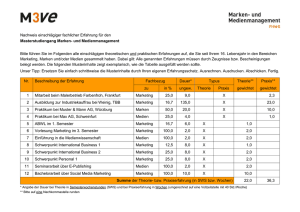
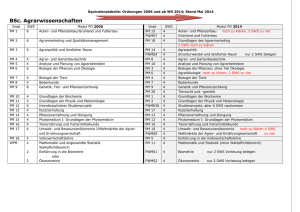
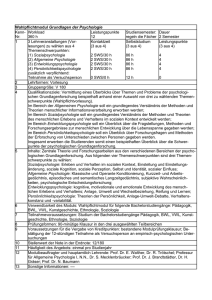
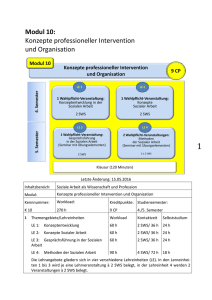
Vorlesung 4
Werbung

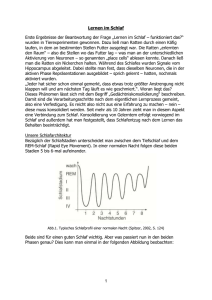
Physiologie 4 Schlaf Zirkadiane Periodik als Grundlage des Wach-Schlaf-Rhythmus Zirkadiane Uhren Die Abfolge der verschiedenen Schlafstadien und des Wachens wird von inneren Uhren gesteuert, die meist eine zirkadiane Periodik besitzen und durch Zeitgeber auf den 24 h-Rhythmus der Außenwelt synchronisiert werden Innere Uhr. Eine freilaufende zirkadiane Periodik bleibt bei Isolation von der Außenwelt über Monate erhalten. Meist ist sie in Isolation länger als 24 h, bei manchen Menschen auch kürzer. Innere Uhren gibt es aber nicht nur für Wachen und Schlafen, sondern auch für viele andere Körperfunktionen. Diese Uhren sind meist untereinander synchronisiert. Ohne äußere Zeitgeber kann es aber auch zur Entkoppelung kommen. Der zirkadiane Rhythmus von Schlafen und Wachen und viele damit einhergehende Rhythmen physiologischer und psychologischer Funktionen werden von endogenen Oszillatoren (inneren Uhren) im Zentralnervensystem (ZNS) gesteuert. Diese inneren Uhren bestehen aus Neuronen, deren Membranstruktur die Membranleitfähigkeit rhythmisch verändert und damit ihre Entladungsraten rhythmisch anordnet. Endogene Oszillatoren I Der Grundrhythmus der endogenen Oszillatoren, von molekularen Uhren gesteuert, wird von äußeren (externen) und inneren (internen) Reizen, die Zeitgeber genannt werden, auf die 24 h-Periodik der Außenwelt synchronisiert. Beim Menschen wirkt helles Licht (700012000 Lux) als stärkster Zeitgeber, aber auch soziale Interaktion hat einen gewissen Einfluss auf die Tag-NachtRhythmik Im Extremfall kann der Wach-Schlaf-Rhythmus in der Isolation besonders lange Werte annehmen zB. 48 hPerioden, also bizirkadiane Rhythmen, dann werden die vegetativen Funktionen völlig abgekoppelt (interne Desynchronisation) und laufen mit der ursprünglichen Periodendauer von etwa 24 h weiter. Die offenbar weniger flexible »Temperaturuhr« kann der neuen, extrem langen Periode der »Aktivitätsuhr« nicht mehr folgen und löst sich daher vom Wach-Schlaf-Rhythmus. Endogene Oszillatoren II Jet-Iag und Schichtarbeit. Wird die zirkadiane Periodik einmalig in ihrem Rhythmus verschoben, z.B. verkürzt durch Flug nach Osten oder verlängert durch Flug nach Westen, so brauchen die zirkadianen Systeme etwa 1 Tag pro 1 h-Zeitzone, um ihre normale Phasenlage zu den äußeren Zeitgebern zurückzugewinnen. Die Resynchronisation erfolgt bei Flügen nach Westen deutlich schneller als bei Flügen nach Osten.Die soziale und berufliche Aktivität lässt sich dem verschobenen Zeitgeber schnell anpassen, Körpertemperatur und andere vegetative Funktionen folgen langsamer. Diese Dissoziation trägt zur vorübergehenden Leistungsminderung nach Langstreckenflügen bei (bezüglich der eventuellen Wirkung des Melatonin bei der Resynchronisation). Nucleus suprachiasmaticus (SCN) I Der zentrale, aber nicht der einzige Schlaf-Wach-Oszillator ist der Nucleus suprachiasmaticus (SCN) Der (SCN) ist im anterioren Hypothalamus direkt über dem Chiasma opticum lokalisiert und stellt die oberste Steuereinheit des zirkadianen Systems dar (master clock). Vom retinohypothalamischen Trakt (RHT) erhält der SCN Information über die Lichtverhältnisse der Umgebung. Der RHT widerum erhält die Licht-DunkelInformation aus spezialisierten bipolaren Ganglienzellen der Retina. Diese enthalten den zirkadianen Photorezeptor Melanopsin, der auf diffuses Licht anspricht und diese Information dann über glutamaterge Synapsen zunächst auf den RHT und dann