Teil VI - Tired Joe
Werbung

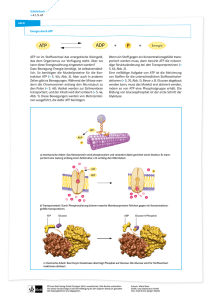
Historische Experimente zur Ernährung der Pflanzen Schon im Jahre 1771 führte der englische Naturforscher und Geistliche Joseph Priestley folgende Experimente durch. Bei ihrer Darstellung wollen wir uns auf die wesentlichsten Daten beschränken. a) Setzte er eine Maus und stellte er eine Kerze in zwei getrennte, luftdichte Glasgefäße, so starb die Maus, und die Kerze erlosch. b) Eine in einem luftdichten Glasgefäß gehaltene Pflanze starb ebenfalls nach einiger Zeit. c) Befanden sich die Maus und Pflanze im selben luftdichten Glasgefäß, so überlebten sie beide. d) Brachte er eine Pflanze in eine Luftmenge, in der eine Kerze erloschen war (a), so gedieh die Pflanze (b), und nach einer Weile konnte eine Kerze wieder gut in dieser Luft brennen (c). Die Ernährungsweise der Pflanzen besteht also darin, dass sie aus energiearmen anorganischen Stoffen (Wasser, Kohlendioxid) energiereiche Nährstoffe synthetisieren und dabei Sauerstoff frei wird. Bedeutung der Photosynthese Autotrophe Pflanzen Stoffaufbau durch Energiebindung Produktion von 170 Billionen t Biomasse/a Ernährungsgrundlage für alle heterotrophen Lebewesen Sauerstoffproduktion für alle aeroben Lebewesen Seite 139 Florian Zeller 07/08 KOHLENSTOFFKREISLAUF bzw. Materiekreislauf Produzenten anorganische Verbindungen organische Substanz CO2 O2 Destruenten Konsumenten abgestorbenes org. Material Ökosysteme: Areal in 106 km2 17,0 322,0 15 Tropischer Regenwald Hochsee Savannen Pflanzenbiomasse in 109 t C 344,0 0,45 27 Nettoprimerproduktion in gC/m3/Jahr 988 56 407 ATP ist das Cosubstrat aller Kinasen ! P EP Glucose – 6 – P ATP Apoenzym Pyruvatkinase ADP ATP Apoenzym Pyruvatkinase BTS ADP Glucose Substrat 1 NADH2 Produkt 2 Apoenzym NAD Produkt 1 NADH2 NAD Apoenzym Substrat 2 NAD ist das Cosubstrat aller Dehydrogenasen Seite 140 Florian Zeller 07/08 Nicotinsäureamid-Adenin-Dinucleotid Enzym 8 NAD+ Wasserstoffübertragendes Coenzym (nur die Bedeutung erkennen!) reaktive Stelle NADP+ Nicotinamid Nicotinamid Ribose P P Ribose Photosynthese Gleichung: 6 CO2 + 6 H2O Licht Chlorophyll C6H12O6 + 6 O2 Formeln von Chlorophyllen und Carotin Porphyringerüst aus 4 Pyrrolringen (I – IV) mit zentralem Magnesiumatom Zeichnen können!! 𝛽-Carotin Seite 141 Florian Zeller 07/08 Zeichnen können!! Chloroplasten im Lichtmikroskop Eine von mehreren Modellvorstellungen zum Aufbau der Thylakoidmembran eines Chloroplasten. Die Quantasomen werden hier als Membranbauteile betrachtet. Bestrahlung mit weißem Licht Rohchlorophyll-Lösung Kontinuierliches Spektrum Aller Spektralfarben reflektiertes Licht kein kontinuierliches Spektrum Spektrum mit Lücken bei bestimmten Wellenlängen absorbiert Licht bestimmten Wellenlängen Eigenfarbe: Mischfarbe aus den nicht absorbierten Spektralbereichen Seite 142 Florian Zeller 07/08 (mehr dazu auf Seite 144) Chlorophyllextraktion - zerkleinerte Blätter Seesand Alkohol Lösungsmittel gemischt Aceton Homogenisieren und Extrahieren Papierchromatographie Die verschiedenen Farbstoffe werden aufgetrennt: Carotinoide Xanthophylle Chlorophylle (a+b) Gemeinsamkeit von Chlorophyll a/b und Carotinoiden: SYSTEM KONJUNGIERTER DOPPELBINDUNGEN -Elektronen leicht anregbar, wichtig für die Lichtabsorption!! Seite 143 Florian Zeller 07/08 Lichtabsorption durch Pigmente Sichtbares Licht ist elektromagnetische Strahlung der Wellenlänge 400 – 750 nm. kontinuierliches Spektrum Werden aus weißem Licht durch Pigmente (Farbstoffe) bestimmte Wellenlängenbereiche Absorbiert (verschluckt), so sehen wir den restlichen reflektierten Wellenlängenbereich bunt. Im sogenannten Spektralphotometer kann man messen, welche Wellenlängenbereiche ein Farbstoff genau absorbiert, z.B. Chlorophyll a und b sowie die Carotinoide. Das Ergebnis ist graphisch darstellbar als Absorptionsspektrum. Chlorophyll a,b: g Absorption im kurzwelligen Blaubereich und im langwelligen