Photorespiration: ein Relikt der Evolution?
Werbung

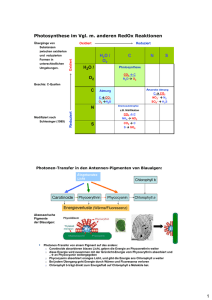
10. Photosynthese 10.1 Pflanzen und andere autotrophe Organismen sind die Primärproduzenten der Biosphäre Jedes Lebewesen muss die organischen Verbindungen, die es zu Gewinnung von Energie und Kohlenstoffgerüsten braucht, auf einem von zwei Wegen gewinnen: durch autotrophe oder heterotrophe Ernährung. Autotrophe Organismen stellen ihre organischen Moleküle aus anorganischen Rohstoffen her, die sie aus der Umwelt beziehen. Deshalb bezeichnet man die autotrophen Organismen auch als Primärproduzenten der Biosphäre. Die Überführung körperfremder Stoffe in körpereigene Substanzen nennt man auch Assimilation. Pflanzen sind autotroph, oder genauer gesagt, photoautotroph: Sie synthetisieren organische Moleküle mithilfe des Sonnenlichts aus CO2 und Wasser. Heterotrophe Organismen verschaffen sich ihr organisches Material mit einer anderen Ernährungsweise. Sie können den lebensnotwendigen Kohlenstoff nicht aus CO2 gewinnen, sondern nur aus organischen Verbindungen, die andere Lebewesen erzeugt haben; deshalb bezeichnet man die heterotrophen Arten als Konsumenten der Biosphäre. Viele heterotrophe Arten leben von der Zersetzung organischer Abfälle – beispielsweise von Kadavern, Kot oder Laub; deshalb nennt man sie auch Destruenten oder „Saprobionten“ (Fäulnisbewohner). 10.2 Chloroplasten sind bei Pflanzen die Orte der Photosynthese Ein Quadratmillimeter der Blattoberfläche enthält etwa eine halbe Million Chloroplasten. Ihre Farbe haben die Blätter vom Chlorophyll, dem grünen Pigment in den Chloroplasten. Die vom Chlorophyll absorbierte Lichtenergie treibt die Synthese der Nährstoffmoleküle in den Chloroplasten an. Die meisten Chloroplasten liegen zwischen oberer und unterer Epidermis in den Zellen des Mesophylls, des Gewebes im Innern der Blätter. Mikroskopisch kleine Poren, die Spaltöffnungen oder Stomata, lassen Kohlendioxid in das Blatt eintreten und Sauerstoff entweichen. Das von den Wurzeln aufgenommene Wasser gelangt über bestimmte Gefässe der Leitbündel in die Blätter. Andersartige Leitbündelröhren dienen zum Transport des Zuckers von den Blättern zu den Wurzeln und anderen nicht photosynthetisch aktiven Teilen der Pflanze. 10.3 Nachdem man wusste, dass Chloroplasten Wassermoleküle spalten, konnte man Atome durch die Photosynthese verfolgen: Wie Forschung funktioniert Unter dem Einfluss von Licht produzieren die grünen Teile der Pflanzen aus Kohlendioxid und Wasser organisches Material und Sauerstoff. In der Sprache der Chemie lässt sich die Photosynthese mit folgender Reaktionsgleichung zusammenfassen: 6 CO2 + 12 H2O + Lichtenergie C6H12O6 + 6 O2 + 6 H2O Wir können die Gleichung vereinfachen, indem wir den Nettoverbrauch an Wasser angeben: 6 CO2 + 6 H2O + Lichtenergie C6H12O6 + 6 O2 Als nächstes kürzen wir die Reaktionsgleichung der Photosynthese durch 6 und bringen sie so in die einfachste Form, die möglich ist: CO2 + H2O CH2O + O2 Die Spaltung des Wassers Einen der ersten Hinweise auf den Mechanismus der Photosynthese erhielt man aus der Entdeckung, dass der Sauerstoff, den die Pflanzen abgeben, nicht aus dem Kohlendioxid, sondern aus dem Wasser stammt. Mit dem schweren Isotop