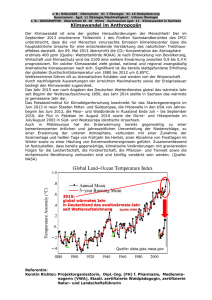
klimawandel und europäische fischerei
Werbung

STUDIE Fachreferat Strukturpolitik und Kohäsion KLIMAWANDEL UND EUROPÄISCHE FISCHEREI FISCHEREI 2007 DE Generaldirektion Interne Politikbereich der Union Fachabteilung Struktur- und Kohäsionspolitik FISCHEREI KLIMAWANDEL UND EUROPÄISCHE FISCHEREI STUDIE IP/B/PECH/IC/2006-199 PE 379.208 24/08/2007 DE Die vorliegende Studie wurde vom Fischereiausschuss des Europäischen Parlaments in Auftrag gegeben. Die vollständige Studie wird in folgenden Sprachen veröffentlicht: - Original: EN; - Übersetzungen: DE, FR, ES, IT. Verfassers: BIPRO GmbH and IFM - GEOMAR Clemmesen, Catriona (IFM-GEOMAR) Potrykus, Alexander (BiPRO GmbH) Schmidt, Jörn (IFM-GEOMAR) Zuständige Beamtin: Eva Casalprim - Calvés Policy Department Structural and Cohesion Policies European Parliament Rue Wiertz 60 B-1047 Brussels E-mail: [email protected] Manuskript abgeschlossen im August 2007. Die Studie ist erhältlich unter folgendem Link: http://www.europarl.europa.eu/activities/expert/eStudies.do?language=EN Brüssel, Europäisches Parlament 2007. Die hier vertretenen Auffassungen geben die Meinung des Verfassers wieder und entsprechen nicht unbedingt dem Standpunkt des Europäischen Parlaments. Nachdruck und Übersetzung - außer zu kommerziellen Zwecken - mit Quellenangabe gestattet, sofern der Herausgeber vorab unterrichtet und ihm ein Exemplar übermittelt wird. Generaldirektion Interne Politikbereich der Union Fachabteilung Struktur- und Kohäsionspolitik FISCHEREI KLIMAWANDEL UND EUROPÄISCHE FISCHEREI STUDIE Inhalt: Der Klimawandel dürfte verschiedene Prozesse auslösen, die sich auf die biologische Produktivität der Meeresökosysteme und die Verbreitung der Meeresressourcen auswirken. In der vorliegenden Studie soll analysiert werden, welche Auswirkungen und Folgen der Klimawandel möglicherweise für die Seefischerei und die Meeresaquakultur der Europäischen Union mit sich bringt. Grundlage der Studie ist eine umfassende Auswertung von etwa 200 relevanten und aktuellen Fundstellen in der Fachliteratur unter besonderer Berücksichtigung der Fischereiressourcen, die für die Fischerei der Europäischen Union im Atlantik und in den angrenzenden Meeren (Mittelmeer, Nord- und Ostsee) von Bedeutung sind. Mit der Fischerei wird ein Sektor von den Klimaveränderungen betroffen, in dem die Ressourcen bereits vollständig genutzt werden. Veränderungen in der Produktivität des Ökosystems werden sich stark auf die Nachhaltigkeit der Fischerei auswirken. Angesichts dieser Lage ist der Klimawandel derzeit Gegenstand sehr umfangreicher Forschungsarbeiten, mit denen abgeschätzt werden soll, wie sich die Fischbestände in der Zukunft entwickeln, und um weitere Fragen mit einem spürbaren Einfluss auf die europäische Fischerei zu bewerten. Es wurden sieben strategische Optionen ermittelt, die in dieser Studie diskutiert werden. IP/B/PECH/IC/2006-199 PE 379.208 DE Klimawandel und europäische Fischerei Zusammenfassung Hintergrund und Zielsetzungen Der Klimawandel dürfte verschiedene Prozesse auslösen, die sich auf die biologische Produktivität der Meeresökosysteme und die Verbreitung der Meeresressourcen auswirken. Mit der Fischerei wird ein Sektor von den Klimaveränderungen betroffen, in dem die Ressourcen bereits vollständig genutzt werden. Veränderungen in der Produktivität des Ökosystems werden sich stark auf die Nachhaltigkeit der Fischerei auswirken. Der Klimawandel ist derzeit Gegenstand sehr umfangreicher Forschungsarbeiten, und die damit verbundenen Erkenntnisse sind von großer Bedeutung für die Einschätzung der künftigen Entwicklungen bei den Fischbeständen. Vor diesem Hintergrund soll in der vorliegenden Studie analysiert werden, welche Auswirkungen und Folgen der Klimawandel möglicherweise für die Seefischerei und die Meeresaquakultur der Europäischen Union mit sich bringt. Grundlage der Studie ist eine umfassende Auswertung der vorhandenen Literatur unter besonderer Berücksichtigung der Fischereiressourcen, die für die Fischerei der Europäischen Union im Atlantik und in den angrenzenden Meeren (Mittelmeer, Nord- und Ostsee) von Bedeutung sind. Die Studie beinhaltet einen konzeptionellen Rahmen mit einer Beschreibung der Schlüsselprozesse, die die Dynamik der Ökosysteme und der Fischereiressourcen bestimmen. Mit dessen Hilfe wurden die Literaturquellen zu den Auswirkungen des Klimawandels auf die für die Fischerei der Europäischen Union wichtigsten Arten systematisch ausgewertet. Auswertung der Fachliteratur Von Januar bis Mai 2007 wurden etwa 200 relevante und aktuelle Fundstellen in der Fachliteratur sowie einschlägige Studien untersucht. Anhand der erzielten Ergebnisse konnten 40 durch den Klimawandel beeinflusste Phänomene ermittelt werden, und es wurden die dazugehörigen Trends und Wirkungen aufgezeigt (siehe Tabelle 1). Jedem dieser Phänomene wurde ausgehend von der Zuverlässigkeit der Literaturquellen und der Häufigkeit ähnlicher Ergebnisse ein bestimmter Zuverlässigkeitsgrad zugeordnet. Schlussfolgerungen und Empfehlungen Treibhausgase tragen zum Klimawandel bei und sind eine Gefahr für die industrielle Fischerei Die in immer größerem Umfang freigesetzten künstlichen Treibhausgase haben einen großen Anteil am Klimawandel und den Veränderungen des Meeresökosystems. Treibhausgase sind eine Gefahr für die bereits geschwächten Fischbestände und sollten daher auf ein Minimum beschränkt werden. Die Wassereigenschaften verändern sich bereits Die Wissenschaftler sind sich darin einig, dass der Klimawandel Realität ist. Die Temperaturen werden ebenso ansteigen wie der Meeresspiegel. Anzeichen für eine Klimaerwärmung sind Veränderungen bei den Oberflächen- und Lufttemperaturen sowie den Temperaturen in den oberen Ozeanschichten bis zu mehreren hundert Meter Tiefe und der Anstieg des iii PE 379.208 Klimawandel und europäische Fischerei Meeresspiegels. Die durchschnittliche globale Temperatur ist während der letzten hundert Jahre um ~0,6 °C angestiegen, der Meeresspiegel um 0,17 m. In diesem Zeitraum haben sich sowohl die Meeres- als auch die Süßwassersysteme erwärmt. Die Prognosen sprechen von einer weiteren Erwärmung des Küstenklimas in Europa im Verlaufe des 21. Jahrhunderts, wobei mit einer Erhöhung der Meeresoberflächentemperatur um 0,2 °C in jedem Jahrzehnt gerechnet wird. Modellschätzungen sagen eine Versauerung der Ozeane voraus( 1). Der pH-Wert an der Meeresoberfläche wird während der nächsten 100 Jahre um 0,3 bis 0,5 Einheiten zurückgehen und in den kommenden 300 Jahren um 0,3 bis 1,4 Einheiten. Die Salinität wird sich verändern. So wird für die nordischen Meere und die Ostsee mit einer Abnahme der Salinität( 2) gerechnet, während sie im Mittelmeer voraussichtlich ansteigen wird. Die atlantische thermohaline Zirkulation( 3) wird schwächer werden. Zwischen der Nordsee und den Gewässern in den arktischen Regionen findet ein Austausch von kalten und warmen Wasserströmungen statt, die durch die atlantische thermohaline Zirkulation bewegt werden. Die Fließintensität wird durch den Klimawandel beeinflusst. Sie dürfte derzeit bereits um 30 % verringert sein. Es ist unwahrscheinlich, dass die Zirkulation einmal gänzlich zum Erliegen kommt, was die derzeitigen Gegebenheiten sehr stark verändern würde (z. B. ein kurzfristiger Anstieg des Meeresspiegels in der Nordsee um 1 m, ein langfristiger Anstieg des globalen Meeresspiegels um 0,5 m, Verschiebung des tropischen Niederschlags( 4)gürtels nach Süden und eine Verringerung der Biomasse im Atlantik um 50 %). Die Schichtung wird sich verstärken( 5). Es wurde nachgewiesen, dass der Klimawandel zu einer verstärkten Schichtung der Ost- und Nordsee und des Mittelmeeres führt. Durch die Schichtung der Wassersäule entstehen natürliche Barrieren, wo es zur Konzentration von Organismen kommen kann oder die von den Organismen passiert werden müssen. Die Schichtung verstärkt sich mit zunehmender Salinität und Temperatur. Eine verstärkte Schichtung wiederum behindert die Vermischung mit Tiefenwasser und führt zu einer geringeren Anreicherung mit Nährstoffen. Durch eine veränderte Zirkulation und Schichtung wird auch die geographische Verbreitung der Organismen verändert. Die Strömungen sind wichtig für den Transport von Organismen wie Plankton( 6) und Fisch über große Entfernungen und können daher zu deren größerer Verbreitung beitragen. Andererseits jedoch wirken die Strömungen auch als biogeographische Barrieren( 7) zwischen den zu beiden Seiten befindlichen Wassermassen. Der Austausch von Organismen über die Strömung hinweg wird dadurch verringert. Durch die Erwärmung könnte es zu einer Abschwächung der Küstenströme kommen, wodurch auch die Verbreitung über diese Ströme geringer wird. Gleichzeitig verschwindet die Barriere zwischen Küsten- und (1) Verringerung des pH-Wertes. Der pH-Wert ist ein Maß für die Stärke der sauren bzw. basischen Wirkung einer Lösung. (2) Die relative Konzentration von Salzen, normalerweise Natriumchlorid, in einem bestimmten Boden oder Gewässer. Auffrischung bezieht sich in diesem Zusammenhang auf eine Verringerung der Salze im Wasser. Ohne Einheit; gelegentlich wird noch die alte Einheit PSU (practical salinity unit) verwendet. (3) Zirkulation im Ozean, die durch temperatur- und salinitätsbedingte Unterschiede in der Meerwasserdichte bewirkt wird. (4) Meteorologie: die auf die Erde fallenden Produkte der Kondensation in der Atmosphäre, wie etwa Regen, Hagel oder Schnee. (5) Das Auftreten von mehr oder weniger horizontalen Wasserschichten im Meer als eine Folge von Unterschieden in der Dichte, d. h. Unterschiede bei Salinität (Halokline) oder Temperatur (Thermokline). (6) Schwebende Organismen, deren Bewegungen mehr oder weniger von den Strömungen abhängig sind. Zwar gibt es beim Zooplankton einige Arten, die sich durch aktive Schwimmbewegungen in einer vertikalen Position halten können, jedoch ist Plankton an sich nicht in der Lage, gegen starke Strömungen anzuschwimmen. (7) Veränderung im Umfeld, wodurch Organismen an einer weiteren Verbreitung gehindert werden. PE 379.208 iv Klimawandel und europäische Fischerei Meeresgewässern. Dadurch könnten sich Organismen, die zuvor auf die küstennahen Bereiche beschränkt waren, weiter ausbreiten. Alle diese Vorgänge wirken sich (positiv oder negativ) auf die Primärproduktion( 8) aus. Der Klimawandel dürfte in halbumschlossenen Meeren noch schwerwiegendere Auswirkungen haben als in offenen Meeren. Die erwarteten Auswirkungen werden die Meeresproduktivität sowohl negativ als auch positiv beeinflussen. Szenarien für den Klimawandel prognostizieren • • • für die Ostsee einen Rückgang der Salinität von 8 bis 50 % und einen Temperaturanstieg beim Meeresoberflächenwasser um 2 bis 4 °C. für die Nordseeregion unterschiedliche Entwicklungen bei der Salinität, d. h. mit Anstiegen bzw. Rückgängen in verschiedenen Gebieten der Nordsee. Die Meeresoberflächentemperaturen sollen in der nördlichen Nordsee zwischen 1,6 °C und 3,0 °C steigen und in der flacheren südlichen Nordsee zwischen 3,0 °C und 3,9 °C. für das Mittelmeer einen Anstieg der Salinität und der Temperaturen. Die Nordatlantik-Oszillation Die Nordatlantik-Oszillation wirkt sich auf das europäische Meeresökosystem aus. Das Winterklima im europäischen Raum wird in vielerlei Hinsicht sehr stark von der Nordatlantik-Oszillation (NAO)( 9) beeinflusst, die die atmosphärischen Vorgänge im Nordatlantik dominiert. Es wurden verschiedene Indizes für die NAO festgelegt, die sich auf alle trophische Ebenen( 10) in den Meeren auswirkt. Die Rekrutierung( 11) der Industriefische hängt vom NAO-Index ab, wie es für die starke Nachwuchsproduktion bei den Kabeljauartigen( 12) (betrifft Kabeljau, Schellfisch, Wittling und Seelachs) in der Nordsee und für die Rekrutierung des Herings und der Sardine im Nordostatlantik aufgezeigt wurde. Die NAO ist so gut wie nicht prognostizierbar, obwohl sie sich aus der Meeresoberflächentemperatur rekonstruieren lässt. Die Untersuchungen des NAO-Index und der Zusammenhang mit beobachteten Auswirkungen auf das marine Ökosystem dürften es möglich machen, Modelle für die Prognostizierung künftiger Auswirkungen zu entwickeln. In vielen Fällen wurden Auswirkungen auf die wichtigsten Industriefischarten beobachtet Temperaturveränderungen können die Verlagerung von Fischpopulationen, das Eindringen fremder Arten und das Verschwinden von Arten zur Folge haben. Bei den Fischen ist die Temperatur eine ganz wesentliche Nischen-Komponente. Fische wählen in der Regel Habitate, (8) Assimilation (brutto) oder Akkumulation (netto) von Energie und Nährstoffen durch Grünpflanzen und Organismen, die sich von anorganischen Verbindungen ernähren. (9) Die Nordatlantik-Oszillation ist ein dominanter atmosphärischer Vorgang im Nordatlantik. Erläuterungen dazu werden in Kapitel 3.1 gegeben. – Sonstige einschlägige Literatur zu den Veränderungen bei den Eigenschaften des Wassers. (10) Einteilung von natürlichen Gemeinschaften oder Organismen entsprechend ihrem Platz in der Nahrungskette. Dabei kann grob zwischen Grünpflanzen (Produzenten), Pflanzenfressern (Konsumenten) und Fleischfressern unterschieden werden. (11) Die Zahl der Fische, die jährlich in einem Fanggebiet zum wirtschaftlich nutzbaren Bestand hinzukommt, und zwar durch Heranwachsen der Jungtiere (auf eine Größe, ab der sie fischbar sind) oder Migration (d. h. Fische dringen in das Fanggebiet vor). (12) Gruppe von Fischarten, die zur Familie Gadidae gehört, z. B. Kabeljau, Schellfisch. v PE 379.208 Klimawandel und europäische Fischerei deren thermische Bedingungen eine größtmögliche Wachstumsrate ermöglichen. Allerdings lassen sich die Auswirkungen von Temperaturveränderungen auf die Fische nur schwer vorhersagen. Abgesehen von der Temperatur entscheiden auch noch das Nahrungsangebot und das Vorhandensein geeigneter Laichplätze über die Verbreitung der Fischbestände. Direkte und indirekte Klimaeffekte können zu einer Verlagerung von Fischpopulationen, zum Eindringen fremder Arten und selbst zum Verschwinden von Arten führen. In verschiedenen Studien wurde ein Zusammenhang zwischen Größe und Verteilung von Fischbeständen und Zooplankton( 13) und einem Anstieg der Meerestemperatur beobachtet. Warmwasserfische dringen in „kalte“ Ökosysteme vor. Verschiedene Arten von Warmwasserfischen sind in „kalte“ Ökosysteme vorgedrungen, während Kaltwasserfische, die in „warmen“ Ökosystemen in relativ großer Zahl vorhanden waren, inzwischen sehr selten geworden oder gänzlich verschwunden sind. So gibt es z. B. im Mittelmeer kaum noch Sprotten und Makrelen, und auch in vielen anderen Fällen wurden Verschiebungen festgestellt. Da jedoch die betreffenden Arten oftmals stark befischt werden, lässt sich nur schwer ein direkter Kausalzusammenhang zwischen Temperatur und Verbreitungsmuster herstellen. Zuverlässige Prognosen zu möglichen Entwicklungen von Fischbeständen, die auf Effekte des Klimawandels zurückzuführen sind, lassen sich nur für einige bislang sehr intensiv untersuchte Arten anstellen (z. B. Kabeljau). Eine Abgrenzung gegenüber anderen Wirkungsfaktoren ist schwierig. Weitere Forschungsarbeiten sind erforderlich. Der Klimawandel beeinflusst Größe und Verbreitung der Industriefischbestände. Das sich verändernde Klima hat einen direkten Einfluss auf Überlebensrate, Ausbreitung, Fertilität und Verhalten der Einzeltiere und damit auf die Größe und Verbreitung der Bestände. Prognosen sind schwierig, da außer dem Klima noch viele andere Faktoren für die Verbreitung der Arten und die Dynamik dieser Veränderungen von großer Bedeutung sind. Veränderungen in der geographischen Streuung haben um Grönland herum zu einer erhöhten Produktivität bei Kabeljau und Schellfisch beigetragen. Das ist eine Reaktion auf eine dramatische Erwärmung in den 20er und 30er Jahren des vorigen Jahrhunderts, die Hauptursache für diese Veränderungen jedoch waren in der Tiefe ausgelöste Prozesse infolge einer vermehrten Phyto- und Zooplanktonbildung. Die mittleren Temperaturen am Meeresboden können die Wachstumsrate eines Fischbestandes wesentlich beeinflussen. Wie simple Simulationen für den Nordseekabeljau gezeigt haben, ist bei steigenden Meeresbodentemperaturen mit negativen Folgen für die Wachstumsrate zu rechnen. Solche Ansätze zeigen, dass sich die Prognosen hinsichtlich des Verhaltens von Fischpopulationen ändern können, wenn bei den entsprechenden Modellen Umweltfaktoren mit einbezogen werden. Simulationen können wertvolle Aufschlüsse für die Bewirtschaftung der Fischbestände geben, und sie machen deutlich, dass dabei die Berücksichtigung von Umweltauswirkungen wahrscheinlich immer wichtiger wird. Der Klimawandel kann sich indirekt sowohl positiv als auch negativ auf die Reproduktion von Industriefischbeständen auswirken, wie an mehreren Beispielen deutlich wird. Im Falle des Norwegischen Frühjahrslaichers (Hering) hat ein klimatisch bedingter Anstieg der Wassertemperatur zur Folge, dass die Fischpopulationen nicht am üblichen Laichprozess teilnehmen. Das wirkt sich indirekt auf das Reproduktionspotenzial aus und hat direkte Folgen für die gewerbliche Fischerei (Bestandsverluste). Der Rote Thun könnte sich während warmer (13) Tierisches Plankton. PE 379.208 vi Klimawandel und europäische Fischerei Perioden auch außerhalb seiner traditionellen Laichgründe vermehren, was der Produktivität des Bestandes förderlich wäre. Klimaveränderungen verursachen Verlagerungen bei den Industriefischpopulationen. Es gibt mehrere Beispiele dafür, dass temperatur- oder salinitätsbedingte Veränderungen bei den Wechselwirkungen (Nahrungsorganismen, Raubfische, Konkurrenten, Reproduktion) zur Verlagerung der Populationen führen. Mit Hilfe dieser Beispiele lassen sich einige Wechselwirkungen und internen Veränderungen in den Ökosystemen erklären. Es müssen jedoch Untersuchungen zur Populations- und Gemeinschaftsebene durchgeführt werden, da sich die direkten klimatischen Auswirkungen für die einzelnen Tiere nicht unmittelbar in Veränderungen in Verbreitung und Größe der Fischpopulationen niederschlagen. Die beim Plankton durch den Klimawandel hervorgerufenen Effekte haben die Verlagerung von Fischpopulationen zur Folge. Viele Industriefischarten sind direkt vom Plankton abhängig. Bei Fischarten, die sich von Plankton ernähren, insbesondere Sardinen und Sardellen, gibt es eine starke natürliche Fluktuation im Zusammenhang mit Klimaveränderungen. Untersuchungen bezüglich der Klimaerwärmung ließen erkennen, dass statt der nördlichen Arten in zunehmendem Maße die südlichen Arten dominieren. Bei der Untersuchung der Grenzen von mehr als 60 verschiedenen Nordseefischarten zeigte sich bei der Hälfte dieser Arten (befischt und nicht befischt) eine Verschiebung der Grenzen, wobei die Tendenz nach Norden geht. Einige Arten dürften ihre Belastbarkeitsgrenze erreicht haben, wie etwa der Kabeljau in der Nordsee, weshalb sich ihre Populationen nach Norden bewegen. Der Rückgang beim Kabeljau wurde mit veränderten Artenzusammensetzungen, Bestandsabnahme und einer kleineren durchschnittlichen Körpergröße des Zooplanktons in Zusammenhang gebracht, was sich möglicherweise auf den Klimawandel zurückführen lässt. Die Verlagerung von Populationen (wie am Beispiel des Kabeljau gezeigt) kann dazu führen, dass in einzelnen Regionen manche Bestände gänzlich verschwinden. Auswirkungen auf die biologische Umwelt betreffen auch die Industriefischarten Der Klimawandel hat Auswirkungen auf die Beutetiere und die natürlichen Feinde von Industriefischen. Wichtige Organismen in der biotischen( 14) Umwelt von Industriefischarten sind ihre Beutetiere und ihre natürlichen Feinde. Die wichtigsten Beuteorganismen für planktonfressende Fischarten sind kleine und große Ruderfußkrebse( 15). In der Ostsee sind kleine Arten wie Pseudocalanus und Acartia wichtige Nahrungsorganismen, wohingegen in der Nordsee und im Nordostatlantik auch große Ruderfußkrebse wie Calanus finmarchicus und C. helgolandicus von Bedeutung sind. Da sie einen wichtigen Stellenwert in der Nahrungskette einnehmen, sind ihre Reaktionen auf Klimaauswirkungen entscheidend für die Produktivität der Industriefischbestände. Invasive Arten treten zunehmend häufiger in europäischen Küstengewässern auf. Die biologische Invasion ist zu einem der auffälligsten Merkmale des globalen Wandels geworden. Durch sie können die biologische Vielfalt und die Funktionen von natürlichen Ökosystemen verändert werden, und sie kann erheblichen wirtschaftlichen Schaden verursachen. Klimatisch bedingte Störungen der Nahrungskette beeinflussen das Überleben und die Produktivität von Industriefischbeständen. Für das Überleben und die Produktivität von (14) Leben und lebende Organismen. (15) Die größte Klasse von Krebstieren. Entweder frei lebend oder als Parasiten. Stellen den größten Teil des Zooplankton, sind jedoch auch bei den Bodenfischarten anzutreffen. vii PE 379.208 Klimawandel und europäische Fischerei Fischbeständen ist es ganz wichtig, dass für die Menge an Fischlarven auch ausreichend Zooplankton in der richtigen Größe vorhanden ist. Aufgrund der klimatischen Veränderungen durchlaufen viele Planktongruppen( 16) ihre jahreszeitlichen Zyklen schneller, und der fehlende Gleichtakt hat schwerwiegende Folgen für das Überleben und die Produktivität der Industriefischbestände. Bei einem weiteren Anstieg der Temperaturen wird es auch künftig zu derartigen Störungen in der marinen Nahrungskette kommen. Das hat Folgen für die Größe und die Verbreitung der Fischbestände, wie an vielen Beispielen veranschaulicht wurde: Diatomeenblüten( 17) wirken sich negativ auf die Organismen in der Nordsee aus, bei denen die Diatomeen die Nahrungsgrundlage darstellen. Beim Kabeljau verringern sich durch den fehlenden Gleichtakt mit der Größe der Beuteorganismen die Überlebenschancen der Larven. Auch der junge Sandaal sieht sich mit einem verringerten Nahrungsangebot konfrontiert, wodurch die Überlebenschancen beeinträchtigt sind. In den Gewässern rund um das Vereinigte Königreich bewirkt eine erhöhte Wassertemperatur eine Verringerung der Rekrutierung bei Scholle und Limande. Eine Verschiebung des Gleichgewichts zwischen Meroplankton( 18) und Holoplankton( 19) und damit zwischen Benthos( 20) und Pelagial wirkt sich auf das Überleben der Fischlarven aus. Wirtschaftliche Auswirkungen für die industrielle Fischerei Der Klimawandel kann schwerwiegende wirtschaftliche Auswirkungen für die industrielle Fischerei haben. Ausgehend davon, dass die industrielle Fischerei unmittelbar mit der marinen Produktion verbunden ist, würde ein Anstieg bzw. Rückgang der Produktivität um 10 % einen wirtschaftlichen Gewinn bzw. Verlust von mehr als 200 Mio. EUR bedeuten.( 21) Die Auswirkungen der Bestandsverlagerungen auf die industrielle Fischerei lassen sich nur schwer beurteilen, da der durch die Verlagerung bedingte Schwund normalerweise mit einer entsprechenden Zunahme anderer Arten einhergeht. Es werden positive und negative wirtschaftliche Auswirkungen für die industrielle Fischerei erwartet. Beim gegenwärtigen Kenntnisstand ist es nicht möglich, die durch den Klimawandel zu erwartenden Veränderungen in der weltweiten marinen Produktion quantitativ vorherzusagen, da hierbei eine Vielzahl von Wechselwirkungen auftreten. Zweifellos jedoch werden Veränderungen der Produktivität und der Saisonmuster die Bewirtschaftung der biologischen Ressourcen der Meere beeinflussen. Wie (nicht sehr zuverlässige) Schätzungen besagen, dürfte es bis 2050 zu einer Zunahme der weltweiten marinen Produktion kommen, jedoch um nicht mehr als 10 %. Im Gegensatz dazu haben Satellitenbeobachtungen und umfangreiche Probenahmen von Plankton gezeigt, dass Phytoplankton( 22) und Chlorophyll in den letzten 20 bis 50 Jahren zurückgegangen sind. Für 74 % der permanent geschichteten Weltmeere ist die Oberflächenerwärmung mit einer Verringerung der Produktivität verbunden. (16) Ein Taxon ist eine Gruppe von Organismen innerhalb der biologischen Systematik. (17) Bildet Bestandteil des Phytoplanktons, daher plötzliche und rasche Zunahme der Biomasse der Planktonpopulation. Saisonale Blüten sind wichtig für die Produktivität des aquatischen Systems. Sporadische Planktonblüten können giftig sein. (18) Temporäres Plankton bestehend aus Organismen in pelagischen Stadien, die auch benthische Stadien haben. Hauptsächlich Larven von sesshaften Organismen. (19) Plankton mit einem vollständig pelagischen Lebenszyklus. (20) Organismen, die am Grund der Gewässer festsitzen bzw. dort kriechen, laufen oder vorübergehend schwimmen oder die auf den Bodensedimenten leben. (21) Bei einem geschätzten Marktwert der Anlandungen der europäischen Länder aus dem Nordostatlantik, dem Mittelostatlantik, dem Mittelmeer und dem Schwarzen Meer in Höhe von 2 Mrd. EUR. Das entspricht etwa einem Drittel des Gesamtmarktwertes aller Anlandungen der EU-Länder, gleich welchen Ursprungs. (22) Planktonpflanze. PE 379.208 viii Klimawandel und europäische Fischerei In den nördlichen Breiten wird mit einer Produktionszunahme gerechnet, da große Gebiete eisfrei werden. Mögliche Auswirkungen auf die Aquakultur Der Klimawandel kann schwerwiegende wirtschaftliche Auswirkungen auf die Aquakultur haben. Die Meeresaquakultur in der EU erreicht einen Marktwert von rund 2 Mrd. EUR, wobei einschneidende wirtschaftliche Veränderungen aufgrund des Klimawandels befürchtet werden. Es werden positive und negative Auswirkungen erwartet. Steigende Wachstumsraten und eine effiziente Futterverwertung bei einigen in der Aquakultur eingesetzten Arten könnten einen Produktivitätszuwachs bewirken. Auch wäre es möglich, neue Arten in die Aquakultur einzuführen. Andererseits wird mit solch negativen Auswirkungen wie Wärmestress für Kaltwasserarten und Wattorganismen, Krankheiten und Krankheitsanfälligkeit gerechnet. Außerdem sind Schäden an Fischzuchtanlagen aufgrund von extremen Wetterereignissen denkbar. Eventuell macht sich in Abhängigkeit von veränderten Umweltbedingungen eine Verlagerung von Produktionsanlagen an geeignetere Standorte notwendig. Darüber hinaus wird befürchtet, dass Hitzewellen insbesondere für Anlagen im küstennahen Flachwasser gravierende wirtschaftliche Folgen haben. Es ist nicht bekannt, ob die positiven Auswirkungen die negativen überwiegen werden oder umgekehrt. Die Auswirkungen der industriellen Fischerei auf die wichtigsten Arten Während der letzten Jahrzehnte wurden bei den kommerziellen Fischbeständen im Atlantik zahlreiche Veränderungen beobachtet. Dabei ist es äußerst schwierig, die aus Veränderungen in der Populationsdichte und der Rekrutierung resultierenden Folgen sowie die regionalen Klimaeffekte von direkten anthropogenen Einflüssen, wie etwa der Fischerei, zu unterscheiden. Die derzeitige Fischereipraxis schwächt die Widerstandsfähigkeit der Fischbestände gegenüber den Auswirkungen des Klimawandels. Die derzeitigen Fangmethoden richten sich auf ältere und größere Fische und bewirken eine Veränderung der Größen- und Altersstruktur der Fischpopulationen. Die Widerstandsfähigkeit gegenüber Umweltauswirkungen wird geschwächt, die genetische Variabilität geht verloren und die Fähigkeit zur Anpassung an Umweltveränderungen wird schwächer. Folglich kann die Fischerei dazu führen, dass die Umweltauswirkungen für die Bestände zu einer immer größeren Gefahr werden. Eine ökologisch nachhaltige Fischerei sollte den Schutz der größeren und älteren Fische zum Ziel haben. Unter dem Gesichtspunkt der Nachhaltigkeit befischte Bestände können besser auf Klimaauswirkungen reagieren. Die Ungewissheit über den künftigen anthropogenen Klimawandel rechtfertigt einen Vorsorgeansatz für die Bestandsbewirtschaftung. Die globale Erwärmung kann sich erheblich – in positiver oder negativer Art und Weise – auf die meisten der kommerziellen Fischbestände auswirken. Dabei sind durch Überfischung drastisch reduzierte Bestände anfälliger gegenüber klimatischen Veränderungen als nachhaltig befischte Bestände. Maßgeblich für die Reaktion der Fischbestände auf Umwelteinflüsse ist die Populationsgröße. Gesunde Bestände können sich besser an Populationsverschiebungen und strukturelle Veränderungen im Ökosystem anpassen. Eine flexible und anpassungsfähige Bestandsbewirtschaftung ist notwendig. Ein mögliches Szenario für die Zukunft der Nordsee beinhaltet einen Anstieg der Temperatur, eine starke NAO und einen verstärkten Zufluss von Atlantikwasser. Folgen davon wären eine geringe ix PE 379.208 Klimawandel und europäische Fischerei Rekrutierung des Kabeljau, eine Nordwärtsverlagerung der jetzigen Fischarten (Kabeljau, Hering und Sprotte) und das Vordringen südlicher Arten (Sardine und Sardelle). Eine wichtige Frage ist die, wie in Anbetracht der klimatischen Auswirkungen auf die Meeresumwelt die industrielle Fischerei künftig zu gestalten ist. Da einige der wichtigsten Industriefischarten (wie Hering und wahrscheinlich andere kleine pelagische( 23) Arten) sehr stark auf sich verändernde hydrographische Bedingungen reagieren, sollte die künftige Bestandsbewirtschaftung kontinuierlich sein, gleichzeitig aber auch flexibel und anpassungsfähig, um den jeweiligen Reaktionen der Bestände auf die Umweltbedingungen Rechnung zu tragen. Insbesondere die ausgesprochenen Wanderarten, die ihre Routen bei Umweltveränderungen ändern, werden besondere Anforderungen an die Bewirtschaftung stellen. Um das Fischereimanagement rechtzeitig auf die Situation einzustellen, müssen durch entsprechende Forschungsarbeiten die Kenntnisse über die Auswirkungen des Klimawandels auf die Fischerei der EU und damit verbundene Prognosen verbessert werden. Strategische Optionen Die in immer größerem Umfang freigesetzten künstlichen Treibhausgase haben einen großen Anteil am Klimawandel. Der Klimawandel wirkt sich direkt wie auch indirekt auf vielerlei Art und Weise auf die industrielle Fischerei und die Meeresaquakultur in Europa aus. Insgesamt steht dabei ein Marktvolumen von mehr als 4 Mrd. EUR auf dem Spiel. Es besteht kein Zweifel, dass die Klimaveränderungen das Meeresökosystem sowie die Fischerei und die Meeresaquakultur der EU positiv und negativ beeinflussen werden. Ungewiss ist jedoch, ob dabei die positiven oder negativen Auswirkungen überwiegen werden. Die Kenntnisse über die Beziehungen und Wechselwirkungen zwischen dem Meeresökosystem, der industriellen Fischerei, der Meeresaquakultur und den Klimaveränderungen und ihr Verständnis sind in vielerlei Hinsicht noch unzureichend. Unter Berücksichtigung verschiedener strategischer Alternativen wurden 7 generelle Optionen ermittelt (die nachfolgend erörtert werden): Option 1. Option 2. Option 3. Option 4. Option 5. Keine Maßnahmen. Verringerung der Treibhausgase. Gesetzgeberische und politische Maßnahmen sowie Unterstützung bestehender Initiativen zur Senkung der Treibhausgasemissionen. Strategien zur Erhöhung der Widerstandsfähigkeit der Fischpopulationen. Entwicklung von Strategien und Konzepten für eine nachhaltige Bewirtschaftung der Fischbestände, wodurch deren Widerstandsfähigkeit gegenüber Umweltauswirkungen erhöht wird. Strategien zur Verbesserung der Bestandsbewirtschaftung. Entwicklung von flexiblen Strategien und Konzepten für die Bestandsbewirtschaftung, die entsprechend den Reaktionen der Bestände auf die Umweltbedingungen angepasst werden können. Strategien zur Verbesserung der Aquakultur. Entwicklung von Strategien und Konzepten, um die negativen Auswirkungen auf die Meeresaquakultur so gering wie möglich zu halten. (23) Fische, die den Großteil ihres Lebens in der Wassersäule schwimmen und den Meeresgrund kaum aufsuchen oder brauchen. Bezieht sich normalerweise auf ausgewachsene Tiere. PE 379.208 x Klimawandel und europäische Fischerei Option 6. Option 7. Förderung der Forschung. Förderung von entsprechenden Forschungsarbeiten und des Austauschs von Kenntnissen. Kompensation negativer Auswirkungen. Bereitstellung von Unterstützung (finanzieller oder anderer Art), um die negativen Auswirkungen des Klimawandels zu kompensieren. Diskussion der Optionen: Option 1) Da nicht erwiesen ist, dass der Klimawandel eine insgesamt negative Auswirkung auf die EU-Fischerei haben wird, wäre es eine mögliche Option, gar keine Maßnahmen zu ergreifen. Dies hätte den Vorteil, dass keinerlei Anschubkosten anfallen. Die positiven Auswirkungen des Klimawandels auf die Fischerei, wie etwa eine erhöhte Produktion des Meeresökosystems aufgrund eines Temperaturanstiegs, werden nicht eingeschränkt. Andererseits erscheint es unakzeptabel, nicht zumindest gegen die negativen Folgen des Klimawandels vorzugehen (einschließlich finanzielle Auswirkungen), da diese ohne irgendwelche Gegenmaßnahmen in vollem Umfang wirksam werden. Option 1 wird nicht empfohlen. Option 2) Diese Option betrifft die Unterstützung einer europäischen Politik zur Bekämpfung des Klimawandels (z. B. Monitoring und Berichterstattung bezüglich der EUTreibhausgasemissionen oder das Emissionshandelssystem) und von geeigneten Maßnahmen, um die Treibhausgaskonzentrationen in der Atmosphäre so gering wie möglich zu halten. Da die Treibhausgasemissionen eine Ursache des Klimawandels sind, werden mit dieser Option die Klimaveränderungen direkt an der Quelle bekämpft. Option 2 wird empfohlen. Option 3) Es hat sich gezeigt, dass die frühere wie auch die jetzige industrielle Fischerei die Widerstandsfähigkeit der Fischbestände gegenüber Umweltveränderungen schwächt. Ziel von Option 3 ist die Entwicklung von Strategien für eine nachhaltige Fischerei, wodurch diese Widerstandsfähigkeit erhöht wird. Dazu ließen sich beispielsweise Möglichkeiten untersuchen, wie der Fang von älteren und größeren Fischen reduziert werden kann, oder es könnten marine Schutzgebiete eingerichtet werden. Mit dieser Option soll die gegenwärtige Fischerei so gestaltet werden, dass sie den Auswirkungen des Klimawandels Rechnung trägt. Das entspricht den im EU-Anpassungsprogramm im Rahmen des Europäischen Programms zur Klimaänderung (ECCP) festgelegten Zielen, wonach Möglichkeiten gefunden werden sollen, um die Resistenz Europas gegen die Folgen des Klimawandels in verschiedenen Sektoren zu stärken. Die Gemeinsame Fischereipolitik (GFP) spielt eine entscheidende Rolle bei der Bewirtschaftung der Fischbestände und sollte dabei mögliche Klimaauswirkungen entsprechend berücksichtigen. Sie könnte auch ein geeignetes Instrument sein, um die Widerstandsfähigkeit der Fischbestände gegenüber den Auswirkungen des Klimawandels durch entsprechende Strategien zu verbessern. Option 3 wird empfohlen. Option 4) Die Fischbestände werden durch spezifisches Verhalten auf den Klimawandel reagieren, z. B. durch zeitliche oder räumliche Veränderungen in der Bestandsgröße oder durch Veränderungen in den Wanderrouten. Bei Option 4 besteht das Ziel darin, die Bestandsbewirtschaftung auf die Reaktionen der Fischbestände einzustellen, um deren effiziente Nutzung zu ermöglichen. Dazu ist ein flexibles Bewirtschaftungsregime erforderlich, da nur so negative wirtschaftliche Folgen vermieden werden können. Diese Option hat das Ziel, die jetzige Bestandsbewirtschaftung an die Auswirkungen des Klimawandels anzupassen und befindet sich daher in Übereinstimmung mit den Zielsetzungen des EU-Anpassungsprogramms im Rahmen des ECCP. Unbedingt müssen dabei auch die Nachhaltigkeitsaspekte im Sinne von Option 2 berücksichtigt werden. Anderenfalls besteht die Gefahr, dass die verbesserte Bewirtschaftung nur für eine maximale Ausbeutung der Meeresressourcen genutzt wird, ohne xi PE 379.208 Klimawandel und europäische Fischerei darauf hinzuwirken, dass die Widerstandsfähigkeit der Fischbestände gegenüber den Umweltauswirkungen erhöht wird. Wie bei Option 3 könnte die GFP maßgeblich zur Umsetzung entsprechender Strategien beitragen. Option 4 wird empfohlen. Option 5) Ähnlich wie bei den Optionen 3 und 4 handelt es sich hier um eine Anpassungsstrategie entsprechend dem EU-Anpassungsprogramm im Rahmen des ECCP. Das Ziel ist die Vermeidung von negativen Auswirkungen auf die Meeresaquakultur. Denkbare Bestandteile der Strategie sind beispielsweise die Verlagerung von Produktionsanlagen Richtung Norden, die Verlagerung von Produktionsstätten in tieferes und kälteres Wasser oder die verstärkte Ausrichtung auf neue Arten. Ähnlich wie bei den Optionen 3 und 4 könnte die GFP maßgeblich zur Umsetzung entsprechender Strategien beitragen. Option 5 wird empfohlen. Option 6) Insbesondere die Optionen 3 bis 5 sind mit einem umfangreichen Forschungsbedarf verbunden. Angemessene Kenntnisse über die Beziehungen und Wechselwirkungen zwischen dem Meeresökosystem, der industriellen Fischerei, der Meeresaquakultur und dem Klimawandel sind unerlässlich. Eine solide Wissensgrundlage ist angesichts der Gefahren durch den Klimawandel eine Voraussetzung für die Entwicklung effizienter Strategien für eine nachhaltige Fischerei, Bestandsbewirtschaftung und Meeresaquakultur. Spezifische Forschungsaufgaben sind in Kapitel 4 aufgeführt. Die EU könnte entsprechende Forschungen anregen und/oder unterstützen, um die Faktengrundlage für ihre politische Entscheidungen und die zu entwickelnden Strategien zu verbessern, und sie könnte zur Verbreitung der gewonnenen Erkenntnisse beitragen. Option 6 wird empfohlen. Option 7) Diese Option betrifft die Bereitstellung von Unterstützung (finanzieller oder anderer Art), um die negativen Auswirkungen des Klimawandels zu kompensieren. Dazu gehören Ausgleichszahlungen für Regionen, die große wirtschaftliche Einbußen als Folge klimatischer Veränderungen erlitten haben (z. B. durch Produktionsausfälle in der Aquakultur nach Hitzewellen oder durch den Zusammenbruch der Fischrekrutierung, wenn eine Fischpopulation aus der Region verschwunden ist). Solche Zahlungen könnten der betreffenden Region kurzfristig helfen. Allerdings tragen Ausgleichsmaßnahmen reaktiven Charakter und sind nicht zukunftsorientiert. Proaktive und wissensbasierte Ansätze dürften hier am besten geeignet sein (z. B. geänderte Strategien für die Aquakulturproduktion oder das Fischereimanagement). Option 7 wird nicht empfohlen. Error! Reference source not found. gibt einen Überblick über die entwickelten strategischen Optionen und deren Vor- und Nachteile PE 379.208 xii Phänomen Thermohaline Zirkulation 12 13 pH-Wert der Meere 11 Ostsee Schichtung Meeresspiegel 10 Küstenerosion Mittelmeer − Zunahme Abnahme Abnahme Globaler Anstieg Anstieg xiii Abnahme der Nährstoffe, Auswirkungen auf die Primärproduktion Schwächere Grenzströme, Temperaturrückgang im Nordostatlantik Verringerte Kalzifizierung ” ” 9 Anstieg/ Rückgang Nordsee − ” 8 Rückgang Anstieg ” ” Mittelmeer − 6 Anstieg (1,6 °C – 3,9 °C) ” Ostsee Nordsee − 5 Anstieg (2 °C-4 °C) ” − Ostsee − 4 Globaler Anstieg Auswirkungen auf die Umweltbedingungen auf allen trophischen Ebenen 7 Temperatur 3 Nicht vorhersagbar ” Nordatlantik-Oszillation 2 Wirkung Anstieg, trägt zur globalen trägt zur globalen Erwärmung bei Erwärmung bei Trend Salinität Treibhausgas 1 Eigenschaften des Meereswassers Lfd. Nr. Zuverlässigkeit BACC 2006 Bryden et al. 2005, WGBU-Bericht 2006 Caldeira und Wickett 2003 IPCC 2007 Eisenreich 2005 ICES 2006 BACC 2006 Hoeppfner et al. 2006 Sheppard 2004 BACC 2006 IPCC 2007 Hurrell et al. 2006 PE 379.208 Mittel Gering Hoch Mittel Mittel Gering Hoch Mittel Hoch Hoch Hoch Gering IPCC 2007, Karl und Hoch Trenberth 2003 Literatur Tabelle 1: Durch den Klimawandel beeinflusste Phänomene mit ihren spezifischen Trends, Wirkungen, Literaturangaben und Zuverlässigkeit Klimawandel und europäische Fischerei Mittelmeer 15 Nordsee − 17 PE 379.208 Reproduktionsmuster Mittelmeer − 21 23 Nordatlantik − 20 Reproduktionsmuster Nordatlantik − 19 22 Nordsee − 18 Verbreitung der wichtigsten Industriefischarten Nordsee − 16 Schwankungen bei der Artendominanz in Abhängigkeit von der NAO Biologie der wichtigsten Arten Nordsee Phänomen 14 Lfd. Nr. Klimawandel und europäische Fischerei Wahl der Muttergewässer in Abhängigkeit von den äußeren Gegebenheiten Spätere Laichzeit Migration in kältere Gewässer Veränderung der Migration Vergrößerung der Verbreitungsgebiete Nordwärtsverlagerung Abnahme Zunahme Zunahme Zunahme Trend Ravier und Fromentin 2004 Änderung der Laichplätze xiv Cushing 1984 Bombace 2001, Dulcic et al. 1999 Sissener und Bjorndal 2005 Drinkwater 2006 Perry et al. 2006, Southward et al. 1996 Cushing 1984 Alheit und Hagen 1997, Beare et al. 2004 Bethoux et al. 2002 Beare 2002 Literatur Gleichtakt/fehlender Gleichtakt mit der Nahrung Verlust von „Kaltwasserbiota“ Zugewinne oder Verluste bei den Beständen Höhere Produktivität Bestandsverluste „Kabeljauboom“ Wechsel von Hering auf Sardine Abnahme der Nährstoffe, Auswirkungen auf die Primärproduktion Abnahme der Nährstoffe, Auswirkungen auf die Primärproduktion Wirkung Mittel Mittel Hoch Mittel Mittel Hoch Hoch Hoch Mittel Mittel Zuverlässigkeit Vordringen südlicher Arten 24 Gestörte Trophodynamik 27 Nördliche Breiten − 29 − Ostsee Verlagerungen bei den Beutearten im Zooplankton Auswirkung der Temperatur Verschiebungen des Anstieg/Rückgang (optimale Temperatur für jede Art) Zunahme Zunahme/Abnahme Zunahme/Abnahme der Sekundär- und Zunahme oder Abnahme der Produktivität Zunahme der Sekundär- und Tertiärproduktion Möllmann et al. 2005 O’Brien et al. 2000, Planque und Fredou 1999 Hoch Hoch Mittel Gering Gering Mittel Gering Hoch Zuverlässigkeit ACIA 2005, Hoepffner et al. 2006 Sarmiento et al. 2004, Behrenfeld et al. 2006 Beaugrand 2003 Geringerer Rekrutierungserfolg Zunahme der Sekundär- und Tertiärproduktion Edwards und Richardson 2004 Helmuth et al. 2006, Nehring 2003 Perry et al. 2005 Literatur Gleichtakt/fehlender Gleichtakt Treten an die Stelle heimischer Arten Veränderung der Zielarten in der Fischerei Wirkung xv PE 379.208 (24) Die Wissenschaft, die sich mit den periodisch wiederkehrenden biologischen Erscheinungen im Hinblick auf das Klima befasst, insbesondere mit saisonalen Veränderungen. 31 30 Rekrutierung Weltweit − 28 Primärproduktivität Zeitliche Vorverlagerung der Frühjahrsblüte Phänologie( 24) 26 Zunehmend geringerer Gleichtakt bei den trophischen Interaktionen Zunahme Vordringen exotischer Arten Zunahme Trend 25 Biologische Umwelt Phänomen Lfd. Nr. Klimawandel und europäische Fischerei Mittelmeer − 33 − Nordsee Auswirkung der Fischerei Auswirkung der Fischerei 39 40 PE 379.208 Quelle: BIPRO/IFM-GEOMAR, 2007. Auswirkung der Fischerei Nordatlantik (Kabeljau) 38 − Erholung der Bestände Fischerei 37 36 Produktionsanlagen − 35 Zusammenbruch der Fischerei Produktivität − 34 Auswirkung auf die Aquakultur Produktivität Nordsee − 32 Klimawandel und europäische Fischerei Verlust der genetischen Variabilität Abnahme der Widerstandsfähigkeit Abnahme der Zahl großer und älterer Fische Ausbleiben aufgrund von Artenverlagerung JA, wenn sich die Arten den Grenzen der Verbreitungsgebiete nähern Verlagerung in nördliche Richtung Zunahme Verschiebungen des Ökosystems Verschiebungen des Ökosystems Ökosystems xvi Geringere Anpassungsfähigkeit an Umweltveränderungen Verringerte Rekrutierung Verringerte Rekrutierung Bestandsverluste Bestandsverluste auf regionaler Ebene Wirtschaftlicher Verlust Wirtschaftlicher Nutzen Zunahme/Abnahme der Sekundär- und Tertiärproduktion Zunahme/Abnahme der Sekundär- und Tertiärproduktion Tertiärproduktion Hauser et al. 2002 Hsieh et al. 2006 Berkeley et al. 2004, Birkeland und Dayton 2005 Drinkwater 2006 Drinkwater 2005 Stenevik und Sundby 2006 Lehtonen 1996 Molinero et al. 2007 Beaugrand 2004 Gering Mittel Hoch Gering Gering Mittel Mittel Mittel Hoch Klimawandel und europäische Fischerei Abkürzungsverzeichnis NAO Nordatlantik-Oszillation ICES Internationaler Rat für Meeresforschung psu Practical Salinity Unit ohne Maßangabe SST Meeresoberflächentemperatur CFP Gemeinsame Fischereipolitik ECCP Europäisches Programm zur Klimaänderung CPR Kontinuierlich messender Plankton-Rekorder PE 379.208 xvii Klimawandel und europäische Fischerei Glossar Abiotische Umwelt Versauerung Advektion Benthisch Benthos Bioklimatische Rahmenbedingungen Biogeographie Biogeographische Barriere Biotisch Biotische Umwelt Blüte Cephalopoden (Kopffüßer) Cladoceren Die nichtlebende Umwelt, bestehend aus allen abiotischen Faktoren, z. B. Temperatur, Salinität Verringerung des pH-Wertes. Der pH-Wert ist ein Maß für die Stärke der sauren bzw. basischen Wirkung einer Lösung. Transport durch den Hauptstrom, im Gegensatz zu Diffusion oder Verbreitung durch Dispersion Bezeichnet Fische und andere Tiere, die auf dem oder im Meeresboden leben Organismen, die am Grund der Gewässer festsitzen bzw. dort kriechen, laufen oder vorübergehend schwimmen oder die auf den Bodensedimenten leben Bestimmt mit Hilfe von Verfahren zur Korrelation der derzeitigen Verbreitung der Arten mit Klimavariablen bzw. durch Ermittlung der physiologischen Reaktion des Einzeltiers auf Klimaänderungen Beschreibt die Verbreitung von Organismen auf der Erde und untersucht die Ursachen der geographischen Verbreitung lebender und ausgestorbener Taxa; Veränderung im Umfeld, wodurch Organismen an einer weiteren Verbreitung gehindert werden Leben und lebende Organismen Die lebende Umwelt, bestehend aus allen biotischen Faktoren, z. B. Nahrung, Fressfeinde Bildet Bestandteil des Phytoplanktons, daher plötzliche und rasche Zunahme der Biomasse der Planktonpopulation. Saisonale Blüten sind wichtig für die Produktivität des aquatischen Systems. Sporadische Planktonblüten können giftig sein. Tiere (Weichtiere) mit am Kopf und um den Mund zusammenlaufenden Tentakeln (Tintenfische, Kuttelfische, Kraken) Gruppe kleiner Plankton-Krebstiere, die sich vor allem in Süßwasser finden, z. B. Daphnien. Es kommen auch marine Arten vor, z. B. Podon-Arten, die zum Zooplankton zählen Konvektion Vertikale Zirkulation in einem Gas oder einer Flüssigkeit unter Bedingungen der Instabilität Ruderfußkrebse Die größte Klasse von Krebstieren. Entweder frei lebend oder als Parasiten. Stellen den größten Teil des Zooplanktons, sind jedoch auch bei den Bodenfischarten anzutreffen. Cyanobakterien Auch Blaualgen genannt. Gruppe von Bakterien Grundfische Leben in enger Beziehung mit dem Meeresgrund und hängen von ihm ab PE 379.208 xviii Klimawandel und europäische Fischerei Diatomeen Mikroskopisch kleine einzellige Algen mit einer äußeren, wunderschön strukturierten Schale aus Silikat, deren zwei Teile mit breiten Gürtelbändern verbunden sind Dinoflagellaten Gruppe planktonischer Algen Euphotische Zone Die obere, durchleuchtete Schicht des Wassers, in der effektive Photosynthese möglich ist Euryhalin Organismus, der bei verschiedenstem Salzgehalt überleben kann Eurytherm Organismus, der bei verschiedensten Temperaturen überleben kann Eutrophierung Im Allgemeinen der natürliche oder vom Menschen ausgelöste Prozess, durch den ein Gewässer mit gelösten Mineralstoffen (vor allem Phospor und Stickstoff) angereichert wird, die das Wachstum von Wasserpflanzen anregen und die organische Produktion im Gewässer erhöhen. Eine übermäßige Anreicherung kann dazu führen, dass der gelöste Sauerstoff aufgebraucht wird, was letztlich das Absterben oder die Verlagerung von Arten zur Folge hat Flagellaten Gruppe, zu denen die Dinoflagellaten gehören, auch Euglenida, Raphidophyta Kabeljauartige Gruppe von Fischarten, die zur Familie Gadidae gehört, z. B. Kabeljau, Schellfisch Gesellig lebend In Bezug auf eine soziale Gruppe Halokline Region unter der Oberflächenschicht eines Meeres oder Sees, in der der Salinitätsgradient sprunghaft zunimmt (d. h. wo die Salinität mit zunehmender Tiefe rapide abnimmt). Holoplankton Plankton mit einem vollständig pelagischen Lebenszyklus Interspezifisch Zwischen den verschiedenen Arten, z. B. Konkurrenzkampf Intraspezifisch Innerhalb einer Art, z. B. Konkurrenzkampf Meroplankton Temporäres Plankton bestehend aus Organismen in pelagischen Stadien, die auch benthische Stadien haben. Hauptsächlich Larven von sesshaften Organismen NAO Die Nordatlantik-Oszillation ist ein dominanter atmosphärischer Vorgang im Nordatlantik; Erläuterungen dazu werden in Fußnote 9 gegeben Neritisch Bezieht sich auf den ozeanischen Bereich oberhalb des Festlandsockels und die Kante des Schelfabhangs. Entspricht den küstennahen Gewässern.. Neritische Arten Arten, deren Lebenszyklus und Verbreitung größtenteils auf den Kontinentalsockel und die obere Neigung beschränkt sind Pelagisch Fische, die den Großteil ihres Lebens in der Wassersäule schwimmen und den Meeresgrund kaum aufsuchen oder brauchen. Bezieht sich normalerweise auf ausgewachsene Tiere PE 379.208 xix Klimawandel und europäische Fischerei Phänologie Phytoplankton Plankton Niederschlag Primärproduktion Strahlungsbilanz (Radiative forcing) Rekrutierung Salinität Sekundärproduktion Nichtteilnahme am Laichprozess SRES-MarkerSzenarien Schichtung (der Wassersäule) Taxon Tertiärproduktion PE 379.208 Die Wissenschaft, die sich mit den periodisch wiederkehrenden biologischen Erscheinungen im Hinblick auf das Klima befasst, insbesondere mit saisonalen Veränderungen Planktonpflanze Schwebende Organismen, deren Bewegungen mehr oder weniger von den Strömungen abhängig sind. Zwar gibt es beim Zooplankton einige Arten, die sich durch aktive Schwimmbewegungen in einer vertikalen Position halten können, jedoch ist Plankton an sich nicht in der Lage, gegen starke Strömungen anzuschwimmen. Meteorologie: die auf die Erde fallenden Produkte der Kondensation in der Atmosphäre, wie etwa Regen, Hagel oder Schnee Assimilation (brutto) oder Akkumulation (netto) von Energie und Nährstoffen durch Grünpflanzen und Organismen, die sich von anorganischen Verbindungen ernähren Siehe Fußnote 31; "Strahlungsbilanz" ist definiert als die Differenz zwischen der ankommenden und der abgehenden Strahlungsenergie in einem bestimmten Klimasystem. Eine positive Bilanz (mehr ankommende Energie) führt in der Regel zu einer Erwärmung des Systems, während eine negative Bilanz (mehr abgehende Energie) eine Abkühlung bewirkt Die Zahl der Fische, die jährlich in einem Fanggebiet zum wirtschaftlich nutzbaren Bestand hinzukommt, und zwar durch Heranwachsen der Jungtiere (auf eine Größe, ab der sie fischbar sind) oder Migration (d. h. Fische dringen in das Fanggebiet vor) Die relative Konzentration von Salzen, normalerweise Natriumchlorid, in einem bestimmten Boden oder Gewässer. Auffrischung bezieht sich in diesem Zusammenhang auf eine Verringerung der Salze im Wasser. Ohne Einheit; gelegentlich wird noch die alte Einheit PSU (practical salinity unit) verwendet Produktionsrate pflanzenfressender Tiere durch Umwandlung der pflanzlichen Nahrung in tierisches Gewebe Fisch, der bereits an der Fortpflanzung teilgenommen hat, aber eine oder mehrere Fortpflanzungsperioden überspringt Spezifische Klimaszenarien, die im Sonderbericht zu den Emissionsszenarien beschrieben werden Das Auftreten von mehr oder weniger horizontalen Wasserschichten im Meer als eine Folge von Unterschieden in der Dichte, d. h. Unterschiede bei Salinität (Halokline) oder Temperatur (Thermokline) Ein Taxon ist eine Gruppe von Organismen innerhalb der biologischen Systematik Produktionsrate fleischfressender Tiere durch Umwandlung der tierischen Nahrung in eigenes Gewebe xx Klimawandel und europäische Fischerei Thermischer Äquator Thermokline Der thermische Äquator wird durch die breitenkreisähnliche Linie repräsentiert, die die im Mittel wärmsten Punkte der Erde auf ihren jeweiligen Längengraden miteinander verbindet Region unterhalb der Oberflächenschicht des Meeres oder eines Sees, die ein steiles Temperaturgefälle aufweist (d. h. in der die Temperaturen mit zunehmender Tiefe sehr stark absinken). Eine Thermokline kann bis an die Oberfläche reichen und zu einer Front werden. Sie stellt normalerweise eine ökologische Barriere dar, und ihre Oszillationen haben erhebliche Auswirkungen auf die Bestandsverteilung und die Produktivität des Meeres. Thermohaline Zirkulation Zirkulation im Ozean, die durch temperatur- und salinitätsbedingte Unterschiede in der Meerwasserdichte bewirkt wird Übergangszone In der Biogeographie eine Zone zwischen zwei deutlich unterschiedlichen Habitaten. Oftmals eine Barriere für die Verbreitung Trophische Ebene Einteilung von natürlichen Gemeinschaften oder Organismen entsprechend ihrem Platz in der Nahrungskette. Dabei kann grob zwischen Grünpflanzen (Produzenten), Pflanzenfressern (Konsumenten) und Fleischfressern unterschieden werden Tropischer Niederschlagsgürtel Der tropische Niederschlagsgürtel erstreckt sich am Äquator zwischen 30° südlicher Breite und 30° nördlicher Breite. Entsprechend dem Zenit der Sonne reicht die Regenzeit im Nordsommer bis an die nördliche Grenze und im Südsommer (Nordwinter) entsprechend bis an die südliche Grenze. Dadurch kommt es zu niederschlagsreichen Zeiten in der Nähe des Zenitpunktes der Sonne und zu trockenen Zeiten weit von diesem Punkt entfernt. Zooplankton Tierisches Plankton PE 379.208 xxi Klimawandel und europäische Fischerei Verzeichnis der Abbildungen Abbildung 1: Maßgebliche Schlüsselprozesse und Kausalketten für die Dynamik der Meeresökosysteme und der Fischereiressourcen............ 