Thieme: Endspurt Vorklinik – Physiologie 2
Werbung

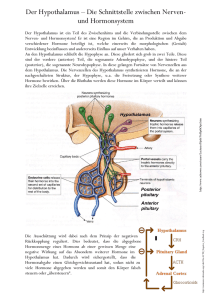
Physiologie 2 | 4 Hormone © PhotoDisc 52 L E R NPAK E T 7 4 Hormone 4.1 Regulation des Hormonsystems und Eigenschaften der Hormone In dem aus mehr als 1014 Zellen bestehenden Körper eines erwachsenen Menschen muss es ein reguliertes Zusammenspiel der einzelnen Zellen und Organe geben, um das innere Milieu aufrechtzuerhalten, auf ändernde Umweltbedingungen reagieren zu können bzw. Wachstum- und Differenzierungsvorgänge zu steuern. Für die Regulation dieser Körperfunktionen sind 2 Systeme vorhanden: Nerven- und Hormonsystem. Sie haben verschiedene Aufgaben, arbeiten aber eng verknüpft. Während das Nervensystem für eine schnelle und räumlich differenzierte Weitergabe der Information sorgt, sind viele v. a. vegetative Funktionen effizienter durch das Hormonsystem über körpereigene chemische Signalstoffe zu regulieren, die Hormone. 4.1.1 Verknüpfung von Nerven- und Hormonsystem Das Bindeglied zwischen Nerven- und Hormonsystem ist der Hypothalamus. Er erhält Signale aus übergeordneten Hirnzentren, dem limbischen System sowie dem Thalamus, und aus der Peripherie über Temperatur- und Osmorezeptoren bzw. nervale Afferenzen. Die Information wird verarbeitet und die entsprechenden Befehle – meist in Form von Releasing-Hormonen – an die Hypophyse weitergegeben. Die Hypophyse stimuliert die Synthese der glandotropen Hormone. Diese wirken an den Hormondrüsen, welche dann wiederum effektorische Hormone in Richtung Zielorgane entlassen. Teilweise stammen diese effektorischen Hormone aber auch direkt aus Hypophyse oder Hypothalamus. Effektorische Hormone. Die effektorischen Hormone stammen größtenteils aus peripheren Hormondrüsen oder verstreut liegenden endokrinen Zellen. Dazu gehören Thyroxin, Aldosteron, Insulin und Progesteron. Ausnahmen sind ADH und Oxytocin, sie stammen aus dem Hypothalamus, sowie Somatotropin und Prolactin, die von der Hypophyse gebildet werden. Die Effektorhormone entfalten am Erfolgsorgan ihre spezifische Wirkung, dabei sind Reichweiten bzw. Wirkorte der effek- torischen Hormone sehr unterschiedlich. Die klassischen Hormone wirken endokrin, sie werden von Hormondrüsen bzw. Drüsenzellen gebildet und auf dem Blutweg im gesamten Körper verteilt. Sie gelangen so auch zu entfernt liegenden Zielorganen. Parakrine Signalstoffe (z. B. Leukotriene, Prostaglandine) dagegen wirken als sog. Gewebshormone lokal auf umliegende Zellen, sie können aber, wenn sie in größeren Mengen gebildet werden, auch endokrine Wirkung entfalten. Autokrine Hormone wirken auf die sezernierende Zelle selbst zurück oder auf benachbarte Zellen des gleichen Typs. Ein Hormon kann sowohl auto- als auch para- oder endokrin wirken. Neben den Hormonen gibt es noch eine weitere Signalgruppe: die Zytokine. Als Zytokine werden Signalmoleküle bezeichnet, die in erster Linie Wachstumsvorgänge, Zellproliferation und -differenzierung regulieren. Man unterscheidet Wachstumsfaktoren wie FGF (= fibroblast growth factor) oder NGF (= nerve growth factor), Hämatopoetine, die die Entwicklung der hämatopoetischen Zellen regulieren, und die Zytokine des Immunsystems. Zytokine sind Proteine, sie werden von zahlreichen Zellen gebildet und wirken v. a. para- und autokrin. Regelkreise. Viele Effektorhormone wirken auf die Hormondrüse, viele glandotropen Hormone auf Hypothalamus und Hypophyse zurück und bremsen dort die weitere Hormonausschüttung. Diesen Effekt bezeichnet man als negative Rückkopplung (negatives Feedback) (Abb. 4.1). So entstehen Regelkreise auf den verschiedenen Ebenen. Im Falle eines einfachen Regelkreises wird die Hormonausschüttung direkt an der Hormondrüse reguliert. Ein Beispiel ist Aldosteron (S. 48) und die Na+-Resorption in der Niere: Bei Na+Mangel wird in der Nebenniere die Ausschüttung von Aldosteron gesteigert, wodurch in der Niere mehr Na+ rückresorbiert wird. Steigt der Na+-Spiegel im Blut, wird