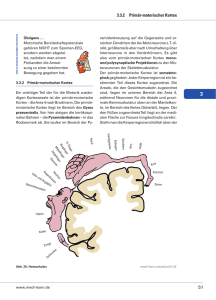
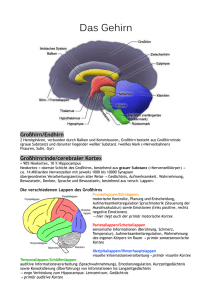
Dokumentvorlage für Studienbriefe
Werbung

Gliederung 1 Gliederung des Kurses Biologische Grundlagen der Psychologie 0 Einführung 3 0.1 Inhaltliche Einführung 3 0.2 Studientechnische Einführung 4 I Biologische Grundlagen 7 I.1 Die Zelle 7 I.2 Grundbegriffe der Genetik 10 I.3 Genetik und Evolution 13 I.4 Biologie, Genetik und Verhalten 14 II Informationsübermittlung im Körper 18 II.1 Nervenzelle und Übertragungsmechanismen 19 II.1.1 Die Nervenzelle 19 II.1.2 Reizleitung 20 II.1.3 Synapsen und synaptische Übertragung 22 II.1.4 Wichtige Transmitter-Rezeptoren-Systeme 26 II.2 Zentrales Nervensystem (ZNS) 29 II.2.1 Das Rückenmark 29 II.2.2 Das Gehirn 32 II.3 Das Vegetative Nervensystem (VNS) 41 II.4 Das Hormonsystem (Endokrines System) 42 III Motorik 51 III.1 Muskelzellen und Muskulatur 52 III.2 Steuerung und Steuerungsmechanismen 55 III.3 Koordination durch das Zentrale Nervensystem 56 IV Sinnessysteme 66 IV.1 Sehen 68 IV.1.1 Auge, Stäbchen und Zapfen 69 IV.1.2 Reizweiterleitung und Reizverarbeitung 72 IV.2 Hören 80 IV.2.1 Ohr, Corti-Organ und Haarzellen 80 IV.2.2 Reizweiterleitung und Reizverarbeitung 84 IV.3 Riechen 86 IV.3.1 Nase, Riechepithel und Riechzellen 86 IV.3.2 Reizweiterleitung und Reizverarbeitung 89 2 Biologische Grundlagen IV.4 Schmecken 91 IV.4.1 Geschmacksknospen und gustatorische Sensoren 91 IV.4.2 Reizweiterleitung und Reizverarbeitung 92 IV.5 Gleichgewicht und Bewegung 94 IV.5.1 Vestibularorgan und Haarzellen 94 IV.5.2 Reizweiterleitung und Reizverarbeitung 97 IV.6 Hautsinne 98 IV.6.1 Haut, Mechanosensoren, Thermosensoren und Schmerz- 99 rezeptoren IV.6.2 Reizweiterleitung und Reizverarbeitung 102 V Erfassungsmethoden der Biologischen Psychologie 109 V.1 Verfahren zur Erfassung zentraler Aktivität 111 V.1.1 Strukturabbildende Verfahren 111 V.1.2 Aktivitäts- und funktionsabbildende Verfahren 112 V.1.2.1 Elektroenzephalogramm (EEG) 112 V.1.2.2 Magnetenzephalographie (MEG) 116 V.1.2.3 Positronenemissionstomographie (PET) 116 V.1.2.4 Funktionelle Magnetresonanztomogrphie (fMRT) 118 V.1.2.5 Transkranielle Magnetstimulation (TMS) 123 V.2 Verfahrung zur Erfassung peripherer Aktivität 124 V.2.1 Elektrokardiogramm (EKG) 124 V.2.2 Blutdruckmessung 125 V.2.3 Photopletysmographie 126 V.2.4 Erfassung elektrodermaler Aktivität 127 V.2.5 Elektromyogramm (EMG) 128 V.2.6 Elektrookulogramm (EOG) 128 V.3 Erfassung humoraler Aktivität 129 VI Ausgewählte Funktionen 133 VI.1 Circadiane Rhythmik, Wachen und Schlafen 134 VI.2 Hunger, Durst, Sättigung 140 VI.3 Sexualität und Fortpflanzung 144 VI.4 Reifung und Altern 148 VI.5 Stress und Immunabwehr 152 VI.6 Emotionen 157 VI.7 Lernen und Gedächtnis 161 Einführung 0 Einführung zum Kurs Biologische Grundlagen der Psychologie Bitte lesen Sie diese Einführung sorgfältig durch! 0.1 Inhaltliche Einführung Körper und Seele des Menschen bilden eine psychophysische Einheit. Im Kurs „Einführung in die Psychologie und ihre Geschichte“ wurde im Abschnitt „Allgemeine Psychologie und ihre biologischen Grundlagen“ auf den biologischen Seelenbegriff verwiesen, wie er von Aristoteles in den Grundzügen geprägt wurde. Es wurde vorgeschlagen, die Seele als Organisation des Körpers zu verstehen. Es sind die seelischen Funktionen wie Wahrnehmen, Gedächtnis, Handeln, körperliche Regungen, die in Funktionskreisen organisiert sind und jeden lebendigen Organismus kennzeichnen. Wenn man den Menschen verstehen will, muss man daher auch den Aufbau seines Körpers (Anatomie) und dessen Funktionsweise kennen lernen. Dazu wird der Bogen von der Psychologie zur Biologie geschlagen. Ausführungen zur Fragestellung und allgemeine Hinweise zur Methodologie der Biologischen Psychologie finden sich im Kurs 03400 Einführung in die Psychologie und ihre Geschichte im Abschnitt 5.1.5. Im Kapitel I werden allgemeine biologische Ansätze und Kenntnisse vermittelt, zunächst die Zelle als Grundbaustein des Lebens. Struktur und Funktion von Zellen werden durch Gene gesteuert. Diese sind der Bereich, in welchem die Mechanismen der (molekularen) Evolution einwirken und sich so indirekt auf Zellaufbau und –funktion auswirken. Über diesen indirekten Weg des Zellaufbaues steuern die Gene auch komplexere Zellorganisationen, die wiederum den Verhalten ermöglichenden Rahmen bilden. Je nach Organismus erlaubt dieser Rahmen bestimmte Freiheitsgrade der Verhaltensplastizität und legt damit Unter- und Obergrenzen der Verhaltensadaptivität fest. Insbesondere für komplexere selbstbewegliche Organismen ist die Disposition zu Lernen genetisch determiniert. Das Kapitel II befasst sich mit Informationssystemen des menschlichen Körpers, die die Aufnahme von Informationen aus der Umgebung und aus dem eigenen Körper sowie deren Verarbeitung ermöglichen. Information ist, vereinfacht gesagt, ein Maß für die Strukturiertheit, für die Ordnung der Umgebung. Die auf die Aufgabe der Informationsverarbeitung spezialisierten Zellen, die Neuronen, werden in ihrem Aufbau und ihrer Funktionsweise ebenso wie die von ihnen gebildeten Verbünde und Systeme, das zentrale und das vegetative Nervensystem, beschrieben. Mit den Hormonen steht ein weiteres Medium zur Verfügung, mit dessen Hilfe Informationen von einem Ort an einen anderen gebracht werden können. Informationsübermittlung ist natürlich nicht Selbstzweck, sondern die Basis für eine wirkungsvolle Auseinandersetzung des Individuums mit seiner Umwelt. Eine Einwirkung des Menschen auf seine Umwelt, sei es eine sprachliche Äußerung 3 4 Biologische Grundlagen oder das Gehen an einen schöneren Ort, ist möglich, weil sich Menschen bewegen und ihre Bewegungen steuern können. Auch die psychologische Erkenntnisbildung bezieht sich in wesentlichen Teilen auf dieses offene, beobachtbare Verhalten. Im Kapitel III wird daher die Motorik behandelt, wobei auch kurz auf die Bewegungen eingegangen wird, die unwillkürlich (nicht willentlich gesteuert) erfolgen. Auch in diesem Abschnitt steht die Informationsübermittlung von den zentralen Steuerungseinheiten hin zu den Effektorzellen im Mittelpunkt. Ein großes und für die Psychologie wichtiges Kapitel sind die Sinnessysteme des Menschen. Mit ihrer Hilfe nehmen Menschen ihre Umwelt wahr, wobei die verschiedenen Sinnessysteme immer nur Ausschnitte aus der Umwelt wahrnehmen können. Im Kapitel IV wird erläutert, durch welche Reize die Sinnesorgane Auge, Ohr, Nase, Zunge und Haut aktiviert werden können und wie aus diesen Aktivierungen Wahrnehmungen werden, die wiederum Verhalten veranlassen und steuern können. Nachdem die Funktionsweise der Sinnes- und der Effektorsysteme beschrieben wurde, stehen im Kapitel V die Methoden im Vordergrund, mit denen die biologisch-physiologischen Korrelate oder Indikatoren innerer Vorgänge erfasst werden. Solche Korrelate sind die physiologischen Größen, die durch einen externen Reiz verändert werden, oder deren Änderung mit bestimmten Bewusstseinzuständen (Erleben) oder Verhalten einhergeht. Im abschließenden Kapitel VI werden – auch im Vorgriff auch die vertiefende Betrachtung in anderen Kursen und Modulen – einige Funktionen oder Prozesse des menschlichen Lebens aus biologisch-physiologischer Sicht beschrieben. Es handelt sich dabei beispielsweise um einfache homöostatische Regelkreise wie den Schlaf-Wach-Rhythmus oder die Regulierung von Hunger und Durst, aber auch um Phänomene wie Emotionen oder Lernprozesse. 0.2 Studientechnische Einführung Jedem Abschnitt des Kurses wurden Lernziele vorangestellt, die erläutern, in welchen Zusammenhängen die im Text beschriebenen Wissensinhalte vermittelt werden sollen. Sie können – am Schluss des Studiums des jeweiligen Abschnitts – als Kontrollfragen verwendet werden. Sie können sie nutzen, um das im jeweiligen Abschnitt dargestellte Wissen mit den anderen, im Verlauf Ihres Studiums erhaltenen Kenntnissen zu verknüpfen beziehungsweise konstruktiv anzuwenden. Jedem Abschnitt folgt unter der Überschrift „Kenntnisse“ ein Stichwortverzeichnis. Sie sollten zu genannten Stichworten Definitionen oder Erläuterungen kennen und sie gegenseitig in Beziehung setzen können. Damit können Sie selbst (aber natürlich auch Ihre Professorinnen und Professoren) überprüfen, ob Sie den Stoff aufgenommen haben. Einführung Und nicht zuletzt wird jedem Abschnitt folgend die Literatur genannt, die gelesen werden sollte (Grundlage), da sie prüfungsrelevant ist. Zur Vertiefung (vertiefend) dient Literatur, die mehr Zusammenhänge herzustellen erlaubt, helfen kann, Stoff zu verfestigen und bei weiterreichendem Interesse nützlich ist. Sonstige zitierte Literatur wird wegen der Zitierpflicht aufgeführt, ist aber auch für Interessierte empfehlenswert. Die in den biologischen Grundlagen dargestellten Zusammenhänge sind meist komplex und verlangen Vorwissen über chemische, biologische und physikalische Sachverhalte. Für den Fall, dass dies seit der Hochschulreife desaktiviert wurde, wird die Nutzung von Nachschlagewerken, aber auch die Erstellung von Übersichten oder Karteikarten angeregt. Auch die eigenhändige Aufzeichnung manches Zusammenhangs mag helfen. Bitte bedenken Sie, dass Wikipedia keine wissenschaftliche Quelle ist, auch wenn viele Einträge durchaus von großem Wert sind. Nutzen Sie bitte Fachlexika. Die Universitätsbibliothek der FernUniversität in Hagen bietet entsprechende Fachlexika an, die elektronisch zugänglich sind. Bitte machen Sie sich mit der Nutzung der Ressourcen der UB vertraut, falls Sie das noch nicht gemacht haben. Die Terminologie der biologischen Grundlagen der Psychologie ist die Terminologie vor allem der Medizin und durch lateinische und altgriechische Bezeichnungen geprägt. Daher wurde der „gebräuchlichste Begriff“ gegenüber einer deutschen Übersetzung (die sich im Zweifelsfall kaum noch an anderer Stelle findet) bevorzugt. Am Rande des Textes finden Sie gelegentlich Marginalien und im Text sind Begriffe unterschiedlich hervorgehoben. Dies soll Ihnen bei der Textstrukturierung helfen. Daraus folgt nicht, dass weniger hervorgehobene Textteile weniger wichtig sind. Wir empfehlen Ihnen sehr, die Inhalte dieses Kurses mit anderen Studierenden zu erarbeiten. Eine Möglichkeit dafür ist die virtuelle Lehr-Lern-Umgebung Moodle, die am Institut für Psychologie der FernUniversität in Hagen standardmäßig eingesetzt wird. Den Zugang finden Sie über den entsprechenden Link auf der Startseite des Instituts. Es ist geplant, die Lernumgebung zu diesem und anderen Kursen des Moduls 3 wird am 03.05.2009 freizuschalten. Bitte informieren Sie sich auf den entsprechenden Webseiten des Instituts für Psychologie. In Moodle werden Sie zu inhaltlichen Fragen des Kurses von MentorInnen und uns unterstützt. Da man aber gut lernt, wenn man anderen selber Fragen beantwortet und gemeinsam versucht, sie zu beantworten, sollten Sie zunächst Fragen im Kreise Ihrer KommilitonInnen beantworten. Wir werden auch kleinere Aufgaben und Kontrollfragen in Moodle einstellen, die Sie bearbeiten können. Studieren ist selbstreguliertes Lernen und Lernen ist Arbeit an einem persönlichen Wissenssystem. Wissen kann nicht nach Art des Nürnberger Trichtes in den Kopf gefüllt werden. Selten versteht man nach einmaligem Durchlesen alles und nicht jede vorgegebene Struktur ist für jeden hilfreich. Deswegen sollte man Lernstoffe 5 6 Biologische Grundlagen eventuell in ein eigenes Skript übertragen. Planen Sie also sorgfältig Ihr lernen. Das heisst vor allem, planen Sie Ihre Zeit so, dass sie regelmäßig lernen. Denken Sie immer wieder darüber nach, wie Sie arbeiten, analysieren Sie realistisch Ihre Stärken und Schwächen und finden Sie Ihre für Sie passenden Lern- und Arbeitstechniken heraus. Und nun viel Freude und viele bildende Erkenntnisse beim Studium des Kurses! Lexika, Nachschlagewerke, die von Studierenden regelmäßig genutzt werden sollten: Dorsch, F. (2004) (hrsg. v. H. O. Häcker & K.-H. Stapf). Psychologisches Wörterbuch (14., vollst. überarb. u. erweiterte Aufl.). Bern etc.: Huber. (Anm. WM: „Klassisches“ Wörterbuch für Psychologen, das eine schnelle Orientierung erlaubt und zahlreiche Tests nachweist). Städtler, T. (1998). Lexikon der Psychologie. Wörterbuch, Handbuch, Studienbuch. Stuttgart. Kröner. (Anm. WM: Auch neuere Aufl. nutzbar). Elektronisch zugänglich für autorisierte Benutzer im Hochschulnetz der FernUniversität in Hagen: Hanser, H. (Red.). (2001) (Elektronische Ressource). Lexikon der Neurowissenschaft. Mannheim: Brockhaus, Duden, Neue Medien. „Elektronisch zugänglich für autorisierte Benutzer im Hochschulnetz der FernUniversität in Hagen“ heißt: Sich in die UB FeU einloggen, Eintrag „Lexikon der Neurowissenschaft“ unter „Suchen“ eingeben und http://lexika.tanto.de/?TANTO_KID=digibib12&TANTO_AGR=133581 klicken. Dort finden sich weitere nützliche Lexika (der Psychologie (s.u.), der Medizin usw.). Wenninger, G. (Hrsg.) (2002) (Elektronische Ressource). Lexikon der Psychologie. Mannheim: Brockhaus, Duden, Neue Medien. Bemerkung zum Copyright der Abbildungen: Die FernUniversität in Hagen dankt allen Rechtsinhabern für die erteilten Abdruckgenehmigungen. Nicht in allen Fällen ist es trotz intensiver Bemühungen gelungen, die Rechtsinhaber bzw. deren Nachfolger zu ermitteln oder zu diesem Kontakt aufzunehmen. Diese werden deshalb gebeten, sich mit der FernUniversität in Hagen, in Verbindung zu setzen. I Biologische Grundlagen I. 7 Biologische Grundlagen Lernziele Die Studierenden sollen den Menschen und damit den „Gegenstand der Psychologie“ als auch biologisches Wesen beziehungsweise biologisches System begreifen lernen, die Basis biologischer Systeme kennenlernen, deren materielle Komponenten, Bau, Funktion, Mechanismen und Grenzen wie o den Aufbau von Zellen und Zellsystemen o Vererbung (Genetik, Evolution) o mögliche Zusammenhänge zwischen Anlage und Umwelt bei der Entstehung und Ausgestaltung des Verhaltens. Ansätze der Biologie kennenlernen, die auf menschliches Erleben und Verhalten mehr oder weniger gut übertragbar sind. I.1 Die Zelle Alle Lebewesen, Pflanzen, Pilze, Tiere und der Mensch, sind aus Zellen zusammengesetzt. Ihre Zellen werden als eukaryote Zellen bezeichnet; sie verfügen im Gegensatz zu den prokaryoten Zellen zum Beispiel der Bakterien über einen Zellkern. (Eukaryote) Zellen bestehen aus einem Zellkern, dem Cytoplasma und der beides umschließenden Plasmamembran. Der Zellkern enthält die genetischen Informationen und ist von einer inneren und äußeren Membran mit Poren umschlossen. Durch die Membran werden Stoffe beziehungsweise vom Zellkern ins Cytoplasma und zurück transportiert. Im Cytoplasma befinden sich eine Reihe von Organellen, darunter die Mitochondrien, die für den Energiehaushalt zuständig sind, das endoplasmatische Retikulum, in dem Proteine synthetisiert werden, und den Golgi-Apparat für die Modifizierung und Reifung der Proteine sowie deren Transport. Die gesamte Zelle ist von einer Zellplasma-Membran umschlossen, die zahlreiche, für die Funktion der Zelle wichtige Proteine enthält. Proteine sind aus Aminosäuren zusammengesetzte Moleküle. In der Zellplasma-Membran gibt es Proteine, die als Transportkanäle dienen, und Strukturproteine, die für Stabilität sorgen. Über Rezeptormoleküle werden Signale aufgenommen oder weitergegeben. Und schließlich sorgen Proteine mit enzymatischer Wirkung, angeregt durch äußere Reize, als Katalysator für chemische Prozesse im Inneren der Zelle. eukaryote Zellen Zelle, ~kern Cytoplasma Zellplasma-Membran Biologische Grundlagen 8 Abb I.1: Aufbau der Zelle und des Zellkerns (aus: Passarge, E. (2004): Taschenatlas der Genetik (2. Aufl.), S. 23, G. Thieme Verl.). Es gibt Lebewesen, die aus einer einzigen Zelle bestehen, hier muss eine Zelle alle Aufgaben für die Aufrechterhaltung ihres Lebens erfüllen. Bei Lebewesen, die wie der Mensch aus einer Vielzahl von Zellen bestehen, spezialisieren sich die Zellen und übernehmen jeweils bestimmte Funktionen. So finden wir beim Menschen mit ungefähr 6·1013 Zellen Muskelzellen, Nervenzellen, Sinneszellen, Knochenzellen, usw. Einige dieser Zelltypen werden im Lauf des Kurses noch besprochen. Organismus als offenes System Ein Organismus ist ein lebendiger Körper, der thermodynamisch gesehen, als ein offenes System zu verstehen ist. Dieses System muss aktiv für eine Ungleichverteilung von Masse und Energie sorgen, damit seine Komponenten sich nicht in kürzester Zeit auflösen und sich gleichmäßig im Raum verteilen (Dissipation). Dies entspricht der Tendenz zum thermodynamischen Gleichgewicht. Daher muss ein Organismus energiereiche Substrate aus der Umgebung aufnehmen (Fette, Kohlenhydrate, Proteine), um aktiv Energiegradienten gegen die Tendenz zur „Unordnung“ (Entropie) aufzubauen. Beispiele für solche Energiegradienten sind I Biologische Grundlagen der Blutdruck und die Kaliumanreicherung in der Nervenzelle. In beiden Fällen beruht dieser Gradient auf chemischer Energie, die mit Hilfe von ATP (Adenosintri-phosphat) gewonnen wird. Leben ist also, unter anderem, ein Prozess, der gegen die Gleichverteilung von Energie gerichtet ist und damit in aktiver Energiegewinnung zur Herstellung und Aufrechterhaltung von Energiegradienten besteht (Negentropie). Im Rahmen der Psychologie ist dabei vor allem interessant, wie diese vielen Zellen mit ihren verschiedenen Strukturen und Funktionen zusammenarbeiten, wie komplexes Verhalten, zum Beispiel das Absolvieren eines Studiums, organisiert werden kann. Diese Zusammenarbeit beruht auf Energiegewinnung und – verwertung sowie auf Informationsverarbeitung. Zu diesem Zweck müssen die Zellen eine Organisation bilden und sich die „Arbeit“ durch funktionelle Differenzierung teilen. Größere Zellgruppen wie Organe übernehmen dann spezialisierte Funktionen wie Verdauung oder Bewegung. So wird die Lokomotion jeder einzelnen Zelle eingeschränkt zugunsten der Bewegung ganzer Zellverbände bis hin zum Organismus als Ganzen. Die Genese der Organisation der Zellen im Sinne einer funktionellen Arbeitsteilung zu verstehen ist bis heute eine der größten Herausforderungen der Biologie (Problem der Selbstorganisation, der Morphogenese, der Emergenz). Bemerkenswert ist auch der Umstand, dass die typische Umgebung der meisten Zellen des menschlichen Organismus andere Zellen sind. Nur die Zellen der äußeren Grenzfläche sind direkt mit der extraorganismischen Außenwelt verbunden. Allerdings wirkt beispielsweise die Gravitationskraft gleichermaßen auf alle Zellen und das Funktionieren des Stoffwechsels ist auf bestimmte Temperaturbandbreiten angewiesen, aber auch auf komplexe energetische Verhältnisse, was An- und Abwesenheit bestimmter Raten und Konzentrationen von Chemikalien und Strahlungen angeht. Das Zusammenspiel von Mikro- und Makroebenen im Organismus lässt sich nur im Rahmen einer Systemtheorie und einer Theorie der variablen Regelung begrifflich verständlich machen, z. B. die Regulation des Blutzuckers, des Blutdrucks oder des Gleichgewichts und der Bewegungsregulation. Ein System ist „ein konkreter Ausschnitt aus der physischen Realität, in dem Interaktionen stattfinden, also Prozesse ablaufen“ (Bischof, 1995, S. 13). Organismen und damit auch Menschen sind lebendige kybernetische Systeme, die ihr inneres Gleichgewicht (Homöostase) gegenüber zahlreichen Einwirkungen aus der Systemumgebung („Außenwelt“) mit Hilfe von zahlreichen Regelungen („Regelkreise“ aus Soll- und Ist-Werten) aufrecht erhalten müssen. Das Ziel dieser Regelung ist die (Wieder-)Herstellung und Aufrechterhaltung eines stabilen Organismus-Umwelt-Verhältnisses, der Anpassung an die Umwelt (äußeres Gleichgewicht). Literatur zur Vertiefung, vor allem für Anspruchsvolle, beispielsweise zum Begriff des Organismus, der (biologischen) Funktion, zu den Begriffen Gen, Biologisches Design, Leben und Lebewesen in Beiträgen bei Krohs und Toepfer (2005). Zur Systemtheorie und Kybernetik vgl. Bischof (1995), Flechtner (1984). 9 10 Biologische Grundlagen I.2 Grundbegriffe der Genetik Gen Genotyp, Phänotyp, Genom DNA Wie oben ausgeführt, spezialisieren sich die Zellen beim Menschen wie bei den meisten anderen Lebewesen. Durch Gene wird festgelegt, welche Proteinarten eine Zelle herstellt und damit Art und Funktion einer Zelle. Prinzipiell kann eine Zelle alle durch den Organismus herstellbaren Proteine produzieren, meistens gibt es jedoch auch hier Spezialisierungen. Gene bestimmen (neben Umwelteinflüssen) die Ausbildung bestimmter Struktur- und Funktionseigenschaften. Mit Genotyp wird die Summe der in den Genen angelegten Erbanlagen bezeichnet, die den Rahmen bilden, in dem in der Auseinandersetzung mit den gegebenen Umweltbedingungen der Phänotyp entwickelt wird. Mit Genom wird die Gesamtheit aller menschlichen Erbinformationen benannt, die vermutlich 30.000 bis 40.000 Gene umfassen. Die Gene bestehen beziehungsweise bilden einen Abschnitt der Desoxyribonukleinsäure (DNS, meist DNA nach englisch Desoxyribonucleinacid). Damit ist die DNA der Träger genetischer Information. Der Aufbau der DNA ist eine „Doppelhelix“, in dem zwei aus Nukleotiden (bestimme Molekülstrukturen) aufgebaute schraubenförmig gedrehte Stränge sich jeweils gegenüberliegen. Es gibt vier verschiedene Nukleotide, die sich durch an ihnen beteiligten Basen unterscheiden. Durch die Reihenfolge der Nukleotide wird die genetische Information verschlüsselt. Dabei entspricht ein Gen einem solchen Abschnitt der DNA, die die Sequenz für die Herstellung eines bestimmten Proteins enthält. Abb. I.2: Aufbau der DNA-Doppelhelix. Die Doppelhelix der DNA setzt sich aus zwei umeinander gewobenen Zucker-Phosphat-Strängen zusammen, zwischen denen die Bindung durch Basenpaare hergestellt wird (aus: Schandry, 2006, S. 12). I Biologische Grundlagen Die Synthese von Proteinen auf der Basis der genetischen Informationen (Genexpression) läuft in verschiedenen Schritten ab. Zunächst wird eine Kopie des betreffenden DNA-Abschnitts auf einen Stoff, die Ribonukleinsäure (RNA) angefertigt, dieser Vorgang wird als Transkription bezeichnet. Die RNA fungiert als Botenstoff und überträgt die Information aus dem Zellkern ins Cytoplasma, wo sie als Vorlage für die Synthese des Proteins im Ribosom wirkt (Translation). 11 Genexpression Abb. I.3: Genexpression und Proteinsynthese. Zum Zweck der Genexpression trennen sich in einem ersten Schritt die beiden DNA-Stränge über eine bestimmte Strecke auf, die der Ausdehnung eines Gens entspricht. Ein Strang Messenger-RNA (mRNA) wird entlang des freien DNAStrangs synthetisiert und übernimmt dabei den genetischen Code. Die mRNA wandert aus dem Kern heraus in die Zellflüssigkeit. Hier lagert sich ein Ribosom an den mRNA-Strang an und bewegt sich entlang des Strangs. Jedes Kodon wird jetzt in eine Aminosäure übersetzt. Schließlich entsteht eine fertige Aminosäurekette, das Protein (aus: Schandry, 2006, S. 17). Die DNA erreicht eine erhebliche Länge (fast 1 Meter) und wird, damit sie in den Zellkern passt, mehrfach gewunden. Die so gebildete Struktur, das Chromosom ist während der Zellteilung im Lichtmikroskop zu erkennen. In (fast) jeder Körperzelle liegt ein Satz von 23 Chromosomen paarig vor (diploider Chromoso- Chromosom 12 Biologische Grundlagen mensatz). Teilt sich eine Zelle, wird zunächst der Chromosomensatz der Zelle verdoppelt (auf 46 Paare) und dann den entstehenden Tochterzellen je ein diploider Chromosomensatz mitgegeben. Bei der Entstehung von Keimzellen (also der Eizelle beziehungsweise der Samenzelle) wird der Chromosomensatz geteilt und es entstehen Keimzellen mit einem einfachem, haploiden Chromosomensatz. Erst bei der Verschmelzung einer Eimit einer Samenzelle entsteht wieder ein vollständiger, also diploider Chromosomensatz. Dabei können die beteiligten DNA-Stränge Gene austauschen, die die gleiche Stelle in einem Chromosom einnehmen (Allele). Durch dieses CrossingOver wird der DNA-Doppelstrang neu zusammengesetzt und es entsteht ein neuer Genotyp, ein neues Individuum. Abb. I.4: Crossing-over zwischen homologen Chromosomen. Beim Crossing-over kommt es zu einer Wechselwirkung zweier homologer Chromosomen, wobei Abschnitte der Chromosomen ausgetauscht werden. Dadurch wird der DNA-Doppelstrang neu zusammengesetzt (aus: Schandry, 2006, S. 21). I Biologische Grundlagen 13 I.3 Genetik und Evolution Sowohl bei der Zellteilung als auch bei der Entstehung von Keimzellen können „Kopierfehler“ (Mutationen) entstehen. Diese können positive oder negative Folgen haben und werden, soweit möglich, durch veschiedene Reparaturmechanismen verhindert. Bei Keimzellen ist es daher von Vorteil, dass diese Mutationen in der Regel rezessiv vererbt werden, d.h. beim Crossing-Over wird sich zunächst nicht das rezessive, sondern das dominante Allel durchsetzen und im Phänotyp sichtbar werden. Eine Mutation kann jedoch auch einen Vorteil hinsichtlich der Überlebens- und Reproduktionsfähigkeit bieten. Ist dies der Fall wird sich dieses Merkmal auf Dauer durchsetzen, die dadurch besser an ihre Umwelt angepassten Individuen werden diese Merkmal an ihre dann auch zahlreichere Nachkommenschaft weitergeben. Mit diesem Mechanismus wird die Evolution erklärt, er bildet aber auch die Grundlage für Tier- und Pflanzenzucht. Schon früh in ihrer Geschichte lernten Menschen, Pflanzen oder Tiere nach von ihnen gewünschten Eigenschaften auszuwählen und gezielt zu vermehren (Zuchtwahl). Dieses Vorgehen wurde im 19. Jahrhundert von Gregor Mendel systematisch beobachtet. Die von ihm formulierten Grundsätze zur Vererbung dominanter beziehungsweise nicht-dominanter Merkmale wurden zur Grundlage der Genetik, auch wenn die biochemischen Grundlagen noch unbekannt waren. (An die Stelle der menschlichen Zuchtwahl setzte Darwin einen impersonalen Selektionsmechanismus. Die Umgebung wählt nicht Organismen aus, sondern die Kopplung aus Organismusstrukturen und Umgebungsstrukturen erhöht entweder die Reproduktionswahrscheinlichkeit der Organismen oder sie erniedrigt diese.) Abb. I.5: Mendel-Erbgang. Im Beispiel ist die Vererbung im dominanten Erbgang eines Merkmals (Blütenfarbe beim Löwenmäulchen) wiedergegeben, das in der Parentalgeneration unterschiedlich ausgeprägt ist. Es sind die Merkmalsausprägungen in der ersten und zweiten Filialgeneration gezeigt. »R« steht für »Rot dominant«, »r« für »Rot rezessiv« (aus: Schandry, 2006, S. 22). Mutation Zuchtwahl 14 natürliche Auslese Evolution Biologische Grundlagen Im Gegensatz zur willentlichen Auslese, die bei der Zucht von Pflanzen oder Tieren getroffen wird, steht die in der Natur vorkommende natürliche Auslese (natürliche Selektion). Sie ist im Vergleich zur Zuchtwahl langsamer und nicht auf die Erzielung eines oder mehrerer gewünschter Merkmale gerichtet, sondern auf die optimale Anpassung an die Umwelt. „Optimal“ ist stets als relativ zu einem Raumzeitbereich zu sehen, in dem Organismen und deren Umgebung in bestimmter Weise wechselseitig aufeinander einwirken. Durch diesen langsamen Anpassungsprozess wird die Evolution erklärt, also die Entwicklung und Diversifizierung der verschiedenen Lebewesen auf der Erde. Evolution ist, so verstanden, keine auf ein endgültiges Ziel hin ausgerichtete Entwicklung. Eine sich verändernde Umwelt erfordert andere Merkmale, ein früherer Vorteil kann zum Nachteil werden und zum Aussterben von Arten führen. „Optimale Anpassung“ ist daher stets relativ zu einer bestimmten naturhistorischen Phase zu sehen. Man geht davon aus, dass vor ungefähr 600 Millionen Jahren die ersten Einzeller entstanden, die Vorfahren aller Lebewesen auf der Erde sind. „Erst“ vor sechs oder sieben Millionen Jahren lebte ein gemeinsamer Vorfahr von Mensch und Schimpanse. Alle heute auf der Erde lebenden Menschen gehören der Art „Homo Sapiens“ an. Der gegenwärtig im Aufbau befindliche Genatlas der Menschheit zeigt auf, dass die genetische Vielfalt der Menschen in Afrika am größten ist. Das bestätigt die These, dass die Menschheit aus Afrika stammt. I.4 Biologie, Genetik und Verhalten Wie hängen nun Gene und (menschliches) Verhalten zusammen? Aus dem bisher Beschriebenen wurde deutlich, dass dieser Zusammenhang ein mittelbarer sein muss. Gene wirken in oder auf Zellen, darunter auch die Nervenzellen. Erst durch das Zusammenspiel der Gene und die Interaktion des Individuums mit seiner Umwelt entsteht der Phänotyp und dessen erfassbare Merkmale beziehungsweise Eigenschaften. Ein Beispiel soll dies verdeutlichen. Geschlecht Das Geschlecht eines Menschen ist genetisch determiniert und bereits vor der Geburt bestimmbar. Das genetisch festgelegte Geschlecht (im Englischen: „sex“) führt zur Bildung der Geschlechtsorgane sowie in der Pubertät zur Ausbildung der sogenannten sekundären Geschlechtsmerkmale (z.B. die Brust bei der Frau, der Bartwuchs beim Mann). Die Pubertät wird hormonell und damit durch die Aktivität von Genen gesteuert. Allerdings gibt es ernährungsbedingte Unterschiede zum Beispiel hinsichtlich des Alters, in dem die Pubertät beginnt. Bereits hier beginnt der Einfluss der jeweiligen Umgebung. I Biologische Grundlagen 15 Über einen Zusammenhang des Geschlechts mit Intelligenz und vor allem deren Struktur, wenn zum Beispiel sprachliche Fähigkeiten eher bei Frauen, räumlichmathematische eher bei Männern gefunden werden, wird auch außerhalb psychologischer Forschung berichtet und diskutiert. Ein Mechanismus, der diesen Zusammenhang vermittelt, wird im Einfluss von Sexualhormonen auf die Nervenzellen des Gehirns gesehen. Und nicht zuletzt ist das Geschlecht auch eine soziale Größe (im Englischen „gender“): Rollenerwartungen und die Interpretation von gezeigtem Verhalten werden durch das Geschlecht bestimmt. Dabei kommt es zu Überlagerungen genetischer und sozialer Einflüsse und die Interpretation hängt auch von der Zielstellung des Forschenden ab. Menschen, Tiere und sogar Pflanzen weisen Ähnlichkeiten auf, hinsichtlich des Aufbaus aus (spezialisierten) Zellen oder bestimmter Prinzipien der Vererbung. Daher kann die Biologie als Nachbarwissenschaft der Psychologie Erkenntnisse liefern, die auch Grundlage der Erforschung menschlichen Lebens, Erlebens und Verhaltens ist. So können Aufbau und Struktur von Zellen oder die Mechanismen, mit denen Informationen innerhalb eines Organismus übertragen werden, an tierischen Zellen oder Tieren studiert werden. Aufschlüsse über menschliches Verhalten oder Erleben sind auch durch die Beobachtung von Tieren zu erhalten. In der Verhaltensforschung oder Ethologie werden durch systematische Beobachtung Prinzipien des Verhaltens von Tieren erforscht. Verhaltensweisen, die so erforscht werden, sind Fortpflanzungs- und Brutpflegeverhalten, aber auch aggressives Verhalten oder die Reaktion auf Belastungen. Eine der Fragen, die dabei beantwortet werden soll, ist die, inwieweit ein Verhalten angeboren, also genetisch determiniert, oder erlernt ist. Anmerkung: Sehr häufig findet sich in der Literatur auch die Gleichsetzung von „Vererbung“ oder „ererbt“ mit „genetisch determiniert“. Das ist aber falsch. Alles, was genetisch determiniert ist, ist auch vererbt, aber nicht alles, was vererbt ist, ist auch genetisch determiniert. Beispiel: Die Anzahl der Finger einer Hand sind beim Menschen genetisch determiniert. In vielen Ethnien tragen Verheiratete einen Ring am vierten Finger der rechten Hand. Da dieser Brauch von Generation zu Generation weitergegeben wird, ist das Merkmal „Verheiratetsein anzeigen durch Tragen eines Ringes am vierten Finger der rechten Hand“ vererbt. Da sich dafür keine Gene finden lassen, ist dieser Brauch nicht genetisch determiniert. Sicher angeboren sind die Reflexe, also schnell, meist unbewusst ablaufende Verhaltensweisen oder Reaktionen. Dazu gehört das Atemholen nach dem Ausatmen, das schnelle Schließen des Lides, wenn sich dem Auge ein Gegenstand nähert oder auch der Rückzug der Extremität, die einen sehr heißen Gegenstand berührt. Aber auch komplexere Verhaltensprogramme sind genetisch determiniert, wie das Brutverhalten oder das eines jungen Kuckucks gegenüber seinen „Pflegegeschwistern“. Und schließlich bestehen viele Verhaltensweisen aus einem definierten Wechselspiel zwischen Angeborenem und Erlerntem, so zum Beispiel das Ethologie Reflexe 16 Sensible Phase Biologische Grundlagen Jagdverhalten von Katzen. Dabei spielt häufig das Alter eine Rolle; in sensiblen Phasen werden Dinge relativ einfach erlernt, deren Erlernen zu einem späteren Zeitpunkt im Lebenslauf nicht mehr oder nur noch schwer möglich ist. Einige der in der Ethologie gewonnenen Erkenntnisse lassen sich sehr gut auf den Menschen übertragen, vor allem wenn es um die Erfüllung primärer Bedürfnisse geht, wie Atmen, Nahrungsaufnahme, aber auch um bestimmte Aspekte des Bindungsverhaltens zwischen Eltern und Kind. Im soziobiologischen Ansatz wird versucht, evolutionstheoretische Prinzipien auf soziales Verhalten zu übertragen. Dieser Ansatz geht davon aus, dass das Ziel eines jeden Individuums die Replikation seiner Gene ist. Soziales Verhalten wird dann dadurch erklärt, dass innerhalb einer verwandten (d.h. genetisch ähnlichen) Gruppe höhere Replikationschancen entstehen, auch wenn sich das einzelne Individuum nicht fortpflanzt. Es wird bisweilen versucht, diesen Ansatz auch auf menschliches Verhalten mit hohen ethischen Ansprüchen (altruistisches Verhalten) zu übertragen. Gegenwärtig wird kaum mehr von Soziobiologie gesprochen, vermutlich wegen des implizierten Reduktionismus, menschliches Sozialverhalten rein biologisch erklären zu können. Die Fragestellungen, Methoden und Ansätze der Soziobiologie finden sich in mehr oder weniger modifizierter Form in der Evolutionären Psychologie wieder. Die Evolutionstheorie ist die zentrale Theorie, um Stabilität und Wandel von Organismen zu erklären, wozu auch das Verhalten derselben gehört. Dazu gehört logischerweise auch das Sozialverhalten, das Verhalten vor allem gegenüber Mitgliedern der eigenen Art. Überlegungen und Beispiele zu Fragestellungen der evolutionären Psychologie im Zusammenhang mit komplexem menschlichen Verhalten sowie weiterführende Literatur finden sich in Hoffrage und Vitouch (2008). I Biologische Grundlagen Studientechnischer Anhang 1. Kenntnisse Zelle und wichtige Bestandteile der Zelle Grundbegriffe der Genetik: Gen, Chromosom, DNA, Vererbung Evolutionsmechanismus: Mutation, Selektion Genetik und Verhalten: Zusammenhänge 2. Literatur Grundlage: Schandry, R. (2006). Biologische Psychologie (2. überarb. Aufl.). Weinheim: Beltz. Kapitel 2: Gene und Verhalten. Vertiefend: Hoffrage, U. & Vitouch, O. (2008) Evolutionäre Psychologie des Denkens und Problemlösens. In J. Müsseler (Hrsg.) Allgemeine Psychologie (2., neu bearb. Aufl.)