Sauerstoff-18 als Markierung verfolgte man das Schicksal der Sauerstoffatome während der Photosynthese. Das von den Pflanzen abgegebene O2 war nur dann mir Sauerstoff-18 markiert, wenn sich die Markierung zuvor in den Wassermolekülen befunden hatte. Brachte man den Sauerstoff-18 mit dem Kohlendioxid in die Pflanzen, tauchte die Markierung im freigesetzten Sauerstoff nicht auf. Das wichtigste Ergebnis der Atomumordnung während der Photosynthese ist die Abspaltung des Wasserstoffs aus dem Wasser und sein Einbau in Zucker. O2, das Abfallprodukt der Photosynthese, ersetzt in der Atmosphäre den Sauerstoff, der durch die Zellatmung verbraucht wird. Wie das Experiment also gezeigt hat, spaltet der Chloroplast Wassermoleküle in Wasserstoff und Sauerstoff (Photolyse), wobei die Elektronen des Wasserstoffs in die Bindungen der Zuckermoleküle eingehen. Die Photosynthese ist also ein Redoxvorgang, bei dem Wasser oxidiert und Kohlendioxid reduziert wird. 10.4 Die Lichtreaktionen und der Calvin-Zyklus wirken zusammen und setzen Lichtenergie in die chemische Energie der Nährstoffe um: Eine Übersicht Die beiden Abschnitte der Photosynthese bezeichnet man als Lichtreaktionen (der PhotoTeil der Photosynthese) und als Calvin-Zyklus (der Synthese-Teil). Die Lichtreaktionen in der Thylakoidmembran erzeugen ATP (sie liefern die Energie für die Bindung einer Phosphatgruppe an ADP, ein Vorgang, den man als Photophosphorylierung bezeichnet) und spalten Wasser; dabei wird Sauerstoff frei, und durch die Übertragung von Protonen und Elektronen vom H2O auf das NADP+ entsteht NADPH. Wie zu beachten ist, entsteht in den Lichtreaktionen kein Zucker; das geschieht erst im Calvin-Zyklus, dem zweiten Abschnitt der Photosynthese. Der Zyklus beginnt, indem CO2 aus der Luft mit organischen Molekülen reagiert, die bereits im Chloroplasten vorhanden sind. Diesen ersten Einbau von Kohlenstoff in organische Verbindungen bezeichnet man als Kohlenstoff-Fixierung. Der so gebundene Kohlenstoff wird dann im Calvin-Zyklus durch Aufnahme von Elektronen zum Kohlenhydrat reduziert. Der Calvin-Zyklus ist also derjenige Prozess, der die Zuckermoleküle erzeugt, aber das ist nur mit Hilfe von NADPH und ATP aus den Lichtreaktionen möglich. Die Stoffwechselreaktionen des Calvin-Zyklus bezeichnet man manchmal auch als Dunkelreaktionen, weil keine davon unmittelbar Licht benötigt. Der Calvin-Zyklus läuft im Stroma ab. 10.5 Die Lichtreaktionen verwandeln Sonnenenergie in die chemische Energie von ATP und NADPH: Eine nähere Betrachtung Die Eigenschaften des Sonnenlichts Licht ist eine Form der sogenannten elektromagnetischen Energie, auch Strahlung genannt. Elektromagnetische Energie wandert in Rhythmischen Wellen. Den Abstand zwischen den Kämmen elektromagnetischer Wellen bezeichnet man als Wellenlänge. Den gesamten Wellenlängenbereich nennt man auch elektromagnetisches Spektrum. Nur einen sehr kleinen Teil davon können wir als sichtbares Licht (380-750 nm) verschiedener Farben wahrnehmen. Das Modell, wonach Licht aus Wellen besteht, erklärt viele seiner Eigenschaften, aber in mancherlei Hinsicht verhält sich das Licht auch so, als bestünde es aus einzelnen Teilchen, die man Photonen nennt. Je kürzer die Wellenlänge, desto