Kapitel 38: Pflanzenreproduktion und Entwicklung
Werbung

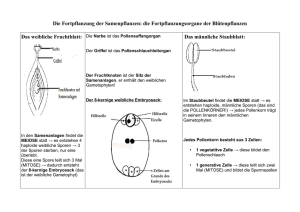
Kapitel 38: Pflanzenreproduktion und Entwicklung Sexuelle Reproduktion (Repetition von Kapitel 30) Lebenszyklus von Angiospermen (Bedecktsamer) und anderen Pflanzen Der Lebenszyklus der Angiospermen und anderen Pflanzen ist alternierend zwischen haploidem und diploidem Stadium. Zudem geht das eine Stadium aus dem anderen hervor. Diploide Pflanzen (Sporophyt) produziert meiotisch haploide Sporen. Die Sporen teilen sich mitotisch zu männlichen oder weiblichen Gametophyten (haploide Pflanzen). Durch mitotische Teilung entstehen aus den Gametophyten die Gameten (Spermium bzw Ei). Durch doppelte Befruchtung (double fertilization) entstehen die Zygoten welche durch Mitose neue Sporophyten bilden. (38.1 S.731) Der Sporophyt ist der dominante Teil der Pflanze, aber nur weil er sichtbar ist. Der Gametophyt hat sich im Laufe der Evolution stark reduziert und sich abhängig vom Sporophyten gemacht. (neu) Entwicklung der Gametophyten Blüten entstehen aus einer gestauchten Sprossachse mit vier Wirteln aus modifizierten Blättern, die durch sehr kurze Internodien (Segment des Pflanzenstengels zwischen zwei aufeinanderfolgenden Blattansätzen) getrennt sind. Das sind von aussen nach innen: Sepal (Kelchblatt): Meist grün und klein, schützt die Knospe. Petal (Kronblatt): Meist auffällig gefärbt, lockt Bestäuber (z.B Insekten) an Stamen (Staubblatt): Langer Faden (Filament) mit Anthere, die Pollenkörner (männl. Gametophyt) enthält. Wichtig für die Fortpflanzung. Carpel (Fruchtblatt): Narbe (Stigma) auf Griffel (Style) und Fruchtknoten (Ovary), der das Ovule (Samenanlage) mit dem Embryosack (weibl. Gametophyt) enthält. Wichtig für die Fortpflanzung. Blütenarten: Vollständige Blüten (complete flowers): enthalten alle vier Blätterarten (z.B. Lilien) Unvollständige Blüten (incomplete flowers): mindestens ein Blatttyp fehlt. (z.B. Gräser; fehlen die Kronblätter) Zwitterblüten (perfect flower): enthalten Staub- und Fruchtblätter Eingeschlechtliche Blüten (imperfect flowers): entweder Staubblätter (staminate) oder Fruchtblätter (carpellate). Sind beide Blütentypen auf derselben Pflanze, dann spricht man von einer einhäusigen (monoecious) Pflanze (z.B. Mais) resp. zweihäusig (dioecious), wenn sie nicht auf der gleichen Pflanze sind (z.B. Dattelpalmen). Die Tiere sind auch „zweihäusig“. Entwicklung der männlichen Gametophyten (Pollen) Jede Anthere besteht aus Theken mit je zwei Pollensäcken (Sporangien), in denen sich die Pollenmutterzellen (Mikrosporozyten) meiotisch teilen. Es entstehen vier haploide Mikrosporen aus jeder Mutterzelle. Die Mikrosporen teilen sich mitotisch weiter in eine generative und eine vegetative (tube) Zelle, die umgeben sind von einer dicken und harten Wand. Aus diesen drei Teilen besteht das Pollenkorn. Es wird geschlechtsreif, wenn das Korn auf eine Narbe trifft, weil dann sich die generative Zelle teilt in zwei Spermien und die vegetative beginnt, den Pollenschlauch zu bilden. Entwicklung der weiblichen Gametophyten (Embryosack) Der Embryosack entwickelt sich im Sporangium aus einer grossen Megasporocyte. Wie sich nun der Embryosack genau entwickelt, ist abhängig von der Pflanzenart. Hier ein Beispiel: Die diploide Megasporocyte teilt sich meiotisch in vier haploide Megasporen, von denen, im Gegensatz zu den Pollen, nur eine überlebt. Aus dieser einen Megaspore entwickelt sich der multizelluläre Gametophyt (sog Embryosack) mit der Eizelle (weibl. Gamet) flankiert von zwei Synergiden auf der einen Seite, mit drei Antipoden auf der anderen Seite und mit zwei Polarkerne in der Mitte. Das Ovule (Samenanlage) besteht jetzt nur noch