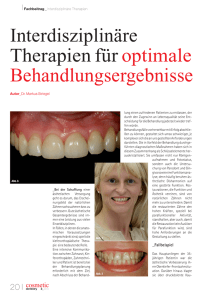
der hand
Werbung

13.9.–22.11. 2009 Der Weg zum Menschen Stadtmuseum Erlangen Martin-Luther-Platz Di/Mi 9 –17 Uhr Do 9 –13 und 17 –20 Uhr Fr 9 –13 Uhr Sa/So 11–17 Uhr Hand und Fuß Der Weg zum Menschen Nichts war für die Menschwerdung so entscheidend wie die Entwicklung von Hand und Fuß. Sein aufrechter Gang, seine Art zu fassen und zu greifen unterscheiden den Menschen von allen anderen Primaten. Diese Eigenschaften bilden auch wichtige Voraussetzungen für die Entwicklung seines Gehirns: Der Weg des Menschen verläuft in der Gattungsgeschichte wie in der Individualgeschichte vom Greifen zum Begreifen. Die Ausstellung zeichnet diesen Aspekt der menschlichen Evolution anhand zahlreicher Exponate aus Sammlungen der Universität sowie aus dem Besitz öffentlicher Museen und privater Leihgeber in einer facettenreichen Präsentation nach. Zugleich bettet sie dieses Kapitel der Evolution in die Geschichte der Evolutionstheorie überhaupt ein und stellt insoweit einen wichtigen Beitrag zum Darwin-Jahr 2009 dar. Ein eigener Ausstellungsteil veranschaulicht die besondere Bedeutung, die der Hand über ihren Werkzeug­c harakter hinaus in allen Kulturen als Zeichen und Symbol zuge­w iesen wurde und wird. Hand und Fuß Der Weg zum Menschen 13.9. – 22.11.2009 Eine Ausstellung des Stadtmuseums Erlangen, der Friedrich-Alexander-Universität Erlangen-Nürnberg (Lehrstuhl für Entwicklungsbiologie, Institut für Anatomie I, Zentralinstitut für Angewandte Ethik und Wissenschafts­kommunikation, Institut für Ur- und Frühgeschichte) und des Naturkunde-Museums Coburg Ausstellungsbereiche 1. Geschichte der Evolutionstheorie Dr. Rudolf Kötter 2. Geschichte von Hand und Fuß Dr. Wolfgang Heimler 3. Stammbäume Dr. Wolfgang Heimler, Dr. Werner Korn 4. Primaten Dr. Wolfgang Heimler 5. Mensch Dr. Wolfgang Heimler, Prof. Dr. Winfried Neuhuber, Dr. Leif Steguweit 6. Sprechende Hände Thomas Engelhardt Mitarbeiter der Ausstellung Leihgeber Dr. Wolfgang Heimler Friedrich-Alexander-Universität Erlangen-Nürnberg Department Biologie, Zoologische Sammlung, Universität Erlangen-Nürnberg Anatomische Sammlung (Wissenschaftliche Leitung) Antikensammlung Thomas Engelhardt Stadtmuseum Erlangen (Ausstellungsleitung) Dr. Rudolf Kötter Zentralinstitut für Angewandte Ethik und Wissenschaftskommunikation, Universität Erlangen-Nürnberg Prof. Dr. Winfried Neuhuber Institut für Anatomie I, Universität Erlangen-Nürnberg Graphische Sammlung der Universität Plastisch- und Handchirurgische Klinik, Universitätsklinikum Erlangen Ur- und Frühgeschichtliche Sammlung Universitätsbibliothek Zoologische Sammlung Kunstmuseum Erlangen Städtische Sammlung Erlangen Marcus Sommer SOMSO Modelle GmbH, Coburg Naturkunde-Museum Coburg, Coburger Landesstiftung Deutsches Medizinhistorisches Museum, Ingolstadt Dr. Leif Steguweit Bayerische Staatsbibliothek, München Institut für Ur- und Frühgeschichte, Universität Erlangen-Nürnberg Bayerische Staatssammlung für Paläontologie und Geologie, München Dr. Werner Korn Zoologische Staatssammlung, München Eberhard-Karls-Universität Tübingen, Institut für Ur- und Frühgeschichte Naturkunde-Museum Coburg Museum Kulturgeschichte der Hand, Wolnzach Grafik: Peter Hörndl Frank Bücher, Alles was schön ist, Erlangen Textredaktion: Dr. Rudolf Kötter, Gertraud Lehmann Öffentlichkeitsarbeit und Projektassistenz: Katharina Gamer Manfred Mayer, Erlangen Dr. Harald Tesan, Nürnberg Völk Orthopädie, Erlangen Videopräsentation: Dr. Harald Tesan (Konzeption) clip...trix, Höchstadt (Realisierung) Präparatorin: Ulrike Neumann, Naturkunde-Museum Coburg Wir danken für die freundliche Unterstützung durch Ausstellungsgestaltung und -aufbau: Claus Theuerkauf, Marcus Sommer Somso Modelle GmbH, Coburg Hans-Jürgen Hippe, Klaus Staudt Arthrex GmbH, Karlsfeld/München Museumspädagogik: Christine Brehm, Lars Hochreuther Studentische Mitarbeiter und Mitarbeiterinnen: Jessica Durlack, Mario Hertlein, Christian Jobst, Isabell Kappel, Edith Merkel, Jürgen Mehl, Oliver Neumann, Elisabeth Obermeier, Tanja Vockenroth, Christian Völk, Bernd Weber, Markus Weinl STADTMUSEUM E R LA N G E N Die Verzeitlichung der Naturgeschichte Abb. 1 D ie moderne Erdgeschichts­ schreibung beginnt mit dem däni­ schen Naturforscher Nicolaus Steno (1638 – 1687), der das „strati­ graphische Prinzip“ formulierte: Die Anordnung von Gesteinsschichten im Raum entspricht einer zeitlichen Ordnung, wobei die tiefsten Schichten die ältesten sind. Außerdem erkannte Steno, dass es sich bei Fos­silien um versteinertes organi­ sches Material handelt, das immer älter ist als das umgebende Gestein. Dieser Gedanke wurde später aufgegriffen und zur relativen Datierung von Gesteinsschichten benutzt: Ge­ steinsschichten, die gleiche Fossi­lien („Leitfossilien“) enthalten, müssen gleich alt sein. Georges Cuvier (1769 – 1838) erkannte durch das Studium von Leitfossilien, dass sich in der Erdgeschichte dramatische Veränderungen voll­zogen haben müssen und postu­ lierte, dass durch katastrophale Ereignisse Teile des Lebens auf der Erde immer wieder vernichtet wor­ den sind. Im Gegensatz dazu vertrat der englische Geowissenschaftler Charles Lyell (1797 –1875) das metho­dische Prinzip des Aktualismus. Aktualistisch dürfen zur Rekonstruktion der Erdgeschichte nur solche Pro­zesse herangezogen werden, die man auch heute noch beobachten kann. Erst auf dem Boden des Aktua­ lismus konnte man die Zeitdauer von geologischen Entwicklungen quantitativ abschätzen. Abb. 2 Abb. 1 Rekonstruktion eines ausgestorbenen Riesenfaultiers aus: Georges Cuvier: Essay on the Theory of the earth Edinburgh 1813 Cuvier rekonstruierte das Riesenfaultier (Megatherium cuvieri) aufgrund von Knochenfunden nach den von ihm aufgestellten Prinzipien der vergleichenden Anatomie. Das Riesenfaultier wurde bis zu 6 m groß und lebte bis in die Bronzezeit in Südamerika. Abb. 2 Schichtenaufbau und geologisches Alter aus: Charles Lyell: Principles of geology, Vol. 3 London 1830 Die Abbildung No.1 zeigt, dass die oberen Gesteinsschichten normalerweise auch die jüngeren sind, und erklärt, wie gelegentlich ältere Schichten (d) über die jüngeren (a – c) zu liegen kommen. Abbildung No. 2 veranschaulicht den geologischen Aufbau des Pariser Beckens. Im Innern der großen geologischen Mulde treten die jüngsten Schichten des Tertiärs zutage. Nach außen hin folgen immer ältere Formationen aufeinander. Zeiträume M it Hilfe der aktualistischen Methode konnte man quanti­tative Abschätzungen der Zeitdauer von geologischen Entwicklungen vornehmen. In diesem Sinne durchgeführte Beobachtungen und Experimente zeigten, dass die Abtragung von Gebirgen durch Erosion und die Bildung neuer Gesteinsschichten durch Sedimentation gewaltige Zeiträume beanspruchen (300 –500 Mio. Jahre). Das war für viele Menschen, die bis dahin noch geglaubt hatten, die Erde sei etwa 6 000 Jahre alt, geradezu unvorstellbar. Folglich mussten frühere Epochen wesentlich wärmer und damit lebens­ feindlicher gewesen sein. Dieser Widerspruch zwischen geologischen und physikalischen Verlaufsmodellen der Erdgeschichte löste sich erst mit der Entdeckung der Radioaktivität (und ihren energetischen Konsequenzen) zu Beginn des 20. Jahrhunderts auf. Altersbestimmung nach der Radio-Carbonmethode Halbwertzeit des Kohlenstoff-Isotops C-14: 5 730 Jahre Innerhalb dieses Zeitraums hat sich die Hälfte des radioaktiven Kohlenstoffs C-14 umgewandelt. 25% Ein gewichtiger Einwand gegen die Vorstellungen der Geologen kam von den Physikern. Sie haben mit Hilfe der klassischen Thermodynamik und ausgehend von Schätzungen der vorhandenen Erdwärme Modelle für die Entwicklung von Sonne und Erde entwickelt (Lord Kelvin, 1824 –1907). Altersbestimmung nach der C-14 Methode Danach musste die Geschichte der Erde relativ kurz sein (deutlich unter 100 Mio. Jahre) und einen Prozess der ständigen Abkühlung darstellen. Zur absoluten Zeitbestimmung erdgeschichtlicher Ereig­ nisse werden heute Zerfallsreihen radioaktiver Isotope heran­ge­zogen. Am bekanntesten ist die zur Altersbestimmung von organischem Material aus erdgeschichtlich jüngerer Zeit benutzte C-14-Methode. Zur Bestimmung größerer Zeiträume verwendet man z. B. die Zerfallsreihe von Kalium-40 zu Argon-40 mit einer Halbwertszeit von 1 300 Mio. Jahren. 