche Konzentration an GPX in der Reifungsphase der Erythrozyten determiniert wird und in den Erythrozyten selbst keine Neubildung erfolgt (PERONA et al. 1978). Als Zeichen für den von der GPX ausgehenden Schutz der Erythrozyten ist festzustellen, dass Erythrozyten mit einer hohen GPX-Aktivität länger leben als welche mit geringer Aktivität (TUCKER et al. 1974). Zur Gesamtaktivität der GPX sind in der Literatur viele unterschiedliche Messergebnisse innerhalb einzelner Tierarten und Messreihen einer Tierart zu finden, obwohl alle Studien die Aktivität nach der Methode von PAGLIA und VALENTINE (1938) bestimmt haben, so dass ein Vergleich zwischen Messergebnissen verschiedener Messreihen schwerfällt (Tab. 6). Tab. 6: GPX-Aktivitäts-Vergleich von Erythrozyten zwischen Rind und Schaf (modifiziert nach ALEKOS 1992) Autor GPX-Aktivität Jahr Tierart THOMPSON et al. 1976 Rind 6 - 41 DOTTA 1979 Rind 0,7 - 78,4 SUZUKI et al. 1985 Rind 122 - 208 ERSKINE 1987 Rind 3 - 35,6 DOTTA 1988 Rind 35,7 HEIKENS 1992 Rind 10,5 - 183,1 DOTTA 1978 Schaf 43,3 - 367 - 32 - (U/gHb) Literatur __________________________________________________________________________________________ DIMANOV 1989 Schaf 21 - 74 HEIKENS 1992 Schaf 88,4 - 442,1 Neuere Untersuchungen zur GPX-Aktivität zeigen, dass ab einer Aktivität von 130 U/mg Hb von einem bedarfsgerechten Selenstatus ausgegangen werden kann (WOLF 1998). Die von SATTLER (2001) untersuchten Kühe mit DA zeigten Aktivitäten von 385 ± 103 U/g Hb. Die von WOLF (1998) festgestellten GPX-Aktivitäten bei gesunden Kühen lagen im selben Bereich. Im jahreszeitlichen Verlauf liegen die Aktivitäten der GPX in den Wintermonaten Dezember bis Februar niedriger ( 355 ± 101 U/mg Hb) als in den Herbstmonaten (411 ± 104 U/mg Hb) (SATTLER 2001). Im zeitlichen Abstand zur Abkalbung steigt die GPX-Aktivität ein bis drei Wochen post partum moderat an (SATTLER 2001). Die Regulation der GPX unterliegt vielen beeinflussenden Faktoren. Eine große Rolle spielt hierbei die Selenverfügbarkeit in der Art einer posttranskriptionalen Regelung (CHADA et al. 1989). Die Beeinflussung durch den Selenstatus bei Tieren ist bei Rind, Schaf, Pferd, Huhn und Ratte nachgewiesen (BACKALL u. SCHOLZ 1979). Bei Rindern und Kälbern gilt eine Selenkonzentration von weniger als 20 µg/l Blut als Mangelzustand (SCHOLZ 1991). Steht dem Körper viel Selen zur Verfügung, so werden stabile Selenoproteine ausgebildet. Bei geringem Selenangebot kommt es nicht nur zum Abbau dieser Selenoproteine, sondern auch ihrer m-RNA. Davon sind besonders die cGPX betroffen, die PHGPX hingegen bleibt unverändert (HILL et al. 1992). Die pGPX liegt zwischen beiden Extremen (AVISSAR et al. 1994). Beeinflusst man den Selenstatus durch Gaben eines Vitamin E-Selen-Gemisches, so kann dies bei Pferden zu einer Verdreifachung der GPX-Aktivität führen (CAPLE et al. 1978, MAYLIN et al. 1980). Dies zeigt auch der nach jeweiliger Selen-Vitamin E-Substitution bei Rindern auftretende Aktivitätsunterschied bei WOLF (1998). Bei oxidativem Stress wie Reperfusion, bei Endotoxinbelastung und bei Interleukin-1-Anstieg steigt die Aktivität der cGPX an, mit Abfall der Stressoren sinkt sie wieder ab (NIWA et al. 1993). Bei Untersuchungen am Rattendarm (BAUER et al. 1995) wurde festgestellt, dass das für die GPX so wichtige Selen im Schock und in Se-Mangelzuständen in bestimmten Geweben angereichert wird. Im Schock wird es nicht in dem Gewebe angereichert, in dem der Schock seinen Ausgang nimmt, sondern in Leber und Niere; in Mangelzuständen wird es in Gehirn, Fortpflanzungs- und endokrinen Organen angereichert. Dies führte in vorliegender Untersuchung zu dem Phänomen, dass die GPX-Aktivität nach Ischämie und Reperfusion am Darm in den Erythrozyten abnahm. Bei Eisenmangel kann sich die Aktivität auf ein Viertel verringern, was bei Kaninchen (CELLERINO et al. 1976) und beim Menschen (HOPKINS u. - 33 - Literatur __________________________________________________________________________________________ TUDHOPE 1973) nachgewiesen wurde. Ein Vitamin E-Defizit führt bei Ratten unter gleichbleibender Katalaseaktivität zu einem Abfall der GPX-Aktivität (CHOW 1977, BOURRE et al. 2000). In Bezug auf das Alter wiesen JORGENSEN et al. (1977) eine mit zunehmendem Alter steigende Aktivität der GPX beim Schwein nach. Glutathionreduktasen Die Glutathionreduktase (E.C. 1.6.4.2.) ist eine Glutathiondisulfid-Oxidoreduktase ((NADCP)H), die in den löslichen Fraktionen von Pro- und Eukaryonten vorkommt. Als Flavoprotein enthält sie ein Flavinadenindinukleotid. Ihr obliegt die Regulation der Konzentrationen von Glutathion (GSH) und Glutathiondisulfid (GSSG) unter Verbrauch von NADPH + H+. GSSG + NADPH + H+ 2 GSH + NADP- Steigt die Konzentration von GSSG, wie es bei einem ungünstigen Redox-Potential der Fall ist, so kommt es nicht mehr zur Regenerierung von GSSG, stattdessen können durch Disulfidbildung Proteine und Coenzym A inaktiviert werden. Darüber hinaus ist GSSG ein Inhibitor der Proteinbiosynthese (KÜHLER 1996). In Bezug auf die Regulation der Glutathionreduktase wurde festgestellt, dass die Glutathionreduktase und der Mono-Hexosephosphatweg als Lieferant des NADPH negativ miteinander korrelieren, so dass bei Ausfall der Regeneration von GSSG über NADPH die Glutathionreduktase einspringt (BRADY et al. 1978). Glutathion-S-Transferase Die Glutathion-S-Transferase (E.C. 2.5.1.18.) besteht aus einem Dimer und kommt im Cytosol und in Mikrosomen von Hepatozyten vor (NAKAZAWA et al. 1996). Beim Menschen besitzt sie drei Untergruppen. Ihre Hauptaufgabe besteht darin, Xenobiotika durch Konju-gation mit Glutathion in ihrer Toxizität abzuschwächen und sie wasserlöslicher zu machen, damit sie leichter ausgeschieden werden können (KÜHLER 1996). So reagieren Ratten, die vor Versuchsbeginn mit Cadmium behandelt worden waren, auf eine intraperitoneale In- jektion mit Ozon und/oder Sauerstoff mit einem Anstieg der GST in Herz, Niere und Leber. Bei Verabreichung von Sauerstoff steigt die Aktivität in Gehirn. Cadmium allein erhöht die Aktivität in allen vier genannten Organen. Auffällig dabei ist, dass - 34 - Literatur __________________________________________________________________________________________ die GPX in diesen Organen unverändert bleibt oder sogar in ihrer Aktivität absinkt (LASZCZYCA et al. 1996). 2.3.3.2 Superoxiddismutasen (SOD) Bei den Superoxiddismutasen (E.C. 1.15.1.1.) handelt es sich um Superoxid- Oxidoreduktasen. Als Metalloenzyme haben sie verschiedene Metalle in ihrem aktiven Zentrum, die auch zu einer Einteilung in drei Gruppen genutzt werden (KÜHLER 1996). I. Cu-Zn-SOD: Kupfer und Zink im aktiven Zentrum II. Fe-SOD: Eisen im aktiven Zentrum III. Mn-SOD: Mangan im aktiven Zentrum Die Cu-Zn-SOD kommt im Cytoplasma, im intermembranösen Raum der Mitochondrien und nur gering in Lysosomen von Eukaryonten vor, die Fe-SOD ist nur in Prokaryonten und in Pflanzen anzutreffen und die Mn-SOD ist in Prokaryonten und in der Mitochondrienmatrix von Eukaryonten zu finden (KÜHLER 1996, NAKAZAWA et al. 1996). Schon 1938 wurde die SOD nach der Entdeckung im Rinderblut erstmals unter dem Namen „Hämocuprein“ erwähnt. 1953 wurde ein ähnliches Protein in der Pferdeleber entdeckt, das „Hepatocuprein“. Es folgte 1959 das menschliche „Cerebrocuprein“ aus dem Gehirn. Die Funktion wurde 1969 erklärt, seitdem gilt die Bezeichnung Cu-Zn-Superoxiddismutase. Sie besteht aus einem Dimer von zwei identischen Untereinheiten und hat ein Molekulargewicht von 32600 (BULKLEY 1987, HARRIS 1992 a). Später folgte auch die Entdeckung der manganhaltigen SOD in E. coli, Streptococcus mutans und in den Mitochondrien der Hühnerleber. Sie blieb im Gegensatz zur Cu-Zn-SOD unter der Behandlung von Chloroform und Ethanol nicht stabil. Zuletzt wurde schließlich in E. coli und in Photobakterien die FESOD entdeckt (KÖNEN 1982). Alle Superoxiddismutasen reduzieren Superoxide unter Verbrauch von Wasserstoffionen zu molekularem Sauerstoff und Wasserstoffperoxid (Abb. 11), welches dann von der Katalase weiter entgiftet wird. Hierbei ist auch die „FENTON chemistry“ von großer Bedeutung (GUTTERIDGE et al. 1990). Die spontane Zerfallsrate der O2--Radikaldismutation beträgt bei pH 7,4 ca. 2 x 105 M-1 sec-1. Unter Einwirkung von SOD erhöht sich die Umwandlungsrate um den Faktor 104 auf 2 x 109 M-1 sec-1 (ZIMMERMANN 1983). - 35 - Literatur __________________________________________________________________________________________ So führt eine gesteigerte Durchblutung mit intakten Erythrozyten und funktionsfähiger Katalase nach einem Infarkt am isolierten Rattenherz zu einer Verminderung des H2O2 im Vergleich zu unbehandelten Tieren (BROWN et al. 1989). Auch bei in-vitro Untersuchungen an isolierten Myozyten von Ratten zeigte sich, dass die Zellen nach Konditionierung sogar in der Lage waren, alle während einer einstündigen Anoxie entstandenen Superoxidradikale zu beseitigen (JUVONEN et al. 1997). Die Verstoffwechslung der Superoxidanionen erfolgt in Mikrosekunden, das Reaktionsgleichgewicht liegt vollständig auf der Seite der Reaktionsprodukte. Sind Superoxidradikale vermehrt vorhanden, so liegt das an der für den Umsatz zu langsamen Diffusion der Radikale in Reichweite der SOD (ANSORGE u. GEMKOW 1991). 2 O2-. + 2 H+ O2 + H2O2 Abb. 11: Beseitigung von Superoxiden durch die SOD Die Unterscheidung der Isoformen des Enzyms erfolgt über die unterschiedliche Cyanidsensitivität. Dabei reagiert die Cu-Zn-SOD im Gegensatz zur Fe-SOD und zur MnSOD sensibel auf Cyanid (KÖNEN 1982, KÜHLER 1996). Die Cu-Zn-SOD kommt ubiquitär in allen Säugetierzellen in Konzentrationen von 0,001-0,05 % vor, gehäuft tritt sie in Hepatozyten, der Niere und den Erythrozyten auf. Sie kommt fast ausschließlich intrazellulär vor, da sie keine Zellmembranen permeieren kann (TURRENS 1991). Ausnahmen hiervon sind nur bei sehr hohen Konzentrationen und in vitro möglich. Eine Ausnahme bildet die extrazelluläre Cu-Zn-SOD, die an Endothelzellen gebunden vorkommt und diesen einen lokalen, wenn auch geringen, Schutz vor Oxidantien bietet (HALLIWELL 1989). Auch die Mn-SOD kann „reactive oxygen species“ beseitigen, Superoxidradikale in nanomolaren und Hydroxylradikale in mikromolaren Konzentrationen. Eine Erhöhung ihrer Aktivität lässt sich durch Zufütterung von Mangandichlorid erreichen. Hierbei gilt allerdings die Neurotoxizität von Mangan in hohen Dosen zu beachten (HUSSAIN u. ALI 1999). Bei der Hepatitis C-Infektion lässt sich in den Monozyten eine erhöhte Konzentration der MnSOD-mRNA feststellen, nicht jedoch in den Hepatozyten selbst. Nach Ausheilung der Erkrankung ist diese erhöhte Konzentration nicht mehr nachweisbar (LARREA et al. 1998). Die SOD-Aktivität kann indirekt durch Messung der Autoxidation eines Substrates wie Adrenalin, Hydroxylamin und Pyrrogallol in Anwesenheit von SOD bestimmt werden. - 36 - Literatur __________________________________________________________________________________________ Mittels der Elektronenspinresonanztechnik kann aber auch eine direkte Messung der übrig bleibenden freien Radikale aus einer definierten Menge in einer definierten Zeit erfolgen (KÜHLER 1996). Eine Möglichkeit zur quantitativen Messung der SOD stellt die „elektrospray ionization mass spectrometry“ (ESIMS) dar, bei der die SOD in einer glykosilierten Form aus Erythrozyten extrahiert wird (SARASWATHI et al. 1999). In letzter Zeit sind auch Testkits erhältlich, die zur Identifizierung der SOD Antikörper verwenden. Bislang können mittels der Antikörper aber nur hohe Konzentrationen der SOD nachgewiesen werden, so dass sie für die Messung der SOD ausscheiden. Dasselbe gilt für die Messung der GPX mittels Western Blot und Immunhistochemie, wobei hier die erhältlichen Antikörper noch nicht vollständig in der Lage sind, die GPX sicher zu identifizieren (YAN et al. 1998). Eine Beeinflussung der Cu-Zn-SOD-Aktivität kann über eine diätetische Kupfersupplementierung erfolgen, was eine Erhöhung der Aktivität bewirkt. Diese Steigerung der Aktivität lässt sich nur diätetisch hervorrufen, da das Kupfer zellgebunden sein muss, um die in allen Zellen enthaltenen geringen Mengen einer „schlafenden“ SOD durch Einbau des fehlenden Kupfers zu aktivieren (HARRIS 1992 a). Bei einer diätetischen Beeinflussung der SOD-Aktivität sind allerdings tierartliche Unterschiede zu beachten. So beeinflusst eine ZnSupplementierung die Plasmakonzentration zwar bei Kamelen nicht, aber bei Kühen. Eine Erhöhung der Kupferkonzentration im Plasma bleibt bei beiden Tierarten ohne Effekt auf die SOD-Aktivität, eine Erhöhung der Plasmakonzentration von Zn erhöht die Aktivität der SOD bei Kühen, bei Kamelen jedoch nicht (BENGOUMI et al. 1998). Bislang bei der Tierart Rind durchgeführte Untersuchungen zur SOD-Aktivität beziehen diese auf den Mililiter Erythrozytenlysat. FÜRLL et al. (1999) sowie SATTLER (2001) stellten bei gesunden Kühen eine Abhängigkeit der SOD-Aktivität vom Abstand zur Abkalbung fest (Tab. 7). Kühe mit DA hatten keine signifikant differierenden Aktivitäten, wobei bei Tieren mit LDA tendenziell geringere Aktivitäten als bei Tieren mit RDA festgestellt wurden. Weitere, bislang unveröffentlichte Untersuchungen zur SOD-Aktivität haben ergeben, dass jahreszeitliche Anpassungen vorliegen. So steigt die Aktivität der SOD in den Frühjahrs- und Sommermonaten an, während sie in den Herbst- und Wintermonaten absinkt (ZAHN u. FÜRLL 2004). Kühe mit SOD-Aktivitäten <7000 U/ml Erythrozytenlysat gelten als stärker stoffwechselbelastet als Kühe mit SOD-Aktivitäten von 7-10.000 U/ml Erythrozytenlysat (SATTLER u. FÜRLL 2002). In Verlaufsuntersuchungen nach operativer Rückverlagerung des verlagerten Labmagens zeugt ein Anstieg der SOD-Aktivität nach Rückverlagerung bei - 37 - Literatur __________________________________________________________________________________________ Kühen mit RDA von deutlichem oxidativem Stress, der aber 24 Stunden post operationem wieder auf die Ausgangsaktivität zurückkehrt. Tab. 7: SOD-Aktivitäten bei gesunden Kühen und bei Kühen mit DA, abgetragen zum Abstand post partum (modifiziert nach SATTLER 2001) Wochen SOD-Aktivität (U/ml Erythrozytenlysat) post partum Gesunde Kühe Kühe mit DA 1 6700 7500 3 7500 5960 >3 8750 Aufgrund der früh entdeckten antioxidierenden und antiinflammatorischen Eigenschaften wird die SOD schon seit 1964 als Medikament eingesetzt. HUBER und SAIFER gewannen „Orgotein“ aus Rinderleberextrakt. Es wird seit langem in der Humanmedizin bei der rheumatischen Arthritis eingesetzt (HUBER et al. 1980). Es hat dabei, im Gegensatz zur körpereigenen SOD, die Aufgabe, Radikale und Entzündungskomponenten im Extrazellulärraum zu reduzieren. Weitere Beispiele für den Einsatz der Superoxiddismutase sind die Verminderung von PMA-induzierten Lungenschäden (ALLISON et al. 1988), Ischämie/Reperfusion während und nach Transplantation von Organen, z. B. Niere (OURIEL et al. 1985). Eine Langzeitbehandlung mit SOD in Tablettenform ist bislang nicht möglich, da sie zu über 90 % wieder über den Kot ausgeschieden wird. 2.3.3.3 Katalase Bei der Katalase (E. C. 1.11.1.6.) handelt es sich um eine Hydrogenperoxid-Oxidoreduktase. Sie besteht aus einem Tetramer, wobei jede Untereinheit eine Hämgruppe trägt. Sie kann vier NADPH-Moleküle binden, um sich gegen eine Inaktivierung durch Wasserstoffperoxid zu schützen. Zu finden ist sie vor allem in aeroben Mikroorganismen, Pflanzen- und Tierzellen, aber auch in einigen Anaerobiern (KÜHLER 1996). Die Aktivität der Katalase verhält sich bei den Säugetieren organspezifisch, so ist sie in Leber, Niere und Erythrozyten sehr hoch, in - 38 - Literatur __________________________________________________________________________________________ Gehirn, Herz und Skelettmuskel dagegen sehr gering (NAKAZAWA et al. 1996). Lokalisiert ist sie in den Peroxisomen der Zellen. Ihre Aufgabe besteht in der Regulation der Wasserstoffperoxidkonzentration und damit im Schutz des Hämoglobins und anderer Proteine mit Sulfhydrylgruppen. Das allgemeine Reaktionsmuster besteht in der Spaltung von Wasserstoffperoxid in Wasser und Sauerstoff und der Bildung von Wasser aus Carboxylgruppen einer organischen Verbindung (R-) unter Beihilfe eines Protonendonors (D) (Abb. 12). Das H2O2 ist gleichzeitig Wasserstoffakzeptor und -donator. (1) 2 H2O2 (2) R-OOH + D-H2 Katalase 2 H2O + O2 Katalase H2O + R-OH + D Abb. 12: Reaktionsschemata der Katalase (modifiziert nach KÜHLER 1996) Welche Reaktion dabei überwiegt, hängt von der Menge an H2O2 bzw. der Menge des Protonendonators ab. Gemessen wird die Katalase photometrisch. Wasserstoffperoxid hat im UV-Bereich eine bestimmte Absorption. Daher kann bei einer Wellenlänge von 240 nm mittels der Extinktionsabnahme direkt die Spaltung des Wasserstoffperoxids verfolgt werden. Die Aktivität der Katalase ergibt sich dann aus der Extinktionsabnahme des H2O2 pro Zeiteinheit (KÜHLER 1996). 2.4 Creatinkinase (CK) In den dreißiger und vierziger Jahren wurden Creatin, Creatinin und die Creatinkinase (CK) als dazugehöriges Enzym entdeckt (WYSS u. KADDURAH-DAOUK 2000). Die Namensgebung erfolgte aufgrund des hohen Gehaltes im Muskelfleisch („kreas“, griech. „Fleisch“). Die CK ist ein Glykoprotein, das aus zwei Untereinheiten besteht. Im Cytosol sind die Untereinheiten „M“ („muscle“) und „B“ („brain“) zu finden. Diese Untereinheiten können miteinander kombiniert auftreten, so dass beim Menschen drei Isoenzyme unterschieden werden können (KOUFEN u. STARK 2000, TAKAGI et al. 2001), im Muskel die CK-MM, im Herzen die CK-MB und im Gehirn die CK-BB. Diese Isoenzyme sind beim Tier ebenso nachweisbar, nur tritt bei diesen die CK-B nicht ins Blut über, so dass die CK als muskelspezifisch angesehen wird. In der Diagnostik kommt die CK-MM bei Muskelerkrankungen wie der Dystrophie, die CK-MB beim Myokardinfarkt und die CK-BB bei Gehirnschäden und Tumoren des Gastrointestinaltraktes zum Einsatz (TAKAGI et al. - 39 - Literatur __________________________________________________________________________________________ 2001). Im Labmagen des Rindes tritt hauptsächlich die CK-BB auf (NAURUSCHAT u. FÜRLL 2002). Außer den genannten Formen existieren auch noch zwei mitochondriale Isoenzyme: CK-“non-sarcomeric“ CK-“sarcomeric” Diese dienen als Marker von Muskelerkrankungen und werden aufgrund ihrer molekularen Größe als „Makro-CK“ bezeichnet (TAKAGI et al. 2001). Die Messung der Aktivität der CK erfolgt im Serum bei 37 °C; sie wird in U/l angegeben. Tab. 8: Physiologische Grenzwerte der Creatinkinaseaktivität bei Rindern post partum (modifiziert nach FÜRLL et al. 2002) Tage post partum Physiologische Grenzwerte der CK-Aktivität (U/l) 3 200 7 125 21 100 Die Aufgabe der CK liegt in der Übermittlung energiereicher Phosphate von Creatinphosphat auf ADP. Creatinphosphat dient vor allem in der Muskulatur als Energiespeicher, da bei hoher Beanspruchung und/oder verminderter Versorgung ATP nicht ausreichend nachgebildet werden kann. Während dieser Phase kann die Regeneration von ADP über Creatinphosphat erfolgen, das dabei entstehende Creatin kann nachfolgend über die Transphosphorylase wieder regeneriert werden (WYSS u. KADDURAH-DAOUK 2000). Die Isoenzyme der CK befinden sich im Cytosol und den Mitochondrien. Eine Freisetzung aus den Zellen und damit ein Anstieg der Aktivität im Serum erfolgt nur bei Schädigung der Zellmembranen (ISHIKAWA et al. 1997). Zum Austritt der CK kommt es auch im Rahmen von oxidativem Stress und Schädigung der Blutgefäße im Rahmen des Ischämie/Reperfusionssyndroms (MAHMOUD u. FORD 1988, REDDY et al. 2000). Zu unterscheiden ist hier zwischen alleiniger Ischämie, die keine Erhöhung der CK nach sich zieht, und Ischämie mit nachfolgender Reperfusion, die schon kurze Zeit nach Beginn der Reperfusion zur Erhöhung bzw. zur Verminderung (s. u.) der Aktivität führt (WYSS u. KADDURAHDAOUK 2000). In Untersuchungen am Hundeherz zeigte sich, dass eine Erhöhung der Aktivität der CK-MB im Plasma auf einen Infarkt am Herzen hinweist (ISHIKAWA et al. 1997). Dabei sind die Anstiege der Enzymaktivität mit histologisch sichtbaren Nekrosen am - 40 - Literatur __________________________________________________________________________________________ Herzen positiv korreliert. Bei Rindern kann auch körperlicher Stress, wie Bewegung und Transport, zu einem Anstieg der Aktivität der CK führen, der auch 24 h nach der Bewegung noch nachweisbar bleibt, während alle anderen Parameter sich wieder normalisiert haben (HILL et al. 1992). Eine Herabsetzung der Enzymaktivität kann durch Freisetzung von Radikalen erfolgen. Hierbei erfolgt durch Wasserstoffperoxid und Superoxidanionen eine dosisabhängige Verringerung der Aktivität, die im Falle des H2O2 durch die Katalase und im Falle der O2.durch die Superoxiddismutase wieder normalisiert werden kann. Die verminderte Aktivität beruht auf der Inaktivierung der Sulfhydrylgruppen der CK (GENET et al. 2000). Durch Zugabe von Radikalfängern und durch Schutz der Sulhydrylgruppen kann die Aktivität der CK bewahrt werden (KOUFEN u. STARK 2000, REDDY et al. 2000). Die Entstehung der Radikale und die Inaktivierung der CK erklären den Abfall der Enzymaktivität im Ischämie/Reperfusionssyndrom. Um den positiven Effekt der CK, die Bereitstellung von Energie, wieder herzustellen, kann reduziertes Glutathion therapeutisch eingesetzt werden (REDDY et al. 2000). Bei Übertragung der Kenntnisse auf die Ätiologie und den Verlauf der Labmagenverlagerung des Rindes ist bei den Tieren die häufig vorausgehende Schwergeburt mit Schädigung im Bereich der Muskulatur, der oxidative Stress bei Geburt und Transport (GROTH u. GRÄNZER 1975) sowie der Einfluss von Operation (AUTEFAGE et al. 2000) und Rückverlagerung des Labmagens im Sinne von Ischämie/Reperfusion als Einflussfaktoren auf die Aktivität der CK zu nennen. So ist bei Kühen mit späterer Labmagenverlagerung am dritten Tag post partum sowie bei Hypocalcämie und Gebärparese eine deutlich höhere Aktivität der CK als bei gesunden Kühen festzustellen (NAURUSCHAT u. FÜRLL 2002). 3. Eigene Untersuchungen: Tiere, Materialien und Methoden 3.1 Tiere Die untersuchten Tiere stammen aus dem Patientengut der Medizinischen Tierklinik der Veterinärmedizinischen Fakultät der Universität Leipzig. Es handelte sich dabei um zweieinhalb bis sechs Jahre alte Kühe der Rasse Holstein-Friesian, meist in einer Kreuzung mit schwarzbunten standorttypischen Tieren aus umliegenden Betrieben, die aufgrund einer DA überwiesen wurden. - 41 - Eigene Untersuchungen __________________________________________________________________________________________ 3.2 Versuchsanordnung, Gruppeneinteilung und Versuchsdurchführung Die in die Medizinische Tierklinik eingewiesenen Kühe wurden klinisch untersucht. Bestand eine DA, wurden die Tiere umgehend der operativen Rückverlagerung des Labmagens zugeführt. Dabei handelte es sich um eine Laparotomie in der rechten Flanke mit lateraler Omentopexie im Bereich des Pylorus. Zur Schmerzausschaltung im Operationsbereich erhielten die Kühe eine Leitungsanästhesie (Paravertebralanästhesie) sowie einen reversen LBlock. Die Gruppeneinteilung der Kühe erfolgte nach Art der DA (linksseitig und rechtsseitig ohne und mit Torsion; LDA, RDA sT, RDA cT) und nach dem aufsteigenden Schweregrad (LDA 2, LDA 3). Bei ausgewählten Parametern erfolgte zusätzlich die Beurteilung der Gruppen nach dem zeitlichen Abstand zum Partus der einzelnen Kühe (Tab. 9). Tab. 9: Gruppeneinteilung der Kühe nach Art und Schweregrad der DA und nach dem Abstand zum Partus ≤ 28 Tage > 28 Tage post partum post partum 37 24 13 LDA 22 17 5 LDA 2 9 8 1 LDA 3 13 9 4 RDA 15 7 8 RDA sT 4 3 1 RDA cT 11 4 7 Gruppe Anzahl der Tiere Alle Tiere Da von allen Tieren Blutproben aus der peripheren V. jugularis und Blutproben aus den viszeralen Venen des Labmagens gewonnen wurden, erfolgte zusätzlich zur Gruppeneinteilung noch die Beschreibung der Herkunft des Blutes in Form von „p“ und „lm“. Zusatzbefunde neben der bestehenden DA wurden entsprechend der Lokalisation erhoben, Befunde am Genitale beinhalteten Puerperalstörungen wie Retentio secundinarum und Endometritiden. Erkrankungen des Euters fassten Mastitiden und Verletzungen des Euters zusammen. Arthritiden, Verletzungen und Entzündungen an den Klauen bildeten die Gruppe der Erkrankungen der Gelenke und der Klauen. Außerdem wurden Erkrankungen der - 42 - Eigene Untersuchungen __________________________________________________________________________________________ Atemwege, wie Pneumonien und sonstige Erkrankungen, wie Ektoparasitosen und tumoröse Umfangsvermehrungen erfasst. 