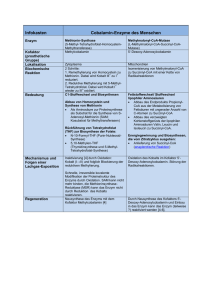
Darstellung und Charakterisierung von Kofaktoren
Werbung

Darstellung und Charakterisierung von Kofaktoren des Elektronentransfers in artifizieller und nativer Proteinumgebung Vom Fachbereich Chemie der Universität Duisburg-Essen Zur Erlangung des akademischen Grades eines Doktors der Naturwissenschaften genehmigte Dissertation von Christoph Breitenstein aus Oberhausen Referent: Prof. Dr. W. Gärtner Korreferent: Prof. Dr. W. S. Veeman Tag der mündlichen Prüfung: 31.05.2006 Diese Arbeit wurde im Zeitraum vom März 2002 bis März 2006 am Max-Planck Institut für Bioanorganische Chemie in Mülheim an der Ruhr unter Anleitung von Herrn Prof. W. Lubitz und Herrn Prof. Dr. W. Gärtner angefertigt. Danksagung Herrn Prof. Wolfgang Lubitz danke ich für die Überlassung der interessanten Themen, sowie die Möglichkeit am Max-Planck-Institut für Bioanorganische Chemie unter seiner Anleitung arbeiten zu dürfen. Außerdem möchte ich mich für die konstruktiven Diskussionen, die zum Erfolg dieser Arbeit beigetragen haben und für seine Flexibilität bedanken, mich in Zusammenarbeit mit Herrn Prof. Wolfgang Gärtner zu betreuen. Herrn Prof. Wolfgang Gärtner möchte ich für die Betreuung danken und die Möglichkeit, diese Arbeit unter seiner Anleitung durchführen zu dürfen. Er war während dieser Zeit stets ein zuverlässiger Ansprechpartner mit konstruktiven Ideen. Für die Zusammenarbeit und die nutzbringenden Diskussionen im Rahmen des Projektes Eisen-Schwefel-Zentren möchte ich Herrn Dr. Mikhail L. Antonkine danken. Außerdem waren seine Kontakte zu Herrn Prof. John H. Golbeck und Prof. Donald A. Bryant von der Pennsylvania State University, Pennsylvania, USA, maßgeblich für den Fortschritt dieses Projektes verantwortlich. Prof. John H. Golbeck stellte die Proben des Photosystems I zur Verfügung und ließ die optischen Messungen am PS I in seinem Labor durchführen. Prof. Donald A. Bryant stellte die Mutanten der Untereinheit PsaC zur Verfügung. Herrn Dr. Maurice van Gastel und Dipl.-Biol. Jens Niklas danke ich für die Unterstützung bei den Triplett-EPR- und ENDOR-Messungen. Frau Gudrun Klihm und Herrn Frank Reikowski danke ich für die technische Betreuung der EPR-Spektrometer und Laser. Herr Dr. Boris Epel hat die meisten Messungen an den Eisen-Schwefel-Peptiden durchgeführt und sich auf diese Weise meinen Dank verdient. Herrn Dr. Hideaki Ogata möchte ich für die Anzucht der E. coli Bakterien danken und die Isolierung des rekombinanten Pottwal-Myoglobins. Außerdem für die unermüdlichen Versuche, Myoglobinkristalle zu züchten. Herr Dr. Sebastian Sinnecker führte die quantenchemischen Berechnungen der Hyperfeinkopplungen durch und trug damit maßgeblich zur Interpretation der erhaltenen Daten bei. Für ihren Einsatz bei den NMR-Messungen möchte ich Herrn Jörg Bitter und Frau Kerstin Sand danken. Sie etablierten in kürzester Zeit das Verfahren, Messungen an wässrigen Lösungen im Institut durchzuführen. Für die Mössbauer Messungen bin ich Herrn Dr. Eckhard Bill und Herrn Bernd Minert zu Dank verpflichtet. Das CD-Spektrometer wurde von Herrn Dr. Frank Neese und Herrn Andreas Göbels zur Verfügung gestellt. Frau Inge Heise möchte ich für die Durchführung der Peptidsynthese und die Wahrung der Ordnung im Labor danken. Ihr Arbeitseinsatz war für mich jederzeit ein Vorbild. Herrn Norbert Dickmann danke ich für die MALDI-TOF Messungen. Herrn Dr. Eberhard Bothe und Herrn Christoph Laurich danke ich für die vielen Hilfestellungen und die gute Zusammenarbeit im Rahmen der elektrochemischen Messungen. Mein Dank gilt ebenfalls Herrn Prof. Jens Ulstrup und Herrn Dr. Tim Albrecht für die Möglichkeit, elektrochemische Messungen in der Bio-Inorganic Group an der Technical University of Denmark, in Kopenhagen durchführen zu dürfen. Frau Manuela Trinoga danke ich für die ausgezeichnete HPLC-Chromatographie. Frau Dipl.-Biol. Petra Kellers danke ich für das Korrekturlesen dieser Arbeit und die Diskussion verschiedener biologischer Sachverhalte. Sämtlichen Mitarbeitern der MS-Abteilung des MPI für Kohlenforschung danke ich für die Unterstützung meiner Arbeit. Frau Helene Steffen und Frau Tanja Berndsen danke ich für die Hilfestellungen bei biologischen Arbeitsmethoden. Frau Tina Buteweg möchte ich für die Durchführung verschiedener Synthesen im Anschluss an ihre Ausbildung danken. Frau Birgit Deckers danke ich für die Hilfe bei der Erstellung einiger Abbildungen. Den Auszubildenden Patricia Malkowski und Melanie Denißen möchte ich für ihren Arbeitseinsatz und die allzeit gute Laune im Labor danken. Besonderer Dank gilt Herrn Dr. Karsten Koppe und Herrn Dr. Hermann Kamperman für die Freundschaft und die gute gemeinsame Zeit während des Studiums. Dem Team der BAC-Dragons unter Leitung von Herrn Bernd Mienert danke ich für die Möglichkeit, Teil dieses Teams zu sein und die schönen Drachenboot-Festivals, die wir im Namen des Institutes bestritten und teilweise gewonnen haben. Allen Mitarbeitern des Institutes, die hier nicht namentlich aufgeführt werden, möchte ich für die freundliche Aufnahme im Haus und das stets gute und freundliche Arbeitsklima danken. Für die finanzielle Unterstützung der Arbeit waren die Deutsch Forschungsgemeinschaft (DFG) und die Max-Planck Gesellschaft (MPG) verantwortlich, auch ihnen gilt mein Dank. Abkürzungsverzeichnis ADP Adenosindiphosphat Apo-Mb Apo-Myoglobin aus Pferdemuskel ATP Adenosintriphosphat Boc tert-Butyloxycarbonyl Da Dalton DCPIP Dichlorphenolindophenol DIPEA Diisopropylethylamin DMF Dimethylformamid DMSO Dimethylsulfoxid DNA (DNS) Desoxyribonukleinsäure FePPIX Eisen-Protoporphyrin (Hämin) ε Extinktionskoeffizient EDTA Ethylendiamintetraacetat EPR Electron Paramagnetic Resonance ENDOR Electron Nuclear Double Resonance ESEEM Electron Spin Echo Envelope Modulation FA Zentrum A in der Untereinheit C des PS I, gleichzeitig Abkürzung des Modellpeptides für dieses Zentrum FB Zentrum B in der Untereinheit C des PS I, gleichzeitig Abkürzung des Modellpeptides für dieses Zentrum Fmoc 9-Fluorenylmethoxycarbonyl FX Zentrum X des Photosystems I zwischen den Untereinheiten PsaA und PsaB HATU N-[(dimethylamino)-1H-1,2,3-triazolo[4,5-b]pyridin-1ylmethylen]-N-methylmethanaminium hexafluorophosphat N-oxid HBTU N-[(1H-benzotriazol-1-yl)(dimethylamino)methylen]-Nmethylmethanaminium hexafluorophosphat N-oxid HiPIP High Potential Iron-Sulfur Protein HOMO Highest Occupied Molecular Orbital HPLC High Performace Liquid Chromatography LUMO Lowest Unoccupied Molecular Orbital MALDI-TOF-MS Matrix Assisted Laser Desoption Ionization – Time of Flight Mass Spectrometry Mb Myoglobin aus Pferdemuskel MePPheid a Pyrophäophorbid a Methylester MePPheid d Pyrophäophorbid d Methylester MO Molekülorbital NAD Nikotinamid-adenin-dinukleotid NADP Nikotinamid-adenin-dinukleotidphosphat n.d. nicht detektiert NMP N-Methylpyrrilidon NOE Nuclear Overhauser Effect ODMR optical detected magnetic resonance P680 Primärer Elektronendonor des Photosystems II P700 special pair des Photosystems I PAL-PEG-PS 5-(4-aminomethyl-3,5-dimethoxyphenoxy)valeryl funktionalisierter Polyethylenglykol-Polystyrol Harz Pheid a Phäophorbid a PPheid a Pyrophäophorbid a PPIX Protoporphyrin PS I Photosystem I PS II Photosystem II PyBOP Benzotriazol-1-yloxytris(pyrrolidino)phosphonium hexafluorophophat RNA (RNS) Ribonukleinsäure SwMb rekombinantes Pottwal-Myoglobin SPPS Solid Phase Peptide Synthesis TBTU N-[(1H-benzotriazol-1-yl)(dimethylamino)methylen]-Nmethylmethanaminium tetrafluoroborat N-oxid TFA Trifluoressigsäure ZnMePPheid a Zink-Pyrophäophorbid a Methylester ZnMePPheid d Zink-Pyrophäophorbid d Methylester ZnPheid a Zink-Phäophorbid a ZnPPheid a Zink-Pyrophäophorbid a ZnPPIX Zink-Protoporphyrin Inhaltsverzeichnis 1. GRUNDLAGEN ........................................................................................1 1.1. Die Struktur von Proteinen ....................................................................................... 1 1.2. Die Struktur der Photoreaktionszentren in der Photosynthese............................. 8 1.2.1. Elektronentransportprozesse in der Photosynthese........................................ 8 1.2.2. Das Photosystem I.......................................................................................... 9 1.3. Die Struktur von Myoglobin ................................................................................... 12 1.4. Die Struktur von Porphyrinen................................................................................ 17 1.4.1. Nomenklatur der Porphyrine........................................................................ 18 1.4.2. Elektronische Struktur der Porphyrine......................................................... 20 1.5. Eisen-Schwefel-Zentren in der Natur..................................................................... 24 1.6. Die Stabilität von Proteinen .................................................................................... 26 1.7. Die Festphasen Peptidsynthese (SPPS) .................................................................. 29 1.8. Grundlagen der EPR- und ENDOR-Spektroskopie ............................................. 34 1.8.1. Grundlagen der EPR-Spektroskopie ............................................................ 34 1.8.2. Grundlagen der ENDOR-Spektroskopie...................................................... 43 1.9. Mössbauer-Spektroskopie ....................................................................................... 48 2. AUFGABENSTELLUNG.......................................................................50 2.1. Metall-Phäophorbide in Myoglobin ....................................................................... 50 2.2. Modelle für [4Fe-4S]-Zentren des Photosystems I ................................................ 51 i 3. ERGEBNISSE UND DISKUSSION ..................................................... 53 3.1. Metall-Phäophorbide in Myoglobin-Matrix...........................................................53 3.1.1. Natives Myoglobin und Apo-Myoglobin .....................................................53 3.1.2. Darstellung der Kofaktoren ..........................................................................56 3.1.3. Komplexbildung und Charakterisierung der Kofaktoren mit Apo-Myoglobin ............................................................................................59 3.1.3.1. Zink-Protoporphyrin Apo-Myoglobin (ZnPPIX Mb) ..................................60 3.1.3.1.1. UV-Vis-Spektroskopie und Stabilitätsmessung ........................60 3.1.3.1.2. NMR-Spektroskopie..................................................................62 3.1.3.1.3. EPR-Spektroskopie....................................................................64 3.1.3.1.4. ENDOR-Spektroskopie von ZnPPIX in Glycerin/DMSO ........68 3.1.3.1.5. ENDOR-Spektroskopie von ZnPPIX Mb in Puffer/DMSO......75 3.1.3.2. Zink-Phäophorbid a Apo-Myoglobin (ZnPheid a Mb) und ZinkPyrophäophorbid a Apo-Myoglobin (ZnPPheid a Mb) ...............................80 3.1.3.2.1. UV-Vis-Spektroskopie und Stabilität........................................80 3.1.3.2.2. NMR-Spektroskopie..................................................................84 3.1.3.2.3. EPR-Spektroskopie....................................................................87 3.1.3.2.4. ENDOR-Spektroskopie von ZnPPheid a in Pyridin/Glycerin ..90 3.1.3.2.5. ENDOR-Spektroskopie von ZnPPheid a SwMb in Puffer/Glycerin ..........................................................................94 3.1.3.2.6. Vergleich der ENDOR-Daten von ZnPPheid a SwMb mit nativen Systemen .......................................................................96 3.1.3.3. Die Komplexe der Zink-Methylpyrophäophorbide mit Apo-Myoglobin.....98 3.1.4. Zusammenfassung ......................................................................................103 3.1.5. Ausblick......................................................................................................105 3.2. Modellpeptide für [4Fe4S]-Zentren ......................................................................106 3.2.1. Literaturübersicht .......................................................................................106 3.2.2. Design der Peptidsequenzen.......................................................................108 3.2.3. Charakterisierung der Modellpeptide .........................................................111 3.2.4. UV-Vis-Spektroskopie von FA und FB .......................................................112 ii 3.2.5. EPR-Spektroskopie der Modellpeptide...................................................... 114 3.2.5.1. Temperaturabhängigkeit des EPR-Signals................................................. 117 3.2.5.2. Leistungsabhängigkeit des EPR-Signals.................................................... 118 3.2.5.3. ESEEM- und ENDOR-Spektroskopie der Modellpeptide FA und FB........ 119 3.2.6. Mössbauer-Spektroskopie.......................................................................... 123 3.2.7. Bestimmung des Redoxpotentials der Modellpeptide FA und FB .............. 126 3.2.8. NMR-Spektroskopie .................................................................................. 129 3.2.9. Bindung an das Photosystem I ................................................................... 130 3.2.9.1. Optische Experimente ................................................................................ 130 3.2.9.2. EPR-Experimente....................................................................................... 132 3.2.10. Zusammenfassung...................................................................................... 134 3.2.11. Ausblick ................................................................................................. 135 4. ZUSAMMENFASSUNG.......................................................................137 5. EXPERIMENTELLER TEIL..............................................................143 5.1. Geräte und Methoden ............................................................................................ 143 5.1.1. NMR-Spektroskopie .................................................................................. 143 5.1.2. UV-Vis-Spektroskopie............................................................................... 143 5.1.3. Infrarot-Spektroskopie ............................................................................... 144 5.1.4. Massenspektrometrie ................................................................................. 144 5.1.5. Säulenchromatographie.............................................................................. 144 5.1.6. Hochdruck-Flüssigkeitschromatographie (HPLC) .................................... 145 5.1.7. Fluoreszenzspektroskopie .......................................................................... 145 5.1.8. Circular Dichroismus- (CD) Spektroskopie............................................... 145 5.1.9. Anaerobe Arbeiten und elektrochemische Titration .................................. 146 5.1.10. Elektron-Paramagnetische Resonanz Spektroskopie (EPR) ...................... 146 5.1.11. Optische Spektroskopie am Photosystem I zur Untersuchung der Elektronentransferkinetik........................................................................... 148 5.1.12. Durchführung der theoretischen Untersuchungen ..................................... 149 iii 5.2. Synthese und Chemikalien.....................................................................................149 5.2.1. Festphasen-Peptidsynthese (SPPS) ............................................................150 5.2.2. Rekombinantes Pottwal-Myoglobin aus E. coli .........................................151 5.2.3. Darstellung von Apo-Myoglobin ...............................................................152 5.2.4. Darstellung von Zink-Protoporphyrin (ZnPPIX) .......................................153 5.3. Darstellung der (Metall-)Phäophorbide ...............................................................154 5.3.1. Allgemeine Arbeitsvorschriften .................................................................154 5.3.1.1. Standardaufarbeitung..................................................................................154 5.3.1.2. Abspaltung des Phytylesters.......................................................................154 5.3.1.3. Metallierung der Phäophorbide ..................................................................154 5.3.2. Gewinnung von Chlorophyll a aus getrockneten Cyanobakterien.............155 5.3.3. Umsetzung von Chlorophyll a zu Phäophorbid a ......................................155 5.3.4. Darstellung von Pyrophäophytin a aus getrockneten Cyanobakterien.......156 5.3.5. Darstellung von Methylpyrophäophorbid a (132-Decarboxymethylphäophorbid a Methylester) .......................................................................157 5.3.6. Darstellung von Methylpyrophäophorbid d ...............................................158 5.4. Rekonstitution von Apo-Myoglobin mit verschiedenen Kofaktoren .................160 5.4.1. Einbau von Zink-Protoporphyrin ...............................................................160 5.4.2. Einbau der Zink-Phäophorbid-Kofaktoren.................................................160 5.5. Darstellung der [4Fe4S]-Modellpeptide ...............................................................161 5.5.1. Die Peptide FA, FB, FX1 und FX2 .................................................................161 5.5.2. Rekonstitution der Peptide .........................................................................162 5.5.3. Herstellung der PS I Proben .......................................................................163 6. LITERATUR......................................................................................... 