weiter auf die Zellen des SCN überträgt. Läsionen des SNC führen zu völliger Arhythmizität vieler Körper- und Verhaltensfunktionen. Transplantierung des SCN auf SCN-läsionierte Tiere erzeugt den zirkadianen Rhythmus des Spendertieres im Empfänger. Auch isolierte Zellen des SCN behalten ihre Rhythmizität bei. SCN II »Sklaven-Oszillatoren«. Der SCN synchronisiert als Schrittmacher efferent verschiedene »SklavenOszillatoren« im Gehirn und der Körperperipherie. Dies geschieht über rhythmische Entladung und rhythmische Sekretion von aktivitätshemmenden und aktivierenden Faktoren, meistens Neuropeptiden. Sowohl die Transmittergesteuerte elektrische als auch die sekretorische Rhythmizität wirken auf die subparaventrikuläre Zone des Hypothalamus (SPVC) und von dort über spezifische Projektionen auf die relevanten Empfängerregionen (z. B. die Schlafanstoßenden Zellen im basalen Vorderhirn, die monoaminergen Zellen des Stammhirns und die REM-anstoßenden cholinergen Regionen). Molekulare Uhren I Die rhythmische Transkription von »Uhr-Genen« ist für die endogenen Rhythmen verantwortlich PER und TIM. (Der Glutamat-induzierte Ca2+-Einstrom in die Zellen des SCN führt während der Lichtphase zum Abbau (Degradierung) eines Proteins PER (von period) im Zytoplasma der SCN-Zellen, wodurch im Laufe des Tages die Transkription von PER durch per-Gene im Nukleus ermöglicht wird. Wenn es dunkel wird, hört die Degradierung von PER-Proteinen im Zytoplasma auf und PER kann wieder akkumulieren. Gegen morgen dringen PER-Proteine in den Nukleus ein und hemmen dort die Transkription der per-Gene in einem negativen Rückmeldekreis. PER allein ist inaktiv und kann nicht in den Zellkern eindringen. Es muss sich daher mit dem Protein TIM (von timeless) zu einem sogenannten Dimer verbinden. Dieses Dimer blockiert die Transkription eines anderen Gens, des sogenannten Clock-Gens (von Circadian Locomotor Output Cycles Kaput). Das Clock-Protein löst die Transkription von per- und tim-Genen im Kern aus. Mutationen auf per, tim oder clock zerstören die zirkadiane Periodik, einige der Mutationen sind letal, andere erhöhen die Krebsinzidenz und reduzieren die Immunkompetenz. Durch eine PER-Mutation werden Gene, welche das unkontrollierte Zellwachstum von Zellen zu Krebs fördern, angeregt. Molekulare Uhren II Frühe Reaktionsgene. Die molekularen Mechanismen der Synchronisation der Zellen des Nucleus suprachiasmaticus werden durch unmittelbare Expression »früher Reaktionsgene« (immediate early Genes) gesteuert. Die frühen Reaktionsgene werden durch Licht aktiviert; bereits nach wenigen Minuten lässt sich in den Zellen des Nucleus suprachiasmaticus die Aktivierung eines c-fos Proto-Onkogens feststellen. Das c-fos-Protein ist ein Transkriptionsfaktor in den frühen Reaktionssystemen, die rasch in die Regulation von Zellproliferation und Membrandifferenzierung eingreifen. Die schnelle Expression des Transkriptionsfaktors wird durch Anstieg der cAMP, oder der Ca2+-Konzentration nach Eintreffen des Nervenimpulses ausgelöst, welche die aktivierende Phosphorylierung des Transkriptionsfaktors bewirken. Zirkadiane Rhythmik Die regelmäßige Abfolge von Wachen und Schlafen entspricht ungefähr (zirka) der Dauer eines Tages (lat. dies) und wird von endogenen Oszillatoren (inneren Uhren) autonom gesteuert. Die Rhythmen werden von molekularen Mechanismen v.a. in Zellen des Nucleus suprachiasmaticus generiert und von äußeren und inneren Reizen (Zeitgebern) auf die 24 h-Periodik synchronisiert. Die Rhythmizität der Zellen im Nucleus suprachiasmaticus wird von molekularen Uhren unter Beteiligung weniger Gene im gesamten Reich des Lebendigen in vergleichbarer