3.3 Material 3.3.1 Blutentnahme aus der V. jugularis Die Kühe erhielten vor der Operation einen Venenverweilkatheter (Splittocan, Fa. BRAUN, MELSUNGEN) in die V. jugularis, aus dem die Blutentnahmen vor, während und nach der Operation erfolgten. Vor der Operation wurden die Blutproben für die klinisch-chemischen Parameter β-HydroxyButyrat (BHB), Cholesterol, Gesamt-Bilirubin, Glutamat-Dehydrogenase (GLDH), AspartatAmino-Transferase (ASAT), Totalprotein sowie Harnstoff und die Ausgangswerte der TEACKonzentrations-Verlaufsuntersuchung gewonnen. Verwendet wurden dazu sterile Glasröhrchen ohne Zusatz zur Serumgewinnung für die klinisch-chemischen Parameter sowie heparinisierte Kunststoffröhrchen zur Plasmagewinnung für die TEAC-Konzentrationen. Unmittelbar vor der Rückverlagerung wurden nachfolgend genannte Proben entnommen: heparinisierte Blutprobe zur Gewinnung von Plasma zur Bestimmung der TEACKonzentrationen, der CK-Aktivitäten, der Lactat- und Chloridkonzentrationen heparinisierte Blutprobe zur Herstellung eines Erythrozytenpellets zur Aktivitätsbestimmung von SOD, GPX und Hämoglobinkonzentration (Hb) im peripheren Blut Nach der Rückverlagerung des Labmagens wurden nachfolgend genannte Blutproben gewonnen: acht Stunden lang halbstündige Entnahme einer heparinisierten Blutprobe aus dem Venenkatheter der V. jugularis zur Verlaufsuntersuchung der TEAC-Konzentration post operationem im peripheren Blut bei LDA zusätzlich 10 und 12 Stunden p. op., bei RDA zusätzlich 10, 12, 14 und 16 Stunden p. op. heparinisierte Blutprobe zur TEAC-Verlaufsuntersuchung 3.3.2 Blutentnahme am Labmagen - 43 - Eigene Untersuchungen __________________________________________________________________________________________ Die Blutentnahme am Labmagen erfolgte durch Punktion der Venen im Bereich des Pylorus (V. gastrica dextra, Rr. gastrici et epiploici der V. gastroepiploica dextra) unmittelbar (<1min) nach Rückverlagerung intra operationem: heparinisierte Blutprobe aus oben genannten Venen zur Gewinnung von Plasma zur Bestimmung der TEAC-Konzentration, CK-Aktivität, Lactat- und Chlorid- konzentrationen heparinisierte Blutprobe zur Herstellung von Erythrozytenpellets und zur Aktivitätsbestimmung von SOD, GPX und Hämoglobinkonzentrationen im venösen Labmagenblut 3.4 Probenaufbereitung Die Probenaufbereitung beschränkte sich bis zur Bearbeitung der Proben auf die Gewinnung von Serum, Plasma und die Herstellung von Erythrozytenpellets. 3.4.1 Gewinnung von Serum Zur Gewinnung des Blutserums wurden die Proben für zehn Minuten bei 3800 g zentrifugiert. Anschließend wurde das überstehende Serum in Eppendorfgefäße abpipettiert. Die Probenbestimmung der klinisch-chemischen Parameter fand direkt im Anschluss an die Entnahme ohne Zwischenlagerung statt. 3.4.2 Gewinnung von Plasma Nach Entnahme der heparinisierten Blutprobe wurde diese etwa 30 Minuten bei 4 °C gekühlt. Anschließend wurde sie zweimal zehn Minuten bei 1450 g zentrifugiert. Das überstehende Plasma wurde in Eppendorfgefäße pipettiert und bis zur Probenbearbeitung bei -21 °C gelagert. 3.4.3 Herstellung von Erythrozytenpellets Die heparinisierten Blutproben wurden bei 4 °C gekühlt und zentrifugiert, wie die zur Gewinnung von Plasma. Anschließend wurde auch hier das Plasma abpipettiert und der - 44 - Eigene Untersuchungen __________________________________________________________________________________________ Leukozytensaum abgesaugt. Das nun entstandene Erythrozytenpellet wurde bis zur weiteren Bearbeitung bei -21 °C gelagert. 3.5 Probenbestimmung Alle Proben wurden vor weiterer Bearbeitung bei Zimmertemperatur aufgetaut und homogenisiert. Aus dem Serum wurden die klinisch-chemischen Parameter BHB, Cholesterol, Gesamt-Bilirubin, GLDH, ASAT, Totalprotein, Harnstoff, Natrium und Kalium bestimmt. Die Messung der Lactat- und Chloridkonzentration, der CK und der TEACKonzentration erfolgte im Plasma. Die Aktivitäten der SOD und der GPX sowie die Konzentration des Hämoglobins wurden im Erythrozytenpellet bestimmt. 3.5.1 Klinisch-chemische Untersuchungsmethoden Die Bestimmung der klinisch-chemischen Parameter im Blutserum sowie die Bestimmung von CK-Aktivität, Lactat- und Chloridkonzentration im Plasma wurden im Labor der Medizinischen Tierklinik der Veterinärmedizinischen Fakultät der Universität Leipzig durchgeführt. Die für die Analysen verwandten klinisch-chemischen Untersuchungsmethoden sind in Tabelle 10 dargestellt. Zusätzlich erfolgte die Angabe der Referenzbereiche, welche zur Auswertung der Untersuchungsmethoden benutzt wurden. Die Labormethoden wurden täglichen laborinternen Kontrollen mittels Precinorm und Precipath (Fa. BOEHRINGER, MANNHEIM) sowie Kontrollseren der Fa. RANDOX, KREFELD hinsichtlich ihrer Präzision unterzogen (relative Streuung 1,3-4,1%). Zusätzlich wurden extern durchgeführte monatliche Ringkontrollen durch das Labor der Veterinärmedizinischen Reichsuniversität Utrecht, Leitung Prof. Dr. Th. WENSING, untersucht. Die Aktivitätsbestimmung der Enzyme erfolgte am Hitachi 704 bei 37°C. Tab. 10: Darstellung der klinisch-chemischen Parameter, deren Bestimmungsmethoden und dafür verwandte Geräte sowie Referenzbereiche der Parameter (KRAFT u. DÜRR 1999) - 45 - Eigene Untersuchungen __________________________________________________________________________________________ Parameter Fettstoff- Einheit Gerät Methode Referenzbereich BHB mmol/l Hitachi 704 UV-Methode 2 bis 0,6 Cholesterol mmol/l Hitachi 704 CHOD-PAP-Methode 1 2,5-4,5 U/l Hitachi 704 Optimierte Standard- 1 bis 80 1 bis 10 wechsel ASAT methode der DGKC Leberstoffwechsel GLDH U/l Hitachi 704 Enzymatischer Test Gesamt- μmol/l Hitachi 704 nach JENDRASSIK u. 2 GROFF Bilirubin Eiweiss- 2-5 TP g/l Hitachi 704 Biuret-Methode 1 68-80 Harnstoff mmol/l Hitachi 704 Kinetischer UV-Test 1 3,3-5,0 Chlorid mmol/l 1* Colometrische Titration 90-110 mmol/l KNa 2 Ionensensitive 135-157 stoffwechsel Mineral- Natrium Elektrode stoffwechsel Kalium mmol/l KNa 2 Ionensensitive 3,5-4,5 Elektrode CK U/l Hitachi 704 2* 1 bis 250 Lactat mmol/l Hitachi 704 Spektrophotometrische 3* 0,5-2,0 Messung 1 = Fa. BOEHRINGER, MANNHEIM, 2 = Fa. RANDOX, KREFELD, 1* = Ciba Corning Chloride Analyser 925, 2* = NAC aktivierte optimierte Standardmethode der DGKC, 3* = Fa. SIGMA ALDRICH, TAUFKIRCHEN 3.5.2 Superoxiddismutase (SOD) Superoxiddismutasen sind Metalloenzyme mit Anteilen von Cupfer und Zink (Cu/Zn), Eisen (Fe) oder Mangan (Mn). Sie katalysieren die Umwandlung von Superoxidanionen O2•- unter Anwesenheit von Kationen H+ zu Wasserstoffperoxid H2O2 und Sauerstoff O2. Bestimmt wurden ihre Aktivitäten mit Hilfe des Testkits SOD 525 der Fa. OXIS/BIOXYTECH, PORTLAND/USA, bestehend aus Reagenz 1 (R1), Reagenz 2 (R2) und Puffer. Hierbei enthält R1 Superoxidanionen in wässriger alkalischer Lösung, die in diesem Fall durch im Erythrozytenpellet vorhandene Cu-Zn-SOD zu einem Chromophor umgewandelt werden, das am Beckman-Photometer bei 525 nm die höchste Absorptionsrate besitzt. Eventuell vorhandene Fe- und Mn-SOD wurden vor der Messung durch Zugabe eines - 46 - Eigene Untersuchungen __________________________________________________________________________________________ Extraktionsgemisches aus Ethanol und Chloroform inaktiviert. Interferenzen im Erythrozytenpellet durch Mercaptane, wie reduziertes Glutathion wurden durch Zugabe von R2 in einer Alkylierungsreaktion beseitigt. Die Messungen fanden am Beckman-UVSpektralphotometer DU 640 des Institutes für Pharmakologie, Pharmazie und Toxikologie der Veterinärmedizinischen Fakultät der Universität Leipzig statt. Im Rahmen der Probenvorbereitung wurden die Pellets bei Zimmertemperatur langsam aufgetaut und gründlich gemischt. 100 µl des Pellets und 400 l etwa 3 °C kaltes Aqua bidest. wurden gemischt. Darauf folgte die Zugabe von 400 µl etwa 3 °C kaltem Extraktionsgemisch zu 250 µl des Erythrozytenlysates. Nach weiterer gründlicher Mischung wurde die Probe bei 3000 g und 4 °C zehn Minuten zentrifugiert. Schließlich wurde der Überstand abpipettiert, der für die Aktivitätsbestimmung maßgeblich ist. Zur Messung wurden jeweils 40 µl Aqua bidest. (zur Bestimmung des Leerwertes Vs; s. u.) bzw. 40 µl des Überstandes (Probenflüssigkeit) mit 900 µl Puffer des Testkits SOD-525 von 37 °C gemischt. Danach erfolgte die Zugabe von 30 µl R2 und eine einminütige Inkubation bei 37 °C, bevor die Probe nach Zugabe von 30 µl R1 sofort im Beckman-Photometer bei 525 2 nm über eine Minute im Fünf-Sekunden-Takt gemessen wurde. Gemessen wird der zeitliche Verlauf der Umwandlung des O2•¯. Als bestehende Aktivität wird der Bereich definiert, in dem eine gleichbleibende Menge der Umwandlung, dargestellt durch konstante Steigung in der Absorptionsänderung in Abhängigkeit von der Zeit, Absorption bei 525 nm stattfindet (Abb. 13). 0,45 0,4 0,35 0,3 0,25 0,2 0,15 0,1 0,05 0 0,31 0,13 Zeit (Minuten) 0 Abb. 13: 1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 Beispiel-Kurve: Absorption zur Zeit im Bereich gleichbleibender/linearer Steigung und daraus resultierendem Vc bzw.Vs Die Umwandlung durch die Probe (Vc) wird im Verhältnis zur Umwandlung durch Wasser (Vs) angegeben. Die Steigung berechnet sich wie folgt. (Y2-Y1) (0,31-0,13) ----------- = -------------- = 0,6 - 47 - Eigene Untersuchungen __________________________________________________________________________________________ (X2-X1) (0,7-0,4) Das Verhältnis Vs zu Vc verhält sich zur Aktivität der SOD wie nachfolgend erläutert. (SOD) Vs/Vc= 1 + ----------------- x (SOD) + Vs = Messergebnis der Probe, die SOD enthält (A/min) Vc = Durchschnitt von zwei Leerwerten (A/min) SOD = SOD-Aktivität der Probe, ausgedrückt in SOD-252 Units = dimensionsloser Koeffizient Erfahrungswert: = 0,073 = Koeffizient mit der Einheit SOD-252 Units hier: = 0,93 SOD Units Löst man die Formel nach der SOD auf, lässt sich die Aktivität berechnen. 0,93 x ( Vs/Vc - 1) (SOD) = --------------------------1,073 x 0,073 x (Vs/Vc) Nach Multiplikation dieser Aktivität mit der Verdünnung des Pellets ergibt sich die Aktivität der Probe in Units pro Liter Erythrozytenpellet (U/l). Um die Messungen für alle Proben bzw. Erythrozytenpellets zu standardisieren, wurde diese Aktivität in Bezug auf das in der Probe vorhandene, gesondert bestimmte Hämoglobin (Hb) angegeben, in U(SOD)/g Hb. Vor Beginn der eigenen Untersuchung wurden Zehnfachbestimmungen durchgeführt. Zum Zeitpunkt dieser Messreihe lag die relative Streuung für die SOD-Aktivitäten bei 3,95 %. 3.5.3 Glutathionperoxidase (GPX) Die GPX wurde nach der UV-Methode nach PAGLIA und VALENTINE (1938) mit Hilfe des Testkits der Firma RANDOX RANSEL KREFELD im Erythrozytenpellet bestimmt. Die Bestimmung der GPX-Aktivitäten in einem bestimmten Substratvolumen erfolgte hierbei durch Absorptionsabnahme nach Zugabe von Cumenhydroperoxid und Reagenz am kinetischen Meßplatz MKE 5081 der Fa. Eppendorf der Medizinischen Tierklinik der Veterinärmedizinischen Fakultät der Universität Leipzig bei 340 nm und 37 °C. Die GPX katalysiert die Oxidation von Glutathion (GSH) zu Cumenhydroperoxid (R-OOH). Dabei entsteht oxidiertes Glutathion (GSSG). Unter Anwesenheit der Glutathionreduktase - 48 - Eigene Untersuchungen __________________________________________________________________________________________ und Nikotinamidadenindinukleotidphosphat (NADPH+) wird dieses GSSG wieder zu Glutathion (GSH) reduziert, wobei NADPH + H+ als Reduktionsmittel dient. Es entsteht NADP+. Daraus resultiert schließlich die Absorptionsabnahme im UV-Meßbereich. Der Reaktionsablauf wird in Abb. 14 schematisiert dargestellt. 2GSH (Reagenz) GPX (Probe) + R-OOH ---------------------- (Cumenhydroperoxid) Glutathionreduktase (Reagenz) + GSSG + NADPH+H ------------------------- (Reagenz) GSSG + ROH + H2O 2GSH NADP+ + Abb.14: Reaktionsablauf der Aktivitätsbestimmung der GPX (nach RANSEL-GlutathionPeroxidase) Zur Vorbereitung Zimmertemperatur der Messung aufgetaut und wurden 30 die sec Erythrozytenpellets gemischt. Danach langsam erfolgte die bei erste Probenverdünnung mit der Verdünnungslösung (100 µl + 3000 µl) mit gründlichem Mischen. Daraufhin wurde die zweite Probenverdünnung mit Verdünnungslösung 1 + 9 durchgeführt und wiederum gemischt. In der Zwischenzeit wurden Reagenz, Kontrolle und Cumenhydroperoxid nach Vorschrift gelöst. Die eigentliche Messung erfolgte in Banden mit jeweils sechs Küvetten. Dabei nahmen in jedem zweiten Durchgang Aqua bidest. (H2O) und Kontrolle die ersten beiden Küvetten in Anspruch. Vor der Messung wurden jeweils 20 µl der zweiten Probenverdünnung in die Küvetten vorgelegt. Daraufhin folgte im Fünf-SekundenTakt die Zugabe von 1000 µl Reagenz und ein Mischen von fünf Sekunden. Eine Minute nach Zugabe des ersten Reagenz wurde mit der Zugabe von 40 µl Cumenhydroperoxid im FünfSekunden-Takt begonnen, woraufhin die Proben dann wieder fünf Sekunden gemischt wurden. Zwei Minuten nach Beginn der Reagenzzugabe erfolgte die Messung im oben genannten Photometer, auch hier im Fünf-Sekunden-Takt. Dabei wurde die Absorptionsänderung in drei Phasen über eineinhalb Minuten verfolgt, woraus schließlich die Differenzen und aus den Differenzen der Mittelwert gebildet wurden. Die GPX-Aktivität, angegeben in U/l, berechnet sich dann wie im Folgenden erläutert. GPX (U/l) = 8576 x VDF 1 x VDF 2 x (E - E0) - 49 - Eigene Untersuchungen __________________________________________________________________________________________ VDF 1 : - Verdünnungsfaktor der ersten Verdünnung hier: VDF 1 = 31 (100 + 3000) VDF 2 : - Verdünnungsfaktor der zweiten Verdünnung hier: VDF 2 = 10 (1 + 9) E : - Mittelwert der Extinktionsänderung der Probe E0 : - Mittelwert der Extinktionsänderung von Wasser Leerwert Um eine feste Bezugsgröße zu haben und um Ungleichmäßigkeiten des Erythrozytenpellets auszugleichen, wird die GPX auf die im Erythrozytenpellet vorhandene Konzentration an Hämoglobin bezogen (s. u.). Die GPX-Aktivität pro Gramm Hämoglobin im Erythrozytenpellet berechnet sich wie folgt: GPX ( U/g Hb) = GPX (U/l) x 0,6207 : ( 10 x Hb (mmol/l)) Vor Beginn der eigenen Untersuchung wurden Zehnfachbestimmungen durchgeführt. Zum Zeitpunkt dieser Messreihe lag die relative Streuung für die GPX-Aktivitäten bei 3,25%. 3.5.4 TEAC-Konzentration Die Bestimmung der „Trolox equivalent antioxidative capacity“ -Konzentration im Blutplasma erfolgte mittels der Extinktionsänderung einer ABTS-Gebrauchslösung durch die Probe im Rahmen einer photometrischen Messung. Die Messungen wurden am UVSpektralphotometer DU 640 der Fa. BECKMAN, KREFELD auf der Grundlage der von MILLER et al. (1996) beschriebenen Methode im Labor des Instituts für Pharmakologie, Pharmazie und Toxikologie der Veterinärmedizinischen Fakultät der Universität Leipzig durchgeführt. Die Gebrauchslösung aus 5 mM ABTS (2,2’-Azino-bis (3-ethylbenz-thiazoline-6sulphonic acid) diammonium salt (Fa. SIGMA-ALDRICH, TAUFKIRCHEN)) mit MnO2 (Manganese Dioxide, Activated, Fa. SIGMA-ALDRICH, TAUFKIRCHEN) und PBS-Puffer (5mM, pH 7,4) besitzt ein ABTS+-Kation und eine tiefgrüne Farbe. Wird das Kation durch die antioxidative Fähigkeit von TROLOX (6-Hydroxy-2,5,7,8-tetramethylchroman-2- carbonsäure) (Fa. SIGMA-ALDRICH) oder der Probe weggefangen („scavenge“), erfolgt eine Aufhellung der Gebrauchslösung, die photometrisch gemessen werden kann. Dabei wird - 50 - Eigene Untersuchungen __________________________________________________________________________________________ zuerst eine Eichkurve erstellt, indem TROLOX in unterschiedlichen Konzentrationen einer 0,0-2,5 mM/l-Lösung in einer Menge von 10 µl zu 990 µl einer ABTS+-Gebrauchslösung, die am Beckman-Photometer bei 30 °C eine Extinktion von 0,7 0,02 nm besitzt, zugegeben wird, 30 Sekunden auf dem Vortex gemischt und nach einer Minute im Photometer gemessen wird. So erhält man für jede Troloxkonzentration eine bestimmte Extinktionsänderung der Gebrauchslösung. Trägt man die Konzentrationen von TROLOX auf der X-Achse und die Werte der Extinktionsänderung auf der Y-Achse ab, so erhält man eine Gerade der Gleichung: y = a x + b (a; b: Variable). Mit Hilfe dieser Gleichung wird es nach Umstellung in die Form x = ( y - b ) : a möglich, jede Probe, die langsam aufgetaut, gemischt und genauso wie zuvor das TROLOX gemessen wurde, einer bestimmten Troloxkonzentration zuzuordnen, einem Trolox-Äquivalent also. Die TEAC-Konzentration wurde angegeben in µmol/l. Vor Beginn der eigenen Untersuchung wurden Zehnfachbestimmungen durchgeführt. Zum Zeitpunkt dieser Messreihe lag die relative Streuung für die TEAC-Konzentrationen bei 2,11%. 3.5.5 Hämoglobin (Hb) Die Bestimmung der Hämoglobinkonzentration im Erythrozytenpellet wurde nach langsamem Auftauen der Pellets bei Zimmertemperatur und deren Verdünnung (1:2) mit Aqua bidest. am Zellcounter M901V der Fa. MÖLAB HILDEN nach der Cyanhämoglobinmethode im Labor der Medizinischen Tierklinik durchgeführt. Die Angabe der Konzentration des Hämoglobins erfolgte standardmäßig in mmol/l . 3.6 Statistische Auswertung Die statistische Auswertung erfolgte mit Hilfe des Computerprogramms „SPSS 10.0“. Dem Test auf Normalverteilung liegen die Verfahren von Kolmogorov-Smirnov und Shapiro-Wilk zugrunde. Für die jeweiligen Parameter wurden insgesamt und in den Gruppen Mittelwert und Standardabweichung bzw. bei fehlender Normalverteilung jeweils Medianwert, 1. und 3. Quartil berechnet. Der U-Test nach Mann-Whitney wurde als nicht-parametrischer Test genutzt, um zu prüfen, ob zwei unabhängige Stichproben aus der gleichen Grundgesamtheit stammen. Dies ist besonders beim Vergleich der Proben aus dem peripheren Blut mit den Proben aus den Venen des Labmagens von Bedeutung. Die Korrelationen in und zwischen den Gruppen wurden nach Spearman berechnet. Signifikante Unterschiede wurden in den graphischen Darstellungen durch Pfeile zwischen den betreffenden Gruppen gekennzeichnet. - 51 - Eigene Untersuchungen __________________________________________________________________________________________ Zur Auswertung der TEAC-Konzentrations-Verläufe wurde die Einzeldarstellung der Tiere gewählt, um die individuellen Abläufe zu veranschaulichen. 4. Ergebnisse Die Tiere der vorliegenden Untersuchung wurden in sechs Gruppen eingeteilt. Neun Tiere bildeten die Gruppe der LDA 2, die Gruppe der LDA 3 bestand aus 13 Tieren, zusammengefasst stellten sie die Gruppe aller LDA (n=22) dar. Die Tiere mit rechtsseitiger - 52 - Ergebnisse __________________________________________________________________________________________ DA wurden nach Verlagerungen ohne Torsion (RDA sT; n=4) und Verlagerungen mit Torsion (RDA cT; n=11) unterteilt. Zusammen bildeten sie die Gruppe aller Tiere mit RDA, die 15 Tiere beinhaltete (Tab. 9). Bei der klinischen Untersuchung fiel auf, dass die Tiere mit LDA außer Indigestionserscheinungen weitestgehend unauffällig waren, während die Tiere mit RDA eine deutlichere Beeinflussung des Allgemeinbefindens und des Kreislaufes (verminderte Hautelastizität, Enophtalmus, verlängerte Kapillarfüllungszeit, vermindertes Blutangebot der V. jugularis) aufwiesen. Elf Kühe zeigten erhöhte Pulsfrequenzen (30 %), innerhalb der LDA sechs Kühe (27 %), innerhalb der RDA fünf Kühe (33 %) (Tab.11). Tab. 11: Pulsfrequenzen, Atmungsfrequenzen und Rektaltemperaturen innerhalb der Gruppen der linksseitigen Verlagerungen 2. (LDA 2) und 3. Grades (LDA 3) sowie der rechtsseitigen Verlagerungen ohne (RDA sT) und mit Torsion (RDA cT) (Median, 1. und 3. Quartil) Gruppen LDA 2 (n=9) LDA 3 (n=13) RDA sT (n=4) RDA cT (n=11) Pulsfrequenz (Schläge pro Minute) 72 (66; 80) 68 (56; 76) 80 (72; 101) 66 (59; 77) Atmungsfrequenz (Züge pro Minute) 24 (22; 32) 24 (22; 32) 22 (20; 27) 20 (20; 24) 39,4 (39,3; 39,5) 38,9 (38,6; 39,7) Rektaltemperatur (°C) 39,1 38,7 (38,8; 39,4) (38,4; 38,9) Neben der Dislocatio abomasi wurden zusätzliche Erkrankungen des Genitaltraktes (59 % aller Tiere) in Form eines gestörten Puerperiums, Mastitiden (16 % aller Tiere), Klauen- und Gelenkserkrankungen (38 % aller Tiere), Erkrankungen der Atemwege (8 % aller Tiere) sowie sonstige Erkrankungen (11 % aller Tiere) wie Ektoparasitenbefall, Dekubitus und Umfangsvermehrungen festgestellt (Tab. 12). Dabei zeigten Kühe mit LDA (73 %) signifikant häufiger (p<0,05) Erkrankungen des Genitale als Kühe mit RDA (40 %). Tab. 12: Anzahl der Kühe, die jeweils zusätzliche Erkrankungen des Genitale, des Euters, der Klauen und Gelenke, der Atemwege oder sonstige Erkrankungen aufwiesen Erkrankungen des LDA 2 LDA 3 (n=9) (n=13) (n=4) (n=11) 8 8 3 3 - 53 - RDA sT RDA cT Ergebnisse __________________________________________________________________________________________ Genitale Erkrankungen des Euters Erkrankungen der Klauen und Gelenke Erkrankungen der Atemwege Sonstige 2 2 1 1 5 5 2 2 1 2 2 Erkrankungen 1 Die Abkalbung lag bei 70 % aller Tiere weniger als 28 Tage zurück. Bei den Tieren mit LDA befanden sich 73 % der Tiere innerhalb der ersten vier Wochen post partum, bei den RDA lag die Geburt bei 33 % auch länger zurück. Der Abstand zum Abkalbedatum und das Alter der Tiere werden bei der Diskussion der einzelnen Parameter berücksichtigt. Während des Zeitraumes dieser Studie (bis 16 h post operationem) traten bei den operierten Tieren keine Komplikationen auf. Bereits am ersten Tag nach der Operation zeigten 23 Kühe eine vollständige Aufnahme der Futterration und Wasseraufnahme, physiologische Pansenbewegungen (8-12 in 5 Minuten), Wiederkäuen und Kotabsatz. Bei sechs Kühen war dies bis zum dritten Tag post operationem der Fall, bei vier Kühen dauerte es bis zu einer Woche nach der Operation. Vier Kühe mussten nach ein bis zwei Wochen wegen ihres schlechten klinischen Zustandes euthanasiert werden. Sie stammten aus den Gruppen LDA 2 (n=2), LDA 3 (n=1) und RDA cT (n=1). Bei ihrer nachfolgenden pathologischen Untersuchung wurde bei allen vier Tieren eine hochgradige degenerative Leberverfettung festgestellt. Hinzu kamen Thromben der Aa. pulmonales, Peritonitiden sowie Ulzerationen des Labmagens und des Duodenums. 4.1 Klinisch-chemische Parameter des Mineralstoffwechsels - 54 - Fett-, Leber- und Eiweiss- und Ergebnisse __________________________________________________________________________________________ Die BHB-Konzentrationen der Gruppen alle Tiere, alle LDA, LDA 2, LDA 3 sowie RDA sT lagen über dem physiologischen Bereich (< 0,6 mmol/l) (Tab. 13), nur die RDA cT zeigten keine erhöhten Konzentrationen. Die Cholesterolkonzentrationen befanden sich in allen Gruppen außer den RDA sT unterhalb des Referenzbereiches (< 2,5 mmol/l). Die Gruppen der Tiere mit RDA hatten höhere Cholesterolkonzentrationen als die Gruppen mit LDA (Tab. 13), die Cholesterolkonzentrationen der LDA 3 waren signifikant niedriger als die der RDA cT (p<0,01). Die Bilirubinkonzentrationen überstiegen bei allen Tieren den physiologischen Bereich (2-5 μmol/l), bei 54 % lagen sie sogar über 15 μmol/l, die Konzentrationen der Gruppe der RDA cT lagen niedriger als die der anderen Gruppen (Tab. 13). Die Aktivitäten der GLDH waren in allen Gruppen erhöht (>50 U/l), zwischen den Gruppen konnten aber keine statistisch gesicherten Unterschiede festgestellt werden (Tab. 13). Die Gruppen der Kühe mit LDA 2 und RDA cT zeigten die höchsten Aktivitäten. Die Aktivitäten der ASAT überschritten ebenfalls in allen Gruppen den Referenzbereich (<80 U/l). Die Tiere der Gruppen LDA 2 sowie RDA sT besaßen höhere Aktivitäten der ASAT als die Gruppe der LDA 3 (Tab. 13). Die Konzentration des Totalproteins (TP) befand sich in allen Gruppen innerhalb des physiologischen Bereiches (68-80 g/l) (Tab. 13), nur 22 % aller Tiere lagen darüber. Die Harnstoffkonzentrationen der Gruppen alle RDA und RDA cT lagen über dem Referenzbereich (3,3-5,0 mmol/l) (Tab. 13), von den Tieren mit LDA hatten nur 35 % erhöhte Harnstoffkonzentrationen, von den Kühen mit RDA cT 73 %. Die Kaliumkonzentrationen lagen nur in der Gruppe aller LDA und der LDA 2 im Referenzbereich (3,5-4,5 mmol/l), in allen anderen Gruppen darunter, die Gruppen der RDA besaßen niedrigere Kaliumkonzentrationen, die Gruppe der RDA cT wies signifikant geringere Konzentrationen als die Gruppe der LDA 2 auf (p<0,01) (Tab. 13). Die Natriumkonzentrationen der Gruppen alle RDA und RDA cT waren erniedrigt, die Gruppen der Tiere mit LDA und RDA sT befanden sich im Referenzbereich (135-157 mmol/l) (Tab. 13). Tab. 13: Darstellung der klinisch-chemischen Parameter β-Hydroxybutyrat (BHB), Cholesterol (Chol.), Gesamt-Bilirubin, Glutamat-Dehydrogenase (GLDH), Aspartat-Amino-Transferase - 55 - Ergebnisse __________________________________________________________________________________________ (ASAT), Totalprotein (TP), Harnstoff, Kalium und Chlorid im peripheren Blut vor Operationsbeginn (Median, 1. und 3. Quartil) alle Tiere alle LDA n=37 BHB LDA 3 n=9 n=13 alle RDA RDA sT n=15 n=4 RDA cT n=11 1,15 1,89 1,91 1,26 (0,32; 2,29) (0,82; 2,73) (1,84; 5,02) (0,67; 2,08) 0,32 (0,2; 1,04) 1,04 0,24 (0,74; 2,26) (0,19; 0,81) 1,67 (1,26; 2,2) 1,31 (0,99; 1,6) 2,29 (1,6; 3,5) 2,6 (1,85; 3,31) 2,29 (1,6; 3,5) Gesamt17,3 18,95 26,0 17,3 Bilirubin (10,9; 28,5) (13,4; 29) (14,4; 32,2) (13,3; 23,7) (μmol/l) 10,9 (5,4; 21) 20,1 (16,6; 23,4) 8,4 (4,5; 19,5) (mmol/l) Chol. (mmol/l) GLDH (U/l) ASAT (U/l) TP (g/l) Harnstoff 1,49 1,72 (1,16; 1,73) (1,56; 2,04) 72 (34; 139) 66 (34; 112) 112 (98; 169) 49 (30; 70) 96 (34; 218) 96 (53; 251) 117 (39; 199) 179 (137; 303) 150 (132; 277) 327 (238; 377) 141 (100; 161) 266 (139; 300) 301 (219; 373) 249 (145; 292) 74,6 73,8 76,8 70,6 74,7 74,9 74,6 (68,2; 79,6) (67,5; 79,4) (68,3; 78,8) (66,9; 79,6) (70,0; 79,7) (74,7; 75,6) (68,3; 80,8) 4,8 (4,0; 6,9) 4,2 (3,6; 5,6) 4,1 (3,7; 4,5) 4,9 (3,6; 6,3) 6,9 (4,5; 12,1) 4,4 (4,1; 7,4) 7,5 (5,3; 12,1) (mmol/l) 3,3 (3,0; 3,6) 3,5 (3,0; 3,7) 3,7 (3,6; 3,7) 3,3 (3,0; 3,5) 3,1 (3,0; 3,4) 3,3 (3,0; 3,4) 3,0 (2,9; 3,4) Natrium (mmol/l) 137 (132; 139) 138 (135; 139) 139 (137; 140) 138 (134; 139) 133 (129; 137) 137 (133; 138) 133 (129; 134) (mmol/l) Kalium 4.2 n=22 LDA 2 Parameter im peripheren und abomasalen Blut Nach dem Test auf Normalverteilung von Kolmogorov-Smirnov waren periphere Hämoglobinkonzentration, periphere TEAC-Konzentration und periphere GPX-Aktivität der einzelnen Probanden nach der Signifikanzkorrektur nach Lillefors normal verteilt. Der Test nach Shapiro-Wilk zeigte eine Normalverteilung von peripherer CK-Aktivität und der aus dem Blut der Venen des Labmagens bestimmten CK-Aktivität, TEAC-Konzentration und GPX-Aktivität. Da bei beiden Tests nicht alle Parameter normal verteilt waren, wurden zur Überprüfung auf Korrelationen für alle Parameter nicht-parametrische Tests verwendet. Zur Darstellung der Ergebnisse kamen aufgrund dieser nicht normalverteilten Daten der Medianwert mit erstem und drittem Quartil. Mit Hilfe des nicht-parametrischen Tests nach Mann-Whitney wurden diese Unterschiede auf ihre Signifikanz überprüft. - 56 - Ergebnisse __________________________________________________________________________________________ Eine aufgegliederte Darstellung der Messergebnisse zu den Graphiken befindet sich im Anhang . 