164 iv Grundlagen 1. Grundlagen 1.1. Die Struktur von Proteinen Proteine sind komplexe Moleküle, die in der Natur sehr spezifische Aufgaben erfüllen und anhand ihrer biologischen Funktion klassifiziert werden können[1]: Enzyme sind hochspezialisierte Proteine, die eine katalytische Aktivität aufweisen. Praktisch alle zellulären chemischen Reaktionen werden durch Enzyme katalysiert. Transportproteine binden bestimmte Moleküle oder Ionen, um sie innerhalb eines Organismus zu transportieren. Das Hämoglobin in den Erythrocyten bindet Sauerstoff und transportiert ihn von der Lunge zum peripheren Gewebe der Organe. Nährstoff- und Speicherproteine dienen der Lagerung von Nährstoffen oder anderer Moleküle und Ionen, die wichtig für die Entwicklung eines Organismus sind. Ferritin, das in einigen Bakterien, sowie Pflanzen und Tieren vorkommt, speichert z. B. Eisen. Casein ist das Hauptprotein der Milch und damit ein wichtiges Nährstoffprotein von Säugetieren. Kontraktile oder motile Proteine sind fähig, ihre Form zu ändern und dienen in vielen Fällen der Fortbewegung. Actin, Myosin und Titin sind die Hauptbestandteile der Skelettmuskulatur. Strukturproteine sind gerüstbildende Filamente, die den Aufbau biologischer Strukturen stabilisieren. Keratin ist der Hauptbestandteil von Haaren, Fingernägeln und Federn. Sehnen und Knorpel werden durch das Faserprotein Kollagen gebildet. Abwehrproteine dienen dem Schutz eines Organismus vor Verletzungen oder eindringenden Organismen. Die Antikörper sind spezialisierte Proteine, die in den Lymphocyten von Wirbeltieren gebildet werden. Sie erkennen eindringende Bakterien, Viren oder artfremde Proteine und neutralisieren diese. Regulatorische Proteine steuern zelluläre und physiologische Vorgänge. Viele Hormone, wie z. B. das Insulin, das den Zuckermetabolismus reguliert, gehören zu dieser Gruppe. Zusätzlich gibt es noch eine Reihe anderer Proteine, die nicht durch diese einfache 1 Grundlagen Klassifizierung beschrieben werden können. Das Blutplasma einiger Fische in der Antarktis enthält z. B. Frostschutzproteine, die ihr Blut vor dem Gefrieren schützen. So unterschiedlich die Funktionen von Proteinen sind, so verschieden sind ihre Molekülstrukturen und –größen. Das bisher größte bekannte menschliche Protein ist das Titin mit einer Größe von etwa 3600 kDa. Es ist neben Myosin und Aktin das wichtigste Muskelprotein. Das Blutzucker-senkende Hormon Insulin ist dagegen ein kleines Protein mit einem Molekulargewicht von lediglich 5,7 kDa. Abbildung 1: Illustration der Winkel Ψ und Φ, durch die die Amidebenen zweier aufeinanderfolgender Aminosäuren gegeneinander verdreht werden können (links). Abhängig von den Winkeln Ψ und Φ werden unterschiedliche Sekundärstrukturelemente gebildet. Neben dem β-Faltblatt (β, roter Bereich) kommen hauptsächlich die rechtshändige α-Helix (α, blauer Bereich) und die linkshändige α-Helix (L, grüner Bereich) in der Natur vor[2]. Die Struktur eines Proteins wird durch seine Zusammensetzung aus den 20 verschiedenen natürlichen Aminosäuren bestimmt. Die lineare Abfolge der Aminosäuren in einem Polypeptid vom N- zum C-Terminus wird als Primärstruktur bezeichnet. Innerhalb einer solchen Kette besitzt eine Aminosäure zwei Bindungen, die mehr oder weniger frei drehbar sind, während die Amidbindung aufgrund ihres partiellen Doppelbindungscharakters nicht frei drehbar ist (Abbildung 1). Die Drehwinkel der Bindungen zwischen Cα und dem Carbonylkohlenstoff und der Bindung zwischen Cα und der Aminogruppe werden durch Ψ und Φ beschrieben. 2 Grundlagen Einige Aminosäuren (z. B. Tryptophan) sind aufgrund ihrer sterisch anspruchsvollen Seitenketten in den Winkeln Ψ und Φ stark eingeschränkt, während Aminosäuren mit kleinen Seitenketten (z. B. Glycin) praktisch jeden Winkel für Ψ und Φ annehmen können. Folgen in einem Polypeptid mehrere Aminosäuren aufeinander, die ähnliche Winkel für Ψ und Φ annehmen, bildet sich ein Sekundärstrukturelement. Abhängig von den Diederwinkeln handelt es sich dabei in den häufigsten Fällen entweder um eine α-Helix oder ein β-Faltblatt. Diese Strukturen werden durch ein Netzwerk von Wasserstoffbrückenbindungen zwischen der Carbonylgruppe einer Aminosäure und der Aminogruppe einer zweiten Aminosäure stabilisiert. Neben der rechtsgängigen α-Helix ist auch die deutlich seltenere linksgängige α-Helix bekannt. Für β-Faltblattstrukturen ist die parallele und die antiparallele Ausrichtung in natürlichen Systemen bekannt. Aufgrund ihrer Struktur besitzt eine α-Helix ein Dipolmoment entlang der Längsachse der Helix, welches vom N-Terminus zum C-terminalen Ende gerichtet ist. Dieses Dipolmoment wirkt in rein α-helicalen Domänen stabilisierend auf die antiparallele Ausrichtung von Helices zueinander und destabilisierend auf eine parallele Ausrichtung. Die Anordnung von mehreren sekundären Strukturelementen räumlich zueinander wird als Tertiärstruktur bezeichnet. Proteine können anhand ihrer Tertiärstruktur in drei Bereiche unterteilt werden, rein α-helikale Strukturen, reine β-Faltblatt-Strukturen und gemischte Strukturen, die sowohl α-Helices als auch β-Faltblätter besitzen. Cytochrom b562, ein Elektronentransferprotein, ist ein Beispiel für ein Protein mit rein α-helikaler Sekundärstruktur. Die Superoxid-Dismutase besitzt dagegen nur β-Faltblätter als Sekundärstrukturelemente. Dieses Enzym katalysiert die Disproportionierung des - Superoxid-Radikals (O2· ) in Wasserstoffperoxid und Sauerstoff. Ein Beispiel für ein Enzym, das sowohl α-Helices als auch β-Faltblattstrukturen besitzt, ist die TryptophanSynthase, die die Reaktion von Indol und Serin zur Aminosäure Tryptophan katalysiert[3]. Viele Proteine bestehen aus mehreren nicht-kovalent verknüpften Polypeptidketten, die sich als Untereinheiten eines Proteinkomplexes zusammenlagern. Die Anordnung verschiedener Untereinheiten zueinander wird als Quartärstruktur bezeichnet. Ein bekanntes Beispiel für ein Protein mit Quartärstruktur ist das Hämoglobin, das aus zwei identischen α-Untereinheiten und zwei identischen β-Untereinheiten besteht und Luftsauerstoff im menschlichen Blut transportiert. 3 Grundlagen Kofaktoren Neben der eigentlichen dreidimensionalen Struktur eines Proteins ist in vielen Fällen entscheidend, wie Kofaktoren im Protein gebunden werden. Unter einem Kofaktor versteht man einen Bestandteil eines Proteins, der nicht aus Aminosäuren besteht. Dabei kann es sich um anorganische Ionen oder ein komplexes organisches oder metallorganisches Molekül handeln. Wird der Kofaktor kovalent an das Protein gebunden, wird er als prosthetische Gruppe bezeichnet. Verschiedene Parameter bestimmen wesentlich die spektralen und funktionellen Eigenschaften eines Kofaktors, wie z. B. das Redoxpotential. Die direkte Umgebung (hydrophil oder hydrophob), die Art der Bindung an das Protein (kovalent, koordinativ oder mittels van-der-Waals Wechselwirkung) sowie zusätzliche Kontakte zur Proteinumgebung (z.B. Wasserstoffbrücken) nehmen Einfluss auf die physikalischen Eigenschaften des Kofaktors. Sofern mehrere Kofaktoren vorhanden sind, ist die relative Ausrichtung der Kofaktoren zueinander entscheidend für die Effizienz eines Energie- oder Elektronentransfers. Faltung Welche Faktoren die Faltung beeinflussen, die für die Funktion eines Proteins nötig ist, ist bis heute weitgehend unverstanden, weil die Faltung eines Proteins ein höchst komplexer Vorgang ist. Ausgehend vom vollständig entfalteten Zustand gibt es eine Vielzahl konformeller Möglichkeiten, die lokale Minima der Energiehyperfläche darstellen, um zu einer „stabilen“ Struktur zu gelangen. Jedoch ist ein Protein einzig in der nativen Struktur in der Lage, die Aufgaben mit der nötigen Effizienz durchzuführen. Bei vielen Proteinen sind spezielle Hilfsstoffe, die sogenannten Chaperone, für die korrekte Faltung eines Proteins nötig. Chaperone sind ebenfalls in vielen Fällen Proteine. Um den Prozess der Faltung besser zu verstehen, bietet es sich an, an kleinen, einfachen Modellen zu ermitteln, welche Primärsequenz besonders gut für ein bestimmtes strukturelles Motiv (z.B. α-Helix) geeignet ist. Als Grundlage dient die rapide wachsende Menge an Kristallstrukturen von nativen Proteinen. Die Zahl der veröffentlichten 4 Grundlagen Strukturen in der seit 1974 bestehenden weltweiten Proteindatenbanka ist von etwa 21000 Strukturen im Juli 2003 auf über 32000 Strukturen im August 2005 angestiegen. Durch Analyse der unterschiedlichen Sekundärstrukturtypen (Helix, Faltblatt) wird versucht, Parameter festzulegen, die diese sekundären Strukturelemente stabilisieren. Abbildung 2: Modell einer Energiehyperfläche für die Proteinfaltung[4]. Ausgehend vom komplett entfalteten Zustand müssen viele lokale Minima durchlaufen werden, bevor der funktionelle, native Zustand erreicht wird. Bei vielen Proteinen sind kleine Moleküle und Proteine, die sogenannten Chaperone, für die native Faltung notwendig. Die Aminosäuren Lysin, Glutamat und Leucin sind zum Beispiel bevorzugte Bausteine von amphiphilen α-Helices, da diese durch die intramolekulare Wasserstoffbrücken zwischen den Seitenketten der beiden polaren Aminosäuren zusätzlich stabilisiert werden. Die hydrophobe Seite wird durch Leucin gebildet. Die in β-Position verzweigten Aminosäuren Valin und Isoleucin, sowie die zyklische Aminosäure Prolin stören die helikale Struktur und werden daher häufig eingesetzt, um dieses Sekundärstrukturelement a RCSB Protein Data Bank, www.pdb.org 5 Grundlagen zu terminieren. Aus amphiphilen Helices wird ein wichtiges Tertiärstrukturelement, das vier-Helix-Bündel, aufgebaut. Dabei lagern sich vier amphiphile Helices in antiparalleler Ausrichtung zusammen. Die Triebkraft ist die hydrophobe Wechselwirkung der Helices untereinander. Daher besitzt ein vier-Helix-Bündel einen stark hydrophoben Kern, wobei es selbst in der Regel gut wasserlöslich ist. Methoden zur Faltungs- und Funktionsaufklärung Für die Funktion eines Enzyms ist die Zugänglichkeit eines aktiven Zentrums für ein Substrat sowie die Polarität und Geometrie der Bindungstasche von essentieller Bedeutung. Im wesentlichen können drei unterschiedliche Wege zur Erforschung von Proteinfaltung und –funktion beschritten werden. a) Molekularbiologisch werden durch ortsspezifische Mutagenese einzelne oder mehrere Aminosäuren eines Proteins verändert und die Änderung der Eigenschaften (z.B. Turn-over Rate, die Substratspezifität eines Enzyms oder spektroskopische Parameter etc.) untersucht und abhängig von der Modifikation am Protein interpretiert. b) In einem Protein mit bekannter Struktur werden neue aktive Zentren konstruiert, wobei das Verständnis der Funktion eines Proteins bzw. der Struktur eines aktiven Zentrums bei dieser Untersuchung im Vordergrund steht. Dabei dient das Protein selbst im Prinzip nur noch als vorgeformte, stabile Abschirmung des neuen aktiven Zentrums. Dieses Verfahren kann nur bei Proteinen durchgeführt werden, die in einem anderen Organismus, z. B. Escherichia coli, durch Einschleusen eines Plasmids überexprimiert werden können (Transformation und heterologe/homologe Expression). Die ursprüngliche Funktion des Proteins geht dabei in der Regel vollständig verloren. c) Alternativ dazu versucht der Ansatz der de novo Synthese, Fragmente eines bestimmten Strukturmotivs (z. B. α-Helices, wie sie in einem vier-Helix-Bündel vorhanden sind) ohne direkten Bezug zu einer natürlich vorkommenden Aminosäuresequenz herzustellen. Diese Methode ist daher auf theoretische Modelle angewiesen. Dieser Ansatz geht besonders stark auf das Verständnis von Faltung und Funktion von Proteinen ein und nutzt dabei native Aminosäuren 6 Grundlagen ebenso wie nicht-native (z.B. α- und β-) Aminosäuren. Bei der de novo Synthese ist zwischen dem rationalen und dem kombinatorischen Design zu unterscheiden. Beim rationalen Design wird die zu synthetisierende Sequenz mit Hilfe theoretischer Modelle entworfen, wobei bestimmte strukturelle Motive berücksichtigt werden. Beim kombinatorischen Design wird eine Bibliothek ähnlicher Modelle synthetisiert. Die einzelnen Modelle werden anschließend anhand von vorher festgelegten Eigenschaften selektiert. Die Selektion stellt häufig ein Problem dar, da eine große Anzahl unterschiedlicher Sequenzen in kurzer Zeit mit eindeutigem Ergebnis bezüglich mindestens einer Eigenschaft untersucht werden muss. Der Syntheseaufwand eines kombinatorischen Ansatzes ist zu Beginn sehr hoch, allerdings ist auch mit sehr guten Ergebnissen zu rechnen, wenn das Problem des „screenings“ zufriedenstellend gelöst werden kann. Die beiden größten Nachteile der de novo Synthese sind der enorme finanzielle Aufwand und die Einschränkung, nur kurze Aminosäureketten bis 60 Aminosäuren mit guter Ausbeute herstellen zu können. Das Ziel der vorgestellten methodischen Ansätze ist es, die Strukturbildung und die Funktion von Proteinen in ihrer Gesamtheit und die Wechselwirkung verschiedener Proteine untereinander zu verstehen. Sollte dies gelingen, so kann entsprechend den Anforderungen eines beliebigen Prozesses ein Protein in silico entworfen und synthetisiert werden. Um dieses Ziel zu erreichen, ist es unabdingbar, einerseits Proteinmodelle zu studieren und andererseits den Einfluss einer bekannten und definierten Proteinumgebung auf einen Kofaktor zu untersuchen. Beide Ansätze werden in dieser Arbeit verfolgt. 7 Grundlagen 1.2. Die Struktur der Photoreaktionszentren in der Photosynthese 1.2.1. Elektronentransportprozesse in der Photosynthese Die pflanzliche Photosynthese ermöglicht durch eine komplexe Reaktion unter Verwendung von Lichtenergie die Umwandlung von Kohlendioxid und Wasser in Kohlenhydrate und liefert damit die chemisch gebundene Energie für das Leben auf der Erde. In grünen Pflanzen findet die Photosynthese in den Chloroplasten statt. Mit Hilfe der Lichtsammelkomplexe werden die Photonen von Chlorophyllen absorbiert und zu zwei Reaktionszentren weitergeleitet. Die Reaktionszentren, in diesem Fall die Photosysteme I und II (Abbildung 3), fungieren als Energieumwandler in den Zellen. Sie absorbieren die Lichtenergie und übertragen diese direkt auf Elektronen, die zum Aufbau energiereicher Stoffe, z. B. NADPH, genutzt werden. Das Photosystem II transferiert die absorbierte Lichtenergie vom P680 über ein Phäophytin auf zwei Plastochinone QA und QB. Durch die Reduktion zum Hydrochinon kann das QB aus dem Protein durch die Membran diffundieren und zwei Elektronen und zwei Protonen an den Cytochrom b6f-Komplex übertragen. Die unbesetzte Bindungstasche im Photosystem II wird durch ein Chinon aus dem Chinonpool besetzt. Während die Protonen in das Thylakoidlumen abgegeben werden und dadurch einen Protonengradienten über die Membran aufbauen, transferiert das Plastocyanin die Elektronen weiter zum PS I. Die Elektronenlöcher, die durch den Abtransport der Elektronen im Photosystem II entstehen, werden durch Oxidation von Wasser wieder gefüllt. Als Koppelprodukte entstehen Sauerstoff, der als Gas die Zelle verlässt und Protonen, die in Form eines Protonengradienten über die Membran beispielsweise von der ATP-Synthase genutzt werden, um Adenosintriphosphat (ATP) aus Adenosindiphosphat (ADP) aufzubauen. Das Photosystem I überträgt nach der Lichtanregung Elektronen an Ferredoxin, das diese an die NADP-Reduktase weitergibt. Die NADP-Reduktase wiederum reduziert mit Hilfe 8 Grundlagen dieser energiereichen Elektronen NADP zu NADPH, das als Reduktionsäquivalent in der Zelle zur Verfügung steht. Abbildung 3: Querschnitt durch eine Thylakoidmembran. Die Lichtsammelkomplexe sind aus Gründen der Übersichtlichkeit nicht gezeigt. 1.2.2. Das Photosystem I Das Photosystem I wird an dieser Stelle detaillierter besprochen, da es als Vorbild für die Modelle der Eisen-Schwefel-Zentren dient, die in dieser Arbeit untersucht werden. Es besteht aus mehr als zehn Untereinheiten (PsaA bis PsaL), von denen A und B ein Homodimer mit jeweils 83 kDa Molekülmasse bilden (Abbildung 4). Sie bestehen aus jeweils elf transmembranen Helices und vier Helices in der Nähe der stromalen Membranoberfläche. Die Untereinheiten C, D und E befinden sich ebenfalls auf der stromalen Seite der Membran. Sekundärstrukturelemente, die Diese meist drei durch Untereinheiten β-Faltblätter besitzen gebildet wenige werden. Die Untereinheiten C und D weisen zusätzlich kurze Helices mit bis zu 15 Aminosäuren auf. In einem Photosystem I sind - abhängig von der Spezies - bis zu 100 Chlorophylle, etwa 22 Carotinoide, zwei Phyllochinone und drei [4Fe4S]-Zentren gebunden. Von dieser Vielzahl an Kofaktoren sind nur elf direkt an der Funktion des Photosystems I, der lichtinduzierten Ladungstrennung, beteiligt. Die restlichen Kofaktoren dienen dem Sammeln von Lichtenergie (Chlorophylle) und der Unterdrückung der reaktiven lichtangeregten Triplettzustände (Carotinoide). Ausgehend von einem Chlorophyll a/a´ Dimer, dem 9 Grundlagen sogenannten special pair P700, existieren in den beiden Untereinheiten PsaA und PsaB zwei symmetrische Elektronentransferwege, der sogenannte A- und der B-Zweig. Sie bestehen aus zwei monomeren Chlorophyll a Molekülen, dem „accessory“ Chlorophyll und dem Akzeptor A0, sowie einem Phyllochinon, dem Akzeptor A1. Die beiden Zweige münden in das erste [4Fe4S]-Zentrum, FX, welches von den Untereinheiten A und B durch eine flexible Region in der Peptidkette gebunden wird[5]. Von FX werden die Elektronen in die Untereinheit PsaC zu den Eisen-Schwefel-Zentren FA und FB übertragen und von Ferredoxin, das an der Oberfläche der Untereinheiten PsaD und PsaE bindet, aufgenommen. A1 A0 Abbildung 4: Schematische Darstellung von sechs Untereinheiten des Photosystems I, sowie der Abfolge der Kofaktoren (links). Kristallstruktur (PDB 1JB0) des oberen Teils des Photosystems I mit den stromalen Untereinheiten C, D und E, sowie den drei [4Fe4S]-Zentren FX, FA und FB (rechts). Die lichtinduzierte Ladungstrennung findet zwischen dem primären Elektronendonor, dem P700, und dem primären Elektronenakzeptor, A0, in etwa 1 ps statt[6]. Anschließend wird das Elektron in 30 ps auf ein Phyllochinon, A1, übertragen und zum FX-Zentrum in 30 ns weitergeleitet. In weniger als 500 ns gelangt das Elektron von dort aus zu den beiden [4Fe4S]-Zentren FA und FB. Der gesamte Elektronentransfer erfolgt von einem Anfangspotential von etwa –1,3 V für den