Art und Weise gesteuert. Dabei kommt es zur rhythmischen Transkription von bestimmten »UhrGenen«. Die Zeitverzögerung im Auf- und Abbau dieser Gene und ihrer Proteinprodukte bestimmen den Rhythmus der Erregbarkeit der Zellmembran von endogenen Oszillatoren. Wach-Schlaf-Verhalten des Menschen Schlafstadien Mit dem Elektroenzephalogramm (EEG) lassen sich die verschiedenen Stadien des Schlafes (REM-, NREM-Schlaf) unterscheiden Mit der Elektroenzephalographie steht eine Methode zur Verfügung, die es erlaubt, den Schlafverlauf fortlaufend aufzuzeichnen, ohne ihn zu stören Den Tiefschlaf bezeichnet man auch als Langsamen-WellenSchlaf (Slow-Wave Sleep, SWS), da er von hochamplitudigen (>1oo mV) Theta- (4-7 Hz) und GammaWellen (0,5-3 Hz) dominiert wird. Dem REM-Schlaf werden alle übrigen Schlafstadien als NREM-Schlaf (Nicht-REM-Schlaf) gegenübergestellt. REM-Schlaf Der REM-Schlaf wird auch als paradoxer Schlaf 'bezeichnet, weil das EEG sich kaum vom Wachzustand unterscheidet, die Person aber regungslos mit geschlossenen Augen liegen bleibt. Es treten dabei sekundenlange Gruppen von 1-4 Hz schnellen Augenbewegungen auf. Im EEG herrschen Beta-Wellen (13-30 Hz), Gamma-Wellen (>3oHz) und eingestreute, kleinamplitudige Theta-Wellen (4-7 Hz) vor. In dieser Zeit wird häufig aktiv-handelnd und emotional geträumt, während in den übrigen Schlafphasen eher abstrakt-gedanklich geträumt wird. Orthographie des Schlafes. Während des Schlafens treten im EEG auch für den Schlaf typische Muster auf. Dazu gehören die Schlafspindeln und K-Komplexe. K-Komplexe geben einen Hinweis darauf, dass das schlafende Gehirn Reize aus der Umwelt wahrnimmt und darauf reagiert. Dieses Wellenmuster tritt nämlich regelmäßig dann auf, wenn dem Schläfer ein Reiz präsentiert wird, z. B. ein Tonsignal. Schlafspindeln sind ebenfalls kurzdauernde Wellenmuster, vor allem der motorischen Areale, welche von hemmenden Interneuronen im somatomotorischen Thalamus erzeugt werden. Es gibt Hinweise darauf, dass Schlafspindeln den Schlaf schützen, indem sie das Gehirn gegen Außenreize abschirmen und die Ruhigstellung der zentralen Motorik ermöglichen. Schlafphasen eines Schlafzyklus I Die Schlafphasen werden unter physiologischen Bedingungen immer in derselben Abfolge von Langsamen-Wellen-Schlaf (SWS) zum REM-Schlaf durchschritten Beim Übergang vom entspannten Wachsein (mit geschlossenen Augen) in das Schlafstadium 1 (S1) verschwinden die AlphaWellen. Die Klarheit des Bewusstseins wird zunehmend eingeschränkt. Viele Menschen erleben in diesem dösenden Übergangszustand zwischen Wachen und Schlafen optische, traumartige Eindrücke. Gleichzeitig beginnen die Augäpfel sich ganz langsam hin- und herzubewegen. Bei manchen Schläfern zeigen sich beim Einschlafen auch feine Zuckungen der Augenlider. Es können aber auch heftige Zuckungen einzelner Gliedmaßen oder des ganzen Körpers auftreten, die wahrscheinlich durch eine Umstellung der motorischen Kontrollsysteme beim Einschlafen bedingt sind. Schlafphasen eines Schlafzyklus II Der Beginn des nachfolgenden Schlafstadiums 2 (S2) ist daher als der eigentliche Zeitpunkt für den Schlafbeginn anzusehen, zumal hier zum ersten Mal Schlafspindeln und K-Komplexe auftauchen. Die Zeitdauer zwischen dem Zubettgehen und dem ersten S2-Schlaf, also die Schlaflatenz, beträgt bei gesunden Erwachsenen etwa 10 bis 15 min. Normalerweise vertieft sich der Schlaf sukzessive aus den Stadien S1 und S2 in die Tiefschlafstadien S3 und S4 .Die Weckschwelle für Reize erhöht sich entsprechend und erreicht ihren höchsten Wert nach etwa einer Stunde. Anschließend nimmt die Weckschwelle wieder ab. Schließlich geht der Tiefschlaf in den