2 Abbildung 2: Biologische Produktivität in Abhängigkeit von den Wassereigenschaften........................................................................................ 3 Abbildung 3: Veränderung der Temperatur, der Meereshöhe und der Schneebedeckung. ............................................................................. 11 Abbildung 4: Prognosen für die Oberflächenerwärmung.......................................... 14 Abbildung 5: Prognose der Versauerung der Ozeane.............................................. 15 Abbildung 6: Temperaturabweichungen................................................................... 18 Abbildung 7: Salinitätsabweichungen....................................................................... 20 Abbildung 8: Meeresströmungen in den nördlichen und subpolaren Becken........... 23 Abbildung 9: Globale Meeresströmungen. ............................................................... 24 Abbildung 10: Die Nordatlantik-Oszillation. ................................................................ 32 Abbildung 11: Auswirkungen von Temperaturextremen............................................. 37 Abbildung 12: Auswirkung der Temperatur auf die Kabeljaubestände....................... 40 Abbildung 13: Die Auswirkungen von Warmperioden auf die Verbreitung des Kabeljau. ............................................................................................. 44 Abbildung 14: Potenzielle ökologische Reaktionen auf den Klimawandel.................. 47 Abbildung 15: Einfluss des Klimawandels auf das Migrationsverhalten. .................... 49 Abbildung 16: Einfluss der Temperatur auf die Fischrekrutierung.............................. 57 Abbildung 17: Kausalzusammenhang zwischen Klima und Fischwachstum (Ostsee) .............................................................................................. 61 Abbildung 18: Kausalzusammenhang zwischen Klima und Fischwachstum (Mittelmeer)......................................................................................... 63 Abbildung 19: Kausalzusammenhang zwischen Klima und Fischwachstum (Nordsee). ........................................................................................... 65 Abbildung 20: Einfluss des Klimas auf die Nahrung für die Fischlarven..................... 67 PE 379.208 xxii Klimawandel und europäische Fischerei Verzeichnis der Tabellen Tabelle 1: Durch den Klimawandel beeinflusste Phänomene mit ihren spezifischen Trends, Wirkungen, Literaturangaben und Zuverlässigkeit..... xiii Tabelle 2: Auswahl von Hauptarten, relevante Verbreitungsgebiete und wirtschaftliche Angaben................................................................................. 6 Tabelle 3: Auswahl der wichtigsten Aquakulturarten, relevante Verbreitungsgebiete und wirtschaftliche Angaben............................................................. 9 Tabelle 4: Auswahl von Hauptarten und deren für die Studie relevanten Verbreitungsgebiete .................................................................................... 34 Tabelle 5: Auswahl wichtiger Arten, die in der Aquakultur eingesetzt werden, und die jeweiligen Gebiete, die im Rahmen der Studie von Bedeutung sind ..... 53 Tabelle 6: Dokumentierung der Ergebnisse der Literaturauswertung ........................ 114 PE 379.208 xxiii Klimawandel und europäische Fischerei Inhalt Zusammenfassung....................................................................................................... iii Glossar...................................................................................................................... xviii 1. Projekthintergrund .................................................................................................... 1 2. Konzeptioneller Rahmen .......................................................................................... 2 3. Literaturauswertung................................................................................................ 11 3.1 Erwartete Veränderungen bei den Wassereigenschaften ................................... 11 3.2 Erwartete Auswirkungen des Klimawandels auf die Hauptarten ......................... 34 3.3 Weitere damit zusammenhängende Fragen ....................................................... 53 4. Dokumentation und kritische Bewertung der Ergebnisse................................... 71 5. Schlussfolgerungen und Empfehlungen .............................................................. 75 ANHÄNGE .................................................................................................................... 83 Anhang 1. Biologische Merkmale der Arten .............................................................. 83 Anhang 2. Dokumentierung der Ergebnisse der Literaturauswertung....................... 97 Bibliography .............................................................................................................. 115 PE 379.208 xxiv Klimawandel und europäische Fischerei 1. Projekthintergrund Der Klimawandel wird aller Wahrscheinlichkeit nach verschiedene Prozesse auslösen, die sich auf die biologische Produktivität der Meeresökosysteme auswirken und Veränderungen in der Verbreitung der Meeresressourcen zur Folge haben können. In der Fischwirtschaft hat man es möglicherweise mit Auswirkungen erheblichen Ausmaßes zu tun, wobei die einzelnen Fischereiarten durch Veränderungen in der biologischen Produktivität der Meeresökosysteme unterschiedlich betroffen sein dürften. Ausschlaggebend sind die spezifischen Umweltveränderungen und die besonderen biologischen Merkmale der einzelnen Arten. Veränderungen in einem bestimmten marinen Umfeld können ein rasches Wachstum dort vorkommender hochwertiger Arten bewirken, während in anderen Fällen möglicherweise genau das Gegenteil eintritt. Mit der Fischerei wird ein Sektor von den Klimaveränderungen betroffen, der durch eine vollständige Nutzung der vorhandenen Ressourcen, eine erhebliche Überkapazität bei vielen Flottensegmenten sowie Konflikte unter den Fischern wie auch zwischen der Fischerei und anderen Nutzern mariner Ökosysteme gekennzeichnet ist. Verändert sich die Produktivität des Ökosystems, wird sich dies spürbar auf die Nachhaltigkeit der Fischerei auswirken, und klimatische Faktoren könnten durch negative Produktivitätsveränderungen die rückläufige Entwicklung verstärkt haben. Sie dürften nun die Erholung der Fischbestände beeinflussen. Klimaveränderungen sind Gegenstand sehr umfangreicher Forschungsarbeiten, die von verschiedenen Gruppen durchgeführt werden, darunter Regierungsstellen, Hochschulen und Schwerpunktgruppen. Die gewonnenen Erkenntnisse können bei der Einschätzung der künftigen Entwicklung der Fischbestände und für die Quantifizierung der Zusammenhänge genutzt werden. Vor diesem Hintergrund ist es das Anliegen dieser Studie, dem Fischereiausschuss in einer klaren und detaillierten Analyse aufzuzeigen, wie sich der Klimawandel in der Fischwirtschaft der Europäischen Union auswirken kann. Grundlage der Studie ist eine Auswertung der vorhandenen einschlägigen Literatur unter besonderer Berücksichtigung der Fischereiressourcen, die für die Fischerei der Europäischen Union im Atlantik und in den angrenzenden Meeren (Mittelmeer, Nord- und Ostsee) von Bedeutung sind. Die Studie beinhaltet einen konzeptionellen Rahmen mit einer Beschreibung der Schlüsselprozesse, die die Dynamik der Ökosysteme und der Fischereiressourcen bestimmen. Mit dessen Hilfe wurden die Literaturquellen zu den Auswirkungen des Klimawandels auf die für die Fischerei der Europäischen Union wichtigsten Arten systematisch ausgewertet. Dabei ging es speziell um solche Elemente wie Habitat, Verfügbarkeit der Nahrung, Fressfeinde, Krankheitserreger und Konkurrenten, die möglicherweise die Ausbreitung von Arten einschränken. Außerdem enthält die Analyse eine Beschreibung von Wassereigenschaften wie Temperatur, Salinität, Gewässerstruktur, Strömungen, Alkalinität (Versauerung der Ozeane) oder Sauerstoffverfügbarkeit. Spezifische Auswirkungen auf halbumschlossene Meere wie die Ostsee, das Mittelmeer, das Ägäische Meer und das nördliche Adriatische Meer finden ebenfalls Berücksichtigung.. 1 PE 379.208 Klimawandel und europäische Fischerei 2. Konzeptioneller Rahmen Der konzeptionelle Rahmen dient als Grundlage für die Festlegung der Schwerpunkte bei einer zielorientierten Literaturdurchsicht, der Dokumentation und Bewertung der Ergebnisse und der Erarbeitung von Schlussfolgerungen. Bei diesem Rahmen wird von den für die Dynamik der Meeresökosysteme und der Fischereiressourcen maßgeblichen Schlüsselprozessen und Kausalketten ausgegangen. Diese werden in Abbildung 1 veranschaulicht. Abbildung 1: Maßgebliche Schlüsselprozesse und Kausalketten für die Dynamik der Meeresökosysteme und der Fischereiressourcen Klimawandel ∆ Temperatur ∆ Niederschlag ∆ Sonneneinstrahlung ∆ Wetterextreme ∆ Meeresspiegelhöhe ∆ Industrielle Fischerei Abiotic Abiotische environmental Umweltbedingungen* conditions Biotische Umweltbedingungen** Kommerziell genutzte Fischpopulation 1 Kommerziell genutzte Fischpopulation 2 bis X Quelle: FAO Statistics 2005 und ICCAT 2005. * ** Die nichtlebende Umwelt, bestehend aus allen abiotischen Faktoren, z. B. Temperatur, Salinität. Die lebende Umwelt, bestehend aus allen biotischen Faktoren, z. B. Nahrung, Fressfeinde. Wichtige Triebkräfte für die Entwicklung der Fischereiressourcen, die im Rahmen der Studie behandelt werden, sind die durch Klimaveränderungen hervorgerufenen spezifischen Umweltbedingungen. Dabei sollte beachtet werden, dass die Auswirkungen von Umweltveränderungen auf Fischpopulationen ganz erheblich von der Größe und der Zusammensetzung der betreffenden Bestände abhängen, weshalb sie also stark von der Fischerei beeinflusst werden. Da sich die Triebkräfte direkt oder indirekt auf kommerziell genutzte PE 379.208 2 Klimawandel und europäische Fischerei Fischpopulationen auswirken, muss die künftige Entwicklung der Fischereiressourcen unter Berücksichtigung möglicher Entwicklungen beim Klima und bei der industriellen Fischerei betrachtet werden. Mit dem Klima verändern sich auch spezifische Umweltbedingungen, insbesondere Temperatur, Niederschlag, Sonneneinstrahlung, Wetterextreme und Meeresspiegelhöhe. Diese spezifischen Umweltbedingungen beeinflussen direkt die abiotischen Bedingungen in Meeresökosystemen. Für das Projekt und im Hinblick auf diese Systeme sind die Wassereigenschaften von besonderer Bedeutung, vor allem Temperatur, Salinität, Alkalinität, Sauerstoffverfügbarkeit, Gewässerstruktur und Strömungen. Die grundlegende Kausalkette, mit der der Klimawandel mit dem Meeresökosystem in Verbindung gebracht wird, lässt sich wie folgt darstellen: Veränderung spezifischer Umweltbedingungen Klimawandel Veränderung abiotischer Bedingungen in Meeresökosystemen Veränderungen in den spezifischen Umweltbedingungen können ermittelt und Veränderungen der abiotischen Bedingungen in Meeresökosystemen, d. h. Wassereigenschaften, in Verbindung gebracht werden. So lassen sich entsprechend Szenarien für die Klimaänderung Prognosen zur künftigen Entwicklung Wassereigenschaften aufstellen. mit den den der Jede Art hat biologische Merkmale hinsichtlich ihres Habitats( 25), die während ihres Lebenszyklus variieren können. In der Meeresumwelt sind die Wassereigenschaften die relevantesten biologischen Merkmale, da jede Fischart an ganz bestimmte Bereiche bei Temperatur, Salinität, Struktur, Strömungen und Sauerstoffverfügbarkeit angepasst ist. Bei diesen Merkmalen werden Bereiche definiert, innerhalb derer eine Art unter Minimal- bis Optimalbedingungen überleben kann. Entsprechend den biologischen Merkmalen der Art gibt es eine optimale Kombination der verschiedenen Wassereigenschaften, bei der ein Maximum an biologischer Produktivität erreicht wird. Anhand der physiologischen Anforderungen der wichtigsten Arten lassen sich die Umweltauswirkungen auf die physiologische Leistung, das Migrationsverhalten und den Reproduktionserfolg einschätzen. Abbildung 2: Biologische Produktivität in Abhängigkeit von den Wassereigenschaften Biologische Produktivität Maximum an biologischer Produktivität Überlebensgrenzen Bereiche bei den Wassereigenschaften 25 Als Habitat wird das physische Umfeld angesehen, das eine Population umgibt. 3 PE 379.208 Klimawandel und europäische Fischerei Ausgehend davon ist eine Ergänzung der grundlegenden Kausalkette möglich: Klimawandel Veränderung spezifischer Umweltbedingungen Veränderung abiotischer Bedingungen in Meeresökosystemen Veränderung der biologischen Produktivität der wichtigsten Arten Die Wassereigenschaften stehen auch in direktem Zusammenhang mit der biologischen Produktivität der biotischen Umwelt( 26), d. h. aller Meeresorganismen einschließlich der befischten Populationen und aller anderen Organismen, die in Wechselwirkung mit den Fischpopulationen stehen, wie etwa Nahrungsorganismen, Konkurrenten, Fressfeinde oder Krankheitserreger. Diese Wechselwirkungen und deren Beeinflussung durch die Fischerei müssen entsprechend berücksichtigt werden, wenn letztendlich die Auswirkungen des Klimawandels auf die industrielle Fischerei beurteilt werden sollen. Die grundlegende Kausalkette, die die Beziehung zwischen Klimawandel und industrieller Fischerei herstellt, kann vervollständigt werden: Klimawandel Veränderung spezifischer Umweltbedingungen Veränderung der biol. Produktivität der biotischen Umwelt Veränderung abiotischer Bedingungen in Meeresökosystemen Veränderung der biologischen Produktivität der wichtigsten Arten Veränderung des Rekrutierungserfolgs der Fischereiressourcnen Fischereitätigkeit Der geographische Erfassungsbereich der Studie ist so gewählt, dass die für die Fischerei der Europäischen Union relevanten Ressourcen im Atlantik und den angrenzenden Meeren (Mittelmeer, Nord- und Ostsee) einbezogen sind. Außerdem sollen spezifische Auswirkungen auf halbumschlossene Meere wie die Ostsee, das Mittelmeer, das Ägäische Meer und das nördliche Adriatische Meer berücksichtigt werden. Innerhalb dieses untersuchten Gebietes variieren die Wassereigenschaften ganz beträchtlich, und auch mögliche Veränderungen bei diesen Eigenschaften werden unterschiedlich ausfallen. Das wiederum wirkt sich auf die geographische Verteilung der Fischressourcen aus. In diesem (26) Die lebende Umwelt bestehend aus allen biotischen Faktoren, z. B. Nahrung, Fressfeinden. PE 379.208 4 Klimawandel und europäische Fischerei Zusammenhang müssen bei den Literaturrecherchen auch folgende Aspekte berücksichtigt werden: • Prozesse in den Grenzbereichen zwischen Warm- und Kaltwassermassen • Veränderungen in der Position der ozeanischen Fronten, wo Wassermassen mit unterschiedlichen Merkmalen aufeinandertreffen, ihre Rolle als Barrieren für die Ausbreitung sowohl der Wassermassen selbst als auch des Planktons und der Fische und die Auswirkungen dieser Veränderungen • Verschiebung von Übergangszonen( 27) und gleichzeitige Diskontinuität beim lokalen Auftreten einiger Arten • Veränderungen in der Abundanz und Verteilung bestehender und neuer Populationen • Veränderungen in der Ausweitung der geographischen Verbreitungsgebiete oder in der geographischen Verteilung von Fischpopulationen im Zusammenhang mit hydroklimatischen Schwankungen und einer regionalen Klimaerwärmung • Effekte wie längere Wachstumsperioden, geringere natürliche Wintersterblichkeit und höhere Wachstumsraten • Nachweise und Prognosen möglicher lokaler Veränderungen bei Produktionszentren und Artenmischungen sowie geographische Verschiebungen und Veränderungen innerhalb der Ökosysteme • Änderung etablierter Reproduktionsmuster, Migrationsrouten und Ökosystembeziehungen • Eintritt exotischer Fische in das Ökosystem und mögliche Veränderungen in der Artenzusammensetzung und der Konkurrenzsituation • Veränderung der Verteilungs- und Migrationsmuster und deren Einfluss auf die Artenmischung, die Nahrungskonkurrenz und die gesamte Nahrungskette sowie die Folgen von Gebietsüberschneidungen bei verwandten Arten • Auswirkungen einer regionalen Klimaerwärmung auf die Rekrutierung durch Veränderungen an der Basis der Nahrungskette, Ermittlung derjenigen von den Hauptarten, die während des pelagischen Larvenstadiums besonders sensibel auf die Verfügbarkeit und Zusammensetzung der planktonischen Nahrung reagieren • Auswirkungen des Klimawandels auf die Aquakultur. Die Ergebnisse der Literaturrecherche werden in Kapitel 3 unter drei Unterüberschriften dokumentiert: • 3.1 Erwartete Veränderungen bei den Wassereigenschaften; • 3.2 Erwartete Auswirkungen des Klimawandels auf die Hauptarten; • 3.3 Sonstige Fragen. Die künftige Entwicklung der Wassereigenschaften im untersuchten Gebiet entscheidet über die künftige Verfügbarkeit und Verteilung der Fischereiressourcen. Folglich lag einer der Schwerpunkte bei der Literaturauswertung auf den Eigenschaften des Wassers (siehe Kapitel 3.1) , wobei insbesondere folgende spezifische Aspekte berücksichtigt wurden: • Prognosen/Modelle für den Klimawandel und Herstellung von Beziehungen zu spezifischen Umweltbedingungen in aquatischen Ökosystemen, vor allem zu den Wassereigenschaften; (27) In der Biogeographie eine Zone zwischen zwei deutlich unterschiedlichen Habitaten. Oftmals eine Barriere für die Verbreitung. 5 PE 379.208 Klimawandel und europäische Fischerei • • • • Auswertung der aus den Modellen erlangten Informationen über Veränderungen der derzeitigen Gegebenheiten, vor allem über die möglichen Prozesse in den Grenzbereichen zwischen den Warm- und Kaltwassermassen; Beschreibung der möglichen Veränderungen in der Position der ozeanischen Fronten, wo Wassermassen mit unterschiedlichen Merkmalen aufeinandertreffen, ihrer Rolle als Barrieren für die Ausbreitung sowohl der Wassermassen selbst als auch des Planktons und der Fische und der Auswirkungen dieser Veränderungen; Untersuchung der Auswirkungen der Erwärmung auf der Grundlage der Erkenntnisse über Verschiebungen von Übergangszonen und gleichzeitige Diskontinuität beim lokalen Auftreten einiger Arten; Spezifische Effekte in halbumschlossenen Meeren. Wie bereits erwähnt, ist die biologische Produktivität befischter Populationen über Veränderungen der Wassereigenschaften und der Merkmale der Fischarten mit dem Klimawandel verbunden. Angesichts der Komplexität dieser Problematik ist es sinnvoll, bei der Literaturauswertung den Schwerpunkt auf einige Hauptarten zu legen, die für die Fischerei in der Europäischen Union von Bedeutung sind. Die Hauptarten sollten das untersuchte Gebiet adäquat repräsentieren, um so mögliche Auswirkungen für die Fischerei einschätzen zu können. Tabelle 3 enthält eine Auswahl wichtiger Industriefischarten, deren Relevanz sich aus den Fangmengen und der geographischen Verbreitung herleitet, sowie einige wirtschaftliche Angaben zu den einzelnen Arten. Daraus geht hervor, dass bei den ausgewählten Arten die Fänge im untersuchten Gebiet einem Marktwert von etwa 2,3 Mrd. EUR entsprachen. Die Preise sind Orientierungspreise der Europäischen Kommission von 2005 und entsprechen nicht unbedingt dem in diesem Jahr erzielten tatsächlichen Marktpreis. Tabelle 2: Angaben Auswahl von Hauptarten, relevante Verbreitungsgebiete und wirtschaftliche Hauptarten Für die Studie relevante Verbreitungsgebiete Anlandunge n 2005( 28) (Tonnen) Orientierung s-preis 2007 (Euro/t) Wert (Mio. EUR) Anteil (%) Hering Nordostatlantik, Nordsee, Ostsee 829.764 260 215,7 9,6 Sprotte Nordostatlantik, Nordsee, Ostsee, Mittelmeer 703.119 160 112,5 5,0 Sardine Nordostatlantik, mittlerer Ostatlantik, Mittelmeer 245.168 587 143,9 6,4 Sardelle Nordostatlantik, mittlerer Ostatlantik, Mittelmeer 111.403 1270 141,5 6,3 Makrele Nordostatlantik, Mittelmeer 289.994 314 91,1 4,1 Roter Thun Atlantik, Mittelmeer 20.727 1500( 29) 31,1 1,4 Blauer Wittling Nordsee, Mittelmeer 462.258 946 437,3 19,5 Kabeljau Nordostatlantik, Nordsee, Ostsee 125.245 1615 202,3 9,0 (28) Anlandungen in den europäischen Ländern aus dem Nordostatlantik, dem mittleren Ostatlantik, dem Mittelmeer und dem Schwarzen Meer; Daten zu den Anlandungen 2005 von FAO Fisheries Department, Fishery Information, Data and Statistics Unit, entnommen aus der Datenbank FISHSTAT Plus (Universal software for fishery statistical time series Version 2.3 2000). (29) Preis von ICCAT (Internationale Kommission für die Erhaltung der Thunfischbestände des Atlantiks), 2005. PE 379.208 6 Klimawandel und europäische Fischerei Hauptarten Für die Studie relevante Verbreitungsgebiete Anlandunge n 2005( 28) (Tonnen) Orientierung s-preis 2007 (Euro/t) Wert (Mio. EUR) Anteil (%) Europäischer Seehecht Nordostatlantik, Nordsee, mittlerer Ostatlantik, Mittelmeer 76.252 3731 284,5 12,7 Sandaal Nordsee 166.755 1000( 30) 166,8 7,4 Schellfisch Nordostatlantik 44.533 983 43,8 1,9 Seelachs Nordostatlantik 59.377 751 44,6 2,0 Scholle Nordostatlantik, Nordsee 66.566 1499 99,8 4,4 30 Ohrensardine mittlerer Ostatlantik, Mittelmeer 90.282 1000( ) 90,3 4,0 Stöcker Nordostatlantik, Mittelmeer 177.345 400(30) 70,9 3,2 Echter Bonito Gelbflossenthun Atlantik, Mittelmeer Atlantik, Mittelmeer Gesamt 29 46.453 630( ) 29,3 1,3 35.214 29 42,5 1,9 2.248 100 3.552.821 1207( ) Quelle: FAO Statistics 2005 und ICCAT 2005. Die Anlandungen der industriellen Fischerei in den EU-15 betrugen 2004 annähernd 4,5 Mrd. t und hatten einen entsprechenden Marktwert von etwa 6 Mrd. EUR (Fakten und Zahlen über die GFP, Europäische Kommission 2006). Diese Zahlen beziehen sich auf alle Anlandungen in europäischen Häfen, unabhängig von ihrem Ursprung. In der vorliegenden Studie werden die wirtschaftlichen Auswirkungen für das untersuchte Gebiet diskutiert (bei dem sich der Marktwert auf rund 2 Mrd. EUR beläuft). Dabei ist jedoch anzumerken, dass der Klimawandel an den Grenzen anderer Fanggebiete nicht Halt macht und entsprechende Auswirkungen folglich auch außerhalb des untersuchten Gebiets anzutreffen sind. Die künftigen Entwicklungen bei den Hauptfischarten lassen sich ausgehend von deren biologischen Merkmalen abschätzen. Je nach ihrer physiologischen Plastizität werden die Arten die Veränderungen tolerieren oder aber ihre Produktivität erhöhen bzw. verringern. Organismen können in solche Gebiete wandern, die für ihre Ansprüche besser geeignet sind. Dadurch kann es zu lokalen Veränderungen bei den Produktionszentren und zu geographischen Verschiebungen kommen, was sich auf die Verteilung der gemeinsamen Fanggebiete auswirkt. Niedrigere Wintertemperaturen, wie sie in den Szenarien zum Klimawandel vorausgesagt werden, könnten die Wachstumsperioden verlängern und damit zu einem Anstieg der Wachstumsrate und der Produktivität führen. Höhere Wintertemperaturen könnten die Wintersterblichkeit verringern und damit ebenfalls einen Produktivitätsanstieg bewirken. Um diese Beziehungen beurteilen zu können, müssen Informationen zu den biologischen Merkmalen der wichtigsten Arten zusammengetragen und analysiert werden. So lässt sich feststellen, ob die betreffende Art bereits ihre Überlebensgrenze erreicht hat und es zu einer veränderten Abundanz und Verteilung kommen wird. Die ökologische Nische einer Fischart hinsichtlich Temperatur, Salinität, Alkalinität oder Sauerstoffverfügbarkeit lässt sich anhand von Kriterien bestimmen, die die mögliche tödliche Wirkung dieser Nische, ihre kontrollierende (Physiologie) und ihre ausrichtende (Verhalten) Wirkung betreffen. Dabei haben die Sterblichkeitskriterien in der Regel breitere Spannen, wohingegen die Kontroll- und insbesondere die Ausrichtungskriterien sehr enge Spannen aufweisen können. Wenn Fische auf Habitate mit ungeeigneten abiotischen Bedingungen treffen, reagieren sie darauf mit einem speziellen Verhalten, indem sie versuchen, (30) Geschätzter Preis. 7 PE 379.208 Klimawandel und europäische Fischerei sich in Gebiete zu begeben, die eher die von ihnen bevorzugten biologischen Merkmale aufweisen. In der Folge wird damit gerechnet, dass durch den Klimawandel ausgelöste Veränderungen in den abiotischen Bedingungen zu Veränderungen bei Abundanz und Verteilung der Industriefischarten führen. Durch Verschiebungen bei der Artenzusammensetzung in einer bestimmten Umgebung können sich auch die Wechselwirkungen innerhalb des Ökosystems verändern. Dies führt zu Veränderungen in den Beziehungen Fressfeind/Beutetier und in der Konkurrenz beispielsweise um Nahrung oder günstige Habitate. Auf derartige Wechselwirkungen wird unter anderem in der Literaturauswertung in Kapitel 3.3 eingegangen. Kapitel 3.2 betrifft die Literaturrecherche zu den erwarteten Auswirkungen des Klimawandels auf die wichtigsten Arten, die für die EU-Fischerei im untersuchten Gebiet von Belang sind. Entsprechend dem konzeptionellen Rahmen werden bei der Recherche die folgenden Aspekte berücksichtigt: • Biologische Merkmale der wichtigsten Industriefischarten; • Abschätzung möglicher Veränderungen in der Größe und Verteilung bestehender und neuer Populationen; • Auswertung der Nachweise und mögliche Veränderungen in der Ausweitung der geographischen Verbreitungsgebiete oder in der geographischen Verteilung von Fischpopulationen im Zusammenhang mit hydroklimatischen Schwankungen und einer regionalen Klimaerwärmung; • Abschätzung und Lokalisierung solch möglicher Effekte wie längere Wachstumsperioden, geringere natürliche Wintersterblichkeit und höhere Wachstumsraten; • Beschreibung der Lokalisierung von Änderungen bei etablierten Reproduktionsmustern, Migrationsrouten und Ökosystembeziehungen; • Beschreibung der Nachweise und Prognosen für mögliche lokale Veränderungen bei Produktionszentren und Artenmischungen sowie geographische Verschiebungen und Veränderungen innerhalb der Ökosysteme. Für die Einschätzung der künftigen Entwicklung der biologischen Produktivität der einzelnen Arten müssen die maßgeblichsten Elemente der biotischen Umgebung wie Nahrungsmittelangebot, Konkurrenten, Fressfeinde oder Krankheitserreger in Betracht gezogen werden. Eine Beurteilung der weiteren Entwicklung der Hauptfischarten und der Elemente der biotischen Umgebung lässt sich auf der Grundlage der biologischen Merkmale der einzelnen Arten (Fische und Arten der biotischen Umgebung) vornehmen. Bei der Literaturauswertung werden noch weitere relevante Punkte berücksichtigt. Die industrielle Fischerei hat direkte Folgen für die biologische Produktivität der Hauptfischarten und deren Fähigkeit zur Anpassung an Umweltveränderungen. Folglich wird die künftige Fischereitätigkeit über die weitere Verfügbarkeit der Fischereiressourcen entscheiden. Infolge des Klimawandels kann es zu Veränderungen der Migrationsrouten sowie zu Veränderungen und Ausweitungen der Verbreitungsgebiete der Arten kommen, wodurch möglicherweise exotische Arten in das Ökosystem eindringen. Durch die Invasion exotischer Arten wird unter Umständen ein Kampf um Lebensraum und Nahrungsmittel einsetzen, bei dem ursprünglich im Habitat angesiedelte Arten verdrängt werden könnten. Es muss mit Veränderungen in der Nahrungskette gerechnet werden. Außerdem beeinflusst der Klimawandel die Verteilung und das Auftreten von Krankheitserregern und Parasiten. Darüber hinaus können sich die Umweltveränderungen auch auf die Arten auswirken, die in der Aquakultur und der damit verbundenen Industrie eingesetzt werden. PE 379.208 8 Klimawandel und europäische Fischerei Tabelle 3 enthält eine Auswahl der wichtigsten Arten in der Aquakultur, deren Relevanz sich aus der Produktion und dem Verbreitungsgebiet herleitet, sowie einige wirtschaftliche Angaben zu den einzelnen Arten. Daraus geht hervor, dass bei den ausgewählten Arten ein Marktwert von etwa 2 Mrd. EUR erzielt wurde. Tabelle 3: Auswahl der wichtigsten Aquakulturarten, relevante Verbreitungsgebiete und wirtschaftliche Angaben Hauptarten Für die Studie relevante Verbreitungsgebiete Produktion 2005 (Tonnen) 144.778,00 70.010,00 Preis Wert (Euro/t) (Mio. Euro) 324,37 446,33 251,71 278,14 Anteil (Wert) (%) 22,47 14,00 Lachs Goldbrasse Nordostatlantik Nordostatlantik, Mittelmeer Pazifische Auster Nordostatlantik, Mittelmeer, Schwarzes Meer 127.150,00 471,17 269,86 13,59 Miesmuschel Wolfsbarsch Nordostatlantik Nordostatlantik, Mittelmeer und Schwarzes Meer 361.399,00 47.982,00 1.372,42 205,31 263,33 233,70 13,26 11,77 Teppischmuschel Nordostatlantik, Mittelmeer und Schwarzes Meer 68.006,00 332,29 204,66 10,31 Mittelmeer-Miesmuschel Mittelmeer und Schwarzes Meer Nordostatlantik Nordostatlantik, Mittelmeer und Schwarzes Meer 108.522,00 1.083,49 100,16 5,04 23.664,00 3.665,00 418,83 89,50 56,50 40,95 2,84 2,06 Steinbutt Große Teppichmuschel Nordostatlantik Nordostatlantik, Mittelmeer und Schwarzes Meer 6.792,00 6.282,00 171,95 206,44 39,50 30,43 1,99 1,53 Europäische Auster Europäischer Aal Nordostatlantik Nordostatlantik, Mittelmeer und Schwarzes Meer 4.647,00 1.122,00 357,19 118,86 13,01 9,44 0,66 0,48 1.986,01 100,00 Regenbogenforelle Roter Thun 974.019,00 Gesamt Quelle: FAO Statistics 2005. 2003 erzielte die Aquakultur in den EU-25 eine Produktion von rund 1,4 Mrd. t, die einem Marktwert von etwa 2,8 Mrd. EUR entsprachen (Fakten und Zahlen über die GFP, Europäische Kommission 2006). Bei diesen Angaben ist die gesamte Aquakulturproduktion, einschließlich Süßwasser, berücksichtigt. Im Rahmen der vorliegenden Studie werden die wirtschaftlichen Auswirkungen für die Meeresaquakultur diskutiert (bei der sich der Marktwert auf rund 2 Mrd. EUR beläuft). Kapitel 3.3 betrifft die Literaturrecherche zu anderen relevanten Fragen im untersuchten Gebiet. Bei der Recherche werden die folgenden Aspekte berücksichtigt: • Biologische Merkmale relevanter Arten aus der biotischen Umgebung wichtiger Industriefischarten (z. B. Nahrungsorganismen, Konkurrenten, Fressfeinde) sowie 9 PE 379.208 Klimawandel und europäische Fischerei • • • • • • • biologische Merkmale möglicherweise in das untersuchte Gebiet vordringender exotischer Fische und der wichtigsten Aquakulturarten. Prognosen zum möglichen Eindringen exotischer Fische in das Ökosystem und zu den wahrscheinlichen Veränderungen in der Artenzusammensetzung und der Konkurrenz. Erwartete Auswirkungen des Klimawandels auf die biologische Produktivität und ihr saisonales Auftreten. Einfluss der Umweltbedingungen und der Fischerei auf den Rekrutierungserfolg bei den Hauptarten. Auswirkungen der künftigen Fischereitätigkeit. Untersuchung der möglichen Veränderungen bei den Verteilungs- und Migrationsmustern und deren Einfluss auf die Artenmischung, die Nahrungskonkurrenz und die gesamte Nahrungskette sowie der Folgen von Gebietsüberschneidungen bei verwandten Arten. Analyse der Auswirkungen einer regionalen Klimaerwärmung auf die Rekrutierung durch Veränderungen an der Basis der Nahrungskette, Ermittlung derjenigen von den Hauptarten, die während des pelagischen Larvenstadiums besonders sensibel auf die Verfügbarkeit und Zusammensetzung der planktonischen Nahrung reagieren. Belege für wahrscheinliche Auswirkungen des Klimawandels auf die Aquakultur. Die Ergebnisse der Literaturrecherche werden zusammengefasst und unter dem Gesichtspunkt der Auswirkungen des Klimawandels auf die europäische Fischerei und die Meeresökosysteme kritisch analysiert. Außerdem wird eingeschätzt, welche Konsequenzen der Klimawandel für die EU-Fischerei haben könnte.. PE 379.208 10 Klimawandel und europäische Fischerei 3. Literaturauswertung 3.1 Erwartete Veränderungen bei den Wassereigenschaften Prognosen für den Klimawandel Der Klimawandel gilt als eine der Gefahren für die biologische Vielfalt sowie die Struktur und Funktionsweise von Ökosystemen (McCarthy et al., 2001; Vitousek, 1994). Obwohl über Ursachen und mögliche Auswirkungen nach wie vor diskutiert wird (Sharp, 2003; Veizer, 2005), ist sich die Wissenschaft einig, dass der Klimawandel Realität ist (Houghton et al. 2001; Karl und Trenberth 2003; King 2004; Walther et al. 2005, IPCC-Bericht 2007). Anzeichen für die Erwärmung des Klimasystems sind Veränderungen der Oberflächen- und Lufttemperaturen, eine Abnahme der Schneebedeckung in der nördlichen Hemisphäre, die Temperaturen in den oberen Ozeanschichten bis zu mehreren hundert Meter Tiefe und der Anstieg des Meeresspiegels (IPCC-Bericht 2007, Abbildung 3), die Versauerung der Ozeane (Caldeira and Wicket 2003, 2005) und Veränderungen der Salinität (MCCIP 2006). Abbildung 3: Veränderung der Temperatur, der Meereshöhe und der Schneebedeckung. Quelle: IPCC 2007. Anmerkungen: Beobachtete Veränderungen bei der (a) globalen Durchschnittsoberflächentemperatur; dem (b) globalen durchschnittlichen Meeresspiegelanstieg anhand von Pegelstandsdaten (blau) und Satellitendaten (rot) und der (c) Schneebedeckung in der nördlichen Hemisphäre im März/April. Alle Veränderungen beziehen sich auf die entsprechenden Durchschnittswerte im Zeitraum 1961-1990. Die geglätteten Kurven veranschaulichen die zehnjährigen mittleren Werte, die Kreise zeigen die jährlichen Werte. Die schattierten Flächen sind die Unsicherheitsintervalle, die anhand einer umfassenden Analyse bekannter Unsicherheiten (a und b) sowie mit Hilfe von Zeitreihen (c) geschätzt wurden.. 11 PE 379.208 Klimawandel und europäische Fischerei Legende: Changes in temperature … Global average temperature Global average sea level Northern hemisphere snow cover Temperature Difference from … Year million Veränderung der Temperatur, der Meereshöhe und der Schneebedeckung in der nördlichen Hemisphäre Globale Durchschnittstemperatur Globale durchschnittliche Meereshöhe Eisbedeckung in der nördlichen Hemisphäre Temperatur Unterschied zu … Jahr Mio. (I) Die Rolle der Treibhausgase Der in letzter Zeit beobachtete Anstieg der globalen Temperatur hängt größtenteils mit dem verstärkten Auftreten verschiedener atmosphärischer Gase zusammen. So haben sich infolge der menschlichen Tätigkeiten die Konzentrationen von Kohlendioxid, Methan und Distickstoffoxid in der Luft seit 1750 überall in der Welt deutlich erhöht und liegen mittlerweile weit über den vorindustriellen Werten, zu deren Ermittlung Eiskerne benutzt wurden, die Aufschluss über Tausende von Jahren geben (Houghton et al. 2001; Hulme et al. 2002; IPCC-Bericht 2007, Raper et al. 1997, Siegenthaler et al. 2005; Spahni et al. 2005). Diese so genannten Treibhausgase kommen von Natur aus in der Atmosphäre vor und sind wichtig für das Leben auf der Erde. Kohlendioxid (CO2) ist das wichtigste anthropogene Treibhausgas, seine Konzentration in der Luft hat sich von einem vorindustriellen Wert von etwa 280 ppm auf 379 ppm im Jahre 2005 erhöht (IPCC 2007). Die Konzentration ist damit deutlich höher als während der vorangegangenen 650 000 Jahre (180 bis 300 ppm), wie anhand von Eiskernen festgestellt wurde (Siegenthaler et al. 2005). Die jährliche Zunahme der Kohlendioxidkonzentration war während der letzten zehn Jahre höher (Durchschnitt 1995–2005: 1.9 ppm pro Jahr) als im gesamten Zeitraum seit Beginn kontinuierlicher Luftmessungen (Durchschnitt 1960–2005: 1,4 ppm pro Jahr, IPCC 2007). Die Treibhausgase bewirken, dass die kurzwellige Sonnenstrahlung die Atmosphäre passiert und die Erde erwärmt, wobei ein Teil der Strahlung absorbiert und nachfolgend von der erwärmten Erdoberfläche wieder abgestrahlt wird (Kiehl und Trenberth 1997). Anthropogene Treibhausgase tragen dazu bei, dass verstärkt Energie zurückbehalten wird (verstärkter Treibhauseffekt), dass sich der Planet erwärmt und die globale Temperatur ansteigt (Karl und Trenberth 2003; Raper et al. 1997; Ruddiman und Thomson 2001). Im Endergebnis ist es also seit 1750 durch die menschlichen Tätigkeiten zu einer weltweiten Erwärmung gekommen, die Strahlungsbilanz( 31) beträgt 1,6 Wm-2. Die Gesamtstrahlungsbilanz liegt wegen des Anstiegs von Kohlendioxid, Methan und Distickstoffoxid bei 2,30 Wm-2. Eine Zuwachsrate wie während des Industriezeitalters dürfte es in mehr als 10 000 Jahren nicht gegeben haben. Beim Kohlendioxid wurde von 1995 bis 2005 eine Zunahme um 20 % festgestellt, das ist die größte Veränderung innerhalb eines Jahrzehnts zumindest in den letzten 200 Jahren (IPCC-Bericht 2007). (31) „Strahlungsbilanz“ ist definiert als die Differenz zwischen der ankommenden und der abgehenden Strahlungsenergie in einem bestimmten Klimasystem. Eine positive Bilanz (mehr ankommende Energie) führt in der Regel zu einer Erwärmung des Systems, während eine negative Bilanz (mehr abgehende Energie) eine Abkühlung bewirkt. PE 379.208 12 Klimawandel und europäische Fischerei Die Emissionen von Treibhausgasen nehmen weiter zu, und obwohl es unklar ist, wie sich das globale Klima in Zukunft verändern wird, werden international mit großem Aufwand Prognosen zu den künftigen Klimabedingungen angestellt (Houghton et al. 2001, Hulme et al. 2002, IPCC 2007). (II) Temperatur Die globalen Durchschnittstemperaturen sind während der vergangenen 100 Jahre um ~0.6 °C angestiegen (Houghton et al. 2001; Hulme et al. 2002). In diesem Zeitraum haben sich sowohl die Meeres- (Levitus et al. 2000) als auch die Süßwassersysteme (Winder und Schindler, 2004) erwärmt. Die Prognosen sprechen von einer weiteren Erwärmung des Küstenklimas in Europa im Verlaufe des 21. Jahrhunderts, wobei mit einer Erhöhung der Meeresoberflächentemperatur um 0,2 °C in jedem Jahrzehnt gerechnet wird (IPCC 2007, Abbildung 4). Ozeanographische Studien dass sich die mittleren jährlichen zur Erforschung der Ostsee zeigen, Meeresoberflächentemperaturen bis zum Ende des 21. Jahrhunderts um 2° bis 4 °C erhöhen könnten (BACC 2006). Im westlichen Mittelmeerbecken wurde während der letzten 20 bis 30 Jahre ein deutlicher Anstieg der durchschnittlichen Wassertemperaturen festgestellt (Francour et al. 1994). Im IPCC-Bericht heißt es auch, dass die tendenzielle Erwärmung auf den Kontinentalschelfen, also dort, wo die kommerziell wichtigen Fischbestände leben, rascher voranschreitet als im offenen Ozean. Der Klimawandel verursacht in den Ozeanen bereits spürbare Veränderungen des pH-Werts, der Temperatur, des Zirkulationsverhaltens und der Nährstoffbelastung infolge von Stürmen (IPCC 2007). Der Sommer 2003 war möglicherweise Europas heißester Sommer seit mehr als 500 Jahren. Temperaturen, die 3 °C-6 °C über den langjährigen Mitteln lagen, und eine jährliche Niederschlagsmenge von nur 50 % des Durchschnitts führten in Europa zu einer Verringerung der Brutto-Primärproduktivität um 30 % (Ciais et al. 2005, Schär et al. 2004). Allein in zwei Wochen im August 2003 starben in Europa 22 000 bis 45 000 Menschen an den Folgen der Hitze (Patz et al. 2005). Diese Hitzewelle übertraf das erwartete Ausmaß bei den Klimaschwankungen bei weitem und ließ sich nicht nur mit natürlichen Triebkräften erklären. (Stott et al. 2004) gelangten zu der Schlussfolgerung, dass eine starke Sommererwärmung durch den Menschen selbst verursacht wird und solche Sommer wie 2003 in Zukunft wahrscheinlich häufiger vorkommen werden. Ihren Prognosen zufolge wird der Sommer 2003 am Ende des Jahrhunderts als ein ungewöhnlich kalter Sommer im Vergleich zu den neuen Klimabedingungen eingestuft werden. 13 PE 379.208 Klimawandel und europäische Fischerei Abbildung 4. Prognosen für die Oberflächenerwärmung. Quelle: IPCC 2007. Anmerkungen: Die durchgehenden Linien zeigen die aus mehreren Modellen gewonnenen globalen Durchschnitte für die Oberflächenerwärmung (in Bezug auf 1980-1999) für die Szenarien A2, A1B und B1, jeweils in Fortsetzung der Simulationen für das 20. Jahrhundert. Die Schattierung kennzeichnet den Bereich der Standardabweichung von plus/minus Eins bei den Jahresdurchschnitten der einzelnen Modelle. Die orangene Linie steht für das Experiment, bei dem die Konzentrationen konstant auf den Werten des Jahres 2000 gehalten wurden. Die grauen Balken rechts veranschaulichen jeweils die bestmögliche Schätzung (waagerechte Linie in jedem Balken) und die mögliche Spanne, die nach Einschätzung für die sechs SRESMarker-Szenarien ( 32) möglich sind. Bei der Festlegung der bestmöglichen Schätzung und der möglichen Spannen (graue Balken) wurden die AOGCM (Kopplung atmosphärischer und ozeanischer Modelle) im linken Teil der Abbildung berücksichtigt, wie auch die Ergebnisse aus einer Hierarchie von unabhängigen Modellen und Beobachtungseinschränkungen. Legende: Multi-model Averages … Global surface warming Year Year 2000 Constant Concentrations 20th century Durchschnitte aus mehreren Modellen und geschätzter Umfang der Oberflächenerwärmung Globale Oberflächenerwärmung Jahr Schadstoffkonzentration bleibt konstant auf Niveau des Jahres 2000 20. Jahrhundert (32) Spezifische Klimaszenarien, die im Sonderbericht zu den Emissionsszenarien beschrieben werden. PE 379.208 14 Klimawandel und europäische Fischerei (III) Anstieg des Meeresspiegels Es besteht ziemliche Gewissheit, dass im 20. Jahrhundert der beobachtete Meeresspiegelanstieg höher war als im Jahrhundert zuvor. Der Gesamtanstieg im 20. Jahrhundert wird auf 0,17 m geschätzt. Weltweit stieg der Meeresspiegel zwischen 1961 und 2003 um durchschnittlich 1,8 mm im Jahr. Zwischen 1993 und 2003 waren es mit jährlich 3,1 mm sogar noch mehr. Es ist unklar, ob sich darin dekadenbezogene Schwankungen widerspiegeln oder aber ein Anstieg in der langfristigen Tendenz. (IV) Versauerung In den letzten 200 Jahren haben die Ozeane die Hälfte des anthropogenen CO2 absorbiert, wodurch der pH-Wert im Oberflächenwasser um 0,1 Einheiten zurückgegangen ist. Dementsprechend kam es bei den Wasserstoffionen zu einem Anstieg um 30 % (The Royal Society 2005). Durch die kontinuierliche Aufnahme von atmosphärischem CO2 wird sich der pH-Wert in den Ozeanen im Verlaufe der nächsten Jahrhunderte voraussichtlich deutlich verringern. Dadurch verschieben sich die Sättigungshorizonte von Aragonit, Kalzit und anderen Mineralien, die für kalzifizierende Organismen wichtig sind (Feely et al. 2004). Experimentell ist nachgewiesen worden, dass es bei einem Anhalten dieser Trends für wichtige Meeresorganismen wie Korallen und einige Planktonorganismen schwierig wird, die sie umhüllenden Kalziumkarbonatskelette aufrecht zu erhalten, wodurch sich wiederum die Struktur und die biologische Vielfalt in den einzelnen Ökosystemen verändern könnten (Orr et al. 2005, Riebesell et al. 2000). Wir wissen sehr wenig über die direkten Auswirkungen der Versauerung auf die Fische. Wahrscheinlich kommt es zu einer Übersäuerung des Körpergewebes und zu einer Beeinträchtigung der Stoffwechselfunktion. Eier und Larven dürften dabei anfälliger sein als erwachsene Tiere. Daher wird mit einem verringerten Reproduktionserfolg als einem der ersten Symptome gerechnet. Modellschätzungen zur pHVerringerung an der Ozeanoberfläche sprechen in Abhängigkeit von den verwendeten Szenarien für die CO2-Emission von 0,3 bis 0,5 Einheiten während der nächsten 100 Jahre und von 0,3 bis 1.4 Einheiten während der nächsten 300 Jahre (Caldeira und Wickett 2003, 2005, Abbildung 5) Abbildung 5: Prognose der Versauerung der Ozeane. 