Orange/Rotbereich, rünes Licht wird durchgelassen. Chlorophylle „sind“ grün. β-Carotin, Xanthophylle: Absorption im blaugrünen Wellenlängenbereich, gelb/oranges Licht wird durchgelassen; diese Farbstoffe „sind“ gelb/orange Anmerkung: Chlorophyll a = Hauptpigment der Photosynthese Chlorophyll b + Carotinoide (Carotin und Xanthophylle) = Hilfspigmente (accessorische Pigmente) Nun kann man auch die Intensität der Photosynthese bei verschiedenem einfarbigem Licht messen. Der Zusammenhang zwischen den Wellenlängen des eingestrahlten Lichtes und den Photosyntheseleistungen der bestrahlten Pflanze wird durch das sog. Wirkungsspektrum wiedergegeben (=Aktionsspektrum) Spektrale Absorption von Photosynthesepigmenten und Wirkungsspektrum der Photosynthese Aus den Kurven ist ersichtlich, dass die Chlorophylle im kurzwelligen blauen und im langwelligen roten Spektralbereich absorbieren. Der mittlere grüne Spektralbereich wird durchgelassen. Carotin absorbiert im blaugrünen Wellenlängenbereich, weshalb es uns in der Komplementärfarbe, in Rot, erscheint. Misst man die Intensität der Photosynthese bei verschiedenfarbigem Licht, so findet man eine ungefähre Übereinstimmung des Wirkungsspektrums mit den Adsorptionsspektren der drei Pigmente, woraus man auf ihre Mitwirkung bei der Photosynthese schließen kann. Seite 144 Florian Zeller 07/08 Modellvorstellung der Lichtabsorption: die Lichtsammelfalle Antennenpigmente: Carotinoide und Chlorophyll b absorbieren Lichtquanten Lichtweiterleitende Pigmente: Chlorophyll a Reaktionszentrum: Chlorophyll a Molekül energie armes e- energie reiches e- Dieses angeregte Elektron wird an ein Empfängermolekül (Acceptor) zur Leistung von chemischer Arbeit abgegeben! Das Chlorophyll a Molekül wurde dabei ionisiert! Grüne Pflanzen haben zwei Typen von Lichtsammelfallen mit Wirkung- bzw. Absorptionsmaxima bei Licht der Wellenlänge 700 nm = Photo- oder Pigmentsystem I, kurz P700 682 nm = Photo- oder Pigmentsystem II, kurz P682 Die unterschiedlichen Eigenschaften von P700 und P682 liegen wohl daran, dass beide unterschiedliche Chlorophyll-Protein-Komplexe darstellen. Ablauf der Photosynthese Gleichung: 1. 6 CO2 + 6 H2O hv C6H12O6 + 6 O2 ∆G‘ = + 2880 kJ/mol Lichtabhängige Reaktion (Primärprozess) am Thylakoidsystrem SPALTUNG von Wasser durch hv-Energie = PHOTOLYSE des WASSERS Lichtreaktion der Photosynthese Kreuzungsexperiment von Kamen und Ruben 1941 Tracer Methode zur Aufklärung der Herkunft des Sauerstoffs. Verwendet wurden Verbindungen, die mit dem schweren Sauerstoffisotop 18O markiert waren „Kreuzung“: in zwei getrennten Experimenten Markierung - einmal von H2O - einmal von CO2 mit schwerem (Achtung: nicht radioaktivem) Sauersoff !!! Licht Experiment 1: 6 CO2 + 12 H218O C6H12O6 + 6 18O2 + 6 H2O Chlorophyll Bei Verwendung von isotopenmarkiertem Wasser tritt das Isotop nur im freiwerdenden Sauersoff auf! Experiment 2: Licht 6 C18O2 + 12 H2O C6H1218O6 + 6 O2 + 6 H218O Chlorophyll Bei Verwendung von isotopenmarkiertem Kohlendioxid tritt das Isotop im Kohlenhydrat und im neu entstandenen Wasser auf. Nun war endgültig bewiesen, dass der Sauerstoff aus der Photolyse des Wassers stammt: 𝟏 H2 O + - 2H + 2e + 𝟏 𝟐 𝟐 O2 O2 H+ e- H+ e- Seite 145 Florian Zeller 07/08 Die Wasserspaltung findet an der Innenseite der Thylakoidmembran statt. Die e- werden blitzschnell entzogen und mittels Elektronentransportketten = Redoxsysteme durch die Membran hindurch weitergeleitet. Die H+ werden in den Thylakoidinnenraum abgegeben O2 verlässt den Chloroplasten - Für den Beweis zweier „unabhängiger“ Reaktionssysteme (Lichtreaktion, Dunkelreaktion) arbeiteten u. a. 1940 Herr Hill und sein Team, 1941 Kamen und Ruben und ihr Team. Hill-Reaktion 1940 Beweis, dass die Sauerstoffproduktion der Photosynthese nicht an das Vorhandensein von Kohlendioxid gebunden ist. O2 stammt nicht aus CO2, sondern aus H2O !! Experiment: Belichtung aus lebenden Zellen