aus Embryosack und Integument (Schutzgewebe). (Bild 38.4 S.734) Bestäubung (Pollenkörner gelangen auf die Narbe) Wie gelangen Pollenkörner auf die Narbe? Wind: Es braucht sehr viele Pollenkörner Tiere: effektiver, Gefahr der Selbstbestäubung! Selbstbestäubung: sehr wenige Pflanzen. Wie wird Selbstbestäubung verhindert? (Ziel: die genetische Vielfalt zu erhalten) - zweihäusige Pflanzen - Verschiedene Reifezeit von Staub- und Fruchtblättern. - Bei tierbestäubten Pflanzen: Anordnung der Staub bzw Fruchtblätter wurde so angelegt, dass gar keine Selbstbestäubung möglich ist: - Selbstinkompatible Pflanzen: Sie haben eine chemische Barriere entwickelt. Sie verhindert das Wachsen des Pollenschlauchs von einem Pollenkern derselben Pflanze. Molekulare Mechanismen der Selbstinkompatibilität Selbstinkompatibilität ist die Fähigkeit der Pflanzen, genetisch sehr nah verwandte Pollenkörner abzuweisen. Es funktioniert ähnlich dem Immunsystem der Tiere, mit dem einzigen Unterschied, nicht das Fremde, sondern das eigene abzuweisen. Die Pflanze kann das eigene erkennen mit Hilfe der S-Lokus. Dort befinden sich über 50 verschiedene Allele. Wenn jetzt ein Allel des Pollenkorns mit dem der Narbe übereinstimmt, kommt es zu keiner Fertilisation. Man unterscheidet zwei Fälle: - gametophytische Selbstinkompatibilität: Das Pollenkorn (Gametophyt) selber wird blockiert. Z. B. in Rosen oder Bohnen führt der Selbsterkennungsmechanismus zur enzymatischen Zerstörung der RNA des in Entwicklung stehenden Pollenschlauchs. - Sporophytische Selbstinkompatibilität: Zellen der Narbe blockieren das Pollenkorn. Z. B. Kreuzblütler: Das Pollenkorn setzt ein chemisches Signal frei, welches mit mindestens einem Allel übereinstimmt mit dem Allel auf dem S-Locus. Diese chemische Substanz bindet an ein Rezeptormolekül, welches an ein Kinasemolekül der Membran andockt. Dieses wiederum löst ein Signalweg (STP=Signal-Transduction-Pathway) aus, was zur Folge hat, dass ein oder mehre Effektorproteine freigesetzt werden und damit das Pollenkorn geblockt wird. (Bilder 38.5 und 38.5 S.735) Doppelte Befruchtung ( double fertilization) Das Pollenkorn produziert aus der generativen Zelle einen Schlauch, der bis zur Eizelle wächst, dort durch die Mikropyle (= Spalt im Integument der Eizelle) in die Eizelle eindringt. Während des Wachstums des Schlauches teilt sich auch die generative Zelle in zwei Spermien. Diese liegen in der Schlauchspitze und werden so hinuntertransportiert. Der Schlauch selber wird möglicherweise mit Hilfe von Kalzium zur Eizelle gesteuert. Nachdem die beiden Spermien eingedrungen sind, befruchtet das erste die Eizelle (wird zur Zygote) und das zweite bildet mit den Polarkernen eine triploide Zelle, welches später zum sekundären Endosperm (Nährgewebe) wird. Vorteil der doppelten Befruchtung: Die Pflanze spart wertvolle Energie, weil das Nährgewebe erst aufgebaut wird, wenn die Eizelle befruchtet worden ist. Die Samenanlage (Ovule) entwickelt sich zum Samen (Seed) Endospermentwicklung Diese Entwicklung beginnt vor der Embryoentwicklung. Das Endosperm entwickelt sich auf zwei Arten: - Monoctyledonen (Einkeimblättrige): brauchen ein Teil des Endosperms für die Entwicklung des Embryos und der andere Teil wird für den Keimling (seedling) nach der Keimung ( germination) gelagert. - Dicotyledonen (Zweikeimblättrige): das ganze Endosperm wird für die Entwicklung des Endosperms gebraucht, das heisst, der reife Samen hat kein Endosperm mehr. Embryoentwicklung Die Zygote teilt sich in eine Terminal- und eine Basalzelle. Aus der Terminalzelle entsteht durch gleichmässige Teilungen der kugelförmige Proembryo. Dieser entwickelt sich weiter zu einem oder zwei Cotyledonen (Keimblätter). Die Basalzelle teilt sich längs weiter und bildet den Suspensor, welcher den Embryo am Integument der Samenanlage befestigt. Bei den Dicotyledonen ist nach der Ausbildung der Hügel, zwischendrin der Keimspross (embryonic shoot) sichtbar und beim Suspensor nach der Befestigung die Keimwurzel (embryonic root). In den Cotyledonen unterscheidet man auch noch das Protoderm, Grundmeristem und Procambium. Diese entwickeln sich zu Abschlussgewebe (dermal tissue), Grundgewebe und Leitgewebe (valscular tissue). Während der Embryo wächst lagert der Samen Proteine, Fette und Stärke solange ein, bis der Embryo keimt. Im letzten Reifestadion dehydriert der Samen. Bild 38.8 S.737 Struktur des reifen Samens Der Embryo ist vor der Keimung umgeben von einer Samenschale (seed coat), die entstanden ist aus dem Integument der Samenanlage (Ovule). Bild 38.9 S. 737 - Hypocotyl: Nennt man den Bereich, der auf der embrionalen Achse unterhalb des Befestigungspunktes der Cotyledonen liegt. Hypocotyl endet in der Keimwurzel (radicle). - Epicotyl: Nennt man den Bereich, der auf der embrionalen Achse oberhalb des Befestigungspunktes liegt. An der Spitze befindet sich die Plumela (Sprossspitze mit 2 kl. Blätter) Gemeine Gartenbohne (Common Bean): Cotyledonen sind fleischig, weil sie Nahrung vom Endosperm absorbiert haben. Rizinus (Castor Bean): Cotyledonen sind sehr dünn, nehmen Nährstoffe erst auf, wenn sie keimen. Monocotyledonen (z.B. Weizen und Mais): Diese haben spezielle Cotyledonen, die man Scutellum nennt. Sie sind sehr dünn, ebenfalls aus dem obengenannten Grund. Das Scutellum ist von zwei Scheiden eingehüllt; Die Coleorhiza und die Coleoptile. Bild 38.9 S.737 Fruchtknoten (Ovary) entwickelt sich zur Frucht Aus dem Fruchtknoten entwickelt sich die Frucht. Dabei wird die Fruchtknotenwand zum Pericarp (= Fruchtwand). Die Frucht hilft bei der Verbreitung und schützt zusätzlich den Samen. Verschiedene Typen von Früchten: Simple Fruit: Entstanden aus einem Fruchtknoten (Kirsche) Aggregate Fruit (Sammelfrucht): Entstanden aus einer Blüte mit mehreren Fruchtblättern (Himbeeren) Multiple Fruit (Fruchtverband): Entstanden aus mehreren Blüten und mehreren Fruchtknoten. (Ananas) Wenn die Früchte reifen, können sie die Farbe ändern, trocknen oder erweichen. Zudem können noch chemische Stoffe freigesetzt werden, wie z. B. Zucker oder Säure. Dies macht dann die Früchte für uns Menschen geniessbar. Evolutionäre Anpassung im Keimungsprozess (seed germination) In jedem Samen steckt schon eine kleine Miniaturpflanze. Manche Samen keimen sobald sie in einer geeigneten Umgebung sind und manche keimen erst, wenn ein bestimmter Reiz (Licht, Regen, Hitze, Kälte, chemische Stoffe) aus der Umwelt eintrifft. Wie lange die Samen „schlafen“ (seed dormancy) hängt von diesen Reizen ab. Im Boden drin gibt es folglich immer solche Schläfer. Dies ist mit ein Grund, warum sich die Vegetation nach einem Brand, Überschwemmung usw schnell wieder aufbaut. Ablauf des Keimens Das Keimen des Samens beginnt mit dem anschwellen des Samens, aufgrund der Wasseraufnahme (imbibition). Gleichzeitig werden Hormone freigesetzt welche die Produktion von Enzymen anregen, die Nahrungsmittel hydrolisieren können. Z.B die AlphaAmylase, die die Stärke in Zucker spaltet. Zucker und andere so hergestellte Nahrungsmittel werden vom Embryo aufgenommen und er beginnt zu wachsen. Zuerst kommt die Keimwurzel. Für das Durchbrechen des Bodens gibt es folgende Methoden: Bohne: Haken im Hypocotyl zieht Epicotyl und Cotyledonen ans Licht. Nach dem alle Vorräte der Cotyledonen aufgebracht wurde, sterben sie ab. Dies ist nicht weiter tragisch den die ersten Blätter beginnen schon sehr schnell Photosynthese