Hydrothermalquellen und der Ursprung des Lebens
Werbung

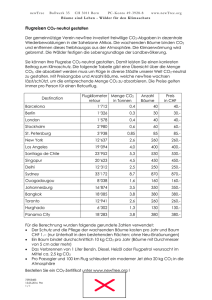
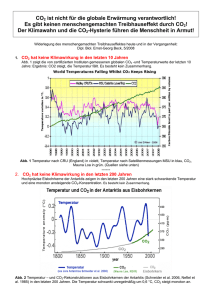
DOI:10.1002/biuz.200910391 Alles hat einen Anfang, auch die Evolution: Hydrothermalquellen und der Ursprung des Lebens W ILLIAM M ARTIN An Tiefsee-Hydrothermalquellen gedeihen ganze Ökosysteme in völliger Finsternis. Am Anfang ihrer Nahrungskette steht nicht die Sonne, sondern chemische Energieträger wie molekularer Wasserstoff, die aus dem Erdinneren strömen. Seit ihrer Entdeckung vor 30 Jahren werden Hydrothermalquellen im Zusammenhang mit dem Ursprung des Lebens diskutiert, weil die Erde dort chemische Energie spendet. Die chemischen Bedingungen dort sind also für die Umwandlung von CO2 in organische Substanzen – die Bausteine des Lebens – förderlich. Stand eine solche Chemie auch am Anfang des Lebens? as Leben ist aus unbelebter Materie entstanden. Wissenschaftliche Theorien für den Ursprung des Lebens müssen daher tragbare und überprüfbare Hypothesen unterbreiten, woher die reduzierten Kohlenstoffverbindungen stammen, aus denen sich das Leben entwickelte,aber auch woher die erforderliche Energie dafür stammte. Beim Thema „Ursprung des Lebens“ denken die meisten Menschen sofort an eine Art „Ursuppe“. Heute ist der Begriff „Ursuppentheorie“ genau soweit verbreitet wie Darwins Evolutionstheorie. Aber es gibt einen entscheidenden Unterschied: Während die Evolutionstheorie nach 150 Jahren in allen wesentlichen Aspekten richtig ist, ist die Ursuppentheorie nach circa 80 Jahren in allen wesentlichen Aspekten überholt. D Weder Ursuppe noch Gärung standen am Anfang Die Vorstellung, das Leben sei aus einer präbiotischen Ursuppe entstanden, welche die Ur-Erde bedeckte, geht etwa auf die Zeit nach der Oktoberrevolution und den russischen Chemiker A. Oparin sowie den britischen Evolutionsbiologen J. B. S. Haldane zurück. Im Wesentlichen besagt die Ursuppentheorie, dass auf der frühen Erde durch die Einwirkung von UV-Strahlung auf CO2, Ammoniak und Wasser die ersten organischen Verbindungen entstanden. Mangels Lebewesen, die diese organischen Moleküle als Nahrung hätten ab166 | Biol. Unserer Zeit | 3/2009 (39) bauen können, reicherten sie sich zu einem ozeanischen Gemisch mit der Konzentration einer dünnen Brühe an. In dieser entstanden dann die ersten Makromoleküle, später dann die ersten Zellen, die mittels Fermentation das Gemisch verzehrten. Als die Ursuppe weg war, wurde die Photosynthese gefordert, und so hat alles angefangen [5]. Mit einem sehr berühmten Experiment zu diesem Thema im Jahre 1953 zeigte Stanley Miller [15], dass aus einem Gemisch von Methan, Ammoniak und Wasserstoff (so hat man sich damals die Uratmosphäre vorgestellt) unter simulierter Blitzeinwirkung einfache organische Moleküle, aber auch wichtige Grundbausteine des Lebens entstehen können. So hat er u.a. Blausäure (HCN), Aldehyde, einfache Aminosäuren, Öl und Teer unter den Reaktionsprodukten gefunden. Spätere Versuche dieser Art führten zur Synthese von weiteren Aminosäuren und Nukleinsäure-Bausteinen wie Adenin und Guanin. Dass Millers erster Versuch zur Synthese einer Ursuppe im Labor prompt gelang, war sensationell. Aber damit diese Bausteine zur weiteren Kondensation und Polymerisierung in Richtung biologische Makromoleküle weiterreagieren, war ein Mechanismus erforderlich, womit sie einkonzentriert werden könnten. Man dachte an austrocknende Gezeitenzonen, Adsorption an Tonerden, die Konzentration im Eis, riesige Ölteppiche, usw. So hat sich im Laufe der Zeit eine Vorstellung verbreitet, wonach die Oparin-Haldane Ursuppentheorie anhand von Miller’s Versuch grundsätzlich richtig sei, lediglich bei den Details gebe es Erklärungsbedarf. Haldanes achtseitiger Aufsatz [5] ist sehr empfehlenswert und bleibt eine Glanzleistung der logischen Ableitung. Bedingt durch den damaligen Stand des Wissens waren jedoch einige seiner Prämissen unwahr. Aus heutiger Sicht gibt es ein unüberwindliches Problem mit der Ursuppentheorie, das man in einem Gedankenexperiment verdeutlichen kann: Wir nehmen eine Dose voller Ursuppe. Dienlich dabei wäre eine Hühnerbrühe, kräftige Gulaschsuppe oder besser noch eine Lösung aus frisch homogenisierten Bakterien, deren Zellstruktur komplett zerlegt wurde (darin befinden sich alle chemischen Bausteine,aus denen das Leben besteht). Wir verschließen die Dosen steril. Wir © 2009 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim HYDROTHERMALQUELLEN | S C H W E R P U N K T E VO LU T I O N S FO R SC H U N G A B B . 