lichtangeregten Singulettzustand des P700 energetisch begünstigt (downhill) über –700 mV für das FX-Zentrum bis zu einem Potential von –590 mV für das Zentrum FB[6]. Einzig der letzte Schritt des 10 Grundlagen Elektronentransfers von FA nach FB ist ein energetisch ungünstiger Transfer (uphill), da FA ein Redoxpotential von –540 mV aufweist. Das Redoxpotential der Eisen-Schwefel-Zentren FX, FA und FB soll hier ausführlicher behandelt werden, da es eine wesentliche Eigenschaft dieser Zentren ist. Das FX-Zentrum ist eines der wenigen Beispiele für ein „interpolypeptid“ Eisen-Schwefel-Zentrum. Im Gegensatz zu den meisten FeS-Zentren wird es von zwei unabhängigen Untereinheiten ligandiert. Es besitzt zudem mit –705 ± 15 mV[7] bzw. –670 mV[8] eines der negativsten Redoxpotentiale. Der negativere Wert wurde durch eine Reduktion an einer Goldoberfläche und Detektion durch EPR bei tiefer Temperatur ermittelt, der positivere Wert ist mit Hilfe transienter optischer Spektroskopie bestimmt worden. Das Redoxpotential für FA liegt im Bereich von –465 mV[9] bis –540 mV[10], dasjenige von FB im Bereich von –440 mV[9] bis –590 mV[10]. Der Grund für diese signifikanten Unterschiede der ermittelten Redoxpotentiale liegt in der verwendeten Messmethode. Die negativeren Redoxpotentiale wurden über eine Redoxtitration mit Dithionit bei pH 10 bei Raumtemperatur durchgeführt. Die Detektion des Titrationsverlaufs erfolgte durch EPR-Spektroskopie bei 20 K. Die positiveren Redoxpotentiale wurden durch die Auswertung der Ladungsrekombinationskinetik nach Anregung durch Blitzlicht ermittelt. Diese Messungen wurden potentialabhängig durchgeführt, wobei das Potential ebenfalls über die Zugabe von Dithionit eingestellt wurde. Bei der Bewertung der beiden Ergebnisse ist zu berücksichtigen, dass die Detektion des Oxidationszustandes von FA und FB durch EPR bei tiefer Temperatur nicht unstrittig ist, weil die Potentiale beider Zentren temperaturabhängig sein können und das Einfrieren eine Änderung des Gleichgewichts bewirken kann. In beiden Studien wurde das Potential von FB in Gegenwart eines reduzierten FA-Zentrums bestimmt, wobei eine gegenseitige Beeinflussung (Kooperativität) beider Zentren untereinander nicht ausgeschlossen werden kann. Außerdem wird bei der Auswertung der Ladungsrekombinationskinetik ein vereinfachtes Modell der möglichen Redoxzustände im PS I angenommen, wobei lediglich die beiden terminalen Elektronenakzeptoren FA und FB, sowie das P700 berücksichtigt werden. Die anderen Kofaktoren, die am Elektronentransfer beteiligt sind, werden bei der Auswertung der Kinetik nicht berücksichtigt. Zusätzlich sind für Photosysteme verschiedener Organismen unterschiedliche Werte für die Redoxpotentiale der einzelnen Eisen-Schwefel Zentren bestimmt worden[11; 12]. 11 Grundlagen Wie durch die Proteinumgebung ein derart negatives Redoxpotential, wie beispielsweise für das Zentrum FX, erzeugt wird, ist weitgehend unverstanden. Eine Wechselwirkung des geladenen [4Fe4S]-Zentrums mit ebenfalls geladenen Aminosäuren in der Umgebung, sowie die Polarisierbarkeit des gesamten Proteins werden zur Erklärung herangezogen. Ein wichtiger Parameter für die Kontrolle des Redoxpotentials ist vermutlich die Zugänglichkeit des Zentrums für das Lösemittel (vgl. Abschnitt 1.5). 1.3. Die Struktur von Myoglobin Myoglobin ist, ebenso wie Hämoglobin, seit Jahrzehnten Ziel intensiver Forschung. Mit einer Sauerstoffaffinität, die zwischen der von Hämoglobin und der von Cytochrom a bzw. a3 (Hauptbestandteil der Cytochrom c-Oxidase) liegt[13], dient Myoglobin in der Zelle als Sauerstoffakzeptor und -speicher aus dem Blut. Weil das Protein in seiner Apo-Form in dieser Arbeit als Matrix für nicht-native Kofaktoren verwendet wird und dadurch seine biologische Funktion verliert, werden im folgenden Abschnitt kurz einige grundsätzliche Eigenschaften und strukturelle Merkmale aufgeführt. Weiterführende Details über die biologische Funktion von Myoglobin sind in verschiedenen Übersichtsartikeln zusammengefasst worden [14-16]. Myoglobin besteht aus 153 Aminosäuren (für Pferdemuskel- und Pottwal-Myoglobin) und ist ein Vertreter der Globin-Proteine. Es ist gut löslich in Wasser (>60 mg/ml) und in seiner nativen Form sehr stabil gegen Denaturierung. In dieser Form liegt der Kofaktor Hämin als Eisen(II)-Spezies vor. Kommerziell ist die inaktive Eisen(III)-Form erhältlich, das sogenannte met-Myoglobin. Myoglobin wurde als erstes Protein vollständig kristallographisch charakterisiert[17]. Im gefalteten Zustand besteht es aus acht α-Helices (A bis H), die von etwa 75 % der Aminosäuren des Proteins gebildet werden. Diese Helices bilden die sogenannte GlobinFaltung, die bei Vertebraten, aber auch bei Pflanzen und Bakterien weit verbreitet ist. Charakteristisches Merkmal der Globin-Faltung ist die Verbindung der Helices über kurze „loops“. Die dreidimensionale Anordnung der Helices bildet eine Tasche, die im Myoglobin durch Hämin besetzt ist. Myoglobin besitzt im nativen Zustand eine hohe Stabilität gegenüber der Entfaltung, die von einer starken Wechselwirkung durch eine 12 Grundlagen dichte Packung der Aminosäurenseitenketten herrührt. Ein typisches Merkmal für die Globin-Faltung ist, dass diese Wechselwirkung nur bei Helices zu beobachten ist, die in der Sequenz nicht direkt aufeinanderfolgen. Eine Ausnahme hiervon stellen die Helices G und H dar, die eine antiparallele Anordnung aufweisen und deshalb durch eine starke Packungswechselwirkung stabilisiert werden. Abbildung 5: Darstellung der Kristallstruktur von Myoglobin (PDB 1A6M). Die Helices A bis F sind durch blaue Zylinder dargestellt. Die Kohlenstoffatome des Kofaktors Hämin, sowie der beiden ligandierenden Histidine sind in türkis, Stickstoffatome in blau und das zentrale Eisenion in gelb gefärbt. Das ursprünglich in der Kristallstruktur enthaltene Sauerstoffmolekül ist zur besseren Übersichtlichkeit nicht abgebildet. Die Aminosäuren des Myoglobins können auf zwei unterschiedliche Arten bezeichnet werden. Entweder wird ihre Position allein durch eine Zahl gekennzeichnet, z. B. Val 68, oder durch einen Buchstaben und eine Zahl, z. B. Val E 11. Bei der ersten Konvention werden die Aminosäuren beim N-Terminus beginnend fortlaufend durchnummeriert. Damit ist die Bezeichnung spezifisch für das Myoglobin einer Spezies, da die Gesamtzahl der Aminosäuren von Spezies zu Spezies variieren kann. Die zweite Möglichkeit 13 Grundlagen bezeichnet die Position der Aminosäure innerhalb einer Helix. Die Bezeichnung E 11 beschreibt die elfte Aminosäure in der Helix E, während CD 1 die erste Aminosäure im „loop“ zwischen den Helices C und D bezeichnet. Diese Art der Positionsangabe ist unabhängig von der Spezies, aus der das Myoglobin isoliert wurde, allerdings erfordert diese Nomenklatur eine präzise Definition der Länge der jeweiligen „loops“ und Helices. In dieser Arbeit werden beide Möglichkeiten der Bezeichnung parallel verwendet, um die Vergleichbarkeit mit älteren Publikationen zu erleichtern. Abbildung 6: Darstellung der Häminbindungstasche im Myoglobin (PDB 1A6M). Neben dem Kofaktor Hämin (grün) mit dem zentralen Eisenion (pink) sind die unmittelbar angrenzenden Aminosäuren dargestellt. Die γ-CH3-Gruppen der Aminosäure Val 68 (E 11) liegen dicht am Ringsystem und stehen in Kontakt mit der π-Elektronenwolke des Kofaktors. Bei Myoglobinen aller Spezies ist die Aminosäure Histidin F 8 konserviert[15]. Dieses proximale Histidin ligandiert das zentrale Eisen(II)ion des Hämins und ist daher essentiell für die Funktion des Proteins. Fast ausschließlich über diese Aminosäure wird die Funktion und Bindungsaffinität des zentralen Eisenions und somit die Funktion des gesamten Proteins gesteuert. Auf der distalen Seite des Hämins sind die Aminosäuren Phenylalanin CD 1, Valin E 11, Leucin B 10 und Histidin E 7 hochkonserviert. Valin E 11, Leucin B 10 und Phenylalanin CD 1 bilden hydrophobe Wechselwirkungen mit dem Kofaktor aus und tragen damit wesentlich zur Stabilisierung des Hämins in der Bindungstasche bei. Die 14 Grundlagen Hauptfunktion des distalen Histidins E 7 ist die Bildung einer Wasserstoffbrückenbindung zum Eisen-koordinierten Sauerstoffmolekül, um dieses zu stabilisieren. In der sauerstofffreien deoxy-Form ist diese Bindungsstelle durch ein Wassermolekül besetzt[18]. Neben dem nativen Kofaktor Hämin ist Myoglobin in der Lage, auch nicht-native Kofaktoren anstelle des Hämins zu binden. Es wurden bereits verschiedene Kobaltporphyrine[19] und Magnesiumporphyrine[20], Zinn(IV)protoprophyrin[21], sowie offenkettige Tetrapyrrole wie Bilinchromophore[22] und verschiedene Chlorophyll- und Bakteriochlorophyllderivate erfolgreich in Myoglobin gebunden[23; 24] . Verschiedene NMR-spektroskopische Untersuchungen zeigen, dass der Einbau des nativen Kofaktors im ersten Schritt maßgeblich durch zwei intermolekulare Salzbrücken zwischen den Propionaten des Hämins und zwei Aminosäuren auf der Oberfläche beeinflusst wird. Dabei handelt es sich um Histidin FG3 und Arginin CD3 bei Pottwal-Myoglobin bzw. Histidin FG3 und Lysin CD3 bei Pferde-Myoglobin[25; 26] . Da das Hämin bezüglich dieser Wechselwirkung symmetrisch ist, kann es bei Rekonstitutionen in vitro in zwei unterschiedlichen Einbaurichtungen im Myoglobin im Verhältnis 1:1 binden. Die eine Einbaurichtung entspricht derjenigen, die in der Kristallstruktur von nativem Myoglobin gefunden wird, während im anderen Fall der Kofaktor 180° um die Achse der α,γ-Methinprotonen gedreht ist (Abbildung 7). Zwar stellt sich in vitro ein Gleichgewicht zwischen beiden Formen ein, wobei die hydrophobe Interaktion der Vinylseitenketten mit der Proteinumgebung die treibende Kraft ist, jedoch wird dieses Gleichgewicht erst nach Tagen oder Monaten erreicht. Für die Umwandlung des einen Isomers in das andere muss der Kofaktor um 180° gedreht werden. Für diesen Vorgang werden zwei unterschiedliche Mechanismen diskutiert[27]. Eine Möglichkeit ist ein unimolekularer Mechanismus, bei dem der Kofaktor die Bindungstasche des Proteins nicht verlässt, sondern sich die Struktur des Proteins derart aufweitet, dass eine Drehung des Kofaktors innerhalb der Bindungstasche möglich ist. Um dies zu erreichen, muss die Eisen-Histidin Bindung gebrochen werden. Der zweite mechanistische Vorschlag ist eine bimolekulare Reaktion, bei der der Kofaktor die Bindungstasche vollständig verlässt, die 180° Drehung ausführt, und anschließend wieder im Protein bindet. Obwohl beide Mechanismen ausführlich in der Literatur diskutiert werden[28], ist die unimolekulare Reaktion wahrscheinlicher. Das Gleichgewicht zwischen beiden Isomeren stellt sich deutlich schneller ein als man unter Berücksichtigung der Dissoziationskonstante 15 Grundlagen von Proteinen und Kofaktor erwartet. Daher kann die Dissoziation des Kofaktors vom Protein bei der Gleichgewichtseinstellung zwischen beiden Isomeren bestenfalls eine untergeordnete Rolle spielen. 5 5 10 10 20 20 15 15 Abbildung 7: Illustration der beiden Einbauisomere am Beispiel von Eisen-Protoporphyrin (FePPIX). Die Isomere können durch eine Rotation um die Achse durch C5 und C15 ineinander überführt werden. Da die Seitenkette der Aminosäure Val 68 bei einem Isomer über dem Ring I zu liegen kommt und bei dem anderen Isomer über dem Ring II, können die beiden Isomere anhand der NMR-Spektren voneinander unterschieden werden. 16 Grundlagen 1.4. Die Struktur von Porphyrinen Die grundlegende Struktur von Chlorophyllen und Protoporphyrin ist das Porphyrin, auch Porphin genannt (Abbildung 8). Es besteht aus vier Pyrrolringen, die durch vier Methinbrücken zu einem Makrozyklus verknüpft sind. Durch teilweise oder vollständige Substitution der acht Protonen in β-Position durch Alkylketten wird die Gruppe der freebase-Porphyrine gebildet. Werden die beiden zentralen Protonen durch ein zweiwertiges Metallion ersetzt, ergibt sich die Klasse der Metalloporphyrine. Der wichtigste Vertreter dieser Gruppe ist das Eisen-Protoporphyrin, welches zu der Klasse der Häm-Proteine gehört. Zu den Häm-Proteinen gehören neben den Sauerstofftransport-Proteinen Hämoglobin und Myoglobin auch die Gruppe der Cytochrome. Die Cytochrome sind sowohl in der Photosynthese als auch in der oxidativen Phosphorylierung wichtige Elektronentransportproteine. Reaktive Sauerstoff-Verbindungen wie Peroxide und Superoxide sind für Organismen schädlich[29] und werden über Katalasen und Peroxydasen beseitigt, die ebenfalls ein Hämgruppen als Kofaktor enthalten. A NH N B 5 3 4 2 1 NH 6 7 N 20 N HN D C 19 N 18 D 17 16 B 8 9 NH N N HN 10 HN 11 15 14 12 13 D E 2 1 13 13 A B C Abbildung 8: Oxidationsstufen des Porphins: A) Porphyrin oder Porphin, Grundstruktur aller Porphyrine mit der Bezeichnung der einzelnen Ringe. B) Durch Absättigen einer Doppelbindung entsteht das sogenannte Chlorin (17,18 Dihydroporphyrin), das Grundgerüst der Chlorophylle. C) Das Absättigen einer weiteren Doppelbindung führt zu Bakteriochlorin (7,8,17,18-Tetrahydroporphyrin) von dem die Bakteriochlorophylle abgeleitet sind. Der für diese Pigmente charakteristische isozyklische Ring E ist gestrichelt gezeichnet. 17 Grundlagen Durch Absättigung einer Doppelbindung eines Pyrrolringes wird die Struktur der Chlorine oder dihydro-Porphine gebildet. Die bekanntesten Vertreter dieser Gruppe sind die Chlorophylle, die zentral ein Magnesium(II)ion enthalten. Bei diesen Pigmenten schließt sich am Ring C ein isozyklischer Ring E an. Wird gegenüberliegend der ersten gesättigten Doppelbindung eine zweite Doppelbindung vollständig gesättigt, entsteht das Grundgerüst der Bakteriochlorine oder o(pposite)-tetra-hydro-Porphine. Die wichtigsten Vertreter dieser Klasse sind die Bakteriochlorophylle, die wie die Chlorophylle Magnesium(II)komplexe der freien Base sind. 1.4.1. Nomenklatur der Porphyrine Die Nomenklatur der Porphyrine nach den Regeln der International Union of Pure and Applied Chemsitry (IUPAC) ist aus historischen Gründen unübersichtlich. Die Nomenklatur nach Fischer, bei der nur die β-Positionen der Pyrrolringe nummeriert wurden, führte bei modifizierten Porphyrinen zu widersprüchlichen Namen und soll daher nicht mehr verwendet werden. Die systematische IUPAC Nomenklatur verursacht bei den verwendeten Ringsystemen jedoch häufig lange und unübersichtliche Namen und wird daher in der Praxis nur in Ausnahmefällen verwendet. Allerdings können die Porphyrine auch semisystematisch nach IUPAC benannt werden. Dabei kommen einige ausgewählte Trivialnamen der Fischer-Nomenklatur zum Einsatz, die von der IUPAC mit einem festgelegten Substitutionsmuster teilweise neu definiert wurden. Zu beachten ist, dass das Nummerierungssystem der systematischen und der semisystematischen Nomenklatur unterschiedlich sind (Abbildung 9). Protoporphyrin trägt in der semisystematischen Nomenklatur die Propionsäuren in den Positionen 13 und 17, während die gleichen Reste in der systematischen Nomenklatur durch die Ziffern 2 und 18 gekennzeichnet sind. Die in Abbildung 9 gezeigte Verbindung wurde nach Fischer Protoporphyrin IX genannt und ist heute nach der semisystematischen Nomenklatur der IUPAC als Protoporphyrin – ohne Nummerierung – zu bezeichnen. In dieser Arbeit wird diese Verbindung mit PPIX bzw. die daraus hergestellten Zinkkomplexe als ZnPPIX abgekürzt, um eine Verwechslung mit anderen Kofaktoren zu vermeiden. 18 Grundlagen H2C H3C CH 8 7 6 H H3C 10 NH 5 3 H2C 4 N 2 1 CH2 1112 13 N 14 OH 3 4 2 1 NH H3C CH H H H 19 17 18 H2C CH2 H2C CH 2 O O HO 5 CH3 6 7 N CH2 8 H HN 11 15 H OH CH 9 10 19 N 18 1716 H3C CH3 H 20 HN 16 20 CH CH3 15 H2C CH2 O 9 H 14 12 13 CH3 H2C CH 2 HO O Abbildung 9: Vergleich der systematischen (links) und der semisystematischen (rechts) Nomenklatur von Protoporphyrin (ehemals Protoporphyrin IX). Die Nummerierungen sind in beiden Systemen unterschiedlich. 3 2 H2C 3 1 CH 3 H3C 2 1 H 4 NH H CH3 5 6 7 N 8 8 9 20 10 19 N HN 11 18 14 12 H3C 16 17 13 15 2 171 2 1 17 13 13 H2C CH2 H 1 8 2 CH3 CH2 H CH3 O O O OPhytyl O CH3 Abbildung 10: Struktur von Phäophytin a. Der Magnesiumkomplex von Phäophytin a ist das Chlorophyll a. Die Chlorophylle werden ebenfalls nicht nach der systematischen Nomenklatur, sondern der semisystematische Nomenklatur nach IUPAC bezeichnet. Die in Abbildung 10 dargestellte Verbindung wird als Phäophytin a bezeichnet. Sie wird durch Demetallierung von Chlorophyll a erhalten und stellt somit die freie Base des Chlorophylls dar. Das Phäophytin a kann seinerseits als Phäophorbid a-Phytylester bezeichnet werden. Die freie Säure trägt daher den Namen Phäophorbid a, wobei der Buchstabe a das 19 Grundlagen Substitutionsmuster beschreibt. Die Metallkomplexe können semisystematisch als Koordinationskomplexe der freien Base benannt werden. Dementsprechend wird der Name Zink-Phäophorbid a (ZnPheid a) für den Komplex von Zink(II) und Phäophorbid a verwendet. Wird die Estergruppe in Position 132 entfernt, sind die Verbindungen als 132-demethoxycarbonyl-Verbindungen zu bezeichnen. In vielen Fällen werden diese Verbindungen PyroVerbindungen genannt, da sie durch eine Esterpyrolyse dargestellt werden. In dieser Arbeit trägt das Molekül, das in der 132-Position keine Esterfunktion mehr besitzt, daher den Namen Pyrophäophorbid a (PPheid a) und der korrespondierende Zinkkomplex den Namen Zink-Pyrophäophorbid a (ZnPPheid a). Wird die Säurefunktion in Position 17 verestert, z. B. als Methylester, wird die Verbindung als Methyl-Pyrophäophorbid a (MePPheid a) bezeichnet. 