die Aldosteronsekretion in der Nebenniere wieder reduziert. Ein Beispiel für einen neuroendokrinen Regelkreis ist die Steuerung der Schilddrüsenhormone (Abb. 4.5): TRH (Thyreotropin-releasing-Hormon) aus dem Hypothalamus bewirkt in der Hypophyse die Ausschüttung von TSH (Thyroidea-stimulierendes Hormon). TSH seinerseits stimuliert in der Schilddrüse die Abgabe der Schilddrüsenhormone T3 und T4. Durch negative Rückkopplung verhindern diese die weitere Freisetzung von TRH und aus: Endspurt Vorklinik – Physiologie 2 (ISBN 9783131534538) © 2015 Georg Thieme Verlag KG 4.1 Regulation des Hormonsystems und Eigenschaften der Hormone 53 oder Hypothalamus Releasing-Hormone Hypophyse glandotrope Hormone Hormondrüse effektorische Hormone Stoffwechselantworten Abb. 4.1 Neuroendokriner Regelkreis. TSH. Sinkt ihre Konzentration jedoch zu weit ab, fällt die hemmende Wirkung auf Hypothalamus und Hypophyse weg und die TRH- und TSH-Synthese nimmt wieder zu. 4.1.2 Eigenschaften der Hormone Hormone gehören unterschiedlichen chemischen Substanzklassen an. Die substanzspezifischen Eigenschaften, z. B. ihre Wasserlöslichkeit, haben Auswirkungen auf den Transport, die zellulären Wirkmechanismen, die Speicherung und die Halbwertszeit der Hormone. Nach ihrer Wasserlöslichkeit lassen sich die hydrophilen Peptidhormone und Aminosäurederivate (mit Ausnahme der lipohilen Schilddrüsenhormone) und die hydrophoben Steroide und Lipidderivate unterscheiden. Peptidhormone. Zu den Peptidhormonen gehören kleinere Oligopeptide (z. B. ADH, TRH), größere Polypeptide (z. B. Insulin, ACTH) und Glykoproteine, die zusätzliche Kohlenhydratketten enthalten (z. B. Erythropoetin, FSH). Da ein Teil der Aminosäuren geladen ist, sind Peptidhormone hydrophil. Aus diesem Grund benötigen sie im Blut keine spezifischen Transportproteine. Sie wirken über extrazelluläre membranständige Rezeptoren, die das Signal an intrazelluläre Botenstoffe, Second Messenger, weitergeben. Die wichtigsten Second Messenger sind Ca2+-Ionen, cAMP, cGMP und IP3. Die Peptidhormone werden am rauen endoplasmatischen Retikulum synthetisiert, im Lumen des ERs bzw. im Golgi-Apparat modifiziert und prozessiert. Die Synthese dauert verhältnismäßig lange. Daher werden Peptidhormone „auf Vorrat“ produziert und in Membranvesikeln gespeichert, aus denen sie auf ein extrazelluläres Signal hin freigesetzt werden. Aminosäurederivate. Zu den Tyrosinderivaten gehören die hydrophilen Katecholamine Dopamin, Noradrenalin und Adrenalin, die enzymatisch aus L-Tyrosin gebildet werden und ihre Wirksamkeit über Ektorezeptoren entfalten. Ihre Halbwertszeit ist sehr kurz (Sekunden bis Minuten). Tyrosinderivate sind auch die Schilddrüsenhormone T3 und T4, die durch Zusammenlagerung zweier iodierter Tyrosinmoleküle entstehen. Sie sind lipo- Steroidhormone. Die lipophilen Steroidhormone (Androgene, Östrogene, Gestagene, Aldosteron, Cortisol, Vitamin-D-Hormon) leiten sich von der Grundstruktur des Cholesterins ab. Ihre Synthese erfordert nur wenige verschiedene Reaktionstypen. Die bei den Umwandlungen stattfindenden Hydroxylierungs- und Oxidationsreaktionen werden bis auf wenige Ausnahmen von Enzymen durchgeführt, die zu der Familie der Cytochrom-P450-Enzyme gehören (Abb. 4.2). Die Synthese von Aldosteron z. B. erfolgt vom Cholesterin ausgehend über Pregnenolon und Progesteron und weitere Zwischenstufen. Vom Pregnenolon zu 17α-OH-Pregnenolon bzw. vom Progesteron zu 17α-Progesteron weiter über 11-Desoxycortisol führt der Weg zu Cortisol. Durch den hydrophoben bzw. lipophilen Charakter ergeben sich einige Eigenschaften der Steroidhormone: ▪ Steroidhormone durchdringen zelluläre Membranen. Sie können daher nicht in membranumgebenen Vesikeln gespeichert werden, sondern werden bei Bedarf neu synthetisiert. ▪ An den Zielzellen gelangen sie durch die Zellmembranen in das Zellinnere. In der Zelle bilden sie mit intrazellulären Rezeptoren Komplexe. Bei den Rezeptoren handelt es sich