4.6 Pflanzliche und bakterielle Stoffwechselprozesse
Werbung

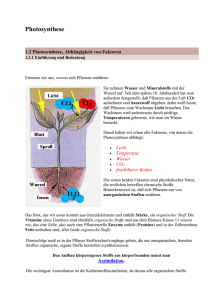
Pflanzliche und bakterielle Stoffwechselprozesse • 4.6 Pflanzliche und bakterielle Stoffwechselprozesse – 4.6.1 Photosynthese – 4.6.2 Chemosynthese – 4.6.3 Calvin-Cyclus Photosynthese • Die Photosynthese ist ein biochemischer Prozess, bei dem Lichtenergie durch Land- und Wasserpflanzen sowie bestimmter Bakterien in einem reduktiven Prozess in chemische Energie umgewandelt wird. • Diese dient der Fixierung von CO2 : Aus energiearmen, anorganischen Stoffen, hauptsächlich CO2 und Wasser H2O, werden dabei energiereiche Kohlenhydrate synthetisiert. • Man unterscheidet zwischen oxygener und anoxygener Photosynthese. Bei der oxygenen wird O2 freigesetzt, bei der anoxygenen nicht. Bei der anoxygenen Photosynthese können statt Sauerstoff andere anorganische Stoffe entstehen, beispielsweise elementarer Schwefel. Teilschritte der Photosynthese • Die Photosynthese kann in der Teilschritte untergliedert werden: • Im ersten Schritt wird die elektromagnetische Engergie in Form geeigneter Wellenlänge unter Verwendung von Farbstoffen absorbiert. • Direkt hieran anschließend erfolgt im zweiten Schritt eine Umwandlung der elektromagnetischen Energie in chemische Energie. • Im letzten Schritt wird diese chemische Energie zur Synthese energiereicher organischer Verbindungen verwendet. Lokalisation • Die Reaktionsfolge der Photosynthese ist membrangebunden. • Photosynthetisch aktive Membranen heißen Thylakoide. • Bei Eukaryonten befinden sich die Thylakoide in den Chlorolasten. Aufbau: Chloroplast Hierbei liegen die Thylakoidmembranen entweder in gestapelter (7) oder in ungestapelter (8) Form vor. 1: äußere Membran; 2: Intermembranraum ; 3: innere Membran; 4: Stroma; 5: Thylakoidlumen; 6: Thylakoidmembran; 7: Granum; 8: Thylakoid; 9: Stärke(körper); 10: plastidäres Ribosom; 11: plastidäre DNS; 12: Plastoglobulus • Die Thylakoide sind die Struktureinheiten der sog. Grana, die wiederum im Stroma der Chloroplasten eingebettet sind. • In den Thylakoiden findet die Lichtreaktion statt: – Lichtabhängige Synthese von NADPH/H+ und ATP • Die im Anschluss stattfindende Synthese von Zuckermolekülen aus NADPH, ATP und CO2 findet im Stroma der Chloroplasten statt (Dunkelreaktion). Photosynthetisch aktive Pigmente • Damit elektromagnetische Energie genutzt werden kann, muss diese zuerst absorbiert werden. • Man unterscheidet drei Gruppen von photosynthetisch aktiven Pigmenten: – Chlorohylle – Carotinoide – Phycobiline Chlorophylle • Chlorophylle sind Chelatkomplexe aus einem derivatisierten Porphyrin-Ring und Mg2+ als Zentralion. Je nach Art des Chlorophylls sind an den Grundkörper verschiedene Seitenketten angehängt. • Chlorophyll ist mit Proteinen nicht kovalent assoziiert. • Läuft die Photosynthese bei Lebewesen ab, die Sauerstoff freisetzen, spricht man allgemein von Chlorophyll. Bei Organismenmit anoxygener Photosynthese bezeichnet man das Chlorophyll als Bakteriochlorophyll. Chlorophylle • Struktur der Chlorophylle Carotinoide • Carotinoide bestehen aus ungesättigten Kohlenwasserstoffketten und deren Oxidationsprodukten. Carotinoide sind formal aus 8 Isopren-Einheiten aufgebaut. Man unterteilt sie in – Carotine, die nur aus Kohlenstoff und Wasserstoff aufgebaut sind und – Xantophylle, sauerstoffhaltige Derivate der Carotine. • Carotinoide kommen in allen photosynthetisch wirksamen Zellen vor Β-Carotin Phycobiline • Phycobiline bestehen aus einem Proteinanteil und einer chromophoren Gruppe, die ähnlich den Porphyrinderivaten (Chlorohyll) aus vier miteinander verbundenen Pyrrolringen bestehen, allerdings nicht in zyklischer, sondern in linearer Anordnung. • charakteristische akzessorische Pigmente von Algen und Cyanobakterien (Blaualgen) • Der Chromophor ist im Gegensatz zu Chlorophyllen und Carotinoiden kovalent an das Protein gebunden, also eine prosthetische Gruppe Phycoerythrobilin Lichtsammelkomplexe • Einfache Pigmente könnten nicht eine effiziente Photosynthese betreiben, weil die Wahrscheinlichkeit für das Einfangen von Licht in der richtigen Wellenlänge und am richtigen Ort äußerst gering ist. • Daher verwenden phototrophe Organismen sogenannte Antennenkomplexe. Diese erhöhen die Auftrittsfläche und das Spektrum, mit der Photonen absorbiert werden. Antennen bestehen aus sogenannten Lichtsammelkomplexe. Diese bestehen in Pflanzen aus einer Zentralantenne (core) und einer äußeren Antenne, allesamt chlorophyllhaltige Proteinkomplexe. • Als Chromophore dienen dabei nicht nur Chlorophyll a, welches für photosynthetische Prozesse essentiell ist, sondern auch aus den sog. akzessorischen Pigmenten (Chlorophylle b-e, Carotinoide und Phycobiline). Diese schützen zum einen die Antenne, falls ein Chlorophyllmolekül einen schädlichen Triplettzustand ausbildet. Zum anderen erhöhen diese Chromophore den Wellenlängenbereich zum Einfangen von Licht. • Die Chromophore leiten durch ihre räumliche Nachbarschaft die Lichtenergie von einem Pigment zum anderen. Diese definierte Menge Anregungsenergie bezeichnet man auch als Exziton. Exzitone gelangen schließlich in wenigen Pikosekunden in das Reaktionszentrum. Bei Pflanzen und Cyanobakterien ist dies ein Chlorophyll a-Molekül. Photosynthetisch wirksame Lichtfrequenzen • Die Absortionsmaxima von Chlorophyll a liegen bei 420 und 600 nm, d.h. die Lichtfrequenzen, die dazwischen liegen, werden von Chlorophyll nicht absorbiert. ( Grünlücke) • Die akzessorischen Pigmente engen diese Lücke ein, da sie in der Lage sind, Lichtfrequenzen in diesem Bereich zu absorbieren. Ablauf der Photosynthese • Die Energieübertragung findet bei der oxygenen Photosynthese in zwei trennbaren Photosystemen statt: – Photosystem II (P680) • Absorptionsmaximum bei 680 nm • Chlorophyll a und b – Photosystem I (P700) • Absorptionsmaximum bei 700 nm • Chlorophyll a • Die beiden Systeme sind durch eine Elektronentransportkette miteinander verbunden (vgl. Atmungskette). Trägt man die Redoxpotentiale der Reaktionsfolge auf, ergibt sich eine Art Zick-Zack-Verlauf. Da dieser an ein zur Seite gedrehtes „Z“ erinnert, spricht man auch von einem Z-Schema Z-Schema der Photosynthese Nichtzyklischer Elektronentransport • Zunächst werden die im Photosystem II befindlichen Chlorophyll-Moleküle (Reaktionszentrum) durch die von den Lichtsammelkomplexen an sie weitergeleitete Lichtenergie (Exziton) auf ein höheres Energieniveau gehoben: Das Chlorophyll ist nach Aufnahme von Lichtenergie in einem angeregten Zustand. • Vom angeregten Chlorophyll a des Photosystem II werden die Elektronen zunächst auf Phaeophytin übertragen. • zurück bleibt oxidiertes P680 mit Elektronenlücke • Photosystem II füllt die Elektronenlücke durch Photolyse von Wasser auf, bei der O2 in einer Mn2+ abhängigen Reaktion generiert wird. – 2 H2O O2 + 4 H+ + 4 e- • Entstandene O2-Moleküle werden dabei an die Atmosphäre abgegeben. • Vom Phaeophytin werden die Elektronen auf QA und QB (mit verschiedenen Proteinen assoziiertes Plastochinon) übertragen. Plastochinon wird zu Plastochinol reduziert. • Redoxsystem Plastochinol/Plastochinon reduziert Cytochrom-b/f-Komplex – Cytochrom-b/f-Komplex enthält Eisen-Schwefel-Proteine, Chinon sowie Cytochrom b und f (Hämoproteine mit Eisen als Redoxcarrier). – dieser gibt Elektron auf Redoxsystem Plastocyanin ab → Cytochrom-b/f-Komplex übernimmt 2 Elektronen vom Plastochinol • Cytochrom-b/f-Komplex reduziert Plastocyanin (ein kupferhaltiges Protein) • reduziertes Plastocyanin überträgt auf Photosystem I (P700) und schließt dadurch die Elektronenlücke, die entsteht, weil durch Lichtabsorption ein Elektron des P700 auf Ferredoxin übertragen wird • Elektronenlücke des P700 wird also durch Elektron aus P680 gefüllt • Das durch Lichtabsorbtion angeregte Photosystem I gibt Elektronen über zwei Redoxcarrier (A1 und A0) Elektronen auf ein Eisen und Schwefel enthaltendes Protein (Ferredoxin) ab • Ferredoxin wird durch Übertragung von Elektronen auf NADP reoxidiert • mit O2 freigesetzte Elektronen werden über Photosystem II und I auf NADP übertragen: → nichtzyklischer Elektronentransport Zyklischer Elektronentransport • Wenn hohes NADPH/NADP-Verhältnis vorliegt wird NADP nicht reduziert, sondern Ferredoxin gibt seine Elektron direkt an das Photosystem I (P700) zurück. • Es kommt dabei zu ATP-Bildung, da Elektronen von einer Komponente mit negativerem Redoxpotential (Ferredoxin) auf Komponenten mit positiveren Potential und schließlich auf p700 zurückfließen (vgl. Atmungskette). → zyklischer Elektronentransport (zyklische Photophosphorylierung) → verläuft ohne Beteiligung von P680 → keine Sauerstoff-Entwicklung ATP-Bildung • ATP-Bildung an den Stromathylakoiden: Proteine, die aufgrund besonders strukturierter prosthetischer Gruppen zum Elektronentransport befähigt sind, bilden Komplexe mit unterschiedlichem Redoxpotential • Durch Elektronenfluss freiwerdende Redoxenergie wird zu einem transmembranen Protonentransport verwendet → Protonengradient an der Membran durch Photolyse des Wassers und durch Protonentransport durch Cytochrom-b/f-Komplex • Protonengradient wirkt bei der Rückführung der Protonen für die ATP-Synthese energieliefernd: Protonen fließen aus dem Innenraum durch die ATP-Synthase zurück ins Stroma • → Photophosphorylierung • ATP-Synthese ist nur indirekt an den Elektronentransport gebunden; sie verläuft über Protonengradientenbeteiligtes Enzym (ATPSynthase), die jedoch kein Glied der eigentlichen Elektronentransportkette darstellt. Photosynthese vs. Atmungskette Teilreaktionen der Photosynthese • Photolyse des Wassers: 2 H2O O2 + 4 H+ + 4 e- • Lichtreaktion A 2 NADP + 4 e- + 2 H+ 2 NADPH • Lichtreaktion B ADP + Pi ATP Calvin-Cyclus • Der Calvin-Zyklus ist eine zyklische Folge von chemischen Reaktionen, durch die CO2 zu Glucose und Wasser reduziert wird. • Man bezeichnet die reduktive Assimilation von Kohlenstoff aus CO2 unter Bildung von Glucose als Dunkelreaktion, weil für diesen Vorgang selbst kein Licht benötigt wird. (Allerdings werden die beteiligten Enzyme durch Licht aktiviert!!!) • Der Calvin-Zyklus läuft bei Pflanzen im Stroma der Chloroplasten ab. Untergliederung • Der Calvin-Cyclus wird in drei Teile gegliedert: – Carboxylierung – Reduktion – Regeneration Carboxylierung • Die Fixierung von CO2 wird durch das Enzym Ribulose-1,5-bisphosphat-Carboxylase katalysiert. • Substrate sind Ribulose-1,5-Bisphoshat und CO2. • Über mehrere