15 PE 379.208 Klimawandel und europäische Fischerei Quelle: Caldeira und Wickett 2005. Anmerkungen: Atmosphärische CO2-Emissionen, in der Vergangenheit aufgetretene CO2-Niveaus in der Atmosphäre und prognostizierte CO2-Konzentrationen (IPCC 2001) sowie Veränderung des pH-Wertes im Ozean in verschiedenen Wassertiefen, abgeleitet aus Modellrechnungen. Legende: Depth GtC/yr Emissions Year Tiefe GtC/Jahr Emissionen Jahr Obwohl sich viele Meeresorganismen im Verlaufe der letzten Millionen Jahre an thermische Schwankungen anpassen konnten, übertreffen die beim pH-Wert erwarteten Veränderungen bei weitem die anhand von Fossilien ermittelten Veränderungen in den letzten 300 Millionen Jahren (Caldeira und Wickett 2003, Feely et al. 2004). Insbesondere für die europäischen Meere dürfte die Versauerung zu einem ernsten Problem werden, da prognostiziert wird, dass 23 % des weltweit von den Meeren aufgenommenen anthropogenen CO2 von den oberflächennahen Wasserschichten des Nordatlantik absorbiert werden (Sabine et al. 2004). Außerdem wird das Problem der Versauerung in halbumschlossenen europäischen Meeren und in den Küstengewässern noch durch den Eintrag zusätzlicher säurehaltiger Verbindungen über die Flüsse und die Atmosphäre verstärkt (Hoepffner et al. 2006). Der Klimawandel wird sich voraussichtlich auch auf die zweischaligen Muscheln auswirken, da durch eine vestärkte Versauerung der Meere der Kalzifizierungsprozess in der Schale beeinträchtigt wird, was sich wiederum auf Wachstum und Reproduktion auswirken könnte. (V) Salinität Generell ist ein Anstieg der Salinität der Ozeane schwer vorhersehbar, jedoch können sich hier Veränderungen beim Niederschlag, bei der Verdunstung, den Ozeanströmungen und der Schneeschmelze potenziell auswirken (MCCIP 2006). Im Nordmeer wurde in den letzten Jahren bereits ein Rückgang der Salinität beobachtet (Curry und Mauritzen 2005, Peterson et al. 2006), wobei dieser Trend nach Modellberechnungen allerdings noch zu schwach sein dürfte, um einen spürbaren Einfluss auf die Atlantikströmungen zu haben. Zukünftig wird ein stärkerer Winterniederschlag erwartet, wodurch die Salinität in der Ostsee tendenziell zurückgehen wird. Im Gebiet des Mittelmeeres hat sich die Niederschlagsmenge während der letzten 100 Jahre um 20 % verringert (Eisenreich 2005). Da die Verdunstungsverluste größer sind als der Eintrag durch Niederschläge und Flusseinleitungen, wird mit einem Anstieg der Salinität im Mittelmeer gerechnet. Entsprechende Daten wurden im Zeitraum 1991-2003 von Malacic et al. (2006) im nördlichsten Teil des Mittelmeeres, im Golf von Triest (nördliche Adria), ermittelt, und sie zeigten einen Anstieg von 0,28-0,34 pro Jahr. Ohne das sehr trockene und heiße Jahr 2003 (siehe Kapitel 3.1 – Prognosen für den Klimawandel) ergibt sich für die Salinitätswerte ein jährlicher Anstieg um 0,22-0,28. Zusammen mit der Überfischung stellt der fortwährende Anstieg der Temperaturen und der vom Menschen verursachten CO2-Emissionen, der eine Versauerung der Weltmeere zur Folge hat, eine Gefahr für die bereits geschwächten Fischbestände dar (WBGU Report 2006). PE 379.208 16 Klimawandel und europäische Fischerei Kernaussage 1 Die Wissenschaftler sind sich darin einig, dass der Klimawandel Realität ist. Die Temperaturen werden ebenso ansteigen wie der Meeresspiegel. Anzeichen für eine Klimaerwärmung sind Veränderungen bei den Oberflächen- und Lufttemperaturen sowie den Temperaturen in den oberen Ozeanschichten bis zu mehreren hundert Meter Tiefe und der Anstieg des Meeresspiegels. Die durchschnittliche globale Temperatur ist während der letzten hundert Jahre um ~0,6 °C angestiegen, der Meeresspiegel um 0,17 m. In diesem Zeitraum haben sich sowohl die Meeres- als auch die Süßwassersysteme erwärmt. Die Prognosen sprechen von einer weiteren Erwärmung des Küstenklimas in Europa im Verlaufe des 21. Jahrhunderts, wobei mit einer Erhöhung der Meeresoberflächentemperatur um 0,2 °C in jedem Jahrzehnt gerechnet wird. Modellschätzungen sagen eine Versauerung der Ozeane voraus. Der pH-Wert an der Meeresoberfläche wird während der nächsten 100 Jahre um 0,3 bis 0,5 Einheiten zurückgehen und in den kommenden 300 Jahren um 0,3 bis 1,4 Einheiten. Die Salinität wird sich verändern. So wird für das Nordmeer und die Ostsee mit einer Abnahme der Salinität gerechnet, während sie im Mittelmeer voraussichtlich ansteigen wird. Die in immer größerem Umfang freigesetzten künstlichen Treibhausgase haben einen großen Anteil am Klimawandel. Sie sind eine Gefahr für die bereits geschwächten Fischbestände und sollten daher auf ein Minimum beschränkt werden. Veränderung in den Strömungsmustern Meeresströmungen, ozeanische Fronten, Auftriebsgebiete (upwelling) wie auch Gebiete mit einer stark abwärts gerichteten Wasserbewegung (downwelling) spielen eine wichtige Rolle für die Verteilung und die Produktion in Meeresökosystemen. Wahrscheinlich werden auch diese Merkmale durch Veränderungen der Temperatur, der Niederschläge, des Abflusses, der Salinität und des Windes beeinflusst werden (Scavia et al. 2002). Bei den Prognosen zur Erwärmung gibt es deutliche regionale Unterschiede, die nördlichen Breiten sind demzufolge jedoch am stärksten betroffen (Houghton et al. 2001). 17 PE 379.208 Klimawandel und europäische Fischerei Abbildung 6: Temperaturabweichungen. Quelle: ICES 2006. Anmerkungen: Abweichungen nach oben bei der Meerestemperatur im Nordatlantik. Die Temperaturen werden als Abweichungen vom langjährigen Mittel angegeben; in Bereichen und an einzelnen Stellen festgestellte Abweichungen werden im Hinblick auf die Standardabweichung normalisiert, ein Wert von +2 bedeutet z .B. 2 Standardabweichungen über Normal (oben - linke Karte (siehe Abbildung 3 für eine leserliche Fassung) und Kurven); die Angaben zu den Meeresoberflächentemperaturen sind Abweichungen in °C (oben - rechte Karte). Die Karten zeigen die Bedingungen im Jahr 2005 (Farbintervalle 0,5, rot bedeutet positiv/warm und blau bedeutet negativ/kalt). PE 379.208 18 Klimawandel und europäische Fischerei Legende: 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. 15. 16. 17. 18. 19. 20. 21. 22. 23. 24. 25. 26. 27. 28. 29. Fram-Straße – Ostgrönlandstrom Grönlandschelf Labradorsee Neufundlandschelf Mid-Atlantic Bight Emerald Bank (Temperatur am Meeresboden) Misaine Bank (Temperatur am Meeresboden) (nicht vorhanden) Nordisland (nicht vorhanden) Südwestisland Rockall Trough (nicht vorhanden) Färöer Strom Färöer-Shetland-Kanal (nicht vorhanden) südliches Europäisches Nordmeer (nicht vorhanden) nördliches Europäisches Nordmeer westliche Barentssee östliche Barentssee (nicht vorhanden) Fram-Straße, Westspitzbergen-Strom Golf von Biskaya nördliche Nordsee (nicht vorhanden) südliche Nordsee Deutsche Bucht Ostsee 19 PE 379.208 Klimawandel und europäische Fischerei Abbildung 7: Salinitätsabweichungen. Quelle: ICES 2006. Anmerkungen: Abweichungen nach oben bei der Salinität im Nordatlantik. Die Salinitätswerte werden als Abweichungen vom langjährigen Mittel angegeben; zum Zwecke der Abstimmung werden Abweichungen im Hinblick auf eine Standardabweichung normalisiert, z .B. ein Wert von +2 bedeutet 2 Standardabweichungen über Normal. Die Karten zeigen die Bedingungen im Jahr 2005 (Farbintervalle 0,5, grün bedeutet positiv/salzig, orange bedeutet negativ/süß). Die Kurven zeigen ausgewählte Zeitreihen. Siehe Abbildung 3 für eine leserliche Fassung der oben dargestellten Karte. PE 379.208 20 Klimawandel und europäische Fischerei Legende: 1) 2) 3) 4) 5) 6) 7) 8) 9) 10) 11) 12) 13) 14) 15) 16) 17) 18) 19) 20) 21) 22) 23) 24) 25) 26) 27) 28) 29) Fram-Straße – Ostgrönlandstrom Grönlandschelf Labradorsee Neufundlandschelf Mid-Atlantic Bight Emerald Bank Misaine Bank (nicht vorhanden) Nordisland (nicht vorhanden) Südwestisland Rockall Trough (nicht vorhanden) Färöer Strom Färöer-Shetland-Kanal (nicht vorhanden) südliches Europäisches Nordmeer (nicht vorhanden) nördliches Europäisches Nordmeer westliche Barentssee östliche Barentssee (nicht vorhanden) Fram-Straße, Westspitzbergen-Strom Golf von Biskaya nördliche Nordsee (nicht vorhanden) südliche Nordsee Deutsche Bucht Ostsee 21 PE 379.208 Klimawandel und europäische Fischerei Auf kontinentaler und regionaler Ebene wie auch in den Meeresbecken wurden zahlreiche langfristige Klimaveränderungen beobachtet (Harrods et al. 2007). Dazu gehören Veränderungen der Temperatur und des Eises in der Arktis, weitverbreitete Veränderungen der Niederschlagsmengen, der Salinität der Weltmeere und der Windmuster wie auch Wetterextreme einschließlich Dürre, Starkregen, Hitzewellen wie im Sommer 2003 und die Intensität von tropischen Wirbelstürmen. Anhand der in den letzten beiden Jahrzehnten gesammelten wissenschaftlichen Beweise lässt sich feststellen, dass Umweltveränderungen auf allen Ebenen auftreten und schwerwiegende Folgen für die europäischen Meere und Küsten haben (IPCC-Bericht 2007). Die an Europa angrenzenden Gewässer, der Nordatlantik, die Arktis und das Mittelmeer, stehen unter dem Einfluss sowohl subtropischer als auch arktischer Regionen, und es wird damit gerechnet, dass es bei deren Strömungsmustern zu regionalen Anpassungen an die durch den Klimawandel bedingten physikalischen Parameter kommt. So wurde während der letzten Dekade eine Erwärmung in den Grenzgebieten beobachtet, die von der ICES-Arbeitsgruppe über ozeanographische Hydrographie während der letzten Jahre analysiert wurde und wozu jährlich eine Aktualisierung vorgenommen wird. Ergebnisse aus dem letzten Bericht werden in dieser Studie vorgestellt (ICES Cooperative Research Report, 2006, Abbildung 6 und Abbildung 7). Die Gewässer der nordatlantischen und arktischen Regionen beheimaten zahlreiche Fischarten, die von großer kommerzieller Bedeutung sind. Der Nordatlantikstrom, der als Fortsetzung zum Golfstrom gehört, befördert von den mittleren Breiten des Nordatlantik warmes und salzhaltiges Wasser nordwärts bis in die Arktis hinein (Abbildung 8) und trägt dazu bei, dass große Gebiete eisfrei bleiben. Dieses Wasser gelangt teilweise als Oberflächenwasser, hauptsächlich jedoch als Tiefenwasser wieder in den Nordatlantik zurück, und zwar als ein durch verschiedene Absenkungen und Kanäle in Richtung Süden fließender kalter Tiefenstrom nahe dem Meeresboden. Er bildet hauptsächlich das nordatlantische Tiefenwasser, von dem die globale thermohaline Zirkulation in den Weltmeeren ganz entscheidend abhängt. Riesige Wassermassen sinken derzeit von der Oberfläche in die großen Tiefen des Nordmeers und der Labradorsee. Von da strömt das Wasser in einer Tiefe von 2-3 km in Richtung südlicher Ozean (Rahmstorf 2002). Um diesen Wasserverlust auszugleichen, fließt warmes Oberflächenwasser aus dem Süden in die nördlichen Breiten (Abbildung 9). PE 379.208 22 Klimawandel und europäische Fischerei Abbildung 8: Meeresströmungen in den nördlichen und subpolaren Becken. Quelle: Curry und Mauritzen 2005. Anmerkungen: Topographische Karte des Nordmeeres und der subpolaren Becken mit einer schematischen Darstellung der Zirkulation der Oberflächenströme (durchgehende Linien) und der Tiefenströme (gestrichelte Linien), die Bestandteil der meridionalen Umwälzzirkulation (MOC) sind . Die Farben der Kurven geben Auskunft über die ungefähren Temperaturen. Die kleine Karte zeigt die Grenzen des Nordmeeres und der subpolaren Becken, wie sie in der volumetrischen Analyse verwendet werden (gestrichelte schwarze Linien). Legende: Nordic Seas Subpolar Basins Labrador Basin Davis Strait Denmark Strait Irminger Basin Iceland Basin Iceland Sea Greenland Sea Fram Strait Norwegian Sea Faroe-Shetland Channel E. Greenland Current North Atlantic Current Norwegian Atlantic Current Temperature Nordmeer subpolare Becken Labrador-Becken Davis-Straße Dänemark-Straße Irmingerbecken Islandbecken Islandsee Grönlandsee Fram-Straße Europäisches Nordmeer Färöer-Shetland-Kanal Ostgrönlandstrom Nordatlantikstrom Norwegischer Atlantikstrom Temperatur Der globale Klimawandel wirkt auf diese Strömung, indem er die Dichte des Meereswassers auf zwei Arten verringert: Zum einen führt der Temperaturanstieg des Wassers zu thermischer Ausdehnung, zum anderen verdünnen verstärkte Niederschläge und Schmelzwasser das Meerwasser mit Süßwasser. Das Absinken des Wassers im nördlichen Atlantik, die so genannte Tiefenwasserbildung, kann durch diesen Rückgang der Salinität und folglich auch der Dichte beeinträchtigt werden. 23 PE 379.208 Klimawandel und europäische Fischerei Abbildung 9: Globale Meeresströmungen. Quelle: WBGU 2006. Anmerkungen: Gezeigt ist die durch Temperatur- und Salinitätsunterschiede ausgelöste thermohaline Zirkulation. Legende: Surface flow Deep flow Bottom flow Deep Water Formation Salinity Oberflächenströmung Tiefenströmung Bodenströmung Tiefenwasserbildung Salinität Im Nordmeer wurde in den letzten Jahren bereits ein Rückgang der Salinität beobachtet (Curry und Mauritzen 2005), wobei dieser Trend nach Modellberechnungen allerdings noch zu schwach sein dürfte, um einen spürbaren Einfluss auf die Atlantikströmungen auszuüben. Es wird darüber diskutiert, ob die von Bryden et al. (2005) vorgelegten Ergebnisse, wonach sich die Umwälzbewegung des Atlantik bereits um 30 % abgeschwächt haben könnte, auch tatsächlich korrekt sind, weil sie weder zu den Modellrechnungen noch zu den Temperaturveränderungen an der Meeresoberfläche passen. Sollte sich jedoch der Trend zu Erwärmung und Salinitätsabnahme in den kommenden Jahrzehnten weiter verstärken, dürfte dies im Verlauf des Jahrhunderts tatsächlich zu einer deutlichen Abschwächung der Atlantikströmung führen, im Extremfall womöglich sogar zu einem völligen Abbrechen der Tiefenwasserbildung (WBGUBericht 2006). Die Folge wäre ein Versiegen des Nordatlantikstroms und des größten Teils des atlantischen Wärmetransports, wodurch die Temperaturverteilung über dem ganzen Atlantikraum stark verändert würde. Als Folge der dynamischen Anpassung der Meeresoberfläche an die veränderten Strömungen würde der Meeresspiegel im Nordatlantik um bis zu 1 m steigen und auf der Südhalbkugel etwas fallen. Diese Umverteilung des Wassers hätte zunächst keinen Einfluss auf den globalen Mittelwert des Meeresspiegels (Levermann et al. 2005). Doch auch im globalen Mittel würde PE 379.208 24 Klimawandel und europäische Fischerei der Meeresspiegel längerfristig zusätzlich um 0,5 m ansteigen, da sich der tiefe Ozean nach Versiegen des Nachschubs an kaltem Wasser allmählich erwärmt. Sehr wahrscheinlich würde sich zudem der tropische Niederschlagsgürtel( 33) verschieben, da der so genannte thermische Äquator( 34) nach Süden wandern würde (Claussen et al. 2003). Ein Abreißen des Nordatlantikstroms ist ein schwer kalkulierbares Risiko. Ein entscheidender Faktor im Simulationsmodell wird die Süßwassermenge sein, die künftig in den nördlichen Atlantik gelangt. Diese wird insbesondere von der Geschwindigkeit abhängen, mit der Grönlands Eisschild abschmilzt. (WBGU 2006). Kernaussage 2 Die an Europa angrenzenden Gewässer, der Nordatlantik, die Arktis und das Mittelmeer, stehen unter dem Einfluss sowohl subtropischer als auch arktischer Regionen. Es wird damit gerechnet, dass es bei deren Strömungsmustern zu regionalen Anpassungen an die durch den Klimawandel bedingten physikalischen Parameter kommt. Die atlantische thermohaline Zirkulation wird schwächer werden. Zwischen der Nordsee und den Gewässern in den arktischen Regionen findet ein Austausch von kalten und warmen Wasserströmungen statt, die durch die atlantische thermohaline Zirkulation bewegt werden. Die Fließintensität wird durch den Klimawandel beeinflusst. Sie dürfte derzeit bereits um 30 % verringert sein. Es ist unwahrscheinlich, dass die Zirkulation einmal gänzlich zum Erliegen kommt, was die derzeitigen Gegebenheiten sehr stark verändern würde (z. B. ein kurzfristiger Anstieg des Meeresspiegels in der Nordsee um 1 m, ein langfristiger Anstieg des globalen Meeresspiegels um 0,5 m, Verschiebung des tropischen Niederschlagsgürtels nach Süden und eine Verringerung der Biomasse im Atlantik um 50 %). Eine Bewertung und Prognose der Auswirkungen ist mit erheblichen Unsicherheiten verbunden. Bislang wurden noch keine Szenarien dazu entwickelt, wie sich mögliche Veränderungen der thermohalinen Zirkulation auf die Meeresökosysteme auswirken könnten. Veränderungen in der Position der ozeanischen Fronten und ihre Rolle als Barrieren Durch die Schichtung der Wassersäule entstehen natürliche Barrieren, an denen es zur Konzentration von Organismen kommen kann oder die von den Organismen passiert werden müssen. Die Intensität der Schichtung korreliert positiv mit Salinität und Temperatur, und eine verstärkte Schichtung unterdrückt den Nährstoffaustausch durch vertikale Durchmischung (Behrenfeld et al. 2006). Die Nordsee weist an ihren östlichen Rändern, entlang der belgischen, niederländischen, dänischen, deutschen und norwegischen Küste, die stärkste vertikale Schichtung auf, was auf die Süßwasserzufuhr aus der Ostsee und den Flüssen zurückzuführen ist. Ein verstärkter Süßwasserzufluss hat eine verstärkte Schichtung zur Folge. Dadurch wird mehr Energie für die vertikale Durchmischung des Wassers notwendig. Die durchmischte Schicht ist folglich flacher und es findet eine geringere Durchmischung mit tieferem Wasser statt, wodurch wiederum die Anreicherung mit Nährstoffen reduziert wird (Scavia et al. 2002). Die Schichtung erreicht jedes Jahr im Spätsommer ihren Höhepunkt, was auf die Erwärmung der Meeresoberfläche durch die Sonne zurückzuführen ist. Ein Temperaturanstieg könnte Zeitpunkt und Umfang der Schichtung verändern (MCCIP 2006). In der südwestlichen Nordsee (33) Der tropische Niederschlagsgürtel erstreckt sich am Äquator zwischen 30° südlicher Breite und 30° nördlicher Breite. Entsprechend dem Zenit der Sonne reicht die Regenzeit im Nordsommer bis an die nördliche Grenze und im Südsommer (Nordwinter) entsprechend bis an die südliche Grenze. Dadurch kommt es zu niederschlagsreichen Zeiten in der Nähe des Zenitpunktes der Sonne und zu trockenen Zeiten weit von diesem Punkt entfernt. (34) Der thermische Äquator wird durch die breitenkreisähnliche Linie repräsentiert, die die im Mittel wärmsten Punkte der Erde auf ihren jeweiligen Längengraden miteinander verbindet. 25 PE 379.208 Klimawandel und europäische Fischerei nahe am Eingang zum Ärmelkanal wird die Wassersäule ständig durchmischt, da sie an dieser Stelle sehr flach ist und sich außerdem der Gezeitenstrom bemerkbar macht. Während der letzten 50 Jahre (1951–1998) wurde die Nordsee westlich von ca. 5° immer stärker geschichtet, während nach Osten hin die Schichtung immer mehr abnahm, worin sich die langfristige Veränderung der Salinität widerspiegelt (Beare et al. 2002). Fangdaten von schottischen Forschungsschiffen vom Januar 1995 zeigen, dass bei den pelagischen Warmwasserarten Sardelle (Engraulis encrasicolus), Sardine (Sardina pilchardus) und Meerbarbe, also bei normalerweise im nördlichen Europa nur selten vorkommenden Arten, die Fangquoten nach 1995 plötzlich stark angestiegen sind (Beare et al. 2004). Eine verstärkte Schichtungsintensität aufgrund abnehmender Windgeschwindigkeit und steigender Temperaturen könnte dazu geführt haben, dass mehr Sardinen- und Sardellenlarven überlebten (Beare et al. 2002). Die Zunahme von Calanus helgolandicus gegenüber Calanus finmarchicus könnte auch einer der Gründe gewesen sein, da Sardinen- und Sardellenlarven bevorzugt auf C. helgolandicus als Nahrung zurückgreifen (Beare et al. 2004). Molinero et al. (2005b) beobachteten eine Verstärkung der Schichtung der Wassersäule im Mittelmeer im Zusammenhang mit abnehmender Windbelastung und hoher Wassertemperatur Mitte bis Ende der 1980er Jahre. Marty und Chiaverini (2002) stellten auch im Zeitraum 19911999 eine solche verstärkte Schichtung aufgrund hoher Wassertemperaturen fest. Wegen der höheren Temperaturen und der geringeren Windbelastung gelangten weniger Nährstoffe in die oberen Schichten, was zu Veränderungen in den Phytoplanktongemeinschaften führte. Mit ihren zusammengestellten Daten zur Phytoplanktonblüte zeigten Bethoux et al. (2002) auf, dass in kalten Jahren mit geringer NAO Diatomeen( 35) in der Phytoplanktonblüte im Frühjahr dominierend sind, während sie in warmen Jahren mit hoher NAO offenbar selten sind und dafür Flagellaten ( 36) in der Blüte vorherrschen. Viele Organismen unterliegen nahe der Grenzen des Ausbreitungsgebietes ihrer Art einem stärkeren Stress, wobei sich die Verbreitung dieser Arten mit den veränderten Umweltbedingungen voraussichtlich ändern wird (Harley et al. 2006). Bei der derzeitig durch äußere Faktoren bestimmen Verbreitung lassen sich viele biogeographische Grenzen in Küstenmeeren festlegen, obwohl es potenziell geeignete Habitate außerhalb dieser Grenzen gibt. Die Ausbreitungsgrenzen der Meeresarten bleiben möglicherweise immer gleich, selbst wenn sich Bedingungen in außerhalb gelegenen Habitaten als geeignet erweisen. Harley et al. (2006) stellen fest, dass durch eine mit der Erwärmung verbundene Abschwächung der Advektion( 37) längs der Küsten tatsächlich einige der biogeographischen Barrieren im Meer beseitigt werden könnten, die gegenwärtig eine Ausweitung der Verbreitungsgebiete verhindern (Gaylord und Gaines 2000). Es muss darauf hingewiesen werden, dass bei Modellen der globalen Zirkulation, die auf Klimaszenarien basieren, die Meeresumwelt und Veränderungen im Meeresklima bislang vereinfacht behandelt werden. Die Modelle werden zwar deutlich besser, jedoch werden die Veränderungen in der Position der ozeanischen Fronten noch nicht mit der erforderlichen Konsequenz untersucht (Brander 2006). (35) Mikroskopisch kleine einzellige Algen mit einer äußeren, wunderschön strukturierten Schale aus Silikat, deren zwei Teile mit breiten Gürtelbändern verbunden sind. (36) Gruppe, zu denen die Dinoflagellaten gehören, auch Euglenida, Raphidophyta. (37) Transport durch den Hauptstrom, im Gegensatz zu Diffusion oder Verbreitung durch Dispersion. PE 379.208 26 Klimawandel und europäische Fischerei Kernaussage 3 Die Schichtung wird sich verstärken. Es wurde nachgewiesen, dass der Klimawandel zu einer verstärkten Schichtung der Ost- und Nordsee und des Mittelmeeres führt. Durch die Schichtung der Wassersäule entstehen natürliche Barrieren, wo es zur Konzentration von Organismen kommen kann oder die von den Organismen passiert werden müssen. Die Schichtung verstärkt sich mit zunehmender Salinität und Temperatur. Eine verstärkte Schichtung wiederum behindert die Vermischung mit Tiefenwasser und führt zu einer geringeren Anreicherung mit Nährstoffen. Durch eine veränderte Zirkulation und Schichtung wird auch die geographische Verbreitung der Organismen verändert. Die Strömungen sind wichtig für den Transport von Organismen wie Plankton und Fisch über große Entfernungen und können daher zu deren größerer Verbreitung beitragen. Andererseits jedoch wirken die Strömungen auch als biogeographische Barrieren zwischen den zu beiden Seiten befindlichen Wassermassen. Der Austausch von Organismen über die Strömung hinweg wird dadurch verringert. Durch die Erwärmung könnte es zu einer Abschwächung der Küstenströme kommen, wodurch auch die Verbreitung über diese Ströme geringer wird. Gleichzeitig verschwindet die Barriere zwischen Küsten- und Meeresgewässern. Dadurch könnten sich Organismen, die zuvor auf die küstennahen Bereiche beschränkt waren, weiter ausbreiten. Alle diese Vorgänge wirken sich (positiv oder negativ) auf die Primärproduktion aus. Verfügbare Modelle der globalen Zirkulation, die auf Klimaszenarien basieren, haben die Meeresumwelt und Veränderungen im Meeresklima bislang in vereinfachter Art und Weise behandelt. Die Modelle werden zwar deutlich besser, jedoch werden die Veränderungen in der Position der ozeanischen Fronten noch nicht mit der erforderlichen Konsequenz untersucht Verschiebung von Übergangszonen Die zum Plankton gehörende Copepodenpopulation Calanus finmarchicus in der Nordsee ist seit dem Ende der 1950er Jahre immer kleiner geworden, während gemäßigte atlantische und neritische Arten( 38) in immer größeren Zahlen anzutreffen sind. Zur gleichen Zeit, da die Population der C. finmarchicus zurückging, war in der östlichen Nordsee eine langfristige Versüßung (d. h. eine Verringerung der Salinität) und Erwärmung zu beobachten, in der westlichen Nordsee hingegen eine langfristige Zunahme der Salinität. Diese Veränderungen lassen sich dadurch erklären, dass das in die Nordsee einströmende Atlantikwasser seit Ende der 1950er Jahre unterschiedlichen Ursprungs ist, was mit dem Klimawandel im Zusammenhang steht (Beare at al. 2002). Mit der Ermittlung klimabedingter Veränderungen in der Ökologie einer Art beschäftigt sich insbesondere die Phänologie, die unter anderem den Zeitpunkt und das Muster biologischer Zyklen untersucht. Molinero et al (2007) analysierten in einer Zeitreihe von 27 Jahren bei den beiden Copepoden Centropages typicus und Temora stylifera zwei im westlichen Mittelmeer vorherrschende Arten. Sie zeigen unterschiedliche saisonale Muster in Abhängigkeit von der Temperatur. C. typicus hat seine Spitzenzeiten im Frühjahr, T. stylifera dagegen im Spätsommer/Herbst. In Jahren mit positiver NAO wies der durchschnittliche Jahreszyklus der C. typicus eine höhere Abundanz mit einer absoluten Spitze im April aus, woran ersichtlich wird, dass es abgesehen von der Zunahme der Abundanz auch zu einer zeitlichen Vorverlagerung kommt. In Jahren mit negativer NAO hat sich die Abundanz von C. typicus nahezu um das Dreifache verringert. Das gegensätzliche Muster wurde bei T. stylifera beobachtet, bei denen in Jahren mit positiver NAO eine geringe Abundanz festzustellen war, (38) Arten, deren Lebenszyklus und Verbreitung größtenteils auf den Kontinentalsockel und die obere Neigung beschränkt sind. 27 PE 379.208 Klimawandel und europäische Fischerei während sie sich in Jahren mit negativer NAO verdoppelte und zu einem frühen Zeitpunkt den Spitzenwert erreichte. Die möglichen Auswirkungen einer erhöhten Salinität in der nördlichen Adria sind nicht vorhersehbar. Entscheidend wird sein, wie tolerant sich die untersuchten Organismen gegenüber einer solchen Veränderung verhalten. Sowohl einzelne Arten als auch Gemeinschaften und Ökosysteme könnten beeinflusst werden. Mobile Organismen können sich einer potenziellen Salinitätsbelastung dadurch entziehen, dass sie das Gebiet verlassen. Wenn sie jedoch bezüglich der Salinität bereits die Toleranzschwelle erreicht haben und das Gebiet nicht verlassen können, kann es zu einer Verringerung des Wachstums und der Fruchtbarkeit und zu Problemen mit der Osmoregulation kommen (siehe allgemeine Ökosystembeziehungen in Kapitel „Reproduktionsmuster, Wanderungswege und Ökosystembeziehungen“. Kernaussage 4 Beispiele zeigen, dass es durch die Verschiebung von Übergangszonen infolge des Klimawandels zu Veränderungen in der Abundanz der Arten kommt und sogar ganze Populationen zusammenbrechen können. Spezifische Auswirkungen in halbumschlossenen Meeren (I) Ostsee In den letzten 100 Jahren hat sich die Lufttemperatur in der Ostseeregion um mehr als 0,7 °C erhöht, was deutlich über dem globalen mittleren Temperaturanstieg von 0,5 °C liegt. Ozeanographischen Studien zufolge könnte sich die durchschnittliche jährliche Temperatur an der Wasseroberfläche bis zum Ende des 21. Jahrhunderts um 2° bis 4 °C erhöhen (BACC 2006). Dieser Temperaturanstieg ist verbunden mit einer Zunahme des Winterabflusses, kürzerer Eisbedeckung und einer geringeren Eisdicke auf den Flüssen und Seen im Baltikum. Für die Zukunft wird mit einer Zunahme der Winterniederschläge gerechnet, wodurch sich die Salinität in der Ostsee tendenziell verringern dürfte. Die in der Vergangenheit beobachteten Temperaturveränderungen waren mit entsprechenden Veränderungen in den terrestrischen Ökosystemen verbunden, wie etwa frühzeitigere phänologische Frühlingsphasen( 39) oder Verlagerungen der Arten nach dem Norden. Im Ökosystem der Ostsee lassen sich diesbezügliche Einschätzungen nur schwer vornehmen, da es starke nichtklimatische Stressfaktoren wie Eutrophierung(40), Fischerei und Schadstoffe gibt. Die erwartete Verringerung der Salinität (Prognosen zufolge zwischen 8 % und 50 %) jedoch wird sich sehr stark auf Verteilung, Wachstum und Reproduktion auswirken. Es wird damit gerechnet, dass sich Süßwasserarten in die Ostsee ausbreiten und Eindringlinge aus wärmeren Meeren wie die Zebramuschel (Dreissena polymorpha) und die Rippenqualle (Mnemiopsis leidyi, Javidpour et al. 2006) ihre Verbreitungsgebiete weiter ausdehnen (BACC 2006). Für die Ostsee werden sich als Folge der prognostizierten Temperaturerhöhung die Wachstumsund Reproduktionsparameter ändern. Es wird für möglich gehalten, dass durch eine erhöhte Temperatur das Wachstum pelagischer Bakterien stärker angeregt wird als die (39) „Phänologische Phasen“ beschreiben den Zeitpunkt und die Merkmale biologischer Zyklen. (40) Im Allgemeinen der natürliche oder vom Menschen ausgelöste Prozess, durch den ein Gewässer mit gelösten Mineralstoffen (vor allem Phospor und Stickstoff) angereichert wird, die das Wachstum von Wasserpflanzen anregen und die organische Produktion im Gewässer erhöhen. Eine übermäßige Anreicherung kann dazu führen, dass der gelöste Sauerstoff aufgebraucht wird, was letztlich das Absterben oder die Verlagerung von Arten zur Folge hat. PE 379.208 28 Klimawandel und europäische Fischerei Primärproduktion, was dazu führt, dass sich der Anteil der bakteriellen Biomasse am Phytoplankton erhöht. Bei den Diatomeenblüten wird es in milden Wintern zu einer Veränderung der Artenzusammensetzung kommen und Diatomeen könnten durch Dinoflagellaten( 41) ersetzt werden. Durch steigende Sommertemperaturen wird es verstärkt zur Blüte von Cyanobakterien( 42) kommen. Höhere Wintertemperaturen könnten die Konvektion( 43) am Ende des Winters und im zeitigen Frühjahr verhindern, so dass die Nährstoffe nicht in die obere euphotische Zone( 44) gemischt werden. In der eigentlichen Ostsee mit einer Salinität von 7 psu tritt die maximale Wasserdichte bei etwa 2,5 °C auf. Bei höheren Temperaturen entwickelt sich eine Thermokline( 45), und es erfolgt keine Umverteilung von Nährstoffen durch Konvektion. Dadurch kann sich möglicherweise im Frühjahr die Artenzusammensetzung verändern (BACC 2006, Pawlak et al. 2007). (II) Nordsee In der Nordsee werden die Temperaturen in den Küstengewässern durch die Wechselwirkungen zwischen Atmosphäre und Ozean und durch Süßwasserabläufe beeinflusst. Langfristig betrachtet sind zwischen Anfang der 1950er und Ende der 1990er Jahre die Meerestemperaturen im Spätwinter (Januar, Februar) in allen Nordseegebieten deutlich angestiegen. Während der letzten 70 bis 100 Jahre ist ein Anstieg der jährlichen Durchschnittstemperatur (Land- und Wasseroberfläche) der Nordsee um 0,6 °C beobachtet worden, der vor allem in den letzten 20 Jahren besonders deutlich ausfiel (Hulme et al. 2002). In der Deutschen Bucht an der Helgoland Reede hat sich die Wassertemperatur während der letzten 40 Jahre um 1,3° erhöht. Kalte Winter mit Meeresoberflächentemperaturen um - 1 °C gab es bis 1944 etwa alle zehn Jahre, seit 1960 jedoch wurde ein solcher Fall nur einmal beobachtet (Wiltshire und Manly 2004). Modellrechnungen von Sheppard (2004) zu den Veränderungen an der Meeresoberfläche in den kommenden 90 bis 100 Jahren prognostizieren etwa 1,6 °C bis 3,0 °C für die nördliche Nordsee und 3,0 °C bis 3.9 °C für die flachere südliche Nordsee. Die niedrigsten Salinitätswerte sind im Norwegischen Küstenstrom und in der südöstlichen Nordsee anzutreffen, was mit den Süßwasserzuflüssen aus Ostsee, Elbe und Rhein zusammenhängt. Saisonal bedingt kommt es zu Salinitätsschwankungen, in der Regel ist diese jedoch im Spätwinter am höchsten (Turrell et al. 1992). Ein genereller Trend ist nach wie vor nicht eindeutig festzustellen (ICES 2006). (III) Mittelmeer, Agäis und nördliche Adria Das Mittelmeer ist ein halbumschlossenes tiefes Becken mittleren Breitengrads, seine beiden großen, in Wechselwirkung stehenden Teilbecken (westliches und östliches Teilbecken) sind durch die Straße von Sizilien miteinander verbunden. Das Mittelmeer wurde als ein „Miniaturozean“ beschrieben und wird aufgrund seiner raschen Reaktionen auf externe Einflüsse als ideales Testgebiet angesehen (Bethoux et al. 1999). (41) (42) (43) (44) (45) Gruppe planktonischer Algen. Synergistische Blaualgen genannt. Gruppe von Bakterien. Vertikale Zirkulation in einem Gas oder einer Flüssigkeit unter Bedingungen der Instabilität. Die obere, durchleuchtete Schicht des Wassers, in der effektive Photosynthese möglich ist. Region unterhalb der Oberflächenschicht des Meeres oder eines Sees, die ein steiles Temperaturgefälle aufweist (d. h. in der die Temperaturen mit zunehmender Tiefe sehr stark absinken). Eine Thermokline kann bis an die Oberfläche reichen und zu einer Front werden. Sie stellt normalerweise eine ökologische Barriere dar, und ihre Oszillationen haben erhebliche Auswirkungen auf die Bestandsverteilung und die Produktivität des Meeres. 29 PE 379.208 Klimawandel und europäische Fischerei Maßgeblich für die Zirkulation des Mittelmeeres sind der Wasser- und Wärmeaustausch mit der Atmosphäre über die Wasseroberfläche sowie der Austausch mit den angrenzenden Meeren über die Meerengen. Die thermohaline Zirkulation im Mittelmeer wird bestimmt durch den vom Auftrieb beeinflussten Austausch und den negativen Wärme- und Süßwasserhaushalt. Die Verdunstungsmenge ist größer als die Menge der Niederschläge und Flussabläufe, wodurch das Wasser eine hohe Dichte erlangt. Über die Straße von Gibraltar strömt aus dem Atlantik leichtes Wasser ein und in geringerem Umfang in den oberflächlichen Schichten auch aus dem Schwarzen Meer. Dichtes salzhaltiges Wasser wird durch Unterwasserströme abtransportiert. Dieses warme und salzige Wasser gelangt in den Atlantischen Ozean und trägt zur nordatlantischen thermohalinen Zirkulation bei (Bethoux et al. 1999, Theocharis et al. 1999). Die wichtigsten Arten der Zirkulationsschwankung und deren Reaktion auf atmosphärische Schwankungen sind noch nicht vollständig geklärt (Tsimplis et al. 2006, Hoepffner et al. 2006). Eine kontinuierliche Beobachtung der Zirkulationsmuster an wichtigen Punkten im Mittelmeer ist daher ganz wichtig, um künftige Veränderungen in den Wassermassen unter unterschiedlichen Erwärmungsbedingungen einschätzen zu können (Hoepffner et al. 2006). In den Wassersäulen des Mittelmeers wurde auf den verschiedenen Ebenen ein Anstieg der Durchschnittstemperatur festgestellt (Bethoux et al. 1990, Francour et al. 1994). Temperaturund Salinitätsdaten wurden von Malacic et al. (2006) im Zeitraum 1991-2003 im nördlichsten Teil des Mittelmeers, im Golf von Triest (nördliches Adriatisches Meer) erfasst. Die Temperatur an der Oberfläche schwankte mit einer jährlichen Amplitude von 8.1 +- 0.4 °C um 16,5 °C, in einer Tiefe von 10 m mit einer Amplitude von 7.0 +- 0.3 °C um 15,5 °C. Die Sommertemperaturen an der Oberfläche stiegen zwischen 0,12 und 0,23 °C jährlich. Ohne das extrem trockene und heiße Jahr 2003 verringert sich dieser jährliche Anstieg auf 0.07-0.09 °C. Die Salinität erhöhte sich um 0,28-0,34 pro Jahr, ohne das Jahr 2003 um 0,22-0,28. Kernaussage 5 Generell lässt sich feststellen, dass die Auswirkungen des Klimawandels in halbumschlossenen Meeren wahrscheinlich noch schwerwiegender sind als in den offenen Meeren und dass es weiterer Forschungen bedarf, um die Klimaeffekte bei Vorhandensein nichtklimatischer Stressfaktoren einschätzen zu können (insbesondere im Hinblick auf die hohe Schadstoffbelastung der Ostsee). Die erwarteten Auswirkungen werden die marine Produktivität negativ wie auch positiv beeinflussen. Für die Ostsee entwickelte Szenarien zum Klimawandel sprechen von Rückgängen bei der Salinität zwischen 8 % und 50 % und von einem Anstieg der Temperatur an der Wasseroberfläche von 2 bis 4 °C. Für die Nordseeregion verändern sich die Salinitätsprognosen entsprechend den erwarteten Anstiegen oder Rückgängen in den verschiedenen Gebieten der Nordsee. Die Temperaturen an der Wasseroberfläche sollen in der nördlichen Nordsee um etwa 1,6 °C bis 3.0 °C ansteigen, in der flacheren südlichen Nordsee um 3,0 °C bis 3,9°. Entsprechend den Szenarien zum Klimawandel werden Salinität und Temperaturen auch im Mittelmeer ansteigen. Sonstige relevante Literatur zu den Veränderungen bei den Wassereigenschaften Das Winterklima in der Europäischen Union wird in vielerlei Hinsicht sehr stark von der so genannten Nordatlantik-Oszillation (NAO) beeinflusst, die die atmosphärischen Vorgänge im Nordatlantik dominiert (Hurrell 1995, Hurrell und Dixon 2004). Die NAO ist ein Wechsel im Druckunterschied zwischen der subtropischen Hochdruckzone mit Zentrum über den Azoren und der Tiefdruckzone über Island (Hurrell 1995, Ottersen et al. 2001). Sie ist ganzjährig das PE 379.208 30 Klimawandel und europäische Fischerei vorherrschende Strömungsmuster im Nordatlantiksektor, ist jedoch im Winter am stärksten ausgeprägt und für mehr als ein Drittel der Unterschiede beim Luftdruck auf Meeresspiegelhöhe (SLP) verantwortlich (Cayan 1992; Hurrell 1995). Für die NAO wurden verschiedene Indizes festgelegt, insbesondere von Rogers (1984), die später dann von Hurrell (1995) und Jones et al. (1997) erweitert wurden. Hurrells Winterindex der NAO (Dezember bis März) basiert auf dem Unterschied bei den normalisierten SLP-Werten zwischen Lissabon, Portugal, und Stykkisholmur, Island, in den Jahren 1864 bis 1995. Ein hoher oder positiver NAO-Index ergibt sich dann, wenn Islandtief und Azorenhoch stark ausgeprägt sind (Abbildung 10). Größere Druckunterschiede führen zu mehr und stärkeren Winterstürmen, die den Atlantik in einer nördlicheren Richtung überqueren. Das geringere Druckgefälle bei niedrigem NAO-Index oder negativer NAO-Phase führt andererseits dazu, dass es weniger und schwächere Winterstürme gibt, die eher in westöstlicher Richtung über den Atlantik ziehen. Die Variabilität bei der Ausrichtung und Stärke der Westwinde bewirkt Schwankungen bei den Wintertemperaturen zwischen den Jahren und Dekaden und beeinflusst das Verhältnis von Niederschlag und Verdunstung über dem Festland zu beiden Seiten des Atlantiks (Rogers 1984; Hurrell 1995). Der Zusammenhang zwischen dem Zustand der NAO und den Temperatur-, Wind- und Niederschlagsmustern ist im nördlichen Europa besonders stark. Die NAO scheint ein guter Ersatzindikator für die Wintertemperaturen an der Wasseroberfläche und die Windstärke in der Nordsee zu sein (Ottersen et al. 2001). 31 PE 379.208 Klimawandel und europäische Fischerei Abbildung 10: Die Nordatlantik-Oszillation. Quelle: Parsons und Lear 2001. Anmerkungen: Schematische Darstellung von hohem NAO-Index (oben) und niedrigem NAO-Index (unten), wobei T=Islandtief, H=Azorenhoch, K=kalte Luftmassen und W=warme Luftmassen. Legende: C L K T Es wurde festgestellt, dass die Druckfelder im Zusammenhang mit der NAO für die Veränderungen des mittleren Meeresspiegels in Europa verantwortlich sind. Der Wasseraustausch in der Straße von Gibraltar (Gomis et al. 2006) sowie die Temperatur- und PE 379.208 32 Klimawandel und europäische Fischerei Salinitätsschwankungen im Tiefenwasser wie auch in den oberen Wasserschichten des Mittelmeers wurden ebenfalls mit der NAO in Verbindung gebracht (Bethoux et al. 1999, Send et al. 1999). Die Zeit der starken Nachwuchsproduktion bei den Kabeljauartigen in der Nordsee in den 1960er Jahren, der so genannte „Kabeljauboom“ (Cushing 1984), fiel zeitlich mit der dauerhaftesten negativen NAO-Phase zusammen. Zu diesen Kabeljauartigen gehörten in der Hauptsache vier Arten (Kabeljau (Gadus morhua), Schellfisch (Melanogrammus aeglefinus), Wittling (Merlangius merlangius) und Seelachs (Pollachius virens)). Die Transporte im Nordatlantikstrom, die Gefälleströmung (slope current) durch den Färöer-Shetland-Kanal und die Zuflüsse in die Nordsee waren alle so niedrig wie seit langem nicht, ebenso die südwestlichen Winde über der Nordsee, die sowohl den Transport als auch die Durchmischung beeinflussen (Parsons und Lear 2001). Außerdem waren die in die Nordsee fließenden Wassermassen kalt und die Wasseroberflächentemperaturen in der Nordsee niedrig. In den 1990er Jahren, als bei den Beständen der Kabeljauartigen in der Nordsee die Rekrutierung generell niedrig war, befand sich die NAO in einer dauerhaften und starken positiven Phase, und die regionalen Umweltbedingungen hatten sich ins Gegenteil verkehrt. Sirabella et al. (2001) zeigten auf, dass eine positive NAO-Phase mit hohen Meerestemperaturen die Kabeljaurekrutierung in der Nordsee ungünstig beeinflusste. Eine Untersuchung der entsprechenden Situation beim Kabeljau in der Irischen See bestätigte dieses Phänomen. Die schlechte Rekrutierung ist vermutlich ein Ergebnis der eingeschränkten Energie aufgrund des erhöhten Stoffwechsels in warmen Jahren (Planque und Fox 1998). Alheit und Hagen (1997) untersuchten die Schwankungen bei den Herings- (Clupea harengus) und Sardinenpopulationen (Sardina pilchardus) im Nordostatlantik. Dabei reichten ihren Ermittlungen zufolge die historischen Höchstwerte für den Hering vor der schwedischen Küste bis ins 10. Jahrhundert zurück, und sie brachten dies mit veränderten atmosphärischen Einflüssen, d. h. der NAO, in Verbindung. Sie konnten aufzeigen, dass die Heringsfänge im Ärmelkanal und in der Bucht von Biskaya bei negativem NAO-Index besonders hoch waren, wenn also im westlichen Europa strenge Winter mit extrem kalter Luft und niedrigen Wassertemperaturen und schwächeren westlichen Winden auftraten. Unter umgekehrten Vorzeichen, also während positiver Phasen des NAO-Index mit einer schwachen Eisbedeckung vor Island, stärkeren westlichen Winden und relativ warmem Wasser in der Nordsee, dem Ärmelkanal und dem Skagerrak, waren der Norwegische Frühjahrslaicher (Hering) und die Sardine vor der Südwestküste Englands und im Kanal vorherrschend. Diese Beobachtung veranlasste Alheit und Hagen (1997) zu der Schlussfolgerung, dass klimatische Schwankungen für den Wechsel von Perioden mit großen und kleinen Herings- und Sardinenvorkommen im Nordostatlantik verantwortlich sind. Eine ähnliche Situation wurde für den Pazifik beschrieben, wo Schwankungen bei den Sardinen- und Sardellenvorkommen nachgewiesenermaßen mit großen Veränderungen der Ozeantemperaturen zusammenhängen. In warmen Jahren ist die Sardine im System vorherrschend, während sich in kalten Jahren die Sardellenpopulationen drastisch vergrößern (Chavez et al. 2003). Es gibt genügend Anhaltspunkte dafür, dass die mit der NAO verbundene Variabilität der atmosphärischen Zirkulation größtenteils auf eine interne, nichtlineare Dynamik der extratropischen Atmosphäre zurückzuführen ist. Hurrell et al. (2006) stellen fest, dass es in der Wissenschaft keine einheitliche Auffassung dazu gibt, welche Prozesse höchstwahrscheinlich für die verstärkte NAO-Variabilität zwischen den Jahren verantwortlich sind. Eine mögliche Ursache für die jüngst beobachtete Tendenz beim NAO-Winterindex ist die Stärke der atmosphärischen Zirkulation in der unteren Stratosphäre über einen längeren Zeitraum hinweg, bedingt durch eine Verringerung des stratosphärischen Ozons und eine Erhöhung der 33 PE 379.208 Klimawandel und europäische Fischerei Treibhausgaskonzentration. Eine andere Theorie besagt, dass die Wechselwirkung zwischen Ozean und Atmosphäre für die jüngste zeitweilige Entwicklung der NAO von Bedeutung ist (Hurrell et al. 2006). Rodwell et al. (1999) führen an, dass Charakteristika der Meeresoberflächentemperatur durch Verdunstung, Niederschlag und Prozesse, die zur Aufheizung der Atmosphäre führen, an die Atmosphäre „übermittelt“ werden, was zu Veränderungen bei der Temperatur, den Niederschlägen und der Sturmintensität über Europa führt. Daher ist es möglich, die NAO-Variabilität anhand der Temperaturen des Oberflächenwassers im Atlantik zu rekonstruieren. Monatliche und jährliche Veränderungen in der Phase und Schwankungsbreite der NAO werden jedoch wahrscheinlich auch weiterhin größtenteils nicht vorhersehbar sein (Hurrell et al. 2006). Kernaussage 6 Die Nordatlantik-Oszillation wirkt sich auf das europäische Meeresökosystem aus. Das Winterklima im europäischen Raum wird in vielerlei Hinsicht sehr stark von der Nordatlantik-Oszillation (NAO) beeinflusst, die die atmosphärischen Vorgänge im Nordatlantik dominiert. Es wurden verschiedene Indizes für die NAO festgelegt, die sich auf alle trophische Ebenen in den Meeren auswirkt. Die Rekrutierung der Industriefische hängt vom NAO-Index ab, wie es für die starke Nachwuchsproduktion bei den Kabeljauartigen (betrifft Kabeljau, Schellfisch, Wittling und Seelachs) in der Nordsee und für die Rekrutierung des Herings und der Sardine im Nordostatlantik aufgezeigt wurde. Die NAO ist so gut wie nicht prognostizierbar, obwohl sie sich aus der Meeresoberflächentemperatur rekonstruieren lässt. Die Ex-post-Untersuchungen des NAO-Index und der Zusammenhang mit beobachteten Auswirkungen auf das marine Ökosystem dürften es möglich machen, Modelle für die Prognostizierung künftiger Auswirkungen zu entwickeln. 