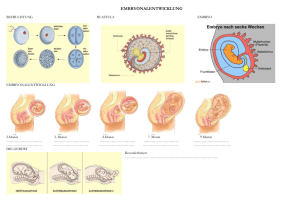
Funktionelle Embryologie - vub
Werbung

Rohen n Lütjen-Drecoll Funktionelle Embryologie Die Entwicklung der Funktionssysteme des menschlichen Organismus 4. Auflage Johannes W. Rohen Elke Lütjen-Drecoll Funktionelle Embryologie 4. Auflage This page intentionally left blank Johannes W. Rohen Elke Lütjen-Drecoll Funktionelle Embryologie Die Entwicklung der Funktionssysteme des menschlichen Organismus 4., überarbeitete und erweiterte Auflage Mit 255 Abbildungen und 9 Tabellen Prof. em. Dr. med. Dr. med. h.c. Johannes W. Rohen Prof. Dr. med. Elke Lütjen-Drecoll Anatomisches Institut (Lehrstuhl II) Universität Erlangen-Nürnberg Universitätsstr. 19, 91054 Erlangen Bibliografische Information der Deutschen Nationalbibliothek Die Deutsche Nationalbibliothek verzeichnet diese Publikation in der Deutschen Nationalbibliografie; detaillierte bibliografische Daten sind im Internet über http://dnb.d-nb.de abrufbar. Besonderer Hinweis: In diesem Buch sind eingetragene Warenzeichen (geschützte Warennamen) nicht besonders kenntlich gemacht. Es kann also aus dem Fehlen eines entsprechenden Hinweises nicht geschlossen werden, dass es sich um einen freien Warennamen handelt. Das Werk mit allen seinen Teilen ist urheberrechtlich geschützt. Jede Verwertung außerhalb der Bestimmungen des Urheberrechtsgesetzes ist ohne schriftliche Zustimmung des Verlages unzulässig und strafbar. Kein Teil des Werkes darf in irgendeiner Form ohne schriftliche Genehmigung des Verlages reproduziert werden. © 2012 by Schattauer GmbH, Hölderlinstraße 3, 70174 Stuttgart, Germany E-Mail: [email protected] Internet: www.schattauer.de Printed in Germany Lektorat: Mira Koerner, Stuttgart Umschlagabbildung: Jörg Pekarsky, Anatomisches Institut, Universität Erlangen-Nürnberg Satz, Druck und Einband: Mayr Miesbach, Druck · Medien · Verlag, Am Windfeld 15, 83174 Miesbach ISBN 978-3-7945-2823-3 V Vorwort zur 4. Auflage Die erfreulicherweise bereits jetzt notwendig gewordene 4. Auflage gibt uns die Möglichkeit, das Bildmaterial erneut zu ergänzen sowie die klinischen Hinweise um die zugehörigen molekularbiologischen Aspekte zu erweitern. Das Einleitungskapitel über die elementaren genetischen Induktionsprozesse bei der frühen Embryonalentwicklung wurde von Herrn Prof. Dr. Klaus von der Mark verfasst und überarbeitet. Im Anhang wurde eine kurze Zusammenfassung über die heutigen Möglichkeiten des somatischen Zellkerntransfers und des therapeutischen Klonens von Herrn Prof. Dr. Michael Wegner eingefügt. Hinsichtlich der Gestaltung und Zielsetzung unseres Buches hat sich jedoch auch bei der Neuauflage nichts Grundsätzliches geändert. Das Buch ist als kurzgefasstes Lehrbuch der Humanembryologie konzipiert, das vor allem durch ein sorgfältig ausgewähltes und anschauliches Bildmaterial die komplizierten Entwicklungsvorgänge der funktionellen Systeme des Organismus ver- ständlich und nachvollziehbar macht. Im Anhang haben wir kurze, zum Teil tabellarische Übersichten zu den Derivaten der Keimblätter, den zeitlichen Abläufen bei der Embryonalentwicklung nach dem CarnegieStadien-System sowie zu den Möglichkeiten der Zwillingsbildungen neu zusammengestellt, um den Studierenden die Rekapitulation der relevanten Entwicklungsereignisse zu erleichtern. Dankenswerterweise hat uns Herr Prof. Dr. Matthias W. Beckmann (UniversitätsFrauenklinik Erlangen) einige Abbildungen aus dem Bereich der Ultraschalldiagnostik zur Verfügung gestellt, die ebenfalls in den Anhang integriert wurden. Wir hoffen, dass unser »kleines Lehrbuch« auch in der jetzt neugestalteten Form seine Freunde behält und neue hinzugewinnt. Erlangen, Herbst 2011 J. W. Rohen E. Lütjen-Drecoll VI Vorwort zur 1. Auflage Bei der Vielzahl der heute vorhandenen Lehrbücher der Embryologie bedarf die Veröffentlichung eines weiteren, wenn auch kurzgefassten Lehrbuches einer besonderen Begründung. Aus langjährigen Unterrichtserfahrungen wissen wir, dass den Studierenden die Vorstellung embryologischer Entwicklungsabläufe immer besonders schwer fällt. Man kann sich die meist flächig und schematisch wiedergegebenen gestaltlichen Vorgänge nicht wirklichkeitsgemäß genug vorstellen und vor allem die räumliche Dimension der entwicklungsgeschichtlichen Veränderungen nicht so erfassen, dass die Vorgänge aus sich heraus verständlich werden. Wir haben daher besondere Sorgfalt auf die Gestaltung des Bildmaterials verwendet, in der Hoffnung, den Studierenden die dreidimensionale Erfassung der Entwicklungsvorgänge und damit das Verständnis der gegenseitigen Beziehungen ihrer induktiven und molekulargenetischen Abhängigkeiten zu erleichtern. Was ist mit »funktioneller Embryologie« gemeint? Alfred Benninghoff hat einmal gesagt, die embryologischen Prozesse sind die gleichen wie die funktionellen des erwachsenen Organismus. So hat er u.a. darauf hingewiesen, dass sich die Spongiosaarchitektur des Knochens embryonal bereits so entwickelt, dass sie später den funktionellen Gegebenheiten entspricht. Hier liegt ein weites Feld zukünftiger Untersuchungen und vieles ist heute noch ungeklärt. Wir haben unter diesen Gesichtspunkten einmal versucht, die funktionellen Zusammenhänge, die auch bei der Embryogenese eine Rolle spielen, herauszustellen und zu zeigen, wie auch bei der Entwicklung der verschiedenen Organsysteme funktionelle Gliederungen erkennbar sind, die das Verständnis der Embryologie nicht nur erleichtern, sondern auch bereichern. So können beispielsweise die Keim- blätter bestimmten Elementarfunktionen des Körpers zugeordnet oder die Entwicklung der Organsysteme unter dem Gesichtspunkt immer enger werdender funktioneller Gliederungen (sozusagen vom Ganzen in die Teile gehend) beschrieben werden. Man kommt dadurch über die heute meist vorherrschende deskriptive oder additive Beschreibung hinaus und findet Systemzusammenhänge, Polaritäten oder Synergismen bei den Gestaltungsprozessen, die ihrerseits wiederum auch das Wesensverständnis der Organe und damit letztlich natürlich auch des Gesamtorganismus erleichtern. Natürlich ist das nur ein bescheidener Anfang, aber vielleicht ein Weg, der über die molekulargenetischen Zusammenhänge hinausgehend das Ganze wieder etwas mehr verständlich machen kann. Da der Student für das ja mehr am Rande liegende Gebiet der Embryologie meist nur wenig Zeit hat, haben wir versucht, nur die wesentlichsten Tatsachen darzustellen, und dies auch in größtmöglicher Kürze. Die bei der Embryonalentwicklung auftretenden induzierenden und steuernden Faktoren und Gene sind heute noch in vielen Bereichen unbekannt oder nur unvollständig aufgeklärt, so dass wir in dieser Auflage nur an den Stellen diejenigen Faktoren mit dargestellt haben, die zum Verständnis der funktionellen Zusammenhänge beitragen können. Die prinzipiellen molekularbiologischen Mechanismen wurden hier nur in einem Anhangskapitel zusammenhängend kurz beschrieben. In folgenden Auflagen soll dieser Teil jedoch noch weiter ergänzt werden. Wir hoffen, dass es gelungen ist, möglichst viele Studierende für dieses so faszinierende Wissenschafts- und Lehrgebiet zu begeistern. Erlangen, Sommer 2002 J. W. Rohen E. Lütjen-Drecoll VII Danksagung Für die Ausarbeitung der nach unseren Entwürfen mit großem Können und Einfühlungsvermögen erstellten Abbildungen möchten wir auch an dieser Stelle Herrn Jörg Pekarsky sowie für die Layout-Arbeiten Herrn Peter Renner herzlich danken. Ganz besonderen Dank schulden wir Frau Lisa Köhler für die sorgfältigen, zuverlässigen und unermüdlichen Arbeiten an dem so oft umgeschriebenen Manuskript. Herrn Prof. Dr. Klaus von der Mark (Nikolaus-Fiebiger-Zentrum für Molekulare Medizin der Universität Erlangen-Nürnberg) möchten wir für das einleitende Kapitel über die genetische Bestimmung des Körperbau- plans und Herrn Prof. Dr. Michael Wegner (Institut für Biochemie der Universität Erlangen-Nürnberg) für das Kapitel über den somatischen Zellkerntransfer und das therapeutische Klonen im Anhang sehr herzlich danken. Auch unseren Mitarbeitern und Studenten möchten wir gerne für die rege Anteilnahme an der Fertigstellung des Buches sowie für die vielen kleinen und größeren, immer sehr hilfreichen Diskussionen unseren Dank aussprechen. J. W. Rohen E. Lütjen-Drecoll VIII Inhalt Einleitung ______________________________ 1 Grundbegriffe ___________________________ 1 Die genetische Bestimmung des Körperbauplans ______________________ 3 Hox-Gene bestimmen den Körperbauplan von Insekten und Wirbeltieren ___ 4 Wie Körperachsen festgelegt werden __ 7 Induktion und Steuerung embryonaler Entwicklungsvorgänge ______________ 8 Molekulare Mechanismen von Induktionsvorgängen ____________ 9 1 1.1 1.2 1.3 2 2.3 Entwicklung von Rückenmark und autonomem Nervensystem _________ 57 2.4 Entwicklung der Extremitäten _______ 61 2.5 Entwicklung des Hautorgans _______ 67 3 Körperhöhlen und Organsysteme _________________ 71 3.1 Perikardhöhle und Herzentwicklung ___________________ 71 Induktion durch lösliche Faktoren __ 10 Induktion durch stationäre Faktoren _______________________ 13 3.1.1 Gefäß- und Blutbildung _____________ 71 3.1.2 Perikardhöhle und primitiver Herzschlauch _____________ 74 Apoptose __________________________ 14 3.1.3 Prozess der Zentralisation ___________ 77 3.1.4 Prozess der Konzentration ___________ 88 Konzeption und Blastemzeit _____________________ 17 3.1.5 Blut- und Lymphgefäßsystem ________ 90 3.1.5.1 Blutgefäßsystem ____________________ 90 3.1.5.2 Lymphgefäßsystem __________________ 93 3.1.6 Fetalkreislauf _______________________ 93 Vorbereitungsprozesse für die Befruchtung – Gametogenese und Menstruationszyklus ___________ 17 3.2 Pleurahöhle und Lungenentwicklung _________________ 97 Befruchtungsvorgang, Morulation und Blastulation ____________________ 25 3.3 Bauchhöhle und Darmentwicklung __ 102 3.3.1 Oberbauchorgane ___________________ 103 Trophoblastentwicklung und Plazentation _______________________ 32 3.3.2 Unterbauchorgane __________________ 108 3.4 Beckenhöhle und Retroperitonealraum – Entwicklung des Urogenitalsystems ______________ 109 3.4.1 Nierenentwicklung und Harnwege ____ 110 Keimblätter und Entwicklung des Embryonalkörpers _______ 43 2.1 Keimblätter ________________________ 43 2.2 Entwicklung des Embryonalkörpers __ 52 3.4.2 Entwicklung der Geschlechtsorgane ___ 114 3.4.2.1 Entwicklung der Keimdrüsen ________ 116 3.4.2.2 Entwicklung der inneren und äußeren Geschlechtsorgane __________ 118 Inhalt IX Anhang ________________________________ 165 4 Kopfbildung 4.1 Gehirnschädel (Desmo- und Neurocranium) _______ 125 Die Embryonalentwicklung im Detail – eine kurze Zusammenfassung _____________ 165 4.2 Gesichtsteil des Kopfes – Branchialskelett und Schlunddarm __ 128 Derivate der Keimblätter __________________ 171 4.2.1 Pharyngealbögen und Schlundtaschenderivate _____________ 128 ___________________ 125 4.2.2 Funktionelle Zusammenhänge _______ 134 4.2.3 Entwicklung von Mund- und Nasenhöhle – Gesichtsentwicklung ____ 135 4.2.4 Zahnentwicklung und Dentitionen ____ 139 4.3 Entwicklung des Nervensystems im Kopfbereich ____________________ 142 4.3.1 Prosencephalon und zugehörige Sinnessysteme (olfaktorisches und visuelles System) __ 4.3.1.1 Vorderhirnentwicklung _____________ 4.3.1.2 Olfaktorisches System ______________ 4.3.1.3 Visuelles System ___________________ Rhombencephalon und Labyrinthorgan ____________________ 4.3.2.1 Entwicklung von Rautenhirn und Medulla oblongata _________________ 4.3.2.2 Kleinhirnentwicklung _______________ 4.3.2.3 Labyrinthorgan ____________________ 146 146 152 152 4.3.2 4.3.3 155 156 158 160 Mesencephalon (Mittelhirn) _________ 163 Die wichtigsten Schritte der Embryonalentwicklung ________________ 172 Zwillingsbildungen ________________________ 173 Ultraschalldiagnostik in der Embryologie _______________________ 174 Somatischer Zellkerntransfer und therapeutisches Klonen ____________________ 175 Literaturhinweise ______________________ 177 Sachverzeichnis _______________________ 178 This page intentionally left blank 1 Einleitung Grundbegriffe Die menschliche Keimesentwicklung findet in der Tuba uterina und der Gebärmutter (intrauterin) statt und dauert von der Befruchtung bis zur Geburt in der Regel 264–268 Tage (38 Wochen = 91/2 Lunarmonate) (Ovulationsalter). In der Klinik berechnet man die Schwangerschaftsdauer in der Regel vom ersten Tag der letzten Menstruation bis zur Geburt, was dann 280 Tage ergibt (40 Wochen = 10 Lunarmonate) (Gestationsoder Menstruationsalter). Man unterscheidet im Allgemeinen eine Blastemzeit, von der Befruchtung bis zur »Gastrulation«, d.h. der Ausbildung der Keimblätter und der ersten axialen Strukturen (16.–19. Tag), eine Embryonalzeit bis zum Ende der 8. Woche, in der im Wesentlichen die Anlagen der großen Organsysteme entstehen, und schließlich eine Fetalzeit, von der 9. Woche bis zur Geburt, in der die Hauptdifferenzierungs- und Reifungsprozesse der Organe, insbesondere des Nervensystems, ablaufen. Wichtige Zäsuren sind die Implantation in die Uterusschleimhaut, die am 6. Tag nach der Befruchtung beginnt, sowie die Ausdifferenzierung der Lunge in der 24.–26. Woche, wonach der Fetus im Falle einer Frühgeburt eine gewisse Überlebenschance hat. Die Fetalperiode geht fließend in die Perinatalperiode über, die vom Ende der 28. Woche bis zum 7. Lebenstag gerechnet wird. In der Perinatalperiode, die noch vor der Geburt beginnt, werden die verschiedenen Organe des Fetus (z.B. Lungen, Herz, Leber, Nieren) nicht nur auf die Geburt selbst vorbereitet, sondern vor allem auch auf die funktionellen Erfordernisse des extrauterinen Lebens eingestellt. In der anschließenden Postnatalperiode ist das 1.–2. Lebensjahr von besonderer Bedeutung, da sich hier nicht nur die Auf- richtung, sondern auch die Ausbildung der Sprachfähigkeit, d.h. elementarer, in dieser ausgeprägten, hochdifferenzierten Form nur dem Menschen eigener Fähigkeiten, vollzieht. Biologisch gesehen ist der Mensch eigentlich ein Nestflüchter, da die vitalen Gehirnfunktionen bei der Geburt schon weitgehend entwickelt und die Sinnesorgane geöffnet sind. Im Gegensatz zu den Nestflüchtern (Huftiere, Wale, Makaken u.a.) ist das menschliche Neugeborene aber noch nicht selbstständig lebens- und bewegungsfähig. Es ist hilflos wie die Neugeborenen von Nesthockern (Nager, Raubtiere usw.), die mit geschlossenen Sinnesorganen und nicht voll funktionsfähigem Bewegungs- und Nervensystem geboren werden. Beim Menschen sind zwar die Augenlider bis etwa zum 7. Monat der Schwangerschaft, wie bei Nesthockern auch, noch geschlossen, öffnen sich aber dann vor der Geburt. In den letzten Schwangerschaftsmonaten erfolgt zwar ein rapides Wachstum des Gehirns, aber die Gliedmaßen und die inneren Organe bleiben im Wachstum zurück. Vergleicht man diese Entwicklung mit der von höheren Säugern und Anthropoiden, müsste die Schwangerschaft beim Menschen nach A. Portmann eigentlich 21–22 Monate dauern. Der Mensch ist also – biologisch gesehen – eine Frühgeburt, die die zweite Hälfte der notwendigen zerebralen Wachstums- und Reifungsperiode erst nach der Geburt absolviert, also gewissermaßen ein »sekundäres Nesthockerstadium« (A. Portmann) durchmacht, für das es im Tierreich sonst nichts Vergleichbares gibt. Die funktionelle Bedeutung dieser Tatsache für die Entwicklung des Menschen liegt darin, dass zusammen mit dem schon embryonal einsetzenden, exzessiven Wachstum des Gehirns, das sich postnatal bis zum Ende des ersten Lebensjahres fortsetzt, die weitere Ausreifung der Organe, 2 Einleitung auch der Sinnesorgane und Gliedmaßen, außerhalb des Uterus in der Umgebung der Familie, d.h. in der sozialen Umwelt, abspielt. Dadurch kann diese Entwicklungsphase, die – biologisch gesehen – eigentlich noch innerhalb des Mutterleibes hätte stattfinden müssen, entscheidend vom sozialen Umfeld mit geprägt werden (Muttersprache, kulturell geprägte Denk- und Verhaltensweisen usw.). Die menschliche Embryonalentwicklung endet also nicht, wie bei den meisten Säugetieren, mit der Fertigstellung eines voll ausgereiften, biologisch überlebensfähigen und vom Verhalten her nicht mehr entwicklungsfähigen Organismus, sondern mit der Geburt eines Wesens, das sich erst im Zusammenwirken mit der sozialen und kulturellen Umwelt zum eigentlich Menschlichen ausgestalten muss. Diese Sonderstellung des Menschen wird auch dadurch unterstrichen, dass der Mensch nicht nur in der Postnatalperiode, sondern auch noch viele Jahre später entwicklungsfähig bleibt und Wachstums- bzw. Reifungsrhythmen zeigt (z.B. der Wachstumsschub in der Pubertät), die bei anderen Spezies in dieser Form nicht vorkommen. Die Sonderstellung des Menschen wird aber nicht nur an den geistigen und körperlichen Entwicklungsfähigkeiten nach der Geburt, sondern auch an den embryonalen Entwicklungsvorgängen selbst ablesbar. Das soll in den folgenden Kapiteln zur Darstellung kommen. Die menschliche Embryonalentwicklung beginnt mit der Befruchtung (Konzeption), d.h. der Vereinigung von Ei- und Samenzelle. Verglichen mit anderen biologischen Prozessen ist dies nicht nur ein äußerst komplexer und umständlich vorbereiteter Vorgang, sondern auch ein staunenswertes, ehrfurchtgebietendes Ereignis. Ei- und Samenzellen sind keine »gewöhnlichen« Körperzellen, die sich auf die Fortpflanzung spezialisiert haben; sie werden auch nicht von den Keim»Drüsen« gebildet, sondern sind als Urgeschlechtszellen in diese Drüsen eingewandert und finden dort ihre für die Befruchtung notwendige Ausgestaltung. Die Urgeschlechtszellen werden von den frühesten embryologischen Entwicklungsstadien (Morula) gewissermaßen beiseite genommen, nehmen an der gesamten Embryonalentwicklung nicht teil und siedeln sich erst relativ spät in den Keimdrüsenanlagen an. Da der Organismus selbst stirbt, das Keimmaterial aber nicht zugrunde geht, sondern von Generation zu Generation weitergegeben wird, spricht man von der Keimbahn (Abb. 1A). Hier liegt also eine funktionelle und – in gewisser Hinsicht – auch materielle Kontinuität vor, die bis in Urzeiten der menschlichen Stammesgeschichte zurückreicht. Würde das genetische Material von Generation zu Generation unverändert weitergegeben, würden keine evolutiven Entwicklungsschritte möglich sein. Bei jeder Befruchtung findet aber eine Neuordnung des genetischen Materials statt, so dass neue Einschläge im Strom des Entwicklungsgeschehens möglich werden, obwohl die Kontinuität erhalten bleibt. Um dies möglich zu machen und das Aussterben der Art zu verhindern, »opfert« der Organismus gewissermaßen einen Teil seiner Organe, nämlich die Geschlechtsorgane, und nutzt sie nicht unmittelbar für eigene Funktionen. Für den Organismus selbst spielen diese Organe keine lebenswichtige Rolle, für die Erhaltung der genetischen Kontinuität und damit für die Erhaltung der Art sind sie aber entscheidend. Individuen Soma Keimplasma Abb. 1A. Schema der Keimbahn. Die Keimbahnzellen (rot) machen die somatische Entwicklung nicht mit und werden von Generation zu Generation weitergegeben, während die Individuen (grau) absterben. Rote Pfeile = jeweils neuer genetischer Einschlag (nach D. Starck). Die genetische Bestimmung des Körperbauplans Die genetische Bestimmung des Körperbauplans Alle Parameter der menschlichen Embryonalentwicklung wie der genaue zeitliche Ablauf, die exakte Koordination der Entwicklung verschiedener Organe und Gewebe, ihre Form, Position, räumliche Ausrichtung und Größe sind – wie bei jeder Spezies – im Genom verankert, sonst wären keine Arterhaltung und identische Reproduktion möglich. Eine der großen Herausforderungen der modernen Entwicklungsbiologie ist die Frage, wie die lineare Information der DNA in die Entwicklung dreidimensionaler Gebilde wie Auge, Wirbelkörper oder Zentrales Nervensystem in ihrer gesamten molekularen und zellulären Komplexität und in die Kontrolle zeitlicher Abläufe umgesetzt wird. Aus Untersuchungen über die Embryonalentwicklung von Fadenwürmern (Nematoden), der Fruchtfliege (Drosophila melanogaster) sowie von Fröschen und Molchen sind in den vergangenen zwei Jahrzehnten bahnbrechende Entdeckungen über Entwicklungskontrollgene hervorgegangen, die in ähnlicher, teilweise fast identischer Form auch die Entwicklung des menschlichen Embryos steuern. Eine der wichtigsten Erkenntnisse war, dass die verschiedenen Stufen der Embryonalentwicklung durch eine Genhierarchie gesteuert werden, in der die sehr früh aktiven Entwicklungskontrollgene, die meist für Transkriptionsfaktoren codieren, Gruppen weiterer Gene (sog. Mastergene) kontrollieren. Diese Kategorie von Genen codiert entweder für weitere Transkriptionsfaktoren oder für parakrine Faktoren, die die weitere Entwicklung steuern (s. S. 10). Am Ende der Hierarchie steht die Regulation von organ- und gewebspezifischen Genen, die für Enzyme, Strukturproteine, Motorproteine oder ribosomale RNA codieren (Abb. 1B). Ein zweiter wichtiger Durchbruch war die Aufklärung von Informationen oder Signalen, die Zellen und Gewebe untereinander austauschen, um so eine exakte zeitliche und räumliche Koordination der Embryonalentwicklung zu gewährleisten. Zu diesen Signalen gehören frei diffundierende Botenstoffe bzw. Signalmoleküle wie Hormone, Wachstumsfaktoren und Zytokine. Sie lösen an ihren zellulären Rezeptoren regulatorische Signalkaskaden in der Zelle aus. Diese Signale sind vor allem für die Kontrolle verschiedener zellulärer Differenzierungsprozesse notwendig, die in koordinierter Form während der Embryonalentwicklung ablau- Entwicklungskontrollgen Transkriptionsfaktor (Mastergen) Promotorregion Myosin MHC Transkriptionsfaktor B (z. B. MyoD) Tropomyosin 3 Aktin Abb. 1B. Hierarchie der Gene in der Regulation der Embryonalentwicklung. Desmin 4 Einleitung fen müssen. Das Gleiche gilt für stationäre Signale, die von Zelloberflächenmolekülen und extrazellulären Matrixmolekülen ausgehen und ebenfalls über spezifische Rezeptoren von benachbarten Zellen erkannt und verarbeitet werden. Obwohl wir noch weit davon entfernt sind, die gesamte Regulationskette während der Entwicklung auch nur eines Organs auf molekularer und zellulärer Ebene lückenlos zu verstehen, gibt es für bestimmte Schlüsselprozesse der Embryonalentwicklung doch schon sehr genaue Modelle, von denen hier exemplarisch einige vorgestellt werden sollen. Ziel ist es, die Wirkungsweise von Entwicklungskontrollgenen, die von ihnen codierten Transkriptionsfaktoren und die spezifischen Funktionen von Wachstumsfaktoren sowie andere extrazelluläre Signale zu entschlüsseln. Dieses Wissen wird unser Verständnis der komplexen morphologischen Veränderungen und Differenzierungsprozesse während der Embryonalentwicklung erleichtern und es uns schließlich ermöglichen, genetisch bedingte Fehlbildungen und andere Erbkrankheiten besser zu verstehen. Hox-Gene bestimmen den Körperbauplan von Insekten und Wirbeltieren Die genetische Analyse von DrosophilaMutanten mit einem extra Beinpaar am Kopf anstelle von Fühlern (Antennapedia) oder einer Mutante mit zwei Paar Flügeln anstelle von einem Paar Flügel und einem Paar Halteren (Bithorax) hat mit zu der bahnbrechenden Entdeckung der homöotischen Gene (Hox-Gene) beigetragen. Sie sind nicht nur für die spezifischen Merkmale der Körpersegmente von Insekten, sondern auch für die von Wirbeltieren verantwortlich. Das Ungewöhnliche an den homöotischen Genen ist, dass sie in der Reihenfolge auf dem Chromosom aneinandergereiht sind, die der Reihenfolge ihrer Expression in den aufeinanderfolgenden Körpersegmenten entspricht, also z. B. den Wirbeln bei den Vertebraten (Abb. 1C). Das bedeutet, dass Hox-Gene, die in terminalen Körpersegmenten exprimiert sind, auch am Ende dieser Gengruppe auf dem Chromosom liegen. Das Neue daran ist auch, dass sich diese Gene, die für Transkriptionsfaktoren codieren, in einer hierarchischen Reihenfolge gegenseitig regulieren. Das hat zur Folge, dass bei Ausfall eines Hox-Gens (z. B. durch Mutation) die benachbarten Hox-Gene auch ausfallen oder geringer aktiviert werden, so dass die betreffenden Körpersegmente die Merkmale eines anderen Segments annehmen können. So führt z. B. die Mutation im HoxGen Ubx bei Drosophila zum Austausch des thorakalen Segments T3 (Merkmal: Stummelflügel oder Haltere) durch T2 (Merkmal: Flügel). Ebenso kann man z. B. in der Maus durch Überexpression des Hox-b3-Gens ein 14. Paar Rippen induzieren. Dieses 14. Paar ist eine Verdoppelung von Paar 13 und kommt durch eine Umwandlung des Lendenwirbels L1 in einen Thorakalwirbel (T14) zustande. Solche Umwandlungen werden homöotische Transformationen genannt. Viele Polysyndaktylien (sechs und mehr Finger) entstehen nicht durch Bildung eines neuen, andersartigen Fingers, sondern durch Verdoppelung eines der fünf Finger. Sie sind auf Mutationen in den Hox-d(9–13)-Genen zurückzuführen, die für die unterschiedlichen Merkmale der fünf Finger bzw. Zehen verantwortlich sind. Im Laufe der Evolution haben sich die zehn Hox-Gene von Drosophila, die auf einem Chromosom liegen, beim Menschen zu vier Hox-Clustern mit je 9–11 Hox-Genen pro Cluster vervielfacht, die jedoch untereinander homolog sind (Abb. 1C). Wie bei Drosophila, sind die am 3’-Ende gelegenen Gene für die anterioren (Kopf-)Segmente verantwortlich, während die am 5’-Ende gelegenen Gene die spezifischen Merkmale der kaudalen Körpersegmente und der Finger bzw. Zehen festlegen. Wie aus Abb. 1D