instabile Zwischenstufe zerfällt das Produkt in zwei Moleküle 3Phosphoglycerinsäure. Reduktion • 3-Phosphoglycerinsäure (3-Phosphoglycerat) ist ein Zwischenprodukt der Glycolyse entstanden. • Im Zuge der Reduktion wird nun der umgekehrte Weg beschritten (analog zur Gluconeogenese) • Unter Verwendung des in der Lichtreaktion gewonnenen ATP wird 3-Phosphoglycerat zu 1,3-Bisphosphogycerat aktiviert. • Das resultierende Säureanhydrid wird unter Verbrauch des ebenfalls in der Lichtreaktion gewonnenen NADPH zum 3Phosphoglycerinaldehyd reduziert. • Im Zuge der weiteren Reaktionen wird Fructose-1,6bisphosphat gebildet, welches auch in andere Hexosen umgewandelt werden kann. • Die Dunkelreaktion beschreibt folgende Nettogleichung: 6 CO2 + 6 H2O + Energie C6H12O6 + 6 O2 Regeneration • Phosphorglycerinaldehyd ist nicht nur Substrat für Synthese von Hexosen, sondern auch Ausgangspunkt für Regeneration des Ribulose1,5-bisphosphats → zyklischer Prozess: Calvin-Zyklus, in dem der Akzeptor des CO2 regeneriert wird • von 12 im Calvin-Zyklus gebildeten Triosen werden 10 zur Regeneration benötigt und nur 2 Triosen für die Glucose-Bildung verwendet • Eines der wichtigsten Charakteristika des Calvin-Cyclus ist sein autokatalytischen Potential! • Darunter versteht man die Fähigkeit, bevorzugt große Mengen von Ribulose-1,5Bisphosphat anstelle von Hexosen zu bilden. • Aus der primären Steigerung des CO2Akzeptormoleküls folgt dann eine sekundäre Steigerung der Hexoseproduktion. Chemosynthese • Die Chemosynthese ist ein der Photosynthese verwandter Prozess: – Bei der Chemosynthese wird Energie in Form von ATP und NADH durch Oxidation anorganischer Substrate gewonnen. ATP und NADH werden anschließend analog zur Photosynthese dazu verwendet, im Rahmen des Calvin-Cyclus CO2 zu fixieren, also Kohlenhydrate zu bilden. • Beispiele für chemoautotrophe Lebewesen sind • Schwefelbakterien (Oxidation von H2S • Nitrifizierende Bakterien, (Oxidation von NH3, bzw. Oxidation von Nitrit NO2−) • Eisenbakterien (Oxidation von zweiwertigem Eisen Fe2+ zu dreiwertigem Eisen Fe3+) • Anaerobe Methanbakterien(Oxidation von Wasserstoff H2 mit CO2 zu H2O und CH4), z. B. in Biogasanlagen Autotrophie • Unter Autotrophie (gr. autotroph – wörtlich: „sich selbst ernährend“ von autos – „selbst“, trophe – „Ernährung“) wird in der Biologie die Fähigkeit von Lebewesen verstanden, ihre Baustoffe (und organischen Reservestoffe) ausschließlich aus anorganischen Stoffen aufzubauen • Der Gegensatz zur Autotrophie ist die Heterotrophie, bei der organische Verbindungen zum Aufbau der Baustoffe verwendet werden. Der Mensch, Tiere, Pilze und die meisten Bakterien und Archaeen sind heterotroph. Sie werden den Konsumenten, spezieller den Herbivoren (Pflanzenfresser), Carnivoren (Fleischfresser) und Omnivoren (Allesfresser) sowie den Destruenten zugeordnet. Photoautotrophie • Photoautotrophie ist die Nutzung von Licht als Energiequelle bei Autotrophie. Lebewesen mit dieser Fähigkeit nennt man photoautotroph. Fast alle Pflanzen und Algen sowie einige Bakterien wandeln im Zuge der Photosynthese Lichtenergie in chemische Energie um die sie zum Aufbau von Bau- und Reservestoffen aus anorganischen Stoffen verwenden. Chemoautotrophie • Chemoautotrophe Organismen benutzen wie die phototrophen Organismen CO2 als einzige Kohlenstoffquelle, aber Energielieferant ist nicht Licht, sondern eine anorganische Verbindung, bei deren Oxidation die nötigen Reduktionsäquivalente bereitgestellt werden.