ersten REM-Schlaf‘ über, mit dem der komplette erste Schlafzyklus abgeschlossen wird. Schlafzyklen im Verlauf der Nacht Im Verlaufe einer Nacht werden die einzelnen Schlafstadien mehrfach durchlaufen; das Maximum des Tiefschlafs liegt dabei im ersten Schlafzyklus; die REM-Episoden nehmen im Verlauf der Nacht an Dauer zu Schlafzyklen. Eine Nacht besteht aus etwa 4-5 Schlafzyklen, die jeweils eine Dauer von etwa 1,5 Stunden haben. Ein kompletter NREM-REM-Zyklus wird als Basic-Rest-Activity-Cycle (BRAC) bezeichnet, da er sich in den wachen Teil des Tages hinein fortzusetzen scheint. Das Maximum des langsamwelligen Schlafs mit den Stadien 3 und 4 liegt im ersten Schlafzyklus, danach nimmt der langsamwellige Schlaf stetig über die Nacht ab. Die Dauer der REM-Phasen nimmt im Laufe der Nacht von ca. 5-10 min bis auf 20-30 min zu. Auch die Augenbewegungsdichte im REM-Schlaf nimmt im Laufe der Nacht zu. Diese Intensivierung des REM-Schlafes gilt auch für viele andere physiologische Prozesse. Mit der Intensivierung des REM-Schlafes geht auch verlängertes und intensiveres Träumen einher. Unterschiede zwischen REM-Schlaf und Wachzustand Physiologisch und psychologisch weisen die REM-Phasen Ähnlichkeit zum Wachzustand auf. Trotzdem bestehen Unterschiede, die auch das psychologisch kaum mit Wachen vergleichbare Träumen erklären. Der zentrale Unterschied besteht in der tonischen Hemmung der spinalen Motoneurone während der REM-Phasen, was zu vollständiger Paralyse der quergestreiften Muskulatur führt. Die spinale Hemmung geht von Kernen in der medialen Medulla oblongata aus und diese benutzen Azetylcholin als Transmitter. Nach Läsion dieser medullären Kerne tritt bei Säugetieren und Menschen REMSchlaf ohne Atonie auf, die Tiere bzw. Menschen agieren motorisch entsprechend dem Trauminhalt (z. B. »fängt« die Katze eine nicht existierende Maus). Ein wesentlicher Unterschied zwischen REM- und Wachzustand ist die Überaktivität cholinerger Synapsen im REM-Schlaf und die veränderte Topographie aktivierter Kortex-Areale Altern und Schlaf I Die Gesamtschlafzeit sinkt im Lauf des Lebens ab, der relative Anteil des SWS-Schlafs (»Tiefschlaf«) wird außerdem erheblich kürzer Altersentwicklung. Die relativen Anteile von Wachen und Schlafen, ebenso wie die Anteile von REM- und NREMSchlaf an der Gesamtschlafzeit machen eine charakteristische Altersentwicklung durch. Insgesamt sinkt im Laufe des Lebens nicht nur die Gesamtschlafzeit ab, sondern es wird auch der relative Anteil des SWSSchlafs (Slow Wave Sleep, Stadium 3 und 4) erheblich kürzer. Das Neugeborene verbringt einen erheblichen Teil des Tages im REM-Schlaf. Dieser Anteil sinkt dann rasch mit der Hirnentwicklung bis um das 14. Lebensjahr von 50% auf ca. 20% ab und bleibt danach konstant. Stadium 1 und 2 nehmen dagegen ab dem 14. Lebensjahr zu, während Stadium 3 und 4 im Erwachsenenalter kontinuierlich abnehmen. Altern und Schlaf II REM-Schlaf als Umweltreiz. Der hohe Anteil des REM-Schlafs bei Säuglingen und Kleinkindern hat zu der Vermutung geführt, dass diese Perioden erhöhter neuronaler Aktivität (desynchronisiertes EEG ähnlich dem bei Aufmerksamkeit) für die ontogenetische Entwicklung des ZNS wichtig sind, da bei diesen Individuen äußere Reize noch weitgehend fehlen: das »Träumen« ersetzt als innere Reizung den mangelnden externen Einstrom. Dagegen spricht allerdings, dass bei Vorschulkindern Traumberichte nach Aufwecken praktisch nicht vorkommen. Es scheint also eher die allgemeine Aktivitätsentwicklung der Hirnrinde im REM-Schlaf für die Hirnentwicklung wichtig zu sein. Schlafstörungen I Primäre Schlafstörungen können als Ein- und Durchschlafstörungen, und