3.2 Erwartete Auswirkungen des Klimawandels auf die Hauptarten Biologische Merkmale der wichtigsten Industriefischarten Schwerpunkt der Studie ist die Fischerei der Europäischen Union im Atlantik und in den angrenzenden Meeren, weshalb auch bei der Literaturrecherche das Hauptaugenmerk auf den Fischarten lag, die für die europäische Fischerei im Atlantik, in der Nordsee, dem Mittelmeer und der Ostsee von besonderer Relevanz sind. Um einen Überblick über die Umweltbedürfnisse der einzelnen Fischarten zu erlangen, wurden die biologischen Merkmale für die folgenden Industriefischarten bestimmt: Tabelle 4: Auswahl Verbreitungsgebiete von Hauptarten Hauptarten und deren für die Studie relevanten Für die Studie relevante Verbreitungsgebiete Hering (Clupea harengus) Nordostatlantik, Nordsee, Ostsee Sprotte (Sprattus sprattus) Nordostatlantik, Nordsee, Ostsee, Mittelmeer Sardine (Sardina pilchardus) Nordostatlantik, Mittelmeer Sardelle (Engraulis encrasicolus) Nordostatlantik, Mittelmeer Makrele (Scomber scombrus) Nordostatlantik Roter Thun (Thunnus thynnus) Atlantik, Mittelmeer Blauer Wittling (Micromesistius poutassou) Nordsee Kabeljau (Gadus morhua) Nordostatlantik, Nordsee, Ostsee PE 379.208 34 Klimawandel und europäische Fischerei Stintdorsch (Trisopterus esmarkii) Nordostatlantik, Nordsee Europäischer Seehecht (Merluccius merluccius) Nordostatlantik, Nordsee, Mittelmeer Großer Sandaal (Hyperoplus lanceolatus) Nordsee Kleiner Sandaal (Ammodytes marinus) Nordsee Schellfisch (Melanogrammus aeglefinus) Nordostatlantik Seelachs (Pollachius virens) Nordostatlantik Scholle (Pleuronecta platessa) Nordostatlantik, Nordsee Ohrensardine (Sardinella aurita) Mittelmeer Stöcker (Trachurus trachurus) Nordostatlantik, Mittelmeer Echter Bonito (Katsuwonus pelamis) Atlantik, Mittelmeer Gelbflossenthun (Thunnus albacares) Atlantik, Mittelmeer Quelle: Basierend auf Fröse und Pauly 2007. Obwohl viele der abiotischen und biotischen Merkmale der Hauptarten durch den Klimawandel beeinflusst werden, so ist doch die Temperatur ein entscheidender Faktor, der die Prozesse von der Zelle bis zum fertigen Organismus steuert. Ein Ziel der integrativen Biologie besteht darin, die Auswirkungen der Temperatur auf die physiologischen Mechanismen auf allen Ebenen der biologischen Organisation zu verstehen, um prognostizieren zu können, wie sich die globale Erwärmung möglicherweise auf die tierische Leistung und die Populationsdynamik auswirkt (Harrods et al. 2007). Da die Fische Wechselblüter sind (Jobling 1997), entspricht ihre Körpertemperatur nahezu der Temperatur der Umgebung. Die Temperatur beeinflusst physiologische Prozesse von der Proteinschädigung bis hin zur Membranflüssigkeit und Organfunktion (Hochachka und Somero 2002), indem sie im Einzelfalle für die enzymatischen, physiologischen und biochemischen Werte maßgeblich ist (Clarke, 1993; Coutant 1987; Regier et al. 1990, Pörtner und Knust 2007). Einige Fische sind in der Lage, extrem geringe Temperaturschwankungen wahrzunehmen und darauf zu reagieren (Brown 2003). Fische wählen ihre thermischen Habitate in der Regel unter dem Gesichtspunkt der Maximierung ihrer Wachstumsrate (Magnuson et al. 1979) und können ihre physiologische, ökologische und reproduktive Leistung optimieren (Coutant, 1987). Der für wärmebezogene Anpassungsprozesse erforderliche Energieaufwand und der allgemeine Energiehaushalt bestimmen maßgeblich das „thermische Fenster“, das die Fische tolerieren können, und beeinflussen Wachstumsleistung, Entwicklung, Fruchtbarkeit und Rekrutierung (Pörtner und Knust 2007) und damit auch die Ökologie und Physiologie der Fische (Brett 1956; Ferguson 1958; Fry 1971; Magnuson et al. 1979). Treffen Fische auf Habitate mit ungeeigneten Wärmeregimes, dann reagieren sie darauf und versuchen in Gebiete zu gelangen, wo die Temperatur eher die von ihnen bevorzugten Werte erreicht. Das hat Konsequenzen für ihre Biogeographie( 46), da sie nicht über die physiologische Fähigkeit der Regulierung ihrer Körpertemperatur verfügen (Jobling, 1997; Sims et al. 2006). Bei der Überschreitung von Temperaturgrenzen kann es zur Schwächung des Immunsystems gestresster Individuen kommen, was letztendlich zum Ausbruch von Krankheiten führt (Harvell et al. 1999). (46) Beschreibt die Verbreitung von Organismen auf der Erde und untersucht die Ursachen der geographischen Verbreitung lebender und ausgestorbener Taxa. 35 PE 379.208 Klimawandel und europäische Fischerei Die thermische Nische einer Fischart lässt sich anhand von Kriterien bestimmen, die die mögliche tödliche Wirkung dieser Nische, ihre kontrollierende und ihre ausrichtende Wirkung betreffen (Fry, 1971). Extreme Temperaturen beispielsweise können bei Überschreitung bestimmter Grenzen tödlich sein (Jobling 1981), die Temperatur richtet jedoch auch das Verhalten aus (Reynolds, 1977) und regelt die individuelle Physiologie (Regier et al. 1990). Eine Vorhersage der Auswirkungen von Temperaturveränderungen auf die Fische ist schwierig, da es bei den Einzeltieren eine enorme Bandbreite möglicher Reaktionen gibt und potenziell noch viele andere physiologische oder ökologische Faktoren Einfluss nehmen können (Burton, 1979; Fry, 1971). Da die Temperatur einen solch großen Einfluss auf die Biologie einzelner Fische nimmt, wird sie natürlich auch ökologische Konsequenzen für Fischpopulationen (z. B. Mills und Mann, 1985) und –gemeinschaften (z. B. Persson, 1986; Southward et al. 1988) haben. Die durch den Klimawandel verursachten Temperaturveränderungen können zu einer Veränderung der Durchschnittstemperatur oder zu einer stärkeren Temperaturvariabilität führen. Extrem hohe und niedrige Temperaturen können sich dabei auf die Artengemeinschaft viel stärker auswirken als eine Veränderung der Durchschnittstemperatur (IPCC-Bericht 2001, Abbildung 11). PE 379.208 36 Klimawandel und europäische Fischerei Abbildung 11: Auswirkungen von Temperaturextremen. Quelle: IPCC 2001 Synthesis Report. Anmerkungen: Schematisches Diagramm zur Darstellung der Auswirkungen auf Temperaturextreme wenn (a) die Durchschnittstemperatur ansteigt (mehr extremheißes Wetter) (b) die Varianz ansteigt und (c)sowohl der Durchschnitt als auch die Varianz ansteigen (viel mehr extremheißes Wetter). 37 PE 379.208 Klimawandel und europäische Fischerei Legende: Probability of occurrence Increase in mean temperature Increase in variance of temperature Increase in mean and variance of temperature Previous climate New climate Cold Hot Average Less/more cold weather More hot weather More record cold/hot weather Much more hot weather Less change for cold weather Wahrscheinlichkeit Anstieg der Durchschnittstemperatur Anstieg der Temperaturvarianz Anstieg des Durchschnitts und der Varianz der Temperatur Früheres Klima Neues Klima Kalt Heiß Durchschnitt Weniger oft/häufiger kaltes Wetter Häufiger heißes Wetter Häufiger extremkaltes/extremheißes Wetter Viel häufiger heißes Wetter Weniger Veränderung zu kaltem Wetter Die wichtigsten industriell genutzten bzw. für die EU-Fischerei wirtschaftlich bedeutsamen Arten repräsentieren eine Reihe unterschiedlicher Life-History-Traits, d. h. es handelt sich um pelagische Arten wie auch um Grundfische( 47), die in gemäßigten bis hin zu subtropischen Habitaten vorkommen. Um die wichtigsten biologischen Merkmale dieser Hauptfischarten darstellen zu können, wurde auf FishBase, die größte Fischdatenbank, (Fröse und Pauly, 2007) zurückgegriffen. Anhang 1-1 enthält für jede Art Informationen zu folgenden Punkten: geographische Verbreitung maximale Länge und Höchstalter Lebensraum (Wasserzone, -art und -tiefe) Temperaturbereich Mindestzeit für die Verdoppelung der Population. Die einzelnen Angaben wurden erforderlichenfalls korrigiert oder ergänzt. Die Karten zu den bekannten Verbreitungsgebieten wurden bei Muus und Nielsen (1999) entnommen. Kernaussage 7 Temperaturveränderungen können die Verlagerung von Fischpopulationen, das Eindringen fremder Arten und das Verschwinden von Arten zur Folge haben. Bei den Fischen ist die Temperatur eine ganz wesentliche Nischen-Komponente. Fische wählen in der Regel Habitate, deren thermische Bedingungen eine größtmögliche Wachstumsrate ermöglichen. Allerdings lassen sich die Auswirkungen von Temperaturveränderungen auf die Fische nur schwer vorhersagen. Abgesehen von der Temperatur entscheiden auch noch das Nahrungsangebot und das Vorhandensein geeigneter Laichplätze über die Verbreitung der Fischbestände. Größe und Verbreitung der bestehenden und neuen Populationen Ein Anstieg der Wassertemperatur beeinflusst das Leben der Meeresorganismen sowohl direkt als auch indirekt. Direkte physiologische Auswirkungen zeigen sich bei Unter- oder Überschreitung der Grenze des Toleranzbereichs für eine Art. Indirekte Auswirkungen der steigenden Wassertemperatur werden beobachtet, wenn die zuvor zeitlich und räumlich verfügbaren, als Nahrung dienenden Organismen nicht mehr vorhanden sind, weil sich aufgrund der Temperaturveränderung die Artenzusammensetzung eines Ökosystems ändert. (47) Leben in enger Beziehung mit dem Meeresgrund und hängen von ihm ab. PE 379.208 38 Klimawandel und europäische Fischerei Über die Verbreitung der Fischbestände entscheiden die Temperatur, das Nahrungsangebot und das Vorhandensein geeigneter Laichplätze. Da Fischarten oder -bestände einen speziellen Temperaturbereich bevorzugen, fällt eine vermehrte bzw. reduzierte Verbreitung einer Art oft mit langfristigen Temperaturveränderungen zusammen. Am auffälligsten sind diese Veränderungen an den nördlichen bzw. südlichen Grenzen des Bereichs einer Art. Die Erwärmung hat zu einer Verschiebung der Verbreitungsgebiete nach Norden geführt, während die Abkühlung des Wassers die Arten in Richtung Süden wandern lässt (Rose 2005). Die Auswirkungen des Klimawandels auf die Phänologie, Abundanz und Verbreitung der terrestrischen Taxa ist gut dokumentiert (Parmesan, 2006; Parmesan und Yohe, 2003; Root et al. 2003; Walther et al. 2002). In einer Metaanalyse berechneten Parmesan und Yohe (2003), dass Frühjahrsereignisse (z. B. der Nestbau der Vögel und das Brüten der Frösche) pro Jahrzehnt im Schnitt 2,3 Tage früher stattfanden und es innerhalb desselben Zeitraums zu einer erheblichen Verschiebung des Verbreitungsgebiets verschiedener Taxa um durchschnittlich 6,1 km in Richtung der Pole kam. Zur Untersuchung der Auswirkungen auf die Fische verwendeten Parmesan und Yohe (2003) Daten von Holbrook et al. (1997) und Southward et al. (1995), die eine Zeitreihe von 70 Jahren im Ärmelkanal analysiert hatten. Sie stellten fest, dass Größe und Verbreitung der Fische und des Zooplanktons sich im Verhältnis zur Erhöhung der Meerestemperatur verändert hatten. In Zeiten ansteigender Meerestemperaturen verzeichneten Warmwasserarten einen Populationsanstieg um das Zwei- bis Dreifache, wobei Verschiebungen in Bezug auf die Breitengrade von bis zu 120 Meilen nordwärts stattfanden. Holbrook et al. (1997) untersuchten darüber hinaus die Zusammensetzung von Fischen in einem gemäßigten Riff und stellten eine enge Verbindung zu einer klimatischen Verschiebung her. Der Artenreichtum ging zurück und die Zusammensetzung änderte sich dahingehend, dass nicht mehr die nördlichen, sondern die südlichen Arten dominierten. Perry et al. (2005) untersuchten die Verschiebungen der Grenzen anhand von mehr als 60 verschiedenen Nordseefischarten. Sie fanden im Wesentlichen heraus, dass sich bei der Hälfte der Arten die Grenzen verschoben hatten, wobei ein Trend in Richtung Norden zu verzeichnen war. Lediglich eine Art, der Stindtdorsch, bildete eine Ausnahme, da er ein wenig Richtung Süden in kältere Gewässer wanderte, um dem Nordatlantikstrom zu entgehen, der warmes Wasser in die nördliche Nordsee bringt. Am stärksten von der Verschiebung betroffen war der Blaue Wittling, dessen südliche Grenze sich um 816 km in Richtung Norden verschob. Nicht jede Art reagierte jedoch mit einer Verschiebung in Bezug auf die Breitengrade. Die Schollen wanderten beispielsweise nur in tiefere, kühlere Gewässer. Hawkins et al. (2003) wiesen zudem eine Verlagerung der pelagischen Arten Hering, Sardine und Makrele nach. 39 PE 379.208 Klimawandel und europäische Fischerei Abbildung 12: Quelle: Anmerkungen: Auswirkung der Temperatur auf die Kabeljaubestände Drinkwater 2005. Zu erwartende Änderungen in Bezug auf die Abundanz der Kabeljaubestände bei einem Temperaturanstieg von (a) 1 C, (b) 2 C, (c) 3 C, und (d) 4 C gegenüber dem derzeitigen Niveau. Legende: North Atlantic Cod Stocks Distribution Spawning Increase No change Decrease Collapse Kabeljaubestände im Nordatlantik Verbreitung Laichgründe Bestandsanstieg Keine Veränderung Bestandsrückgang Bestandszusammenbruch Fangdaten von schottischen Forschungsschiffen vom Januar 1995 zeigen, dass bei den pelagischen Warmwasserarten Sardelle (Engraulis encrasicolus), Sardine (Sardina pilchardus) und Meerbarbe, also bei normalerweise im nördlichen Europa nur selten vorkommenden Arten, PE 379.208 40 Klimawandel und europäische Fischerei die Fangquoten nach 1995 plötzlich stark angestiegen sind (Beare et al. 2004). Dies steht nach Ansicht der Autoren mit dem Anstieg der Meerestemperaturen in Zusammenhang, obwohl der genaue kausale Mechanismus nicht klar ist. Da all diese Arten stark befischt werden, ist der Einfluss der Wassererwärmung auf die Schwankungen der Abundanz unklar und rein spekulativ und erweist sich eine Unterscheidung der direkten Kausalzusammenhänge zwischen Temperatur des Lebensraums und Verbreitungsmuster als schwierig (Jensen 2003). Dennoch bewiesen Maravelia et al. (1995) sowie Misund et al. (1998) den Einfluss von Wassertemperatur und Salinität auf die Migration des Herings, machten jedoch beide geltend, dass der Einfluss aufgrund einer Verlagerung des Beuteangebots auch indirekter Natur sein könnte. Wichtig ist auch, dass die Untersuchung von Perry et al. (2005) bei den befischten wie auch den nicht befischten benthonischen und benthisch-pelagischen Arten zu ähnlichen Ergebnissen gelangte, was darauf hindeutet, dass die Reaktion auf den Temperaturanstieg nicht von der Tatsache beeinflusst wurde, ob die Art befischt wurde oder nicht. Hannesson (2007) analysierte den Kabeljau-, Hering-, Makrelen-, Sardellen- und Sardinenfang in der Nordsee und dem Europäischen Nordmeer. Ihm zufolge ist es aufgrund der starken Befischung von Hering und Kabeljau in der Nordsee schwer festzustellen, ob die Erwärmung sich auf die Größe beider Artenbestände auswirkt. Ein starker Anstieg der Makrelenanlandungen Ende der 1960er Jahre wurde auf veränderte Fangtechniken zurückgeführt. Dennoch wurde ein darauffolgender Bestandsanstieg in den 1970er Jahren mit der Erhöhung der Wassertemperatur in Verbindung gebracht. Bei den Sardellen ließ sich kein entsprechender Zusammenhang feststellen, während die Zunahme des Sardinenbestands mit der Wassertemperatur in Verbindung gebracht werden konnte. Die wärmere Wassertemperatur wirkte sich offenbar auch positiv auf die Rekrutierung des Kabeljaus in norwegischen Gewässern, jedoch negativ auf die Nordsee aus. Eine Wanderung des Kabeljaus in Richtung Norden wurde aufgrund eines nicht vorhandenen Anstiegs der Fangmengen um Europäischen Nordmeer verworfen. Arnott und Ruxton (2002) prognostizieren eine Verschiebung der südlichen Grenze der Sandaalpopulation in der Nordsee in Richtung Norden, sollten die Temperaturen ansteigen. Eines der wenigen Beispiele einer Prognose zur möglichen Entwicklung der Kabeljaubestände bei einem Temperaturanstieg wurde von Drinkwater (2005, Abbildung 12) angeführt. Er verband die bestehenden Erkenntnisse zu den Auswirkungen der Klimaschwankungen auf den Kabeljau (Gadus morhua) mit Prognosen zu den künftigen klimatischen Veränderungen, um die Verschiebung der Verbreitung des Kabeljaubestands im Verhältnis zum Anstieg der Wassertemperatur vorherzusagen. Für seine Untersuchungen wählte er den Kabeljau, da dieser zu den am meisten erforschten Arten im Nordatlantik zählt und über den Lebenszyklus und die Biologie dieser Art mehr bekannt ist als über die der meisten anderen Fischarten. Oberhalb einer Durchschnittstemperatur von 12 °C am Meeresboden wird der Kabeljau nur noch selten angetroffen (Dutil & Brander 2003). Steigt die Temperatur am Meeresboden auf über 12 °C, wird der Kabeljau verschwinden, indem er in kühlere Gewässer wandert oder stirbt. Für die Gewässer des Festlandsockels, in denen verschiedene Kabeljaubestände leben, wird bis zum Jahr 2100 ein Anstieg der durchschnittlichen Jahrestemperatur von 2 °C bis 6 °C vorhergesagt, wobei die Kabeljaubestände in der Nordostarktis die größte Zunahme verzeichnen werden. Für den Kabeljau in der Nordsee wird sich die Temperatur um 2 °C bis 3 °C, für den Kabeljau in der Ostsee unter Umständen um 3 °C bis 4 °C erhöhen. Die Kabeljaubestände in der Nordsee haben heute ihren temperaturbezogenen Toleranzbereich erreicht, was dazu führt, dass die Population sich in Richtung Norden bewegt (Drinkwater 2005, Abbildung 12). 41 PE 379.208 Klimawandel und europäische Fischerei Wie Pörtner und Knust (2007) anhand von Datensätzen aus dem Zeitraum 1954-1989 untersuchten, hat die relative Abundanz der nicht wandernden Aalmutterpopulationen (Zoarces viviparus) in der Nordsee im Verhältnis zur Erwärmung abgenommen, wobei die Fische in heißen Sommern eine hohe Mortalitätsrate aufwiesen. Da diese Fischart keine Wanderungen durchführt, kann sie, wenn die Temperatur ihren Toleranzbereich überschreitet, nicht entkommen und stirbt. Genner et al. (2004) untersuchten die Auswirkungen des regionalen Klimawandels auf die Artenzusammensetzung anhand zweier unabhängiger Datensätze aus dem Ärmelkanal (19132002) und dem Kanal von Bristol (1981-2002). Sie konnten beweisen, dass die klimatischen Veränderungen sich dramatisch auf die Zusammensetzung der Fischgemeinschaft auswirkten, da es zu Verschiebungen in Richtung Norden kam. Es trat jeweils eine Teilmenge dominanter Arten auf, deren Abundanz in strengem Zusammenhang mit der Jahresdurchschnittstemperatur der Meeresoberfläche stand. Der breitengradbezogene Bereich der Arten eignete sich nicht als gute Prognose für die Reaktionen in Bezug auf den Artenbestand, da ein und dieselbe Art in den verschiedenen Gebieten keine übereinstimmenden Entwicklungen aufwies. Daher weisen Genner et al. (2004) darauf hin, dass Populationen derselben Art innerhalb einer Region unterschiedlich auf die klimatischen Veränderungen reagieren können, was möglicherweise eine Reaktion auf zusätzliche lokale Umweltfaktoren, interspezifische(48) ökologische Interaktionen und die Ausbreitungsfähigkeit darstellt. Das erschwert die Vorhersage, wie sich die Arten innerhalb geographisch unterschiedlicher Gemeinschaften in Bezug auf ihren Bestand entwickeln werden. Mehrmals wurden Warmwasserfischarten in der „kalten Biota“ des Mittelmeers gefangen (Francour et al. 1994). Aus dem östlichen und südlichen Mittelmeer stammende Fischarten wurden in den nördlichen Bereichen des Tyrrhenischen Meeres (Golfe du Lion und Korsika) gesichtet. Umgekehrt sind Kaltwasserarten wie die Sprotte (Sprattus sprattus) und Makrele (Scomber scombrus), die in den nördlichen Bereichen des Mittelmeers (Nordadria und Golfe du Lion) relativ weit verbreitet waren, sehr selten geworden oder ganz verschwunden (Bombace 2001, Dulcic et al. 1999). Anfang bis Mitte der 1990er Jahre, als die Wasseroberflächentemperaturen in der Adria bei +0,30 °C über den normalen Temperaturen lagen, traten dort die meisten neuen Arten erstmals auf. In den letzten 25 Jahren wurden in der Adria elf subtropische und tropisch-thermophile Arten beobachtet, deren Bestand kontinuierlich zugenommen hat (Dulcic et al. 1999). Kernaussage 8 Direkte und indirekte Klimaeffekte können zu einer Verlagerung von Fischpopulationen, zum Eindringen fremder Arten und selbst zum Verschwinden von Arten führen. In verschiedenen Studien wurde ein Zusammenhang zwischen Größe und Verteilung von Fischbeständen und Zooplankton und einem Anstieg der Meerestemperatur beobachtet. Viele Industriefischarten sind direkt vom Plankton abhängig. Bei Fischarten, die sich von Plankton ernähren, insbesondere Sardinen und Sardellen, gibt es eine starke natürliche Fluktuation im Zusammenhang mit Klimaveränderungen. Untersuchungen bezüglich der Klimaerwärmung ließen erkennen, dass statt der nördlichen Arten in zunehmendem Maße die südlichen Arten dominieren. Bei der Untersuchung der Grenzen von mehr als 60 verschiedenen Nordseefischarten zeigte sich bei der Hälfte dieser Arten (befischt und nicht befischt) eine Verschiebung der Grenzen, wobei die Tendenz nach Norden geht. Einige Arten dürften ihre Belastbarkeitsgrenze erreicht haben, wie etwa der Kabeljau in der Nordsee, (48) Zwischen den verschiedenen Arten, z. B. Konkurrenzkampf. PE 379.208 42 Klimawandel und europäische Fischerei weshalb sich ihre Populationen nach Norden bewegen. Der Rückgang beim Kabeljau wurde mit veränderten Artenzusammensetzungen, Bestandsabnahme und einer kleineren durchschnittlichen Körpergröße des Zooplanktons in Zusammenhang gebracht, was sich möglicherweise auf den Klimawandel zurückführen lässt. Die Verlagerung von Populationen (wie am Beispiel des Kabeljau gezeigt) kann dazu führen, dass in einzelnen Regionen manche Bestände gänzlich verschwinden. Warmwasserfische dringen in „kalte“ Ökosysteme vor. Verschiedene Arten von Warmwasserfischen sind in „kalte“ Ökosysteme vorgedrungen, während Kaltwasserfische, die in „warmen“ Ökosystemen in relativ großer Zahl vorhanden waren, inzwischen sehr selten geworden oder gänzlich verschwunden sind. So gibt es z. B. im Mittelmeer kaum noch Sprotten und Makrelen Auch in vielen anderen Fällen wurden Verschiebungen festgestellt. Da jedoch die betreffenden Arten oftmals stark befischt werden, lässt sich nur schwer ein direkter Kausalzusammenhang zwischen Temperatur und Verbreitungsmuster herstellen. Zuverlässige Prognosen zu möglichen Entwicklungen von Fischbeständen, die auf Effekte des Klimawandels zurückzuführen sind, lassen sich nur für einige bislang sehr intensiv untersuchte Arten anstellen (z. B. Kabeljau). Eine Abgrenzung gegenüber anderen Wirkungsfaktoren ist schwierig. Weitere Forschungsarbeiten sind erforderlich. Geographische Verbreitungsgebiete oder die Verbreitung von Fischpopulationen Änderungen bei den geographischen Verbreitungsgebieten der Arten hängen von den „bioklimatischen Rahmenbedingungen“ oder „Klimaräumen“ ab, die mit Hilfe von Verfahren zur Korrelation der derzeitigen Verbreitung der Arten mit Klimavariablen bzw. durch Ermittlung der physiologischen Reaktion des Einzeltiers auf Klimaänderungen bestimmt werden (Pearson und Dawson 2003). Es wird angenommen, dass der Klimawandel einen direkten Einfluss auf Überlebensrate, Ausbreitung, Fertilität und Verhalten der Einzeltiere und damit auf die Größe und Verbreitung der Bestände hat (Walther et al. 2002) hat. Um die Bewegung einer Art vorhersagen zu können, sind Erkenntnisse über ihre klimatischen Rahmenbedingungen erforderlich, die dann auf Szenarien künftiger Klimaänderungen angewandt werden. Davon ausgehend kann die potenzielle Neuverbreitung der betreffenden Art abgeschätzt werden. Die Gültigkeit eines solchen auf den bioklimatischen Rahmenbedingungen beruhenden Ansatzes( 49) wird in Frage gestellt, da außer dem Klima noch viele andere Faktoren für die Verbreitung der Arten und die Dynamik dieser Veränderungen von großer Bedeutung sind. Biotische Interaktionen, der evolutionäre Wandel und die Fähigkeit zur Dispersion müssen berücksichtigt werden. Daher kann das auf den bioklimatischen Rahmenbedingungen beruhende Modell die potenziellen Auswirkungen des Klimawandels nur annähernd darstellen. Es gilt zu berücksichtigen, dass die Komplexität des natürlichen Systems den Modellierungsstrategien grundlegende Grenzen setzt. Eine Schwäche korrelativer Ansätze besteht darin, dass die betreffenden Korrelationen möglicherweise künftig nicht gelten, da die Bedingungen, insbesondere die Beziehungen zwischen den Arten sich ändern können (Pearson und Dawson 2003). (49) Wobei die entsprechenden Rahmenbedingungen durch Korrelation der derzeitige Verteilung der Arten mit Klimavariablen bzw. durch Ermittlung der physiologischen Reaktion des Einzeltiers auf Klimaänderungen bestimmt werden. 43 PE 379.208 Klimawandel und europäische Fischerei Um ausgehend von früheren Trends Aussagen über wahrscheinliche künftige Entwicklungen zu treffen, unterzog Drinkwater (2006) die Auswirkungen eines dramatischen Anstiegs der Luftund Wassertemperaturen im nördlichen Nordatlantik und der hohen Arktis in den 1920er und 1930er Jahren einer erneuten Analyse. Eine verstärkte Rekrutierung und ein größeres Wachstum führten in vielen Regionen des Nordatlantiks zu einem Anstieg der Biomasse von kommerziell bedeutsamen Arten wie Kabeljau und Hering. Andererseits wanderten Kaltwasserarten wie Lodde nicht mehr so weit südwärts. Es wurde ein verstärktes Aufkommen von atlantischem Kabeljau und Heilbutt (Hippoglossus hippoglossus) entlang der Westküste Grönlands als Reaktion auf das veränderte Ozeanklima dokumentiert (Jensen 1939, zitiert in Drinkwater 2006, Abbildung 13). Es wird angenommen, dass das plötzliche Auftreten von Kabeljau darauf zurückzuführen ist, dass infolge der Zunahme des Irmingerstroms eine verstärkte Larvendrift von Island zu verzeichnen war und dass die Larven, nachdem sie grönländische Gewässer erreicht hatten, besser überleben konnten. Abbildung 13: Kabeljau. Die Auswirkungen von Warmperioden auf die Verbreitung des Quelle: Drinkwater 2006. Anmerkungen: Die Verbreitung des Kabeljau (rot gekennzeichnet) entlang der westgrönländischen Küste von 1900 bis Ende der 1930er Jahre; während der warmen Periode ist eine Ausweitung in Richtung Norden zu verzeichnen. Legende: Greenland - Grönland PE 379.208 44 Klimawandel und europäische Fischerei Die Schellfischfänge vor Westgrönland sind ein weiterer Beleg für einen verstärkten Larventransport, da diese Art nicht westlich von Grönland laicht und von Island stammen muss (Dickson und Brander 1993). Während der Warmperioden in den 1920er und 1930er Jahren breiteten sich Katfisch (Anarchichas minor) Hering (Clupea harengus) sowie Miesmuscheln (Mytilus edulis) und Seesterne (Asterias) nach Norden hin aus und vermehrten sich erfolgreich nördlich ihrer vorherigen Ausbreitungsgebiete. Das Migrationsverhalten von in wärmerem Wasser lebenden Arten hat sich ebenfalls geändert, so dass die Ankunft nunmehr früher erfolgt und die Rückwanderung zu einem späteren Zeitpunkt angetreten wird. Diese Studien zeigen, dass der Temperaturanstieg nicht nur neue Gebiete für Fische attraktiv gemacht hat, sondern dass der Klimawandel darüber hinaus zu einer Änderung bei den bestehenden Systemen geführt hat, die sich auf die Larventransportrouten auswirkten. Die Hauptursache für diese Veränderungen waren wahrscheinlich in der Tiefe ausgelöste Prozesse infolge einer vermehrten Phyto- und Zooplanktonbildung in verschiedenen Gebieten. Im Ergebnis einer Untersuchung der Reaktion auf den Temperaturanstieg in den 1920er und 1930er Jahren im Vergleich zur heutigen Situation kann festgestellt werden, dass sich einige der beobachteten Veränderungen wiederholen. Als die Temperatur im Nordatlantik in den 1960er Jahren absank, stellten sich wieder die vorherigen Umweltbedingungen ein. In einigen Regionen haben sich jedoch neue Systeme angesiedelt. So nahm beispielsweise die Garnelenbiomasse vor Westgrönland zu und die Garnelenfischerei erlangte vorherrschende wirtschaftliche Bedeutung und trat an die Stelle der Kabeljaufischerei (Drinkwater 2006). Ähnliche Tendenzen mit Blick auf die Kabeljaufischerei sind in der kanadischen Provinz Neuschottland (Nova Scotia) zu beobachten, wo die vormals von Kabeljau dominierte kommerzielle Fischerei nunmehr vorrangig auf Garnelen und Arktische Seespinnen ausgerichtet ist. Verschiedene Bewirtschaftungsmaßnahmen (Schließung der Fischerei, neuartige Fangmethoden zur Rettung des Kabeljau), mit denen der Trend umgekehrt und das System in seinen ursprünglichen Zustand zurückgeführt werden sollte, sind gescheitert. Auf die Frage, ob die jüngsten Veränderungen umkehrbar sind, gibt es nach wie vor keine Antwort (Frank et al. 2005). Kernaussage 9 Der Klimawandel beeinflusst Größe und Verbreitung der Industriefischbestände. Das sich verändernde Klima hat einen direkten Einfluss auf Überlebensrate, Ausbreitung, Fertilität und Verhalten der Einzeltiere und damit auf die Größe und Verbreitung der Bestände. Prognosen sind schwierig, da außer dem Klima noch viele andere Faktoren für die Verbreitung der Arten und die Dynamik dieser Veränderungen von großer Bedeutung sind. Veränderungen in der geographischen Streuung haben um Grönland herum zu einer erhöhten Produktivität bei Kabeljau und Schellfisch beigetragen. Das ist eine Reaktion auf eine dramatische Erwärmung in den 20er und 30er Jahren des vorigen Jahrhunderts. Die Hauptursache für diese Veränderungen jedoch waren in der Tiefe ausgelöste Prozesse infolge einer vermehrten Phyto- und Zooplanktonbildung. Wachstumszeiten, Wintersterblichkeit und Wachstumsraten Die Umwelt wirkt sich maßgeblich auf das Wachstum von Fischen aus. Brander (1994, 1995) gelang der Nachweis, dass die beobachteten Unterschiede bei der Wachstumsrate zwischen verschiedenen Beständen von Atlantischem Kabeljau im Nordatlantik zu 90 % auf die mittleren Temperaturen am Meeresboden zurückzuführen sind, wobei Kabeljau aus wärmeren Gewässern höhere Wachstumsraten aufweist. Die Temperatur ist jedoch nicht nur für Unterschiede hinsichtlich der Wachstumsraten zwischen den Beständen, sondern auch dafür ausschlaggebend, dass innerhalb eines Bestands von Jahr zu Jahr Änderungen zu verzeichnen sind (Brander 2001), d. h. sie kann eine höhere Produktivität der Kabeljaubestände bedingen (Dutil und Brander 2003). 45 PE 379.208 Klimawandel und europäische Fischerei Um die wahrscheinlichen Auswirkungen des Klimawandels auf die Fischbestände zu untersuchen, sind großangelegte Klimamodelle in Verbindung mit Simulationen der Fischbestände erforderlich. Clark et al. (2003) haben Projektionen aus dem allgemeinen Zirkulationsmodell von Hadley für den Zeitraum 2000-2050 verwendet, um die wahrscheinlichen Auswirkungen des Klimawandels auf die Kabeljaubestände in der Nordsee abzuschätzen. Die Auswirkungen der Temperatur wurden mittels des glockenförmigen Verhältnisses Temperatur – Wachstum für die verschiedenen Größenklassen unter Verwendung der von Bjornsson und Steinarsson (2002) bereitgestellten Parameter berücksichtigt. Die mittlere Temperatur am Boden der Nordsee betrug im Zeitraum 1998-2000 etwa 8,67 °C. Da die optimale Temperatur für das Wachstum des Kabeljau bei ungefähr 8,5 °C liegt (Bjornsson und Steinarsson 2002), wird ein Anstieg der Umgebungstemperatur über die derzeit in der Nordsee beobachteten Werte hinaus nicht zu einem höheren Wachstum führen und keine Erhöhung der Rekrutierung bzw. der Biomasse der Laichbestände bewirken (Clark et al. 2003). Obwohl Clark et al. (2003) darauf hinweisen, dass die für die Nordsee entwickelten Simulationen auf relativ simplen Ansätzen beruhen und zahlreiche weitere potenziell bedeutsame Prozesse außer Acht lassen, veranschaulichen die Ergebnisse, dass die Einbeziehung von Umweltfaktoren in Populationsmodelle die Prognosen hinsichtlich des Verhaltens von Fischpopulationen ändern können. Diese Simulationen können wertvolle Aufschlüsse für die Bewirtschaftung der Fischbestände geben, und sie machen deutlich, dass dabei die Berücksichtigung von Umweltauswirkungen immer wichtiger wird (Jurado-Molina und Livingstone 2002). Kernaussage 10 Die mittleren Temperaturen am Meeresboden können die Wachstumsrate eines Fischbestandes wesentlich beeinflussen. Wie simple Simulationen für den Nordseekabeljau gezeigt haben, ist bei steigenden Meeresbodentemperaturen mit negativen Folgen für die Wachstumsrate zu rechnen. Solche Ansätze zeigen, dass sich die Prognosen hinsichtlich des Verhaltens von Fischpopulationen ändern können, wenn bei den entsprechenden Modellen Umweltfaktoren mit einbezogen werden. Simulationen können wertvolle Aufschlüsse für die Bewirtschaftung der Fischbestände geben, und sie machen deutlich, dass dabei die Berücksichtigung von Umweltauswirkungen wahrscheinlich immer wichtiger wird. Reproduktionsmuster, Wanderungswege und Ökosystembeziehungen Die unmittelbaren Auswirkungen des Klimawandels betreffen die Leistung der Einzeltiere in verschiedenen Lebensphasen durch Änderungen in Physiologie, Morphologie und Verhalten (Harley et al. 2006). Klimaauswirkungen machen sich auch auf der Ebene der Populationen bemerkbar, durch Änderungen bei den Transportprozessen, die einen Einfluss auf Ausbreitung und Rekrutierung haben. Die Auswirkungen auf Gemeinschaftsebene werden durch interagierende Arten vermittelt und umfassen klimabedingte Änderungen bei der Abundanz und der Stärke der Interaktion der Arten (Abbildung 14). PE 379.208 46 Klimawandel und europäische Fischerei Abbildung 14: Potenzielle ökologische Reaktionen auf den Klimawandel. Quelle: Harley et al. 2006. Anmerkungen: Der Lebenszyklus einer generischen Meeresart ist in Grün dargestellt. Abiotische Veränderungen in der Umwelt haben direkte Auswirkungen (gelbe Boxen) auf Ausbreitung und Rekrutierung und auf die individuelle Leistung in den verschiedenen Phasen des Lebenszyklus. Weitere Auswirkungen sind auf Gemeinschaftsebene durch Änderungen in der Populationsgröße und Pro-Kopf-Auswirkungen bei interagierenden Arten zu verzeichnen (in Blau). Somit umfassen die unmittelbaren ökologischen Auswirkungen des Klimawandels Verlagerungen bei der Leistung von Einzeltieren, der Dynamik von Populationen und der Struktur von Gemeinschaften. Insgesamt führen diese Auswirkungen dazu, dass sich neue Muster - wie zum Beispiel Änderungen in Verbreitung, Biodiversität, Produktivität der Arten und mikroevolutionäre Prozesse - ergeben. Legende zu Abbildung 14 Proximate ecological responses Larvae or propagules Physiological, morphological and behavioral effects Transport processes Changes in per capita interaction strengths Changes in population size of interacting species Adults Emergent ecological responses Distributional shifts Changes in diversity Changes in productivity Microevolutionary change Annähernde ökologische Reaktionen Larven oder Propagulae Physiologische, morphologische und verhaltensbezogene Auswirkungen Transportprozesse Veränderungen bei der Stärke der Pro-KopfInteraktionen Veränderungen bei der Populationsgröße interagierender Arten Geschlechtsreife Fische Sich ergebende ökologische Reaktionen Verlagerungen bei der Verbreitung Veränderungen hinsichtlich der Vielfalt Veränderungen hinsichtlich der Produktivität Mikroevolutionärer Wandel 47 PE 379.208 Klimawandel und europäische Fischerei Um den Ausgleich zwischen Fortpflanzung, Wachstum und Überleben in Gegenwart und Zukunft zu gewährleisten, zahlt sich die Teilnahme am Laichen, was die Tauglichkeit betrifft, nur dann aus, wenn die Fische groß genug und in einem ausreichend guten Zustand sind. Im Falle des Norwegischen Frühjahrslaichers (Hering) wurde beobachtet, dass Fischpopulationen nicht am Laichprozess teilnahmen (Engelhard und Heino 2006), und es wurde festgestellt, dass dies nicht nur mit der Größe der Fische und ihrem Zustand, sondern auch mit klimatischen Faktoren und den Meeresoberflächentemperaturen zusammenhing. Engelhardt und Heino (2006) gelangten zu der Erkenntnis, dass in kälteren Jahren, wenn C. finmarchicus, die wichtigste Nahrungsquelle des Herings, in reichlicherem Maße vorhanden ist (Fromentin und Planque 1996), bessere Futterbedingungen, bessere Überlebenschancen und eine stärkere Teilnahme an der Fortpflanzung zu verzeichnen sind. Ein klimatisch bedingter Anstieg der Wassertemperatur hat zur Folge, dass Fischpopulationen nicht am Laichprozess teilnehmen( 50) und kann daher indirekte Auswirkungen auf das Fortpflanzungspotenzial der betreffenden Populationen haben und für die kommerzielle Heringsfischerei in der Barentssee von Bedeutung sein. Darüber hinaus fanden Sissener und Bjoerndal (2006) Belege für Änderungen beim Migrationsverhalten des Norwegischen Frühjahrslaichers (Hering), die auf klimatische Einflüsse zurückzuführen sind (Abbildung 15). (50) Fisch, der bereits an der Fortpflanzung teilgenommen hat, aber eine oder mehrere Fortpflanzungsperioden überspringt. PE 379.208 48 Klimawandel und europäische Fischerei Abbildung 15: Einfluss des Klimawandels auf das Migrationsverhalten. Quelle: Vilhjalmsson 1997, Drinkwater 2006. Anmerkungen: (a) Migrationsmuster des Norwegischen Frühjahrslaichers (Hering), 1950; (b) Migrationsmuster des Norwegischen Frühjahrslaichers (Hering), 1965–1966; (c) Migrationsmuster des Norwegischen Frühjahrslaichers (Hering), 1972–1986; (d) Migrationsmuster des Norwegischen Frühjahrslaichers (Hering), 1995–1999. Legende: Approximate boundary of arctic water Maturing area Main feeding area May–Sep Feeding migration Small and irregular sprawning areas Wintering area Oct-Dez Sprawning migration Jan-Feb Sprawning areas Feb-Mar Annähernde Grenzlinie arktische Gewässer Aufwuchsgebiet Hauptweidegebiet Mai-Sep Weidewanderung Kleine, nicht reguläre Laichgebiete Überwinterungsgebiete Okt-Dez Laichwanderung Jan-Feb Laichgebiete Feb-Mär Feeding area Aug-Sep Maturing area Wintering area Feeding area May Sprawning migration Weidegebiet Aug-Sep Aufwuchsgebiet Überwinterungsgebiet Weidegebiet Mai Laichwanderung 49 PE 379.208 Klimawandel und europäische Fischerei Sprawning areas Laichgebiete Maturing area Wintering area Feeding area Aufwuchsgebiet Überwinterungsgebiet Weidegebiet Adult stock feeding area Weidegebiet für geschlechtsreife Bestände Das Wanderungsverhalten des Norwegischen Frühjahrslaichers (Hering) umfasst vier Phasen: Laichen, Aufzucht, Weiden und Überwintern. Im Januar machen sich die geschlechtsreifen Heringe auf den Weg in die Laichgründe. Die Laichzeit dauert von Februar bis April. Anschließend wandern die geschlechtsreifen Tiere zu den Weidegründen, während die Larven zu den Aufzuchtgründen driften. Im Anschluss an die Weidezeit, die bis Mitte November dauert, wandern die geschlechtsreifen Bestände zu den Überwinterungsgebieten. Diese Phasen laufen normalerweise nach einem festen Rhythmus ab, während es hinsichtlich der geographischen Standorte mit der Zeit zu Verlagerungen gekommen ist (Sissener und Bjorndal 2005). Die Grenzlinie zu den arktischen Gewässern hat Einfluss auf das Wanderungsverhalten des Herings, der sich normalerweise östlich dieser Grenzlinie aufhält. Die Fische kehren entweder in das Gebiet zurück, in dem sie geschlüpft sind, oder wählen ein Laichgebiet aus, dessen Umweltbedingungen ihrer Brut gute Überlebenschancen bieten. Während kalter Perioden beschränken sich die Fortpflanzungsaktivitäten des Roten Thun offenbar auf die angestammten Laichgründe im Mittelmeer. Da die letzten Stadien der Gonadenreifung beim Roten Thun innerhalb eines kurzen Zeitraums temperaturgesteuert ablaufen (Medina et al. 2002), können geeignete Temperaturen die letzten Stadien der Gemetogenese und somit die Fortpflanzung auslösen, was ein Grund dafür sein könnte, dass bei steigenden Temperaturen mehr Standorte entlang den Migrationsrouten des Roten Thun geeignete Laichgründe darstellen können. Das würde bedeuten, dass die Population des Roten Thun sich in warmen Perioden auch außerhalb ihrer angestammten Laichgründe fortpflanzen kann. Das zunehmende Laichen außerhalb der angestammten Laichgründe kann eine Verringerung der Rekrutierung innerhalb der angestammten Laichgründe bewirken. Die von Ravier und Fromentin (2004) analysierten Daten deuten auf eine Art „Opportunistic homing“ - d. h. Wahl der Muttergewässer in Abhängigkeit von den äußeren Gegebenheiten - hin, wobei eine Kombination des „Natal homing“ – d. h. des Laichens in den angestammten Gebieten - mit der opportunistischen Fortpflanzungsstrategie des Roten Thun erfolgt. Kernaussage 11 Der Klimawandel kann sich indirekt sowohl positiv als auch negativ auf die Reproduktion von Industriefischbeständen auswirken, wie an mehreren Beispielen deutlich wird. Im Falle des Norwegischen Frühjahrslaichers (Hering) hat ein klimatisch bedingter Anstieg der Wassertemperatur zur Folge, dass die Fischpopulationen nicht am üblichen Laichprozess teilnehmen. Das wirkt sich indirekt auf das Reproduktionspotenzial aus und hat direkte Folgen für die gewerbliche Fischerei (Bestandsverluste). Der Rote Thun könnte sich während warmer Perioden auch außerhalb seiner traditionellen Laichgründe vermehren, was der Produktivität des Bestandes förderlich wäre. Örtliche Verlagerungen bei Produktionszentren und Mischung von Arten; geographische Verlagerungen und interne Veränderungen in den Ökosystemen Es wurde nachgewiesen, dass Verlagerungen innerhalb der Ökosysteme mit klimatischen Einflüssen zusammenhängen, aber auch auf anthropogene Einflüsse wie starke Befischung oder Umweltverschmutzung zurückzuführen sein können (Steele 2004, Collie et al. 2004, Chavez et al. 2003, Frank et al. 2005). Eine Systemänderung in der Meeresökologie ist eine ständige radikale Änderung der typischen Größe bzw. des typischen Produktivitätsniveaus mehrerer PE 379.208 50 Klimawandel und europäische Fischerei wichtiger Komponenten der Struktur der biologischen Meeresgemeinschaft. Sie erfolgt auf mehreren trophischen Ebenen und in einem geographischen Rahmen, der vom Umfang her zumindest regional ist. Eine veränderte Verbreitung ist oftmals charakteristisch für eine Systemänderung (Drinkwater 2006). Wahrscheinlich führen Temperaturänderungen und andere abiotische Faktoren zu Veränderungen bei den interspezifischen Interaktionen, welche darüber hinaus die Reaktion von Fischen und anderen Taxa auf den Klimawandel beeinflussen (Davis et al. 1998). Da einzelne Taxa unterschiedlich auf den Klimawandel reagieren, ist es offensichtlich möglich, dass durch den Klimawandel bedingte Verlagerungen bei den Interaktionen auf Gemeinschaftsebene (z. B. Räuber-Beute-Beziehungen, Konkurrenz und Parasitismus) die Ökologie der Fische und anderer Taxa beeinflussen (Harley et al. 2006). Klimaänderungen können auf verschiedenen Ebenen Auswirkungen auf Fische haben, d. h. auf der zellularen und der individuellen Ebene sowie auf der Ebene der Lebensstadien, der Population, der Arten, der Gemeinschaft und des Ökosystems. Harley et al. (2006) weisen darauf hin, dass Studien sowohl auf Populations- als auch auf Gemeinschaftsebene durchgeführt werden sollten, da direkte klimatische Auswirkungen auf Einzeltiere sich nicht unmittelbar durch Veränderungen hinsichtlich der Verbreitung und Größe bemerkbar machen. Die Empfindlichkeit des marinen Ökosystems der Ostsee gegenüber Temperaturänderungen wurde nachgewiesen. Es wurde festgestellt, dass die jährlichen Spitzenaufkommen der am zahlreichsten auftretenden Cladoceren in der nördlichen Ostsee in Abhängigkeit von der Wasseroberflächentemperatur schwanken. In den 1990er Jahren gingen höhere Temperaturen in Verbindung mit einer geringeren Salinität mit einer Verlagerung der Dominanz innerhalb der Copepoden-Gemeinschaft auf offener See von Pseudocalanus auf Acartia einher (Möllmann et al. 2003). Bei Sprotten- und Heringspopulationen waren erhöhte Produktions- und Überlebensraten nachweislich von hohen Temperaturen und hohen NAO-Werten begleitet. Die Ende der 1970er Jahre in der nördlichen Ostsee beobachtete Verringerung der Salinität spiegelte sich in einem Rückgang bei den großen neritischen Copepodenarten und in einer Zunahme von Süßwasser-Cladoceren( 51) wider. Dagegen nahm die Populationsgröße bei temperaturempfindlichen Arten (z. H. Acartia) zu. Das rückläufige Wachstum bei Hering und Sprotte hängt mit einer salinitätsbedingten Veränderung in der Copepoden-Gemeinschaft zusammen. Der Bestand des wichtigsten Raubfischs in der pelagischen Nahrungskette der Ostsee, des Dorschs, der normalerweise die Sprotten- und Heringsbestände reguliert, ist zurückgegangen. Darüber hinaus benötigen Dorscheier für den hydrostatischen Auftrieb eine Mindestsalinität von 11,5 PSU, die sie normalerweise in den Regionen der Halokline( 52) in den tiefen Ostseebecken vorfinden (Wieland et al. 1994). Aufgrund der geringen Salinität und der ebenfalls geringen Sauerstoffkonzentration in diesen tiefen Becken sind die Dorscheier häufig letalen Sauerstoffbedingungen ausgesetzt, die das Überleben der Dorschbrut und die anschließende Rekrutierung beeinträchtigen. Der verstärkte Fortpflanzungserfolg der Sprotten hat dazu geführt, dass nunmehr nicht mehr der Dorsch, sondern die Sprotte im System dominiert, was auf eine Systemänderung hindeutet (Köster et al. 2005, Alheit et al. 2005). Die Anlandungen von Anchovis und Sardinen und die Produktivität des Pazifischen Ozeans waren im Laufe der letzten 50 Jahre Schwankungen unterworfen. Mitte der 1970er Jahre ging der Pazifik von einem kalten, vor allem für Sardellen (Anchovis) geeigneten System zu einem warmen System über, in dem Sardinen am stärksten vertreten sind, während Mitte bis Ende der 1990er Jahre wiederum eine Abkühlung erfolgte, die ein höheres Aufkommen von Sardellen zur Folge hatte. (Chavez et al. 2003). (51) Gruppe kleiner Plankton-Krebstiere, die sich vor allem in Süßwasser finden, z. B. Daphnien. Es kommen auch marine Arten vor, z. B. Podon-Arten, die zum Zooplankton zählen. (52) Region unter der Oberflächenschicht eines Meeres oder Sees, in der der Salinitätsgradient sprunghaft zunimmt (d. h. wo die Salinität mit zunehmender Tiefe rapide abnimmt). 51 PE 379.208 Klimawandel und europäische Fischerei Kernaussage 12 Klimaänderungen können auf verschiedenen Ebenen Auswirkungen auf Fische haben, d. h. auf der zellularen und der individuellen Ebene sowie auf der Ebene der Lebensstadien, der Population, der Arten, der Gemeinschaft und des Ökosystems. Wahrscheinlich führen Temperaturänderungen und andere abiotische Faktoren zu Veränderungen bei den interspezifischen Interaktionen, welche darüber hinaus die Reaktion von Fischen und anderen Taxa auf den Klimawandel beeinflussen. Klimaänderungen verursachen Verlagerungen bei den Industriefischpopulationen. Es gibt mehrere Beispiele dafür, dass temperatur- oder salinitätsbedingte Veränderungen bei den Wechselwirkungen (Nahrungsorganismen, Raubfische, Konkurrenten, Reproduktion) zur Verlagerung der Populationen führen. Mit Hilfe dieser Beispiele lassen sich einige Wechselwirkungen und internen Veränderungen in den Ökosystemen erklären. Es müssen jedoch Untersuchungen zur Populations- und Gemeinschaftsebene durchgeführt werden, da sich die direkten klimatischen Auswirkungen für die einzelnen Tiere nicht unmittelbar in Veränderungen in Verbreitung und Größe der Fischpopulationen niederschlagen. Weitere einschlägige Literaturquellen zu den wichtigsten Arten Pörtner et al. (2002, 2004, 2005) haben verschiedene Aspekte des Sauerstofftransports und des Stoffwechsels untersucht und Pejustemperaturen (Verschlechterung) ermittelt, bei deren Überschreitung die Fähigkeit der Tiere zum aeroben Stoffwechsel vermindert ist. Dies wird anhand der Abnahme des aeroben Bereichs deutlich, der als die proportionale Differenz zwischen dem verbleibenden und dem größtmöglichen Sauerstoffverbrauch definiert wird. Der Unterschied zwischen der unteren und der oberen Pejustemperatur ist wesentlich geringer als bei den kritischen Temperaturen, bei deren Überschreitung die Tiere nur kurze Zeit überleben, d. h. die Temperaturtoleranz gegenüber Veränderungen ist sogar noch geringer als anhand der kritischen Temperaturwerte prognostiziert. Wang und Overgaard (2007) haben nachgewiesen, dass das kardiale Auswurfvolumen( 53) bei höheren Temperaturen vermindert ist und nicht proportional zur Verstärkung des Stoffwechsels ansteigt, die bei erhöhten Temperaturen einsetzt. Daher wird der aerobe Bereich wahrscheinlich durch das Herz begrenzt. Weil die Population offenbar schrumpft, noch bevor die Temperatur das Überleben des Einzeltiers bedroht, wird der Populationsrückgang anscheinend eher durch Verschlechterung der Wachstums- und Fortpflanzungsbedingungen als durch das wärmebedingte Sterben verursacht. Verstärkt werden diese Probleme noch durch die Tatsache, dass die Konzentration von physikalisch gelöstem Sauerstoff im Wasser mit zunehmender Temperatur abnimmt (Weiss 1970). Temperaturänderungen haben Auswirkungen auf die Chemie aquatischer Systeme (Arnell 1998; Harley et al. 2006). Chemische Reaktionen laufen bei hohen Temperaturen schneller ab und die Mobilität von Kontaminanten kann sich infolge von Klimaänderungen erhöhen (Moore et al. 1997). Kernaussage 13 Temperaturänderungen beeinflussen die Chemie aquatischer Systeme (unter anderem auch die Verfügbarkeit von Sauerstoff) sowie die Stoffwechsel- und Sauerstofftransportsysteme bei Fischen, was möglicherweise Auswirkungen auf die Abundanz und die Verbreitung von Fischarten haben kann. (53) Kardiales Auswurfvolumen = Blutstrom aus dem Herzen. PE 379.208 52 Klimawandel und europäische Fischerei 3.3 Weitere damit zusammenhängende Fragen Biologische Merkmale einschlägiger Arten der biotischen Umwelt und wichtiger in der Aquakultur eingesetzter Arten Die wichtigsten Organismen für die biotische Umwelt der bedeutendsten Fischarten sind ihre Beute und natürlichen Feinde. Als Beuteorganismen der planktonfressenden Fischarten spielen vor allem kleine und große Copepoden eine wichtige Rolle. In der Ostsee sind kleine Arten wie Pseudocalanus und Acartia wichtige Nahrungsorganismen, wohingegen in der Nordsee und im Nordostatlantik auch große Ruderfußkrebse wie Calanus finmarchicus und C. helgolandicus von Bedeutung sind. Einschlägige Informationen in Bezug auf die grundlegenden biologischen Merkmale der einzelnen Arten, die sich in der biologischen Umwelt der wichtigsten Industriefischarten finden, sind in Anhang 1-2 dargestellt. Kernaussage 14 Der Klimawandel hat Auswirkungen auf die Beutetiere und die natürlichen Feinde von Industriefischen. Wichtige Organismen in der biotischen Umwelt von Industriefischarten sind ihre Beutetiere und ihre natürlichen Feinde. Die wichtigsten Beuteorganismen für planktonfressende Fischarten sind kleine und große Ruderfußkrebse( 54). In der Ostsee sind kleine Arten wie Pseudocalanus und Acartia wichtige Nahrungsorganismen, wohingegen in der Nordsee und im Nordostatlantik auch große Ruderfußkrebse wie Calanus finmarchicus und C. helgolandicus von Bedeutung sind. Da sie einen wichtigen Stellenwert in der Nahrungsmittelkette einnehmen, sind ihre Reaktionen auf Klimaauswirkungen entscheidend für die Produktivität der Industriefischbestände. Es ist erforderlich, die wahrscheinlichen Reaktionen der betreffenden Organismen auf den Klimawandel zu erforschen. Die wichtigsten Arten, die in der Meeresaquakultur eingesetzt werden Tabelle 5: Auswahl wichtiger Arten, die in der Aquakultur eingesetzt werden, und die jeweiligen Gebiete, die im Rahmen der Studie von Bedeutung sind Wichtigste Arten Gebiete, die im Rahmen der Studie von Bedeutung sind Atlantischer Lachs Nordostatlantik Goldbrasse Nordostatlantik, Mittelmeer Pazifische Felsenauster Nordostatlantik, Mittelmeer, Schwarzes Meer Miesmuschel Nordostatlantik Wolfsbarsch Nordostatlantik, Mittelmeer and Schwarzes Meer Japanische Teppichmuschel Nordostatlantik, Mittelmeer and Schwarzes Meer Mittelmeer-Miesmuschel Mittelmeer and Schwarzes Meer Regenbogenforelle Nordostatlantik (54)Die größte Klasse von Krebstieren. Entweder frei lebend oder als Parasiten. Stellen den größten Teil des Zooplankton, sind jedoch auch bei den Bodenfischarten anzutreffen. 