1 Ein schwarzer Raucher. Dieser Schlot mit dem Namen „Sully“ liegt im Main Endeavour Field, Endeavour Segment, Juan de Fuca Ridge. Sully liegt bei einer Tiefe von ca. 2200 m. Im Main Endeavour Field sind über 100 aktive Schlote bekannt, teilweise umgeben von Sulfidablagerungen, die einen Durchmesser von 50 m haben und circa 20 m hoch sind. Zum Zeitpunkt dieser Aufnahme (2005) war das austretende Wasser von Sully ca. 360° C heiß und er war Heimat für eine dichte Kolonie von Röhrenwürmern, die ca. 1–1,5 cm im Durchmesser groß sind. Sully war nicht immer so dicht bewachsen. Im Jahre 1999–2000, kurz nach einem Magma-Ausbruch, war Sully gänzlich unbewachsen und führte noch heißeres Wasser. Bild und Informationen zu Sully: J. Delaney und D. Kelley, University of Washington. © 2009 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim www.biuz.de 3/2009 (39) | Biol. Unserer Zeit | 167 stellen die Dosen bei beliebiger Temperatur hin und warten beliebig lang.Wir entnehmen in Abständen Proben, um zu sehen, ob darin neues Leben entstanden ist. Es ist aus heutiger Sicht intuitiv verständlich, dass sich in diesen Dosen voller perfekter Ursuppe unter gar keinen Umständen neues Leben bilden wird. Schwieriger wird es jedoch mit der Frage, warum dort kein neues Leben entsteht. Der Grund ist, dass sich der Kohlenstoff, der Stickstoff, der Sauerstoff und der Wasserstoff, aus denen unsere Ursuppe besteht, im chemischen Gleichgewicht befinden; d.h., der chemische Inhalt dieser Dosen voller Ursuppe hat schon reagiert, er hat kein chemisches Potenzial um weiterzureagieren, weil er sich im – oder sehr nah am – Gleichgewicht befindet. Lebende Systeme befinden sich stets weit ab vom Gleichgewicht. Alle Lebensformen müssen sich ständig ernähren, ihren Stoffwechsel „anfeuern“, um ihre bioenergetische Hauptreaktion aufrecht zu erhalten, und sich somit vom chemischen Gleichgewicht fernzuhalten. Man kann die Dosen auch gerne mit Blitzeinschlag behandeln, um etwas gegen den Gleichgewichtszustand zu unternehmen, aber die Situation bessert sich nicht: bei Blitzeinwirkung erhöht sich die Temperatur des Doseninhalts binnen Bruchteilen einer Sekunde auf Temperaturen, die Sand zu Glas verschmelzen lassen, und die organischen Substanzen verwandeln sich dabei nicht in Leben, sondern in CO2. Das Leben ist eine chemische Reaktion, eine überaus komplizierte, aber letztendlich eine chemische Reaktion, bei der Energie stufenweise frei wird. Bei allen Lebensformen gibt es eine zentrale Hauptreaktion im Stoffwechsel, aus der die Energie für das Leben gewonnen wird.Beim Menschen ist die Hauptreaktion die Verbrennung von Zuckern in unseren Mitochondrien, den Kraftwerken unserer Zellen. Bei allen Lebensformen, ohne Ausnahme, gibt es eine solche Hauptreaktion, die als energetische Triebfeder für alle anderen Lebensvorgänge dient. Bei Pflanzen ist es die Photosynthese, die Nutzung der Sonnenenergie für die Bildung von Zuckern aus CO2. Bei der Gärung von fermentierenden Zellen, wie Hefe, ist die Hauptreaktion die Zerlegung von Zucker in CO2 und Ethanol – eine Disproportionierungs-Reaktion, wobei der Kohlenstoff im Zucker in einen höheren und einen niedrigeren Oxidationszustand versetzt wird. Bei der Gärung wird auch Energie frei (die Sonnenenergie, die im Zucker gespeichert ist). Meistens ist es so, dass aus der physiologischen Hauptreaktion zugleich die wichtigsten Grundbausteine für Biosynthesen im Stoffwechsel bereitgestellt werden. Auch die Vorstellung, dass die Gärung die ursprünglichste Form der Energiegewinnung war,ist heute noch sehr weit verbreitet. Aber genau wie die Ursuppentheorie ist auch diese Vorstellung untauglich. Warum? Wenn man Gärung als energetische Urreaktion am Anfang des Lebens diskutiert, so muss man auch fragen 168 | Biol. Unserer Zeit | 3/2009 (39) www.biuz.de „Und woher kam der Zucker?“ – und auf diese Frage gibt es keine befriedigende Antwort. Die Geologen, die sich heute mit der Ur-Erde befassen, sagen uns, dass die Meere anfangs (vor circa vier Milliarden Jahren) etwa 10 km tief waren, es gab gar keine Landmasse, wo sich ein „warmer Tümpel“ wie es Darwin einst formulierte, hätte bilden können, es gab ständig Kometen- und Meteoriten-Einschläge und es gab keinen Sauerstoff zum Atmen [18]. Die Ur-Erde war aus unserer heutigen Sicht ein sehr unangenehmer Ort, ganz ungeeignet für das Leben, so wie wir es in unserer alltäglichen Erfahrung kennen. Der springende Punkt dabei ist „unsere alltägliche Erfahrung“, weil es durchaus viele heutige Lebensformen gibt, die sich auf einer solchen Ur-Erde rundum wohl gefühlt hätten,und sie hätten sogar ein sprudelndes Überangebot an Nährstoffen gehabt, weil ihre Nährstoffe sehr einfache chemische Verbindungen sind: H2 und CO2, die es auf der Ur-Erde nach dem heutigem Stand des Wissens reichlich gab [18]. Sie heißen Chemolithoautotrophe. Sowohl bei den Archaebakterien als auch bei den Eubakterien gibt es solche Organismen, die bei ihrer