um ligandenabhängige Transkriptionsfaktoren, die sich als Komplex mit dem jeweiligen Hormon an spezifische DNA-Abschnitte im Zellkern anlagern und dadurch die Transkription bestimmter Gene beeinflussen. So fördert Cortisol bspw. die Transkription der Schlüsselenzyme für die Glukoneogenese. Neben den genomischen Effekten können Steroidhormone intrazellulär Einfluss auf verschiedene Transportmechanismen und verschiedene Proteinkinasen haben. ▪ Aufgrund ihrer schlechten Wasserlöslichkeit werden Steroidhormone im Blut an Transportproteine gebunden. So liegt z. B. Cortisol im Blut zu 90 % an Plasmaeiweiß gebunden vor (75 % an Transcortin = Corticosteroid-bindendes Globulin = CBG, 10 % an Albumin). Die Bindung an die Transportproteine schützt die Hormone vor einem schnellen Abbau. Wirkdauer und Inaktivierung Die Plasmahalbwertszeit gibt an, nach welcher Zeit 50 % der Hormonmenge aus dem Plasma eliminiert sind. Lipophile Hormone (Steroidhormone, Schilddrüsenhormone) sind durch ihre hohe Plasmaeiweißbindung vor einem schnellen Abbau geschützt, ihre Halbwertszeit ist daher deutlich höher (Stunden bis Tage) als die der hydrophilen Hormone (Peptidhormone: Minuten bis Stunden, Katecholamine: Sekunden). Insulin hat wegen einer Reihe hochaktiver Enzyme eine sehr kurze Halbwertszeit von nur 5–15 Minuten. Die Inaktivierung der Hormone erfolgt meist entweder direkt im Erfolgsorgan oder in der Leber. Peptidhormone werden durch Proteolyse in Aminosäuren gespalten, die dann im Stoffwechsel weiterverwendet werden. Steroidhormone werden durch Hydrierung der Doppelbindung in den Hepatozyten inaktiviert. Anschließend werden sie z. B. mit Glucuronsäure und Sulfat verestert und damit wasserlöslicher, sodass sie über die Galle oder mit dem Urin ausgeschieden werden können. Schilddrüsenhormone werden zunächst deiodiert und dann ebenfalls sulfatiert oder glucuronidiert. Katecholamine werden durch Desaminierung und Methylierung inaktiviert. aus: Endspurt Vorklinik – Physiologie 2 (ISBN 9783131534538) © 2015 Georg Thieme Verlag KG L E R NPAK E T 7 phil und können daher wie die Steroidhormone die Plasmamembran durchdringen und binden v. a. an intrazelluläre Transkriptionsfaktoren als Rezeptoren. höhere Zentren Bewertung: metabolischer Status, Energiestatus, Stress etc. 54 Physiologie 2 | 4 Hormone 21 20 22 18 17 19 1 3 HO Cholesterin 1 CH3 CH3 C O C O O OH 6a 6b 7 HO HO HO Pregnenolon 2 C CH3 2 O C O O 17α-Hydroxyprogesteron CH2 OH C O 6b O O Progesteron DHEA-Sulfat 2 OH 6a 3 DHEA 17α-Hydroxypregnenolon CH3 3 O CH2 OH C Androstendion Zona reticularis O OH Enzym/Aktivität O 5 2 3β-Hydroxysteroid-Dehydrogenase 11-Desoxycortisol 11-Desoxycorticosteron CH2 OH 4 C 4 O HO 4 11β-Hydroxylase (P450c11) C 5 Aldosteronsynthase, besitzt drei Aktivitäten: 11β-Hydroxylase, 18-Hydroxylase, 18-Oxidase 5 HO O O OH O Cortisol Corticosteron 3 21α-Hydroxylase (P450c21) CH2 OH HO O O 1 Desmolase (P450scc) O 6a P450c17, besitzt zwei Aktivitäten: 6b a) 17α-Hydroxylase b) 17, 20-Lyase/Desmolase 7 Sulfokinase CH2 OH HC C O Aldosteron Zona glomerulosa Zona fasciculata Abb. 4.2 Übersicht über die Steroidhormonsynthesen in der Nebennierenrinde. Jede der 3 Rindenschichten besitzt zur Synthese der verschiedenen Steroidhormongruppen eine unterschiedliche Enzymausstattung. [aus Behrends et al., Duale Reihe Physiologie, Thieme 2010] APROPOS Als Ursache einer überschießenden oder mangelnden Hormonproduktion kann eine primäre oder eine sekundäre Störung vorliegen. Bei primären Störungen ist die periphere Hormondrüse selbst betroffen und produziert zu viel Hormon (z. B. NNR-Adenom) oder zu wenig (z. B. nach hämorrhagischer Infarzierung beider Nebennieren). Die glandotropen Hormone und Releasing-Hormone sind aufgrund der Rückkopplung durch die jeweiligen Hormonspiegel entsprechend supprimiert oder erhöht. Bei sekundären Störungen ist der Überschuss bzw. Mangel an Effektorhormon auf eine gestörte Stimulation der peripheren Hormondrüse zurückzuführen, daher sind sowohl die Konzentration des Effektorhormons als auch die des glandotropen Hormons verändert. Zwischen primären und sekundären Ursachen lässt sich durch einen Stimulationsversuch mit den entsprechenden glandotropen Hormonen oder Releasing-Hormone unterscheiden. Bei sekundären Ursachen steigt die Konzentration des Effektorhormons nach exogener Zufuhr des Tropins an, bei primären Störungen bleibt sie unverändert niedrig. aus: Endspurt Vorklinik – Physiologie 2 (ISBN 9783131534538) © 2015 Georg Thieme Verlag KG 4.3 Hormone der Nebennierenrinde – ! Oxytocin wird in Zellen des Hypothalamus synthetisiert. – ! Insulin hat mit 5–15 Minuten eine kurze Halbwertszeit. 4.2 Hypothalamus- und Hypophysenhormone noch glandotrope Hormone, sondern ADH und Oxytocin wirken direkt auf periphere Organe. ▪ ADH (S. 49), auch als Adiuretin oder Vasopressin bezeichnet, induziert den Einbau von Aquaporinen in die Sammelrohrwand und bewirkt so eine vermehrte Wasserrückresorption. ▪ Oxytocin (S. 75) steigert die Uteruskontraktiliät (Wehen) und löst Kontraktionen der myoepithelialen Zellen der Milchdrüsen aus (Milchejektion). 4.2.1 Hormone des Hypothalamus FAZIT – DAS MÜSSEN SIE WISSEN Der Hypothalamus verarbeitet die Signale aus den übergeordneten Zentren und schickt die entsprechenden Befehle an neurosekretorische Zellen im medialen und rostralen Hypothalamus: – ! Dopamin ist der bedeutsamste Hemmer der hypophysären Sekretion von Prolactin. – ! Oxytocin steigert die Kontraktion des Uterus. Medialer Hypothalamus. Im medialen Hypothalamus produzieren neurosekretorische Zellen Releasing- und Inhibiting-Hormone, die über die Axone der Neurone zum Hypophysenstiel (Infundibulum) transportiert werden. Dort gelangen die Hormone in die Portalgefäße, die sich im Bereich des Hypophysenvorderlappens zu einem zweiten Kapillarsystem aufzweigen (Pfortadersystem). In der Adenohypophyse werden die Hormone durch das gefensterte Endothel aufgenommen und beeinflussen dort die Bildung der glandotropen und nicht glandotropen Hormone. Releasing-Hormone (Liberine) fördern die Ausschüttung des entsprechenden glandotropen Hormons, z. B. CRH (Corticotropin-releasing-Hormon), TRH (Thyreotropin-releasing-Hormon) oder GnRH (Gonadotropin-releasing-Hormon), Inhibiting-Hormone (Statine) hemmen sie. Rostraler Hypothalamus. Im rostralen Hypothalamus synthetisieren neurosekretorische Zellen des Ncl. supraopticus und des Ncl. paraventricularis Antidiuretisches Hormon (ADH, Adiuretin, Vasopressin) und Oxytocin. ADH und Oxytocin sind sich strukturell sehr ähnlich: Sie bestehen beide aus 9 Aminosäuren mit jeweils 2 Cysteinmolekülen, die eine Disulfidbrücke bilden. Trotz ihrer Ähnlichkeit kann eine Zelle aber jeweils nur entweder ADH oder Oxytocin synthetisieren. In beiden Kerngebieten des Hypothalamus findet man jedoch beide Zelltypen. Die Hormone gelangen über axonalen Transport in die Neurohypophyse (Hypophysenhinterlappen). 4.2.2 Hormone der Hypophyse Die Hypophyse besteht aus der Adenohypophyse (Hypophysenvorderlappen) und der Neurohypophyse (Hypophysenhinterlappen). Adenohypophyse. Die Adenohypophyse (Hypophysenvorderlappen, HVL) bildet unter dem Einfluss der hypothalamischen Releasing- und Inhibiting-Hormone glandotrope Hormone (Tropine), die über das Blut zu den endokrinen Drüsen gelangen (= Zielorgan) und dort die Freisetzung von effektorischen Hormonen steuern (Tab. 4.1). Dazu gehören z. B. ACTH (Adrenocorticotropes Hormon), TSH (Thyroidea-stimulierendes Hormon) und FSH (follikelstimulierendes Hormon). Neurohypophyse. Die aus dem Hypothalamus stammenden Hormone ADH und Oxytocin gelangen durch axonalen Transport in die Neurohypophyse (Hypophysenhinterlappen, HHL). Dort werden sie in den axoterminalen Strukturen gespeichert (vergleichbar mit Neurotransmittern in synaptischen Vesikeln). Ein Aktionspotenzial der Nervenzelle führt zur Freisetzung der Hormone ins Blut. ADH und Oxytocin sind effektorische Hormone. Im ADH- und Oxytocin-System gibt es also weder Releasing- 4.3 Hormone der Nebennierenrinde LERNTIPP Zielorgan der Mineralocorticoide ist die Niere. Diese Überschneidung können Sie zum themenübergreifenden Lernen nutzen (Kap. 3.2). Die Nebenniere besteht aus funktionell weitgehend unabhängigen Teilen: der Nebennierenrinde (NNR) und dem Nebennierenmark (NNM). Die Nebennierenrinde lässt sich histologisch und funktionell von außen nach innen in 3 Schichten gliedern (Abb. 4.2): ▪ Die Zona glomerulosa produziert Mineralocorticoide (v. a. Aldosteron), die der Regulation des Elektrolyt- und Wasserhaushalts und des Blutdrucks dienen. ▪ In der Zona fasciculata werden Glucocorticoide (v. a. Cortisol) gebildet, die Einfluss auf fast alle Stoffwechselvorgänge haben und v. a. der Energiebereitstellung in Stresssituationen dienen. ▪ Die Zona reticularis setzt Androgene (v. a. Dehydroepiandrosteron und Androstendion) frei, sie sind Vorstufe für die Synthese von Testosteron und Östrogenen (Kap. 5.3). Im Nebennierenmark sind der vorherrschende Zelltyp die chromaffinen Zellen. Diese modifizierten Ganglienzellen produzieren die Amine Adrenalin und Noradrenalin. Sie werden bei Stresssituationen ins Blut freigesetzt. Für Adrenalin ist das NNM die wichtigste Quelle, Noradrenalin wird zusätzlich in postganglionären sympathischen Neuronen gebildet, wo es als Transmitter wirkt. 4.3.1 Mineralocorticoide Das in der Zona glomerulosa produzierte Mineralocorticoid Aldosteron (S. 48) ist das wichtigste Hormon für die Regulation des Na+- und K+-Haushaltes. Zielorgan von Aldosteron ist hauptsächlich die Niere, wo es die Na+-Rückresorption und die K+- und H+Sekretion steigert. Dem Na+ folgend wird vermehrt Cl− und Wasser resorbiert, was eine Zunahme des Extrazellulärvolumens und einen Anstieg des Blutdrucks zur Folge hat. Mineralocorticoide spielen damit eine wichtige Rolle bei der Regulation des Elektrolyt- und Wasserhaushalts und damit des Blutdrucks. Zusätzlich zur Niere wird auch im Magen-Darm-Trakt und in den Schweißdrüsen die Na+-Resorption unter Aldosteron-Einfluss gesteigert. Die Aldosteronfreisetzung wird in erster Linie über das Renin-Angiotensin-Aldosteron-System (S. 49) in Abhängigkeit von der Plasmaosmolalität und dem Blutdruck reguliert. Ein wichtiger Stimulus an den Zellen der Zona glomerulosa ist Angiotensin II. Außerdem beeinflussen die Na+- und K+-Konzentrationen im Plasma die Zellen direkt: aus: Endspurt Vorklinik – Physiologie 2 (ISBN 9783131534538) © 2015 Georg Thieme Verlag KG L E R NPAK E T 7 FAZIT – DAS MÜSSEN SIE WISSEN 55 56 Physiologie 2 | 4 Hormone ▪ Eine niedrige Na+- (Hyponatriämie, < Normalwert: 145 mmol/l) und hohe K+-Konzentration (Hyperkaliämie, > Normalwert: 4,5 mmol/l) steigern die Aldosteron-Synthese. ▪ Eine hohe Na+- und niedrige K+-Konzentration hemmen die Aldosteron-Synthese. Auch der pH-Wert spielt eine wenn auch nur untergeordnete Rolle: ▪ Bei Azidose wird vermehrt Aldosteron ausgeschüttet. ▪ Bei Alkalose wird vermindert Aldosteron ausgeschüttet. Tab. 4.1 Hypothalamus- und Hypophysenhormone. (nach Silbernagl/Despopoulos) Abkürzung Name Wirkung Hypothalamus (Releasing-Hormone, Liberine) TRH Thyreotropin-releasing-Hormon, Thyroliberin fördert die Freisetzung von: ▪ Thyreotropin (TSH) ▪ Prolactin (Prl) CRH Corticotropin-releasing-Hormon, Corticoliberin fördert die Freisetzung von: ▪ Corticotropin (ACTH, Vorstufe: POMC) GnRH Gonadotropin-releasing-Hormon, Gonadoliberin fördert die Freisetzung von: ▪ Luteotropin (LH) ▪ Follikotropin (FSH) GHRH Growth-Hormone-releasing-Hormon, Somatoliberin fördert die Freisetzung von: ▪ Somatotropin (STH = GH) Hypothalamus (Inhibiting-Hormone, Statine) GHIH, SIH Somatostatin, Growth-Hormone-inhibiting-Hormon (GHIH) hemmt die Freisetzung von: ▪ Somatotropin (STH = GH) ▪ Thyreotropin (TSH) PIH Dopamin, Prolactin-inhibiting-Hormon hemmt die Freisetzung von: ▪ Prolactin (Prl) Adenohypophyse (Glandotrope Hormone, Tropine) ACTH Adrenocorticotropes Hormon, Corticotropin wirkt auf: ▪ Nebenniere (Corticoide, insbesondere Glucocorticoide) TSH Thyroidea-stimulierendes Hormon, Thyreotropin wirkt auf: ▪ Schilddrüse (T3 und T4, Iodaufnahme und Schilddrüsenwachstum) FSH Follikelstimulierendes Hormon, Follikotropin wirkt auf: ▪ Ovar (Follikelreifung, Östrogenfreisetzung) ▪ Hoden (Spermatogenese) LH Luteinisierendes Hormon, Luteotropin wirkt auf: ▪ Ovar (Ovulation, Progesteronfreisetzung) ▪ Hoden (Testosteronfreisetzung) MSH Melanozytenstimulierendes Hormon, Melanotropin wirkt auf: ▪ Pigmentbildung in den Melanozyten effektorische Hormone des Hypohysenvorderlappens Prl Prolactin bewirkt: ▪ Milchbildung (Brustdrüse) ▪ Hemmung der GnRH-Freisetzung STH (= GH) Somatotropin, Growth Hormone bewirkt: ▪ Körperwachstum ▪ Blutzucker ↑ ▪ Lipolyse ↑ ▪ Freisetzung von Insulin-like growth factor (IGF-I) effektorische Hormone des Hypohysenhinterlappens ADH Antidiuretisches Hormon, Adiuretin, Vasopressin wirkt auf: ▪ Niere (Antidiurese) – Oxytocin wirkt auf: ▪ Uterus ▪ Brustdrüse aus: Endspurt Vorklinik – Physiologie 2 (ISBN 9783131534538) © 2015 Georg Thieme Verlag KG 4.3 Hormone der Nebennierenrinde LERNTIPP Was bei einem Hormonmangel bzw. -überschuss passiert, ist klinisch wichtig. Besonders Conn-Syndrom, Cushing-Syndrom, Morbus Addison sind wichtige IMPP-Themen. Es gibt zahlreiche Fragen zu den Befunden beim Hyperaldosteronismus, Hyper- und Hypokortisolismus. Schauen Sie sich dazu auch ggf. nochmals das Aldosteronkapitel (S. 48) bei der Nierenfunktion an. FAZIT – DAS MÜSSEN SIE WISSEN – ! Bei Hyperaldosteronismus kommt es zur Hypokaliämie. 4.3.2 Glucocorticoide Die vorwiegend in der Zona fasciculata, aber auch in der Zona reticularis gebildeten Glucocorticoide (Cortisol, Cortison, Corticosteron) beeinflussen zahlreiche Stoffwechselprozesse, die der Energiebereitstellung in Stresssituationen dienen. In Ruhe haben sie einen Einfluss auf das Immunsystem. Das biologisch wirksamere Glucocorticoid ist Cortisol, es macht 85 % der gesamten Glucocorticoidaktivität aus. Cortisol wird enzymatisch durch die 11β-Hydroxysteroid-Dehydrogenase 2 in das biologisch weniger aktive Cortison umgewandelt (Abb. 4.3). Die umgekehrte Reaktion – die Bildung von Cortisol CH2 OH C HO O Wirkungen Da alle Körperzellen Glucocorticoidrezeptoren besitzen, hängt die Wirkung von der jeweiligen Funktion der Zelle ab und die Wirkungen der Glucocorticoide sind entsprechend vielfältig. Sie spielen insbesondere für den Stoffwechsel eine zentrale Rolle: ▪ Anstieg des Blutzuckerspiegels durch vermehrte Glukoneogenese und Senkung des Glucoseverbrauchs in der Peripherie. ▪ Steigerung der Lipolyse. Die Konzentration der Fettsäuren, die z. T. in Ketonkörper umgewandelt werden, steigt. ▪ Katabole Wirkung auf den Proteinstoffwechsel, besonders auf die Muskulatur, mit negativer Stickstoffbilanz (die dabei anfallenden Aminosäuren werden zum großen Teil zur Glukoneogenese verwendet). Bei hohen Glucocorticoidkonzentrationen kann der Proteinkatabolismus zur Muskelschwäche führen. Ebenso werden Fibroblasten und die Kollagensynthese im Fibroblasten gehemmt. ▪ Mineralocorticoide Wirkung: Na+-Retention und K+- und H+Sekretion führen zum Blutdruckanstieg. ▪ Abbau von Knochensubstanz ▪ Sensibilisierung verschiedener Organe (z. B. Herz und Gefäße) für die Wirkung der Katecholamine (Vasokonstriktion, Blutdrucksteigerung), z. B. durch Stimulierung der Rezeptorbildung. ▪ Glucocorticoide hemmen Immunprozesse wie z. B. die Bildung von Lymphozyten und eosinophilen und basophilen Granulozyten. Sie stimulieren die Bildung von neutrophilen Granulozyten und bewirken eine verminderte Zytokinfreisetzung, bei längerer Anwendung wird auch die Antikörperbildung herabgesetzt. Dieser Effekt wird zur Unterdrückung unerwünschter Immunreaktionen (z. B. Verhindern einer Abstoßung nach Transplantation, bei Asthma bronchiale, chronisch-entzündlichen Darmerkrankungen) genutzt. CH2 OH 11β-Dehydrogenase 2 (11β-HSD2) OH aus Cortison – wird von der NADPH-abhängigen 11β-Hydroxysteroid-Dehydrogenase 1 katalysiert. Die beiden Enzyme werden gewebsspezifisch exprimiert. 11β-Hydroxysteroid-Dehydrogenase 1 wird v. a. in Haut, Leber, Fettgewebe und ZNS gebildet, während die 11β-Hydroxysteroid-Dehydrogenase 2 in der Niere in hohen Konzentrationen vorkommt. Cortisol hat eine hohe Affinität zum Aldosteronrezeptor, während Cortison nur schlecht bindet. Die Umwandlung des im Blut zirkulierenden Cortisols in der Niere in Cortison verhindert die falsche Besetzung der Aldosteronrezeptoren. Als lipophile Steroidhormone wirken Glucocorticoide über intrazelluläre ligandenaktivierte Transkriptionsfaktoren. Der Transkriptionsfaktor-Hormon-Komplex bindet an zelluläre DNA und beeinflusst die Transkription spezifischer Gene. Dabei ist die Spezifität der Rezeptoren nicht absolut, sodass Glucocorticoide eine leichte mineralocorticoide Wirkung haben und umgekehrt. C O Abb. 4.3 Interkonvertierung von Glucocorticoiden. [aus Behrends et al., Duale Reihe Physiologie, Thieme 2010] O OH 11β-Dehydrogenase 1 (11β-HSD1) O O biologisch aktive 11-Hydroxysteroide – Cortisol – Corticosteron biologisch inaktive 11-Ketosteroide – Cortison – 11-Dehydrocorticosteron aus: Endspurt Vorklinik – Physiologie 2 (ISBN 9783131534538) © 2015 Georg Thieme Verlag KG L E R NPAK E T 7 Das hypophysäre ACTH, das die Zona fasciculata zur Produktion von Glucocorticoiden anregt, hat auch eine schwache stimulierende Wirkung auf die Aldosteronproduktion, allerdings in sehr viel geringerem Ausmaß als auf die Cortisolfreisetzung. Bei einer übermäßigen Aldosteronproduktion (Hyperaldosteronismus) kommt es aufgrund der verstärkten Ausscheidung von K+ zu einer Hypokaliämie, aufgrund der verstärkten Na+-Rückresorption, der Wasser nachfolgt, zu einer Zunahme der Extrazellulärflüssigkeit und dadurch zu einer Blutdrucksteigerung. Die Hypernatriämie bedeutet einen Anstieg der Blutosmolarität, sodass es über die Osmorezeptoren im Hypothalamus zu einer vermehrten Sekretion des antidiuretischen Hormons (ADH) kommt. ADH sorgt für eine weitere Zunahme der Wasserresorption in der Niere und damit für eine weitere Steigerung des Flüssigkeits- und Blutvolumens und des Blutdrucks. Häufig leiden die Patienten unter Kopfschmerzen, Obstipation, Muskelschwäche, Polyurie und -dipsie, EKG-Veränderungen und einer metabolischen Alkalose mit Parästhesien. Ein primärer Hyperaldosteronismus (Conn-Syndrom) kann z. B. durch einen Aldosteron produzierenden Tumor in der Nebennierenrinde (NNRAdenom) verursacht sein. Eine anhaltend starke Stimulation der NNR, z. B. durch Überstimulation des RAAS bei Nierenarterienstenose, führt zu einem sekundären Hyperaldosteronismus. 57 58 Physiologie 2 | 4 Hormone ▪ Beeinflussung des ZNS mit Steigerung der Erregbarkeit gegenüber sensorischen Reizen, euphorisierende oder auch depressionsauslösende Wirkung, Senkung der Krampfschwelle. ▪ Stimulierung der Säuresekretion im Magen. Stress und eine exogene Cortisonzufuhr begünstigen die Entstehung von Magengeschwüren. Regulation der Glucocorticoidsekretion Das aus dem Hypothalamus stammende CRH (Corticotropin-releasing-Hormon) stimuliert in der Hypophyse die Ausschüttung von ACTH (Adrenocorticotropes Hormon). Allerdings wird ACTH nicht direkt synthetisiert, sondern ist Bestandteil eines Vorläuferproteins, des sog. POMC (Proopiomelanocorticotropin). POMC wird posttranslational durch Proteasen in mehrere Peptide gespalten. Dabei entstehen: β-Endorphin („Opio“), α- und γ-MSH („Melano“), ACTH („Cortico“) und γ-LPH (lipotropes Hormon). Eine stark vermehrte ACTH-Sekretion geht daher mit einer vermehrten Sekretion auch der anderen hormonal wirkenden Peptide einher, insbesondere β-Endorphin und α-MSH (Melanozytenstimulierendes Hormon). Letzteres führt z. B. beim Morbus Addison zu verstärkter Pigmentierung der Haut. ACTH wirkt auf die Zona fasciculata der Nebennierenrinde und regt