als schlafstadiengebunde Störungen auftreten.Schlafstörungen, die nicht als Folge von organischen Erkrankungen auftreten, werden als primäre bezeichnet. Ein- und Durchschlafstörungen (Insomnia): Ideopathische Insomnia bezeichnet subjektiv erlebte und objektiv, d. h. mit polygrafischen Aufzeichnungen im Schlaflabor verifizierbaren Störungen im Schlafprofil. Diese können zahlreiche Ursachen haben, z.B. zu viel oder zu wenig körperliche Aktivität, chronischer Stress, Reisen und exzessives Essen oder Fasten. Pseudoinsomnia äußert sich durch subjektive Störungen des Ein- und Durchschlafens, wobei das Schlafprofil aber altersgerecht ist. Pseudoinsomnia liegt vor, wenn die subjektiven Erwartungen an die Schlafgüte nicht mit dem objektiv vorhandenen Schlafprofil übereinstimmen. Dies ist häufig bei alten Menschen der Fall, die sich nicht an die zunehmende »Leichtigkeit« des Schlafes gewöhnen können. Schlafmittelmissbrauch ist eine der häufigsten Ursachen für Schlafstörungen. Alle bekannten Schlafmittel führen bei längerer Einnahme zu einer Veränderung des natürlichen Schlafprofils und bei Absetzen der Einnahme zu erheblichen Schlafstörungen. Schlafmittelmissbrauch ist die häufigste iatrogene Erkrankung (von Medizinern verursachte Krankheit). Schlafstörungen II Hypersomnia. Der Prototyp einer hypersomnischen Erkrankung ist die Narkolepsie. Ihr Leitsymptom ist die gesteigerte Tagesmüdigkeit mit unkontrollierbaren Schlafattacken (Dauer von wenigen Sekunden bis 30 min). Zur Narkolepsie gehören auch die Kataplexie, d. h. ein meist durch affektive Reize ausgelöster Tonusverlust, sowie Schlaflähmungen und hypnagoge Halluzinationen. Diese Symptome können als das »Eindringen« von REM-Episoden in den Wachzustand aufgefasst werden, denn Kataplexie und Schlaflähmung sind mit der Atonie des REMSchlafes eng verwandt, hypnagoge Halluzinationen mit den traumgenerierenden Prozessen dieses Schlafzustandes. Narkolepsie und REM-Schlaf.( Der Azetylcholinspiegel im Hirnstamm ist dauerhaft wie im REM-Schlaf stark erhöht. Die Tiere weisen eine Mutation am »Carnac«-Gen auf, welches den Rezeptor für das Neuropeptid Orexin bildet. (Orexin-knock-out Mäuse sind narkoleptisch und zeigen profunde Störungen der Nahrungsaufnahme, daher der Name. Orexin wirkt vom Hypothalamus auf alle Schlafregulierenden Strukturen des Hirnstamms, besonders auch die REM-anstoßenden cholinergen Systeme.) Eine der häufigsten Hypersomnien, vor allem bei übergewichtigen Rauchern, ist die Schlaf-Apnoe. Schlafstörungen III Schlafstadiengebundene Störungen: Somnambulismus (Schlafwandeln) ist ein motorischer Automatismus, der beim Übergang vom Tiefschlafstadium 4 in das Stadium 2 auftritt, und zwar besonders bei Kindern und Jugendlichen, sowie bei Erwachsenen unter Stressbelastung. Die Augen des Schlafwandlers sind weit geöffnet, er ist nicht ansprechbar, nach dem Aufwecken desorientiert und kann sich nicht an Träume erinnern. Bettnässen (Enuresis nocturna) kommt bei rund 10 % aller Kinder nach dem 2. Lebensjahr vor. Es tritt praktisch immer aus dem NREM-Schlaf auf. Der kindliche Pavor nocturnus kann ähnliche Ursachen haben und kommt zwischen dem 3. und 8. Lebensjahr, selten später, vor. Plötzlich, während des Schlafes, setzt sich das Kind auf und fängt an zu schreien. Das Gesicht ist bleich und schweißbedeckt, der Atem geht schwer. Nach kurzer Zeit wacht das Kind auf, erkennt seine Umwelt und schläft oft wieder ein. Neuronale Schlafsteuerung I Der langsame Wellen-Schlaf (SWS) ist homöostatischer Natur und wird durch die Akkumulation von bestimmten »Schlafsubstanzen« ausgelöst Der langsame Wellen-Schlaf (SWS, Tiefschlaf) hat weniger rhythmischen, sondern