Hauptreaktion während der Energiegewinnung H2 und CO2 als Substrat benutzen. Methanogene (Archaebakterien) bilden daraus Methan [21], Acetogene (Eubakterien) bilden daraus Essigsäure [20]. Weder Methanogene noch Acetogene sind Fermentierer. Stattdessen benutzen sie einen fast universellen Mechanismus der Energiegewinnung, die Chemiosmose: Darunter versteht man den Aufbau und die Nutzung von Ionen-Gradienten zur Gewinnung chemisch nutzbarer Energie in der Form von ATP. Chemiosmose Heute ist die universelle Währung chemischer Energie unter allen Lebensformen Adenosintriphosphat (ATP). Es gibt nur zwei Grundmechanismen zur Synthese von ATP zwecks Gewinnung (genauer gesagt,Speicherung) von chemischer Energie zur Deckung des zellulären Energiehaushalts: Substratstufenphosphorylierung und Chemiosmose. Unter Substratstufenphosphorylierung versteht man die direkte enzymatische Phosphorylierung von ADP durch die Überführung eines energiereichen Substrats in ein energieärmeres Reaktionsprodukt. Ein Teil der freiwerdenden Energie aus der Reaktion bleibt in der chemisch nutzbaren Währung von ATP gespeichert. Das klassische Beispiel für Substratstufenphosphorylierung findet man in der Glykolyse: In mehreren enzymatischen Schritten wird bei diesem wichtigsten Abbauweg der Kohlenhydrate ein „aktivierter“ Phosphatrest auf ADP übertragen, um ATP zu bilden, und 3-Phospho-D-Glycerat wird freigesetzt. Es gibt viele enzymatische Variationen der Substratstufenphosphorylierung, aber das Prinzip bleibt gleich: die in einer energiereichen Bindung eines Substratmo© 2009 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim HYDROTHERMALQUELLEN leküls vorhandene chemische Energie wird durch ein Enzym direkt genutzt, um ATP als Endprodukt der katalysierten Reaktion zu bilden. Der andere Mechanismus zur Konservierung von Energie in der Form von ATP heißt Chemiosmose, so benannt in 1961 durch seinen Entdecker Peter Mitchell [16]. Unter Chemisomose versteht man den Aufbau eines Gradienten von Protonen (oder auch anderer Ionen, beispielsweise Na+) über die Cytoplasma-Membran, und die Nutzung der in jenem Gradienten gespeicherten Energie zur Phosphorylierung von ADP zu ATP. Der Protonengradient kann entweder mit Hilfe des Lichts (Phototrophie) oder durch die Kopplung von einer Elektronenübertragung vom Donor zum Akzeptor (eine Redox-Reaktion, Chemotrophie) mit dem Pumpen von Protonen erfolgen. Bei der Redox-Reaktion werden Elektronen von einem Elektronen-Donor zu einem Elektronen-Akzeptor meist über einige Stationen geleitet. Dies geschieht mit der Hilfe von Membranproteinen, indem Protonen aus dem Inneren der Zelle hinaus gepumpt werden; der Elektronentransport und das Pumpen von Protonen sind gekoppelt. Der so aufgebaute Gradient stellt eine Ladungstrennung und zugleich potenzielle Energie dar, der vom Prinzip her genau so genutzt wird, wie das Wasser in einem Staudamm. Wie das Wasser in einem Staudamm talabwärts abfließen „will“, so „wollen“ die aufgestauten Protonen zurück in das Cytoplasma fließen. Der Wiedereintritt der Protonen in die Zelle erfolgt nicht wahllos,sondern durch ganz bestimmte Proteine, ATP-Synthasen oder Kopplungsfaktoren genannt, die erstaunlich ähnlich wie die Turbinen in einem Wasserkraftwerk arbeiten. Der Flux von wiedereintretenden Protonen bringt das Herzstück der ATP-Synthase zum Rotieren (wörtlich,es dreht sich wie das Blattwerk einer Turbine). Diese Rotationsenergie wird auf einen anderen Bestandteil der ATP-Synthase übertragen, der die energiereiche Bindung zwischen ADP und Phosphat knüpft, um das ATP zu bilden. Bei Fermentierern ist die Substratstufenphosphorylierung der einzige Weg zur Synthese von ATP. Obwohl die Fermentation als Weg der Energiegewinnung allgemein besser bekannt ist, vermutlich weil sehr viel einfacher zu erklären,ist sie eigentlich eher die Ausnahme, wenn man das volle Spektrum der bekannten Organismen betrachtet. Betrachtet man alle Lebewesen, ist Chemiosmose die Norm. Und weil alle Fermentierer letztendlich von Primärproduzenten wie Pflanzen, Algen oder CO2-fixierenden Prokaryoten abhängig sind, die letztendlich alle Substrate zur Fermentation liefern, können die ersten Organismen eigentlich kaum Fermentierer gewesen sein. Anders ausgedrückt: die Fermentierer können erst dann entstanden sein, als Autotrophe bereits etwas zu fermentieren gebildet haben, und alle Autotrophe nutzen das Prinzip der Chemiosmose.Was sind die Alternativen? © 2009 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim | S C H W E R P U N K T E VO LU T I O N S FO R SC H U N G Zwei Typen von Hydrothermalquellen Hydrothermalquellen bieten in vielerlei Hinsicht eine konzeptionelle Alternative zur Ursuppe. Sie katalysieren sogar chemische Reaktionen, die