4.2.1 Vergleich der Chloridkonzentrationen im peripheren und abomasalen Blut Die Chloridkonzentrationen aller Tiere lagen zwischen 83 und 102 mmol/l. Zwischen den Entnahmestellen des Blutes bestanden in den Gruppen keine signifikanten Unterschiede. Die Gruppe der RDA cT besaß die niedrigste Chloridkonzentration (Abb. 15). Chlorid-Konzentration (mmol/l) **=1,0 110 **=1,0 **=1,0 100 **=1,0 90 80 70 60 50 40 1 alle Tiere Abb. 15: alle LDA LDA 2 LDA 3 alle RDA RDA sT RDA cT Vergleich der Chloridkonzentrationen (p) (grün) zu den Chloridkonzentrationen (lm) (rot) innerhalb der Gruppen aller Tiere (alle Tiere), der Tiere mit linksseitiger Verlagerung (alle LDA), davon mit Verlagerung zweiten (LDA 2) und dritten (LDA 3) Grades und der Tiere mit rechtsseitiger Verlagerung (alle RDA), davon ohne (RDA sT) und mit (RDA cT) Torsion (Median, 1.- 3. Quartil; **: Korrelationen der Chloridkonzentrationen (p) zu den Chloridkonzentrationen (lm) mit p ≤ 0,01) 4.2.2 Lactatkonzentration 4.2.2.1 Lactatkonzentrationen im peripheren Blut Die Lactatkonzentrationen aller Tiere bewegten sich im peripheren Blut zwischen 1,49 und 6,14 mmol/l. In der Gruppe mit RDA cT bestanden im Vergleich zu den Tieren mit LDA 2 (p<0,05) und mit LDA 3 (p<0,01) signifikant höhere Lactatkonzentrationen (Abb. 16). - 57 - Ergebnisse __________________________________________________________________________________________ Lactatkonzentration (mmol/l) 9 8 7 6 5 4 3 2 1 0 alle LDA 2 LDA 3 LDA Abb. 16: 1 alle RDA RDA sT RDA cT Gruppenvergleich der Lactatkonzentrationen (p): Vergleich der Lactatkonzentrationen im peripheren Blut bei allen Tieren mit linksseitiger Labmagenverlagerung (alle LDA), aufgeteilt in zweiten (LDA 2) und dritten (LDA 3) Grad mit den Lactatkonzentrationen aller Tiere mit rechtsseitiger Labmagenverlagerung (alle RDA), aufgeteilt in Verlagerung ohne (RDA sT) und mit (RDA cT) Torsion (Median, 1.- 3. Quartil) 4.2.2.2 Lactatkonzentrationen im abomasalen Blut Die Lactatkonzentrationen aller Tiere im abomasalen Blut betrugen zwischen 1,10 und 6,04 mmol/l. In der Gruppe RDA cT lagen im Vergleich zu den Tieren mit LDA 2 und LDA 3 die signifikant höheren Lactatkonzentrationen vor (p<0,01; Abb. 17). Lactatkonzentration (mmol/l) 10 8 6 4 2 0 alle LDA LDA 2 LDA 3 Abb. 17: 1 alle RDA RDA sT RDA cT Gruppenvergleich der Lactatkonzentrationen (lm): Vergleich der Lactatkonzentrationen im abomasalen Blut bei allen Tieren mit linksseitiger Labmagenverlagerung (alle LDA), aufgeteilt in zweiten (LDA 2) und dritten (LDA 3) Grad, mit den Lactatkonzentrationen aller Tiere mit rechtsseitiger Labmagenverlagerung (alle RDA), aufgeteilt in Verlagerung ohne (RDA sT) und mit (RDA cT) Torsion (Median, 1.- 3. Quartil) 4.2.2.3 Vergleich der Lactatkonzentrationen im peripheren und abomasalen Blut - 58 - Ergebnisse __________________________________________________________________________________________ Zwischen den Entnahmestellen des Blutes bestanden in der Lactatkonzentration keine signifikanten Unterschiede (Abb. 18). Lactatkonzentration (mmol/l) 9 8 7 6 5 4 3 2 1 0 **=1,0 **=1,0 **=1,0 alle Tiere Abb. 18: **=1,0 alle LDA LDA 2 1 3 LDA alle RDA RDA sT RDA cT Vergleich der Lactatkonzentrationen (p) (grün) zu den Lactatkonzentrationen (lm) (rot) innerhalb der Gruppen aller Tiere (alle Tiere), der Tiere mit linksseitiger Verlagerung (alle LDA), davon mit Verlagerung zweiten (LDA 2) und dritten (LDA 3) Grades und der Tiere mit rechtsseitiger Verlagerung (alle RDA), davon ohne (RDA sT) und mit (RDA cT) Torsion (Median, 1.- 3. Quartil; **: Korrelationen der Lactatkonzentrationen (p) zu den Lactatkonzentrationen (lm) mit p ≤ 0,01) 4.2.3 Creatinkinase-Aktivität (CK-Aktivität) 4.2.3.1 Creatinkinase-Aktivitäten im peripheren Blut (CK-Aktivität (p)) Die Minimal- und Maximalaktivitäten der CK (p) aller Tiere lagen zwischen 94,2 und 739,9 U/l. Der Median betrug 380 U/l, erstes bzw. drittes Quartil 213,8 bzw. 793,7 U/l (Abb.19). Beim Gruppenvergleich der einzelnen Medianwerte zeigte sich, dass die CK-Aktivitäten der Gruppe der LDA 2 signifikant höher waren als die CK-Aktivitäten der Gruppe der LDA 3 (p<0,05). - 59 - Ergebnisse __________________________________________________________________________________________ CK-Aktivität (U/l) 1200 1000 800 600 400 200 0 Abb. 19: alle LDA 2 LDA LDA 3 1 alle RDA RDA sT RDA cT Gruppenvergleich der CK-Aktivitäten (p): Vergleich der CK-Aktivitäten im peripheren Blut bei allen Tieren mit linksseitiger Labmagenverlagerung (alle LDA), aufgeteilt in zweiten (LDA 2) und dritten (LDA 3) Grad, mit den CK-Aktivitäten aller Tiere mit rechtsseitiger Labmagenverlagerung (alle RDA), aufgeteilt in Verlagerung ohne (RDA sT) und mit (RDA cT) Torsion (Median, 1.- 3. Quartil) 4.2.3.2 Creatinkinase-Aktivitäten im abomasalen Blut (CK-Aktivität (lm)) Die Aktivitäten der CK (lm) lagen zwischen 55,9 und 1666,5 U/l. Der Median aller Proben lag bei 383,8 U/l, erstes und drittes Quartil betrugen 189,1 und 902,8 U/l (Abb. 20). Im Vergleich der einzelnen Gruppen zeigte sich, dass die Aktivitäten der CK in der Gruppe der LDA 3 signifikant niedriger waren als in den Gruppen der LDA 2 (p<0,01), RDA sT (p<0,05) und RDA cT (p<0,05). CK-Aktivität (U/l) 1200 1000 800 600 400 200 0 alle LDA LDA 2 LDA 3 1 alle RDA RDA sT RDA cT Abb. 20: Gruppenvergleich der CK-Aktivitäten (lm): Vergleich der CK-Aktivitäten im abomasalen Blut bei allen Tieren mit linksseitiger Labmagenverlagerung (alle LDA), aufgeteilt in zweiten (LDA 2) und dritten (LDA 3) Grad, mit den CK-Aktivitäten aller Tiere mit rechtsseitiger Labmagenverlagerung (alle RDA), aufgeteilt in Verlagerung ohne (RDA sT) und mit (RDA cT) Torsion (Median, 1.- 3. Quartil) 4.2.3.3 Vergleich der Creatinkinase-Aktivitäten im peripheren und abomasalen Blut - 60 - Ergebnisse __________________________________________________________________________________________ Zwischen den Aktivitäten der CK aus dem peripheren und dem abomasalen Blut der einzelnen Gruppen bestanden keine signifikanten Unterschiede (Abb. 21). Tendenziell waren die CK-Aktivitäten in der Gruppe der LDA 3 im peripheren Blut höher als im abomasalen. CK-Aktivität(U/l) 1200 **=0,98 **=0,92 **=0,92 **=1,00 1000 800 **=0,89 **=0,87 600 **=0,73 400 200 0 alle Tiere alle LDA LDA 2 1 LDA 3 alle RDA RDA sT RDA cT Abb. 21: Vergleich der CK-Aktivitäten (p) (grün) zu den CK-Aktivitäten (lm) (rot) innerhalb der Gruppen aller Tiere (alle Tiere), der Tiere mit linksseitiger Verlagerung (alle LDA), davon mit Verlagerung zweiten (LDA 2) und dritten (LDA 3) Grades und der Tiere mit rechtsseitiger Verlagerung, davon ohne (RDA sT) und mit (RDA cT) Torsion (Median, 1.- 3. Quartil; **: Korrelationen der CK-Aktivitäten (p) zu den CK-Aktivitäten (lm) mit p ≤ 0,01) 4.2.4 Superoxiddismutase-Aktivität (SOD-Aktivität) 4.2.4.1 Superoxiddismutase-Aktivitäten im peripheren Blut (SOD-Aktivität (p)) Die auf die Hämoglobinkonzentration bezogene Aktivität der SOD (p) lag minimal bei 1371 U/g Hb und maximal bei 3979 U/g Hb. Der Medianwert von allen Tieren betrug 2384 U/gHb, erstes und drittes Quartil 2075 U/g Hb und 2517 U/g Hb (Abb. 22). Bei der Überprüfung auf signifikante Unterschiede der SOD-Aktivitäten im peripheren Blut zwischen den Gruppen wurden höhere Aktivitäten aller Tiere mit LDA im Vergleich zu allen Tieren mit RDA deutlich (p<0,05). Auffällig wurde dies vor allem zwischen der Gruppe mit LDA 2 und der Gruppe mit RDA sT (p<0,05) sowie zwischen der Gruppe mit LDA 2 und der Gruppe mit RDA cT (p<0,01). Entsprechend lagen auch die SOD-Aktivitäten aller Tiere mit LDA im Vergleich zu den Tieren mit RDA cT signifikant höher (p<0,05). - 61 - Ergebnisse __________________________________________________________________________________________ SOD-Aktivität (U/g Hb) 4000 3500 3000 2500 2000 1500 alle LDA 2 LDA Abb. 22: LDA 3 1 alle RDA RDA sT RDA cT Gruppenvergleich der SOD-Aktivitäten (p): Vergleich der SOD-Aktivitäten im peripheren Blut bei allen Tieren mit linksseitiger Labmagenverlagerung (alle LDA), aufgeteilt in zweiten (LDA 2) und dritten (LDA 3) Grad, mit den SOD-Aktivitäten aller Tiere mit rechtsseitiger Labmagenverlagerung (alle RDA), aufgeteilt in Verlagerung ohne (RDA sT) und mit (RDA cT) Torsion (Median, 1.- 3. Quartil) Im Vergleich der Parameter im peripheren Blut in der Gruppe aller Tiere bestand eine negative Korrelation mit p<0,01 zwischen Hämoglobin und SOD-Aktivitäten (r = -0,44), die aus der Angabe der SOD-Aktivität in „Units je Gramm Hämoglobin“ (U/g Hb) resultierte. Diese Korrelation trat auch in der Gruppe aller Tiere mit LDA (r = -0,56) und in der Gruppe der Tiere mit LDA 3 (r = -0,81) mit p<0,01 auf. Des weiteren waren die SOD-Aktivitäten in der Gruppe der Tiere mit RDA sT negativ mit den TEAC-Konzentrationen im peripheren (r = -1,00) und abomasalen Blut (r = -1,00) korreliert (p<0,001). 4.2.4.2 Superoxiddismutase-Aktivitäten im abomasalen Blut (SOD-Aktivität (lm)) Die SOD-Aktivitäten lagen zwischen 1386 und 3507 U/g Hb auf. Der Median aller Werte betrug 2164 U/gHb, erstes bzw. drittes Quartil erstreckten sich bis 1888 bzw. 2284 U/g Hb (Abb. 23). Alle Tiere mit LDA und die Gruppe der LDA 2 lagen signifikant höher als die Gruppe der RDA sT (p<0,05). - 62 - Ergebnisse __________________________________________________________________________________________ SOD-Aktivität (U/g Hb) 3000 2500 2000 1500 1000 alle LDA 2 LDA Abb. 23: LDA 3 1 alle RDA RDA sT RDA cT Gruppenvergleich der SOD-Aktivitäten (lm): Vergleich der SOD-Aktivitäten im abomasalen Blut bei allen Tieren mit linksseitiger Labmagenverlagerung (alle LDA), aufgeteilt in zweiten (LDA 2) und dritten (LDA 3) Grad, mit den SOD-Aktivitäten aller Tiere mit rechtsseitiger Labmagenverlagerung (alle RDA), aufgeteilt in Verlagerung ohne (RDA sT) und mit (RDA cT) Torsion (Median, 1.- 3. Quartil) Die abomasalen SOD-Aktivitäten in der Gruppe aller Tiere mit LDA (r = -0,66) und der LDA 3 (r = -0,78) waren mit der Hämoglobinkonzentration aus dem peripheren Blut mit p<0,01 negativ korreliert. In der Gruppe der Tiere mit LDA 3 (r = -0,64) waren die abomasalen Hämoglobinkonzentrationen und die abomasalen SOD-Aktivitäten mit p<0,05 negativ miteinander korreliert. 4.2.4.3 Vergleich der Superoxiddismutase-Aktivitäten im peripheren und abomasalen Blut Im Vergleich von peripherem zu abomasalem Blut ergaben sich in Bezug auf die SODAktivität keine signifikanten Unterschiede. In der Gruppe aller LDA und der LDA 2 wurde die Signifikanz nach dem Mann-Whitney-Test jedoch fast erreicht (p = 0,05), die SODAktivitäten zeigten sich im peripheren Blut höher als in dem aus dem Labmagen gewonnenen (Abb. 24). Ein weiterer, nicht signifikanter Unterschied ließ sich bei den RDA sT feststellen, wobei die SOD-Aktivitäten im peripheren Blut höher als im abomasalen waren. - 63 - Ergebnisse __________________________________________________________________________________________ SOD-Aktivität (U/g Hb) 4000 **=0,64 3000 **=0,66 **=0,81 *=0,53 2000 1000 0 Abb. 24: alle Tiere alle LDA LDA 2 1 3 LDA alle RDA RDA sT RDA cT Vergleich der SOD-Aktivitäten (p) zu den SOD-Aktivitäten (lm) innerhalb der Gruppen aller Tiere (alle Tiere), der Tiere mit linksseitiger Verlagerung (alle LDA), davon mit Verlagerung zweiten (LDA 2) und dritten (LDA 3) Grades, und der Tiere mit rechtsseitiger Verlagerung, davon ohne (RDA sT) und mit (RDA cT) Torsion (Median, 1.- 3. Quartil; * / **: Korrelationen der SOD-Aktivitäten (p) zu den SOD-Aktivitäten (lm) mit p ≤ 0,05 / 0,01) Tab. 14: Vergleich der SOD-Aktivitäten im peripheren (SOD (p)) und abomasalen (SOD (lm)) Blut, abhängig vom Abstand zur Abkalbung (Tage, d) in den Gruppen LDA 3 und RDA cT LDA 3 (n=13) Tage (d) post partum SOD (p) RDA cT (n=11) < 28 d (n=9) > 28 d (n=4) < 28 d (n=4) > 28 d (n=7) 2536 1921 2222 2039 (U/g Hb) (2438; 2723) (1783; 2059) (1922; 2416) (1793; 2449) SOD (lm) 2247 1703 2117 2019 (U/g Hb) (2183; 2469) (1466; 1997) (2039; 2259) (1849; 2471) Bei den LDA 3 fiel auf, dass die SOD-Aktivität bei den Kühen mit LDA 3 innerhalb der ersten vier Wochen post partum signifikant (p<0,05m; p+lm) höher lag als bei Kühen, die erst nach vier Wochen und später an LDA 3 erkrankten. Diese Differenz zeigte sich bei den RDA cT nur gering ausgeprägt (Tab. 14). - 64 - Ergebnisse __________________________________________________________________________________________ 4.2.5 Glutathionperoxidase-Aktivität (GPX-Aktivität) 4.2.5.1 Glutathionperoxidase-Aktivitäten im peripheren Blut (GPX-Aktivität (p)) Die höchste gemessene Aktivität der GPX, bezogen auf die Hämoglobinkonzentration, betrug 635,4 U/g Hb, die niedrigste 202,1 U/g Hb. Der Medianwert lag bei 509,1 U/g Hb, erstes und drittes Quartil bei 427,9 und 563,9 U/g Hb (Abb. 25). Die GPX-Aktivitäten aus dem peripheren Blut der Tiere mit LDA 3 waren signifikant niedriger als die der Gruppe mit LDA 2 (p<0,05) und die der Gruppe mit RDA cT (p<0,01), wobei die GPX-Aktivitäten der Gruppe der RDA cT signifikant höher lagen als die aller Tiere mit LDA (p<0,01), alle Tiere mit RDA hatten signifikant höhere GPX-Aktivitäten als alle Tiere mit LDA (p<0,01). GPX-Aktivität (U/g Hb) 700 600 500 400 300 200 alle LDA LDA 2 LDA 3 1 alle RDA RDA sT RDA cT Abb. 25: Gruppenvergleich der GPX-Aktivitäten (p): Vergleich der GPX-Aktivitäten im peripheren Blut bei allen Tieren mit linksseitiger Labmagenverlagerung (alle LDA), aufgeteilt in zweiten (LDA 2) und dritten (LDA 3) Grad, mit den GPX-Aktivitäten aller Tiere mit rechtsseitiger Labmagenverlagerung (alle RDA), aufgeteilt in Verlagerung ohne (RDA sT) und mit (RDA cT) Torsion (Median, 1.- 3. Quartil) 4.2.5.2 Glutathionperoxidase-Aktivitäten im abomasalen Blut (GPX-Aktivität (lm)) Die Aktivitäten der aus dem Labmagen gewonnenen GPX-Proben lagen minimal bei 206,3 und maximal bei 672,4 U/g Hb. Der Medianwert befand sich bei 484,1, erstes und drittes Quartil betrugen 426,3 und 525,5 U/g Hb (Abb. 26). Die GPX-Aktivitäten aller Tiere mit LDA waren signifikant niedriger als die aller Tiere mit RDA (p<0,05), besonders auch signifikant niedriger als die der Gruppe der RDA cT (p<0,05). - 65 - Ergebnisse __________________________________________________________________________________________ Innerhalb der Gruppen lagen die Aktivitäten der Gruppe mit LDA 3 signifikant niedriger als die der Gruppe mit RDA cT (p<0,05). GPX-Aktivität (U/g Hb) 700 600 500 400 300 200 alle LDA 2 LDA Abb. 26: LDA 3 1 alle RDA RDA sT RDA cT Gruppenvergleich der GPX-Aktivitäten (lm): Vergleich der GPX-Aktivitäten im abomasalen Blut bei allen Tieren mit linksseitiger Labmagenverlagerung (alle LDA), aufgeteilt in zweiten (LDA 2) und dritten (LDA 3) Grad, mit den GPX-Aktivitäten aller Tiere mit rechtsseitiger Labmagenverlagerung (alle RDA), aufgeteilt in Verlagerung ohne (RDA sT) und mit (RDA cT) Torsion (Median, 1.- 3. Quartil) 4.2.5.3 Vergleich der Glutathionperoxidase-Aktivitäten im peripheren und abomasalen Blut Tab. 15: Vergleich der GPX-Aktivitäten im peripheren (GPX (p)) und abomasalen (GPX (lm)) Blut, abhängig vom Abstand zur Abkalbung (Tage, d) in den Gruppen LDA 3 und RDA cT LDA 3 (n=13) Tage (d) post partum GPX (p) RDA cT (n=11) < 28 d > 28 d < 28 d > 28 d (n=9) (n=4) (n=4) (n=7) 428 305 563 571 (U/g Hb) (347; 504) (236; 371) (547; 580) (530; 625) GPX (lm) 435 331 527 522 (U/g Hb) (374; 490) (283; 380) (468; 591) (499; 606) Bei den LDA 3 warenGPX-Aktivitäten bei Kühen, die innerhalb der ersten vier Wochen an LDA 3 erkrankten, im peripheren Blut signifikant höher (p<0,05) als die der Kühe, die erst später erkrankten. Bei den RDA cT trat diese Differenz nicht auf (Tab. 15). - 66 - Ergebnisse __________________________________________________________________________________________ Zwischen den Aktivitäten der aus dem Labmagen bzw. aus peripherem Blut stammenden Proben bestanden keine signifikanten Unterschiede, nur bei den RDA cT deutete sich ein Unterschied an. Hier lagen die GPX-Aktivitäten im peripheren Blut höher als im abomasalen Blut (Abb. 27). GX-Aktivität (U/g Hb) 700 600 **=0,80 **=0,83 **=0,80 **=0,84 **=1,00 **=0,85 500 **=0,86 400 300 200 100 0 alle Tiere Abb. 27: alle LDA 1 LDA 3 LDA 2 alle RDA RDA sT RDA cT Vergleich der GPX-Aktivitäten (p) zu den GPX-Aktivitäten (lm) innerhalb der Gruppen aller Tiere (alle Tiere), der Tiere mit linksseitiger Verlagerung (alle LDA), davon mit Verlagerung zweiten (LDA 2) und dritten (LDA 3) Grades, und der Tiere mit rechtsseitiger Verlagerung, davon ohne (RDA sT) und mit (RDA cT) Torsion (Median, 1.- 3. Quartil; **: Korrelationen der GPX-Aktivitäten (p) zu den GPX-Aktivitäten (lm) mit p ≤ 0,01) 4.2.6 TEAC-Konzentration 4.2.6.1 TEAC-Konzentrationen im peripheren Blut (TEAC-Konzentration (p)) Die gemessenen TEAC-Konzentrationen variierten von 11 bis 726 µmol/l. Der Medianwert befand sich bei 284 µmol/l, das erste Quartil bei 210 µmol/l und das dritte bei 375 µmol/l (Abb. 28). Die TEAC-Konzentrationen der Gruppe mit LDA 2 zeigten sich im peripheren Blut signifikant höher als die der Gruppe mit LDA 3 (p<0,05). - 67 - Ergebnisse __________________________________________________________________________________________ TEAC-Konzentration (µmol/l) 450 400 350 300 250 200 150 Abb. 28: alle LDA LDA 2 LDA 3 1 alle RDA RDA sT RDA cT Gruppenvergleich der TEAC-Konzentrationen (p): Vergleich der TEAC-Konzentrationen im peripheren Blut bei allen Tieren mit linksseitiger Labmagenverlagerung (alle LDA), aufgeteilt in zweiten (LDA 2) und dritten (LDA 3) Grad, mit den TEAC-Konzentrationen aller Tiere mit rechtsseitiger Labmagenverlagerung (alle RDA), aufgeteilt in Verlagerung ohne (RDA sT) und mit (RDA cT) Torsion (Median, 1.- 3. Quartil) Die peripheren TEAC-Konzentrationen waren in der Gruppe aller Tiere mit LDA sowohl mit den Aktivitäten der Creatinkinase (p) (r = 0,57) als auch (lm) (r = 0,60) positiv korreliert (p<0,01). 4.2.6.2 TEAC-Konzentrationen im abomasalen Blut (TEAC-Konzentration (lm)) Die Konzentrationen der TEAC (lm) lagen zwischen 117 und 795 µmol/l. Der Medianwert betrug 267 µmol/l, erstes bzw. drittes Quartil betrugen 227 bzw. 342 µmol/l (Abb. 29). Ein signifikanter Unterschied zwischen den Gruppen bestand nur zwischen der Gruppe der LDA 3 und der Gruppe mit RDA cT (p<0,05). - 68 - Ergebnisse __________________________________________________________________________________________ TEAC-Konzentration (µmol/l) 400 360 320 280 240 200 alle LDA 2 LDA 3 LDA Abb. 29: Gruppenvergleich der 1 alle RDA RDA sT RDA cT TEAC-Konzentrationen (lm): Vergleich der TEAC- Konzentrationen im abomasalen Blut bei allen Tieren mit linksseitiger Labmagenverlagerung (alle LDA), aufgeteilt in zweiten (LDA 2) und dritten (LDA 3) Grad, mit den TEACKonzentrationen aller Tiere mit rechtsseitiger Labmagenverlagerung (alle RDA), aufgeteilt in Verlagerung ohne (RDA sT) und mit (RDA cT) Torsion (Median, 1.- 3. Quartil) 4.2.6.3 Vergleich der TEAC-Konzentrationen im peripheren und abomasalen Blut Zwischen den Konzentrationen der TEAC aus dem peripheren und dem Labmagenblut bestanden keine signifikanten Unterschiede. Tendenziell lagen bei den Gruppen mit LDA und der Gruppe der RDA sT im peripheren Blut höhere TEAC-Konzentrationen als im Labmagenblut vor, bei den RDA cT kehrte sich dies jedoch um (Abb. 30). TEAC-Konzentration (µmol/l) 500 **=0,42 **=1,00 *=0,45 400 300 200 100 0 alle Tiere Abb. 30: alle LDA LDA 2 1 LDA 3 alle RDA RDA sT RDA cT Vergleich der TEAC-Konzentrationen (p) zu den TEAC-Konzentrationen (lm) innerhalb der Gruppen aller Tiere (alle Tiere), der Tiere mit linksseitiger Verlagerung (alle LDA), davon mit Verlagerung zweiten (LDA 2) und dritten (LDA 3) Grades, und der Tiere mit rechtsseitiger Verlagerung, davon ohne (RDA sT) und mit (RDA cT) Torsion (Median; 1.- 3. Quartil; * / **: Korrelationen der TEAC-Konzentrationen (p) zu den TEAC-Konzentrationen (lm) mit p ≤ 0,05 / 0,01) - 69 - Ergebnisse __________________________________________________________________________________________ Tab. 16: Vergleich der TEAC-Konzentrationen im peripheren (TEAC-Konz. (p)) und abomasalen (TEAC-Konz. (lm)) Blut, abhängig vom Abstand zur Abkalbung (Tage, d) in den Gruppen LDA 3 und RDA cT LDA 3 (n=13) RDA cT (n=11) Tage (d) post partum TEAC-Konz. (p) < 28 d > 28 d < 28 d > 28 d (n=9) (n=4) (n=4) (n=7) 236 250 346 266 (μmol/l) (200; 309) (226; 276) (289; 388) (229; 281) TEAC-Konz. (lm) 227 264 350 327 (μmol/l) (205; 238) (207; 420) (308; 369) (257; 376) Bei den Kühen mit LDA 3 waren die TEAC-Konzentrationen mit zunehmendem Abstand zum Partus geringfügig höher. Bei den RDA cT dagegen lagen die TEAC-Konzentrationen innerhalb der ersten vier Wochen post partum im peripheren Blut signifikant höher als bei Tieren die später erkrankten (p<0,05; Tab. 16). 