53 PE 379.208 Klimawandel und europäische Fischerei Roter Thun Nordostatlantik, Mittelmeer and Schwarzes Meer Steinbutt Nordostatlantik Teppichmuschel Nordostatlantik, Mittelmeer and Schwarzes Meer Europäische Auster Nordostatlantik Europäischer Aal Nordostatlantik, Mittelmeer and Schwarzes Meer Quelle: Fröse und Pauly 2007 und FAO-Daten. „FAO Fisheries and Aquaculture Departement, Cultured Aquatic Species fact sheets“. Einschlägige Informationen in Bezug auf die grundlegenden biologischen Merkmale der wichtigsten in der marinen Aquakultur eingesetzten Arten sind in Anhang 1-3 dargestellt. Das mögliche Vordringen exotischer Fischarten ins Ökosystem Die biologische Invasion ist zu einem der auffälligsten Merkmale des globalen Wandels geworden. Durch sie können die biologische Vielfalt und die Funktionen natürlicher Ökosysteme verändert werden, und sie kann erheblichen wirtschaftlichen Schaden verursachen. Die Invasion in terrestrische und aquatische Systeme hat gezeigt, dass das erfolgreiche Eindringen exotischer Arten ehemals stabile Systeme aus dem Gleichgewicht bringen und unvorhersagbar machen kann. Allerdings ist nur eine begrenzte Zahl exotischer Arten in der Lage, sich in den Aufnahmeökosystemen anzusiedeln und sich deren Bedingungen anzupassen. Aufgrund der Rolle, die die Veränderlichkeit der Umwelt spielt, wenn es darum geht, die Inversionsergebnisse festzustellen, ist es schwierig, den Ansiedelungserfolg und die Folgen der Invasionen vorherzusagen (Helmuth et al. 2006). Klimaänderungen wirken sich unmittelbar auf systemspezifische Attribute aus und fungieren daher als Filter zur Modulierung des Invasionsrisikos und der entsprechenden Reaktionen. Es wird davon ausgegangen, dass bislang keine neuen Seefischarten in den Nordostatlantik und die angrenzenden Meeresgebiete gelangt sind (Nehring 2003). Dagegen wurden in die Süßwassersysteme während des letzten Jahrhunderts fast 70 nicht heimische Arten eingebracht, in den meisten Fällen absichtlich. Nehring stellt ferner die Hypothese auf, dass, obgleich sich keine neuen Meeresarten in den nördlichen Gewässern angesiedelt haben, bei den südlichen Arten, die in der Vergangenheit selten anzutreffen waren, eine Zunahme zu verzeichnen ist. In die Levantinische See (südöstliches Mittelmeer) wurden durch die Öffnung des Suezkanals mehr als 60 aus dem Indischen/Pazifischen Ozean stammende Fischarten eingetragen, von denen einige bereits einheimische Arten ersetzt haben (Goren und Galil 2005). Auch wenn dieses Gebiet keine direkten Auswirkungen auf die europäische Fischerei hat, so zeigt dies, wie dramatisch eine Artenverschiebung ein Ökosystem verändern kann. Es wurde nachgewiesen, dass die biologische Vielfalt im Mittelmeer infolge anthropogener und klimatischer Einflüsse erhebliche Veränderungen erfahren hat. Mehr als 300 neue Arten (Fische und andere) sind aus dem Roten Meer über den Suezkanal ins Mittelmeer eingewandert und haben sich dort angesiedelt. Diese im Hinblick auf Fische und Phytoplankton beobachteten Veränderungen haben vor allem wegen der Ausbildung toxischer Blüten schwerwiegende wirtschaftliche Folgen für Fischereiindustrie und Tourismus (Bethoux et al. 1999). Invasive Arten sind in den europäischen Küstengewässern häufiger anzutreffen. Allerdings wird die Dynamik dieser Invasionen nur unzureichend registriert und verstanden. Steigende Temperaturen können die Ansiedelung und Verbreitung absichtlich oder zufällig eingeführter PE 379.208 54 Klimawandel und europäische Fischerei Arten erleichtern (Wonham et al. 2000) und könnten es möglich machen, dass ein breiteres Spektrum von Arten eindringt und sich ansiedelt (MCCIP 2006). Kernaussage 15 Invasive Arten treten zunehmend häufiger in europäischen Küstengewässern auf. Die biologische Invasion ist zu einem der auffälligsten Merkmale des globalen Wandels geworden. Durch sie können die biologische Vielfalt und die Funktionen natürlicher Ökosystemen verändert werden, und sie kann erheblichen wirtschaftlichen Schaden verursachen. Allerdings wird die Dynamik dieser Invasionen nur unzureichend registriert und verstanden. Steigende Temperaturen können die Ansiedelung und Verbreitung absichtlich oder zufällig eingeführter Arten erleichtern und könnten es möglich machen, dass ein breiteres Spektrum von Arten eindringt und sich ansiedelt. Auswirkungen des Klimawandels auf die biologische Produktivität und ihre Saisonalität Der Synchronismus zwischen einer hohen Phytoplanktonabundanz und den entsprechenden Fischlarvenstadien ist für das Überleben der Fischbestände von maßgeblicher Bedeutung (Cushing 1990) und ein fehlender Gleichtakt kann schwerwiegende Folgen für den Energietransfer auf höhere trophische Ebenen haben (Stenseth et al. 2002). Im Hinblick auf die marine Phänologie wurde festgestellt, dass viele Planktongruppen ihre jahreszeitlichen Zyklen schneller durchlaufen (Edwards und Richardson 2004). Gemäßigte Meeresumgebungen sind besonders empfindlich gegenüber phänologischen Änderungen, die durch die Klimaerwärmung hervorgerufen werden, da der Rekrutierungserfolg höherer trophischer Ebenen vom Gleichtakt mit der pulsierenden Planktonproduktion abhängt. Dies gilt auch für die Fischlarven und deutet darauf hin, dass sich die Klimaerwärmung möglicherweise durch den mangelnden Gleichtakt bei trophischen Interaktionen nachteilig auf kommerziell genutzte Fischarten auswirkt (Edwards und Richardson 2004). Ferner sei erwähnt, dass nicht bekannt sind, inwieweit sich Fische und Planktongemeinschaften genetisch an die regionale Klimaerwärmung anpassen können und wie rasch eine solche Anpassung erfolgen kann (Hoepffner 2006). Zur Abschätzung der künftigen Auswirkungen der Klimaänderung muss auf Modellversuche zurückgegriffen werden, aber es ist beim gegenwärtigen Kenntnisstand nicht möglich, die klimabedingten Veränderungen in der weltweiten marinen Produktion quantitativ vorherzusagen, da hierbei eine Vielzahl von Wechselwirkungen auftritt. In einer komparativen Studie mit unterschiedlichen Ansätzen für globale Ozeanmodelle gehen Sarmiento et al. (2004) davon aus, dass es bis 2050 zu einer Zunahme der Produktion kommen kann, jedoch um nicht mehr als 10 %. Allerdings stufen sie diese Schätzungen als nicht sehr zuverlässig ein. Im Gegensatz dazu haben Satellitenbeobachtungen und umfangreiche Probenahmen von Plankton gezeigt, dass Phytoplankton und Chlorophyll in den letzten 20 bis 50 Jahren zurückgegangen sind, was mit den Folgen einer verringerten Nährstoffversorgung im Zusammenhang mit der Verstärkung der vertikalen Dichtegradienten im Einklang steht (Brander. 2006). Für 74 % der permanent geschichteten Weltmeere geht die Oberflächenerwärmung mit einer Verringerung der Produktivität einher und ist eng mit den Klimaschwankungen verbunden (Behrenfeld et al. 2006). In den höheren Breiten ist das Gegenteil der Fall. Empirischen Modellen zufolge wird es zu einer Primärproduktionszunahme kommen, da große Gebiete infolge des Temperaturanstiegs eisfrei werden (ACIA 2005). Die höhere Wasserstabilität wird sich trotz des verringerten Nährstoffangebots positiv auf die Produktion auswirken, da das Phytoplankton nicht länger unterhalb der Kompensationstiefe fixiert sein wird (d. h. in einer Tiefe, in der der Respirationsverlust größer ist als der Photosynthesegewinn (Brander 2006). In der arktischen 55 PE 379.208 Klimawandel und europäische Fischerei Zone hat der Klimawandel das Abfließen von Wasser mit geringer Salinität aus der Arktis gefördert, was generell zu einem Anstieg des Frischwasseranteils in den Schelfgewässern führt, der Veränderungen der Abundanz und der jahreszeitlichen Zyklen von Phytoplankton, Zooplankton und Fischpopulationen bewirkt (Greene und Pershing 2007, Smetacek und Nicol 2005). Diese Veränderungen der Produktivität werden die biologische Vielfalt und die Tragfähigkeit der Systeme sowie die Bewirtschaftung der biologischen Ressourcen der Meere beeinflussen (Hoepffner et al. 2006). Kernaussage 16 Klimatisch bedingte Störungen der Nahrungsmittelkette beeinflussen das Überleben und die Produktivität von Industriefischbeständen. Für das Überleben und die Produktivität von Fischbeständen ist es ganz wichtig, dass für die Menge an Fischlarven auch ausreichend Zooplankton in der richtigen Größe vorhanden ist. Aufgrund der klimatischen Veränderungen durchlaufen viele Planktongruppen ihre jahreszeitlichen Zyklen schneller, und der fehlende Gleichtakt hat schwerwiegende Folgen für das Überleben und die Produktivität der Industriefischbestände. Beim gegenwärtigen Kenntnisstand ist es nicht möglich, die durch den Klimawandel zu erwartenden Veränderungen in der weltweiten marinen Produktion quantitativ vorherzusagen, da hierbei eine Vielzahl von Wechselwirkungen auftreten. Zweifellos jedoch werden Veränderungen der Produktivität und der Saisonmuster die Bewirtschaftung der biologischen Ressourcen der Meere beeinflussen. Wie (nicht sehr zuverlässige) Schätzungen besagen, dürfte es bis 2050 zu einer Zunahme der weltweiten marinen Produktion kommen, jedoch um nicht mehr als 10 %. Im Gegensatz dazu haben Satellitenbeobachtungen und umfangreiche Probenahmen von Plankton gezeigt, dass Phytoplankton und Chlorophyll in den letzten 20 bis 50 Jahren zurückgegangen sind. Für 74 % der permanent geschichteten Weltmeere ist die Oberflächenerwärmung mit einer Verringerung der Produktivität verbunden. In den nördlichen Breiten wird mit einer Produktionszunahme gerechnet, da große Gebiete eisfrei werden. Einfluss der Umweltbedingungen und der Fischerei auf den Rekrutierungserfolg Wenngleich während der letzten Jahrzehnte bei den kommerziellen Fischbeständen im Atlantik zahlreiche Veränderungen beobachtet wurden, ist es äußerst schwierig, die aus Veränderungen in der Populationsdichte und der Rekrutierung resultierenden Folgen sowie die regionalen Klimaeffekte von direkten anthropogenen Einflüssen, wie etwa der Fischerei, zu unterscheiden. Die Fischerei ist als nachteiliger anthropogener Einfluss von erheblicher Bedeutung anzusehen (Pauly et al. 2005, IPCC 2007). Strukturelle Veränderungen innerhalb des Ökosystems können von oben nach unten durch einen Zusammenbruch der Raubfischpopulation durch Überfischung (Worm und Myers 2003, Frank et al. 2005) oder durch den Klimawandel (Polovina 2005) ausgelöst werden und sich über trophische Kopplung bis in die unteren Schichten des Nahrungsnetzes auswirken. Es sei daran erinnert, dass der Fischereiaufwand darauf gerichtet ist, die älteren und größeren Tiere der Population zu fangen, und dass kürzlich nachgewiesen wurde, dass dieser selektive Fischereidruck auf ältere und größere Fische zu einer raschen Verringerung der Körpergröße und der Fruchtbarkeit (Olsen et al. 2004) sowie zu einem Verlust der genetischen Variabilität führt (Hauser et al. 2002). Da die Fischbrut älterer Fische ein höheres Wachstums- und Überlebenspotenzial aufweist (Berkeley et al. 2004), gilt es mit Blick auf die Erhaltung der Produktivität einer Population, ihrer Fähigkeit zur Anpassung an veränderte Umweltbedingungen und ihrer Fruchtbarkeit, diese älteren und größeren Fische langlebiger Fischarten zu schützen, statt nur die Gesamtfangzahlen für die Population festzulegen (Berkeley et al. 2004, Birkeland und Dayton, 2005). Die Rekrutierung des Kabeljau im Nordatlantik steht bei Beständen, die an den Breitengradgrenzen für die Verbreitung der betreffenden Art angesiedelt sind, offenbar im Zusammenhang mit den Meerestemperaturen (O’Brien et al. 2000). Auf der Nordhalbkugel sind PE 379.208 56 Klimawandel und europäische Fischerei steigende Temperaturen günstig für Bestände an den höchsten Breitengraden, während sie sich auf Bestände an den südlichen Grenzen nachteilig auswirken. Die Kabeljaubestände in der Nordsee sind nahe der südlichen Grenze ihres Verbreitungsgebiets angesiedelt, und starke Jahresklassen wurden von jeher damit in Verbindung gebracht, dass die Temperaturen während des ersten Halbjahres unter dem Durchschnitt lagen (O’Brien et al. 2000). Ein weiterer Faktor neben den gestiegenen Wassertemperaturen in der Nordsee ist die Tatsache, dass die Struktur der Kabeljaubestände von unreifen, weniger als fünf Jahre alten Jungfischen dominiert wird. Planque und Frédou (1999) haben den Zusammenhang zwischen Temperatur und Rekrutierung für die Kabeljaubestände im Nordatlantik analysiert und festgestellt, dass das Verhältnis zwischen Wasseroberflächentemperatur und Rekrutierung für Kaltwasserbestände mit geschlechtsreifen Tieren, die in Gebieten mit Bodentemperaturen von <= 6 °C leben, im Allgemeinen positiv ist. Ein negatives Verhältnis ergab sich bei Kabeljaubeständen in Warmwassergebieten mit Bodentemperaturen von >=9 °C. Bei Beständen, die in Gebieten leben, in denen die Bodentemperaturen im mittleren Bereich (7-8 °C) liegen, gab es normalerweise kaum einen bzw. gar keinen Zusammenhang zwischen Wasseroberflächentemperatur und Rekrutierung (Abbildung 16). Abbildung 16: Einfluss der Temperatur auf die Fischrekrutierung. Quelle: Geändert ausgehend von Planque and Frédou (1999), Drinkwater 2005. Anmerkungen: Das Verhältnis zwischen Log2 der Rekrutierungsanomalie und der Anomalie der Wasseroberflächentemperatur in °C für verschiedene Kabeljaubestände. Die Achsenbeschriftung in der unteren Mitte der Darstellung gilt für alle Diagramme. Die Zahl unter den Diagrammen entspricht der mittleren jährlichen Bodentemperatur für die betreffenden Bestände. Es sei darauf hingewiesen, dass die Bestände nach der Bodentemperatur – von links nach rechts steigend – dargestellt sind. Für Kaltwasserbestände ist das Verhältnis zwischen Wasseroberflächentemperatur und Rekrutierung im Allgemeinen positiv, während es für Warmwasserbestände negativ ist. Im mittleren Temperaturbereich besteht kein Zusammenhang. 57 PE 379.208 Klimawandel und europäische Fischerei Legende: Warmer Temperatures increase Recruitment Warmer Temperatures decrease Recrutiment Newfoundland W. Greenland Barents Sea Iceland Faroes Georges Bank North Sea Irish Sea Celtic Sea Höhere Temperaturen – verstärkte Rekrutierung Höhere Temperaturen – verminderte Rekrutierung Neufundland Westgrönland Barentssee Island Färoer Georges Bank Nordsee Irische See Keltische See Log2 Recruitment Anomaly SST Anomaly Log2 Rekrutierungsanomalie Anomalie der Wasseroberflächentemperatur Hsieh et al. (2006) haben den Einfluss der Befischung und der Umweltvariabilität auf die Larven befischter und nicht befischter Arten im kalifornischen Strömungssystem in einer Zeitreihe analysiert (1951-2002). Sie konnten nachweisen, dass die befischten Arten eine höhere zeitliche Variabilität bei der Abundanz aufwiesen als die nicht befischten Arten. Es wird angenommen, dass die höhere Variabilität der befischten Arten durch fischereibedingte Einschnitte in die Altersstruktur verursacht wird. Man geht davon aus, dass sich durch die Verringerung des Durchschnittsalters und der Durchschnittslänge einzelner Fische innerhalb einer Population die Rekrutierungsvariabilität erhöht, da die Fähigkeit, kurzfristige ungünstige Umweltbedingungen auszuhalten, abnimmt. Viele Fischarten nutzen Strategien der Risikostreuung („bet-hedging strategies“), um die Überlebensrate unter unwirtlichen und unbeständigen Umweltbedingungen zu erhöhen. Solche Hedging-Strategien sind mit breitgefächerten Altersstrukturen verbunden und schließen altersbedingte Unterschiede hinsichtlich der Laichplätze und –zeiten sowie eine höhere Menge und Qualität der von den älteren (erfahrenen) bzw. größeren Fische produzierten Eier ein (Marteinsdottir und Steinarsson 1998, Berkeley et al. 2004). Diese Ergebnisse haben unmittelbare Auswirkungen auf das Fischereimanagement und zeigen, dass Fischerei nicht nur einen Rückgang der Abundanz bewirken, sondern auch zu einer größeren Variabilität bei den befischten Beständen führen und auf diese Weise die Widerstandsfähigkeit schwächen und die Gefahr eines Zusammenbruchs der betreffenden Fischerei aufgrund von Umwelteinflüssen erhöhen kann (Hsieh et al. 2006). Kernaussage 17 Während der letzten Jahrzehnte wurden bei den kommerziellen Fischbeständen im Atlantik zahlreiche Veränderungen beobachtet. Dabei ist es äußerst schwierig, die aus Veränderungen in der Populationsdichte und der Rekrutierung resultierenden Folgen sowie die regionalen Klimaeffekte von direkten anthropogenen Einflüssen, wie etwa der Fischerei, zu unterscheiden. Die derzeitige Fischereipraxis schwächt die Widerstandsfähigkeit der Fischbestände gegenüber den Auswirkungen des Klimawandels. Die derzeitigen Fangmethoden richten sich auf ältere und größere Fische und bewirken eine Veränderung der Größen- und Altersstruktur der Fischpopulationen. Die Widerstandsfähigkeit gegenüber Umweltauswirkungen wird geschwächt, die genetische Variabilität geht verloren und die Fähigkeit zur Anpassung an Umweltveränderungen wird schwächer. Folglich kann die Fischerei dazu führen, dass die Umweltauswirkungen für die Bestände zu einer immer größeren Gefahr werden. Eine ökologisch nachhaltige Fischerei sollte den Schutz der größeren und älteren Fische zum Ziel haben. Auswirkungen der künftigen Fischereitätigkeit Eines der wichtigsten Ergebnisse des Weltgipfels 2002 in Johannesburg war die Verpflichtung, das Konzept des höchstmöglichen Dauerertrags für die Fischerei der Europäischen Union bis PE 379.208 58 Klimawandel und europäische Fischerei 2015 umzusetzen. Ein Weg in Richtung auf eine bessere Bewertung der Fischbestände unter Berücksichtigung der Umweltaspekte ist der Ökosystemansatz im Fischereimanagement (EAFM; ESA 1998, Pope und Symes 2000), der anlässlich des Weltgipfels 1992 in Rio propagiert wurde. Die Europäische Union engagiert sich nachdrücklich für den Schutz der Ozeane und Meere. Die Kommission hat 2005 eine thematische Strategie für den Schutz und die Erhaltung der Meeresumwelt vorgelegt, in der sie unter anderem eine Richtlinie über eine Meeresstrategie vorschlägt. Die Richtlinie soll sicherstellen, dass bis 2021 alle Meeresgewässer der EU ökologisch gesund sind, sodass die Europäer Ozeane und Meere Nutzen können, die sicher, sauber und reich an biologischer Vielfalt sind. Die Ernährungs- und Landwirtschaftsorganisation der Vereinten Nationen (FAO) nennt den künftigen anthropogen bewirkten Klimawandel als ein Beispiel für Unsicherheiten, die einen Vorsorgeansatz für das Fischereimanagement rechtfertigen (FAO, WBGU-Bericht 2000). Im Lagebericht zur Fischerei (FAO, WGBU-Bericht 2002) wird festgestellt, dass die globale Erwärmung deutliche – positive oder negative – Auswirkungen auf die meisten kommerziellen Fischbestände haben könnte. Die Autoren der Studie kommen zu dem Schluss, dass durch Überfischung drastisch reduzierte Bestände anfälliger für Klimaänderungen sind als nachhaltig genutzte Bestände (FAO, 2004, Hsieh et al. 2006). Darüber hinaus hängt die Reaktion der Fischbestände auf Umwelteinflüsse von ihrer Populationsgröße ab. Gesunde Bestände mit großer Produktion von Fischlarven können sich besser an Populationsverschiebungen und Veränderungen der Ökosystemstrukturen anpassen (Hsieh 2006). Dementsprechend reagieren durch Überfischung stark verkleinerte Bestände gegenüber Umwelteinflüssen besonders empfindlich, weil der für die Reproduktion erforderliche Mindestbestand leichter unterschritten wird (MA 2005 b, WGBU). Ein mögliches Szenario für die Zukunft der Nordsee beinhaltet einen Anstieg der Temperatur, eine starke NAO und einen verstärkten Zufluss von Atlantikwasser (Stenevik und Sundby 2007). Folgen davon wären eine geringe Rekrutierung des Kabeljau, eine Nordwärtsverlagerung der jetzigen Fischarten (Kabeljau, Hering und Sprotte) und das Vordringen südlicher Arten (Sardine und Sardelle) (Stenevik and Sundby 2007). Da der Hering und wahrscheinlich auch andere kleine pelagische Arten sehr stark auf sich verändernde hydrographische Bedingungen reagieren, sollte die künftige Bestandsbewirtschaftung kontinuierlich, gleichzeitig aber auch flexibel sein (Miller und Munro 2004). Insbesondere die ausgesprochenen Wanderarten, die ihre Routen bei Umweltveränderungen ändern, werden besondere Anforderungen an die Bewirtschaftung stellen, da wahrscheinlich ein Übergang von der alleinigen zur gemeinsamen Bewirtschaftung der Bestände erfolgen wird (Miller und Munro 2004, Miller 2007). Kernaussage 18 Unter dem Gesichtspunkt der Nachhaltigkeit befischte Bestände können besser auf Klimaauswirkungen reagieren. Die Ungewissheit über den künftigen anthropogenen Klimawandel rechtfertigt einen Vorsorgeansatz für die Bestandsbewirtschaftung. Die globale Erwärmung kann sich erheblich – in positiver oder negativer Art und Weise – auf die meisten der kommerziellen Fischbestände auswirken. Dabei sind durch Überfischung drastisch reduzierte Bestände anfälliger gegenüber klimatischen Veränderungen als nachhaltig befischte Bestände. Maßgeblich für die Reaktion der Fischbestände auf Umwelteinflüsse ist die Populationsgröße. Gesunde Bestände können sich besser an Populationsverschiebungen und strukturelle Veränderungen im Ökosystem anpassen. Eine flexible und anpassungsfähige Bestandsbewirtschaftung ist notwendig. Ein mögliches Szenario für die Zukunft der Nordsee beinhaltet einen Anstieg der Temperatur, eine starke NAO und einen verstärkten Zufluss von Atlantikwasser. Folgen davon wären eine geringe Rekrutierung des Kabeljau, eine Nordwärtsverlagerung der jetzigen Fischarten (Kabeljau, Hering und Sprotte) und das Vordringen südlicher Arten (Sardine und Sardelle). Eine wichtige Frage ist die, wie in Anbetracht der 59 PE 379.208 Klimawandel und europäische Fischerei klimatischen Auswirkungen auf die Meeresumwelt die industrielle Fischerei künftig zu gestalten ist. Da einige der wichtigsten Industriefischarten (wie Hering und wahrscheinlich andere kleine pelagische Arten) sehr stark auf sich verändernde hydrographische Bedingungen reagieren, sollte die künftige Bestandsbewirtschaftung kontinuierlich sein, gleichzeitig aber auch flexibel und anpassungsfähig, um den jeweiligen Reaktionen der Bestände auf die Umweltbedingungen Rechnung zu tragen. Insbesondere die ausgesprochenen Wanderarten, die ihre Routen bei Umweltveränderungen ändern, werden besondere Anforderungen an die Bewirtschaftung stellen. Um das Fischereimanagement rechtzeitig auf die Situation einzustellen, müssen durch entsprechende Forschungsarbeiten die Kenntnisse über die Auswirkungen des Klimawandels auf die Fischerei der EU und damit verbundene Prognosen verbessert werden. Kausalketten zwischen Klimaeinflüssen – Hydrographie – trophischen Ebenen und Fischproduktivität Ostsee Die physikalischen Bedingungen in der Ostsee reagieren auf Klimaänderungen durch direkte Wechselbeziehung zwischen Luft und Meer, das Ausmaß des Frischwasserabflusses und Interaktionen mit dem Ozean an den offenen Rändern. Diese Bedingungen sind eng mit atmosphärischen Einflüssen verbunden. Ein hoher NAO-Index und starke Westwinde seit Ende der 1980er Jahre haben zu einem Anstieg der Wassertemperaturen und zu einer Verringerung der Salinität geführt (Hänninen et al. 2000). Diese Änderungen der Temperatur und der Salinität bewirkten eine Verlagerung der Dominanz bei den wichtigsten Copepodenarten von Pseudocalanus sp. auf Temora longicornis und Acartia spp. (Möllmann et al. 2003). Ähnlich wie bei der Copepodengemeinschaft verschob sich in der Fischgemeinschaft der mittleren Ostsee die Dominanz vom Dorsch (Gadus morhua), der während der 1980er Jahre zahlenmäßig am stärksten vertreten war, auf die Sprotte (Sprattus sprattus), die ihrerseits in den 1990er Jahren die Fischgemeinschaft dominierte (Köster et al. 2003). Darüber hinaus wies der kommerziell bedeutsame Hering (Clupea harengus) ein geringeres Wachstum auf. Möllmann et al. 2005 konnten nachweisen, dass der Zustand der Heringsbestände auf die Veränderung des Nahrungsangebots für diese Fischart in Verbindung mit der verstärkten Konkurrenz durch die Sprotte zurückzuführen ist, während der Zustand der Sprottenbestände offenbar vorrangig von der intraspezifischen( 55) Konkurrenz abhängt (Abbildung 17). (55) Innerhalb ein und derselben Art, z. B. Konkurrenz. PE 379.208 60 Klimawandel und europäische Fischerei Abbildung 17: Kausalzusammenhang zwischen Klima und Fischwachstum (Ostsee) Quelle: Möllmann 2005. Anmerkungen: Schematische Darstellung des Zusammenhangs zwischen Klima, Copepoden und dem Wachstum pelagischer Fische: linke Seite: Zusammenhang zwischen den Variablen; rechte Seite: die sich ergebenden idealisierten zeitlichen Trends; die grauen Linien und Pfeile stellen temperaturbedingte Prozesse dar, während die schwarzen Linien und Pfeile salinitätsbedingte Prozesse darstellen. Legende: 1970s/1980s 1990s BSI Temperature Salinity Pseudocalanus Temora Acartia Sprat stock Herring condition Sprat condition 1970er/1980er Jahre 1990er Jahre BSI (Baltic Sea Index) Temperatur Salinität Pseudocalanus Temora Acartia Sprottenbestand Zustand des Heringsbestands Zustand des Sprottenbestands Mittelmeer Molinero et al. (2007) haben die ökologischen Auswirkungen des Klimawandels auf pelagische Copepoden, Chaetognathen und Quallen im Mittelmeer untersucht. Diese Organismen spielen im Materie- und Energiefluss von Meeresökosystemen eine Schlüsselrolle. Während die Copepoden die meisten Nahrungsnetze und die biologische Kohlenstoffpumpe unterstützen, durch die Kohlenstoff in die Tiefsee befördert wird, können Chaetognathen und Quallen 61 PE 379.208 Klimawandel und europäische Fischerei Auswirkungen auf die Stärke der Top-down-Steuerung von Planktongemeinschaften haben. Molinero et al. (2007) haben nachgewiesen, dass extreme Ereignisse in der langfristigen Variabilität dieser Funktionsgruppen im nordwestlichen Mittelmeer eng mit den Veränderungen der klimatischen Einflüsse im nordatlantischen Sektor zusammenhängen. Überregionale klimatische Einflüsse haben durch Änderungen in den biologischen Interaktionen sowie bei Konkurrenz und Prädation die Dynamik des pelagischen Nahrungsnetzes verändert. In warmen Wintern sind die Wärme- und Wasserverluste gering, und es wird eine Verringerung des Transports mit der Wasserströmung vom Tyrrhenischen Meer zum Ligurischen Meer im Kanal zwischen Korsika und Italien erwartet. Diese hydrologischen Systeme sind mit hohen klimatischen Werten im Nordatlantik verbunden und beeinflussen die Temperatur und die Schichtung des Wassers, die sich ihrerseits auf die Zusammensetzung des Phytoplanktons auswirken (Dominanz der Diatomeen – keine Dominanz der Diatomeen). Anschließend können die hohen Temperaturen und die Änderungen beim Nahrungsangebot (größere Verfügbarkeit anderer Nahrung als Diatomeen) die Eiproduktion und Rekrutierung von Copepoden beeinflussen und möglicherweise auch die Nahrungsverfügbarkeit für Chaetognathen verringern. Darüber hinaus begünstigen die höhere Wassertemperatur, die geringere Windbelastung und weniger Niederschläge und die daraus resultierende stärkere Schichtung das Überleben und die verstärkte Fortpflanzung von Quallen und können somit zu Quallenplagen führen. Letztere können die Chaetognathen beeinträchtigen und eine höhere Sterblichkeit bei den Sommer/Herbst-Copepoden bewirken, was wiederum Auswirkungen auf die pelagischen Fischarten wie Sardellen und Sardinen hat, die sich von Copepoden ernähren (Abbildung 18). PE 379.208 62 Klimawandel und europäische Fischerei Abbildung 18: (Mittelmeer). Kausalzusammenhang zwischen Klima und Fischwachstum . Quelle: Literatur: Aus Molinero et al. (2007): „Mechanisms through which the NAC (positive phase) affect Ligurian Sea hydroclimate and zooplankton long-term changes“. 1. Hurrell, 1995; 2. Trigo et al. 2000; 3. Rixen et al. 2005; 4. Fernandez et al. 2003; 5. Molinero et al. 2005b; 6. Astraldi 1995; 7. Vignudelli et al. 1999; 8. Tsimplis und Josey, 2001; 9.Molinero et al. 2005a; 10. Bethoux et al. 2002; 11. Goffart et al. 2002; 12. Gomez und Gorsky, 2003; 13. Ianora et al. 2003; 14. Carotenuto et al. (Presseveröffentlichung); 15. Halsband-Lenk et al. 2001; 16. Goy et al. 1989; 17. Buecher 1999. 18. Menard et al. 1994, 1997. 63 PE 379.208 Klimawandel und europäische Fischerei Legende: Atmospheric processes associated to high NAC high atmospheric pressure high air temperature low precipitation low wind stress low moisture transport Atmospherische Prozesse im Zusammenhang mit einer starken Nordatlantikströmung hoher Luftdruck hohe Lufttemperatur geringe Niederschläge geringe Windbelastung geringer Feuchtigkeitstransport Air-sea interactions low heat loss sea level changes Interaktionen zwischen Luft und Meer geringer Wärmeverlust Veränderungen des Meeresspiegels Hydrographic Changes lower water mass transport through the Corsica Channel, change in the strength of the Northern current Hydrographische Veränderungen Transport geringerer Wassermassen durch den Kanal zwischen Korsika und Italien, geänderte Stärke der Nordströmung high water temperature, ….. hohe Wassertemperatur, verringerte Durchmischung der Wassersäule und Ausdehnung der Schichtung Interspezifische Modifizierung des Phytoplanktons (Dominanz der Diatomeen/keine Dominanz der Diatomeen) Interspecific modification of ….. Ecological changes associated to high NAC forcing higher abundance of gelatinous zooplankton (salps, doliolids, jellies), higher frequence of gelatinous outbreaks Bottom up control modifications on copepods Top down control chaetognaths abundance of copepods pelagic fish (anchovy, sardine) Ökologische Veränderungen im Zusammenhang mit einem starken Einfluss der Nordatlantikströmung höhere Abundanz von gelatinösem Zooplankton (Salpen, Dolioliden, Quallen), größere Häufigkeit von durch gelatinöses Zooplankton verursachten Plagen In der Tiefe ausgelöste Prozesse Änderungen bei der Nahrung der Copepoden und daraus resultierende Änderungen bei der Eiproduktion und der Rekrutierung von Copepoden Von oben nach unten gerichtete Prozesse Chaetognathen Abundanz von Copepoden pelagische Fischarten (Sardelle, Sardine) Nordsee Beaugrand (2004) untersuchte die Einflüsse, die 1982 und 1988 einen Umbruch im Ökosystem der Nordsee hervorriefen. Die Änderungen im Ökosystem reichten vom Phytoplankton bis zu den Fischen. Der Anstieg der Meeresoberflächentemperatur und die Änderungen bei der Windstärke und –richtung lösten die Verschiebung einer biogeographischen ozeanischen Grenze entlang der europäischen Kontinentalküste aus. Die Zusammensetzung und Vielfalt der calanoiden Copepodenarten änderte sich. So war in der Nordsee nach 1986 bei den Warmwasserarten eine Zunahme zu verzeichnen, während sich der Anteil der Kaltwasserarten verringerte. Während einer Phase, die von starken Westwinden und einer verringerten Salinität gekennzeichnet war, wurde eine erhöhte Rekrutierung von Plattfischen beobachtet, während die Rekrutierung des Kabeljau durch einen Anstieg der Meeresoberflächentemperatur und ein verstärktes Auftreten von calanoiden Copepodenarten der gemäßigten Zonen (Abbildung 19), beeinträchtigt wurde. Hier zeigte sich, dass ein und demselber klimatische Stressfaktor für eine Spezies vorteilhaft ist, während er bei der anderen Spezies zu einem Rückgang der Abundanz führt. PE 379.208 64 Klimawandel und europäische Fischerei Abbildung 19: (Nordsee). Kausalzusammenhang zwischen Klima und Fischwachstum Quelle: Beaugrand 2004. Anmerkungen: Langfristige Veränderungen bei der Abundanz von Fischen im Zusammenhang mit den Jahresschwankungen bei der Zusammensetzung der calanoiden Copepoden und den hydrometeorologischen Einflüssen. (a) Langfristige Veränderungen bei der Fischrekrutierung (Hering, Kabeljau, Schellfisch, Scholle, Seezunge). Die wichtigste Komponente auf der linken Seite (37,18 % des gesamten Artenspektrums) besteht vor allem aus Plattfischen, d. h. Schollen und Seezungen. Die zweitwichtigste Komponente auf der rechten Seite (31,36 % des gesamten Artenspektrums) besteht hauptsächlich aus Dorschartigen (Gadiformes), d. h. Schellfisch und Kabeljau. (b) Langfristige Veränderungen bei der Zusammensetzung der calanoiden Copepodenarten. Es wurde keine Übereinstimmung zwischen den Veränderungen bei den Calanoiden und den Plattfischen festgestellt. (c) Veränderungen bei den hydrologischen Variablen (Salinität auf der linken Seite und Meeresoberflächentemperatur auf der rechten Seite). (d) Veränderungen bei den meteorologischen Einflüssen (Stärke des Westwinds über der Nordsee auf der linken Seite und Temperaturanomalien auf der Nordhalbkugel auf der rechten Seite). Legende: First principal component (37,18% of the total variability) (a) Fish (flatfish recruitment) (b) Calanoid copepods Wichtigste Komponente (37,18 % des gesamten Artenspektrums) (a) Fische (Rekrutierung der Plattfische) (b) Calanoide Copepoden Second principal component (31,36%) (a) Fish (gadaod recruitment) Zweitwichtigste Komponente (31,36 %) (a) Fische (Rekrutierung der Dorschartigen (Gadiformes)) (schwarz: Arten der gemäßigten Zonen; grau: subarktische Arten) (in black: temperate species; in grey: subarctic species) 65 PE 379.208 Klimawandel und europäische Fischerei No match for any of the calanoid copepod assemblages Mean number of species per assemblage Bei keiner der Gruppen von calanoiden Copepoden ist eine Übereinstimmung festzustellen. Mittlere Anzahl von Arten pro Gruppe Salinity (central North Sea) (c) Hydrological variables SST (central North Sea) Salinität (mittlere Nordsee) (c) Hydrologische Variablen Meeresobeflächentemperatur (mittlere Nordsee) Westerly wind intensity (d) Climatological variables Years (1958-1999) Stärke der Westwinde (d) Klimatologische Variablen Jahre (1958-1999) Im Ergebnis ihrer Untersuchung der im Nordostatlantik gesammelten Daten des CPRPlanktonrekorders stellten Kirby et al. (2006) eine Zunahme der Abundanz der Großen Schlangennadel (Entelurus aequoreus) fest, die auf den Einfluss der wärmeren Meerestemperaturen auf die Fortpflanzung und das Überleben dieser Art zurückgeführt wurde. Zwischen 1958 und 2002 kamen Jungfische dieser Art nur gelegentlich in den Proben des CPRPlanktonrekorders vor. Seit 2002 sind sie regelmäßig in den Proben vertreten. Dieser signifikante Anstieg bei Larven und Jungfischen der Gattung Große Schlangennadel wurde möglicherweise durch eine Zunahme der durchschnittlichen Wassertemperatur bewirkt, die im Zeitraum 2002-2005 um 0,5 °C höher war als im Zeitraum 1958-1972. Ein Grund könnte darin bestehen, dass es der Temperaturanstieg den männlichen Tieren ermöglicht, sich häufiger fortzupflanzen, da die Entwicklungszeit kürzer ist. Kernaussage 19 Studien in der Nordsee, der Ostsee und im Mittelmeer zeigen, dass Kausalzusammenhänge zwischen Klima – Hydrographie – niedrigeren und höheren trophischen Ebenen – Rekrutierung der Fische bestehen, deuten darauf hin, dass die Interaktionen von großer Bedeutung sind, und unterstreichen die Tatsache, dass die Klimaänderung multifaktoriell auf verschiedenen Ebenen zum Tagen kommt. Gemeinsam ist den beobachteten Phänomenen die Tatsache, dass die Klimaänderung zu Verschiebungen in den Meeresökosystemen führt (z. B. Temperatur- und Salinitätsänderungen) die sich auf die Produktivität der im Plankton vertretenen Beutearten der wichtigsten Fischarten auswirken und somit Änderungen bei der Produktivität der entsprechenden Fischbestände zur Folge haben. Änderungen an der Basis des Nahrungsnetzes Einige der überzeugendsten Beweise für die biologische Reaktion auf regionale Klimaschwankungen kommen von der Basis des marinen Nahrungsnetzes, insbesondere von den Phytoplankton- und den Zooplanktongemeinschaften. Die Diatomeenblüten im Frühjahr (d. h. zu Beginn des jahreszeitlichen Zyklus der pelagischen Organismen) finden nach wie vor zu relativ festen Zeiten statt, da sie eher vom Tageslicht bzw. von der Lichtintensität als von temperaturvermittelten physiologischen Reaktionen abhängen. Organismen, die im Hinblick auf die Stimulierung physiologischer Entwicklungen und die Freisetzung von Larven von der Temperatur abhängen, durchlaufen ihre jahreszeitlichen Zyklus temperaturbedingt schneller (Edwards und Richardson 2004). Die Reaktion auf Klimasignale kann bei verschiedenen Funktionsgruppen und trophischen Ebenen unterschiedlich sein. Dies hat dazu geführt, dass sich die aufeinanderfolgenden trophischen Ebenen nicht im Gleichtakt befinden und es zu Verschiebungen hinsichtlich der Gleichzeitigkeit der Abläufe zwischen PE 379.208 66 Klimawandel und europäische Fischerei Primär-, Sekundär-( 56) und Tertiärproduktion ( 57) in der Nordsee gekommen ist (Edwards und Richardson 2004). Änderungen bei der Primär- und Sekundärproduktion haben über das Nahrungsnetz Auswirkungen auf die Fischereiproduktion. Hjort hat bereits im Jahr 1914 erkannt, dass die interannuellen Schwankungen bei der Rekrutierung des Kabeljau damit zusammenhängen, inwieweit die Produktion der Nahrung für die Larven zur entsprechenden Zeit erfolgt. Cushing (1990) (Abbildung 20) schlug die „Match-Mismatch-Hypothese“ in Bezug auf die klimatisch bedingte Koppelung/Entkoppelung dieser phänologischen Zusammenhänge vor. Wichtig sind nicht nur das zeitliche Zusammentreffen und die Menge, wahrscheinlich ist auch die jahreszeitliche und interannuelle Nahrungsqualität eine wichtige mechanistische Verbindung zwischen den Klimaschwankungen und den beobachteten Veränderungen beim Rekrutierungserfolg der Fische (Beaugrand et al. 2003, Malzahn et al. 2007). Abbildung 20: Einfluss des Klimas auf die Nahrung für die Fischlarven.. Quelle: Abgewandelte Abbildung von Leggett und Deblois 1994. Schematische Darstellung der von Cushing vorgeschlagenen Hypothese des Gleichtakts/fehlenden Gleichtakts. Legende: Frequency Eggs Larvae Zooplankton Match Mismatch Time Frequenz Eier Larven Zooplankton Gleichtakt Fehlender Gleichtakt Zeit Der Atlantische Kabeljau wird seit Ende der 1960er Jahre in der Nordsee überfischt, und es besteht große Sorge mit Blick auf den Rückgang der Biomasse und der Rekrutierung (Cook et (56) Produktionsrate pflanzenfressender Tiere durch Umwandlung der pflanzlichen Nahrung in tierisches Gewebe. (57) Produktionsrate fleischfressender Tiere durch Umwandlung der tierischen Nahrung in eigenes Gewebe. 67 PE 379.208 Klimawandel und europäische Fischerei al. 1997). Abgesehen von den Auswirkungen der Überfischung haben Schwankungen beim Plankton durch in der Tiefe ausgelöste Prozesse zu langfristigen Veränderungen bei der Rekrutierung des Kabeljau geführt, da der effiziente Transfer der pelagischen Produktion auf höhere trophische Ebenen wie kommerziell bedeutsame Fischarten vom zeitlichen Gleichtakt der Phasen abhängt, in denen die Produktion jeweils am höchsten ist. Es hat sich erwiesen, dass das Überleben der Kabeljaularven im Wesentlichen von drei biologischen Parametern abhängt: der mittleren Größe, dem jahreszeitlichen Timing und der Abundanz. Nach Ansicht von Beaugrand et al. (2003) ist von einem Mechanismus auszugehen, der eine „Match-MismatchSituation“ einschließt, bei der Temperaturschwankungen das Überleben der Kabeljaularven beeinflussen. Infolge der seit Mitte der 1980er Jahre steigenden Temperaturen in der Nordsee haben sich die Überlebenschancen von Kabeljaularven verringert, da es zu einer Veränderung des Planktonsystems gekommen ist, die zu einem Ungleichgewicht zwischen der Größe der Beute und der Größe des Kabeljau geführt hat. Der Kabeljauboom (ein sprunghafter Anstieg der Abundanz einer Reihe von Kabeljauartigen Ende der 1960er bis Mitte der 1980er Jahre) war eine Folge der für Kabeljaularven außerordentlich günstigen Planktonsituation (Beaugrand et al. 2003). Edwards und Richardson (2004) gehen davon aus, dass die marine Trophodynamik sich bereits radikal verändert hat und sich bei einer fortgesetzten Erwärmung weiterhin verändern wird. Die herkömmlichen Zielarten der Nordseefischerei, wie zum Beispiel Kabeljau, werden wahrscheinlich weiterhin zurückgehen und möglicherweise durch Arten wie Rote Meerbarbe, Sardine und Sardelle ersetzt werden (Hoepffner 2006). Ein negativer Zusammenhang zwischen der Rekrutierung und dem NAO-Winterindex, der während der Ei- und Larvenstadien des Nordsee-Sandaals die Meerestemperatur beeinflusst, wurde von Arnott und Ruxton (2002) aufgezeigt. Überdurchschnittlich warme Meerestemperaturen gingen mit einer unterdurchschnittlichen Rekrutierung einher, was sich am stärksten nahe der südlichen Grenze des Verbreitungsgebiets dieser Art am stärksten bemerkbar machte. Es gab einen positiven Zusammenhang zwischen der Rekrutierung und der Abundanz von calanoiden Copepoden in bestimmten Lebensstadien etwa zu der Zeit, in der die Larven schlüpfen, der darauf hindeutet, dass die Verfügbarkeit dieser Beuteart für das Überleben junger Sandaale von Bedeutung ist (Arnott und Ruxton 2002). In den Gewässern um das Vereinigte Königreich verringert sich die Rekrutierung von Scholle und Limande bei steigender Meerestemperatur (Henderson 1998, Fox et al. 2000). Auch Veränderungen in der trophischen Struktur des Ökosystems im Hinblick auf die Bedingungen für die Fütterung der Larven (in der Tiefe ausgelöste Reaktionen) oder die Gefährdung durch Prädatoren (Gefährdung von oben) können die Rekrutierung beeinträchtigen. Kirby et al. (2007) haben Änderungen in der Planktongemeinschaft und deren Saisonbedingtheit aufgezeigt. Die Zunahme des Meroplanktons( 58) in der Nordsee, insbesondere der Larven eines benthischen( 59) Echinoderms, des Echinocardium cordatum, sind darauf zurückzuführen, dass die Meerestemperatur seit 1987 schrittweise angestiegen ist und zu einer früheren Zeit des Jahres wärmere Bedingungen auftreten. Die Korrelationsanalyse hat ergeben, dass eine Zunahme der Abundanz von Echinodermlarven positiv mit einer Erhöhung der Anzahl der Tage korreliert ist, an denen eine Temperatur von mehr als 6 °C herrscht. Diese im Sommer zu verzeichnende Dominanz der Larven von benthischen Echinodermen im Plankton stellt eine erhebliche Veränderung des Gleichgewichts zwischen Meroplankton und Holoplankton dar und (58) Meroplankton besteht aus Organismen, die nur einen Teil ihres Lebenszyklus, in der Regel das Larvenstadium, im Pelagial verbringen. Zum Meeresplanktum gehören beispielsweise die Larven des Seeigels, des Gemeinen Seesterns, von Krebstieren, von Meereswürmern sowie der meisten Fischarten. (59) Bezeichnet Fische und andere Tiere, die auf dem oder im Meeresboden leben. PE 379.208 68 Klimawandel und europäische Fischerei deutet auf eine Verschiebung bei der Ressourcenaufteilung zwischen Benthos und Pelagial hin, die sich möglicherweise auf die Fischlarven auswirkt. Kernaussage 20 Änderungen bei der Primär- und Sekundärproduktion haben über das Nahrungsnetz Auswirkungen auf die Fischereiproduktion. Der Klimawandel hat bereits dazu geführt, dass sich die aufeinanderfolgenden trophischen Ebenen nicht im Gleichtakt befinden, was eine Verringerung der Sekundär- und Tertiärproduktion bewirkt hat. Bei einem weiteren Anstieg der Temperaturen wird es auch künftig zu derartigen Störungen in der marinen Nahrungskette kommen. Das hat Folgen für die Größe und die Verbreitung der Fischbestände, wie an vielen Beispielen veranschaulicht wurde: − Diatomeenblüten wirken sich negativ auf die Organismen in der Nordsee aus, bei denen die Diatomeen die Nahrungsgrundlage darstellen. − Beim Kabeljau verringern sich durch den fehlenden Gleichtakt mit der Größe der Beuteorganismen die Überlebenschancen der Larven. − Der junge Sandaal sieht sich mit einem verringerten Nahrungsangebot konfrontiert, wodurch die Überlebenschancen beeinträchtigt sind. − In den Gewässern rund um das Vereinigte Königreich bewirkt eine erhöhte Wassertemperatur eine Verringerung der Rekrutierung bei Scholle und Limande. − Eine Verschiebung des Gleichgewichts zwischen Meroplankton und Holoplankton und damit zwischen Benthos und Pelagial wirkt sich auf das Überleben der Fischlarven aus. Belege für wahrscheinliche Auswirkungen des Klimawandels auf die Aquakultur Die Aquakultur ist nach wie vor umstritten, gibt sie doch angesichts der erheblichen potenziellen Auswirkungen der intensiven Fischzucht auf die Eutrophierung der Küstengewässer (Naylor et al. 2000) Anlass zu großen ökologischen Bedenken. Darüber hinaus werden bei der Aufzucht von Flossenfischen (= ein Begriff, mit dem „echte“ Fische von Schalentieren, Krebstieren und Quallen unterschieden werden) große Mengen Fischmehl verbraucht, so dass sie vom Wildfischfang abhängig ist (Naylor et al. 2000). Allerdings erzeugen die größten Aquakulturproduktionen Mollusken, die nicht von Fischmehl abhängen, die jedoch von den direkten Auswirkungen des Temperaturanstiegs betroffen sein könnten. Obgleich sich ein Anstieg der Wassertemperatur als vorteilhaft erweisen und die Aufzucht neuer Arten in ehemals zu kalten Gebieten sowie eine Produktionssteigerung bei den bestehenden Kulturen ermöglichen könnte, ist die marine Aquakultur verschiedenen Gefahren ausgesetzt. Es gibt Informationen, nach denen Goldbrasse und Wolfsbarsch unter hohen Temperaturen leiden und die Nahrungsaufnahme einstellen (Anhaltspunkte dafür stammen aus Produktionsanlagen in Griechenland, in denen die Aufzucht in Netzgehegen erfolgt). Es kann zu einem Anstieg der Sterblichkeit kommen, der zu erheblichen Produktionsverlusten führt. Allerdings kann der mögliche wirtschaftliche Verlust in gewissem Umfang durch eine Erhöhung des Marktpreises ausgeglichen werden. Daher haben Hitzewellen wie der extrem heiße Sommer 2003 bislang nicht zu großen wirtschaftlichen Verlusten geführt. Erhöhte Wassertemperaturen an den Produktionsanlagen können jedoch zu Produktionsausfällen mit erheblichen wirtschaftlichen Folgen führen. Eine mögliche Strategie zur Vermeidung solcher wirtschaftlichen Verluste kann darin bestehen, Produktionsanlagen vom küstennahen Flachwasser in tieferes und kühleres Wasser zu verlagern. Alternativ dazu könnten für die Aquakultur auch andere Arten eingesetzt werden. Mögliche Auswirkungen des Klimawandels könnten in steigenden Wachstumsraten und einer effizienteren Futterverwertung infolge des Temperaturanstiegs bestehen (Lehtonen 1996). Andererseits wird mit Wärmestress für Kaltwasserarten und Wattorganismen wie Schalentiere gerechnet. Darüber hinaus könnte die Temperaturänderung zu Krankheiten und einer erhöhten 69 PE 379.208 Klimawandel und europäische Fischerei Krankheitsanfälligkeit bei Aquakulturarten führen. Das vermehrte Auftreten von schädlichen Algen und Quallenblüten könnte zum Absterben von Fischen und zur Schließung von Fanggebieten für Schalentiere führen (Kent und Poppe 1998). Sturmschäden an Fischzuchtanlagen Netzgehege-Kulturen können dazu führen, dass verstärkt Tiere entweichen und dass weniger geeignete Gebiete verfügbar sind, und somit eine rückläufige Entwicklung der Aquakultur bewirken (MCCIP 2006). Für die Produktion von Atlantischem Lachs entlang der norwegischen Küste wird, wenn die Temperatur steigt, damit gerechnet, dass die Produktionsanlagen nordwärts verlagert werden (Stenevik und Sundby 2006). Kernaussage 21 Es werden positive und negative Auswirkungen auf die marine Aquakultur erwartet. Steigende Wachstumsraten und eine effiziente Futterverwertung bei einigen in der Aquakultur eingesetzten Arten könnten einen Produktivitätszuwachs bewirken. Auch wäre es möglich, neue Arten in die Aquakultur einzuführen. Andererseits wird mit solch negativen Auswirkungen wie Wärmestress für Kaltwasserarten und Wattorganismen, Krankheiten und Krankheitsanfälligkeit gerechnet. Außerdem sind Schäden an Fischzuchtanlagen aufgrund von extremen Wetterereignissen denkbar. Eventuell macht sich in Abhängigkeit von veränderten Umweltbedingungen eine Verlagerung von Produktionsanlagen an geeignetere Standorte notwendig. Darüber hinaus wird befürchtet, dass Hitzewellen insbesondere für Anlagen im küstennahen Flachwasser gravierende wirtschaftliche Folgen haben. Es ist nicht bekannt, ob die positiven Auswirkungen die negativen überwiegen werden oder umgekehrt. PE 379.208 70 Klimawandel und europäische Fischerei 4. Dokumentation und kritische Bewertung der Ergebnisse Von Januar bis Mai 2007 wurden relevante und aktuelle Fundstellen in der Fachliteratur sowie einschlägige Studien von erfahrenen Sachverständigen des Projektteams untersucht. Die Ergebnisse der Literaturstudie wurden so dokumentiert, dass die relevanten Fundstellen zu jedem einzelnen den Leser interessierenden Thema (Auswirkungen des Klimawandels auf die „Wassereigenschaften“, die „wichtigsten Fischarten“ und „sonstige damit zusammenhängende Fragen“), sowie die betreffenden Unterkapitel (siehe Tabelle 6) leicht auffindbar sind. Das Kapitel „Bibliographie“ enthält ein alphabetisches Verzeichnis der verwendeten Literatur. Anhand der erzielten Ergebnisse konnten 40 durch den Klimawandel beeinflusste Phänomene ermittelt werden. Diese Phänomene sind in Tabelle 1 aufgelistet. Sie sind den Themen „Eigenschaften des Meereswassers“, „Biologie der wichtigsten Arten“, „biologische Umwelt“, „Produktivität“ und „Fischerei“ zugeordnet. Es sei darauf hingewiesen, dass eine eindeutige Zuordnung zu den entsprechenden Themen nicht immer möglich ist. So hängen beispielsweise die Auswirkungen im Zusammenhang mit der Biologie der wichtigsten Industriefischarten in der Regel auch mit der Produktivität der industriellen Fischerei zusammen. Jedem dieser Phänomene wurde ausgehend von der Zuverlässigkeit der Literaturquellen und der Häufigkeit ähnlicher Ergebnisse ein bestimmter Zuverlässigkeitsgrad zugeordnet. Eine kritische Bewertung der Ergebnisse aus der Literaturstudie lässt sich wie folgt zusammenfassen: Die in immer größerem Umfang freigesetzten künstlichen Treibhausgase haben einen großen Anteil am Klimawandel und den Veränderungen des Meeresökosystems. Treibhausgase sind eine Gefahr für die bereits geschwächten Fischbestände und sollten daher auf ein Minimum beschränkt werden. Der Klimawandel hat nachweislich Auswirkungen auf die europäischen marinen Ökosysteme. Die direkten Auswirkungen hängen insbesondere mit Temperaturänderungen, dem Anstieg des Meeresspiegels, der Versauerung und der Salinität zusammen. Die erwarteten Auswirkungen werden die Meeresproduktivität und die industrielle Fischerei in Europa sowohl negativ als auch positiv beeinflussen. Der Klimawandel dürfte in halbumschlossenen Meeren noch schwerwiegendere Auswirkungen haben als in offenen Meeren. Abgesehen von Temperatur und Salinität entscheiden auch noch das Nahrungsangebot und das Vorhandensein geeigneter Laichgründe über die Verbreitung der Fischbestände. Direkte und indirekte Klimaeffekte können zu einer Verlagerung von Fischpopulationen, zum Eindringen fremder Arten und selbst zum Verschwinden von Arten führen. Es wurden erhebliche Auswirkungen auf die wichtigsten Industriefischarten und deren biologische Umwelt beobachtet. Veränderungen bei der Primär- und Sekundärproduktion werden über das Nahrungsnetz Folgen für die Fischereiproduktion haben. Der Klimawandel hat bereits zu einem fehlenden Gleichklang zwischen den aufeinanderfolgenden trophischen Ebenen und somit zu einer Verringerung der Sekundär- und Tertiärproduktion geführt. Wenn die Temperaturen weiterhin steigen, wird die Störung der marinen Trophodynamik anhalten. Beim gegenwärtigen Kenntnisstand ist es nicht möglich, die wirtschaftlichen Auswirkungen des Klimawandels auf die europäische Fischereiindustrie und auf die Aquakultur abzuschätzen. Gemäß den bestehenden Szenarien für den Klimawandel deutet eine Reihe erwarteter Auswirkungen auf die Industriefischarten auf eine Zunahme der Meeresproduktivität und der betreffenden Populationen hin, während andere Auswirkungen offenbar zu einem 71 PE 379.208 Klimawandel und europäische Fischerei entsprechenden Rückgang beitragen. Es ist nicht bekannt, ob die positiven Auswirkungen die negativen überwiegen werden oder umgekehrt. Eine wichtige Frage ist die, wie in Anbetracht der klimatischen Auswirkungen auf die Meeresumwelt die industrielle Fischerei künftig zu gestalten ist. Um das Fischereimanagement rechtzeitig auf die Situation einzustellen, müssen durch entsprechende Forschungsarbeiten die Kenntnisse über die Auswirkungen des Klimawandels auf die Fischerei der EU und damit verbundene Prognosen verbessert werden. Für verschiedene Aspekte wurde der Forschungsbedarf ermittelt: Forschungsbedarf im Zusammenhang mit den Auswirkungen auf die Wassereigenschaften und die wichtigsten Fischarten: − Die verfügbaren globalen Zirkulationsmodelle, die auf Klimaszenarien beruhen, behandeln die Meeresumwelt und die Veränderungen des Meeresklimas auf eine vereinfachte Weise. Es gilt die Methoden dahingehend weiterzuentwickeln, dass die globalen Modelle auf die Ebene regionaler Modelle reduziert werden. In diesem Zusammenhang werden weitere Studien über das Verhalten der thermohalinen Zirkulation infolge des Klimawandels benötigt. − Sterblichkeitskriterien sind in der Regel nutzlos, wenn es darum geht, die direkten Auswirkungen auf Fische zu bewerten. Wenn überhaupt, können Kontroll- oder Ausrichtungskriterien angewandt werden. In dieser Hinsicht besteht Forschungsbedarf. − In vielen Fällen wurden Verschiebungen bei der Verbreitung aufgezeigt, aber da die betreffenden Arten oftmals stark befischt werden, lässt sich nur schwer ein direkter Kausalzusammenhang zwischen Temperatur und Verbreitungsmuster herstellen, der die Voraussetzung für eine zuverlässige Prognose wäre. Zuverlässige Prognosen zu wahrscheinlichen Entwicklungen von Fischbeständen, die auf Effekte des Klimawandels zurückzuführen sind, lassen sich nur für einige sehr intensiv untersuchte Arten anstellen (z. B. für den Atlantischen Kabeljau). Eine Abgrenzung gegenüber anderen Wirkungsfaktoren ist schwierig. Es besteht Forschungsbedarf hinsichtlich der Bewertung von Klimaauswirkungen in Anwesenheit nichtklimatischer Stressfaktoren. − Selbst wenn verschiedene Beispiele für temperatur- oder salinitätsinduzierte Wechselwirkungen (Nahrungsorganismen, Raubfische, Konkurrenten, Reproduktion) beobachtet wurden, die zu einer Populationsverlagerung führen und auf deren Grundlage sich Wechselwirkungen und interne Veränderungen in den Ökosystemen erklären lassen, müssen Untersuchungen zur Populations- und Gemeinschaftsebene durchgeführt werden, da sich die direkten klimatischen Auswirkungen für die einzelnen Tiere nicht unmittelbar in Veränderungen bei der Verbreitung und Größe der Fischpopulationen niederschlagen. − Besondere Aufmerksamkeit sollte der Erforschung halbumschlossener Gebiete gewidmet werden, da diese am stärksten von klimabedingten Veränderungen betroffen sein werden und daher hochanfällig sind. Forschungsbedarf im Zusammenhang mit den Auswirkungen auf die biologische Umwelt von Industriefischarten: − Da sie einen wichtigen Stellenwert im Nahrungsnetz einnehmen, sind die Reaktionen der Arten in der biologischen Umwelt der Fische auf Klimaauswirkungen entscheidend für die Produktivität der Industriefischbestände. Es gilt die wahrscheinlichen Reaktionen der betreffenden Organismen auf den Klimawandel zu erforschen. − Invasive Arten treten zunehmend häufiger in europäischen Küstengewässern auf, aber die Dynamik dieser Invasionen wird nur unzureichend registriert und verstanden. Es gilt PE 379.208 72 Klimawandel und europäische Fischerei zu untersuchen, welche Auswirkungen die Invasion im Kontext der Wechselbeziehungen innerhalb der Gemeinschaften und des Ökosystems hat. − Beim gegenwärtigen Kenntnisstrand ist es nicht möglich, die durch den Klimawandel zu erwartenden Veränderungen in der weltweiten marinen Produktion quantitativ vorherzusagen, da es hierbei zu einer Vielzahl von Wechselwirkungen kommt. − Um die Auswirkungen des Klimawandels auf die Meeresumwelt zu untersuchen, müssen langfristig Daten erhoben und Zeitreihen analysiert werden. Diese langfristigen Erhebungen sollten neben der Überwachung auch Prozessstudien einschließen, um ein besseres Verständnis der Konzepte zu erlangen, die den Reaktionsmustern der wichtigsten Arten innerhalb des Systems zugrunde liegen. Dem Zugang zu marinen Daten für Zeitreihenanalysen und ihrer Verwertung sollte hohe Priorität eingeräumt werden. − Es muss untersucht werden, wie sich die Versauerung der Meere auf die Meeresbiota und deren Physiologie auswirkt, und zwar insbesondere auf die biologische Produktivität und den Fischnachwuchs, da diese am stärksten gefährdet sind. Prozessstudien, experimentelle Arbeiten und Feldstudien sollten in biogeochemische, Zirkulations- und Klimamodelle zur Bewertung der künftigen Auswirkungen der Versauerung der Meere eingebunden werden. Forschungsbedarf im Zusammenhang mit den Auswirkungen der Fischerei: − Informationen über die Entwicklung der Fische in der marinen Umwelt, die Veränderung der genetischen Diversität und das Anpassungsmuster liegen kaum vor. Es müssen Maßnahmen zur Anpassung an den Klimawandel entwickelt und angewandt werden. − Gebiete, die sich für eine räumliche Absperrung eignen (Meeresschutzgebiete), sind zu überwachen und zu bewerten. − Es müssen Strategien für die Erholung stark befischter Arten festgelegt und bewertet werden; in diesem Bereich besteht noch Forschungsbedarf. − Es besteht Forschungsbedarf mit Blick auf eine nachhaltige und angemessene Bewirtschaftung der Fischbestände, die die Anpassungsfähigkeit der Fischbestände an veränderte Umweltbedingungen erhöht. − Der Ökosystemansatz für die Bestandsbewirtschaftung erfordert Kenntnisse über die Infrastruktur von Fischbeständen und ihre Wirkungsweise. Daher sind detaillierte Informationen über die Ökologie wichtiger Meeresarten, ihre Lebenszyklen und ihr Wanderungsverhalten sowie über ihren biologischen Hintergrund und ihre Wechselbeziehungen mit der biotischen und abiotischen Umwelt( 60) erforderlich. Es gilt fortgeschrittene Ökosystemmodelle sowie die multidisziplinäre Forschung weiterzuentwickeln, um die Methoden zur Erkennung, Vorhersage und Vorausschätzung von Reaktionen des Ökosystems auf den Klimawandel zu verbessern und eine wirksame Bewirtschaftung sowie eine nachhaltige Nutzung der Ressourcen zu ermöglichen. (60) Die nicht lebendige Umwelt, die alle abiotischen Faktoren umfasst, z. B. Temperatur und Salinität. 73 PE 379.208 Klimawandel und europäische Fischerei PE 379.208 74 Klimawandel und europäische Fischerei 5. Schlussfolgerungen und Empfehlungen Treibhausgase tragen zum Klimawandel bei und sind eine Gefahr für die industrielle Fischerei Die in immer größerem Umfang freigesetzten künstlichen Treibhausgase haben einen großen Anteil am Klimawandel und den Veränderungen des Meeresökosystems. Treibhausgase sind eine Gefahr für die bereits geschwächten Fischbestände und sollten daher auf ein Minimum beschränkt werden. Die Wassereigenschaften verändern sich bereits Die Wissenschaftler sind sich darin einig, dass der Klimawandel Realität ist. Die Temperaturen werden ebenso ansteigen wie der Meeresspiegel. Anzeichen für eine Klimaerwärmung sind Veränderungen bei den Oberflächen- und Lufttemperaturen sowie den Temperaturen in den oberen Ozeanschichten bis zu mehreren hundert Meter Tiefe und der Anstieg des Meeresspiegels. Die durchschnittliche globale Temperatur ist während der letzten hundert Jahre um ~0,6 °C angestiegen, der Meeresspiegel um 0,17 m. In diesem Zeitraum haben sich sowohl die Meeres- als auch die Süßwassersysteme erwärmt. Die Prognosen sprechen von einer weiteren Erwärmung des Küstenklimas in Europa im Verlaufe des 21. Jahrhunderts, wobei mit einer Erhöhung der Meeresoberflächentemperatur um 0,2 °C in jedem Jahrzehnt gerechnet wird. Modellschätzungen sagen eine Versauerung der Ozeane voraus. Der pH-Wert an der Meeresoberfläche wird während der nächsten 100 Jahre um 0,3 bis 0,5 Einheiten zurückgehen und in den kommenden 300 Jahren um 0,3 bis 1,4 Einheiten. Die Salinität wird sich verändern. So wird für die nordischen Meere und die Ostsee mit einer Abnahme der Salinität() gerechnet, während sie im Mittelmeer voraussichtlich ansteigen wird. Die atlantische thermohaline Zirkulation wird schwächer werden. Zwischen der Nordsee und den Gewässern in den arktischen Regionen findet ein Austausch von kalten und warmen Wasserströmungen statt, die durch die atlantische thermohaline Zirkulation bewegt werden. Die Fließintensität wird durch den Klimawandel beeinflusst. Sie dürfte derzeit bereits um 30 % verringert sein. Es ist unwahrscheinlich, dass die Zirkulation einmal gänzlich zum Erliegen kommt, was die derzeitigen Gegebenheiten sehr stark verändern würde (z. B. ein kurzfristiger Anstieg des Meeresspiegels in der Nordsee um 1 m, ein langfristiger Anstieg des globalen Meeresspiegels um 0,5 m, Verschiebung des tropischen Niederschlagsgürtels nach Süden und eine Verringerung der Biomasse im Atlantik um 50 %). Die Schichtung wird sich verstärken Es wurde nachgewiesen, dass der Klimawandel zu einer verstärkten Schichtung der Ost- und Nordsee und des Mittelmeeres führt. Durch die Schichtung der Wassersäule entstehen natürliche Barrieren, wo es zur Konzentration von Organismen kommen kann oder die von den Organismen passiert werden müssen. Die Schichtung verstärkt sich mit zunehmender Salinität und Temperatur. Eine verstärkte Schichtung wiederum behindert die Vermischung mit Tiefenwasser und führt zu einer geringeren Anreicherung mit Nährstoffen. Durch eine veränderte Zirkulation und Schichtung wird auch die geographische Verbreitung der Organismen verändert. Die Strömungen sind wichtig für den Transport von Organismen wie Plankton() und Fisch über große Entfernungen und können daher zu deren größerer Verbreitung beitragen. Andererseits jedoch wirken die Strömungen auch als biogeographische Barrieren zwischen den zu beiden Seiten befindlichen Wassermassen. Der Austausch von Organismen über die Strömung hinweg wird dadurch verringert. Durch die Erwärmung könnte es zu einer Abschwächung der Küstenströme kommen, wodurch auch die Verbreitung über diese Ströme geringer wird. Dadurch könnten sich Organismen, die zuvor auf die küstennahen 75 PE 379.208 Klimawandel und europäische Fischerei Bereiche beschränkt waren, weiter ausbreiten. Alle diese Vorgänge wirken sich (positiv oder negativ) auf die Primärproduktion aus. Der Klimawandel dürfte in halbumschlossenen Meeren noch schwerwiegendere Auswirkungen haben als in offenen Meeren. Die erwarteten Auswirkungen werden die Meeresproduktivität sowohl negativ als auch positiv beeinflussen. Szenarien für den Klimawandel prognostizieren für die Ostsee einen Rückgang der Salinität von 8 bis 50 % und einen Temperaturanstieg beim Meeresoberflächenwasser um 2 bis 4 °C. Für die Nordseeregion wird mit unterschiedlichen Entwicklungen bei der Salinität gerechnet, d. h. mit Anstiegen bzw. Rückgängen in verschiedenen Gebieten der Nordsee. Die Meeresoberflächentemperaturen sollen in der nördlichen Nordsee zwischen 1,6 °C und 3,0 °C steigen und in der flacheren südlichen Nordsee zwischen 3,0 °C und 3,9 °C. Den Szenarien für den Klimawandel zufolge werden Salinität und Temperatur im Mittelmeer ansteigen. Die Nordatlantik-Oszillation Die Nordatlantik-Oszillation wirkt sich auf das europäische Meeresökosystem aus. Das Winterklima im europäischen Raum wird in vielerlei Hinsicht sehr stark von der so genannten Nordatlantik-Oszillation (NAO) beeinflusst, die die atmosphärischen Vorgänge im Nordatlantik dominiert. Es wurden verschiedene Indizes für die NAO festgelegt, die sich auf alle trophische Ebenen in den Meeren auswirkt. Die Rekrutierung der Industriefische hängt vom NAO-Index ab, wie es für die starke Nachwuchsproduktion bei den Kabeljauartigen (betrifft Kabeljau, Schellfisch, Wittling und Seelachs) in der Nordsee und für die Rekrutierung des Herings und der Sardine im Nordostatlantik aufgezeigt wurde. Die NAO ist so gut wie nicht prognostizierbar, obwohl sie sich aus der Meeresoberflächentemperatur rekonstruieren lässt. Die Ex-Post-Untersuchungen des NAO-Index und der Zusammenhang mit beobachteten Auswirkungen auf das marine Ökosystem dürften es möglich machen, Modelle für die Prognostizierung künftiger Auswirkungen zu entwickeln. In vielen Fällen wurden Auswirkungen auf die wichtigsten Industriefischarten beobachtet Temperaturveränderungen können die Verlagerung von Fischpopulationen, das Eindringen fremder Arten und das Verschwinden von Arten zur Folge haben. Bei den Fischen ist die Temperatur eine ganz wesentliche Nischen-Komponente. Fische wählen in der Regel Habitate, deren thermische Bedingungen eine größtmögliche Wachstumsrate ermöglichen. Allerdings lassen sich die Auswirkungen von Temperaturveränderungen auf die Fische nur schwer vorhersagen. Abgesehen von der Temperatur entscheiden auch noch das Nahrungsangebot und das Vorhandensein geeigneter Laichplätze über die Verbreitung der Fischbestände. Direkte und indirekte Klimaeffekte können zu einer Verlagerung von Fischpopulationen, zum Eindringen fremder Arten und selbst zum Verschwinden von Arten führen. In verschiedenen Studien wurde ein Zusammenhang zwischen Größe und Verteilung von Fischbeständen und Zooplankton und einem Anstieg der Meerestemperatur beobachtet. Warmwasserfische dringen in "kalte" Ökosysteme vor. Verschiedene Arten von Warmwasserfischen sind in "kalte" Ökosysteme vorgedrungen, während Kaltwasserfische, die in "warmen" Ökosystemen in relativ großer Zahl vorhanden waren, inzwischen sehr selten geworden oder gänzlich verschwunden sind. So gibt es z. B. im Mittelmeer kaum noch Sprotten und Makrelen, und auch in vielen anderen Fällen wurden Verschiebungen festgestellt. Da jedoch die betreffenden Arten oftmals stark befischt werden, lässt sich nur schwer ein direkter Kausalzusammenhang zwischen Temperatur und Verbreitungsmuster herstellen. Zuverlässige PE 379.208 76 Klimawandel und europäische Fischerei Prognosen zu möglichen Entwicklungen von Fischbeständen, die auf Effekte des Klimawandels zurückzuführen sind, lassen sich nur für einige bislang sehr intensiv untersuchte Arten anstellen (z. B. Kabeljau). Eine Abgrenzung gegenüber anderen Wirkungsfaktoren ist schwierig. Weitere Forschungsarbeiten sind erforderlich. Der Klimawandel beeinflusst Größe und Verbreitung der Industriefischbestände. Das sich verändernde Klima hat einen direkten Einfluss auf Überlebensrate, Ausbreitung, Fertilität und Verhalten der Einzeltiere und damit auf die Größe und Verbreitung der Bestände. Prognosen sind schwierig, da außer dem Klima noch viele andere Faktoren für die Verbreitung der Arten und die Dynamik dieser Veränderungen von großer Bedeutung sind. Veränderungen in der geographischen Streuung haben um Grönland herum zu einer erhöhten Produktivität bei Kabeljau und Schellfisch beigetragen. Das ist eine Reaktion auf eine dramatische Erwärmung in den 1920er und 1930er Jahren des vorigen Jahrhunderts, die Hauptursache für diese Veränderungen jedoch waren in der Tiefe ausgelöste Prozesse infolge einer vermehrten Phytound Zooplanktonbildung. Die mittleren Temperaturen am Meeresboden können die Wachstumsrate eines Fischbestandes wesentlich beeinflussen. Wie simple Simulationen für den Nordseekabeljau gezeigt haben, ist bei steigenden Meeresbodentemperaturen mit negativen Folgen für die Wachstumsrate zu rechnen. Solche Ansätze zeigen, dass sich die Prognosen hinsichtlich des Verhaltens von Fischpopulationen ändern können, wenn bei den entsprechenden Modellen Umweltfaktoren mit einbezogen werden. Simulationen können wertvolle Aufschlüsse für die Bewirtschaftung der Fischbestände geben, und sie machen deutlich, dass dabei die Berücksichtigung von Umweltauswirkungen wahrscheinlich immer wichtiger wird. Wie an mehreren Beispielen deutlich wird, kann sich der Klimawandel indirekt sowohl positiv als auch negativ auf die Reproduktion von Industriefischbeständen auswirken. Im Falle des Norwegischen Frühjahrslaichers (Hering) hat ein klimatisch bedingter Anstieg der Wassertemperatur zur Folge, dass die Fischpopulationen nicht am üblichen Laichprozess teilnehmen. Das wirkt sich indirekt auf das Reproduktionspotenzial aus und hat direkte Folgen für die gewerbliche Fischerei (Bestandsverluste) Der Rote Thun könnte sich während warmer Perioden auch außerhalb seiner traditionellen Laichgründe vermehren, was der Produktivität des Bestandes förderlich wäre. Klimaveränderungen verursachen Verlagerungen bei den Industriefischpopulationen. Es gibt mehrere Beispiele dafür, dass temperatur- oder salinitätsbedingte Veränderungen bei den Wechselwirkungen (Nahrungsorganismen, Raubfische, Konkurrenten, Reproduktion) zur Verlagerung der Populationen führen. Mit Hilfe dieser Beispiele lassen sich einige Wechselwirkungen und internen Veränderungen in den Ökosystemen erklären. Es müssen jedoch Untersuchungen zur Populations- und Gemeinschaftsebene durchgeführt werden, da sich die direkten klimatischen Auswirkungen für die einzelnen Tiere nicht unmittelbar in Veränderungen in Verbreitung und Größe der Fischpopulationen niederschlagen. Die beim Plankton durch den Klimawandel hervorgerufenen Effekte haben die Verlagerung von Fischpopulationen zur Folge. Viele Industriefischarten sind direkt vom Plankton abhängig. Bei Fischarten, die sich von Plankton ernähren, insbesondere Sardinen und Sardellen, gibt es eine starke natürliche Fluktuation im Zusammenhang mit Klimaveränderungen. Untersuchungen bezüglich der Klimaerwärmung ließen erkennen, dass statt der nördlichen Arten in zunehmendem Maße die südlichen Arten dominieren. Bei der Untersuchung der Grenzen von mehr als 60 verschiedenen Nordseefischarten zeigte sich bei der Hälfte dieser Arten (befischt und nicht befischt) eine Verschiebung der Grenzen, wobei die Tendenz nach Norden geht. Einige Arten dürften ihre Belastbarkeitsgrenze erreicht haben, wie etwa der Kabeljau in der 77 PE 379.208 Klimawandel und europäische Fischerei Nordsee, weshalb sich ihre Populationen nach Norden bewegen. Der Rückgang beim Kabeljau wurde mit veränderten Artenzusammensetzungen, Bestandsabnahme und einer kleineren durchschnittlichen Körpergröße des Zooplanktons in Zusammenhang gebracht, was sich möglicherweise auf den Klimawandel zurückführen lässt. Die Verlagerung von Populationen (wie am Beispiel des Kabeljau gezeigt) kann dazu führen, dass in einzelnen Regionen manche Bestände gänzlich verschwinden. Auswirkungen auf die biologische Umwelt betreffen auch die Industriefischarten Der Klimawandel hat Auswirkungen auf die Beutetiere und die natürlichen Feinde von Industriefischen. Wichtige Organismen in der biotischen Umwelt von Industriefischarten sind ihre Beutetiere und ihre natürlichen Feinde. Die wichtigsten Beuteorganismen für planktonfressende Fischarten sind kleine und große Ruderfußkrebse. In der Ostsee sind kleine Arten wie Pseudocalanus und Acartia wichtige Nahrungsorganismen, wohingegen in der Nordsee und im Nordostatlantik auch große Ruderfußkrebse wie Calanus finmarchicus und C. helgolandicus von Bedeutung sind. Da sie einen wichtigen Stellenwert in der Nahrungskette einnehmen, sind ihre Reaktionen auf Klimaauswirkungen entscheidend für die Produktivität der Industriefischbestände. Invasive Arten treten zunehmend häufiger in europäischen Küstengewässern auf. Die biologische Invasion ist zu einem der auffälligsten Merkmale des globalen Wandels geworden. Durch sie können die biologische Vielfalt und die Funktionen von natürlichen Ökosystemen verändert werden, und sie kann erheblichen wirtschaftlichen Schaden verursachen. Klimatisch bedingte Störungen der Nahrungskette beeinflussen das Überleben und die Produktivität von Industriefischbeständen. Für das Überleben und die Produktivität von Fischbeständen ist es wichtig, dass für die Menge an Fischlarven ausreichend Zooplankton in der richtigen Größe vorhanden ist. Aufgrund der klimatischen Veränderungen durchlaufen viele Planktongruppen ihre jahreszeitlichen Zyklen schneller, und der fehlende Gleichtakt hat schwerwiegende Folgen für das Überleben und die Produktivität der Industriefischbestände. Bei einem weiteren Anstieg der Temperaturen wird es auch künftig zu derartigen Störungen in der marinen Nahrungskette kommen. Der Klimawandel beeinflusst damit auch die Abundanz und Verbreitung der Industriefischbestände. Das hat Folgen für die Größe und die Verbreitung der Fischbestände, wie an vielen Beispielen veranschaulicht wurde: Diatomeenblüten wirken sich negativ auf die Organismen in der Nordsee aus, bei denen die Diatomeen die Nahrungsgrundlage darstellen. Beim Kabeljau verringern sich durch den fehlenden Gleichtakt mit der Größe der Beuteorganismen die Überlebenschancen der Larven. Auch der junge Sandaal sieht sich mit einem verringerten Nahrungsangebot konfrontiert, wodurch die Überlebenschancen beeinträchtigt sind. In den Gewässern rund um das Vereinigte Königreich bewirkt eine erhöhte Wassertemperatur eine Verringerung der Rekrutierung bei Scholle und Limande. Eine Verschiebung des Gleichgewichts zwischen Meroplankton und Holoplankton und damit zwischen Benthos und Pelagial wirkt sich auf das Überleben der Fischlarven aus. Wirtschaftliche Auswirkungen für die industrielle Fischerei Der Klimawandel kann schwerwiegende wirtschaftliche Auswirkungen für die industrielle Fischerei haben. Ausgehend davon, dass die industrielle Fischerei unmittelbar mit der marinen Produktion verbunden ist, würde ein Anstieg bzw. Rückgang der Produktivität um 10 % einen wirtschaftlichen Gewinn bzw. Verlust von mehr als 200 Mio. EUR bedeuten. Die Auswirkungen der Bestandsverlagerungen auf die industrielle Fischerei lassen sich nur schwer beurteilen, da PE 379.208 78 Klimawandel und europäische Fischerei der durch die Verlagerung bedingte Schwund normalerweise mit einer entsprechenden Zunahme anderer Arten einhergeht. Es werden positive und negative wirtschaftliche Auswirkungen für die industrielle Fischerei erwartet. Beim gegenwärtigen Kenntnisstand ist es nicht möglich, die durch den Klimawandel zu erwartenden Veränderungen in der weltweiten marinen Produktion quantitativ vorherzusagen, da hierbei eine Vielzahl von Wechselwirkungen auftreten. Zweifellos jedoch werden Veränderungen der Produktivität und der Saisonmuster die Bewirtschaftung der biologischen Ressourcen der Meere beeinflussen. Wie (nicht sehr zuverlässige) Schätzungen besagen, dürfte es bis 2050 zu einer Zunahme der weltweiten marinen Produktion kommen, jedoch um nicht mehr als 10 % Im Gegensatz dazu haben Satellitenbeobachtungen und umfangreiche Probenahmen von Plankton gezeigt, dass Phytoplankton und Chlorophyll in den letzten 20 bis 50 Jahren zurückgegangen sind. Für 74 % der permanent geschichteten Weltmeere ist die Oberflächenerwärmung mit einer Verringerung der Produktivität verbunden. In den nördlichen Breiten wird mit einer Produktionszunahme gerechnet, da große Gebiete eisfrei werden. Mögliche Auswirkungen auf die Aquakultur Der Klimawandel kann schwerwiegende wirtschaftliche Auswirkungen auf die Aquakultur haben. Die Meeresaquakultur in der EU erreicht einen Marktwert von rund 2 Mrd. EUR, wobei einschneidende wirtschaftliche Veränderungen aufgrund des Klimawandels befürchtet werden. Es werden positive und negative Auswirkungen erwartet. Steigende Wachstumsraten und eine effiziente Futterverwertung bei einigen in der Aquakultur eingesetzten Arten könnten einen Produktivitätszuwachs bewirken. Auch wäre es möglich, neue Arten in die Aquakultur einzuführen. Andererseits wird mit solch negativen Auswirkungen wie Wärmestress für Kaltwasserarten und Wattorganismen, Krankheiten und Krankheitsanfälligkeit gerechnet. Außerdem sind Schäden an Fischzuchtanlagen aufgrund von extremen Wetterereignissen denkbar. Eventuell macht sich in Abhängigkeit von veränderten Umweltbedingungen eine Verlagerung von Produktionsanlagen an geeignetere Standorte notwendig. Darüber hinaus wird befürchtet, dass Hitzewellen insbesondere für Anlagen im küstennahen Flachwasser gravierende wirtschaftliche Folgen haben. Es ist nicht bekannt, ob die positiven Auswirkungen die negativen überwiegen werden oder umgekehrt. Die Auswirkungen der industriellen Fischerei auf die wichtigsten Arten Während der letzten Jahrzehnte wurden bei den kommerziellen Fischbeständen im Atlantik zahlreiche Veränderungen beobachtet. Dabei ist es äußerst schwierig, die aus Veränderungen in der Populationsdichte und der Rekrutierung resultierenden Folgen sowie die regionalen Klimaeffekte von direkten anthropogenen Einflüssen, wie etwa der Fischerei, zu unterscheiden. Die derzeitige Fischereipraxis schwächt die Widerstandsfähigkeit der Fischbestände gegenüber den Auswirkungen des Klimawandels. Die derzeitigen Fangmethoden richten sich auf ältere und größere Fische und bewirken eine Veränderung der Größen- und Altersstruktur der Fischpopulationen. Die Widerstandsfähigkeit gegenüber Umweltauswirkungen wird geschwächt, die genetische Variabilität geht verloren und die Fähigkeit zur Anpassung an Umweltveränderungen wird schwächer. Folglich kann die Fischerei dazu führen, dass die Umweltauswirkungen für die Bestände zu einer immer größeren Gefahr werden. Eine ökologisch nachhaltige Fischerei sollte den Schutz der größeren und älteren Fische zum Ziel haben. 79 PE 379.208 Klimawandel und europäische Fischerei Unter dem Gesichtspunkt der Nachhaltigkeit befischte Bestände können besser auf Klimaauswirkungen reagieren. Die Ungewissheit über den künftigen anthropogenen Klimawandel rechtfertigt einen Vorsorgeansatz für die Bestandsbewirtschaftung. Die globale Erwärmung kann sich erheblich - in positiver oder negativer Art und Weise - auf die meisten der kommerziellen Fischbestände auswirken. Dabei sind durch Überfischung drastisch reduzierte Bestände anfälliger gegenüber klimatischen Veränderungen als nachhaltig befischte Bestände. Maßgeblich für die Reaktion der Fischbestände auf Umwelteinflüsse ist die Populationsgröße. Gesunde Bestände können sich besser an Populationsverschiebungen und strukturelle Veränderungen im Ökosystem anpassen. Eine flexible und anpassungsfähige Bestandsbewirtschaftung ist notwendig. Ein mögliches Szenario für die Zukunft der Nordsee beinhaltet einen Anstieg der Temperatur, eine starke NAO und einen verstärkten Zufluss von Atlantikwasser. Folgen davon wären eine geringe Rekrutierung des Kabeljau, eine Nordwärtsverlagerung der jetzigen Fischarten (Kabeljau, Hering und Sprotte) und das Vordringen südlicher Arten (Sardine und Sardelle) Eine wichtige Frage ist die, wie in Anbetracht der klimatischen Auswirkungen auf die Meeresumwelt die industrielle Fischerei künftig zu gestalten ist. Da einige der wichtigsten Industriefischarten (wie Hering und wahrscheinlich andere kleine pelagische Arten) sehr stark auf sich verändernde hydrographische Bedingungen reagieren, sollte die künftige Bestandsbewirtschaftung kontinuierlich sein, gleichzeitig aber auch flexibel und anpassungsfähig, um den jeweiligen Reaktionen der Bestände auf die Umweltbedingungen Rechnung zu tragen. Insbesondere die ausgesprochenen Wanderarten, die ihre Routen bei Umweltveränderungen ändern, werden besondere Anforderungen an die Bewirtschaftung stellen. Um das Fischereimanagement rechtzeitig auf die Situation einzustellen, müssen durch entsprechende Forschungsarbeiten die Kenntnisse über die Auswirkungen des Klimawandels auf die Fischerei der EU und damit verbundene Prognosen verbessert werden. Strategische Optionen Die in immer größerem Umfang freigesetzten künstlichen Treibhausgase haben einen großen Anteil am Klimawandel. Der Klimawandel wirkt sich direkt wie auch indirekt auf vielerlei Art und Weise auf die industrielle Fischerei und die Meeresaquakultur in Europa aus. Insgesamt steht dabei ein Marktvolumen von mehr als 4 Mrd. EUR auf dem Spiel. Es besteht kein Zweifel, dass die Klimaveränderungen das Meeresökosystem sowie die Fischerei und die Meeresaquakultur der EU positiv und negativ beeinflussen werden. Ungewiss ist jedoch, ob dabei die positiven oder negativen Auswirkungen überwiegen werden. Die Kenntnisse über die Beziehungen und Wechselwirkungen zwischen dem Meeresökosystem, der industriellen Fischerei, der Meeresaquakultur und den Klimaveränderungen und ihr Verständnis sind in vielerlei Hinsicht noch unzureichend. Unter Berücksichtigung verschiedener strategischer Alternativen wurden sieben generelle Optionen ermittelt (die nachfolgend erörtert werden): Option 1. Keine Maßnahmen. Option 2. Verringerung der Treibhausgase. Gesetzgeberische und politische Maßnahmen sowie Unterstützung bestehender Initiativen zur Senkung der Treibhausgasemissionen. Option 3. Strategien zur Erhöhung der Widerstandsfähigkeit der Fischpopulationen. Entwicklung von Strategien und Konzepten für eine nachhaltige Bewirtschaftung der PE 379.208 80 Klimawandel und europäische Fischerei Option 4. Option 5. Option 6. Option 7. Fischbestände, wodurch deren Widerstandsfähigkeit gegenüber Umweltauswirkungen erhöht wird. Strategien zur Verbesserung der Bestandsbewirtschaftung. Entwicklung von flexiblen Strategien und Konzepten für die Bestandsbewirtschaftung, die entsprechend den Reaktionen der Bestände auf die Umweltbedingungen angepasst werden können. Strategien zur Verbesserung der Aquakultur. Entwicklung von Strategien und Konzepten, um die negativen Auswirkungen auf die Meeresaquakultur so gering wie möglich zu halten. Förderung der Forschung. Förderung von entsprechenden Forschungsarbeiten und des Austauschs von Kenntnissen. Kompensation negativer Auswirkungen. Bereitstellung von Unterstützung (finanzieller oder anderer Art), um die negativen Auswirkungen des Klimawandels zu kompensieren. Diskussion der Optionen: Option 1) Da nicht erwiesen ist, dass der Klimawandel eine insgesamt negative Auswirkung auf die EU-Fischerei haben wird, wäre es eine mögliche Option, gar keine Maßnahmen zu ergreifen. Dies hätte den Vorteil, dass keinerlei Anschubkosten anfallen. Die positiven Auswirkungen des Klimawandels auf die Fischerei, wie etwa eine erhöhte Produktion des Meeresökosystems aufgrund eines Temperaturanstiegs, werden nicht eingeschränkt. Andererseits erscheint es unakzeptabel, nicht zumindest gegen die negativen Folgen des Klimawandels vorzugehen (einschließlich finanzielle Auswirkungen), da diese ohne irgendwelche Gegenmaßnahmen in vollem Umfang wirksam werden. Option 1 wird nicht empfohlen. Option 2) Diese Option betrifft die Unterstützung einer europäischen Politik zur Bekämpfung des Klimawandels (z. B. Monitoring und Berichterstattung bezüglich der EUTreibhausgasemissionen oder das Emissionshandelssystem) und von geeigneten Maßnahmen, um die Treibhausgaskonzentrationen in der Atmosphäre so gering wie möglich zu halten. Da die Treibhausgasemissionen eine Ursache des Klimawandels sind, werden mit dieser Option die Klimaveränderungen direkt an der Quelle bekämpft. Option 2 wird empfohlen. Option 3) Es hat sich gezeigt, dass die frühere wie auch die jetzige industrielle Fischerei die Widerstandsfähigkeit der Fischbestände gegenüber Umweltveränderungen schwächt. Ziel von Option 3 ist die Entwicklung von Strategien für eine nachhaltige Fischerei, wodurch diese Widerstandsfähigkeit erhöht wird. Dazu ließen sich beispielsweise Möglichkeiten untersuchen, wie der Fang von älteren und größeren Fischen reduziert werden kann, oder es könnten marine Schutzgebiete eingerichtet werden. Mit dieser Option soll die gegenwärtige Fischerei so gestaltet werden, dass sie den Auswirkungen des Klimawandels Rechnung trägt. Das entspricht den im EU-Anpassungsprogramm im Rahmen des Europäischen Programms zur Klimaänderung (ECCP) festgelegten Zielen, wonach Möglichkeiten gefunden werden sollen, um die Resistenz Europas gegen die Folgen des Klimawandels in verschiedenen Sektoren zu stärken. Die Gemeinsame Fischereipolitik (GFP) spielt eine entscheidende Rolle bei der Bewirtschaftung der Fischbestände und sollte dabei mögliche Klimaauswirkungen entsprechend berücksichtigen. Sie könnte auch ein geeignetes Instrument sein, um die Widerstandsfähigkeit der Fischbestände gegenüber den Auswirkungen des Klimawandels durch entsprechende Strategien zu verbessern. Option 3 wird empfohlen. Option 4) Die Fischbestände werden durch spezifisches Verhalten auf den Klimawandel reagieren, z. B. durch zeitliche oder räumliche Veränderungen in der Bestandsgröße oder durch Veränderungen in den Wanderrouten. Bei Option 4 besteht das Ziel darin, die 81 PE 379.208 Klimawandel und europäische Fischerei Bestandsbewirtschaftung auf die Reaktionen der Fischbestände einzustellen, um deren effiziente Nutzung zu ermöglichen. Dazu ist ein flexibles Bewirtschaftungsregime erforderlich, da nur so negative wirtschaftliche Folgen vermieden werden können. Diese Option hat das Ziel, die jetzige Bestandsbewirtschaftung an die Auswirkungen des Klimawandels anzupassen und befindet sich daher in Übereinstimmung mit den Zielsetzungen des EU-Anpassungsprogramms im Rahmen des ECCP. Unbedingt müssen dabei auch die Nachhaltigkeitsaspekte im Sinne von Option 2 berücksichtigt werden. Anderenfalls besteht die Gefahr, dass die verbesserte Bewirtschaftung nur für eine maximale Ausbeutung der Meeresressourcen genutzt wird, ohne darauf hinzuwirken, dass die Widerstandsfähigkeit der Fischbestände gegenüber den Umweltauswirkungen erhöht wird. Wie bei Option 3 könnte die GFP maßgeblich zur Umsetzung entsprechender Strategien beitragen. Option 4 wird empfohlen. Option 5) Ähnlich wie bei den Optionen 3 und 4 handelt es sich hier um eine Anpassungsstrategie entsprechend dem EU-Anpassungsprogramm im Rahmen des ECCP. Das Ziel ist die Vermeidung von negativen Auswirkungen auf die Meeresaquakultur. Denkbare Bestandteile der Strategie sind beispielsweise die Verlagerung von Produktionsanlagen Richtung Norden, die Verlagerung von Produktionsstätten in tieferes und kälteres Wasser oder die verstärkte Ausrichtung auf neue Arten. Ähnlich wie bei den Optionen 3 und 4 könnte die GFP maßgeblich zur Umsetzung entsprechender Strategien beitragen. Option 5 wird empfohlen. Option 6) Insbesondere die Optionen 3 bis 5 sind mit einem umfangreichen Forschungsbedarf verbunden. Angemessene Kenntnisse über die Beziehungen und Wechselwirkungen zwischen dem Meeresökosystem, der industriellen Fischerei, der Meeresaquakultur und dem Klimawandel sind unerlässlich. Eine solide Wissensgrundlage ist angesichts der Gefahren durch den Klimawandel eine Voraussetzung für die Entwicklung effizienter Strategien für eine nachhaltige Fischerei, Bestandsbewirtschaftung und Meeresaquakultur. Spezifische Forschungsaufgaben sind in Kapitel 4 aufgeführt. Die EU könnte entsprechende Forschungen anregen und/oder unterstützen, um die Faktengrundlage für ihre politische Entscheidungen und die zu entwickelnden Strategien zu verbessern, und sie könnte zur Verbreitung der gewonnenen Erkenntnisse beitragen. Option 6 wird empfohlen. Option 7) Diese Option betrifft die Bereitstellung von Unterstützung (finanzieller oder anderer Art), um die negativen Auswirkungen des Klimawandels zu kompensieren. Dazu gehören Ausgleichszahlungen für Regionen, die große wirtschaftliche Einbußen als Folge klimatischer Veränderungen erlitten haben (z. B. durch Produktionsausfälle in der Aquakultur nach Hitzewellen oder durch den Zusammenbruch der Fischrekrutierung, wenn eine Fischpopulation aus der Region verschwunden ist). Solche Zahlungen könnten der betreffenden Region kurzfristig helfen. Allerdings tragen Ausgleichsmaßnahmen reaktiven Charakter und sind nicht zukunftsorientiert. Proaktive und wissensbasierte Ansätze dürften hier am besten geeignet sein (z. B. geänderte Strategien für die Aquakulturproduktion oder das Fischereimanagement. Option 7 wird nicht empfohlen. Mit der Tabelle „Überblick über die strategischen Optionen und deren Vor- und Nachteile“ in der Zusammenfassung kann der Leser rasch einen Überblick über die entwickelten strategischen Optionen und deren Vor- und Nachteile erlangen. PE 379.208 82 Klimawandel und europäische Fischerei ANHÄNGE Anhang 1. Biologische Merkmale der Arten Anhang 1-1: Biologische Merkmale der wichtigsten industriellen Fischarten Hering (Clupea harengus L. 1758) Nordostatlantik, Nordsee, Ostsee Maximale Länge Höchstalter Lebensraum (Wasserzone, -art und -tiefe) Temperaturbereich 45 cm 25 Jahre Pelagial, Meer und Brackwasser; 0-200 m 1 °C – 18 °C Die Verdoppelung der Population dauert mindestens 1,4 – 4,4 Jahre (mittlere Dauer). Der Hering zählt nach wie vor zu den wichtigsten kommerziell genutzten Arten im Nordatlantik. Er bildet Schwärme und führt umfangreiche Nahrungs- und Laichwanderungen durch. Heringe führen täglich vertikale Wanderungen durch, wobei sie sich tagsüber im tiefen Wasser aufhalten und nachts in Richtung Oberfläche schwimmen, um der vertikalen Bewegung ihrer Beute zu folgen. Sie ernähren sich von planktonischen Organismen wie Ruderfußkrebsen und im Pelagial lebenden Schnecken. Ausgewachsen stellen Heringe die wichtigste Beute von Raubfischen wie Makrelen und Kabeljauen dar. Zum Laichen wandern sie zum Meeresgrund und legen ihre Eier in das Substrat in Küstennähe. Das Überleben der Eier und Larven hängt von der Anwesenheit von Raubfischen und den abiotischen Umweltbedingungen ab. In Europa existieren mehrere verschiedene Bestände, die sich in ihren Wanderrouten und Laichzeiten unterscheiden. Sprotte (Sprattus sprattus L. 1758) Nordostatlantik, Nordsee, Ostsee, Mittelmeer Maximale Länge Höchstalter 16 cm 6 Jahre Lebensraum (Wasserzone, -art und -tiefe) Pelagial, Meer und Brackwasser; 10-150 m Temperaturbereich gemäßigt Die Verdoppelung der Population dauert weniger als 15 Monate (kurz). Wie die Heringe bilden Sprotten Schwärme und wandern ebenfalls zwischen ihren Nahrungs- und Laichgründen hin und her. Tagsüber halten sie sich im tiefen Wasser auf und folgen dann nachts ihrer Beute an die Wasseroberfläche. Sie ernähren sich hauptsächlich von kleinem Krebsplankton. Sprotten laichen im freien Wasser und können während der gesamten Laichzeit bis zu 14 000 Eier produzieren (Muus und Nielsen 1999). Ausgewachsen stellen sie die wichtigste Beute von Raubfischen (d. h. Kabeljau, Makrele) dar. 83 PE 379.208 Klimawandel und europäische Fischerei Sardine (Sardina pilchardus Walbaum 1792) Nordostatlantik, Mittelmeer Maximale Länge Höchstalter 25 cm 15 Jahre Lebensraum (Wasserzone, -art und -tiefe) Pelagial, Meer und Brackwasser; 10-100 m Temperaturbereich subtropisch Die Verdoppelung der Population dauert mindestens 1,4 – 4,4 Jahre (mittel). Sardinen bilden Schwärme und führen vertikale Wanderungen durch, wobei sie tagsüber in 100 m Tiefe leben und nachts ihrer planktonischen Beute folgen und auf 20 m aufsteigen. Sie können bis zu 50 000 Eier produzieren, die sie in das freie Wasser abgeben (Muus and Nielsen 1999). Sardelle (Engraulis encrasicolus L. 1758) Nordostatlantik, Mittelmeer Maximale Länge Höchstalter 20 cm 3 Jahre Lebensraum (Wasserzone, -art und -tiefe) Pelagial, Meer, Brackwasser und Süßwasser; 0-400 m Temperaturbereich subtropisch Die Verdoppelung der Population dauert weniger als 15 Monate (kurz). Die Sardelle lebt überwiegend an der Küste und ist im Frühling (Laichzeit) regelmäßig auch in Flussmündungen und sogar Seen anzutreffen. In den Sommermonaten wandern die Sardellen in Richtung Norden und geben von April bis November in Tiefen von bis zu 50 m Eier in das freie Wasser ab. Im Winter bleiben sie im tiefen Wasser (bis zu 150 m) (Whitehead et al. 1988). Die planktonfressenden Sardellen ernähren sich sowohl von Zooplankton als auch von Phytoplankton (James 1988). Makrele (Scomber scombrus L. 1758) Nordostatlantik, Mittelmeer Maximale Länge Höchstalter 60 cm 17 Jahre PE 379.208 Lebensraum (Wasserzone, -art und -tiefe) Pelagial, Meer und Brackwasser; 0-1000 m 84 Temperaturbereich gemäßigt Klimawandel und europäische Fischerei Die Verdoppelung der Population dauert mindestens 1,4 – 4,4 Jahre (mittel). Makrelen leben in kaltem Wasser und gemäßigten Riffgebieten. Im Nordostatlantik wird zwischen zwei verschiedenen Beständen unterschieden, dem Nordseebestand (im Osten) und dem Bestand der Britischen Inseln (im Westen). Markelen leben in riesigen Schwärmen nahe der Wasseroberfläche. Sie überwintern im tiefen Wasser und nähern sich im Frühling mit dem Anstieg der Wassertemperatur der Küste. Eier und Larven leben im freien Wasser. Ausgewachsene Makrelen ernähren sich von Zooplankton und kleinen Fischen. Roter Thun (Thunnus thynnus L. 1758) Atlantik, Mittelmeer Maximale Länge Höchstalter Lebensraum (Wasserzone, -art und -tiefe) Temperaturbereich 460 cm 15 Jahre Pelagial, Meer und Brackwasser; 0-200 m 3 °C – 29 °C Die Verdoppelung der Population dauert mindestens 4,5 – 14 Jahre (lang). Der Rote Thun ist ein sehr großer Fischfresser. Bei der Verfolgung seiner Beute, die überwiegend aus Schwarmfischen besteht, legt er sehr weite Strecken zurück. Die meiste Zeit des Jahres hält er sich im Ozean auf und bewegt sich während der Laichzeit im Sommer (Juni) in die Nähe der Mittelmeerküste. Die Weibchen können pro Laichzeit bis zu zehn Millionen Eier produzieren und legen diese im offenen Meer ab. Die natürlichen Feinde des Roten Thuns sind vor allem verschiedene Haiarten, Schwertwale und bestimmte Vögel. Blauer Wittling (Micromesistius poutassou Risso 1827) Nordsee Maximale Länge Höchstalter 50 cm 20 Jahre Lebensraum (Wasserzone, -art und -tiefe) Pelagial, Meer Temperaturbereich gemäßigt Die Verdoppelung der Population dauert mindestens 1,4 – 4,4 Jahre (mittel). Der Blaue Wittling lebt am Festlandabhang und -sockel in bis zu 1000 m, überwiegend jedoch in 300-400 m Tiefe. Tagsüber hält er sich in der Nähe des Meeresgrunds auf, um sich dann nachts an die Oberfläche zu begeben. Die Jungfische ernähren sich überwiegend von Krebsplankton, die ausgewachsenen Fische von kleinen Fischen und Kopffüßern ( 61). Seine Eier legt der Blaue Wittling im Frühling an den Küsten Großbritanniens, Islands und Norwegens in das freie Wasser ab. (61) Tiere (Weichtiere) mit am Kopf und um den Mund zusammenlaufenden Tentakeln (Tintenfische, Kuttelfische, Kraken). 85 PE 379.208 Klimawandel und europäische Fischerei Kabeljau (Gadus morhua L. 1758) Nordostatlantik, Nordsee, Ostsee Maximale Länge Höchstalter 200 cm 25 Jahre Lebensraum (Wasserzone, -art und -tiefe) Benthopelagial, Meer und Brackwasser; 0-600 m Temperaturbereich -1 °C – 10 °C Der Verdoppelung der Population dauert mindestens 1,4 – 4,4 Jahre (mittel). Der Kabeljau ist eine weit verbreitete Art. Er lebt und ernährt sich nahe des Meeresbodens (von wirbellosen Tieren und Fischen), bildet Gruppen und begleitet mitunter andere Arten. Er führt Wanderungen von den Nahrungs- zu den Laichgründen durch. Die Laichzeit liegt zwischen Frühling (Nordsee) und Spätsommer (östliche Ostsee). Die Eier werden in das freie Wasser abgegeben. Europäischer Seehecht (Merluccius merluccius L. 1758) Nordostatlantik, Nordsee, Mittelmeer Maximale Länge Höchstalter 140 cm 20 Jahre Lebensraum (Wasserzone, -art und -tiefe) Meeresgrund, Meer; 30-1000 m Temperaturbereich gemäßigt Die Verdoppelung der Population dauert mindestens 4,5 – 14 Jahre (lang). Der Europäische Seehecht lebt normalerweise in 70 bis 370 m Tiefe, wobei die Jungfische in größeren Tiefen leben und sich mit steigendem Alter in die Küstengewässer bewegen. Die Jungfische ernähren sich von Krebstieren (vor allem Euphausiiden und Amphipoden), die ausgewachsenen Fische überwiegend von anderen Fischen. Seehechte führen Laichwanderungen von den Nahrungsgründen nahe der Küste zu den Laichgründen in tieferem Gewässer durch. Die Laichzeit dauert von Frühling bis Sommer. Die Eier werden in das freie Wasser abgegeben. Sandaal (Hperoplus lanceolatus Le Sauvage 1824) Nordsee Maximale Länge Höchstalter 40 cm 25 Jahre Lebensraum (Wasserzone, -art und -tiefe) Meeresgrund, Meer und Brackwasser; 6-30 m Die Verdoppelung der Population dauert mindestens 1,4 – 4,4 Jahre (mittel). PE 379.208 86 Temperaturbereich gemäßigt Klimawandel und europäische Fischerei Sandaale werden gemeinhin mit der Art der Ammodyten in Verbindung gebracht. Sie ernähren sich im Wesentlichen von Zooplankton und kleinen Fischen. Sie laichen in den Sommermonaten und legen klebrige Eier am Meeresgrund ab. Kleiner Sandaal (Ammodytes marinus Raitt 1934) Nordsee Maximale Länge Höchstalter 25 cm 10 Jahre Lebensraum (Wasserzone, -art und -tiefe) Benthopelagial, Meer und Brackwasser Temperaturbereich gemäßigt Die Verdoppelung der Population dauert mindestens 1,4 – 4,4 Jahre (mittel). Der Kleine Sandaal bildet Schwärme und lebt meist nahe der Küste. In Zeiten geringer Lichtintensivität (Nacht und Winter) vergräbt er sich im sandigen Untergrund. Er ernährt sich im Wesentlichen von Zooplankton. Er laicht im Winter und vergräbt seine Eier im Sand. Schellfisch (Melanogrammus aeglefinus L. 1758) Nordostatlantik, Nordsee Maximale Länge Höchstalter 100 cm 20 Jahre Lebensraum (Wasserzone, -art und -tiefe) Meeresgrund, Meer; 10-450 m Temperaturbereich 4 °C - 10 °C Die Verdoppelung der Population dauert mindestens 1,4 – 4,4 Jahre (mittel). Der Schellfisch lebt meist in 80 bis 200 m Tiefe über Felsen, Sand, Kies oder Muscheln. Er ernährt sich hauptsächlich von kleinen am Meeresgrund lebenden Organismen wie Krebstieren, Weichtieren, Stachelhäutern, Würmern und Fischen. Er legt seine Eier in einer Tiefe von 50-150 m in das freie Wasser ab. Seelachs (Pollachius virens L. 1758) Nordostatlantik, Nordsee Maximale Länge Höchstalter Lebensraum (Wasserzone, -art und -tiefe) 87 Temperaturbereich PE 379.208 Klimawandel und europäische Fischerei 130 cm 25 Jahre Meeresgrund, Meer; 0-200 m gemäßigt Die Verdoppelung der Population dauert mindestens 1,4 – 4,4 Jahre (mittel). Der Seelachs ist ein aktiver, gesellig lebender( 62) Fisch, der in Küstennähe und im offenen Meer lebt. Normalerweise hält er sich im Frühling in Küstengewässern auf und kehrt dann im Winter in tiefere Gewässer zurück. Die jüngeren Tiere leben in Küstengewässern und ernähren sich von kleinen Krebstieren (Copepoden, Amphipoden, Euphausiiden) und kleinen Fischen, während die älteren Fische überwiegend andere Fische fressen. Scholle (Pleuronectes platessa L. 1758) Nordostatlantik, Nordsee Maximale Länge Höchstalter 100 cm 50 Jahre Lebensraum (Wasserzone, -art und -tiefe) Meeresgrund, Meer und Brackwasser; 0-200 m Temperaturbereich 2 °C – 15 °C Die Verdoppelung der Population dauert mindestens 4,5 – 14 Jahre (lang). Die Scholle lebt auf gemischtem Untergrund, ist nachtaktiv und vergräbt sich tagsüber im Sand. Die Jungfische halten sich in Küstennähe in seichtem Gewässer auf, während die älteren Fische in größeren Tiefen leben. Schollen ernähren sich überwiegend von Weichtieren und Vielborstern. Obwohl oft davon ausgegangen wird, dass Schollen immer am selben Ort bleiben, wurde anhand von Kennzeichnungsexperimenten nachgewiesen, dass auch sie Wanderungen über große Strecken durchführen können. Sie laichen im Spätwinter und Frühling und geben ihre Eier in das freie Wasser ab. Ohrensardine (Sardinella aurita Valenciennes 1847) Mittelmeer Maximale Länge Höchstalter Lebensraum (Wasserzone, -art und -tiefe) Temperaturbereich 31 cm 7 Jahre Riffnähe, Meer und Brackwasser; 0-350 m subtropisch Die Verdoppelung der Population dauert weniger als 15 Monate (kurz). Ohrensardinen leben in Schwärmen in Küstengewässern, sowohl nah an der Küste als auch am Rand des Festlandsockels. Die Jungfische halten sich in Küstennähe in den Nahrungsgründen auf und schließen sich nach Erreichen der Geschlechtsreife ihren ausgewachsenen Artgenossen an. Ohrensardinen ernähren sich von benthischen Krebstieren und Zooplankton. Sie laichen das (62) In Bezug auf eine soziale Gruppe. PE 379.208 88 Klimawandel und europäische Fischerei ganze Jahr über, wobei es keinen speziellen Zeitraum gibt, in dem sie mehr laichen als in einem anderen. Ihre Eier legen sie im offenen Meer ab. Stöcker (Trachurus trachurus L, 1758) Atlantik, Mittelmeer Maximale Länge Höchstalter Lebensraum (Wasserzone, -art und -tiefe) Temperaturbereich 70 cm --- Jahre Pelagial, Meer (ozeanodrom); 0-1050 m 15 °C – 31 °C Die Verdoppelung der Population dauert mindestens 4,5 – 14 Jahre (lang). Der Stöcker bildet große Schwärme in Küstengebieten und ernährt sich hauptsächlich von Fisch, Krebstieren und Kopffüßern. Echter Bonito (Katsuwonus pelamis L. 1758) Atlantik, Mittelmeer Maximale Länge Höchstalter 100 cm 12 Jahre Lebensraum (Wasserzone, -art und -tiefe) Pelagial, Meer (ozeanodrom); 0-260 m Temperaturbereich 15 °C – 30 °C Die Verdoppelung der Population dauert mindestens 1,4 – 4,4 Jahre (mittel). Der Echte Bonito lebt vor allem im offenen Meer und bildet gern Schwärme nahe der Wasseroberfläche. Man findet ihn häufig bei im Wasser treibenden Gegenständen. Er laicht im offenen Meer. Die Larven beschränken sich auf Gewässer mit Oberflächentemperaturen von 15 °C bis 30 °C. Die natürlichen Feinde des Echten Bonito sind vor allem verschiedene Haiarten, Schwertwale und bestimmte Vögel. 89 PE 379.208 Klimawandel und europäische Fischerei Gelbflossenthun (Thunnus albacares Bonnaterre, 1788) Atlantik, Mittelmeer Maximale Länge Höchstalter 240 cm 8 Jahre Lebensraum (Wasserzone, -art und -tiefe) Pelagial, Brackwasser und Meer (ozeanodrom); 0-250 m Temperaturbereich 15 °C – 31 °C Die Verdoppelung der Population dauert mindestens 1,4 – 4,4 Jahre (mittel). Der Gelbflossenthun lebt im Meer. Die einzelnen Tiere schließen sich je nach Größe zu Schwärmen zusammen. Man findet die Schwärme häufig in der Nähe von im Wasser treibenden Gegenständen. Der Gelbflossenthun ist bei der Zubereitung von Sashimi sehr beliebt. PE 379.208 90 Klimawandel und europäische Fischerei Anhang 1-2: Biologische Merkmale des biologischen Lebensraums der wichtigsten industriellen Fischarten Pseudocalanus acuspes, Ostsee Der P. acuspes ist eine boreale Art und kann sich daher an niedrige Temperaturen und einen hohen Salzgehalt anpassen (Renz und Hirche 2006). Er profitiert von kalten Wintern und dem Zustrom von stark salzigem Nordseewasser in die Ostsee (Möllmann u. a. 2000, 2003). Acartia spp. Ostsee, Nordsee Die Gattung Acartia besteht aus mehreren Arten, die in allen Gebieten vorkommen. In der Ostsee dominieren die Arten A. bifilosa und A. longiremis. Beide Arten sind euryhalin( 63), d. h. sie vertragen große Schwankungen im Salzgehalt. Darüber hinaus kommen ihnen die höheren Temperaturen zugute (Möllmann u. a. 2000, 2003). In der Nordsee existieren darüber hinaus die A. clausi und A. tonsa. Die A. clausi wird als typische marine Art beschrieben (Hansen 1960), während es sich bei der A. tonsa um eine (euryhaline und eurytherme( 64)) Brackwasserart handelt. Calanus finmarchicus und Calanus helgolandicus Nordostatlantik, Nordsee Der C. finmarchicus stellt eine an kaltes Wasser angepasste marine Art dar und ist ein Indikator für subarktisches Wasser. Der C. helgolandicus hat sich an ein gemäßigtes Umfeld angepasst und ist sowohl in ozeanischem als auch in neritischem ( 65)Wasser anzutreffen. Beide Arten führen täglich vertikale Wanderungen über mehrere hundert Meter durch. Centropages typicus Nordsee, Mittelmeer Bei dem C. typicus handelt es sich um eine Warmwasserart, die sowohl für die Nordsee als auch das Mittelmeer typisch ist (Clark 1933). Die für ihre Entstehung erforderliche Mindesttemperatur beträgt 8 °C-12 °C (Bigelow 1926). Temora stylifera Nordsee, Mittelmeer Die T. stylifera stellt eine neritische Art dar, die auch in einem ozeanischen Lebensraum angetroffen werden kann. Sie bevorzugt niedrigere Temperaturen (Mazza 1961). In Europa gezüchtete größere Organismen sind die Miesmuschel, die Regenbogenforelle und der Lachs. Nordeuropa konzentriert sich auf die Salmoniden, während im Mittelmeerraum mehr als 40 verschiedene Arten gezüchtet werden (Hoepffner 2006). Die nachfolgende Tabelle enthält ausschließlich marine Organismen. (63) Organismus, der bei verschiedenstem Salzgehalt überleben kann. (64) Organismus, der bei verschiedensten Temperaturen überleben kann. (65) Bezieht sich auf den ozeanischen Bereich oberhalb des Festlandsockels und die Kante des Schelfabhangs. Entspricht den küstennahen Gewässern. 91 PE 379.208 Klimawandel und europäische Fischerei Anhang 1-3: Biologische Merkmale der wichtigsten Arten der marinen Aquakultur Lachs (Salmo salar L. 1785) Maximale Länge Höchstalter 150 cm 13 Jahre Lebensraum (Wasserzone, -art und -tiefe) Pelagial, Süßwasser, Meer und Brackwasser (anadrom); 0-210 m Temperaturbereich 2 °C – 9 °C Die Verdoppelung der Population dauert mindestens 1,4 – 4,4 Jahre (mittel). Lachs wird vor allem in Norwegen gezüchtet. In freier Wildbahn wachsen die Lachse im Meer auf und wandern in die Flüsse zurück, um dort zu laichen. Die Jungfische bleiben ein bis sechs Jahre im Süßwasser, bevor sie ins Meer wandern. Ausgewachsene Tiere, die sich der Geschlechtsreife nähern, nehmen im Süßwasser keine Nahrung zu sich. Goldbrasse (Sparus aurata L. 1785) Maximale Länge Höchstalter 70 cm 11 Jahre Lebensraum (Wasserzone, -art und -tiefe) Meeresgrund, Meer und Brackwasser (anadrom); 0-150 m Temperaturbereich subtropisch, 4 °C > Die Verdoppelung der Population dauert mindestens 1,4 – 4,4 Jahre (mittel). Goldbrassen werden vor allem in Spanien, Italien und Griechenland gezüchtet. Sie sind euryhalin und eurytherm, können also bei verschiedenstem Salzgehalt und unterschiedlichsten Temperaturen überleben. Obwohl in der Literatur nur von einer unteren Temperaturgrenze gesprochen wird, liegt die obere Temperaturgrenze, oberhalb der sie nichts mehr fressen und sich die Mortalität dramatisch erhöht, wohl bei ca. 26 °C. PE 379.208 92 Klimawandel und europäische Fischerei Meerbarsch (Dicentrarchus labrax L. 1785) Maximale Länge Höchstalter 100 cm 15 Jahre Lebensraum (Wasserzone, -art und -tiefe) Meeresgrund, Süßwasser, Meer und Brackwasser (anadrom); 10-100 m Temperaturbereich subtropisch, 8-24 °C Die Verdoppelung der Population dauert mindestens 1,4 – 4,4 Jahre (mittel). Meerbarsche werden vor allem in Spanien, Italien und Griechenland gezüchtet. Sie sind euryhalin und eurytherm, können also bei verschiedenstem Salzgehalt und unterschiedlichsten Temperaturen überleben. Miesmuschel (Mytilus edulis L. 1785) Maximale Länge Höchstalter cm 24 Jahre Lebensraum (Wasserzone, -art und -tiefe) Benthos, Meer und Brackwasser, Tidebereich bis zum Sublitoral Temperaturbereich 5 °C – 20 °C Die M. edulis ist weit verbreitet, was vor allem darauf zurückzuführen ist, dass sie großen Schwankungen in Bezug auf Salzgehalt, Austrocknung, Temperatur und Sauerstoffspannung standhalten kann. Obwohl Miesmuscheln eine Lebenserwartung von 18 bis 24 Jahren haben können, werden die meisten gezüchteten Muscheln nach weniger als zwei Jahren verarbeitet. 93 PE 379.208 Klimawandel und europäische Fischerei Blaubart-/Seemuschel (Mytilus galloprovincialis Lamark 1819) Maximale Höchstalter Lebensraum (Wasserzone, -art und -tiefe) Länge --- --- Benthos, Meer und Brackwasser, Tidebereich bis zum Sublitoral Temperaturbereich 5 °C – 20 °C Bis Anfang der 1990er Jahre wurden alle Muscheln aus Westeuropa als Mytilus edulis bezeichnet. Mittlerweile weiß man, dass die Muscheln aus dem Gebiet zwischen der südlichen Bretagne (Frankreich) und dem Mittelmeer M. galloprovincialis sind. Portugiesische Auster (Crassostrea gigas Thunberg 1793) Maximale Länge Höchstalter Lebensraum (Wasserzone, -art und -tiefe) Temperaturbereich --- --- Benthos, Meer und Brackwasser, 0-40 m -1.8 °C – 35 °C Der für die Portugiesische Auster optimale Salzgehalt liegt zwischen 20 % und 25 %, obwohl sie auch bei einem Salzgehalt von unter 10 % vorkommen und in einem Salzgehalt von über 35 % überleben kann, in dem sie sich jedoch wahrscheinlich nicht fortpflanzt. Obwohl die Auster in einem großen Temperaturbereich überleben kann, laicht sie üblicherweise bei Temperaturen von über 20 °C und selten bei 15–18 °C. Unter guten Bedingungen wächst sie sehr schnell; ihre Marktgröße erreicht sie in 18 bis 30 Monaten. PE 379.208 94 Klimawandel und europäische Fischerei Japanische Teppichmuschel (Ruditapes philippinarum Adams & Reeve, 1850) Maximale Länge Höchstalter --- --- Lebensraum (Wasserzone, -art und -tiefe) Meer Temperaturbereich subtropisch leicht boreal bis Obwohl die Japanische Teppichmuschel in einem großen Temperaturbereich überleben kann, laicht sie üblicherweise bei Temperaturen zwischen 20 °C und 25 °C. Die untere Temperaturgrenze liegt bei 12 °C. Unter guten Bedingungen wächst sie sehr schnell; ihre Marktgröße erreicht sie in 18 bis 36 Monaten. Regenbogenforelle (Oncorhynchus mykiss Walbaum , 1792) Maximale Länge Höchstalter 120 cm 11 Jahre Lebensraum (Wasserzone, -art und -tiefe) Benthopelagial, Süßwasser, Meer und Brackwasser (anadrom); 0-200 m Temperaturbereich 0 °C – 27 °C Die Verdoppelung der Population dauert mindestens 1,4 – 4,4 Jahre (mittel). Die Züchtung von Regenbogenforellen ist sehr weit verbreitet. Regenbogenforellen können in einem großen Temperaturbereich überleben, obwohl die optimale Laich- und Wachstumstemperatur zwischen 9 °C und 14 °C liegt. Die Zuchttemperatur liegt üblicherweise unter 21 °C. Steinbutt (Psetta maxima L , 1758) Maximale Länge Höchstalter 100 cm 25 Jahre Lebensraum (Wasserzone, -art und -tiefe) Meersgrund, Meer und Brackwasser (ozeanodrom); 20-70 m Temperaturbereich gemäßigt Die Verdoppelung der Population dauert mindestens 1,4 – 4,4 Jahre (mittel). Obwohl der Steinbutt überwiegend in Europa gezüchtet wird, wurde die Art in Chile und kürzlich auch in China eingeführt. Die optimale Zuchttemperatur liegt zwischen 14 °C und 18 °C. Marktgröße kann der Steinbutt in 24 bis 30 Monaten erreichen. 95 PE 379.208 Klimawandel und europäische Fischerei Große Teppichmuschel (Ruditapes decussatus L, 1758) Maximale Länge Höchstalter --- --- Umgebung (Wasserzone, -art und -tiefe) Meer Temperaturbereich subtropisch leicht boreal bis Die Große Teppichmuschel wird an der Atlantikküste Frankreichs, Spaniens und Portugals und dem Mittelmeerbecken gezüchtet. Zum Überleben benötigt sie einen sandigen Lebensraum. Marktgröße kann sie in 18 bis 36 Monaten erreichen. Europäische Auster (Ostrea edulis L, 1758) Maximale Länge Höchstalter --- --- Umgebung (Wasserzone, -art und -tiefe) Brackwasser, Meer Temperaturbereich Gemäßigt, bis 25 °C 10 °C Die Europäische Auster stammt aus Europa. Sie wird von Großbritannien bis Griechenland gezüchtet und kann in einem Salzgehalt von lediglich 15 % überleben. Für das Wachstum der Larven ist ein Salzgehalt von über 20 % optimal. Aal (Anguilla anguilla L , 1758) Maximale Länge Höchstalter 130 cm 85 Jahre Umgebung (Wasserzone, -art und -tiefe) Temperaturbereich Meeresgrund, Süßwasser, Meer und Brackwasser (katadrom); 0-700 m Gemäßigt, 4 °C – 24 °C Die Verdoppelung der Population dauert mehr als14 Jahre (sehr lange Zeit). Die drei Länder, die in der Aalzucht momentan führend sind, sind die Niederlande, Italien und Dänemark. Bei der Aalzucht werden in freier Wildbahn Glasaale gefangen, die dann für die weitere Aufzucht verwendet werden. Die extensive Aalzucht unter natürlichen Bedingungen ist mittlerweile auf ein Ausmaß zurückgegangen, das keine kommerzielle Bedeutung hat. Zur gängigen Zuchtmethode hat sich die intensive Zucht mithilfe der Rezirkulation entwickelt, bei der die Wassertemperatur konstant bei ca. 24 °C gehalten wird. PE 379.208 96 1169 1711 1747 1923 1773 1773 1988 1814 1805 1805 1980 1818 1876 1843 Alheit et al. 2005 Araujo & New 2007 Araujo et al. 2005 Arnell 1998 Arnott & Ruxton 2002 Arnott & Ruxton 2002 Astraldi et al. 1995 Attrill & Power 2002 Attrill et al. 2007 Attrill et al. 2007 BACC 2006 Basilone et al. 2006 Beare et al. 2002 Beare et al. 2004a PE 379.208 1821 Lfd. Nr. 1981 Alheit & Hagen 1997 ACIA 2005 Verweis 4.1.3 4.1.2 Wasser Sonstiges Fisch 4.2.5 4.1.5 4.3.7 4.3.1 4.2.2 4.3.6 4.3.7 4.2.3 4.2.7 4.2.3 4.2.3 4.2.6 4.1.6 Unterkapitel 4.3.3 Fisch Wasser Fisch Fisch Fisch Sonstiges Sonstiges Fisch Sonstiges Fisch Fisch Fisch Wasser Sonstiges Bezug zu Temperatur Calanus, Temperatur, Salinität, Schichtung, Zufluss Temperatur 97 Temperaturveränderung, Salinität Quallen Quallen NAO Artenverbreitung Verbreitung Laichzeitpunkt Anstieg Anstieg Änderung Nordsee Nordsee Straße von Sizilien Ostsee Nordsee Nordsee Mündungsgebiet der Themse Mittelmeer Nordsee Verschiebung der Verbreitung; Rekrutierung in Abhängigkeit vom Calanus-Vorkommen Fischrekrutierung, NAO Änderung Nordsee Verschiebung der Verbreitung; Rekrutierung in Abhängigkeit vom Calanus-Vorkommen Fischrekrutierung, NAO Nordsee, Vereinigtes Königreich Änderung Wasserressourcen, Klimaauswirkungen weltweit weltweit Nordsee, Ostsee Nordatlantik, Nordsee Nordatlantik Gebiet Änderung Änderung Änderung Wechsel Anmerkung Temperatur Temperatur Systemänderung NAO, Hering, Sardine Primärproduktion und Klima Stichwort Anhang 2. Dokumentierung der Ergebnisse der Literaturauswertung allgemein Calanus Sardelle allgemein Qualle Qualle allgemein Sandaal Sandaal allgemein allgemein Kabeljau, Sprotte, Hering, Ruderfußkrebs Hering, Sardine allgemein Art Klimawandel und europäische Fischerei 1842 1924 1802 374 374 374 374 2061 2061 575 1905 1920 1998 Beare et al. 2004a Beare et al. 2004b Beaugrand 2004 Beaugrand et al. 2002 Beaugrand et al. 2003 Beaugrand et al. 2003 Beaugrand et al. 2003 Beaugrand et al. 2003 Behrenfeld et al. 2006 Behrenfeld et al. 2006 Berkeley et al. 2004 Bethoux et al. 1999 Bethoux et al. 2002 Bigelow 1928 PE 379.208 Verweis Lfd. Nr. 1843 4.3.4 4.3.7 Fisch, Sonstiges Fisch, Sonstiges Sonstiges Wasser 4.3.1 4.1.3 4.1.5 4.3.4 Fisch, Sonstiges Sonstiges 3.3.3 Sonstiges 3.1.3 4.3.3 Fisch, Sonstiges Wasser 4.2.6 4.3.7 Zooplankt on Fisch 4.3.6 4.1.3 Unterkapitel 4.2.2 Fisch Fisch Fisch Bezug zu Klimawandel und europäische Fischerei Temperaturanstieg; Invasion neuer Arten Verschiebung der Verbreitung von Sardine und Sardelle in der Nordsee Gleichtakt/fehlender Gleichtakt; in der Tiefe ausgelöste Prozesse Gleichtakt/fehlender Gleichtakt; in der Tiefe ausgelöste Prozesse Gleichtakt/fehlender Gleichtakt; in der Tiefe ausgelöste Prozesse Rekrutierung beim Kabeljau, Zooplankton, Überleben der Kabeljaularven Rekrutierung beim Kabeljau, Zooplankton, Überleben der Kabeljaularven Rekrutierung beim Kabeljau, Zooplankton, Überleben der Kabeljaularven Temperatur Auswirkung der NAO auf das Phytoplankton, Verschiebung 98 Auswertung des besonderen Systems im Mittelmeerraum Bedeutung der Muttertiere Produktivität in hohen Breitengraden Zucht Erhaltung von Alttieren Rückgang Erderwärmung Gleichtakt/fehlender Gleichtakt; in der Tiefe ausgelöste Prozesse Rekrutierung beim Kabeljau, Zooplankton, Überleben der Kabeljaularven Schichtung Anstieg Temperatur Systemänderung Verbreitung Anmerkung Temperatur Stichwort Golf von Maine Mittelmeer weltweit weltweit weltweit Nordsee Nordsee Nordsee Nordsee Nordsee Nordsee Nordsee Nordsee Gebiet Centropages Plankton Steinbeißer Kabeljau Kabeljau Kabeljau Kabeljau Ruderfußkrebs allgemein Sardine, Sardelle allgemein Art 1837 1886 1778 2017 1891 1925 1925 1752 1926 1726 1927 1824 2002 1928 1855 Birkeland & Dayton 2005 Bjornsson & Steinarsson 2002 Bombace 2001 Brander 1994 Brander 1995 Brander 2006 Brander 2006 Brander et al. 2003 Brett 1956 Browman et al. 2000 Brown 2003 Bryden et al. 2005 Buecher 1999 Burton 1979 Caldeira & Wickett 2003 PE 379.208 Lfd. Nr. Verweis Sonstiges Fisch Sonstiges Wasser 4.1.1 4.2.1 4.3.6 4.1.2 4.2.1 4.2.7 Fisch, Sonstiges Fisch 4.2.1 4.3.7 Zooplankt on Fisch 4.3.3 4.1.3 4.2.2 4.2.4 4.2.2 Sonstiges Fisch Fisch Fisch Fisch 4.2.4 4.3.4 Fisch, Sonstiges Fisch Unterkapitel Bezug zu weltweit Reaktion während der Lebensdauer, phänotypische Plastizität, Selektion im Hinblick auf schnelleres Wachstum Versauerung der Ozeane 99 weltweit weltweit stärkere Änderung der pH-Werte als bei Fossilien Mittelmeer Nordostatlantik Physiologie Änderung langfristige Schwankungen beim Vorkommen von Quallen Temperatur und Belüftung Rückgang der Zirkulation weltweit weltweit Reaktion auf Temperaturveränderungen weltweit weltweit Atlantik, Nordsee Nordsee Nordsee Physiologie Gebiet Mittelmeer weltweit weltweit Anstieg Bewertung anhand von Modellen thermohaline Zirkulation Temperaturnachweis beim Fisch evolutionäre Änderungen aufgrund von Fischerei Temperatur, Physiologie, Ökologie Temperatur Temperatur und Schichtung Bewertung anhand von Modellen Änderung Temperatur und Wachstum, Temperatur am Meeresboden Temperatur und Schichtung Wachstumsrate Änderung glockenförmiges Verhältnis Erhaltung von Alttieren Anmerkung Temperatur Verschiebung der Verbreitung Versuche: Temperatur, Wachstum Bedeutung der Muttertiere Stichwort allgemein in Ästuaren lebende Fische Qualle thermohaline Zirkulation Hai allgemein allgemein Ruderfußkrebs allgemein allgemein Kabeljau Kabeljau Sprotte, Makrele Kabeljau allgemein Art Klimawandel und europäische Fischerei 1854 1997 1801 1982 2054 2054 2057 1751 1863 1929 1826 1914 1730 1930 1823 Caldeira & Wickett 2005 Carotenuto et al. 2006 Casini et al. 2006 Cayan 1992 Chavez et al. 2003 Chavez et al. 2003 Ciais et al. 2005 Ciannelli et al. 2005 Clark et al. 2003 Clarke 1993 Claussen et al. 2003 Collie et al. 2004 Cook et al. 1997 Coutant 1987 Curry & Mauritzen 2005 PE 379.208 Lfd. Nr. Verweis Wasser Fisch Fisch Fisch Wasser Fisch Fisch Fisch Wasser Wasser Fisch Wasser Fisch Sonstiges Sonstiges Bezug zu Klimawandel und europäische Fischerei 4.1.1 4.2.1 4.3.7 4.2.6 4.1.2 4.2.1 4.2.4 4.2.7 3.1.1 3.1.6 3.2.6 4.1.6 4.2.5 4.3.6 4.1.1. Unterkapitel Änderung Systemänderungen Rückgang der Kabeljaubestände Verschiebung des thermischen Äquators interne Veränderungen in den Ökosystemen Überfischung, Kollaps der Kabeljaubestände Nordatlantik Rückgang des Salzgehalts im Nordatlantik, Salinität 100 weltweit Temperaturwahl, Präferenzverhalten in Abhängigkeit von den thermischen Verhältnissen Nordsee weltweit weltweit weltweit Physiologie Pazifik Europa Pazifik Pazifik weltweit Ostsee Temperatur und Verbreitungsgrenze, Gefahr des Aussterbens Änderung Gebiet Mittelmeer weltweit Nordsee Anstieg Hitzewelle, Klimawandel Änderung des Ökosystems Änderung des Ökosystems Änderung prognostizierte Änderung der pHWerte Anmerkung Modellierung Änderung Kabeljaubestände und Klimawandel, Nordsee Temperatur Temperatur Temperatur Systemänderung Auswirkungen der Thermodynamik auf die Temperatur an der Meeresoberfläche Beutetiere Populationsdynamik bei Ruderfußkrebsen Versauerung der Ozeane Stichwort allgemein Kabeljau allgemein allgemein allgemein Kabeljau Pazifischer Pollack Sardelle, Sardine NAO Hering, Sprotte Ruderfußkrebs allgemein Art 1931 1932 1869 1869 1867 1781 788 1719 1719 1803 1804 2011 1838 Davis et al. 1998 Dickson & Brander 1993 Drinkwater 2005 Drinkwater 2005 Drinkwater 2006 Dulcic et al. 1999 Dutil & Brander 2003 Edwards & Richardson 2004 Edwards & Richardson 2004 Edwards et al. 2001 Edwards et al. 2002 Eisenreich 2005 Engelhard & Heino 2004 PE 379.208 1844 Lfd. Nr. 1887 Cushing 1990 Cushing 1984 Verweis 4.3.7 Zooplankt on Fisch 4.2.5 4.1.1. 4.3.7 Phytoplankton Wasser 4.3.3 4.3.7 4.2.2. 4.2.2 4.2.3 4.3.4 4.2.2 4.2.3 4.2.6 4.3.3 Unterkapitel 4.1.6 Plankton Sonstiges Fisch Fisch Fisch Fisch Fisch Fisch Fisch Sonstiges Sonstiges Bezug zu Atlantik hohe Temperatur; nicht erfolgende Reproduktion; norwegischer Hering Temperatur, Zustand der Fische, nicht erfolgende Reproduktion 101 Mittelmeer Nordsee Nordsee Nordsee Änderung Änderung anthropogene Ursachen Anstieg Nordsee Salinität und Klima NAO Invasion Temperatur Phänologie, fehlender Gleichtakt zeitliche Verschiebung Nordsee, Nordatlantik Auswirkungen auf die Verbreitung optimale Temperatur für Kabeljau Mittelmeer Verschiebung der Verbreitung Nordatlantik Rückgang der Kaltwasserarten, Invasion von neuen und Warmwasserarten Vergleich der prognostizierten Kabeljaubestände Kabeljaurekrutierung, Verbreitung und Temperatur Nordatlantik Atlantik Vergleich der prognostizierten Kabeljaubestände Kabeljaurekrutierung, Verbreitung und Temperatur Nordatlantik Gebiet Migration 1920-1930, Systemänderung Änderung Verbreitung der Kabeljaubestände, Umweltauswirkungen Nordsee Wechselwirkung zwischen Plankton und Fisch Änderung Nordsee NAO Anmerkung Klimahülle, Interaktion der Arten Gleichtakt/fehlender Gleichtakt „Kabeljauboom“ Stichwort Hering Plankton Coscinodiscus wailesii Plankton Phyto-, Zoo-, Meroplankton Kabeljau Sprotte, Makrele Kabeljau Kabeljau Kabeljau Kabeljau Drosophila allgemein Kabeljauartige Art Klimawandel und europäische Fischerei 1838 1878 1921 1853 1933 1987 2020 1779 1104 2017 1934 2017 1935 1444 1857 1841 Engelhard & Heino 2004 Engelhard & Heino 2006 ESA Feely et al. 2004 Ferguson 1958 Fernandez et al. 2003 Fox et al. 2000 Francour et al. 1994 Frank et al. 2005 Froese & Pauly 2007 Fromentin & Planque 1996 Fry 1947 Fry 1971 Garcia et al. 2003 Gaylord & Gaines 2000 Genner et al. 2004 PE 379.208 Lfd. Nr. Verweis Fisch Sonstiges Fisch Fisch Fisch Fisch Fisch Fisch Fisch Fisch Sonstiges Fisch Sonstiges Fisch Fisch Fisch Bezug zu Klimawandel und europäische Fischerei 4.2.2. 4.2.3 4.3.5 4.2.1 4.2.1 4.2.5 4.2.1 4.3.4 4.2.2 4.3.7 4.3.6 4.2.1 4.1.1 4.3.5 4.2.5 4.2.7 Unterkapitel weltweit Auswirkungen der Zunahme des Gehalts an CO2 und der Abnahme des pH-Werts weltweit Reaktion auf Temperaturveränderungen 102 Nordsee Temperatur, Änderung der Verbreitung von Fischarten Auswirkungen der Wechselwirkung weltweit weltweit weltweit Reaktion auf Temperaturveränderungen Fischereiwesen Nordsee Änderung weltweit Transport – Temperatur: Bedeutung für die Verbreitung Klimawandel Temperatur - Physiologie, Ökologie Temperatur - Physiologie, Ökologie Zusammenhang zwischen NAO, Temperatur und Ruderfußkrebsen Fischdatenbank Nordatlantik Auswirkungen des Wegfalls der wichtigsten natürlichen Feinde, Umgestaltung des Nahrungsnetzes Auswirkungen der Fischerei Mittelmeer Nordsee Mittelmeer stehende und fließende Gewässer weltweit Rekrutierungsvariabilität Gebiet Nordatlantik Atlantik Fischereiwesen hohe Temperatur; nicht erfolgende Reproduktion; norwegischer Hering Anmerkung Warmwasserfische in Kaltwasserbiota Temperatur Änderung des Meeresspiegels Auswirkungen der Temperatur auf die Ökologie und Physiologie Versauerung der Ozeane Klima Klima, nicht erfolgendes Ablaichen Temperatur, Zustand der Fische, nicht erfolgende Reproduktion Stichwort viele Arten allgemein allgemein allgemein allgemein allgemein Kabeljau Scholle allgemein allgemein Hering Hering Art Lfd. Nr. 1993 1994 1911 1807 2001 2055 1817 1999 1799 234 1995 1840 1840 1936 1936 Goffart et al. 2002 Gomez & Gorsky 2003 Gomis et al. 2006 Goren & Galil 2005 Goy et al. 1989 Greene & Pershing 2007 Guisande et al. 2004 Halsband-Lenk & Hirche 2001 Hannesson 2007 Hänninen et al. 2000 Hansen 1960 Harley et al. 2006 Harley et al. 2006 Harrods et al. 2007 Harrods et al. 2007 PE 379.208 Verweis 4.2.7 Fisch, Sonstiges Fisch 4.2.1 4.1.2 4.2.6 Fisch, Sonstiges Wasser 4.3.1 4.3.6 4.2.2 4.3.6 4.3.3 3.3.3 4.3.6 4.3.2 4.1.6 4.3.6 Unterkapitel 4.3.6 Sonstiges Sonstiges Fisch Sonstiges Fisch Sonstiges Sonstiges Fisch Wasser Sonstiges Sonstiges Bezug zu 103 Klima und Fischerei, britische Inseln Klima und Fischerei, britische Inseln Auswertung Auswertung Bedeutung der Wechselwirkung, Schlussfolgerungen Auswertung der Auswirkungen des Klimawandels auf die marinen Ökosysteme in Küstengebieten unter besonderer Beachtung der Bodenfischarten Nordsee, Nordatlantik Nordsee, Nordatlantik weltweit weltweit Bedeutung der Wechselwirkung, Schlussfolgerungen Auswertung der Auswirkungen des Klimawandels auf die marinen Ökosysteme in Küstengebieten unter besonderer Beachtung der Bodenfischarten Ostsee Nordostatlantik Nordsee Mittelatlantik, Mittelmeer Europäisches Nordmeer Änderung Anstieg Änderung Änderung Arktis Mittelmeer Levantinisches Meer Mittelmeer Mittelmeer Mittelmeer Gebiet Verbreitung Temperatur, Salinität, NAO Temperatur jahreszeitlich bedingte Produktionszyklen Temperatur Zunahme Änderung langfristige Schwankungen beim Vorkommen von Quallen Produktivität in hohen Breitengraden anthropogene Ursachen Änderung Änderung Anmerkung Invasion NAO, Variabilität des Meeresspiegels, Salinität Zyklen des Mikrozooplanktons Phytoplanktonblüte Stichwort allgemein allgemein allgemein allgemein Acartia Kabeljau, Makrele, Sardine Ruderfußkrebs Sardine Qualle Fisch Phytoplankton Art Klimawandel und europäische Fischerei 1877 1754 1937 1753 1753 1740 1740 1852 2019 1813 1798 1798 23 1856 1938 1938 1938 Harvell et al. 1999 Harwood & Stokes 2003 Hauser et al. 2002 Hawkins et al. 2003 Hawkins et al. 2003 Hays et al. 2005 Hays et al. 2005 Helmuth et al. 2006 Henderson 1998 Herrick et al. 2007 Hjermann et al. 2007 Hjermann et al. 2007 Hjort 1914 Hochachka & Somero 2002 Hoepffner et al. 2006 Hoepffner et al. 2006 Hoepffner et al. 2006 PE 379.208 Lfd. Nr. Verweis 4.3.4 Fisch, Sonstiges 4.3.7 Zooplankt on Wasser Sonstiges Sonstiges 4.1.5 4.3.7 4.3.1 4.2.1 4.3.7 Fisch, Sonstiges Sonstiges 4.3.1 4.2.2 Schlussf. 4.3.7 Fisch Fisch Fisch Fisch 4.3.2 4.3.3 Zooplankt on Sonstiges 4.3.7 Allgemein 4.2.2 Schlussf. allgemeine r Zshg. Allgemein 4.2.7 Unterkapitel Sonstiges Bezug zu Klimawandel und europäische Fischerei Auswertung Klimawandel – Folgen für marine Systeme und Küstengebiete 104 Auswertung Klimawandel – Folgen für marine Systeme und Küstengebiete europäische Gewässer europäische Gewässer europäische Gewässer Auswertung Klimawandel – Folgen für marine Systeme und Küstengebiete Nordsee , Nordatlantik Nordostatlantik Nordostatlantik Pazifik Nordsee intertidale Ökosysteme Nordostatlantik Nordostatlantik Ärmelkanal Ärmelkanal Neuseeland weltweit Gebiet weltweit Rekrutierungsvariabilität Änderung Änderung Änderung Rekrutierungsvariabilität Rolle der Variabilität der Umwelt Anstieg Anstieg Änderung Änderung Verlust der Vielfalt Anmerkung biochemische Anpassungen an die Temperatur Gleichtakt/fehlender Gleichtakt Temperatur Temperatur Klima Temperatur biologische Invasion Temperatur Temperatur Temperatur Temperatur Auswirkungen der Überfischung auf die genetische Vielfalt Komplexität Klima und das Auftreten von Krankheiten Stichwort allgemein Kabeljau Kabeljau Kabeljau Sardine Limande Zooplankton Zooplankton alle alle Schnapper Modelle allgemein Art 1939 1940 1723 1941 2012 1943 1942 1996 2006 2007 1835 1990 1979 1867 Holbrook et al. 1997 Houghton et al. 2001 Hsieh et al. 2006 Hulme et al. 2002 Hurrell et al. 2006 Hurrell & Dickson 2004 Hurrell 1995 Ianora et al. 2003 ICES 2006 IPCC-Bericht 2007 Iversen et al. 2002 James 1988 Javidpour et al. 2006 Jensen 1939 wie in Drinkwater 2006 zitiert PE 379.208 1938 Lfd. Nr. Hoepffner et al. 2006 Verweis Fisch Sonstiges Fisch Fisch Wasser Wasser Sonstiges Wasser Wasser Wasser Wasser Sonstiges Wasser Fisch Sonstiges Bezug zu 4.2.3 4.1.5 4.2.1 4.2.5 4.1.1. 4.1.2 4.3.6 4.1.6 4.1.6 4.1.6 4.1.1. 4.3.4 4.1.1. 4.2.2. 4.3.3 Unterkapitel Verschiebung der Verbreitung Invasion Beutetiere Klima Prognosen des Klimawandels 105 Ostsee Nordatlantik Nordwärtsverschiebung in Abhängigkeit von Temperatur und Beutetieren Mittelmeer Nordsee Änderung Änderung weltweit Nordatlantik, Nordsee, Ostsee Änderung Bedingungen an den Grenzen der Verbreitungsgebiete weltweit Auswirkungen der NAO auf Temperatur und Niederschlag weltweit weltweit Auswirkungen der NAO auf Temperatur und Niederschlag Auswirkungen von Diatomeen auf die Reproduktion von Ruderfußkrebsen NAO-Index NAO-Index weltweit Auswirkungen der NAO auf Temperatur und Niederschlag weltweit Tatsache des Klimawandels, Zunahme der Treibhausgasemissionen NAO Kalifornienstrom Einfluss der Befischung und der Umweltvariabilität Nordatlantik weltweit Änderung Verschiebung der Verbreitung von Fischarten in Riffen in gemäßigten Zonen europäische Gewässer Gebiet Tatsache des Klimawandels, Zunahme der Treibhausgasemissionen Auswertung Anmerkung Klimawandel – Folgen für marine Systeme und Küstengebiete Stichwort Kabeljau Qualle Sardelle Bastardmakrele allgemein allgemein allgemein allgemein Fischlarven allgemein allgemein Art Klimawandel und europäische Fischerei 1944 1948 1865 1972 2022 1947 1946 1775 1839 1748 1750 271 Jobling 1997 Jones et al. 1997 Jurado-Molina & Livingstone 2002 Karl & Trenberth 2003 Kent & Poppe 1998 Kiehl & Trenberth 1997 King 2004 Kirby et al. 2006 Kirby et al. 2007 Klyashtorin 1998 Klyashtorin 2001 Köster et al. 2003 PE 379.208 1945 Lfd. Nr. 2023 Jobling 1981 Jensen 2003 Verweis Sonstiges Fisch 4.3.6 4.3.5 Schlussf. 4.3.7 Fisch, Sonstiges Fisch 4.3.4 4.1.1 4.1.1 4.3.8 4.1.1 4.2.4 4.1.6 4.2.1 4.2.1 Unterkapitel 4.2.2 Fisch Wasser Wasser Fisch Wasser Fisch Wasser Fisch Fisch Fisch Bezug zu Klimawandel und europäische Fischerei weltweit Auswirkungen der NAO auf Temperatur und Niederschlag Berücksichtigung der Auswirkungen auf die Umwelt NAO-Index Simulation von Populationsmodellen, Rekrutierung, Klima als auslösender Faktor weltweit Nordostatlantik Nordsee Analyse und Prognose des Risikos größeres Vorkommen seit dem Jahr 2002 Zunahme des Meroplanktons (Echinocardium) aufgrund gestiegener Temperaturen während des Winters und des Frühjahrs weltweiter Temperaturanstieg Temperatur und Vorkommen, höhere Überlebensraten bei Fischlarven und Jungfischen Änderung Temperatur, Salinität, Artenzusammensetzung 106 Änderung Temperatur Index der atmosphärischen Zirkulation Temperatur – Kopplung Benthos/Pelagial Ostsee weltweit weltweit weltweit verstärkte Absorption der Strahlung Anstieg der Treibhausgasemissionen weltweit Krankheiten Klima weltweit weltweit weltweit Auswirkungen der Temperatur auf das Wachstum weltweit Wechselblüter, Temperaturregulierung, Temperaturwahl weltweiter Temperaturanstieg Gebiet Nordamerika Temperaturtoleranz Verbreitung und Wachstum Anmerkung letale Temperaturgrenzen Temperatur Stichwort Fisch, Zooplankton allgemein allgemein Mero-, Holo-, Zooplankton Schlangennadel allgemein allgemein allgemein allgemein Grundfischarten allgemein allgemein allgemein Lachs Art 1430 1894 2005 1825 1973 1815 1974 1780 2063 1741 1741 1741 699 1919 2000 1746 2008 2008 Köster et al. 2005 Lehodey et al. 2006 Lehtonen 1996 Levermann et al. 2005 Levitus et al. 2000 Maes et al. 2005 Magnuson et al. 1979 Malacic et al. 2006 Malzahn et al. 2007 Maravelias & Reid 1995 Maravelias & Reid 1995 Maravelias & Reid 1995 Marteinsdottir & Steinarsson 1998 Marty & Chiaverini 2002 Mazza 1961 McCarty 2001 MCCIP 2006 MCCIP 2006 PE 379.208 Lfd. Nr. Verweis Sonstiges 4.3.2 4.1.3 Schlussf. allgemeine r Zshg. Wasser 4.3.1 4.1.3 4.3.4 4.2.3 4.2.2 4.2.1 3.3.7 4.1.1 4.2.1 4.2.1 4.1.1 4.1.2 4.3.8 Schlussf. 4.2.6 Unterkapitel Sonstiges Wasser Sonstiges Fisch Fisch Fisch Sonstiges Fisch Fisch Fisch Wasser Wasser Sonstiges Fisch, Sonstiges Fisch Bezug zu Invasion von Organismen Schichtung, Nordsee Klima Temperatur 107 Auswirkungen der NAO auf das Phytoplankton; Verschiebung größere Fische, größere Eier Temperatur Temperatur Temperatur Gleichtakt/fehlender Gleichtakt Temperatur, Änderung der Salinität Auswahl des thermischen Habitats Zunahme Zunahme Änderung Auswirkungen auf die Rekrutierung Änderung Änderung Änderung Lebensmittelqualität Strategie zur Maximierung des Wachstums Änderung Änderung Erwärmung der marinen und der Süßwassersysteme Futterverwertung Schlussfolgerungen Änderung Temperatur Anmerkung Veränderungen bei Plankton und Fischabundanz sowie den entsprechenden Zusammenhängen Meeresspiegel weltweit Temperatur Fischerei auf Fische, Klima Systemänderung Stichwort Nordsee Nordsee weltweit Mittelmeer Mittelmeer Island Nordsee Nordsee Nordsee Ostsee Mittelmeer Nordsee weltweit weltweit Ostsee weltweit Ostsee Gebiet allgemein Temora Plankton Kabeljau Hering Hering Hering Hering Fisch Hering allgemein allgemein Monoporeia allgemein Plankton, Fisch Art Klimawandel und europäische Fischerei 1811 1896 2004 2003 2021 1812 1975 1738 1738 1783 1784 1784 1983 1983 205 277 1949 1986 McGoodwin 2007 Medina et al. 2002 Menard & Fromentin 1997 Menard et al. 1994 Miller & Munro 2004 Miller 2007 Mills & Mann 1985 Misund et al. 1998 Misund et al. 1998 Molinero et al. 2005a Molinero et al. 2005b Molinero et al. 2005b Molinero et al. 2007 Molinero et al. 2007 Möllmann et al. 2000 Möllmann et al. 2003 Moore et al. 1997 Muus & Nielsen 1999 PE 379.208 2008 Lfd. Nr. MCCIP 2006 Verweis Fisch Fisch Sonstiges Sonstiges Sonstiges Wasser Sonstiges Sonstiges Sonstiges Fisch Fisch Fisch Fisch Fisch Sonstiges Sonstiges Fisch Fisch Sonstiges Bezug zu Klimawandel und europäische Fischerei 4.2.1 4.2.7 4.3.6 4.3.1 4.3.6 4.1.4 4.3.6 4.1.3 4.3.6 4.2.5 4.2.2 4.2.1 4.3.5 4.3.5 4.3.6 4.3.6 4.2.5 Schlussf. 4.3.8 Unterkapitel Änderung temporale Schwankungen beim Vorkommen von Salpen Verändertes Nahrungsnetz ökologische Auswirkungen des Klimawandels 108 Nordostatlantik Süßwasserökosysteme, Mittelatlantik Temperaturanstieg, stärkere Mobilität von Kontaminanten Verbreitung Ostsee Ostsee Mittelmeer Mittelmeer Mittelmeer Mittelmeer Mittelmeer Nordsee Nordsee Pazifik Pazifik Mittelmeer Mittelmeer Mittelmeer Alaska weltweit Gebiet veränderte Ruderfußkrebsabundanz Temperatur Phänologie und Produktivität Änderung Änderung Änderung Änderung Änderung Bewirtschaftung veränderte Ruderfußkrebsabundanz NAO, Plankton NAO, Plankton Phänologie des Zooplanktons Temperatur Temperatur Auswirkung der Temperatur auf Populationen, ökologische Auswirkungen Klimavariabilität Bewirtschaftung Änderung Klimavariabilität Anmerkung Variabilität temporale Schwankungen beim Vorkommen von Doliolida Gonadenreifung, Temperatur Klima Auswirkungen des Klimas auf die Aquakultur Stichwort allgemein Plankton Pseudocalanus, Acartia allgemein Plankton Plankton Plankton Plankton Hering Hering Thunfisch Lachs Salpen Thunfisch allgemein allgemein Art 1913 1806 1950 221 2052 1736 1731 1951 1830 1820 2051 1348 2062 1866 1718 1718 1952 2060 1889 Naylor et al. 2000 Nehring 2003 O'Brien et al. 2000 Olsen et al. 2004 Orr et al. 2005 Ottersen et al. 2001 Parmesan & Yohe 2003 Parmesan 2006 Parmesan et al. 2005 Parsons & Lear 2001 Patz et al. 2005 Pauly et al. 2005 Pawlak et al. 2007 Pearson & Dawson 2003 Perry et al. 2005 Perry et al. 2005 Persson 1986 Peterson et al. 2006 Planque & Fox 1998 PE 379.208 Lfd. Nr. Verweis Fisch Wasser Fisch Fisch Fisch Sonstiges Wasser Fisch Wasser Wasser 4.1.6 3.1.1 4.2.1 4.2.3 4.2.2 4.2.3 3.1.5 4.3.4 3.1.1 4.1.6 Schlussf. allgemeine r Zshg. 4.2.2 Fisch, Sonstiges 4.2.2 4.1.6 Fisch, Sonstiges Sonstiges 3.1.1 4.3.4 4.3.4 4.3.2 4.3.8 Unterkapitel Wasser Fisch Fisch Sonstiges Sonstiges Bezug zu Temperatur Salinität Temperatur Klima Klima bioklimatische Hülle Temperatur, Salinität Fischerei Temperatur NAO Klima 109 Phänologie, Größe, Verbreitung Metaanalyse: Verschiebung bei indifferenten Arten NAO, ökologische Auswirkungen Versauerung Arktis Nordatlantik höherer Stoffwechselumsatz, geringere Rekrutierung stehende Gewässer Nordsee Nordsee weltweit Rückgang Folgen für die Gemeinschaft Änderung Änderung Änderung der Artenverbreitung Ostsee weltweit negative Auswirkungen auf Fische Erderwärmung Europa Nordatlantik weltweit weltweit weltweit weltweit weltweit Hitzewelle, Klimawandel Auswirkungen auf die Biosphäre Änderung Änderung Nordwärtsverschiebung der Verbreitung, früheres Frühjahr Zunahme Nordatlantik Befischung verringert Größe und Fruchtbarkeit Nordsee, Ostsee weltweit Gebiet Nordsee anthropogene Ursachen Anmerkung Auswirkungen der Temperatur auf die Rekrutierung beim Kabeljau Invasion Auswirkungen der Aquakultur auf die Eutrophierung Stichwort Kabeljau Plötze, Barsch allgemein allgemein allgemein Fisch Fisch allgemein allgemein allgemein allgemein Kabeljau Kabeljau allgemein allgemein Art Klimawandel und europäische Fischerei 1816 1832 2018 1899 1953 1954 1861 1744 1822 1955 1892 1956 1064 1957 2053 1985 1728 Poloczanska et al. 2004 Polovina 2005 Pope & Symes Pörtner & Knust 2007 Pörtner 2002 Pörtner et al. 2004 Pörtner et al. 2005 Rahmstorf 2000 Rahmstorf 2002 Raper et al. 1997 Ravier & Fromentin 2004 Regier et al.1990 Renz & Hirche 2006 Reynolds & Casterlin 1977 Riebesell et al. 2000 Rixen et al. 2005 Rodwell et al. 1999 PE 379.208 1978 Lfd. Nr. Planque & Fredou 1999 Verweis Wasser Sonstiges Wasser Fisch Sonstiges Fisch Fisch Wasser Wasser Wasser Fisch Fisch 4.1.1 4.3.6 3.1.1 4.2.1 4.3.1 4.2.1 4.2.5 4.1.1 4.1.2 4.1.1 4.2.1 4.2.7 4.2.7 4.2.1 Fisch, Sonstiges Fisch 4.3.5 4.3.4 4.3.5 4.3.4 Unterkapitel Fisch Fisch Fisch Fisch Bezug zu Klimawandel und europäische Fischerei NAO Auswirkungen der NAO Versauerung Temperatur Salinität Temperatur Temperatur 110 Anstieg der Treibhausgasemissionen Klimawandel Temperatur der Meeresoberfläche Zunahme Verhalten Physiologie Wahl der Muttergewässer in Abhängigkeit von den äußeren Gegebenheiten Änderung der Zirkulation Nordatlantik Mittelmeer weltweit weltweit Ostsee weltweit Atlantik Britische Inseln Nordatlantik Nordatlantik Änderung of thermohaline Zirkulation Klima weltweit Schlussfolgerungen kombinierte Auswirkungen von Temperatur, pH-Wert, Physiologie weltweit nördliche Breitengrade Atmung Stoffwechsel Auswirkungen auf die Verbreitung Nordpazifik Systemänderung aufgrund des Einflusses auf die stärksten natürlichen Feinde weltweit Nordsee Nordatlantik, Nordsee Gebiet Änderung Anmerkung Temperatur Temperatur Temperatur und Stoffwechsel Ökosystemansatz für das Fischereiwesen Klimawandel Fischerei Zusammenhang zwischen Temperatur und Rekrutierung Stichwort Phytoplankton allgemein Pseudocalanus allgemein Thunfisch allgemein allgemein allgemein allgemein allgemein Sandaal Kabeljau Art 1918 1960 1908 1737 1862 1862 2058 1808 1903 1961 1900 1962 1963 1888 1745 2056 1819 1964 1834 1965 Rose 2005 Ruddiman & Thomson 2001 Sabine et al. 2004 Sarmiento et al. 2004 Scavia et al. 2002 Scavia et al. 2002 Schär et al. 2004 Schrank 2007 Send et al. 1999 Sharp 2003 Sheppard 2004 Siegenthaler et al. 2005 Sims et al.2006 Sirabella et al. 2001 Sissener & Bjoerndal 2005 Smetacek & Nicol 2005 Somarakis et al. 2006 Southward et al. 1988 Southward et al. 1995 Spahni et al. 2005 PE 379.208 1959 Lfd. Nr. 1598 Root et al. 2003 Rogers 1984 Verweis Wasser Fisch Fisch Fisch Sonstiges Fisch Fisch Fisch Wasser Sonstiges Fisch Wasser Fisch Wasser Wasser Wasser Sonstiges Wasser Wasser Fisch Fisch Wasser Bezug zu 4.1.1 4.2.2 4.2.1 4.3.3 3.3.3 4.2.5 4.1.6 4.2.1 4.1.1 4.1.5 4.1.1 4.1.6 Schlussf. 3.1.1 4.1.3 4.1.2 4.3.3 4.1.1 4.1.1 4.2.2 4.2.2 Unterkapitel 4.1.1 111 Anstieg der Treibhausgasemissionen Temperatur Temperatur Klima Produktivität in hohen Breitengraden Klima NAO Temperatur Anstieg der Treibhausgasemissionen Temperaturanstieg in der Nordsee Klimawandel NAO Klima Temperatur Klimawandel Klimawandel Klimawandel Versauerung Anstieg der Treibhausgasemissionen Verschiebung bei der Verbreitung Temperatur NAO Stichwort Erderwärmung Anstieg Fischgemeinschaft Änderung Zunahme Änderung Kabeljaurekrutierung Physiologie Erderwärmung Zeitreihe 1871-2099 regionale Fischereien Temperatur und Salinität Änderung Hitzewelle, Klimawandel weltweit Nordsee Nordsee Mittelmeer Arktis, Antarktis Nordsee Nordatlantik weltweit weltweit weltweit Mittelmeer Arktis Europa weltweit weltweit Änderung wichtiger ozeanographischer Merkmale weltweit weltweit Nordatlantik weltweit weltweit Schichtung Gebiet Nordatlantik Produktivität Zunahme Erderwärmung Auswirkungen der Temperatur Änderung der Verbreitung Index Anmerkung allgemein Hering, Pilchard Sardine Hering Kabeljau Katzenhai allgemein allgemein Nährstoffe Phytoplankton Fisch allgemein Art Klimawandel und europäische Fischerei 1809 1809 1729 1742 2059 1966 1907 1749 1755 1984 1992 1909 1967 1968 1991 1976 1739 1969 1743 1970 Stenevik & Sundby 2007 Stenevik & Sundby 2007 Stenseth et al. 2002 Stenseth et al. 2003 Stott et al. 2004 The Royal Society 2005 Theocharis et al. 1999 Tietenberg 2003 Tonn 1990 Trigo et al. 2000 Tsimplis & Josey 2001 Tsimplis et al. 2006 Turrell et al. 1992 Veizer, 2005 Vignudelli et al. 1999 Vilhjalmsson 1997 Visbeck et al. 2001 Vitousek, 1994 Walther et al. 2002 Walther et al. 2005 PE 379.208 1898 Lfd. Nr. Steele 2004 Verweis Wasser Fisch Wasser Wasser Fisch Sonstiges Wasser Wasser Wasser Sonstiges Sonstiges Fisch Fisch Wasser Wasser Wasser Wasser Sonstiges Fisch Fisch Fisch Bezug zu Klimawandel und europäische Fischerei 4.1.1 Schlussf. 4.1.1 4.1.1 4.2.5 4.3.6 4.1.1 4.1.5 4.1.5 4.3.6 4.3.6 4.2.5 4.3.5 4.1.5 4.1.1 3.1.1 4.1.1 4.3.3 4.3.8 4.2.4 4.2.6 Unterkapitel Klimawandel Klima Klimawandel Klima Klimawandel Zirkulation, NAO Klimawandel Salinität Klimawandel Zirkulation, NAO Anstieg der Salinität Klima Klima Temperatur, Salinität Versauerung der Meere Temperatur Klima Klimawandel Klima Klima Fischerei Stichwort 112 Änderung Änderung von Ökosystemen Änderung bei der NAO Migration Änderung Treibhausgasemissionen im Gegensatz zur verstärkten Sonneneinstrahlung Meeresspiegel Änderung Änderung Änderung Bildung von Tiefenwasser Rückgang der Kalzifizierung Hitzewelle, Klimawandel Änderung Phänologie und Produktivität Änderung Änderung Verschiebung des Ökosystems bzw. Systemänderung Anmerkung weltweit weltweit weltweit Nordatlantik Nordostatlantik Mittelmeer weltweit Nordsee Mittelmeer Mittelmeer Mittelmeer stehende Gewässer weltweit Mittelmeer weltweit Europa weltweit Nordatlantik und Pazifik Nordsee Nordsee weltweit Gebiet allgemein Hering allgemein allgemein Phytoplankton Klimaindizes allgemein allgemein allgemein Art 1770 2009 2009 1977 1989 267 1901 1971 1858 1831 Wang & Overgaard 2007 WBGU-Bericht 2006 WBGU-Bericht 2006 Weiss 1970 Whitehead et al.1988 Wieland et al. 1994 Wiltshire & Manly 2004 Winder & Schindler 2004 Wonham et al. 2000 Worm & Myers 2003 PE 379.208 Lfd. Nr. Verweis Fisch Fisch Wasser Sonstiges Fisch Fisch Sonstiges Sonstiges 4.3.4 4.3.2 4.1.1 4.1.5 4.2.6 4.2.1 4.2.7 4.3.5 4.1.2 4.2.7 Fisch, Sonstiges Wasser Unterkapitel Bezug zu 113 Rückgang der Bestände invasive Arten Einführung neuer Fischarten über das Ballastwasser Überfischung Süßwasser Nordseewasser Schwimmfähigkeit des Laiches Auswirkungen der Temperatur mögliche Änderung Verschlechterung der Temperatur, thermische Grenzwerte, Sauerstofftransportsystem Anmerkung Temperatur Temperaturanstieg Salinität und Sauerstoff Verbreitung Sauerstofflöslichkeit Auswirkungen der Überfischung thermohaline Zirkulation Temperatur und physiologische Mechanismen Stichwort weltweit weltweit weltweit Nordsee Ostsee Mittelmeer weltweit Nordatlantik Nordatlantik weltweit Gebiet allgemein allgemein Kabeljau Sardelle allgemein allgemein Art Klimawandel und europäische Fischerei PE 379.208 2009 2009 1977 1989 267 1901 1971 1858 1831 WBGU Report 2006 WBGU Report 2006 Weiss 1970 Whitehead et al. 1988 Wieland et al . 1994 Wiltshire & Manly 2004 Winder & Schindler 2004 Wonham et al. 2000 Worm & Myers 2003 water other other fish fish others water fish fish 1970 water 1770 Fish, Other Walther et al. 2005 Wang & Overgaard 2007 other fish Water water Fish 1991 1976 1739 1969 1743 1966 water 60 1907 water 60 1749 Fish 1755 Fish 1984 other 1992 other 1909 water 1967 water 1968 water serial related to 4.1.2 4.3.5 4.2.7 4.2.1 4.2.6 4.1.5 4.1.1 4.3.2 4.3.4 4.1.1 4.2.7 4.3.6 4.2.5 4.1.1 4.1.1 conclusion subchapter 4.1.1 4.1.5 4.3.5 4.2.5 4.3.6 4.3.6 4.1.5 4.1.5 4.1.1 114 thermohaline circulation effects of overfishing oxygen solubility distribution salinity and oxygen temperature increase temperature Fish introduction via ballast water overfishing climate change temperature and physiological mechanisms circulation, NAO climate change climate climate change climate ocean acidification temperature and salinity climate climate salinity increase circulation, NAO climate change salinity climate change keyword Dokumentierung der Ergebnisse der Literaturauswertung Vignudelli et al. 1999 Vilhjalmsson 1997 Visbeck et al. 2001 Vitousek, 1994 Walther et al. 2002 Tonn 1990 Trigo et al. 2000 Tsimplis & Josey 2001 Tsimplis et al . 2006 Turrell et al. 1992 Veizer, 2005 The Royal Society 2005 Reference Tabelle 6: Klimawandel und europäische Fischerei Area buoyancy of eggs North sea water freshwater Invasive species stock decline temperature effect possible change pejus temperature thermal limits, oxygen transport system change migration change in NAO ecosystem change change North Atlantic North Atlantic global Mediterranean Baltic Sea North Sea global global global global global Mediterranean North East Atlantic North Atlantic global global global mediterranean global lake Mediterranean change Mediterranean sea level mediterranean North Sea greenhouse gas vs. solar increase global decrease in calcification deep water formation change change remark general general anchovy cod general general general herring general feneral phytoplankton Species Klimawandel und europäische Fischerei Bibliography 1. ACIA (2005) Artic Climate Impact Assessment. Cambridge University press 1042 pp. 2. Alheit, J, Hagen, E (1997) Long-term climate forcing of European herring and sardine populations. Fish Oceanogr 6(2): 130-139. 3. Alheit, J, Möllmann, C, Dutz, J, Kornilovs, G, Loewe, P, Mohrholz, V, Wasmund, N (2005) Synchronous ecological regime shifts in the central Baltic and the North Sea in the late 1980s. ICES Journ Mar Sci (62): 1205-1215. 4. Araujo, MB, New, M (2007) Ensemble forecasting of species distributions. Trends Ecol Evol 22(1): 42-47. 5. Araujo, MB, Pearson, RG, Thuiller, W, Erhard, M (2005) Validation of species-climate impact models under climate change. Glob Change Biol 11(9): 504-1513. 6. Arnell, NW (1998) Climate change and water resources in Britain. Clim Change 39(1): 83-110. 7. Arnott, SA, Ruxton, GD (2002) Sandeel recruitment in the North Sea: Demographic, climatic and trophic effects. Mar Ecol Prog Ser 238: 199-210. 8. Astraldi, M-F, Bianchi, CN, Gasparini, GP, Morri, C (1995) Climatic fluctuations, current variability and marine species distribution: A case study in the Ligurian Sea (north-west Mediterranean). Oceanol Acta 18(2): 139-149. 9. Attrill, MJ, Power, M (2002) Climatic influence on a marine fish assemblage. Nature 417(6886): 275-278. 10. Attrill, MJ, Wright, J, Edwards, M (2007) Climate-related increases in jellyfish frequency suggest a more gelatinous future for the North Sea. Limnol Oceanogr 52(1): 480-485. 11. BACC (2006) Assessment of climate change for the Baltic Sea basin. 16 pp. 12. Basilone, G, Guisande, C, Patti, B, Mazzola, S, Cuttitta, A, Bonanno, A, Vergara, AR, Maneiro, I (2006) Effect of habitat conditions on reproduction of the European anchovy (Engraulis encrasicolus) in the Strait of Sicily. Fish Oceanogr 15(4): 271-280. 13. Beare, DJ, Batten, S, Edwards, M, Reid, DG (2002) Prevalence of boreal Atlantic, temperate Atlantic and neritic zooplankton in the North Sea between 1958 and 1998 in relation to temperature, salinity, stratification intensity and Atlantic inflow. J Sea Res 48(1): 29-49. 14. Beare, DJ, Burns, F, Greig, A, Jones, EG, Peach, K, Kienzle, M, McKenzie, E, Reid, DG (2004a) Long-term increases in prevalence of North Sea fishes having southern biogeographic affinities. Mar Ecol Prog Ser 284: 269-278. 15. Beare, D, Burns, F, Jones, E, Peach, K, Portilla, E, Greig, T, Mckenzie, E, Reid, D (2004b) An increase in the abundance of anchovies and sardines in the north-western North Sea since 1995. Global Change Biol 10(7) 1209-1213. 16. Beaugrand, G (2004) The North Sea regime shift: evidence, causes, mechanisms and consequences. Prog Oceanogr 60(2-4): 245-262. 17. Beaugrand, G, Reid, PC, Ibanez, F, Lindley, JA, Edwards, M (2002) Reorganization of North Atlantic marine copepod biodiversity and climate. Science 296(5573): 1692-1694. 18. Beaugrand, G, Brander, KM, Lindley, JA, Souissi, S, Reid, PC (2003) Plankton effect on cod recruitment in the North Sea. Nature 426(6967): 661-664. 115 PE 379.208 Klimawandel und europäische Fischerei 19. Behrenfeld, MJ, et al. (9 co-authors) (2006) Climate-driven trends in contemporary ocean productivuíty. Nature 444: 752-755. 20. Bethoux, JP, Gentili, B, Morin, P, Nicolas, E, Pierre, C, Ruiz-Pino, D (1999) The Mediterranean Sea: a miniature ocean for climatic and environmental studies and a key for the climatic functioning of the North Atlantic. Prog Oceanogr 44(1-3): 131-146. 21. Bethoux, JP, Morin, P, Ruiz-Pino, DP (2002) Temporal trends in nutrient ratios: chemical evidence of Mediterranean ecosystem changes driven by human activity. Deep-Sea Res (II Top Stud Oceanogr) 49(11): 2007-2016. 22. Bigelow, HB (1928) Exploration of the waters of the gulf of Maine. Geogr Rev 18(2): 232-260. 23. Birkeland, C, Dayton, PK (2005) The importance in fishery management of leaving the big ones. Trends Ecol Evol 20(7): 356-358. 24. Björnsson B, Steinarsson A (2002) The food-unlimited growth rate of Atlantic cod (Gadus morhua). Can J Fish Aquat Sci 59: 494-502. 25. Bombace, G.B. (2001) Influence of climatic changes on stocks, fish species and marine ecosystems in the Mediterranean Sea. Archo Oceanogr Limnol 22: 67-72. 26. Brander, KM (1994) The location and timing of cod spawning around the British-Isles. ICES J Mar Sci 51(1): 71-89. 27. Brander, KM (1995) The effect of temperature on growth of Atlantic cod (Gadus morhua L.). ICES J Mar Sci 52(1): 1-10. 28. Brander, KM (2006) Assessment of possible impacts of climate change on fisheries 29. Brander, KM, Dickson, RR, Edwards, M (2003) Use of Continuous Plankton Recorder information in support of marine management: applications in fisheries, environmental protection, and in the study of ecosystem response to environmental change. Prog Oceanogr 58(2-4): 175-191. 30. Brett, JR (1956) Some principles in the thermal requirements of fishes. Q Rev Biol 31(2): 75-87. 31. Browman, HI, Hutchings, JA, Conover, DO, Stokes, K, Law, R, Walters, C (2000) 'Evolution' of fisheries science. Mar Ecol Prog Ser 208: 301-309. 32. Brown, BR (2003) Neurophysiology - Sensing temperature without ion channels. Nature 421(6922): 495. 33. Bryden, HL, Longworth, HR, Cunningham, SA (2005) Slowing of the Atlantic meridional overturning circulation at 25 degree N. Nature 438(7068): 655-657. 34. Buecher, E (1999) Appearance of Chelophyes appendiculata and Abylopsis tetragona (Cnidaria, Siphonophora) in the Bay of Villefranche, northwestern Mediterranean. J Sea Res 41(4): 295-307. 35. Burton, DT (1979) Ventilation frequency compensation responses of three eurythermal estuarine fish exposed to moderate temperature increases. J Fish Biol 15(5): 589–600. 36. Caldeira, K, Wickett, ME (2003) Oceanography - Anthropogenic carbon and ocean pH Nature 425(6956): 365. 37. Caldeira, K, Wickett, ME (2005) Ocean model predictions of chemistry changes from carbon dioxide emissions to the atmosphere and ocean. J Geophys Res 110(C9): 12 pp. PE 379.208 116 Klimawandel und europäische Fischerei 38. Carotenuto Y, Ianora A, Di Pinto M, Sarno D, Miralto A (2006) Annual cycle of early developmental stage survival and recruitment in the copepods Temora stylifera and Centropages typicus. Mar Ecol Prog Ser 314: 227–238. 39. Casini, M, Cardinale, M, Hjelm, J (2006) Inter-annual variation in herring, Clupea harengus, and sprat, Sprattus sprattus, condition in the central Baltic Sea: what gives the tune? Oikos 112(3): 638-650. 40. Cayan, DR (1992) Latent and sensible heat flux anomalies over the northern oceans: Driving the sea surface temperature. J Phys Oceanogr 22(8): 859-881. 41. Ciais et al. (32 co-authors) (2005) Europe-wide reduction in primary productivity caused by the heat and drought in 2003: Nature 437: 529-533. 42. Chavez, FP, Ryan, J, Lluch-Cota, SE, Niquen, MC (2003) From anchovies to sardines and back: Multidecadal changes in the Pacific Ocean. Science 299: 217-221. 43. Ciannelli, L, Bailey, KM, Chan, K-S, Belgrano, A, Stenseth, NC (2005) Climate change causing phase transitions of walleye pollock (Theragra chalcogramma) recruitment dynamics. Proc R Soc Lond, Ser B: Biol Sci 272(1573): 1735-1743. 44. Clark, RA, Fox, CJ, Viner, D, Livermore, M (2003) North Sea cod and climate change modelling the effects of temperature on population dynamics. Global Change Biol 9(11): 1669-1680. 45. Clarke, A (1993) Temperature and extinction in the sea: A physiologist's view. Paleobiol 19(4): 499-518. 46. Claussen, M, Ganopolski, A, Brovkin, V, Gerstengarbe, FW, Werner, P (2003) Simulated global-scale response of the climate system to Dansgaard /Oeschger and Heinrich events. Clim Dyn 21(5-6): 361-370. 47. Collie, JS, Richardson, K, Steele, JH (2004) Regime shifts: can ecological theory illuminate the mechanisms? Prog Oceanogr 60(2-4): 281-302. 48. Cook, RM, Sinclair, A, Stefansson, G (1997) Potential collapse of North Sea cod stocks. Nature 385(6616): 521-522. 49. Coutant, CC (1987) Thermal preference: When does an asset become a liability? Environ Biol Fish 18(3): 161-172. 50. Curry, R, Mauritzen, C (2005) Dilution of the Northern North Atlantic Ocean in Recent Decades. Science 308(5729): 1772-1774. 51. Cushing, DH (1984) The gadoid outburst in the North Sea. ICES J Mar Sci 41(2): 159166. 52. Cushing, DH (1990) Plankton production and year-class strength in fish populations: An update of the match/mismatch hypothesis. Adv Mar Biol 26: 249-294. 53. Davis AJ, Jenkinson, LS, Lawton, JH, Shorrocks, B, Woods, S (1998) Making mistakes when predicting shifts in species in response to global warming. Nature 391: 783-786. 54. Dickson, RR, Brander, KM (1993) Effects of a changing windfield on cod stocks of the North Atlantic. Fish Oceanogr 2(3-4): 124-153. 55. Drinkwater, KF (2005) The response of Atlantic cod (Gadus morhua) to future climate change. ICES J Mar Sci 62(7): 1327-1337. 56. Drinkwater, KF (2006) The regime shift of the 1920s and 1930s in the North Atlantic. Prog Oceanogr 68(2-4): 134-151. 117 PE 379.208 Klimawandel und europäische Fischerei 57. Dulcic, J, Grbec, B, Lipej, L (1999) Information on the Adriatic ichthyofauna - effect of water warming? Acta Adriat 40(2): 33-43. 58. Dutil, J, Brander, K (2003) Comparing productivity of North Atlantic cod (Gadus morhua) stocks and limits to growth production. Fish Oceanogr 12(4-5): 502-512. 59. Ecological Society of America (ESA) (1998) Ecosystem management for sustainable marine fisheries. Ecol Appl 8(Suppl. 1): 174 pp. 60. Edwards, M, Richardson, AJ (2004) Impact of climate change on marine pelagic phenology and trophic mismatch. Nature 430(7002): 881-884. 61. Edwards, M, John, AWG, Johns, DG, Reid, PC (2001) Case history and persistence of the non-indigenous diatom Coscinodiscus wailesii in the north-east Atlantic. J Mar Biol Assoc UK 81(2): 207-211. 62. Edwards, M, Beaugrand, G, Reid, PC, Rowden, AA, Jones, MB (2002) Ocean climate anomalies and the ecology of the North Sea. Mar Ecol Prog Ser 239: 1-10. 63. Eisenreich, SJ (2005) Climate Change and the European Water Dimension. European Reports 21553: 253 pp. 64. Engelhard, GH, Heino, M (2004) Dynamics in frequency of skipped reproduction in Norwegian spring-spawning herring. ICES Mar Sci Sympos K:43. 65. Engelhard, GH, Heino, M (2006) Climate change and condition of herring (Clupea harengus) explain long-term trends in extent of skipped reproduction. Oecologia 149: 593603. 66. European Commission (2006) Facts and figures on the CFP. Luxembourg: Office for Official Publications of the European Communities, 37pp. 67. Feely, RA, Sabine, CL, Lee, K, Berelson, W, Kleypas, J, Fabry, VJ, Millero, FJ (2004) Impact of Anthropogenic CO2 on the CaCO3 System in the Oceans. Science 305(5682): 362-366. 68. Ferguson, RG (1958) The prefered temperature of fish and their midsummer distribution in temperate lakes and streams. J Fish Res Bd Canada 15: 607-624. 69. Fernandez J, Saenz J, Zorita E (2003) Analysis of wintertime atmospheric moisture transport and its variability over southern Europe in the NCEP Reanalysis. Climate Research 23: 195-215. 70. Fox, CJ, Planque, BP, Darby, CD (2000) Synchrony in the recruitment time-series of plaice (Pleuronectes platessa L) around the United Kingdom and the influence of sea temperature. J Sea Res 44(1-2): 159-168. 71. Francour, P, Boudouresque, CF, Harmelin, JG, Harmelin-Vivien, ML, Quignard, JP (1994) Are the Mediterranean waters becoming warmer? Information from biological indicators. Mar Pollut Bull 28(9): 523-526. 72. Frank, Kenneth T, Petrie, Brian, Choi, Jae S, Leggett, William C (2005) Trophic Cascades in a Formerly Cod-Dominated Ecosystem. Science 308: 1621-1623. 73. Froese, R, Pauly, D (eds) (2007) FishBase. www.fishbase.org. 74. Fromentin, J-M, Planque, B ( 1996) Calanus and environment in the eastern North Atlantic. 2. Influence of the North Atlantic Oscillation on C. finmarchicus and C. helgolandicus. Mar Ecol Prog Ser 134(1-3): 111-118. PE 379.208 118 Klimawandel und europäische Fischerei 75. Fry, FEJ (1947) Effects of the environment on animal activity. Publ Ont Fish Res Lab 68: 1-62. 76. Fry FEJ (1971) The effect of environmental factors on the physiology of fish. Fish Phys 6: 1-98. 77. Garcia, SM, Zerbi, A, Aliaume, C, Do Chi, T, Lasserre, G (2003) The ecosystem approach to fisheries. Issues, terminology, principles, institutional foundations, implementation and outlook. FAO Fish Tech Pap 443, 71 pp. 78. Gaylord, B, Gaines, SD (2000) Temperature or transport? range limits in marine species mediated solely by flow. Am Nat 155(6): 769-789. 79. Genner, MJ, Sims, DW, Wearmouth, VJ, Southall, EJ, Southward, AJ, Henderson, PA, Hawkins, SJ (2004) Regional climatic warming drives long-term community changes of marine fish. Proc R Soc Lond, Ser B: Biol Sci 271(1539): 655-661. 80. Goffart, A, Hecq, J-H, Legendre, L (2002) Changes in the development of the winterspring phytoplankton bloom in the Bay of Calvi (NW Mediterranean) over the last two decades: a response to changing climate? Mar Ecol Prog Ser 236: 45-60. 81. Gomez F, Gorsky G (2003) Microoplankton cycle in the Northwestern Mediterranean. J Plankton Res 25: 323-329. 82. Gomis, D, Tsimplis, MN, Martin-Miguez, B, Ratsimandresy, AW, Garcia-Lafuente, J, Josey, SA (2006) Mediterranean Sea level and barotropic flow through the Strait of Gibraltar for the period 1958-2001 and reconstructed since 1659. J Geophys Res 111 C11. 83. Goren, M, Galil, BS (2005) A review of changes in the fish assemblages of Levantine inland and marine ecosystems following the introduction of non-native fishes. J Appl Ichthyol 21(4): 364-370. 84. Goy, J, Morand, P, Etienne, M (1989) Long-term fluctuations of Pelagia noctiluca (Cnidaria, Scyphomedusa) in the Western Mediterranean Sea. Prediction by climatic variables. Deep-Sea Res I 36(2A): 269-279. 85. Greene, CH, Pershing, AJ (2007) Climate drives sea change. Science 315: 1084-1085. 86. Guisande, C, Vergara, AR, Riveiro, I, Cabanas, JM (2004) Climate change and abundance of the Atlantic-Iberian sardine (Sardina pilchardus). Fish Oceanogr 13(2): 91-101. 87. Halsband-Lenk, C, Hirche, HJ (2001) Reproductive cycles of dominant calanoid copepods in the North Sea. Mar Ecol Prog Ser 209: 219-229. 88. Hannesson, R (2007) Geographical distribution of fish catches and temperature variations in the northeast Atlantic since 1945. Mar Pol 31(1): 32-39. 89. Hansen, VK (1960) Investigations on the quantitative and qualitative distribution of zooplankton in the southern part of the Norwegian Sea. Med. Daninarks Fiskeri-og Havundersogelser 2(23): 1-53. 90. Harley, CDG, Randall Hughes, A, Hultgren, KM, Miner, BG, Sorte, CJB, Thornber, CS, Rodriguez, LF, Tomanek, L, Williams, SL (2006) The impacts of climate change in coastal marine systems. Ecol Lett 9(2): 228-241. 91. Harrods, C, Graham, C, Mallela, J (2007) Climate change and the fishes of Britain and Ireland Briefing Paper 4: 1-52. 119 PE 379.208 Klimawandel und europäische Fischerei 92. Harvell, CD, Kim, K, Burkholder, JM, Colwell, RR, Epstein, PR, Grimes, DJ, Hofmann, EE, Lipp, EK, Osterhaus, ADME, Overstreet, RM, Porter, JW, Smith, GW, Vasta, GR (1999) Emerging marine diseases-climate links and anthropogenic factors. Science 285: 1505-1510. 93. Harwood, J, Stokes, K (2003) Coping with uncertainty in ecological advice: lessons from fisheries. Trends Ecol Evol 18(12): 617-622. 94. Hauser, L, Adcock, GJ, Smith, PJ, Ramirez, JHB, Carvalho, GR (2002) Loss of microsatellite diversity and low effective population size in an overexploited population of New Zealand snapper (Pagrus auratus). Proc Natl Acad Sci USA 99(18): 11742-11747. 95. Hawkins, SJ, Southward, AJ, Genner, MJ (2003) Detection of environmental change in a marine ecosystem - evidence from the western English Channel. Sci Total Environ 310(13): 245-256. 96. Hays, GC, Richardson, AJ, Robinson, C (2005) Climate change and marine plankton. Trends Ecol Evol 20(6): 337-344. 97. Hänninen, J, Vuorinen, I, Hjelt, P (2000) Climatic factors in the Atlantic control the oceanographic and ecological changes in the Baltic Sea. Limnol Oceanogr 45(3): 703-710. 98. Helmuth, B, Mieszkowska, N, Moore, P, Hawkins, SJ (2006) Living on the Edge of Two Changing Worlds: Forecasting the Responses of Rocky Intertidal Ecosystems to Climate Change. Annu Rev Ecol, Evol Syst 37: 373-404. 99. Henderson, PA (1998) On the variation in dab Limanda limanda recruitment: a zoogeographic study. J Sea Res 40(1-2): 131-142. 100. Herrick, J, Samuel. F., Norton, JG, Mason, JE, Bessey, C (2007) Management application of an empirical model of sardine-climate regime shifts. Mar Pol 31(1): 71-80. 101. Hjermann, DO, Bogstad, B, Eikeset, AM, Ottersen, G, Gjosaeter, H, Stenseth, NC (2007) Food web dynamics affect Northeast Arctic cod recruitment. Proc R Soc B-Biol Sci 274(1610): 661-669. 102. Hjort, J. (1914) Fluctuations in the great fisheries of Northern Europe - viewed in the light of biological research. Rapp Proc Verb Cons Int Expl Mer 20: 1-288. 103. Hochachka, PW, Somero, GN (2002) Biochemical Adaptation: Mechanism and process in physiological evolution. Oxford University Press, New York. 104. Hoepffner, N et al. (2006) Marine and coastal dimension of climate change in Europe. A report to the Europena Water Directors. 1-100. 105. Holbrook, SJ, Schmitt, RJ, Stephens, JSJ (1997) Changes in an assemblage of temperate reef fishes associated with a climate shift. Ecol Appl 7(4): 1299-1310. 106. Houghton, JT, Ding, Y, Griggs, DJ, Noguer, M, van der Linden, PJ, Dai, X, Maskell, K, Johnson, CA (2001) Climate Change 2001: the Scientific Basis. Contributions of Working Group 1 to the third assessment report of the Intergovernmental Panel on Climate Change. 107. Hsieh, C-H, Reiss, CS, Hunter, JR, Beddington, JR, May, RM, Sugihara, G (2006) Fishing elevates variability in the abundance of exploited species. Nature 443(7113): 859-862. 108. Hulme, M, Jenkins, GJ, Xianfu, L, Turpenny, JR, Mitchell, TD, Jones, GR, Lowe, J, Murphy, JM, Hassell, D, Boorman, P, McDonald, R, Hill, S (2002) Climate change scenarios for ther United Kingdom. The UKCIP02 Scientific Report. PE 379.208 120 Klimawandel und europäische Fischerei 109. Hurrell JW (1995) Decadal trends in the North Atlantic Oscillation: regional temperatures and precipitations. Science 269: 676-679. 110. Hurrell, JW, Dickson, RR (2004) Climate variability over the North Atlantic. Marine Ecosystems and Climate Variation: 15-32. 111. Hurrell, JW, Kushnir, Y, Visbeck, M (2001) Perspectives on Climate: The North Atlantic Oscillation. Science 291(5504): 603-605. 112. Hurrell, JW, Visbeck, M, Busalacchi, A, Clarke, RA, Delworth, TL, Dickson, RR, Johns, WE, Koltermann, KP, Kushnir, Y, Marshall, D, Mauritzen, C, McCartney, MS, Piola, A, Reason, C, Reverdin, G, Schott, F, Sutton, R, Wainer, I, Wright, D (2006) Atlantic climate variability and predictability: A CLIVAR perspective. J Clim 19(20): 5100-5121. 113. Hutchings, JA (2000) Collapse and recovery of marine fishes. Nature 406(6798): 882-885. 114. Ianora A, Poulet S, Miralto A (2003) The effect of diatoms on copepod reproduction: a review. Phycologia 42: 351-363. 115. ICES (2006) ICES Working group on Oceanic Hydrography (ICES Cooperative Research Report 280) 53pp. 116. IPCC (2007) Climate Change 2007: The Physical Science Basis: Summary for Policymakers. 1-21. 117. Iversen, SA, Skogen, MD, Svendsen, E (2002) Availability of horse mackerel (Trachurus trachurus) in the north-eastern North Sea, predicted by the transport of Atlantic water. Fish Oceanogr 11(4): 245-250. 118. James, A (1988) Are clupeid microphagists herbivorous or omnivorous - a review of the diets of some commercially important clupeids. South Afr J Mar Sci 7: 161-177. 119. Javidpour, J, Sommer, U, Shiganova T (2006) First record of Mnemiopsis leidyi A. Agassiz 1865 in the Baltic Sea. Aquat Invas 1(4): 299-302. 120. Jensen, AJ (2003) Atlantic salmon (Salmo salar) in the regulated River Alta: Effects of altered water temperature on parr growth. River Res Appl 19(7): 733-747. 121. Jobling, M (1981) Temperature tolerance and the final preferendum - rapid methods for the assessment of optimum growth temperatures. J Fish Biol 19(4): 439-455. 122. Jobling, M (1997) Temperature and growth: Modulation of growth rate via temperature change. In: Global Warming: Implications For Freshwater And Marine Fish: 225-253. 123. Jones, PD, Jonsson, T, Wheeler, D (1997) Extension to the north atlantic oscillation using early instrumental pressure observations from Gibraltar and south-west Iceland. Int J Climatol 17(13): 1433-1450. 124. Jurado-Molina, J, Livingston, P (2002) Climate-forcing effects on trophically linked groundfish populations: implications for fisheries management. Can J Fish Aquat Sci 59(12): 1941-1951. 125. Karl, TR, Trenberth, KE (2003) Modern global climate change. Science 302: 1719-1723. 126. Köster, FW, Hinrichsen, H-H, Schnack, D, St. John, MA, MacKenzie, BR, Tomkiewicz, J, Möllmann, C, Kraus, G, Plikshs, M, Makarchouk, A, Aro, E (2003). Recruitment of Baltic cod and sprat stocks: identification of critical life stages and incorporation of environmental variability into stock-recruitment relationships. Sci Mar 67(Suppl. 1): 129154. 121 PE 379.208 Klimawandel und europäische Fischerei 127. Köster, FW, Möllmann, C, Hinrichsen, H-H, Wieland, K, Tomkiewicz, J, Kraus, G, Voss, R, Markarchouk, A, MacKenzie, BR, St.John, MA, Schnack, D, Rohlf, N, Linkowski, T, Beyer, JE (2005) Baltic cod recruitment - the impact of climate variability on key processes. ICES J Mar Sci 62(7): 1408-1425. 128. Kent, ML, Poppe, TT (1998) Diseases of Seawater Netpen-Reared Salmonid Fishes. Fisheries and Oceans Canada, Nanaimo, Canada 138 pp. 129. Kiehl, JT, Trenberth, KE (1997) Earth's annual global mean energy budget. Bull Am Met Soc 78(2): 197-208. 130. King, DA (2004) Climate change science: adapt, mitigate, or ignore? Science 303(5655): 176-177. 131. Kirby, RR, Johns, DG, Lindley, JA (2006) Fathers in hot water: rising sea temperatures and a Northeastern Atlantic pipefish baby boom. Biol Lett 2(4): 597-600. 132. Kirby, RR, Beaugrand, G, Lindley, JA, Richardson, AJ, Edwards, M, Reid, PC (2007) Climate effects and benthic-pelagic coupling in the North Sea. Mar Ecol Prog Ser 330: 3138. 133. Klyashtorin, LB (1998) Long-term climate change and main commercial fish production in the Atlantic and Pacific. Fish Res 37(1-3): 115-125. 134. Klyashtorin, LB (2001) Climate change and long-term fluctuations of commercial catches. The possibility of forecasting. FAO Fish Tech Rep. 135. Lehodey, P, Alheit, J, Barange, M, Baumgartner, T, Beaugrand, G, Drinkwater, K, Fromentin, J, Hare, SR, Ottersen, G, Perry, RI, Roy, C, van der Lingen, CD, Werner, F (2006) Climate Variability, Fish, and Fisheries. J Clim 19(20): 5009-5030. 136. Lehtonen, KK (1996) Ecophysiology of the benthic amphipod Monoporeia affinis in an open-sea area of the northern Baltic Sea: Seasonal variations in body composition, with bioenergetic considerations. Mar Ecol Prog Ser 143(1-3): 87-98. 137. Levermann, A, Griesel, A, Hofmann, M, Montoya, M, Rahmstorf, S (2005) Dynamic sea level changes following changes in the thermohaline circulation. Clim Dyn 24(4): 347354. 138. Levitus, S, Antonov, JI, Boyer, TP, Stephens, C (2000) Warming of the world ocean. Science 287(5461): 2225-2229.Lloret, J, Lleonart, J (2002) Recruitment dynamics of eight fishery species in the northwestern Mediterranean Sea. Sci Mar 66(1): 77-82. 139. Maes, J, Limburg, KE, Van de Putte, A, Ollevier, F (2005) A spatially explicit, individualbased model to assess the role of estuarine nurseries in the early life history of North Sea herring, Clupea harengus. Fish Oceanogr 14(1): 17-31. 140. Magnuson, JJ, Crowder, LB, Medvick, PA (1979) Temperature as an ecological resource. Int Comp Biol 19(1): 331-343. 141. Malacic, V, Celio, M, Cermelj, B, Bussani, A, Comici, C (2006) Interannual evolution of seasonal thermohaline properties in the Gulf of Trieste (northern Adriatic) 1991-2003. J Geophys Lett 111 C8. 142. Malzahn, AM, Aberle, N, Clemmesen, C, Boersma, M (2007) Nutrient limitation of primary producers affects fish condition. Limnol. & Oceangr. 52(5): 11pp. PE 379.208 122 Klimawandel und europäische Fischerei 143. Maravelias, CD, Reid, DG (1995) Relationship between herring (Clupea harengus, L.) distribution and sea surface salinity and temperature in the northern North Sea. Sci Mar 59(3-4): 327-343. 144. Marteinsdottir, G, Steinarsson, A (1998) Maternal influence on the size and viability of Iceland cod Gadus morhua eggs and larvae. J Fish Biol 52(6): 1241-1258. 145. Marty, J-C, Chiaverini, J (2002) Seasonal and interannual variations in phytoplankton production at DYFAMED time-series station, northwestern Mediterranean Sea. Deep-Sea Res (II Top Stud Oceanogr) 49(11): 2017-2030. 146. Mazza, J (1961) Remarques sur la répartition qualitative et quantitative des copépodes en Méditerrannée. Rapp. P. V. Réun. Comm. Int. Explor. Sci. Mer Médit 16: 157-164. 147. Möllmann, C, Kornilovs, G, Sidrevics, L (2000) Long-term dynamics of main mesozooplankton species in the central Baltic Sea. J Plankton Res 22(11): 2015-2038. 148. Möllmann, C, Kornilovs, G, Fetter, M, Köster, F, Hinrichsen, H (2003) The marine copepod, Pseudocalanus elongatus, as a mediator between climate variability and fisheries in the Central Baltic Sea. Fish Oceanogr 12(4-5): 360-368. 149. McCarty, JP (2001) Ecological consequences of recent climate change. Conserv Biol 15(2): 320-331. 150. MCCIP (2006) Marine climate change impacts, annual report card 2006 8pp. 151. McGoodwin, JR (2007) Effects of climatic variability on three fishing economies in highlatitude regions: Implications for fisheries policies. Mar Pol 31(1): 40-55. 152. Medina, A, Abascal, FJ, Megina, C, Garcia, A (2002) Stereological assessment of the reproductive status of female Atlantic northern bluefin tuna during migration to Mediterranean spawning grounds through the Strait of Gibraltar. J Fish Biol 60(1): 203217. 153. Menard, F, Fromentin, JM (1997) Temporal fluctuations of doliolid abundance in the Bay of Villefranche-sur-Mer (northwestern Mediterranean Sea) from 1967 to 1990. Oceanol Acta 20(5): 733-742. 154. Menard, F, Dallot, S, Thomas, G, Braconnot, JC (1994) Temporal fluctuations of two Mediterranean salp populations from 1967 to 1990. Analysis of the influence of environmental variables using a Markov chain model. Mar Ecol Prog Ser 104(1-2): 139152. 155. Miller, KA (2007) Climate variability and tropical tuna: Management challenges for highly migratory fish stocks. Mar Pol 31(1): 56-70. 156. Miller, KA, Munro GR (2004) Climate and Cooperation: A New Perspective on the Management of Shared Fish Stocks. Mar Res Econ 19(3): 367-393. 157. Mills, CA, Mann RHK (1985) Environmentally induced fluctuations in year -class strength and their implications for management. J Fish Biol 27(Suppl. A): 209-226. 158. Misund, OA, Vilhjalmsson, H, Jakupsstovu, SH, Rottingen, I, Belikov, S, Asthorsson, O, Blindheim, J, Jonsson, J, Krysov, A, Malmberg, SA, Sveinbjornsson, S (1998) Distribution, migration and abundance of Norwegian spring spawning herring in relation to the temperature and zooplankton biomass in the Norwegian Sea as recorded by coordinated surveys in Spring and Summer 1996. Sarsia 83(2): 117-127. 123 PE 379.208 Klimawandel und europäische Fischerei 159. Molinero, JC, Ibanez, F, Nival, P, Buecher, E, Souissi, S (2005a) North Atlantic climate and northwestern Mediterranean plankton variability. Limnol Oceanogr 50(4): 1213-1220. 160. Molinero, JC, Ibanez, F, Souissi, S, Chifflet, M, Nival, P (2005b) Phenological changes in the Northwestern Mediterranean copepods Centropages typicus and Temora stylifera linked to climate forcing. Oecologia 145(4): 640-649. 161. Moore, MV, Pace, ML, Mather, JR, Murdoch, PS, Howarth, RW, Folt, CL, Chen, CY, Hemond, HF, Flebbe, PA, Driscoll, CT (1997) Potential effects of climate change on freshwater ecosystems of the New England/Mid-Atlantic region. In: Special Issue: Freshwater Ecosystems And Climate Change: 925-947. 162. Muus, BJ, Nielsen JG (1999) Die Meeresfische Europas in Nordsee, Ostsee und Atlantik. Kosmos Verlag 336 pp. 163. Naylor, RL, Goldburg, RJ, Primavera, JH, Kautsky, N, Beveridge, MCM, Clay, J, Folke, C, Lubchenco, J, Mooney, H, Troell, M (2000) Effect of aquaculture on world fish supplies Nature 405(6790): 1017-1024. 164. Nehring, Stefan (2003) Alien species in the North Sea: invasion success and climate warming. Ocean Chall 13(3): 12-16. 165. O'Brien, CM, Fox, CJ, Planque, B, Casey, J (2000) Climate variability and North Sea cod Nature 404(6774): 142. 166. Olsen, EM, Heino, M, Lilly, GR, Morgan, MJ, Brattey, J, Ernande, B, Dieckmann, U (2004) Maturation trends indicative of rapid evolution preceded the collapse of northern cod. Nature 428(6986): 932-935. 167. Orr JC et al. (26 co-authors) (2005) Anthropogenic ocean acidification over the twentyfirst century and its impacts on calcifying organisms. Nature 437: 681-686. 168. Ottersen, G, Planque, B, Belgrano, A, Post, E, Reid, PC, Stenseth, NC (2001) Ecological effects of the North Atlantic Oscillation. Oecologia 128(1): 1-14. 169. Parmesan, C (2006) Ecological and evolutionary responses to recent climate change Annu Rev Ecol Evol Syst 37: 637-669. 170. Parmesan, C, Gaines, S, Gonzalez, L, Kaufman, DM, Kingsolver, J, Townsend Peterson, A, Sagarin, R (2005) Empirical perspectives on species borders: from traditional biogeography to global change. Oikos 108(1): 58-75. 171. Parmesan, C, Yohe, G (2003) A globally coherent fingerprint of climate change impacts across natural systems. Nature 421(6918): 37-42. 172. Parsons, LS, Lear, WH (2001) Climate variability and marine ecosystem impacts: a North Atlantic perspective. Prog Oceanogr 49(1-4): 167-188. 173. Patz, JA, Campbell-Lendrum, D, Holloway, T, Foley, JA (2005) Impact of regional climate change on human health. Nature 438: 310-317. 174. Pauly, D, Watson, R, Alder, J (2005) Global trends in world fisheries: impacts on marine ecosystems and food security. Phil Trans R Soc B 360: 5-12. 175. Pawlak, JF et al. (2007) Climate change in the Baltic Sea area. Draft HELCOM thematic assessment in 2007, 53 pp. 176. Pörtner, HO (2002) Physiological basis of temperature-dependent biogeography: tradeoffs in muscle design and performance in polar ectotherms. J Exp Biol 205(15): 22172230. PE 379.208 124 Klimawandel und europäische Fischerei 177. Pörtner H O, Knust R (2007) Climate change affects marine fishes through the oxygen limitation of thermal tolerance. Science 315: 95-97. 178. Pörtner, HO, Mark, FC, Bock, C (2004) Oxygen limited thermal tolerance in fish? Answers obtained by nuclear magnetic resonance techniques. Respir Physiol Neuro 141(3): 243-260. 179. Pörtner, HO, Langenbuch, M, Michaelidis, B (2005) Synergistic effects of temperature extremes, hypoxia, and increases in CO2 on marine animals: From Earth history to global change. J Geophys Res 110 C9. 180. Pearson RG & Dawson TP (2003) Predicting the impacts of climate change on the distribution of species: are bioclimate envelope models useful? Glob Ecol Biogeogr 12: 361-371. 181. Perry, AL, Low, PJ, Ellis, JR, Reynolds, JD (2005) Climate Change and Distribution Shifts in Marine Fishes. Science 308(5730): 1912-1915. 182. Persson, L (1986) Temperature-induced shift in foraging ability in two fish species, roach (Rutilus rutilus) and perch (Perca fluviatilis) - implications for coexistence between poikilotherms. J Anim Ecol 55(3): 829-839. 183. Peterson, BJ, McClelland, J, Curry, R, Holmes, RH, Walsh, JE, Aagaard, K (2006) Trajectory shifts in the Arctic and Subarctic freshwater cycle. Science 313: 1061-1066. 184. Planque, B, Fox, CJ (1998) Interannual variability in temperature and the recruitment of Irish Sea cod. Mar Ecol Prog Ser 172: 101-105. 185. Planque, B, Fredou, T (1999) Temperature and the recruitment of Atlantic cod (Gadus morhua). Can J Fish Aquat Sci 56(11): 2069-2077. 186. Poloczanska, ES, Cook, RM, Ruxton, GD, Wright, PJ (2004) Fishing vs. natural recruitment variation in sandeels as a cause of seabird breeding failure at Shetland: a modelling approach. ICES J Mar Sci 61(5): 788-797. 187. Polovina, JJ (2005) Climate Variation, Regime Shifts, and Implications for Sustainable Fisheries. Bull Mar Sci 76(2): 233-244. 188. Pope, JG, Symes, D (2000) An ecosystem based approach to the Common Fisheries Policy: defining the goals. Eng Nat: 40 pp. 189. Rahmstorf, S (2000) The thermohaline ocean circulation: a system with dangerous thresholds? Clim Change 46(3): 247-256. 190. Rahmstorf, S (2002) Ocean circulation and climate during the past 120,000 years. Nature 419(6903): 207-214. 191. Raper, SCB: Viner, D, Hulme, M, Barrow, EM (1997) Global warming and the British Isles 454pp. 192. Ravier, C, Fromentin, J (2004) Are the long-term fluctuations in Atlantic bluefin tuna (Thunnus thynnus) population related to environmental changes? Fish Oceanogr 13(3): 145-160. 193. Riebesell, U, Zondervan, I, Rost, B, Tortell, PD, Zeebe, RE, Morel, FMM (2000) Reduced calcification of marine plankton response to increased atmospheric CO2. Nature 407: 364367. 194. Regier, H, Holmes, J, Pauly, D (1990) Influence of temperature-changes on aquatic ecosystems - an interpretation of empirical-data. Trans Am Fish Soc 119(2): 374-389. 125 PE 379.208 Klimawandel und europäische Fischerei 195. Renz, J, Hirche, H-J (2005) Life cycle of Pseudocalanus acuspes Giesbrecht (Copepoda, Calanoida) in the Central Baltic Sea: I. Seasonal and spatial distribution. Mar Biol 148(3): 567 – 580. 196. Reynolds, W, Casterlin, M (1977) Temperature preferences of four fish species in an electronic thermoregulatory shuttlebox. Prog Fish Cult 39(3): 123-125. 197. Rixen M, Beckers JM, Levitus S, Antonov J, Boyer T, Maillard C, Fichaut M, Balopoulos E, Iona S, Dooley H, Garcia MJ, Manca B, Giogetti A, Manzella G, Mikhailov N, Pinardi N, Zavatarelli M (2005) The Western Mediterranean deep water: aproxy for climate change. Geophys Res Lett 32. 198. Rodwell, MJ, Rowell, DP, Folland, CK (1999) Oceanic forcing of the wintertime North Atlantic Oscillation and European climate. Nature 398(6725): 320-323. 199. Rogers, J (1984) The association between the North-Atlantic Oscillation and the Southern Oscillation in the Northern hemisphere. Month Weath Rev 112(10): 1999-2015. 200. Root, TL, Price, JT, Hall, KR, Schneider, SH, Rosenzweig, C, Pounds, JA (2003) Fingerprints of global warming on wild animals and plants. Nature 421(6918): 57-60. 201. Rose, GA (2005) On distributional responses of North Atlantic fish to climate change. ICES J Mar Sci 62(7): 1360-1374. 202. Ruddiman, WF, Thomson, JS (2001) The case for human causes of increased atmospheric CH4. Quat Sci Rev 20(18): 1769-1777. 203. Sabine, CL, Feely, RA, Gruber, N, Key, RM, Lee, K, Bullister, JL, Wanninkhof, R, Wong, CS, Wallace, DWR, Tilbrook, B, Millero, FJ, Peng, T-H, Kozyr, A, Ono, T, Rios, AF (2004) The oceanic sink for anthropogenic CO2. Science 305(5682): 367-371. 204. Sarmiento, JL, Slater, R, Barber, R, Bopp, L, Doney, SC, Hirst, AC, Kleypas, J, Matear, R, Mikolajewicz, U, Monfray, P, Soldatov, V, Spall, SA, Stouffer, R (2004) Response of ocean ecosystems to climate warming. Global Biogeochem Cycles 18(3) 205. Scavia, D, Field, JC, Boesch, DF, Buddemeier, RW, Burkett, V, Cayan, DR, Fogarty, M, Harwell, MA, Howarth, RW, Mason, C, Reed, DJ, Royer, TC, Sallenger, AH, Titus, JG (2002) Climate Change Impacts on U.S. Coastal and Marine Ecosystems Estuaries. 25(2): 149-164. 206. Schär, C, Vidale, PL, Lüthi, D, Frei, C, Häberli, C, Liniger, MA, Appenzeller, C (2004) The role of increasing temperature variability in European summer heatwaves. Nature 427, 332-336. 207. Schrank, WE (2007) The ACIA, climate change and fisheries. Mar Pol 31(1): 5-18. 208. Send, U, Font, J, Krahmann, G, Millot, C, Rhein, M, Tintore, J (1999) Recent advances in observing the physical oceanography of the western Mediterranean Sea. Prog Oceanogr 44(1-3): 37-64. 209. Sharp, GD (2003) Future climate change and regional fisheries: a collaborative analysis. FAO Fish Tech Pap 452 75p. 210. Sheppard, C (2004) Sea surface temperature 1871-2099 in 14 cells around the United Kingdom. Mar Pollut Bull 49(1-2): 12-16. 211. Siegenthaler, U, Monnin, E, Kawamura, K, Spahni, R, Schwander, J, Stauffer, B, Stocker, TF, Barnola, JM, Fischer, H (2005) Supporting evidence from the EPICA Dronning Maud Land ice core for atmospheric CO2 changes during the past millennium. Tellus Ser B- PE 379.208 126 Klimawandel und europäische Fischerei Chem Phys Meteorol: 57(1): 51-57. 212. Sims, DW, Wearmouth, VJ, Southall, EJ, Hill, JM, Moore, P, Rawlinson, K, Hutchinson, N, Budd, GC, Righton, D, Metcalfe, J, Nash, JP, Morritt, D (2006) Hunt warm, rest cool: bioenergetic strategy underlying diel vertical migration of a benthic shark. J Anim Ecol 75(1): 176-190. 213. Sirabella, P, Giuliani, A, Colosimo, A, Dippner, JW (2001) Breaking down the climate effects on cod recruitment by principal component analysis and canonical correlation. Mar Ecol Prog Ser 216: 213-222. 214. Sissener, EH, Bjoerndal, T (2005) Climate change and the migratory pattern for Norwegian spring-spawning herring, implications for management. Mar Policy 29(4): 299-309. 215. Smetacek V, Nicol, S (2005)Polar ocean ecosystems in a changing world. Nature 437: 362-368. 216. Somarakis, S, Ganias, K, Siapatis, A, Koutsikopoulos, C, Machias, A, Papaconstantinou, C (2006) Spawning habitat and daily egg production of sardine (Sardina pilchardus) in the eastern Mediterranean. Fish Oceanogr 15(4): 281-292. 217. Southward, A, Boalch, G, Maddock, L (1988) Fluctuations in the herring and pilchard fisheries of devon and cornwall linked to change in climate since the 16th-Century. J Mar Biol Assoc UK 68(3): 423-445. 218. Southward, A, Hawkins, S, Burrows, M (1995) 70 years observations of changes in distribution and abundance of zooplankton and intertidal organisms in the Western English-Channel in relation to rising sea temperature. J Therm Biol 20(1-2): 127-155. 219. Spahni, R, Chappellaz, J, Stocker, TF, Loulergue, L, Hausammann, G, Kawamura, K, Fluckiger, J, Schwander, J, Raynaud, D, Masson-Delmotte, V, Jouzel, J (2005) Atmospheric methane and nitrous oxide of the late Pleistocene from Antarctic ice cores. Science 310(5752): 1317-1321. 220. Steele, JH (2004) Regime shifts in the ocean: reconciling observations and theory. Prog Oceanogr 60(2-4): 135-141. 221. Stenevik, EK, Sundby, S (2007) Impacts of climate change on commercial fish stocks in Norwegian waters. Mar Pol 31(1): 19-31. 222. Stenseth, NC, Mysterud, A, Ottersen, G, Hurrell, JW, Chan, K-S, Lima, M (2002) Ecological Effects of Climate Fluctuations. Science 297(5585): 1292-1296. 223. Stenseth, NC, Ottersen, G, Hurrell, JW, Mysterud, A, Lima, M, Chan, K-S, Yoccoz, NG, Aadlandsvik, B (2003) Review article. Studying climate effects on ecology through the use of climate indices: the North Atlantic Oscillation, El Nino Southern Oscillation and beyond. Proc R Soc Lond, Ser B: Biol Sci 270(1529): 2087-2096. 224. Stott, PA, Stone, DA, Allen, MR (2004) Human contributions to the heatwave of 2003. Nature 432: 610-614. 225. The Royal Society (2005) Ocean acidification due to increasing atmospheric carbon dioxide 12/05 68 pp. 226. Theocharis, A, Nittis, K, Kontoyiannis, H, Papageorgiou, E, Balopoulos, E (1999) Climatic changes in the Aegean Sea influence the Eastern Mediterranean thermohaline circulation (1986-1997). Geophys Res Lett 26(11): 1617-1620. 127 PE 379.208 Klimawandel und europäische Fischerei 227. Tietenberg, T (2003) The tradable-permits approach to protecting the commons: Lessons for climate change. Oxf Rev Econ Policy 19(3): 400-419. 228. Tonn, WM (1990) Climate change and fish communities: A conceptual framework. Trans Am Fish Soc. 229. Trigo I, Davies T, Bigg, G. (2000) Decline in Mediterranean rainfall caused by weakening of Mediterranean cyclones. Geophys Res Lett 27: 2913-2916. 230. Tsimplis M, Josey SA (2001) Forcing on the Mediterranean Sea by atmospheric oscillations over the North Atlantic. Geophys Res Lett 28: 803-806. 231. Tsimplis, MN, Alvarez-Fanjul, E, Gomis, D, Fenoglio-Marc, L, Perez, B (2005) Mediterranean Sea level trends: Atmospheric pressure and wind contribution. Geophy Res Lett 32(20): 1-4. 232. Turrell, W, Henderson, E, Slesser, G, Payne, R, Adams, R (1992) Seasonal-Changes In The Circulation Of The Northern North-Sea. Cont Shelf Res 12(2-3): 257-286. 233. Veizer, J (2005) Celestial climate driver: A perspective from four billion years of the carbon cycle. Geosci Can 32(1): 13-28. 234. Vignudelli S, Gasparini G, Astraldi M, Schiano ME (1999) A possible influence of the North Atlantic Oscillation on the circulation of the Western Mediterranean Sea. Geophys Res Lett 26: 623-626. 235. Vilhjalmsson, H (1997) Climatic variations and some examples of their effects on the marine ecology of Icelandic and Greenland waters, in particular during the present century. Rit Fisk J Mar Res Inst 15(1): 7-29. 236. Visbeck, MH, Hurrell, JW, Polvani, L, Cullen, HM (2001) The North Atlantic Oscillation: Past, present, and future. Proc Nat Am Soc 98(23): 12876-12877. 237. Vitousek, P (1994) Beyond global warming - ecology and global change. Ecology 75(7): 1861-1876. 238. Walther, G-R, Post, E, Convey, P, Menzel, A, Parmesan, C, Beebee, TJC, Fromentin, J-M, Hoegh-Guldberg, O, Bairlein, F (2002) Ecological responses to recent climate change. Nature 416(6879): 389-395. 239. Walther, GR, Hughes, L, Vitousek, P, Stenseth, NC (2005) Consensus on climate change. Trends Ecol Evol 20(12): 648-649. 240. Wang, T, Overgaard, J (2007) The heartbreak of adapting to global warming. Science 315(5808): 49-50. 241. Weiss RF (1970) The solubility of nitrogen, oxygen and argon in water and seawater. Deep-Sea Res 17: 721-735. 242. WBGU Report (2006) The future Oceans- Warming up, rising high, turning sour. 110pp. 243. Whitehead, PJP, Nelson, GJ, Wongratana, T (1988) FAO species catalogue. Vol. 7. Clupeoid fishes of the world (Suborder Clupeoidei). An annotated and illustrated catalogue of the herrings, sardines, pilchards, sprats, shads, anchovies and wolf-herrings. Part 2 – Engraulididae. FAO fish synop 125(7/2): 305-579. 244. Wieland, K, Waller, U, Schnack, D (1994) Development of Baltic cod eggs at different levels of temperature and oxygen content. Dana 10: 163-177. 245. Wiltshire KH, Manly BFJ (2004) The warming trneds at Helgoland Roads, North Sea: Phytoplankton response. Helgol Mar Res 58: 269-273. PE 379.208 128 Klimawandel und europäische Fischerei 246. Winder, M, Schindler, DE (2004) Climate change uncouples trophic interactions in an aquatic system. Ecology 85(11): 3178-3178. 247. Wonham, MJ, Carlton, JT, Ruiz, GM, Smith, LD (2000) Fish and ships: Relating dispersal frequency to success in biological invasions. Mar Biol 136(6): 1111-1121. 248. Worm, B, Myers, RA (2003) Meta-analysis of cod-shrimp interactions reveals top-down control in oceanic food webs. Ecology 84(1): 162-173. 129 PE 379.208 Notizen BA-30-08-216-DE-C