auffällige Ähnlichkeiten mit den bioenergetischen Hauptreaktionen mancher Mikrooganismen zeigen. Die zuerst entdeckten Hydrothermalquellen wurden als „schwarze Raucher“ bezeichnet. Sie beherbergten hochdiversifizierte Ökosysteme,deren Energiequelle größtenteils vom Vulkanismus am Meeresboden stammt. Ihre Schlote führten 360° C heißes Wasser und ihre Sulfid-Schornsteine boten das Bild eines Urhabitats auf der frühen Erde, mit reaktiven Gasen, gelösten Übergangsmetallen und thermischen sowie chemischen Gradienten. Bald nach ihrer Entdeckung wurden sie vom amerikanischen Biologen John Baross konkret in Zusammenhang mit dem Ursprung des Lebens diskutiert [4], weil diese Systeme geochemisch äußerst reaktive Bedingungen bieten, eine wichtige Voraussetzung für präbiotische Synthesen. Anhänger der Ursuppentheorie blieben aber extrem skeptisch, dass Hydrothermalsysteme für den Ursprung des Lebens überhaupt relevant sein könnten [3]. Immerhin ist das Wasser an der Austrittstelle von schwarzen Rauchern bis zu 400° C heiß, während der Temperaturrekord für mikrobielles Wachstum „nur“ bei 121° C liegt.Erst im Jahre 2000 wurde von Deborah Kelley und ihren Kollegen jedoch ein völlig neuer Typ von Tiefsee-Hydrothermalquellen entdeckt, dessen austretendes Wasser eine Temperatur um die 40° C bis 90° C besaß [9].Der erste Vertreter dieser Klasse wurde „Lost City“ genannt. Systeme wie Lost City haben sehr günstige Eigenschaften, wenn es um den Ursprung des Lebens geht. Die Hydrothermalsysteme vom Typ schwarzer Raucher (Abbildung 1) und vom Typ Lost City (Abbildung 2) sind geochemisch sehr verschieden, wie der nachfolgende Kurzvergleich zeigt. Schwarze Raucher Die bisher bekannten schwarzen Raucher (engl: black smokers), liegen in der Regel direkt über den Spreizungszonen am mittelozeanischen Rücken. Dort, wo die Kontinente auseinanderdriften, sitzt circa 1–3 km unterhalb des Meeresbodens eine Magmakammer. Die Schlote der schwarzen Raucher führen extrem heißes (bis zu ~400° C), chemisch modifiziertes Meerwasser, das durch die Erdkruste zirkuliert [1]. Durch Risse im Meeresboden in der Nähe der Spreizungszone fließt kaltes Meerwasser kilometerweit in die Kruste hinab, wo es in unmittelbare Nähe der Magmakammer (circa 800–1200° C) gelangt. Das erhitzte Wasser steigt dann in einem Konvektionsstrom auf und tritt am schwarzen Raucher wieder ins Meer ein. Das austretende Wasser an schwarzen Rauchern ist typischerweise sehr sauer (pH 2–3) und führt große Mengen gelöster Metallionen wie Fe(II) and Mn(II) sowie Wasserstoff (H2: 0.1– 50 mmol·kg–1) mit. www.biuz.de 3/2009 (39) | Biol. Unserer Zeit | 169 Das Hydrothermalwasser fließt in unmittelbarer Nähe der Magmakammer, daher sind hohe Konzentrationen von Kohlendioxid (CO2: 4–215 mmol·kg–1) und Schwefelwasserstoff (H2S: 3–110 mmol·kg–1) üblich. Unterschiedliche Mengen an Methan (CH4: 0,05–4.5 mmol·kg–1) noch ungeklärten Ursprungs wird auch im Wasser von schwarzen Rauchern gefunden. Die gelösten Gase (H2, CO2, H2S) bilden die Lebensgrundlage von mikrobiellen Gemeinschaften, die am Anfang der Nahrungskette der Ökosysteme bei schwarzen Rauchern stehen [10]. Man kennt auch fossile schwarze Raucher, die bis zu 3,2 Milliarden Jahre alt sind. steigende Meerwasser auf etwa 150–200° C erhitzt, beim Wiedereintritt ins Meer beträgt seine Temperatur etwa 40–90° C. Ein weiterer wichtiger Unterschied ist, dass das Hydrothermalwasser bei Lost City durch Gestein fließt, das sehr viel Mg2+ und Fe2+ und sehr wenig Kieselsäure enthält. Die Folge ist, dass das austretende Wasser bei Lost City sehr alkalisch ist (pH 9–11). Auf den alkalischen pH-Wert kommen wir am Ende dieses Beitrags zurück. Im Gegensatz zu den schwarzen Rauchern, die eher kurzlebig sind, ist Lost City seit circa 100.000 Jahren aktiv. Das Austrittswasser bei Lost City enthält sehr viel H2, CH4, sowie andere kurzkettige (bis C5) Kohlenwasserstoffe,aber so gut wie kein CO2 [17]. Lost City Ein sehr wichtiger Unterschied zwischen dem Lost City-System und den schwarzen Rauchern ist, dass Lost City einige Kilometer von der Spreizungszone entfernt liegt – mit der Folge, dass das Hydrothermalwasser zwar auch kilometertief durch die Kruste in einem Konvektionsstrom zirkuliert, aber nicht in Kontakt mit einer Magmakammer gerät. In der Kruste wird das ab- Serpentinisierung Die Frage nach der Herkunft von Methan, Wasserstoff und den kurzen Kohlenwasserstoffen im austretenden Hydrothermalwasser von Lost City führt uns direkt zu einem überaus wichtigen und interessanten, aber bei Biologen (noch) kaum bekannten Prozess, der für den Ursprung des Lebens hochrelevant sein könnte:die Ser- A B B . 