1.4.2. Elektronische Struktur der Porphyrine Das absorptionsspektroskopische Verhalten der Porphyrine, Chlorophylle und Bakteriochlorophylle lässt sich qualitativ gut mit der 4-Orbital-Modell von Gouterman[30] beschreiben. Dieses Modell wurde zunächst für hochsymmetrische Metalloporphyrine (D4h-Symmetrie) entwickelt und später für Chlorophyll- und Bakteriochlorophyllgerüste mit geringerer Symmetrie modifiziert. Man erhält zwei vollständig besetzte Orbitale a2u(π) und a1u(π) (HOMO und HOMO–1) und zwei unbesetzte Orbitale egx(π*) und egy(π*) (LUMO und LUMO+1). Die Absorptionsbanden im UV-Vis- und NIR-Bereich entstehen durch Übergänge zwischen diesen Orbitalen. Es kommt zu vier möglichen Übergängen. Zwei dieser Übergänge sind in x-Richtung (a1u(π) → egy(π*), a2u(π) → egx(π*)) polarisiert und die beiden anderen sind in y-Richtung (a1u(π) → egx(π*), a2u(π) → egy(π*)) polarisiert. Die hochenergetischen Übergänge a2u(π ) → egy(π*) (By) und a1u(π) → egy(π*) (Bx) werden aus historischen Gründen mit B gekennzeichnet, die niederenergetischen Übergänge werden dagegen mit Q bezeichnet [a2u(π) → egx(π*) (Qx) und a1u(π) → egx(π*) (Qy)]. Die vier Molekülorbitale (MO) für Porphyrin sind in Abbildung 11 dargestellt. In der D4h-Symmetrie liegen die HOMOs nah beieinander und die LUMOs sind entartet. Daher 20 Grundlagen sind die Übergänge der Bx- und der By-Bande erlaubt, die Übergänge der Qx- und der Qy-Bande jedoch verboten. Das Resultat ist eine intensive Soret-Bande und zwei schwache Banden, die durch Schwingungsverzerrung des Moleküls teilweise erlaubt sind. Abbildung 11: Elektronendichteverteilung der einzelnen Bakteriochlorophyll (rechts) nach dem 4-Orbital-Modell [31] Orbitale von Porphyrin (links) und . Wird eine Doppelbindung des Makrozyklus abgesättigt, so sind die MOs betroffen, die an dieser Position Elektronendichte aufweisen. Bei den Chlorophyllen sind dies die MOs a1u und egy, die in Folge dessen energetisch höher liegen. Dies bedeutet, dass die Übergänge mit einem Übergangsdipolmoment entlang der x-Achse eine starke Bx-, aber eine schwache Qx-Bande erzeugen. Die Übergänge mit einem Übergangsdipolmoment entlang der y-Achse erzeugen dagegen eine By- und eine Qy-Bande mit vergleichbarer Intensität. Die Bx- und die By-Bande überlagern sich und bilden gemeinsam die intensive SoretBande. Der Übergang für die Qx-Bande ist verboten, daher ist sie schwach. Wird zusätzlich eine zweite Bindung hydriert, wird dieser Effekt verstärkt. Dadurch kann die Verschiebung der B-Bande (Soret) ins Blaue und der Qy-Bande ins Rote mit zunehmender Zahl an reduzierten Doppelbindungen erklärt werden. 21 Grundlagen Abbildung 12: Die Energie der Molekülorbitale ändert sich abhängig von der Zahl der Doppelbindungen im Porphyrin, Chlorophyll und Bakteriochlorophyll. Durch die aufgehobene Entartung (eg) wird der Übergang der Qy-Bande erlaubt wodurch im UV-Vis-Spektrum eine intensive Bande auftritt. Die freien Basen der Porphyrine sind vierzähnige Chelatliganden, die nach ihrer Deprotonierung zum zweifach negativ geladenen Anion Metallionen binden können. Wesentlich für die Bindung eines Metallions ist der Ionenradius, da das planare Porphyrin Ionen mit einem Radius von 60 bis 70 pm optimal binden kann. Ist das Metallion kleiner, wird es lediglich schwach koordiniert. Ist es größer als 70 pm kann das Metallion nicht in der Ringebene gebunden werden, wodurch eine sogenannte out-of-plane-Koordination verursacht wird. Durch diese asymmetrische Koordination wird das Ringsystem verzerrt, wobei drei Formen zu unterscheiden sind. Eine Wölbung des gesamten Ringsystems in einer Richtung („doming“), ein sattelförmiger Makrozyklus, bei dem die Stickstoffe zweier Fünfringe nach oben und zwei Stickstoffe nach unten aus der Ebene gedreht werden, und eine Verdrehung („ruffling“) des Makrozyklus[32]. Weil die biologische Funktion eines Prophyrins maßgeblich durch das zentrale Metallatom bestimmt wird, werden im folgenden die Eigenschaften der Metalle Eisen, Magnesium und Zink und die daraus resultierenden Unterschiede in der Funktion exemplarisch vorgestellt. Eisen ist eines der wichtigsten Metalle, die durch den Prophyrin-Makrozyklus gebunden werden. Eisenporphyrine kommen außer in Myoglobin und Hämoglobin auch in Cytochromen, Katalasen und Peroxidasen vor. Im gebundenen Zustand existiert es in vier 22 Grundlagen unterschiedlichen Formen. In den beiden Oxidationszuständen +2 und +3 treten abhängig von den Liganden jeweils der high-spin und der low-spin Zustand auf[33]. Der Vergleich der Ionenradien zeigt, dass die Oxidationszahl +2 in der high-spin Form mit 78 pm deutlich größer als die „Lücke“ im Porphyrinsystem ist und daher nur in der out-of-planeStruktur gebunden werden kann, wie es in nativem desoxy-Myoglobin der Fall ist. Durch Bindung von Sauerstoff geht das Eisen in die low-spin Form über, wobei allerdings bis heute nicht genau bekannt ist, ob es im Oxidationszustand +2 oder +3 vorliegt[34; 35]. In beiden Oxidationszuständen ist der Ionenradius mit 61 pm (Fe(II) low-spin) und 55 pm (Fe(III) low spin) deutlich kleiner als in der desoxy-Form, wodurch das Metallion in die Ringebene wandert und die Bindung des Sauerstoffmoleküls durch eine elektrostatische Wechselwirkung mit dem distalen Histidin des Proteins stabilisiert wird. Für ein Elektronentransport-Protein wie das Cytochrom c wäre eine unbesetzte Bindungsstelle am zentralen Eisen nachteilig. Für eine schnelle Redoxreaktion ist eine Änderung der Struktur ungünstig, da hierfür eine Reorganisationsenergie aufgewendet werden muss, die die Effizienz des Elektronentransfers reduziert. Daher findet man bei den meisten redox-aktiven Häm-Proteinen ein koordinativ gesättigtes Eisen im low-spinZustand vor und das Eisen ist aufgrund seines kleinen Ionenradius in der Ringebene gebunden. Durch eine ein-Elektronen-Übertragung ändert sich der Ionenradius nur um etwa 6 pm von 55 pm (Fe3+) auf 61 pm (Fe2+), dadurch tritt keine Strukturänderung bzw. keine Reorganisationsenergie bei einer Elektronenübertragung auf. Entsprechend ist eine hohe Effizienz für den Elektronentransfer die Folge. In der pflanzlichen Photosynthese kann Eisen kein Ersatz für Magnesium sein, weil es nicht redox-inert ist und daher die Energieweiterleitung bzw. die Ladungstrennung in den Photoreaktionszentren durch einen intramolekularen Elektronentransfer verhindert würde. Das ideale Metallion für ein Porphyrinsystem, das an photosynthetischen Prozessen beteiligt ist, muss also redox-inert und passend für das Porphyrin sein. Von den Hauptgruppenelementen besitzt einzig Mg2+ eine entsprechende Größe von 72 pm. Es unterscheidet sich von Zn2+ (74 pm) nur geringfügig bezüglich des Ionenradius, besitzt aber eine geringere Atommasse. Aufgrund der daraus resultierenden geringeren SpinBahn-Kopplung für das Mg2+-Ion tritt die Bildung eines Triplettzustandes nach Lichtanregung durch „inter-system crossing“ (ISC) deutlich seltener auf als für das Zn2+Ion. Der langlebige Triplettzustand verlangsamt die notwendigerweise sehr schnellen 23 Grundlagen Primärereignisse der Photosynthese (Lichtsammlung und Ladungstrennung) und erzeugt damit eine Konkurrenzreaktion von unerwünschter Licht- bzw. Wärmeabgabe. Ist umgekehrt eine möglichst hohe Triplettausbeute das Ziel der Lichtanregung, wie es in dieser Arbeit der Fall ist, ist die Verwendung von Zink als zentrales Metallion vorteilhaft. 1.5. Eisen-Schwefel-Zentren in der Natur Eisen-Schwefel Proteine kommen in allen Lebewesen vor[36]. Am häufigsten sind sie an Elektronentransferprozessen beteiligt wie z. B. in den Ferredoxinen. Es sind allerdings auch die katalytische Aktivität in der Aconitase und Sensorfunktionen für NO, Sauerstoff und Eisen bekannt[37]. In der Endonuklease III besitzt das Eisen-Schwefel-Zentrum ausschließlich eine strukturelle Funktion. Die Eisen-Schwefel Komplexe bestehen aus Eisen und anorganischem Schwefel in Form von Sulfidionen. Diese Strukturen werden durch Cysteine an das Protein gebunden. Von den einfachsten Eisen-Schwefel Komplexen sind gegenwärtig vier Formen bekannt. Die Rubredoxine enthalten einen einkernigen [FeCys4] Komplex. Zweikernige [2Fe2SCys4]-Zentren kommen in den pflanzlichen Ferredoxinen vor, zu denen auch der sogenannte Rieske-Typ der Eisen-Schwefel-Zentren zu zählen ist, bei dem zwei der Cysteinliganden durch Histidin ersetzt sind [2Fe2SCys2His2]. Ein Eisen-Schwefel-Zentrum vom Rieske-Typ liegt zum Beispiel im Cytochrom b6f-Komplex vor. Außerdem sind noch drei- und vierkernige Komplexe mit der Struktur [3Fe4SCys4] und [4Fe4SCys4] bekannt, die einzeln oder gemeinsam in Ferredoxinen und katalytisch aktiven Proteinen als redox-aktive Kofaktoren vorkommen. Die Hydrogenase, sowie die Nitrogenase, besitzen sowohl [3Fe4S] als auch [4Fe4S]Zentren. In allen Komplexen ist das Eisen tetraedrisch durch Schwefel koordiniert, die zweikernigen Komplexe besitzen die Struktur von kantenverknüpften Tetraedern, während die drei- und vierkernigen Komplexe eine kubische Struktur besitzen (Abbildung 13), wobei bei den dreikernigen Komplexen eine Ecke des Kubus nicht besetzt ist. Neben der eigentlichen Ligandierung des Kubus und seiner dreidimensionalen Struktur, sind Wasserstoffbrückenbindungen zwischen dem Amidproton einer Peptidbindung und einem 24 Grundlagen Schwefelatom des Kubus ein wichtiges Merkmal dieser Kofaktoren[38]. Die Zahl und Länge dieser Bindungen ist ausschlaggebend für die Art des Eisen-Schwefel-Zentrums, das vom Protein gebunden wird, und dafür, welches Redoxpaar der [4Fe4S]-Zentren im nativen Zustand zugänglich ist. In der Regel werden Abstände von 2,3 Å bis 2,8 Å in unterschiedlichen Ferredoxinen gefunden. 2+ + Fe2.5+ Fe3+ Fe2+ Fe3+ Fe2+ + 1e- 1e- S=0 Fe2+ Fe2.5+ Fe2+ S = 1/2 Abbildung 13: Die Struktur eines [4Fe4S]-Zentrums im oxidierten und reduzierten Zustand zeigt die Positionen der gemischt-valenten Eisenpaare. Die Eisenatome (schwarze Kugeln) und Sulfidatome (graue Kugeln), sowie die β-Kohlenstoffe der Cysteinliganden (kleine schwarze Kugeln) sind gezeigt. Mit Ausnahme der [4Fe4S]-Zentren besitzen die Eisen-Schwefel-Zentren zwei Redoxzustände, bei denen im oxidierten Zustand alle Eisenatome formal in der Oxidationsstufe +3 vorliegen und durch die Reduktion des Komplexes ein Eisenatom in die Oxidationsstufe +2 übergeht. Die [4Fe4S]-Zentren können drei unterschiedliche Redoxzustände annehmen, wobei in nativen Proteinen grundsätzlich nur zwei Oxidationsstufen zugänglich sind. In oxidierter Form besitzt der Komplex formal die Ladung [4Fe4S]3+. Dabei liegen drei Eisenatome in der Oxidationsstufe +3 und eins in der Stufe +2 vor. Durch eine Ein-Elektronen-Reduktion geht der Komplex in die Oxidationsstufe [4Fe4S]2+ über, in der zwei Eisenatome die Ladung +2 und zwei die Ladung +3 tragen. Durch eine weitere Reduktion kann der Komplex in seine niedrigste Oxidationsstufe überführt werden, in der formal drei Kerne die Ladung +2 und einer die Ladung +3 tragen. Anhand von Mössbauer- und EPR-Studien konnte jedoch gezeigt werden, dass die Zentren besser durch zwei Kerne in der Oxidationsstufe +2 und zwei Kerne in der Oxidationsstufe +2,5 beschrieben werden können (Abbildung 13). 25 Grundlagen Welches der beiden Redoxpaare - 3+/ 2+ oder 2+/ 1+ - in nativen Proteinen zugänglich ist, wird maßgeblich von der Proteinstruktur bestimmt[39]. Die Redoxpotentiale werden durch eine Kombination aus Coulomb-Wechselwirkung des geladenen Komplexes und geladener Aminosäuren in unmittelbarer Nähe, der Polarisierbarkeit des Proteins und der Wechselwirkung des Komplexes sowie des Proteins mit dem Lösemittel bestimmt[40-42]. Die Redoxpotentiale von Eisen-Schwefel-Zentren decken einen Bereich von +400 mV bis –700 mV ab[7; 42] . Die Proteine, deren Eisen-Schwefel-Zentren im Redoxpaar 3+/ 2+ zugänglich vorliegen, besitzen in der Regel ein positives Redoxpotential, weshalb sie als High Potential Iron-Sulfur Proteins, HiPIP´s, bezeichnet werden. Der Grundzustand eines [4Fe4S]2+-Zentrums ist diamagnetisch durch eine antiferromagnetische Kopplung der Eisenatome untereinander, woraus ein effektiver Gesamtspin von S=0 resultiert. Im reduzierten Zustand ergibt sich aus dieser Kopplung ein effektiver Gesamtspin von S=1/2[43]. Obwohl formal drei Fe3+ und ein Fe2+ vorliegen, ist das ungepaarte Elektron über zwei Eisenatome in einem gemischt-valenten Oxidationszustand +2,5 delokalisiert[44]. In gefrorenen Lösungen ist ein rhombisches EPRSpektrum typisch für ein [4Fe4S]1+-Zentrum. 1.6. Die Stabilität von Proteinen Die Stabilität von Proteinen gegenüber der Denaturierung liefert wichtige Hinweise über deren strukturelle Eigenschaften, insbesondere bei Veränderungen gegenüber dem nativen Zustand, wie zum Beispiel dem Einbau eines nicht-nativen Kofaktors. Erhält man nach einer Modifikation identische Stabilitäten, kann von einer ähnlichen Faltung ausgegangen werden. Ändert sich die Stabilität eines Proteins jedoch, muss von einer signifikanten Änderung in der Struktur des Proteins ausgegangen werden. Die Änderung der Freien Gibbs-Enthalpie während der Entfaltung gibt den Energieunterschied (∆G) zwischen dem gefalteten und dem entfalteten Zustand wieder, während die Änderung von ∆G mit der Konzentration des Denaturierungsreagenzes ein Maß für die Kooperativität des Entfaltungsprozesses ist. In dieser Arbeit wird das chaotrop wirkende Guanidiniumhydrochlorid als Denaturierungsreagenz verwendet, das die Wasserstoffbrückenbindungen der sekundären 26 Strukturelemente zerstört. Die Grundlagen Entfaltungskurven werden durch ein Zwei-Zustandsmodell beschrieben. Es wird das Gleichgewicht zwischen gefaltetem Protein und entfaltetem Protein betrachtet. Die Freie Entfaltungsenthalpie ist dann gegeben durch Gleichung (1). ∆G = − RT ln K mit K = [entfaltet] 1 − f = [gefaltet] f (1) Zur Auswertung muss der Anteil an gefaltetem Protein f den gemessenen Spektren entnommen werden. Dabei sind zwei Bedingungen zu beachten. a) Im Ausgangszustand muss das Protein in einem vollständig gefalteten Zustand vorliegen und b) am Ende der Entfaltung muss das Protein tatsächlich vollständig entfaltet sein. Während die zweite Bedingung überprüft werden kann, indem die Konzentration an Denaturierungsreagenz kontinuierlich erhöht wird, bis konstante Messwerte erhalten werden, ist die erste Bedingung eine nicht überprüfbare Annahme. Weiterhin muss angenommen werden, dass die Freie Entfaltungsenthalpie linear von der Konzentration an Denaturierungsreagenz abhängt[45; 46]. ∆G = ∆G0 − m ⋅ [GdnHCl ] (2) In Gleichung 2 entspricht ∆G0 der Energie, die aufgebracht werden muss, um das Protein in wässriger Lösung ohne Zusatz von Denaturierungsreagenz zu entfalten. Die Kooperativität m gibt an, wie stark die Stabilität eines Proteins im Verlauf der Denaturierung von der bereits vorliegenden Entfaltung beeinflusst wird. Bei einer stark kooperativen Entfaltung (mit großen Werten für m) führen bereits kleine Störungen der Struktur zu einer vollständigen Entfaltung. Durch Kombination von Gleichung 1 und 2 ergibt sich Gleichung 3 wie folgt. ln K = − ∆G0 + m[GdnHCl ] RT 1− f ⎛ − ∆G0 + m[GdnHCl ] ⎞ = exp⎜ ⎟ f RT ⎝ ⎠ 27 Grundlagen f = 1 ⎛ − ∆G0 + m[GdnHCl ⎞ exp⎜ ⎟ +1 RT ⎝ ⎠ (3) Der Anteil an gefaltetem Protein f im Gleichgewichtszustand kann nach Gleichung 4 berechnet werden. f = xobs − x entf (4) x gef − xentf Dabei stehen xentf und xgef für die Messgröße im vollständig entfalteten bzw. gefalteten Zustand und xobs für den Messwert bei einer bestimmten Konzentration an Denaturierungsreagenz. Die beobachtete Messgröße richtet sich nach der verwendeten Methode. Bei Detektion durch Fluoreszenzspektroskopie wird häufig die Verschiebung der Fluoreszenzbande der Aminosäure Tryptophan beobachtet, die bei nativen Proteinen in einem Bereich von 330 nm im gefalteten Zustand bis 360 nm im entfalteten Zustand variiert. Allerdings sind auch kleinere Verschiebungen von 8 nm ausreichend, um aussagekräftige Werte für eine Entfaltungsstudie zu erhalten. Bei der CircularDichroismus- (CD) Spektroskopie wird das Signal eines Sekundärstrukturelements, z. B. einer α-Helix bei 208 und 222 nm, detektiert. Entfaltet sich die Sekundärstruktur des Proteins, nimmt das CD-Signal auf Null ab. Mit Hilfe der UV-Vis-Spektroskopie kann eine beliebige Absorptionsbande, die ausreichend sensibel auf eine Umgebungsänderung des Kofaktors reagiert, gewählt werden. Bei Tetrapyrrolen ist dies beispielsweise die SoretBande. Die ermittelten Messpunkte werden nach Gleichung 3 interpoliert, wodurch Werte für ∆G0 und m bestimmt werden können. 28 Grundlagen 1.7. Die Festphasen Peptidsynthese (SPPS) Die SPPS (solid phase peptide synthesis) wird in dieser Arbeit verwendet, um die Modelle der Eisen-Schwefel-Zentren zu synthetisieren. Mit ihr können auf einer funktionalisierten, polymeren festen Phase Polypeptide bis zu einer Länge von 60 Aminosäuren mit guten Ausbeuten synthetisiert werden. Sie wurde ursprünglich von Merrifield entwickelt[47] und ist seitdem in vielerlei Hinsicht optimiert worden. Im wesentlichen werden zwei verschiedene Verfahren angewendet. Die Fmoc-Strategie (Fmoc steht für 9-Fluorenylmethoxycarbonyl), bei der N-terminale Aminogruppen durch die Fmoc-Gruppe geschützt werden und die Boc (tert-Butoxycarbonyl) Strategie, bei der die Säure-labile Boc-Gruppe zum Schutz der N-terminalen Aminogruppen verwendet wird. Obwohl die Boc-basierte Synthese häufig bessere Ausbeuten liefert und auch mehr Möglichkeiten zur Modifikation der Peptide bietet, ist die Fmoc-Synthese in den letzten Jahren häufiger eingesetzt worden. Dies liegt vor allem darin begründet, dass für die Peptidsynthese im Rahmen der Fmoc-Strategie keine Flusssäure (HF) zur Gewinnung der entschützten Peptide verwendet werden muss, wie dies in der Boc-Synthese der Fall ist. Durch den verstärkten Einsatz der Fmoc-Synthese und dem daraus resultierenden ökonomischen Zwang, ein vergleichbares Methodenregister zur Verfügung zu haben, sind die meisten Nachteile der Fmoc-Synthese in den letzten Jahren gelöst worden. Eine gute Übersicht über die gesamte Fmoc-Synthese bieten W.C. Chan et al.[48]. P. LloydWilliams et al.[49] zeigen Unterschiede und Gemeinsamkeiten zwischen Fmoc- und BocStrategie auf. Im Folgenden soll ausschließlich auf die in dieser Arbeit verwendete FmocStrategie näher eingegangen werden. Zu Beginn der Synthese wird zunächst die polymere feste Phase, das sogenannte Harz, im Lösemittel gequollen, um eine optimale Zugänglichkeit der funktionellen Gruppen zu erreichen. Anschließend erfolgt die Abspaltung der Fmoc-Gruppe durch die Base Piperidin (Abbildung 14), die die Schutzgruppe an der Position 9 deprotoniert, wobei ein anionisches System mit aromatischem Charakter entsteht, welches durch Umlagerung zum Dibenzofulven stabilisiert wird. Als zweites Spaltprodukt bildet sich eine Carbaminsäure, welche durch Decarboxylierung die Aminogruppe freisetzt. Diese Reaktion verläuft innerhalb von 15 Minuten quantitativ. 