isolierte Chloroplasten Wasser reduzier bare Substanz (Fe3+-Ionen) Ergebnis: viel O2 entsteht! Erklärung: Wasser wird durch Licht und mit Hilfe der Chloroplasten in Protonen, Elektronen und Sauerstoff zerlegt. Die reduzier bare Substanz nimmt dabei die Elektronen auf und wird zur reduzierten Substanz. H2 O Licht Chlorophyll Elektronenacceptor Gesamt: = H2O + 2 Fe3+ 2 H+ + 2 e- + reduzier bare Substanz (hier Fe3+) 𝟏 𝟐 + O2 reduzierte Substanz (hier 2 Fe2+) 2 e- 2 Fe2+ + 2 H+ + 𝟏 𝟐 O2 Im Hill-Versuch wurden als reduzier bare Substanz Fe3+-Ionen verwendet, bei der Photosynthese ist die reduzier bare Substanz NADP (eigentlich NADP+) !!! Seite 146 Florian Zeller 07/08 Gesamtgleichung der Photosynthese 6 CO2 + 12 H2O Licht Chlorophyll C6H12O6 + 6 O2 + 6 H2O Elektronentransport und Photophosphorylierung zeichnen können!!! Anmerkung: Redoxsysteme = Enzyme = e --Transprotketten Bilanz: 12 H2O 24 H+ + 24 e- + 6 O2 + 12 NADP + 24 H + 24 e12 NADPH2 (Reduktionsäquivalente) 18 ADP + 18 P 18ATP (Energieäquivalente) 12 H2O + 12 NADP + 18 ADP + 18 P 12 NADPH2 + 18 ATP + 6 O2 Seite 147 Florian Zeller 07/08 Lichtabhängige Reaktion am Fotosystem I (P700) Durch Lichtabsorption wird das Reaktionszentrum in P 700 angeregt Chlorophyll a Molekül gibt Elektronen ab, diese werden auf einen Zwischenempfänger = Stoff Y gehoben. Das Empfängermolekül überträgt die Elektronen auf den endgültigen Akzeptor NADP. Lichtabhängige Reaktion am Fotosystem II (P682) Durch die Elektronenabgabe am P700 ist dort eine Elektronenlücke (Kationencharakter nachweisbar!) entstanden, diese muss gefüllt werden. Die Füllung der Lücke erfolgt durch das Fotosystem II: Wiederum Absorption von Lichtenergie Anregung des Reaktionszentrums P682 Chlorophyll a Molekül gibt Elektronen ab werden auf den Zwischenakzeptor Stoff X gehoben und von dort über eine besondere Elektronentransportkette in das P700 geleitet Lücke gefüllt! Fotolyse des Wassers Nun ist eine Elektronenlücke im P682!! Dies wird gefüllt durch chemische Spaltung des Wassers in Sauersoff, Protonen und Elektronen! Der Sauerstoff wird abgegeben. Die Elektronen werden zum P682 weitergeleitet und füllen dort die Lücke. Die Protonen werden auf andere Weise wie die Elektronen zum NADP transportiert! Fotophosphorylierung bzw. chemoiosmotische ATP-Bildung Die ATP-Bildung erfolgt ähnlich wie in der Endoxidation an einem ATP-Syntheseprotein in der Thylakoidmembran. Die Energie zur ATP-Bildung wird aus dem Konzentrationsgradienten der Protonen zwischen Thylakoidinnenraum und Matrix (= Stroma) geschöpft. Messungen des pH-Wertes im Thylakoidinnenraum ergaben eine Differenz von 2 pH-Einheiten zum Stroma, d.h. innen ist die Konzentration der H+ um das 100-fache höher! Folge: H+ diffundieren von innen nach außen durch das ATP-Syntheseprotein, die dabei frei werdende Energie wird zur ATP-Synthese genutzt. Die Aufrechterhaltung des Protonengradienten erfolgt 1. durch das Fotosystem II, das ständig auf das Wasser einen so starken Elektronensog ausübt, dass Wasser gespalten wird H+-Nachschub 2. Auf dem Weg der e- von X zu P700 wird Energie frei, diese nutzt ein Stoff namens Plastochinon zum Rücktransport der ausgetretenen H+ in den Thylakoidinnenraum H+-Nachschub. Da 1. und 2. Nur bei Belichtung funktionieren, also letztendlich das Licht die Triebkraft der ATP-Bildung darstellt, kann man auch von Fotophosphorylierung sprechen = „Nicht zyklische Fotophosphorylierung“ „Zyklische Fotophosphorylierung“ Die Lichtanregung am P700 wird ausgenützt. Die Elektronen wandern vom Stoff Y aus nicht zum NADP, sonder über andere Redoxsysteme und das Plastochinon zurück zum P700. Plastochinon nutzt die energie und pumpt H+ nach innen, H+-Gradient wird erhöht, H+ diffundieren durch das ATP-Syntheseprotein nach außen, ATP kann gebildet werden. Hierin liegt wohl der Sinn der zyklischen Fotophosphorylierung! Zusätzliche ATP-Bildung!!! Achtung: hier läuft nur dann die Lichtreaktion