2 Das Bild zeigt die Spitze eines der Schlote bei Lost City, aufgenommen mit dem Tiefsee-Tauchboot „Alvin“ im Jahr 2003. Die fingerähnlichen Carbonat-Ablagerungen sind Heimat für dichtbewachsene Kolonien von Eubakterien und Lost City-Methanosarcinales. Der Schlot befindet sich auf der Ostseite von Lost City bei einer Wassertiefe von ca. 750 m. Der Roboterarm von Alvin ist links im Bild zu sehen. Bild und Informationen: D. Kelley, University of Washington. 170 | Biol. Unserer Zeit | 3/2009 (39) www.biuz.de © 2009 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim HYDROTHERMALQUELLEN pentinisierung. Serpentinisierung ist eine geochemische Reaktionensabfolge in der Erdkruste, die durch Hydrothermalsysteme in Gang gesetzt wird [2]. Benannt wird der Prozess nach dem Mineral Serpentinit (Mg2.85Fe0.15Si2O5(OH)4), das dabei gebildet wird. Fasst man das Geschehen auf das Wesentliche zusammen, wird ein Teil des Fe2+ im Olivin (Mg1.8Fe0.2SiO4) durch Wasser zu Fe3+ oxidiert, das dabei zum Bestandteil vom Magnetit (Fe3O4) wird.Ein Teil des Sauerstoffs im Wasser bleibt im Gestein als Eisenoxid zurück, und die vom Fe2+ spendierten Elektronen verlassen das Gestein als H2. Das Entscheidende dabei ist die Freisetzung von H2, der mit dem Hydrothermalwasser an die Oberfläche davongetragen wird [1]. Im Zusammenhang mit dem Ursprung des Lebens ist die H2-Bildung der Serpentinisierung deshalb so wichtig, weil H2 für viele Mikroben, insbesondere für Chemolithoautotrophe, eine lebenswichtige Form von chemischer Energie ist. Serpentinisierung findet auch in den Hydrothermalsystemen unter schwarzen Rauchern statt, aber unser Fokus bleibt bei Lost City, weil diese alkalisch und nicht zu heiß sind. Man geht davon aus, dass die Serpentinisierung genau so alt ist, wie die Ozeane auf der Erde selbst [19]. Anders ausgedrückt, Hydrothermalsysteme vom Lost City-Typ hat es schon immer auf der Erde gegeben. Die globale Dimension von Serpentinisierung ist beachtlich. Ein Kubikmeter Olivin liefert im Laufe der Serpentinisierung etwa 500 Mol H2.Der Löwenanteil der ozeanischen Erdkruste besteht aus Olivin und ähnlichen Mineralien, die zur Serpentinisierung beitragen können. Man schätzt, dass das Volumen der Ozeane etwa alle 100.000 Jahre durch Hydrothermalsysteme fließt, und dass die massenhaften Reserven an Fe2+ in der Erde,die für die Serpentinisierung zur Verfügung stehen, mehr oder weniger unerschöpflich sind [19]. Vor kurzem haben Deborah Kelley und ihre Mitarbeiter mittels Isotopenuntersuchungen gezeigt, dass das Methan im Wasser von Lost City nicht biologischen Ursprungs, sondern geochemischen Ursprungs ist, woraus zu schließen ist, dass das Methan aus der Serpentinisierung stammt [17]. Die geologische Methanbildung bei Lost City und die biologische Methanbildung von Archaebakterien laufen im Wesentlichen gemäß der gleichen Gesamtreaktion ab, lediglich die chemischen Zwischenverbindungen sind verschieden (Abbildung 3). Aber während bei der geologischen Methanbildung die freie Enthalpie der Reaktion zur Gänze als Wärme frei wird, wird bei der biologischen Methanbildung ein Teil der Energie zwischenzeitlich als ATP konserviert. Möglicherweise liefert die geologische Methanbildung bei Lost City wichtige Hinweise auf den Übergang von geochemischen Prozessen zu biochemischer Evolution beim Ursprung des Lebens. Der aus Serpentinisierung stammende H2 reduziert CO2 kontinuierlich und in einer © 2009 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim | S C H W E R P U N K T E VO LU T I O N S FO R SC H U N G geologisch stabilen Umgebung [17], was grundlegend neue Perspektiven in der Frage nach der Herkunft organischer Verbindungen am Anfang des Lebens eröffnet. Hydrothermalquellen und die Chemie ursprünglicher Lebensformen Die CO2-Fixierung steht am Anfang aller Ökosysteme: das Leben besteht aus reduzierten Kohlenstoffverbindungen, die alle letztendlich aus CO2 stammen. Heute gibt es nur zwei Gruppen von Organismen, die CO2 in den globalen biologischen Kohlenstoff-Kreislauf hineinbringen: Photoautotrophe und Chemoautotrophe. Erstere nutzen die Energie des Sonnenlichts, um CO2 zu fixieren. Letztere nutzen rein chemische Energie, beispielsweise in der Form von H2, der als zentraler Elektronen-Donor im Stoffwechsel vieler Prokaryoten dient.Wenn wir Gärungen am Anfang des Lebens ausschließen können, formulieren wir die Frage lieber so: Welcher ist unter den heutigen Stoffwechselwegen zur CO2-Fixierung der Ursprünglichste? Es gibt nur fünf bekannte Stoffwechselwege der CO2-Fixierung. Darunter ist der Acetyl-CoA-Weg, auch Wood-Ljungdahl-Weg genannt, der einzige Weg der CO2-Fixierung, bei dem Zellen Energie in der Form von ATP gewinnen; bei allen anderen Wegen der CO2-Fixierung müssen Zellen Energie investieren. Abbildung 3 zeigt diesen Weg in vereinfachter Darstellung. Eine