29 Grundlagen O H NH O O H2 N H N H2 N NH O O NH O H 2N + CO2 N Abbildung 14: Entschützen des N-Terminus während der Peptidsynthese. Durch die Base Piperidin wird die Fmoc-Gruppe deprotoniert und die Aminogruppe freigesetzt. Die Kopplung der nächsten Aminosäure mit dem freien N-Terminus erfordert eine Aktivierung, da sonst aufgrund der pKS-Werte der Amino- und der Carboxylgruppe lediglich ein Carboxylatanion und eine protonierte Aminogruppe gebildet werden. Die Aktivierung erfolgt durch Aminiumbenzotriazol-Derivate (vgl. Abbildung 16), wie in Abbildung 15 gezeigt. Zunächst greift die Aminosäure nukleophil das Aminiumbenzotriazolderivat an und bildet das Addukt, welches durch die Base Diisopropylethylamin (DIPEA) deprotoniert wird und anschließend in das AminosäureHarnstoff-Addukt und Benzotriazol-N-oxid zerfällt[50]. Das Benzotriazol-N-oxid ist ein starkes Nukleophil und verdrängt den Harnstoff, der eine gute Abgangsgruppe darstellt. Der Ester der Aminosäure und des Benzotriazol-N-oxids ist reaktiv genug, so dass er nukleophil durcheine freie Aminogruppe angegriffen werden kann, wodurch letztendlich die Peptidbindung gebildet wird. Das resultierende Amid ist relativ unreaktiv und wird dadurch dem Gleichgewicht entzogen. Die Ausbeute pro Kopplungsschritt liegt über 98 %. Im nächsten Synthesezyklus wird erneut der N-Terminus entschützt und mit einer weiteren Aminosäure gekoppelt. Nach der kompletten Synthese wird das Peptid mit Trifluoressigsäure (TFA) vom Harz gespalten, wobei zeitgleich die Seitenketten der 30 Grundlagen Aminosäuren entschützt werden. Die bei dieser Reaktion entstehenden Carbeniumionen und Radikale werden durch verschiedene schwefelhaltige Reagenzien abgefangen (z.B. Thioanisol, Ethandithiol). NHFmoc (CH 3)2N R OH N(CH3) 2 (CH 3)2N N O R O H O N FmocHN NHFmoc R O +DIPEA - DIPEAH+ N(CH3) 2 N N O (CH3)2N + N O N(CH3) 2 N N N O N O NHFmoc R NHFmoc O N O OH R N + N O +H2N N N N O + (CH3)2N N(CH3) 2 N Abbildung 15: Reaktionsmechanismus von des Kopplungsreagenz HBTU mit einer Aminosäure zur Bildung eines aktivierten Esters und Kopplung an einen freien Peptid N-Terminus. O O Cl OMe Cl NH2 OH OMe Wang Resin Trialkoxybenzylamine (PAL) Resin 4-Chlorotrityl chloride Resin PF6(H3C)2N O P N N(CH3) 2 N N N 3 N N N - PyBOP O TBTU (H3C)2N N(CH3) 2 N BF4- N PF6- N N OHATU Abbildung 16: Strukturen einiger gebräuchlicher Harze (oben) und eine Auswahl häufig verwendeter Kopplungsreagenzien (unten) mit ihren üblichen Abkürzungen (vgl. Abkürzungsverzeichnis). 31 Grundlagen Der N-Terminus des Peptides liegt nach der Synthese als freie Aminogruppe vor. Die Aminogruppe kann durch Acetanhydrid acetyliert werden, um die Ausbildung einer positiven Ladung an dieser Position zu vermeiden, wodurch in synthetischen Peptiden häufig eine Stabilisierung der gewünschten Tertiärstruktur erreicht wird. Der C-Terminus kann durch die Wahl des Harzes modifiziert werden. Zur Zeit ist eine große Auswahl unterschiedlicher Harze kommerziell erhältlich, die verschiedene Modifikationen ermöglichen (Abbildung 16). Auf diese Weise kann der C-Terminus des Peptides als Carbonsäure, Carbamid, Ester oder Thioester dargestellt werden. In dieser Arbeit wurde ein PAL-PEG-PS Harz verwendet, weshalb der C-Terminus nach der Abspaltung des Peptides amidiert vorliegt. Die Seitenketten der Aminosäuren, die Heteroatome oder funktionelle Gruppen enthalten, müssen mit Schutzgruppen versehen werden, um Nebenreaktionen und Verzweigungen der Peptide während der Synthese zu vermeiden. Dazu stehen eine Vielzahl orthogonaler und nicht orthogonaler Schutzgruppen zur Verfügung. Die in dieser Arbeit verwendeten Schutzgruppen sind alle labil gegen TFA und daher nicht orthogonal (Tabelle 1). 32 Grundlagen Tabelle 1: Übersicht über die in dieser Arbeit verwendeten Schutzgruppen für die Seitenketten der Aminosäuren. Aminosäuren, die nicht in dieser Liste enthalten sind, wurden ungeschützt eingesetzt. Alle Schutzgruppen sind gegen TFA labil. Aminosäure Abkürzung der Schutzgruppe Schutzgruppe 2,2,4,6,7-pentamethyldihydrobenzofuran-5sulfonyl Arginin O R Pbf S O O Asparagin Trityl Cystein Trityl Histidin Trityl Lysin Boc Triphenylmethyl R tert-Butoxycarbonyl O Tryptophan Boc Tyrosin tert-Butyl Aspartat tert-Butyl Serin tert-Butyl Threonin tert-Butyl R O tert-Butyl R 33 Grundlagen 1.8. Grundlagen der EPR- und ENDORSpektroskopie Eine kurze Einführung in die Grundlagen der Elektronenspinresonanz (EPR)- und die Electron Nuclear Double Resonance (ENDOR)-Spektroskopie soll die wesentlichen Merkmale dieser spektroskopischen Methoden darlegen. Ausführliche theoretische Grundlagen sind in verschiedenen Lehrbüchern detailliert beschrieben[51; 52]. Die EPR-Spektroskopie setzt voraus, dass ungepaarte Elektronen in der zu untersuchenden Verbindung vorliegen. Die EPR wird zur Untersuchung und Charakterisierung von Radikalen oder paramagnetischen Übergangsmetallkomplexen verwendet. Bei der Untersuchung von Triplettzuständen ist die EPR-Spektoskopie und die eng verwandte ENDOR-Spektroskopie äußerst nützlich, da beide Methoden unmittelbar Informationen über die Verteilung der ungepaarten Elektronen auf dem Molekülgerüst geben. Zunächst wird der einfachste Fall, ein System mit nur einem ungepaarten Elektron (Elektronenspinquantenzahl S=1/2), betrachtet. In erster Näherung liegt dieser Fall bei den Eisen-Schwefel-Peptiden vor, die in dieser Arbeit untersucht wurden. Im weiteren Verlauf soll auf die wesentlichen Unterschiede zum Triplettzustand, wie er in den untersuchten Myoglobin-Modellsystemen auftritt, kurz eingegangen werden. Insbesondere werden die Besonderheiten eines Triplettzustandes für die ENDOR-Messungen kurz zusammengefasst. 1.8.1. Grundlagen der EPR-Spektroskopie Ein freies Elektron besitzt in einem äußeren Magnetfeld genau zwei Ausrichtungsmöglichkeiten, entweder parallel oder antiparallel, zu einem Magnetfeld. Die Energie E der beiden unterschiedlichen Ausrichtungen sowie ihre Energiedifferenz ∆E wird durch Gleichung (5) beschrieben. 1 E ± = ± gβ e B0 ⇒ ∆E = gβ e B0 2 (5) 34 Grundlagen Dabei steht βe für das Bohrsche Magneton, B0 für das externe Magnetfeld und g für eine Proportionalitätskonstante, den g-Wert. Für ein freies Elektron liegt der g-Wert bei ge = 2,0023. Definitionsgemäß ist die antiparallele Ausrichtung des Elektronenspins energetisch günstiger. Der Besetzungsunterschied beider Energieniveaus wird durch die Boltzmann-Verteilung (Gleichung 6) beschrieben. n n + 1 2 =e − gβ e B0 kT (6) 1 − 2 Die energetische Aufspaltung zwischen diesen beiden Energieniveaus wird als ElektronZeeman-Aufspaltung bezeichnet. In einem EPR-Experiment induziert die Einstrahlung elektromagnetischer Wellen Übergänge zwischen den Spinniveaus, wenn die Resonanzbedingung entsprechend Gleichung (7) erfüllt ist. ∆E = hν = gβ e B0 (7) Bei einem externen Magnetfeld von 350 mT und einem g-Wert, der dem des freien Elektrons entspricht (g = ge) liegt die Resonanzfrequenz bei etwa 9,5 GHz, also im X-Band des Mikrowellenbereichs (Wellenlänge λ ≈ 3 cm). Die experimentellen Ergebnisse einer EPR-spektroskopischen Charakterisierung werden häufig durch den Formalismus des Spin-Hamilton-Operators beschrieben, der alle Wechselwirkungen, die zu einem EPR Spektrum beitragen, zusammenfasst. Die einzelnen Terme des Spin-Hamilton-Operators werden im Folgenden kurz vorgestellt. Elektron-Zeeman-Wechselwirkung In den EPR-Spektren von Übergangsmetallkomplexen findet man in der Regel deutliche Abweichungen vom g-Wert des freien Elektrons. Zusätzlich wird beobachtet, dass die Elektron-Zeeman-Aufspaltung von der Orientierung der Probe im Magnetfeld abhängig ist. Dies ist durch das Bahnmoment L eines Elektrons begründet, welches ein eigenes magnetisches Moment besitzt. Dessen Beitrag zur Elektron-Zeeman-Aufspaltung hängt 35 Grundlagen von der Spin-Bahn-Kopplung und der energetischen Lage der Orbitale ab. Der Term für die Elektron-Zeeman-Wechselwirkung im Spin-Hamilton-Operator lautet r r H EZ = β e B0 g Sˆ . (8) Wobei Ŝ der Operator des effektiven Spins ist. Der g-Tensor kennzeichnet die Symmetrie und die elektronische Struktur eines Übergangsmetallkomplexes. Abbildung 17: Elektron-Zeeman-Aufspaltung eines ungepaarten Elektronenspins S=1/2 in einem statischen Magnetfeld B0. Bei der Einstrahlung elektromagnetischer Wellen der Energie ∆E erfolgt eine Absorption, da das energetisch tiefere Niveau nach der Boltzmann-Gleichung stärker populiert ist. Nullfeldaufspaltung Liegen mehrere ungepaarte Elektronen vor, so kommt es bereits ohne externes Magnetfeld zu einer Aufspaltung der Energieniveaus des elektronischen Grundzustandes. In diesem Fall spricht man von einer Nullfeldaufspaltung (zero field splitting, zfs). Der entsprechende Term im Hamilton-Operator wird durch Gleichung (9) wiedergegeben. H ZFS r r = SˆDSˆ (9) 36 Grundlagen Der Nullfeldtensor D kann auch in Form der Nullfeldparameter D und E ausgedrückt werden. H ZFS r 1 ˆ2 2 ˆ = D( S z − S ) + E ( Sˆ y2 − Sˆ x2 ) 3 (10) Bei Triplettzuständen von organischen Molekülen wird die Nullfeldaufspaltung in erster Linie durch die dipolare Wechselwirkung der ungepaarten Elektronen bestimmt. Die Nullfeldparameter D und E können deswegen zur Charakterisierung der Elektronenspinverteilung in der Triplettwellenfunktion herangezogen werden. Hyperfeinwechselwirkung Liegen in der zu untersuchenden Verbindung Kerne mit einem Kernspin von I ≠ 0 vor, so kann eine Wechselwirkung zwischen Elektronenspins und Kernspins auftreten, die als Hyperfeinwechselwirkung bezeichnet werden. Der Term des Spin-Hamilton-Operator für die Hyperfeinwechselwirkung ist durch Gleichung (11) beschrieben. H HFC r r = SˆAIˆ . (11) In dieser Gleichung steht Iˆ für den Vektoroperator des Kernspins und A für den Hyperfeintensor. Der Hyperfeintensor kann formal in einen isotropen Anteil aiso und einen anisotropen Anteil A´ zerlegt werden. Die isotrope Kopplungskonstante aiso wird durch die Fermi-Kontakt-Wechselwirkung hervorgerufen und ist proportional zur Spindichte des ungepaarten Elektrons am Kern |ψns(0)|. E Kontakt = − 8π 2 ψ ns (0) µ e µ N 3 (12) bzw. H Kontakt rr rr 8π 2 ˆ ˆ = ( ) g e β e g N β N ψ ns (0) S I = aiso Sˆ Iˆ 3 37 (13) Grundlagen Der Term für die isotrope Hyperfeinwechselwirkung ist nicht winkelabhängig und seine absolute Größe hängt maßgeblich von der Größe des s-Charakters der jeweiligen Wellenfunktion ab. Für reine p- und d-Orbitale ist die Aufenthaltswahrscheinlichkeit am Kern gleich Null, so dass keine isotrope Hyperfeinwechselwirkung auftreten kann. In der Praxis wird jedoch auch für viele Verbindungen ein isotroper Anteil der Hyperfeinwechselwirkung gefunden, die diese nicht aufweisen sollten. Dies kann durch eine Konfigurationswechselwirkung oder eine Spinpolarisation hervorgerufen werden. Beide Mechanismen bewirken eine endliche Aufenthaltswahrscheinlichkeit des ungepaarten Elektrons am Kernort. Die anisotrope Hyperfeinwechselwirkung liegt in der Wechselwirkung zwischen den magnetischen Momenten der Elektronen und der Kerne begründet. Sie kann daher analog zur klassischen Dipol-Dipol-Wechselwirkung hergeleitet werden (vgl. Gleichung 14). E Dipol = r r µeµ N r3 r r r r 3( µ e r )( µ N r ) − r5 (14) r wobei r der Radiusvektor von µe zu µN ist und r der Abstand zwischen Kern und Elektron ist. In Operatorschreibweise ergibt sich daraus Gleichung (15). H Dipol rr r r ⎡ IˆSˆ 3( Iˆrr )( Sˆrr ) ⎤ ⎥ = −ge βe g N β N ⎢ 3 − r5 ⎢r ⎥ ⎣ ⎦ (15) Dieser Term ist abhängig vom Winkel des äußeren Magnetfeldes relativ zum untersuchten Molekül. Kern-Zeeman-Wechselwirkung Der Kern-Zeeman-Term des Spin-Hamilton-Operators wird analog zum Elektron-ZeemanTerm gebildet. H NZ r r = − β N g N B0 Iˆ (16) 38 Grundlagen Der g-Faktor des Kernspins gN kann in erster Näherung immer als isotrope Konstante angesehen werden. Lediglich, wenn Effekte höherer Ordnung auftreten, muss ein g-Tensor für den Kernspin berücksichtigt werden. Kern-Quadrupol-Wechselwirkung Die Kern-Quadrupol-Wechselwirkung ist auf Kerne mit einem Spin I ≥ 1 beschränkt. Dabei handelt es sich um einen elektrischen Feldgradienten am Ort des Kerns, der mit dem elektrischen Quadrupolmoment des Kerns wechselwirkt. Der entsprechende Term im SpinHamilton-Operator lautet r r H Q = IˆQIˆ (17) Der Quadrupoltensor Q enthält das skalare Quadrupolmoment und die erste Ableitung des Feldgradienten des jeweiligen Kerns. Fasst man die Gleichungen 8, 9, 11, 16 und 17 zusammen, so erhält man Gleichung (18), die den kompletten Spin-Hamilton-Operator darstellt, der ein EPR-Spektrum beschreibt. r r r r r r r r r r H = H EZ + H ZFS + H HFC + H NZ + H Q = β e B0 gSˆ + SˆDSˆ + SˆAIˆ − β N g N B0 Iˆ + IˆQIˆ (18) Der Triplettzustand Bei der Betrachtung von Triplettzuständen muss zwischen organischen Molekülen mit einem Elektronenspin von S = 1 und Metallkomplexen (S = 1) unterschieden werden. In beiden Fällen tritt bereits ohne äußeres Magnetfeld eine Aufspaltung der drei Energieniveaus auf, die sogenannte Nullfeldaufspaltung (Abbildung 18). Bei Metallkomplexen ist die Spin-Bahn-Kopplung der bestimmende Term für die Nullfeldaufspaltung, während bei organischen Molekülen die dipolare Wechselwirkung (Gleichung 9) überwiegt. Die Spin-Bahn-Kopplung wird durch den Hamiltonoperator gemäß Gleichung (19) mit der Spin-Bahn-Kopplungskonstante λ beschrieben. H SP rr = λLˆ Sˆ (19) 39 Grundlagen Abbildung 18: Darstellung der Nullfeldaufspaltung für Nullfeldparameter D>E>0. Die Nullfeldsublevel Tx, Ty und Tz sind im lichtangeregten Triplettzustand nicht nach der Boltzmann-Gleichung besetzt. Bei Porphyrinen sind zwei unterschiedliche Wege, der „inter-system-crossing“Mechanismus (ISC) und der Radikalpaar-Mechanismus (RP), zu unterscheiden, über die der angeregte Triplettzustand gebildet werden kann. Der RP-Mechanismus erfordert nach der Bildung des ersten angeregten Singulettzustands (S1) eine temporäre Ladungstrennung[53] in je ein Kation- (D·+) und ein Anionradikal (A·-) und wird daher bevorzugt in den nativen Reaktionszentren beobachtet. Aufgrund der Trennung der beiden Elektronenspins auf zwei unterschiedlichen Molekülen mit unterschiedlichen magnetischen Umgebungen wird ein Triplettradikalpaar gebildet (1[D·+ A·-] → 3[D·+ A·-]), welches durch Rekombination zu einem Triplettzustand, der auf einem Molekül lokalisiert ist, führen kann. Weil die Lebensdauer des Radikalpaares im Verhältnis zur Elektronenspindynamik kurz ist, wird bei der Rekombination ausschließlich das Sublevel ms = 0 besetzt[54]. Der Triplettzustand ist spinpolarisiert und befindet sich nicht im thermischen Gleichgewicht nach der Boltzmann-Verteilung. Im Gegensatz zum RP-Mechanismus findet der Übergang in den Triplettzustand nach dem ISC-Mechanismus auf einem einzigen Molekül statt[55]. Nach der Anregung in den Singulettzustand S1 findet aufgrund der Spin-Bahn-Kopplung ein Übergang in den Triplettzustand statt. Weil die Spin-Bahn-Kopplung von der Richtung des resultierenden Elektronenspins der beiden ungepaarten Elektronen abhängig ist, werden die 40 Grundlagen Triplettsublevel auch in diesem Fall nicht nach der Boltzmann-Verteilung besetzt. Dieser Mechanismus tritt bei den in dieser Arbeit untersuchten Modellverbindungen auf. Aufgrund der Spinpolarisation treten im Spektrum absorptive und emissive Linien auf. Weil jeweils ein Übergang zwischen den Energieniveaus |0> und |1> bzw. |0> und |-1> auftritt, werden jeweils zwei Linien pro kanonische Orientierung im Spektrum beobachtet (vgl. Abbildung 19). Anhand der absorptiven und emissiven Linien kann auf die Besetzungsunterschiede der einzelnen Sublevel |-1>, |0> und |+1> geschlossen werden, indirekt können damit auch die Besetzungsunterschiede der Nullfeldsublevel berechnet werden. Aus dem EPR-Spektrum eines Triplettzustandes können direkt die Nullfeldparameter, die die Geometrie der Triplettwellenfunktion beschreiben, abgelesen werden (Abbildung 20). Jedoch kann durch dieses Verfahren nur der Betrag der Nullfeldparameter ohne ihr absolutes Vorzeichen bestimmt werden. Abbildung 19: Darstellung der Energieniveaus eines Triplettzustandes (D>0, E>0). Die Sublevel Tx, Ty und Tz sind durch die Nullfeldaufspaltung nicht mehr energetisch entartet. Ihr Besetzungsgrad richtet sich aufgrund der Spin-Bahn Kopplung nicht nach der Boltzmannverteilung. Wird ein externes Magnetfeld angelegt, werden die Sublevel |+1> und |-1> durch Mischung zweier Nullfeldsublevel gebildet und besetzt. So kommt es abhängig von der Orientierung des Moleküls im Magnetfeld zu jeweils einer absorptiven (a) und einer emissiven (e) Linie im EPR-Spektrum. Die detektierten Linien sind durch die farbigen Pfeile angedeutet. Ein Spektrum, das aus einer solchen Besetzung der Nullfeldsublevel resultiert, hat die Polarisation aaa eee, wie es für das ZnPPIX der Fall ist. 41 Grundlagen Abbildung 20: Beispiel für das Spektrum eines lichtangeregten Triplettzustandes. Aus dem Linienabstand können direkt die Nullfeldparameter abgelesen werden. Die aus einem EPR-Spektrum erhaltenen Werte für D und E in Gauss können durch Umformen von Gleichung (7) in Wellenzahlen ausgedrückt werden. Aus dem abgebildeten Spektrum (Abbildung 20) wird für 2D ein Wert von 825 G erhalten, was 82,5 mT entspricht. Durch Einsetzen in Gleichung (7) ergibt sich die folgende Gleichung (20) wobei ein g-Wert von ge = 2,0023 für die Berechnung verwendet wird. ν= 2,0023 ⋅ 82,5mT ⋅ 9,27408 ⋅ 10 −24 JT −1 = 1156 MHz 2 ⋅ 6,62618 ⋅ 10 −34 Js (20) Aus dieser Gleichung wird ein Wert für D in der Einheit Hertz erhalten, der nach Gleichung (21) in Wellenzahlen umgerechnet werden kann, wobei c der -1 Lichtgeschwindigkeit in cm s entspricht. 