(S. 631-679). Berlin, Heidelberg: Spektrum Verl. Vertiefend zu Grundbegriffen der Biologie wie Biologisches Design, Systemeigenschaften, Systemtheoretische Organismuskonzeptionen, Organismen als Entwicklungssysteme, Leben, Lebewesen, Gen und genetische Information, Evolutionstheorien (auch alternative): Krohs, U. & Toepfer, G. (2005). Philosophie der Biologie. Eine Einführung. Frankfurt a. M.:Suhrkamp. Vertiefend zur Systemtheorie und Kybernetik: Bischof, N. (1995). Struktur und Bedeutung. Eine Einführung in die Systemtheorie für Psychologen. Bern etc.: Huber. Flechtner, H. J. (1984). Grundbegriffe der Kybernetik: eine Einführung. München:dtv. 17 18 Biologische Grundlagen II Informationsübermittlung im Körper Lernziele Die Studierenden sollen das Grundprinzip der Informationsübermittlung im Organismus durch elektrochemische und biochemische Prozesse verstehen lernen, basale Bestandteile wie die Nervenzelle und deren Funktionsweise, aber auch wichtige chemische Botenstoffe (Transmitter) kennen lernen, Aufbau und Funktion des zentralen Nervensystems als wichtigster Steuerungseinheit des Organismus kennen lernen, das vegetative Nervensystem und das Hormonsystem als weitere wichtige, Informationen austauschende und Reaktionen steuernde Systeme in Aufbau und Funktion kennen lernen, Verständnis für die Steuerung und Integration in und durch diese Systeme entwickeln. Psychologie befasst sich mit dem Erleben und Verhalten des Menschen. Eine wichtige Basis für diese Funktionen ist die Kommunikation, die zwischen den verschiedenen Zellen und Zelltypen des menschlichen (aber auch tierischen) Organismus, aber auch zwischen dem Organismus und seiner (physischen und sozialen) Umwelt stattfindet. Umgebungsveränderungen (Reize) führen zu sensorischen Erregungen, die je nach Stärke und Bedeutung für den Menschen zu einer (bewussten) Wahrnehmung dieses Reizes und gegebenenfalls zu einer motorischen Reaktion, für die verschiedene Teile des Körpers abgestimmt werden müssen, führen. Die biologische Grundlage für diesen Austausch ist vor allem das Nervensystem, das in den folgenden Abschnitten in seinen Bestandteilen (Nervenzelle als Baustein, Übertragungsmechanismen, anatomische und funktionelle Struktur) dargestellt wird. Mit dem Hormonsystem wird ein weiteres System vorgestellt, mit dem Informationen innerhalb des Körpers weitergeleitet werden und das Überschneidungen mit dem Nervensystem aufweist und mit diesem zusammenwirkt. II Informationsübermittlung im Körper II.1 Nervenzelle und Übertragungsmechanismen II.1.1 Die Nervenzelle Nervenzellen oder Neurone sind spezialisierte Zellen, die ebenso wie andere Zellen aus einem Zellkern, dem Zellplasma und der Zellmembran bestehen. Ihre Aufgabe ist es, Informationen, zum Beispiel Steuerungssignale an Muskeln („Befehle“) im Organismus weiterzuleiten. Dazu verfügen sie über Auswüchse, die kurzen, baumartig verzweigten Dendriten sowie das langgestreckte, bis zu einem Meter lange Axon, das Kollaterale (Seitenäste) ausbilden kann und bei bestimmten Nervenzellen von einer Myelinscheide umgeben ist. Den Zellkörper eines Neurons bezeichnet man als Zellkörper oder Soma. Die Kontaktstelle zwischen zwei Nervenzellen oder einer Nervenzelle mit einem Effektororgan bezeichnet man als Synapse. Abb. II.1: Schema-Zeichnung einer Nervenzelle (aus: Schandry, 2006, S. 37). Die Formen, die Nervenzellen annehmen können, sind trotz grundlegender Gemeinsamkeiten recht vielfältig, ebenso wie die Bezeichnungen. So handelt es sich bei Pyramidenzellen um pyramidenförmige Neurone, die im Neokortex, einem Teil des Hirns, vorkommen. In diesem Zusammenhang soll auf die Gliazellen verwiesen werden. Es handelt sich um Zellen, die diffus im Nervengewebe verteilt sind. Sie übernehmen Hilfsfunktionen für die Nervenzellen (Ernährung, Stützfunktion). Auch sie sind in ver- 19 Myelinscheide 20 Biologische Grundlagen schiedenen Formen vorhanden. Die Schwann-Zellen genannten Gliazellen bilden die Myelin- oder Markscheide um die Axone von Neuronen aus. Ruhepotential Im Ruhezustand besteht zwischen dem Inneren eines Neurons und der Zellumgebung eine elektrische Spannung von –70 mV. Dies beruht auf der Verteilung bestimmter Ionen (elektrisch geladener Teilchen) innerhalb und außerhalb der Nervenzelle. Dieses Ruhepotential wird durch verschiedene elektrochemische Mechanismen aufrechterhalten. So können bestimmte Proteine in der Membran der Nervenzellen Moleküle entgegen ihrer elektrochemischen Gradienten transportieren (z.B. „Natrium-Kalium-Pumpe“). II.1.2 Reizleitung Wie oben beschrieben, wird in der Nervenzelle durch Membranströme ein Gleichgewicht zwischen dem Zellinneren und der Umgebung der Zelle aufrechterhalten, das sich als Ruhemembranpotential von ca. – 70 mV darstellt. Kommt es nun in der Umgebung der Nervenzelle zu einer Veränderung (einem Reiz), so kann ein zusätzlicher Membranstrom zu einer Verschiebung des Membranpotentials in Richtung positiverer Werte führen. Diese Depolarisation setzt sich entlang der Zellmembran fort und wird durch elektrochemische Mechanismen abgeschwächt und schließlich ausgeglichen. Aktionspotential Erreicht die Depolarisation jedoch einen kritischen Schwellenwert von ca. - 40 mV, kommt es zu einer überschießenden Reaktion, das Potential erreicht sogar positive Werte von etwa +,30 mV. Dieses Aktionspotential folgt einem Allesoder-Nichts-Prinzip, das heißt, unabhängig davon, wieweit die Schwelle der Depolarisation überschritten wird, wird das Aktionspotential ausgelöst; Reizintensitäten werden durch die Frequenz von Aktionspotentialen, nicht durch die Höhe deren Spannung kodiert (Impulsfrequenzkodierung). Dabei zeigt sich ein typischer Verlauf des Spannungsverhältnisses: Nach dem Überschreiten der Schwelle erfolgt ein schneller Anstieg hin zum Maximum, danach erfolgt die Repolarisation, die im Nachpotential das Ruhemembranpotential unterschreitet, bevor das Ruhepotential wieder erreicht wird. Während einer Refraktärphase ist der Bereich nicht (wieder) erregbar. Der gesamte Prozess von Depolarisation bis zur Wiedererreichung des Ruhemembranpotentials läuft in wenigen Millisekunden an. II Informationsübermittlung im Körper 21 Abb. II.2: Verlauf Aktionspotential (aus: Schandry, 2006, S. 63). Durch ein Aktionspotential werden benachbarte Membranbereiche ebenfalls überschwellig erregt, wodurch sich das Aktionspotential über die gesamte Nervenzelle verbreitet. Die Ausbreitung erfolgt linear in eine Richtung, da die Membranbereiche, die sich in der Refraktärphase befinden, nicht wieder erregt werden und die Erregung nicht zurück laufen kann. Die Ausbreitungsgeschwindigkeit nimmt mit der Dicke der Nervenfasern zu. Die dicksten Nervenfasern haben einen Durchmesser von 13 – 20 μm (= Mikrometer oder 10-6m oder eintausendstel mm), die dünnsten 0.2 – 1.5 μm, die dicksten ermöglichen Leitungsgeschwindigkeiten von 80 – 120 m/s, die dünnsten 0.5 – 1.5 m/s (zum Vergleich: Ein Schlag mit einem Baseballschläger auf den Baseball beschleunigt diesen auf eine Geschwindigkeit von 40 m/s). Bis auf die dünnsten Nervenfasern sind alle anderen von einer Myelinscheide umgeben, so dass diese höhere Geschwindigkeiten erreichen als marklose Fasern. Dies wird durch die saltatorische Erregung der myelinisierten Fasern erreicht. An einigen Stellen ist die Myelinscheide von Einschnürungen unterbrochen (Ranviersche Schnürringe); die Erregung „springt“ von Einschnürung zu Einschnürung und erreicht dadurch saltatorische Erregung 22 Biologische Grundlagen die hohen Geschwindigkeiten. Die Klassifikation der Nervenfasern erfolgt nach den drei korrelierten Kriterien Dicke, Geschwindigkeit, Myelinscheide mit Schnürringen. Verwirrenderweise finden sich in der Literatur verschiedene Klassifikationen. In der Abb. II.3 bezeichnen die Großbuchstaben A bis C die sensorischen Nerven, die römischen Ziffern I bis IV dienen zur Klassifikation motorischer Nerven, die hier aber nicht aufgeführt sind. Vielmehr soll nur deutlich werden, welche Faserdicke und Leitungsgeschwindigkeit welcher Klasse motorischer Nerven entspricht. Es kommt hier aber nur darauf an zu wissen, dass es ein Klassifikationssystem gibt, welches es ermöglicht, Nervenfasern nach Dicke und Leitungsgeschwindigkeit zu klassifizieren. Fasertyp Funktion z.B. Mittlerer Faserdurchmesser Mittlere Leitungsgeschwindigkeit [µm] [m/s] Aα (I) Primäre Muskelspindelafferenzen, motorisch zu Skelettmuskeln 15 100 (70-120) Aβ (II) Hautafferenzen für Berührung und Druck 8 50 (30-70) Aγ (III) Motorisch zu Muskelspindeln 5 20 (15-30) Aδ Hautafferenzen für Temperatur <3 15 (12-30) Sympathisch präganglionär 3 7 (3-15) Hautafferenzen für Nozizeption, sympathische postganglionäre Efferenzen 1 1 (0,5-2) B (IV) C Abb. II.3: Klassifikation der Nervenfasern (nach verschiedenen Autoren; angelehnt an Schmidt & Schaible, 2006, S. 221f.). II.1.3 Synapsen und synaptische Übertragung Bislang wurde die Reizleitung innerhalb einer Nervenzelle betrachtet. Nun geht es darum, wie die Übertragung eines Reizes von einer Nervenzelle auf eine weitere Zelle stattfindet. Dieser Mechanismus ermöglicht die Informationsleitung auch zwischen weit auseinander liegenden Körperzellen und die Steuerung von Bewegungen (Reaktionen) über die Muskelzellen. II Informationsübermittlung im Körper Die Verbindungs- oder Schaltstelle zwischen einer Nervenzelle und einer weiteren Nervenzelle oder einem Effektororgan (=Ausführungsorgan) wird als Synapse bezeichnet. Man findet zwei verschiedene Formen von Synapsen im menschlichen Körper. Die elektrische Synapse, an der ein nur geringer Zwischenraum zwischen den beiden Zellen besteht, die Breite beträgt ca. 2 nm (= Nanometer oder 10-9m oder einmillionstel mm). Dieser Zwischenraum ist durch Kontaktmoleküle überbrückt, über die geladene Teilchen von einer Zelle in die andere und auch zurück wandern können. Bei der chemischen Synapse wird der synaptische Spalt von 20 bis 50 nm Breite durch chemische Botenstoffe (Moleküle), die (Neuro-) Transmitter überbrückt. Die Information wird nur in eine Richtung übertragen, daher können die präsynaptische Endigung des informationssendenden Neurons und der postsynaptische Membranbereich des informationsempfangenden Neurons unterschieden werden. Dieser Synapsentyp ist der erheblich häufigere. Abb. II.4: Chemische Synapse. Die chemische Synapse besteht aus einer präsynaptischen Endigung mit transmittergefüllten Vesikeln sowie einer postsynaptischen Membran mit spezifischen Rezeptoren für die jeweilige Transmittersubstanz. Zwischen beiden liegt der synaptische Spalt (aus: Schandry, 2006, S. 72). Erreicht ein Aktionspotential die präsynaptische Endigung, werden die dort in Bläschen (Vesikeln) gespeicherten Transmitter in den synaptischen Spalt freigesetzt, diffundieren in diesen und lagern sich an speziellen Empfängermolekülen des postsynaptischen Bereichs, den Rezeptoren, an. Die Rezeptoren (Empfängermoleküle) reagieren nach dem Schlüssel-Schloss-Prinzip auf den für sie spezifischen Botenstoff oder diesem sehr ähnliche Stoffe. Hat der Ersatzstoff eine 23 Synapse Transmitter 24 Biologische Grundlagen ähnliche Wirkung wie der spezifische Transmitter, spricht man von einem Agonisten. Anhand der Bindungsfähigkeit verschiedener, agonistischer Transmitter lassen sich Subtypen von Rezeptoren unterscheiden. Löst der Ersatzstoff jedoch keine Wirkung aus, blockiert den Rezeptor aber, spricht man von einem Antagonisten. Die Wirkung von Curare und anderer Gifte beruht auf diesem Mechanismus. Abb. II.5 Synaptische Übertragung. Ein Aktionspotential läuft ein. Es kommt zu einem massiven Einstrom von Kalziumionen. Die Lipidmembranen der Vesikel verbinden sich mit der Lipiddoppelschicht der Zellmembran. Die Vesikel öffnen sich zum Extrazellulärraum hin, der Transmitter wird in den synaptischen Spalt ausgeschüttet (aus: Schandry, 2006, S. 73). Der Teil des Transmitters, der nach der Freigabe in den synaptischen Spalt nicht an Rezeptoren gebunden wurde, diffundiert aus dem Spalt, wird enzymatisch abgebaut oder enzymatisch aufbereitet wieder in die abgebende Zelle aufgenommen (Re-Uptake). Diese Prozesse lassen sich pharmakologisch beeinflussen, wenn zum Beispiel der enzymatische Abbau verzögert beziehungsweise gestoppt oder der Re-Uptake verhindert wird. Die Wirkung einer synaptischen Übertragung ist entweder eine Hyperpolarisation des Empfängerneurons (d.h. dessen Membranpotential wird in Richtung eines stärker negativen Werts verschoben) oder eine Depolarisation des Empfängerneurons (d.h. dessen Membranpotential wird in Richtung eines stärker positiven Werts verschoben). Da bei der Hyperpolarisation die Erregungsschwelle deutlich erhöht wird, spricht man von einem inhibitorischen postsynaptischen Potential II Informationsübermittlung im Körper (= IPSP). Im Fall der Depolarisation wird die Erregungsschwelle deutlich gesenkt, üblicherweise spricht man von einem exzitatorischen postsynaptischen Potential (= EPSP). 25 IPSP, EPSP Die Zelle ist im Regelfall von zahlreichen Synapsen mit dicht angeordneten Rezeptoren überzogen. So trifft eine Vielzahl von Informationen (Impulsfrequenzen) auf eine Zelle. Ob sich im Zielneuron ein Aktionspotential ausbildet, hängt davon ab, ob sich die elektrischen Potentiale summieren oder gegenseitig aufheben. Für das Zusammenwirken von Neuronen wurden verschiedene Mechanismen gefunden. Bei der räumlichen Summation werden über mehrere Synapsen erregende Impulse abgegeben bzw. exzitatorische postsynaptische Potentiale ausgelöst, die sich gegenseitig verstärken und ein Aktionspotential im Zielneuron auslösen. Voraussetzung dafür ist die Konvergenz, also das Zusammentreffen mehrerer Nervenfasern auf einem Zielneuron. Bei der zeitlichen Summation werden in so schneller Folge postsynaptische Potentiale erzeugt, dass die Depolarisation beziehungsweise die Hyperpolarisation zunimmt und der Effekt verstärkt wird, unter Umständen bis zur Auslösung eines Aktionspotentials. Diese zeitliche Summation kann durch Prozesse im prä- oder postsynaptischen Bereich begründet sein. Bisweilen kann durch eine schnell wiederholte synaptische Aktivierung jedoch auch der umgekehrte Fall eintreten, dass auf Grund des dann eintretenden Mangels an Transmittersubstanz eine zeitlich begrenzte, reduzierte Erregbarkeit besteht. Die Erregung einer Nervenzelle kann sich auch auf mehrere oder viele andere Zellen verteilen. Voraussetzung dafür ist die Divergenz von Neuronen, also die Ausbildung entsprechender Kontakte einer mit mehreren anderen Zellen. Divergenzen sind bei Neuronen zu beobachten, die Muskelfasern innervieren (Motoneurone). Im Bereich wenig differenzierter Muskeln zum Beispiel im Bereich des Rückens kann ein Motoneuron einige tausend Muskelfasern innervieren. Auch die von Sinneszellen wegleitenden Neurone (Afferenzen) können divergieren; dadurch wird Redundanz geschaffen, indem ein wichtiger Sinneseindruck über mehrere Kanäle vermittelt wird. Bei der Vorwärtshemmung wird durch die Aktivität eines Neurons die Aktivität eines anderen Neurons gehemmt. Möglich wird dies durch den Mechanismus der präsynaptischen Hemmung, wobei das hemmende Neuron eine Synapse kurz vor der axonalen Synapse des erregenden Neurons auf dem Zielneuron hat. Das hemmende Neuron löst ein IPSP an der Synapse des erregenden Neurons aus und hemmt dadurch die Weitergabe eines Reizes des erregenden Neurons an das Zielneuron. Eine Rolle spielt dies bei der Aktivität von Muskeln, die antagonistisch wirken und daher nicht gleichzeitig aktiviert werden sollten, wie der Beuger und der Strecker des Unterarms. räumliche Summation Konvergenz zeitliche Summation Divergenz Vorwärtshemmung 26 laterale Hemmung Biologische Grundlagen Mit lateraler Hemmung wird schließlich ein Mechanismus beschrieben, bei dem sich benachbarte Zellen über ein zwischen ihnen befindliches Neuron (Interneuron) gegenseitig hemmen. Wenn ein Neuron aktiv ist, wird die Erregbarkeitsschwelle seines Nachbarn erhöht. Dieser Mechanismus ist vor allem bei Sinneszellen zu beobachten, er verstärkt Kontraste und lässt Übergänge deutlicher erscheinen. II.1.4 Wichtige Transmitter-Rezeptoren-Systeme Im Folgenden sollen nun einige wichtige Transmitter-Rezeptorensysteme beschrieben werden. Acetylcholin (ACh) ist Transmitter bei der Übertragung von Nerven- auf Muskelzellen, im vegetativen Nervensystem, bei Nervenzellen, die auf Drüsen wirken, und im Gehirn. Zugehörige Rezeptoren sind entweder nikotinerg (= Nikotin wirkt als Agonist, d.h. aktiviert auch den ACh-Rezeptor) mit einer erregenden Wirkung auf Muskelzellen und einer entspannenden auf das Zentrale Nervensystem, oder muskarinerg (= Muskarin wirkt als Agonist). Sie befinden sich im vegetativen Nervensystem, in der Großhirnrinde, dem Striatum und dem Hippocampus. Die Wirkung ist je nach Subtyp des Rezeptors erregend oder hemmend. Die Transmitter Dopamin, Noradrenalin und Adrenalin werden unter den Begriff Katecholamine zusammengefasst. Eng damit verwandt ist das Serotonin. Rezeptoren, die auf Dopamin reagieren, finden sich im Gehirn eher selten, allerdings sind dopaminerge Neuronen weit verzweigt. Ihre Wirkung entfalten sie vor allem bei der Willkürmotorik. Adrenalin ist ein Hormon, das im Nebennierenmark gebildet wird; als Transmitter wirkt es vor allem im Hirnstamm. Noradrenalin wird ebenfalls im Nebennierenmark, aber auch in Neuronen gebildet. Noradrenalin wirkt bei der Übertragung von sympathischen Nerven auf die Erfolgsorgane, darunter innere Organe (Herz, Niere, Bauchspeicheldrüse und Gefäße), Muskelzellen und Zellen des Fettgewebes. Im Gehirn findet sich Noradrenalin im Locus coeruleus. Bei den Rezeptoren, die auf Adrenalin und Noradrenalin reagieren (adrenerge Rezeptoren) unterscheidet man verschiedene Haupttypen mit unterschiedlichen Affinitäten (α1, α2, β1, β2, usw.) und unterschiedlicher, auch gegensätzlicher Wirkung. Diese Rezeptorentypen spielen eine wichtige Rolle im vegetativen Nervensystem, das weiter unten besprochen wird. II Informationsübermittlung im Körper Abb. II.6: Effekte der Aktivierung von Sympathikus und Parasympathikus (aus: Schmidt & Schaible (Hrsg.) (2006). Neuro- und Sinnesphysiologie (5., neu bearb. Aufl.), S.138, mit Genehmigung von © Springer). Serotonin wird in den sogenannten Raphe-Kernen produziert, serotonerge Verbindungen finden sich im gesamten Gehirn und Rückenmark. Serotonin reguliert den Schlaf-Wach-Rhythmus, die emotionale Befindlichkeit, die Schmerzwahrnehmung sowie die Wahrnehmung von Hunger und Durst. Die Aminosäuren Glutamat, GABA (Gamma-Amino-Buttersäure) sowie Glycin sind im ZNS häufiger als Noradrenalin oder Dopamin. Glutamat ist eine exzitatorische Transmittersubstanz, der Wirkung bei Lernvorgängen zugeschrieben wird. GABA hat hemmende Wirkung und wird für beruhigende Medikamente (Tranquillantien) genutzt. Glycin wirkt inhibitorisch. Weiterhin gibt es Hormone, die auch als Neurotransmitter wirken (Neuropeptide): Endorphin, die an Morphinrezeptoren bindenden Enkephaline, Substanz P, Oxytocin, Neuropeptid Y. Sie haben meistens keinen direkten Effekt, sondern 27 28 Biologische Grundlagen modulieren die Wirkung von Transmittern, das heißt sie vermindern oder erhöhen dessen Wirkung. Lernfähigkeit Die Verbindung von Nervenzellen miteinander oder mit Effektorzellen ist nicht statisch, sondern veränderbar. Anders ließen sich Gewöhnungs- oder Lerneffekte nicht erklären. So kann die Reaktion von Rezeptoren bei wiederholter Reizung abnehmen, die Dichte bestimmter Rezeptoren wird herauf- oder heruntergesetzt oder die Zahl der Synapsen, die an einem Vorgang beteiligt sind, nimmt zu oder ab. neuronale Plastizität neuronales Netz Auch kann durch anhaltende Aktivierung das Rezeptormolekül unempfindlich gegenüber dem Transmitter werden (Desensitivierung). Ist ein Transmitter im Überfluss vorhanden, z.B. durch die Einnahme bestimmter Substanzen, stellt das Neuron unter Umständen die Produktion dieses Transmitters ein (DownRegulation). Und schließlich können sich neue Verbindungen zwischen Neuronen bilden, es können durch Aussprossung neue Dendriten „wachsen“ und sich neue Synapsen bilden (neuronale Plastizität). Zusammenfassend lässt sich festhalten, dass im Nervensystem Informationen in Form elektrischer Zustände und Zustandsänderungen codiert, weitergeleitet (Transduktion) und verändert (Transformation) werden. Das Ruhepotential einer Nervenzelle bildet einen „Off-Zustand“ ab, der durch ständige, chemische Prozesse aufrechterhalten wird. Der Off-Zustand lässt sich im dualen Zahlensystem mit Null kodieren und entspricht einem geöffneten Schalter. Informationen treffen auf ein Neuron in Form eines Aktionspotentials, das zu einer elektrochemischen Veränderung an den Synapsen führt, die zu einem Aktionspotential, zum „OnZustand“ des Neurons führen können. Dies würde man im dualen Zahlensystem mit einer Eins kodieren, was einem geschlossen Schalter und damit einer Energieleitung entsprechen würde, die Informationen weiter leiten kann. Die Weitergabe dieser Aktivierung und damit der Informationen an andere Nervenzellen erfolgt ebenfalls auf elektrochemischem Weg über die Synapsen zu anderen Zellen (Nervenzellen oder Effektorzellen) über Transmitter und dazu passende Rezeptoren. Die Koppelung von aktivierten Neuronen bildet ein temporäres neuronales Netz (assembly) und ist ein Erregungsmuster, das wiederum als Ganzes größere Informationseinheiten kodieren kann. Auf einer höheren Ebene können wiederum aktive Nervennetze mit einander gekoppelt werden, so dass temporäre aktivierte Nervennetz-Netze entstehen. Diese dynamische hierarchische Musterbildung ist wahrscheinlich die Grundlage unterschiedlicher Phasen der Integration von Teilinformationen zu einer Struktur, die man phänomenal beispielsweise als bewussten Wahrnehmungseindruck beschreiben könnte. II Informationsübermittlung im Körper 29 II.2 Zentrales Nervensystem (ZNS) Bislang wurden einzelne Neurone und deren Kommunikation mit ihrer Umwelt beziehungsweise mit ihren Nachbarzellen beschrieben. Allerdings stehen die einzelnen Neuronen miteinander in einem großen Informationsverbund, dem Nervensystem. Das Nervensystem erfüllt vielfältigste Aufgaben und erstreckt sich mit seinen Subsystemen in alle Bereiche des Körpers. Eine der möglichen Einteilungen des Nervensystems ist die nach einem peripheren und einem zentralen, steuernden Anteil. Im Folgenden soll nun der steuernde Teil des Nervensystems betrachtet werden. Das Zentrale Nervensystem (ZNS) besteht aus dem Gehirn und dem Rückenmark. II.2.1 Das Rückenmark Das Rückenmark (RM) durchzieht den Wirbelkanal und besteht aus Neuronen. In der schmetterlingsförmigen grauen Substanz befinden sich die Zellkörper von Neuronen, in der sie umgebenden weißen Substanz auf- und absteigende Fasern. Unter den absteigenden Fasern ist besonders die Pyramidenbahn zu nennen: die Axone dieser Bahn sind bis zu einem Meter lang und stellen eine nahezu direkte Verbindung zwischen dem Ursprung dieser Bahn im Cortex (s. Großhirn) und den Motoneuronen dar. Umgeben sind diese Neuronen von der Rückenmarksflüssigkeit und Häuten (Meningen). Jeweils zwischen den Wirbeln verlassen die Spinalnerven das Rückenmark. Die Spinalnerven enthalten sowohl afferente Fasern, also solche, mit denen Informationen aus der Peripherie aufgenommen und zum zentralen Nervensystem geleitet werden, als auch efferente Fasern, mit denen Reaktionen vom zentralen Nervensystem an die ausführenden Organe vermittelt werden. Die afferenten Fasern nehmen die sensorische Informationen auf und treten von der Rückenseite her kommend (dorsal) in das Rückenmark ein. Bauchwärts (ventral) verlassen die efferenten Fasern das Rückenmark und geben die Information für die Muskeln weiter, weswegen sie motorische Nerven genannt werden. Nach ihren Ausgangsoder Zielorganen richtet sich die Unterscheidung in sensorische und motorische Nerven. ZNS 30 Biologische Grundlagen Abb. II.7: Innervation der Haut und Dermatome des Menschen. A. Die afferenten Fasern der Spinalnerven ziehen durch die Hinterwurzeln in das Rückenmark. Die sensorisch innervierten Hautbezirke einzelner Hinterwurzeln (Dermatome) überlappen sich. B. Segmentale Anordnung der sensorischen Innervation. C: Zervikalsegmente, T: Thorakalsegmente, L: Lumbalsegmente, S: Sakralsegmente. C. Anordnung und Ausdehnung einiger Dermatome. Die Überlappung der Dermatome ist für die Berührungsempfindung größer als für die Temperatur- und Schmerzempfindung (aus: Klinke, Pape & Silbernagl (2005), Physiologie (5.Aufl.), S.642, Thieme-Verl.). Die Spinalnerven versorgen (afferent-sensorisch und efferent-motorisch) jeweils einen bestimmten Bereich des Körpers, ein Dermatom. Die Dermatome werden anhand der Wirbelabschnitte des Rückenmarks bezeichnet, beginnend mit den Wirbeln des Halses (Cervicalsegmente 1 bis 8), über die Segmente des Brustraums (Thorakelsegmente 1 bis 12), die Lenden- (Lumbal-)Segmente 1 bis 5 und schließlich die Sakralsegmente (1 bis 5). II Informationsübermittlung im Körper 31 Man unterscheidet drei Gruppen von Neuronen im Rückenmark, Motoneurone, sensible Neurone und Interneurone. Abb. II.8: Typen von Neuronen und Verbindungen im Rückenmark. A Querschnitt (halbseitig) mit den wichtigsten Neuronentypen, grauer (zellreicher) und weißer (zellarmer, Axone) Substanz. Segmentaler Eingang in Hinterwurzeln (afferent), Ausgang durch die Vorderwurzeln (efferent). Absteigende motorische Bahnen vorwiegend ventromedial bzw. Motoneurone dorsolateral. Spinozerebelläre Neurone sind ein Beispiel für aufsteigende Bahnen. B Anordnung der Motoneuronenkerne (und zugehöriger Interneurone, links) im Querschnitt. Motoneurone zu axialen Muskeln ventromedial, zu distalen lateral gelegen (aus: Klinke, Pape & Silbernagl (2005), Physiologie (5.Aufl.), S.739, Thieme-Verl.). Motoneuronen aktivieren die Muskeln des Bewegungsapparates sowie die Muskulatur der inneren Organe und Drüsen. Sie verlassen das Rückenmark in Richtung Bauch (ventral). Sie werden ihrerseits direkt durch efferente Neurone oder durch absteigende Fasern aus den höheren Hirnregionen aktiviert. Sensible Neuronen empfangen Reize aus der Peripherie und leiten diese dorsal (von der Rückenseite her kommend) ins Rückenmark; dort sind sie mit aufsteigenden Neuronen, mit Interneuronen oder mit Motoneuronen verschaltet. Die Interneuronen haben keine Fortsätze aus dem Rückenmark heraus; ihre Aufgabe ist die der Weiterleitung von afferenten oder efferenten Aktivierungen oder deren Verschaltung. Aus der Aktivität der Nervenzellen des Rückenmarks lässt sich dessen Aufgabe im Organismus insgesamt beschreiben. Es ist die Weitergabe von Informationen, von der Peripherie in das Gehirn über die verschiedenen Strukturen des Gehirns (s.u.), die Weiterleitung von Aktivierungen aus dem Gehirn in die Peripherie, aber sensible Neurone Interneurone 32 Biologische Grundlagen auch die direkte Verarbeitung von Informationen aus der Peripherie durch die (nahezu) unmittelbare Verschaltung afferenter mit efferenten Neuronen. Reflex Bei der Verschaltung afferenter Informationen über eine oder nur wenige Synapsen zu den Effektoren, also Muskeln oder Drüsen, ohne den „Umweg“ über das Hirn zu nutzen, spricht man von einem Reflex. Mit Eigenreflex wird ein solcher Reflex bezeichnet, bei denen das Organ der Reizentstehung auch das Organ der reflektorischen Aktivität ist. Eigenreflexe laufen sehr schnell ab und ermüden wenig. Der wohl bekannteste Eigenreflex ist der Patella-Reflex, wobei ein Schlag auf die Sehne unterhalb der Kniescheibe (Patella) deren Aktivität und damit das „Kicken“ des Beines auslöst. Bei einem Fremdreflex sind Reiz- und Effektororgan nicht identisch, es sind mehrere Neuronen an der Entstehung beteiligt, wodurch die Reaktionszeit (im Vergleich zum Eigenreflex) länger wird. Die Reaktionszeit ist die Zeit von der Veränderung des Rezeptors (=Reizung) bis zur Veränderung des Endorgans (Effektors) (=Reaktion). Generell werden Reflexe automatisch ausgelöst, die Reflexreaktion erfolgt sehr schnell und die Bahnung zwischen Reizorgan und Reaktionsorgan ist angeboren und relativ starr angelegt. Ein solcher Reflex ist zum Beispiel der Hustenreflex: ein „Kratzen“ oder trockenes Gefühl im Hals (festgestellt durch Rezeptoren in den Schleimhäuten) führt zu einer motorischen Reaktion (vom Räuspern bis hin zum Husten). Dieser kann bewusst unterdrückt oder abgeschwächt, aber auch gelernt oder verstärkt werden. Husten oder das schnelle Schließen der Augenlider sind Schutzreflexe. Die Bahn vom Reizorgan (Rezeptor) zum Erfolgsorgan (Effektor) wird als Reflexbogen bezeichnet. Diese Grundstruktur eines einfachen neuronalen Schaltkreises (Rezeptor – Afferenz – Efferenz – Effektor) wiederholt sich vielfach im Nervensystem. Einfach ist dieser Schaltkreis deshalb, weil es nur wenige Interneurone zwischen afferenter und efferenter Struktur gibt. Bei vielen Nerven oder gar Nervennetzen zwischen afferenten und efferenten Strukturen kann nicht mehr von einem Reflexbogen gesprochen werden. Es finden vielfache Transformationen statt und es ist meist nicht möglich, einen „Schaltplan“ dieser Zwischenprozesse zu ermitteln. II.2.2 Blut-Hirn-Schranke Das Gehirn Das Gehirn besteht aus Nervenzellen und Gliazellen. An einigen Stellen, den sogenannten Kernen, findet man dicht gepackte Ansammlungen von Nervenzellkörpern. Die Gehirnmasse ist stark gefurcht und ebenso wie das Rückenmark von Flüssigkeit (Liquor cerebrospinalis) umgeben, die auch die Hohlräume (Ventrikel) ausfüllt. Gehirnmasse und Liquor sind von Hirnhäuten (Meningen) umgeben und in den knöchernen Schädel eingepasst. Die Blutversorgung des Gehirns wird über vier große Arterien gesichert. Dabei verhindert die sogenannte Blut-HirnSchranke, dass bestimmte Giftstoffe mit dem Blut zu den Nervenzellen des Gehirns gelangen können. II Informationsübermittlung im Körper 33 Üblicherweise wird das Gehirn von „unten“ nach „oben“ als vom verlängerten Rückenmark bis hin zur „obersten“ Struktur, dem Großhirn (Telencephalon) mit der Großhirnrinde (Kortex), beschrieben. Dieser Logik entspricht auch – in ganz groben Zügen – die Funktionalität des Gehirns und dessen phylogenetische Entwicklung. Während in den unteren Strukturen „einfache“, unbewusst ablaufende, aber für die Lebenserhaltung notwendige Prozesse gesteuert werden (man erinnere sich an die einfachen Reiz-Reaktionsmuster der Reflexe), werden in den „oberen“ Strukturen integrative, unter kognitiver, sogar willentlicher Beteiligung des Menschen ablaufende Kontroll- und Steuerungsmechanismen vermutet. Die räumliche Anordnung der Hauptfunktionskreise des Gehirns, die sensorischen und die motorischen Funktionen, entspricht in den Grundzügen der Anordnung des Rückenmarks. Nach vorne (ventral) befinden sich eher die motorischen Kontrollfunktionen angeordnet, rückwärtig (dorsal) eher die sensorischen Verarbeitungsbereiche. Im Gegensatz zum Rückenmark befinden sich aber zahlreiche komplexe neuronale Netzwerke zwischen diesen beiden Hauptverarbeitungsbereichen. Abb. II.9: Hirnnerven (aus: Schandry, 2006, S. 119). Wie bei der Beschreibung des Rückenmarks deutlich wurde (vgl. Dermatome), wird der menschliche Organismus weitestgehend über Nerven versorgt, die vom oder zum Rückenmark gehen. Darüber hinaus gibt es zwölf Hirnnerven, die nicht 12 Hirnnerven 34 Biologische Grundlagen durch das Rückenmark verlaufen und afferent oder efferent wirken (vgl. Abb. II.9). Sie sind im Wesentlichen für die sensorische und motorische Versorgung von Kopf und Hals und der dort angesiedelten Sinne und Reaktionen zuständig. Darunter ist der Nervus Trigeminus (5. Hirnnerv) zu benennen, der für Sinnesempfindungen in Gesicht und Mund (Zähne!) und für motorische Efferenzen beim Kauen und der Drüsen des Gesichts (Tränen) zuständig ist. Der Nervus vagus (10. Hirnnerv) wirkt über den Bereich von Kopf und Hals hinaus, er ist sowohl sensorisch als auch motorisch für die Eingeweide, motorisch für die Herzmuskeln zuständig. Das Gehirn und Rückenmark gehen anatomisch ineinander über. Vom Rückenmark beginnend werden folgende Strukturen oder Hirnteile unterschieden: − − − − − − Hirnstamm Verlängertes Mark (Medulla oblongata), Brücke (Pons), Mittelhirn (Mesenzephalon), Zwischenhirn (Dienzephalon), Kleinhirn (Zerebellum), Großhirn (Telencephalon). Unter dem Begriff Hirnstamm werden bisweilen Medulla oblongata, Brücke, Mittelhirn und Zwischenhirn zusammengefasst. Wichtig an dieser Stelle ist der Hinweis, dass die Bezeichnungen und „Einteilungen“ des Gehirns vor allem aus dessen anatomisch unterscheidbaren Strukturen, nicht so sehr aus dessen Funktionen abgeleitet werden. Außerdem schließt die oben bereits erwähnte neuronale Plastizität eine 1:1-Struktur- und Funktionsübereinstimmung aus. Und schließlich werden Oberbegriffen wie „Hirnstamm“ oder „Zwischenhirn“ von verschiedenen Autoren teilweise unterschiedlich Substrukturen zugeordnet. Abb. II.10: Die Hauptabschnitte des Gehirns (aus: Schandry, 2006, S. 108). II Informationsübermittlung im Körper 35 Abb. II.11: Gliederungssystematik des Gehirns (aus: Schandry, 2006, S. 109). Im Folgenden sollen wichtige Strukturen des Gehirns kurz beschrieben und ihre Funktion erläutert werden. In der Medulla oblongata kreuzt die Pyramidenbahn, also derjenige Nervenstrang, der sich weiter in das Rückenmark erstreckt und für motorische Efferenzen zuständig ist, größenteils auf die andere Seite. Aus der Medulla oblongata treten sieben der zwölf Hirnnerven aus, darunter der Nervus vagus. Wichtige Funktionen, die hier verortet sind, sind die Steuerung von Atmung und Kreislauffunktion, aber auch Reflexe wie Erbrechen, Schlucken oder Husten sowie die Steuerung des Wach- und Schlafrhythmusses. Medulla oblongata Die Brücke (Pons) besteht zum Teil aus Faserbündeln, die von hier in das Zerebellum ziehen. Es ist Ursprung von vier Hirnnerven, die den Kopfbereich sensorisch und motorisch versorgen, darunter der Nervus trigeminus. Brücke (Pons) Im Mittelhirn (Mesencephalon) befinden sich Schaltstellen des optischen Systems sowie der akustischen und der Schmerzwahrnehmung. Außerdem ist das Mittelhirn an der Steuerung der Bewegung, der Willkürmotorik beteiligt. Mittelhirn Eine sehr interessante Struktur durchzieht den gesamten Hirnstamm: die Formatio retikularis, die ihren Namen ihrer netzartigen Struktur verdankt. Innerhalb dieser Struktur befinden sich die Raphe-Kerne, die Afferenzen aus dem Hypothalamus empfangen und Efferenzen in verschiedene Hirnregionen entsenden. Die Raphe-Kerne sind bei der Schmerzempfindung beteiligt, sie steuern den SchlafWach-Rhythmus und eventuell auch aggressives, emotionales Verhalten. In diesem Gebiet findet sich Serotonin als Transmitter. Eine weitere Substruktur der Formatio retikularis ist der Locus coeruleus, dem eine global aktivierende Funk- (Mesencephalon) Formatio retikularis 36 Biologische Grundlagen tion zugeschrieben wird; hier ist Noradrenalin ein vorherrschender Transmitter. Weitere Teile der Formatio retikularis leiten Impulse nach oben und unten weiter; sie üben eine integrierende Funktion bei basalen vegetativen und grobmotorischen Funktionen aus. Die Formatio retikularis kann als aktivierend-deaktivierendes Zentrum verstanden werden, das entsprechende Impulse nach oben beziehungsweise nach unten weitergibt und integrierend verarbeitet. Es wird angenommen, dass sie bei der Bewusstwerdung peripherer Reize als Filter fungiert. Zerebellum Zwischenhirn (Dienzephalon) Das Zerebellum (Kleinhirn) bildet eine prägnante, fast in sich geschlossene Struktur mit stark gefalteter Oberfläche (Kleinhirn = „kleines Hirn“). Es weist Verbindungen zu den Regionen des Gehirns auf, die mit Motorik befasst sind, da es über Kollaterale („Seitenäste“ / Seitenfasern) mit auf- und absteigenden motorischen Fasern verbunden ist. Es erhält sowohl Informationen aus der Großhirnrinde als auch aus der Peripherie über Lage und Bewegungszustand der Gliedmaße, den Muskeltonus sowie über den Gleichgewichtszustand. Seine Aufgabe liegt vor allem in der Integration dieser Informationen, um Feinabstimmung, körperliche Koordination und zeitliche Taktung vornehmen zu können. Es wird vermutet, dass das Kleinhirn auch an Lernprozessen sowie der Steuerung vegetativer Reaktionen beteiligt ist. Auch das Zwischenhirn (Dienzephalon) wird in verschiedene Strukturen unterteilt: − − − − Thalamus Hypothalamus Thalamus und Methalamus, Epithalamus und Epiphyse, Subthalamus, Hypothalamus und Hypophyse. Der Thalamus steht in Verbindung sowohl mit dem Kortex (also der Rinde des Großhirns) als auch mit dem Hirnstamm. Er sortiert sensorische Informationen (außer denen des Geruchs) vor und leitet sie an die Strukturen des Kortexes weiter, die Informationen aus der Peripherie so verarbeiten, dass sie bewusst werden können. Der Thalamus wird daher etwas poetisch als „Tor zum Bewusstsein“ bezeichnet. Er moduliert den Informationsfluss zwischen den motorischen Zentren des Gehirns und erhält durch abzweigende Nerven eine sogenannte „Efferenzkopie“, das heißt Informationen über efferente Signale aus dem Gehirn an die Peripherie. Vereinfacht gesprochen ist die Efferenzkopie die Kopie eines Bewegungsprogrammes, das mit der Bewegungsprogrammausführung über eine Feedbackschleife verglichen wird. So kann der Organismus feststellen, ob eine Bewegung auf ihn selbst zurückgeht oder nicht. Schließlich wird dem Thalamus Einfluss auf die sogenannten höheren psychischen Funktionen wie Emotion, Motivation und Kognition zugeschrieben. Die Funktion des Hypothalamus besteht in der Steuerung vegetativer Funktionen (siehe auch Kapitel Vegetatives Nervensystem), darunter die Atmung, die Kreislaufaktivität, die Nahrungs- und Flüssigkeitsaufnahme sowie die Körpertempera- II Informationsübermittlung im Körper tur. Die für die Aktivierung der inneren Organe zuständigen Nervensysteme, das parasympathische und das sympathische, werden ebenso durch den Hypothalamus gesteuert wie die Hormonausschüttung. Die dem Hypothalamus untergeordnete Hypophyse (Hirnanhangdrüse) erhält sowohl über Nervenverbindungen als auch durch vom Hypothalamus ausgeschütteten Releasing- bzw. Inhibiting Factors (das sind hormonelle Botenstoffe) Befehle zur Ausschüttung bzw. NichtAusschüttung von Hormonen, die dann direkt oder indirekt wirken (siehe auch Kapitel Hormonsystem). Zwei Hormone, ADH (Adiuretisches Hormon = Vasopressin) und Oxytocin, werden im Hypothalamus selbst gebildet. 37 Hypophyse Bei der Beschreibung des Zwischenhirns soll nun noch auf die Epiphyse (= Zirbeldrüse) hingewiesen werden, die das Hormon Melatonin produziert, dem eine wichtige Bedeutung bei der Regulierung des Schlaf-Wach-Rhythmusses zugeschrieben wird. Das Großhirn (Endhirn, Telenzephalon) besteht aus Großhirnrinde und Großhirnmark. Letzteres ist vor allem für die Modulation der Impulse aus dem Kortex zuständig und ermöglicht koordinierte Bewegungen. Das Großhirn überwächst die anderen Hirnregionen. Es ist durch eine große Furche in zwei Hälften (Hemisphären) geteilt und zeichnet sich durch zahlreiche Sulci (Furchen) und Gyri (Windungen) aus. Der Kortex (Großhirnrinde) ist die flächenhaft ausgebreitete graue Substanz an der Oberseite von Endhirn und Kleinhirn. Großhirn (Telenzephalon) Kortex Abb. II.12: Die wichtigsten Windungen und Furchen der Großhirnrinde (aus: Schandry, 2006, S. 129). Ein Teil des Kortex mit den Substrukturen Hippocampus, Gyrus cinguli, Amygdala und einigen weiteren wird zum limbischen System zusammengefasst. Dieses System ist sowohl mit der Vorverarbeitung sensorischer Informationen als auch der Steuerung von Bewegungen oder vegetativer Funktionen befasst. Darüber hinaus wird ihnen eine Beteiligung an psychischen Vorgängen wie Gedächtnisprozessen, dem Lernen, aber auch bei Emotionen zugeschrieben. limbisches System 38 Biologische Grundlagen Abb. II.13: Limbisches System in zwei Ansichten (aus: Schandry, 2006, S. 136). Neokortex Die oberste Struktur des Kortex ist der Neokortex. Dieser wird üblicherweise in vier Bereiche, die sogenannten Lappen, eingeteilt, denen jeweils spezifische Funktionen zuordenbar sind. Dabei ist diese Zuordnung nicht unveränderbar – sie kann von Individuum zu Individuum variieren und sich (z.B. durch Übung oder nach Verletzungen) auch bei einem Menschen verändern. Auch hier wieder der Hinweis auf die Plastizität des Gehirns, also auf die funktionale Veränderbarkeit von Strukturen. Der Neokortex kann als diejenige Struktur verstanden werden, mit der die anspruchvollsten geistigen Leistungen - sowohl sensorisch-wahrnehmend als auch motorisch-handelnd - gesteuert werden. Zum Neokortex gehören folgende Bereiche: Frontallappen (= Stirnlappen) Parietallappen (= Scheitellappen) Temporallappen (=Schläfenlappen) Okzipitallappen (= Hinterhauptslappen) II Informationsübermittlung im Körper 39 Abb. II.14: Die vier Lappen der Hirnrinde (aus: Schandry, 2006, S. 146). Der Okzipitallappen verarbeitet Informationen aus dem optischen System. In der dem Okzipitallappen zugehörigen primären Sehrinde werden die visuellen Reize zunächst nach physikalischen Eigenschaften wie Farbe, Helligkeit oder Kontrast verarbeitet. In der sekundären Sehrinde werden diese Informationen weiterverarbeitet und visuelles Material „erkannt“ oder klassifiziert, also mit Gelerntem verknüpft. Der Temporallappen übt ähnliche Funktionen für das akustische System aus. In der primären Hörrinde entsteht ein erster Höreindruck, der in der sekundären Hörrinde weiter zu Wörtern, Melodien etc. verarbeitet wird. Einen Großteil der sekundären Hörrinde bildet das Wernicke-Sprachzentrum, das es ermöglicht, dass Sprachsignale wahrgenommen bzw. verstanden werden (Sprachrezeption). Wernicke-Sprachzentrum Der Parietallappen umfasst zu einem großen Teil den primären somatosensorischen Kortex. Zu ihm führen afferente Fasern aus Haut, Sehnen, Gelenken und Muskeln. Der somatosensorische Kortex ist somatotop gegliedert, das heißt, dass jedem Bereich dieses Kortexareals ein bestimmter Bereich des Körpers zugeordnet ist. Dabei werden jedoch nicht die räumlichen Größenverhältnisse abgebildet, sondern das Ausmaß der Sensibilität. So ist die Zunge als ausgesprochen sensibles Organ durch ein fast ebenso großes Areal repräsentiert wie der gesamte Rumpf. Im Frontallappen findet man im motorischen Kortex eine Entsprechung zum sensorischen Kortex; er ist ebenfalls entsprechend der funktionellen Bedeutung im motorischen System somatotop gegliedert. Von hier gehen Bewegungsimpulse an den gesamten Körper. Der prämotorische Kortex bereitet Bewegungen vor und steuert deren sequenzielle Abfolge. Im frontalen Augenfeld werden Augenbewegungen veranlasst. Das Broca-Sprachzentrum bildet die motorische Entsprechung zum Wernicke-Sprachzentrum: Im Broca-Zentrum wird das komplexe Zusammenwirken von Lippen, Zunge, Kehlkopf, Atmung etc. bei der Generierung, Produktion von Sprachsignalen kontrolliert (Sprachproduktion). Der präfrontale Broca-Sprachzentrum 40 Biologische Grundlagen Kortex ist schließlich die Struktur, die mit höheren kognitiven Prozessen in Zusammenhang gebracht wird. Hier ist das Kurzzeit- bzw. Arbeitsgedächtnis verortet, aber auch die Vorbereitung komplexer Handlungen hat ihr neuronale Korrelate. Durch den präfrontalen Kortex werden emotionale und motivationale Impulse sowie Konzentration und Antrieb gesteuert. Und nicht zuletzt lassen sich hier die neuronalen Korrelate der Fähigkeit des Menschen zu ethisch-moralischer Einsicht und Handlungsfähigkeit verorten. Brodmann-Areale Neben der Einteilung des Kortex anhand der Grobstruktur in Lappen wird zunehmend eine Einteilung der Hirnrinde verwendet, die sich an Gestalt und Struktur der Nervenzellen orientiert. Der Neuroanatom Brodmann hat den Kortex schon um 1900 in 47 Areale eingeteilt, die ihm zu Ehren Brodmann-Areale genannt werden und die bis heute verwendet werden, um Funktionsbereiche im Neokortex zu lokalisieren. Diese Einteilung stimmt großenteils mit der Einteilung überein, die durch die Furchen und Windungen vorgegeben ist, ist aber feiner und für die heutigen Erfassungsmethoden der Hirnaktivität daher besser geeignet (siehe auch Kapitel Messmethoden). Abb. II.15 Brodmann-Areale. Oben: Ansicht lateral von links. Unten: Ansicht medial von links (aus: Schandry, 2006, S. 146). II Informationsübermittlung im Körper 41 II.3 Das Vegetative Nervensystem (VNS) Das viszerale, autonome oder vegetative Nervensystem (VNS) ist für die Steuerung der inneren Organe wie Herz, Lunge oder Verdauungsorgane sowie Drüsen und Blutgefäße zuständig. Die Bezeichnung „autonom“ weist darauf hin, dass ein Großteil dieser Aktivität unbewusst und ohne willentliche Steuerung abläuft. Das vegetative Nervensystem ist, wie oben bereits beschreiben, kein vom zentralen Nervensystem unabhängiges System, sondern wird durch Strukturen des Gehirns gesteuert. Durch Hormone und Nerven steuert der Hypothalamus vegetative Prozesse. Er steht in Verbindung mit einer ganzen Reihe weiterer Hirnstrukturen, darunter kortikale Strukturen, Kleinhirn, Formatio retikularis und limbisches System. Der 10. Hirnnerv, der Nervus vagus, der der Medulla oblongata entspringt, leitet Informationen aus dem Verdauungstrakt in das Hirn und aktiviert Eingeweide, Herz und Kehlkopf. Die Leitungsgeschwindigkeit im VNS ist im Gegensatz zum willkürlichen Nervensystem geringer, da die Nervenfasern nicht myelinisiert sind, häufiger verschaltet werden und längere Übertragungszeiten an den Synapsen aufweisen. Das vegetative Nervensystem besteht aus einem sympathischen und einem parasympathischen Anteil (Sympathikus und Parasympathikus), die sich hinsichtlich des Verlaufs der Nerven und ihrer Funktion unterscheiden. Daneben wird das Darmnervensystem zunehmend als eigenständiger Teil des VNS betrachtet. Aufgabe des Darmnervensystems ist die Steuerung des Verdauungssystems mit der Kontrolle über die Darmbewegungen und die Sekretions- und Absorptionsprozesse in Leber und Bauchspeicheldrüse. Die meisten, vom VNS gesteuerten Organe werden sowohl von sympathischen als auch von parasympathischen Nerven versorgt, Ausnahmen bilden die Schweißdrüsen und die meisten Blutgefäße, die ausschließlich sympathisch gesteuert werden. Wesentliche Transmitter in diesen Systemen sind Noradrenalin und Acetylcholin mit muskarinergen und nikotinergen Rezeptoren. Das Katecholamin Noradrenalin entsteht nicht nur an den Synapsen, sondern wird auch vom Nebennierenmark ausgeschüttet und unterliegt damit auch nicht-zentralen Steuerungsmechanismen. Eine Aktivierung des Sympatikus führt zum Anstieg der Herzleistung, des Blutdrucks, der Körpertemperatur, des Blutzuckerspiegels und einer Erweiterung der Pupillen. Da damit die Voraussetzungen für körperliche Aktivität geschaffen werden, spricht man von einem ergotropen (leistungsvorbereitenden) Zustand. Der Parasympathikus führt hingegen zu einem trophotropen (erholungsstützenden) Zustand, wobei die Pulsfrequenz sinkt und die Magen-Darm-Motilität steigt. Die beiden Systeme ergänzen sich, arbeiten jedoch nicht antagonistisch (vergleiche Tabelle II.6 Effekte der Aktivierung von Sympathikus und Parasympathikus). Sympathikus, Parasympathikus 42 Biologische Grundlagen Effektor des vegetativen Nervensystems ist unter anderem die glatte Muskulatur der Gefäße sowie des Magen-Darmtrakts. Ihr Aufbau und ihre Funktionsweise werden im Kapitel „Muskelzelle“ beschrieben. Fast alle Fasern des vegetativen Systems verfügen auch über Afferenzen, die meist zur Ebene der Medulla oblongata führen. Projektionen in höhere Gebiete des Gehirns führen zu Hypothalamus, Thalamus sowie zum somatosensorischen Kortex. Dabei geschieht es, dass bei der bewussten Wahrnehmung von Schmerzen z.B. im Darmsystem diese auf die Körperoberfläche projiziert und dort zu motorischen Reaktionen führen (z.B. dem Anspannen der Bauchdecke). Diesen Effekt kann man umgekehrt in der Behandlung nutzen, wenn durch eine Wärmflasche Bauchschmerzen gelindert werden. Aus dem bisherigen wird deutlich, dass das vegetative Nervensystem die inneren Organe zu dem Zweck steuert, Energie bereitzustellen oder verbrauchte Energie wieder aufzubauen, kurz ein Gleichgewicht (Homöostase) zwischen Verbrauch und Bereitstellung von Energie zu halten. Vegetative Funktionen und psychische Prozesse sind miteinander verknüpft. Dass starke Gefühle (Emotionen) „auf den Magen schlagen“ können, ist bereits im Alltagswissen verankert, die Verbindungen des Vegetativen Nervensystems mit Strukturen des Gehirns und dem Hormonsystem bilden das somatische Korrelat. Auch das Phänomen „Stress“ (siehe auch Kapitel Funktionen) zeigt den Zusammenhang psychischer mit vegetativen Funktionen. Veränderungen im VNS wirken sich schließlich auch auf das Immunsystem aus, da an der Immunabwehr beteiligte Organe wie Milz oder Lymphknoten beeinflusst werden oder die Transmitter des VNS direkt an Immunreaktionen beteiligt sind. Durch diesen Zusammenhang können die Beziehungen zwischen (somatischen) Erkrankungen und psychischen Ursachen beziehungsweise Auslösern begründet werden. II.4 Das Hormonsystem (Endokrines System) Bei der Besprechung des Nervensystems wurde an verschiedenen Stellen bereits das Hormonsystem erwähnt, so bei der Beschreibung des Hypothalamus als zentralem Steuerungsorgan des Hormonsystems und bei der Darstellung der synaptischen Übertragung, wobei Hormone als Transmitter wirken können. Als Neuromodulatoren werden Hormone bezeichnet, wenn sie nicht als Transmitter wirken, sondern die Wirkung von Transmittern verstärken oder dämpfen. Hormone werden in bestimmten Zellen synthetisiert, ins Blut abgegeben und über den Blutkreislauf zu den Zielzellen befördert. Anhand ihrer chemischen Struktur lassen sich zwei Gruppen von Hormonen unterscheiden. Lipophile Hormone können direkt in die Zielzelle gelangen; hydrophile Hormone docken an der II Informationsübermittlung im Körper 43 Membran einer Zelle an und aktivieren dort weitere Vermittlungsstoffe (sog. Second Messenger). Auch hinsichtlich ihrer Übertragungswege werden hormonelle Botenstoffe unterschieden. So können Botenstoffe Signale an die sezernierende Zelle selbst (autokrine Übertragung) oder benachbarte Zellen senden (parakrine Übertragung). Mit endokriner Übertragung wird der Vorgang beschrieben, wenn die Hormone von bestimmten Zellen sezerniert und über das Blut in weit entfernte Zielzellen gelangen. Und schließlich gibt es die Möglichkeit, dass Nervenzellen Empfänger oder sogar Sender von Hormonen sind (neuroendokrine Übertragung). Der Hypothalamus als oberstes Steuerorgan setzt Hormone frei, die als Releasinghormone wiederum die Freisetzung von Hormonen aus dem Hypophysenvorderlappen bewirken oder als inhibitorische Releasinghormone deren Freisetzung hemmen. Die Hormone des Hypophysenvorderlappens wirken ihrerseits entweder direkt auf Zielzellen (nicht-glandotrope Hormone) oder sie regen die Tätigkeit endokriner Drüsen an (glandotrope Hormone). Der Hypophysenhinterlappen wird durch den Hypothalamus neuronal gesteuert. Dort werden vom Hypothalamus produzierte Hormone gespeichert und bei Bedarf ausgeschüttet. Abb. II.16: Regulation der endokrinen Drüsen. a Halbanatomische Darstellung. HVL Hypophysenvorderlappen (Adenohypophyse). HHL Hypophysenhinterlappen (Neurohypophyse). b Regulation der Konzentration eines endokrinen Hormons im Blut durch den Hypothalamus (graue Pfeile) und die neuronale Beeinflussung dieser Regulation durch andere ZNS-Bereiche (schwarze Pfeile). Beachte, dass periphere Hormone auch zentrale Neurone außerhalb des Hypothalamus beeinflussen. RH Releasing-Hormon, HVL-Hormon Hypophysenvorderlappenhormon (aus: Schmidt & Schaible (Hrsg.) (2006). Neuro- und Sinnesphysiologie (5., neu bearb. Aufl.), S.178, mit Genehmigung von © Springer). endokrine Übertragung Releasinghormone 44 Biologische Grundlagen Nach der Art eines Regelkreises mit negativer Rückkopplung werden im Hypothalamus Informationen über den Hormonstatus spezifischer Hormone verarbeitet. Bei einem Absinken der Konzentration wird das Hormonsystem angeregt, die Produktion und Sezernierung zu fördern, bis der Sollwert wieder erreicht ist. Die eigentliche Produktion von Hormonen geschieht durch Synthese in den produzierenden Zellen oder die Verwandlung von Vorgängerstoffen, die z.B. aus der Nahrung aufgenommen werden. Die Verfügbarkeit von Hormonen im Organismus kann durch verschiedene Mechanismen verändert werden. Die produzierenden Zellen können aktiviert beziehungsweise desaktiviert werden oder die Aktivität von Syntheseenzymen kann verändert werden. Durch die Bindung von Hormonen an Speicher- oder Transportproteine wird die Verfügbarkeit erhöht. Und schließlich kann die Abbaugeschwindigkeit durch die Aktivierung oder Desaktivierung von Abbauenzymen verändert werden. Hypophysenhormone Im Folgenden werden nun die wichtigsten Hormone des menschlichen Körpers vorgestellt, zunächst die beiden nicht-glandotropen Hormone des Hypophysenvorderlappens, das Wachstumshormon und das Prolaktin. Abb. II.17: Übersicht über Hypophysenvorderlappenhormone (aus: Schmidt & Schaible (Hrsg.) (2006). Neuro- und Sinnesphysiologie (5., neu bearb. Aufl.), S.179, mit Genehmigung von © Springer). II Informationsübermittlung im Körper Das in dem Hypophysenvorderlappen gebildete Wachstumshormon (Somatotropin) ist verantwortlich für die menschlichen Wachstumsprozesse durch seine Wirkung vor allem auf die Knorpelzonen des Körpers, Muskeln und Knochen. Beim Erwachsenen beeinflusst das Wachstumshormon die Energiebereitstellung. Das die Ausschüttung von Somatotropin inhibierende Hypothalamus-Hormon Somatostatin wird als Neuromodulator wirksam. Die Wirkung des Hypophysenvorderlappen-Hormons Prolaktin besteht in der Anregung des Brustwachstums bei der Frau in der Pubertät sowie in der Stimulation der Milchdrüsen nach der Geburt. Bei Stress kann der Prolaktinspiegel ansteigen, eine Auswirkung auf das Immunsystem wird diskutiert. Die gonadotropen Hormone, die vom Hypophysenvorderlappen gebildet werden, sind das follikelstimulierende Hormon (FSH) und das Luteinisierungshormon (LH). Sie wirken auf die Keimdrüsen von Männern und Frauen, aber auch auf Nervenzellen vor allem des limbischen Systems und des Hypothalamus. Sie steuern damit das Sexualverhalten beziehungsweise dessen Voraussetzungen. Das follikelstimulierende Hormon bewirkt bei der Frau die Reifung des Follikels und (gemeinsam mit dem Luteinisierungshormon) die Biosynthese des Östrogens. Beim Mann regt es die Spermienproduktion an. Das Luteinisierungshormon aktiviert bei der Frau die Östrogenproduktion. Es löst den Eisprung aus und regt die Gelbkörperbildung an. Beim Mann führt es zur Synthese und Ausschüttung der Androgene (männlichen Sexualhormone), darunter das Testosteron. Menstruationszyklus Der Verlauf des weiblichen Menstruationszyklus wird durch die beiden Hormone beeinflusst. In der Follikelphase (bis zum 12. Tag des Zyklus) fördert das follikelstimulierende Hormon die Reifung einer Eizelle: Es bildet sich eine Schicht um das Ei, die Östrogene, v.a. Östradiol, erzeugt. Es folgt eine vermehrte Freisetzung des follikelstimulierenden sowie des Luteinisierungshormons, wobei letzteres schließlich dazu führt, dass der Follikel platzt (Eisprung). In der Lutealphase (16. bis 28. Zyklustag) wird der geplatzte Follikel zum Gelbkörper umgebildet und produziert Progesteron und Östrogene. Kam es nicht zu einer Befruchtung, werden schließlich die Gebärmutterschleimhaut sowie die Eizelle abgestoßen. Ein weiteres, vom Hypophysenvorderlappen gebildetes, glandotropes Hormon ist das Adrenocorticotrope Hormon (ACTH). Es wirkt auf die Nebennierenrinde und regt dort die Synthese der Nebennierenrindenhormone wie Kortisol an. ACTH weist eine deutliche circadiane Rhythmik auf und wird bei Stress vermehrt ausgeschüttet; Zusammenhänge mit depressiven Erkrankungen werden diskutiert. 45 46 Biologische Grundlagen Und schließlich wird vom Hypophysenvorderlappen das Thyreotropin sezerniert, das das Wachstum der Schilddrüse, deren Hormonproduktion und Ausschüttung anregt. Bei den beiden nun zu besprechenden Hormonen des Hypophysenhinterlappens handelt es sich um Adiuretin und Oxytocin. Das Adiuretin oder Vasopressin verhindert die Wasserausscheidung durch die Niere und lässt die glatte Muskulatur kontrahieren, was den Blutdruck erhöht sowie die Darmperistaltik verstärkt. Vasopressin wird auch Einfluss auf Lern- und Gedächtnisprozesse zuerkannt. Auch dem anderen Hypophysenhinterlappenhormon, dem Oxytocin, wird eine Rolle bei Gedächtnis- und Lernleistungen zugeschrieben. Daneben erhöht es das Brutpflege- und Paarungsverhalten. Unmittelbare Wirkung zeigt das Oxytocin bei der Geburt: Es leitet die Geburt durch die Anregung der Uteruskontraktionen (Wehen) ein und fördert die Milchabgabe aus der Brustdrüse. Im Folgenden werden die Hormone beschrieben, die in endokrinen Drüsen produziert werden, bevor abschließend auf einige Hormone eingegangen wird, die nicht in eigenständigen Organen produziert werden. Abb. II.18: Endokrine Drüsen und diffuses endokrines System (aus: Klinke, Pape & Silbernagl (2005), Physiologie (5.Aufl.), S.513, Thieme-Verl.). II Informationsübermittlung im Körper Hormone der Bauchspeicheldrüse Die Bauchspeicheldrüse (Pankreas) produziert unter anderem Insulin, das die Glukosekonzentration im Blut reduziert und die Proteinsynthese fördert. Damit wird Insulin zu einem wichtigen anabolen Enzym, also zu einem Stoff, der dafür zuständig ist, vom Körper verbrauchte Energie wieder aufzubauen. Der direkte Gegenspieler des Insulins, das Glukagon, wird ebenfalls in der Bauchspeicheldrüse produziert. Die Aktivität von Insulin und Glukagon wird nicht durch Hormone gesteuert, sondern, vermittelt über zentralnervöse Prozesse, durch die Verfügbarkeit von Blutzucker im Blut. Hormone der Schilddrüse In der Schilddrüse werden Thyroxin und Trijodthyronin aus der Aminosäure Tyrosin und Jod gebildet, die beide zu einer Aktivierung des Organismus führen. Sie erhöhen den Zucker- und Fettabbau, steigern Herz- sowie Atemaktivität und regen die Synthetisierung von Enzymen und Proteinen an. Damit steigern sie den Grundumsatz des Organismus. Hormone des Nebennierenmarks Im Nebennierenmark, das aus modifizierten Nervenzellen besteht, werden die beiden Katecholamine Adrenalin und Noradrenalin gebildet. Wie bereits beschrieben vermitteln sie die Wirkung des Sympathikus (siehe auch das Kapitel Vegetatives Nervensystem). Diese Stoffe steigern die Leistung des HerzKreislaufsystems, erweitern die Bronchien und senken die Aktivität des MagenDarm-Trakts. Zusammenfassend wird die Funktion der Katecholamine als Reaktion auf Gefahrensituationen interpretiert, mit der eine Flucht oder ein Kampf vorbereitet wird. Hormone der Nebennierenrinde In der Nebennierenrinde werden drei Gruppen von Hormonen gebildet, Die Glukokortikoide, darunter das Kortisol, erhöhen die Blutzucker-Konzentration, hemmen Entzündungen und können, bei längerer Ausschüttung, immunsuppressiv wirken. Die Mineralkortikoide fördern die Wasserrückresorption durch die Niere, wodurch Blutvolumen und Blutdruck steigen. Und schließlich werden hier, 47 48 Biologische Grundlagen wenn auch in deutlich geringerem Umfang als durch die Keimdrüsen, männliche Geschlechtshormone (Androgene) produziert. Hormone der Keimdrüsen Die Hormone der Keimdrüsen, darunter das Östrogen und das Testosteron, bewirken die Ausbildung der primären und sekundären Geschlechtsmerkmale während der Pubertät. Bei beiden Geschlechtern wächst die Schambehaarung. Bei den Mädchen entwickelt sich die Brust, die Hüften werden breiter und die erste Monatsblutung (Menarche) findet statt. Jungen kommen in den Stimmbruch, die Schultern werden breiter und der Bartwuchs setzt ein. Ab etwa dem 45. Lebensjahr kommt es auf Grund des Rückgangs der Östrogenproduktion durch die Eizellen bei der Frau zum Klimakterium, das durch eine letzte Monatsblutung, die Menopause, gekennzeichnet wird. Beim Mann geht etwa ab dem 55. Lebensjahr die Produktion von Testosteron zurück. Die Keimdrüsenhormone sind wesentliche Voraussetzung für das Sexual- und Fortpflanzungsverhalten. Da diese Hormone die Blut-Hirn-Schranke passieren können, wird hier ein Weg gesehen, mit dem spezifisch männliches oder weibliches Verhalten begründet werden kann. Hormone, die nicht in eigenständigen Organen produziert werden Abschließend sollen nun einige weitere Hormone vorgestellt werden, die nicht in eigenständigen Hormondrüsen produziert werden. So werden im Magen-DarmBereich verschiedene Stoffe hergestellt, die Verdauungs- und Resorptionsprozesse ermöglichen. In der Niere wird Erythropoetin sezerniert, das die Bildung roter Blutkörperchen, die für den Sauerstofftransport im Blut zuständig sind, fördert. Das in der Nebenschilddrüse gebildete Parathormon fördert die Verfügbarkeit von Kalzium, der Gegenspieler, das Kalzitonin, wird in der Schilddrüse produziert. Nochmals sei an dieser Stelle auf die Epiphyse, eine Verlängerung des Zwischenhirns, hingewiesen, die Melatonin produziert, dem eine wichtige Funktion bei der Steuerung des Tag-Nacht-Rhythmusses zugeschrieben wird. Gewebehormone, also Hormone, die in verschiedenen Geweben produziert werden, spielen eine Rolle bei Entzündungsreaktionen, bei der Blutgerinnung oder auch beim Fettabbau. Auch das Histamin wird in Geweben produziert, das als Entzündungsmediator oder Schmerzstoff z.B. bei der Reaktion auf Insektenstiche aktiv wird und unter Umständen Asthmaanfälle auslösen kann. II Informationsübermittlung im Körper Studientechnischer Anhang 1. Kenntnisse Nervenzelle o Aufbau und Bestandteile o Funktionsweise der Nervenzelle (Ruhepotential, Aktionspotential und dessen Verlauf, Alles-oder-Nichts-Prinzip) Synaptische Übertragung o Aufbau chemischer Synapse o Transmitter und Rezeptoren Zusammenwirken von Neuronen o Mechanismen (EPSP, IPSP) o Formen (Summation, Konvergenz, Divergenz, Hemmung) Rückenmark (als Bestandteil des ZNS) o Aufbau und Bestandteile o Reflexe (Mechanismus und Formen) Zentrales Nervensystem - Gehirn o Aufbau und Unterteilungsmöglichkeiten (Brodmann-Areale) o Teile sowie Strukturen des Gehirns und deren Funktion (Medulla Oblongata, Hirnnerven, Kleinhirn, Brücke, Mittelhirn, Formation Retikularis, Thalamus, Hypothalamus, Hypophyse, Kortex, limbisches System, Neokortex) Vegetatives Nervensystem o Bestandteile: Sympathikus, Parasympathikus, Darmnervensystem o Transmitter (Adrenalin, Noradrenalin, etc.) und Funktionen Hormonsystem o Regulation und Regulationsmechanismen (Hypothalamus, Hypophyse, Releasing-Hormone, Prinzip der negativen Rückkopplung) o Hormongruppen (Hypophysenhormone, Hormone endokriner Drüsen, weitere Hormone) o Beispiele für Hormone (Wachstumshormon, Gonadotrope Hormone, Hormone der endokrinen Drüsen, darunter Sexualhormone) und deren Wirkung 49 50 Biologische Grundlagen 2. Literatur Grundlage: Schandry, R. (2006). Biologische Psychologie (2. überarb. Aufl.). Weinheim: Beltz. Kapitel 3: Bausteine des Nervensystems – Neuronen und Gliazellen, bes. Abschnitt 3.1.2 Kapitel 4: Die zelluläre Basis der Informationsverarbeitung im Nervensystem, bes. Abschnitte 4.1 und 4.5 Kapitel 5: Zusammenwirken von Nervenzellen – Informationsübertragung und –verarbeitung, bes. Abschnitte 5.1, 5.2 und 5.4 Kapitel 6: Aufbau und Funktion des Nervensystems Kapitel 7: Die Steuerung vegetativer Funktionen, bes. Abschnitte 7.2, 7.3 und 7.4 Kapitel 8: Das Hormonsystem, bes. Abschnitt 8.5 Vertiefend: Dudel, J. (2006) Innerneurale Homöostase und Kommunikation, Erregung. In R.F.Schmidt & H.-G. Schaible (Hrsg.), Neuro- und Sinnesphysiologie (S. 14-42). Heidelberg:Springer. Dudel, J. (2006) Synaptische Übertragung. In R.F.Schmidt & H.-G. Schaible (Hrsg.), Neuro- und Sinnesphysiologie (S. 43-64). Heidelberg:Springer. Jänige, W. J. (2006). Vegetatives Nervensystem. In R.F.Schmidt & H.-G. Schaible (Hrsg.), Neuro- und Sinnesphysiologie (S. 132-181). Heidelberg:Springer. III Motorik 51 III Motorik Lernziele Die Studierenden sollen den Aufbau von Muskeln kennen lernen, das Zusammenwirken von Nerven und Muskeln kennen lernen, einfache und komplexe Steuerungsmechanismen und die zugehörigen Strukturen kennen lernen, Verständnis für das Zusammenwirken steuernder, kontrollierender und taktender Prozesse bei der Ausführung von Bewegungen entwickeln, Verständnis für die Komplexität der Bewegungssteuerung mit der Abfolge von Aktion – Ruhe, Grob- und Feinmotorik, Planung und Ausführung unter Berücksichtigung der Umgebung entwickeln. Bewegung ist Grundvoraussetzung und Begleiterscheinung des menschlichen Lebens. Bewegung beginnt mit einfachen Reflexen und reicht hin zu den komplexen Mustern, die ein Tänzer ausführt, wenn er ein Lied singt und dazu tanzt. Ohne aktive Bewegungen könnten Menschen sich nicht im Raum von Ort zu Ort bewegen und nicht miteinander kommunizieren, nicht miteinander sprechen. Bewegung heißt, dass die Masse des Körpers in eine bestimmte Richtung beschleunigt werden muss. Es muss also Kraft (= Masse * Beschleunigung; Einheit= Newton; 1 N = kg*m*s-2) ausgeübt werden. Ohne Kraftaufwand kann weder Arbeit (= Kraft * Weg; Einheit Joule) noch Leistung (= Arbeit pro Zeit; Einheit Watt) erbracht werden. Die Kraft wird in den Muskeln erzeugt, die über die Sehnen auf die (starren) Knochen wirkt. Menschen sind Selbstbeweger, d. h. die Kraft wird in einigen Körperteilen erzeugt und wirkt von da aus auf die Umgebung, die als reactio nach den Gesetzen der Mechanik zurückwirkt und so die Ortsveränderung (Lokomotion) erlaubt. Die Bewegungen des Körpers, die grundsätzlich willentlich beeinflussbar sind, werden als Willkürmotorik bezeichnet. Bei der Beschreibung der Steuerung einzelner Bewegungen wird deutlich werden, warum diese Bezeichnung nur teilweise zutrifft. Aufgrund der Struktur der Muskelzellen wird die Muskulatur dieses Systems als gestreifte Muskulatur bezeichnet. Diese Muskeln sind über Sehnen mit dem Skelett verbunden und bewegen einzelne Knochen, die an Gelenken, meist nur in vorgegebenen Richtungen, gegeneinander bewegt werden können. Daher wird für diesen Teil des Muskelsystems auch der Überbegriff Skelettmuskulatur verwendet. Willkürmotorik 52 unwillkürliche Motorik motorische Koordination Biologische Grundlagen Neben dem System der Willkürmotorik gibt es ein weiteres Bewegungssystem, das sich einer willentlichen Beeinflussung weitestgehend entzieht. Es handelt sich dabei um die Bewegungen, die von den Blutgefäßen oder Magen und Darm ausgeführt werden. Bei der Besprechung des Vegetativen Nervensystems wurde dieses System und seine Steuerung bereits vorgestellt; es wird in diesem Kapitel nicht vertiefend betrachtet, lediglich der Aufbau seiner Muskelzellen wird beschrieben. Im Unterschied zur oben genannten gestreiften Muskulatur sind die Muskelzellen der unwillkürlichen Motorik glatt und man spricht von der glatten Muskulatur. Es wird also nicht in einem punktuellen Zentrum des Körpers Kraft erzeugt und ausgeübt, sondern verschiedene Kraftquellen müssen so in Raum und Zeit koordiniert werden, dass komplexe Bewegungen wie Aufstehen, Klettern, Greifen oder Sprechen möglich werden. Es gibt also zahlreiche Körperbewegungen, die nicht der Lokomotion (Ortsveränderung) dienen wie bspw. das Sprechen oder das Schreiben. Auch die Einnahme einer stabilen Haltung wie Stehen oder den Kopf aufrecht halten, aber auch das Bewegen der Augen erfordert Kraft. Diese Bewegungsprobleme werden unter dem zentralen Begriff der motorischen Koordination zusammengefasst. Unter motorischer Koordination versteht man „die reibungslose, aufeinander abgestimmte Durchführung von Bewegungen. Bei jeder normalen Bewegung ist eine Vielzahl von Muskeln beteiligt, deren Spannung und Kraft in einem zeitlich genau aufeinander abgestimmten Muster variieren muss. Die zeitliche Koordination von Anfang und Ende des Kontraktionsgrads verschiedener Muskelanteile ist Voraussetzung für jede komplexe Bewegung. Verantwortlich für die motorische Koordination ist das Kleinhirn (…)“ (Hanser, H. (2001) (Red.), Lexikon der Neurowissenschaft, Eintrag zum Stichwort motorische Koordination). III.1 Muskelzellen und Muskulatur Muskelzellen sind Körperzellen, die so aufgebaut sind, dass sie ihre Form ändern können. Möglich wird dies durch Aktin- und Myosinfilamente, die sich gegeneinander verschieben können und dabei die Kontraktion eines Muskels erzeugen oder diese lösen. Anhand ihrer Form und Anordnung unterscheidet man die gestreifte und die glatte Muskulatur. Muskelzellen der quergestreiften Muskulatur Mit der quergestreiften oder Skelettmuskulatur werden Kräfte über Sehnen auf unsere Knochen übertragen und damit Bewegung erzeugt. Häufig arbeiten Muskeln antagonistisch, das heißt, ein Muskel bewegt einen Knochen in eine, sein Antagonist in die andere Richtung, zum Beispiel der Beuger und der Strecker des Arms. III Motorik 53 Ein Skelettmuskel besteht aus einer Reihe von auch mit dem bloßen Auge zu erkennenden Fleischfasern. Diese sind jeweils aus 0,1 bis 0,01 mm breiten Muskelfasern, den Muskelzellen, zusammengesetzt. Abb. III.1: Feinbau des Muskels (aus: Schandry, 2006, S. 195). Das Innere einer Muskelzelle der quergestreiften Muskulatur besteht größtenteils aus Myofibrillen. In den langgestreckten Myofibrillen sind Aktin- und Myosinfilamente jeweils so angeordnet, dass sie sich gegeneinander verschieben und damit den Muskel kontrahieren, also zusammenziehen bzw. verkürzen können. Die Myofibrillen weisen eine charakteristische Bänderung auf, die im Mikroskop zu erkennen ist, und geben der quergestreiften Muskulatur ihren Namen. Motorische Einheiten Die Muskelfasern der quergestreiften Muskulatur werden durch Motoneurone innerviert. Die α-Motoneurone sind Motoneurone, deren Axone die Vorderwurzel des Rückenmarks verlassen. Ein Motoneuron kann nur eine, aber auch bis zu 1000 einzelne Muskelfasern versorgen; mit motorischer Einheit wird die Gesamtheit aller von einem Motoneuron versorgten Muskelfasern bezeichnet. Je kleiner eine motorische Einheit ist, desto feinere Bewegungen werden damit möglich. Die Synapse des Motoneurons auf der Muskelfaser ist die motorische Endplatte, motorische Einheit 54 Biologische Grundlagen Transmitter hier ist das Acetylcholin. Jede Muskelfaser wird nur von einem Motoneuron innerviert. Ist ein Motoneuron aktiv, aktiviert es alle Muskelfasern seiner motorischen Einheit, Abstufungen der Intensität werden über die Veränderung in der Zahl der aktiven Motoneurone und die Frequenz deren Entladungen abgebildet. Muskelzellen der glatten Muskulatur Auch die Muskelzellen der glatten Muskulatur, zum Beispiel der Blutgefäße oder des Darms, verfügen über Aktin- und Myosinfilamente. Diese Muskelzellen sollen daher an dieser Stelle beschrieben werden; die Steuerung und Funktion dieses Systems wurde im Kapitel „Vegetatives Nervensystem“ dargestellt. In der glatten Muskulatur sind die einzelnen Muskelzellen kurz und spindelförmig. Im Gegensatz zur faserförmigen Bündelung in der gestreiften Muskulatur ist die Anordnung der Muskelzellen der glatten Muskulatur netzartig. Bei Aktivierrung ziehen sich die Aktin- und Myosinzellen so zusammen, dass die Muskelzelle insgesamt kontrahiert (und kugelförmiger wird). Abb. III.2 Glatte Muskelzelle bei verschiedenen Kontraktionszuständen. Schematische Darstellung; a relaxiert. b lokale Kontraktion rechts. c im Ganzen kontrahiert. hellgrau eingezeichnet, Myosin (aus: Schmidt & Schaible (Hrsg.) (2006). Neuro- und Sinnesphysiologie (5., neu bearb. Aufl.), S.88, mit Genehmigung von © Springer). Die Muskelzellen der glatten Muskulatur können lange anhaltend kontrahieren, wodurch sie plastisch formbar werden. Verformungen können über längere Zeit aufrechterhalten bleiben und kehren unter Umständen nicht aktiv zu ihrer Ausgangsform zurück. Auch ohne exogene Reizung weisen glatte Muskelzellen einen eigenständigen Kontraktionsrhythmus auf (myogener Rhythmus oder Eigenrhythmik). Bei einer passiven Dehnung des glatten Muskels kann es zu einer reflektorischen Tonuserhöhung kommen. Die glatte Muskulatur ist aber auch über vegetative Nerven aktivierbar (neurogener Tonus). III Motorik 55 III.2 Steuerung und Steuerungsmechanismen Die folgenden Abschnitte beziehen sich auf die Skelett- bzw. gestreifte Muskulatur. Afferenzen Eine wichtige Voraussetzung für die Steuerbarkeit des Muskelsystems sind Afferenzen, also Informationen über den (Dehnungs-)Zustand der Skelettmuskeln und damit über die Haltung des Körpers. Sie werden über zwei Wege vermittelt, die Muskelspindeln und die Golgi-Sehnenorgane. Bei den Muskelspindeln handelt es sich um spezielle Muskelfasern, die zwischen den Muskelfasern der gestreiften Muskulatur liegen. Sie reagieren sensibel auf Dehnungen des Muskels und können über die γ-Motoneurone durch Vordehnung in ihrer Empfindlichkeit gesteuert werden. Die Golgi-Sehnenapparate liegen am Übergang vom Muskel zur Sehne und übermitteln Informationen zum Spannungszustand des Muskels. Afferenzen Afferenzen informieren damit nicht nur über einen Zustand, sondern sie unterstützen auch die Ausführung von Bewegungen in der sogenannten Reafferenz, eines Abgleichs zwischen Bewegungsplanung und tatsächlicher Bewegungsausführung, welche zurückgemeldet (feedback) und mit dem Bewegungsplan (Efferenzkopie) verglichen wird. Differenzen können so erkannt und korrigiert werden oder es kann, falls die Ausführung durch externe Ursachen beeinträchtigt wird, die Planung angepasst werden. Steuerungsmechanismen auf spinaler Ebene Die einfache Art der motorischen Steuerung auf spinaler Ebene durch Reflexe wurde bei der Besprechung des Rückenmarks bereits beschrieben. Dabei wird auf eine bestimmte Reizung hin eine immer gleiche, motorische Antwort gegeben, ohne dass übergeordnete Hirnstrukturen eingebunden werden. Ein Eigenreflex ist ein Reflex, bei dem das Organ der Reizentstehung auch das Organ der reflektorischen Aktivität ist. Bei einem Fremdreflex sind Reiz- und Effektororgan nicht identisch, es sind mehrere Neuronen an der Entstehung beteiligt, die sowohl erregende als auch hemmende Impulse weitergeben können. Reflexe 56 Biologische Grundlagen Abb. III.3: Patellarsehnenreflex. Bei Aktivierung der Muskelspindel eines Muskels, z.B. durch Schlag mit dem Reflexhammer auf die Patellarsehne, aktiviert eine Ia-Afferenz über eine direkte synaptische Verschaltung das α-Motoneuron desselben Muskels, der daraufhin kontrahiert (aus: Schandry, 2006, S. 203). Reflex und Hemmung Zum Beispiel bei Fremdreflexen, aber auch bei bewussten Bewegungen werden durch die reziproke Hemmung antagonistisch arbeitende Muskeln gesteuert. Bei der Erregung eines Muskels, zum Beispiel des Muskels, der die Beugung des Beines vollzieht, wird über eine spinale Verschaltung durch ein Interneuron der antagonistisch arbeitende Muskel, in unserem Beispiel der Strecker, gehemmt. Diese Hemmung ist notwendig, damit die Bewegung vollzogen und nicht durch die Antagonistenaktivität verhindert wird. Ein anderer Mechanismus wird in der Rückwärtshemmung wirksam. Hemmende Interneurone wirken (über Axonkollaterale) auf das aktive Motoneuron selber oder auf andere agonistische Motoneuronen hemmend ein. Damit wird die Aktivität des Muskels reduziert. Diese Interneurone können weiterhin die Aktivität eines Antagonisten bewirken (über die Hemmung hemmender Interneurone). Dieser zunächst paradox erscheinende Mechanismus hat seinen Sinn bei der Aufrechterhaltung bestimmter Körperhaltungen, zum Beispiel dem aufrechten Stehen, wo ein ständiger Abgleich antagonistisch arbeitender Muskeln notwendig ist, um Ausgleichsbewegungen, die sich als Zittern bemerkbar machen würden, und damit einen unnötigen Energieverbrauch zu verhindern. III.3 Koordination durch das Zentrale Nervensystem Bei der Besprechung des Gehirns wurden bereits einige Strukturen besprochen, die für die Steuerung der Motorik zuständig sind. Im Folgenden sollen nun einige Areale genannt werden, denen eine wichtige Rolle bei der Bewegungsplanung und –ausführung zugeschrieben wird. III Motorik 57 Dabei ist zu bedenken, dass an der Bewegungssteuerung eine Vielzahl von kortikalen und subkortikalen Strukturen beteiligt ist. Dazu gehören auch Informationszuflüsse, die Bewegungen (Handlungen) auslösen oder als Reafferenz den Erfolg einer bereits laufenden Bewegung rückmelden. Kortexareale Unter der Bezeichnung Motorkortex werden einige Strukturen zusammengefasst, die maßgeblich an der Steuerung einer Bewegung beteiligt sind: der primäre motorische Kortex, der supplementär-motorische Kortex sowie der prämotorische Kortex. Sie sind miteinander und über Fasern auch mit der jeweils gegenüberliegenden Hemisphäre des Gehirns verknüpft. Sie arbeiten parallel und abgestuft. Bereits vor Ausführung einer Bewegung kann in einer Planungsphase eine Aktivierung der motorischen Kortexareale gemessen werden, das sogenannte Bereitschaftspotential. Auch bei der Vorstellung von Bewegungen, der Imagination, zeigt sich in diesen Kortexarealen eine Aktivität. Diese fand sich aber auch dann, wenn Affen (Makakken) bewegungslos bestimmte Bewegungen anderer Artgenossen beobachten, die denen der eigenen Bewegungsausführung ähnlich sind (siehe auch Kapitel Messmethoden). Diese Neurone wurden etwas reißerisch als „Spiegelneurone“ bezeichnet, welche von einigen Forschenden als neuronale Korrelate des Mitfühlens und des „Sich-in-jemanden-anderen-hineinversetzenKönnens“ (Empathie) angesehen werden. Die Forschungslage ist jedenfalls keineswegs so klar, dass man dies so allgemein sicher behaupten kann, zumal über verschiedene Primatenarten generalisiert wird. Abb. III.4: Motorische Kortexareale. Die in der Abbildung angegebenen Ziffern sind die Areale nach Brodmann (aus: Schandry, 2006, S. 213). Informationen erhalten die motorischen Rindenareale aus dem sensorischen Kortex; thalamokortikale Projektionen liefern außerdem Informationen aus dem Bereitschaftspotential „Spiegelneurone“ 58 Biologische Grundlagen Kleinhirn, den Basalganglien sowie der Körperperipherie. Efferenzen gehen zu den subkortikalen Kernen, dem Rückenmark oder in kortikospinalen Bahnen (Tractus corticospinalis) zu den spinalen Motoneuronen. Der primäre motorische Kortex steuert Bewegungen sehr direkt, da er über die kortikospinalen Verbindungen die Effektororgane rasch erreicht. Insbesondere die feinmotorischen Bewegungen der Finger werden von hier aus gesteuert. Willkürlich initiierte Bewegungssequenzen und deren Erinnerung werden durch den supplementär motorischen Kortex vorbereitet. Auch beidhändige Bewegungen werden hier koordiniert; der supplementär-motorische Kortex ist für beide Körperhälften zuständig (im Gegensatz zu den anderen Strukturen, die sich jeweils auf eine Körperhälfte beziehen.) Weitere Strukturen wie der prämotorische Kortex und der posterior-parietale Kortex (letzterer wird nicht zu motorischen Hirngebieten gezählt) spielen eine Rolle in der Startphase einer Bewegung und bei Bewegungen, die durch sensorische Informationen gesteuert werden. Mit großer Wahrscheinlichkeit wird in diesem Bereich auch das Wissen über Objekte mit Bewegungsplänen integriert. Abb. III.5 Ziel- und Greifbewegung. a Formierung des Griffs bei Annäherung der Hand an das Objekt. b Kortikaler Verarbeitungsweg: Der posterior-parietale Kortex erhält Informationen aus den visuellen und somatosensorischen Kortizes. Er projiziert zum prämotorischen Kortex, der wiederum eng mit dem primär motorischen Kortex verknüpft ist (reprinted from Trends in Neurosciences, Vol.18, No. 7., pp. 314-320, 1995, Jeannerod, M., Arbib, M. A., Rizzolatti, G. and Sakata H.; Grasping objekts: the cortical mechanisms of visuomotor transformation, © 2009, with persmission from Elsevier). III Motorik 59 Weitere, an der Bewegungsausführung beteiligte Hirnareale Die Rolle des Kleinhirns bei der Bewegungssteuerung wurde im Zusammenhang mit der Darstellung des zentralen Nervensystems bereits angesprochen. Es erhält Afferenzen aus dem Rückenmark und kann ausgehende Motorikbefehle mit den Konsequenzen, also dem Erfolg der Bewegung abgleichen. Efferenzen des Kleinhirns gehen zu Kerngebieten (Vestibularkerne, Nucleus ruber, Thalamuskerne) und tragen zur Feinabstimmung von Bewegungen bei, erhöhen die Stabilität beim Gehen und Stehen und modulieren Muskeltonus und Bewegungsabläufe. Eine wichtige Rolle übernimmt das Kleinhirn beim Erlernen reflexhaft ablaufender Reaktionen sowie bei der Zeitabstimmung. Letzteres ist notwendig, um Rhythmen zu erkennen und nachzuvollziehen oder auch, um akustische Reize anhand des Zeitunterschieds beim Auftreffen in den beiden Ohren lokalisieren zu können. Kleinhirn und Feinabstimmung der Bewegung Der Nucleus ruber, der Afferenzen aus dem Kleinhirn empfängt, wirkt über den Tractus rubrospinalis auf die motorischen Neurone des Rückenmarks ein. Seine Aufgabe ist es, über die Aktivierung der Beugemuskeln und die Hemmung der Strecker Zielbewegungen der Extremitäten fein, ohne Zittern (Tremor), abzustimmen. Die Vestibularkerne steuern die aufrechte Körperhaltung. Sie erhalten durch den 8. Hirnnerv (Nervus vestibularis) Informationen aus dem Gleichgewichtsorgan und geben diese in das Rückenmark, an die Augenmuskeln sowie zu Thalamus und Kleinhirn weiter. In ihnen werden Muskelreflexe generiert, mit denen der Körper in einer gewünschten ruhigen Körperhaltung gehalten wird (Stehreflexe), mit denen der Körper aus einer ungewöhnlichen Lage in eine normale Körperstellung gebracht wird (Stellreflexe), und solche, die bei Bewegungen für die Erhaltung des Gleichgewichts in Form von Ausgleichsbewegungen sorgen (statokinetische Reflexe). statokinetische Reflexe 60 Biologische Grundlagen Abb. III.6: Vestibuläre und propriozeptive Komponenten von Stell- und Haltereflexen. Medialer und lateraler Vestibulariskern sind Ursprung von medialem (bilateral) bzw. lateralem (ipsilateral) vestibulospinalem Trakt. Sie projizieren zu zervikalen bzw. lumbalen Inter- und Motoneuronen. Deszendierende (inferiore) und superiore Kerne haben supraspinale Ziele (u.a. Okulomotorik, vestibulookulärer Reflex). Vestibuläre Information läuft zu allen Kernen; Verschaltungen der propriozeptiven Afferenzen aus Halsmuskeln und Wirbelgelenken sind nicht genau bekannt (aus: Klinke, Pape & Silbernagl (2005), Physiologie (5.Aufl.), S.754, Thieme-Verl.). Emotion, „Trieb“ und Bewegung Zu den Basalganglien werden eine Reihe von Kerngebieten gezählt. Sie erhalten unter anderem Afferenzen vom Kortex und geben Informationen über den Thalamus an diesen zurück. Die Funktion dieser, hier sehr vereinfacht beschriebenen Feed-Back-Schleife ist die Abstimmung von einzelnen Befehlen zur Bewegungsausführung mit dem Gesamtsystem. Daher wird hier auch die Verbindung von (motorischem) Verhalten mit emotionalen und motivationalen Kontexten angenommen. Das sind z.B. Bewegungen, die mit Annäherung (im Extrem Aggression, Angriff) oder Vermeidung (im Extrem Flucht) zu tun haben. Hinsichtlich ihrer Plus- und Minus-Charakteristik (hin-weg, stark-schwach, schnell-langsam usw.) sind dies Bewegungsarten, die sich unter den Oberbegriff „Trieb“ subsumieren lassen. III Motorik Abb. III.7: Absteigende motorische Bahnsysteme und Komponenten der Dezerebrierungsstarre. Ursprungsneurone verschiedener Trakte im Kortex bzw. Hirnstamm und ihre Funktionen. Flexorenfördernde, dorsolaterale Verbindungen im Rückenmark links, extensorenfördernde, ventromediale rechts dargestellt (im Organismus bilateral-symmetrisch vorhanden). Interkollikuläre Hirnstammdurchtrennung führt zu Dezerebrierungsstarre: übermäßige Aktivität der spinalen Extensormotoneurone durch Fortfall retikulospinaler und zerebellofugaler Hemmung sowie durch aufsteigende Aktivierung der exzitatorischen pontinen Retikulärformation (aus: Klinke, Pape & Silbernagl (2005), Physiologie (5.Aufl.), S.754, Thieme-Verl.). Im Hirnstamm, insbesondere in der Formatio retikularis, werden Informationen aus Muskeln und Gelenken, dem Gleichgewichtsorgan sowie den Augen integriert. Absteigende Bahnen (= retikulospinale Bahnen) erreichen die Motoneurone der Rumpfmuskulatur. Über diese Struktur wird die Haltung des Körpers vor allem über Ausgleichsbewegungen und Muskelregulationen des gesamten Körpers reguliert. 