4.3 TEAC-Konzentrationsverlauf post operationem Für die Verlaufsuntersuchungen der TEAC-Konzentration wurden einzelnen Tieren vor, während und nach der Rückverlagerung des Labmagens Blutproben aus der V. jugularis entnommen, die sich bei den linksseitigen auf einen Zeitraum von zwölf Stunden und bei den rechtsseitigen auf einen von sechzehn Stunden erstreckten. Der erste Messwert entsprach dabei immer dem Status vor Operationsbeginn. Der zweite und zum Teil auch der dritte wurden intra operationem entnommen, die nachfolgenden zeigten den Zustand post operationem. Bei den linksseitigen Verlagerungen wurde der Verlauf bei fünf Tieren bestimmt (Abb. 31- 35), bei den rechtsseitigen bei neun Tieren (Abb. 37-46). Zur besseren Demonstration der Ergebnisse wurden im Folgenden Einzeldarstellungen von rechts- und linksseitiger Verlagerung gezeigt. Zu jedem einzelnen Verlauf wurden noch Alter des Tieres, letzter Abkalbetermin, Lactat- und Harnstoffkonzentrationen im peripheren Blut in einer Tabelle neben der Graphik und zusätzliche Erkrankungen aufgeführt, um die einzelnen Krankheitsverläufe einordnen zu können. Innerhalb der Kühe mit linksseitigen Verlagerungen war ein einheitlicher Trend angedeutet (Abb. 36), innerhalb der rechtsseitigen Verlagerungen nicht. - 70 - Ergebnisse __________________________________________________________________________________________ 4.3.1 TEAC-Konzentrationsverlauf bei Tieren mit LDA Bei allen Tieren mit LDA, bei denen postoperativ der TEAC-Konzentrationsverlauf gemessen wurde, handelte es sich um Verlagerungen dritten Grades. Alle Kühe konnten nach der erfolgreichen operativen Rückverlagerung nach Hause entlassen werden. TEAC-Konzentrationsverlauf Kuh 7 Diagnose LDA3 800 Alter (a) 6 600 Abstand zur letzten Abkalbung (d) Lactatkonzentration (mmol/l) Harnstoffkonzentration (mmol/l) 30 TEAC-Konzentration (µmol/l) 400 200 0 Zeit (h) 0 Abb. 31: 2 4 6 8 10 12 2,19 5,7 TEAC-Konzentrationsverlauf post operationem, klinische Daten und Lactat- sowie Harnstoffkonzentrationen im peripheren Blut von Kuh 7 (LDA 3) Diese Kuh wies neben der DA eitrig-nekrotische Dekubitusstellen an den Hintergliedmaßen, hochgradig gerötete Kronsäume (Klauenrehe) sowie eine katarrhalisch-eitrige Endometritis auf. TEAC-Konzentrationsverlauf Kuh 8 TEAC-Konzentration (µmol/l) 600 500 400 300 200 100 Zeit (h) 0 0 Abb. 32: 2 4 6 8 10 Diagnose LDA 3 Alter (a) Abstand zur letzten Abkalbung (d) Lactatkonzentration (mmol/l) Harnstoffkonzentration (mmol/l) 3 7 1,75 6,3 12 TEAC-Konzentrationsverlauf post operationem, klinische Daten und Lactat- sowie Harnstoffkonzentrationen im peripheren Blut von Kuh 8 (LDA 3) - 71 - Ergebnisse __________________________________________________________________________________________ Zusätzlich zur LDA 3 hatte die Kuh wässrig-stinkenden Durchfall und eine katarrhalischeitrige Endometritis. TEAC-Konzentrationsverlauf Kuh 14 TEAC-Konzentration (µmol/l) 800 600 400 200 Zeit (h) 0 0 Abb. 33: 2 4 6 8 10 Diagnose LDA 3 Alter (a) Abstand zur Abkalbung (d) Lactatkonzentration (mmol/l) Harnstoffkonzentration (mmol/l) 4,5 60 3,55 2,8 12 TEAC-Konzentrationsverlauf post operationem, klinische Daten und Lactat- sowie Harnstoffkonzentrationen im peripheren Blut von Kuh 14 (LDA 3) Diese Kuh wies keine zusätzlichen Erkrankungen auf. TEAC-Konzentrationsverlauf Kuh 18 TEAC-Konzentration (µmol/l) 800 600 400 200 0 Zeit (h) 0 Abb. 34: 2 4 6 8 10 Diagnose LDA 3 Alter (a) Abstand zur letzten Abkalbung (d) Lactatkonzentration (mmol/l) Harnstoffkonzentration (mmol/l) 2,5 30 1,10 4,2 12 TEAC-Konzentrationsverlauf post operationem, klinische Daten und Lactat- sowie Harnstoffkonzentrationen im peripheren Blut von Kuh 18 (LDA 3) Diese Kuh zeigte zusätzlich entzündliche, fluktuierende Umfangsvermehrungen an den Tarsalgelenken (Bursitis der Schleimbeutel) sowie eine katharrhalische Pneumonie. - 72 - Ergebnisse __________________________________________________________________________________________ TEAC-Konzentrationsverlauf Kuh 21 TEAC-Konzentration (µmol/l) 800 600 400 200 0 Zeit (h) 0 Abb. 35: 2 4 6 8 10 Diagnose LDA 3 Alter (a) Abstand zur letzten Abkalbung (d) Lactatkonzentration (mmol/l) Harnstoffkonzentration (mmol/l) 4 20 3,56 3,4 12 TEAC-Konzentrationsverlauf post operationem, klinische Daten und Lactat- sowie Harnstoffkonzentrationen im peripheren Blut von Kuh 21 (LDA 3) Bei dieser Kuh bestand zusätzlich im vorderen linken Viertel eine katarrhalische Mastitis, der Uterus war 30 Tage post partum geringgradig vergrößert. 4.3.1.1. Zusammenfassende Darstellung der TEAC-Konzentrationsverläufe bei Kühen mit LDA 3 TEAC-Konzentrationsdifferenz (µmol/l) 2,5-3 100 50 0,5-1 3,5-4 4,5-5 6,5-7 7,5-8 10,5-12 1,5-2 0 5,5-6 8,5-10 Zeitintervalle 1 -50 -100 -150 -200 Abb. 36: TEAC-Konzentrationsverläufe post operationem: bei den Kühen 7, 8, 14, 18 und 21 abgetragene Änderungen von ihrem jeweiligen TEAC-Ausgangsniveau (Median, 1.-3. Quartil) - 73 - Ergebnisse __________________________________________________________________________________________ Nach Abtragen der Einzelverläufe als jeweilige Differenz vom Ausgangsniveau fielen zu Beginn die individuellen Schwankungen auf. Nur sechs und acht Stunden post operationem stiegen die TEAC-Konzentrationen bei allen Kühen mit LDA um 150 bzw. 100 µmol/l an. Danach fielen sie wieder zum jeweiligen Ausgangsniveau ab (Abb. 36). 4.3.2 TEAC-Konzentrationsverlauf bei Tieren mit RDA Bei allen Tieren mit RDA, bei denen postoperativ der TEAC-Konzentrationsverlauf gemessen wurde, handeltees sich bis auf die Kühe 24 und 31 um Tiere mit RDA cT. TEAC-Konzentrationsverlauf Kuh 19 RDA cT TEAC-Konzentration (µmol/l) Diagnose 600 500 400 300 200 100 0 Zeit (h) 0 Abb. 37: 2 4 6 8 10 12 14 16 Alter (a) Abstand zur letzten Abkalbung (d) Lactatkonzentration (mmol/l) Harnstoffkonzentration (mmol/l) (270°) 4 60 10,29 7,5 TEAC-Konzentrationsverlauf post operationem, klinische Daten und Lactat- sowie Harnstoffkonzentrationen im peripheren Blut von Kuh 19 (RDA cT) Bei dieser Kuh bestanden keine zusätzlichen Erkrankungen. TEAC-Konzentrationsverlauf Kuh 24 TEAC-Konzentration (µmol/l) Diagnose RDA sT Alter (a) Abstand zur letzten Abkalbung (d) Lactatkonzentration (mmol/l) Harnstoffkonzentration (mmol/l) 4 800 600 400 200 Zeit (h) 0 0 2 4 6 8 10 12 14 16 - 74 - 19 2,00 4,5 Ergebnisse __________________________________________________________________________________________ Abb. 38: TEAC-Konzentrationsverlauf post operationem, klinische Daten und Lactat- sowie Harnstoffkonzentrationen im peripheren Blut von Kuh 24 (RDA sT) Neben der RDA sT bestand bei dieser Kuh eine hochgradige Puerperalstörung. TEAC-Konzentrationsverlauf Kuh 25 RDA cT TEAC-Konzentration (µmol/l) 800 Diagnose 600 Alter (a) 2,5 400 Abstand zur letzten Abkalbung (d) Lactatkonzentration (mmol/l) Harnstoffkonzentration (mmol/l) 20 200 Zeit (h) 0 0 Abb. 39: 2 4 6 8 10 12 14 16 (>360°) 6,14 10,2 TEAC-Konzentrationsverlauf post operationem, klinische Daten und Lactat- sowie Harnstoffkonzentrationen im peripheren Blut von Kuh 25 (RDA cT) Diese Kuh zeigte bei Aufnahme einen hochgradigen Enophthalmus sowie weitere Anzeichen eines hochgradig gestörten Kreislaufes. Zusätzlich zur DA bestand eine hochgradige Puerperalstörung mit massivem Scheidenausfluss. In der Operation wies die Labmagenwand eine zyanotische Färbung auf, auffällig war zudem eine massive Flüssigkeitsansammlung im Labmagen. Obwohl die Kuh zwei Tage post operationem Futteraufnahme, Wiederkauen und dünnflüssigen Kotabsatz zeigte, verschlechterte sich das klinische Bild bis zum fünften Tag post operationem. Das Abdomen war hochgradig umfangsvermehrt, die Kuh nahm kein Futter mehr auf und zeigte Schmerzäußerungen, der Kot war schwarz gefärbt. Nach weiterer Verschlechterung des Kreislaufes wurde die Kuh euthanasiert. In der pathologischhistologischen Untersuchung wurden eine mittel- bis hochgradige fibrinöse Peritonitis, eine hochgradige Labmagenüberladung sowie eine fokale, mittelgradige ulzerative Abomasitis im Bereich des Pylorus festgestellt. - 75 - Ergebnisse __________________________________________________________________________________________ TEAC-Konzentrationsverlauf Kuh 31 RDA sT TEAC-Konzentration (µmol/l) 800 Diagnose 600 400 200 0 Zeit (h) 0 Abb. 40: 2 4 6 8 10 12 14 16 (Sand) Alter (a) 4 Abstand zur letzten Abkalbung (d) Lactatkonzentration (mmol/l) Harnstoffkonzentration (mmol/l) 65 4,46 16,0 TEAC-Konzentrationsverlauf post operationem, klinische Daten und Lactat- sowie Harnstoffkonzentrationen im peripheren Blut von Kuh 31 (RDA sT) Bei dieser Kuh bestanden keine zusätzlichen Erkrankungen. In der Operation war die Labmagenwand zyanotisch gefärbt. Nach der Operation bestanden zwischen Phasen normaler Futteraufnahme intermittierend Digestionsstörungen mit Tympanien. Die Kuh konnte erst zwei Wochen post operationem entlassen werden. TEAC-Konzentrationsverlauf Kuh 32 RDA cT TEAC-Konzentration (µmol/l) 800 Diagnose 600 Alter (a) 4 Abstand zur letzten Abkalbung (d) Lactatkonzentration (mmol/l) Harnstoffkonzentration (mmol/l) 180 400 200 0 Zeit (h) 0 Abb. 41: 2 4 6 8 10 12 14 16 (180°) 8,66 19,4 TEAC-Konzentrationsverlauf post operationem, klinische Daten und Lactat- sowie Harnstoffkonzentrationen im peripheren Blut von Kuh 32 (RDA cT) Bei dieser Kuh konnte die vor der Operation gewonnene Blutprobe nicht verwertet werden, der Verlauf beginnt also erst nach der Rückverlagerung. Neben der RDA cT bestanden keine zusätzlichen Erkrankungen. - 76 - Ergebnisse __________________________________________________________________________________________ TEAC-Konzentrationsverlauf Kuh 33 RDA cT TEAC-Konzentration (µmol/l) 400 Diagnose 300 Alter (a) 5 Abstand zur letzten Abkalbung (d) Lactatkonzentration (mmol/l) Harnstoffkonzentration (mmol/l) 60 200 100 Zeit (h) 0 0 Abb. 42: 2 4 6 8 10 12 14 16 (180°) 2,33 7,5 TEAC-Konzentrationsverlauf post operationem, klinische Daten und Lactat- sowie Harnstoffkonzentrationen im peripheren Blut von Kuh 33 (RDA cT) In der Operation fielen die zyanotische Färbung der Labmagenwand, eine Verwachsung des Labmagens mit dem Netz und dem Peritoneum und eine mittelgradige Aufgasung des Zäkums auf. TEAC-Konzentrationsverlauf Kuh 34 RDA cT TEAC-Konzentration (µmol/l) Diagnose 800 (180°) 600 Alter (a) 6 400 Abstand zur letzten Abkalbung (d) Lactatkonzentration (mmol/l) Harnstoffkonzentration (mmol/l) 7 200 0 Zeit (h) 0 Abb. 43: 2 4 6 8 10 12 14 16 9,26 6,9 TEAC-Konzentrationsverlauf post operationem, klinische Daten und Lactat- sowie Harnstoffkonzentrationen im peripheren Blut von Kuh 34 (RDA cT) Neben der RDA cT hatte die Kuh eine mittelgradige Puerperalstörung. - 77 - Ergebnisse __________________________________________________________________________________________ TEAC-Konzentrationsverlauf Kuh 35 RDA cT TEAC-Konzentration (µmol/l) Diagnose 800 600 400 200 0 Zeit (h) 0 Abb. 44: 2 4 6 8 10 12 14 16 Alter (a) Abstand zur letzten Abkalbung (d) Lactatkonzentration (mmol/l) Harnstoffkonzentration (mmol/l) (270°) 4 100 7,77 4,2 TEAC-Konzentrationsverlauf post operationem, klinische Daten und Lactat- sowie Harnstoffkonzentrationen im peripheren Blut von Kuh 35 (RDA cT) Die Kuh zeigte keine zusätzlichen Erkrankungen. TEAC-Konzentrationsverlauf Kuh 37 RDA cT TEAC-Konzentration (µmol/l) 200 Diagnose 150 100 50 0 Zeit (h) 0 Abb. 45: 2 4 6 8 10 12 14 16 Alter (a) Abstand zur letzten Abkalbung (d) Lactatkonzentration (mmol/l) Harnstoffkonzentration (mmol/l) (270°) 6 140 3,85 13,6 TEAC-Konzentrationsverlauf post operationem, klinische Daten und Lactat- sowie Harnstoffkonzentrationen im peripheren Blut von Kuh 37 (RDA cT) Neben der RDA cT lahmte die Kuh hinten links hochgradig, der Besitzer wünschte keine weitere Diagnostik und Therapie dieser Begleiterkrankung. - 78 - Ergebnisse __________________________________________________________________________________________ 5. Diskussion 5.1 Gruppeneinteilung der Tiere Die Einteilung der Tiere mit DA erfolgte entsprechend der gängigen Unterteilung in DA nach links (LDA) und DA nach rechts (RDA) (BÜCKNER 1993, KLEISER u. FÜRLL 1998, u.v.a.). Das hat für den Labmagen und den Gesamtorganismus sehr unterschiedliche Folgen. Handelt es sich bei den LDA um einen Zustand, der permanent oder nur zeitweise als sogenannter „pendelnder Labmagen“ auftreten kann, so sind die RDA mit Volvolus als akute Erkrankung anzusehen, die ohne operativen Eingriff binnen weniger Tage zum Tode führen (MEYLAN 1999, FÜRLL et al. 1999). Innerhalb der LDA wird eine weitere Unterteilung in 1.-3. Grad nach der Höhe des Aufsteigens des Labmagens vorgenommen. Bei Verlagerungen dritten Grades besteht eine größere Beeinflussung der Zirkulation und der Wand des Labmagens als bei Verlagerungen zweiten Grades (WITTEK u. FÜRLL 2002). Bei der RDA wird zwischen Verlagerungen ohne (RDA sT) und mit (RDA cT) Torsion unterschieden (MEYLAN 1999). Eine Torsion zieht eine sofortige Einschränkung des Blutflusses und somit eine Unterversorgung im Sinne einer Ischämie nach sich. Dies konnte auch durch pulsoximetrische Untersuchungen und Bestimmung der Lactatkonzentration an den Tieren mit RDA dieser Studie nachgewiesen werden (WITTEK et al. 2004). Eine rechtsseitige Verlagerung ohne Torsion ist hinsichtlich der Beeinträchtigung des Gesamtorganismus und des Labmagens durch die Aufgasung eher mit einer linksseitigen Verlagerung zu vergleichen als mit einer Verlagerung mit Torsion. 5.2 Methode der Blutentnahme am Labmagen Die Blutentnahme aus den Venen des Labmagens ermöglicht, die Verhältnisse direkt am Labmagen unmittelbar nach Beginn der Reperfusion zu erfassen. Vergleicht man diese mit den Parametern aus dem peripheren venösen Blut, so hat man die Möglichkeit, die Vorgänge am Labmagen, die direkt durch Verlagerung, Ischämie und Reperfusion verursacht wurden, mit der Reaktion des Gesamtorganismus auf die Verlagerung des Labmagens und auf begleitende Umstände wie Transport und Operation zu vergleichen. Um die Tiere vor und nach der Operation in ihrem antioxidativen Status einzuschätzen, muss sichergestellt sein, dass die Verhältnisse im peripheren Blut auch denen am Ort des Geschehens, dem Labmagen, - 79 - Diskussion __________________________________________________________________________________________ entsprechen. Eine Blutentnahme vor und nach der Operation war nur peripher möglich, so dass für eine praktikable Diagnostik sichergestellt sein muss, dass keine signifikant abweichende Veränderung des antioxidativen Status auf dem Weg vom Labmagen zur peripheren Blutentnahme erfolgt. 5.3 Ausgewählte klinische Befunde bei Kühen mit DA Die Pulsfrequenzen der untersuchten Kühe waren bei 30 % aller Tiere erhöht, innerhalb der LDA bei 27 %, innerhalb der RDA bei 33 % (Tab. 11). Entsprechende Ergebnisse erhielt auch MÜLLER (1992). Erhöhte Pulsfrequenzen sind Zeichen einer Kreislaufinsuffizienz (FUBINI et al. 1991), die bei Kühen mit RDA häufiger besteht, was sich auch am klinischen Verlauf bestätigt (DIRKSEN 1961, KÜMPER 1995). Bei 70 % der Kühe lag die letzte Abkalbung weniger als 28 Tage zurück (Tab. 9). Dies entspricht der Bezeichnung der DA als peripartale Erkrankung. Ursache dafür sind nach der Geburt geänderte Platzverhältnisse im Abdomen der Kühe (FÜRLL u. KRÜGER 2000). KÜMPER (1995) sieht im geringen Füllungszustand der Vormägen die Ursache für das vermehrte Auftreten von LDA in dieser Zeit. Als häufigste zusätzliche Erkrankungen traten Veränderungen am Genitale auf (57 % aller Tiere, Tab. 12). Dies entspricht den Ergebnissen von REHAGE et al. (1996), die bei Kühen mit LDA zu 50 % Begleiterkrankungen feststellten, wobei die Endometritis an erster Stelle stand. Kühe mit LDA wiesen signifikant (p<0,05) häufiger Erkrankungen des Genitale auf als Kühe mit RDA. Erkrankungen des Genitale, wie der Endometritis oder der Retentio secundinarum, treten am häufigsten nach der Geburt auf, je länger die Geburt zurückliegt, desto seltener werden diese Erkrankungen (Grunert 1986). Da die LDA häufiger im peripartalen Zeitraum auftritt als die RDA (KÜMPER 1995), erklärt sich dieser Sachverhalt. 5.4 Messergebnisse und Vergleich Die Lagerung der Proben erfolgte bei -21 °C über einen maximalen Zeitraum von einem Monat. In einer Studie von KÜHLER (1996) wird dies als Zeitraum angegeben, in dem die Aktivitäten von GPX und SOD nahezu unverändert bleiben. Energie- und Fettstoffwechsel bei Kühen mit DA - 80 - Diskussion __________________________________________________________________________________________ Die untersuchten Parameter (BHB, Cholesterol, Bilirubin) stellen eine repräsentative Auswahl zur Beurteilung des Energie- und Fettstoffwechsels dar (FÜRLL et al. 2002). Der Medianwert der BHB-Konzentration im peripheren Serum lag bei allen Kühen mit 1,15 mmol/l oberhalb des Referenzbereiches gesunder Kühe (Tab. 10, Tab. 13). Kühe mit LDA zeigten höhere BHB-Konzentrationen als Kühe mit RDA. Kühe mit RDA sT wiesen ähnliche BHB-Konzentrationen wie Kühe mit LDA auf. Kühe mit RDA cT zeigten BHBKonzentrationen im Referenzbereich. Die Cholesterolkonzentrationen lagen bei allen Kühen unter dem Referenzbereich (Tab. 10, Tab. 13). Dabei waren die Cholesterolkonzentrationen der RDA cT signifikant (p<0,01) höher als die der Tiere mit LDA 3. Dies entspricht den Ergebnissen von KÜMPER (1995), BAJCSY et al. (1997) und FÜRLL et al. (1997), die bei LDA auch bestehende Ketosen beobachteten. Zu erklären sind gesteigerte BHBKonzentrationen durch den Abbau von Körperfett bei gesteigertem Energiebedarf und verringerter Futteraufnahme in der Endgravidität und der Frühlaktation (MUDRON et al. 1997). Niedrige Cholesterolkonzentrationen treten bei verminderter Futteraufnahme (ROSSOW u. STAUFFENBIEL 1983) sowie bei geringerer Synthese in der Leber auf (MUDRON et al. 1998). Die Unterschiede zwischen LDA und RDA erklären sich durch die zumeist längere Krankheitsdauer bei Kühen mit LDA und den bei den Kühen mit RDA zum Teil länger zurück liegenden Abkalbeterminen (SATTLER 2001). Ein weiterer diagnostischer Parameter stellt die Konzentration an freien Fettsäuren im Blut dar, die während Energiemangel und Ketose durch Lipidabbau im Fettgewebe vermehrt freigesetzt werden (MUDRON et al. 1991). Die Bilirubinkonzentrationen im Serum waren bei allen Kühen erhöht (Tab. 10, Tab. 13). Die Bilirubinkonzentrationen der Kühe mit RDA cT lagen niedriger als aller anderen Gruppen. Bilirubinkonzentrationen zwischen fünf und 15 können durch Interaktion freier Fettsäuren mit dem Transportprotein des Bilirubins und durch mangelnde Stimulation der Gallensekretion bei verminderter Futteraufnahme hervorgerufen werden (FÜRLL u. SCHÄFER 1992). Darüber liegende Konzentrationen sprechen für eine Schädigung der Leberzellen, wie sie durch Verfettung hervorgerufen wird. Eine solche Verfettung stellten REHAGE et al. (1996) bei 72 % aller untersuchten Kühe mit DA fest. Davon muss auch in dieser Untersuchung bei 54 % der Kühe ausgegangen werden, da sie Bilirubinkonzentrationen über 15 μmol/l besaßen. Leberenzymaktivitäten Die Aktivitäten der GLDH lagen in allen Gruppen deutlich über dem Referenzbereich (Tab. 10, Tab. 13). In neueren Untersuchungen werden erst Aktivitäten > 50 IU/l als erhöht - 81 - Diskussion __________________________________________________________________________________________ angesehen (FÜRLL et al. 2002), die in der Gruppe der LDA 3 nicht erreicht wurden. Ein einheitliches Schema in der Erhöhung der GLDH konnte nicht festgestellt werden, so zeigten sich die Gruppen der LDA 2 und der RDA cT mit den höchsten Aktivitäten, die deutlich über 100 IU/l liegen (Tab. 13). Da die GLDH intramitochondrial lokalisiert ist, kommt es erst bei schwereren Leberzellschädigungen zur Erhöhung ihrer Aktivität im Serum (BEKELE 1997). Neben Schädigungen der Leber kann allerdings auch eine Nierenschädigung in Betracht gezogen werden, da die GLDH in Organanalysen dort in ähnlichem Umfang wie in der Leber nachgewiesen werden konnte (FÜRLL et al. 2002). Anhand der anderen Parameter des Fettstoffwechsels, die bei den RDA cT eine geringere Belastung als bei den LDA aufwiesen, ist der Grund für die Erhöhung der GLDH auch teilweise extrahepatisch zu suchen. Die Aktivität der ASAT war in allen Gruppen deutlich erhöht (Tab. 13). Besonders herausragend waren die Gruppen der RDA und der LDA 2. Erhöhungen der ASAT gelten als sensible Indikatoren für das Vorliegen von Leberzellschädigungen (HERDT u. GERLOFF 1982). Bei der Wertung der Aktivitäten muss berücksichtigt werden, dass Kühe mit geburtsnaher DA signifikant höhere Aktivitäten aufweisen (GEISHAUSER 1999). Dies erklärt die hohen Aktivitäten der Gruppe der LDA 2 und RDA sT, die bis auf eine Kuh innerhalb von 4 Wochen nach der Geburt an einer DA erkrankten (Tab. 9). Neben der Belastung der Leber durch Fett- und Energiestoffwechsel kann die Aktivität der ASAT auch durch Muskelschäden erhöht werden, wie sie vor allem nach der Geburt im Uterus auftreten können (SOBIRAJ 1992). Dies erklärt auch die Abhängigkeit der ASAT-Aktivität zum Abstand zum letzten Abkalbetermin. Untersuchungen bei Kühen mit gestörtem Puerperium und Endometritiden haben ergeben, dass die ASAT-Aktivität bei diesen Kühen signifikant höher liegt als bei gesunden (EULENBERGER 1984, SATTLER 2001). Obwohl aus Untersuchungen des Blutes allein nicht auf den Leberfettgehalt geschlossen werden kann (STAUFFENBIEL 1993), so ist doch aus der Summe der Konzentrations- und Aktivitätsänderungen von BHB, Cholesterol, Gesamtbilirubin, GLDH und ASAT auf eine unterschiedlich ausgeprägte Leberverfettung (MERTENS 1992, REHAGE et al. 1996) vorrangig in den Gruppen der LDA, aber auch in der Gruppe der RDA sT zu schließen. Eiweissstoffwechsel Die Konzentration des Totalproteins lag in allen Gruppen im physiologischen Bereich (Tab. 10, Tab. 13), nur 22 % aller Tiere überschritten den Referenzbereich. Eine erhöhte Konzentration an Totalprotein im Serum ist Indikator für verstärkte Abwehrprozesse, wie sie - 82 - Diskussion __________________________________________________________________________________________ im Rahmen einer Entzündung auftreten (ROUSSEL et al. 1981). Ein gruppengebundenes vermehrtes Auftreten war nicht festzustellen. Die Harnstoffkonzentration war nur in der Gruppe der RDA cT erhöht (Tab. 13), die Konzentrationen der RDA sT lagen auf demselben Niveau wie die der LDA. Kühe im peripartalen Zeitraum unterliegen einem verstärkten Eiweißkatabolismus, was sich durch Anstieg des Abbauproduktes Harnstoff äußert (FÜRLL et al. 2002). Desweiteren können erhöhte Konzentrationen an Harnstoff bei Kühen mit RDA als Zeichen einer prärenalen Nierenfunktionsstörung oder einer verminderten Perfusion der Nieren (prärenales Nierenversagen) durch Dehydratation angesehen werden (Morrow 1975). Die Dehydratation zeigte sich bereits bei der klinischen Untersuchung in Form des Enophthalmus und des verringerten Blutangebotes. Kühe mit RDA sind von der Dehydratation stärker betroffen als Kühe mit LDA (FUBINI et al. 1991). Mineralstoffwechsel Die verringerten Kaliumkonzentrationen in den Gruppen der LDA 3, RDA sT und RDA cT (Tab. 13) entsprechen den von FÜRLL et al. (1998) nachgewiesenen Veränderungen der Kaliumkonzentration bei Kühen, die nachfolgend an DA erkrankten, am dritten Tag post partum. Eine Verringerung der Kaliumkonzentration tritt laut CARLSON (1990) unter Stresseinwirkung, wie einer Schwergeburt auf. Da Kühe mit DA häufig auch geringere Futtermengen aufnehmen und unter weiteren Digestionsstörungen wie Diarrhoe leiden, müssen zusätzlich auch eine verringerte Kaliumaufnahme und ein erhöhter Kaliumverlust über den Darm mit einbezogen werden (JÜNGER u. FÜRLL 1998). Folgeerscheinungen werden durch Hypokaliämie vor allem in Organen mit glatter Muskulatur hervorgerufen, da die Erregungsleitung der Muskelzellen gestört wird. Dies ist bei der Ätiologie der gestörten Labmagenmotorik (EICHER et al. 1998) und dem Wiederingangkommen derselben nach Reposition des Labmagens bei Kühen mit RDA cT zu beachten, da diese in dieser Untersuchung signifikant (p<0,01) geringere Kaliumkonzentrationen als Kühe mit LDA 2 aufwiesen. Die verringerten Natriumkonzentrationen in der Gruppe der RDA cT (Tab. 13) sind mit Ausdruck der verringerten Futteraufnahme der Tiere (JÜNGER u. FÜRLL 1998), aber auch Ausdruck des Energiemangels bei Ischämie, was zum Aussetzen der Na-K-Pumpe führen kann (CHAUDRY 1983). So konnten Geishauser et al. (1996 a) bei Kühen mit RDA im Labmagensaft signifikant höhere Natriumkonzentrationen als bei Kühen mit LDA feststellen. - 83 - Diskussion __________________________________________________________________________________________ Chloridkonzentration Nur in der Gruppe der RDA cT lagen durchschnittlich verringerte Chloridkonzentrationen im Blutplasma vor (Abb. 15). Ähnliche Ergebnisse erhielten auch KLEISER u. FÜRLL (1998) bei Entwicklung eines Screenings zur Früherkennung der DA. Sie erklären die verringerten Chloridkonzentrationen durch die gestörte Labmagenpassage und die daraus resultierende verringerte Resorption. In Untersuchungen des Labmagensaftes durch GEISHAUSER et al. (1996 a) waren die Chloridkonzentrationen im Labmagen bis zum dritten Tag post operationem bei Kühen mit RDA noch immer signifikant erhöht, während sie bei den Kühen mit LDA direkt nach der Reposition wieder abfällt. Lactatkonzentration Die Lactatkonzentrationen der Tiere mit RDA cT waren signifikant höher als der Tiere mit LDA 2 ( p<0,05) und die der Tiere mit LDA 3 (p<0,01; Abb. 16, 17). Das Lactat entsteht in gesteigertem Umfang in ischämischen Bezirken, wenn die Zellen nicht mehr ausreichend mit Sauerstoff versorgt werden und auf die anaerobe Glykolyse zur Energiegewinnung zurückgreifen müssen (ÖHLENSCHLÄGER 1993, TILL 1999). Somit ist der Anstieg der Lactatkonzentration bei den RDA cT als Anzeichen einer verringerten Durchblutung (Ischämie) der Labmagenwand durch die Torsion anzusehen. Bei den Kühen mit RDA sT liegen kaum Perfusionsstörungen vor, das Lactat steigt bei ihnen nicht an. Daher können die RDA sT mit den LDA verglichen werden. Die Perfusionsstörung bei den RDA cT konnten durch WITTEK et al. (2004) auch mittels Pulsoximetrie und Sauerstoffsättigung im Labmagenblut bestätigt werden. Creatinkinase-Aktivität (CK-Aktivität) Die gemessenen Aktivitäten der CK in der Gruppe aller Tiere lagen sowohl im peripheren (380 U/l) als auch im abomasalen Blut (384 U/l) deutlich über dem Referenzbereich. (Abb. 19, 20). An dieser Erhöhung können viele Faktoren beteiligt sein. Die Aktivität der CK steigt beim Transport von Rindern (GROTH u. GRÄNZER 1975, WITT et al. 1992) und bei chirurgischen Eingriffen (AUTEFAGE et al. 2000). Des weiteren ist sie bei Kühen, die nachfolgend an DA erkrankten, am 3. Tag p. p. im Vergleich zu Kühen ohne spätere Verlagerung erhöht (NAURUSCHAT u. FÜRLL 2002). Auch die zusätzlichen Erkrankungen, besonders am Genitale und an den Klauen können zur Aktivitätssteigerung beitragen (KLEISER u. FÜRLL 1998). Auch wenn die erhöhten CK-Aktivitäten nicht allein durch die DA hervorgerufen werden, ist doch davon auszugehen, dass sie einen Hauptteil dieser Erhöhung ausmacht. So konnten NAURUSCHAT u. FÜRLL (2002) bei Kühen mit DA - 84 - Diskussion __________________________________________________________________________________________ höhere Aktivitäten der CK als bei gesunden Tieren und Kühen mit Retentio secundinarum, Endometritiden und Gebärparese feststellen. Im Vergleich der Gruppen zeigte sich, dass die Gruppe der LDA 2 signifikant höhere Aktivitäten als die der LDA 3 (p<0,05) besaß, auch die Gruppe der RDA sT wies höhere Aktivitäten als die der RDA cT auf. SATTLER (2001) konnte feststellen, dass Kühe bis vier Wochen post partum signifikant höhere Aktivitäten der CK als Kühe vier bis sechs Wochen post partum haben. Die Berücksichtigung des Abkalbedatums zeigt, dass sich in der Gruppe RDA sT 75 % und in der Gruppe der LDA 2 72 % der Kühe innerhalb der ersten vier Wochen post partum befanden (Tab. 9), in den anderen Gruppen ist der Anteil wesentlich geringer. Die höheren Aktivitäten bei den Tieren sind demzufolge auf den geringeren Abstand zur Abkalbung zurückzuführen, nicht auf höheren Stress durch die DA. Dennoch fällt auf, dass die Gruppe der LDA 3 signifikant niedrigere Aktivitäten als die Gruppen der RDA (p<0,05) hatten. Als mögliche Erklärungen hierfür sind die Schädigung der Labmagenmuskulatur bei den RDA cT (GEISHAUSER 1995) mit Austritt der CK aus dem Cytosol ins Blut (WYSS u. KADDURAH-DAOUK 2000) und die Inaktivierung der CK bei andauerndem oxidativem Stress zu berücksichtigen. REDDY et al. (2000) stellten hierzu fest, dass die CK in Phasen oxidativen Stresses mit oxidiertem Glutathion reagiert. Reduziertes Glutathion zählt zur Klasse der nicht-enzymatischen Scavenger von Radikalen, das durch Abfangen von Radikalen oxidiert wird. In seiner oxidierten Form kann es die CK reversibel inaktivieren. Eine Reaktivierung der CK kann nur durch reduziertes Glutathion erfolgen. Dieses ist erst wieder vorhanden, wenn die Phase des oxidativen Stresses vorbei ist und das oxidierte Glutathion wieder vollständig regeneriert wurde. Da bei Tieren mit LDA die DA schon über längere Zeit bestehen kann, bevor Diagnose und Operation erfolgen, kann auch die Dauer des oxidativen Stresses, der zur Inaktivierung führen kann, für die Tiere nicht nachvollzogen werden. Schlussfolgernd für die präoperative Diagnostik der DA ist festzustellen, dass bei der Beurteilung der CK-Aktivität zugleich der Uterusbefund zu erheben ist, da dieser in Bezug auf Abkalbetermin (SATTLER 2001) und zusätzliche Erkrankungen wie Endometritiden (PAYNE et al. 1993) selbst zur Erhöhung beitragen kann. Geringere Aktivitäten sind nicht gleichzusetzen mit einem geringeren oxidativen Stress. WITTEK u. FÜRLL (2002) stellten bei Kühen mit LDA 3 und RDA cT mittels Pulsoximetrie am verlagerten Labmagen Störungen der Perfusion fest, die aber nur bedingt zum messbaren Anstieg der CK-Aktivität führten. - 85 - Diskussion __________________________________________________________________________________________ Superoxiddismutase-Aktivität (SOD-Aktivität) Die SOD zählt zu den enzymatischen Antioxidantien und gilt mit der GPX zusammen als Indikator für die Leistungsfähigkeit des antioxidativen Systems (WINNEFELD 1996). In Untersuchungen bei gesunden Kühen stellte SATTLER (2001) eine Abhängigkeit der SODAktivität zum Abkalbetermin fest. Eine Woche post partum lagen die Aktivitäten bei 6700 U/ml Erythrozytenlysat, vier Wochen post partum bei 7500 U/ml Erythrozytenlysat. Die niedrigen Aktivitäten eine Woche post partum sind auf Geburtsstress und verminderte Futteraufnahme in der Hochgravidität, die zu einer Schwächung des antioxidativen Systems führen, zurückzuführen (WEBER u. BRUCH 1992). Vier Wochen post partum ist die Futteraufnahme angestiegen und damit die Aufnahme an nichtenzymatischen Antioxidantien, was die Leistungsfähigkeit des antioxidativen Systems fördert (YOUNG et al. 2000) und zu höheren SOD-Aktivitäten führt (SATTLER 2001). Bei längerer Beanspruchung des antioxidativen Systems nimmt die SOD-Aktivität ab (WEBER u. BRUCH 1992), so bei Tumoren im Magen-Darm-Trakt (TOH et al. 2000) und bei oxidativem Stress (FUJI u. TANIGUCHI 1999). Auch nach Induktion einer Ischämie bei Rattennieren konnten verminderte SOD-Aktivitäten festgestellt werden (DAVIES et al. 1995). Als Verfeinerung der bisherigen Messmethodik wurde die SOD-Aktivität, die im Erythrozytenpellet bestimmt wird, auf die im Pellet enthaltene Hämoglobinkonzentration bezogen, um die bei der Herstellung des Pellets auftretenden Konzentrationsschwankungen des Erythrozytengehaltes zu berücksichtigen. Eine Vergleich ist über die Beziehung von Hämoglobin und Hämatokrit im Vollblut möglich. Die bislang in der Literatur angegebenen Aktivitäten beziehen sich auf die Messung der SOD-Aktivität im Vollblut (KÖNEN 1982), auf die Aktivität bezogen auf das Gesamtprotein der Probe (ZIMMERMANN 1983, HUSSAIN u. ALI 1999) oder auf die Aktivität bezogen auf den Hämatokrit (FÜRLL et al. 1999). Der Bezug auf das in der Probe enthaltene Hämoglobin wird auch von ZIMA et al. (1996) bei Untersuchungen zur Lungenreife von Frühgeburten und von BENGOUMI et al. (1998) bei Kühen und Kamelen propagiert. Zu beachten beim direkten Vergleich ist der Zeitpunkt der Aktivitätsbestimmung (siehe 3.5.2). Im Vergleich der Aktivitäten der SOD bei Tieren mit LDA mit den Aktivitäten aus der Gruppe der Tiere mit RDA wiesen alle Tiere mit LDA signifikant höhere Aktivitäten als alle Tiere mit RDA auf (p<0,05) (Abb. 22, 23). Auch bei dieser Diskussion muss das Abkalbedatum berücksichtigt werden, das bei den RDA sT in 75 % weniger als vier Wochen zurückliegt und somit Ursache für die geringeren SOD- 86 - Diskussion __________________________________________________________________________________________ Aktivitäten sein kann. Bei den RDA cT kann anhand der erhöhten Lactatkonzentrationen (Abb. 16, 17) von einer deutlichen Ischämie ausgegangen werden, die auch WITTEK et al. (2004) anhand von Pulsoximetrie und Sauerstoffsättigung am Labmagen bei Kühen mit RDA cT vorfand. Diese Ischämie kann Ursache für geringere SOD-Aktivitäten sein (DAVIES et al. 1995). Dabei kann es durch Ansammlung von Wasserstoffperoxid zur Inaktivierung der SOD kommen (WEBER u. BRUCH 1992). Die Inaktivierung ist reversibel, sie kann durch Beseitigung des Wasserstoffperoxides durch die Katalase (siehe 2.3.3.3) wieder regeneriert werden. Die Aktivitäten der Gruppen mit LDA entsprechen den von SATTLER (2001) festgestellten höheren Aktivitäten bei Kühen mit DA in Abhängigkeit vom Abkalbedatum und im Vergleich zu gesunden Kühen. Bei Kühen mit LDA innerhalb von vier Wochen post partum liegen höhere Aktivitäten vor als bei Kühen mit größerem Abstand zur Geburt (Tab. 14). Ursache dafür kann die verspätete Diagnose sein, die LDA kann schon über längere Zeit bestehen und gemeinsam mit verringerter Futteraufnahme und erhöhter Stoffwechselbelastung zu einem beanspruchten antioxidativen System mit niedrigeren SODAktivitäten führen. Bei den Kühen mit RDA cT, bei denen die Diagnose direkt erfolgte, sind nur geringe Unterschiede zwischen den Abständen zur Abkalbung aufzuweisen, die dem Abfall nach der Geburt und erst in der folgenden Trächtigkeit wieder ansteigende Aktivitäten bei Kühen mit DA entsprechen, wie er von SATTLER (2001) beschrieben wurde. Höhere SOD-Aktivitäten bei Kühen mit LDA im Vergleich zu gesunden Kühen können durch eine erhöhte Belastung infolge oxidativem Stress und durch Endotoxine hervorgerufen werden (ALLEN u. VENKATRAJ 1992, TSAN 1993, MAULIK et al. 1995). Frühere Untersuchungen zur Adaptation an Endotoxine und oxidativen Stress zeigten erhöhte Enzymaktivitäten, die bei starker, aber plötzlich auftretender Belastung nicht erreicht werden (ZÜLLKE 1993). Glutathionperoxidase-Aktivität (GPX-Aktivität) Die gemessenen GPX-Aktivitäten lagen auf einem Niveau von etwa 500 U/g Hb (Abb. 25, 26). Die Aktivität der selenhaltigen erythrozytären GPX ist abhängig von der Selenkonzentration im Blutserum (GEISHAUSER et al. 1995). GPX-Aktivitäten < 130 U/g Hb weisen in erster Linie auf ein Defizit in der Selen-Versorgung der Tiere hin. Höhere Aktivitäten wie in dieser Untersuchung setzen eine bedarfsgerechte Selen-Versorgung voraus (>70 g/l) (FLOHÉ 1997). Die GPX-Aktivitäten waren in allen Gruppen erhöht, die von SATTLER (2001) bei gesunden Kühen festgestellten Aktivitäten wurden wie alle in der Literatur beschriebenen Aktivitäten - 87 - Diskussion __________________________________________________________________________________________ bei gesunden Kühen deutlich überschritten (Tab. 5). Da die GPX mit der SOD als Indikator für die Leistungsfähigkeit des antioxidativen Systems angesehen wird (WINNEFELD 1996), kann von einer Belastung der GPX durch die DA ausgegangen werden. Eine Erhöhung der GPX-Aktivität wird dabei als Adaptation an oxidativen Stress angesehen (VAIDOVICH et al. 1993, BOR et al. 1999). Im Vergleich der rechts- und linksseitigen LMV waren die Aktivitäten der Tiere mit RDA cT signifikant höher (p<0,05) als die der Tiere mit LDA (Abb. 25, 26). Da Tiere mit RDA cT einem momentan höheren Stress ausgesetzt sind, d.h. dass bei einer verminderten Blutzufuhr mit nachfolgender Reperfusion massive Radikalfreisetzung und Gewebsschädigungen erfolgen, ist die höhere Aktivität der GPX bei den Tieren mit RDA als Anpassung an diesen oxidativen Stress anzusehen. Andere Faktoren, wie Transport, Alter (BAUMGARTNER 1983), Operation etc. als Grund für die Erhöhung scheiden aus, da alle Tiere mit LMV den gleichen Einflüssen unterlagen. Auffällig ist, dass die GPX-Aktivität sich gegenläufig zur SOD-Aktivität verhielt, die durch die Inaktivierung bei hohem oxidativem Stress nicht als Spiegel desselben dienen kann (WEBER u. BRUCH 1992, MAO et al. 1993). Bei den linksseitigen Verlagerungen, die für die Kühe einen längeren Zustand des oxidativen Stresses infolge der längeren Krankheitsdauer bedeuten, liegen die Aktivitäten der GPX zwar auch über dem Normbereich, sind aber nicht so deutlich erhöht wie die der SOD (Abb. 22, 23, 25, 26). Weiterführende Untersuchungen bestätigten, dass die GPX-Aktivität bei Kühen mit LDA p. op. weiter ansteigt, während dies bei der SOD nicht der Fall war (SATTLER u. FÜRLL 2002). Bei den Kühen mit RDA sT traten ähnliche GPX-Aktivitäten wie bei den Tieren mit LDA auf. Dies lässt darauf schließen, dass bei Kühen mit LDA und RDA sT das gleiche Ausmaß an oxidativem Stress vorliegt, die RDA sT also in dieser Beziehung den LDA zuzuordnen sind. Dies wird durch die Befunde der klinisch-chemischen Parameter (Tab. 13) und durch die Untersuchungen von WITTEK et al. (2004) anhand von Pulsoximetrie und Sauerstoffsättigung am Labmagen bestätigt. Der zeitliche Abstand zur Geburt wirkt sich nur bei den LDA 3 auf die GPX-Aktivität aus. Erkrankten Kühe später als einen Monat an LDA 3, hatten sie geringere GPX-Aktivitäten als Kühe innerhalb eines Monats post partum (Tab. 15). Dies lässt mit auf die Beanspruchung der GPX durch zusätzliche Erkrankungen des Genitale schliessen, die im ersten Monat post partum besonders häufig auftreten. Bei den Kühen mit RDA cT zeigten sich keine Unterschiede im zeitlichen Abstand zur Abkalbung, die hier auftretenden GPX-Aktivitäten sind auf die DA zurückzuführen (Tab. 15). - 88 - Diskussion __________________________________________________________________________________________ Trotz der scheinbaren Gegenläufigkeit von SOD und GPX, die unter anderem von SATTLER (2001) statistisch nachgewiesen wurde, kann nicht grundsätzlich von einer direkten negativen Korrelation in der Regulation der Enzyme ausgegangen werden. Hierzu durchgeführte Untersuchungen bei Mensch, Hase und Schwein ergaben, dass beide Enzyme unabhängig voneinander reguliert werden. Auftretendes gleichartiges oder gegenläufiges Verhalten ist auf die betreffenden Erkrankungen und deren Pathomechanismen zurückzuführen, ergibt aber keine ursächlichen Korrelationen. Es ist somit als „zufällig“ zu betrachten, da beiden Enzymen ein Bindeglied in der gegenseitigen Regulation fehlt (GODIN u. GARNETT 1992). Dem entgegengesetzt sind Katalase-Aktivität und GPX-Aktivität bei den genannten Tierarten negativ korreliert, Katalase-Aktivität und Glutathionreduktase sind positiv korreliert. Diese Korrelationen kommen durch das gemeinsame Bindeglied NADPH zustande, das durch Bindung an die Katalase zum Wegfall der Hemmung der Glutathionreduktase führt, deren Aktivität daraufhin steigt. Dies wiederum führt in den Zellen zum Anstieg von reduziertem Glutathion (GODIN u. GARNETT 1992). Die Aktivität der Katalase wiederum ist eng mit der SOD-Aktivität verbunden. Bei Inaktivierung der Katalase kommt es durch den Anstieg von Wasserstoffperoxid zur Inaktivierung der SOD. Dies bietet die Erklärung für eine scheinbare Korrelation von SOD-Aktivität und GPX-Aktivität (ZÜLLKE 1993). TEAC-Konzentration Die Konzentrationen der TEAC, gemessen in peripherem und abomasalem Blut, bewegten sich in einem Bereich von 11 bis 795 µmol/l (Abb. 28, 29). In einer zeitgleich durchgeführten Studie bei Kühen unterschiedlicher Leistungsebenen traten mit ähnlichen Schwankungen Konzentrationen im peripartalen Zeitraum von ca. 300 mol/l auf (WILKEN u. FÜRLL 2002). Dabei stiegen die Konzentrationen post partum kontinuierlich an. Auffällig war bei der Untersuchung die signifikant geringere TEAC-Konzentration bei Mutterkühen im Vergleich zu den Hochleistungskühen. Die individuellen TEAC-Konzentrationen weisen darauf hin, dass nur wenige Tiere einen ähnlichen antioxidativen Status besitzen. Dieses individuelle Basisniveau der totalen antioxidativen Kapazität ist auch beim Menschen bekannt. Neben geschlechtsspezifischen Variationen geht man hier bei Untersuchungen zur antioxidativen Kapazität vom Basislevel der einzelnen Probanden aus (KAIKKONEN et al. 2002). Beim Rind erklärt sich das individuelle Niveau über die unterschiedliche Fütterung, auch jahreszeitlich abhängig, in den verschiedenen Betrieben, den Abstand zur Geburt und zusätzliche Erkrankungen wie Endometritiden und Gelenkserkrankungen. Auch beim Menschen hängt der antioxidative Status von der Nahrungsaufnahme ab (CAO et al. 1998). In einer Studie von BASKIN et al. (2000) wurden Schlittenhunde einen Monat mit Vitamin E, - 89 - Diskussion __________________________________________________________________________________________ Carotin und Lutein supplementiert und wiesen nachfolgend signifikant erhöhte Konzentrationen dieser Antioxidantien im Serum auf. Auf äußere Einflüsse reagiert der Organismus aber von seinem individuellen Ausgangsniveau aus auf ähnliche Art und Weise. Innerhalb der Gruppen der LDA waren die TEAC-Konzentrationen der LDA 3 niedriger als die der LDA 2, im peripheren Blut war dieser Unterschied signifikant (p<0,05; Abb. 28, 29). Eine höhere TEAC-Konzentration bei Tieren mit LDA 2 im Vergleich zu Tieren mit LDA 3 kann bedeuten, dass bei einer LDA 2 eine geringere Beanspruchung der nichtenzymatischen Antioxidantien vorliegt oder vermehrt Antioxidantien aus Speichern mobilisiert worden sind. Setzt man die TEAC-Konzentrationen der Tiere mit LDA 2 in Bezug zu SOD- und GPXAktivität innerhalb dieser Gruppe, so fällt auf, dass auch beide Enzyme bei den LDA 2 höhere Aktivitäten als bei den LDA 3 aufwiesen. Dies lässt darauf schließen, dass bei den LDA 2 eine höhere Beanspruchung der antioxidativen Kapazität vorliegt und nichtenzymatische Antioxidantien aus Speichern mobilisiert worden sind. In Untersuchungen zu den Quellen der Mobilisation von Antioxidantien bei Marathonläufern wurde eine positive Korrelation von Glycerol und Vitamin E deutlich, die erst bei trainierten Läufern während des Marathons auftritt und auf die Fettmobilisation zurückzuführen ist (HUTLER et al. 2000). Dabei kann auch die Hochregulation der Antioxidantien über das oxyR-Gen eine Rolle spielen, die bei längerer Belastung des Körpers einsetzt. Die Aktivierung dieses Gens erfolgt zum Teil bereits nach zehn Minuten (HARRIS 1992 b). Die Ursache für die Mobilisation muss allerdings nicht die DA sein, da bei den LDA 2 89 % der Kühe innerhalb des ersten Monats post partum an DA erkrankten (Tab. 9) und der Anteil an Puerperalstörungen innerhalb dieser Gruppe genauso hoch war (Tab. 12). Ein weiteres Indiz stellt die Erhöhung der CK in dieser Gruppe dar (Abb. 19, 20). Innerhalb der Gruppe der LDA 3 erkrankten 69 % der Tiere innerhalb des ersten Monats post partum an DA, der Anteil der Tiere mit Puerperalstörung lag bei 62 %. Die größere Perfusionsstörung bei Tieren mit LDA 3 als bei Tieren mit LDA 2 (WITTEK u. FÜRLL 2002) mit erhöhter Radikalbildung als Ursache für den Konzentrationsunterschied scheidet aus, da hierbei auch unterschiedliche Lactatkonzentrationen auftreten müssten (Abb. 16, 17). Die Gruppe der RDA sT zeigte wie bei den SOD- und GPX-Aktivitäten ähnliche TEAC-Konzentrationen wie die Gruppe der LDA 3, die Zuordnung der RDA sT zu den LDA erscheint weiterhin sinnvoll. Bei den RDA cT war die TEAC-Konzentration im abomasalen Blut signifikant höher als die der LDA 3, was auf die gestörte Perfusion zurückzuführen ist, die sich auch anhand der Lactatkonzentrationen nachvollziehen lässt (Tab. 13). Hinzu kommt, dass den RDA cT eine kurze Erkrankungsdauer vorangeht und der Abstand zur Geburt bei 64 % der Kühe mit RDA cT mehr als vier Wochen betrug. WILKEN und FÜRLL (2002) - 90 - Diskussion __________________________________________________________________________________________ konnten eine ansteigende Tendenz der TEAC-Konzentration post partum aufzeigen. Dieser Trend zeigte sich auch innerhalb der LDA 3 (Tab. 16). Die positive Korrelation zwischen TEAC-Konzentration (p) und CK-Aktivität (p + lm) in der Gruppe aller Tiere mit LDA (siehe 4.2.6.1) zeigt, dass der Körper auch mittels nichtenzymatischer Antioxidantien in der Lage ist, sich an oxidativen Stress anzupassen, auch wenn dieser, wie im Fall der LDA, über längere Zeit andauert. In einer Studie von PIERCY et al. (2000) zeigten Schlittenhunde bei Belastung ebenfalls einen gleichgerichteten Anstieg der CK und der VitaminC-Konzentration, unabhängig von einer vorher erfolgten Supplementierung mit Vitamin C, E und -Carotin. 5.5 Vergleich der Ergebnisse nach der Herkunft des Blutes Die wiederkehrenden positiven Korrelationen der aus der V. jugularis und der V. epigastrica stammenden Messergebnisse der einzelnen Parameter legen zugrunde, dass im peripheren Blut grundsätzlich die gleichen oxidativen Belastungen wie am Labmagen vorherrschen. Treten allerdings bei den einzelnen Parametern in den Gruppen Unterschiede zwischen den Entnahmestellen der Proben auf, weisen sie auf Veränderung des Blutes auf dem Weg vom Labmagen zur peripheren Entnahme hin. Lactatkonzentration Die Lactatkonzentrationen im peripheren und abomasalen Blut korrelierten innerhalb der Gruppen LDA 2, LDA 3, RDA sT und RDA cT signifikant (p0,01, Abb. 18). Die zu erwartenden Unterschiede in der Gruppe der RDA cT, die im abomasalen Blut höhere Lactatkonzentrationen aufzeigten, traten nicht auf. Demzufolge muss ein Austausch zwischen peripherem und abomasalem Blut auch während der Verlagerung noch stattfinden. Creatinkinase-Aktivität Veränderungen zwischen peripherem und abomasalem Blut bei Kühen mit DA sind in Bezug auf die CK nur in geringem Umfang zu erwarten, da die CK-Aktivität, wie auch die ASATAktivität signifikant mit dem pathologischen Befund des Uterus korreliert (EULENBERGER 1984, KLEISER u. FÜRLL 1998, SATTLER 2001). Da in dieser Studie 54 % (Tab. 12) der Kühe von pathologischen Veränderungen des Genitale betroffen waren, die zur Freisetzung der CK aus den Zellen des Endometriums führen, muss zumindest von einer Überlagerung der durch die DA hervorgerufene Erhöhung der CK-Aktivität durch Freisetzung aus geschädigter Labmagenmuskulatur (GEISHAUSER 1995) ausgegangen werden. Ursachen für die - 91 - Diskussion __________________________________________________________________________________________ Schädigung der Labmagenmuskulatur können bei allen Kühen Überdehnung, aber auch Ischämie und Reperfusion sein (MAHMOUD u. FORD 1988, REDDY et al. 2000), wie sie bei RDA cT nachweislich auftreten (WITTEK u. FÜRLL 2002, WITTEK et al. 2004). Innerhalb der anderen Gruppen ist in Bezug auf die CK-Aktivität nicht mit einer Dominanz der Veränderungen am Labmagen über die Veränderungen am Genitale zu rechnen. Aber selbst in der Gruppe der RDA cT war dieser Unterschied in der vorliegenden Untersuchung nur angedeutet (Abb. 21), die CK-Aktivitäten im abomasalen Blut lagen bei 40 % deutlich über den Aktivitäten im peripheren Blut. Dabei muss allerdings beachtet werden, dass die CK-Aktivitätserhöhungen durch Transport (GROTH u. GRÄNZER 1975) sowie Operationsstress (AUTEFAGE et al. 2000) bei den Tieren mit RDA cT am Labmagen aufgrund des verringerten venösen Abflusses mit Rückstau verzögert ankommen und so im peripheren Blut den Veränderungen am Labmagen gegenüberstehen. In weiterführenden Untersuchungen könnte anhand der Herkunft der CK auch auf die Ursache der Aktivitätserhöhung geschlossen werden. Erste Untersuchungen haben ergeben, dass in der Labmagenwand hauptsächlich CK-BB zu finden ist, das bei Kühen mit DA bisher aber nur in einzelnen Fällen im Serum nachgewiesen werden konnte (NAURUSCHAT u. FÜRLL 2002). Der Hauptanteil der bei DA gemessenen CK-Aktivität entfällt zu zwei Dritteln auf die CKMM und zu einem Drittel auf die atypische CK. Superoxiddismutase-Aktivität Im Vergleich zur Herkunft des Blutes bestanden bei der SOD-Aktivität zwar keine signifikanten Unterschiede, aber dafür signifikante Korrelationen zwischen den Entnahmestellen in den Gruppen alle Tiere (p ≤ 0,01), alle LDA (p ≤ 0,01), LDA 3 (p ≤ 0,01) und alle RDA (p ≤ 0,05). Bei den rechtsseitigen Verlagerungen mit Torsion bestand diese Korrelation nicht (Abb.24). Hier ist davon auszugehen, dass am Labmagen durch Ischämie und Reperfusion zwar genug Gründe zur Steigerung der SOD vorliegen, diese aber nicht im Einklang mit der Aktivitätssteigerung des restlichen Körpers stehen müssen. Tendenzielle Unterschiede bestanden in der Gruppe der LDA 2 und der RDA sT, wobei die Enzymaktivitäten im peripheren Blut höher lagen (Abb. 24). Am Labmagen liegt während der Verlagerung ein verminderter venöser Blutabfluss vor. Dies führt zu einer Ansammlung von Oxidantien. Für das nach der Reposition neu durchfließende Blut bedeutet dies, neben der zusätzlichen Neuentstehung von Radikalen, eine Konfrontation mit diesen Oxidantien (Ischämie-/Reperfusionssyndrom) (COGHLAN et al. 1991).. Die SOD wird durch Wasserstoffperoxid ab einer bestimmten Konzentration allerdings inaktiviert - 92 - Diskussion __________________________________________________________________________________________ (WEBER u. BRUCH 1992). Dies erklärt die am Labmagen selbst etwas geringeren Aktivitäten der SOD. Glutathionperoxidase-Aktivität Die allgemein deutlich erhöhten Aktivitäten der GPX unterschieden sich nur bei den RDA cT, wobei abomasal niedrigere Aktivitäten vorlagen als peripher (Abb. 27). Zwischen den Entnahmestellen des Blutes bestand in allen Gruppen eine positive Korrelation (p ≤ 0,01) (Abb. 26). Der bei den RDA cT auftretende Unterschied zwischen peripherem und abomasalem Blut lässt sich durch die Hämostase während der Verlagerung erklären. So kann die oxidative Belastung von Transport und Operation (GROTH u. GRÄNZER 1975) im peripheren Blut schon zur Steigerung der GPX-Aktivität beitragen. TEAC-Konzentration Im Vergleich der TEAC-Konzentration (p) mit der TEAC-Konzentration (lm) zeigten die Kühe mit LDA und RDA sT peripher tendenziell höhere Konzentrationen, während bei den Kühen mit RDA cT abomasal höhere TEAC-Konzentrationen bestanden (Abb. 30). Peripher höhere Konzentrationen der TEAC bei Kühen mit LDA und RDA sT sind aus verschiedenen Gründen zu erklären. Zum Einen entstehen am Labmagen selbst mehr Radikale, die dort zu einem erhöhten Verbrauch an Antioxidantien führen. Zum anderen passiert das Blut auf dem Weg zur peripheren Entnahme die Leber und intakte Zirkulationsbereiche, wo es verdünnt und zum Teil bereinigt wird. Allerdings sollte dieser Unterschied zum Ort des Geschehens sehr viel deutlicher ausfallen. Beispielsweise konnte in einer Untersuchung bei Schlittenhunden nach Trainingsrennen gezeigt werden, dass bereits fünf Minuten nach Beginn der Reperfusion im peripheren Plasma signifikant höhere antioxidative Kapazitäten als am Ort der Ischämie/Reperfusion zu finden waren (JUVONEN et al. 1997). Dies deutet darauf hin, dass weder Ischämie noch Reperfusion bei Kühen mit LDA bzw. RDA sT in dem Maße ablaufen, als dass die zu erwartenden Konsequenzen eintreten. Dies wird auch anhand der Studie von WITTEK et al. (2004) deutlich, der nur bei Kühen mit RDA cT am Labmagen eine verminderte Sauerstoffsättigung und erhöhte Lactatansammlung als Zeichen einer geringeren Durchblutung und einer Hämostase feststellen konnte. Treten Ischämie-/Reperfusionssyndrom und Hämostase nur bei den RDA cT stärker in Kraft, so können kurz nach Rückverlagerung am Labmagen selbst tatsächlich noch höhere Konzentrationen an Antioxidantien als im peripheren Blut vorliegen, da zum Einen die - 93 - Diskussion __________________________________________________________________________________________ Radikalbildung noch nicht ihr volles Ausmaß erreicht hat und die Kommunikation mit dem restlichen Kreislauf, der von der Operation beeinträchtigt ist (AUTEFAGE et al. 2000), noch nicht wieder komplett hergestellt ist. 5.6 TEAC-Konzentrations-Verlauf post operationem Die Darstellung des TEAC-Konzentrationsverlaufes bei Kühen mit DA (Abb. 31-45) kann aufgrund der vielfältigen Beeinflussungen des antioxidativen Systems durch Herkunft der Kühe (WILKEN u. FÜRLL 2002), Futteraufnahme (CAO et al. 1993), Jahreszeit (SATTLER 2002, ZAHN u. FÜRLL 2004), Abstand zur Geburt (WILKEN u. FÜRLL 2002), Kreislaufsituation und zusätzliche Erkrankungen nur begrenzt zu einem einheitlichen Verlauf zusammengefasst werden. Die verschiedenen Einflussmöglichkeiten fügen sich bei jeder Kuh zu einem individuellen Profil zusammen, das bei der TEAC-Konzentration seine Auswirkungen zeigt. So werden bei jeder Kuh Abkalbedatum, zusätzliche Erkrankungen, Lactatkonzentration im peripheren Blut zur Einschätzung der Perfusionsstörung sowie periphere Harnstoffkonzentration zur Beurteilung der Kreislaufsituation aufgeführt und daraus folgende Veränderungen der TEACKonzentration abgeleitet. TEAC-Konzentrations-Verlauf bei Tieren mit LDA Bei den LDA (Abb. 31-35) gilt zu beachten, dass sie über längere Zeit bestanden und der Körper damit die Möglichkeit hatte, sich auf ein gesteigertes Maß an oxidativem Stress einzustellen (HUTLER et al. 2000, PIERCY et al. 2000, DAWSON et al. 2002). Nach bisherigen Erkenntnissen reagiert der Organismus auf erhöhten oxidativen Stress auch mit einem Anstieg der plasmatischen nichtenzymatischen Antioxidantien, solange die Reserven nicht erschöpft oder verbraucht sind. Die Kühe 7, 8 und 18 (Abb. 31, 32, 34) zeigten neben der LDA jeweils erhebliche Erkrankungen des Genitale, die zum Verbrauch der nichtenzymatischen Antioxidantien führen können. Die TEAC-Konzentrationen dieser Kühe stiegen auch nach der Operation nicht über 400 mol/l. Im Vergleich hierzu fällt auf, dass bei den Kühen 14 und 21 (Abb. 33, 35) keine Erkrankungen des Genitale vorlagen, die Geburt länger als vier Wochen zurücklag und die TEAC-Konzentrationen postoperativ deutlich über 400 mol/l steigen konnten. Dies kann mit als Anzeichen für den Verbrauch nichtenzymatischer Antioxidantien bei Kühen durch Erkrankungen des Genitale wie Endometritiden gewertet werden. Die Lactatkonzentrationen der Kühe mit LDA waren nur gering erhöht (<4 mmol/l, Abb. 31-35). - 94 - Diskussion __________________________________________________________________________________________ Es kann daher von geringen Perfusionsstörungen bei diesen Kühen ausgegangen werden, die nach Reposition folgende Perfusion bleibt ohne sichtbaren Effekt auf den antioxidativen Status. Die Harnstoffkon-zentrationen der Kühe waren ebenfalls nur gering erhöht (<6,5 mmol/l, Abb. 31-35), was eine Hämokonzentration mit daraus folgender Erhöhung der TEAC-Konzentration ausschließen lässt. Da die Kühe dieser Untersuchung aus unterschiedlichen Betrieben mit verschiedenen Fütterungsregimen und Haltungsbedingungen stammen, besitzt jede Kuh zu Beginn der Verlaufsuntersuchung ein „individuelles“ Niveau der TEAC-Konzentration. Um die Verläufe miteinander vergleichen zu können, wurden nur die Änderungen vom jeweiligen Ausgangsniveau betrachtet (Abb. 36). Individuelle Beeinflussungen durch zusätzliche Erkrankungen und Kreislaufdefizite wurden hierbei nicht berücksichtigt. Dabei deutet sich an, dass in den ersten fünf Stunden nach der Operation im peripheren Blut nur minimale Schwankungen der TEAC-Konzentration zu verzeichnen sind. Auch wenn der Abfall zu Beginn und der darauf folgende Anstieg aus Sicht der Anpassung des antioxidativen Systems mit einem erhöhten Verbrauch während und kurz nach der Operation und nachfolgender Regeneration gut nachzuvollziehen sind, sind diese Änderungen der antioxidativen Kapazität < 50 µmol/l und damit eher unbedeutend. In dieser Zeit, nach Beendigung der Operation, befanden sich die Tiere weitestgehend ungestört in Einzelboxen, versorgt mit intravenösen Infusionen (NaCl, 0,9%ig, + 760 g Glucose) und Antibiotika. Diese Behandlung, ohne zusätzliche Supplementierung von Antioxidantien oder Glucocorticoiden, reichte bei den untersuchten Kühen aus, um den Organismus bei der Regeneration zu unterstützen. Sie wirkt sich zudem fördernd auf die antioxidative Kompensation aus, so dass die Antioxidantien nicht erschöpft werden. Auch bei Hunden mit Magendrehung erwies sich postoperativ die Versorgung mit Kochsalzlösung (NaCL, 0,9%ig) mit als entscheidendes Kriterium für die Bewältigung der Auswirkung des Ischämie-/Reperfusionssyndroms (LANTZ et al. 1992). Erst nach fünf Stunden stieg die TEAC-Konzentration um mehr als 100 µmol/l an, kehrte aber innerhalb von zwei Stunden auf das Ausgangsniveau zurück. Eine Stunde später war wiederum ein Anstieg um fast 100 µmol/l zu verzeichnen, der bis zum Ende der Messzeit wieder ausgeglichen wurde. Diese Veränderungen können mit auf die Aktivierung der körpereigenen Abwehr (s. 2.1.1.4) zurückgeführt werden, die durch die Lyse von Zellbestandteilen zur Produktion von Radikalen beiträgt. Dabei gilt zu beachten, dass sowohl am Labmagen durch die Verlagerung als auch durch die Operation an sich entzündliche Vorgänge in Gang gesetzt werden. Die Anpassung an oxidativen Stress durch plasmatische Antioxidantien ist auch ohne vorbereitende Maßnahmen möglich. Bei Untersuchungen an - 95 - Diskussion __________________________________________________________________________________________ Marathonläufern zeigte sich, dass auch nur gering trainierte Menschen auf die enorme Belastung mit einer Erhöhung der plasmatischen antioxidativen Kapazität von ca. 15 % reagieren können, bevor diese nicht mehr in der Lage ist, den oxidativen Stress zu kompensieren (KAIKKONEN et al. 2002). Die Rückkehr auf das „individuelle“ Niveau der einzelnen Tiere ist vielfach in anderen Studien zum Verlauf der nichtenzymatischen Antioxidantien nach Phasen oxidativen Stresses beschrieben, so auch bei Untersuchungen zum TRAP im zeitlichen Verlauf nach Ischämie und Reperfusion am Rattendarm (SLAVIKOVA 1998). Um die postoperative Regeneration günstig zu beeinflussen, können den Kühen nach der Operation nichtenzymatische Antioxidantien, wie Vitamin C verabreicht werden, die ein Absinken der TEAC-Konzentration und damit der antioxidativen Kapazität verringern oder verhindern könnten. TEAC-Konzentrations-Verlauf bei Tieren mit RDA Bei den rechtsseitigen Verlagerungen ist die gemeinsame Diskussion der TEACKonzentration nicht vorteilhaft. Dagegen spricht der unterschiedliche Grad der Verlagerung (0° bis >360°), der im Gegensatz zu den linksseitigen Verlagerungen deutliche Auswirkungen auf die Blutzirkulation und die Ansammlung von Lactat hat (WITTEK et al. 2004, Abb. 3745). Die Kühe 19, 25, 32, 34 und 35 (RDA cT, Abb. 37, 39, 41, 43, 44) zeigten nach der Reposition des Labmagens einen deutlichen Anstieg der TEAC-Konzentration von bis zu 400 mol/l. Bei ihnen fielen zudem deutlich erhöhte Lactatkonzentrationen (>5mmol/l) auf, die auf Perfusionsstörungen des Labmagens durch die Torsion (180 °) zurückzuführen sind. Bei der Reperfusion nach der Reposition kommt es zur Radikalbildung (siehe 2.1.2.1), woraufhin die TEAC-Konzentration ansteigt. Dieser Anstieg fällt nachfolgend wieder ab, was den Zusammenhang Lactatkonzentrationen mit der zeigten die Reperfusion Tiere bis verdeutlicht. auf Kuh Neben 35 (Abb. den 44) erhöhten erhöhte Harnstoffkonzentrationen, die das Kreislaufdefizit der Tiere verdeutlichen. Durch diese Hämokonzentration werden die z.T. extremen Anstiege der TEAC-Konzentration noch potenziert. Zusätzliche Erkrankungen, wie Endometritiden fallen bei diesen RDA cT weniger ins Gewicht, da die Geburt bis auf Kuh 7 länger zurücklag. Kuh 24 (Abb. 38) gliederte sich als RDA sT in den Verlauf der LDA ein, bei ihr waren weder Lactat- noch Harnstoffkonzentration erhöht, die Endometritis trat als einziger störender Faktor auf. Der Anstieg der TEAC-Konzentration nach sechs bis acht Stunden entspricht dem Verlauf der LDA 3. - 96 - Diskussion __________________________________________________________________________________________ Kuh 31 (Abb. 40) als RDA sT durch Obstipation mit Sand zeigte wie oben genannte RDA cT erhöhte Lactat- und Harnstoffkonzentrationen, die auf Perfusionsstörungen und ein Kreislaufdefizit hinweisen. Dementsprechend erfolgte auch bei ihr nach Reposition des Labmagens durch die Reperfusion ein Anstieg der TEAC-Konzentration. Die nach der Operation auftretenden Digestionsstörungen deuten auf Dilatation und Atonie des Labmagens hin, wie sie von GEISHAUSER 1995 beschrieben wurden und durch Radikalbildung mit beeinflusst werden. Bei Kuh 37 (Abb. 45) fiel das durchweg niedrige Niveau der TEAC-Konzentration auf, das trotz lang zurückliegender Geburt und erhöhten Lactat- und Harnstoffkonzentrationen bestand. Bei ihr musste die zusätzliche Erkrankung in Form der hochgradigen Lahmheit berücksichtigt werden, von der nicht bekannt ist, wie lange sie schon bestand und was sie ausgelöst hatte. Da keinerlei äußere Anzeichen mehr auffielen, die nur auf kurze Erkrankungsdauer hindeuteten, muss davon ausgegangen werden, dass die Kuh über einen längeren Zeitraum weniger Futter aufgenommen haben kann und die Entzündung zum Verbrauch der nichtenzymatischen Antioxidantien führte. Kuh 25 (Abb. 39) zeigte den Verlauf einer RDA cT, der zum Verlust des Tieres führte. Die TEAC-Konzentration zeigte nach Reposition den typischen Anstieg, aber auch nachfolgend weitere Anstiege, die zwar auf die in der pathologischen Untersuchung festgestellten Ulzerationen und die Peritonitis hinweisen können, auf die aber aufgrund des TEACVerlaufes nicht geschlossen werden kann. Wie bei den Kühen mit LDA kommt es bei den RDA cT (Abb. 37-45) sechs bis zehn Stunden nach der Operation wieder zum Anstieg der TEAC-Konzentration vom Ausgangsniveau, die zwar zeitlich verzögert zu denen der Tiere mit LDA auftraten und auch weitaus höhere Schwankungen darstellen, die aber ebenso als Zeichen einer Entzündungsreaktion gewertet werden können. Dass die Schwankungen höher ausfallen als bei den Tieren mit LDA, kann auf einen zusätzlichen Entzündungsherd hinweisen, wobei auch hier der Labmagen als stärker betroffenes Organ nahe liegt. Nachfolgend ist wie bei den linksseitigen Verlagerungen eine Rückkehr zum Ausgangsniveau gegen Ende der Messzeit zu verzeichnen. Zusammenfassend für den Verlauf der TEAC-Konzentration bei Kühen mit RDA ist zu sagen, dass sich die starke Belastung mit oxidativem Stress infolge von Ischämie und Reperfusion in Form von Schwankungen der TEAC-Konzentration ausdrückt, die z.T. ein mehrfaches des Ausgangsniveaus betrugen. Daher kann eine Unterstützung mittels nichtenzymatischer Antioxidantien nur befürwortet werden. Die intravenöse Verabreichung von nichtenzymatischen Antioxidantien wie Vitamin C und E kurz vor Entgasung und - 97 - Diskussion __________________________________________________________________________________________ Rückverlagerung ist aufgrund der Verlaufsuntersuchung als günstig anzusehen. Im weiteren Regenerationsverlauf erscheint eine weitere Gabe nicht zwingend notwendig. 6. Zusammenfassung Untersuchungen zum antioxidativen Status bei verschiedenen Formen der Dislocatio abomasi des Rindes im Blut der V. jugularis und der V. epigastrica Gregor Dinges Medizinische Tierklinik der Veterinärmedizinischen Fakultät der Universität Leipzig März 2004 (125 Seiten, 45 Abbildungen, 16 Tabellen, 246 Literaturangaben, 1 Seite Anhang) Die Entstehung und Auswirkungen von Radikalen am Labmagen bei Kühen mit Dislocatio abomasi (DA) gelten mit als zentraler Faktor für Störungen der Labmagenmotorik und damit für die postoperative Regeneration der Kühe. Ziel der vorliegenden Arbeit war, die Auswirkungen der verschiedenen Formen der Labmagenverlagerung des Rindes auf den antioxidativen Stoffwechsel des Labmagens und des Gesamtorganismus unter Berücksichtigung der Stoffwechsellage und zusätzlicher Erkrankungen zu vergleichen. Zur Einschätzung des anti-oxidativen Status dienten die Aktivitäten der Superoxiddismutase (SOD) und der Glutathionperoxidase (GPX) sowie die TEAC-Konzentration („TROLOX equivalent antioxidative capacity“ -Konzentration) zur Beurteilung der nichtenzymatischen wasserlöslichen Antioxidantien im Plasma. Weiterhin wurden die klinischen und die Stoffwechselbefunde der Kühe erhoben. Versuchsanordnung: Bei 22 Kühen mit links- und 15 Kühen mit rechtsseitiger Labmagenverlagerung (LDA bzw. RDA) wurden vor der operativen Reposition des - 98 - Zusammenfassung __________________________________________________________________________________________ Labmagens die klinischen Befunde erhoben und Blut aus der V. jugularis zur Bestimmung der ß-Hydroxybutyrat- (BHB-), Cholesterol-, Gesamtbilirubin-, Totalprotein-, Harnstoff- , Natrium- und Kaliumkonzentrationen sowie der Aktivitäten der GLDH und der ASAT genommen. Die Tiere mit LDA wurden nach dem Schweregrad in LDA 2 und LDA 3 eingeteilt, die Tiere mit RDA in RDA ohne (RDA sT) und mit Torsion (RDA cT). Unmittelbar vor und nach Reposition des Labmagens wurden intraoperativ peripher (p) und direkt am Labmagen (lm) in Ausläufern der V. epigastrica dextra venöse Blutproben entnommen. Aus dem daraus gewonnenen Plasma wurden Chlorid-, Lactat- und TEACKonzentrationen sowie die Aktivitäten der Creatinkinase (CK) ermittelt. Aus Erythrozytenpellets der gleichen Probenentnahme wurden die Aktivitäten von GPX und SOD sowie die Hämoglobinkonzentrationen in den Pellets erfasst. Zusätzlich wurde der Verlauf der TEAC-Konzentration im peripheren Plasma 12 (LDA) bzw. 16 (RDA) Stunden p. op. verfolgt. Ergebnisse: Fett- und Leberstoffwechsel bei Kühen mit LDA und RDA sT waren stärker belastet als bei Kühen mit RDA cT. Dabei unterlagen die Tiere mit LDA 2 in vorliegender Studie den größten Beeinflussungen. Sie hatten zudem den größten Anteil zusätzlicher Erkrankungen und befanden sich zu 89 % im ersten Monat post partum. Bei den Kühen mit RDA cT dagegen war der Mineralstoffwechsel stärker belastet als bei Kühen mit LDA und RDA sT. Hier traten zudem die höchsten Harnstoff- (5,3; 7,5; 12,1 mmol/l) und Lactatkonzentrationen (3,4; 6,14; 8,6 mmo/l) auf. Die CK zeigte in den Gruppen der LDA 2 (380; 489; 1110 U/l) und der RDA sT (316; 424; 814 U/l) die höchsten Aktivitäten. Die SOD-Aktivitäten der Kühe mit LDA (2194; 2191; 2309 U/g Hb) waren höher als die der Kühe mit RDA (1667; 2061; 2449 U/g Hb), wobei in Bezug auf den zunehmenden Abstand zum Partus die Aktivitäten in der Gruppe der LDA 3 abnahmen. Bei der GPX lagen innerhalb der RDA (508; 560; 592 U/g Hb) höhere Aktivitäten vor, auch hier lagen bei Kühen mit größerem Abstand zum Partus in der Gruppe der LDA 3 niedrigere Aktivitäten vor. Die TEAC-Konzentrationen waren in den Gruppen der LDA 2 (302; 361; 403 µmol/l) und der RDA cT ( 235; 270; 346 µmol/l) am höchsten, Kühe mit LDA 3 zeigten mit zunehmendem Abstand zum Partus höhere, Kühe mit RDA cT niedrigere TEAC-Konzentrationen. Im Vergleich von peripherem zu abomasalem Blut konnten keine signifikanten Unterschiede festgestellt werden. Im postoperativen Verlauf stiegen die TEAC-Konzentrationen bei Kühen mit LDA 3 ca. fünf und acht Stunden post operationem an, kehrten aber wieder auf das Ausgangsniveau zurück. Bei Kühen mit RDA, die deutlich erhöhte Lactat- und Harnstoffkonzentrationen hatten, war - 99 - Zusammenfassung __________________________________________________________________________________________ zusätzlich nach der Rückverlagerung ein Anstieg der TEAC-Konzentration zu verzeichnen, der nachfolgend wieder abfiel. Schlussfolgerungen: Der antioxidative Status bei Kühen mit DA unterliegt nicht nur der Beeinflussungen durch die DA, sondern auch durch zusätzliche Erkrankungen und durch den Abstand zum Partus. Dies zeigt sich deutlich in den Gruppen der LDA 2 und der LDA 3. Die Kühe mit RDA sT sind hinsichtlich ihrer oxidativen Belastungen den LDA vergleichbar. Bei den RDA cT kommt es zu Perfusionsstörungen, die für die Ansammlung von Lactat verantwortlich sind. Das antioxidative System zeigt bei ihnen höhere Aktivitäten der GPX und höhere Konzentrationen der TEAC, die geringere SOD-Aktivität kann Ausdruck einer vorübergehenden Inaktivierung sein. Im TEAC-Verlauf post operationem kommt es bei den RDA cT mit den größten Perfusionsstörungen nach der Reposition des Labmagens zu einem Anstieg, der auf den Anstrom von Radikalen während der Reperfusion zurückzuführen ist. 7. Summary Studies on antioxidative metabolism in cows with different forms of the dislocatio abomasi (DA) in blood samples from jugular and epigastric vein Gregor Dinges Clinic for Large Animal Internal Medicine Faculty of Veterinary Medicine of the University of Leipzig March 2004 (125 pages, 45 figures, 16 tables, 246 references, 1 page appendix) The formation and the effects of radicals on the abomasum in cows with dislocatio abomasi (DA) are regarded as one of the main factors for disturbed abomasal motility and the postoperative regeneration. The aim of this study was to compare the effects of the different forms of DA in cows on the abomasal antioxidative metabolism and the whole organism under consideration of the metabolic status and additional diseases. To asses the antioxidative status the activities of superoxide dismutase and glutathione peroxidase, and the concentration of the TEAC (“TROLOX equivalent antioxidative capacity”) for the non-enzymatic, water soluble plasmatic antioxidants were determined. Additionally, clinical and metabolic parameters were measured. - 100 - Summary __________________________________________________________________________________________ Methods: Before surgical reposition of the abomasum 22 cows with left and 15 cows with right displaced abomasum (LDA or RDA) were physically examined. Blood samples were taken from the jugular vein to measure the concentrations of β-hydroxybutyrate (BHB), cholesterol, total bilirubin, total protein, urea, sodium, potassium and the activities of glutamatedehydrogenase (GLDH) and aspartate-amino-transferase (ASAT). The animals with LDA were divided into groups depending on the degree of displacement (LDA 2 and LDA 3), the animals with RDA into RDA without (RDA sT) and with abomasal volvulus (RDA cT). Immediately before and after surgical reposition of the abomasum, blood was taken from the jugular and the epigastric vein. The plasma concentrations of chloride, lactate and TEAC and the activities of the creatine kinase (CK) were measured. The activities of GPX and SOD and the concentrations of haemoglobin were determined in erythrocyte pellets of these samples. Moreover, the time course of the concentration of the TEAC was observed in the peripheral plasma over 12 (LDA) or 16 (RDA) hours. Results: In cows with LDA and RDA sT, the lipid and liver metabolism were more disturbed in comparison to cows with RDA cT. In this study cows with LDA 2 were heavily influenced. They had the highest portion of additional diseases and 89% of them were in the first month after parturitions. In the cows with RDA cT the metabolism of minerals was more influenced. Additionally, they showed the highest concentrations of urea (5.3; 7.5; 12.1 mmol/l) and lactate (3.4; 6.14; 8.6 mmo/l). The highest activities of the CK were found in the groups with LDA 2 (380; 489; 1110 U/l) and RDA sT (316; 424; 814 U/l). In cows with LDA the activities of the SOD (2194; 2191; 2309 U/g Hb) were higher than in cows with RDA (1667; 2061; 2449 U/g Hb). The cows with LDA 3 which were longer in milk had lower activities of the SOD. The cows with RDA (508; 560; 592 U/g Hb) had higher GPX activities than cows with LDA, but the activities decreased with increasing lactation days. The groups with LDA 2 (302; 361; 403 µmol/l) and RDA cT (235; 270; 346 µmol/l) showed the highest concentrations of the TEAC. The concentrations of TEAC increased with increasing time after parturition in cows with LDA 3. In contrast the concentration of TEAC decreased with the time after parturition in cows with RDA cT. There were no significant differences comparing the peripheral and the abomasal blood parameters The TEAC concentration increased five and eight hours post operationem in the group of the LDA 3, but decreased to the basic level later. In cows with RDA cT, there was an additional transient increase immediately after the reposition of the abomasums. - 101 - Summary __________________________________________________________________________________________ Conclusions: In cows with DA the antioxidative status was not only influenced by the DA but concurrent diseases and the duration of lactation had additional effects. This was shown in the groups with LDA 2 and LDA 3. Cows with RDA sT were similar to cows with LDA. Only cows with RDA cT suffered from disturbances of the abomasal perfusion, which resulted in increased lactate concentrations. In these cows the activities of GPX and the concentrations of the TEAC were higher. It is believed that inactivation was reason for the lower activities of the SOD in cows with RDA cT. The highest values of TEAC concentration after reposition of the abomasum were measured in cows which had the strongest disturbances of the perfusion. The influx of radicals during the phase of reperfusion after reposition of the abomasums is considered as reason. 8. Literaturverzeichnis AKGÜR, F. M., M. OLGUNER, O. YENICI, M. GÖKDEN, T. AKTUG, M. YILMAZ u. G. ATAC (1996): The effect of allopurinol pretreatment on intestinal hypoperfusion encountered after correction of intestinal volvolus. J. Pediatr. Surg. 31, Nr. 9, 1205 - 1207 ALEKOS, T. (1992): Der Einfluss der Selenkonzentration in der Nahrung auf die Aktivität der GPS-Transferase und der Glutathionperoxidase. Berlin, Freie Univ., Fachber. Veterinärmed., Diss. AL-GOUHL, W. M., R. B. MEEKER u. R. S. GREENWOOD (1995): Kindling induces a longlasting increase in brain nitric oxide synthase activity. Neuroreport 6, 457 - 460 ALLEN, R.G. u. V. S. VENKATRJ (1992): Oxidants and antioxidants in development and differentiation. J. Nutr. 122, 631 - 635 ALLISON, R. C., E. M. HERNANDEZ, V. R. PRASAD, M. B. GRISHAM u. A. E. TAYLOR (1988): Protective effects of O2 radical scavengers and adenosine in PMA-induced lung injury. J. Appl. Physiol. 64, 2175 - 2182 ANSORGE, K. u. B. - O. GEMKOW (1992): - 102 - Literaturverzeichnis __________________________________________________________________________________________ Zum Einfluss von Superoxiddismutase, Oxypurinol und Iloprost auf die Wirkung freier Sauerstoffradikale bei Konservierung und Reperfusion isolierter Dünndarmtransplantate. Eine tierexperimentelle Studie. Berlin, Univ., Med. Fak., Diss. AUPPERLE, H., H. A. SCHOON u. M. FÜRLL (1998): Pathologisch-anatomische Befunde bei Dislocatio abomasi und deren klinische Konsequenzen. Internationaler Workshop Ätiologie, Pathogenese, Diagnostik, Prognose, Therapie und Prophylaxe der Dislocatio abomasi, Leipzig 1998, Proceed., 195 - 197 AUTEFAGE, A., H. P. LEFEBVRE, F. ALVANITAKIS u. J. P. BRAUN (2000) : Non-invasive quantifikation of muscle damage during surgery in the dog. Rev. Med. Vet. 151, Nr. 7, 661 AVISSAR, N., E. A. KERL, S. S. BAKER u. H. J. COHEN (1994): Extracellular glutathione peroxidase mRNA and protein in human cell lines. Arch. Biochem. Biophys. 309, Nr. 2, 239 - 246 BAADER, W. J., P. ROMOFF, H. P. CARAVIELLO u. E. L. BASTOS (2000): A novel method for the evaluation of natural products antioxidant activity. Clin. Lab. 7+8, 388 BACKALL, K. A. u. R. W. SCHOLZ (1979): Reference values for a field test to estimate inadequate glutathione peroxidase activity and selenium status in blood of cattle. Am. J. Vet. Res. 30, Nr. 5, 733 - 738 BAJCSY, Á. C.,J. REHAGE, H. SCHOLZ u. O. SZENCI (1997): Changes in blood ionized calcium and some other blood parameters before and after replacement of a left-sided displaced abomasum in dairy cattle. Dtsch. Tierärztl. Wschr. 104, Nr. 12, 527 - 529 BALTHAZARY, S. T., H.-P. SALLMANN u. H. FUHRMANN (1999): The determination of TBA- reactive substances and alkenals in the presence of antioxidants. Acta Veterinaria Hungarica 47, Nr. 2, 155 - 159 BARTLEY, J. C. (1989): Lipid metabolism and its diseases. In: Clinical Biochemistry of Domestic Animals (Ed. by J. J. KANEKO), Academic Press, Inc., San Diego (California), 106 - 141 BASKIN, C. R., K. W. HINCHCLIFF, R. A. DISILVESTRO, G. A. REINHART, M. G. HAYEK, B. P. CHEW, J. R. BURR u. R. A. SWENSON (2000): Effects of dietary antioxidant supplementation on oxidative damage and resistance to oxidative damage during prolonged exercise in sled dogs. Am. J. Vet. Res. 61, Nr. 8, 886 - 891 BAUER, P., F. BELLEVILLE-NABET, F. WATELET, F. DUBOIS u. A. LARCAN (1995): - 103 - Literaturverzeichnis __________________________________________________________________________________________ Selenium, oxygen-derived free radicals, and ischemia-reperfusion injury. An experimental study in the rat. Biol. Trace. Elem. Res. 47, Nr.1 - 3, 157 - 163 BAUMGARTNER, M. (1983): Untersuchungen zur Aktivität der Glutathionperoxidase im Plasma von Pferden. Berlin, Univ., Fachber. Veterinärmed., Diss. BECVAR, O., J. ILLEK, J. STERC u. R. BARTOS (2001): Experience in surgical treatment of abomasal displacement in cattle. In: Ref. III. Mitteleurop. Buiatr. Kongr.: Gesundheitsproblematik der Wiederkäuer, Milovy, S. 163 - 168 BEKELE, M. (1997): Untersuchungen zur postpartalen Leberverfettung beim Rind. Leipzig, Univ., Vet.-Med. Fak., Diss. BELL, J. G. u. C. B. COWEY (1982): Comparative studies on glutathione peroxidase from the liver of rainbow trout (Salmo gairdnerii). Biochem. Soc. Trans. 10, 451 BENGOUMI, M., K. ESSAMADI, J.-P. CHARCONAC, J.-C. TRESSOL u. B. FAYE (1998): Comparative relationship between copper-zinc plasma concentrations and superoxide dismutase activity in camels and cows. Inra/Elsevier, Paris BERGHAUS, K. (1997): Superoxidanionenbildung, Stoffwechsel- und Funktionsalterationen in der isolierten Rattenleber während Minderperfusion und anschließender regionaler Reperfusion sowie ihre Beeinflussung durch Superoxiddismutase. Giessen, Univ., Fachber. Veterinärmed., Diss. BIELEFELDT, K. u. J. L. CONKLIN (1997): Intestinal motility during hypoxia and reoxygenation in vitro. Dig. Dis. Sci. 42, Nr. 5, 878 - 84 BOR, M. V., C. CEVIK, I. USLU, F. GUNERAL u. E. DUZGUN (1999): Selenium levels and glutathione peroxidase levels in patients with acute myocardial infarction. Acta. Cardiol. 54, Nr. 5, 271 - 276 BOROS, M., L. BAKO u. S. NAGY (1991): Effect of antioxidant therapy on cyclooxygenase-derived eicosanoid release during intestinal ischemia-reperfusion. Eur. Res. Surg. 23, Nr. 3-4, 141 - 150 BOROS, M., J. KASZAKI u. S. NAGY (1989): - 104 - Literaturverzeichnis __________________________________________________________________________________________ Oxygen free radical-induced histamine release during intestinal ischemia and reperfusion. Eur. Surg. Res. 21, Nr. 6, 297 - 304 BOURRE, J.-M., O. DUMONT, M. CLEMENT, L. DINH, M.-T. DROY-LEFAIX u. Y. CHRISTEN (2000): Vitamin E deficiency has different effects on brain and liver phospholipid hydroperoxide glutathione peroxidase activities in the rat. Neurosci. Lett. 286, 87 - 90 BRADY, P. S., L. J. BRADY, D. E. ULLREY u. D. R. ROMSOS (1978): Interrelationships of erythrocyte glutathione peroxidase system enzymes in the fasted beagle. Am. J. Vet. Res. 39, Nr. 2, 267 - 270 BRIGELIUS-FLOHÉ, R. (1999): Tissue-specific functions of individual glutathione peroxidases. Free Radic. Biol. Med. 27, Nr. 9 - 10, 951 - 965 BROWN, J. M., M. A. GROSSO, L. S. TERADA, C. J. BEEHLER, K. M. TOTH, G. J. WHITMANN, A. H. HARKEN u. J. E. REPINE (1989): Erythrocytes decrease myocardial hydrogen peroxide levels and reperfusion injury. Am. J. Physiol. 256, H584 - H588 BULKLEY, G. B. (1987): Free radical-mediated reperfusion injury: a selective review. Br. J. Cancer 55, Nr.8, 66 - 73 BÜCKNER, R.(1993): Operation bei linksseitiger Labmagenverlagerung. Tierärztl. Prax. 21, 507 - 510 CAO, G., H. M. ALESSIO u. R. G. CUTLER (1993): Oxygen-radical absorbance capacity assay for antioxidants. Free Radic. Biol. Med. 14, Nr. 3, 303 - 311 CAO, G., R. M. RUSSELL, N. LISCHNER u. R. L. PRIOR (1998): Antioxidant capacity is increased by consumption of strawberries, red wine or Vitamin C in elderly women. J. Nutr. 128, 2383 - 2390 CAO, G., C. P. VERDON, A. H. WU, H. WANG u. R. L. PRIOR (1995): Automated assay of oxygen radical absorbance capacity with the COBAS FARA II. Clin. Chem. 41,12 Pt 1, 1738 - 1744 CAPLE, I. W., S. J. EDWARDS, W. M. FORSYTH, P. WHITELEY, R. H. SELTH u. L. J. FULTON (1978): Blood glutathione peroxidase activity in horses in relation to muscular dystrophy and selenium nutrition. Aust. Vet. J. 54, Nr. 2, 57 - 60 - 105 - Literaturverzeichnis __________________________________________________________________________________________ CARLSON, G. P. (1990): Clinical chemistry tests. In: SMITH, B. P.: Large animal Internal Medicine. C.V. Mosby Company, St. Louis, Baltimore, Philadelphia, Toronto CELLERINO, R., G. GUIDI u. G. PERONA (1976): Plasma iron and erythrocytic glutathione peroxidase activity. A possible mechanism for oxidative haemolysis in iron deficiency anemia. Scand. J. Haematol. 17, Nr. 2, 111 - 116 CHADA, S., C. WHITNEY u. P. E. NEWBURGER (1989): Post-transcriptional regulation of glutathione peroxidase gene expression by selenium in the HL-60 human myeloid cell line. Blood 74, Nr. 7, 2535 - 2541 CHAUDRY, I. H. (1983): Cellular mechanisms in shock and ischemia and their correction. Am. J. Physiol. 245, R117 - R134 CHOW C. K. (1977): Dietary vitamin E and levels of reduced glutathione, glutathione peroxidase, catalase and superoxide dismutase in rat blood. Int. J. Vitam. Nutr. Res. 47, Nr. 3, 268 - 273 CHU, F. F., J. H. DOROSHOW u. R. S. ESWORTHY(1993): Expression, characterization, and tissue distribution of a new cellular selenium-dependent glutathione peroxidase, GSHPx-GI. J. Biol. Chem. 268, Nr. 4, 2571 - 2576 COCHRANE, C. G. (1991): Mechanisms of oxidant injury of cells. Molec. Aspects. Med. 12, 137 - 147 COGHLAN, J. G., W. D. FLITTER, C. D. ILSLEY, A. REES u. T. F. SLATER (1991): Reperfusion of infarcted tissue and free radicals. The Lancet 338, 1145 - 1146 CONSTABLE, P. D., G. Y. MILLER, G. F. HOFFSIS, B. L. HULL u. D. M. RINGS (1992): Risc factors for abomasal volvolus and left abomasal displacement in cattle. Am. J. Vet. Res. 53, 1184 - 1192 CORBUCCI, G. G., A. GASPARETTO, M. ANTONELLI, M. BUFI u. R. A. DE BLASI (1986): Changes in levels of coenzyme Q homologues, -tocopherol and Malondialdehyde in human tissue during the course of circulatory shock. Int. J. Tiss. Reac. VIII, Nr. 5, 421 - 430 CORBUCCI, G. G., A. GASPARETTO, A. M. CANDIANI, G. CRIMI, M. ANTONELLI, . BUFI, R. A. DE BLASI, M. B. COOPER u. K. GOHLI (1985): Shock-induced damage to mitochondrial function and some cellular antioxidant mechanisms in humans. - 106 - Literaturverzeichnis __________________________________________________________________________________________ Circulatory Shock 15, 15 - 26 DAVIES, S. J., S. Y. REICHARDT-PASCAL, D. VAUGHAN u. G. I. RUSSELL (1995) : Differential effect of ischemia-reperfusion injury on antioxidant enzyme activity in the rat kidney. Exp. Nephrol. 3, Nr. 6, 348 – 354 DAWSON, B., G. J. HENRY, C. GOODMAN, I. GILLAM, J. R. BEILBY, S. CHING, V. FABIAN, D. DASIG, P. MORLING u. B. A. KAKULUS (2002): Effect of Vitamin C and E supplementation on biochemical and ultrastructural indices of muscle damage after 21 km run. Int. J. Sports. Med. 23, Nr. 1, 10 - 15 DELGADO-LECAROZ, R., L. D. WARNICK, C. L.GUARD, M. C. SMITH u. D. A. BARRY (2000): Cross-sectional study of the association of abomasal displacement or volvolus with serum elctrolyte and mineral concentrations in dairy cows. Can. Vet. J. 41, Nr. 4, 301 - 305 DEL MAESTRO R. F. (1982): Role of superoxide anion radicals in microvascular permeability and leukocyte behaviour. Can J Physiol. Pharmacol. 60, Nr. 11, 1406 - 1414 DE WAZIER, I. u. R. ALBRECHT (1987): The effect of vitamin A nutritional status on glutathione, glutathione transferase and glutathione peroxidase activities in rat intestine. Experimentia. 43, 394 - 395 DE WINTER, B. Y., G. E. BOECKXSTAENS, J. G. DE MAN, T. G. MOREELS, A. G. HERMAN u. P. A. PELCKMANS (1997): Effect of adrenergic and nitergic blockade on experimental ileus in rats. Br. J. Pharmacol. 120, 464 - 468 DIRKSEN, G. (1961) : Vorkommen, Ursachen und Entwicklung der linksseitigen Labmagenverlagerung des Rindes. Dtsch. Tierärztl. Wschr. 68, 8 - 12 DIRKSEN, G. (1987): Differentialdiagnostik und Therapie von Vormagen- und Labmagenerkrankungen bei Kalb und Jungrind. Tierärztl. Umschau 36, 674 - 680 DIRKSEN, G. (1995): Displacement of the abomasum. IXth Internat. Conference on Production Diseases in Farm animals, Berlin, 11.-14. Sept., 63 67 DORMANDY, T. L. (1978): Free radical oxidation and antioxidants. The Lancet Mar. 25, 647 - 650 - 107 - Literaturverzeichnis __________________________________________________________________________________________ DORMANDY, T. L. (1983): An approach to free radicals. The Lancet Oct. 29, 1010 - 1014 EICHER, R., L. AUDIGE, U. BRAUN, J. BLUM, A. STEINER (1998): Epidemiologie und Risikofaktoren der Dislocatio abomasi bei der Milchkuh. Internationaler Workshop, Leipzig 14. Okt.: „Ätiologie, Pathogenese, Diagnostik, Prognose, Therapie und Prophylaxe der Dislocatio abomasi“, 61 - 71 ENGELHART, K. (1999): Parenteraler Einsatz lipidlöslicher Antioxidantien bei Ischämie/Reperfusion. Hohenheim, Univ., Fak. Allg. Angew. Nat., Diss. EULENBERGER, K. (1984): Physiologische und pathophysiologische Grundlagen für Maßnahmen zur Steuerung des Puerperiums beim Rind. Leipzig, Univ., Vet.-Med. Fak., Habil.-schr. FLOHE, L. (1997): Selenium in peroxide metabolism. Med. Klin. 92, Suppl. 3, 5 - 7 FLOHÉ, L., W. A. GUNZLER u. H. H. SCHOCK (1973): Glutathione peroxidase: a selenoenzyme. FEBS Lett. 32, Nr. 1, 132 - 134 FONG, K.- L., P. B. MCCAY u. J. L. POYER (1973): Evidence, that peroxidation of lysosomal membranes is initiated by hydroxyl free radicals produced during flavin enzyme activity. J. Biolog. Chem. 248, Nr. 22, 7792 - 7797 FUBINI, S. L., Y. T. GRÖHN u. D. F. SMITH (1991): Right displacement of the abomasum and abomasal volvolus in dairy cows: 458 cases (19801987). J. Am. Vet. Med. Assoc. 198, Nr. 3, 460 - 464 FÜRLL, M. (2002): Damit der Stoffwechsel auch in kritischen Phasen nicht verrückt spielt. In: Stoffwechselstörungen bei Wiederkäuern: Erkennen – Behandeln –Vorbeugen, Leipzig 2002, ISBN 3-00-009263-3, 2 - 12 FÜRLL, M., N. BIALEK, L. JÄCKEL u. E. SCHMIDT (1997): Dislocatio abomasi beim erwachsenen Rind in den neuen Bundesländern: Inzidenz, Ätiologie und Prophylaxe. Prakt. Tierarzt, coll. vet. XXVII, 81 - 86 FÜRLL, M., M. N. DABBAGH, B. FÜRLL u. T. SATTLER (2004): Verhalten der Superoxiddismutase (SOD) im Serum von Kühen mit Dislocatio abomasi (DA). Dtsch. Tierärztl. Wschr. 111, Nr. 1, 7 - 13 - 108 - Literaturverzeichnis __________________________________________________________________________________________ FÜRLL, M., B. HIEBL u. C. NAURUSCHAT (2002): Wie „leberspezifisch“ ist die GLDH beim Rind? In: Stoffwechselstörungen bei Wiederkäuern: Erkennen – Behandeln –Vorbeugen, Leipzig 2002, ISBN 3-00-009263-3, 45 - 46 FÜRLL, M., A. KASTNER, D. FRITSCHE, L. KLEISER, L. JÄCKEL, V. RICHTER u. M. KRÜGER (1998): Fettstoffwechsel-Endotoxin-Interaktionen bei Kühen und deren mögliche pathogenetische Bedeutung für die Entstehung der Dislocatio abomasi. Samstagsakademie, Leipzig, 25. April.: „Stoffwechselbelastung, -diagnostik und – stabilisierung beim Rind“, ISBN 3-00-002818-8, 105 – 116 FÜRLL, M., L. KLEISER u. T. SATTLER (2002): CK-Grenzwerte beim Rind-biologische Aspekte. In: Stoffwechselstörungen bei Wiederkäuern: Erkennen – Behandeln –Vorbeugen, Leipzig 2002, ISBN 3-00-009263-3, 47 FÜRLL, M. u. M. KRÜGER (1999): Labmagen-Verlagerung erfolgreich verhindern. Top Agrar 4, R4 - R7 FÜRLL, M., T. SATTLER, M. N. DABBAGH, C. SPIELMANN u. B. FÜRLL (1999): Ätiologie und Prophylaxe von Reperfusionsschäden. Dtsch. Tierärztlich. Wschr. 106, 389 - 393 FÜRLL, M., T. SATTLER u. T. WITTEK (2001): Bewertung der Therapie der Leberverfettung bei Kühen. In: Ref. III. Mitteleurop. Buiatr. Kongr.: Gesundheitsproblematik der Wiederkäuer, Milovy, 119 - 123 FÜRLL, M. u. M. SCHÄFER (1992): Lipolyse und Hyperbilirubinämie - Beitrag zur Pathogenese des Ikterus. Monatsh. Veterinärmed. 47, 181 - 185 FUHRMANN, H. u. H. P. SALLMANN (1999): Amount and type of unsaturated aldehydes in chicken plasma and tissues depend more on dietary lipids than on vitamin E status. Lipids 34 Suppl., 253 - 254 FUJI, J. u. N. TANIGUCHI (1999): Down regulation of superoxide dismutase and glutathione peroxidase by reactive oxygen and nitrogen species. Free Radic. Res. 31, Nr. 4, 301 - 308 GEISHAUSER, T. (1995): Untersuchungen zur Labmagenmotorik von Kühen mit Labmagenverlagerung. Gießen, Univ., Fachber. Veterinärmed., Habil.-Schr. GEISHAUSER T. (1999): - 109 - Literaturverzeichnis __________________________________________________________________________________________ Labmagenverlagerung bei Milchkühen Herdenüberwachung. Prakt. Tierarzt., coll. vet. XXIX, 59 - 62 – Vorbeuge, Früherkenneund und GEISHAUSER, T., K. LESLIE u. T. DUFFIELD (2000): Metabolic aspects in the etiology of displaced abomasum. Vet. Clin. North. Am. Food Anim. Pract. 16, Nr. 2, 255 - 265 GEISHAUSER, T., D. REICHE, C. SEEH, B. THÜNKER, S. VERWOHLT u. K. FAILING (1996): Ammonia in ruminal and abomasal contents of cows with abomasal displacement. Dtsch. Tierärztl. Wschr. 103, 465 - 468 GEISHAUSER, T., D. REICHE, C. SEEH, B. M. THÜNKER, S. VERWOHLT u. K. FAILING (1996): pH, Natrium, Kalium, Magnesium, Kalzium, Phosphat und Chlorid im Pansen- und Labmageninhalt von Kühen mit Labmagenverlagerung. Dtsch. Tierärztl. Wschr. 103, 16 - 20 GEISHAUSER, T., K. LESLIE, T. DUFFIELD, D. SANDALS u. V. EDGE (1998): The association between selected metabolic parameters and left abomasal displacement in dairy cows. Zentralbl. Veterinärmed. A 45, Nr. 8, 499 - 511 GEISHAUSER, T., M. WEISER u. K. – L. ZIBELL (1995): Indirekte und direkte Bestimmung des Selengehaltes im Blut von Kühen mit linksseitiger Labmagenverlagerung. Tierärztl. Prax. 23, 443 - 445 GENET, S., R. K. KALE u. N. Z. BAQUER, N.Z. (2000): Effects of free radicals on cytosolic creatine kinase activities and protection by antioxidant enzymes and sulfhydryl compounds. Mol. Cell. Biochem. 210, Nr.1 - 2, 23 - 28 GHISELLI, A., M. SERAFINI, G. MAIANI, E. AZZINI u. A. FERRO-LUZZI (1995): A fluorescence-based method for measuring total plasma antioxidant capability. Free Radic. Biol. Med. 18, Nr. 1, 29 - 36 GODIN, D. V. u. M. E. GARNETT (1992): Species related variations in tissue antioxidant status---I. Differences in antioxidant enzyme profiles. Comp. Biochem. Physiol. 103 B, Nr. 3, 737 - 742 GOERRES, A. u. M. FÜRLL (2002) : Antioxidativer- (TEAC) und Endotoxin-Status bei kranken Kühen In: 11. Jahrestagung d. Dtsch. Veterinärmed. Ges., Fachgr. Innere Med. und Klin. Lab.-Diag., München, 2002, 36 - 38 GRANGER, D., N. G. RUTILI u. J. M. McCORD (1981): Superoxide radicals in feline intestinal ischemia. Gastroenterology 81, Nr. 1, 22 – 29 - 110 - Literaturverzeichnis __________________________________________________________________________________________ GRANGER, D. N., J. M. McCORD, D. A. PARKS u. M. E. HOLLWARTH (1986): Xanthine oxidase inhibitors attenuate ischemia-induced vascular permeability changes in the cat intestine. Gastroenterology 90, Nr. 1, 80 - 84 GROGAARD, B., D.A. PARKS, N. GRANGER, J. M. MC CORD u. J. O. FORSBERG (1982): Effects of ischemia and oxygen radicals on mucosal albumin clearance in intestine. Am. J. Physiol. 242, G448 - G454 GROTH, W. u. W. GRÄNZER (1975): The influence of transport stress on serum GOT, GPT, LDH and CPK activity in calves. Zentralbl. Veterinaermed. A 22, Nr. 1, 57 - 75 GRUNERT, E. (1986): Ursachen und Behandlungsmöglichkeiten der Endometritis beim Rind. Prakt. Tierarzt, coll. vet. XVII, 43 - 47 GÜNEL, E., F. CAGLAYAN, O. CAGLAYAN, A. DILSIZ, S. DUMAN u. M. AKTAN (1998): Treatment of intestinal reperfusion injury using antioxidative agents. J. Pediatr. Surg. 33, 1536 - 1539 GUTTERIDGE, M.C., L. MAIDT u. L. POYER (1990): Superoxide dismutase and Fenton chemistry. Biochem. J. 269, 169 - 174 HALLIWELL, B. (1989): Superoxide, iron, vascular endothelium and reperfusion injury. Free. Rad. Res. Comms. 5, 315 - 318 HALLIWELL, B. u. J. M. C. GUTTERIDGE (1984): Lipid peroxidation, oxygen radicals, cell damage, and antioxidant therapy. The Lancet June 23, 1396 - 1397 HALLIWELL, B. u. J. M. C. GUTTERIDGE (1990): The antioxidants of human extracellular fluids. Arch. Biochem. Biophys. 280, Nr. 1, 1 - 8 HALLIWELL, B., J. M. C. GUTTERIDGE u. C. E. CROSS (1992): Free radicals, antioxidants, and human diseases: Where are we now? J. Lab. Clin. Med. 119, Nr. 6, 598 - 620 HARRIS, E.D. (1992): Copper as a cofactor and regulator of copper, zinc superoxide dismutase. J. Nutr. 122, 636 - 640 HARRIS, E.D. (1992): Regulation of antioxidant enzymes. J. Nutr. 122, 625 - 626 - 111 - Literaturverzeichnis __________________________________________________________________________________________ HERDT, T.H., B.J. GERLOFF (1982): Hepatic lipidosis and liver function in 49 cows with displaced abomasum. Proc. XIIth World Congress on disease of cattle, Amsterdam, Netherlands, 12./13, Aug., 522 526 HERMANN, H. (2000): Schnell und automatisierbar Antioxidantien analysieren. Lab. Prax. Februar, 24 - 27 HILL, BD., A. C. MCMANUS, N. N. BROWN, C. L. PLAYFORD u. J. W. NOBLE (1992): A bovine stress syndrome associated with exercise-induced hyperthermia. Aust. Vet. J. 78, Nr. 1, 38 - 43 HOPKINS, J. u. G. R. THUDHOPE (1973): Glutathione peroxidase in human red cells in health and disease. Br. J. Haematol. 25, Nr. 5, 563 - 575 HUBER, W., K. B. MENANDER-HUBER, M. G. SAIFER u. L. D. WILLIAMS (1980): Bioavailibility of superoxide dismutase: implications for the anti-inflammatory action mechanism of orgotein. Agents Actions Suppl. 7, 185 - 195 HUSSAIN, S. u. S. F. ALI (1999): Manganese scavenges superoxide and hydroxyl radicals: An in vitro study in rats. Neuroscience Letters 261, 21 - 24 HUTLER, M., S. WOWERIES, R. LEITHAUSER, D. BONING u. R. BENEKE (2001): Exercise-induced changes in blood level of alpha-tocopherol. Eur. Appl. Physiol. 85, Nr. 1 - 2, 151 - 156 ISHIKAWA, Y., J. E. SAFFITZ, T. L. MEALMAN, A. M. GRACE u. R. ROBERTS (1997): Reversible myocardial ischemic injury is not associated with increased creatine kinase activity in plasma. Clin. Chem. 43, Nr. 3, 467 - 475 JAIN, S. K., K. KANNAN u. G. LIM (1998): Ketosis (Acetoacetate) can generate oxygen radicals and cause increased lipid peroxidation and growth inhibition in human endothelial cells. Free Rad. Biol. Med. 25, Nr. 9, 1083 - 1088 JANOWITZ, H. (1998): Laparascopic reposition and fixation of the left displaced abomasum in cattle. Tierärztl. Praxis, Ausg. G, Großtiere, Nutztiere, 26, Nr. 6, 308 – 313 JENTZSCH, A. M. (1997): Untersuchungen zu Rolle und Bedeutung Ischämie/Reperfusionssyndrom. Hohenheim, Univ., Allg. Angew. Nat., Diss. reaktiver Sauerstoffmetaboliten JOHNSON, K. J., J. C. FANTONE, J. KAPLAN u. P. A. WARD (1981): - 112 - im Literaturverzeichnis __________________________________________________________________________________________ In vivo damage of rat lungs by oxygen metabolites. J. Clin. Invest. 67, 983 - 993 JOHNSTON, R. B. u. J. E. LEHMEYER (1977): Elaboration of toxic oxygen by-products by neutrophils in a model of immune complex disease J. Clin. Invest. 57, Nr. 4, 836 - 841 JORGENSEN, P. F., J. HYLDGAARD-JENSEN u. J. MOUSTGAARD (1977): Glutathione peroxidase activity in porcine blood. Acta vet. Scand. 18, Nr. 3, 323 – 334 JÜNGER, C. u. M. FÜRLL (1998): Akute und chronische Störungen des Kaliumstoffwechsels beim Rind. In: „Stoffwechselbelastung, -diagnostik und –stabilisierung beim Rind, Leipziger Samstagsakademie, Med. Tierklinik, Vet.-Med. Fak., 25. April, ISBN 3-00-002818-8, 10 - 26 JUVONEN, P. O., H. E. PAAJANEN, A. A. HEINO, E. J. KOSKI, M. MERASTO, J. E. HARTIKAINEN, V. M. KOSMA, E. M. ALHAVA u. TAKALA (1997): Lipid peroxidation products and antioxidant capacity in portal venous and systemic arterial plasma during gradual intestinal ischemia and reperfusion in pigs. Eur. Surg. Res. 30, Nr. 2, 95 - 101 KAIKKONEN, J., E. PORKKALA-SARATOHO, T. P. TUOMAINEN, K. NYYSSONEN, L. KOSONEN, U. RISTONMAA, H. M. LAKKA, R. SALONEN, H. KORPELA u. J. T. SALONEN (2002): Exhaustive exercise increases plasma/serum total oxidation resistance in moderately trained men and women, whereas their VLDL + LDL lipoprotein fraction is more susceptible to oxidation. Scand. J. Clin. Lab. Invest. 62, Nr. 8, 599 - 607 KARATZIAS, H. (1998): Epidemiology and risk factors of abomasal displacement in dairy cows. Internationaler Workshop Ätiologie, Pathogenese, Diagnostik, Prognose, Therapie und Prophylaxe der Dislocatio abomasi, Leipzig 1998, Proceed., 73 - 81 KEELER, R., J. SWINNEY, R. M. TAYLOR u. P. R. ULDALL (1966): The problem of renal preservation. Br. J. Urol. 38, 653 - 656 KLEISER, L. u. M. FÜRLL (1998): Screening zur Früherkennung einer Disposition für die Dislocatio abomasi bei Kühen. Internationaler Workshop Ätiologie, Pathogenese, Diagnostik, Prognose, Therapie und Prophylaxe der Dislocatio abomasi, Leipzig 1998, Proceed., 275 – 286 KÖNEN, H. J. (1982): Klinisch-chemische Untersuchungen über die Superoxiddismutase mit einer Cytochrom-CZentrifugalanalysator- und Chemilumineszenzmethode. Aachen, Techn. Hochsch., Med. Fak., Diss. KOLB, E. (1989): - 113 - Literaturverzeichnis __________________________________________________________________________________________ Lehrbuch der Physiologie der Haustiere: Teil I. 5. Aufl. Gustav Fischer Verlag, Jena, S. 370 - 424 KOUFEN, P. u. G. STARK (2000): Free radical induced inactivation of creatine kinase: sites of interaction, protection, and recovery. Biochim. Biophys. Acta 1501, Nr. 1, 44 - 50 KRAFT, W. u. U. DÜRR (1999): Klinische Labordiagnostik in der Tiermedizin. Verlag Schattauer, New York, 5. Auflage KÜHLER, S. (1996): Einfluss der Lagerungstemperatur und Lagerungszeit auf die katalytischen Aktivitäten der Superoxiddismutase, Katalase, Glutathionperoxidase, Glutathionreduktase und Glutathion-STransferase in Leber- und Nierenhomogenaten von Ratten und deren Referenzwertermittlung. Berlin, Univ., Med. Fak., Diss. KÜHN, K. (1997): Bestimmungsmethodische und In-vitro-Untersuchungen der Aktivität erythrozytärer Glutathionperoxidase unterschiedlicher Tierarten unter Einwirkung verschiedener Umweltschadstoffe. Berlin, Univ., Fachber. Veterinärmed., Diss. KÜMPER, H. (1995): Die rechtsseitige Labmagenverlagerung des Rindes: 1.Teil: Entstehungsweise, klinischer Verlauf und Prognose. Tierärztl. Prax., 23, 351 - 359 KÜMPER, H. (1995): Die rechtsseitige Labmagenverlagerung des Rindes: 2.Teil: Neuere Erkenntnisse zur operativen Behandlung. Tierärztl. Prax. 23, 437 - 442 LAIPPLE, A. (2000): Sauerstoffradikal-induzierte Lipidperoxidation und Leukozytenaktivierung während KarotisThrombendarteriotomie. Heidelberg, Univ., Med. Fak., Diss. LAMBOTTE, L. (1977): Effect of anoxia and ATP depletion on the membrane potential and permeability of dog liver. J. Physiol. 269, 53 – 76 LAMONT, J., J. CAMPBELL u. P. FITZGERALD (1997): Measurement of individual versus total antioxidants. Clin. Chem. 43, Nr. 5, 852 - 3 + 4 - 5 LANDS, L. C., V. L. GREY u. C. GRENIER (2000): Total plasma antioxidant capacity in cystic fibrosis. Pediatr. Pulmonol. 29, Nr. 2, 81 – 87 - 114 - Literaturverzeichnis __________________________________________________________________________________________ LANTZ, G. C., S. F. BADYLAK, M. C. HILES u. T. E. ARKIN (1992): Treatment of reperfusion injury in dogs with experimentally induced gastric dilatationvolvolus. Am. J. Vet. Res. 53, Nr. 9, 1594 - 1598 LARREA, E., O. BELOQUI, M. – A. MUNOZ-NAVAS, M. – P. CIVEIRA u. J. PRIETO (1998): Superoxide dismutase in patients with chronic Hepatitis C virus infection. Free Radic. Biol. Med. 24, Nr. 7/8, 1235 - 1241 LASZCZYCA, P., E. KAWKA-SERWECINSKA, I. WITAS, B. DOLEZYCH, B. FALKUS, A. MEKAIL, B. ZIOLKOWSKA, P. MADEJ u. P. MIGULA (1996): Lipid peroxidation and activity of antioxidative enzymes in the rat model of ozone therapy. Mater Med. Pol. 28, Nr. 4, 155 - 160 LEAF, A. (1973): Cell swelling: A factor in tissue injury. Circulation 48, 455 - 458 LEHMANN, C., B. LUTHER u. W. J. KOX (1996): Pharmakologische Beinflussbarkeit des intestinalen Reperfusionsschadens im Tierexperiment. Zentral. Chir. 121, 70 - 76 LEHR, H. A., C. HUBNER, B. FINCKH, S. ANGERMULLER, D. NOLTE, U. BEISIEGEL, A. KOHLSCHUTTER u. K. MESSMER (1991): Role of leukotrienes in leukocyte adhesion following systemic administration of oxidatively modified human low density lipoprotein in hamsters. J. Clin. Invest. 88, Nr. 1, 9 - 14 LUTHER, B., C. LEHMANN, E. – D. SCHWILDEN u. A. HOLZÄPFEL (1996): Perioperative intraarterielle Spülperfusion zur adjuvanten Therapie akuter intestinaler Durchblutungsstörungen. Zentralbl. Chir. 121, 61 - 66 MAHMOUD, O. M. u. E. J. FORD (1988): The activity of diagnostic enzymes and the concentration of lipids in the blood vessels of cattle. Vet. Res. Commun. 12, Nr. 2 - 3, 233 - 5 MAIORINO, M., COASSIN, A. ROVERI u. F. URSINI (1989): Microsomal lipid peroxidation: effect of vitamin E and its functional interaction phospholipid hydroperoxide glutathione peroxidase. Lipids 24, Nr. 8, 721 - 726 MAO, G. D., P. D. THOMAS, G. D. LOPASCHUK, u. M. J. POZNANSKY (1993): Superoxide dismutase (SOD)-Catalase conjugates. Biol. Chem. 268, Nr. 1, 416 - 420 MARKUSFELD, O. (1989): - 115 - Literaturverzeichnis __________________________________________________________________________________________ Possible association of vitamin A deficiency with displacement of the abomasum in dairy heifers. J. Am. Vet. Med. Assoc. 195, Nr. 8, 1123 - 1124 MARTENS, H. (1998): Beziehungen zwischen Fütterung, Physiologie der Vormägen und Pathogenese der Dislocatio abomasi. Ätiologie, Pathogenese, Diagnostik, Prognose, Therapie und Prophylaxe der Dislocatio abomasi: Proceedings; Internationaler Workshop Leipzig 1998, Proceed., S. 81 - 101 MARTIG, J. (1996): Hochtönendes Perkussionsgeräusch (PING) als Leitsymptom. Schweiz. Arch. Tierheilk. 138, 351 - 352 MAULIK, N., M. WATANABE, D. T. ENGELMAN, R. M. ENGELMAN u. D. K. DAS (1995): Oxidative stress adaptation improves postischemic ventricular recovery. Mol. Cell. Biochem. 144, Nr. 1, 67 - 74 MAYLIN, G. A., D. S. RUBIN u. D. H. LEIN (1980): Selenium and vitamin E in horses. Cornell. Vet. 70, Nr. 3, 272 - 289 MENGER, M. D., S. PELIKAN, D. STEINER u. K. MESSMER (1992): Microvascular ischemia-reperfusion injury in striated muscle: significance of „reflow paradox“. Am. J. Physiol. 263, Nr. 6 Pt 2, H1901 - H1906 MERTENS, M. (1992): Leberbiopsie beim Rind – Histologische und enzymatische Auswertung von Bioptaten aus Kühen mit Dislocatio abomasi sinistra. Hannover, Univ., Tierärztl. Hochschule, Diss. MEYLAN, M. (1999): Prognostic indicators in cattle with right-sided displacement of the abomasum and abomasal volvolus. Schweiz. Arch. Tierheilk. 141, Nr. 9, 413 - 418 MILLER, N. J., J. SAMPSON, L. P. CANDEIAS, P. M. BRAMLEY u. C. A. RICE-EVANS (1996): Antioxidant activities of carotenes and Xanthophylls. FEBS Letters 384, 240 - 242 MORROW, D. A. (1975): Fat cow syndrome. J. Dairy Sci. 59, Nr. 9, 1625 - 1628 MORTENSEN, A., L. H. SKIBSTED, J. SAMPSON, C. A. RICE-EVANS u. S. A. EVERETT (1997): - 116 - Literaturverzeichnis __________________________________________________________________________________________ Comparative mechanisms and rates of free radical scavenging by carotenoid antioxidants. FEBS Letters 418, 91 - 97 MUDRON, P., J. REHAGE, K. QUALMANN, H. P. SALLMANN u. H. SCHOLZ (1999): A study of lipid peroxidation and vitamin E in dairy cows with hepatic insufficiency. Zentralbl. Veterinärmed. A 46, Nr. 4, 219 - 224 MUDRON, P., J. REHAGE, H. P. SALLMANN, M. MERTENS, H. SCHOLZ u. G. KOVACN (1997): Plasma and liver alpha-tocopherol in dairy cows with left abomasal displacement and fatty liver Zentralbl. Veterinärmed. A 44, Nr. 2, 91 - 97 MUDRON, P., H. P. SALLMANN, J. REHAGE, M. HÖLTERSHINKEN, G. KOVAC, P. BARTKO u. H. SCHOLZ (1994): Auswirkungen einer operativen Reposition der linksseitigen Labmagenverlagerung auf Parameter des Energiestoffwechsels bei Milchkühen. Dtsch. tierärztl. Wschr. 101, 376 - 378 MÜLLER, U. (1992): Untersuchungen über das Vorkommen und die Diagnostik der Labmagenverlagerung beim Rind. Leipzig, Univ., Vet.-Med. Fak., Diss. MURPHY, S., Q. HUI-QI, T. SAKAI, D. E. DEPACE u. J. D. FONDACARO (1997): Ischemia/reperfusion injury in the rat colon. Inflammation 21, Nr. 2, 173 - 190 MURTHY, K. S., K. M. ZHANG, J. G. GRIDER u. G. M. MAKHLOUF (1993): VIP-mediated G protein-coupled Ca2+ influx activates a constitutive NOS in dispersed gastric muscle cells. Am. J. Physiol. 265, G660 – G 671 NAKAMURA, M., M. OZAKI, S. FUCHINOUE, S. TERAOKA u. K. OTA (1997): Ascorbic acid prevents ischemia-reperfusion injury in the rat small intestine. Transpl. Int. 10, Nr. 2, 89 - 95 NAKAZAWA, H., C. GENKA u. M. FUJISHIMA (1996): Pathological aspects of active oxygens / free radicals. Jap. J. Physiol. 46, 15 - 32 NAURUSCHAT, C. u. M. FÜRLL (2002): Zur diagnostischen Bedeutung von CK-Isoenzymen bei Kühen. In: 11. Jahrestagung d. Dtsch. Veterinärmed. Ges., Fachgr. Innere Med. und Klin. Lab.-Diag., München, 2002, S. 38 - 39 NINFALI, P. u. G. ALUIGI (1998): Variability of oxygen radical absorbance capacity (ORAC) in different animal species. Free Rad. Res. 29, 399 - 408 - 117 - Literaturverzeichnis __________________________________________________________________________________________ NIWA, Y., O. IIZAWA, K. ISHIMOTO, H. AKAMUTSU u. T. KANOH (1993): Age-dependent basal level and induction capacity of copper-zinc and manganese superoxide dismutase and other scavenging enzyme activities in leukocytes from young and elderly adults. Am. J. Pathol. 143, Nr. 1, 312 - 320 NOCKELS, C. F., K. G. ODDE u. A. M. CRAIG (1996): Vitamin E supplementation and stress affect tissue alpha-tocopherol content of beef heifers. J. Anim. Sci. 74, Nr. 3, 672 - 7 ÖHLENSCHLÄGER, G. (1993): Aktivierte Sauerstoffstufen, Freie Radikale, Oxidativer Stress – Krankheitsverursachende Prinzipien in lebenden Systemen. J. Orthomol. Med. 1, 36 - 55 ÖHLENSCHLÄGER, G. (1993): Scavenger-Radikalfänger-Antioxidantien, Prävention und Reparatur in lebenden Systemen. J. Orthomol. Med. 2, 49 - 63 OH, S. H., H. E. GANTHER u. W. G. HOEKSTRA (1974): Selenium as a component of glutathione peroxidase isolated from ovine erythrocytes. Biochemistry 13, Nr. 9, 1825 - 1829 OIKAWA, S., N. KATOH, F. KAWAWA u. Y. ONO (1997): Decreases serum apolipoprotein B-100 and A-I concentrations in cows with ketosis and left displacement of the abomasum. Am. J. Vet. Res. 58, 121 - 125 OK, M., I. SEN, K. TURGUT u. K. IRMAK (2001): Plasma gastrin activity and the diagnosis of bleeding abomasal ulcers in cattle. J. Vet. Med. A. Physiol. Pathol. Clin. Med. 48, Nr. 9, 563 - 568 OURIEL, K., N. SMEDIRA u. J. J. RICOTTA (1985): Protection of the kidney after temporary ischemia: Free radical scavengers. J. Vasc. Surg. 2, 49 - 53 PAGLIA, D. E. u. W. N. VALENTINE (1938): Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 70,158 - 169 PAPPARELLA, A., F. G. DELUCA, C. E. OYER, H. PINAR u. B. S. STONESTREET (1997): Ischemia –Reperfusion injury in the intestines of newborn pigs. Pediatr. Res. 42, Nr. 2, 180 - 188 PARKS, D. A. (1989): Oxygen radicals: Mediators of gastrointestinal pathophysiology. Gut 30, 293 - 298 - 118 - Literaturverzeichnis __________________________________________________________________________________________ PARKS, D. A., G. B. BULKLEY u. D. N. GRANGER (1983): Role of oxygen free radicals in shock, ischemia, and organ preservation. Surgery 94, 428 - 432 PAYNE, R. M., D. L. FRIEDMAN, J. W. GRANT, M. B. PERRYMAN u. A. W. STRAUSS (1993): Creatine kinase isoenzymes-highly regulated during pregnancy in rat uterus and placenta. Am. J. Physiol. 265, Nr. 4 Pt 1, E624 - E635 PEHRSON, B. u. L. STENGÄRDE (1998): Labmagenverlagerung: Klinische Charakteristika sowie Ergebnisse unterschiedlicher Behandlungsmaßnahmen. Internationaler Workshop Ätiologie, Pathogenese, Diagnostik, Prognose, Therapie und Prophylaxe der Dislocatio abomasi, Leipzig 1998, Proceed., 49 - 60 PERONA, G., G. C. GUIDI, A. PIGA, R. CELLERINO, R. MENNA u. M. ZATTI (1978): In vivo and in vitro variations of human erythrocyte glutathione peroxidase activity as result of cells ageing, selenium availibility and peroxide activation. Br. J. Haematol. 39, Nr. 3, 399 - 408 PIERCY, R. J., K. W. HINCHCLIFF, R. A. DISILVESTRO, G. A. REINHART, C. R. BASKIN, M. G. HAYEK, J. R. BURR u. R. A. SWENSON (2000): Effect of dietary supplements containing antioxidants on attenuation of muscle damage in exercising sled dogs. Am. J. Vet. Res. 61, Nr. 11, 1438 - 1445 POIKE, A. u. M. FÜRLL (1998): Zur Epidemiologie der Labmagenverlagerung (dislocatio abomasi) in Mitteldeutschland. Internationaler Workshop Ätiologie, Pathogenese, Diagnostik, Prognose, Therapie und Prophylaxe der Dislocatio abomasi, Leipzig 1998, Proceed., 29 - 40 POPOV, I. u. G. LEWIN (2000): Photosensibilisierte Chemolumineszenz bei der Quantifizierung von Antioxidantien. BIOforum 1-2, 46 - 48 REDDY, S., A. D. JONES, C. E. CROSS, P. S. WONG, u. A. VAN DER VLIET (2000): Inactivation of creatine kinase by S-glutathionylation of the active-site cysteine residue. Biochem. J. 347 Pt 3, 821 - 827 REES, R., D. SMITH, T. D. LI, B. CASHMER, W. GARNER, J. PUNCH u. D. J. JR. SMITH (1994): The role of xanthine oxidase and xanthine dehydrogenase in skin ischemia. J. Surg. Res. 56, Nr. 2, 162 - 167 REHAGE, J., M. MERTENS, N. STOCKHOFE-ZURWIEDEN, M. KASKE u. H. SCHOLZ (1996): - 119 - Literaturverzeichnis __________________________________________________________________________________________ Post surgical convalescence of dairy cows with left abomasal displacement in relation to fatty liver. Schweiz. Arch. Tierheilk. 138, 361 - 368 REICHE, D., T. GEISHAUSER u. M. SCHEMANN (1998): Labmagenverlagerung: Physiologie und Pathophysiologie der Labmagenmotorik. Internationaler Workshop Ätiologie, Pathogenese, Diagnostik, Prognose, Therapie und Prophylaxe der Dislocatio abomasi, Leipzig 1998, Proceed., 17 - 29 REID, I. M. (1980): Incidence and severity of fatty liver in dairy cows. Vet. Rec. 107, 281 - 284 ROMAY, C., C. PASCUAL u. E. A. LISSI (1996): The reaction between ABTS radical cation and antioxidants and its use to evaluate the antioxidant status of serum samples. Braz. J. Med. Biol. Res. 29, 175 - 183 ROSSOW N. u. R. STAUFFENBIEL (1983): Störungen des Lipidstoffwechsels bei der Milchkuh unter besonderer Berücksichtigung der Lipolyse. Monatsh. Veterinärmed. 38, 404 - 409 ROUSSEL, J. D., T. J. ARANAS u. S. H. SEYBT (1981) : Metabolic profile testing in holstein cattle in Louisiana: Reference values. Am. J. Vet. Res. 43, Nr. 9, 1658 –1660 ROVERI, A., A. CASASCO, M. MAIORINO, P. DALAN, A. CALLIGARO u. F. URSINI (1992): Phospholipid hydroperoxide glutathione peroxidase of rat testis. Gonadotropin dependence and immunocytochemical identification. J. Biol. Chem. 267, Nr. 9, 6142 - 6146 ROVERI, A., F. URSINI, L. FLOHE u. M. MAIORINO (2001): PHGPx and spermatogenesis. Biofactors 14, Nr. 1 - 4, 213 - 222 SALZMAN, A. L. (1995): Nitric oxide in the gut. New. Horiz. 3, 352 - 364 SATTLER, T. (2001): Untersuchungen ausgewählter Parameter des antioxidativen und Stoffwechselstatus bei Kühen mit Labmagenverlagerung. Leipzig, Univ., Veterinärmed. Fak., Diss. SATTLER, T. u. M. FÜRLL (2002): Einfluss von Antioxidantien auf den postoperativen Verlauf bei Kühen mit Dislocatio abomasi. - 120 - Literaturverzeichnis __________________________________________________________________________________________ In: 11. Jahrestagung d. Dtsch. Veterinärmed. Ges., Fachgr. Innere Med. und Klin. Lab.-Diag., München, 2002, S. 32 - 33 SATTLER, N., G. FECTEAU, P. HELIE, J. M. LAPOINTE, L. CHOUINARD, M. BABKINE, A. DESROCHERS, Y. COUTURE u. P. DUBREUIL (2000): Etiology, forms and prognosis of gastrointestinal dysfunction resembling vagal indigestion occuring after surgical correction of right abomasal displacement. Can. Vet. J. 41, Nr. 10, 777 - 785 SCHELLER, W. (1999): In vivo-Messung von Superoxid-anionen im tierexperimentellen IschämieReperfusionsmodell. Berlin, Univ., Med. Fak., Diss. SCHILLER, H. J., P. M. REILLY u. G. B. BULKLEY, G. B.(1993): Tissue perfusion in critical illness. Antioxidant therapy. Crit. Care. Med. 21(2 Suppl), S92 - 102 SCHOENBERG, M. H. u. H. G. BEGER (1993): Reperfusion injury after intestinal ischemia. Crit. Care Med. 21, Nr. 9, 1376 - 86 SCHOLZ, H. (1991): Selen-Vitamine: Bedeutung und Versorgung beim Kalb. Tierärztl. Umschau 46, 194 - 202 SHEPPARD, A. D. u. K. R. MILLAR (1981): Stability of glutathione peroxidase in ovine blood samples under various storage conditions and the response of this enzyme to different methods of selenium supplementation. New Zealand vet. J. 29, Nr. 5, 77 - 80 SLAVIKOVA, H., A. LOJEK, J. HAMAR, M. DUSKOVA, L. KUBALA, J. VONDRACEK u. M. CIZ (1998): Total antioxidant capacity of serum increased in early but not late period after intestinal ischemia in rats. Free Radic. Biol. Med. 25, Nr. 1, 9 – 18 SOBIRAJ, A. (1992): Untersuchungen zur Morphologie sowie zur Histochemie und Biochemie des Uterus bei Rindern in der frühen postpartalen Periode. Gießen, Univ. Vet.-Med. Fak., Habil.-Schr. STAUFFENBIEL, R. (1993): Energie- und Fettstoffwechsel des Rindes unter besonderer Berücksichtigung der Messung der Rückenfettdicke und der Untersuchung von Fettgewebe. Berlin, Univ., FU, Habil.-Schr. STEINER, A. (1996): Die chirugische Behandlung der linksseitigen Labmagenverlagerung bei der Kuh: Eine Literaturübersicht. Schweiz. Arch. Tierheilk. 138, 353 - 360 - 121 - Literaturverzeichnis __________________________________________________________________________________________ STÖBER M. u. G. DIRKSEN (1982): Das Lipomobilisationssyndrom (Verfettungssyndrom) der Milchkuh. Prakt. Tierarzt, coll. vet. XII 63, 79 - 88 STOHRER, M., A. EICHINGER, M. SCHLACHTER u. M. STANGASSINGER (1998): Protective effect of Vitamin E in rat model of focal cerebral ischemia. Z. Naturforsch. (C) 53, Nr. 3 - 4, 273 – 278 SUZUKI, T., N. S. AGAR u. M. SUZUKI (1985): Red blood cell metabolism in experimental animals: pentose phoshate pathway, antioxidant enzymes and glutathione. Jikken Dobutsu 34, Nr. 4, 353 – 366 SZABO, A., M. BOROS, J. KASZAKI u. S. NAGY (1997): The role of mast cells in mucosal permeability changes during ischemia-reperfusion injury of the small intestine. Shock 8, Nr. 4, 284 - 291 TAKAGI, Y., T. YASUHARA u. K. GOMI (2001): Creatine kinase and its isoenzymes. Rinsho. Byori. Suppl. 116, 52 – 61 TAKAHASHI, K., N. AVISSAR, J. WHITIN u. H. COHEN (1987): Purification and characterization of human plasma glutathione peroxidase: a selenoglycoprotein distinct from the known cellular enzyme. Arch. Biochem. Biophys. 256, Nr. 2, 677 – 686 THOMPSON, R.H. (1976): The levels of selenium and glutathione peroxidase activity in blood of sheep, cows and pigs Res. Vet. Sci. 20, 229 - 231 TILL, U.(1999): Pathophysiologie / Pathobiochemie systematisch 1.Aufl. UNI-MED-Verl., Bremen, 146 - 160 TITHOF, P. K. u. W. C. REBHUN (1986): Complications of blind.stitch abomasopxy: 20 cases (1980-1995). J. Am. Vet. Med. Assoc. 189, Nr. 11, 1489 - 1492 TOH, Y., S. KUNINAKA, T. OSHIRO, Y. IKEDA, H. NAKASHIMA, H. BABA, S. KOHNOE, T. OKAMURA, M. MORI u. K. SUGIMACHI (2000): Overexpression of manganese superoxide dismutase mRNA may correlate witrh aggressiveness in gastric and colorectal adenocarcinomas. Int. J. Oncol. 17, Nr. 1, 107 - 112 TSAN, M. F. (1993): Superoxide dismutase and pulmonary oxygen toxicity. Proc. Soc. Exp. Biol. Med. 203, Nr. 3, 286 - 290 - 122 - Literaturverzeichnis __________________________________________________________________________________________ TUBARO, F., A. GHISELLI, P. RAPUZZI, M. MAIORINO u. F. URSINI (1998): Analysis of plasma antioxidant capacity by competition kinetics. Free Rad. Biol. Med. 4, 1228 - 1234 TUCKER, E. M. (1974): A shortened life span of sheep red cells with a glutathione deficiency. Res. Vet. Sci. 16, Nr. 1, 19 - 22 TURRENS, J. F. (1991) : The potential of antioxidant enzymes as pharmacological agents in vivo. Xenobiotica 21, Nr. 8, 1033 - 1040 URSINI, F., M. MAIORINO u. C. GREGOLIN (1985): The selenoenzyme phospholipid hydroperoxide glutathione peroxidase. Biochim. Biophys. Acta 839, Nr. 1, 62 - 70 VAIDOVICH P. T. GAÁL u. A. SZILÁGYI (1993): Changes of lipid peroxidation parameters in dogs with alloxan diabetes. Acta Physiol. Hung. 81, Nr. 4, 317 - 326 VAN WINDEN, S. C., K. E. MULLER, R. KUIPER u. J. P. NOORDHUIZEN (2002): Studies on the pH value of abomasal contents in dairy cows during the first 3 weeks after calving. J. Vet. Med. A. Physiol. Pathol. Clin. Med. 49, Nr. 3, 157 - 160 WARD, J. L., D. F. SMITH, S. L. FUBINI u. D. M. DEUEL-AROMANDO (1994): Evaluation of abomasal outflow as an experimental model of hypochloremic, hypokalemic metabolic alkalosis in lactating cows. Can. J. Vet. Res. 58, 13 - 19 WARREN, J. S., S. KUNKEL, K. J. JOHNSON u. P. A.WARD (1987): In vitro activation of rat neutrophils and alveolar macrophages with Ig A and Ig G immune complexes: Implications for immune complex-induced lung injury. Am. J. Pathol. 129, 578 - 588 WEBER, G. F. u. H. P. BRUCH (1992): The pharmacology of superoxide dismutase. Pharmazie 47, Nr. 3, 159 - 167 WENDEL, A. u. P. CIKRYT (1980): The level and half-life of glutathione in human plasma. FEBS Lett. 120, Nr. 2, 209 - 211 WILKEN, H. u. M. FÜRLL (2002): Antioxidativer (TEAC) und Endotoxin-Status bei gesunden Kühen unterschiedlicher Leistungsebenen. In: 11. Jahrestagung d. Dtsch. Veterinärmed. Ges., Fachgr. Innere Med. und Klin. Lab.-Diag., München, 2002, S. 34 - 35 WILKINS, P. A., N. G. DUCHARME, J. E. LOWE, W. S. SCHWARK, C. MESCHTER u. H. N. ERB (1994): - 123 - Literaturverzeichnis __________________________________________________________________________________________ Measurements of blood flow and xanthine oxidase activity during postischemic reperfusion of the large colon of ponies. Am. J. Vet. Res. 55, Nr. 8, 1168 - 77 WINNEFELD, K. (1996): Antioxidantien und Radikale: Analytik und klinische Bedeutung. J. Lab. Med. 20, Nr. 4, 199 - 204 WITT, E. H., A. Z. REZNICK, C. A. VIGUIE, P. STARKE-REED u. L. PACKER (1992): Exercise, oxidative damage and effects of antioxidant manipulation. J. Nutr. 122, 766 - 773 WITTEK, T., P. D. CONSTABLE u. M. FÜRLL (2004): Comparison of abomasal luminal gas pressure und volume and perfusion of the abomasum in dairy cows with left displaced abomasum or abomasal volvolus. J. Am. Vet. Res., zur Veröffentlichung angenommen am 10. Okt. 2003 WITTEK, T. u. M. FÜRLL (2002): Pulsoximetrische Messung der Sauerstoffsättigung des Labmagens bei der Dislocatio Abomasi des Rindes. In: 11. Jahrestagung d. Dtsch. Veterinärmed. Ges., Fachgr. Innere Med. und Klin. Lab.-Diag., München, 2002, S. 48 - 49 WOLF, V., H. HAMANN, H. SCHOLZ u. O. DISTL (2001): Influences on the occurence of abomasal displacement in German Holstein cows. Dtsch. tierärtzl. Wochenschr. 108, Nr. 10, 403 – 408 WOLF, C. (1998): Selenversorgung bei Kühen –Mangel oder Überschuss? 2. Mitteilung: Die Entwicklung der Selen-Glutathionperoxidase-Aktivität von Milch- und Mutterkühen in MecklenburgVorpommern von 199-1996. Prakt. Tierarzt 79, Nr. 8, 755 - 759 WOODFORD, F. P. u. T. P. WHITEHEAD (1998): Is measuring serum antioxidant capacity clinical useful? Ann. Clin. Biochem. 35, 48 - 56 WYSS, M. u. R. KADDURAH-DAOUK (2000): Creatine and Creatinine metabolism. Physiol. Rev. 80, Nr. 3, 1107 - 1181 YAN, T., X. JIANG, H. J. ZHANG, S. LI u. L. W. OBERLEY (1998): Use of commercial antibodies for detection of the primary antioxidant enzymes. Free Rad. Biol. Med. 25, Nr. 6, 688 - 693 YOSHIDA, W. B., T. ALASIO, R. MAZZIOTTA, F. QIN, M. KASHANI, S. LEE u. H. DARDIK (1998): Effect of -tocopherol, taurine and selenium on the attenuation of ischemia/reperfusion injury of splanchnic organs. - 124 - Literaturverzeichnis __________________________________________________________________________________________ Cardiovasc. Surg. 6, Nr. 2, 178 - 87 YOUNG, J. F., S. E. NIELSEN, J. HARALDSDOTTIR, B. DANESHVAR, S. T. LAURIDSEN, P. KNUTHSEN, A. CROZIER, B. SANDSTROM u. L. O. DRAGSTED (2000): Polyphenic antioxidants in fruit juice. Urinary excretion and effects on biological markers for antioxidative status. UGESKR. Laeger. 162, Nr. 10, 1388 - 1392 ZADNIK, T., M. KLINKON, M. NEMEC, M. MESARIC u. T. MODIC (2001): Anterior displacement of abomasum-Our clinical and laboratory experience. In: Ref. III. Mitteleurop. Buiatr. Kongr.: Gesundheitsproblematik der Wiederkäuer, Milovy, S. 159 - 163 ZAHN, N. u. M. FÜRLL (2004): Jahreszeitlicher Verlauf der SOD-Aktivität bei Rindern. Unveröffentlichte Mitteilung aus der Med. Tierklinik der Vet.-Med. Fak. der Univ. Leipzig ZIMA, T., S. STIPEK, J. CRKOVSKA, D. DOUDOVA, A. MECHUROVA u. P. CALDA (1996): Activity of the antioxidant enzymes superoxide dismutase and glutathione peroxidase in fetal erythrocytes. Prenatal. Diagn. 16, 1083 - 1085 ZIMMERMANN, B. J., D. J. GUILLORY, M. B. GRISHAM, T. S. GAGINELLA u. D. N. GRANGER (1990): Role of leukotriene B4 in granulocyte infiltration into the postischemic feline intestine. Gastroenterol. 99, 1358 – 1363 ZIMMERMANN, G. (1983): Superoxiddismutase ( Cu/Zn ): Aktivitätsbestimmung und Isolierung. Frankfurt, Univ., Fachber. Humanmed., Diss. ZÜLLKE, C. (1993): Veränderungen der Enzymaktivitäten der Radikalscavenger Superoxiddismutase und Katalase, sowie von plasmatischen Scavengersystemen im Rahmen der Beatmungstherapie der akuten respiratorischen Insuffizienz. München, Techn. Univ., Med. Fak., Diss. ZULAUF, M., C. SPRING, R. EICHER, M. MEYLAN, G. HIRSBRUNNER, G. SCHOLTYSIK u. A. STEINER (2002): Spontaneous in vitro contractile activity of specimens from the abomasal wall of healthy cows and comparison among dairy breeds. Am. J. Vet. Res. 63, Nr. 12, 1687 – 1694 - 125 - Literaturverzeichnis __________________________________________________________________________________________ - 126 - Tab. I: Vergleichende Darstellung der Hämoglobinkonzentration (Hb-Konz.), der Creatinkinase-Aktivität (CK-Aktivität), der TEACKonzentration (TEAC-Konz.), der Glutathionperoxidase-Aktivität (GPX-Aktivität) und der Superoxiddismutase-Aktivität (SOD-Aktivität) an den Entnahmestellen V. Jugularis (p) und den Venen des Labmagens (lm) innerhalb der Gruppen der linksseitigen Verlagerungen (LDA) zweiten (LDA 2) und dritten (LDA 3) Grades sowie der rechtsseitigen Verlagerungen (RDA) ohne (RDA sT) und mit (RDA cT) Torsion (Median, 1. und 3. Quartil) alle LDA (n=22) LDA 2 (n=9) LDA 3 (n=13) alle RDA (n=15) RDA sT (n=4) RDA cT (n=11) Hb-Konz. (p) (mmol/l) 18,0 (16,9; 19,1) 18,0 (17,1; 19,8) 18,0 (16,2; 18,6) 18,0 (17,3; 18,5) 18,2 (17,0; 19,3) 18,0 (17,4; 18,2) Hb-Konz. (lm) (mmol/l) 17,7 (16,9; 18,3) 18,3 (17,4; 19,2) 17,1 (16,5; 18,0) 18,9 (17,3; 19,2) 17,3 (17,0; 17,9) 19,2 (18,3; 19,4) CK-Aktivität (p) (U/l) 282 (209; 543) 489 (380; 1110) 233 (179; 305) 438 (316; 912) 619 (379; 886) 424 (316; 814) CK-Aktivität (lm) (U/l) 242 (174; 535) 484 (438; 1055) 176 (134; 224) 466 (280; 903) 611 (411; 825) 458 (283; 990) TEAC-Konz. (p) (µmol/l) 293 (215; 361) 361 (302;403) 239 (200; 309) 270 (213; 387) 303 (206; 415) 270 (235; 346) TEAC-Konz. (lm) (µmol/l) 241 (220 ;319) 317 (243; 343) 233 (205; 267) 325 (230; 359) 245 (226; 276) 334 (267; 373) SOD-Aktivität (p) (U/g Hb) 2491 (2194; 2814) 2718 (2443; 3501) 2438 (2055; 2550) 2061 (1667; 2449) 1969 (1677; 2232) 2061 (1793; 2449) SOD-Aktivität (lm) (U/g Hb) 2229 (2081; 2309) 2302 (2192; 2414) 2188 (1876; 2332) 2019 (1700; 2219) 1585 (1556; 1730) 2073 (1918; 2416) GPX-Aktivität (p) (U/g Hb) 429 (351; 520) 509 (444; 522) 395 (275; 429) 560 (508; 592) 505 (475; 537) 569 (539; 614) GPX-Aktivität (lm) (U/g Hb) 446 (359; 511) 490 (450; 519) 426 (308; 460) 521 (470; 570) 496 (470; 522) 522 (476; 598) Danksagung Mein besonderer Dank gilt Herrn Dr. habil. M. Fürll für die Überlassung des interessanten Themas, für seine Betreuung und die stets freundliche fachliche und geduldige Unterstützung bei der Anfertigung dieser Arbeit. In gleicher Weise bedanke ich mich bei Herrn Dr. T. Wittek für die Einarbeitung in das Themengebiet, das zielorientierte Arbeiten und die Betreuung. Außerdem danke ich dem Labor der Medizinischen Tierklinik unter der Leitung von FDC H. Kirbach für geistige Unterstützung bei der langwidrigen Probenbearbeitung. Ich danke für stets offene Augen und Ohren gegenüber Neulingen in ihrem Reich. Den Klinikmitarbeitern sei die Hilfe bei der Bändigung widerspenstiger Tiere und die zuvorkommende unkomplizierte Hilfsbereitschaft gedankt. Dem Team der Pathologie, im besonderen Herrn Prof. H.-A. Schoon, Frau Dr. H. Aupperle und Frau Dr. C. Ellenberger danke ich für die nette Zusammenarbeit auch über die erste Hürde hinaus. Auch wenn sie nicht direkten Anteil an der Promotion hatten, möchte ich besonders meinen Arbeitgebern der letzten vier Jahre danken, allen voraus Frau S. Fuchs, Frau Dr. M. Wiegand, Herrn H. Schnelle und Herrn A. Schneider. Herzlichen Dank an meine Freunde, besonders Ricci, die mir beigestanden haben, an meine Eltern, die dies immer getan haben und auch weiterhin werden, an meinen Hund und an meine Brüder.