dort die Ausschüttung von Cortisol an. Über negative Rückkopplung hemmt Cortisol die Synthese und Freisetzung von ACTH (Abb. 4.4). Die Freisetzung von Cortisol unterliegt einer ausgeprägten zirkadianen Rhythmik. Sie folgt der pulsatilen Freisetzung des CRH, die in den frühen Morgenstunden am intensivsten ist. Daher ist auch der Cortisolspiegel am frühen Morgen am höchsten, er sinkt bis Mitternacht wieder auf etwa ein Viertel ab. Zu diesem Tag-Nacht-Rhythmus kommt die bedarfsangepasste Freisetzung, dabei ist der stärkste Stimulus für die Cortisolausschüttung körperliche oder psychische Belastung („Stress“), wodurch die Sekretion auf das 10-Fache gesteigert werden kann. Corticoliberin (CRH) FAZIT – DAS MÜSSEN SIE WISSEN – ! Über ihre mineralocorticoide Wirkung (Na+-Retention und K+und H+-Sekretion) führen Glucocorticoide zu einem Blutdruckanstieg. – !! Glucocorticoide wirken am Knochen katabol. – ! Sie hemmen die Lymphozytenbildung. – ! Bei Morbus Addison tritt u. a. Hyperpigmentierung der Haut auf. – ! Der Cortisolspiegel ist morgens höher als nachts (zirkadiane Rhythmik). Hyper- und Hypokortisolismus Hyperkortisolismus (Cushing-Syndrom). Die Symptome eines Hyperkortisolismus (Cushing-Syndrom) sind aufgrund der unterschiedlichen Glucokortikoideffekte vielfältig. ▪ Die gesteigerte Glukoneogenese begünstigt eine diabetische Stoffwechsellage mit erhöhtem Blutglucosespiegel („Steroiddiabetes“). ▪ Durch die Umverteilung des Fettgewebes entwickeln sich eine Stammfettsucht, Stiernacken und Vollmondgesicht. ▪ Gleichzeitig sind die Extremitäten auffallend dünn, was durch den Muskelschwund (Proteinkatabolismus!) verstärkt wird. ▪ An der Haut sieht man neben einer Atrophie Striae distensae und Purpura. ▪ Die Immunabwehr ist herabgesetzt (Immunsuppression). ▪ Die Anzahl der T-Lymphozyten im peripheren Blut ist verringert. ▪ Die Wirkung auf das ZNS kann zu einem endokrinen Psychosyndrom führen. Cortisol hat eine leichte mineralocorticoide Wirkung, sodass sich Symptome wie beim Hyperaldosteronismus (s. o.) ausbilden: Hypokaliämie und Hypernatriämie, was eine Hypervolämie nach sich zieht. Der Blutdruck ist erhöht (arterielle Hypertonie) und ebenso kann durch die verstärkte H+-Ausscheidung in den Nieren eine Alkalose auftreten. Als endogene Ursachen für ein Cushing-Syndrom kommen Störungen der Nebennierenrinde (z. B. NNR-Adenom) oder erhöhte ACTH- oder CRH-Sekretion (z. B. ektope ACTH-Sekretion bei kleinzelligem Bronchialkarzinom) infrage. Weitaus häufiger ist jedoch das exogene, iatrogene Cushing-Syndrom durch Langzeitbehandlung mit Steroiden (z. B. zur Immunsuppression nach Transplantation oder bei Autoimmunkrankheiten). Dabei kann eine langdauernde exogene Cortisolzufuhr zu einer Atrophie der Nebennierenrinde führen. POMC MSH ACTH (Corticotropin) γ-Lipotropin β-Endorphin Nebennierenrinde, Zona fasciculata Cortisol Abb. 4.4 Regelkreis der Cortisolfreisetzung. Hypokortisolismus. Ein Mangel an Glucocorticoiden manifestiert sich mit Hypotonie, Schwäche und rascher Ermüdung, Adynamie, Gewichtsverlust. Auch beim Hypokortisolismus unterscheidet man zwischen primären (NNR-Insuffizienz) und sekundären (Insuffizienz von Adenohypophyse oder Hypothalamus) Störungen, die man bereits klinisch unterscheiden kann. Eine NNR-Insuffizienz (Morbus Addison) ist durch den Mangel an Glucocorticoiden und Mineralocorticoiden (Aldosteron) geprägt. Da Aldosteron die Rückresorption von Na+ und die Sekretion von K+ und H+ fördert, finden sich beim Mangel eine Hyponatriämie einhergehend mit Flüssigkeitsmangel und erniedrigtem Blutdruck, eine Hyperkaliämie und eine metabolische Azidose. Wegen der fehlenden negativen Cortisol-Rückkopplung ist die ACTH-Produktion deutlich gesteigert. Als Nebenprodukt fällt bei der ACTH-Synthese (Vorstufe POMC) immer auch MSH (= Melanozyten-stimulierendes Hormon) an, die Haut und die Schleim- aus: Endspurt Vorklinik – Physiologie 2 (ISBN 9783131534538) © 2015 Georg Thieme Verlag KG