grösser die Energie eines Photons der betreffenden elektromagnetischen Strahlung. Der Teil des Spektrums, den wir sehen können – das sichtbare Licht – treibt die Photosynthese an. Blau und Rot, die beiden Wellenlängen, die das Chlorophyll am wirksamsten absorbiert, sind auch die Farben, die für die Lichtreaktionen am nützlichsten sind. Photosynthesepigmente: die Lichtrezeptoren Wenn Licht auf Materie trifft, kann es reflektiert, durchgelassen oder absorbiert werden. Substanzen, die sichtbares Licht bestimmter Wellenlängen absorbieren, nennt man Pigmente. Ein Blatt erscheint grün, weil das Chlorophyll rotes und blaues Licht absorbiert und grünes Licht reflektiert beziehungsweise durchlässt. Die Kurve, die sich ergibt, wenn die Lichtabsorption eines Pigments gegen die Wellenlänge aufgetragen wird, bezeichnet man als Absorptionsspektrum. Genauer als das Absorptionsspektrum gibt das Wirkungsspektrum die Leistungsfähigkeit der einzelnen Wellenlängen wieder. Ein Wirkungsspektrum erhält man, wenn man die Chloroplasten mit Licht verschiedener Wellenlängen bestrahlt und dann die Wellenlängen gegen die gemessene Photosyntheserate aufträgt. Nur Das Chlorophyll a ist unmittelbar an den Lichtreaktionen beteiligt, durch die das Sonnenlicht in chemische Energie umgesetzt wird, aber auch andere Pigmente können Licht absorbieren und die Energie dann auf das Chlorophyll a übertragen, das die Lichtreaktionen in Gang setzt. Eines dieser Hilfspigmente oder „akzessorischen Pigmente“ ist das Chlorophyll b, das sich nur geringfügig vom Chlorophyll a unterscheidet. Absorbiert das Chlorophyll b ein Photon des Sonnenlichts, überträgt es die Energie auf das Chlorophyll a, das sich dann genauso verhält, als habe es das Photon direkt aus dem Sonnenlicht aufgenommen. Weiterhin befinden sich in den Chloroplasten die Carotinoide, eine Familie von Hilfspigmenten in verschiedenen Farbvarianten von gelb und orange. Diese Moleküle sind zusammen mit den beiden Chlorophylltypen in die Thylakoidmembran eingelagert. Anregung des Chlorophylls durch Licht Energie kann nicht verschwinden; wenn ein Molekül mein Photon absorbiert, wird eines seiner Elektronen auf ein höheres Niveau gehoben. Solange das Elektron sich auf dem niedrigeren Energieniveau befindet, ist das Pigmentmolekül im Grundzustand. Hat ein absorbiertes Photon das Elektron jedoch auf das energiereichere Niveau verschoben, spricht man vom angeregten Zustand des Pigments. In isolierten Pigmenten kann das Elektron dort aber nicht lange bleiben, denn der angeregte Zustand ist wie alle hohen Energieniveaus instabil. Wenn Pigmente Licht absorbieren, fallen ihre angeregten Elektronen im Allgemeinen schon nach einer Milliardstelsekunde wieder auf das Niveau des Grundzustandes zurück, und die Energiedifferenz wird als Wärme frei. Manche Pigmente, so auch das Chlorophyll, senden nach der Absorption von Photonen nicht nur Wärme, sondern auch Licht aus. Das Elektron springt in einen energiereicheren Zustand, und wenn es zurückfällt, wird ein Photon abgegeben. Ein solches Nachleuchten bezeichnet man als Fluoreszenz. Die Photosysteme: Lichtsammelkomplexe in der Thylakoidmembran Die Anregung des Chlorophylls in den Chloroplasten hat völlig andere Folgen, als