wichtige Rolle spielt dabei das interessante, bifunktionelle Enzym Kohlenmonoxid Dehydrogenase/AcetylCoA-Synthase (CODH/ACS). Wichtige Unterstützung für die These, dass der Acetyl-CoA-Weg ein sehr primitiver Stoffwechselweg ist, kommt aus dem chemischen Labor. Einige der wesentlichen Grundreaktionen des Acetyl-CoA-Weges laufen im Reagenzglas ohne Enzyme ab [6, 7]. Solche Versuche zeigen, dass unter chemischen Bedingungen, wie sie an manchen Hydrothermalquellen existieren, Reaktionen, wie sie im AcetylCoA-Weg ablaufen, grundsätzlich möglich sind. Man darf jedoch nicht darüber hinwegsehen, dass zwischen solchen einfachen chemischen Reaktionen in Hydrothermalquellen und der Vorstellung einer „RNA-Welt“ [8], d.h. einer Synthese und Polymerisierung der Grundbausteine der Nukleinsäuren zu selbstreplizierenden und katalytisch aktiven Molekülen, noch eine große Lücke klafft. Aber auch für die Synthese der Bausteine in einer RNA-Welt waren Energie und eine chemisch reaktive Umgebung erforderlich. In Hydrothermalquellen, wo H2 aus der Serpentinisierung und CO2 im Meer aufeinandertreffen, wäre für das energetische Problem prinzipiell eine Lösung in Sicht. Hydrothermalquellen und Chemiosmose Der dienstälteste Verfechter der These, dass das Leben an Hydrothermalquellen entstand, der Geochemiker Michael J. Russell, hat stets betont, dass auf der Urerde www.biuz.de 3/2009 (39) | Biol. Unserer Zeit | 171 ABB. 3 a) b) Acetogenese Methanogenese +H2 +MF +3.5 –MF +H4MPT Formyl-H4F CODH/ACS +6 CO2 Methenyl-H4F +H2 +H2 –H2O –23 Acetat –H4F * +H2O –HSCoA +SCoA– –Ni[E] CH3CO~SCoA +3.5 CO +H2O –16.4 Ameisensäure +H2 –H2O +32.5 –4.2 Formaldehyd Methenyl-H4MPT +H2 –10 +H2 –53.8 Methylene-H4MPT +H2 CH3CO–Ni +20 –3.5 –H2O +Ni[E] –41 Methyl-H4F +H2 Formyl-H4MPT +20 CO Methylene-H4F +H2 +16 Formyl-MF +22.5 –H2O CO2 CO2 Ameisensäure +H4F Ohne Kofaktoren +H2, –H2O CO2 +H2 c) Methanol –14 Methyl-H4MPT –H4MPT +H2 * +H2 –H2O –112.6 –115 Methan Methan Biosynthesen Bei Acetogenen und Methanogenen ist der Vorgang der CO2-Fixierung über den Acetyl-CoA-Weg direkt mit dem Energiestoffwechsel (Netto-ATP-Synthese) verbunden. Bei Acetogenen wird im Zuge der Zuführung von Elektronen aus H2 zur Methylgruppe ein chemiosmotisches Potential (Ionengradienten) aufgebaut. Bei Methanogenen wird das chemiosmotische Potenzial dadurch aufgebaut, dass ein Teil der gebildeten Methylgruppen zur Methanbildung herangezogen wird. Bei beiden Gruppen wird das chemiosmotische Potenzial durch eine ATP-Synthase genutzt. Bei beiden Gruppen werden Acetyl-CoA und ATP aus der CO2-Fixierung gewonnen. Des geht deshalb, weil die bei der CO2-Reduktion freiwerdende Energie an die Erzeugung von chemiosmotischem Potenzial gekoppelt wird. Aus Acetyl-CoA können Acetogene, aber auch Vertreter der Methanogene etwas ATP bilden (aber keine Netto ATP-Synthese für das Wachstum betreiben), und zwar über ein energiereiches Intermediat, das gemischte Anhydrid Acetylphosphat, das aus der einfachen Phosphorolyse der Thioesters hervorgeht. Wenn mittels einer ähnlichen Chemie Acetylthioester an einer Hydrothermalquelle längst vergangener Zeiten hätten gebildet werden können, so könnte folglich Acetylphosphat als der chemische Vorläufer von ATP als universelle Energie-Währung gedient haben [13]. Acetylphosphat ist chemisch sehr viel einfacher als ATP und kann noch besser als ATP andere Moleküle phosphorylieren. a) CO2-Fixierung bei Acetogene (für Details siehe [12, 20]). b) CO2-Fixierung bei Methanogene (für Details siehe [12, 21]). c) Die chemischen Zwischenstufen auf dem Wege der nichtenzymatischen CO2-Reduktion zu Methan, das bei Lost City gebildet wird [17]. Die Zahlen neben den Reaktionspfeilen geben die Änderungen (ca.) der freien Enthalpie, ΔG 0’, unter physiologischen Bedingungen (25° C und pH 7) in kJ · mol–1an. Das gestrichelte Oval weist auf das Enzym bifunktionelle Kohlenmonoxid Dehydrogenase/ Acetyl-CoA-Synthase (CODH/ACS) hin, das bei Methanogenen und Acetogenen konserviert ist. H4F, Tetrahydrofolsäure; MF, Methanofuran; H4MPT, Tetrahydromethanopterin; Ni[E], ein Fe-Ni-S Cluster im CODH/ACS; HSCoA, Coenzym-A. Ein Asterisk (*) neben der Konversion von Methyl-H4MPT zu Methan zeigt an, dass einige der an diesem Geschehen beteiligten enzymatischen Schritte hier nicht gezeigt sind (siehe [12, 21]), das Gleiche gilt für die Konversion von Acetyl-CoA zu Acetat bei Acetogenen (siehe [12, 20]). Für Acetogene und Methanogene ist das Reaktionsgeschehen, das zur Netto-ATP-Gewinnung beiträgt, zusammengefasst; graue Pfeile bedeuten, dass ein Teil des C-Fluxes zur Deckung des zellulären Bedarfs an reduziertem Kohlenstoff „abgezweigt“ wird. 172 | Biol. Unserer Zeit | 3/2009 (39) www.biuz.de © 2009 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim HYDROTHERMALQUELLEN milde, alkalische Hydrothermalquellen vom Lost CityTyp besonders reich an FeS und NiS als anorganische Katalysatoren gewesen sein mussten. Das ist deshalb der Fall, weil theoretisch das austretende Wasser an solchen Quellen sehr viel H2S aus der Kruste hätte führen müssen, während das Meer sehr große Mengen an gelöstem Fe2+ (neben etwas Ni2+) enthielt. Am Schlot bilden sich daher FeS und NiS-Niederschläge – genau dort,wo H2 aus der Serpentinisierung und CO2 im Meer aufeinandertreffen, um ihrer natürlichen Neigung zur chemischen Reaktion nachzugehen. Das sind sehr gute Voraussetzungen für Reaktionen. Aber was nutzen Reaktionen, wenn alle Reaktionsprodukte ins Meer diffundieren? Fossile Hydrothermalquellen, aus FeS bestehend, zeigen beeindruckende Labyrinthe von Mikrokompartimenten in der Größenordnung von 1–100 µm [13]; solche natürlich entstehenden Mikrokompartimente könnten durchaus als Konzentrierungsmechanismus gedient haben.Lost City bildet auch solche Mikrokompartimente, wenngleich der Schlot aus Karbonatgestein statt FeS besteht [11]. Durch geochemisch entstehende Mikrokompartimente könnten Reaktionsprodukte an einer urzeitlichen Hydrothermalquelle am Ort ihrer Synthese von der Diffusion zurückgehalten werden und Konzentrationen erreichen, die für weitere Reaktionen und letztendlich für Polymerisierung erforderlich wären. Bis auf die reinen Fermentierer nutzen alle Zellen die Chemiosmose zur Netto-ATP-Synthese. Die reinen Fermentierer sind aber aus chemiosmotisch lebenden Zellen hervorgegangen, weil erstere ohne letztere, welche die fermentierbaren Substrate bereitstellen, nicht hätten entstehen können. Demnach waren auch die ersten freilebenden Zellen zur Chemiosmose fähig. Aber die chemiosmotische ATP-Bildung besteht stets aus zwei Komponenten: einem Mechanismus zum Pumpen von Protonen,und einer ATP-Synthase,die den Gradienten in chemische Energie umwandelt. Aus evolutionärer Sicht ist dies ein gewaltiges Hühnchen-oderEi Problem, da die eine Komponente ohne die andere nichts nützt – es sei denn, das Leben ist an einem alkalischen Hydrothermalsystem entstanden, weil dort der Protonengradient geochemisch umsonst geliefert wird [13, 18]. Wie das? Bei einer alkalischen Hydrothermalquelle (pH 9–10, wie Lost City) ist ein natürlicher Protonengradient vorhanden. Heute hat Meerwasser ein pH von circa 8.1, der pH im Urmeer lag aufgrund der damals höheren CO2 Konzentrationen eher bei circa pH 6–7, [18]. Zwischen dem austretenden Hydrothermalwasser und dem Meereswasser lag also ein auf geochemischen Wege entstandenes chemiosmotisches Potenzial vor, das in der gleichen Größenordnung liegt, wie das der heutigen Zellen (etwa 3 pH Einheiten). Es kann daher sein, dass die Vorstufen der freilebenden Zellen dieses geochemisch gebildete chemiosmotische Potential zuerst mit Hilfe einer primitiven ATP© 2009 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim | S C H W E R P U N K T E VO LU T I O N S FO R SC H U N G Synthase lange Zeit genutzt haben, bevor sie die sehr viel komplizierteren Mechanismen erfunden haben,die zur Erzeugung eines eigenen Protonengradienten erforderlich sind. Die ATP-Synthasen bei allen Organismen,auch Acetogenen und Methanogenen,ähneln sich im Aufbau und Struktur stark, d.h. sie haben einen gemeinsamen Vorfahren.Dagegen haben die Proteine und Kofaktoren, mit denen moderne Zellen ihre Protonengradienten aufbauen, eine schwindelerregende Vielfalt. Die Universalität der Nutzung von Protonengradienten mittels ATP-Synthasen stimmt mit der Vorstellung gut überein, dass zuerst ein geochemischer Protonengradient am Anfang vorhanden war und genutzt wurde. Aber der letzte und entscheidende Schritt – der Übergang zur freilebenden Zelle – setzte voraus, dass die Reduktion von CO2 mit Elektronen aus H2 mit dem Aufbau eines eigenständigen, selbst erzeugten Protonengradienten gekoppelt wurde. Diejenigen Urzellen, die das geschafft haben,konnten ihrem geochemischen Brutkasten entkommen, und der Zugang zur Evolution als freilebende Zellen wäre geschafft gewesen. Demnach wären die ersten Lebewesen hinsichtlich Größe und Form von modernen Prokaryoten kaum verschieden und ihrem biochemischen Wesen nach den acetogenen Eubakterien und den methanogenen Archaebakterien ähnlich gewesen [13]. Zusammenfassung An Tiefsee-Hydrothermalquellen gedeihen ganze Ökosysteme in völliger Finsternis. Am Anfang ihrer Nahrungskette steht nicht die Sonne, sondern chemische Energieträger wie molekularer Wasserstoff, die aus dem Erdinneren strömen. Seit ihrer Entdeckung vor 30 Jahren werden Hydrothermalquellen im Zusammenhang mit dem Ursprung des Lebens diskutiert, weil die Erde dort chemische Energie spendet. Manche Hydrothermalquellen