100% C-14-Gehalt nach 0 Jahren 50% C-14-Gehalt nach 5 730 Jahren C-14-Gehalt nach 11 460 Jahren „I have shown that most probably the sun was sensibly hotter a million years ago than he is now. Hence, geological speculations assuming somewhat greater extremes of heat, more violent storms and floods, more luxuriant vegetation, and hardier and coarser-grained plants and animals, in remote antiquity, are more probable than those of the extreme quietist, or ‚uniformitarian‘ school. ... it is impossible that hypotheses assuming an equability of sun and storm for 1.000.000 years, can be wholly true.“ Lord Kelvin (William Thomson), 1864 Ähnlichkeit und Verwandtschaft Als wichtiges Prinzip der vergleichenden Anatomie galt das Gesetz der „Kor­relation der Teile“, wonach Organe und Skelettteile immer funk­ ti­o­nal aufeinander bezogen sind. Auf­grund dieses Gesetzes lassen sich z. B. aus einzelnen Knochenfunden ganze Skelette rekonstruieren. Abb. 1 A n der Wende vom 18. zum 19. Jahrhundert wurde durch Biologen wie Georges Cuvier (1769 – 1838), Étienne Geoffroy Saint-Hilaire (1772 –1844) und Jean-Baptiste de Lamarck (1744 –1829) die ver­ gleichende Ana­tomie und Morpho­ logie begründet. Deren grund­legende Idee war, dass sich die Verwandtschaft von Arten über Gemeinsamkeiten in ihren ana­tomischen bzw. morphologischen Bauplänen konstru­ieren ließen: Der Grad der Verwandt­schaft lässt sich an der Ausdifferenzie­rung eines abstrakten Grundbau­plans ablesen. Umstritten waren unter den Theo­re­ tikern der damaligen Zeit insbesondere, (a) ob es für die verschiedenen Lebensformen unterschiedliche Grundbaupläne gibt (Cuvier), oder ob diesen wiederum ein gemeinsamer „Master-Plan“ zugrundeliegt (Geoffroy Saint-Hilaire, mit dem auch Goethe sympathisierte) und (b) ob die Aus­differenzierung in den Bauplänen auf eine historische Entwicklung verweist (bejaht von Lamarck, abgelehnt von Cuvier). Abb. 2 Abb. 1 Die Bewegungsapparate von Lemur, Vogel Strauß und Eidechse aus: Georges Cuvier: Vorlesungen über vergleichende Anatomie. Erster Theil, welcher die Organe der Bewegung enthält. Leipzig 1809 Hier werden Skelette von Gorilla (o. links), Vogel Strauß (o. rechts) und Eidechse (unten) als Repräsentanten verschiedener Tier­gruppen zum Vergleich ihres Bewegungsapparats dargestellt. Abb. 2 Idealmuster oder Urtyp des Wirbeltier-Skeletts nach: Richard Owen: On the nature of Limbs. London 1849 Die vergleichende Anatomie beruht auf der Grundannahme, dass die höchst unterschiedlichen Baupläne der Arten einer bestimmten Gruppe (z. B. der Wirbeltiere) Konkretisierungen eines gemeinsamen, abstrakten Bauplan-Musters sind. Richard Owen (1804 – 1892) hat einen solchen lange Zeit akzeptierten „Master-Plan“ für das Wirbeltierskelett erstellt. Verwandtschaft und Abstammung haben. Er ging davon aus, dass sich zu allen Zeiten (auch heute noch!) primitives Leben aus unbeleb­ten Vorformen bilde und sich dieses Leben dann im Laufe der Zeit den sich wechselnden Umweltbedingungen an­passe. Abb. 1 Diese Anpassungen würden dann an die nächste Generation weiterge­ geben, wodurch immer komplexere Lebensformen entstehen. Je komplexer eine rezente Art ist, desto älter ist sie, d. h. jede Art hat ihre eigene, von anderen Arten getrennte Entwicklungsgeschichte. Damit vertrat La­marck eine Transformationstheorie der Arten, aber noch keine Abstammungstheorie; dies wird häufig missverstanden. Z Wirbellose Tiere Infusiorien Polypen Strahltiere Annelieden Cirripedien Mollusken Insekten Arachniden Crustaceen Wirbeltiere Fische Reptilien Amphibische Säugetiere Vögel Monotremen Cetaceen Säugetiere Zeit u Beginn des 19. Jahrhunderts ver­teidigte vor allem Georges Cuvier (1769 – 1838) vehement die Über­ zeugung, dass in der Naturgeschichte die Konstanz der Arten herrsche, d. h. die Arten sich seit der Schöpfung in ihren strukturellen Eigenarten erhalten haben, wenn sie nicht durch Katastrophen vernichtet worden sind. wiederholte Urzeugung Entwicklung zur „Vervollkommnung” Abb. 2 Ihm gegenüber vertrat vor allem JeanBaptiste de Lamarck (1744 – 1829) die An­sicht, dass der Ausdifferenzierung von einfachen Grundbauplänen in komplexe­re Baupläne ein historischer Prozess entspreche, bei dem sich gegenwärtige Arten aus einfachsten Urformen entwickelt Abb. 1 Titelblatt der Erstausgabe von Lamarcks „Philosophie zoologique“ Paris 1809 Die „Philosophie zoologique“ ist das Hauptwerk Lamarcks. Ihre erste deutsche Übersetzung erschien 1876 als „Philo­ sophische Zoologie“. Abb. 2 Lamarcks Modell: Fortschreitende Evolution ohne gemeinsame Abstammung Jean-Baptiste de Lamarck (1744 – 1829) vertrat die Idee, dass sich das Leben im Laufe der Zeit zwar aus primitiven zu höheren Formen weiter entwickele, dieser Prozess aber nicht einmalig sei, sondern sich ständig wiederhole. Wir finden demnach zu jeder Zeit Lebewesen auf unterschiedlichen Entwicklungsstufen vor, die eine mehr oder weniger lange Entwicklungsgeschichte vor sich haben. Charles Darwin: Person entwickelte er sein Hauptwerk „On the Origin of Species“, das 1859 erschien. Gleich­zeitig mit Darwin (wenn nicht sogar etwas vor ihm) hat der engli­sche Naturforscher Alfred R. Wallace (1823 – 1913), ebenfalls nach der Lektüre von Malthus, seine der Darwinschen Theorie sehr ähn­ liche Evolutionstheorie entworfen und niedergeschrieben. Abb. 1 C harles Robert Darwin wurde 1809 in Shrewsbury geboren, studierte Medizin und Theologie und schloss 1831 sein Theologiestudium mit dem Bachelor-Grad ab. Kurz danach bekam er die Gelegenheit, an der Welt­­ umseglung des Captain FitzRoy auf der HMS Beagle teilzunehmen. 1836 kehrte er mit einer gewaltigen Samm­ lung an Präparaten und Materialien zurück, deren Auswertung ihn über Jahre hinweg beschäftigte. Im Gegensatz zu Wallace wurde Darwin durch sein Buch ungeheuer populär, allerdings zu dem Preis, dass viele seiner weiteren wichtigen und überaus interessanten Arbeiten nicht die öffentliche Aufmerksamkeit erhielten, die sie eigentlich verdient hätten. Darwin starb 1882 hoch ge­ehrt in seinem Heimatort Down bei London. Während dieser Zeit und angeregt durch die geologischen Arbeiten von Charles Lyell sowie durch den „Essay on the Principles of Population“ von Thomas R. Malthus (1798) Abb. 2 Abb. 1 Charles Darwin im Alter von 40 Jahren Lithographie von T. H. Maguire, 1849 London, National History Museum Abb. 2 Charles Darwin im Alter von 71 Jahren Fotografie, 1880 London, Natural History Museum Abb. 3 Titelseite der Erstausgabe von „On the Origin of Species“ London 1859 Das Buch erlebte zu Darwins Lebzeiten sechs Auflagen. Eine erste deutsche Übersetzung ist schon im Jahre 1860 erschienen. Abb. 3 Charles Darwin: Theorie oder -nachteil (Selektion). Auf Dauer setzen sich in einer Population die Träger mit „positiven“ Merkmals­ ausprägungen durch (Anpassung). 3. Wird eine Population durch äußere Umstände räumlich getrennt, so greifen dadurch unterschiedliche Selektionsmechanismen, was dazu führt, dass die Teilpopulationen sich im Laufe der Zeit in ihren Merkmals­ ausprägungen immer stärker voneinander unterscheiden, bis sie schließ­ lich nicht mehr als zu einer Art ge­­hörig erfasst werden können: Auf diese Weise entstehen neue Arten. Abb. 1 D ie wesentlichen Grundlagen der Evolutionstheorie lassen sich in vier Punkten zusammenfassen: 4. Die Evolutionsgeschichte des Lebens beschreibt, wie heutige Arten aus früheren, diese wieder aus vor­ gän­gigen usw. entstanden sind. Damit gibt der Stammbaum des Lebens nicht nur die Verwandtschaft der Arten wieder, sondern zeigt auch den Weg ihrer historischen Entwicklung auf. 1. Im Gegensatz zur Transfor­ma­ti­ons­­ theorie von Lamarck ist die Evo­lu­ti­ons­ theorie eine Abstammungstheorie, die zeigt, wie sich aus einer Stamm­ art neue Arten entwickeln. 