wenn isoliertes Chlorophyll Lichtenergie absorbiert. In der natürlichen Umgebung der Thylakoidmembran ist das Chlorophyll zusammen mit anderen Molekülen in Photosystemen organisiert. Ein Photosystem besitzt einen lichtsammelnden Antennenkomplex (auch „LHC“, vom englischen light harvesting complex) aus mehreren hundert Chlorophyll-a-, Chlorophyll-b- und Carotinoidmolekülen. Wenn eines dieser „Antennenpigmente“ ein Photon absorbiert, wird dessen Energie zum nächsten Antennenpigment weitergeleitet, bis sie zwei ganz bestimmte Chlorophyll-a-Moleküle erreicht. Das Besondere an diesen ist nicht ihre Struktur, sondern ihre Position. Nur diese beiden Chlorophyllmoleküle befinden sich im Reaktionszentrum, jenem Bereich des Photosystems, in dem die erste lichtgetriebene chemische Reaktion der Photosynthese stattfindet. Neben dem speziellen Chlorophyll-a-Paar befindet sich im Reaktionszentrum ein weiteres Molekül, der primäre Elektronenakzeptor. An ihn verliert das Chlorophyll-a-Paar in einer Redoxreaktion eines seiner Elektronen. Die von der Sonne angetriebene Elektronenübertragung vom Chlorophyll zum primären Elektronenakzeptor ist die erste Stufe der Lichtreaktionen. Die Thylakoidmembran ist mit zwei Typen von Photosystemen besetzt, die in den Lichtreaktionen der Photosynthese zusammenwirken. Sie werden in der Reihenfolge ihrer Entdeckung als Photosystem I und Photosystem II bezeichnet. Jedes von ihnen besitzt ein charakteristisches Reaktionszentrum mit einem typischen primären Elektronenakzeptor und einem Chlorophyll-a-paar (diese Chlorophyll-a-Paare (Pigmente) werden P700 bzw. P680, nach ihrem besten Absorptionsbereich, genannt), das mit spezifischen Proteinen assoziiert ist. Als nächstes wollen wir uns ansehen, wie die beiden Photosysteme durch ihr Zusammenwirken ATP und NADPH erzeugen, die Hauptprodukte der Lichtreaktionen. Nichtzyklischer Elektronentransport Beim nichtzyklischen Elektronentransport, der während der Lichtreaktionen der Photosynthese vorherrscht, kehren die von den Chlorophyllmolekülen abgegebenen Elektronen nicht mehr zu ihrem Ursprungsort zurück, sondern sie werden vom primären Elektronenakzeptor eingefangen. Das so oxidierte Chlorophyll wird dabei zu einem sehr starken Oxidationsmittel, indem es bestrebt ist, seine Elektronenlücke wieder zu füllen. Wenn das Photosystem II Licht absorbiert, werden im Reaktionszentrum die auf ein höheres Energieniveau gebrachten Elektronen des P680 vom primären Elektronenakzeptor eingefangen. Ein Enzymkomplex, der wasserspaltende Komplex, entzieht einem Wassermolekül zwei Elektronen und überträgt sie auf die beiden P680-Moleküle, wo sie die nach der Absorption der Lichtenergie verlorengegangenen Elektronen ersetzen. Diese lichtinduzierte Wasserspaltung wird als Photolyse bezeichnet. Sie setzt in der Photosynthese das O2 frei. Die angeregten Elektronen fliessen über eine Elektronentransportkette vom primären Elektronenakzeptor des Photosystems II zum Photosystem I. Diese Kette ähnelt stark derjenigen der Zellatmung. In den Chloroplasten besteht sie zunächst aus einem Elektronenüberträger namens Plastochinon (Pq), dann folgt ein Komplex aus Zwei Cytochromen und schliesslich ein kupferhaltiges Protein namens Plastocyanin (Pc). Wenn die Elektronen die Energiekaskade der Transportkette passieren, nutzt die Thylakoidmembran ihren exergonischen Fluss zur Produktion von ATP. Diese Art der ATPSynthese nennt man Photophosphorylierung, weil sie von Lichtenergie angetrieben wird. Genauer bezeichnet man die Produktion von ATP durch nichtzyklischen Elektronentransport als nichtzyklische Photophosphorylierung. Das in den Lichtreaktionen gebildete ATP liefert die Energie für die Zuckerproduktion im Calvin-Zyklus, dem zweiten Abschnitt der Photosynthese. Wenn die Elektronen das untere Energieniveau der Transportkette erreichen, füllen sie Elektronenlücken im P700 auf, dem Chlorophyll-a-Paar im Reaktionszentrum des Photosystems I. Sie ersetzen dort diejenigen Elektronen, welche die Lichtenergie vom P700 zum primären Elektronenakzeptor des Photosystems I getrieben hat. Der primäre Elektronenakzeptor des Photosystems I gibt die angeregten Elektronen an das Ferredoxin (Fd) weiter, ein eisenhaltiges Protein. Ein Enzym namens NADP+-Reductase überträgt die Elektronen dann von Fd auf NADP+. Durch diese Redoxreaktion werden die energiereichen Elektronen im NADPH gespeichert, jenem Molekül, welches im CalvinZyklus das Reduktionsmittel für die Zuckersynthese darstellt. Mit Sonnenenergie werden in den Lichtreaktionen ATP und NADPH regeneriert; ATP ist die chemische Energiequelle und NADPH das Reduktionsmittel für die Zuckersynthese im Calvin-Zyklus. Zyklischer Elektronentransport Unter bestimmten Bedingungen können die angeregten Elektronen einen anderen Weg einschlagen, den zyklischen Elektronentransport, an dem nur das Photosystem I beteiligt ist, nicht aber das Photosystem II. Dabei wird weder NADPH produziert noch Sauerstoff freigesetzt. Der zyklische Elektronentransport erzeugt aber ATP. Man spricht deshalb auch von zyklischer Photophosphorylierung im Gegensatz zur nichtzyklischen Photophosphorylierung. Welche Funktion erfüllt der zyklische Elektronentransport? Beim nichtzyklischen Prozess entstehen ATP und NADPH in ungefähr äquimolaren Mengen, aber der Calvin-Zyklus verbraucht mehr ATP als NADPH, wie wir noch sehen werden. Den Unterschied gleicht der zyklische Elektronentransport aus. Die Chemiosmose der Chloroplasten und Mitochondrien: Ein Vergleich Chloroplasten und Mitochondrien erzeugen ATP durch den gleichen grundlegenden Mechanismus: die Chemiosmose. Eine in die Membran integrierte Elektronentransportkette pumpt Protonen durch diese Membran, und gleichzeitig durchlaufen Elektronen eine Energiekaskade hin zu immer elektronegativeren Transportmolekülen. Die Elektronentransportkette wandelt also im Verlauf von Redoxreaktionen freigesetzte chemische Energie in protonenmotorische Kraft um, in potentielle Energie, die sich aus dem H+-Gradienten und dem Membranpotential zusammensetzt. In dieselbe Membran ist ein ATP-Synthase-Komplex eingelagert, der die durch die protonenmotorische Kraft getriebene Diffusion der Protonen mit der Phosphorylierung von ADP koppelt. Einige Elektronenüberträger, so die eisenhaltigen Proteine aus der Familie der Cytochrome, sind sich in Chloroplasten und Mitochondrien sehr ähnlich. Auch die ATP-Synthase-Komplexe der beiden Organellentypen ähneln sich stark. Aber es gibt auch auffällige Unterschiede zwischen der oxidativen Phosphorylierung in den Mitochondrien und der Photophosphorylierung in den Chloroplasten. In den Mitochondrien wurden die energiereichen Elektronen, welche die Transportkette durchlaufen, den Nährstoffmolekülen durch Oxidation entzogen. Chloroplasten brauchen keine Nährstoffe, um ATP zu erzeugen; ihre Photosysteme fangen Lichtenergie ein und heben mit ihrer Hilfe die Elektronen energetisch an den Anfang der Transportkette. Mit anderen Worten: Mitochondrien übertragen chemische Energie von den Nährstoffmolekülen auf ATP, Chloroplasten dagegen wandeln Lichtenergie in chemische Energie um. Auch in der räumlichen Anordnung der Chemiosmose unterscheiden sich Chloroplasten und Mitochondrien. (Abb. 10.14) 10.6 Im Calvin-Zyklus dienen ATP und NADPH dazu, Zucker aus CO2 herzustellen: Eine nähere Betrachtung Bei dem Kohlenhydrat, das im Calvin-Zyklus unmittelbar entsteht, handelt es sich nicht um Glucose, sondern um Glycerinaldehyd-3-phosphat (G3P), einen Zucker mit drei Kohlenstoffatomen. Damit unter dem Strich ein Molekül dieses Zuckers synthetisiert wird, muss der Zyklus dreimal ablaufen, so dass drei CO2-Moleküle fixiert werden. Der Calvin-Zyklus ist in drei Phasen zerlegt: Phase 1: Kohlenstoff-Fixierung. Der Calvin-Zyklus nimmt ein CO2-Molekül auf, indem er es an das Ribulose-1,5-bisphosphat (RuBP) anlagert, einen Zucker mit fünf Kohlenstoffatomen. Das Enzym, das diesen ersten Schritt katalysiert, heisst RuBP-Carboxylase oder kurz Rubisco. Das Produkt der Reaktion, ein Molekül mit sechs Kohlenstoffatomen, ist so instabil, dass es sofort zu zwei Molekülen des 3-Phosphoglycerats zerfällt. Phase 2: Reduktion. An jedes Molekül des 3-Phosphoglycerats wird eine weitere Phosphatgruppe angehängt. Ein Enzym überträgt das Phosphat vom ATP, und als Produkt entsteht 1,3-Bisphosphoglycerat. Im nächsten Schritt reduziert ein vom NADPH beigesteuertes Elektronenpaar das 1,3-Bisphosphoglycerat zu G3P. Am Anfang des Zyklus standen 15 Kohlenstoffatome in Form dreier Moleküle der Pentose RuBP, und am Ende sind es 18 Kohlenstoffatome in Form von sechs Molekülen der Triose G3P. Eines davon verlässt den Zyklus und kann von der Pflanzenzelle anderweitig verwendet werden, aber die übrigen fünf dienen zur Regeneration der drei RuBP-Moleküle. Phase 3: Regeneration des CO2-Akzeptors (RuBP). In den letzten Schritten des CalvinZyklus werden die Kohlenstoffgerüste der fünf G3P-Moleküle in einer komplizierten Reaktionsfolge so umgeordnet, dass wieder drei Moleküle RuBP entstehen. Dazu verbraucht der Zyklus nochmals drei ATP-Moleküle. Anschliessend kann das RuBP erneut CO2 aufnehmen, und der Zyklus läuft ein weiteres Mal ab. Um insgesamt ein Molekül G3P zu erzeugen, verbraucht der Zyklus neun ATP- und sechs NADPH-Moleküle. Das G3P, welches den Calvin-Zyklus verlässt, dient als Ausgangssubstanz für Stoffwechselwege, auf denen andere organische Verbindungen synthetisiert werden, darunter auch Glucose und weitere Kohlenhydrate. Weder die Lichtreaktionen noch der Calvin-Zyklus können für sich allein Zucker aus CO2 herstellen. Die Fähigkeit zur Photosynthese erwächst aus dem Gesamtgeschehen im Chloroplasten, das beide Prozesse in sich vereinigt. 10.7 In heissen und trockenen Lebensräumen haben sich alternative Mechanismen der Kohlenstoff-Fixierung entwickelt Oft werden in der Biologie Probleme durch einen Kompromiss gelöst. Ein Beispiel ist das Verhältnis zwischen der Photosynthese und der Vermeidung übermässigen Wasserverlustes bei den Pflanzen. Das für die Photosynthese benötigte CO2 gelangt durch die Spaltöffnungen in das Blatt. Diese sind aber auch der wichtigste Weg der Wasserabgabe, denn durch sie verdunstet das Wasser aus den Blättern. Bei heissem, trockenen Wetter schliessen die meisten Pflanzen ihre Spaltöffnungen, um Wasser zu sparen. Aber diese Reaktion vermindert auch die Photosyntheseleistung, weil die verfügbare CO2-Menge sinkt. Diese Bedingungen setzen im Blatt einen offenbar verschwenderischen Prozess in Gang: die Photorespiration. Photorespiration: ein Relikt der Evolution? In den meisten Pflanzen erfolgt die anfängliche Kohlenstoff-Fixierung durch die Rubisco, jenes Enzym des Calvin-Zyklus, das CO2 an Ribulose-1,5-bisphosphat bindet. Solche Pflanzen bezeichnet man als C3-Pflanzen, weil das erste Produkt der Kohlenstoff-Fixierung bei ihnen das 3-Phosphoglycerat ist, eine Verbindung mit drei Kohlenstoffatomen. Bei heissem, trockenem Wetter schliessen die meisten Pflanzen ihre Spaltöffnungen, um Wasser zu sparen. Daraufhin sammelt sich der Sauerstoff aus den Lichtreaktionen im Blatt an. Wenn O2 statt CO2 am aktiven Zentrum der Rubisco gebunden wird, bildet sich das Zwischenprodukt Glycolsäure, die aus dem Chloroplasten transportiert wird und in den Peroxisomen weiter oxidiert. Schliesslich wird in den Mitochondrien ein Molekül CO2 abgespalten. Den Gesamtprozess nennt man Photorespiration oder Lichtatmung, weil er im Licht abläuft, O2 verbraucht und CO2 produziert. Anders als bei der Zellatmung entsteht bei der Lichtatmung kein ATP, und im Gegensatz zur Photosynthese erzeugt sie auch keine Nährstoffe. Im Gegenteil: Die Photorespiration vermindert die Photosyntheseleistung, weil sie organisches Material aus dem Calvin-Zyklus abzieht. Ob die Photorespiration den Pflanzen in irgend einer Form nützt, ist noch nicht ganz klar. Eine Reihe von Pflanzenarten haben Alternativmethoden der Kohlenstoff-Fixierung, um sich an extreme Bedingungen anzupassen. Die beiden wichtigsten derartigen Modifikationen finden sich bei den C4-Pflanzen und den CAM-Pflanzen. C4-Pflanzen Die C4-Pflanzen heissen so, weil ihrem Calvin-Zyklus eine andere Art der KohlenstoffFixierung vorgeschaltet ist, bei der als erstes Produkt eine Verbindung mit vier Kohlenstoffatomen entsteht. Landwirtschaftlich wichtige C4-Pflanzen sind unter anderem Zuckerrohr und Mais, die zu den Gräsern gehören. Der Mechanismus des C4-Dicarbonsäurewegs ist an einen besonderen anatomischen Aufbau der Blätter gebunden. C4-Pflanzen besitzen zwei Typen photosynthetisch aktiver Zellen: die Bündelscheidenzellen der Leitbündel und die Mesophyllzellen. Die Bündelscheidenzellen sind als dichter Kranz um die Leitbündel angeordnet. Zwischen diesen Leitbündelscheide und der Blattepidermis (äusseres Abschlussgewebe) liegen die lockerer gepackten Mesophyllzellen. Um die Photorespiration zu verhindern wird die Reaktion von dem Enzym PEP-Carboxylase katalysiert, das im Vergleich zu