bilden sogar heute noch Methan aus CO2 auf rein geochemischem Wege. Die chemischen Bedingungen dort sind also für die Umwandlung von CO2 in organische Substanzen – die Bausteine des Lebens – förderlich. Summary Deep sea hydrothermal vents harbour rich ecological communities that thrive independent of sunlight. The energy underpinning such communities is of chemical nature, of particular importance is molecular hydrogen in the hydrothermal vent effluent. Since their discovery 30 years ago, hydrothermal vents have come to bear heavily on thinking about the origin of life. Some contemporary hydrothermal vents even generate methane through geochemical means. The chemical environments at hydrothermal vents are conducive to the conversion of CO2 into reduced organic compounds and the building blocks of life. Schlagworte Tiefsee-Hydrothermalquellen, Ursprung des Lebens www.biuz.de 3/2009 (39) | Biol. Unserer Zeit | 173 Literatur [1] W. Bach, Submarine Hydrothermalquellen – Treffpunkte von Geologie, Geochemie und Biologie http://www.g-v.de/content/view/ 260/59/ [2] W. Bach, H. Paulick, C.J. Garrido, B. Ildefonse, W.P. Meurer, S. E. Humphris, Unraveling the sequence of serpentinization reactions: petrography, mineral chemistry, and petrophysics of serpentinites from MAR 15ºN (ODP Leg 209, Site 1274), Geophysical Research Letters 2006, 33, L13306. [3] J.L. Bada, A. Lazcano, Some like it hot, but not the first biomolecules, Science 2002, 296, 1982–1983. [4] J.A. Baross, S.E. Hoffman, Submarine hydrothermal vents and associated gradient environments as sites for the origin and evolution of life, Origins Life Evol. Biosphere 1985, 15, 327–345. [5] J.B.S. Haldane, The origin of life, Rationalist Annual 1929, 148, 3–10. [6] W. Heinen, A.M. Lauwers, Organic sulfur compounds resulting from the interaction of iron sulfide, hydrogen sulfide and carbon dioxide in an anaerobic aqueous environment. Origins Life Evol. Biosphere 1996, 26, 131–150. [7] C. Huber, G. Wächtershäuser, Activated acetic acid by carbon fixation on (Fe,Ni)S under primordial conditions, Science 1997, 276, 245–247. [8] G. F. Joyce, The antiquity of RNA-based evolution, Nature 2002, 418, 214–221. [9] D. S. Kelley et al. An off-axis hydrothermal vent field near the MidAtlantic Ridge at 30° N. Nature 2001 412, 145–149. [10] D.S. Kelley, J.A. Baross, J. R. Delaney, Volcanoes, fluids, and life at mid-ocean ridge spreading centers. Annu. Rev. Earth Planet. Sci. 2002, 30, 385–491. [11] D.S. Kelley, et al. A serpentinite-hosted ecosystem: The Lost City hydrothermal field, Science, 2005 307, 1428–1434. [12] B. E. H. Maden, Tetrahydrofolate and tetrahydromethanopterin compared: functionally distinct carriers in C1 metabolism. Biochem. J. 2000, 350, 609–629. [13] W. Martin, M.J. Russell, On the origin of biochemistry at an alkaline hydrothermal vent. Phil. Trans Roy. Soc. Lond. B 2007, 367, 1187– 1925. [14] W. Martin, J. Baross, D. Kelley, M.J. Russell: Hydrothermal vents and the origin of life, Nature Rev. Microbiol. 2008, 6, 805–814. 174 | Biol. Unserer Zeit | 3/2009 (39) www.biuz.de [15] S.L. Miller, A production of amino acids under possible primitive Earth conditions. Science 1953, 117, 528–529. [16] P. Mitchell, Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature 1961, 191, 144–148. [17] G. Proskurowski et al. Abiogenic hydrocarbon production at Lost City Hydrothermal Field, Science 2008, 319, 604–607. [18] M.J. Russell, First life, American Scientist 2006, 94, 32–39. [19] N.H. Sleep, A. Meibom, T. Fridriksson, R.G. Coleman, D.K. Bird, H2-rich fluids from serpentinization: Geochemical and biotic implications, Proc. Natl. Acad. Sci. USA 2004, 101, 12818–12823. [20] S. W. Ragsdale, E. Pierce, Acetogenesis and the Wood-Ljungdahl pathway of CO2 fixation, Biochim. Biophys. Acta 2008, 1784, 1873–1898. [21] R.K. Thauer, A.K. Kaster, H. Seedorf, W. Buckel, R. Hedderich, Methanogenic archaea: ecologically relevant differences in energy conservation, Nature Rev. Microbiol. 2008, 6, 579–591. Der Autor William Martin, Jahrgang 1957, studierte Biologie an der Texas A&M Universität und an der Technischen Universität Hannover. Er promovierte 1989 am Max-Planck-Institut für Züchtungsforschung in Köln, forschte anschließend als Postdoktorand an der Technischen Universität Braunschweig auf dem Gebiet der molekularen Evolution und ist seit 1999 Professor der Botanik an der Universität Düsseldorf. Seine Forschungsinteressen sind Endosymbiose, mikrobielle Evolution und frühe Evolution. Korrespondenz: Prof. Dr. William Martin Institut für Botanik III Heinrich-Heine-Universität Düsseldorf Universitätsstraße 1 40225 Düsseldorf Email: [email protected] © 2009 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim