Wie Axone ihren Weg durch den Körper finden
Werbung

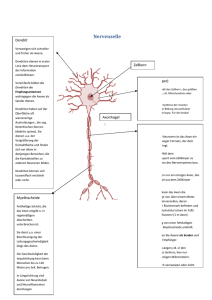
P O L I T I K MEDIZINREPORT Neuronale Führungsmoleküle Wie Axone ihren Weg durch den Körper finden Wie finden Axone, die bis zu zwei Meter langen Fortsätze von Nervenzellen, ihren Weg durch verschiedenste Körpergewebe? Weltweit arbeiten Neurobiologen daran, die Wegweiser für wachsende Neurone zu identifizieren. Nach dem aktuellen Stand der Forschung steuern vier Arten von Signalen das Wachstum von Axonen: Über kleine Distanzen wirken Zelladhäsionsmoleküle an der Oberfläche von Nervenzellen, deren Existenz Forscher bereits in den achtziger Jahren nachgewiesen haben. Diese Moleküle können als Lockstoffe fungieren. Als solche veranlassen sie Neuronen, sich bei Kontakt mit anderen Zellen an diese zu haften. Andere Zelladhäsionsmoleküle lösen den Rückzug von Nervenzellen aus; derartige Proteine wurden auf der Membran embryonaler Gewebezellen nachgewiesen. Daneben gibt es Lock- und Abwehrstoffe größerer Reichweite. Sie werden von Neuronzielzellen freigesetzt und diffundieren ins umliegende Gewebe. Als ersten Lockstoff dieser Art entdeckten Neurobiologen 1994 das Protein Netrin-1. Es wird von Zellen der Bodenplatte produziert, einem Gewebestreifen, der längs der ventralen (unteren) Oberfläche des sich entwickelnden Rückenmarks verläuft. Dort dient das Protein als spezifisches Führungsmolekül für Axone der Kommissurenneurone, die Schmerz- und Temperaturreize zum Gehirn leiten: Durch den Lockeffekt von Netrin-1 finden die Axone dieser Nervenzellen, die an der dorsalen (oberen) Oberfläche des Rückenmarks entstehen, ihren Weg zur ventralen Oberfläche, wo sie die Bodenplatte durchqueren und zum Gehirn hin weiterwachsen. Der erste Nachweis freier Abwehrstoffe gelang 1993 mit Kollagengelkulturen von Nervenzellen, wobei die Identität der Stoffe zu- nächst unklar war. Inzwischen sind einige Vertreter dieser Gruppe von Führungsmolekülen identifiziert. Semaphorin III steuert die Axone bestimmter sensorischer Neuronen der Ratte. Am Entstehungsort, der ventralen Hälfte des Rückenmarks, wirkt es wie ein Sieb auf Axone, die durch die dorsale Oberfläche ins Rückenmark eintreten: Nervenzellfortsätze sensorischer Neuronen, die Schmerzund Temperaturreize übermitteln und in der dorsalen Region enden, stößt es ab. Ein neurales Führungsmolekül, Semaphorin III, stößt ein sensorisches Neuron ab (oben), während (unten) ein anderer Typ des sensorischen Neurons unangegriffen bleibt und sich an den Zellen ausbreiAbbildung: Science, Vol 268, Mai 1995 ten kann. Axone von Nervenzellen hingegen, die zur ventralen Region des Rückenmarks verlaufen und Informationen über Muskellage und Muskelstreckung weiterleiten, beeinflußt Semaphorin III nicht. Semaphorin III ist auch unter der Bezeichnung Collapsin bekannt. Eine andere Forschergruppe identifizierte es nämlich A-2452 (32) Deutsches Ärzteblatt 93, Heft 39, 27. September 1996 parallel als neuronales Führungsmolekül, das Hühneraxonspitzen kollabieren läßt. Semaphorin II, ein weiteres nichtmembrangebundenes Abwehrmolekül, weist einen anderen Wirkungsmechanismus auf: Bestimmte Neurone bilden mit Muskelzellen, die Semaphorin II produzieren, keine Synapsen. Interessanterweise gibt es auch bifunktionelle Führungsmoleküle wie UNC-6, nachgewiesen in einer Nematodenart. Das Protein steuert zwei Arten von Axonen, die in gegensätzliche Richtungen wachsen. Während es die eine Axongruppe anlockt, stößt es die andere ab. Diese Push-pull-Wirkung konnten Neurobiologen auch für Netrin-1 nachweisen, das bereits als Lockstoff für Kommissurenneurone genannt wurde. Netrin-1 lockt Neurone, die die Bodenplatte durchqueren sollen, an; solche, die sie nicht durchqueren sollen, stößt es ab. Vermutlich spielt das Protein auch bei der Führung von Neuronen im Gehirn eine Rolle. Die Erkenntnisse über die Funktion von Netrin-1 und Semaphorin III sind abgeleitet von Neuronkulturen. Untersuchungen am Tier, die endgültig nachweisen, ob das Entfernen von Genen, die für ein bestimmtes Führungsmolekül kodieren, zu den erwarteten neuronalen Führungsdefekten führt, fehlen bisher. Daß Semaphorin-Gene überraschenderweise auch in Geweben der Lunge oder des Immunsystems exprimiert werden, läßt auf Funktionen in neuen Bereichen des Nervensystems schließen. Außerdem verfügt jedes Semaphorin über ein spezifisches Expressionsmuster, wobei in verschiedenen Geweben unterschiedliche Gene an- oder abgeschaltet sind. Dies legt nahe, daß jedes Semaphorin eine andere Neurongruppe lenkt. Zu den wichtigsten Forschungsinhalten zählt, herauszufinden, welche Neurone auf welche freien Führungsmoleküle ansprechen. Denn die neuen Erkenntnisse über neuronale Führungsmechanismen sollen auch klinische Anwendung finden: Neuronlockstoffe könnten eingesetzt werden, um sich regenerierende Neuronen bei Rückenmarksverletzungen oder anderen Nervenschäden an ihr Ziel zu dirigieren. Birgit Strohmaier