eher homöostatischen Charakter: Er hängt stark von der vorausgegangenen Aktivität (Müdigkeit), Nahrungsaufnahme, Hirntemperatur und anderen Faktoren ab. Man nimmt an, dass die Akkumulation einer oder mehrerer »Schlafsubstanzen« während des Wach-Seins als Ursache für den Beginn von SWS dient. Eine wichtige Schlafsubstanz ist das Purin Adenosin, das neben motorischen und motivationalen Funktionen auch in neuronalen Schlafstrukturen als Signalmolekül wirkt. Es akkumuliert während des Tages und hemmt vor allem über seine A-Rezeptoren die cholinergen exzitatorischen Neurone des basalen Vorderhirns. Das basale Vorderhirn mit dem Nucleus praeopticus des Hypothalamus ist eine Struktur, deren elektrische Reizung oder Erwärmung zu SWS führt. Jene Teile des basalen Vorderhirns, welche bei Reizung SWS auslösen, sind räumlich klar von den cholinergen, REM-bewirkenden Regionen getrennt. Die SWS-Regionen liegen in der Nachbarschaft zu den Kernen des vorderen Hypothalamus. Neuronale Schlafsteuerung II SWS wird aber offensichtlich auch durch periphere Peptide, wie z.B. Muramyl-Peptide, angestoßen, die in subkortikalen Gliazellen und den Gliazellen des basalen Vorderhirns die Produktion von Interleukin-1 stimulieren. Dabei handelt es sich um ein Peptid, das mit der Immunabwehr befasst ist. Fieber nach Infektionen und der Anstieg der Körper- und Hirntemperatur sind daher potente Reize für SWS. Immunkompetenz. Die restaurativen Prozesse im homöostatischen Non-REMSchlaf finden vor allem in den ersten drei Nachststunden mit einem Maximum an SWS statt. Das Hypothalamus-Nebennieren-Stress-System ist in dieser Zeit gehemmt, die Kortisolproduktion auf einem Minimum und die Produktion immunkompetenter Zellen auf einem Maximum. REM-Schlaf Gleichzeitig mit der Aktivierung der cholinergen REM-Zellen werden die im langsamen-Wellen-NREM-Schlaf aktiven aminergen Zellen in Nucleus raphe (Serotonin) und Locus coeruleus (Noradrenalin) blockiert. Umgekehrt hemmt die Aktivierung von Raphe und Coeruleus die cholinergen Kerne. Im REM-Schlaf wird also ein primär »cholinerges Klima« erzeugt, was sich deutlich vom Wachzustand unterscheidet, in dem auch die Produktionsstätten der aminergen Transmitter aktiv sind. Schlafverhalten Die verschiedenen Schlafstadien lassen sich durch die Registrierung des Elektroenzephalogramms (EEG) und der Augenbewegungen (Elektrookulogramm, EOG) erfassen: Wir unterscheiden vier Stadien zunehmender Schlaftiefe mit zunehmend langsamen Wellen im EEG. Das Tiefschlafstadium (Stadium 4) wird auch Langsamer Wellen-Schlaf genannt, da es im EEG hochamplitudige Wellen zeigt. Der Langsame-Wellen-Schlaf (SWS, »Tiefschlaf«) geht in ein dem Wachzustand vergleichbares Stadium mit schnellen Augenbewegungen (REM) und WachEEG über; diese REM-Perioden werden im Laufe der Nacht länger. Eine Abfolge von Nicht-REM-Schlaf (NREM) und REM-Schlaf wird als Basic-RestActivity-Cycle, BRAC, bezeichnet. Eine Nacht besteht aus 4-5 solcher Schlafzyklen. Die Dauer der einzelnen Schlafstadien ändert sich im Laufe des Lebens: Während Neugeborene und Kleinkinder erhebliche Teile des Tages und der Nacht im REM-Schlaf verbringen, bleibt der REM-Anteil nach der Pubertät konstant. Im späten Erwachsenenalter und im hohen Alter nimmt auch der Anteil des tiefsten SWS kontinuierlich ab. Neuronale Steuerung SWS wird von präoptischen Regionen des Hypothalamus und Teilen des basalen Vorderhirns erzeugt. Die Regulation erfolgt homöostatisch durch Akkumulation von Schlafsubstanzen während der aktiven Zeit. REM wird von cholinergen Kernen des Mittelhirns und basalen Vorderhirns erzeugt und hängt von zirkadianen und ultradianen Oszillatoren ab. Die