Biochemie_Breitling_Stryer_Kap4_1
Werbung

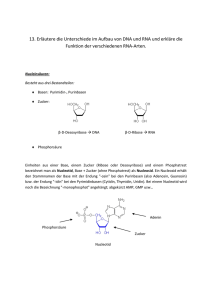
Vertretung durch Frank Breitling (Institut für Mikrostrukturtechnik (IMT), Campus Nord; www.imt.kit.edu/529.php) Vorlesungsdoppelstunde am 26.06.2014 Basis der Vorlesung: Stryer, Biochemie, 6. Auflage, Kapitel 4 DNA, RNA und der Fluss der genetischen Information Wie funktioniert das mit der Vererbung? Jean-Baptiste de Lamarck: Begründer der Evolutionstheorie: gerichtete Höherentwicklung von Organismen, durch die die Artenvielfalt entsteht; dabei werden erworbene Eigenschaften vererbt. Charles Darwin: Gerichtete Höherentwicklung von Organismen, durch die die Artenvielfalt entsteht; dabei werden aus zufälligen Mutationen diejenigen ausgewählt, die verbesserte Eigenschaften vermitteln (natürliche Selektion). Träger der Erbinformation: Ist die Deoxyribonukleinsäure (DNA). Heute wissen wir, dass Lamarck zu einem kleineren Teil auch Recht hatte, denn auch erworbene Informationen können weiter gegeben werden (Epigenetik). Wie wird die Erbinformation umgesetzt? Erbinformation: Gene, in dem Speichermedium DNA („Doppelhelix“, Watson & Crick). Transkription: Kopieren und vervielfältigen der einzelnen Gene in messenger RNA (mRNA; „RNA-Polymerasen“). Translation: Übersetzen und vervielfältigen der einzelnen Geniformationen in Proteine durch eine Matrizen-gesteuerte (= mRNA) nanoskalige Werkzeugmaschine (das „Ribosom“). DNA Transkription RNA Translation Protein Struktur von Nucleinsäuren Nukleinsäuren sind lineare, nicht verzweigte Polymere, die in sich gerichtet sind. Ribose und Desoxyribose Die Atome in den Zuckern sind nummeriert und zur Unterscheidung von den Atomen der Basen mit Strichen versehen Wieso wird die Energie-reiche Phosphodiesterbindung des DNARückgrats nicht sofort hydrolisiert (siehe nächste Folie)? Das Rückgrat von DNA und RNA Das Rückgrat beider Nucleinsäuren wird durch 3'→5‚ Phosphodiesterbindungen gebildet. Eine Zuckereinheit ist rot hervorgehoben, eine Phosphatgruppe blau. Das Rückgrat von DNA und RNA Die negativen Ladungen stoßen Nucleophile wie z.B. ein Hydoxidion ab. Das Fehlen der 2‘-OH-Gruppe bei DNA erhöht deren Resistenz gegen Hydrolyse noch weiter. Das ist sehr wichtig für die Stabilität der Erbinformation! Purine und Pyrimidine Die Basenatome werden ohne Striche durchnummeriert. In RNA kommt Uracil anstelle von Thymin vor. Im DNA- oder RNA-Polymer ist immer das N-9-Atom eines Purins sowie das N-1-Atom eines Pyrimidins ist mit dem C-1‘-Atom des Zuckers verknüpft. Die β-glykosidische Bindung in einem Nucleosid Schreibt man die Strukturformel des Nucleosids (= eine Einheit von Base + Zucker) in der Standardorientierung, liegt die Base oberhalb des Zuckers (= bglykosidische Bindung). Die Nucleotide Adenosin-5'-triphosphat (5'-ATP) und Desoxyguanosin-3'-monophosphat (3'-dGMP) Ein Nucleotid ist ein Nucleosid, das über eine Esterbindung mit einer oder mehreren Phosphatgruppen verbunden ist. Natürlich vorkommende Nucleotide sind fast immer über die Hydroxylgruppen des C-5‘-Atoms des Zuckers verestert. Beispiel: ATP = Adenosin-5‘-triphosphat. Struktur eines DNA-Stranges Ein DNA-Strang verfügt über ein 5'-Ende, an das in der Regel ein Phosphat gebunden ist, und ein 3'-Ende mit einer freien Hydroxylgruppe. Abkürzung dafür: pApCpG bzw. ACG. Elektronenmikroskopische Aufnahme des E. coli Genoms DNA-Stränge können sehr, sehr lang sein. Das Genom von E. coli umfasst ein einziges DNA-Molekül, das aus 2x 4,6 Millionen Nucleotiden besteht. DR GOPAL MURTI/SCIENCE PHOTO LIBRARY Der menschliche Chromosomensatz Quelle: NATIONAL HEALTH MUSEEUM, USA DNA-Stränge können sehr, sehr lang sein. Das Genom des Menschen umfasst 2x 3 Milliarden Nucleotide, die auf 46 Chromosomen aufgeteilt sind. Zusammen genommen und aneinander gereiht sind diese DNA-Stränge etwa 2m lang. Röntgenbeugungsbild einer hydratisierten DNA-Faser Das Kreuz in der Mitte ist für eine helikale Struktur charakteristisch. Die starken Bögen auf dem Meridian rühren von der Stapelung der Basenpaare her, die 0,34 nm voneinander entfernt sind. Watson & Crick formulierten auf Grund der Daten von Rosalind Franklin und von Maurice Wilkins das Modell der DNA-Doppelhelix. Watson-Crick-Modell einer DNADoppelhelix Der eine Polynucleotidstrang ist blau, der andere rot dargestellt. Die Purin- und Pyrimidinbasen sind heller gefärbt als das Zucker-PhosphatGerüst. A) Axiale Ansicht. Entlang der senkrechten Helixachse wiederholt sich die Struktur alle 3,4 nm, was zehn Basenresten auf jeder Kette entspricht. B) Radiale Ansicht, Blick von oben auf die Helixachse Strukturformeln der von Watson und Crick vorgeschlagenen Basenpaare Wasserstoffbrücken: Spezifische Basenpaarungen zwischen A und T sowie zwischen G und C stabilisieren die Doppelhelix. Hydrophober Effekt: Die hydrophoben Basen lagern sich im Inneren der Doppelhelix zusammen. Van-der-Waals-Wechselwirkungen: Eine große Zahl von Atomen der aufeinander gestapelten Aromaten hat die ideale Kontaktdistanz. Elektrostatische Kräfte: Die gleich geladenen Phosphate befinden sich möglichst weit voneinander entfernt außen an der Doppelhelix. Axiale Aufsicht auf eine Doppelhelix Die Basenpaare sind innerhalb der Helix fast genau eins auf das andere gestapelt. Dient immer ein Strang als Matrize für den anderen Strang? Methode um das nachzuweisen: Auftrennung von 14N-DNA und 15N-DNA durch Dichtegradientenzentrifugation A) Eine Aufnahme der UV-Absorption einer Zentrifugen-zelle zeigt zwei getrennte DNA-Banden. B) Densitometrische Auswertung der Absorptionsfotografie. Nachweis der semikonservativen Replikation bei E. coli durch Dichtegradientenzentrifugation Die Lage einer DNA-Bande hängt von ihrem Gehalt an 14N und 15N ab. Nach einer Generation sind alle DNAMoleküle Hybride mit gleichem 14N- und 15N-Gehalt. (Meselson M, Stahl FW (1958) Proc Natl Acad Sci USA 44: 671–682.) Schematische Darstellung der semikonservativen Replikation Die Eltern-DNA ist blau gezeichnet, die neu synthetisierte DNA rot. (Meselson M, Stahl FW (1958) Proc Natl Acad Sci USA 44: 671–682.) Hypochromie A) Doppelsträngige DNA absorbiert Licht weniger effizient als einzelsträngige DNA. B) Die Extinktion einer DNA-Lösung bei einer Wellenlänge von 260nm nimmt zu, wenn die Doppelhelix zu Einzelsträngen schmilzt. Durch Renaturieren lässt sich der Verwandschaftsgrad verschiedener einzelsträngiger DNA-Lösungen sehr einfach bestimmen. Elektronenmikroskopische Aufnahmen von ringförmiger mitochondrialer DNA A) Entspannte Form. B) Superhelix-(supercoiled-)Form. (Mit freundlicher Genehmigung von Dr. David Clayton.) DNA kann in einer superhelikalen Form sehr kompakt verpackt werden. Stamm-Schleife-Strukturen Einzelsträngige DNA- und RNA-Moleküle können Stamm-Schleife-Strukturen ausbilden. Damit können DNA- und RNA-Moleküle auch viele verschiedene 3D-Strukturen einnehmen. DNA-Origami DNA- und RNA-Moleküle können mit sich selbst hybridisieren und so viele verschiedene und vorhersagbare 3D-Strukturen einnehmen. Insofern ist es nicht überraschend, dass auch RNA-Moleküle enzymatische Aktivität zeigen können. Komplex organisiertes RNA-Molekül Einzelsträngige RNA kann sich zu einer komplexen Struktur falten. A) Die Nucleotidsequenz weist in Stamm-Schleife-Strukturen Watson-Crick- und andere, unübliche Basenpaarungen auf. B) Die dreidimensionale Struktur und eine wichtige Wechselwirkung zwischen drei weit auseinander liegenden Basen. Bei der 3D Struktur (links) sind Cytidinnucleotide blau, Adenosin rot, Guanosin schwarz und Uridin grün dargestellt. Die Wasserstoffbrücken innerhalb des Watson-Crick-Basenpaars sind als schwarz gestrichelte Linien gezeichnet, zusätzliche Wasserstoffbrücken als grün gestrichelte Linien. Die von DNA-Polymerasen katalysierte Polymerisation (DNA)n Reste + dNTP (DNA)n+1 Reste + PPi DNA wird durch Polymerasen repliziert, die ihre Instruktionen von Matrizen beziehen (analog zu einer „Matrizen-gesteuerten Werkzeugmaschine“). Die Energie dafür kommt aus der Hydrolyse von PPi: eine Energie-reiche Verbindung wird geknüpft während zwei Energie-reiche Bindungen hydrolysiert werden). Kettenverlängerungsreaktion Die Bildung einer Phosphodiesterbrücke wird von DNA-Polymerasen katalysiert. Dazu wird benötigt: (1) alle vier aktivierten Vorstufenmoleküle dATP, dGTP, dCTP, dTTP und Mg2+ (2) eine Matrize und (3) einen Primer. Außerdem können (4) viele Polymerasen auch noch Fehler korrigieren „proof-reading“. Informationsfluss von RNA zu DNA bei Retroviren Das RNA-Genom eines Retrovirus wird durch die Reverse Transkriptase in DNA überführt. Das Enzym wird durch die infizierenden Viruspartikel in die Zelle gebracht. Die Reverse Transkriptase besitzt mehrere Aktivitäten und katalysiert die Synthese eines komplementären DNA-Stranges, den Abbau der RNA und die anschließende Synthese des zweiten DNA-Stranges. Die in der DNA enthaltene Information muss umgesetzt („exprimiert“) werden. Dabei spielen unterschiedliche Arten von RNA eine Rolle: Typ rel. Menge (%) ribosomale RNA (rRNA) 80 Transfer RNA (tRNA) Messenger-RNA (mRNA) 15 5 Sed. koeff. (S) Masse Anzahl der Nucleotide 23 16 5 4 1,2 x 103 0,55 x 103 3,6 x 101 2,5 x 101 3700 1700 120 75 heterogen Tabelle: RNA-Moleküle in E. coli Die ribosomale DNA ist der Hauptbestandteil der Ribosomen, in denen die Basenabfolge in eine Abfolge von Proteinen übersetzt wird. Die Transfer RNAs transportieren Aminosäuren in aktivierter Form zu den Ribosomen. Für jede Aminosäure gibt es mindestens eine tRNA. Die Messenger-RNAs dienen den Ribosomen als Matrize für die Proteinsynthese. Sie tragen die Informationen der verschiedenen Gene in sich. Es gibt noch einige weitere unterschiedliche Arten von RNA 4. Kleine Kern-RNA-Moleküle („small nuclear RNA“) sind am spleißen der RNA-Exons beteiligt. 5. Der RNA-Proteinkomplex des Signalerkennungspartikels. Dieser Komplex „verschickt“ neu gebildete Proteine, die ein Signalpeptid tragen, durch die Membran des endoplasmatischen Retikulums z.B. für den Export aus der Zelle heraus. 6. Mikro-RNA (miRNA) ist eine Klasse von kleinen (etwa 20 Nucleotide lang) nicht-codierenden RNAs, die an komplementäre mRNAs binden und deren Translation blockieren. 7. Small interfering RNAs (siRNA) ist eine Klasse von kleinen RNAs, die an komplementäre mRNAs binden und deren Abbau bewirken. 8. Der RNA-Bestandteil der Telomerase. Dieses Enzym bewirkt, dass die Telomere (Enden) der Chromosomen bei der DNA-Replikation erhalten bleiben. RNA-Polymerase Dieses große Enzym besteht aus vielen Untereinheiten, darunter β- (rot) und β'-Einheiten (gelb), die eine Art Zange bilden, von der die zu transkribierende DNA festgehalten wird. Man beachte, dass das Aktivitätszentrum in der Mitte der Struktur ein Mg2+-Ion (grün) enthält. Die gewundenen Schläuche, aus denen das Protein in der Darstellung besteht, entsprechen dem Rückgrat der Polypeptidkette. (Zeichnung nach IL9Z.pdb.) Transkriptionsmechanismus der Kettenelongationsreaktion, die von der RNA-Polymerase katalysiert wird Die Bildung einer Phosphodiesterbrücke wird von RNA-Polymerasen katalysiert. Dazu wird benötigt: (1) alle vier aktivierten Vorstufenmoleküle ATP, GTP, CTP, UTP und Mg2+ (2) und eine Matrize. Einen Primer benötigen RNAPolymerasen nicht. Fehler korrigieren („proof-reading“) können sie auch nicht. Komplementarität zwischen mRNA und DNA Die Basensequenz der mRNA (rot) ist das Gegenstück zur Sequenz der DNA-Matrize (blau). Die hier abgebildete Sequenz stammt aus dem Tryptophanoperon, einem DNA Segment, das Gene für fünf Enzyme enthält, welche die Tryptophansynthese katalysieren. Der andere DNAStrang (schwarz) wird als codierender Strang bezeichnet, weil er dieselbe Sequenz wie das RNA-Transkript aufweist (wobei U durch T ersetzt ist). Promotoren für die Transkription von A) Prokaryoten und B) Eukaryoten Angegeben sind die Consensussequenzen. Das erste transkribierte Nucleotid wird mit +1 bezeichnet. Das benachbarte Nucleotid auf seiner 5'-Seite (upstream) trägt die Nummer –1. Die angegebenen Sequenzen sind die des codierenden DNA-Stranges. Es gibt noch viele weiter regulatorische DNA-Sequenzen, z.B. Enhancer, die ziemlich weit entfernt sein können von dem regulierten Gen. Die Basensequenz am 3'-Ende eines mRNA-Transkripts in E. coli Einer stabilen Stamm-Schleife-Struktur folgt eine Sequenz aus Uridin-(U-)Resten. Was anfängt, muss auch aufhören. Die RNA-Polymerase „stolpert“ nach getaner Arbeit am 3‘-Ende regelrecht über Terminator-Sequenzen. Die Modifikation von mRNA Bei Eukaryoten wird Messenger-RNA nach der Transkription modifiziert: Das 5'- Ende wird mit einer „Kappe“ oder Cap versehen, das 3'-Ende mit einem Poly(A)-Schwanz. Der codierende Bereich liefert die Anheftungsstellen für die >20 verschiedenen tRNAs (über spezifische Wasserstoffbrücken), die dadurch die Abfolge der Nucleotide in eine Abfolge von Aminosäuren übersetzen. Bindung einer Aminosäure an ein tRNA-Molekül Die Aminosäure (blau) ist mit der 3'Hydroxylgruppe des endständigen Adenylats der tRNA verestert. Dadurch wird die Aminosäure chemisch aktiviert (für die Bildung einer Peptidbindung muss Energie investiert werden). Auf der anderen Seite der tRNA befindet sich das Anticodon mit der die tRNA an die mRNA Matrize angepasst wird. Allgemeine Darstellung einer Aminoacyl-tRNA Die Aminosäure ist an das 3'Ende der RNA gebunden. Das Anticodon ist die Matrizenerkennungsstelle. Man beachte die Kleeblattstruktur der tRNA mit zahlreichen Wasserstoffbrücken (grüne Punkte) zwischen den Basen. Die spezifische Verknüpfung der 20 verschiedenen Aminosäuren an „ihre“ tRNA wird durch Aminoacyl-tRNASynthetasen katalysiert. Der genetische Code 1. Je drei Basen codieren eine Aminosäure. 2. Der Code ist nicht überlappend. 3. Es gibt keine Zeichensetzung, außer „Start“ und „Stopp“. 4. Der Code ist degeneriert. Startsignal: Formyl-Methionin Die erste Aminosäure eines Proteins, die bei E. coli eingebaut wird, ist immer ein formyliertes Methionin. Die entsprechende tRNA erkennt das Starttriplett AUG (manchmal auch GUG). Bei Eukaryonten wird nichtformyliertes Methionin an der ersten Position eingebaut. Die Initiation der Proteinsynthese Sowohl in Prokaryoten (A) als auch in Eukaryoten (B) ist für die Initiation der Proteinsynthese ein besonderes Startsignal erforderlich Der genetische Code ist nahezu universell Codon Standard Code mitochondrialer Code UGA UGG AUA AUG AGA AGG Stop Trp Ile Met Arg Arg Trp Trp Met Arg Stopp Stopp Tabelle: Besondere Codons in den Mitochondrien des Menschen Der genetische Code stimmt in allen Lebewesen überein, er ist universell, allerdings hat man einige kleiner Änderungen in mindestens 16 Lebewesen gefunden. Meist ist dabei die Bedeutung der dritten, am wenigsten wichtigen Base des Tripletts verändert worden. Die meisten eukaryontischen Gene sind Mosaike aus Exons und Introns Ein durchschnittliches menschliches Gen enthält 8 Introns. Einige Gene können aber auch 100 Introns enthalten, die 50 bis 10,000 Nucleotide lang sein können. Die Identifizierung von nichtcodierenden Introns mithilfe der Elektronenmikroskopie Ein mRNAMolekül (rot) wird mit genomischer DNA hybridisiert, die das ge-suchte Gen enthält. A) Bei einem kontinuierlichen Gen ist eine einzige Schleife aus einzelsträngiger DNA (blau) zu erkennen. B) Enthält das Gen ein Intron, sind zwei Schleifen von einzelsträngiger DNA (blau) und eine Schleife von doppelsträngiger DNA (blau und grün) zu sehen. Zusätzliche Schleifen zeigen dementsprechend weitere Introns an. Transkription des β-GlobinGens und Prozessierung der RNA Die Transkription des Gens ergibt zunächst das Primärtranskript, das dann durch Hinzufügen der Cap-Struktur und der Poly(A)-Sequenz modifiziert wird. Introns werden herausgeschnitten und es entsteht die mRNA. Consensussequenz für das Spleißen von mRNA-Vorläufern Komplexe aus Proteinen und kleinen RNAs, die sog. Spleißosomen, erkennen Signale im Primärtranskript und entfernen auf die Base exakt das Intron. Das Spleißen ist ein evolutionär sehr, sehr altes Verfahren (mindestens 1 Milliarde Jahre). Zellextrakte von Säugerzellen können Hefe mRNA spleißen. Exon shuffling Durch Rekombination lassen sich Exons beliebig mischen, wodurch sich das genetische Repertoire erweitert. Alternatives Spleißen Alternatives Spleißen erzeugt mRNA-Moleküle, die Matrizen für unterschiedliche Formen eines Proteins sind – etwa für einen membrangebundenen Antikörper auf der Oberfläche eines Lymphocyten (A) und für sein lösliches Gegenstück, das von der Zelle sezerniert wird (B). Der membrangebundene Antikörper ist in der Plasmamembran mit einem helikalen Segment (gelb) verankert, das durch ein eigenes Exon codiert wird.