zu machen. Erbse: Haken im Epicotyl zieht die Plumula mit den ersten Laubblättern ans Licht, die Cotyledonen bleiben im Boden. Mais (Monocots): Coleoptile gräbt sich senkrecht ans Licht. Anschliessend kann die Pflanze weiterwachsen dank dieses Tunnels. In diesem Stadium im Lebenszyklus der Pflanze ist sie vielen Gefahren ausgesetzt. Das Überlebensproblem löst sie aber mit einer grossen Anzahl Nachkommen ( =>Erhöhung der Überlebenschancen). Bild 38.12 S.740 Asexuelle Reproduktion Viele Pflanzen können sich klonen. Asexuelle Reproduktion = vegetative Reproduktion Bei den Pflanzen funktioniert die asexuelle Reproduktion, weil sie ein meristematisches Gewebe(Gewebe aus undiff. Zellen, die sich unendlich teilen können) haben und weil sie mit den Parenchymzellen (Zellen, die sich teilen und sich in verschiedene Typen von speziellen Zellen differenzieren können), die sie überall in ihrem Körper haben, kaputte Teile regenerieren können. Fragmentation: Aufteilung einer Adultpflanze in mehrere Teile, aus welchen dann wieder neue Pflanzen entstehen. Bild 38.13 S.741 Kalanchoe („Kindlibaum“) als Beispiel. Er bildet an den Blattenden Jungpflanzen, die abfallen und heranwachsen zu Adultpflanzen. Eine andere Möglichkeit besteht darin, in den Wurzeln Ausläufer zu bilden, die dann einen neuen Spross bilden. Beispiel: Espe. Apomixis: (=Selbstbefruchtung der nicht befruchteten Blüten) Z. B. Löwenzahn: Eine diploide Zelle in der Samenanlage erzeugt einen Embryo. Dank dieses Vorganges haben diese Pflanzen den Vorteil, sich mit dem Wind zu verbreiten, was für sie neue Gebiete erschliessen lässt. Vegetative Vermehrung in der Landwirtschaft. Arten der Vermehrung - Mit Stecklingen: Man schneidet von einer Pflanze Sprossteile ab. Diese bilden dann zuerst viele undiff. Zellen, den Callus, aus diesem wiederum entstehen Wurzeln und anschliessend eine neue Pflanze. - Mit Reisern: Reiser sind Zweige, die auf Wurzelstöcke von anderen Arten daraufgesetzt (grafted) werden. Der untere Teil ist die Unterlage (Stock), der obere Teil Pfropfreis (scion). Das genetische Material vom scion vermischt sich nicht mit dem stock. Damit kann man z.B Super-Weintrauben konstruieren: sehr resistente Wurzeln mit den guten französischen Weintrauben verbinden. - Im Reagenzglas: Aus einzelnen Parenchymzellen oder kleinen Gewebestückchen kann man in einem geeigneten Nährmedium vollständige Pflanzen züchten. Wichtig für die Gentechnologie. Mit Hilfe einer DNA-Kanone kann fremde DNA in die DNA von einer Pflanze hineingeschossen werden. Eine andere Methode ist die Protoplastenfusion: Man fusioniert zwei Protoplasten (Pflanzenzellen ohne Zellwand) zu einer Hybridpflanze und züchtet diese anschliessend. Vor- und Nachteile der Monokultur Vorteile: - Die Pflanzen reifen gleichzeitig - Ermöglicht den Bauern die Menschheit zu ernähren Nachteile: Anfällig gegen Krankheitserreger. Es brauchte nur einen einzigen, um das ganze Feld zu zerstören. Beispiele: Irland ( Kartoffel) Lösung: Gendatenbanken, um die Vielfalt zu erhalten. Sexuelle und asexuelle Reproduktion sind komplementär in der Lebensgeschichte Vorteile sexuelle Reproduktion - Variation in der Population bleibt erhalten - zeitlich und örtlich flexibler Vorteile asexuelle Reproduktion - Sehr schnelle Vermehrung - Weniger anfällige Jungpflanzen Zelluläre Mechanismen in der Pflanzenentwickllung Entwicklung ist die Summe aller Veränderungen, die einen Organismus verändert haben. Überblick über die Pflanzenentwicklung Drei Prozesse spielen eine wichtige Rolle bei der Entwicklung vom Embryo zur Pflanze - Wachstum (growth): Der Embryo macht unzählige Zellteilungen durch und vergrössert sich dadurch. - Morphogenese (Formwerdung): Gibt der