I ab keine Wasserspaltung, daher keine O2-Bildung keine NADPH2-Bildung! Seite 148 Florian Zeller 07/08 2. Lichtunabhängige Reaktion = REDUKTION von KOHLENDIOXID zu TRAUBENZUCKER Dunkelreaktion der Photosynthese Voraussetzungen für die Reduktion von CO2 zum energiereichen Kohlenhydrat: 1. Reduktionsmittel: NADPH2 = Wasserstoffübertragendes Coenzym 2. Energie: ATP als Speicher und Überträger von Energie Adenosin – P ~ P ~ P P = Phosphatgruppe ~ = energiereiche Bindung Umwandlung von Kohlendioxid zu Kohlenhydraten Diese Reaktionen laufen normalerweise im Licht neben den Lichtreaktionen ab, können aber im Dunklen noch weiterlaufen, solange der Vorrat an ATP und NADPH2 aus der Lichtreaktion noch reicht. Die Reaktionen können im Dunkeln ständig ablaufen, wenn man künstlich ATP und NADPH2 zugibt. Mevin Calvin (Nobelpreis 1961) hat experimentell den Weg des CO2 von der Aufnahme durch die Pflanze bis zum fertigen Kohlenhydrat durch das Verfahren der Autoradiographie verfolgt: Grünalgen der Gattung Chlorella wurde *CO2 ( * = mit radioaktivem 14-C markiert) angeboten, bei Belichtung bauen die Algen dies ein. Unterbrochen wird die Photosynthese-Tätigkeit zu verschiedenen Zeitpunkten durch Übergießen mit siedendem Alkohol. Nun kann man in verschiedenen zeitabständen die Photosynthese-Zwischenprodukte, die ja *14 C enthalten, isolieren und untersuchen. 1. 2. 3. Extraktion der Intermediärprodukte Chromatographie der Intermediärprodukte: Die verschiedenen Substanzen laufen verschieden weit Auftrennung; Substanzen farblos, man sieht nichts Sichtbarmachen der IMP: Auflegen eines Filmstreifens, Stellen mit radioaktivem Verbindungen (*C!) schwärzen den Film, nun weiß man, wo die IMP auf dem Chromatogramm sind! Seite 149 Florian Zeller 07/08 4. 5. Auswaschen der IMP aus dem Chromatogramm Chemische Identifikation (nicht den Filmstreifen) Bereits 5 Sekunden nach Zugabe von *CO2 sind viele verschiedene Substanzen entstanden, z.B. Saccharose und andere Zucker. Calvin verkürzte schrittweise die Versuchszeit, bei einem Stopp bereits nach 1,7 Sekunden erhielt er einen einzigen *C-haltigen Stoff, das Glycerinsäure-3-Phospaht bzw. 3-Phosphoglycerinsäure = 3-PGS. Dies ist also das erste fassbare IMP der Photosynthese. Wo aber kommt die Phosphoglycerinsäure mit ihren 3 C-Atomen her? Weitere Versuche zeigten, dass der Primäre Acceptor für CO2 ein C5-Körper mit dem klangvollen Namen RuDP ist, d.h. Ribulose-1,6-Diphosphat, welches in der Dunkelreaktion in einem Kreisprozess, dem Calvin-Zyklus ständig nachgeliefert wird. 3-PGS Temperaturabhängigkeit der Photosynthese ATP NADPH2 In den Lichtreaktionen wird so wenig „Rohmaterial“ hergestellt, dass dessen völlige „Aufarbeitung“ bereits bei niedrigen Temperaturen erfolgt. Eine gesteigerte Aktivität in der Dunkelabteilung bringt nichts ein. Anders liegen die Verhältnisse bei Starklicht: ATP NADPH2 Seite 150 Florian Zeller 07/08 Fotochemischer Wirkungsgrad Wie viele Elektronenanregungen sind zur Herstellung von einem Mol Glucose notwendig? Pro Bildung von einem NADPH2 müssen an zwei Lichtsammelfallen je zwei Elektronen angeregt werden 4 Elektronenanregungen notwendig Da 12 NADPH2 24 Elektronenanregungen durch Licht notwendig. Energie von einem Lichtquant: Energie von 48 Lichtquanten: ca. 200 kJ/mol ca. 9600 kJ/mol In einem Mol Glucose sind aber nur 2872 kJ/mol Energie gespeichert. Wie viel % der eingesetzten Energie sind das? 9600 kJ = 100 % 2872 kJ = x 𝑥= Wie groß ist der fotochemische Wirkungsgrad? Der maximale Wirkungsgrad 𝜂 = 1; 2872 𝑘𝐽 ×100% 9600 𝑘𝐽 = 𝟐𝟗, 𝟗𝟐 % 9600 kJ = 1 2872 kJ = x 𝑥 = 2872 ÷ 9600 = 𝟎, 𝟑 𝜂 = 0,3 Die Dunkelreaktion der Photosynthese ist also enzymgesteuert und temperaturabhängig! - die Prozesse laufen im Stroma ab - lichtunabhängig - jedoch werden bei der Pflanze die Energieäquivalente und die Reduktionsäquivalente – also die 18 ATP und die 12 NADPH2 – aus der Lichtreaktion gebraucht. Die Dunkelreaktion lässt sich in drei Phasen gliedern: 1. Fixierungsphase: 2. Reduktionsphase: 3. Regenerationsphase: Bindung des CO2 an einen Acceptor (C5-Körper), über ein instabiles Zwischenprodukt entstehen Glycerinsäurephosphatmoleküle. Glycerinsäurephosphat wird unter ATP-Verbrauch und mit Hilfe des Reduktionsmittels NADPH2 zu Glycerinaldehydphosphat reduziert. NADPH2 wird dabei oxidiert zu NADP. Der Acceptor wird im Kreisprozess unter erneutem ATP-Verbrauch regeneriert. Bilanzmäßig lässt sich 1 Molekül energiereicher Glucose abzapfen. 6 CO2 6 H2O 6 C5 Ribulose 1,5 di – P 6 [C6 ] 12 ATP 12 C3 Glycerinsäure-P 12 ADP + 12 P 12 H2O 12 C3 12 NADPH2 12 NADP 6 ADP + 6 P C6 Glucose CALVIN-BENSON-ZYKLUS 6 ATP 6 C5- P 6 CO2 + 18 ATP + 12 NADPH2 Saccharose 10 C3- P Stärke, Cellulose C6H12O6 + 18 ADP + 18 P + 12 NADP + 6 H2O Seite 151 Florian Zeller 07/08 Zentraler Reduktionsschritt: 12 ATP 12 ADP + 12 P 12 NADPH2 12 NADP Glycerinsäure-3- P 3 PGS 6 CO2 + 12 H2O* Glycerinaldehyd-3- P 3 PGA C6H12O6 + 6 O2* + 6 H2O Beeinflussung der Photosyntheseleistung durch Außenfaktoren Photosyntheserate ist abhängig von 1. Lichtintensität (Beleuchtungsstärke) 2. Lichtqualität (𝜆 des Lichts = Farbe des Lichts) 3. Kohlendioxidgehalt der Luft 4. Temperatur 5. Luftfeuchtigkeit zu 1. Lichtintensität: Man unterscheidet zwischen Brutto- und Nettophotosyntheserate Brutto-Photosynthese-Rate = Photosynthesegesamtleistung Netto-Photosynthese-Rate = Bruttophotosyntheserate – Glucoseverbrauch bei der Atmung Tatsächlicher Glucosegewinn (Gewinn an Biomasse) Einfluss der Lichtintensität: - im Dunkeln: Pflanze betreibt Atmung - bei bestimmter Lichtintensität: Lichtkompensationspunkt K erreicht, d.h. Atmung (Glucoseverbrauch) und Bruttophotosynthese (Glucoseentstehung) kompensieren sich (heben sich auf). - Lichtintensität darüber hinaus: Nettogewinn setzt ein !! Diagramm 1: Abhängigkeit der Nettophotosynthese von der Beleuchtungsstärke bei Schattenpflanzen und Sonnenpflanzen Seite 152 Florian Zeller 07/08 zu 2. Lichtqualität vergleiche Wirkungsspektrum der Photosynthese zu 3. Kohlendioxidgehalt der Luft ist mit 0,03 % der sog. Minimumfaktor (= jener abiotischer Umweltfaktor, der die Photosyntheseleistung unter natürlichen Bedingungen begrenzt). Diagramm 2: Abhängigkeit des Pflanzenwachstums vom CO2-Gehalt der Luft; Unterschiedliche CO2-Toleranz verschiedener Pflanzen Jede Pflanze zeigt ein - CO2-Maximum (sehr hohe Konzentrationen nicht tolerierbar) - CO2-Optimum ( Konzentration etwas über dem normalen CO2-Gehalt der Luft) - CO2-Minimum: Bohne: stellt bei erniedrigter CO2-Konzentration das Wachstum bald ein. Mais: toleriert geringe CO2-Konzentration, verlangsamt lediglich das Wachstum!! Ursache: Mais, andere Gräser, Kakteen (allg. Pflanzen trockener Standorte) sind Wegen hoher Außentemperaturen und damit verbundener Gefahr zu hoher Verdunstung oftmals gezwungen, die Stomata zu schließen wenig CO2-Aufnahme möglich !! Trotzdem können diese Pflanzen Photosynthese betreiben: Ihr Stoffwechsel ist an die verringerte Aufnahmemöglichkeit von CO 2 angepasst. Das bei der Atmung entstehende CO2 kann chemisch gespeichert werden und in die Photosynthese eingeschleust werden. Seite 153 Florian Zeller 07/08 zu 4. Temperatur Diagramm 3: Temperatur –Toleranz Die Reaktionsgeschwindigkeit nimmt allgemein bei enzymatischen Reaktionen mit steigender Temperatur zu, bis Hitzedenaturierung der Enzyme für Abfall der Reaktionsgeschwindigkeit sorgt, schließlich kommt das gesamte Stoffwechselgeschehen zum erliegen! Allgemein: Zunahme der Photosynthese mit steigender Temperatur, Einstellung der Photosynthese bei zu hohen Temperaturen Jede Pflanze hat Temperaturminimum, -optimum, -maximum! Jenseits von Minimum und Maximum stellt die Pflanze die Stoffproduktion ein und stirbt. zu 5. Luftfeuchtigkeit Luftfeuchtigkeit muss genügend vorhanden sein. Erhöhung kann sich positiv auf die Photosyntheserate auswirken! Analyse der Photosynthesefaktoren Summarisch lässt sich der Photosyntheseprozess in folgender Bruttogleichung darstellen: Pro Mol erzeugter Glucose wird eine Energiemenge von 2825 kJ benötigt. C6H12O6 Chlorophyll Photosynthese (Schema) Chlorophyll a und b Seite 154 Florian Zeller 07/08 Seite 155 Florian Zeller 07/08 Vorgehensweise: kurze: 6 CO2 + 6 H2O erweiterte: 6 CO2 + 12 H2O Lichtreaktion Dunkelreaktion Licht Chlorophyll Licht Chlorophyll C6H12O6 + 6 O2 C6H12O6 + 6 O2 + 6 H2O Lichtreaktion: 12 H2O 24 H+ + 24 e- + 6 O2 + 12 NADP + 24 H + 24 e12 NADPH2 18 ADP + 18 P 18 ATP 12 H2O + 12 NADP + 18 ADP + 18 P 12 NADPH2 + 18 ATP + 6 O2 Dunkelreaktion: 6 CO2 + 18 ATP + 12 NADPH2 ausgleichen: 6 CO2 + 12 H2O Es ist schön, autotroph zu sein … C6H12O6 + 18 ADP + 18 P + 12 NADP + 6 H2O C6H12O6 + 6 O2 + 6 H2O … , zeitweise wenigstens. autotrophe Organismen (autotroph = „sich selbst ernährend“) = Organismen, die energiereiche organische Stoffe durch Photosynthese aus einfacheren anorganischen Verbindungen aufbauen können und damit von energiereichen organischen Nährstoffen unabhängig sind. heterotrophe Organismen (heterotroph = „sich von anderen ernährend“) = Organismen, die hinsichtlich ihres Stoffwechsels auf die Zufuhr energiereicher organischer Stoffe angewiesen sind. Diese werden entweder in körpereigene Stoffe umgewandelt oder unter Energiefreisetzung abgebaut. Seite 156 Florian Zeller 07/08 Die Farben kennzeichnen die Reaktion der Verdauungssäfte: grün = neutral rot = sauer blau = alkalisch Organ Abbau Enzym Mundhöhle Stärke Maltose ∝-Amylase (Ptyalin) Magen Proteine Polypeptide Pepsin, Kathepsin (Schleim, HCl) Stärke Maltose Maltose Glucose ∝-Amylase Maltase Nucleiosom Nucleotide Nucleasen Proteine Polypeptide Fette Polypeptide Aminosäuren Glycerin + Fettsäuren Trysin, Chymotrypsin Proteasen Lipasen Dünndarm Seite 157 Florian Zeller 07/08 Energiefreisetzung durch Abbauvorgänge AEROBER STOFFABBAU Mitochondrium unter dem Elektronenmikroskop mit O2 Bau eines Mitochondriums (zeichnen können!) Übersicht über die Vorgänge beim STOFFABBAU = Dissimilation Glucose C6 1. GLYCOLYSE Brenztraubensäure 2 x C3 CO2 2. Oxidative Decarboxylierung Aktivierte Essigsäure C2 3. Citronensäurecyclus CO2 CO2 NADH2 FADH2 4. Sauerstoff vom Atmen braucht man nur bei Schritt 4 !!! 2H 1 O 2 2 H2O Endoxidation: Hauptsächliche Energiequelle ! ATP !! Seite 158 Florian Zeller 07/08 anaerober Abbau Glykolyse aerober Abbau C6H12O6 Milchsäure-Gärung Oxid. Decarboxil. BTS 2 BTS 2 BTS Milchsäure 2 NADH2 2 ATP 2 Acetyl-CoA + 2 CO2 Rückgewinnung NAD 2 NADH2 alkoh. Gärung Citratcyclus BTS 2 Acetyl-CoA Ethanol + CO2 4 CO2 Rückgewinnung NAD 6 NADH2 2 FADH2 2 GTP Endoxydation 10 NADH2 2 FADH2 + 6 O2 12 H2O 34 ATP Seite 159 Florian Zeller 07/08 Bilanzgleichung für den Oxydativen Abbau C6H12O6 + 6 O2 38 ADP + 38 P 1. 6 CO2 + 6 H2O 38 ATP Glycolyse Reaktionskette ohne O2-Verbrauch im Zytoplasma. Sehr alter Stoffwechsel !!! - Läuft bei allen Lebewesen gleich ab - Ablauf im Zytoplasma, denn zu Zeiten der Prokaryonten gab es noch keine Mitochondrien. - Anaerober Ablauf der Glykolyse, denn ursprünglich kein O2 in der Atmosphäre O Bilanz: H3C – C – COOH + 2 ATP + 2 NADH2 BTS 2 C3-Körper = Pyruvat C6H12O6 + 2 NAD + 2 ADP + 2 P Vereinfachte Darstellung der Glycolyse (können müssen!) Glucose C6H12O6 PHOSPHORYLIRUNG und ISOMERISIERUNG ATP C6 Körper ADP Clucose – 6 – P C6 ~ P Fructose – 6 – P C6 ~ P ATP ADP Fructose – 1,6 – diphosphat P ~ C6 ~ P 2 Glycerinaldehyd – 3 – P 2 C3 Körper ~ P SPALTUNG 2 ADP 2 H2 O 2P 2 ATP 2 NAD OXIDATION 2 NADH2 2 C3 ~ P 2 Glycerinsäure – 3 – P ( 2 H2O) 2 Phosphoenolbrenztraubensäure 2 ADP 2 C3 ~ P 2 ATP 2 x Brenztraubensäure 2 C3 Körper Seite 160 Florian Zeller 07/08 Formulierung des zentralen Oxidationsschritts mit Strukturformel 2 NAD 2 2 NADH2 + 2 H2O 2 ADP + 2 P 2 ATP (nur zum Verständnis!!!) Bilanz Glycolyse: C6H12O6 + 2 NAD + 2 ADP + 2 P O 2 CH3 – C – COOH + 2 ATP + 2 NADH2 Seite 161 Florian Zeller 07/08 2. Oxydative Decarboxylierung Die BTS wandert in die Mitochondrien (durch eine Doppelmembran aber nur noch halb so groß!). Aus dem C3-Körper (BTS) entstehen C2-Körper (Essigsäure) unter Abspaltung von CO2 wird mit Hilfe eines Enzyms, dem HS-Coenzyms A in Schwung gebracht, d.h. aktiviert „aktivierte Essigsäure“ = Acetyl-Coenzym A (spielt im GESAMTEN Zellstoffwechsel eine GANZ entscheidende Rolle!!!) GEWINN von 2 NADH2 !!!! Bilanz: O 2 CH3 – C – COOH + 2 NAD + 2 H - S CoA 2 CH3 – C O + 2 CO2 + 2 NADH2 S CoA BTS (= Pyruvat) 3. Acetyl-Coenzym A Zitronensäurezyklus = Tricarbonsäurecyclus (TCC) = Krebs-Martius-Cyclus - Reaktionsort in den Mitochondrien Abbau des C2-Körpers Acetyl-Coenzyms A zum CO2-Molekülen Bildung von Reduktions- und Energieäquivalenten (NADH2) (ATP) Beginn: Der Acetylrest der aktivierten Essigsäure wird auf einem C4-Körper (Oxalessigsäure) übertragen C6-Körper entsteht = Zitronensäure. (können müssen!) Seite 162 Florian Zeller 07/08 Da beim Abbau von 1 Molekül Glycose 2 Acetyl-CoA entstehen: Bilanz: 2 + 2 ADP + 2 P + 6 NAD + 2 FAD + 6 H2O 4 CO2 + 2 ATP + 6 NADH2 + 2 FADH2 + 2 HSCoA (nur zum Verständnis!) Seite 163 Florian Zeller 07/08 4. Endoxidation = ATMUNGSKETTE = Oxidative Phosphorylierung In der Endoxidation wird „gewechselt“: pro NADH2 FADH2 3 ATP 2 ATP Die reduzierten Koenzyme NADH2 und FADH2 schleusen ihren Wasserstoff H in die Endoxidation ein. NAD und FAD werden dabei zurückgewonnen. Spaltung des Wasserstoffs in H+ und e- ; die e- werden über Elektronentransportketten zum endgültigen Acceptor Sauerstoff O2 übertragen 2 H + + 2 e- + 𝟏 𝟐 O2 H2O Anmerkung: Bei der Endoxidation spielen ähnlich wie bei der Lichtreaktion der Photosynthese ein Membrangebundener Protonengradient und ein ATP-Syntheseprotein eine Rolle. ATP-Bilanz für 1 Molekül Glycose in der Endoxidation 1. Glycolyse: 2 NADH2 6 ATP 2. Oxid. Dec.: 2 NADH2 6 ATP 3. Zitronenzy.: 6 NADH2 2 FADH2 18 ATP 6 ATP direkt 2 ATP 2 ATP (=GTP) = 38 ATP andere Ausführung: pro NADH2 3 ATP; pro FADH2 2 ATP; da 10 NADH2 da 2 FADH2 30 ATP aus Atmungskette 4 ATP 2 ATP direkt aus Glycolyse 2 ATP direkt aus Citratzyklus 38 ATP Die Enzyme für die Atmungskette sitzen in der inneren Mitochondrienmembran. Was passiert nun? Der in den reduzierten Coenzymen NADH2 und FADH2 gebundene Wasserstoff (also kein Wasserstoffgas H2) wird auf Sauerstoff übertragen, Wasser wird gebildet. Die dabei stufenweise frei werdende Energie (nicht schlagartige, explosive Freisetzung von Wärmeenergie wie bei der Knallgasreaktion) kann zum großen Teil zur Herstellung von ATP aus ADP + P genutzt werden. Ähnlich wie bei der Lichtreaktion der Photosynthese spielen ein H+-Gradient, – hier an der inneren Mitochondrienmembran und Membranporen (= ATP-Syntheseproteine) eine Rolle. Seite 164 Florian Zeller 07/08 NADH2 lädt den Wasserstoff unter Spaltung in 2 H+ und 2 e- auf Redoxsystem 1 ab, die Protonen und die Elektronen wandern zum Redoxsystem 2, auf dem FADH2 nach Art des NADH2 sein Wasserstoff ablädt. Die reduzierten Coenzyme sind jetzt wieder oxidiert und können in Glycolyse, oxidativer Phosphorylierung oder im Citratzyklus neu beladen werden, d.h. wieder reduziert werden. Das Redoxsystem 2 pumpt die e- zu Redoxsystem 3, wo sie dann für die Wassersynthese zur Verfügung gestellt werden; alle H+ werden in den Intermembranraum (= Lumen) gepumpt, es kommt zur Anreicherung der Protonen im Lumen. In der Matrix sind vergleichsweise wenig Protonen. Daher können nun die H + durch die Membranporen vom Lumen in die Matrix diffundieren, dabei stellt die ATP-Synthase ATP her. Die H+ reagieren zusammen mit den e- und Sauerstoff zu Wasser. Stoffbilanz der Atmungskette: 10 NADH2 + 2 FADH2 + 6 O2 + 34 ADP + 34 P 10 NAD + 2 FAD + 12 H2O + 34 ATP aerob !!!!! Die Änderung der freien Enthalpie bei der Endoxidation beträgt pro NADH 2 etwas über 210 kJ/mol. Da für die Synthese von 1 mol ATP ca. 30 kJ notwendig sind, könnten rein rechnerisch 7 mol ATP entstehen. Tatsächlich gebildet werden aber nur 3 mol ATP/NADH2, die Restwärme kann z.B. zur Aufrechterhaltung der Körpertemperatur genutzt werden. Die Bildung von 3 mol ATP entspricht einem Wirkungsgrad von ca. 40 % technische Wärmekraftwerke haben einen Wirkungsgrad zwischen 20 und 35 %. Anmerkung: (folgende Begriffe muss man können!!) Oxidierte Coenzyme (photosyn) NAD FAD NADP reduzierte Coenzyme NADH2 FADH2 NADPH2 energiereiche Triephosphate = ATP Reduktionsäquivalenten = NADH2 & FADH2 Seite 165 Florian Zeller 07/08 Übersicht zum Nachschauen Seite 166 Florian Zeller 07/08 Übersicht des aeroben Glucoseabbaus Glucose (C6) 1. Schritt: Glycolyse 2 ATP 2 NADH2 2 x BTS (C3) 2. Schritt: Oxidative Decarboxylierung 2 CO2 2 NADH2 in Mitochondrien-innenraum 2 x „aktivierte Essigsäure“ (C2) 6 NADH2 2 FADH2 3. Schritt: Citronensäurecyclus 4. Schritt: Endoxidation 2 x 2 CO2 restloser Abbau der Glucose ! 2 ATP 10 NADH2 2 FADH2 = Atmungskette Innenmembran der Mitochondrien werden eingeschleust zum ATP-Gewinn !!! 10 NADH2 30 ATP 2 FADH2 4 ATP ATP-Bilanz zwei aus Glycolyse zwei aus Citronensäurecyclus 38 ATP 34 aus ATmungskette Seite 167 Florian Zeller 07/08 GÄRUNGEN Stoffabbau = Dissimilation Die Grundprozesse laufen bei Mensch und Tier gleich ab. Bedeutung: ATP-Gewinn aus energiereichen organischen Molekülen wie Glucose. 2 Wege: anaerob = ohne O2 = Gärung, sehr alter Stoffwechsel aerob = mit O2 = Atmung, tritt später in der Evolution auf Hier: alkoholische Gärung Anmerkung: Um die restliche Luft oben im Rundkolben zu verdrängen, kann man z.B. Stickstoff einleiten! Summengleichung: C6H12O6 + 2 ADP + 2 P C2H5OH + 2 CO2 + 2 ATP Ethanol (Alkohol) Beim anaeroben Abbau ist der Weg bis zur Brenztraubensäure (BTS) gleich, da kein O 2-Verbrauch bei der Glycolyse; Im Cytoplasma; Gewinn pro Glucose-Molekül: 2 ATP; der Wasserstoff der beiden NADH2 kann nicht „Endoxidiert“ werden. Die Arten der Gärungen sind nach ihren Endprodukten benannt! Seite 168 Florian Zeller 07/08 1. Milchsäuregärung 1. Schritt: Glycolyse Traubenzucker (C6) 2 x Brenztraubensäure (C3) C6H12O6 + 2 ADP + 2 P + 2 NADP O 2 H3C – C – COOH + 2 ATP + 2 NADH2 BTS Der Energiegewinn = 2 ATP ist also bei den Gärern mit der Glycolyse abgeschlossen. Jedoch muss das NAD regeneriert (= zurückgewonnen) werden, sonst käme die Glycolyse zum Stillstand (und damit der lebensnotwendige ATP-Gewinn!) daher zweiter Abbauschritt notwendig, der sich nun je nach Gärungstyp verschieden gestaltet. 2. Schritt: Regeneration von NAD 2 C3H4O3 + 2 NADH2 2 C3H6O3 O + 2 NAD (Summengleichung) OH 2 H3C – C – COOH + 2 NADH2 2 H3C – C – COOH + 2 NAD H Milchsäure Bilanz: C6H12O6 + 2 ADP + 2 P Milchsäurebakterien 2 C3H6O3 + 2 ATP Milchsäure OH C6H12O6 + 2 ADP + 2 P 2 H3C – C – COOH + 2 ATP H Bedeutung von Milchsäuregärung: - beim sauerwerden der Milch Joghurt, Quark - Skelettmuskel bei Sauerstoffmangel - Sauerkrautherstellung - Herstellung von Silofutter 2. Alkoholische Gärung 1. Schritt: Glycolyse C6H12O6 + 2 ADP + 2 P + 2 NADP O 2 H3C – C – COOH + 2 ATP + 2 NADH2 Seite 169 Florian Zeller 07/08 2. Schritt: Regeneration von NAD 2 C3H4O3 + 2 NADH2 2 C2H5OH + 2 CO2 + 2 NAD O (Summengleichung) O 2 H3C – C – COOH 2 H3 C – C + 2 CO2 H OH O 2 H3 C – C 2 H3C – C – H + 2 NADH + 2 NADH2 H H Ethanol Bilanz: C6H12O6 + 2 ADP + 2 P Hefepilze 2 C2H5OH + 2 CO2 + 2 ATP Ethanol Bedeutung der alkoholischen Gärung: - Herstellung alkoholischer Getränke - Konservierung ACHTUNG: Hefen können – falls O2 vorhanden – auch atmen !!!! Aerober Abbau von Glucose (innere Atmung) C6H12O6 + 6 O2 38 ADP + 38 Pi 6 CO2 + 6 H2O 38 ATP Pi i = inorganic = anorganisches Phosphat Seite 170 Florian Zeller 07/08 Beim aeroben Abbau wird die in der Glycolyse gebildete Brenztraubensäure (C3) in die Mitochondrien aufgenommen. Seite 171 Florian Zeller 07/08 Seite 172 Florian Zeller 07/08 Seite 173 Florian Zeller 07/08