physiologischen Aufgaben der Schlafstadien Träumen, Mentale Prozesse im Schlaf. Mentale Prozesse sind während der gesamten Schlafzeit vorhanden, in NREM-Pha-sen sind sie eher abstrakt, gedankenartig. Die aktiven, halluzinatorischen, geschichtenartigen Traumphänomene, die wir eigentlich meinen, wenn wir von Träumen reden, sind während der phasischen REM-Aktivitäten (z. B. Augenbewegungen) am stärksten. Sie sind während der ersten Nachthälfte eher Erinnerungen an Ereignisse des vergangenen Tages und werden gegen Morgen zunehmend emotionaler. Traumnetzwerke. Bildgebende Untersuchungen während des Schlafes ergaben, dass bei SWS die Hirndurchblutung drastisch absinkt. Wenn das EEG desynchronisiert und lebendige Träume berichtet werden - was nicht unbedingt, aber oft mit REM-Phasen korreliert - werden die cholinergen Systeme aktiv (lebendiges Erleben), die primären sensorischen und motorischen Projektionsareale gehemmt (Abschluss von Außenwelt), limbische und dienzephale Regionen aktiv (Gefühls- und Trieberlebnisse) und der dorsale Frontalkortex gehemmt (Kontrollverlust, Gedächtniskonsolidierung). Die Assoziationsareale sind je nach Trauminhalt aktiv, wodurch die lebendigen Szenenabfolgen, oft mit Erinnerungen durchmischt, erklärt werden können Kernschlaf Nur ein Teil des Schlafes ist wirklich vital notwendig: Kernschlaf; er umfasst in etwa die ersten 3 Schlafzyklen einer Nacht Trotz allen Fortschritts blieb die Bedeutung der Schlafphasen bis heute offen. Klar ist nur, dass beide (REM und NREM) überlebenswichtig sind. Totale Schlafdeprivation über längere Zeit führt zum Tod bei Mensch und Tier. Beim Menschen sind die ersten 2-3 SWS-REMPhasen offensichtlich essenziell, sie werden daher Kernschlaf genannt. Eine Deprivation der letzten 3 Schlafstunden führt kaum zu merkbaren Störungen (Optional- oder Füllschlaf). Die psychischen und gesundheitlichen Auswirkungen auch langer Schlaflosigkeit (z. B. 10 Tage und Nächte) beim erwachsenen Menschen sind allerdings relativ gering. Nach 3-4 Nächten treten bei einigen Personen Wahrnehmungsverzerrungen und ein leichtes Nachlassen von Vigilanz (Daueraufmerksamkeit) auf. Nach nur wenigen Stunden Erholungsschlaf tritt völlige Erholung ein. Bereits nach wenigen Nächten »holen« sich die Versuchspersonen durch extrem kurze, aber zunehmend häufiger werdende Mikroschlafepisoden »ihren« Schlaf. Die Aufgaben des Tiefschlafs (SWS) I Der homöostatische SWS hängt mit restaurativen Funktionen zusammen Stoffwechselenergie. Nach Schlafdeprivation wird zuerst SWS nachgeholt, was für die Energie konservierende Funktion von SWS spricht. Adenosin ist ein wichtiger Vorläufer für ATP und kommt häufig als hemmender Neuromodulator im ZNS vor. Während des Tages und bei Anstrengung oder Schlaflosigkeit steigt die Konzentration von Adenosin im Extrazellulärraum kontinuierlich an, vor allem in den SWS-anstoßenden HirnStrukturen. Koffein u.a. Weckmittel blockieren die Ax und A2A Adenosinrezeptoren. Endokrinologie. Während der SWS-Phasen zu Beginn des Schlafes wird vor allem bei Körperwachstum das Wachstumshormon (GH, Growth Hormon) ausgeschüttet und die Ausschüttung der Stresshormone Kortisol und ACTH gehemmt. Extremer Stress führt zu Schlafstörungen und zu Wachstumsstörungen bei Kindern bis hin zu psychosozialem Zwergwuchs. Da GH auch am Wachstum und der Verbindung von Nervenzellen beteiligt ist, werden auch die kognitive Entwicklung und die Lernfähigkeit durch Stress und SWS-Mangel gestört. Bei der Depression ist ebenfalls der zirkadiane Gipfel abgeflacht und der relative Anteil von Kortisol erhöht. Dabei ist der REM-Schlaf, vor allem die REM-Latenz (Zeit bis zur ersten REMPhase) verkürzt. Die