1 ν 1156 ⋅ 10 6 s −1 ~ ν = = = = 385 ⋅ 10 − 4 cm −1 10 −1 λ c 3 ⋅ 10 s cm (21) 42 Grundlagen 1.8.2. Grundlagen der ENDOR-Spektroskopie Die ENDOR-Spektroskopie ist eine Doppelresonanzmethode, in der die Übergänge der Kernspins über die EPR-Spektroskopie detektiert werden. Das Prinzip der ENDORSpektroskopie soll hier am einfachsten System mit einem Elektronenspin S = 1/2 und einem Kernspin I = 1/2 kurz erläutert werden. Wenn sowohl die Elektron-ZeemanWechselwirkung als auch die Hyperfeinwechselwirkung als isotrop angenommen werden, so vereinfacht sich der Spin-Hamilton-Operator dieses Systems aus Gleichung (18) folgendermaßen. rr r r r r ˆ ˆ ˆ H = β e B0 gS + aiso S I − β N g N B0 Iˆ (22) In der Hochfeldnäherung1 ergibt sich für die Energie-Niveaus folgende Gleichung. E (ms , mI ) = ν e ms −ν N mI + aiso ms mI h (23) mit νe = g e β e B0 und h νN = g N β N B0 h (24) r Hierbei steht B0 für den Betrag des Vektors ( B0 = B0 ). Für die erlaubten EPR- und NMRÜbergänge ergeben sich im einfachsten Fall jeweils zwei Linien. ν EPR = 1 ν e ± aiso 2 und ν NMR = ν N ± aiso 2 (25) In der Hochfeldnäherung überwiegt der Term der Elektron-Zeeman-Aufspaltung deutlich gegenüber den anderen Termen des Spin-Hamilton-Operators (HEZ>>HZFS, HHFC). 43 Grundlagen Um die Hyperfeinkopplungskonstanten zu messen, sind sowohl cw- als auch PulsENDOR-Verfahren entwickelt worden. In dieser Arbeit wurde eine Puls-ENDOR-Sequenz nach Davies[56] eingesetzt. Dieses Verfahren beruht auf dem Transfer und der Detektion von Spinpolarisation. Die Spinpolarisation resultiert aus den Besetzungsunterschieden zwischen verschiedenen Energieniveaus von EPR- und NMR-Übergängen. EZ EZ+NZ EZ+NZ+ HFC Abbildung 21: Schema der Energieniveaus für ein System mit S=1/2 und I=1/2. Gezeigt sind ElektronZeeman-Aufspaltung (EZ), die Kern-Zeeman-Aufspaltung (NZ) und die Hyperfeinwechselwirkung (HFC). Die erlaubten EPR-Übergänge (schwarz) und die erlaubten NMR-Übergänge (rot) sind durch die Pfeile eingezeichnet. Die Davies-Puls-ENDOR-Sequenz ist in drei Phasen aufgeteilt (vgl. Abbildung 22). Die Detektion der Spinpolarisation erfolgt in diesem Experiment mit einer Hahn-EchoSequenz. Durch einen 180°-Puls in der Präparationsphase wird das Besetzungsverhältnis eines EPR-Überganges invertiert. Ohne zusätzlichen Puls wird auf Grund der Inversion der Spinpolarisation ein emissives Spin-Echo-Signal detektiert. Wird jedoch zwischen Präparations- und Detektionsphase ein Radiofrequenzpuls eingestrahlt, erfolgt ein Übergang innerhalb der Kernspinniveaus. Dadurch kommt es zu einem Ausgleich der Polarisation der EPR-Übergänge. In der Detektionsphase wird nun kein Echo beobachtet. 44 Grundlagen In einem Puls-ENDOR-Experiment wird die Amplitude des invertierten Echo-Signals in Abhängigkeit von der Frequenz des Radiofrequenzpulses aufgezeichnet. In der hier vorliegenden Arbeit wurde die Puls-ENDOR-Spektroskopie eingesetzt, um die Hyperfeinwechselwirkung des Elektronenspins in Modellen für [4Fe4S]-Zentren und des lichtangeregten Triplettzustandes verschiedener Zink-Porphyrine zu detektieren. Abbildung 22: Darstellung der Davies-Puls-ENDOR Sequenz. In der Präparation wird das Besetzungsverhältnis eines EPR-Übergangs durch einen 180° Puls invertiert (schwarze Rechtecke => hohe Besetzung, weiße Rechtecke => geringe Besetzung). Wird bei der Einstrahlung eines 180° RF-Pulses keiner der beiden NMR-Übergänge invertiert (νRF ≠ νNMR), wird in der Detektionsphase mit der Hahn-Echo Sequenz das invertierte EPR-Signal beobachtet. Wird jedoch einer der beiden NMR-Übergänge durch den RF-Puls invertiert (νRF = νNMR), wird der Besetzungsunterschied des EPR-Übergangs ausgeglichen und das Spin-Echo verschwindet. 45 Grundlagen Für einen Triplettzustand (S = 1) liegen in der ENDOR-Spektroskopie einige Besonderheiten vor, auf die im folgenden kurz eingegangen werden soll. Das ENDORSpektrum eines Triplettzustandes zeigt keine symmetrische Verteilung der einzelnen Signale um die Larmor-Frequenz νH der ungekoppelten Protonen, sondern die Signale mit gleichem Vorzeichen der Kopplungskonstante liegen auf einer Seite des Spektrums, während die Signale mit dem entgegengesetzten Vorzeichen auf der anderen Seite zu liegen kommen. mS +1 Az>0 -1/2 νH - Az ZI 0 -1 ZII mS mI +1 +1/2 -1/2 νH +1/2 -1/2 νH + Az B0 || Z D>0 0 -1 +1/2 Az<0 ZI νH mI -1/2 νH - Az +1/2 -1/2 ZII +1/2 -1/2 νH + Az +1/2 Abbildung 23: Diagramm der Hyperfeinaufspaltung der drei Triplettsublevel in Abhängigkeit vom Vorzeichen der Hyperfeinkopplungskonstante. Die ENDOR-Messung wird bei einem festen Wert des Magnetfeldes durchgeführt und die RF-Frequenz variiert. Für die erlaubten NMR-Übergänge gilt die Auswahlregel ∆mI = ±1 und für die erlaubten EPR-Übergänge ∆mS = ±1. Findet kein EPR-Übergang statt, wird ein Signal bei der Larmor-Frequenz der Protonen entsprechend dem angelegten äußeren Magnetfeld detektiert (blauer Pfeil in Abbildung 23,Abbildung 24). Wird das ENDORSpektrum in der ZI-Position aufgenommen (schwarzer Pfeil in Abbildung 23, vgl. Abbildung 20), werden nur EPR-Übergänge zwischen den Triplettleveln |0> und |+1> detektiert. Im Spektrum treten die Kopplungen jeweils bei νRF = νH-AZ (26) 46 Grundlagen auf, wobei die Kopplungskonstante AZ ein negatives oder ein positives Vorzeichen besitzen kann. Besitzt die Kopplungskonstante ein negatives Vorzeichen, werden die Kopplungen bei einer RF-Frequenz beobachtet, die größer ist als die Larmor-Frequenz der ungekoppelten Protonen, während bei einem positiven Vorzeichen der Kopplungskonstante die Signale bei kleineren Frequenzen als νH beobachtet werden (Abbildung 24). Wird das ENDOR-Spektrum in der ZII-Position aufgenommen, werden die EPR-Übergänge zwischen den Triplettleveln |0> und |-1> detektiert. In diesem Fall treten die Signale der mit dem Elektronenspin wechselwirkenden Protonen bei Frequenzen von νRF = νH+AZ (27) auf. Entsprechend Gleichung 27 werden Kopplungen mit positivem Vorzeichen bei RF-Frequenzen beobachtet, die größer sind als νH und Kopplungen mit negativem Vorzeichen bei Frequenzen kleiner als νH (vgl. Abbildung 24). Abbildung 24: ENDOR-Spektren von ZnPPIX Mb in der ZI- und der ZII-Position. Aufgrund des unterschiedlichen Magnetfeldes bei den beiden unterschiedlichen Positionen liegt die Larmor-Frequenz der ungekoppelten Protonen νH (blauer Pfeil) bei unterschiedlichen Frequenzen. Die Kopplungskonstanten der gekoppelten Protonen behalten in beiden Spektren ihre Position relativ zu νH bei. 47 Grundlagen Beide Spektren enthalten in erster Näherung die gleiche Information und sind lediglich um die Larmor-Frequenz invertiert. Für die Kopplung der Stickstoffatome gilt die gleiche Betrachtungsweise, allerdings liegt die Larmor-Frequenz von 14 N-Atomen bei den verwendeten Magnetfeldern bei etwa 1,5 MHz, so dass die Inversion der Signale um die Larmor-Frequenz der ungekoppelten Stickstoffatome in den beiden Spektren nicht so offensichtlich ist, wie bei den Signalen der Protonen. 1.9. Mössbauer-Spektroskopie Die Mössbauer-Spektroskopie ist eine Kernabsorptions−/Kernemissions−Spektroskopie mit γ−Quanten[57; 58] . Die geringe relative Linienbreite von γ−Quanten ermöglicht den Nachweis sehr kleiner Effekte in der chemischen Umgebung (Elektronenhülle) des absorbierenden Kerns. Über die Dopplerverschiebung von relativ zueinander bewegten γ−Emitter und Absorber wird die Isomerieverschiebung (δ) relativ zu einem Standard gemessen. Die Isomerieverschiebungn entspricht der Verschiebung des Signalschwerpunktes zum Nullpunkt des Spektrums, der wiederum durch das Signal eines Standards definiert wird (Abbildung 25). Die Ursache für die Isomerieverschiebung ist die elektrische Wechselwirkung zwischen dem Atomkern und dem s-Elektron am Kernort, so dass aus der Isomerieverschiebung vor allem Rückschlüsse auf den Oxidationszustand gezogen werden können[59]. Durch die Wechselwirkung zwischen dem elektrischen Quadrupolmoment des Kerns und einem inhomogenen elektrischen Feld kommt es zur Quadrupolaufspaltung (∆EQ) der Resonanzlinien, woraus Daten über die Symmetrie und die Koordination des Atoms gewonnen werden können. Die Quadrupolaufspaltung entspricht dem Abstand der Linien im Spektrum. Die magnetische Dipolwechselwirkung zwischen dem magnetischen Dipolmoment des Kerns und einem magnetischen Feld am Kernort führt zur Hyperfeinaufspaltung des Mössbauer-Spektrums und liefert Informationen über den magnetischen Zustand (z.B. Ferromagnetismus). Um die Verbreitung der Spektrallinien durch den Rückstoß der Atomkerne bei Emission und Absorption zu vermeiden, müssen Emitter− und Absorberatome in einen Festkörper eingebaut werden. Ein Nachteil dieser Methode besteht darin, dass sie nur für Elemente mit geeigneten Isotopen zugänglich ist. 48 Grundlagen Am weitesten verbreitet ist die 57 Fe−Mössbauer-Spektroskopie, die auch in dieser Arbeit verwendet wurde. Abbildung 25: Beispiel eines Mössbauer-Spektrums mit der Isomerenverschiebung (δ) und der Quadrupolaufspaltung (∆EQ). 49 Aufgabenstellung 2. Aufgabenstellung 2.1. Metall-Phäophorbide in Myoglobin Chlorophylle und ihre Derivate sind die wesentlichen funktionellen Bestandteile der Photosynthese. Ihre physikalischen Eigenschaften werden in den nativen Systemen durch die Proteinumgebung stark beeinflusst, wodurch gleiche Moleküle verschiedene Aufgaben erfüllen können. Chlorophyll a fungiert in den Lichtsammelkomplexen einzig als Lichtabsorber, während es in den Reaktionszentren der Photosysteme I und II die Rolle des Elektronendonors und –akzeptors nach der Lichtanregung übernimmt. Allerdings sind die natürlichen Systeme aufgrund ihrer Komplexität nur schwer vollständig zu charakterisieren. Werden Chlorophylle aus einem Protein herausgelöst, neigen sie zur Aggregation. Durch die enge Wechselwirkung innerhalb der Aggregate („π-stacking“) werden die Eigenschaften, insbesondere nach Lichtanregung, drastisch verändert. Der lichtangeregte Zustand eines monomeren Chlorophylls ist derzeit nur unzureichend charakterisiert, obwohl dieser essentiell für das Verständnis des Zusammenspiels mehrerer dieser Moleküle ist. Daher ist es von besonderem Interesse, ein monomeres Chlorophyllmolekül, das keine Wechselwirkungen zu anderen photoaktiven Verbindungen aufweist, möglichst vollständig zu charakterisieren. Der lichtangeregte Triplettzustand ist aufgrund seiner Reaktivität und seiner langen Lebensdauer schädlich für photosynthetisch aktive Organismen und muss daher effizient durch Carotinoide gelöscht werden (Triplett quenching). In vitro bietet er die Möglichkeit, das Molekülorbital, in dem sich das angeregte Elektron befindet (LUMO), mit Hilfe der EPR- und ENDOR-Spektroskopie zu untersuchen. Dieses Molekülorbital wird auch im lichtangeregten Singulettzustand, welcher die entscheidende Rolle in den Energiesammel- und Ladungstrennungsprozessen der Photosynthese spielt, durch ein ungepaartes Elektron besetzt. Aufgrund der kurzen Lebensdauer des Singulettzustandes und seines inhärenten Diamagnetismus ist er jedoch für die Spektroskopie nur schwer zugänglich. 50 Aufgabenstellung Ein Ziel dieser Arbeit ist, die in der Literatur beschriebenen Rekonstitutionsverfahren für Porphyrine in Apo-Myoglobin zu etablieren und die auf diese Weise dargestellten Komplexe umfassend zu charakterisieren. Insbesondere sollen die Unterschiede des lichtangeregten Triplettzustandes des Kofaktors in der Proteinmatrix und in einem Lösemittel unter Verwendung der EPR- und ENDOR-Spektroskopie untersucht werden. Zu diesem Zweck soll zunächst kommerziell erhältliches Myoglobin aus Pferdemuskel mit dem diamagnetischen Zink-Komplex des nativen Kofaktors rekonstituiert und charakterisiert werden. Darauf aufbauend werden Zink-Komplexe von Chlorophyll aDerivaten, die eine möglichst geringe Änderung zu den Ausgangsverbindungen durchlaufen haben, untersucht. Der Phytylester des Chlorophyll a muss für den Einbau entfernt werden. Änderungen im Substitutionsmuster ermöglichen den Zugang zu verschiedenen Chlorophyll-Derivaten, z. B. Chlorophyll d. Sobald das Rekonstitutionsverfahren etabliert ist, sollen die gleichen Komplexe mit Myoglobin vom Pottwal (SwMb) dargestellt werden. Da die Bindungstasche hochkonserviert ist, wird eine ähnliche Wechselwirkung zwischen Kofaktor und Protein, wie im Fall des Myoglobins aus Pferdemuskel, erwartet. Der Vorteil des PottwalMyoglobins ist die Möglichkeit, Einkristalle von ausreichender Größe für eine EPR- und ENDOR-spektroskopische Analyse züchten zu können. 2.2. Modelle für [4Fe-4S]-Zentren des Photosystems I Eisen-Schwefel-Zentren sind in der Natur weit verbreitet und dienen als redox-aktive Kofaktoren des Elektronentransfers. Zwischen den verschiedenen Arten der FeS-Zentren variiert nicht nur die Zahl an Eisen- und Sulfidionen des Systems, sondern auch das Redoxpotential. Dies ist die wesentliche physikalische Eigenschaft eines redox-aktiven Kofaktors und die FeS-Zentren umfassen einen Potentialbereich von +400 mV bis –700 mV. Bisher sind nur wenige Peptid-basierte Modelle für FeS-Zentren entwickelt worden, um den Einfluss der Proteinumgebung auf die physikalischen Eigenschaften, wie das Redoxpotential und die magnetischen Eigenschaften, zu untersuchen. Die meisten der 51 Aufgabenstellung bereits existierenden Modelle waren auf die Modellierung von bakteriellen [4Fe4S]Ferredoxinen ausgerichtet, die einen Potentialbereich von –100 mV bis –500 mV abdecken. Um die Mechanismen der Kontrolle des Redoxpotentials zu verstehen, werden zusätzlich Modelle benötigt, die möglichst dicht an die Extremwerte bezüglich des Redoxpotentials gelangen. Die FeS-Zentren des Photosystems I bieten einen guten Ansatzpunkt für derartige Modelle, da ihre Redoxaktivität einen Potentialbereich von –500 mV bis –700 mV umfasst. Zusätzlich ist das native System durch Kristallstrukturanalyse charakterisiert und von der Untereinheit PsaC existiert eine NMR-Struktur in Lösung. Außer dem Einfluss der Proteinumgebung auf das Redoxpotential ist die Kontrolle über die magnetischen Eigenschaften von FeS-Zentren, insbesondere der [4Fe4S]-Zentren, noch nicht vollständig verstanden. Abhängig vom Redoxzustand können in verschiedenen [4Fe4S]-Zentren jeweils zwei lokalisierte Paare aus je zwei Eisenkernen gleicher Oxidationsstufe auftreten. Inwieweit die Position der Paare im kubischen [4Fe4S]-Zentrum durch das Protein kontrolliert wird, und ob auf diese Weise Einfluss auf andere physikalische Eigenschaften ausgeübt wird, ist weitgehend unbekannt. Wahrscheinlich ist die Geometrie des kubischen FeS-Zentrums der entscheidende Faktor für die Position der Paare mit unterschiedlicher Oxidationsstufe. Modelle, die einfach darzustellen und zu variieren sind, können detaillierte Einsicht in diese Problematik geben. Ein weiteres strukturelles Merkmal von kubischen FeS-Zentren ist die Ausbildung von Wasserstoffbrücken zwischen den Sulfidionen des Zentrums und Amidprotonen der Polypeptidkette. Über die Anzahl der Wasserstoffbrückenbindungen werden die zugänglichen Oxidationsstufen (3+/2+ oder 2+/1+) des FeS-Zentrums festgelegt. Die Entwicklung von Peptid-basierten Modellsystemen, die in der Lage sind, kubische [4Fe4S]-Zentren zu binden, ist daher eine wesentliche Grundlage für die Aufklärung dieser Fragestellungen. Aus diesem Grund sollen in einem zweiten Teil dieser Arbeit synthetische Modelle für [4Fe4S]-Zentren dargestellt werden, um den Einfluss der Proteinumgebung auf die Eigenschaften dieser wichtigen Kofaktoren des Elektronentransfers zu untersuchen. Als Vorbilder sollen die Eisen-Schwefel-Zentren des Photosystems I, FA, FB und FX, dienen, weil sie stark negative Redoxpotentiale aufweisen und bereits gut charakterisiert sind. Nach der Herstellung und vollständigen Charakterisierung wird untersucht, inwieweit die synthetischen Modelle in der Lage sind, die Funktion ihrer nativen Vorbilder im Elektronentransfer zu übernehmen. 52 Ergebnisse und Diskussion 3. Ergebnisse und Diskussion 3.1. Metall-Phäophorbide in Myoglobin-Matrix 3.1.1. Natives Myoglobin und Apo-Myoglobin Myoglobin aus Pferdemuskeln ist kommerziell (Sigma) in der biologisch inaktiven metForm (Fe3+) des Hämins erhältlich. Um nicht-native Kofaktoren in das Protein Myoglobin einbauen zu können, muss zunächst der native Kofaktor, das Hämin, entfernt werden. Zu diesem Zweck wurde das Protein zunächst denaturiert, indem der pH-Wert der Proteinlösung durch Zugabe von Salzsäure auf einen Wert kleiner als zwei gesenkt wurde. Abbildung 26: UV-Vis-Spektren von met-Myoglobin (schwarz), denaturiertem met-Myoglobin bei pH = 1,5 (rot) und Apo-Mb (grün). Während das Spektrum von met-Myoglobin von der Soret-Bande dominiert wird, liegt der Kofaktor Hämin bei einem pH-Wert von 1,5 im ungebundenen Zustand vor. Nach der Extraktion kann kein Kofaktor mehr nachgewiesen werden. Die Konzentrationen der Proben sind unterschiedlich. 53 Ergebnisse und Diskussion Liegt das Protein schließlich in denaturierter Form vor, ist der native Kofaktor nicht mehr stabil gebunden und kann durch ein organisches Lösemittel, in diesem Fall 2-Butanon, extrahiert werden[60]. Die saure Lösung des Apo-Proteins (Protein ohne Kofaktor) wurde durch Dialyse gegen Natriumhydrogencarbonat (50 mg/L) neutralisiert, wodurch eine Rückfaltung des Proteins erreicht wurde. Anschließend wurde gegen 0,1 mM EDTALösung dialysiert, um gelöste Metallionen zu komplexieren, die andernfalls im ApoProtein gebunden werden könnten. Abschließend wurde gegen destilliertes Wasser dialysiert, um die niedermolekularen Bestandteile der Lösung zu entfernen. Nach vollständiger Extraktion des Kofaktors dominierte die Soret-Bande bei 408 nm das Spektrum nicht mehr, allerdings blieb die Absorptionsbande der aromatischen Aminosäuren bei 280 nm erhalten. Für die weiteren Untersuchungen wurden ausschließlich Proben verwendet, bei denen der Restgehalt an Hämin unter 1 % lag. Die Entfernung des Kofaktors wirkt sich deutlich auf die Stabilität des Proteins aus. Liegt die Stabilität des nativen Myoglobins bei einem Wert von 30 kJ mol-1, so geht der Wert nach Entfernung des Kofaktors auf 10 kJ mol-1 zurück. Die Kooperativität sinkt bedingt durch die Extraktion des Kofaktors ebenfalls von 20 kJ mol-1 M-1 auf 10 kJ mol-1 M-1. Abbildung 27: Stabilitätsmessung von Apo-Myoglobin (●) und met-Myoglobin (▲). Die Entfaltung von Apo-Mb wurde durch Fluoreszenz-, die Entfaltung des met-Myoglobins durch CD-Spektroskopie verfolgt. Der Übergang von vollständig gefaltetem zum entfalteten Zustand ist bei met-Myoglobin steiler und findet bei einer höheren Konzentration von Guanidiniumhydrochlorid statt. 