Rubisco eine wesentlich höhere Affinität zum CO2 hat. Deshalb kann die PEP-Carboxylase den Kohlenstoff auch dann sehr wirksam fixieren, wenn Rubisco dazu nicht in der Lage ist – das heisst, wenn bei Hitze der CO2-Partialdruck im Blatt sinkt und der O2-Partialdruck steigt. Nachdem die C4-Pflanze das Co2 fixiert hat, leiten die Mesophyllzellen ihre Produkte – Dicarbonsäure-Anionen mit vier Kohlenstoffatomen – über die Plasmodesmen an die Bündelscheidenzellen weiter. In diesen Zellen setzen die C4Dicarbonsäuren dann wieder CO2 frei, das von der Rubisco über den Calvin-Zyklus in organischen Verbindungen fixiert wird. CAM-Pflanzen Diese Pflanzen öffnen die Spaltöffnungen nachts und schliessen sie am Tage, genau umgekehrt wie die anderen Pflanzen. Wenn sie die Spaltöffnungen tagsüber geschlossen halten, vermindert sich der Wasserverlust, aber gleichzeitig kann auch kein CO2 in die Blätter gelangen. Nachts nehmen die Pflanzen durch die geöffneten Spaltöffnungen CO2 auf und bauen es in verschiedene Carbonsäuren ein, hauptsächlich in Malat. Diesen Weg der Kohlenstoff-Fixierung nennt man diurnalen Säurerhythmus oder, weil man ihn bei Sukkulenten aus der Familie der Crassulaceen entdeckt hat, Crassulaceen-Säurestoffwechsel (Crassulacean acid metabolism, CAM). Die Mesophyllzellen solcher CAM-Pflanzen speichern die Carbonsäuren, die sie nachts produzieren, bis zum Morgen in grossen Vakuolen und schliessen dann ihre Spaltöffnungen. Tagsüber, wenn die Lichtreaktionen ATP und NADPH für den Calvin-Zyklus liefern, wird das CO2 aus den in der Nacht zuvor gebildeten Carbonsäuren in denselben Zellen wieder freigesetzt, so dass es in den Chloroplasten in Zuckermoleküle eingebaut werden kann. Der CAM-Reaktionsweg ähnelt dem der C4-Pflanzen: In beiden Fällen wird das Kohlendioxid zunächst in organische Zwischenprodukte eingebaut und gelangt erst dann in den Calvin-Zyklus. Der Unterschied besteht darin, dass die ersten Schritte der KohlenstoffFixierung bei den C4-Pflanzen räumlich vom Calvin-Zyklus getrennt sind, während bei CAM-Pflanzen beide Prozesse zu unterschiedlichen Zeiten ablaufen. 10.8 Die Photosynthese ist die Stoffwechselgrundlage der Biosphäre: Eine Übersicht Wie sieht das weitere Schicksal der Photosyntheseprodukte aus? Der in den Chloroplasten erzeugte Zucker wird enzymatisch weiter zu Saccharose (Transportform) und Stärke (Speicherform) aufgebaut und versorgt die gesamte Pflanze mit chemischer Energie und Kohlenstoffgerüsten für die Synthese aller organischen Verbindungen durch die Zellen. Etwa die Hälfte des organischen Materials, das durch die Photosynthese entsteht, wird in den Mitochondrien der Pflanzen als Betriebsstoff für die Zellatmung verbraucht. Ein Teil der Produkte geht oft durch die Photorespiration verloren. Genaugenommen sind die grünen, chlorophyllhaltigen Zellen die einzigen autotrophen Teile der Pflanzen. Alle übrigen Teile sind auf die organischen Moleküle angewiesen, die über die Leitbündel aus den Blättern abtransportiert werden. Überschüssiges organisches Material wird in Form von Stärke, Ölen und Proteinen in Blättern, Wurzeln, Knollen, Samen und Früchten gespeichert. Einen grossen Teil dieses organischen Materials verbrauchen die heterotrophen Lebewesen.