Aufgaben des Tiefschlafs (SWS) II Immunologie. Stress und Kortisolanstieg hemmen die Immunabwehr. Daher geht ein SWS-Mangel auch mit Störungen des Immunsystems einher. Der Verlust von SWS im Alter trägt zum vermehrten Auftreten von Krankheiten bei, welche von Immunfaktoren »in Schach« gehalten wurden. Der Schlaf-Wach-Rhythmus wird von immunaktiven Substanzen ebenso beeinflusst wie umgekehrt der Schlaf zum restaurativen Aufbau von immunkompetenten Zellen notwendig ist. Interleukine, z.B. 11-1, die von T-Helferzellen abgegeben werden und das Lymphozytenwachstum beschleunigen, haben schlafanstoßende Wirkung im Gehirn. Chronische Schlafdeprivation im Tierversuch führt umgekehrt zu raschem Absinken der Immunkompetenz mit Anstieg von Neoplasien (krebsartiger Entartung), Infektionen und Tod des Tieres. Zirkadiane Rhythmusstörungen wie Nachtarbeit und Zeitzonen überfliegen (»JetIag«) erhöhen ebenfalls die Infektionsanfälligkeit. Die Auswirkungen des Schlafens auf das Immunsystem scheinen u.a. von der zirkadianen Rhythmik des Zirbeldrüsenhormons Melatonin bewirkt zu werden. Melatonin ist während des Schlafes erhöht, seine Konzentration im Kindesalter ist hoch und sinkt mit der Dauer des Tiefschlafes im Alter ab. Extern vor dem Einschlafen verabreicht, reduziert es Belastungseffekte (»Stress«) und kann anscheinend bei Jet-Iag den Rhythmus resynchronisieren. Melatonin bewirkt in Antigen-aktivierten THelferzellen die Ausschüttung kleiner Mengen endogener Opioide. Die Aufgaben des REM-Schlafs I Der Anteil des REM-Schlafs pro Schlafzyklus hängt mit der Nahrungsaufnahme und der Gedächtniskonsolidierung zusammen REM-Schlaf und Nahrungsaufnahme. REM-Schlaf weist eine enge Beziehung zur Nahrungsaufnahme auf: Übergewicht geht mit erhöhtem REM-Anteil einher, Patienten mit Magersucht (Anorexie) erhöhen REM-Schlaf, wenn sie ihr Gewicht normalisieren. Personen, die an Narkolepsie leiden, haben erhöhtes Körpergewicht. Extremes Fasten und Hungern geht mit REM-Unterdrückung einher. Dies wird als evolutionärer Mechanismus zur Maximierung von Wachzeiten interpretiert, um Futtersuche zu ermöglichen. REM signalisiert möglicherweise den hypothalamischen Esszentren die Energiebalance.Diese Veränderungen hängen mit dem Orexin-System zusammen. Das Orexin-System des lateralen Hypothalamus erhöht seine Aktivität während Wachheit und bei Hunger. Die Aufgaben des REM-Schlafs II REM-Schlaf und Gedächtnis. Schlaf fördert die Fähigkeit zur Einprägung und Wiedergabe von gelerntem Material, dies gilt für beide Schlaftypen SWS und REM. Für die REMGedächtnis-Beziehung sprechen Untersuchungen zur RNA- und DNA-Synthese im Gehirn während des Schlafes. DNA-Synthese in der Entwicklung wird durch REMDeprivation reduziert. Schlafentzug im Säuglings- und Kindesalter wirkt besonders destruktiv auf kognitive Funktionen, Körper- und Gehirnwachstum. Ein weiteres korrelatives Indiz für die Rolle von REM-Schlaf im Konsolidierungsprozess ist die Gegenwart von Hippokampus-Theta-Rhythmus während des Übergangs von Kurzzeit- ins Langzeitgedächtnis. Physiologische Aufgaben des Schlafs Sowohl SWS (Slow WaveSleep) als auch REM-Schlaf sind zum Überleben notwendig. 2-3 SWS-REM-Phasen sind für den Menschen essenziell, sie werden daher als »Kernschlaf« bezeichnet. SWS wird nach Schlafdeprivation als erstes nachgeholt, dürfte also für die körperinternen Homöostasen (Hirntemperatur?) Vorrang haben. REM-Schlaf könnte mit Gedächtnisspeicherung und damit Wachstum und Aktivitätsniveau plastischer Synapsen zusammenhängen. Nahrungssuche und REM-Schlaf sind eng korreliert, und das Neuropeptid Orexin des Hypothalamus scheint REM-Schlaf und Nahrungssuche zu regeln.