54 Ergebnisse und Diskussion Für alle dargestellten Komplexe wurde die Entfaltung durch CD-, UV-Vis-, und Fluoreszenz-Spektroskopie verfolgt. Die UV-Vis-Spektroskopie lieferte in allen Fällen die schlechtesten Ergebnisse, da sie im Vergleich zu den beiden anderen Methoden deutlich zu große Werte lieferte. Diese Tatsache kann darauf zurückgeführt werden, dass diese Methode direkt die spektralen Eigenschaften des Kofaktors wiedergibt, die wesentlich durch die Ligandierung bestimmt werden und erst in zweiter Linie durch die Proteinumgebung. Die CD- und die Fluoreszenz-Spektroskopie sind dagegen Methoden, die im untersuchten Messbereich ausschließlich die Struktur der Polypeptidkette wiedergeben können. Bei der CD-Spektroskopie wurde das Signal der α-helikalen Sekundärstrukturelemente bei 222 nm untersucht, während die Fluoreszenz-Spektroskopie verwendet wurde, um die Verschiebung des Fluoreszenzmaximums der Aminosäure Tryptophan abhängig vom Entfaltungszustand zu untersuchen. Die Daten dieser Methoden waren im Rahmen des experimentellen Fehlers identisch. Lediglich das paramagnetische met-Myoglobin zeigte eine zu schwache Fluoreszenz, weshalb keine Auswertung der Daten möglich war. Die Schlussfolgerung aus dieser Beobachtung ist, dass der Kofaktor zumindest teilweise noch durch das Protein koordiniert wird, wenn die Tertiär- und Sekundärstrukturelemente bereits entfaltet sind. Das auf die oben beschriebene Weise dargestellte Apo-Myoglobin befindet sich in einem nicht vollständig gefalteten Zustand und weist daher eine geringere Stabilität auf. Durch Zugabe von Hämin zu einer Apo-Myoglobin-Lösung kann das Myoglobin derart rekonstituiert werden, dass es von kommerziell erhältlichem Protein nicht zu unterscheiden ist. Sowohl das UV-Vis-Spektrum als auch die Stabilität des so hergestellten Myoglobins sind identisch mit met-Myoglobin. Deshalb kann davon ausgegangen werden, dass die Aminosäurekette während der Extraktion des Kofaktors nicht beschädigt wurde. Außerdem ist das auf diese Weise hergestellte Apo-Myoglobin in der Lage, einen Kofaktor zu binden. 55 Ergebnisse und Diskussion 3.1.2. Darstellung der Kofaktoren In dieser Arbeit wurden die Kofaktoren Zink-Protoporphyrin, Zink-Phäophorbid a, ZinkPyrophäophorbid a, Zink-Pyrophäophorbid a Methylester, Zink-Pyrophäophorbid d Methylester und 3-Devinyl-3-hydroxymethyl-132-demethoxycarbonylphäophorbid a Methylester verwendet. Das Zink-Protoporphyrin wird durch Umsetzen der kommerziell erhältlichen freien Base Protoporphyrin mit Zinkacetat in Essigsäure erhalten. Die restlichen Kofaktoren stellen Derivate des Chlorophyll a dar und können über kurze Synthesewege ausgehend von Chlorophyll a beziehungsweise von einem ungereinigten Algenrohextrakt dargestellt werden. In allen Fällen wurde der Phytylester verseift, da diese C20-Einheit die Bindung des Kofaktors in der Proteinbindungstasche erschweren würde. An Stelle des Phytylesters lag entweder die freie Carbonsäure oder der korrespondierende Methyl-Ester vor. Als zentrales Metallion wurde grundsätzlich Zink verwendet, da das in Chlorophyll a natürlich vorkommende Magnesiumion während der Synthese verloren geht. Das Zink(II)Ion hat vergleichbare Eigenschaften wie das Magnesium(II)-Ion in Bezug auf den Ionenradius. Er liegt bei 72 pm für Mg2+ und 74 pm für Zn2+ bei Koordinationszahl sechs. Damit sind beide Ionen etwas zu groß für die Bindungsstelle im Porphyrin, die einen diagonalen Kern-Kern Abstand der Stickstoffatome von 200 pm aufweist (Stickstoff VDW-Radius 74 pm). Als Konsequenz daraus zeigen die Porphyrinkomplexe beider Metallionen eine starke Präferenz für eine verzerrte quadratisch-pyramidale Struktur, in der die vier äquatorialen Ligandenstellen durch den Ring besetzt sind und das Metallion außerhalb der Ringebene liegt[61]. Gleichzeitig weisen die Zinkkomplexe eine höhere Stabilität aufgrund der größeren Elektronegativität des Zinkions auf. Die elektronischen Strukturen der Zink- und Magnesiumkomplexe haben sich als ähnlich erwiesen[62], wodurch der Einsatz von Zink als zentrales Metallion für die durchgeführten EPR- und ENDOR-Messungen unproblematisch ist. Da die Zinkkomplexe leichter zu synthetisieren sind, werden sie in dieser Arbeit verwendet[63]. 56 Ergebnisse und Diskussion 3 2 H 2C 3 1 4 2 CH3 5 NH 1 H H CH 3 H 3C 3 6 7 N 8 20 1 8 3 H 3C 19 N HN 11 18 14 12 H 3C 17 16 13 15 1 2 2 1 17 17 13 13 H 2C CH2 H Collidin CH3 O O OPhytyl O H H 5 4 2 NH 1 180°C 3 1 CH CH3 H 10 3 2 CH2 8 9 2 H 2C CH3 6 7 N 20 8 10 19 N HN 11 18 14 12 H 3C 16 17 13 15 1 2 2 1 17 17 13 13 H 2C CH2 H O 1 8 3 CH3 CH2 1 CH 2 3 H 3C MeOH H 2SO4 H 4 2 H H CH3 5 6 7 NH 1 N 20 8 8 9 2 CH3 O H O 8 CH3 H 10 19 N HN 11 18 14 12 17 16 13 15 2 171 2 1 17 13 13 H 2C CH2 H OPhytyl 1 CH2 H 3C CH3 O H O 8 9 2 H 2C OMe CH3 Phäophytin a im Algenrohextrakt Pyrophäophytin a 3 3 3 H 3C 2 1 H 3 1 4 NH CH3 5 6 7 N 8 8 9 20 10 19 N HN 11 18 14 12 17 16 13 15 2 171 2 1 17 13 13 H 2C CH2 H H 3C 1 8 OH O 3 CH3 1 3 H 3C CH2 2 1 H H O H CH 2 5 4 NH CH3 6 7 N 20 8 8 9 10 19 N HN 11 18 14 12 17 16 13 15 2 171 2 1 17 13 13 H 2C CH2 H H 3C CH3 O O 2 H 2C H CH OsO 4 NaIO4 TFA TFA 2 H 2C Methylpyrophäophorbid a H O O O 1 8 2 CH3 CH2 H 3C 4 2 H H CH3 5 6 7 NH 1 N 20 8 8 9 1 8 2 CH3 CH2 H 10 19 N HN 11 18 14 12 17 16 13 15 2 171 2 1 17 13 13 H 2C CH2 H H 3C CH3 O H O OH 1 3 H CH3 3 CH OMe CH3 Phäophorbid a Pyrophäophorbid Methylpyrophäophorbid d HO 3 1 CH2 3 H 3C CH3 HO CH3 CH3 CH3 CH3 Phytol 4 2 1 H tertButylamin boran H 5 NH CH3 6 7 N 20 8 8 9 10 19 N HN 11 18 14 12 16 H 3C 17 13 15 1 2 2 1 17 17 13 13 H 2C CH2 H H O 1 8 2 CH3 CH2 H CH3 O OMe Methyl 3-devinyl-3-hydroxymethyl132-demethoxycarbonylphäophorbid a Abbildung 28: Darstellung des Syntheseweges der in dieser Arbeit untersuchten Kofaktoren und vollständige Struktur des Phytols, das in Chlorophyllen in Form eines Esters an der Propionsäure gebunden ist. 57 Ergebnisse und Diskussion Setzt man Chlorophyll a mit Trifluoressigsäure (TFA) um, so erhält man aus dem Phytylester nahezu quantitativ die freie Carbonsäure[64] (Abbildung 28). Der Methylester in Position 132 wird jedoch nicht hydrolysiert. Diese Tatsache ist auf die Molekülstruktur zurückzuführen. Die Trifluoressigsäure greift die Carbonylgruppe der Esterfunktion an und induziert dadurch eine Vinyl-Umlagerung, die letztendlich zur Esterspaltung führt. Ein vergleichbarer Angriff auf den Methylester ist nicht möglich. Bei der kurz gewählten Reaktionszeit von etwa 10 Minuten wird die Hydrolyse des Methylesters fast völlig unterdrückt. Da durch die aciden Bedingungen der Komplex das Magnesiumion verliert, erhält man Phäophorbid a. Die Synthese des Pyrophäophorbids geht vom Algenrohextrakt aus. Zunächst wird der Methylester in der 132-Position durch Kochen am Rückfluss in Collidin bei 180 °C unter Argon pyrolysiert[65]. Durch Spaltung des Methylesters wird eine β-Ketocarbonsäure erzeugt, die daraufhin decarboxyliert wird. Nach der säulenchromatographischen Reinigung der Reaktionsmischung erhält man Pyrophäophytin. Der Phytylester wird wie bei Chlorophyll a durch Reaktion mit Trifluoressigsäure in die freie Carbonsäure umgewandelt. Der Vorteil dieses Kofaktors liegt darin, dass das Stereozentrum in der 132-Position zerstört wird und dadurch das Epimeren-Gleichgewicht a/a´ aufgehoben wird. Der Pyrophäophorbid a Methylester wird analog dargestellt, jedoch wird zur Spaltung des Phytylesters nicht Trifluoressigsäure verwendet, sondern es wird mit Schwefelsäure/Methanol umgeestert. Der Methylester kann durch Kristallisation aus CH2Cl2/Hexan gereinigt werden. Der Methylester des Pyrophäophorbids d wird aus Pyrophäophorbid a Methylester dargestellt. Dazu wird die Vinylgruppe in der 31-Position durch Osmiumtetraoxid/Natriumperjodat zum entsprechenden Aldehyd gespalten[66]. Alle auf diesem Weg dargestellten Kofaktoren liegen zunächst als freie Base vor, können aber mit einem Überschuss an Zinkacetat in Eisessig metalliert werden. Der Aldehyd Pyrophäophorbid d kann unter Verwendung von tert-Butylaminboran selektiv zum Alkohol reduziert werden. Hierbei wird die Ketogruppe in Position 131 aufgrund ihrer geringeren Reaktivität nur zu einem kleinen Teil reduziert. Die Metallierung in Essigsäure ist nicht möglich, sondern erfolgt in Dichlormethan mit 3-4 % Methanol. Alle analytischen Daten stimmen mit den Literaturdaten überein. Die analytischen Daten sind im experimentellen Teil bei der jeweiligen Verbindung aufgelistet. 58 Ergebnisse und Diskussion 3.1.3. Komplexbildung und Charakterisierung der Kofaktoren mit Apo-Myoglobin Nachdem nachgewiesen wurde (vgl. Abschnitt 3.1.1), dass das Apo-Myoglobin intakt und in der Lage ist, Kofaktoren verschiedener Art einzubauen, wurden die dargestellten Kofaktoren einer Einbauuntersuchung unterzogen. Der ideale Kofaktor sollte a) einen 1:1-Komplex mit dem Protein bilden können, b) eine eindeutige Einbaurichtung in der Bindungstasche des Proteins aufweisen und c) diamagnetisch sein. Die letzte Bedingung wird von allen dargestellten Kofaktoren erfüllt. Insgesamt wurde in dieser Arbeit der Einbau von sechs verschiedenen Kofaktoren untersucht. Als Beispiel für den nativen Kofaktor wurde das diamagnetische Derivat des Hämins, Zinkprotoporphyrin, verwendet. Außerdem wurde der Einbau von fünf Chlorophyll-Derivaten in das Apo-Myoglobin untersucht. In den spektroskopischen Untersuchungen wird der Kofaktor-Myoglobinkomplex mit dem Kofaktor ohne Proteinkomplexierung verglichen. Letzterer wird als freier bzw. unkomplexierter Kofaktor bezeichnet, wobei die Wortwahl „frei“ nicht suggerieren soll, dass die vakanten Koordinationsstellen am Zink nicht besetzt sind. Der freie Kofaktor ist entweder symmetrisch und/oder schwach durch das Lösemittel komplexiert. Eine Ausnahme stellt das Zink-Pyrophäophorbid a dar, welches in Pyridin als Lösemittel fünffach koordiniert vorliegt. Im Protein werden alle Kofaktoren lediglich durch ein Histidin koordiniert. Das Zink ist daher fünffach koordiniert (ggf. sechsfach durch ein Wassermolekül) und der Kofaktor unterliegt einer Verzerrung, weil das zentrale Metallatom aus der Ringebene herausgezogen wird. Aufgrund dieser Wechselwirkung wird die ursprünglich planare Struktur des Porphyrins zu einer gewölbten Anordnung, ähnlich dem Ausschnitt aus einer Kreisoberfläche, verformt. 59 Ergebnisse und Diskussion 3.1.3.1. Zink-Protoporphyrin Apo-Myoglobin (ZnPPIX Mb) 3.1.3.1.1. UV-Vis-Spektroskopie und Stabilitätsmessung Zink-Protoporphyrin ist gut löslich in DMSO und besitzt in diesem Lösemittel ein Absorptionsspektrum, das durch vier Absorptionsbanden bei 586 nm (α-Bande), 547 nm (β-Bande), 417 nm (Soret-Bande) und 347 nm (Hyper-Bande) gekennzeichnet ist (Abbildung 29). Wird dieser Kofaktor in das Apo-Myoglobin eingebaut, so verschieben diese Absorptionsbanden nach 595 nm, 555 nm, 428 nm und 350 nm. Die Analyse der UV-Vis-Daten des Zink-Protoporphyrin (ZnPPIX) erfolgte anhand von Literaturdaten[67]. Weiterhin ist zu beobachten, dass die Soret-Bande deutlich schmaler wird und an Intensität gewinnt. Der Extinktionskoeffizient für Myoglobin, isoliert aus Pferdemuskel, bei 280 nm beträgt 15,2 mM-1 cm-1, während der Extinktionskoeffizient der Soret-Bande des gebundenen Zink-Protoporphyrins mit einem Wert von 152,6 mM-1 cm-1 angegeben wird[67]. Daraus ergibt sich ein Bandenverhältnis von 10 für einen 1:1 Komplex. In dieser Arbeit wurden für die weiteren Messungen nur solche Proben verwendet, die ein Bandenverhältnis von ≥ 9 aufwiesen. Abbildung 29: UV-Vis-Spektren von ZnPPIX in DMSO (schwarz) und vom ZnPPIX Mb Komplex (rot) in unterschiedlichen Konzentrationen. Durch den Einbau in das Protein werden alle Banden ins Langwellige (bathochrom) verschoben. 60 Ergebnisse und Diskussion Abbildung 30: Stabilitätsmessung von ZnPPIX Mb detektiert durch CD-Spektroskopie (■) im Vergleich zu Apo-Mb (■). Die Interpolation der Messpunkte liefert für die freie Entfaltungsenthalpie einen Wert von 31,2 kJ mol-1 und für die Kooperativität einen Wert von 16,1 kJ mol-1 M-1. Apo-Myoglobin weist im Vergleich zu met-Myoglobin durch den fehlenden Kofaktor eine deutlich geringere Stabilität auf (vgl. Abschnitt 3.1.1). Der Einbau eines Kofaktors sollte zu einer teilweisen Wiederherstellung der ursprünglichen Stabilität führen. In welchem Ausmaß dies geschieht, hängt von der Struktur des Kofaktors ab. Für ZnPPIX ist die freie Base des Kofaktors Protoporphyrin (PPIX) und damit identisch mit der freien Base des nativen Kofaktors. Lediglich das zentrale Eisen-Metallion wurde durch ein Zink(II)ion ersetzt. Daher wird eine vollständige bzw. nahezu vollständige Wiederherstellung der ursprünglichen Stabilität erwartet. Diese Erwartung konnte experimentell nachgewiesen werden (Abbildung 30). Für die Stabilität wurde ein Wert von 31,2 ± 6,6 kJ mol-1 ermittelt, der damit im Rahmen des Fehlers mit der Stabilität von met-Myoglobin übereinstimmt (30,6 ± 4,5 kJ mol-1). Für die Kooperativität wurde ein etwas kleinerer Wert von 16,1 ± 3,3 kJ mol-1 M-1 gemessen (met-Myoglobin 19,8 ± 2,9 kJ mol-1 M-1, siehe Abschnitt 3.1.1). Diese Abweichung der Kooperativität könnte auf die unterschiedlichen Bindungseigenschaften von Hämin und ZnPPIX im Bezug auf die hier vorliegende Histidinkomplexierung zurückgeführt werden. Die Histidin-Eisen Bindung ist deutlich stärker als die Histidin-Zink Bindung. Im Rahmen der Fehler wurden für die Entfaltungsenthalpie identische Werte erhalten. 61 Ergebnisse und Diskussion 3.1.3.1.2. NMR-Spektroskopie Nachdem gezeigt wurde, dass ein stabiler und stöchiometrischer Komplex von ZnPPIX und Apo-Myoglobin gebildet wurde, muss nun festgestellt werden, ob eine einheitliche Einbau-Orientierung des Kofaktors im Protein vorliegt (vgl. Abschnitt 1.3). Aufgrund der strukturellen Ähnlichkeit mit nativem Hämin wird auch ein ähnliches Verhalten für ZnPPIX erwartet, d. h., zwei Einbauisomere, die im Gleichgewicht miteinander stehen. Die Protonen der γ-CH3-Gruppen der Aminosäure Valin 68 liegen in der π-Elektronenwolke des Kofaktors und werden daher durch den Ringstromeffekt in der NMR-Spektroskopie bei negativen ppm-Werten detektiert[68]. Daher sind sie leicht von den anderen Protonen innerhalb des Proteins zu unterscheiden und können einfach zugeordnet werden. Innerhalb von 24 Stunden nach erfolgter Rekonstitution werden zwei Einbauisomere im Verhältnis 1:1 gefunden, die miteinander im Gleichgewicht stehen. Die Signale der γ-CH3-Gruppen der Aminosäure Valin 68 liegen bei einer Messtemperatur von 8°C bei –3,9 ppm (Peak I) und –1,1 bzw. –1,0 ppm (Peak II) (Abbildung 31). Diese Werte unterscheiden sich signifikant von den publizierten Werten von –3,6 ppm (Peak I) und –0,9 ppm (Peak II)[69]. Diese Abweichung kommt durch die unterschiedliche Messtemperatur zustande, die in diesem Experiment bei 8°C und für die Vergleichswerte aus der Literatur bei 25°C lag. Wird die gleiche Probe nach 13 Monaten Lagerzeit bei 4°C erneut bei einer Messtemperatur von 25°C gemessen, so werden Werte von –3,5 ppm (Peak I) und –0,9 ppm bzw. –0,7 ppm (Peak II) erhalten (Abbildung 31), wobei zu berücksichtigen ist, dass sich die Signale der beiden Einbauisomere bei –3,5 ppm überlagern. Das Peakverhältnis liegt im Gleichgewichtszustand etwa bei 1:3. Ähnlich wie beim nativen Kofaktor Hämin werden zwei unterschiedliche Einbauisomere beobachtet, die miteinander im Gleichgewicht stehen. Während jedoch das Verhältnis der Isomere im Gleichgewicht für das Hämin bei 9:1 liegt[28], wird für das ZnPPIX lediglich ein Verhältnis von 3:1 gefunden. Die Triebkraft der Gleichgewichtseinstellung ist die hydrophobe Wechselwirkung der Vinyl-Substituenten des Kofaktors mit den unpolaren Aminosäuren der Bindungstasche. Sowohl bei Hämin, als auch bei ZnPPIX sollte diese Wechselwirkung identisch sein. Demzufolge sollte im Gleichgewichtszustand auch ein ähnliches Verhältnis der beiden Isomere vorliegen. Nach 13 Monaten wird aber lediglich ein Verhältnis der beiden Isomere von 1:3 im NMR-Spektrum nachgewiesen. Daher kann 62 Ergebnisse und Diskussion man davon ausgehen, dass der Gleichgewichtszustand auch nach 13 Monaten noch nicht ppm (t1)-0.50 -2.50 -3.00 -3.50 -1.50 -2.00 -2.50 -3.00 -3.50 -3.94 -3.90 -2.00 -4.00 -4.50 -4.00 -4.50 -3.51 -1.50 -0.88 -1.00 -0.71 ppm (t1)-0.50 -1.12 -0.95 erreicht wurde. -1.00 Abbildung 31: Ausschnitt aus dem 1H-NMR-Spektrum (400 MHz) vom ZnPPIX Apo-Mb Komplex bei 8°C direkt nach der Rekonstitution (oben) und bei 25°C nach 13 Monaten Lagerzeit (unten). Unmittelbar nach der Rekonstitution werden zwei Einbauisomere im Verhältnis 1:1 gefunden, die miteinander im Gleichgewicht stehen. Nach 13 Monaten Lagerzeit liegen die Isomere im Verhältnis 1:3 vor. Die unterschiedlichen Signalpositionen liegen in der unterschiedlichen Messtemperatur begründet (siehe Text). Tabelle 2: Zusammenfassung der NMR-Daten des Komplexes ZnPPIX Mb. ZnPPIX Mb ZnPPIX Mb Literaturangabe [69] Messtemperatur Peak I Peak II 8°C -3,90, -3,94 ppm -1,12 ppm / -0,95 ppm 25°C -3,51 ppm -0,88 ppm / -0,71 ppm 25°C -3,6 ppm -0,9 ppm 63 Ergebnisse und Diskussion Aus den Signallagen bei unterschiedlichen Messtemperaturen kann die temperaturabhängige Verschiebung der Signale berechnet werden. Diese Abhängigkeit gilt nur in einem relativ kleinen Temperaturbereich, aber es bietet sich die Möglichkeit, die Signalpositionen bei verschiedenen Messtemperaturen miteinander zu vergleichen. Die Änderung der Signallage liegt bei 0,02 ppm/°C für Peak I bzw. bei 0,012 ppm/°C für Peak II. 3.1.3.1.3. EPR-Spektroskopie Die transiente EPR-Spektroskopie am lichtangeregten Triplettzustand von ZnPPIX in ApoMyoglobin wurde bereits von Hoffman et al.