2. Ausgegangen wird von dem Faktum, dass die Individuen einer Art über unterschiedliche Merkmals­ ausprä­gungen verfügen, die das Leben unter gegebenen Umwelt­ bedingungen erleichtern oder erschweren. Dadurch erhalten ihre Träger einen Vermehrungsvorteil Abb. 3 Abb. 1Darwins erster Stammbaum-Entwurf aus: Charles Darwin: Notebook B, 1837/38 Cambridge University Library Die erste Skizze eines Stammbaums von Darwin enthält schon die Idee, dass Menschenaffen und Menschen auf einen gemeinsamen Vorfahren zurückgehen. Abb. 2 Darwinfinken aus: Charles Darwin: Journal of researches. 2. Auflage. London 1845 Während der Forschungsreise der Beagle studierte Darwin auf den Galapagos-Inseln die dort lebenden Finken-Arten. Die verschiedenen Schnabelformen dieser eng verwandten Galapagos-Finken hat Darwin als Anpassung an verschiedene Lebensräume mit unterschiedlichem Futterangebot verstanden. Charles Darwin und der Darwinismus Abb. 1 D arwins Theorie wurde schon im 19. Jahrhundert von Ideologen und Politikern aus dem liberalistischen und später auch aus dem faschistischen Lager aufgegriffen und zur Rechtfertigung sozialer, ökonomischer, kultu­ reller und rassistischer Auslese missbraucht („Sozialdarwinismus“). Der Grund­gedanke war, dass eine Gesellschaftsordnung den naturgewollten Kampf ums Dasein, bei dem der Stärkere sich durchzusetzen hat, nicht behindern dürfe. Vielmehr müsse es wichtigste Aufgabe des Staates sein, dafür zu sorgen, dass die Schwachen und Minderwertigen nicht den Starken in ihrem Fortkommen hinderlich werden. Dabei wurden immer verdeckt oder offen weltanschauliche Kriterien herangezogen, um vorab zu bestimmen, was denn den „Stär­keren“ ausmache. So wurde der Verlauf der „natürlichen“ Evo­ lution, der ja keine Richtung kennt, bei den Ideologen geradezu in sein Gegenteil verkehrt. Abb. 2 Vorreiter einer Politisierung der Evo­lutionstheorie waren in England H. Spencer und H. St. Chamberlain, in Frankreich J. A. Gobineau und G. Vacher de Lapouge und in Deutschland vor allem E. Haeckel und später H. F. K. Günther, der die Grundlagen der nationalsozia­ listischen Rassenkunde schuf. Abb. 3 Abb. 1 Darwin-Karikatur, um 1880 Farblithographie von André Gill Cambridge University Library Die Karikatur erschien um 1880 als Titelseite der Pariser satirischen Zeitschrift „La Petite Lune“. Dargestellt ist ein Affe mit übergroßem Darwinkopf, der im „Baum der Erkenntnis“ turnt. Der Gedanke, dass der Mensch abstammungsgeschichtlich der nächste Verwandte des Menschenaffen ist, wurde damals von vielen noch als Provokation empfunden. Abb. 2 Verwandtschaft von Schimpanse, Gorilla, Orang und „Neger“, 1874 aus: Ernst Haeckel: Anthropogenie oder Entwicklungs­ geschichte des Menschen. Leipzig 1874 Hier zeigt sich ganz unverhohlen, wie wenig Distanz selbst Wissenschaftler gegenüber dem herrschenden Rassismus ihrer Zeit einzunehmen vermochten. Abb. 3 Propagandadruck zur Rassenkunde, 1938 aus: Alfred Vogel: Erblehre und Rassenkunde in bildlicher Darstellung. Stuttgart 1938 Auf so perfide Weise wollte man während der Nazi-Zeit schon Kindern beibringen, dass die Ausrottung der Schwachen und Kranken im Grunde genommen ganz im Einklang mit der Natur stehe. Die Evolution von Hand und Fuß gungsapparates war eine wesentliche Voraussetzung für den Landgang. Nach diesem Prinzip kommen bis heute die Kriechtiere vorwärts. Säuger und Vögel hingegen bewegen sich nur mit den nach unten gerichteten Extremitäten fort. Die Rumpfwirbelsäule wurde bei ihnen zu einem geraden Stab, auf den die Antriebs­ kräfte übertragen werden. Abb. 1 V or mehr als 400 Mio. Jahren – beim Übergang vom Strudler zum aktiven Räuber – sind die paarigen Extremi­tä­ ten der Wirbeltiere entstanden. Die Brustflossen fungieren als Tragflächen und Steuerruder, die Schwanzflosse besorgt den Vortrieb, die übrigen Extremitäten dienen als Stabilisatoren. Bei Haien hat sich diese Konstruktion fast unverändert erhalten. Die mit den Vierfüßern eng verwand­ten Quastenflosser und Lungenfische besitzen beinartige Flossen. Einige devo­ nische Quastenflosser sind mit ihrer Hilfe am Gewässergrund „gelaufen“. Neu gebildete Finger- und Zehenstrah­ len haben ihre Flossenfächer er­setzt. Die so veränderten Extremi­täten unterstützen als seitliche Hebel das Schlängeln des Körpers und verringern die Reibung durch das Anheben des Bauches. Dieser Aufbau des Bewe- Die überaus bewegliche Greifhand der Primaten behielt ihren anatomischen Aufbau und gleicht immer noch stark den vorderen Extremitäten der ursprünglichen Vierfüßer. Eusthenopteron Acanthostega Ichthyostega Abb. 3 Abb. 1 Bewegungsformen der Wirbeltiere aus: Kardong (1998) obere Reihe: Schlängelschwimmen bei kieferlosen Wirbeltieren (Neunauge) mittlere Reihe: Schlängelschwimmen mit paarigen Ruder- und Steuerflossen (Hai) untere Reihe: Schlängellaufen mit seitlichen Extremitäten bei Lurchen und Reptilien Abb. 2 Vierfüßer – Konstruktionen aus: Kardong (1998) oben: Grundkonstruktion. Die seitlich gestellten Extremitäten verbessern die Effizienz beim Schlängeln. unten: Säugerkonstruktion. Die nach unten gerichteten Extremitäten übernehmen allein die Fortbewegung, die Wirbelsäule ist bogenförmig. Abb. 3 Entstehung von Hand und Fuß bei devonischen Quastenflossern und Amphibien aus: Linder (2006) Abb. 2 obere Reihe: Eusthenopteron mit Flossenfächer mittlere Reihe: Acanthostega mit Fingerstrahlen zum Laufen auf Gewässergrund untere Reihe: Ichthyostega mit kräftigen Laufbeinen und Schwanzflosse Abstammung des Menschen nach Darwin und Haeckel Im Unterschied zu Darwin sah er den Ursprung der menschlichen Entwicklungslinie eher in Südost-Asien mit den Gibbons und Orangs als nächsten Verwandten. Hoch spekulativ ist seine Annahme eines versunkenen Kontinents „Lemuria“ im Indischen Ozean als mögliche Heimat des Urmenschen. Erstmals versuchte Haeckel die tat­säch­ lichen Verwandtschaftsverhältnisse in Form eines Stammbaums bildlich darzustellen. Da damals noch keine vormenschlichen Fossilien bekannt waren, überbrückte er die Lücken durch angenommene Zwischenformen wie den Pithecanthropus alalus und Homo stupidus. Abb. 1 C harles Darwin (1809 – 1882) sah die Entstehung des Menschen wie die aller Arten als Evolutionsprozess. Grundlage für Darwins Annahmen waren morphologische, anatomische und physiologische Vergleiche rezenter Arten einschließlich ihrer Embryonalstadien und Jugendentwicklung. Dabei fand er die größte Übereinstimmung zwischen dem Menschen und den afrikanischen Menschenaffen und vermutete richtig die Wiege der Menschheit in Afrika. Der Zoologe Ernst Haeckel (1834 – 1919) war einer der bedeutendsten Ver­ fechter von Darwins Evolutionstheorie im 19. Jahrhundert, die er in seinen popu­lären Schriften mit missiona­­ rischem Eifer vertrat. Abb. 2 Abb. 1 Stammbaum der Primaten nach Darwin Handzeichnung, „drawn by Darwin on April 21, 1868” Cambridge University Library. Darwins Stammbaumskizze stellt ausschließlich die Verwandschaftsverhältnisse lebender Arten dar. Nach Darwin zweigt die Menschenlinie („men“, links) vor dem Ursprung der Menschenaffenlinien („Gorilla + Chimp.“, „Orang-utan“, „Hylobates“, Mitte) vom Hauptstamm ab. Ganz rechts sind die noch früher abzweigenden Tieraffen (Paviane und Makaken) dargestellt. Abb. 2 Stammbaum der Primaten nach Haeckel Handzeichnung, 1868 aus: Heberer (1968) Der Stammbaum Haeckels bezieht erstmals fossile Formen ein. Vorhandene Lücken werden mit spekulativen Zwischenformen ausgefüllt. Die Menschenlinie beginnt mit einem Urgibbon (Protohylopates atavus) und führt über den sprachlosen Affenmenschen (Pithecanthropus alalus) über den stupiden Mensch (Homo stupidus) zum weisen Menschen (Homo sapiens). Abstammung des Menschen nach Heberer Pongiden (Menschenaffen) Hominiden (Menschen) Gorilla Schimpanse Orang-Utan Homo sapiens Australopithecinen P A Holozän 1 000 000 Pleistozän Homininen 3 000 000 Pliozän Tier-Mensch-Übergangsfeld 6 000 000 Brachiatoren BrachiatorenHypothese 9 000 000 12 000 000 Miozän 15 000 000 18 000 000 PräbrachiatorenHypothese 21 000 000 Dryopithecinen ProtocatarrhinenHypothese Proconsul 24 000 000 Präbrachiatoren Propliopithecinen 27 000 000 Jahre Oligozän Ancient member Die „Brachiatoren“-Hypothese vermutet einen gemeinsamen Vorfahren von Mensch und Menschen­affen mit einer Trennung der Entwicklungslinien vor 6 – 12 Mio. Jahren. Obwohl zeitlich den heutigen Erkennt­nissen ganz nahe, verwarf Heberer auch diese Hypothese, da er die Transformation zum Menschen in so kurzer Zeit für unmöglich hielt. Nach der von ihm favorisierten „Prä­ brachiatoren“-Hypothese liegt eine lange evolutionäre Distanz von rund 25 Mio. Jahren zwischen dem Homo sapiens und dem letzten gemeinsamen Vorfahren von Mensch und Menschenaffe. Abb. 1 D er deutsche Anthropologe Gerhard Heberer (1901 – 1973) prägte mit seiner starken Betonung der Sonderentwicklung des Menschen lange Zeit die Vorstellungen über unsere Evolution. Damals wurden drei Hypo­ thesen diskutiert: Nach der „Protocatarrhinen“-Hypo­ these beginnt der Eigenweg des Menschen vor Entstehung der Alt­ welt­affen (ca. 40 Mio. Jahre) – eine Hypothese, die Heberer rasch wieder verwarf, weil ihm eine so lange Parallelentwicklung unwahrscheinlich erschien. Heberer erweiterte die Methoden Haeckels um die Untersuchung der Chromosomen, ignorierte aber die längst erkannte enge Blutgruppen­ verwandtschaft zwischen Schimpanse und Mensch. Trotz Kenntnis der absoluten Datierung favorisierte er einen völlig falschen Zeitrahmen. Offensicht­ lich vertraute er auf den klassischmorphologischen Ansatz mehr als auf die damals noch neuen molekularen Methoden. Abb. 1 Die Evolution des Menschen nach Heberer aus: Heberer (1968) Das Schaubild stellt die drei damals erörterten Hypothesen zur menschlichen Abstammung dar, wobei die beiden „weniger wahrscheinlichen“ blau gezeichnet sind. Gemeinsam ist ihnen die Annahme eines „Tier-Mensch-Übergangsfeldes“ (oberer Kreis) vor ca. 6 Mio. Jahren. Nach aktuellen Vorstellungen fällt dieser Übergang mit dem Auftreten der Gattung Homo und der Werkzeugherstellung vor ca. 2,5 Mio. Jahren zusammen. Abstammung des Menschen Aktueller Stand Evolutionsstammbaum H. sapiens H. neanderthalensis 1 Millionen Jahre vor heute 0 H. floresiensis H. heidelbergensis H. erectus H. ergaster P. boisei P. robustus P. aethiopicus 2 H. habilis Die Molekularbiologie gibt allerdings keinen Einblick in den Evolutionsverlauf. Noch immer kann nur die Interpretation der Fossilien zeigen, wann Merkmale wie der aufrechte Gang oder die Greifhand aufgetreten sind. A. garhi A. africanus A. afarensis A. anamensis 4 Ardipithecus Homo 5 Paranthropus Orrorin tugenensis Australopithecus frühe Hominiden Altersspektrum der Hominiden 6 Sahelanthropus tchadensis mutmaßliche Verwandtschaftsbeziehungen zwischen den Hominidenarten Verwandtschaftsbeziehungen zwischen den Hauptgruppen der Hominiden 7 Abb. 1 W ährend die Evolutionsbiologie sich früher nur auf vergleichende Merkmals­ analysen von lebenden und fossilen Organismen stützen konnte, stehen ihr heute molekularbiologische Hilfsmittel wie die Genom- und Proteom-Analyse zur Verfügung. Ein wichtiges Hilfsmittel ist auch das Konzept der „molekularen Uhr“, das von einer bekannten Mutationsrate ausgeht und es erlaubt, aus der Anzahl der molekularen Unterschiede auf den dafür benötigten Zeitraum zu schließen. Danach erfolgte die Trennung vom Schimpansenvorfahren vor 6 – 8 Mio. Jahren. Ebenso wurde der Ursprung der Primaten auf die Zeit vor ca. 80 Mio. Jahren zurückverlegt, obwohl die bisher ältesten sicheren Fossilien nur 55 Mio. Jahre alt sind. Aktuelle Stammbaumdarstellungen wie die von Friedemann Schrenk (*1956) veranschaulichen die geo­ grafische und zeitliche Verortung der Fossilien, ohne sich auf eindeutige Verwandtschaftslinien festzulegen. Der Evolutionsverlauf ist inzwischen gut belegt, so dass wesentliche Ände­ rungen hier nicht mehr zu erwarten sind. Dagegen steht die Erforschung der evolutionsgenetischen Mechanismen erst am Anfang. Stammbaum der Evolution der Primaten Mensch Schimpansen Zeit (in Millionen Jahren vor heute) 3 Gorillas Orang-Utans Gibbons Altweltaffen 0 5 10 15 20 25 30 35 Abb. 2 Abb. 1 Evolutionsstammbaum nach Zimmer nach: Zimmer (2006), Umzeichnung Peter Hörndl Aus aufrecht gehenden Menschenaffen (blau) gehen zierliche Vormenschen (Australopithecus, grün) hervor, die einerseits große Pflanzen essende Vormenschen (Paranthropus, gelb) und andererseits werkzeug­herstellende Menschen (Homo, rötlich) hervorbrachten. Abb. 2 Beispiel einer molekularen Uhr aus Jones, Martin, Pealbeam (1994) Aus dem Vergleich der DNA-Sequenzen von Mitochondrien lebender Arten konnten die Aufspaltungszeitpunkte berechnet werden. Die Trennung von Mensch und Schimpanse erfolgte danach vor 6 Mio. Jahren. Ähnliche Ergebnisse brachte die Untersuchung anderer molekularer Merkmale. Slow Loris – „Slow Food“ Greifzangenkletterer Als Anpassung an diese Bewegungs­ art sind die Gelenke der Loris hochmobil konstruiert, damit die Tiere mit jeder Extremität ihr Ziel erreichen können. Loris besitzen auch, bezogen auf ihre Größe, eher kurze Extremi­ täten. Der Schwerpunkt der Tiere liegt daher näher am Ast und erhöht die Sicherheit beim Bewegen. Abb. 1 D Hände und Füße der Loris sind als Greifzangen ausgebildet. Durch die Verkürzung des zweiten Strahls wird die Hebelwirkung verbessert, so dass sie extrem fest zugreifen können. Auch ihre Muskulatur ist stärker für Geschicklichkeit und Greiffähigkeit ausgebildet – Loris springen nie! ie meisten Loris sind auf langsame und gut getarnte oder giftige Beutetiere wie Schmetterlingslarven und Tausendfüßer aus. Zum Aufspüren dieser Nahrung benötigen sie einen guten Geruchssinn und die Fähigkeit zum Entgiften, wozu ihnen ihr niedriger Grundumsatz hilft. Aus dieser Nahrungsspezialisierung hat sich eine energiesparende Fort­ bewegung entwickelt, die einzigartig unter den Primaten ist: das „Slow Climbing“ oder Greifzangenklettern. Dabei umklammern immer minde­stens drei Extremitäten den Ast, während die vierte langsam neuen Halt sucht. Ein solch vorsichtiges Vor­ankommen macht es Fressfeinden schwer, Loris zu entdecken (vgl. Faultiere). Abb. 2 Abb. 3 Abb. 1 Plumplori (Nycticebus coucang), Borneo aus: Cubit/Payn (1994) Die großen Augen mit Licht reflektierendem Augen­ hintergrund (Tapetum) sind charakteristisch für Nachttiere. Abb. 2 Skelett des Plumploris in Greifhaltung aus: Fleagle (1999) Abb. 3 Greifzangen – Hand und Fuß des Plumploris aus: Geissmann (2003) Die verkürzte 2. Strahl trägt am Fuß die für Halbaffen charakteristische Putzkralle, die anderen Zehen bzw. Finger haben flache Fingernägel. Das Totenkopfäffchen – ein vierhändiger Baumläufer und Verankerung, die Füße besorgen den Vortrieb. Großzehen und Daumen sind beim Umgreifen des Astes unentbehrlich. Die Krallen sind wie bei allen Primaten zu flachen Fingernägeln umgebildet, die zusammen mit den Fingerlei­ sten die Sensibilität steigern. Abb. 1 Der Schwanz der Totenkopf­äffchen ist aber auch ein gutes Beispiel für Rückentwicklung in der Evolution, da er nur noch bei Jungtieren als Greifschwanz funktioniert. D ie Totenkopfäffchen (Familie Kapuzineraffen) bilden nach den Klammeraffen die zweitgrößte Familie der Neuweltaffen. Ihre Vertreter sind alle von mittlerer Größe, ernähren sich von pflanzlicher und tierischer Nahrung und bewegen sich auf sehr vielseitige Weise. Durch ihre Greiffüße und Greif­ hände klettern sie gewandt und zügig durch die Äste. Ihr langer Schwanz fungiert dabei als Balancierstange. Dass die Art ihrer Fortbewegung noch stark dem vierbeinigen Säugergrundtyp ähnelt, zeigt sich auch in ihrem unspezia­ lisierten anatomi­schen Bau. So sind Arme und Beine etwa gleich­lang, wodurch der Schwerpunkt gleichmäßig verteilt ist. Die Arme dienen dabei der Steuerung Abb. 2 Abb. 3 Abb. 1 Totenkopfäffchen (Saimiri sciureus), Amazonasgebiet aus: Bloom (1999) Abb. 2 Vierhändiges Laufen auf dem Ast unter Einsatz von abspreizbaren Daumen und Großzehen aus: Fleagle (1999) Abb. 3 Hände und Füße des Totenkopfäffchens mit gut entwickelten, aber nicht opponierbaren Daumen aus: Fleagle (1999) Der Klammerschwanzaffe – vier Finger, fünf Hände! Auf der Innenseite ihres Schwanz­ endes ist, wie auf den Innenseiten unserer Hände, eine haarlose, hoch sensible Tasthaut ausgebildet, mit deren Hilfe die Affen sicher zugrei­fen können. Auch die Muskulatur und die Wirbelsäule des Greifschwanzes sind sehr kräftig ausgebildet, so dass die Tiere längere Zeit mit ihrem gesamten Körpergewicht unter einem Ast hängen können. Die Hände sind extrem an dieses „Unter-dem-AstHängen“ angepasst. Der Daumen ist nahezu vollständig reduziert, da die Hand als Greifhaken wirkt. Abb. 1 D ie Klammerschwanzaffen besitzen mit ihrem Greifschwanz eine vollwertige fünfte Extremität. Am Schwanz kopfunter an Ästen hängend können sie ihren Aktionsradius zum Erreichen von Nahrungs­quellen nahezu verdoppeln und so Früchte an dünnen Astspitzen erlangen, die ihnen wegen ihres relativ hohen Gewichts von 4 – 10 kg sonst nicht zugänglich wären. Abb. 3 Beim Laufen auf dem Ast wird der Schwanz meist zur Sicherung an einem höheren Ast eingehakt und wie eine Sicherungsschlaufe mitgeführt. Wie bei anderen die Arme zum Hangeln einsetzenden Affen der alten Welt (Gibbons, Orangs) sind die Arme der Klammerschwanz­affen im Vergleich zum Säuger- bzw. Affengrundbauplan verlängert und die Schultergelenke sehr beweglich. Abb. 1 Klammerschwanzaffe (Ateles geofroyi) und Wollaffe (Lagothrix lagotricha) nur am Greifschwanz aufgehängt aus: Macdonald (2004) Abb. 2 Skelett des Klammerschwanzaffen aus: Schultz (1972) Abb. 2 Die extrem verlängerten Handglieder bilden einen Greif­ haken beim Hangeln, der Daumen ist extrem reduziert. Abb. 3 Tastfelder auf der Greiffläche des Klammerschwanzes aus: Schultz (1972) Auch kleine Gegenstände wie Erdnüsse können mit dem Schwanz ergriffen und transportiert werden – eine fünfte Hand. Der Gibbon – keiner schwingt schneller Die filigrane Anatomie verschafft dem Gibbon Vorteile beim Beschaffen von Nahrung. Das Zusammenspiel der Schulter- und Handgelenke beim Hangeln erlaubt es ihm, die Arme vollständig hinter den Kopf zu bringen und den Körper um 90 Grad zu drehen. Durch die optimale Ge­ wichts­verteilung erntet er Astspitzen geschickter ab als die auf Ästen sitzenden oder stehenden Primaten. Makaken, die im Laufen Nahrung an Astspitzen zu erreichen versuchen, scheitern, da sie den Ast von sich wegdrücken. Dagegen beugt sich der Ast, an dem der Gibbon hängt, in dessen Richtung, so dass ihm die Ernte erleichtert wird. Abb. 1 K ein Affe schwingt schneller durch die Bäume als der Gibbon. Nur er beherrscht, einem Trapezflieger vergleichbar, das Schwinghangeln mit freier Flugphase (Brachiation). Diese Kunst ermöglichen ihm ein feinglied­ riger Körperbau, eine an das Hangeln angepasste bewegliche Schulterpartie, eine kurze Lendenregion, die läng­ sten Arme aller Primaten und fest zupackende Hände. Häufiger als andere Primaten geht der Gibbon aufrecht. Dabei stabili­ siert er sich ganz anders als der Mensch. Während der EinbeinstandPhase erreicht sein Körperschwerpunkt die tiefste und nicht die höchste Lage. Er beugt die Kniegelenke stark und hält die Arme zum Aus­ balancieren vor und hinter dem Kopf hoch. Abb. 2 Abb. 1 Weißhandgibbon (Hylobates lar), Südostasien aus: Bloom (1999) Abb. 2 Greifraum eines Gibbon aus: Geissmann (2003) Durch Hängen unter dem Ast können Gibbons ihren Greifradius stark erweitern und so auch die Kronenregion der Bäume mit schwachen Zweigen erreichen („terminal branch feeding“). Abb. 3 Schwing-Hangeln aus: Geissmann (2003) Beim Hangeln kann der Gibbon wie ein Pendel schwingen, bis er beinahe die Ausgangshöhe erreicht hat. Bei schneller Brachiation tritt eine Phase des freien Fluges vor dem Ergreifen des nächsten Halts auf. Abb. 3 Die großen Menschenaffen Die Hände der Knöchelgänger sind zwar zum Greifen und Werkzeug­ gebrauch geeignet, lassen aber nur wenige Präzisionsgriffe zu. An der Außenseite ihrer Handknöchel tragen sie zusätzliche Fingerlinien. Die enge Verwandtschaft von Mensch und Schimpanse legt nahe, den Knöchelgang auch für die Vorfahren des Menschen anzunehmen. Jüngste Untersuchungen stellen diese Hypo­ these infrage. In jedem Fall waren aber unsere Urahnen Baumbewohner. Abb. 1 Alle Menschenaffen bauen Schlafnester auf Bäumen, selbst dann, wenn sie, wie die Gorillas, viel Zeit am Boden verbringen. Hier bewegen sie sich – auf die verlängerten Arme gestützt – auf allen Vieren vorwärts. Dabei stützen sich die Orangs auf ihre Faust (Faustgang), die afrikani­ schen Menschenaffen aber auf die Handknöchel (Knöchelgang). Nur kurze Strecken laufen die Menschenaffen – auf den Außenkanten ihrer Füße – aufrecht, was energe­ tisch sehr aufwendig ist. Vierbeiniger Stand Phalangen 2 und 3 ie die meisten Primaten leben die großen Menschenaffen in Wäl­ dern auf Bäumen. Ihr Körperbau hat sich an dieses Dasein angepasst und weist sie als gemäßigte Arm­ kletterer aus. Metacarpus Carpus W Phalangen 2 und 3 Abb. 2 Die aufrecht gehenden Vormenschen (Lucy) zeigten noch vor 3 Mio. Jahren Merkmale eines evolutionären Zwischenstadiums, wie abspreizbare Klettergroßzehen und Klettergreifhände mit verlängerten gebogenen Zehen und Fingern. Abb. 1 Berggorilla (Gorilla beringei) – „Siberrücken“, Ruanda aus: Bloom (1999) Wegen ihres hohen Gewichts halten sich ausgewachsene männliche Gorillas meist am Boden auf, wo sie sich auf die Arme gestützt im Knöchelgang fortbewegen, Abb. 2 Schimpanse (Pan troglodytes) – Weibchen in Knöchelganghaltung aus: Geissmann (2003) Details: Handskelett beim Aufstützen auf die Außenseite der Fingerknochen, rechts: Finger mit Hautleistenmuster auf der Oberseite Der aufrechte Gang muskulatur von außen zu umfassen und um die Adduktoren (Spann­ muskeln) der Oberschenkel mit dem Schambein zu verbinden. Schimpanse Brustregion und Arme bleiben, abgesehen von deren relativer Verkürzung, menschenaffenartig, nur die Greifhand wird für den präziseren Griff verändert: Der Daumen ist verstärkt und verlängert, die übrigen Finger sind verkürzt und gerade. Die senkrechte Wirbelsäule mit der charakteristischen Doppel-S-Form balanciert den Kopf, der ohne die kräftige Nackenmuskulatur der Menschenaffen auskommt. Mensch gebogene Phalangen großes Erbsenbein zum Kopf hin orientiertes Schultergelenk Australopithecus afarensis lange gebogene Phalangen relativ kurze Hintergliedmaßen Abb. 1 I m Vergleich mit Menschenaffen sind beim Menschen die Beine deutlich länger als die Arme. Der Fuß ist ein Lauffuß mit verstärktem Großzehstrahl, der parallel zu den übrigen Zehen den Körper beim Gehen vom Boden abstößt. Aufgesetzt wird der Fuß mit der Ferse, danach wird der Fuß abgerollt und wieder mit der Groß­ zehspitze abgestoßen. Das Becken ist verkürzt und verbreitert, um das Hüftgelenk mit der aufgefächerten GesäßAbb. 3 Abb. 1 Fortbewegung bei Menschenaffen, Vormenschen und Mensch aus Lewin (1995) Die Menschenaffen sind gemäßigte Armkletterer mit Greif­ füßen und leben überwiegend auf Bäumen. Die Vormenschen sind mit ihren Lauffüßen und verändertem Becken sowohl zum Klettern als auch zum aufrechten Gang befähigt. Der Mensch mit seinen verlängerten Beinen und verkürzten Armen hat sich völlig vom Baumleben emanzipiert. Abb. 2 Abb. 2 Becken und Oberschenkel des Menschen aus: Franzen (1997) Die Abbildung zeigt die Bänderbefestigung des Hüftgelenks (dunkel) und die Adduktoren zwischen Oberschenkel und Schambein. Das breite und kurze Becken des Menschen ist der Dreh- und Angelpunkt bei der Fortbewegung. Hier setzen die Beinbewegungs- und Haltemuskeln an. Abb. 3 Vergleich der Beinstellung bei Gorilla und Mensch aus: Franzen (1997) Die leicht nach innen geneigten Oberschenkel des Menschen benötigen weniger Muskelmasse als die extremen O-Beine des Gorillas. Der Pfeil symbolisiert die Lage des Schwerpunkts. Der Fuß dient als Lauffläche, die Fußgewölbe sorgen für die Federung. Zehen und Fuß werden über Sehnenzüge von der Unterschenkelmuskulatur bewegt, die Fußmuskulatur verspannt das Gewölbe, nur die große Zehe behält eine Eigenbeweglichkeit. Ein bindegewebiges Fettpolster bildet ein druckverteilendes Fußsohlen-Polster. er menschliche Fuß ist stärker umkonstruiert worden als die Hand, so dass er sich vom handähnlichen Fuß der Menschenaffen deutlich unterscheidet. Das Laufen verlangt eine hohe Stabi­ lität in den mechanischen Verbindungen der Beine und erfordert wegen der stereotypen Bewegungsabfolge eine Einschränkung der Bewegungsfreiheitsgrade. Im Unterschied zu den meisten guten Läufern ist der Mensch kein Zehen-, sondern ein Sohlengänger, der mit der Ferse aufsetzt, den verlängerten Fuß abrollt und sich mit dem verstärkten Großzehenstrahl abstößt. Die rechtwinklig abgeknickte Fußsohle Schienbein D Achillessehne Abb. 1 Beim Laufen speichern lange Sehnenverbindungen wie die Achillessehne einen Teil der Bewegungsenergie, die beim Bewegen wieder freigesetzt wird. Die Energiebilanz verbessert sich dadurch zwar nicht so stark wie bei guten Läufern (Huftieren, Straußenvögeln), doch erhöht sie die Ausdauer. So konnte der frühe Mensch auch schnellere Beute tagelang jagen und zur Strecke bringen. Querschnitt durch das Fußgewölbe Abb. 3 Abb. 1 Abrollbewegung beim Gehen Foto: Dr. med. Hans Zollinger, Zürich Last Hubkraft Abb. 2 Hebelkräfte beim Gehen aus: Liem (2001) Der Vergleich mit einer Schubkarre zeigt die Hebelwirkung beim Abstoßen des Fußes. Drehpunkt Abb. 3 Fußgewölbe aus: Franzen (1997) Elastische Sehnenverbindungen speichern beim Gehen einen Großteil der Bewegungsenergie und verbessern so die Effizienz. Kraftarm Lastarm Körpergewicht Muskel-Zugkraft Wadenmuskel Schienbein Drehpunkt Lastarm Kraftarm Abb. 2 Die Hand Bei Menschenaffen und Vormenschen ist diese Gelenkfuge noch gerade, und die Finger sind verlängert und gekrümmt, um besser Äste umfassen zu können. Ihre Hände sind auf den Kraftgriff spezialisiert, wie auch die starken Ansätze der Fingersehnen zeigen. Dagegen sind die tastempfind­ lichen Fingerspitzen kleiner als beim Homo sapiens. Die menschlichen Merk­ male sind schon beim Homo habilis, dem ersten Werkzeughersteller, fest­ stellbar. Bis zum Neandertaler waren die Muskelansätze allerdings kräftiger als beim Homo sapiens. Offenbar besteht zwischen der Anatomie der Hand und dem Werkzeug­gebrauch ein evolutionärer Zusammenhang. Abb. 1 D ie Hand des Menschen ist mit ihren relativ kurzen Fingern und dem beweglichen verlängerten Daumen als universelles Greiforgan ausgebildet. Entwicklungsgeschichtlich haben sich Arm und Hand weniger verändert als der Fuß, da die Bewegungsabläufe ähnlich geblieben sind, auch wenn sie für die Fortbewegung keine Rolle mehr spielen. Wie alle Primaten können Menschen greifen und tragen, aber sie haben eine wesentlich bessere Feinmotorik der Hand als diese. Anatomisch zeigt sich das besonders an Daumen und kleinem Finger, die beide zur Handfläche hin gekrümmt werden können. Die Gelenkbasis des Mittelfingers ist schräg gestellt, um ihn gegenüber dem Daumen zu stabilisieren. Abb. 3 Abb. 1 Sehnen und Muskeln des Handrückens aus: Toldt (1911) Die Finger werden von der Unterarmmuskulatur über lange Sehnenzüge bewegt, so dass die Hand kraftvoll zugreifen kann. Die eigentlichen Handmuskeln besorgen nur die Feineinstellung. Besonders ausgeprägt ist die Muskulatur des Daumens, da sie beim Präzisionsgriff unerlässlich ist. Abb. 2 Anatomie des Präzisionsgriffs aus: Benninghoff (1997) Die Abbildung zeigt die Strecker- und Beuger-Sehne des Zeigefingers (weiß und grün) und die beteiligte Handmuskulatur. Abb. 3 Kraftgriff und Präzisionsgriff aus: Andreose (1978) Abb. 2 Tasten, Greifen und Begreifen (150 pro Fingerbeere) zurück, mit denen die Oberflächen-Textur mit einer ReizAuflösung ab 2 mm erfühlt wird. Hier sind auch die Rezeptoren für Tempe­ ratur-, Druck-, Schmerz- und Vibrations-Wahrnehmungen konzentriert. Die Hände sind nur das Werkzeug für die manuellen Handlungen, die vom Gehirn erfasst und gesteuert werden. Die überragende Bedeutung der Hand zeigen die überproportional vergrößerten Projektionsfelder für Sensorik und Motorik in der Großhirnrinde. Der „Homunculus“ mit seinen riesigen Händen visualisiert diese Repräsentanz der Handfunktionen im menschlichen Gehirn. Abb. 1 D ie menschliche Hand kann sowohl zupacken und halten (Kraftgriff) als auch subtil tasten und fassen (Präzisionsgriff). Sie bewältigt das Einfädeln in ein Nadelöhr ebenso wie das Heben schwerer Lasten. Ihre motorischen Fähigkeiten sind unübertroffen. Mit Händen Erkunden, Ergreifen führt zum Begreifen. Die Muskelkraft wird mit Sehnen­ zügen vom Unterarm auf die Finger übertragen. Das sprichwörtliche „Fingerspitzengefühl“ geht auf die in den Fingerkuppen besonders zahl­ reichen Meissner-Tastkörperchen Abb. 2 Abb. 1 Meissner-Tastkörperchen aus: Krstic (1988) Die unmittelbar unter der Oberhaut gelegenen Meissner-Tastkörperchen stehen über Faserverbindungen mit den Tastleisten der Hand in Kontakt und sorgen für die große Berührungsempfindlichkeit der Fingerspitzen und aller anderen Hautbereiche, die mit Hautleisten ausge­stattet sind. Abb. 2 Sensorischer Homunculus aus: Kell et. al. (2005) Die symbolische Darstellung der sensorischen und motorischen Felder der Großhirnrinde verdeutlicht die große Bedeutung der Hand. Schädel und Gebiss Auch ein größerer Anteil tierischer Nahrung begünstigt diese Rück­ bildung, wie man an den lebenden Menschenaffen gut sehen kann. Orang-Utans und Gorillas, die fast nur harte pflanzliche Nahrung zu sich nehmen, besitzen riesige Backenzähne sowie Schädelkämme und Über­ augen­wülste zum Abfangen der hohen Kaudrücke. Abb. 1 I m Laufe der Evolution des Menschen haben sich Schädel und vor allem Gebiss stark verändert. Zeigen Aus­ tralopithecus und Paranthropus noch große Kiefer, stark gebaute Zähne, ausgeprägte Überaugenwülste und Schädelkämme, bilden sich diese Formen auf dem Weg zum Homo sapiens schrittweise zurück. Schimpansen und Menschen als „Allesfresser“ zeigen solche Merkmale kaum. Während Gorillas und Orang-Utans große Kauflächen für ihre Pflanzenkost brauchen, haben die Menschen deutlich kleinere Backenund Mahlzähne wie auch kleinere an Schneidezähne erinnernde Eckzähne. Scheitelkamm Schläfenmuskel ausladender Jochbogen Grund dafür ist der einsetzende Werkzeuggebrauch. Die Nahrung kann immer besser aufbereitet werden, es sind also keine so hohen Kaudrücke mehr nötig. Schädelkämme und große Jochbögen als Ansatz für kräftige Kaumuskeln werden über­ flüssig. Backenmuskel vergrößerte Backenzähne Abb. 3 Abb. 1 Oberkiefer bei Schimpanse, Australopithecus und Mensch obere Reihe aus: Lewin (1991) untere Reihe aus: British Museum: Our fossil relatives (1983) Im Vergleich zu Menschenaffen und Vormenschen hat der Mensch kleine Zähne. Sein Kiefer ist parabelförmig, nicht parallel. Die Eckzähne ähneln Schneidezähnen, wodurch die Affenlücke (s. Pfeile) entfällt. Da die Kieferbewegung nicht mehr durch große Eckzähne eingeschränkt wird, lassen sich auch mahlende Querbewegungen ausführen. Abb. 2 Kräfte beim Kauen aus: Franzen (1997) Abb. 2 Wie das Beispiel des Pekingmenschen (Homo erectus pekinensis) illustriert, entspricht die Schädelform dem Verlauf der Kaudrücke, die hier durch Pfeile symbolisiert sind. Abb. 3 Kauapparat des Paranthropus aus: Butcher (2006) Hominiden mit starkem Gebiss wie Paranthropus benötigen kräftige Knochenwülste über den Augen, die den Schädel vor starken Kaudrücken schützen. Der Schädelkamm auf der Kopfoberseite bildet sich als Ansatz für die Schläfenmuskeln. Gehirnentwicklung Pan troglodytes Australopithecus africanus Homo habilis Homo erectus Homo neanderthalensis Homo sapiens Abb. 1 Für diese Entwicklung gibt es verschiedene Erklärungsversuche: Nach der älteren „technologischen“ Hypo­ these entstand aus der Freisetzung der Hände durch den aufrechten Gang ein Selektionsdruck in Richtung Gehirnvergrößerung. Die „soziale“ Hypothese sieht in der zunehmenden Gruppengröße mit dem Zwang zu intensiverer Kommunikation und Kooperation den Auslöser für die Gehirnentwicklung. Als weitere Voraussetzung wird die verbesserte Ernäh­rung ins Feld geführt, die auch komplexere Sozialgefüge ermöglichte. D ie Gehirngröße der Vormenschen (Australopithecus) lag mit 420 – 520 cm³ noch im Bereich der Menschenaffen. Erst mit der Werkzeugherstellung vor 2,5 Mio. Jahren begann ein schubweises Wachstum der Gehirngröße: von 680 – 800 cm³ beim Homo habilis steigerte sich das Volumen bis zum Homo heidelbergensis auf 1000 – 1200 cm³. Mit über 1600 cm³ wird dann beim Neandertaler das Maximum erreicht. Beim modernen Menschen liegt das Gehirnvolumen dagegen bei ca. 1400 cm³. Wie der Vergleich mit dem Neandertaler zeigt, ist die Leistungsfähigkeit des Gehirns nicht nur von seiner absoluten bzw. relativen Größe (Cerebralisation) abhängig, sondern auch von seiner Feinstruktur (Ence­ phalisation, z. B. Furchung der Großhirnrinde, Zunahme der Neuronen und deren Verschaltungen). durchschnittliches Gehirnvolumen in Kubikzentimetern 1 600 H. neanderthalensis Gattung Australopithecus Gattung Paranthropus Gattung Homo Gattung unklar H. sapiens 1 200 1 000 H o o m r H. ergaste 800 e e r c u t s ergensis H. heidelb H. rudolfensis A. (H.?) habilis 600 P. robustus P. boisei 400 A. africanus A. afarensis 4 Abb. 1 Schädelformen der Hominiden aus: Zimmer (2005) Bereits an den Schädelformen ist die Vergrößerung des Gehirns erkennbar, die mit einer gleichzeitigen Verkleinerung des Kieferapparats einhergeht. Abb. 2 Gehirnvolumen von Vormenschen und Menschen aus: Bild der Wissenschaft, Juli 2009 Im Unterschied zu den nichtmenschlichen Hominiden verdoppelte sich das Gehirnvolumen bei Homo erectus innerhalb von ca. 1,6 Mio. Jahren. In den letzten 300 000 Jahren kam es zu einer sprunghaften Entwicklung zur heutigen Größe. Homo erectus – der Aufsteiger 1 400 Abb. 3 3 2 Schimpanse 1 Millionen Jahre vor heute Abb. 2 Abb. 3 Primatengehirne aus: Dr. Robert Dahnke, Universität Jena, Psychatrie Die Grafik zeigt nicht nur die Größenverhältnisse der Gehirne, sondern auch die unterschiedlich starke Ausbildung der Hirnrindenareale bei „Altweltaffen“. Werkzeuge der Altsteinzeit Abb. 1 W erkzeuge als „verlängertes Gebiss“ definieren die Gattung Homo, die sich vor rund 2,5 Mio. Jahren aus den frühen Australo­ pithe­cinen Ost- und Südafrikas entwickelte. Die Hand ist dabei der Überträger: Werkzeuge müssen handlich sein. Etwa vor 1,6 Mio. Jahren wurde der Faustkeil erfunden, das erste und zugleich erfolgreichste Universalwerkzeug. Faustkeile gibt es bis an das Ende der Neandertalerzeit. Die jüngsten Exemplare sind etwa 50 000 Jahre alt, wie die Stücke aus Le Moustier (Dordogne), der namengebenden Fundstelle der Moustérien-Kultur. Schon der Neandertaler hat seine Keilmesser in Griffschäfte eingeklebt und Blattspitzen in Speere geschäftet. Mit der Einwanderung des Homo sapiens nach Europa vor etwa 40 000 Jahren steigt die Vielfalt der Werkzeuge an. Neu sind filigrane, in Holzschäfte eingeklebte Messerchen oder Wurfgeräte wie Bumerang und Speerschleuder. Bereits „Chopping tools“ aus der Olduvai-Schlucht (Tansania) zeigen, dass eine Spitze oder Schneidkante die dürftigen Zähne und Krallen des Menschen ersetzt, während die Gegenseite in der Handfläche ruht. Grazile Urmenschen, die Werk­ zeuge herstellten, waren bei der Wahl und Aufbereitung ihrer Nahrung flexibler als pflanzenessende Vormenschen mit robustem Gebiss. So starben die robusten Arten vor etwa 1 Mio. Jahren aus. Abb. 2 Abb. 3 Abb. 1 Faustkeile, Blattspitze, Chopper Abb. 2 Blattspitze, als Speerspitze geschäftet Abb. 3 Abwurfbild Speerschleuder Rechts- und Linkshändigkeit in der Urgeschichte memetisch überlieferten Fähigkei­ ten. Durch Hunderttausende von Jahren einer rechtshändig domi­ nierten Werkzeugtradition wurde dieses Mem im Genpool der Mensch­heit verankert. An Werkzeugen lässt sich Händigkeit erst sehr spät in der Mensch­ werdung nachweisen. Hatten Steinwerkzeuge der Neandertaler noch oft symmetrische Arbeitskanten, so sind erst Stichel und Bohrer des Homo sapiens eindeutig asymmetrisch und meist eindeutig für Rechtshänder hergestellt. Abb. 1 E twa 90% der heutigen Menschen unterschiedlichster Zivilisationen sind Rechtshänder. Das bedeutet, ihre genetisch dominante Werk­ zeughand ist die rechte. „Sind Affen Rechtshänder?” – fragte sich bereits Charles Darwin in einem seiner Notizbücher. Gorillas sind es, Schimpansen sind es nicht … So einfach ist es also nicht! Folgt man jedoch Statistiken über Verletzungen fossiler Menschen, dann war bereits beim Ur­mensch und Neandertaler eine Tendenz zur Rechtshändigkeit angelegt. Vorteil der Spezialisierung auf eine Hand ist die höhere Prä­ zision bei der Ausübung der Hand­ arbeit. Kulturleistungen des Menschen („Meme“) sind Informationen, die nicht über „Gene“ an die nächste Generation weitergegeben werden. Der seit mehr als 2 Mio. Jahren praktizierte Umgang mit Werkzeugen gehört somit zu den Abb. 2 Abb. 3 Abb. 1 Harpunen aus Rengeweih, Repliken vom Typ „spätes Magdalénien bis Mittelsteinzeit”, ca. 14 000 – 8 000 v. Chr. Die Harpune ist ein Zeugnis für das handwerkliche Geschick, das erforderlich ist, um hartes Knochenmaterial so fein zu bearbeiten. Abb. 2 Keilmesser vom Neandertaler-Fundplatz Sesselfelsgrotte (bei Neuessing, Lkr. Kelheim, Bayern) Das Keilmesser liegt gut in der Hand. Da die flache Schneidenseite unten liegt, ist die Rechtshändigkeit des Benutzers sehr wahrscheinlich. Abb. 3 Elfenbeinschnitzen, Stichel des Jungpaläolithikums, 40 000 – 10 000 v. Chr. Experiment von Wulf Hein Die scharfe Kante ist nur von einem Rechtshänder sinnvoll zu benutzen. Prothesen In jüngster Zeit wurden Erfahrungen mit sogenannten „Gehirn-Maschinen“ oder „Gehirn-Computer-Schnitt­ stellen“ gesammelt. Sie erlauben es einem gelähmten Patienten, über die Registrierung seiner mit der Vorstellung einer Handbewegung einher­ gehenden Gehirnaktivität eine Hand­ prothese zu steuern. Abb. 1 P Neue Begriffe, wie „Cyborg“ („cybernetic organism“) und „Ersatzteil­ chirurgie“, reflektieren zwar die viel­ versprechende Entwicklung, lassen aber den bleibenden Verlust eines Körper­teils, das Einzelschicksal, außer Betracht. rothesen als Gliedmaßenersatz haben eine lange Tradition. Legendär sind die „Eiserne Hand“ des Götz von Berlichingen oder die Hakenhand des Käpt’n Hook in „Peter Pan“. Insbesondere die zahllosen Versehrten der Kriege und des Straßenverkehrs stellen eine ständige Herausforderung an die Prothesentechnik dar. Während des Ersten Weltkriegs entwickelte der Chirurg Sauerbruch bewegliche, vom Träger mechanisch steuerbare Prothe­ sen. Seit den 1950/60er Jahren werden bioelektrische Prothesen gebaut, bei denen Muskelströme, die vom Amputationsstumpf abgeleitet werden, kleine Elektromotoren in der Prothese steuern. Inzwischen werden solche Prothesen auch mit Sensoren aus­ gestattet. Endoprothesen werden zum Ersatz erkrankter Gelenke, meist der Hüfte oder des Knies, aber auch kleiner Hand- und Fußgelenke, eingesetzt. Abb. 2 Abb. 3 Abb. 1 Die „Zweite Hand“ aus Florenz, 15. – 16. Jahrhundert Foto: Museum Kulturgeschichte der Hand, Wolnzach Vier Finger der Hand können durch eine Druckvorrichtung gemeinsam bewegt werden, während der ebenfalls angewinkelte Daumen starr ist. Die Mechanik ermöglicht einen Zangengriff. Abb. 2 Krukenberghand, 1951/52 und Sauerbruch­ prothese, um 1940 aus: Prothesen von Kopf bis Fuß. Ausstellungskatalog 2003 Die Krukenberghand (links), ein beweglicher Greifarm mit einer Zange, wurde während des Ersten Weltkriegs für beidseitig Unterarmamputierte entwickelt. Die nach dem Chirurgen Ferdinand Sauerbruch benannte Handprothese (rechts) besitzt einen doppelten Seilzug, von denen der eine zum Beugen, der andere zum Strecken der Finger benutzt wird. Abb. 3 Bioelektrische, sensorgesteuerte Handprothese der Fa. Otto Bock aus: Myobock-Armprothesen. Firmenkatalog 2007 Der in Daumen und Zeigefinger der Prothese eingepasste Greifmechanismus wird durch am Oberarm des Trägers abgeleitete Muskelströme angetrieben und über Sensoren in der Prothese dosiert gesteuert. Hand in Hand: der HandChirurg als Hand-Heiler die Erfindungsgabe und manuelle Geschicklichkeit