61 62 Biologische Grundlagen Absteigende Bahnen Bereits an einigen Stellen wurden die Bahnen (Trakte) erwähnt, die dafür zuständig sind, die Informationen aus dem Gehirn in das Rückenmark weiterzuleiten, von wo sie zu den Effektororganen geht. Die Pyramidenbahn (Tractus corticospinalis) zieht paarig von der Hirnrinde zum Rückenmark. Ihre Nervenfasern sind sehr lang und können vom Kortex bis zu den spinalen Motoneuronen reichen, die sie, meist über Interneurone, aktivieren. Während ihres Verlaufs durch die Medulla oblongata bildet die Bahn eine pyramidenförmige Wölbung aus, die ihr den Namen gab. Unterhalb dieser Pyramide kreuzen ungefähr 80% ihrer Fasern auf die jeweils gegenüberliegende Seite und ziehen dann im Seitenstrang des Rückenmarks nach unten. Die ungekreuzten Fasern laufen auf der Vorderseite des Rückenmarks nach unten. Die Pyramidenbahn aktiviert die Muskeln der Unterarme und Hände sowie der Unterschenkel und Füße (distale Extremitäten). Abb. III.8: Projektionen aus Kortex und Hirnstamm a Die kortikalen Efferenzen ziehen als dickes Faserbündel durch die Capsula interna in die subkortikalen Gebiete des Hirnstamms und in das Rückenmark. Dabei nimmt die Zahl der Axone ab. Die aufgeführten Trakte bezeichnen die Projektionsgebiete der kortikalen Efferenzen: 1, kortikostriatal und kortiko-thalamisch; 2, kortikorubral; 3, kortikopontin; 4, kortikoretikulär; 5, kortikoolivär; 6, kortikocuneatus und kortikogracilis; 7, kortikospinalis lateralis; 8, kortikospinalis ventralis. (Mod. nach Phillips & Porter, 1977). b Mediale und laterale Bahnsysteme. Auf der linken Seite ist das laterale Bahnsystem im dorsolateralen Funikulus schematisiert dargestellt. Diese Trakte kreuzen im Hirnstamm. Die Axone treten in den lateralen Teil des intermediären Rückenmarksgraus ein und innervieren die dort liegenden Interneurone sowie die Motornuklei zur Extremitätenmuskulatur (zur Vereinfachung ist die Projektion in das Hinterhorn weggelassen). Auf der rechten Seite ist das mediale Bahnsystem in seinem Verlauf im ventralen Funikulus schematisiert dargestellt. Die Axone treten von medial in das Vorderhorn ein und innervieren die dort gelegenen Interneurone, sowie die Motornuklei zur Stammmuskulatur. Sie kreuzen zum Teil auf die kontralaterale Seite (aus: Schmidt & Schaible (Hrsg.) (2006). Neuro- und Sinnesphysiologie (5., neu bearb. Aufl.), S.104, mit Genehmigung von © Springer). III Motorik Neben der Pyramidenbahn gibt es weitere deszendierende Bahnen aus dem Hirn ins Rückenmark und dort auf Motoneurone. Der rubrospinale Trakt aus dem Nukleus ruber aktiviert distale Flexoren. Der vestibulospinale Trakt ist für die Aufrechthaltung des Gleichgewichts zuständig, gemeinsam mit dem retikulospinalen und tektospinalen Trakt versorgt er die Muskeln des Rumpfs und der proximalen (also nahe am Rumpf gelegenen) Muskeln. Aufgrund ihrer Anordnung im Rückenmark werden bisweilen die im Seitenstrang des Rückenmarks verlaufenden Fasern der Pyramidenbahn sowie der rubrospinale Trakt zum lateralen Bahnsystem und der vestibulospinale, der retikulospinale sowie der tektospinale Trakt zum medialen Bahnsystem zusammengefasst. Zusammenfassung Durch Bewegung wird es dem Menschen möglich, auf seine Umwelt gestaltend einzuwirken und sich in ihr eigenständig zu bewegen. Bewegung wird durch das Zusammenwirken von steuernden Impulsen aus dem zentralen Nervensystem sowie der Ausführung durch spezifische Körperzellen, den Muskelzellen, die in Muskeln angeordnet sind, erzeugt. Die Muskelzellen der Willkürmuskulatur können sich nur in einer Richtung zusammenziehen. Komplexe Bewegungen, die verschiedene Richtungen, Rotationen und unterschiedlichen Kraftaufwand erfordern, entstehen durch das Zusammenwirken mehrerer Muskeln und Muskelgruppen, das durch das zentrale Nervensystem gesteuert wird. Dabei entsteht ein Wirkungsgefüge aus Afferenzen und Efferenzen, Efferenzkopien und Feedback-Schleifen, das darüber hinaus zeitlich geordnet (getaktet) werden muss. Dies ist im weitesten Sinne als (sensumotorisches) Koordinationsproblem zu verstehen. 63 64 Biologische Grundlagen Studientechnischer Anhang 1. Kenntnisse Muskeln, Aufbau und Funktion o glatte Muskulatur, gestreifte Muskulatur: Unterscheidung o Muskelzelle / Muskelfaser, Aktin- und Myosinfilamente o Motorische Einheit Schnittstelle Nervensystem – Muskeln o Efferenzen: Motoneuron und motorische Endplatte o Afferenzen: Muskelspindeln und Golgi-Sehnenapparate Steuerungsmechanismen o Afferenz und Reafferenz o Reflexe: Eigenreflex, Fremdreflex o reziproke Hemmung, Rückwärtshemmung Gehirn-Strukturen der Bewegungssteuerung und deren Funktionen o primärer motorischer Kortex, supplementär-motorischer Kortex, prämotorischer Kortex und deren Funktionen o Kleinhirn, Kerne (Vestibularkerne, Nucleus ruber, Thalamuskerne) Basalganglien, Formatio retikularis und deren Funktionen Bahnen und Rückenmark sowie deren Funktion o Pyramidenbahn, laterales und mediales Bahnsystem: Verlauf und Funktion o Aufbau Rückenmark sowie dessen Funktion III Motorik 2. Literatur Grundlage: Schandry, R. (2006). Biologische Psychologie (2. überarb. Aufl.). Weinheim: Beltz. Kapitel 9: Bewegung Vertiefend: Hanser, H. (2001) (Red.). Lexikon der Neurowissenschaft. (Elektronische Ressource der FernUniversität in Hagen). Brockhaus Duden Neue Medien. Illert, M. & Kuhtz-Buschbeck, J.P. (2006) Motorisches System. In R.F.Schmidt & H.-G. Schaible (Hrsg.), Neuro- und Sinnesphysiologie (S. 94-130). Heidelberg:Springer. Jänig, W (2006) Vegatives Nervensystem. In R.F.Schmidt & H.-G. Schaible (Hrsg.), Neuro- und Sinnesphysiologie (S. 132-181). Heidelberg:Springer. Abschnitt 6.2: Der glatte Muskel: ein Effektor des peripheren vegetativen Nervensystems Abschnitt 6.4: Spinaler, vegetativer Reflexbogen und Harnblasenregulation Rüdel, R. & Brinkmeier, H. (2006) Muskelphysiologie. In R.F.Schmidt & H.-G. Schaible (Hrsg.), Neuro- und Sinnesphysiologie (S. 65-93). Heidelberg:Springer. Sonstige zitierte Literatur: Phillips, C. G. & Porter, R. (1977). Corticospinal neurons. Their role in movement. London: Academic Press. Jeannerod, M., Arbib, M. A., Rizzolatti, G. and Sakata H. (1995). Grasping objekts: the cortical mechanisms of visuomotor transformation. Trends in Neurosciences, 18, 314 – 320. 65 66 Biologische Grundlagen IV Sinnessysteme Lernziele Die Studierenden sollen Sinneszellen adäquater Reiz Einteilung der Sinne die verschiedenen Sinnessysteme des Menschen kennen lernen, die Sinneszellen sowie deren Funktionsweise kennen lernen, die jeweils adäquaten Reize kennen lernen, die Weiterleitung der Informationen von den Sinneszellen in das Gehirn und die dabei verwendeten Verarbeitungsmechanismen kennen lernen, die Umwandlung physikalisch-chemisch definierbarer Reize in (bewusste) Wahrnehmungen verstehen. Unsere Sinne ermöglichen es uns, unsere Umwelt, aber auch unseren Körper wahrzunehmen. Empfindungen der verschiedenen Sinnesmodalitäten wie Sehen, Riechen, Hören, aber auch die Schmerzwahrnehmung und die Wahrnehmung der Haltung unserer Muskeln beruhen auf der Tätigkeit von Sinnesorganen sowie der Aktivierung von Sinneszellen. Primäre Sinneszellen geben erhaltene Informationen direkt weiter, bei den sekundären Sinneszellen handelt es sich um modifizierte Gewebszellen, über die Informationen in Neurone weitergegeben werden müssen. Spezielle Sinne verfügen über ein eigenes Organ, wie das Sehen (Auge), das Hören (Ohr) oder der Geruch (olfaktorische Membran in der Nase). Der jeweils adäquate Reiz führt dann zum entsprechenden Sinneseindruck, also Energie spezifischer Wellenlänge (Licht) führt zum Sehen mit dem Auge (aber nicht zum Hören mit dem Ohr). Andere Sinne müssen sich ein Organ „teilen“, die Haut ist das Organ, durch das Berührung beziehungsweise Druck genauso empfunden wird wie Kälte, Wärme oder Schmerz. Sinne können auch danach unterschieden werden, ob die Reizung, die zu einer Sinnesempfindung führt, fern, nah oder sogar innerhalb des Körpers stattfindet. Hören, Sehen und Riechen sind Fernsinne, ihre Sinneszellen Telezeptoren. Die Hautsinne nehmen Reize wahr, die direkt die Haut betreffen, aber außerhalb des Körpers liegen (Exterozeption). Mit Propriozeption und Interozeption werden die Sinne bezeichnet, die Reize innerhalb des Körpers wahrnehmen. Propriozeption beschreibt die Empfindung der Körperhaltung durch die, bereits im Kapitel „Motorik“ besprochenen Muskelspindeln und Golgi-Sehnenapparate. Zur Interozeption gehört der Geschmackssinn, aber auch die Viszerozeption, also die Ableitung der Zustände der Gefäße (z. B. Dehnung) sowie bestimmter chemischer Zustände (z. B. Blutzuckerkonzentration). IV Sinnessysteme 67 Sinnesmodalität Sinnesorgan Einteilung Sinne Rezeptoren Gesicht Auge Gehör Ohr Geruch olfactorische Membran Teleceptor spezielle Sinne (Chemoreceptor) Geschmack Geschmacksknospen (Chemoreceptor) Winkelbeschleunigung Bogengänge Linearbeschleunigung Utriculus Schmerz Freie Nervenendigungen Interoceptor (Nociceptor) Berührung Meissner-Körperchen etc. Druck Pacini Körperchen Wärme Ruffini Endorgane Kälte Krause-Endkolben Muskeldehnung Golgi-Sehnenorgane Muskeldehnung Muskelspindeln Gelenkslage Nervenendigungen um Gelenke Arterieller Blutdruck Wand von Carotissinus u. Aortenbogen Exteroceptoren Proprioceptoren ‚zentraler’ Venendruck, Wand der großen Venen, Vorhöfe Lungendehnung Vagusendigungen in Lunge Bluttemperatur im Kopf O²-Partialdruck Hypothalamuszellen Carotis- u. Aortenkörperchen Liquor-pH (Chemoreceptor) Osmot. Druck d. Plasmas Receptoren an der ventralen Oberfläche der Med. oblong. Arterio-venöse Blutzuckerdifferenz Receptoren im vorderen Hypothalamus Hautsinne viscerale Sinne Interoceptoren (Visceroceptoren) (Glucostaten) Abb. IV.1 Übersicht über Sinnesmodalitäten und –organe. Aus: Dorsch, F. (Hrsg. v. H. O. Häcker & K.-H. Stapf) (2004), Psychologisches Wörterbuch (14. vollst. überarb. u. erweiterte Aufl.), S. 865, Huber. 68 Biologische Grundlagen Welchen Effekt hat nun die Aktivität der Sinneszellen? Sie kann unbewusst bleiben. Viszerale Afferenzen lösen beispielsweise bestimmte Reaktionen aus, werden aber „nicht bewusst“, das heißt, selbst bei einer intensiven Selbstbeobachtung wird man nichts über den Dehnungszustand seiner Venen aussagen können. Anders ist es zum Beispiel bei der Propriozeption, hier kann man durch Selbstbeobachtung oder spezifische Methoden wie das Biofeedback Aussagen über den Dehnungszustand von Muskeln und die Haltung von Gelenken machen. Empfindung, Wahrnehmung Ein weiterer Aspekt ist die Bedeutung, die einem Reiz zugewiesen wird, die mit Gelerntem oder Erfahrungen verknüpft ist. Wird eine Sinneszelle durch einen Reiz aktiviert, wird diese Aktivierung an höhere Regionen des Gehirns weitergegeben und dort verarbeitet; dabei treten Informationen oder auch Bewertungen wie „wichtig“ oder „nicht-wichtig“ zu dem Reiz. Man unterscheidet daher die Empfindung eines physikalisch oder chemisch definierbaren Reizes von der Wahrnehmung dieses Reizes, die von Verarbeitungs- und Bewertungsmechanismen bestimmt wird. Im Folgenden sollen nun die verschiedenen Sinne mit Ausnahme der Interozeption und Propriozeption besprochen werden (diese beiden „Sinne“ wurden bei der Besprechung des vegetativen Nervensystems beziehungsweise der Motorik beschrieben). Dabei werden jeweils zunächst Sinnesorgane und Sinneszellen, anschließend die Reizverarbeitung und Bewertung beschrieben. IV.1 Sehen Der adäquate Reiz für das Sehen ist Licht beziehungsweise Strahlungsenergie des Wellenlängenbereichs zwischen 380 nm und 760 nm. Dabei wird die Farbe durch die Wellenlänge, die Helligkeit durch die Intensität (Amplitude der Welle) bestimmt. Farbe und Helligkeit sind Interpretationen bestimmter physikalischer Eigenschaften der Wellen. Selbstleuchter sind jedoch eher selten als visuelle Reizquelle wirksam. Vielmehr gelangt meistens dasjenige Licht ins Auge, das nach dem Auftreffen auf Objekte der Umwelt reflektiert wird. Das reflektierte Licht weist eine andere Verteilung der Wellenlängen auf als das auftreffende Licht, von dem ein Teil absorbiert und in Wärme oder chemische Energie umgewandelt wird. Bei nichtfarbigen Objekten ist eine gleichförmige Abnahme der Intensität für alle Wellenlängen charakteristisch, bei Schwarz ist sie erheblich, bei Weiß relativ gering. Alle farbigen Objekte reflektieren eine andere Verteilung der Wellenlängen als die auftreffende. Das Licht, das ein Selbstleuchter abstrahlt, wird Beleuchtungsstärke (Illuminanz) genannt, die Maßeinheit ist lumen/m2 oder lux. Die Intensität des Lichts, das von einer definierten Fläche zurückgeworfen wird, wird als Leuchtdichte (Luminanz) bezeichnet, die Einheit ist candela pro m2 (cd/m2). IV Sinnessysteme 69 IV.1.1 Auge, Stäbchen und Zapfen Das Auge ist aus verschiedenen Bestandteilen zusammengesetzt, die die Aufnahme und Weiterleitung optischer Reize ermöglichen. Es besteht aus einem mechanisch-optischen Apparat, der für die Ausrichtung des Sinnesorgans auf interessierende Objekte zuständig ist, und einer Membran, in der Sehrezeptoren sitzen, die die Lichtmuster aufnehmen und weiterleiten. Das Auge hat eine kugelige Form und ist von verschiedenen Häuten umgeben. Im Inneren des Auges befindet sich der Glaskörper, eine durchsichtige, gallertartige Masse. Die kugelige Gestalt wird durch den Innendruck des Auges erreicht. Am vorderen Teil des Auges befindet sich die Linse. Das Auge wird durch sechs Augenmuskeln, die an den das Auge umgebenden Häuten ansetzen, innerhalb der Augenhöhle bewegt. Die Augenmuskeln werden durch den Nervus oculomotorius, den Nervus trochlearis und den Nervus abducens innerviert und bewegen das Auge in horizontaler, vertikaler oder rollender Richtung. Zweck der verschiedenen Augenbewegungen ist es, interessierende Gegenstände zu fixieren (das heißt, deren Abbild möglichst im Bereich des schärfsten Sehens zu halten, der Sehgrube, s.u.). Bei Sakkaden bewegt sich das Auge bewusst oder unbewusst von einem Fixpunkt zum nächsten. Sakkaden sind beispielsweise beim Lesen gut zu beobachten. Wenn sich ein interessierendes Objekt langsam bewegt, führen die Augen Folgebewegungen aus. Bewegt sich dieses Objekt schneller, kann es zum optokinetischen Nystagmus kommen, bei dem (langsame) Folgebewegungen mit Sakkaden abwechseln. Der optokinetische Nystagmus kann auch bei schneller Bewegung einsetzen, wenn man beispielsweise beim Zugfahren aus dem Fenster hinaussieht. Optokinetische Antworten gleichen Bewegungen des Kopfes aus, dazu werden Informationen aus dem Gleichgewichtsorgan herangezogen. Im Gegensatz zu den bislang genannten Augenbewegungen, die in beiden Augen jeweils gleichsinnig erfolgen, bewegen sich bei Vergenzbewegungen die Augen gegensinnig zueinander. Nähert sich ein Gegenstand den Augen, bewegen sich die Linsen aufeinander zu (Konvergenz) und voneinander weg, wenn er sich entfernt (Divergenz). Auch innerhalb des Auges befinden sich Muskeln. Der ringförmige Ziliarmuskel wölbt durch Kontraktion die Linse stärker und verändert dadurch die Brechkraft der Linse: damit können naheliegende Gegenstände scharf gesehen werden. Der Ziliarmuskel wird durch parasympathische Fasern des 3. Hirnnervs (Nervus oculomotorius) versorgt. Mit Akkommodation wird die Fähigkeit beschrieben, das Auge auf die jeweilige Sehentfernung einzustellen, so dass ein interessierender Gegenstand „scharf“ gesehen wird. Mit dem Alter reduziert sich die Brechkraft der Linse. Kann ein 10jähriger Gegenstände mit einem Abstand von 7 cm vom Sakkaden 70 Biologische Grundlagen Auge noch deutlich sehen, erhöht sich dieser Abstand beim 50jährigen auf 50 cm (Altersweitsichtigkeit). Durch die Pupille fällt Licht in das Auge. Die Pupille wird von der Iris umschlossen, die durch zwei Muskeln bewegt wird. Der sympathisch innervierte Musculus dilatator pupillae erweitert die Pupillenöffnung, der parasympathisch innervierte Musculus sphincter pupillae verengt sie. Damit wird die Intensität des Lichteinfalls in das Auge gesteuert. Abb. IV.2 Schnitt durch das menschliche Auge (aus: Schandry, 2006, S. 243). Photorezeptoren Fovea centralis Stäbchen, Zapfen Diese bislang beschriebenen Strukturen, die den optischen Apparat des Auges ausmachen, steuern die Blickrichtung, die Akkomodation und den Lichteinfall. Damit regulieren sie die physische Reizung, die auf die Sinnesrezeptoren trifft. Die Sinnesrezeptoren des Auges, Zapfen und Stäbchen, sitzen im hinteren Teil der Retina, der inneren Haut des Auges. Bei diesen Photorezeptoren handelt es sich um primäre Sinneszellen. Zapfen und Stäbchen sind nicht gleichmäßig über die Retina verteilt: In der Sehgrube (Fovea centralis), der Stelle des schärfsten Sehens, befinden sich ausschließlich Zapfen. An der Stelle, an der der Sehnerv das Auge verlässt, fehlen naturgemäß die Rezeptoren. Dies ist die Stelle, die den blinden Fleck erzeugt. Die Stäbchen sind für das Sehen in der Dämmerung (skotopisches Sehen), die Zapfen sind für das Sehen am Tage (photopisches Sehen) sowie das Farbensehen zuständig. Es gibt drei Typen von Zapfen, die unterschiedliche Empfindlichkeitsmaxima für verschiedene Wellenlängen haben, Blau-Zapfen bei 440 nm, Grünzapfen bei 535 nm und Rotzapfen bei 565 nm. Über Bipolarzellen geben die Photorezeptoren ihre Informationen an die Ganglienzellen weiter, deren Axon das erste Neuron der zentralen Sehbahn bildet. Horizontalzellen und amakrine Zellen verbinden IV Sinnessysteme Photorezeptoren beziehungsweise Bipolarzellen untereinander und mit den Ganglienzellen. Da die verbindenden beziehungsweise weiterleitenden Neurone über den in die Haut eingebetteten Photorezeptoren liegen (also dem Glaskörper näher), spricht man von der Inversion der Retina. Abb. IV.3: Elemente der Netzhaut und deren Verschaltungen. Es ist der mehrschichtige Aufbau der Retina erkennbar. Die Zelltypen, die bei der Verarbeitung von Netzhautbildern beteiligt sind, sind eingezeichnet. Sie werden im Text besprochen. (IB = invaginierende Bipolarzellen, FB = flache Bipolarzellen, SB = Stäbchen-Bipolarzellen) (aus: Schandry, 2006, S. 246). Bei Lichteinfall werden die in den Photorezeptoren eingelagerten Photopigmente umgewandelt und es erfolgt (über mehrere Zwischenschritte) eine Hyperpolarisation. (Im Gegensatz zu bisher beschriebenen Prozessen entspricht die Aktivierung eines Photorezeptors einer Hyperpolarisierung, die erst bei der Weitergabe an das nächste Neuron die für ein Aktionspotential typischen Depolarisationen auslöst. Stäbchen und die für die Wahrnehmung der verschiedenen Farben zuständigen Zapfen unterscheiden sich in der Art ihres Pigments. 71 72 Biologische Grundlagen IV.1.2 Reizweiterleitung und Reizverarbeitung Wie bereits erwähnt, sind die Photorezeptoren, Bipolarzellen und die weiterleitenden Ganglienzellen der Retina miteinander in vielfältiger Weise verbunden. Durch dieses retinale Neuronennetzwerk werden einfache Verarbeitungen möglich – so werden durch amakrine Zellen die Stäbchen während des photopischen Sehens gehemmt. rezeptive Feld Jede Ganglienzelle fasst die Informationen aus verschiedenen Sinneszellen zusammen, die über ein bestimmtes Netzhautareal, das rezeptive Feld, verteilt sind. Die rezeptiven Felder des Auges überlappen sich. Ihre Ausdehnung auf der Netzhaut ist in der Sehgrube am geringsten, nach außen nimmt sie zu. Anhand von Größe und Funktion lassen sich drei Grundtypen von Ganglienzellen unterscheiden: M-Zellen (Zellen vom magnozellulären Typ) verfügen über einen großen Zellkörper und große rezeptive Felder; sie sind kontrast- und bewegungsempfindlich. Die wesentlich häufiger vorkommenden, etwa 80% der Zellpopulation ausmachenden P-Zellen (Zellen vom parvozellulären Typ) sind für die räumliche Auflösung, die Formwahrnehmung und die Farbwahrnehmung zuständig. Die Zellen vom blauempfindlichen koniozellulären Typ leiten Informationen weiter zu höheren Hirnregionen, die unter anderem visuelle Reflexe auslösen. Die rezeptiven Felder am Auge sind in ein Zentrum und ein Umfeld aufgeteilt. Es gibt Ganglienzellen, bei denen ein Licht-Punkt im Zentrum des rezeptiven Feldes zu einer Erregung führt (Ganglienzelle mit einem On-Zentrum). Fällt der Lichtpunkt in das Umfeld, also den äußeren Teil des rezeptiven Feldes, wird diese Ganglienzelle gehemmt. Eine Ganglienzelle mit einem Off-Zentrum wird hingegen durch einen Lichtpunkt in das Umfeld des rezeptiven Feldes aktiviert. Werden sowohl Umfeld als auch Zentrum des rezeptiven Feldes einer Ganglienzelle beleuchtet, erfolgt eine schwächere Aktivierung. Diese Mechanismen werden unter anderem durch die Horizontalzellen vermittelt. IV Sinnessysteme 73 Abb. IV.4: On- und Off-Zentrum-Antworten im retinalen Netzwerk und Verschaltungen des antagonistischen Umfelds. Links: Oben ist das rezeptive Feld mit Zentrum und Umfeld auf der Netzhaut dargestellt. Der Photorezeptor hyperpolarisiert bei Licht im Zentrum. Die invaginierende, hemmende Synapse zur On-Bipolarzelle invertiert das Signal. Rechts: Licht im Umfeld wirkt auf den Photorezeptor im Zentrum über die laterale Horizontalzellverschaltung hemmend; das Signal wird invertiert und führt zu einer Depolarisation am Photorezeptor. Diese wirkt im On-Kanal hemmend, im Off-Kanal erregend (aus: Schandry, 2006, S. 251). Diese komplexen Mechanismen ermöglichen es, Kontraste wahrzunehmen und sind gemeinsam mit der Dichte der Photozeptoren die Grundlage für die Sehschärfe, das räumliche Auflösungsvermögen des Auges. Bestimmt wird die Sehschärfe durch den Visus, den Kehrbruch des minimalen Winkels, in dem das Auge zwei Lichtpunkte voneinander diskriminieren kann und nicht als einen Punkt wahrnimmt. So können beispielsweise zwei Lichtpunkte, die sich 10 m von unserem Auge entfernt befinden, dann noch diskriminiert werden, wenn sie einen Abstand von nur 2 mm haben. Die größte Sehschärfe wird in der Fovea erzielt und die Sehschärfe nimmt rapide ab, wenn ein Reiz außerhalb der Fovea dargeboten wird. Des Weiteren weist das skotopische System (Stäbchen) eine höhere Leuchtdichteempfindlichkeit auf als das photopische (Zapfen). Für die Beziehung zwischen Reizeigenschaften und Sehschärfe erweisen sich folgende Reizeigenschaften als wichtig: Reizwellenlänge, Reizintensität (Leuchtdichte), Reizdauer, Reizgröße und Reizkontrast. Der Kontrast ist das Verhältnis der Leuchtdichtedifferenz zwischen Hintergrund und Objekt, ΔL = Lh – Lo, zur Leuchtdichte des helleren Reizes L, also ΔL/L. Er ist sehr hoch beim Verhältnis von Schwarz und Weiß. Mit Reizgröße ist nicht die absolute Größe des Reizes gemeint, sondern die Sehschärfe Kontrast 74 Sehwinkel Biologische Grundlagen Größe des Reizes auf der Netzhaut. Diese Größe wird leicht missverständlich als Sehwinkel bezeichnet, denn es ist eben nicht der Winkel gemeint, der Auge und Objekt einschließt, sondern die Größe der Reizprojektion auf der Netzhaut. Der Sehwinkel wird in Grad oder Winkelminuten (meist als Bogenminuten bezeichnet) (1°=60 Winkelminuten) oder Winkelsekunden (1 Winkelminute = 60 Winkelsekunden) ausgedrückt und wie folgt berechnet, wenn G = Größe des Reizobjektes, D = Distanz zwischen Auge und Reizobjekt bedeutet, dann gilt: Sehwinkel α° = (G/(2π D)) * 360°. Farbensehen Auch das Farbensehen wird durch retinale Vernetzung ermöglicht. Durch die drei Zapfentypen werden drei Farbbereiche repräsentiert. Dazu, wie durch diese drei Farben die integrierte Farbwahrnehmung des Menschen entsteht, wurden zwei Theorien entwickelt. Die trichromatische Farbtheorie geht davon aus, dass sich durch die Mischung von drei Grundfarben je nach deren Intensität alle möglichen Farben herstellen lassen. So kann man sich vorstellen, dass ein Farbeindruck durch die unterschiedliche Aktivität der beteiligten Zapfen entsteht. Die Gegenfarbtheorie geht von den Farb-Gegensatzpaaren Schwarz-Weiß, Rot-Grün und Gelb-Blau aus. Die Farben eines Paars werden im Zentrum beziehungsweise im Umfeld des rezeptiven Feldes einer On- oder Off-Ganglienzelle abgebildet. Durch die Verrechnung der Informationen aus den verschiedenen Varianten der farbempfindlichen Ganglienzellen kommt die Farbempfindung zustande. In der KriesZonentheorie werden beide Farbtheorien vereint, indem in den zentralen Bereichen des Sehens (an der Sehgrube) eher gegenfarblich organisiertes, in der Peripherie eher trichromatisches Farbensehen angenommen wird. Sehbahn Nachdem bereits auf der Ebene der Retina einfache Verarbeitungsschritte visueller Reize stattgefunden haben, werden visuelle Reize in die höheren Regionen des Gehirns geleitet und dort weiterverarbeitet. Wie oben beschrieben bilden die Axone der Ganglienzellen der Retina die Axone des Sehnerven (Nervus opticus), der an der den blinden Fleck erzeugenden Stelle das Auge verlässt. Im Chiasma opticum kreuzen jeweils die Hälfte der Nervenfasern auf die andere Seite, und zwar jeweils die Fasern, die von der der Nase zugewandten Hälfte der Retina stammen. Danach ziehen die Fasern als Tractus opticus weiter. Ein Teil der Fasern zieht in den Nucleus suprachiasmaticus des Hypothalamus und stellt eine Verbindung zur Epiphyse her. Ein von den M-Zellen stammender Teil der Ganglienzellaxone zweigt zu den Colliculi superiores im Mittelhirn ab. Sie reagieren besonders auf Bewegungen, IV Sinnessysteme 75 stimmen die visuellen mit somatosensitiven und akustischen Signale ab und haben Einfluss auf die Blickmotorik. So werden hier Reflexe ausgelöst, mit dem die Augen durch Sakkaden auf neuartige Reize fokussiert werden. Der größte Teil der Fasern des Tractus opticus (circa zwei Drittel) zieht in das Corpus geniculatum laterale im Thalamus, das die synaptische Schaltstelle zwischen Retina und visuellem Kortex darstellt. Die Projektion von der Retina in das Corpus geniculatum laterale (sowie auch weiter in den visuellen Kortex) erfolgt retinotop, das heißt nebeneinander liegende Retinaareale werden auch in nebeneinander liegende Areale projiziert, wobei der Bereich um die Sehgrube disproportional groß repräsentiert wird. Auch die Aufgabenteilung zwischen magnozellulärem und parvozellulärem System bleibt erhalten. Die Aufgabe des Corpus geniculatum laterale kann vereinfachend als die einer Verstärkung der komplexen retinalen Reize verstanden werden. Darüber hinaus erreichen das Corpus geniculatum laterale Projektionen aus anderen Hirnarealen, zum Beispiel aus dem Hirnstamm zur Anpassung der visuellen Signalübertragung oder aus dem visuellen Kortex zur selektiven Beeinflussung der Übertragung aus bestimmten Hirnregionen. In der Sehstrahlung (Radiatio optica) werden die Informationen vom Corpus geniculatum laterale zum visuellen Kortex gebracht. Abb. IV.5: Verlauf der Sehbahn. Für das rechte Auge ist das Gesichtsfeld, unterteilt in eine nasale und eine temporale Hälfte, eingezeichnet. Die efferenten Verbindungen zwischen Sehrinde und subkortikalen Gebieten sind hier nur auf der rechten Seite eingezeichnet (aus: Schandry, 2006, S. 259). retinotop 76 Biologische Grundlagen Visueller Kortex Im Okzipitallappen der Hirnrinde befindet sich der primäre visuelle Kortex (Sehrinde). Auch dieser Teil des visuellen Systems ist retinotop aufgebaut. Die rezeptiven Felder der visuellen Kortexneurone reagieren allerdings auf komplexe Muster oder bestimmte Anordnungen, nicht auf das Vorhandensein oder das Nicht-Vorhandensein von Lichtreizen wie die rezeptiven Felder der Retina. Dies wird auch dadurch möglich, dass der primäre visuelle Kortex Afferenzen aus anderen Hirnarealen als dem Corpus geniculatum laterale erhält. orientierungsspezifische Kortexneurone kortikaler Modul Was-System, Wo-System Eine solche optische Struktur ist beispielsweise die Orientierung eines visuellen Reizes im Raum, auf die orientierungsspezifische Kortexneuronen reagieren. Bewegungs- oder richtungsspezifische Neuronen werden bei Bewegungen jeweils spezifischer Richtung aktiv. Und längenspezifische Neuronen erlauben die Identifikation von Ecken und Konturen, da sie gehemmt werden, wenn ein Objekt eine bestimmte Länge überschreitet. Der primäre visuelle Kortex (V1) besteht (wie die anderen Kortexareale) aus sechs übereinanderliegenden Schichten, wobei übereinanderliegende Neuronen jeweils den gleichen Bereich der Netzhaut repräsentieren und als Säule bezeichnet werden. Die Zellen in okularen Dominanzsäulen werden vorzugsweise durch ein Auge (rechtes oder linkes) aktiviert. Innerhalb der okularen Dominanzsäulen findet man Orientierungssäulen, die orientierungsspezifische Neuronen enthalten. Zwischen je zwei Dominanzsäulen sind ebenfalls säulenförmige farbempfindliche Blobs. Die beiden okularen Dominanzsäulen jeweils des rechten und linken Auges werden zu Hypersäulen zusammengefasst. In etwas idealisierender Weise kann man okulare Dominanzsäulen, Orientierungssäulen und Blobs zu einem kortikalen Modul zusammenfassen. Eine 2 Kubikmillimeter großer „Würfel“ aus V1 sei notwendig und hinreichend, um das Abbild eines punktförmigen Objektes im Raum zu verarbeiten, so Hubel und Wiesel (nach Bear, Conners und Paradiso, 2007, S. 332). Notwendig deswegen, weil der Ausfall eines kortikalen Moduls zu einem blinden Fleck im Gesichtsfeld führen würde und hinreichend, weil er alle neuronalen Korrelate zur visuellen Analyse enthalte. Im primären visuellen Kortex erfolgt eine Parallelverarbeitung von Form, Farbe, Tiefe und Bewegung. Dann trennen sich die Wege. Im inferotemporalen „WasSystem“ werden Farbe, Form und Details, im parietalen „Wo-System“ Ort, Bewegung und Tiefe analysiert. Dabei werden neben den visuellen Kortexarealen auch andere beteiligt, was sich in der Namensgebung der beiden System abbildet. IV Sinnessysteme 77 Abb. IV.6: Zusammenfassung der verschiedenen Leistungen der mit visueller Informationsverarbeitung befassten Strukturen. Die Verarbeitung im magnozellulären System und parvozellulären System ist getrennt dargestellt (aus: Schandry, 2006, S. 265). Verarbeitungsmechanismen im visuellen System Auf der Ebene der kortikalen Bearbeitung visueller Reize können weitere Fähigkeiten bzw. Eigenschaften des visuellen Systems verortet werden. Hierbei werden neben den unmittelbaren Empfindungen auch Erfahrungen herangezogen, um visuelle Empfindungen wahrzunehmen. Mit Farbkonstanz wird die Tatsache beschrieben, dass die Farbe eines bestimmten Objekts immer identisch wahrgenommen wird, unabhängig von der Beleuch- Farbkonstanz 78 Biologische Grundlagen tung und daraus resultierend der Energie und Wellenlänge, die das Objekt abstrahlt. räumliches Sehen Am räumlichen Sehen kann man sehr gut demonstrieren, wie Informationen unterschiedlichster Ebene und Herkunft zusammenwirken, um eine Empfindung, eine Wahrnehmung zu erzeugen. Zum räumlichen Sehen werden vor allem die Informationen aus beiden Augen miteinander verglichen. Befindet sich ein Gegenstand nahe am Auge, liefern die beiden korrespondierenden Photorezeptoren unterschiedliche Informationen. Mit korrespondierend werden jeweils die Rezeptoren bezeichnet, die, denkt man sich die Netzhäute übereinander gelegt, an der gleichen Stelle sitzen. Die Abweichung wird quasi verrechnet und einerseits ausgeglichen (man sieht keine Doppelbilder), andererseits für die Abschätzung des Abstands vom Auge verwendet. Abb. IV.7: Korrespondierende Netzhautbereiche. Korrespondierende Rezeptoren werden aktiviert, wenn ein unendlich weit entfernter Lichtpunkt (= weiter als 6 m entfernt) auf die Netzhäute beider Augen fällt. Bei einem nahe gelegenen punktförmigen Objekt werden auf den beiden Netzhäuten Punkte getroffen, die deutlich voneinander abweichen (aus: Schandry, 2006, S. 268). Bei der Analyse der Querdisparation wird die Projektion eines nicht fixierten Gegenstands auf die Netzhaut verwendet. Liegt der Gegenstand hinter dem Fixpunkt, wird dieser stärker nach innen (Richtung Nase) auf die Netzhaut projiziert. IV Sinnessysteme Abb. IV.8: Querdisparation. Wird ein Punkt fixiert, so erschließt sich die relative Tiefe anderer Punkte im Raum aus der Querdisparation (aus: Schandry, 2006, S. 269). Doch auch beim Sehen mit einem Auge werden Informationen zur Tiefe beziehungsweise zu Entfernungen vermittelt. Verdeckt ein Objekt ganz oder teilweise ein zweites, so wird das erste Objekt als näher liegend wahrgenommen (Verdeckung, Interposition). Größe (eines uns in seiner Größenausdehung bekannten Objektes) und Perspektive (z.B. paralle Linien werden als zusammenlaufend gesehen) werden aufgrund unserer Erfahrung als Hilfsmittel zur räumlichen Wahrnehmung verwendet. Auch der Farbton sowie die Verteilung von Licht und Schatten (Texturen) können als Hinweis auf die Entfernung genutzt werden. Und schließlich können wir durch eigene Bewegung auf die Entfernung schließen, denn je weiter ein Objekt von uns entfernt ist, desto langsamer wird es sich auf unserer Netzhaut verschieben (Bewegungsparallaxe). Dies wird noch deutlicher bei passiver Bewegung, wenn man beispielsweise aus dem Fenster eines fahrenden Zuges sieht. 79 80 Biologische Grundlagen IV.2 Hören Der adäquate Reiz für das Hören ist Schall. Entstandene Schallwellen können anhand ihrer Amplitude (der Differenz zwischen maximalem und minimalem Druck, die dem Schalldruck entspricht) und ihrer Frequenz (als Zahl der Schwingungen pro Sekunde) beschrieben werden. Menschen können Frequenzen zwischen 20 und 16.000 Hertz hören. Ein Schall mit nur einer Frequenz wird als Ton bezeichnet. Gemeinsam erklingende Töne, die jeweils ein ganzzahliges Vielfaches einer Grundfrequenz abbilden sind Klänge. Meist hören wir Geräusche, die aus verschieden frequenten Tönen (und Klängen) zusammengesetzt sind. Als weißes Rauschen bezeichnet man ein Geräusch, bei dem viele Töne und Klänge mit jeweils gleichem Anteil gleichzeitig vorkommen. Beim Schalldruckpegel wird ein gegebener Schalldruck logarithmiert und auf einen festgelegten Bezugsschalldruck bezogen; diese Größe wird in dezibel (db) angegeben. Sie wurde entwickelt, um den menschlichen Hörbereich in handhabbaren Zahlen abbilden zu können. So entspricht das Rascheln von Blättern einem Schalldruckpegel von 20 db, ein Gespräch einem Schalldruckpegel von 60 db und das Starten eines Propellerflugzeugs einem von 120 db. Ab ungefähr 140 db beginnen Geräusche zu schmerzen. IV.2.1 Ohr, Corti-Organ und Haarzellen Das Ohr besteht aus Außen-, Mittel- und Innenohr. Das Außenohr bilden die Ohrmuschel, in der Schallwellen gebündelt werden, und der äußere Gehörgang bis zum Trommelfell. Das sich daran anschließende Mittelohr ist ein luftgefüllter Raum, in dem sich die drei Gehörknöchelchen Hammer, Amboss und Steigbügel befinden. Der Hammer setzt auf dem Trommelfell auf und ist gelenkartig mit dem Amboss verbunden, der seinerseits gelenkartig mit dem Steigbügel verbunden ist. Der Steigbügel setzt auf dem ovalen Fenster auf, das das Mittelohr zum Innenohr hin abschließt. Die Gehörknöchelchen haben die Aufgabe, auf das Trommelfell auftreffende Schallwellen zu verstärken und in das Innenohr weiterzuleiten. Das Innenohr besteht aus dem Gleichgewichtsorgan (siehe unten) und dem Hörorgan, der Kochlea (Schnecke); auf Grund der Form dieser Organe wird der gesamte Aufbau als Labyrinth bezeichnet. IV Sinnessysteme Abb. IV.9: Aufbau des menschlichen Ohres. In der eingesetzten Abbildung ist die Lage des äußeren Ohrs, Mittelohrs und Innenohrs im Kopf dargestellt (aus: Schandry, 2006, S. 273). Die Kochlea kann man sich als einen eingerollten Schlauch vorstellen, in dem eine Trennwand verläuft, die den mit Flüssigkeit (Perilymphe) gefüllten Innenraum in zwei Gänge, die Scala vestibuli und die Scala tympani, trennt. Am äußersten Ende der Kochlea, dem Helikotrema, sind diese beiden verbunden, am anderen Ende werden die Gänge durch das ovale Fenster (Scala vestibuli) beziehungsweise das runde Fenster (Scala tympani) begrenzt. In der kochleären Trennwand findet man einen weiteren, jedoch kleineren flüssigkeitsgefüllten Hohlraum, die Scala media. Auf der Basilarmembram, die diesen Hohlraum von der Scala tympani trennt, befindet sich das Corti-Organ, das durch die Tektorialmembram abgedeckt wird. 81 82 Biologische Grundlagen Abb. IV.10: Schematische Darstellung des Ohres. a Längsschnitt durch äußeren Gehörgang, Mittelohr und Cochlea. Die zweieinhalb Windungen der Cochlea sind in diesem Schema „entrollt“. Auslenkung der kochleären Trennwand nach unten aufgrund einer Einwärtsschwingung des Steigbügels. Gleichzeitig findet zum Ausgleich eine Auswärtsbewegung des runden Fensters statt. b Querschnitt durch die Cochlea. Die Scala media mit positivem endokochleärem Potential und zusätzlich auffällig hoher Kaliumkonzentration in der Endolymphe ist Teil der kochleären Trennwand, die unten von der Basilarmembran und oben von der Reissner-Membran begrenzt wird. Das apikale Ende der Haarzellen ragt in die Scala media hinein (aus: Schmidt & Schaible (Hrsg.) (2006). Neuro- und Sinnesphysiologie (5., neu bearb. Aufl.), S.292, mit Genehmigung von © Springer). Corti-Organ Das reizaufnehmende Organ des Ohrs, das Corti-Organ besteht aus Haarzellen, an deren Spitze sich feine Härchen, die Stereozilien, befinden, wobei die längeren Stereozilien an der Tektorialmembram heften. Die Haarzellen sind sekundäre Sinneszellen, die Synapsen mit dem Hörnerv bilden. Trifft nun ein Schall beziehungsweise der mit Schall verbundene Druck auf das Ohr, wird er über Trommelfell und die Gehörknöchelchen des Mittelohrs auf das ovale Fenster und damit auf Perilymphe weitergeleitet. Die dadurch entstehende Bewegung der Perilymphe führt zu einer Auslenkung der kochleären Trennwand und zu einer Verschiebung der Tektorialmembram gegenüber der Basilarmembram. Die Stereozilien werden ebenfalls ausgelenkt und damit das elektrische Potential der Haarzelle verändert. Über die synaptische Verschaltung mit Fasern des Hörnervs werden dort gegebenenfalls Aktionspotentiale ausgelöst. IV Sinnessysteme 83 Abb. IV.11: Deflektion der Stereozilien. Abscherung der Stereozilien äußerer Haarzellen durch die Abwärts- und Aufwärtsbewegungen der kochleären Trennwand (aus: Schmidt & Schaible (Hrsg.) (2006). Neuro- und Sinnesphysiologie (5., neu bearb. Aufl.), S.295, mit Genehmigung von © Springer). Aufgrund ihrer Schwingungseigenschaften (Resonanz) gerät die Basilarmembram je nach Frequenz des eingehenden Schalls unterschiedlich in Schwingung. Bei Tönen höherer Frequenz schwingen die mittelohrnahen Bereiche, bei tiefen Frequenzen die Bereiche am Helikotrema. Tonhöhen werden daher über den Ort, an dem sie eine Aktivierung auslösten, kodiert (Ortsprinzip). Abb. IV.12: Frequenzkodierung längs der Basilarmembran. Für jede Frequenz gibt es längs der Membran einen optimalen Bereich, innerhalb dessen sie besonders leicht in Schwingungen versetzt werden kann. Hier findet Resonanz statt, die Basilarmembran schwingt hier mit maximaler Amplitude. Die Position dieses Ortes der maximalen Schwingung dient zur Umkodierung von Tonhöhen in neuronale Entladungsfrequenzen (Frequenzortsabbildung) (aus: Schandry, 2006, S. 279). Ortsprinzip 84 Biologische Grundlagen IV.2.2 Reizweiterleitung und Reizverarbeitung Bereits auf der Ebene der Fasern des Hörnervs (Nervus cochlearis), die in der Basilarmembran an die Haarzellen anschließen, werden erste Informationen „verarbeitet“. Das Ortsprinzip der Frequenzkodierung wurde bereits besprochen. Die Dauer eines Hörreizes wird über die Aktivierungsdauer, die Intensität (also der Schalldruckpegel) wird über die Entladungsfrequenz der jeweiligen Faser kodiert. Die Fasern des Hörnervs ziehen von der Kochlea in die Nuclei cochleares der Medulla oblongata, wo eine erste synaptische Umschaltung stattfindet und ein Großteil der Fasern auf die gegenüberliegende Seite kreuzt. Akustische Signale ziehen dann im Lemniscus lateralis weiter in die Colliculi inferiores und das Corpus geniculatum mediale des Thalamus. Schließlich ziehen sie in der Hörstrahlung vom Thalamus in die primäre Hörrinde des Temporallappens. Von dort werden die Informationen in sekundäre auditorische und Assoziationsareale weitergeleitet sowie in die Areale, die für Spracherkennung (Wernicke-Zentrum) und Sprachproduktion (Broca-Zentrum) zuständig sind. Im Verlauf dieser hier natürlich vereinfacht dargestellten Hörbahn werden bereits auf der Ebene der Olivenkerne, im Verlauf des Lemniscus laterale und den Colliculi inferiores Informationen aus beiden Ohren verarbeitet, zum Beispiel durch rückkreuzende Fasern. Des Weiteren sind Kollaterale zum Kleinhirn und zur Formatio retikularis ausgebildet; vor allem letzteres scheint wichtig, um akustische Signale und Aktivierung zu verknüpfen. Abb. IV.13: Schematische Darstellung der zentralen Hörbahn (aus: Schmidt & Schaible (Hrsg.) (2006). Neuro- und Sinnesphysiologie (5., neu bearb. Aufl.), S.304, mit Genehmigung von © Springer). IV Sinnessysteme 85 Im Verlauf der Hörbahn erfolgen weitere Verarbeitungen der Reize. Schon auf der Ebene der Nuclei cochleares werden Informationen wie Reizbeginn und Reizende sowie Frequenzänderungen identifiziert. Zeitliche Merkmale werden weitergehend in den Colliculi inferiores analysiert. Sowohl in den Nuclei cochleares als auch in den Colliculi inferiores werden die Mechanismen verortet, die die Voraussetzung für eine zweite Art der Tonhöhenunterscheidung, die Periodizitätsanalyse, bilden. Hierbei wird die Tatsache genutzt, dass die Ausbreitung einer Welle über die Basiliarmembram dauert und in Abhängigkeit von der Tonhöhe zu bestimmten zeitlichen Mustern bei der Erregung der Fasern des Hörnervs führt. Wenn diese miteinander „verrechnet“ werden, kann dadurch die Tonhöhe bestimmt werden. Periodizitätsanalyse Abb. IV.14: Periodizitätsanalyse. Die verschiedenen Fasern (a-e) werden von derselben wandernden Welle erregt, und zwar aufgrund der räumlichen Anordnung der Sinneszellen in der Kochlea nacheinander. Die Sinneszellen sind im dargestellten Fall in ihrer Empfindlichkeit so eingestellt, dass sie nur am Maximum der Schwingung zu einem Aktionspotential in der nachgeschalteten Faser führen. Das heißt mit dem Fortschreiten der Welle können immer neue Fasern rekrutiert werden. Aus dem zeitlichen Abstand der Entladungen dieser Faserpopulation (kollektive Antwort) kann im Gehirn auf die Periodendauer und damit auf die Frequenz zurückgerechnet werden. Es ist ein Beispiel gezeigt, in dem Schallwelle B die halbe Frequenz von Schallwelle A hat. Man erkennt, dass sich in der kollektiven Antwort genau diese Verdopplung der Periode von A nach B widerspiegelt (aus: Schandry, 2006, S. 282). Auch die räumliche Ortung von Schallquellen stellt einen Verarbeitungsmechanismus dar. Hierzu werden verschiedene Mechanismen verwendet. Zunächst werden Unterschiede in den Empfindungen beider Ohren analysiert. Liegt eine Schallquelle nicht genau vor oder hinter dem Hörenden, trifft der Schall in zeitli- räumliche Ortung 86 Biologische Grundlagen chem Abstand und mit unterschiedlicher Intensität an den Ohren ein. Die Analyse der Laufzeitunterschiede und Intensitätsunterschiede erlauben die örtliche Zuweisung. Die Faltung der Ohrmuschel, mit der Geräusche auf das Trommelfell reflektiert werden, führt zu Verzerrungen in Abhängigkeit davon, ob sich die Schallquelle vor uns, hinter uns oder über uns befindet. Zusammenfassend lässt sich festhalten, dass das Prinzip der Reizweiterleitung und Reizerkennung im auditiven System darin besteht, wichtige Informationen aus Schalldruckmustern zu identifizieren und weiter zu leiten sowie unwichtige Informationen auszufiltern. Wichtige Informationen werden anhand von Merkmalsmustern des Schalls identifiziert, nicht anhand einzelner Merkmale wie Dauer, Frequenz und Frequenzänderung und Intensität. Auf der kortikalen Ebene findet sich dabei die ausgeprägteste Form der Schallmuster-Erkennung, die zur (bewussten) Wahrnehmung und Interpretation von auditiven Reizen und gegebenenfalls zu Reaktionen führt. Dabei sind insbesondere die Spracherkennung und das Verstehen der gesprochenen Sprache von Bedeutung für den Menschen. IV.3 Riechen Der für das Riechen adäquate Reiz sind in der Luft enthaltene Moleküle eines Geruchsstoffs (Geruchsmoleküle). Diese müssen gaslöslich sein und mit dem Sekret der Nasenschleimhaut in Lösung gehen. Allerdings genügen sehr geringe Konzentrationen, um eine Geruchsempfindung auszulösen. Der Mensch kann ca. 10.000 Gerüche voneinander unterscheiden, wobei es sich bei den einzelnen Gerüchen jeweils um Mischgerüche handelt, die aus verschiedenartigen Geruchsmolekülen zusammengesetzt sind. IV.3.1 Nase, Riechepithel und Riechzellen Im hinteren oberen Teil der Nasenhöhle befindet sich das Riechepithel, die für das Riechen spezialisierte Zellschicht, das eigentliche Riechorgan. In der Luft gelöste Geruchsmoleküle gelangen mit der Atemluft in die Nase. Beim normalen Atmen strömt nur wenig Luft an das Riechepithel, erst wenn durch Schnüffeln der Atemstrom umgelenkt wird, gelangen Geruchsmoleküle in höherer Zahl an das Riechepithel. IV Sinnessysteme Abb. IV.15: a. Längsschnitt durch die menschliche Nase. Die Luftströmung bei der Einatmung ist durch Pfeile gekennzeichnet. Man erkennt das Riechepithel und den Bulbus olfactorius. Von hier aus führt der Tractus olfactorius zu den primären olfaktorischen Zentren. b. Riechepithel. Die bipolaren Riechzellen sind von Stützzellen umgeben. Die Bowman-Drüsen bilden das Sekret, das die Riechschleimhaut bedeckt. c. Eine einzelne Riechzelle (aus: Schandry, 2006, S. 293). Das Riechepithel besteht aus verschiedenen Zelltypen und wird durch ein Sekret aus den Bowman-Drüsen überzogen. Man unterscheidet Stützzellen, Basalzellen und Riechsinneszellen. Die Riech(sinnes)zellen sind die olfaktorischen Sensoren, es handelt sich um primäre Sinneszellen. Sie haben eine nur kurze Lebensdauer von einem Monat und werden durch Teilung der Basalzellen nachgebildet. Die Stützzellen haben, wie ihr Name bereits sagt, stützende Funktion für die Riechzellen. 87 88 Biologische Grundlagen Abb. IV.16: Aufbau der Riechschleimhaut mit den zentralen Verbindungen. Die Riechschleimhaut setzt sich aus den Riechsinneszellen, Stützzellen, Basalzellen und Drüsenzellen zusammen. Am apikalen dendritischen Ende der Riechsinneszellen ragen die ziliären Fortsätze in den Schleim. Jede Riechsinneszelle sendet einen Nervenfortsatz zu den Mitralzellen im Riechkolben. Die periglomerulären Zellen stellen meist inhibitorische Verbindungen zwischen den Mitralzellen her. Daneben findet man noch die ebenfalls inhibitorisch wirkenden Körnerzellen, die dendrodendritische Kontakte ausbilden. Der Erregungseingang steht auch zusätzlich unter Kontrolle efferenter Fasern von anderen Gehirnregionen (aus: Schmidt & Schaible (Hrsg.) (2006). Neuro- und Sinnesphysiologie (5., neu bearb. Aufl.), S.341, mit Genehmigung von © Springer). Die Riechzellen sind bipolar aufgebaut. Ihre Dendriten sind zu einem Endkolben mit jeweils 5 bis 20 Zilien (Riechhärchen) verdickt. Auf den Zilien sitzen Rezeptorproteine, ungefähr 350 verschiedene beim Menschen. Bindet ein Geruchsmolekül an sein korrespondierendes Rezeptorprotein werden intrazelluläre Verstärkungsmechanismen in Gang gesetzt, die ein Aktionspotential der Riechzelle auslösen können, das über deren Axon weitergeleitet wird. Weitere Geruchsrezeptoren Neben dem Riechepithel existiert, zumindest rudimentär, auch beim Menschen das vomeronasale Organ. Bei Tieren dient dieses Organ der Aufnahme von Pheromonen, also der Duftstoffe, mit denen Tiere miteinander kommunizieren. Da beim Menschen ein Funktionieren dieses Organs nicht nachgewiesen werden konnte, wird es nicht weiter behandelt. IV Sinnessysteme Ein weiteres „Geruchssystem“ steht mit den freien Endigungen des Nervus trigeminus (des 5. Hirnnerven) in der Riechschleimhaut zur Verfügung. Diese reagieren auf Schmerzreize, aber auch auf Duftstoffe in hohen, als unangenehm empfundenen Konzentrationen wie stechend-beißend oder brennend-scharf. IV.3.2 Reizverarbeitung und Reizweiterleitung Die Axone der Riechzellen (Fila olfactoria) ziehen als Nervus olfactorius in Bündeln zu den Glomeruli im Bulbus olfactorius. Die Glomeruli sind eine komplexe Umschaltstation. Dort konvergieren bis zu 1000 Fasern auf einer Mitralzelle, die den Reiz weiterleitet. Periglomeruläre und Körnerzellen bewirken laterale Hemmungen, auch durch eingehende Efferenzen höherer Hirnregionen. Es wird angenommen, dass die Glomeruli spezifisch für jeweils einen Rezeptortyp beziehungsweise einen Duftstoff sind. Durch einen Geruch (der ja aus verschiedensten Geruchsmolekülen besteht) wird ein spezifisches Glomeruli-Muster aktiviert. Abb. IV.17: Musterbildung und Verschaltung des neuronalen Netzwerks im Bulbus olfactorius. A Verstreut liegende Riechsinneszellen mit gleicher Rezeptorspezifität projizieren in gleiche Glomeruli. Die Aktivität der Fasern des Tractus olfactorius bildet ein komplexes Muster, welches die unterschiedlich starke Aktivierung der Riechrezeptoren durch eine Substanz widerspiegelt. B Afferente Verschaltung und laterale Hemmung im Glomerulus. Die apikalen dendriten der Mitralzellen erhalten Afferenzen vieler gleichartiger Sinneszellen. Die laterale Hemmung durch Körnerund periglomeruläre Zellen wird teilweise über dendro-dendritische Synapsen vermittelt (aus: Klinke, Pape & Silbernagl (2005), Physiologie (5.Aufl.), S.723, Thieme-Verl.). 89 90 Biologische Grundlagen Die Axone der Mitralzellen ziehen als Tractus olfactorius weiter; ein Hauptast kreuzt zum Bulbus olfactorius der anderen Hirnseite, die anderen Fasern ziehen zum Riechhirn. Zum Riechhirn (primärer olfaktorischer Kortex) werden Tuberculum olfactorium, Areale der Amygdala sowie der präpiriforme Kortex gezählt. Fasern aus diesem Bereich bilden eine Verbindung zur Formatio reticularis. Vom Riechhirn wird die Information in das limbische System, Hypothalamus und Hippocampus getragen; ein anderer Weg führt in den Thalamus und (entweder direkt oder über den Thalamus vermittelt) in den Neokortex (orbitofrontaler Kortex und Insel). Abb. IV.18: Zentrale Weiterverarbeitung der Geruchsinformation (aus: Schandry, 2006, S. 295). Die eingebundenen Gehirnstrukturen erlauben Rückschlüsse auf die Funktionen des Geruchssinns. So zeigt die Verbindung zur Formatio retikularis, dass Gerüche Aktivierungen bis hin zu Weckreizen auslösen können. Die emotionale Bewertung, die mit Geruch häufig verbunden ist („Ich kann ihn nicht riechen“), wird über das limbische System vermittelt, sowie die Anregung des Appetits durch Geruch über den Hypothalamus. IV Sinnessysteme Gerüche bleiben sehr gut im Gedächtnis haften, hier spielt vielleicht die Beziehung zu Hippocampus-Strukturen eine Rolle. Und schließlich sorgt der orbitofrontale Kortex für die bewusste Wahrnehmung und Bewertung eines Geruchs. IV.4 Schmecken Den für das Schmecken adäquaten Reiz liefern spezifische, wasserlösliche Moleküle. Sie werden im Speichel gelöst und zu den für Geschmacksempfindung verantwortlichen Rezeptoren gebracht. Menschen können mindestens vier Geschmacksqualitäten „süß“, „salzig“, „sauer“ und „bitter“ unterscheiden. Hinzu kommt die 1908 vom japanischen Forscher Kikunae Ikeda beschriebene Geschmacksrichtung „umami“ (jap., fleischig und herzhaft, wohlschmeckend), die einen eiweißartigen Geschmack beschreibt, und eventuell noch weitere. Die Konzentration der Geschmacksmoleküle im aufgenommenen Stoff bestimmt die Intensität des Geschmacks, aber anscheinend auch die Geschmacksqualität. Auch Temperatur und Einwirkdauer des aufgenommenen Stoffes wirken sich auf den Geschmackseindruck aus. Hinsichtlich der Sensibilität für die Geschmacksqualitäten und die Adaptation unterscheiden sich die Menschen. IV.4.1 Geschmacksknospen und gustatorische Sensoren In verschieden geformten Papillen vor allem auf der Zunge, am weichen Gaumen sowie am Eingang der Speiseröhre befinden sich Geschmacksknospen. Die Zahl der Geschmacksknospen nimmt mit dem Alter ab; bei Kindern liegt die Zahl bei 8000 bis 10000, beim Erwachsenen zwischen 3000 und 8000, bei Hochbetagten bei nur noch 2000. Eine Geschmacksknospe besteht aus Versorgungs-, Stütz- und Basalzellen sowie ungefähr 50 gustatorischen Sensoren, also den Sinneszellen, die Geschmackseindrücke aufnehmen. Diese verschiedenen Zellen bilden die Form einer Knospe mit einer Öffnung, dem Porus, am oberen Ende. Über den Porus erreichen die im Speichel gelösten Geschmacksmoleküle die Sinneszellen. Die gustatorischen Sinneszellen verfügen an ihrem oberen, in den Porus ragenden Ende über kleine Fortsätze, die Mikrovilli. An diesen befinden sich spezifische Proteine, die Geschmacksrezeptormoleküle. Die gustatorischen Sinneszellen 91 92 Biologische Grundlagen werden ständig erneuert, nach circa 10 Tagen sterben sie ab und werden durch eine von der Basalzelle neu gebildete ersetzt. Abb. IV.19 Geschmacksknospen auf der Zunge a Lage der Geschmacksknospen auf den drei verschiedenen Typen von Geschmackspapillen. b Bau und Innervation einer Geschmacksknospe. Die 3 Zellelemente: Sinneszellen, Stützzellen und Basalzellen sind knospenartig angeordnet und gegenüber der Epitheloberfläche etwas versenkt, somit entsteht ein flüssigkeitsgefüllter Raum, in den die Mikrovilli der Sinneszelle ragen. Die Geschmackssinneszellen werden durch afferente Nervenfasern innerviert. Die Verbindung hat alle Eigenschaften einer chemischen Synapse. Einzelne afferente Fasern können mehrere Sinneszellen versorgen (aus: Schmidt & Schaible (Hrsg.) (2006). Neuro- und Sinnesphysiologie (5., neu bearb. Aufl.), S.329, mit Genehmigung von © Springer). Dockt nun ein Geschmacksmolekül an das Geschmacksrezeptormolekül einer Sinneszelle an, kommt es zu einer Depolarisation der Zelle, die zu einem Aktionspotential in der afferenten Nervenzelle führen kann, die mit der Sinneszelle über eine Synapse verbunden ist. Die gustatorischen Sinneszellen sind also sekundäre Sinneszellen. Die einzelnen Sinneszellen scheinen auf alle vier Grundgeschmacksqualitäten ansprechen zu können, allerdings mit unterschiedlichen Intensitäten. IV.4.2 Reizverarbeitung und Reizweiterleitung Mehrere Sinneszellen, meist sogar mehrere Geschmacksknospen werden von einem afferenten Nerv versorgt. Ein ableitendes Neuron ist in der Regel nicht spezifisch durch den Geschmack einer Qualität erregbar, sondern weist ein Geschmacksprofil mit mehr oder weniger hoch ausgeprägten Empfindlichkeiten für die vier Geschmacksqualitäten auf. Die von diesen Neuronen versorgten rezeptiven Felder überlappen sich. IV Sinnessysteme Erst die Zusammenführung der Informationen aus verschiedenen Nervenfasern beziehungsweise den von ihnen gemeldeten Erregungsmustern erlaubt die Beurteilung eines Geschmacks hinsichtlich Qualität und Intensität. Die Geschmacksbahn Die Neuronen, die die Erregung von den gustatorischen Sinneszellen aufnehmen, werden in drei Nerven weitergeleitet, dem Nervus glossopharyngeus (9. Hirnnerv), dem Nervus facialis (7. Hirnnerv) und dem Nervus vagus (10. Hirnnerv). Im Nucleus tractus solitarii der Medulla oblongata findet die Umschaltung auf das zweite Neuron der Geschmacksbahn statt. Aus diesem Bereich werden Informationen an viszeromotorische und sekretorische Kerne weitergeleitet. Hierüber lassen die verschiedenen Reflexe des Verdauungsapparates wie Speichelfluss, Magensaftsekretion oder Speiseröhrenperistaltik erklären. Die Zahl der zweiten Neuronen der Geschmacksbahn ist geringer als die der ersten (Konvergenz). Im weiteren Verlauf kreuzen sie auf die Gegenseite und erreichen den Hypothalamus, den Thalamus und im limbischen System denselben Bereich wie die olfaktorischen Afferenzen. Mit der Beteiligung des Hypothalamus und des limbischen Sytems können die vegetativen und affektiven Begleiterscheinungen einer gustatorischen Reizung erklärt werden. Vom Thalamus werden gustatorische Informationen schließlich in die primäre Geschmacksrinde gebracht. Diese befindet sich neben den Projektionsfeldern der Mundhöhle im sensorischen Kortex. An dieser Stelle wird die ausdifferenzierte Wahrnehmung von Geschmacksnuancen angenommen. 93 94 Biologische Grundlagen Abb. IV.20 Geschmacksbahn von den Sinneszellen der Zunge bis zur Hirnrinde (aus: Schandry, 2006, S. 301). IV.5 Gleichgewicht und Bewegung Als adäquaten Reiz für die Gleichgewichtssinne kann man prinzipiell die physikalischen Größen Schwerkraft und Drehmoment bezeichnen. Mit diesen Sinnen werden Informationen darüber verarbeitet, in welcher Lage (in Bezug auf die Schwerkraft) sich der Mensch befindet und ob und in welche Richtung er sich bewegt. Dabei ist es zunächst nicht von Interesse, ob eine Bewegung aktiv (also selbsterzeugt) oder passiv ist. IV.5.1 Vestibularorgan (Makulaorgane, Bogengänge) und Haarzellen Vestibularorgan Gemeinsam mit der Kochlea bildet das Vestibularorgan (Gleichgewichtsorgan) das Labyrinth des Innenohrs. Das Vestibularorgan besteht aus zwei funktionellen Teilen, den beiden Makulaorganen sowie den Bogengängen, und ist, wie die Kochlea, mit Endolymphe gefüllt. IV Sinnessysteme Abb. IV.21: Schema der Bogengangsorgane und der Maculaorgane des Innenohrs. Endolymphe (dunkel) und Perilymphe (hell) stehen mit Endolymphe und Perilymphe der Cochlea in Verbindung (aus: Schmidt & Schaible (Hrsg.) (2006). Neuro- und Sinnesphysiologie (5., neu bearb. Aufl.), S.313, mit Genehmigung von © Springer). Die beiden Makulaorgane, Sacculus und Utriculus, erfassen Informationen über Lage und Lageveränderungen des Menschen im Raum. Beide enthalten Sinnesepithelien, in denen sich die Sinneszellen des Vestibularorgans, die Haarzellen, befinden. An deren oberen Ende befinden sich je 60 bis 100 Sinneshärchen (Stereozilien), deren längstes Kinozilium genannt wird. Die Stereozilien ragen in die Otolithenmembran, eine gallertartige Masse, die durch kleinste Kalkkristalle, die Otolithen, beschwert wird. Bei einer linearen Beschleunigung (Translationsbeschleunigung), wie sie zum Beispiel bei einem Sprung oder Sturz, beim Anfahren oder Bremsen eines Autos entsteht, werden die mit dem Körper fest verbundenden Sinneszellen mitbewegt. Die Otholitenmembram reagiert träger und bleibt zurück, wodurch die Stereozilien ausgelenkt werden. Die Auslenkung führt je nach Richtung zu einer Erregung oder Hemmung der ableitenden Nerven. Auch in der Ruhe verbleibt eine gewisse Grundaktivität. 95 96 Biologische Grundlagen Abb. IV.22: Transduktion in den Stereozilien. Bei einer Auslenkung der Stereozilien in Richtung auf das Kinozilium kommt es zu einer Dehnung der tip links, Kaliumkanäle öffnen sich und Kaliumionen strömen aus der Endolymphe in die Haarzelle. Es wird vermehrt Glutamat an der Synapse zur afferenten Faser ausgeschüttet, die Aktionspotentialfrequenz steigt an. Werden die Stereozilien in die entgegengesetzte Richtung ausgelenkt, nimmt entsprechend die Aktivität in der Afferenz ab (aus: Schandry, 2006, S. 286). Bei einer aufrechten Kopfhaltung befindet sich der Sacculus ungefähr in senkrechter Stellung, es kommt zu einer ständigen Reizung der Haarzellen des Sacculus. In dieser Kopfhaltung liegt der Utriculus nahezu waagrecht, seine Sinneszellen sind nicht gebogen. Über die beiden Organe können daher Lage und Lageveränderungen im Raum vollständig abgebildet werden. Die drei Bogengänge (vorderer, hinterer und seitlicher Bogengang) verfügen ebenfalls über ein Sinnesepithel, deren Haarzellen beziehungsweise Stereozilien in eine gallertartige Masse hineinragen. In den Bogengängen wird diese Masse Cupula genannt; sie bildet gemeinsam mit den Haarzellen eine Art Sperrwand in jedem der Bogengänge. Die Cupula ist nicht beschwert und reagiert daher nicht bei Translationsbeschleunigung, sondern auf Drehbeschleunigungen, wie sie bei Kopfdrehungen entstehen. Bei Kopfdrehungen machen die Bogengänge die Bewegung mit, die darin befindliche Endolymphe bleibt zunächst zurück. Dadurch entsteht Druck auf die Cupula, was zu deren Auslenkung und damit zu einer Auslenkung der Stereozilien führt. Der ableitende Nerv wird je nach Richtung der Auslenkung aktiviert. Die drei Bogengänge haben jeweils verschiedene Ausrichtungen, so dass sie in der Lage sind, die Gesamtheit aller möglichen Drehbewegungen vollständig abzubilden IV Sinnessysteme 97 Abb. IV.23: Schema eines Bogenganges mit Cupula und Haarzellen. Wird der Kopf gedreht (Pfeil), wird auch der Bogengang gedreht. Cupula und Endolymphe hingegen bleiben zurück. Dadurch werden die Stereozilien ausgelenkt (aus: Schmidt & Schaible (Hrsg.) (2006). Neuro- und Sinnesphysiologie (5., neu bearb. Aufl.), S.319, mit Genehmigung von © Springer). IV.5.2 Reizweiterleitung und Reizverarbeitung Die Haarzellen des Vestibularorgans sind sekundäre Sinneszellen, die eine Aktivierung der Fasern des afferenten Nervus vestibularis bewirken. Einige der Fasern des Nervus vestibularis ziehen direkt zum Kleinhirn, der größte Teil zu den Vestibularkernen der Medulla oblongata, wo auch Afferenzen aus dem visuellen System und den Propriozeptoren eingehen. Über die Medulla oblangata erreichen Informationen aus dem Gleichgewichtsorgan das Kleinhirn, das seinerseits Informationen in die Vestibularkerne zurücksendet. Von den Vestibularkernen gibt es Verbindungen zu den Augenmuskelkernen im Mittelhirn sowie den Motoneuronen des Rückenmarks. Hierüber werden Ausgleichsbewegungen der Augen und des Körpers eingeleitet. Bei Drehbewegungen des Kopfes werden durch vestibulookuläre Reflexe reflektorisch die Augen so bewegt, dass sich das Gesichtsfeld nicht verschiebt. Mit den vestibulospinalen Reflexen werden die Bewegungen zum Beispiel beim Übergang vom Stehen zum Gehen so koordiniert, dass man nicht stürzt. Und schließlich erreichen die Informationen aus den Vestibulariskernen über den Thalamus den Kortex und dort die sensumotorischen Areale, wo die Informationen aus den verschiedenen Systemen integriert werden. So kann man sich, auch bei geschlossenen Augen, die Körper- und Kopfhaltung jederzeit bewusst machen. vestibuläre Reflexe 98 Biologische Grundlagen Abb. IV.24: Die wichtigsten Verbindungen der Vestibulariskerne (aus: Schandry, 2006, S. 288). IV.6 Hautsinne Die Haut ist das größte Organ des Menschen. Unterscheiden lassen sich die behaarte Haut, die den größten Teil des Körpers bedeckt und von zum Teil sehr dünnen Haaren bedeckt ist, und die unbehaarte Haut, die sich an den Handflächen, den Fingerspitzen, den Fußsohlen und den Genitalien befindet. Über die Haut werden eine Reihe von Sinneseindrücken vermittelt. Mechanorezeptoren nehmen die verschiedenen Arten taktiler Reize wie Berührungen, Kitzeln, Vibration, Druck oder Spannung auf. Thermorezeptoren werden bei Temperaturänderungen aktiv und Schmerzrezeptoren bei noxischen Reizen, also Reizen, die gewebsschädigend oder potentiell gewebsschädigend sind. Abb. IV.25: Schnitt durch die behaarte und unbehaarte Haut mit den wichtigsten sensorischen Endigungen. Freie Nervenendigungen sind wesentlich häufiger als hier dargestellt. Die Funktion der schnell adaptierenden Meißner-Körperchen, die nur in der unbehaarten Haut vorkommen, wird in der behaarten Haut von den Haarfollikelrezeptoren übernommen. Statt der langsam adaptierenden Merkel-Endigungen finden sich in der behaarten Haut die in Struktur und IV Sinnessysteme Funktion ähnlichen Tastscheiben (aus: Klinke, Pape & Silbernagl (2005), Physiologie (5.Aufl.), S.630, Thieme-Verl.). IV.6.1 Haut, Mechanosensoren, Thermosensoren und Schmerzrezeptoren Mechanosensoren Mechanosensoren oder Mechanorezeptoren nehmen die verschiedenen taktilen Reize auf. Alle verschiedenen Typen der Mechanosensoren sind primäre Sinneszellen. Freie Nervenendigungen sind die dünnen, unmyelinisierten Enden von afferenten, myelinisierten Nervenfasern, die in die Haut hineinragen. Die Nervenendigungen können den Schaft von Haarwurzeln umwickeln und reagieren, wenn sich das Haar aufgrund von Zug oder Druck verformt. Sie übernehmen damit die Funktion eines Berührungssensors. Bei den Merkel-Tastzellen hat sich das Ende eines Nervs oval verformt und reagiert mit hoher Empfindlichkeit auf Druck. Die Meissner-Tastkörperchen weisen am Ende einen lamellenförmigen Aufbau auf; sie reagieren auf die Geschwindigkeit von Verformungen und adaptieren schnell. Auch die Vater-PaciniLamellenkörperchen weisen am Ende eine lamellenförmige Struktur auf, die allerdings konzentrisch angeordnet ist. Sie reagieren auf Druckänderungen beziehungsweise Vibrationen. Bei den Ruffini-Körperchen ist das Nervenende spindelförmig verformt, sie reagieren auf Dehnung. Die verschiedenen Mechanosensoren unterscheiden sich nicht nur hinsichtlich ihres Aufbaus, sondern auch hinsichtlich ihrer Adaptationsgeschwindigkeit und ihrer Ruheaktivität. So adaptieren Merkel-Tastkörperchen und RuffiniKörperchen relativ langsam und sind so in der Lage, auch andauernde Reizung zu indizieren. Doch während die Merkel-Tastkörperchen nur bei einer Reizung aktiv sind, weisen die Ruffini-Körperchen auch ohne Reizung eine gewisse Spontanaktivität auf, die sich bei Reizung verstärkt. Bei Arten sind jedoch in der Lage, die Intensität einer Reizung zu kodieren. 99 100 Biologische Grundlagen Abb. IV.26: Rezeptive Felder und Reizantworten von Mechanosensoren (aus: Schandry, 2006, S. 232). Jeder afferente Nerv hat mehrere Aussprossungen, die gemeinsam sein rezeptives Feld versorgen. Die Größe der rezeptiven Felder ist sehr unterschiedlich, sie hängt von der Art des Mechanosensors ab und dessen Lage im Körper. So weisen Meissner-Tastkörperchen und Merkel-Tastzellen relativ kleine Felder auf und sind in relativ großer Zahl an den Fingerspitzen zu finden. Zweipunktschwelle Da die ableitenden Nerven nicht konvergieren, setzt sich diese hohe Auflösung bis in höhere Hirnstrukturen fort. Erfasst wird dies durch die Zweipunktschwelle, also den Abstand, den zwei simultane Berührungen haben müssen, um als zwei (und nicht nur als eine) wahrgenommen zu werden; die Zweipunktschwelle wird üblicherweise mit zwei Zirkelspitzen geprüft. Die Funktion der verschiedenen Mechanosensoren kann auch aktiv genutzt werden, wenn zum Beispiel durch das Bestreichen von Oberflächen mit den Fingern Informationen unterschiedlicher Art erzeugt werden, z.B. wenn eine Münze in der Hosentasche hinsichtlich Größe und Randgestaltung identifiziert wird. Thermosensoren Temperatur wird durch zwei Typen von Thermosensoren erfasst. Die Kaltsensoren bestehen meist aus unmyelinisierten, zum Teil auch aus myelinisierten Nervenendigungen. Ihre langsame Spontanaktivität steigt schnell an, wenn es zu einer Abkühlung kommt. Sie adaptieren jedoch auch relativ schnell. Das Maximum ihrer Empfindlichkeit liegt bei 25° Celsius. Auch die Warmsensoren mit einem Empfindlichkeitsmaximum von 50° Celsius adaptieren schnell. Es gibt etwa 3 bis 5mal so viele Kalt- wie Warmsensoren; beide Typen von Thermosensoren sind ungleich über die Körperoberfläche verteilt. Je schneller sich die Temperatur än- IV Sinnessysteme dert, desto eher wird sie diese Veränderung und damit die Temperatur wahrgenommen. Weiterhin nimmt die Wahrscheinlichkeit einer bewussten Temperaturempfindung mit der Größe des betroffenen Körperareals zu. Nozizeptoren Nozizeptoren sind freie Nervenendigungen, die auf noxische Reize reagieren. Sie sitzen in der Haut, den Muskeln, Gelenken, inneren Organen, inneren Hohlräumen und den Hirnhäuten, kurz in allen Körpergeweben mit Ausnahme des Hirngewebes und des Leberparenchyms. Man unterscheidet nach den auslösenden Reizen Nozizeptoren für mechanische Reize, die auf starken (schmerzhaften) Druck reagieren, Thermonozizeptoren, die auf Hitze ab 45° Celsius reagieren und chemosensible Nozizeptoren, die auf bestimmte chemische Substanzen reagieren. Die meisten Nozizeptoren sind polymodal, das heißt, sie reagieren sowohl auf mechanische als auch auf thermische und chemische Schmerzreize. Es gibt jedoch eine Gruppe mechanoinsensitiver Nozizeptoren, die weder auf mechanische noch thermische Reize reagieren. Die Hyperalgesie (gesteigerte Schmerzempfindlichkeit) bei Entzündungen entsteht durch die Anreicherung bestimmter, schmerzauslösender Substanzen, den sogenannten Entzündungsmediatoren. Gleichzeitig werden jedoch auch vermehrt Rezeptoren für schmerzhemmende Substanzen wie Opioide gebildet. Bei den Fasern, die die Erregung weiterleiten, handelt es sich um langsamleitende C-Fasern (unmyelinisiert) und in geringerer Zahl ebenfalls relativ langsam leitende, schwach myelinisierte Aδ–Fasern. Der „helle“ erste Schmerz, der zum Beispiel der bei einem Schnitt zunächst entsteht, wird über die Aδ–Fasern vermittelt und löst Schutzreflexe aus. Es folgt der über die C-Fasern vermittelte, dumpfe zweite Schmerz, der lange anhalten kann. 101 102 Biologische Grundlagen Abb. IV.27 Unterteilung der somatoviszeralen Sensibilität und der zugehörigen Sinnesrezeptoren (aus: Klinke, Pape & Silbernagl (2005), Physiologie (5.Aufl.), S.629, Thieme-Verl.). IV.6.2 Reizweiterleitung und Reizverarbeitung Die (sensiblen) Fasern von Mechanosensoren, Thermosensoren und Nozizeptoren treten über das Hinterhorn in das Rückenmark ein. Ein Teil der von den Mechanosensoren ableitenden Fasern ist bereits im Rückenmark mit Interneuronen oder Motoneuronen verschaltet und bewirkt unmit- IV Sinnessysteme telbare Reflexe auf Druck- oder Berührungsreize. Der andere Teil steigt im Hinterstrang zu den Hinterstrangkernen der Medulla oblongata auf, wo die Leitungsbahn nach einer synaptischen Verschaltung auf die Gegenseite kreuzt (Lemniscus medialis). Von dort laufen die Bahnen in den ventrobasalen Teil des Thalamus und schließlich in den primären somatosensorischen Kortex. Mechanosensorische Informationen aus dem Kopf- und Halsbereich werden über den Nervus trigeminus in das zentrale Nervensystem geleitet; die afferenten Nervenfasern werden in Mittelhirn synaptisch verschaltet bevor sie auf die Gegenseite kreuzen und ebenfalls über den Thalamus in den primären somatosensorischen Kortex ziehen. Wegen der Kreuzung (der Schleife) in der Medulla oblongata wird dieses System der Somatosensorik als lemniscales System bezeichnet. Abb. IV.28: Zentralnervöse Weiterleitung der Somatosensorik. Die Information aus der Haut wird im Wesentlichen über das Vorderseitenstrangsystem sowie das Hinterstrangsystem zum Gehirn geleitet. Beide Systeme haben als Umschaltstelle den Thalamus und als Zielgebiete den primären und sekundären somatosensorischen Kortex (aus: Schandry, 2006, S. 237). 103 104 Biologische Grundlagen Die von Thermosensoren und Nozizeptoren ableitenden Fasern kreuzen bereits auf der Ebene des Rückenmarksegments, in das sie in das Rückenmark eintreten, bisweilen auch auf der nächsten oder übernächsten Ebene. Sie ziehen dann im Vorderseitenstrang zur Formatio retikularis und größtenteils zum ventrobasalen Thalamus und schließlich zum primären somatosensorischen Kortex. Auch in diesen Systemen wird der Kopf- und Halsbereich durch den Trigeminusnerv versorgt, der über eine Verschaltung und Kreuzung im Mittelhirn zum primären somatosensorischen Kortex weiterleitet. In Abgrenzung vom lemniscalen wird hier vom extralemniscalen System gesprochen. Für die kortikale Bewertung der Informationen kann man die beiden Systeme etwas genauer ansehen. Die Fasern des lemniscalen Systems sind myelinisiert und schnellleitend. Sinn dieses Systems ist eine schnelle und genaue Identifikation der verursachenden Reize hinsichtlich ihrer Intensität und Lokalisation. Anders ist es beim extralemniscalen System, die Fasern leiten Informationen langsamer und weniger präzise weiter. Hier sind in der Regel auch „gröbere“ Reaktionen nützlich. Ein Beispiel soll dies verdeutlichen: Nimmt man ein öliges, aber sehr dünnes Glas in die Hand, wird über die verschiedenen Mechanosensoren angezeigt, dass dieses aus der Hand zu rutschen droht. Ein zu festes Zugreifen könnte jedoch das Glas eindrücken, so dass in abgestimmten Bewegungen und Rückmeldungen die richtige Krafteinwirkung gefunden wird. Ist das Glas jedoch sehr heiß und eine Gewebsschädigung droht, wird man es fallen lassen, ohne „darüber nachzudenken“. Besonders bei Schmerzen werden neben dem primären somatosensorischen Kortex auch weitere Gebiete des Kortex einbezogen. Sie sind für emotionale Bewertung, gedächtnismäßige Verarbeitung und Auslösung komplexer Verhaltensmuster zur Schmerzbekämpfung verantwortlich. IV Sinnessysteme Studientechnischer Anhang 1. Kenntnisse Auge o Optischer Apparat (Aufbau des Auges, Augenmuskeln, Linse, Pupille, Iris und zugehörige Muskeln) o Netzhaut und Aufbau Sinneszellen des Auges o adäquater Reiz: Licht und Wellenlänge o Photoceptoren (Stäbchen, Zapfen) o Zelltypen der Retina (Bipolarzellen, Horizontalzellen, Ganglienzellen und Varianten, amakrine Zellen) Rezeptive Felder o Definition o Formen: On- und Off-Zentren Farbensehen o Arten von Zapfen o Farbtheorien: trichromatisch, Gegenfarbtheorie, KriesZonentheorie Reizleitung im visuellen System o Sehnerv, Sehbahn, Sehstrahlung o Verlauf der Reizleitung (Chiasma opticum, Colliculi superioes, Corpus geniculatum laterale, visueller Kortex) Organisation und Funktionen im visuellen Kortex o Rezeptive Felder der Kortexneurone (Orientierung, Bewegung, Länge) o Dominanzsäulen (Orientierungssäulen, Blobs) und Hypersäulen Verarbeitungsmechanismen im visuellen System o Farbkonstanz, Größenkonstanz o räumliches Sehen (korrespondierende Netzhautareale und Querdisparation, Verdeckung, Größe, Perspektive, Parallaxe, Färbung) Ohr o Aufbau des Ohrs mit Außenohr, Mittelohr, Innenohr o Aufbau Mittelohr: Hammer, Amboss, Steigbügel o Aufbau Innenohr: Kochlea, Scalae tympani, media, vestibuli, Helikotrema, Basilar- und Tektorialmembram Sinneszellen des Ohrs o adäquater Reiz: Schalldruck, hörbare Frequenz- und SchalldruckBereiche o Corti-Organ o Haarzellen und Stereozilien Reizleitung im auditiven System o Hörnerv, Hörbahn, Hörstrahlung o Verlauf der Reizleitung (Nuclei cochleares, Lemniscus lateralis, Colliculi inferores, Corpus geniculatum mediale) Auditiver Kortex o primärer auditiver Kortex, sekundärer auditiver Kortex 105 106 Biologische Grundlagen o Wernicke- und Broca-Sprachzentrum Verarbeitungsmechanismen im auditiven System o Ortsprinzip der Frequenzkodierung o Periodizitätsanalyse o räumliches Hören (Laufzeit- und Intensitätsunterschiede, Verzerrungen) o Prinzip der Mustererkennung und Filterung Nase o Atemstrom und „Schnüffeln“ o Riechepithel (Riechzellen, Basalzellen, Bowman-Drüsen) Sinneszellen des Riechepithels o adäquater Reiz: Geruchsmoleküle (Duftstoffe und Düfte) o Riechsinneszelle mit Zilien o Geruchsrezeptoren außerhalb des Riechepithels (vomeronasales Organ, N. trigeminus) Reizleitung des Geruchssinns o Mitralzellen, Glomeruli im Bulbus Olfactorius und deren Funktion o Nervus olfactorius, Tractus olfactorius o Weg der Geruchsinformation (Nervus olfactorius, Tractus olfactorius, Riechhirn, Formatio retikularis, limbisches System, Thalamus und Neokortex) Riechhirn,übergeordnete Hirnstrukturen und deren Funktion o Riechhirn (präpiriformer Kortex, Tuberculum olfactorium, Nucleus corticalis amygdalae) o Formatio retikularis, limbisches System, Hypothalamus, Hippocampus, orbitofrontaler Kortex Zunge o Papillen der Zunge, Gaumen, Speiseröhre o Geschmacksknospen (Aufbau, gustatorische Sensoren, Stütz- und Basalzellen) Gustatorische Sensoren o adäquater Reiz: Geschmacksmoleküle (süß, sauer, salzig, bitter, umami) o Aufbau gustatorischer Sensoren, Mikrovilli (sekundäre Sinneszellen) Reizleitung des Geschmackssinns: Stationenen und Funktion o N. glossopharyngeus, N. facialis, N. vagus o weiterer Weg der Geschmacksinformation: Nucleus tractus solitarii, Lemniscus medialis, Thalamus, primäre Geschmacksrinde o Einbindung: Hypothalamus, limbisches System Vestibularorgan o Makulaorgane und Otholitenmembran o Bogengänge und Cupula Haarzellen o adäquater Reiz: Schwerkraft, Beschleunigung, Drehung o Haarzellen und Sterozilien (Kinozilium) IV Sinnessysteme Reizleitung o Nervus vestibularis o Vestibularkerne und deren Verbindung zu Kleinhirn, Motorik, Kortex Haut o Haut: behaart, unbehaart Rezeptoren der Haut und jeweils adäquate Reize o Mechanorezeptoren: freie Nervenendigungen, Merkel-Tastzellen, Meissner-Tastkörperchen, Vater-Pacini-Lamellenkörperchen, Ruffini-Körperchen o Thermosensoren o Nozizeptoren Reizleitung; Stationen und Funktion o Afferente Fasern: Größe der rezeptiven Felder und Geschwindigkeit der Reizleitung o Lemnicales System der Mechanosensoren o Extralemnicales System der Thermo- und Nozizeptoren Weitergehende Verarbeitung o Lemniscales System o Extralemniscales System o Weitergabe von Information aus primärem sensorischen Kortex: Bewertungen 107 108 Biologische Grundlagen 2. Literatur Grundlage: Schandry, R. (2006). Biologische Psychologie (2. überarb. Aufl.). Weinheim: Beltz. Kapitel 12: Das visuelle System Kapitel 13: Gehör Kapitel 15: Riechen, Schmecken und der allgemeine chemische Sinn Kapitel 14: Gleichgewichts-, Bewegungs- und Lagesinn Kapitel 11: Somatosensorik Kapitel 16: Schmerz, v.a. Abschnitt 16.1 Das nozizeptive System Vertiefend: Bear, M. F., Connors, B. W., & Paradiso, M. A. (2007). Neuroscience. Exploring the brain (3d Ed.). Philadelphia etc.:Lippincott Williams & Wilkins. Eysel, U. (2006). Sehen. In R.F.Schmidt & H.-G. Schaible (Hrsg.), Neuro- und Sinnesphysiologie (S. 243-286). Heidelberg: Springer. Handwerker, H.O. (2006). Somatosensorik. In R.F.Schmidt & H.-G. Schaible (Hrsg.), Neuro- und Sinnesphysiologie (S. 203-228). Heidelberg:Springer. Handwerker, H.O. & Schaible, H.-G.(2006). Somatosensorik. In R.F.Schmidt & H.-G. Schaible (Hrsg.), Neuro- und Sinnesphysiologie (S. 229-242). Heidelberg:Springer. Hatt, H. (2006). Geruch. In R.F.Schmidt & H.-G. Schaible (Hrsg.), Neuro- und Sinnesphysiologie (S. 340-351). Heidelberg:Springer. Hatt, H. (2006). Geschmack. In R.F.Schmidt & H.-G. Schaible (Hrsg.), Neuround Sinnesphysiologie (S. 328-339). Heidelberg:Springer. Zenner, H.P. (2006). Hören. In R.F.Schmidt & H.-G. Schaible (Hrsg.), Neuround Sinnesphysiologie (S. 287-311). Heidelberg:Springer. Zenner, H.P. (2006). Gleichgewicht. In R.F.Schmidt & H.