Pflanze die Form, sonst würde es nur ein Zellhaufen geben und keine Pflanze. - Zelldifferenzierung: Ist wichtig, damit die verschiedenen Funktionen der Pflanze überhaupt durchgeführt werden können. Das Cytoskelett gibt die Geometrie der Zellteilung und Expansion vor Pflanzenzellen haben eine Zellwand, die keine Bewegung ermöglicht. Damit haben sie einen grossen Nachteil gegenüber den Tieren. Deshalb ist es für sie enorm wichtig, ein organisiertes Wachstum zu haben. Verantwortlich dafür ist das Cytoskelett. Bei der Zellteilung bildet sich in der G2-Phase ein Band aus Mikrotubuli, das sog. Preprophase Band. Dieses entscheidet wo sich die Zelle teilt. Je nach Lage des Bandes teilt sich die Zelle horizontal oder vertikal. Dieses Band verschwindet aber noch vor der Metaphase. Es dient lediglich zur Festlegung des Ortes für die Mikrotubuli. Zusätzlich legt das Cytoskelet mit seiner Anordnung fest, in welche Richtung die Zellverlängerung stattfindet, da es die Orientierung der Mikrotubuli kontrolliert. Beim Zellwachstum: In der innersten Zellwandschicht gibt es viele parallele Mikrofibrillen aus Zellulose. Das Zellwachstum verläuft senkrecht zu diesen Fasern, indem die Säure aktivierten Enzyme Querverbindungen zwischen den Mikrofibrillen auflösen, die nun schwächere Zellwand dem Turgor in die gewünschte Richtung nachgibt und Wasser in die Zelle fliesst. Im Gegensatz wachsen Tiere nicht indem sie Wasser einlagern, sondern indem sie das Cytoplasma synthetisieren. Differenzierung der Zelle ist abhängig von der Kontrolle der Genexpression Wie sich eine Zelle schlussendlich differenziert, ist abhängig von der Genexpression. Je nachdem werden zu bestimmten Zeitpunkten gewisse Gene aktiviert, die dann dazuführen, dass die Zelle Enzyme produziert, die bewirken, dass sich die Zelle zum Beispiel zu einer Xylemzelle differenziert. Dies geschieht, wenn das Enzym Lignin (härtet die Zellwand) produziert wurde. Ist es nicht vorhanden, gibt es eine Schutzzelle (guard cells) mit einer flexiblen Zellwand. Wie die Aktivierung genau funktioniert weiss man noch nicht. Man konnte aber zeigen, dass jede Zelle das ganze Genom enthält und damit sich zu jedem Zelltyp differenzieren kann. Musterbildung (pattern formation) Musterbildung: Die Entwicklung von bestimmten Strukturen an bestimmten Orten. Wichtig für die Musterbildung der Pflanze ist die Lageinformation, das heisst die Zelle muss Informationen über, wo sie hingehört, erhalten. Hier hat man die Vermutung, dass der Konzentrationsgradient von bestimmten Molekülen (meistens Proteine) eine entscheidende Rolle spielt. Für die genaue Orientierung bräuchte es zum Beispiel nur Informationen von der Zelle zuvorderst im Apikalmeristem ( Nord-Süd Orientierung) und der äussersten Zelle des Sprosses (West-Ost Orientierung). Man versuchte auch herauszufinden ob jede Zelle einen vorbestimmten Weg hat, das zum Beispiel diese Zelle an diesem Ort des Meristem zu einer Zelle im Gewebe des Blattes wird. Diese Frage kann man nicht allgemein beantworten, da der Ort an der die undiff. Zelle gelandet ist entscheidet, welche Differenzierung die Zelle erfährt. Genetische Basis der Musterbildung am Beispiel einer Blütenentwicklung Faktoren, die die Blütenbildung aktivieren: Tageslänge und Hormone (mehr dazu in Kapitel 39) Nach der Aktivierung sind zwei Typen von Genen wichtig. Meristemidentitätsgene ( induzieren die Blütenbildung) und die Organidentitätsgene ( organisieren die Bildung von den Blütenblättern). Ohne Organidentitätsgene gäbe es zum Beispiel nur Kelchblätter und keine Kronblätter. Bild 38.21 S 748: In diesem Beispiel legen drei Organidentitätsgene (A,B,C) fest, welche Art von Blütenblätter es schlussendlich gibt.