des Hand-Chirurgen. So ist die moderne Handchirurgie ein interdisziplinäres Fach, welches feinstes Chirurgen-Handwerk mit Zellbiologie, Neurologie, Werkstoffwissenschaft und Medizintechnik vereint zum Wohle des Patienten. Abb. 1 V erletzungen der Hand oder Läh­mun­gen ihrer Muskeln zählen zu den folgenschwersten Erkrankungen, da sie den Patienten im privaten Alltag und in seiner Berufsausübung oft massiv beeinträchtigen. Ihre Behandlung ist die Domäne des Handchirurgen, der sie heute mehr und mehr in speziellen Kliniken oder Zentren ausübt. Abb. 2 Abb. 3 Entscheidend für den Behandlungs­ erfolg sind die exakte Kenntnis der Anatomie von Skelett, Muskulatur, Blut- und Nervenversorgung der Hand, die Verfügbarkeit subtiler, oft mikros­kopischer Operationsmethoden und des zugehörigen Instrumentariums, das Verständnis plastischer und regene­rativer Vorgänge in den Geweben der Hand und im Nervensystem – und natürlich vor allem Abb. 4 Abb. 1 Handchirurgen bei der Arbeit am Operations­ mikroskop Abb. 2 Operationsergebnis nach unfallbedingter Abtrennung der linken Hand im Bereich des Unterarms Die durchtrennten Skelettteile werden durch einen äußeren Halteapparat (sog. Fixateur externe) stabilisiert, bis die Heilung erfolgt. Abb. 3 Die Patientin mehrere Monate nach erfolgreicher Operation und Anschluss-Heilbehandlung Form und Funktion der Hand sind beide wiedergewonnen! Abb. 4 Hand in Hand: der Handchirurg und sein Patient Fotos: Plastisch- und Handchirurgische Klinik, Klinikum der Universität Erlangen-Nürnberg, Direktor: Prof. Dr. med. Raymund Horch Ontogenese von Hand und Fuß Das Fingerleistenmuster entsteht in der 19. SW. Wie andere Primaten besitzen Neugeborene noch einen Greifreflex zum Festhalten an der Mutter. Abb. 1 Der Fuß ist bei der Geburt noch ein sehr affenähnlich gestalteter „Platt­ fuß“ mit seitlich abstehender Greif­ zehe. Der Umbau zum Lauffuß wird in den ersten beiden Lebensjahren mit der Ausbildung der Fußgewölbe und der Parallelstellung des Großzehenstrahls abgeschlossen. Die endgültige Ausformung der Fußkonstruktion hängt sehr stark von äußeren Faktoren ab. Optimal wäre ständiges BarfußGehen. Unsere heutige schmale, lange Fußform geht auf zu enges Schuhwerk zurück. B eim Embryo bildet sich zunächst der Kopf mit der Kiemenregion aus, danach entstehen die Arme aus Abfaltungen der Hals- und Brustregion. In der 5. Schwangerschaftswoche (SW) erscheinen paddelförmige Hand­ anlagen, die Füße folgen in der 7. SW. Die Hände gehen den Füßen voraus, da der Kopf des Embryos wegen des stark wachsenden Gehirns besser mit Blut und Nährstoffen versorgt wird als der Rest. Die Finger werden durch „Apoptose“ – programmierten Zelltod – in den Zwischenräumen während der 5. bis 6. SW freigestellt, wobei sie anfangs durch „Schwimmhäute“ verbunden bleiben. Erste aktive Bewegungen sind in der 13. SW zu erkennen. Abb. 2 Abb. 1 Embryo, Abfaltung der Extremitäten aus: Rohen/Lütjen-Drecoll (2006) Die vom Rückenmark her einwachsenden Arm- und Handnerven zeigen die Entstehung der Hand aus Auswüchsen der Hals- und Brustregion. Abb. 2 Vergleich Neonatus Mensch – Rhesus aus: Schulz (1974) Beim Neugeborenen (links) ist der Fuß, wie bei anderen Primaten, anfangs noch als Greiffuß mit abspreizbarer Großzehe ausgebildet. Abb. 3 Entstehung der Hand beim Embryo Rasterelektronenmikroskopische Aufnahmen aus: Hinrichsen (1990) Nach dem Aussprossen der Armknospen mit 26 Tagen bilden sich die paddelförmigen Handtelleranlagen aus. Anschließend werden die Finger über ein flossenartiges Zwischenstadium freigestellt. Nachdem sich die Finger­ strahlen getrennt haben, beginnt das Armwachstum. Abb. 3 Sprechende Hände Künstler haben der Hand seit jeher ihr besonderes Augenmerk geschenkt. Bereits in den frühesten Zeugnissen der Höhlenmalerei finden sich Handdarstellungen als erste Form der bildnerischen Selbstthematisierung des Menschen. Figurative Malerei und Plastik reflektieren bis heute die komplexe Sprache menschlicher Gesten. Auch in der politischen Propaganda spielt die Hand eine wichtige Rolle. Bildzeichen wie der solidarische Handschlag oder die geballte Faust sind zu zentralen Symbolen der politischen Linken geworden. Abb. 1 D ie Hand ist nicht nur unser erstes und unentbehrlichstes Werkzeug, sondern auch unser wichtigstes nonverbales Verständigungsmittel. Mit der „Sprache der Hände“ können wir unsere Empfindungen ausdrücken, Nähe signalisieren und ermuntern, aber auch drohen und zurückweisen oder einfach nur sachliche Informationen übermitteln. Manche unserer Gesten – wie die geballte Faust – sind als archaische Zeichen über unseren Kulturkreis hinaus verständlich. Schließlich spiegelt auch unsere Sprache in zahlreichen Wortbildungen und Redewendungen die immense Kulturbedeutung der Hand. Abb. 3 Eine große Bedeutung haben Gesten in den Religionen. Zum christlichen Ritus gehören die gefalteten Hände der Gläubigen und die segnende Hand des Priesters. In der katholischen Liturgie sind Berührungen wie die Handauflegung ein wichtiger Teil der Sakramentenordnung. Abb. 1 Martha Magdalena Mezger, 1767 Johann Eberhard Ihle (1727 – 1814) Öl auf Leinwand Kunstsammlung der Universität Die Witwe des Nürnberger Goldhändlers Paul Christoph Mezger präsentiert sich reich gekleidet und Schmuck überladen dem Betrachter. In der Linken hält sie das weiße Seidenfutter ihres rosafarbenen Umhangs prätentiös zwischen Daumen und Zeigefinger, um auf dessen Kostbarkeit hinzuweisen. Das Gemälde veranschaulicht so auch die Bedeutung der Hand für die Kunst des Porträts. Abb. 2 Winston Churchill in typischer Pose mit dem Victory-Zeichen Fotografie, um 1941 International bekannt wurde das Victory-Zeichen während des Zweiten Weltkrieges durch Winston Churchill. Mit dieser Geste stärkte der britische Premierminister die Siegeszuversicht und den Widerstandswillen des britischen Volkes gegen die nationalsozialistische Eroberungspolitik. Abb. 3 „Die gefesselten Hände“ Holzskulptur von Hans Gügel Foto: Rudi Stümpel, 1953 Stadtarchiv Abb. 2 Ein Erlanger Beispiel für die politische Symbolik der gefessel­ ten Hand: das Mahnmal zum Gedenken an die nicht aus Russland zurückgekehrten Kriegsgefangenen am Bohlenplatz. Die gefesselten, aber zur Faust geballten Hände bringen wie die Inschrift Protest zum Ausdruck: „Lasst unsere Kriegsgefangenen frei“. Die Skulptur wurde1953 vom „Verband der Heimkehrer“ aufgestellt und 1979 entfernt. Heute befindet sie sich, neu gedeutet, in einer Anlage in Alterlangen. Redensarten in jemandes Hand stehen die Hand über jemanden halten die Oberhand behalten sein Schicksal in die Hand Gottes legen in guten Händen sein jemanden um die Hand seiner Tochter bitten jemanden an die Hand nehmen nicht aus der Hand geben jemandem die Hand reichen etwas in die Hand nehmen sich in der Hand haben unter den Händen zerrinnen freie Hand haben jemandem sind die Hände gebunden etwas aus erster Hand haben in andere Hände übergehen mit leeren Händen dastehen mit warmer Hand schenken zu treuen Händen mit beiden Händen zugreifen die Hände davon lassen sich von seiner Hände Arbeit ernähren beide Hände voll zu tun haben gut von der Hand gehen jemandem zur Hand gehen etwas zur Hand haben eine glückliche Hand haben in die Hände spucken die Hände in den Schoß legen jemanden auf den Händen tragen jemandes rechte Hand sein zwei linke Hände haben eine Politik der starken Hand die öffentliche Hand die unsichtbare Hand des Marktes alle Trümpfe in der Hand haben die Hand im Spiel haben unter der Hand jemandem auf die Hände sehen von der Hand in den Mund leben jemandem aus der Hand fressen an der ausgestreckten Hand ver­ hungern lassen jemandem in die Hände fallen Hand an sich legen die Hand gegen jemanden erheben für jemanden die Hände ins Feuer legen seine Hände in Unschuld waschen sich die Hände reiben die Hände über dem Kopf zusammenschlagen die Hände ringen im Handumdrehen kurzer Hand von langer Hand Hand in Hand auf der Hand liegen etwas ist mit Händen zu greifen sich mit Händen und Füßen wehren Hand und Fuß haben