-G. Schaible (Hrsg.), Neuro- und Sinnesphysiologie (S. 312-325). Heidelberg:Springer. V Erfassungsmethoden der Biologischen Psychologie V 109 Erfassungsmethoden der Biologischen Psychologie Lernziele Die Studierenden sollen verschiedene (physiologische) Methoden kennen lernen, mit denen im Rahmen psychologischer Forschung die körperlichen Begleiterscheinungen (Korrelate) psychischer Prozesse erfasst werden, Ansatz und Logik dieser Methoden kennen lernen, Verständnis für die ethischen und erkenntnis- und wissenschaftstheoretischen Grenzen dieser Methoden entwickeln. Grundsätzlich geht es in der biologischen Psychologie um die Zusammenhänge zwischen Erleben und Verhalten auf der einen Seite und biologischen Prozessen auf der anderen Seite. Dabei können verschiedene Forschungsansätze verwendet werden. Zunächst können spontane Veränderungen in den biologischen beziehungsweise physiologischen Variablen beobachtet werden. Eine solche Veränderung ist zum Beispiel die Tagesrhythmik, bei der sich Schlafen und Wachen, aber auch Phasen höherer und geringerer Aktivierung abwechseln. Diese Phasen lassen sich sowohl psychisch („munter und voll fit“ versus „müde, fast schon eingeschlafen“) als auch physiologisch in der elektrischen Aktivität des Gehirns nachweisen. Ein weiteres Beispiel hierfür ist der Menstruationszyklus der Frau, wo die hormonellen Schwankungen und psychisches Erleben aufeinander bezogen werden. Ein anderer Ansatz variiert in Studien oder Experimenten physiologische oder biologische Parameter, um deren Einfluss auf Erleben und Verhalten zu beobachten. So werden die Auswirkungen psychotroper Substanzen (wie Alkohol) auf die psychische Leistungsfähigkeit (wichtig für die Fahrtüchtigkeit) untersucht oder die kognitive und emotionalen Bewertungen verschiedener Schmerzreize. Diesen Studien sind beim Menschen natürlich ethische Grenzen gesetzt. Und schließlich können die in Folge von psychischen Veränderungen herbeigeführten biologischen Veränderungen erfasst werden, zum Beispiel die Aktivität des Gehirns beim Lösen einer Mathematikaufgabe oder der Anstieg der HerzKreislauf-Aktivität bei heftigen Emotionen. Bei der Interpretation von Biosignalen ist zu beachten, dass diese bei verschiedenen Personen unterschiedlich ausgeprägt sind, das heißt stark interindividuell variieren. Um diese Ausgangslagenproblematik zu beachten, bezieht man Signale, die z.B. in Folge eines Reizes aufgetreten sind, auf einen individuell bestimmten Grundwert. Ein kleines Beispiel: Ein Puls von 85 (Schlägen in der Mi- Interpretation von Biosignalen 110 Biologische Grundlagen nute) in Folge eines emotional belastenden Reizes ist bei einem Menschen mit einem Ruhepuls von 60 anders zu interpretieren als bei einer Person mit einem Ruhepuls von 80. Auch die Reaktivität der Messung sowie Artefakte sind zu beachten. Physiologische Messmethoden leiten physikalisch, bisweilen auch chemisch bestimmbare Größen ab. Dazu werden Apparate benötigt, die die Reaktionsweise einer Person beeinflussen können. So kann schon die Messung des Blutdrucks mit einer Blutdruckmanschette während einer Leistungsaufgabe ablenken und Ergebnisse verfälschen. Die von Messapparaten aufgenommen Signale müssen verstärkt und/oder gemittelt werden, auch hier verstecken sich möglicherweise verzerrende Effekte (Messartefakte, statistische Artefakte). nicht-invasive vs. invasive Verfahren In diesem Kurs werden einige der Messmethoden vorgestellt, die verwendet werden, um biologische Kenngrößen zu erfassen. Dabei werden in der psychologischen Forschung in der Regel nicht-invasive Verfahren verwendet, also Verfahren, die keine körperliche Verletzung oder Schädigung oder körperliche Gefährdung für die zu untersuchende Person bedeuten. Zu invasiven Verfahren zählt bereits die kleine Verletzung, die entsteht, wenn Blut abgenommen wird. Der Einsatz invasiver Messmethoden muss aus ethischen Gründen bestens überlegt und der mögliche Einsatz alternativer Verfahren nicht-invasiver Art gründlich geprüft werden. Des Weiteren ist beim Einsatz invasiver Verfahren ein Arzt oder eine Ärztin zu beteiligen und die betroffene Person muss dem invasiven Eingriff zustimmen, nach dem sie entsprechend informiert worden ist. Im Forschungskontext muss darüber hinaus die Zustimmung einer Ethikkommission eingeholt werden, wenn Forschungsprojekte an Menschen, aber auch an Tieren, mit invasiven Eingriffen verbunden sind. V Erfassungsmethoden der Biologischen Psychologie V.1 Verfahren zur Erfassung zentraler Aktivität Das Gehirn als zentrale Verarbeitungsstelle für Reize und Steuerungsorgan für die Reaktionen des Menschen wird durch verschiedenste Messmethoden in seiner Struktur und Funktion beschrieben, wobei für die psychologische Erkenntnisbildung vor allem die funktionsabbildenden Verfahren wichtig sind. V.1.1 Strukturabbildende Verfahren Röntgenbild Erste Aufschlüsse über Form und Struktur des Gehirns liefert das Röntgenbild. Röntgenstrahlen durchdringen das Gehirn. Da die unterschiedlichen Gewebsarten des Schädels beziehungsweise die flüssigkeitsgefüllten Hohlräume die Strahlungsenergie unterschiedlich stark absorbieren, ergibt sich ein zweidimensionales Abbild der durchleuchteten Struktur. Durch verschiedene Aufnahmewinkel lassen sich Rückschlüsse auch auf die dreidimensionale Struktur erzielen. Allerdings ist die Strahlenbelastung relativ hoch. Des Weiteren sind die Aufnahmen relativ kontrastarm. Röntgenaufnahmen werden vor allem in der Klinik eingesetzt, um Verletzungen darzustellen. Computertomographie (CT) Eine Weiterentwicklung der Röntgentechnik ist die Computertomographie (CT). Dabei wird eine Röntgenröhre im 360°-Winkel um den Kopf herumgeführt und Schnittbilder im Abstand von 1 bis 10 mm erstellt. Durch diese spezielle Aufnahmetechnik entsteht ein kontrastreiches, räumliches Bild der Strukturen des Gehirns. Aber auch hier wird die zu untersuchende Person mit Strahlung belastet. Auch die Computertomographie des Gehirns wird vor allem in der medizinischen Klinik verwendet, zum Beispiel bei der Darstellung degenerativer Prozesse des Gehirns bei demenziellen Erkrankungen. Abb. V.1: Es ist ein CT des Gehirns zu sehen (Horizontalschnitt). Man erkennt in der rechten Bildhälfte einen weißen Bereich infolge eines Hämatoms (Bluterguss), aufgenommen am Tage der Blutung (aus: Schandry, 2006, S. 580). 111 112 Biologische Grundlagen V.1.2 Aktivitäts- und funktionsabbildende Verfahren V.1.2.1 Elektroenzephalogramm (EEG) Das Elektroenzephalogramm (EEG) zeichnet hirnelektrische Vorgänge auf. Dazu werden Elektroden auf die Schädeloberfläche aufgesetzt, die Potentialschwankungen (Spannungsschwankungen) aufzeichnen. Diese Schwankungen entstehen durch exzitatorische oder inhibitorische Prozesse an den Neuronen des Kortex, die in jeweils relativ großen Neuronenpopulationen synchron und gleichartig ablaufen. Dadurch ändern sich aufgrund der Dipoleigenschaften der Neuronen die um sie aufgebauten elektrischen Felder, was wiederum durch Elektroden erfasst werden und auf einem Zeitstrahl abgebildet werden kann. Eine Elektrode erfasst dabei Spannungsänderungen, die von 100.000 bis 1.000.000 Neuronen verursacht werden. Für die Erstellung von Elektroenzephalogrammen werden in der Regel Standardableitungen genutzt, das sind Festlegungen der Stellen auf dem Schädel, an denen die Elektroden platziert werden. Abb. V.2 Grundlagen des EEG. A Registrierung des EEG mit Hilfe von Elektroden, die nach dem internationalen 10-20-System an definierten Standorten an der Schädeloberfläche angebracht werden. Schema einer bipolaren Messung zwischen zwei differenten Elektroden und einer unipolaren Messung zwischen einer differenten und einer indifferenten Referenzelektrode. B Zelluläre Mechanismen der Entstehung des EEG. Durch aktive Synapsen in kortikalen Neuronen werden elektrische Felder generiert, deren Summe mit einer EEG-Elektrode an der Schädeloberfläche registriert wird (nach 1) (aus: Klinke, Pape & Silbernagl (2005), Physiologie (5.Aufl.), S.837, Thieme-Verl.). V Erfassungsmethoden der Biologischen Psychologie 113 Spontan-EEG Das Spontan-EEG wird ohne spezielle Reizung abgeleitet und zeigt die dauernd bestehende Grundaktivität des Gehirns. Das Spontan-EEG zeigt rhythmische Potenzialänderungen mit Frequenzen zwischen 0,5 und 50 Hz, die Amplituden der Potentiale liegen zwischen 1 und 200 μV. Bestimmte Frequenzbänder (Frequenzbereiche) treten häufiger miteinander auf und lassen sich verschiedenen Aktivierungszuständen zuordnen. Im entspannten Wachzustand, besonders bei geschlossenen Augen, zeigt sich das synchronisierte Wach-EEG, bei dem die Alpha-Wellen (α-Wellen) im Frequenzbereich von 8 bis 13 Hz und einer Amplitude zwischen 5 und 100 μV dominieren. Die Beta-Wellen (β-Wellen) im Frequenzbereich von 14 bis 30 Hz und Amplituden zwischen 2 und 20 μV entstehen bei mentaler und körperlicher Aktivität. Gamma-Wellen (γ-Wellen, 30 bis 100 Hz, 2 bis 10 μV) zeigen anscheinend vor allem neuronale Aktivität bei der Repräsentation und Analyse von Wahrnehmungsobjekten an. Nur im Tiefschlaf finden sich Delta-Wellen (δ-Wellen; 0,5 bis 4 Hz, 20-200 μV). Und Theta-Wellen (θ-Wellen; 5 bis 7 Hz, 5 bis 100 μV) entstehen in der Entspannungsphase vor dem Einschlafen, aber auch bei Konzentration. Abb. V.3: Beispiele für die vier häufigsten Arten rhythmischer Aktivität im Spontan-EEG (aus: Schandry, 2006, S. 566). Für die Entstehung der α-Wellen wird die Schrittmacherfunktion des Thalamus verantwortlich gemacht, der seinerseits von der Formatio retikularis beeinflusst wird. β-Frequenzen werden auf intrakortikale Neuronennetzwerke zurückgeführt, θ-Wellen auf die Tätigkeit neuronaler Netzwerke mit Hippocampus-Anteilen. γWellen bilden exzitatorische und inhibitorische Verschaltungen im Kortex, auch über große Entfernungen, ab. Und δ-Wellen beruhen auf der maximalen Synchronisation großer Neuronenpopulationen. EEG-Frequenzbänder 114 Frequenzanalysen Biologische Grundlagen Ein Spontan-EEG kann durch verschiedene Frequenzanalysen ausgewertet werden. Im Frequenzhistogramm wird die prozentuale Häufigkeit des Auftretens der verschiedenen Frequenzbänder aufgetragen. Das Powerspektrum beruht auf der Amplitude der EEG-Frequenzen und macht Aussagen über die Verteilung der Amplitudenhöhen („power“ in Watt pro Hz) pro Frequenz oder Frequenzband. Die Frequenzverteilung im Spontan-EEG ist eine intraindividuell stabile Größe. Frequenzanalysen von EEGs werden genutzt, um Aussagen über die kortikale Aktiviertheit von Personen zu machen, wobei ein desynchronisiertes EEG für höhere Aktiviertheit spricht. In der Schlafforschung wird das EEG verwendet, um Aussagen über die Schlaftiefe beziehungsweise die verschiedenen Schlafphasen zu machen. Null-Linien-EEG Zeigt die EEG-Ableitung keine Aktivität mehr (Null-Linien-EEG) wird der Hirntod festgestellt, der als Kriterium für den Tod eines Menschen gilt. Elektroenzephalogramme mit Bezug zu Ereignissen Neben dem Spontan-EEG werden bei verschiedenen Ereignissen oder in Vorbereitung von Bewegungen bestimmte Veränderungen in den Potentialverläufen des Kortex festgestellt. Bereitschaftspotential Das Bereitschaftspotential, eine Verschiebung zu negativen Werten, entsteht ungefähr eine Sekunde vor der motorischen Aktivität über motorischen Kortexarealen und zeigt die Bewegungsplanung an. CNV Die kontingente negative Variation (contingent negative variation = CNV) ist eine langsame Potentialverschiebung zu negativen Werten. Sie wird in der Zeit beobachtet, wenn ein erster Reiz einen zweiten Reiz angekündigt hat, auf den eine Reaktion, nicht unbedingt motorischer Art, erfolgen soll. (Wie die rot-gelbKombination einer Ampel das baldige Aufleuchten des Grüns ankündigt). Diese Veränderung wird als Vorbereitung interpretiert: durch die Depolarisation wird die Auslösung von Aktionspotentialen begünstigt. evozierte Potentiale Bei ereigniskorrelierten oder evozierten Potentialen handelt es sich um typische Potentialverläufe, die in Folge von äußeren Ereignissen oder Reizen auftreten. Ungefähr 100 ms nach der Reizapplikation kommt es zu einer ersten Positivierung (P1) im EEG, gefolgt von einer ersten Negativierung (N1), danach je eine weitere Positivierung und Negativierung (P2 und N2) und schließlich die dritte und größte positive Auslenkung (P3 oder P300, da sie ungefähr 300 ms nach dem Reiz auftritt). Da die Amplituden des ereigniskorrelierten Potentials im Vergleich zum Spontan-EEG sehr gering sind, wird zur Darstellung ein Mittelungsverfahren verwendet, bei dem der Reiz mehrfach appliziert und die jeweils entstehenden Potentialverläufe rechnerisch übereinandergelegt werden. V Erfassungsmethoden der Biologischen Psychologie Ereigniskorrelierte Potentiale werden abgeleitet, um die Funktionsfähigkeit sensorischer Systeme zu überprüfen. Auch die Bedeutung von Reizen spiegelt sich in Verlauf und Amplitude der ereigniskorrelierten Potentiale wieder, wobei die ersten Komponenten exogen determinierte Eigenschaften des Reizes wie dessen Intensität, die späteren Komponenten (insbesondere die P300) endogen bestimmte Eigenschaften wie die emotionale Bewertung oder die zugeschriebene Bedeutung reflektieren. Abb. V.4: Akustisch evoziertes Potential. »R« markiert den Reizeinsatz. P1, N1, P2, N2, P3 sind die Bezeichnungen für die wichtigsten Komponenten (aus: Schandry, 2006, S. 568). Brainmapping Das Brain-Mapping ist eine Darstellungsmethode, in der die Aktivität des Gehirns in Bilder übersetzt wird. Dazu werden zum Beispiel die Amplituden abgeleiteter Potentiale von allen Elektroden zu einem bestimmten Zeitpunkt erfasst und auf eine Schema-Zeichnung eines Gehirns übertragen. Durch die Verbindung von Werten gleicher Höhe (ähnlich wie bei den Höhenlinien einer Landkarte oder den Isobaren einer Wetterkarte) und die Einfärbung ähnlich aktivierter Bereiche entsteht so ein Bild der verschiedenen Aktivierungsstufen des Kortex zum Beispiel bei einer Denkaufgabe. 115 116 Biologische Grundlagen V.1.2.2 Magnetenzephalographie (MEG) Die Magnetenzephalographie (MEG) erfasst neuronale Aktivitäten des Gehirns durch die Erfassung elektromagnetischer Feldlinien, die durch die Aktivität kortikaler Neuronen entstehen. Die Erfassung kann präziser als im EEG erfolgen, da die Ausbreitung von Magnetfeldern nicht durch die unterschiedlichen Gewebsarten des Gehirns beeinträchtigt wird, und die Sensoren nicht auf die Kopfhaut aufgesetzt werden, sondern sich in einem geringen Abstand von 10 bis 15 mm davon befinden. So wird durch die MEG eine räumliche Lokalisation aktiver Neuronengruppen mit einer Genauigkeit von 1 bis 2 mm möglich. Allerdings ist das elektromagnetische Signal sehr schwach. Es muss zur Erfassung hoch verstärkt und die Umgebung optimal gegenüber magnetischen Einflüssen abgeschirmt werden. Neben klinischen Einsatzbereichen wird die MEG verwendet, um die Verarbeitung somatosensorischer Reize zu studieren. Auch die Aktivität des Gehirns bei kognitiven Prozessen (Denken, Lernen, Aufmerksamkeit) kann abgebildet und die Plastizität des Gehirns erfasst werden. V.1.2.3 Positronenemissionstomographie (PET) Die Positronenemissiontomographie (PET) ist das erste bildgebende Verfahren, mit dem sich die Stoffwechselaktivität im Gehirn darstellen ließ. „Bildgebend“ heißt ähnlich wie beim Brainmapping, dass Gehirnaktivitätsunterschiede in eine Hirndarstellung mit unterschiedlichen Farben eingetragen werden. Jede Interpretation, es handele sich um eine Art von fotografischer Abbildung von Hirnaktivitäten ist also falsch. Das Färbungsprinzip entspricht damit demjenigen der Darstellung beispielsweise geologischer Verhältnisse einer Landkarte. Die PET erlaubt die räumliche Darstellung der Hirnaktivität hinsichtlich der Durchblutung und neurochemischer Prozesse, also der Aktivität der beteiligten Neurotransmitter und Rezeptoren. Die PET beruht auf der Erfassung der Energieemissionen beim Zerfall von Positronen, den Betaplusteilchen. Dazu werden zu Positronen zerfallende, radioaktive Substanzen in den Körper gebracht. Das ausgestrahlte Positron verbindet sich im Körper sofort mit den vorhandenen Elektronen und zerstrahlt in Form von zwei Gammaquanten, die in genau entgegengesetzte Richtungen emittiert werden. Diese als Vernichtungsstrahlen bezeichnete Gammastrahlung wird von ringförmig um den Kopf angebrachten Detektoren registriert, wenn zwei sich direkt gegenüberliegende Detektoren zur gleichen Zeit von Gammastrahlen getroffen werden. Ein Hochleistungsprogramm erlaubt den Ort des Zerfalls zu errechnen und ermöglicht die Berechnung eines Querschnittbildes (Gassen, 2008, S. 83). PET-Studien sind also mit einer Strahlenbelastung für die Untersuchungsperson verbunden. Mit der PET werden die Dichte bestimmter Rezeptorentypen in den verschiedenen Hirnarealen bestimmt und die Folgen von Erkrankungen (vor allem von Demenz V Erfassungsmethoden der Biologischen Psychologie oder Epilepsien) auf die Funktionen des Gehirns abgebildet. Auch die Veränderung der Durchblutung verschiedener Hirnareale im Zusammenhang mit Sprechen, Gedächtnisleistungen und anderen psychischen Funktionen wie Wahrnehmung oder Aufmerksamkeit werden mit Hilfe der PET erfasst. Abb. V.7: Messung von Durchblutung, Stoffwechsel und Morphologie des Gehirns bei einem Patienten mit Hirninfarkt. Durchblutung, O2-Verbrauch und Glucosestoffwechsel wurden mit Hilfe der Positronenemissionstomographie (PET) gemessen. Die Morphologie wurde anhand von Magnetresonanztomographie bestimmt. Das Gehirn ist in einer horizontalen Schnittebene dargestellt. Die Messwerte sind farbkodiert: niedrig = schwarz-blau-grün-gelb-rot = hoch. Die Sauerstoffextraktion entspricht dem Quotienten Sauerstoffverbrauch/ Sauerstofftransport. Die Funktionsparameter Hirndurchblutung, Sauerstoffverbrauch und Glucosestoffwechsel zeigen Ausfälle im Infarktbereich (links anterior). Die Morphologie ist hingegen im Infarktbereich in diesem Fall nicht sichtbar verändert. (Die Aufnahme wurde von Prof. W.-D. Heiss, Max-Planck-Institut für Neurologische Forschung, Köln, zur Verfügung gestellt.) (aus: Klinke, Pape & Silbernagl (2005), Physiologie (5.Aufl.), S.857, Thieme-Verl.). Ähnlich funktioniert die Single-Photon-Emissions-Computer-Tomographie (SPECT), für die allerdings handelsübliche und daher günstigere Markierungssubstanzen verwendet werden können. Auch mit der SPECT kann die regionale Hirndurchblutung bei kognitiven und psychischen Prozessen abgebildet werden. 117 118 Biologische Grundlagen V.1.2.4 Funktionelle Magnetresonanztomographie (fMRT) Die Magnetresonanztomographie (MRT) oder Kernspintomographie ist ein Verfahren, das ohne Strahlenbelastung, in frei wählbaren Schnittebenen und in hoher Auflösung Strukturen des Gehirns abbildet. Allerdings entwickeln sich bei der Methode starke Geräusche und die zu untersuchende Person befindet sich in einer Röhre. Abb. V.5: Schematische Darstellung vom Aufbau eines MRT. Der außen liegende Magnet erzeugt das statische und das Gradientenrohr das wechselnde Magnetfeld. Die Kopfspule erzeugt die Hochfrequenzpulse zur Anregung der ausgerichteten Atome (aus: Rose & Büchel (2008); in Gauggel & Herrmann (Hrsg.), S. 200, Hogrefe). Basis für diese Erfassungsmethode sind die magnetischen Momente bestimmter Atomkerne wie des Wasserstoffkerns, die im menschlichen Körper vorkommen. Durch das Anlegen eines Magnetfelds werden diese Dipole parallelisiert und kippen in ihre Ausgangslage zurück, wenn das Magnetfeld wieder abgeschaltet wird. Die dabei abgegebenen Radiowellen werden mit geeigneten Detektionsvorrichtungen erfasst. Hämodynamik Das Ziel funktioneller Verfahren ist es, Gehirnaktivität „bei der Arbeit“ zu erfassen. „Bei der Arbeit“ heißt, möglichst synchron zu bestimmten Arten psychischer Aktivität. Für die Darstellung momentan aktiver Hirnareale in der sogenannten funktionellen Magnetresonanztomographie (fMRT) macht man sich die Tatsache zunutze, dass der Sauerstoff- und Glukosebedarf in aktivierten Hirnregionen steigt. Sauerstoff wird über das Blut transportiert und sobald Neurone aktiver werden, brauchen sie mehr Sauerstoff und sind damit auf eine höhere Blutzufuhr angewiesen. Dabei ist zu bedenken, dass keine Nervenzelle weiter als ein tausendstel Millimeter von einem Blutgefäß entfernt ist (Gassen, 2008). Man misst also die Veränderung der Durchblutung (Hämodynamik) in lokal begrenzten Bereichen des Gehirns. Grundlage dafür ist die Chemie des Hämoglobins, das den Sauerstoff transportiert. Das Hämoglobin kommt in zwei Formen vor: einmal mit Sauerstoff als Oxyhämoglobin und einmal nach Abgabe des Sauerstoffs als Desoxyhämoglobin. Beide besitzen unterschiedliche magnetische Eigenschaften, so V Erfassungsmethoden der Biologischen Psychologie dass sie nach Menge und Ort voneinander unterschieden werden können. Bei zunehmender Aktivität des Gehirns geht die Konzentration des Desoxyhämoglobins in den Gefäßen zurück und dieser Konzentrationsabfall ist das, was mit fMRT gemessen wird. Dies führt zu einer leichten Erhöhung des MR-Signals. Dieses Signal ist also abhängig vom Niveau der Blutsauerstoffkonzentration und wird als BOLD-Signal (= Blood-Oxygene-Level-Dependent-Signal) bezeichnet. Mit dem BOLD-Signal erhält man gute räumliche und zeitliche Auflösungen. Die räumliche Auflösung liegt im Bereich von wenigen Millimetern, die zeitliche hängt von der Durchblutungsänderung ab, die bei ca. 2-5 s liegt. Man kann jedoch die zeitliche Auflösung erhöhen. Mit einer sogenannten „event-related“ fMRT – Messung ist es möglich, die Latenz zweier Aktivierungen im Bereich von 100 – 200 ms zu differenzieren. Dazu wird nach einem Trigger, z. B. durch einen akustischen Reiz, mittels wiederholter Messungen der genaue Anstieg des Signals bestimmt (vgl. Abb. V.6). Durch abwechselnde BOLD-Messungen der interessierenden Hirnregion während einer Experimental- und einer Kontrollbedingung (z. B. Stimulation vs. Ruhe) erhält man den Effekt der experimentellen Bedingung durch Subtraktion der Aktivierungmuster der Kontrollbedingung von demjenigen der Experimentalbedingung. Die signifikanten Differenzaktivierungen werden in ein Standardgehirn farbkodiert eingezeichnet. Die Signalerhöhungen sind jedoch meist sehr schwach und müssen mit relativ komplexen Filterungsverfahren aus dem Hintergrundrauschen herausgerechnet werden. Statistisch gesehen hat man das Problem, aus der Aktivitätsverteilung des Blutsauerstoffs eines Gehirns signifikante Teil-Verteilungen zu identifizieren. Ein Problem ist, wie man die Hämodynamik in einem Gehirn statistisch modellieren soll. Standardmäßig werden multivariate statistische Verfahren angewandt, wobei die theoretischen Annahmen, die dem multiplen abhängigen Testen zugrunde liegen, keineswegs selten klar expliziert werden. Zur weitreichenden Problematik der statistischen Modellierung des komplexen kybernetischen Systems „Gehirn“ sei auf den kritischen Artikel von Vul, Harris, Winkielman und Pashler (2009) mit dem bezeichnenden Titel „Voodoo correlations in social neuroscience“ hingewiesen. 119 BOLD-Signal event-related Problematik der statistischen Analyse von BOLD-Signalen 120 Biologische Grundlagen Abb. V.6: In einem Areal im visuellen Kortex erhaltene BOLD-Antwort (gestrichelte Linien zeigen 90% Konfidenzintervalle). Die dargestellte HRF erhält man, indem nach mehrfacher Reizdarbietung zu vielen unterschiedlichen Zeitpunkten das MR-Signal gemessen wird (Messzeitpunkte exemplarisch durch die Punkte veranschaulicht) (aus: Rose & Büchel (2008); in Gauggel & Herrmann (Hrsg.), S. 206, Hogrefe). Vorteile fMRT gegenüber PET Nachteile fMRT Mit dem fMRT kann man auch komplexere mentale Prozesse neuronalen Aktivierungen zuordnen, z. B. Arbeitsgedächtnis, Raumgedächtnis, sprachliche Prozesse usw. Gegenüber PET hat fMRT einige Vorteile. Durch die Magnetfelder entsteht nach heutigem Wissensstand keine schädliche Wirkung und die Messung kann recht oft wiederholt werden, z.B. um den Verlauf einer Rehabilitation nach einer Hirnläsion zu messen. Die Lokalisation mit fMRT im Bereich weniger Millimeter ist höher als mit der PET, bei der die Auflösung maximal 4 – 6 mm beträgt. Auch die zeitliche Auflösung ist mit einigen Sekunden höher als mit der PET, bei der die integrale Aktivität über 60 – 90 s bestimmt wird. Ein Nachteil der fMRT ist, dass Patienten mit Metallimplantaten oder Herzschrittmachern nicht untersucht werden können. Die jetzigen engeren Geräte können auch Platzangst verursachen. Die Untersuchungsbedingung in einer engen Röhre erlaubt nicht die Arbeit mit vor allem räumlich aufwendigeren Paradigmen oder Paradigmen, die komplexere Bewegungen als Knopfdrücken erforderlich machen. V Erfassungsmethoden der Biologischen Psychologie Auswertung und Studiendesigns (nach Rijntjes & Weiler, 2003, S. 8) Das Ergebnis einer fMRT-Messung ergibt pro Bildpunkt (Voxel = Volume element) eine Zeitreihe. Die Korrelation dieser gemessenen Zeitreihe mit einer aus dem Design abgeleiteten Zeitreihe wird statistisch untersucht. Voxel, die eine signifikante Aktivierung anzeigen, können farbkodiert auf ein anatomisches Hirnbild gelegt werden, um die Lokalisation zu bestimmen. Sowohl einzelne Individuen als auch Gruppen können untersucht werden. Für eine Gruppenstudie ist es jedoch noch erforderlich, die Gehirnbilder der einzelnen Probanden in einen Standardraum zu transformieren, damit die anatomisch unterschiedlichen Gehirne miteinander verglichen werden können. Es gibt verschiedene Möglichkeiten, eine Messung zu untersuchen, wobei wichtig ist, dass vorher die Kontrollbedingung festgelegt wird. Typische Versuchspläne von fMRT-Studien sind die folgenden: Kategorisch: Vergleich der Aktivierung gegen eine Ruhebedingung (z. B.: Was aktiviert Fingeropposition mehr als Ruhe?) Hierarchischer Vergleich einer Aktivierung gegen eine andere Aktivierung (z.B.: Was aktiviert das Lesen von Wörtern mehr als das Lesen von Buchstaben?) Faktoriell: Interaktion (z.B.: Was ist der Beitrag des Seheindrucks beim Lesen von Wörtern zur beobachteten Aktivierung?). Vergleich von Aktivierungen zwischen Gruppen (z. B. was aktiviert eine Gruppe von Patienten mit Halbseitenlähmung mehr als eine Gruppe von gesunden Probanden, wenn sie die rechte Hand bewegen?), d. h. Gruppe x Aufgabe – Interaktion. Haupteffekt (z. B.: Wo finden die Aktivierungen von Fingerbewegungen und Fußbewegungen gemeinsam statt?) Konjunktionsanalyse (z. B.: Was haben Fingerbewegungen und Fußbewegungen gemeinsam, unter Ausschluss der Interaktion?) Parametrisch: Korrelation (z. B.: Wie korreliert die Aktivierung mit der zunehmenden Geschwindigkeit der Wortpräsentation?) 121 122 Biologische Grundlagen Zu einigen Zielen von fMRT – Studien, zur Interpretation von Ergebnissen derselben und zu deren prinzipiellen Grenzen (die auch die Neurowissenschaft als Ganze betreffen) Zuordnung psychischer zu neuronalen Prozessen Ein wichtiges Ziel der funktionellen Bildgebung besteht nicht länger darin, eine spezifische Region im Gehirn zu lokalisieren, sondern die komplex verteilten neuronalen Netzwerke zu kartieren, die im gesunden Gehirn aktiv sind, wenn bestimmte mentale Aktivitäten stattfinden. Dennoch müssen die „Hirnbilder“ vorsichtig interpretiert werden. Wenn eine bestimmte Hirnregion signifikante BOLDSignale anzeigt, dann folgt daraus nicht, dass man diesem Ort eine bestimmte psychische Aktivität zuordnen kann. Wie oben dargelegt, sind diese BOLDSignale Differenzmaße. Das bedeutet, dass es sich um einen signifikanten Teil der Differenz von Aktivierungsverteilungen handelt. Daraus folgt nicht, dass die subtrahierten Teile für die in Rede stehende psychische Funktion bedeutungslos sind. Deswegen müssen psychische Aktivitäten unter verschiedenen Bedingungen untersucht werden, um eine einigermaßen reliable und valide Struktur-FunktionsZuordnung vornehmen zu können. Ein anderer Aspekt ist das Problem der Zuordnung eines psychischen Prozesse zu neuronalen Prozessen, genauer zu Aktivitätsänderungen. Sicherlich kann man nicht aus BOLD-Signalen ablesen, was genau für psychische Prozesse ablaufen und welchen semantischen Gehalt sie haben (Stichwort: Problematik der physiologischen Lügendetektoren). Vielmehr ist es umgekehrt. Ohne Kenntnis der mentalen Aufgaben, der Art und Weise, wie diese bearbeit werden, der Kenntnis, ob diese Aufgabenbearbeitung intra- und interindividuell gleich vorgenommen wird, ist es nicht möglich, psychische Prozesse neuronalen Aktivitätsmustern zuzuordnen. Wenn man eine Veränderung der Blutsauerstoffkonzentration in einem großen Muskel misst, dann weiß man auch nur, dass Teile des Muskels aktiver sind. Man weiß aber nicht, warum das so ist, wenn man nicht die Aktivität des gesamten Organismus berücksichtigt. Man muss dem Muster der Blutsauerstoffverteilung ein Bewegungsmuster zuordnen können, sonst kann man nichts über die funktionelle Relevanz der Aktivitätsänderung sagen. Ähnlich ist es mit dem BOLD-Signal im Gehirn. Man muss sicherstellen, dass man weiß, mit welcher psychischen Funktion das BOLD-Signal korreliert ist, ob es also überhaupt ein Signal ist. Hat man dies sichergestellt, dann bewegt man sich allerdings auf der Ebene der Korrelation und nicht der Kausalität. Was man von der psychischen Seite nicht weiß, wird einem die physiologische nicht mitteilen, es sei denn, man befragt die physiologische Seite. Das Gehirn gibt keine Antwort auf die Frage, was z. B. Denken ist. Es ist vielmehr so, dass hier keine Zuordnung von der Art einer isomorphen Punkt-zu-Punkt-Zuordnung möglich ist. Die Wahrnehmungs- und Handlungspsychologie gewinnt in enger Zusammenarbeit mit Neuropsychologie und Neurologie Kenntnisse über die neurophysiologischen Gesetzmäßigkeiten der Informationsverarbeitung. Es ist von einer nomischen Korrelation zwischen neuronalen und psychischen Zuständen auszugehen, wenn man mit Zuständen meint, dass es sich um Vorkommnisse in Zeit und Raum handelt. Fragwürdig ist allerdings, ob sich der semantische Gehalt V Erfassungsmethoden der Biologischen Psychologie wie „2 + 2 = 4“ von psychischen Zuständen ebenfalls nur als raum-zeitliches Vorkommnis verstehen lassen muss. Mit nomischer Korrelation ist gemeint, dass kein psychisches Vorkommnis ohne ein neuronales Vorkommnis auftritt. Es wird des Weiteren mit der Annahme nomischer Korrelation keine Annahme über das Kausalverhältnis zwischen neuronalen und psychischen Vorkommnissen getroffen, sondern vielmehr betont, dass beide gleichzeitig vorkommen wie beispielsweise Druck und Temperatur. Für Handeln und Verhalten sind also stets psychoneurale Paare relevant, die nomische Korrelation wird demzufolge nicht reduktiv zugunsten eines Paarlings kausal aufgelöst, weil dies in Aporien führt. Methodologisch ist jedoch die Situation die, dass der neuronale Paarling sich nur mit Hilfe des psychischen identifizieren lässt. Die ausschließliche Kenntnis der neuronalen Vorkommnisse erlaubt es nicht, mit Sicherheit auf die psychischen zu schließen. So müsste man eine Überzeugung wie „Glauben-dass-der-angebotene-Weinvergiftet ist“ zum Zeitpunkt t bei der Person P1 aus deren neuronalen Vorkommnissen 1 ,…, n „ablesen“ können, ohne dass der Beobachter P2 zum Zeitpunkt t weiß, dass P1 diese Überzeugung hat. De facto geht die Hirnforschung aber gerade so nicht vor. Andere Probleme sind empirisch ungeklärt, wie die Annahme, dass diese Überzeugung von P1 nicht nur zum Zeitpunkt t mit den neuronalen Vorkommnissen 1, …, n verpaart ist, sondern auch zu anderen Zeitpunkten, ganz zu schweigen, ob sich diese Paarung auch bei anderen Personen findet. Dabei war noch keine Rede davon, dass Überzeugungen stets mit anderen und dem allgemeinen Weltwissen verbunden sind, so dass es eigentlich um die nomische Korrelation nicht eines psychischen Vorkommnisses mit einem neuronalen, sondern um die zwischen einem semantischen und einem neuronalen Netz geht. An dieser Stelle wird deutlich, dass es sich um theoretische Probleme handelt, die durch Beobachtungsdaten mit Sicherheit keine vollständige Lösung erwarten lassen. Dies sollte keineswegs als Argument gegen die Hirnforschung verstanden werden. Vielmehr sollte man sich vor schlechter Philosophie hüten und hinsichtlich der unklaren Behauptung skeptisch sein, dass Mentales durch die Neurowissenschaften erklärt werden könnte (für Anspruchsvolle sei auf das Buch von Bennett & Hacker, 2003, verwiesen). V.1.2.5 Transkranielle Magnetstimulation (TMS) Anders als mit den bislang besprochenen Verfahren, wobei die Aktivität des Gehirns in Abhängigkeit von externen Reizen oder bestimmten Tätigkeiten abgebildet wurden, wird mit der transkraniellen Magnetstimulation (TMS) die kortikale Funktion durch ein von außen angelegtes Magnetfeld beeinflusst. Durch die nur 1 ms dauernde, erregende oder hemmende Stimulation können die zugeordneten Funktionen hervorgerufen oder verhindert werden. Durch die TMS kann damit die Lokalisation und Funktion auch kleinster Hirnareale beschrieben werden; von der Bewegung des kleinen Fingers durch die Stimulation des korrespondierenden motorischen Kortexareals bis zur Bewegungserkennung oder der Sprachproduktion. 123 nomische Korrelation psychoneurale Paare 124 Biologische Grundlagen V.2 Verfahren zur Erfassung peripherer Aktivität Eine Reihe von Biosignalen, die indikativ für psychische Funktionen und Reaktionen sind, können mit relativ geringem Aufwand peripher und nicht-invasiv abgeleitet werden. Eine große Rolle spielen dabei die Maße, die die Durchblutung des Körpers anzeigen. Mit dem Blut werden Sauerstoff und Energie, aber auch Hormone transportiert, daher sind diese Maße geeignet, die potentiellen oder tatsächlichen Leistungen des Körpers abzubilden. Andere Maße weisen auf motorische Aktivität oder die Erregung des vegetativen Nervensystems hin. V.2.1 Elektrokardiogramm (EKG) Mit dem Elektrokardiogramm (EKG) werden Spannungsveränderungen aufgezeichnet, die sich durch die Aktivität der Herzkammern ergeben und durch Elektroden erfasst werden. Die Elektroden werden auf die Haut aufgesetzt, wobei es standardisierte Ableitungen gibt, bei denen die Aufsatzorte festgelegt sind. Die abgeleiteten Spannungsänderungen zeigen einen typischen Verlauf, dessen Berge, Spitzen und Täler mit den Buchstaben P bis U bezeichnet werden. Abb. V.8: Elektrokardiogramm (EKG). Das EKG ist durch verschiedene typische Kurvenanteile (P- bis U-Welle) und die dazwischen liegenden Zeitstrecken gekennzeichnet. Die Wellen repräsentieren unterschiedliche Phasen im Aktionszyklus des Herzens (aus: Schandry, 2006, S. 588). V Erfassungsmethoden der Biologischen Psychologie Da die R-Zacke des EKG besonders gut zu identifizieren ist, wird sie zur Bestimmung der Herzfrequenz herangezogen, indem die Dauer des RR-Intervalls in die für die Herzfrequenz übliche Einheit „Schläge pro Minute“ umgerechnet wird. Andere Möglichkeiten zur Bestimmung der Herzfrequenz ist das wenig genaue Pulsfühlen oder die Erfassung der rhythmischen Änderungen des Blutvolumens zum Beispiel am Finger. Die Herzfrequenz ist über die Zeit hinweg variabel und stabilisiert sich bei aktivierenden Bedingungen. Damit wird die Herzfrequenzvariabilität ein gutes Maß für länger dauernde (tonische) Aktivierungen. Herzfrequenzänderungen, die in Folge eines Reizes auftreten, werden als phasische Herzfrequenzänderungen bezeichnet. Diese wird als Differenz der Herzfrequenz nach einem Reiz zu einer vorher erfassten Basisfrequenz bestimmt. Bei nahezu allen psychischen und physischen Anforderungen ändert sich die Herzfrequenz. Sie nimmt bei Schmerz- oder Angstreizen zu, bei Entspannung, der Orientierung auf nicht-aversive Reize oder Aufmerksamkeit nimmt sie ab. Eine systematische Veränderung der Herzfrequenz ist die respiratorische Sinusarrhythmie (RSA). Beim Einatmen schlägt das Herz schneller, beim Ausatmen reduziert sich die Herzfrequenz. Um die RSA in ihrem Einfluss auf die Herzfrequenz zu kontrollieren, ist es notwenig, parallel zum EKG auch die Atemfrequenz zu erfassen. Die respiratorische Sinusarrythmie wird durch den Nervus vagus verursacht: je stärker dieser aktiv ist, desto höher ist die Amplitude dieser Veränderung. So ist die respiratorische Sinusarrythmie im Schlaf oder bei tiefer Entspannung stark ausgeprägt, bei hoher Anspannung wenig. Die Amplitude der T-Welle des EKG (T-Wellen-Amplitude, TWA) verändert sich mit der Sympathikus-Aktivität. Sie nimmt bei Erhöhung der SympathikusAktivität ab und nimmt zu, reduziert sich dessen Aktivität. So kann die TWA als Indikator für mentale und emotionale Belastungen verwendet werden, vor allem bei kognitiven Anforderungen reagiert sie sensibler als zum Beispiel die Herzfrequenz. V.2.2 Blutdruckmessung Herkömmlicherweise wird der Blutdruck durch das Manschettendruck-Verfahren nach Riva-Rocci gemessen. Durch eine aufblasbare Manschette wird auf die Arterie im Oberarm Druck ausgeübt, der den Blutfluss unterdrückt. Der Druck wird kontinuierlich reduziert, bis zunächst stoßweise (Systole), dann kontinuierlich (Diastole) wieder Blut fließt. Die beiden zugeordneten Druckwerte [in mmHg] bilden den Blutdruck ab, wobei ein Blutdruck mit einem diastolischen Wert von 80 mmHG und einem systolischen Wert von 130 mmHg als Normblutdruck definiert ist. Da nach diesem Verfahren jedoch nur diskrete Blutdruckmessungen in gewissen Abständen möglich sind, wird alternativ die kontinuierlich mögliche 125 126 Biologische Grundlagen Erfassung des Blutdrucks in einem Blutgefäß zum Beispiel des Fingers verwendet. Der Blutdruck reagiert sehr schnell auf psychische Belastungen, wobei die Veränderungen auch stark ausfallen können. Abb. V.9 Indirekte Blutdruckmessung nach Riva-Rocci. Geräusche entstehen, wenn der Manschettendruck M den systolischen Blutdruck (PS) unterschreitet (B), und werden wieder leiser oder verschwinden, wenn der diastolische Blutdruck (PD) unterschritten wird (D) (aus: Klinke, Pape & Silbernagl (2005), Physiologie (5.Aufl.), S.186, Thieme-Verl.). V.2.3 Photopletysmographie Mit Hilfe der Photopletysmographie wird die periphere Durchblutung erfasst. Dazu werden eine Lichtquelle und ein Photosensor auf die Haut aufgebracht. In Abhängigkeit von der Durchblutung wird mehr oder weniger des Lichts aus der Lichtquelle reflektiert und mit dem Photosensor aufgenommen. Bezogen auf einen individuellen Basiswert lassen sich Aussagen über die periphere Durchblutung machen. Diese ist durch die Vasomotorik bestimmt und daher ein guter Indikator für die Sympathikusaktivierung, der die periphere Durchblutung reduziert. Da sich die Gefäße auch stark erweitern können, gilt die periphere Durchblutung als guter Entspannungsindikator. Die Photopletysmographie wird auch verwendet, um über die Durchblutung der Genitalien Aussagen zur sexuellen Erregung machen zu können. V Erfassungsmethoden der Biologischen Psychologie V.2.4 Erfassung elektrodermaler Aktivität Durch das Anlegen einer kleinen Spannung und der Erfassung des Stromflusses auf der Haut wird die Hautleitfähigkeit als Indikator der elektrodermalen Aktivität (EDA) abgeleitet. Die elektrodermale Aktivität beruht auf der Aktivität der Schweißdrüsen der Haut, die ausschließlich durch den Sympathikus erregt werden. Meist wird die EDA von der Handinnenfläche abgeleitet, deren Haut viele Schweißdrüsen enthält. Es werden verschiedene Maße der Hautleitfähigkeit verwendet, zunächst das Grundniveau. Dabei kommt es immer wieder zu kurzzeitigen Erhöhungen der Leitfähigkeit, den sogenannten Spontanfluktuationen, deren Zahl erfasst wird. Mit einer gewissen Latenz auf externe Reize folgen kurzfristige Erhöhungen der Leitfähigkeit (Elektrodermale Reaktionen) deren Latenzzeit und Amplitude erfasst werden können. Grundsätzlich gilt, dass bei einer Aktivierung Leitfähigkeit und Spontanfluktuationen zunehmen. Dies entspricht der Alltagsbeobachtung, dass in Angst- und Anspannungssituationen die Hände feucht, sogar schweißnass werden können. Abb. V.10 Veränderung der Hautleitfähigkeit auf einen Reiz. Es sind die wichtigsten Kenngrößen der Reaktion eingezeichnet: L = Latenz der Reaktion, A = Anstiegszeit, G = Gipfelzeit, E = Erholungszeit, H = Amplitudenhöhe (aus: Schandry, 2006, S. 591). 127 128 Biologische Grundlagen V.2.5 Elektromyogramm (EMG) Mit dem Elektromyogramm (EMG) wird der Spannungszustand von Muskeln bestimmt. Dazu werden Elektroden auf die Hautoberfläche aufgebracht, die die Muskelaktionspotenziale ableiten. Vor allem die Muskeln an Unterarm und Stirn sind gute Indikatoren des Spannungszustands jener. Das EMG wird daher auch verwendet, um dauerhafte Spannungszustände („Stress“) abzubilden. Beim Biofeedback wird das EMG genutzt, um Personen den Spannungszustand ihrer Muskulatur bewusst zu machen, z. B. über Töne, und sie in der Entspannung zu unterstützen. V.2.6 Elektrookulogramm (EOG) Das Elektrookulogramm (EOG) zeichnet Augenbewegungen auf. Dabei macht man sich die Tatsache zu nutze, dass zwischen Netzhaut und Hornhaut ein Polaritätsunterschied besteht und damit das Auge einen Dipol bildet. Bei der Bewegung der Augen können durch Elektroden, die in der Nähe der Augen auf die Haut geklebt werden, resultierende Spannungsverschiebungen und damit die Augenbewegungen erfasst werden. Diese Technik wird zum Beispiel angewendet, um das Blickverhalten beim Lesen zu erfassen. Abb. V.11: Prinzip der Elektrookulographie (EOG). Der Augapfel verhält sich wie ein Dipol. In der Ruhelage liegen die Pole symmetrisch zwischen den Elektroden, die links und rechts des Auges auf die Haut aufgeklebt sind. Es ergibt sich keine Spannung zwischen den Elektroden. Wandern die Augen zu einer Seite, wird entweder die linke Elektrode positiver (Blickbewegung nach links) oder die rechte (Blickbewegung nach rechts) (aus: Schandry, 2006, S. 592). V Erfassungsmethoden der Biologischen Psychologie V.3 Erfassung hormoneller Aktivität Der Vollständigkeit halber soll abschließend kurz auf die Bestimmung von Hormonen hingewiesen werden. Wie beschrieben, werden Hormone über den Blutkreislauf im Körper verteilt und damit zu den Effektororganen gebracht. Daher wird die Konzentration des jeweils interessierenden Hormons im Blut bestimmt (Hormonkonzentration im Blut). Dabei handelt es sich allerdings um ein invasives Vorgehen, das außerdem verschiedenen Einflüssen ausgesetzt ist. So ist die Hormonkonzentration im Blut außerordentlich gering, was deren Bestimmung fehleranfällig macht. Außerdem unterliegt die Hormonsekretion diversen Schwankungen, zum Beispiel durch den Tagesrhythmus bestimmt oder länger dauernden Zyklen wie dem Menstruationszyklus der Frau. Eine weitere, weniger störanfällige, dabei spezifische Methode nutzt Antikörper gegen Hormone zur Bestimmung, wobei jedoch radioaktiv markierte Hormone verwendet werden. Sehr störanfällig ist die Erfassung von Cortisol (als Stressindikator) im Speichel. Sie ist zwar nicht-invasiv und relativ einfach durchzuführen, doch unterschiedlichsten Störeinflüssen ausgesetzt. 129 130 Biologische Grundlagen Studientechnischer Anhang 1. Kenntnisse Methodik in der Biologischen Psychologie o Forschungsmethoden: Beobachtung spontaner Variation, willkürliche Variation physiologischer Parameter, willkürliche Variation psychischer Veränderungen o Probleme der Messung: Interindividuelle Variation, Ausgangslage, Reaktivität, Artefakte o invasive versus nicht-invasive Verfahren Verfahren zur Erfassung der Gehirnaktivität und deren jeweilige Grundlage o strukturabbildende Verfahren: Röntgen, CT o aktivitätsabbildende Verfahren: EEG (Spontan-EEG, Frequenzbänder, Evozierte Potentiale), MET, fMRT, PET, TMS Verfahrung zur Erfassung peripherer Aktivität und deren jeweilige Grundlage o EKG: Herzfrequenz, RR-Abstand, Herzfrequenzvariabilität, RSA, TWA o Blutdruck: Manschettendruckverfahren, Normblutdruck, Diastole, Systole o Photopletysmographie o EDA: Spontanfluktuationen, elektrodermale Reaktionen o EMG: Biofeedback o EOG Erfassung humoraler Aktivität o Hormonkonzentration im Blut o andere Verfahren V Erfassungsmethoden der Biologischen Psychologie 2. Literatur Grundlage: Schandry, R. (2006). Biologische Psychologie (2. überarb. Aufl.). Weinheim: Beltz. Kapitel 26: Methoden der Biologischen Psychologie (besonders Abschnitte 26.2 ff.) Vertiefend: Fehr, T. (2008). Magnetenzephalographie. In S. Gauggel & M. Herrmann (Hrsg.). Handbuch der Neuro- und Biopsychologie (S. 185-189). Göttingen: Hogrefe. Gassen, H. G. (2008). Das Gehirn. Darmstadt: WBG. Gauggel, S. Knops, A. & Städtgen, M. (2008). Transkranielle Magnetstimulation. In S. Gauggel & M. Herrmann (Hrsg.). Handbuch der Neuro- und Biopsychologie (S. 220-227). Göttingen: Hogrefe. Rijntjes, M. & Weiller, C. (2003). Funktionelle Bildgebung. In H.-O. Karnath & P.Thier (Hrsg.), Neuropsychologie (S. 7-8). Berlin etc.: Springer. Rose, M. & Büchel, Chr. (2008). Magnetresonanztomographie (MRT) und funktionelle Magnetresonanztomographie (fMRT). In S. Gauggel & M. Herrmann (Hrsg.). Handbuch der Neuro- und Biopsychologie (S. 198-210). Göttingen: Hogrefe. Rüsseler, J. & Münte, T. F. (2008). Elektroenzephalogramm (EEG) und ereigniskorrelierte Potenziale. In S. Gauggel & M. Herrmann (Hrsg.). Handbuch der Neuro- und Biopsychologie (S. 173-184 ). Göttingen: Hogrefe. Thiel, A. (2008). Positronenemissionstomographie (PET) und Single-PhotonEmissions-Computertomographie (SPECT). In S. Gauggel & M. Herrmann (Hrsg.). Handbuch der Neuro- und Biopsychologie (S. 190-197 ). Göttingen: Hogrefe. Vögele, C. (2008). Elektrodermale Aktivität. In S. Gauggel & M. Herrmann (Hrsg.). Handbuch der Neuro- und Biopsychologie (S. 157-163). Göttingen: Hogrefe. 131 132 Biologische Grundlagen Vögele, C. (2008). Kardiovaskuläre Aktivität. In S. Gauggel & M. Herrmann (Hrsg.). Handbuch der Neuro- und Biopsychologie (S. 164 - 172). Göttingen: Hogrefe. Voigt, K. (2005). Endokrines System. In R. Klinke, H.-Chr. Pape, H.-Chr. & S. Silbernagl, (2005). Physiologie (S. 509-559). Thieme: Stuttgart Vul, E., Harris C., Winkielman, P., & Pashler, H. (2009). Voodoo correlations in social neuroscience. Perspectives on Psychological Science (in press). (Download möglich unter: http://www.edvul.com/voodoocorr.php (30.01.2009) (Preprint); dort finden sich auch kritische Erwiderungen und weiterführende Diskussionen). Vertiefend für Anspruchsvolle, besonders philosophisch Interessierte: Bennett, M.R. & Hacker, P.M.S. (2003). Philosophical foundations of neuroscience. Oxford: Blackwell. VI Ausgewählte Funktionen VI Ausgewählte Funktionen Lernziele Die Studierenden sollen eine Übersicht über wichtige, psychologisch-biologische Funktionen erhalten, die Funktion und ihre biologisch-physiologischen Ursachen und Begleiterscheinungen sowie ihre Bedeutung für das menschliche Leben kennen lernen, Verständnis für den Zusammenhang zwischen psychologischen und biologischen Prozessen (Variablen) entwickeln, eine Grundlage für die vertiefende Befassung mit psychologischen Prozessen und Konstrukten erhalten. Im abschließenden Teil dieses Kurses werden nun einige biologischphysiologische Funktionen beschrieben. Diese Funktionen sind komplexer Art und betreffen eine Reihe von biologischen und psychologischen Subsystemen. Gemeinsam ist ihnen, dass ohne sie menschliches Leben nicht möglich ist. Nach einer kurzen Beschreibung der jeweiligen Funktion werden deren biologisch-physiologischen Korrelate erläutert und schließlich die Bedeutung für den Menschen diskutiert. Eine Reihe der hier besprochenen Funktionen werden im Laufe Ihres Studiums in anderen Kursen vertiefend behandelt. 133 134 Biologische Grundlagen VI.1 Circadiane Rhythmik, Wachen und Schlafen Ungefähr ein Drittel seines Lebens schläft der Mensch, in Kindheit und Jugend mehr, im Alter weniger. Schlaf- und Wachzeiten verteilen sich mehr oder weniger regelhaft über den 24 Stunden dauernden Tag. Der Schlaf-Wachrhythmus ist damit der deutlichste endogene Rhythmus, den der menschliche Organismus aufweist. Er trennt die Ruhephase des Schlafes mit deutlich reduziertem Bewusstsein von der Phase des Wachseins, in dem Menschen ihre Umwelt bewusst wahrnehmen (können). Circadiane Rhythmik Neben der Verteilung von Schlaf- und Wachzeit gibt es eine Reihe weiterer Funktionen und Parameter, die in einem 24stündigen Rhythmus variieren. Mit circadianer Rhythmik wird dieser Sachverhalt beschrieben. So sinkt die Körpertemperatur in den Abendstunden ab und erreicht am Nachmittag maximale Werte. Die Leistungsfähigkeit ist nicht den ganzen Tag gleich, sondern am Vormittag und nochmals am Nachmittag höher. Auch die Empfindlichkeit für Sinnesempfindungen und der Spiegel der verschiedenen Hormone variieren über den 24-StundenTag. Abb. VI.1: Tagesperiodische Schwankungen einiger Variablen. Zirkadiane Schwankungen der körperlichen Aktivität, der Körpertemperatur (rektal gemessen), der Rechengeschwindigkeit und der Zeitschätzung (aus: Schandry, 2006, S. 383). VI Ausgewählte Funktionen Abb. VI.2: Extremwerte verschiedener Phänomene innerhalb 24 Stunden (aus: Schandry, 2006, S. 384). Diese Rhythmik ist endogen gesteuert und stellt sich auch ohne äußere Taktgeber wie Tag-Nacht-Helligkeitsunterschiede oder Uhren ein, allerdings auf einen Rhythmus von ungefähr 25 Stunden Länge. Andererseits können exogene Bedingungen diese Rhythmik leicht verändern oder stören, wie jeder weiß, der wegen einer Feier über seine übliche Schlafenszeit hinaus munter bleibt. Auch ist eine Umstellung, wie sie zum Beispiel bei Schichtarbeit oder Reisen in andere Zeitzonen notwendig wird, schwierig und mit Beeinträchtigungen der Rhythmik der Aktiviertheit und damit auch der psychophysischen Leistungsfähigkeit verbunden. Vermittelt wird die circadiane Rhythmik über den Nucleus suprachiasmaticus des Hypothalamus, der über Kollaterale der Sehbahn Informationen über die HellDunkel-Verhältnisse erhält. Der Nucleus suprachiasmaticus wirkt als endogener Taktgeber und bewirkt über gepulste Freisetzungen von Hormonen und rhythmische Entladungen seiner Neuronen die Rhythmizität anderer Hirnstrukturen. Mittelbar wird auch die Zirbeldrüse zur Ausschüttung von Melatonin angeregt. Dieses Hormon verursacht verschiedene periphere Begleiterscheinungen des circadianen Rhythmus’. 135 136 Biologische Grundlagen Welche Bedeutung hat nun der circadiane Rhythmus? Zunächst einmal oszillieren alle lebenden Systeme um ihren Sollwert. Daher findet man auch im menschlichen Organismus eine Reihe unterschiedlicher Rhythmen, die sich überlagern. Neben dem circadianen existiert mit dem ein- bis zweistündigen Basic Rest Activity Cycle (BRAC) ein weiterer, stabiler Rhythmus verschiedener Vitalfunktionen, zum Beispiel des Bedürfnisses nach Nahrung oder Flüssigkeit, der Magenbewegungen oder des Harndrangs. Auch psychomotorische Leistungen scheinen diesem Rhythmus zu unterliegen. Der circadiane Rhythmus überlagert die verschiedenen Einzelrhythmen und stellt die Anpassung des Organismus an den 24-Stunden-Tag dar und legt die notwendigen Ruhezeiten in die Phase der Dunkelheit. Wachen Ein Mensch ist die meiste Zeit des Tages wach, das heißt, er reagiert auf Reize aus seinem Körper oder der Umwelt. (Im Folgenden wird „Reiz“ für alle Signale oder Informationen im oder zum menschlichen Organismus verwendet, die über die diversen Sinne (Exterozeption) und körperinneren Rezeptoren (Interozeption) zum ZNS geleitet werden). Er kann sich, sofern er dies möchte, seinen körperlichen Zustand bewusst machen und aktiv werden. Ob ein Reiz oder ein Ereignis bewusst wird, hängt von verschiedenen Umständen ab. Mit Aufmerksamkeit wird der Selektionsmechanismus beschrieben, mit dem aus der Vielzahl der Reize, mit denen der Mensch jeden Moment konfrontiert ist, der- oder diejenigen ausgewählt werden, die als wichtig oder relevant bewertet werden. Auch die Aufmerksamkeit ist nicht den ganzen Tag über auf gleich hohem Niveau, sondern schwankt tonisch über den Tag hinweg beziehungsweise phasisch auf bestimmte Anforderungssituationen hin. Bei der Erwartung eines Reizes wird Aufmerksamkeit top-down auf diesen gerichtet. Übersteigt ein unerwarteter Reiz eine bestimmte Intensitätsschwelle oder ist von vitaler Bedeutung für das Individuum, wird die Aufmerksamkeit bottom-up auf diesen gelenkt. An diesen Prozessen sind eine Reihe von Hirnstrukturen beteiligt. So hat die Formatio reticularis eine zentrale Bedeutung für den Wachzustand. Durch absteigende Bahnen zu den spinalen Motoneuronen hält sie die tonische Muskelaktivität aufrecht, die eines der Kennzeichen für den Wachzustand ist. Von der Formatio reticularis aufsteigende Bahnen (aufsteigendes retikuläres Aktivierungssystem = ARAS) erreicht sie fast alle kortikalen Hirnbereiche, insbesondere den Nucleus reticularis des Thalamus. Dieser wird mit der selektiven Aufmerksamkeitsfunktion in Verbindung gebracht, in dem er weitere Kerne des Thalamus aktiviert oder hemmt, während seine tonische Aktivität die allgemeine Aktivierungsfunktion bedingt. VI Ausgewählte Funktionen Es ist Aufgabe dieser Struktur, das für eine optimale Leistung notwendige, mittlere Aktivierungsniveau aufrechtzuerhalten und nur wichtige Reize an höhere Regionen weiter zu leiten (Gate-Funktion). Die Reiz-Bewertung erfolgt anhand von Zuflüssen aus dem präfrontalen und orbitofrontalen Kortex sowie dem limbischen System. Abb. VI.3: (a) Das aufsteigende retikuläre Aktivierungssystem ARAS. Links: Stark schematisierte Darstellung des aufsteigenden retikulären Aktivierungssystems im Affengehirn, ohne Berücksichtigung der genauen Verbindungen zwischen Hypothalamus, Thalamus und limbischem Kortex. Angedeutet die multisynaptischen retikulären Neurone und Kollateralen aus den spezifischen Bahnen (blau). Rechts: Stimulation vieler kortikaler Areale führt zu Potentialen in der Formatio, was eine kortikoretikuläre Verbindung und eine funktionelle Kontrolle der Aufmerksamkeitssteuerung nahelegt (aus: Schmidt & Schaible (Hrsg.) (2006). Neuro- und Sinnesphysiologie (5., neu bearb. Aufl.), S.385, mit Genehmigung von © Springer). (b) Erhöhter Blutfluss in der mesenzephalen Retikulärformation (FR), gemessen mit PET bei 10 Personen (gemittelt) während einer visuellen Aufmerksamkeitsaufgabe. Neben der Aktivierung in der FR auch Aktivierungen (rot, weiß, gelb) in visuellen Arealen (rechts) (printed with permission from Kinomura et al. (1996), Science, 271, p. 512, © 2009 AAAS). In den verschiedenen Funktionsmaßen des Gehirns lassen sich die Reizverarbeitungsprozesse erkennen. Während des Wachseins zeigt sich im EEG das β-Band, das eine weitestgehend desynchronisierte Aktivierung des Kortex indiziert. Vor einem (erwarteten) Reiz kommt es zu einer Depolarisation der beteiligten Hirnareale, die dann zu einer Positivierung mit γ-Aktivität im EEG während der Verarbeitung führt. In den ersten 100 ms nach dem Reiz findet die noch unbewusste, primäre Reizverarbeitung statt, die gegebenenfalls zu einer Bewusstwerdung des Reizes führen und damit eine bewusste Reaktion vorbereiten kann. 137 138 Biologische Grundlagen Im Wachzustand ist der Mensch in der Lage, Reize zu erkennen und darauf zu reagieren. Dafür wird eine gewisse Grundaktivierung benötigt, die mit gewissen Schwankungen über die ganze Wachzeit hinweg bereit steht. Um eine Reizüberflutung zu verhindern, die Reaktionen oder Handeln unmöglich machen würde, müssen Reize nach ihrer Bedeutung selegiert werden. Die Selektion erfolgt „unbewusst“ durch verschiedene Selektionsmechanismen oder auch aktiv, wenn Reize erwartet werden. Schlafen Während des Schlafs ist der Mensch nicht aktiv und reagiert nicht auf externe Reize, es sei denn, diese sind so stark, dass sie ihn aufwecken. Der Schlaf ist jedoch nicht gleichförmig, sondern lässt sich in verschiedene Stadien einteilen. Diese können durch verschiedene Parameter beschrieben werden, darunter die Stärke des Weckreizes, der notwendig ist, einen schlafenden Menschen aufzuwecken. Vor allem wird jedoch die Wellenaktivität im Elektroenzephalogramm (EEG) verwendet, um die verschiedenen Schlafstadien voneinander zu unterscheiden. Während im Wachzustand im EEG β-Wellen vorherrschen und im entspannten Wachzustand α-Wellen, verändert sich beim Übergang vom Wachen zum Schlafen das Bild. Im Schlafstadium 1 zeigen sich nur noch wenige α-Wellen, dafür aber niedrigamplitudige β- und θ-Wellen. Dieses Stadium lässt sich als Übergang vom Wachen zum Schlafen bezeichnen. Im Stadium 2 finden sich neben den θWellen im EEG auch so genannte Schlafspindeln, das sind kurze, hochfrequente Veränderungen, sowie plötzliche Veränderungen hoher Amplituden (KKomplexe). Die Stadien 3 und 4 sind die so genannten Tiefschlafstadien; wobei im Stadium 3 zwischen 20% und 50% der EEG-Aktivität aus δ-Wellen besteht, im Stadium 4 über 50%. Während des Schlafes kommt es wiederholt zu Phasen, in denen sich die Augen bei geschlossenen Lidern rasch hin und her bewegen (Rapid Eye Movements = REM). Die REM-Phasen haben Ähnlichkeit mit dem Schlafstadium 1, im Unterschied dazu ist jedoch die Weckschwelle hoch. Außer den Augenbewegungen ist der Muskeltonus deutlich herabgesetzt, wobei kleine Muskelzuckungen auftreten können. VI Ausgewählte Funktionen Abb. VI.4: EEG in verschiedenen Schlafstadien. Ausschnitte aus einem Schlaf-EEG, die typisch für verschiedene Schlafstadien sind (aus: Schandry, 2006, S. 398). Im Schlaf laufen die Schlafstadien in einer gewissen Regelhaftigkeit ab, wobei die Länge der einzelnen Schlafzyklen mit circa 90 Minuten den BRAC (Basic Rest Activity Cycle) widerspiegelt. Sowohl während des REM-Schlafes als auch während der anderen Schlafstadien kommt es zu Träumen, wobei bisweilen beobachtet wird, dass während des REM-Schlafes mehr geträumt wird. Träume werden als Versuch kortikaler Assoziationsfelder erklärt, inkohärente Informationen aus den verschiedenen Kanälen zu deuten. Beispielsweise werden motorische Impulse ausgesandt, die jedoch wegen des mangelnden Muskeltonus nicht umgesetzt werden. In einem Traum könnte diese Diskrepanz durch zum Beispiel eine unerklärliche Lähmung „erklärt“ werden. Schlafen und die Zeit des Schlafens werden sowohl durch das Schlafbedürfnis als auch die circadiane Rhythmik gesteuert. Bei Schlafentzug, vor allem bei ei- 139 140 Biologische Grundlagen nem Mangel an Tiefschlafphasen (Stadien 3 und 4) wird das Schlafbedürfnis immer stärker. Bereits bei der Besprechung der circadianen Rhythmik sowie des Wachzustandes wurde auf die Bedeutung der Formatio retikularis hingewiesen, die den Aktivitätszustand (Arousal) der Hirnrinde bedingt. Das ebenfalls bereits erwähnte, von der Zirbeldrüse ausgeschüttete Hormon Melatonin wirkt hingegen aktivitätssenkend und schlaffördernd. Die verschiedenen Schafstadien scheinen die Aktivität unterschiedlicher Hirnregionen abzubilden. Der REM-Schlaf wird durch Strukturen der Formatio retikularis „an“- und „ausgeschaltet“, während die langsamen Wellen des Tiefschlafs die Schrittmacherfunktion des Thalamus abbilden. Schlaf kann als Erholungs- und Reparaturphase interpretiert werden, in der zahlreiche, die Vitalfunktionen erhaltenden Prozesse wie Zellteilung oder Verdauung ablaufen. Auch kortikale Verarbeitungsprozesse und Konsolidierungsprozesse laufen während des Schlafes ab. Vor allem während der REM-Phasen scheinen sich gelernte Inhalte zu verfestigen, und besonders Inhalte des prozeduralen Wissens. Wird Schlaf entzogen, kommt es zu immer stärkerer Müdigkeit, zu Einbußen in der psychischen und physischen Leistungsfähigkeit und zu Schlafattacken („Sekundenschlaf“). Ein dauerhafter Schlafentzug führt zum Tod. VI.2 Hunger, Durst, Sättigung Hunger und Durst werden als Triebe bezeichnet. Sie bilden den Anreiz zu überlebensnotwendigen Verhaltensweisen, dem Trinken und dem Essen. Der Organismus ist bestrebt, zwischen Wasser- und Energieverbrauch des Körpers auf der einen Seite und der Aufnahme von Nahrung und Wasser auf der anderen Seite, ein Gleichgewicht zu erstellen, man spricht daher von homöostatischen Trieben. Durst Durst entsteht schon, wenn nur 0,5% des Körpergewichts an Wasser verloren wird. Über verschiedene Mechanismen wird der Wassermangel vermittelt. Der hypovolämische Durst wird über die Veränderung des Blutvolumens vermittelt. Über Barorezeptoren wird der Druckverlust über mehrere Stationen an den Hypothalamus gemeldet, der den Hypophysenhinterlappen zur Freisetzung von Vasopressin (=Adiuretisches Hormon ADH) anregt. Vasopressin erhöht die Rückresorption von Wasser in der Niere. Durch einen erhöhten Sympathikotonus sowie die Reduktion des arteriellen Drucks und des Blutflusses werden die Nieren außerdem angeregt, Renin auszuschütten. Dieses wird in mehreren Zwischenschritten in Angiotensin II verwandelt, das mittelbar den Nucleus medianus praeopticus des Hypothalamus stimuliert, der das Trinkverhalten einleitet. VI Ausgewählte Funktionen Auch der osmotische Durst führt zur Einleitung von Trinkverhalten durch den Nucleus medianus praeopticus. Er wird durch Osmosensoren ausgelöst, die auf die Erhöhung der intrazellulären Salzkonzentration im Bereich des Hypothalamus reagieren. Beim Trinken in Folge von Durst spricht man von primärem Trinken, das den Sollwert der Flüssigkeitskonzentration im Körper wieder herstellen soll. Da der Prozess der Aufnahme und der Verteilung der Flüssigkeit im Körper eine gewisse Zeit dauert, setzt beim Trinken schon vorwegnehmend eine Sättigung ein (antizipatorische Sättigung). Dies führt dazu, dass bereits deutlich vor der Erreichung des Sollwerts mit dem Trinken aufgehört wird. Das Trinken in Folge von Durst ist jedoch eher eine Ausnahme. In der Regel nimmt man vorsorglich ausreichend Flüssigkeit zu sich, sowohl in Form von Getränken als auch mit der Nahrung. Dieses sekundäre Trinken kann durch Geschmack und durch andere emotionale Faktoren sehr stark beeinflusst werden. Abb. VI.5 Hypovolämischer und osmotischer Durst bei Wassermangel (aus: Schmidt & Schaible (Hrsg.) (2006). Neuro- und Sinnesphysiologie (5., neu bearb. Aufl.), S.425, mit Genehmigung von © Springer). 141 142 Biologische Grundlagen Hunger Auch Hunger, also das Bedürfnis nach Nahrungsaufnahme, wird über verschiedene Mechanismen gesteuert. Über homöostatische Glukosesignale aus der Leber und Sättigungssignale aus dem Magen-Darm-Trakt, die den Nucleus tractus solitarius erreichen, wird die Kurzzeitregulation des Hungers vermittelt. Diese Signale sind zwar schnell, aber unpräzise. Über die Hormone Insulin und Leptin wird die Langzeitregulation des Hungers vermittelt. Sie sinken bei Nahrungsdeprivation. Leptin ist ein Hormon aus dem Fettgewebe, das sowohl bei langfristiger Entleerung der Fettspeicher als auch bei kurzfristiger Nahrungsdeprivation absinkt. Ein niedriger Insulinspiegel führt dazu, dass der Körper Energie konserviert. Die Hirnregionen, die den katabolen Gewichtsverlust (also den abbauenden Stoffwechsel) verursachen, werden gehemmt. Und letztendlich wird über die Aktivierung weiterer Hirnregionen die Nahrungsaufnahme eingeleitet. Der Nucleus tractus solitarius spielt auch bei der Verarbeitung von Sättigungssignalen aus der Leber sowie dem gastrointestinalen Trakt (Mund, Magen und Darm) über den Nervus vagus eine vermittelnden Rolle. Über efferente Verbindungen zum ventromedialen Hypothalamus wird bei Sättigung die Nahrungsaufnahme beendet. Abb. VI.6: Weg der Sättigungssignale. Neuroanatomische Verbindungen, über die Sättigungssignale aus der Leber und dem Darm (gastrointestinaler Trakt, GT) sowie Fettsättigung anzeigende Adipositassignale aus Pankreas und Fettgewebe mit den zentralnervösen Steuerstrukturen des autonomen Nervensystems (Nucleus tractus solitarius, NTS) zusammenwirken, um die Menge und Zusammensetzung der aufgenommenen Nahrung zu bestimmen. Abkürzungen: ARC, Nucleus arcuatus des Hypothalamus, LH, lateraler Hypothalamus; PFA, peri- und subfornikuläres Organ; POMC, Pro-opiomelanokortin; SNS, sympathisches Nervensystem (nach Schwartz et al., 2000). VI Ausgewählte Funktionen Ebenso wie beim Durst findet beim Hunger eine präresorptive Sättigung statt, das heißt, noch bevor die Sättigungssignale das Wiedererreichen des Sollwerts anzeigen, wird die Nahrungsaufnahme beendet. Dabei spielen auch mechanische Dehnungsrezeptoren in der Magenwand eine Rolle, die den Füllungszustand des Magens indizieren. Das Essverhalten wird jedoch nicht nur durch diese chemisch-physikalischen Signale des Körpers, sondern im hohen Maße auch durch emotionale und gelernte Einflüsse gesteuert. So kann der appetitliche Geruch von Speisen zur Nahrungsaufnahme anregen, auch wenn der Organismus satt ist. Hinsichtlich dessen, was appetitlich ist oder nicht, vielleicht sogar als eklig bezeichnet wird, gibt es interkulturelle Unterschiede, was auf Lernerfahrungen hinweist. Vermittelt werden diese über das mesolimbische Dopaminsystem. Zusammenfassung und Bedeutung Hunger und Durst bezeichnen Mangelzustände, den Bedarf an Nahrung beziehungsweise Flüssigkeit. Dabei werden für die Identifikation dieser Zustände mehrere Kanäle genutzt (hypovolämischer und osmotischer Durst beziehungsweise die Systeme Insulin und Leptin sowie Glukose). Auch die Wiederherstellung des gewünschten Sollwerts wird durch unterschiedliche Mechanismen erreicht, durch die Aufnahme von Nahrung und Flüssigkeit, aber auch durch Veränderungen im Stoffwechsel (Erhöhung der Rückresorption von Flüssigkeit durch die Niere, Hemmung des katabolen Gewichtsverlusts). Der Mechanismus der präresorptiven Sättigung ist nötig, um die Aufnahme von Flüssigkeit und Nahrung zu beenden, wenn ausreichend Menge zu sich genommen wurde und nicht zu warten, bis die biochemischen Sättigungssignale das Wiedererreichen des Sollwerts anzeigen. Diese basalen, das Überleben des Organismus sichernden Mechanismen sind stark durch emotionale und gelernte Faktoren überformt. So wird mit sekundärem Trinken die Tatsache bezeichnet, dass in der Regel auch ohne Durstgefühl ausreichend Flüssigkeit zu sich genommen wird. Man spricht zwar nicht vom sekundären Essen, doch sind durch die in unserem Kulturkreis üblichen, regelmäßigen „Mahlzeiten“ ähnliche Mechanismen anzunehmen. Geruch und Geschmack von Getränken und Speisen führen ebenfalls dazu, dass diese unter Umständen auch bei einem ausreichenden Sättigungszustand aufgenommen werden oder der Mechanismus der präresorptiven Sättigung ausgesetzt wird. Vermittelt wird dies über das limbische System (emotionale Bedeutung) beziehungsweise den Orbitofrontalkortex. In der „Überlernung“ des Hunger-Sättigungssystems wird eine der Ursachen für die verschiedenen Essverhaltensstörungen gesehen (Adipositas = Fettsucht, Anorexia = Magersucht). Im Rahmen der Klinischen und Gesundheitspsychologie 143 144 Biologische Grundlagen werden die Zusammenhänge zwischen vor allem Essverhalten und bestimmten psychischen Zuständen erforscht ebenso wie das Lernen schädlichen Essverhaltens als Basis für dessen Therapie. VI.3 Sexualität und Fortpflanzung Sexualität und Fortpflanzung sind Funktionen, die nicht das Überleben des Individuums sichern, sondern das Überleben einer Art. Sexualität ist ein Trieb, der sich daher nicht an einem Ideal- oder Sollwert ausrichtet (nicht-homöostatischer Trieb). Sexualität Mit dem Begriff Sexualität werden sowohl einfache, reflexhafte sexuelle Reaktionen als auch das komplexe, diese Reaktionen einschließende Sexualverhalten beschrieben. Sexualverhalten ist typischerweise ein soziales Verhalten zwischen zwei Menschen, das durch erlernte Verhaltensweisen zum Beispiel bei der Annäherung an einen möglichen Sexualpartner und individuelle Vorlieben zum Beispiel bei der Partnerwahl geprägt wird. Die Partnerwahl scheint außerdem durch Pheromone (also der von anderen abgegebenen Duft- oder Lockstoffen) beeinflusst zu werden. Das Interesse an sexueller Aktivität (sexuelle Appetenz oder Libido) kann erheblich variieren, sowohl zwischen Menschen (interindividuell) als auch über den Lebenslauf hinweg (intraindividuell). Der eigentliche Geschlechtsakt (Koitus) wird in vier Phasen eingeteilt: Erregungsphase, Plateau-Phase, Orgasmus und Entspannungsphase. Die Erregungsphase ist subjektiv durch Lustempfindung gekennzeichnet. Diese Erregung kann durch Vorstellungen, Wahrnehmungen oder sensorische Stimulation ausgelöst werden. Die Durchblutung der Sexualorgane erhöht sich, der Penis des Mannes erigiert und die Vagina befeuchtet sich (Lubrifikation). In der Plateau-Phase stabilisiert sich dieser Zustand; Herzfrequenz, Blutdruck, Atmung und Muskeltonus erhöhen sich. Kurz vor dem Höhepunkt wird beim Mann aus Drüsen in der Nähe der Prostata ein Sekret abgesondert, bei der Frau verengt sich die Vaginalöffnung zur orgiastischen Manschette. Die Orgasmusphase ist durch rhythmische Kontraktionen der Prostata und des Penis beziehungsweise der orgiastischen Manschette und der Gebärmutter gekennzeichnet. Die Kontraktionen führen beim Mann schließlich zur Ejakulation, nach der für einige Zeit (zwischen Minuten und Stunden) beim Mann kein weiterer Orgasmus möglich ist (Refraktärphase). Während der Entspannungsphase geht das Blutvolumen in den Genitalien zurück und die Aktivierung erreicht wieder normale Werte. VI Ausgewählte Funktionen Abb. VI.7 Bahnen der Kohabitationsreflexe bei Mann (links) und Frau (rechts). Die gestrichelten Bahnen des Rückenmarks deuten die Einflüsse von höheren Zentren auf das Erektionszentrum und das Ejakulationszentrum an. Genitale Stimulation, afferent zum Erektionszentrum geleitet, bewirkt durch efferente Impulse die Erektion. Dazu bedarf es auch bahnender Impulse, die von höheren Zentren stammen. Weitere Stimulation führt im Ejakulationszentrum zu einer Aktivierung efferenter Nerven und damit zur Ejakulation bzw. zum Orgasmus. Auch hierbei wirken hemmende oder bahnende Einflüsse aus höheren Zentren mit (aus: Klinke, Pape & Silbernagl (2005), Physiologie (5.Aufl.), S.568, Thieme-Verl.). Das Sexualverhalten wird wesentlich durch Sexualhormone gesteuert, die sowohl langfristige als auch kurzfristige Prozesse bedingen. Wie bei der Besprechung des Hormonsystems bereits erläutert ist, bewirken sie die Ausbildung der Geschlechtsorgane sowie die Produktion und Reifung der Gameten (Ei- und Samenzellen). Sexualhormone wirken direkt auf die Geschlechtsorgane, sind aber auch in den Nervenzellen des limbischen Systems und des Hypothalamus zu finden. Vor allem den Hormonen Östrogen und Testosteron wird eine wichtige Rolle bei der Entwicklung sexueller Appetenz zugeschrieben, die über die zentralen Strukturen vermittelt ist. Die körperlichen Veränderungen während der Erregungs- und Plateauphase des Geschlechtsakts werden durch den parasympathischen Ast des vegetativen Nervensystems vermittelt und über deszendierende Bahnen aus den zentralen Regionen als auch über parasympthische Afferenzen angeregt. Der Orgasmus hingegen ist durch den Sympathicus vermittelt und läuft reflektorisch über das Rückenmark ab. 145 146 Biologische Grundlagen Bedeutung der Sexualität Die sexuelle Fortpflanzung mit Hilfe der Aufteilung zweier Gametentypen auf zwei räumlich unverbundene Organismen erhöht im Vergleich zu Organismen, die beide Gametentypen haben, die Wahrscheinlichkeit von Veränderungen (Mutationen) im genetischen Material und bescheunigt damit die Selektion günstiger Mutationen (vgl. Kapitel Genetik und Evolution). Die Aufteilung zweier Gamatentypen auf zwei Organismentypen führte in der großen Mehrzahl zum Sexualdimorphismus. Damit ist gemeint, dass sich die beiden Organismentypen in ihrem Erscheinungsbild in der Regel unterscheiden. Damit ist nicht (nur) das unterschiedliche Aussehen der Geschlechtsorgane gemeint, sondern Körperbau und –größe, Behaarung, Färbung (z. B. bei Vögeln) oder es finden sich die äußere Erscheinung unterscheidende besondere Körperteile wie z. B. das Geweih bei Hirschen. Bei Menschen ist der Sexualdimorphismus Ausgangspunkt und Kristallisationspunkt einer komplexen Kulturbildung. Die Existenz zweier Geschlechter war wohl schon früh Gegenstand des Nachdenkens von Menschen, wie sich an diversen Ursprungsmythen zeigen lässt. Der Mensch ist das einzige Wesen mit einer komplexen Kultur der Kleidung, die auch eine besondere Rolle bei der Markierung sexueller Differenzen und Interessen spielt. Theorien zur sexuellen Selektion und zu den Geschlechtsunterschieden finden sich in Kultur- und Naturwissenschaften, vor allem in der Psychologie, der Evolutionstheorie und der Ethologie. Empfehlenswert zur Psychologie der Geschlechtsunterschiede ist die Arbeit von Bischof-Köhler (2006). Neben diesem evolutionsbiologischen Vorteil hat Sexualität eine hohe emotionale und soziale Bedeutung. Die Verbindung von Sexualität beziehungsweise Sexualverhalten mit positiven Gefühlen begünstigt und verstärkt die Bindung zwischen Partnern, kann aber auch zu negativen Gefühlen (Eifersucht, Scham) führen. Damit wird das Zusammenleben von Menschen in Familien und Gruppen unterstützt, vielleicht sogar aufgebaut. Die soziale Komponente der Sexualität zeigt sich weiterhin darin, dass dieser Bereich wesentlich stärker als beispielsweise die homöostatischen Triebe Hunger und Durst sozial geregelt und reglementiert ist (z. B. Inzesttabu). Schwangerschaft und Geburt (Fortpflanzung) Der weibliche Zyklus mit dem Eisprung und Aufbau von Gebärmutterschleimhaut schafft eine optimale Voraussetzung für eine Schwangerschaft. Nach dem Eisprung gelangt die Eizelle in den Eileiter, wo sie für circa 12 Stunden verbleibt. In diesem Zeitraum ist eine Befruchtung möglich. Da Spermien im Milieu von Scheide und Gebärmutter für ungefähr drei Tage befruchtungsfähig bleiben, ergibt sich ein Zeitraum von vier bis fünf fruchtbaren Tagen, während derer eine Schwangerschaft entstehen kann. VI Ausgewählte Funktionen Mit der Verschmelzung von Ei- und Samenzelle entsteht die Zygote, die sofort mit der Zellteilung beginnt, während sie zur Gebärmutter transportiert wird. In diesem Stadium wird der Keim Morula genannt, der sich durch Wassereinlagerung zur Blastozyte wandelt, die sich sechs bis sieben Tage nach der Befruchtung in die Gebärmutterschleimhaut einnistet (Nidation). Bereits einige Tage nach der Befruchtung beginnt die Produktion schwangerschaftsbegleitender Hormone durch Nachbarzellen der Zygote und die Plazenta. Letztere unterstützen die Ernährung des Ungeborenen. Auch die Einleitung der Geburt ist im Wesentlichen hormonell ausgelöst. Begleitet durch Bewegungs- und Druckreize, die vom Embryo ausgehen, beginnt die Geburt mit der Ausschüttung von Oxytocin und Prostaglandinen. Die Geburt beginnt mit einer Eröffnungsperiode, die durch regelmäßige Wehen gekennzeichnet ist und in deren Verlauf die Fruchtblase platzt. Mit Austreibungsperiode bezeichnet man den Zeitraum von der vollständigen Öffnung des Muttermundes bis zur Geburt. Nach der Durchtrennung der Nabelschnur wird das Neugeborene durch die nun fehlende Sauerstoffversorgung angeregt, selbstständig zu atmen. In der Nachgeburtsperiode löst sich schließlich die Plazenta und wird abgestoßen. Der Körper der Mutter beginnt mit der Produktion der Milch. Wird das Kind gestillt, steigt der Prolaktinspiegel der Mutter dauerhaft an. Bedeutung von Schwangerschaft und Geburt Eine Schwangerschaft bietet dem werdenden Menschen eine optimale Umgebung, in der er sich bis zu einer eigenständigen Lebensfähigkeit entwickeln kann. Die körperlichen Begleiterscheinungen werden im Wesentlichen hormonell vermittelt und gehen mit psychischen Veränderungen bei der werdenden Mutter einher. Neben positiven Gefühlen der Freude können auch Angst und Traurigkeit bis hin zur Depression auftreten, vor allem mit der relativ plötzlichen Hormonumstellung während der Geburt. Schwangerschaft, Geburt und eine sich gegebenenfalls anschließende Stillzeit sind damit ein wesentliches Lebensereignis für Frauen und eine der Grundlagen für die Bindung beziehungsweise die Beziehung zwischen Mutter und Kind. 147 148 Biologische Grundlagen VI.4 Reifung und Altern Unter Reifung versteht man alle Veränderungen des Körpers, des Verhaltens und der Leistungsfähigkeit, die keimhaft angelegt sind, das heißt durch das Genom bestimmt und genetisch verursacht werden. (Biologische) Reifungsprozesse finden während des gesamten Lebens eines Menschen statt, natürlich vor allem in der Zeit vor und in den Jahren nach der Geburt. Aber auch das Altern mit seinen typischen Veränderungen ist genetisch programmiert. Reifungsprozesse laufen damit quasi automatisch ab, werden jedoch in unterschiedlichem Umfang durch die soziale und physische Umwelt beeinflusst. So sind ausreichende Ernährung, Zuwendung und eine anregende Umgebung notwendig, um die körperliche und geistige Reifung stattfinden zu lassen. Auch die vorgegebene Alterung kann durch ungesunde Lebensweisen beschleunigt werden. Unter dem Begriff der Entwicklung werden alle Veränderungen der psychophysischen Struktur und des Verhaltens, Handelns und Erlebens von Menschen über die Lebensspanne hinweg betrachtet. Neben den Reifungsprozessen gehören hierzu die sozialen Prozesse der Individualisierung (des Sich-selbst-bewußt-werdens, der Selbstgenese) sowie der Übernahme von Verantwortung für sich selbst und für andere, das Lernen und das Sammeln und Verwerten von Erfahrungen. Vorgeburtliche Entwicklung Bereits mit der Verschmelzung von Ei- und Samenzelle ist das Genom des Keims, also die genetische Ausstattung mit Merkmalen wie Geschlecht oder Blutgruppe, festgelegt. Noch nach den ersten Zellteilungen sind alle Zellen omnipotent, das heißt, aus ihnen kann sich prinzipiell jede Struktur und Funktion entwickeln (embryonale Stammzellen). Deutlich wird dies, wenn sich durch eine Zellteilung zu diesem frühen Zeitpunkt eigenständige Lebewesen mit gleichem Genom entwickeln (eineiige Zwillinge). Mit den weiteren Zellteilungen entsteht die genetisch festgelegte zunehmende Differenzierung von Strukturen und Funktionen der Zellen und Zellgruppen hin zu verschiedenen Organen und Gewebstypen. Die Differenzierung verläuft nicht gleichmäßig, sondern in einer bestimmten Abfolge. Kommt es in diesen sensiblen Phasen der Reifung zu Störungen z.B. durch Gifte, resultieren jeweils spezifische Ausfälle. Ab der 10.Schwangerschaftswoche bilden sich die bis dahin unspezifischen Gonaden (Keimdrüsen) unter dem Einfluss von Testosteron bei männlichen Embryonen zu Hoden, die ihrerseits wieder Testosteron abgeben. Bei weiblichen Embryonen entwickeln sich die Ovarien. VI Ausgewählte Funktionen Ebenfalls in der 10. Schwangerschaftswoche beginnt die Gehirndifferenzierung bis in der 28. Woche die endgültige Zahl an Neuronen im Gehirn, allerdings noch nicht das Ende der Hirnentwicklung erreicht wird. Die Myelinisierung, also der Prozess der Ausbildung von Markscheiden um Nervenzellen, beginnt in der 24. Schwangerschaftswoche, um sich dann bis in 6., eventuell sogar bis ins 15. Lebensjahr hinweg fortzusetzen. Auch die Gestalt des Ungeborenen verändert sich. Bis zum 3. Monat der Schwangerschaft ist der Kopf größer als der Rumpf; bis zu dieser Zeit spricht man vom Embryo. Zum Zeitpunkt der Geburt circa 38 Wochen nach der Befruchtung ist der Rumpf dreimal so lang wie der Kopf. In der Zeit vom 4. bis zum 9. Monat spricht man vom Ungeborenen als Fetus. Kindheit Die Kindheit zwischen Geburt bis hin zur Pubertät ist durch Reifungsprozesse wesentlich geprägt. Die weitere Ausdifferenzierung des Gehirns und die zunehmende Myelinisierung der Neuronen bildet die Grundlage für die Entwicklung von einem sich unbeholfen bewegenden Neugeborenen, das sich nur wenig differenziert äußern kann, hin zu einem sprach- und denkfähigen, sich koordiniert bewegenden Schulkind. Abb. VI.8 Entwicklungsneuropsychologie funktioneller Systeme (aus: Gauggel & Herrmann (Hrsg.) (2008), S. 310, Hogrefe). 149 150 Biologische Grundlagen Auch die äußere Gestalt verändert sich, wobei sich die Länge vergrößert und sich die Proportionen von Kopf, Rumpf und Gliedmaßen jeweils zueinander verschieben sich. Mit ersten Gestaltwandel bezeichnet man die recht deutliche Veränderung vom rundlichen Kleinkind hin zum schlanken Schulkind um das 6. Lebensjahr herum. Wesentlich für das Längenwachstum verantwortlich ist das Wachstumshormon (Somatotropes Hormon = STH). Pubertät Etwa ab dem 7. bis 10. Lebensjahr bei Mädchen, bei Jungen etwas später, werden über die Hypothalamus-Hypophysenachse gonadotrope Hormone freigesetzt und damit die Pubertät ausgelöst. Es entwickeln sich die sekundären Geschlechtsmerkmale: Schambehaarung, bei den Mädchen die Brüste, bei Jungen Bartwuchs und tiefe Stimme (Stimmbruch). Die Proportionen verändern sich erneut. Zum abschließenden, durch STH stimulierten Längenwachstum tritt die geschlechtsspezifische Formung mit breiteren Hüften bei Mädchen und breiteren Schultern bei Jungen (zweiter Gestaltwandel). Die Keimdrüsen beginnen mit der Bildung von Spermien beziehungsweise der Ausreifung von Eizellen. Dieser genetisch programmierte Ablauf hängt jedoch vom Ernährungszustand beziehungsweise Gewicht der Kinder beziehungsweise Jugendlichen ab. Erwachsenenalter Zwischen dem 20. und 30. Lebensjahr erreichen Menschen den Gipfel ihrer körperlichen und geistigen Leistungsfähigkeit, damit sind die Reifungsprozesse abgeschlossen. Danach folgt eine Zeit der relativen Stabilität. Doch auch in dieser Zeit verändert der Organismus ständig. Zellen sterben ab und werden von neuen, durch Zellteilung entstandenen Zellen ersetzt. Zwei Mechanismen des Zelltods können unterschieden werden: Nekrose und Apoptose. Bei der Nekrose sterben die Zellen ab, weil die Zufuhr an Sauerstoff, Glukose oder Blutversorgung gestört oder unterbrochen ist. Die Apoptose kann als programmierter Zelltod verstanden werden, bei der sich eine Zelle durch Freisetzung bestimmter Stoffe selbst auslöscht, ohne dass die Gefahr besteht, Entzündungen auszulösen. Diese ständigen Veränderungen dienen der Anpassung an externe Gegebenheiten und werden durch Instandhaltungs- beziehungsweise Regulierungsvorgänge des Körpers überwacht. Beispielsweise erkennen und reparieren so genannte Caretaker-Gene DNA-Defekte, die während der Zellteilung entstanden sind. VI Ausgewählte Funktionen Alter Bereits mit der Beendigung der Reifung beginnen die verschiedenen Alterungsprozesse, die schließlich beim alten Menschen zu Altersschwäche und zum Tod führen. Altern kann als Störung der Homöostase der oben genannten Regulierungsprozesse verstanden werden. Die Zellteilungen erfolgen nicht mehr in dem benötigten Umfang, da bei manchen Zelltypen die Zahl der möglichen Zellteilungszyklen beschränkt bleibt. Auch nimmt die Zahl schadhafter Mutationen und damit schädlicher Stoffe (z. B. freie Radikale) im Organismus zu. Altern führt damit zu einer Reduktion vieler Körperfunktionen und einer verminderten Widerstandsfähigkeit gegenüber Umweltanforderungen wie Infektionen oder Umweltgiften. Die Muskel- und Knochenmasse nehmen zugunsten von Körperfett ab. Abb. VI.9: Altern. (A) Im Alter nehmen zahlreiche Körperfunktionen ab, neben den im Blut gezeigten auch die glomeruläre Filtrationsrate, die maximale O2-Aufnahme u.a.m. (B) Kultiviert man Lungenfibroblasten von Neugeborenen und von 100-jährigen Probanden, so endet die Teilungsfähigkeit der Zellen bei letzteren früher als bei den Kindern. Noch wesentlich früher endet die Teilungsfähigkeit der Zellen von Patienten mit Werner-Syndrom, bei denen das Gen mutiert ist, das für die DNA-Helicase kodiert. Sie altern vorzeitig: Progeria adultorum (aus: Klinke, Pape & Silbernagl (2005), Physiologie (5.Aufl.), S.46, Thieme-Verl.). Ein deutlich erkennbarer Einschnitt ist bei Frauen das Klimakterium. Um das 50. Lebensjahr herum reduzieren die Ovarien die Follikelreifung bis hin zur vollständigen Aufgabe. Damit wird die Produktion von Östrogenen reduziert beziehungsweise eingestellt, was zu einer Reihe weiterer körperlicher Veränderungen führt. Die Reduktion der Testosteronproduktion beim Mann verläuft langsamer und beginnt meist erst um das 55. Lebensjahr. 151 152 Biologische Grundlagen Im hohen Alter nehmen bei ansonsten bestehender Gesundheit unter anderem die Muskelkraft und die Sauerstoffaufnahme deutlich ab, die Reflexe verlangsamen sich und der Gleichgewichtssinn ist gestört. Es kommt zu einem unsicheren Gang und der Möglichkeit von Stürzen, die aufgrund der geringeren Knochendichte dann auch häufiger zu Knochenbrüchen führen. Man geht davon aus, dass die maximal mögliche Lebenszeit eines Menschen genetisch determiniert ist und ungefähr 115 Jahre beträgt. Die mittlere Lebenserwartung liegt in den westlichen Industrienationen bei 80 bis 85 Jahren und damit noch deutlich unter diesem Maximum. Hier spielen die jeweiligen Lebensumstände und Lebensweisen eine Rolle. VI.5 Stress und Immunabwehr Wie bereits mehrfach ausgeführt, befindet sich der Organismus des Menschen in einem steten Austausch mit seiner Umwelt. Die Umwelt stellt alles bereit, was der Organismus an lebenserhaltenden und förderlichen Substanzen und Anregungen benötigt, beginnend von der Luft zum Atmen, über Nahrung bis hin zu sozialen und emotionalen Anregungen aus der Beziehung zu Eltern und anderen Menschen. Die Umwelt ist jedoch nicht nur Quelle förderlicher, sondern auch schädlicher Reize. Im folgenden Abschnitt werden nun Reaktionsweisen beschrieben, mit denen der Organismus auf negative Einflüsse reagiert. Beide Reaktionsweisen stehen, obwohl auf unterschiedlichen Ebenen angesiedelt, in einem gewissen Zusammenhang und sollen daher in einem Abschnitt beschrieben werden, zunächst das basale System der Immunabwehr. Immunsystem Das Immunsystem schützt den Körper vor schädlichen Substanzen, die aus der Umwelt, aber auch aus dem eigenen Körper stammen können. Dabei werden unspezifische und spezifische Arten der Abwehr unterschieden. Die genetisch angelegte, unspezifische Immunabwehr wird zunächst durch physikalisch-chemische Barrieren vermittelt. Haut und Schleimhäute bilden Barrieren, die das Eindringen fremder, möglicherweise schädlicher Stoffe verhindern. Dabei wirken mechanische, aber auch chemische und physikalische Mechanismen protektiv, wie der Säuremantel der Haut oder die Fieber- beziehungsweise Entzündungsreaktion, durch die bereits eingedrungene Mikroorganismen geschädigt werden. Weitere Formen der unspezifischen Immunabwehr basieren auf der Aktivität bestimmter Zellen, der Makrophagen. Diese nehmen eingedrungene Erreger in sich auf („fressen“ sie quasi) und bauen sie dadurch ab. Durch chemische VI Ausgewählte Funktionen Komplementsysteme werden fremde Zellmembranen zerstört und die Zellen letztendlich aufgelöst. Und schließlich werden durch verschiedene körpereigene Substanzen fremde Mikroorganismen zerstört, zum Beispiel virusinfizierte Zellen durch Interferone. Abb. VI.10: Phagozytose durch neutrophile Granulozyten oder Makrophagen. Phase 1: Der mit Antikörpern (z.B. IgG) oder dem Komplementärfaktor C3b bedeckte (opsonisierte) Fremdkörper ist durch die entsprechenden Rezeptoren der Phagozyten (Fc- und C3b-Rezeptoren) als etwas Fremdes „gekennzeichnet“. Phase 2: Nach Kontaktaufnahme mit dem Fremdkörper bilden die Phagozyten Pseudopodien, mit denen sie den Fremdkörper „umarmen“. Phase 3: Nach vollständiger Aufnahme des Fremdkörpers (Phagozytose im eigentlichen Sinn) kommt es zur Bildung von Phagosomen. Phase 4: Die hydrolasenreichen Lysosomen verschmelzen mit Phagosomen und bilden Phagolysosomen, in denen der Fremdkörper verdaut wird. Phase 5: Unverdaubares Material wird nach außen abgegeben; auf der Zelloberfläche erscheinen wieder die Fc- und C3bRezeptoren, die vor der Bildung der Phagosomen abgespalten worden sind (Recycling) (aus: Klinke, Pape & Silbernagl (2005), Physiologie (5.Aufl.), S.237, Thieme-Verl.). Um fremde von den eigenen Zellen unterscheiden zu können, bedient sich der Organismus bestimmter Proteine, die fast jede Zelle auf ihrer Oberfläche trägt, die MHC-Proteine (MHC = major histocompatibility complex). Die Spezifizierung dieser Proteine ist genetisch festgelegt und für das jeweilige Individuum einzigartig (mit Ausnahme z.B. eineiiger Zwillinge, die über dieselbe genetische Ausstattung verfügen). 153 154 Biologische Grundlagen Die spezifische Immunabwehr wird im Gegensatz zu der unspezifischen Abwehr erst nach Erfahrungen mit bestimmten schädlichen Substanzen aufgebaut, sozusagen „erlernt“. Gelangt ein Organismus mit einer Substanz in Kontakt, kann sich eine Art des Gedächtnisses bilden, das bei der erneuten Kontaktierung mit dieser (oder hoch ähnlichen), dann Antigen genannten Substanz zu einer schnelleren und damit effektiveren Abwehrreaktion führt. Diesen Mechanismus nutzt man beispielsweise bei Schutzimpfungen. Abb. VI.11 Klonale Selektion und Differenzierung von B-Lymphozyten. Es sind drei verschiedene Typen von Lymphozyten dargestellt, die durch jeweils einen spezifischen IgG-Rezeptor (Paratop) gekennzeichnet sind (Zellklone 1, 2, 3). Nur der Zellklon 2 hat den zu dem antigenen Epitop passenden Rezeptor. Diese spezifische Merkmalserkennung führt zur klonalen Selektion mit nachfolgender Vermehrung des Zellklons 2 (klonale Expansion). Die nachfolgende Differenzierung des expandierten Klons bewirkt die Bildung von Antikörper bildenden Plasmazellen und von B-Gedächtniszellen. Die Plasmazellen sezernieren Immunglobuline mit einem dem B-ZellRezeptor identischen Paratop (s. Ausschnittsvergrößerung des sezernierten Immunglobulins). Die B-Gedächtniszellen speichern die Information über den stattgefundenen Antigen-AntikörperKontakt, so dass bei einer zweiten Exposition mit diesem Antigen eine schnellere und verstärkte Antikörperbildung stattfindet (aus: Klinke, Pape & Silbernagl (2005), Physiologie (5.Aufl.), S.239, Thieme-Verl.). Für die Vernichtung schädlicher Substanzen wie Viren, virusinfizierter Zellen oder auch Tumorzellen sind bei der spezifischen Immunabwehr verschiedene immunkompetente Zellen wie B-Lymphozyten, T-Lymphozyten und natürliche Killerzellen verantwortlich. Lymphozyten werden im Knochenmark gebildet und „warten“ in so genannten sekundären Organen wie den Lymphknoten oder den Rachen- und Gaumenmandeln auf Antigene, die die spezifischen Abwehrmechanismen auslösen. VI Ausgewählte Funktionen Stress Das beschriebene System der Immunabwehr ist weitgehend angeboren, funktioniert insgesamt ohne kognitive Beteiligung und ist also nicht bewusst steuerbar. Hingegen wird mit Stress eine Art der Auseinandersetzung mit der Umwelt beschrieben, die alle Systeme des Körpers betreffen kann. Stress beschreibt eine Interaktion des Individuums mit seiner Umwelt: An das Individuum werden Forderungen gestellt, die es zu bewältigen versucht. Diese Interaktion bedeutet ein Auslenken aus dem idealerweise bestehenden Gleichgewicht körperlicher und psychischer Funktionen. Die Anforderungen aus der Umwelt, die eine Stressreaktion hervorrufen, werden Stressoren genannt. Solche Stressoren können sehr unterschiedlich sein, von plötzlichen Temperaturschwankungen über Nahrungsmangel bis hin zu Arbeiten unter Zeitdruck oder kognitiven Leistungsanforderungen. Die Reaktion auf Stressoren, die Stressreaktion oder der Stress im engeren Sinne ist multidimensional und zeigt sich sowohl in körperlichen (neurologischen und endokrinen) als auch in psychischen (Verhaltens- und Erlebens-)Komponenten. Da diese Reaktion unabhängig von der Art des Stressors immer ähnlich verläuft, bezeichnet man Stress auch als generalisierte Anpassungsreaktion. Abb. VI.12: Die zwei Systeme der Stressreaktion. Die Abbildung zeigt das Zusammenspiel der HPA-Achse (links) und des sympathischen Nervensystems (rechts) bei der Stressreaktion. CRH = Kortikotropin-releasing-Hormone, ADH = antidiuretisches Hormon, ACTH = adrenokortikotropes Hormon. Die Arbeit der HPA-Achse wird durch zahlreiche Feedbackschleifen sehr eng reguliert (unterbrochene Linien). So besitzen v.a. die Hypophyse und der Hypothalamus Chemosensoren zur Registrierung der im Blut vorhandenen Konzentrationen verschiedener Hormone (aus: Schandry, 2006, S. 336). 155 156 Biologische Grundlagen Die körperliche Komponente der Stressreaktion ist im Wesentlichen durch die Achse Hypothalamus–Hypophyse–Nebennierenrinde (= hypothalamus– pituitary–adrenocortical –axis oder HPA-Achse) vermittelt, wobei sowohl das sympathische Nervensystem als auch das Hormonsystem beeinflusst werden. Vermittelt über die zentralen Steuerungseinheiten Kortex, limbisches System und Hypothalamus reagiert das sympathische Nervensystem mit einer Aktivierung im Sinne der Vorbereitung einer (körperlichen) Leistung: Die Aktivität des Herzens wird gesteigert, die Bronchien werden weit gestellt und die Arbeitsmuskulatur besser durchblutet. Diese Aktivierung wird durch die Ausschüttung der Transmitter Adrenalin und Nordadrenalin vermittelt, die außerdem zu einer Energiefreisetzung über den Glukosestoffwechsel führen, aber auch als Transmitter in zentralen Regionen emotional-kognitive Bewertungen anregen und modulieren. Die hormonelle Komponente der Stressreaktion besteht insbesondere in einer, durch verschiedene Releasing-Faktoren und –Hormone vermittelten Freisetzung von Glukokortikoiden, darunter vor allem des Kortisols aus der Nebennierenrinde. Die Glukokortikoide regen die Produktion von Glukose in der Leber sowie den Abbau von Muskelproteinen an und hemmen die Synthese von Muskelproteinen. Durch diese Prozesse wird die Energie für mögliche Reaktionen auf Stressoren bereit gestellt. Weiterhin wird durch Glukokortikoide das Immunsystem beeinflusst: Entzündungsprozesse werden gehemmt und das gesamte Immunsystem gedämpft (Immunsuppression). Und nicht zuletzt werden durch Glukokortikoide auch zentrale Prozesse wie Emotionen und Kognitionen beeinflusst. Gleichzeitig mit den körperlichen Reaktionen laufen daher erlebens- und verhaltensmäßige Reaktionen ab. Stressoren werden kognitiv-emotional bewertet und lösen zum Beispiel Emotionen wie Angst oder Aggression aus, die ihrerseits wiederum mögliche Verhaltensreaktionen beeinflussen und das Individuum zu einem Rückzug oder zu einem Angriff bewegen. Bedeutung von Stress Stress beziehungsweise die Stressreaktion kann als generalisierte Anpassungsreaktion auf Anforderungen aus der Umwelt verstanden werden. Der Organismus bereitet sich auf eine Reaktion vor, indem er die notwendige Energie bereitstellt. Daneben werden die Mechanismen gedrosselt, die für eine solche Reaktion nicht benötigt werden (z.B. Immunsuppression). Die Reaktion auf einen Stressor kann in einer aktiven Bewältigung, einer Auseinandersetzung mit oder sogar in einer Beseitigung des Stressors, aber auch in Flucht oder Rückzug bestehen (sog. fight-flight – response). Wirkt der Stressor nicht mehr, kehrt der Organismus wieder zu seinem Gleichgewicht zurück. VI Ausgewählte Funktionen Problematisch wird es allerdings, wirkt der Stressor weiter, weil er nicht beseitigt oder ihm nicht ausgewichen werden kann. Hier können sich die kurzfristig positiven Folgen, die Stress auf die verschiedenen Systeme hat, ins Negative verändern bis hin zum Zusammenbruch des Immunsystems oder einer dauerhaften Erhöhung der Herzleistung oder Herz-Kreislauf-Erkrankungen. Auch kognitiv-emotional ergeben sich bei länger dauerndem Stress (oder auch bei einmaligen, starken Stressoren) negative Auswirkungen wie Gedächtnis- und Konzentrationsstörungen, die Deregulation des Schlaf-Wachzyklus und der psychischen Ausgeglichenheit. VI.6 Emotionen Der Begriff Emotionen bezieht sich auf Gefühlszustände, die bewusst erlebt werden, wegen ihrer Intensität eine Auslenkung aus dem üblichen Gleichgewicht bedeuten und eine gewisse Zeit dauern. Damit werden Emotionen von Stimmungen und Befindlichkeiten, aber auch von kurzfristigen Affekten abgegrenzt. Beispiele für Emotionen sind Angst, Ärger, Freude, Traurigkeit. Emotionen können auf zwei Dimensionen beschrieben werden, einmal nach dem Grad der sie begleitenden Aktivierung (aktivierend versus desaktivierend) und ihrer Valenz (angenehm versus unangenehm). Emotionen sind immer ein mehrdimensionales Geschehen. Auf der körperlichen Ebene verändert sich der physiologische Erregungszustand, wenn beispielsweise bei Ärger der Blutdruck steigt. Die subjektive Bewertung beziehungsweise das Erleben des Zustandes beschreibt die Qualität der Emotion, also ob es sich um Ärger oder um Freude handelt. In Mimik und Gestik finden Emotionen ihren Ausdruck und können sich anderen mitteilen. Und schließlich kann eine Emotion von spezifischem Verhalten begleitet werden, zum Beispiel dem Fliehen aus einer Furcht und Angst erregenden Situation. Ausgelöst werden Emotionen durch Wahrnehmung und Bewertung spezifischer Reize oder Reizkonstellationen, auch die Wahrnehmung von Emotion bei anderen. Emotionen können umgekehrt aber auch Einfluss auf die Wahrnehmung beziehungsweise die Selektion von Reizen nehmen. Innerhalb der psychologischphysiologischen Modellbildung wird diskutiert, in welcher zeitlichen und kausalen Verbindung die dynamische und die evaluative Komponente der Emotionen stehen oder, anders ausgedrückt, ob die physiologische Erregung der Bewertung vorausgeht oder ob die Bewertung die physiologische Aktivierung bewirkt. Emotionen manifestieren sich immer als multidimensionales Geschehen, das alle körperlichen Prozesse und Funktionen umfasst. Auf der Ebene des Gehirns spielt das limbische System mit den zugehörigen Strukturen der Amygdala und des Hippokampus eine zentrale Rolle. Die Funktion der Amygdala ist vor allem eine auslösende und integrierende, was durch Affe- 157 158 Biologische Grundlagen renzen aus dem sensorischen Kortex sowie Efferenzen zum Thalamus und weiteren Hirnstrukturen belegt wird. Die Gedächtnisfunktion des Hippokampus erklärt das Erlernen der emotionalen Bedeutung von Reizen. Abb. VI.13: Konditionierte emotionale Furchtreaktion und Amygdala. Der Fluchtreiz wird schnell und stereotyp über die thalamo-amygdalären Verbindungen und langsamer über die kortikalen Verbindungen zur Amygdala erzeugt. Die sensorische Information vom Thalamus zur Amygdala ist schemenhaft und auf den biologischen Sachverhalt reduziert (z.B. grobe Konturen einer Schlange in der Mitte der Abb.), die vom Kortex ist präzise. Exekutive Funktionen werden über das Cingulum und den Frontallappen aktiviert (aus: Schmidt & Schaible (Hrsg.) (2006). Neuro- und Sinnesphysiologie (5., neu bearb. Aufl.), S.445, mit Genehmigung von © Springer). VI Ausgewählte Funktionen Abb. VI.14: Schematische Darstellung der in VI.13 gezeigten Vorgänge. Der laterale Kern der Amygdala erhält Informationen aus den sensorischen Kernen des Thalamus (1), Neokortex (2) und höheren Assoziationskortizes (3) und der basale Kern vom Hippocampus (4). Bei einfachen Hinweisreizen, die keine Diskrimination erfordern, kann die Konditionierung schon über 1 erfolgen. 2, 3 und 4 sind notwendig, wenn das Ereignis von anderen Ereignissen genau diskriminiert und Bewertungen und Erwartungen erfolgen. Die Amygdala projiziert zu vielen kortikalen Arealen und zum Hippokampus zurück. Die somatomotorischen, endokrinen und vegetativen Reaktionen während der Furchtkonditionierung werden über den zentralen Kern der Amygdala und die entsprechenden Kerngebiete im Hypothalamus und Hirnstamm ausgelöst und der parietale Kortex erhält eine Rückmeldung darüber („Interoception“). Die Weckreaktion des Kortex wird über den zentralen Kern der Amygdala und den Nucleus basalis vermittelt (aus: Schmidt & Schaible (Hrsg.) (2006). Neuro- und Sinnesphysiologie (5., neu bearb. Aufl.), S.446, mit Genehmigung von © Springer). Die Bewertung des erlebten Zustandes, also das Erleben einer spezifischen Emotion, sowie die resultierende Verhaltensplanung werden im Präfrontalkortex gesteuert. Der anteriore Gyrus cinguli scheint dabei nicht nur den Reiz, sondern auch mögliche Verhaltensresultate zu bewerten. Auf der neurochemischen Seite wird eine Reihe von Transmittern im Zusammenhang mit Emotionen erforscht. So scheint Noradrenalin bei negativen Emotionen eine wichtige Rolle zu spielen, bei positiven Gefühlen werden vor allem Dopamin und Endorphine genannt. Deren Ausschüttung vermittelt die verschiedenen physiologischen Veränderungen im Zusammenhang mit Emotionen, wie die Veränderung der Herzfrequenz bei Angst oder auch der durch positive Emotionen geschaffene Anreiz, die auslösende Situation wieder aufzusuchen. 159 160 Biologische Grundlagen Bei eindeutigen und starken Emotionen imponiert die Ebene des Ausdrucks, insbesondere der sprachliche Ausdruck aber auch die Mimik. Der Gesichtsausdruck bei starkem Ärger und die Bedeutung eines Lächelns werden interkulturell verstanden. Das resultierende Verhalten wird durch Emotionen gesteuert, aber auch durch kognitive Bewertungen beeinflusst. Abb. VI.15: Prozentualer Anteil richtiger Zuordnungen von Gesichtsausdrücken und Emotionswörtern in fünf verschiedenen Kulturen (aus: Ekman, P. (1976) Ein Lächeln ist ein Lächeln ist ein Lächeln, Psychologie Heute, 3, 31-35). Bedeutung von Emotionen Emotionen helfen, Verhalten zu steuern. Durch einfache, schnelle Bewertungsprozesse wird, ohne durchgängige detaillierte kognitive Analyse, mögliches Verhalten vorbereitet und ausgeführt. Die schnelle Klassifikation von Reizen bzw. Situationen in bedrohlich – ungefährlich, angenehm – unangenehm, wichtig - unwichtig und interessant – uninteressant-neutral, kann auch als eine basale wertende und damit kognitive Leistung verstanden werden. Bei angenehmen Gefühlen wird eine Suche nach den auslösenden Situationen beziehungsweise der Versuch stehen, diese möglichst zu verlängern. Unangenehme Gefühle führen zu Flucht oder Vermeidung der auslösenden Reizkonstellation. Diese Bewertung ist jedoch nicht nur auf die eigene Person bezogen. Emotionen sind eines der unmittelbar wirksamsten Kommunikationssysteme. Das Erkennen der Emotion eines anderen, vor allem einer wichtigen Bezugsperson kann lebensnotwendig sein, sei es, um möglichen Gefahren zu entgehen, um Angriffen auszuweichen oder verstärkende Situationen zu erkennen. Über diese Systeme werden Bindungen aufgebaut und aufrechterhalten. VI Ausgewählte Funktionen VI.7 Lernen und Gedächtnis Mit Lernen wird in der Psychologie der erfahrungsbasierte Änderungsprozess bezeichnet, der die Wahrscheinlichkeit des Auftretens bestimmter Verhaltensweisen verändert. Anders ausgedrückt bedeutet Lernen, dass sich auf der Basis von Erfahrungen Veränderungen in der Verbindung zwischen Reizen und Reaktionen, allgemein gesprochen, im Verhaltens- und speziell im Handlungsrepertoire ergeben. Abzugrenzen ist Lernen von der Reifung, die nicht erfahrungsbasiert, sondern genetisch determiniert ist und die Bandbreite der Interaktionsmöglichkeiten mit der Umwelt und damit der Verhaltensplastizität festlegt. Lernen ist neben Reifung ein zweiter basaler Prozess der sich ständig verbessernden Anpassung eines Individuums an seine Umwelt. Im Folgenden werden die psychologischen Aspekte von Lernen und Gedächtnis nur in Grundzügen dargestellt. Im Kurs 03413 Lernen, Gedächtnis, Wissen, Sprache, Denken und Problemlösen wird diese Thematik vertieft. Im Lexikon der Psychologie von Städtler (1998) sollten entsprechende Stichworte durchgelesen werden. Auch der Terminus Gedächtnis bezieht sich auf Prozesse, hier jedoch auf die Prozesse der Enkodierung von Informationen (also deren Aufbereitung in eine speicherfähige Einheit), deren Speicherung sowie deren Abruf. Zwei Arten des Lernens werden unterschieden: die nicht-assoziative und die assoziative Variante. Nicht-assoziatives Lernen beschreibt Prozesse der Gewöhnung und Anpassung an Reize oder Reizkonstellationen. Der elementarste Lernmechanismus ist die Unterscheidung eines neuen von einem bekannten Reiz. Eine plötzlich auftretende, unerwartete Veränderung in der Umgebung, z. B. ein Rauschen in der Heizung, löst eine Orientierungsreaktion aus. Hält dieses Rauschen gleichmäßig an, dann tritt die Habituation der Orientierungsreaktion ein, also die Gewöhnung an einen Reiz, wenn dieser wiederholt auftritt. Dies ist erkennbar an der Abschwächung der Reaktion. Dieser Gewöhnungsprozess kann, zum Beispiel durch einen weiteren Reiz unterbrochen werden. So könnte das Rauschen im Heizkörper gelegentlich durch ein Knacken unterbrochen werden. Es kommt zur Dishabituation, also zum Wiederauftreten einer, wenn auch schwächeren Reaktion. Oder, falls der weitere Reiz störend oder aversiv ist, zu einer Sensitivierung, also zu einer erneuten Reaktion auf den Ursprungsreiz, deren Stärke sogar das Ursprungsniveau übersteigen kann. Assoziatives Lernen bezieht sich auf die Lernvarianten, bei denen verschiedene Reize miteinander verknüpft (assoziiert) werden. In der einfachen Variante des Klassischen Konditionierens wird ein Reiz, der einen Reflex auslöst (= unkonditionierter Reiz), so häufig mit einem anderen Reiz kombiniert, dass schließlich auch der zweite Reiz diesen Reflex (das reflektorische Verhalten) auslöst. Der zweite Reiz ist der konditionierte Reiz, der kurz vor dem unkonditionieren Reiz auftreten muss (dieses kurze Intervall zwischen konditioniertem und unkonditio- 161 162 Biologische Grundlagen niertem Reiz wird als Kontiguität bezeichnet). Beim operanten beziehungsweise instrumentellen Konditionieren spielen die Effekte der Reaktion auf einen Reiz hin eine Rolle. Sind diese angenehm, wird der Reiz häufiger zu dem Verhalten führen. Man bezeichnet dies als Lernen durch Verstärkung (Belohnung) oder Lernen am Erfolg. Weitere Formen des assoziativen Lernens sind das Wahrnehmungs- oder Imitationslernen, wo über die Beobachtung anderer ein Verhalten oder über eigene Erfahrungen Reaktions- und Handlungshäufigkeiten verändert werden. Das Klassische Konditionieren ist das Lernen von Reizassoziationen (Reizassoziationslernen, Lernen von Stimuluskontingenzen) und das Operante Konditionieren das Lernen von Assoziationen zwischen Verhaltensweisen und Effekten (vgl. law of effect Thorndikes, Lernen am Erfolg). Beim Gedächtnis werden anhand von Art und Dauer der Speicherung von Informationen verschiedene Bestandteile des Gedächtnisprozesses unterschieden. Der sensorische Speicher bezieht sich auf die einzelnen Wahrnehmungsmodalitäten; in ihm sind sensorische Eindrücke (Töne, Bilder) für kurze Zeit (1 bis 2 Sekunden) präsent, bevor sie gelöscht oder in das Kurzzeitgedächtnis übertragen werden. Im Kurzzeit- und Arbeitsgedächtnis sind Informationen (wie der Name bereits sagt), ebenfalls nur für kurze Zeit präsent, meist so lange bis sie benötigt werden. So prägt man sich eine Telefonnummer, die einem ein Bekannter nennt, ein, bis man sie gewählt hat. Im Kurzzeitgedächtnis können sich Informationen für mehrere Minuten halten. Das Langzeitgedächtnis, in dem Informationen über Jahre hinweg verbleiben können, ist schließlich das Gedächtnis, auf das man sich im Alltag meist bezieht, wenn man einer Person ein „gutes Gedächtnis“ zuschreibt. Die biologisch-physiologischen Grundlagen für Lern- und Gedächtnisvorgänge lassen sich auf der Ebene einzelner Zellen sowie in größeren neuronalen Netzen und Regionen des Gehirns finden. Für die zelluläre Ebene lässt sich die von Donald Hebb 1949 aufgestellte Regel zumindest ansatzweise bestätigen. Die Hebb-Regel formuliert eine Assoziationsstärkeregel für zwei Neurone: Eine Verbindung zwischen zwei Neuronen wird immer dann gestärkt, wenn beide zum gleichen Zeitpunkt aktiviert sind (gemeinsame Aktivierung in Raum und Zeit = Kontiguität der Aktivierung). Bei gleichem Vorzeichen der Aktivierung wird die Verbindungsstärke zwischen zwei Neuronen erhöht, bei ungleichem Vorzeichen der Aktivierung wird die Verbindungsstärke erniedrigt. Damit lässt sich die Bildung von neuronalen Netzen verständlich machen, wie sie in der Theorie künstlicher neuronaler Netze simuliert wird. Die Hebbsche Lernregel ist ein wichtiger Bestandteil des neuronalen Konnektionismus und stammt aus der Tradition des Assoziationismus (vgl. Einträge zu den Stichwörtern „Konnektionismus“ und „Assoziationismus“ in Städtler, 1998 oder Eintrag zur Hebb-Regel im Lexikon der Neurowissenschaft, 2001). VI Ausgewählte Funktionen Für natürliche neuronale Netze bedeutet dies, dass die wiederholte, erfolgreiche Erregungsübertragung von einer Nervenzelle auf eine andere zu einem Wachstumsprozess oder einer metabolischen Veränderung in einer oder beiden dieser Zellen führt. Dieser Wachstumsprozess beziehungsweise die metabolische Veränderung macht die Erregungsübertragung zwischen den beiden Neuronen effizienter. Eine solcherart gestärkte Verbindung führt zu einer schnelleren und effizientern Erregungsübertragung, wenn der gleiche Reiz wieder wahrgenommen wird (Bahnung). Dieses, in der Hebb-Regel postulierte Phänomen wird als Langzeitpotenzierung (Long term potentiation = LTP) oder die längerfristige Erleichterung oder Verstärkung der Informationsübertragung von einem Neuron auf ein anderes beschrieben (vgl. Eintrag zum Stichwort „Assoziationsgesetze“ in Städtler, 1998). Abb. VI.16: Strukturveränderung der Synapsen an dentritischen Spines nach LangzeitPotenzierung (LTP). Dendritische Spines sind Ausstülpungen der postsynaptischen (subsynaptischen) Membranabschnitte von Dendriten, deren Größe durch die präsynaptische Aktivität bestimmt wird (aus Schmidt & Schaible (Hrsg.) (2006). Neuro- und Sinnesphysiologie (5., neu bearb. Aufl.), S.421, mit Genehmigung von © Springer). Die dem assoziativen Lernen entsprechenden Habituations- und Sensitivierungsphänomene lassen sich damit bereits auf zellulärer Ebene finden. Kurzfristige Lerneffekte werden durch membranphysiologische Mechanismen erklärt, längerfristige durch den Umbau von Zellen mit dem Aufbau beziehungsweise dem Untergang von Synapsen. Besonders an den Neuronen des für Gedächtnisprozesse wichtigen Hippocampus wurde die Langzeitpotenzierung erforscht, wobei neben den Potenzierungseffekten von Interesse ist, dass Signale aus verschiedenen Quellen hierbei verknüpft werden können. Ein wichtiger Transmitter hier ist das Glutamat. Für den nicht-assoziativen Lerntyps des operanten Konditionierens müssen weitere Systeme in Aktion treten, wobei insbesondere das ventrale tegmentale Areal wichtig zu sein scheint. Diese Systeme wirken als „Belohnungssystem“, die das Auftreten einer gefundenen Reiz-Reaktionsstruktur häufiger (wahrscheinlicher) macht. Gemeinsam ist ihnen, dass hier Dopamin als Transmitter zu finden ist. 163 164 Biologische Grundlagen Die Bedeutung des Hippocampus für Lern- und Gedächtnisprozesse lässt sich auch bei der Betrachtung des Gedächtnisses finden. Während sich die kurzfristigen Gedächtnisphänomene im parahippokampalen und im präfrontalen Kortex (Enkodierung) beziehungsweise im Präfrontalkortex allein (Kurzzeitgedächtnis und Arbeitsgedächtnis) abspielen, wird bei der Konsolidierung und Langzeitspeicherung von Gedächtnisinhalten ebenfalls der Hippokampus aktiv. Daneben werden bei unterschiedlichen Gedächtnisinhalten auch weitere Strukturen eingebunden. Wird eine sensomotorische Fertigkeit wie das Radfahren gespeichert, sind motorische Kortexareale und natürlich das Kleinhirn involviert. Abb. VI.17: Kortikale Strukturen für Lernen und Gedächtnis. a Ventrale Ansicht des Affengehirns mit den verschiedenen Läsionsorten, die im Tiermodell zur Amnesie führten. Amygdala (A) und Hippokampus (H) sind punktiert eingezeichnet und die benachbarten kortikalen Regionen in Farbe. Angegeben sind der perirhinale Kortex (Area 35 und 36), der periamygdaloide Kortex (Area 51), der entorhinale Kortex (Area 28) und der parahippokampale Kortex (Areale TH und TF). Schematischer Aufbau des Gedächtnissystems des medialen Temporallappens. Der entorhinale Kortex projiziert in den Hippokampus, wobei zwei Drittel der kortikalen Afferenzen in den entorhinalen Kortex aus den benachbarten perirhinalen und parahippokampalen Kortizes entspringen. Diese wiederum erhalten Projektionen von unimodalen und polymodalen kortikalen Arealen im frontalen, temporalen und parietalen Bereich. Der entorhinale Kortex erhält darüber hinaus direkte Afferenzen vom orbitalen Frontalkortex, dem Gyrus cinguli, dem insulären Kortex und dem oberen Temporallappen. Alle diese Projektionen sind reziprok (printed with permission from Squire & Zola-Morgan (1991), Science, 253, p. 1380, © 2009 AAAS). Wie oben bereits ausgeführt, sind Lern- und damit verbundene Gedächtnisprozesse neben der Reifung der zweite wichtige Prozess, der die Entwicklung des Individuums und dessen Anpassung an seine Umwelt (auch im Sinne einer Veränderung seiner Umwelt) ermöglicht. Die oben beschriebenen physiologischen Grundlagen des Lernens lassen sich über den gesamten Lebenslauf hinweg finden und bilden damit die Basis für lebenslanges Lernen. VI Ausgewählte Funktionen Studientechnischer Anhang 1. Kenntnisse Rhythmen des (menschlichen) Lebens o circadiane Rhythmik, BRAC o Steuerung, beteiligte Hirnareale und Hormone o betroffene Subsysteme Wachsein und Aufmerksamkeit o Aufmerksamkeit (Auslösung, beteiligte Hirnstrukturen) o Reizselektion, Reizerkennung und Reizverarbeitung o Bedeutung für den Menschen Schlafen o Schlafstadien und deren Kennzeichnung (EEG-Frequenzbänder) o „aktive“ Hirnregionen o Bedeutung des Schlafs Durst o hypovolämischer Durst, osmotischer Durst o beteiligte Systeme und Vermittlung (Barorezeptoren, Osmosensoren, Vasopressin, Renin und Angiotensin, Nucleus medianes praeopticus, Hypothalamus) Trinken o primäres und sekundäres Trinken o antizipatorische Sättigung Hunger o Kurzzeitregulation und Langzeitregulation o beteiligte Systeme und Vermittlung (Glukose, Sättigungssignale aus Magen-Darm-Trakt, Insulin, Leptin, Nucleus tractus solitarius, Hypothalamus) Essen und Sättigung o präresorptive Sättigung o Lernerfahrungen und Bewertungen Sexualität o Bedeutung (Trieb, Arterhaltung, Bindung) o sexuelle Reaktion, Sexualverhalten Koitus o Phasen des Koitus o Steuerung (Sexualhormone, VNS) Schwangerschaft und Geburt o Befruchtung und Keimentwicklung (hormonelle Steuerung) o Geburt (hormonelle Steuerung) Reifung o Abgrenzung: Reifung, Entwicklung, Altern o Reifungs- und Alterungsprozesse: hormonelle Steuerung, Zellteilung und Zelltod 165 166 Biologische Grundlagen Reifung und Entwicklung über die Lebensspanne: Körperliche Veränderungen und Korrelate o vorgeburtliche Entwicklung: Nidation, Embryonalstadium, Fetus o Kindheit: Gehirnentwicklung, Längenwachstum o Pubertät: Wachstum, Geschlechtsreife o Erwachsenenalter o Alter: genetische Programmierung, Lebenserwartung Immunabwehr und beteiligte Strukturen o unspezifische Immunabwehr: physikalisch-chemische Barrieren, Makrophagen, Komplementsysteme, MHC-Proteine o spezifische Immunabwehr: Antigene, B- und T-Lymphozyten, Killerzellen Stress o Stress, Stressoren und Stressreaktion: generalisierte Anpassungsreaktion, Bewältigung o Stressreaktion: HPA-Achse, VNS, Hormone, Immunsuppression o Stressreaktion: Ebene des Verhaltens und Erlebens o längerdauernder Stress und Folgen Emotionen o Gefühle (gewisse Intensität, überdauernd) vs. Affekt, Befindlichkeit, Stimmung o Dimensionen von Emotionen (Aktivierung, Valenz) o Bedeutung (Bewertung, Kommunikation) Emotionsgeschehen o Ebenen des Emotionsgeschehens (körperliche Ebene, Erleben, Ausdruck, Verhalten) o Auslösung von Emotionen o beteiligte Strukturen (limbisches System, Präfrontalkortex) und Stoffe (Transmitter) Lernen o Definition o nicht-assoziatives Lernen (Habituation und Orientierungsreaktion, Dishabituation, Sensitivierung) o assoziatives Lernen (klassische Konditionierung, instrumentelles und operantes Konditionieren) o Lernen als erfahrungsbasierte Verhaltensänderung (vgl. Reifung) Gedächtnis o Definition (Enkodierung, Speicherung, Abruf) o Arten: sensorischer Speicher, Kurzzeit- oder Arbeitsgedächtnis, Langzeitgedächtnis Physiologische Grundlagen (Strukturen und Prozesse) o Hebb-Regel, Langzeitpotenzierung o Parahippokampaler Kortex, Präfrontalkortex, Hippocampus, ventrales tegmentales Areal o Transmitter: Glutamat und Dopami VI Ausgewählte Funktionen 2. Literatur Lexika, Nachschlagewerke Städtler, T. (1998). Lexikon der Psychologie. Wörterbuch, Handbuch, Studienbuch. Stuttgart. Kröner. Elektronisch zugänglich für autorisierte Benutzer im Hochschulnetz der FernUniversität in Hagen: Hanser, H. (Red.). (2001) (Elektronische Ressource). Lexikon der Neurowissenschaft. Mannheim: Brockhaus, Duden, Neue Medien. „Elektronisch zugänglich für autorisierte Benutzer im Hochschulnetz der FernUniversität in Hagen“ heißt: Sich in die UB FeU einloggen, Eintrag „Lexikon der Neurowissenschaft“ unter „Suchen“ eingeben und http://lexika.tanto.de/?TANTO_KID=digibib12&TANTO_AGR=133581 klicken. Dort finden sich weitere nützliche Lexika (der Psychologie (s.u.), der Medizin usw.). Wenninger, G. (Hrsg.) (2002) (Elektronische Ressource). Lexikon der Psychologie. Mannheim: Brockhaus, Duden, Neue Medien. Grundlage: Schandry, R. (2006). Biologische Psychologie (2. überarb. Aufl.). Weinheim: Beltz. Kapitel 19: Rhythmen des Verhaltens Kapitel 20: Schlaf und Traum (Abschnitte 20.1 bis 20.6) Kapitel 18: Sexualität und geschlechtsspezifisches Verhalten (Abschnitte 18.1, 18.2, 18.5, 18.6.3 und 18.6.4) Kapitel 17: Stress Kapitel 23: Emotionen Kapitel 24: Lernen und Gedächtnis Birbaumer, N. & Schmidt, R.F. (2006). Motivation und Emotion. In R.F. Schmidt & H.-G. Schaible (Hrsg.). Neuro- und Sinnesphysiologie (S. 424-448). Heidelberg: Springer. (Abschnitt 18.1: Homöostatische Triebe: Durst und Hunger) 167 168 Biologische Grundlagen Heubrock, D. (2008). Entwicklung kognitiver Systeme und Entwicklungsneuropsychologie. In S. Gauggel & M. Herrmann (Hrsg.). Handbuch der Neuround Biopsychologie (S. 305-317). Göttingen: Hogrefe. (Abschnitte 1 und 2) Vertiefend: Bauer, C. & Walzog, B. (2005). 9.4 Abwehrmechanismen des Körpers. In R. Klinke, H.-Chr. Pape & S. Silbernagl, S. (Hrsg.). Physiologie (S. 233-245). Thieme: Stuttgart Birbaumer, N. & Schmidt, R.F. (2006). Wachen, Aufmerksamkeit und Schlafen. In R.F. Schmidt & H.-G. Schaible (Hrsg.). Neuro- und Sinnesphysiologie (S. 374-401). Heidelberg: Springer. Birbaumer, N. & Schmidt, R.F. (2006). Motivation und Emotion. In R.F. Schmidt & H.-G. Schaible (Hrsg.). Neuro- und Sinnesphysiologie (S. 424-448). Heidelberg: Springer. Birbaumer, N. & Schmidt, R.F. (2006). Lernen und Gedächtnis. In R.F. Schmidt & H.-G. Schaible (Hrsg.). Neuro- und Sinnesphysiologie (S. 402-423). Heidelberg: Springer Bischof-Köhler, D. (2006). Von Natur aus anders. Die Psychologie der Geschlechtsunterschiede (3., überarb. u. erw. Aufl.). Stuttgart: Kohlhammer. Klinke, R., Pape, H.-Chr. & Silbernagl, S. (Hrsg.). Physiologie (5. Aufl.) (S. 4551). Thieme: Stuttgart Leichtweiß, H.-P., Schröder, H.J. & Singer, D. (2005). Sexualfunktionen, Schwangerschaft und Geburt. In R. Klinke, H.-Chr. Pape & S. Silbernagl, S. (Hrsg.). Physiologie (S. 561-588). Thieme: Stuttgart Schmidt, R.F. & Schaible, H.-G. (Hrsg.) (2006). Neuro- und Sinnesphysiologie (5., neu bearb. Aufl.). Heidelberg: Springer. Silbernagl, S. (2005). 2.8 Altern und Zelltod. In R. Klinke, H.-Chr. Pape & S. Silbernagl (Hrsg.). Physiologie (5. Aufl.) (S. 45-51). Thieme: Stuttgart Sturm, W. (2008). Aufmerksamkeit. In S. Gauggel & M. Herrmann (Hrsg.). Handbuch der Neuro- und Biopsychologie (S. 329-336). Göttingen: Hogrefe. Zulley, J. & Hajek, G. (2008). Schlaf und circadiane Rhythmik. In S. Gauggel & M. Herrmann (Hrsg.). Handbuch der Neuro- und Biopsychologie (S. 295302). Göttingen: Hogrefe. VI Ausgewählte Funktionen Sonstige zitierte Literatur: Ekman, P. (1976). Ein Lächeln ist ein Lächeln ist ein Lächeln. Psychologie Heute, 3, 31-35. Schwartz, M.W., Woods, S., Porte, D., Seeley, R., Baskin, D. (2000). Central nervous system control of food intake. Nature, 404, 661-670. Squire, L. R. & Zola-Morgan, S. (1991). The medial temporal lobe memory system. Science, 253, 1380-1384. 169