[70] durchgeführt und bietet daher die Möglichkeit, die hier erhaltenen Ergebnisse zu überprüfen. Aus den transienten Messungen können die Nullfeldparameter des Triplettzustandes unmittelbar abgelesen werden. Die Nullfeldparameter D und E sind ein Maß für die Geometrie der Wellenfunktion des Triplettzustandes. Ist D = E = 0 liegt eine kubische Symmetrie vor, bei D ≠ 0, aber E = 0 liegt axiale Symmetrie vor und bei D ≠ 0 und E ≠ 0 liegt eine Symmetrie vor, die niedriger als axial ist. Der Vergleich der Messergebnisse des ungebundenen Kofaktors mit dem Proteinkomplex gibt also unmittelbar Aufschluss darüber, in wie weit die elektronische Struktur des Triplettzustands durch die Proteinumgebung beeinflusst wird. Für beide Nullfeldparameter ist bekannt, dass sie im Fall des Protoporphyrins positive Vorzeichen besitzen, so dass die Werte im Folgenden mit absoluten Vorzeichen angegeben werden können. Die transienten Spektren von ZnPPIX Mb und dem freien Kofaktor (Abbildung 32) zeigen mit zunehmendem Magnetfeld drei absorptive Linien, gefolgt von drei emissiven Linien. Im Spektrum des freien Kofaktors tritt zusätzlich bereits zu Beginn der Messungen das Signal eines Radikals auf. Die Simulation der Spektren liefert Werte für die Nullfeldparameter D und E von 357 ± 4·10-4 cm–1 bzw. 67 ± 4·10-4 cm–1 für den Proteinkomplex und 350 ± 4·10-4 cm–1 bzw. 63 ± 4·10-4 cm–1 für den freien Kofaktor (Tabelle 3). Im Rahmen des Fehlers stimmen die ermittelten Nullfeldparameter mit den Literaturwerten überein. Der Fehler der experimentellen Werte ist mit ± 4·10-4 cm–1 groß. Dieser Wert ist 64 Ergebnisse und Diskussion auf das derzeitige Entwicklungsstadium des Simulationsprogramms zurückzuführen. Gegenwärtig muss jeder Parameter vollständig manuell an das experimentelle Spektrum angepasst werden. Eine halbautomatische Anpassung der Simulation an das experimentelle Spektrum befindet sich in der Entwicklung und wird die Präzision der Simulationen erhöhen. A B * Abbildung 32: Transiente EPR Spektren (X-Band, 9,5 GHz) von ZnPPIX Mb (A) und ZnPPIX in DMSO (B) bei 50 K. Die aufgenommenen Spektren sind in schwarz und die entsprechende Simulation in rot dargestellt. Beide Spektren weisen drei absorptive Linien, gefolgt von drei emissiven Linien auf (aaa eee). Das EPR-Signal eines Radikals im Spektrum von ZnPPIX ist durch * gekennzeichnet. Tabelle 3: Zusammenfassung der experimentellen Nullfeldparameter D und E von ZnPPIX Mb und freiem ZnPPIX und Vergleich mit den Literaturwerten. Die Literaturwerte für den freien Kofaktor beziehen sich auf das Lösemittel Pyridin (*), während in dieser Arbeit ein Gemisch aus DMSO und Glycerin 40/60 verwendet wurde. ZnPPIX Mb Nullfeldparameter (50 mM Phosphat, pH 7,0, 60 % ZnPPIX (DMSO/Glycerin) Glycerin) D (experimentell) 357 ± 4·10-4 cm–1 350 ± 4·10-4 cm–1 E (experimentell) 67 ± 4·10-4 cm–1 63 ± 4·10-4 cm–1 D (Literatur)[70] 349·10-4 cm–1 350·10-4 cm–1 * E (Literatur)[70] 69·10-4 cm–1 66·10-4 cm–1 * 65 Ergebnisse und Diskussion Im Vergleich zum simulierten Spektrum wird die Asymmetrie des experimentellen Spektrums, das vom Proteinkomplex gemessen wurde, besonders deutlich. Die Intensität der emissiven Linien ist deutlich größer als die der absorptiven Linien. Dies kann nicht auf eine apparative Ursache zurückgeführt werden, sondern ist ein Merkmal des untersuchten Komplexes. Obwohl dieses Resultat schon bei anderen Verbindungen beobachtet wurde, ist bis heute unverstanden. Eine weitere in diesem System bisher nicht verstandene Beobachtung ist die g-Faktor Anisotropie, die in den meisten Fällen erst bei sehr hohen Magnetfeldern bzw. Mikrowellenfrequenzen (z.B. im W-Band bei 95 GHz) detektiert werden kann. Alle in dieser Arbeit untersuchten Systeme zeigten jedoch bereits im Q-Band bei 35 GHz eine deutliche g-Faktor-Anisotropie. Um sicherzustellen, dass nur der energetisch niedrigste Triplettzustand während der Messungen besetzt wird, wurde die Wellenlängenabhängigkeit des transienten EPRSpektrums des ZnPPIX Mb Komplexes untersucht. Obwohl man bei einer Anregungswellenlänge von 585 nm, also in der β-Bande des UV-Vis-Spektrums, davon ausgehen muss, dass nur der S1-Zustand und damit auch nur der T1-Zustand zugänglich ist, wurden auch Spektren bei kürzeren Wellenlängen aufgenommen. Sollte der zweite angeregte Triplettzustand ebenfalls besetzt sein, müsste dies einen Effekt auf die Nullfeldparameter des EPR-Spektrums haben, da der T2-Zustand eine andere elektronische Struktur hat und zusätzlich schneller relaxiert. Im Bereich der α- und der β-Bande wurden in Intervallen von 10 nm transiente EPR-Spektren aufgenommen. Da die Soret-Bande eine Anregung in den S2-Zustand ermöglicht, wurde zusätzlich ein weiteres Spektrum bei einer Anregungswellenlänge von 440 nm aufgenommen. Bei diesen Messungen konnte keine Wellenlängenabhängigkeit der Nullfeldparameter festgestellt werden. Diese Ergebnisse zeigen eindeutig, dass nur der erste angeregte Triplettzustand unter den verwendeten Messbedingungen besetzt wird. Da die durchgeführten ENDOR-Messungen (s. Abschnitt 3.1.3.1.4) nicht auf cw-EPR, sondern auf Puls-EPR beruhen, wurden ebenfalls Puls-EPR-Spektren des Komplexes in einem Puffer/Glycerin-Gemisch (50 mM Phosphatpuffer, pH 7,0), sowie des freien Kofaktors in einem DMSO/Glycerin-Gemisch aufgenommen. Anhand dieser Spektren wurden die Feldpositionen für die ENDOR-Messungen gewählt, wobei zwei Kriterien die Auswahl bestimmten. Das EPR-Signal an der zu untersuchenden Feldposition muss möglichst intensiv sein, um ein gutes Signal zu Rausch-Verhältnis zu erreichen und es 66 Ergebnisse und Diskussion muss nach Möglichkeit nur eine Orientierung des Moleküls zum Magnetfeld selektiert werden können. Beide Spektren lassen keine Selektion einer einzigen Molekülorientierung durch eine bestimmte Magnetfeldstärke zu, wie es für das Spektrum einer gefrorenen Lösung typisch ist. Daher wurden die ENDOR-Spektren an den Feldpositionen der Z-, Yund X-Orientierung aufgenommen. A B Abbildung 33: Puls EPR-Spektren des ZnPPIX Apo-Mb Komplexes im X-Band Spektrometer (A) und des freien Kofaktors in DMSO/Glycerin im Q-Band Spektrometer (B) bei 10 K. Im Spektrum des Komplexes sind die induzierten Übergänge zwischen den Subleveln gekennzeichnet (für D>0 und E>0). Im Spektrum des freien Kofaktors ist deutlich das scharfe Radikalsignal bei 12000 G zu erkennen. Aus diesen experimentellen Puls-EPR-Spektren wurden die gleichen Nullfeldparameter erhalten wie aus den transienten Spektren. Auch hier ist der deutliche Intensitätsunterschied zwischen emissiven und absorptiven Linien des Proteinkomplexes sichtbar. Ebenso wie bei den transienten Spektren ist der Intensitätsunterschied der Linien im Spektrum des freien Kofaktors nicht so groß wie im Spektrum des Proteinkomplexes. Das Puls-EPR-Spektrum von ZnPPIX Mb (X-Band) wird durch Modulationen der PyrrolStickstoffatome beeinflusst, wodurch die Intensität der einzelnen Übergänge stark von den Pulsabständen des 90°- und 180°-Pulses abhängt. Die Modulationen sind deshalb im XBand-Spektrum stark, weil die Kern-Zeeman-Aufspaltung und die Hyperfeinwechselwirkung für 14N-Kerne bei einer Magnetfeldstärke von 3500 G (0,35 T) ungefähr die gleiche Größe besitzen, wodurch die drei Hyperfeinsublevel geringe Energieunterschiede aufweisen. Auf diese Weise werden alle Kernübergänge teilweise 67 Ergebnisse und Diskussion erlaubt, weil alle teilweise mI = 0 Charakter enthalten. Während der Zeit zwischen den Mikrowellenpulsen evolviert das System in jedem Triplettlevel (l) mit der Phase der Kernspinns (e-ie(l)t2π/h), wodurch die Modulation hervorgerufen wird. Bei einer Magnetfeldstärke von etwa 1,2 T (Q-Band) ist die Kern-Zeeman-Aufspaltung der 14 N-Kerne etwa dreimal größer als bei 0,35 T, da sie proportional mit der Magnetfeldstärke zunimmt. Die Größe der Hyperfeinkopplungen dagegen ist unabhängig vom Magnetfeld und ändert seine Größe nicht. Daher treten in einem Puls-EPR-Spektrum im Q-Band keine Modulationen durch die Pyrrol-Stickstoffatome auf. 3.1.3.1.4. ENDOR-Spektroskopie von ZnPPIX in Glycerin/DMSO Die ENDOR-Spektroskopie dient dazu, die Hyperfeinkopplungskonstanten von Kernen mit einem Kernspin von I ≠ 0 zu bestimmen. Die Größe der Hyperfeinkopplungen ist ein Maß für die Elektronenspindichte am Kernort, mit dem er wechselwirkt. In den untersuchten Porphyrinsystemen werden die Kopplungen der Elektronenspins mit den Methinprotonen (α-Protonen) und den an das Ringsystem angrenzenden Methyl- und Methylengruppen (β-Protonen) beobachtet. Dabei gilt für die Kopplungen der α-Protonen die folgende Näherung, die von einem isolierten sp2-C-H Fragment abgeleitet werden kann. Liegt die z-Achse des Systems parallel zum pz-Orbital des Kohlenstoffs, die x-Achse entlang der C-H-Bindungsachse und die y-Achse senkrecht zu dieser und zur z-Achse, können die beobachteten Kopplungen durch die isotrope Hyperfeinkopplungskonstante Aiso folgendermaßen angenähert werden: AZ ≈ 1·Aiso, AY ≈ 1,5·Aiso, AX ≈ 0,5·Aiso[71; 72], wobei eine die isotrope Hyperfeinkopplungskonstante Aiso mit einem Wert von –60 MHz als eine Spindichte von 1 am Kern eines α-Protons interpretiert wird. Für β-Protonen, insbesondere die Methylgruppen, wird eine geringe Anisotropie von etwa 10 % von Aiso erwartet, da sie zwei Bindungen vom ungepaarten Elektronenspin entfernt sind. Die Analyse, welche Kernspins mit dem Elektronenspin des angeregten Triplettzustands in welchem Ausmaß wechselwirken, gibt wertvolle Informationen über die Verteilung des Tripletts auf dem Molekülgerüst. Damit steht mit der ENDOR-Spektroskopie eine Methode zur Verfügung, die direkt Aufschluss Triplettwellenfunktion gibt. 68 über die Ausdehnung der Ergebnisse und Diskussion Anhand des Puls-EPR-Spektrums des ZnPPIX Mb Komplexes (X-Band, Abbildung 33) kann eine gute Orientierungsselektion erreicht werden. Bei dem Magnetfeldwert der maximalen Mikrowellenabsorption einer Orientierung liegen nur geringe Absorptionsbeiträge der beiden anderen Orientierungen vor. Dadurch sind die Signale der jeweils nicht selektierten Orientierungen in den ENDOR-Spektren aufgrund der geringen Absorption wahrscheinlich nicht detektierbar. Für den freien Kofaktor ist dies nicht der Fall. Die EPR-Absorptionen überlagern sich stark, daher ist die Orientierungsselektion insbesondere bei der Feldposition des YII-Überganges unmöglich. Dies erschwert die Interpretation der erhaltenen ENDOR-Spektren deutlich, da nicht erkennbar ist, auf welchen EPR-Übergang die Hyperfeinkopplung zurückzuführen ist. Die aussagekräftigsten ENDOR-Spektren werden bei einer Orientierungsselektion entlang der Z-Achse erhalten, da in diesem Fall praktisch keine Beiträge der anderen EPRÜbergänge zu erwarten sind. Dennoch liefern die Spektren in der X- und Y-Position ebenfalls Hinweise, die zur Interpretation wichtig sind. Anhand dieser Spektren kann untersucht werden, ob ein Signal anisotrop ist oder nur eine geringe Richtungsabhängigkeit zeigt. Daraus ergibt sich eine zuverlässige Zuordnung der Signale. Zusätzlich wird die Zuordnung der Signale durch quantenchemische Rechnungen an unterschiedlichen Modellsystemen unterstützt. Die verwendete Methode zur Berechnung der Hyperfeinkopplungen (vgl. Abschnitt 5.1.12) lieferte bereits bei der Analyse der ENDOR-Spektren von Chlorophyllradikalen gute Ergebnisse und wird in dieser Arbeit erstmals mit den ENDOR-Spektren der Triplettzustände von Chlorophyllderivaten verglichen werden. Die berechneten Hyperfeinkopplungen des nicht-ligandierten Kofaktors wurden mit den Ergebnissen des freien Kofaktors verglichen. In der Rechnung für den Proteinkomplex wurde ein fünfter Ligand in Form von Imidazol zum Kofaktor hinzugefügt. Obwohl dieser Ligand keinen direkten Kontakt zum delokalisierten Triplett auf dem Molekülgerüst hat, weil das zentrale Metallatom keine nennenswerte Spindichte trägt, ändert er die Geometrie des Kofaktors. Die ursprünglich planare Struktur von ZnPPIX wird zu einer gewölbten Struktur verzerrt (Abbildung 34), ähnlich dem Ausschnitt aus einer Kugeloberfläche. Dadurch wird eine Änderung der Hyperfeinkopplungen beobachtet. 69 Ergebnisse und Diskussion Abbildung 34: Darstellung des ZnPPIX mit Imidazol als fünftem Liganden. Deutlich zu erkennen ist die Verzerrung der ursprünglich planaren Struktur des Kofaktors zu einer gewölbten Struktur. Die Vinylgruppe in Position 3 ist leicht aus der Ebene gedreht (ca. 20°), während die Vinylgruppe in Position 8 in der Ebene liegt. Die Rotationsbarrieren beider Gruppen sind sehr gering. Die ENDOR-Spektren des freien Kofaktors (Abbildung 35) weisen in der Z-Orientierung fünf eindeutig definierte Signale auf, von denen zwei zu kleinen Kopplungen gehören (Signal 3 und 4). Ferner werden eine große Kopplung mit negativem Vorzeichen (Signal 1) und zwei mittlere Kopplungen detektiert, von denen eine ein negatives Vorzeichen (Signal 2) und die andere positives Vorzeichen aufweist (Signal 5). In der Y- und der X-Orientierung tritt ein weiteres Signal auf, welches sich durch eine sehr große Kopplung mit positivem Vorzeichen auszeichnet. Anhand der quantenchemischen Rechnung (Tabelle 4) kann das breite Signal 1 um –9 MHz den vier Methinprotonen des Ringsystems zugeordnet werden. Alle vier Protonen weisen berechnete Kopplungen von –7,9 MHz bis –9,7 MHz auf. Die Methinprotonen erzeugen durch ihre Anisotropie häufig breite und schwache Signale, wodurch die Detektion erschwert wird. Beim Übergang zur Y-Orientierung tritt das Signal 1 bei –6,9 MHz auf. Diese starke Verschiebung bestätigt die Zuordnung der Signale zu den Methinprotonen. Es ist allerdings unwahrscheinlich, dass bei der Y-Orientierung das Signal noch immer von allen vier Protonen verursacht wird. Wahrscheinlich tragen hier nur die Signale der Protonen 5 und 15 zum Signal bei, weil bei diesen Protonen die 70 Ergebnisse und Diskussion Komponente AY der Hyperfeintensors entlang der Bindungsachse liegt und daher theoretisch den kleinsten Kopplungswert erreichen. Für die Protonen 10 und 20 liegt die Komponente AY innerhalb der Ringebene senkrecht zur Bindungsachse. Die Kopplung müsste daher ihren maximalen Wert annehmen. Ein solches Signal konnte nicht gemessen werden. Dies ist wahrscheinlich auf die oben erwähnte Signalverbreiterung zurückzuführen. In der X-Orientierung ist das Gegenteil der Fall. Hier liegt das äußere Magnetfeld entlang der Bindungsachsen der Protonen 10 und 20, während es senkrecht zur Bindungsachse der Protonen 5 und 15 steht. Bei X-Orientierung wird das Signal bei –6,9 MHz daher von den Protonen 10 und 20 hervorgerufen, während für die Protonen 5 und 15 keine Kopplung nachgewiesen werden kann. Abbildung 35: ENDOR-Spektren (Q-Band) des Kofaktors ZnPPIX in Glycerin/DMSO bei 10 K. Es wurden Spektren bei drei unterschiedlichen Feldpositionen aufgenommen (ZII – grün, YII – rot, XII – schwarz). Die Verschiebung der einzelnen Signale ist durch gestrichelte Linien angedeutet. Das Signal 2 bei –2,9 MHz wurde der Methylengruppe in Position 82 zugeordnet, da nur für diese Gruppe eine ausreichend negative Kopplungskonstante berechnet wurde. Dieses Signal wurde nur in der Z-Orientierung detektiert. Obwohl die Gruppe 82-CH2 weit vom Ringsystem entfernt ist, ist aufgrund der Konjugation eine Wechselwirkung mit dem Elektronenspin des Triplettzustandes naheliegend. Die isotrope Hyperfeinkopplungs71 Ergebnisse und Diskussion konstante ist mit –5,3 MHz berechnet worden und weicht stark vom experimentellen Ergebnis ab. Allerdings ist hierbei zu berücksichtigen, dass die Energiebarriere für die Rotation dieser Gruppe aus der Ebene heraus klein ist. Schon bei kleinen Winkeln können dadurch relativ große Abweichungen in der berechneten Kopplungskonstante entstehen. Tabelle 4: ENDOR-Kopplungen des Kofaktors ZnPPIX in Glycerin/DMSO. Alle Angaben in MHz mit Vorzeichen relativ zu D (D>0). Signalnr. Position im Molekül 1 5-H, 10-H, 15-H, Exp. Exp. Exp. Kopplung Kopplung Kopplung Berechnete Kopplung (Aiso) in Z in Y in X -9 (br) -6,9 -6,9 -9,4; -9,7; -7,9; -8,9 20-H 2 82-CH2 -2,9 n.d. n.d. -5,3 3 2-CH3, 12-CH3, -0,5 -0,4 n.d. -1,2; -0,4; -0,3 32-CH2 4 171-CH2, 31-CH +0,6 +0,6 n.d. +0,4; +0,3 5 7-CH3, 18-CH3, +2,9 +2,4 +2,5 +2,2; +4,7; +1,7 n.d. +9,5 +14,4 - 81-CH 6 - Das Signal 3 bei –0,5 MHz wird den Methylgruppen in Position 2 und 12 zugeordnet. In der Y-Orientierung liegt das Signal bei –0,4 MHz. Es ist daher eindeutig isotrop und muss durch β-Protonen verursacht werden. In der X-Orientierung ist eine eindeutige Zuordnung schwierig, weil die EPR-Signalintensität deutlich geringer ist als bei den anderen Orientierungen. Daraus resultieren erheblich längere Messzeiten zur Aufnahme der ENDOR-Spektren, um ein ausreichendes Signal zu Rausch Verhältnis zu erzielen. Der für die Kopplung der 2-CH3 Gruppe berechnete Wert (-1,2 MHz) ist größer als der experimentell ermittelte Wert (-0,5 MHz). Dabei darf nicht außer acht gelassen werden, dass die verwendete Rechenmethode bisher in der Vorhersage der Hyperfeinkopplungen von Chlorophyllradikalen sehr gute Übereinstimmungen mit den experimentellen Daten gezeigt hat[73], jedoch wurde das Potential der Rechnung bisher nicht zur Interpretation von 72 Ergebnisse und Diskussion Protoporphyrin verwendet[74]. Möglicherweise ist die Präzision der Rechnung im Fall des Protoporphyrins etwas geringer. Die 32-CH2 Gruppe weist eine berechnete Kopplung von –0,4 MHz auf und wird zunächst ebenfalls diesem Signal zugeordnet. Um präzisere Aussagen über die Zuordnung aller drei Gruppen zu diesem Signal machen zu können, sind zusätzliche Messungen nötig, die idealerweise an einem Einkristall durchgeführt werden müssten. Die Zuordnung von Kopplungen mit geringer Größe ist bei Messungen in gefrorenen Lösungen generell schwierig. Dem Signal 4 bei +0,6 MHz wurden die 171-CH2 Gruppe und die 31-CH Gruppe zugeordnet. Für diese beiden Gruppen wurden Kopplungen von +0,4 MHz und +0,3 MHz berechnet. In der Y-Orientierung liegt dieses Signal ebenfalls bei +0,6 MHz und es kann, ähnlich wie das Signal 3, in der X-Orientierung nicht beobachtet werden. Die Methylengruppe in Position 171 ist zwar nicht mehr mit dem π-System konjugiert, aber durch Hyperkonjugation kann ein geringer Anteil an Spindichte auf diese Position übertragen werden, woraus eine kleine Kopplung resultiert, die zudem stark vom Winkel der Methylengruppe zur Ringebene abhängt. Für die konjugierte 31-Vinylposition ist der gleiche Effekt wie für die 82-Position zu berücksichtigen. Zwar kann nicht die gesamte CH-Einheit aus der Ebene gedreht werden, aber die Ausrichtung der CH-Bindungsachse relativ zum Molekül ändert sich abhängig vom Drehwinkel der Vinylgruppe. Die Richtung dieser Achse ist entscheidend dafür, welcher Wert des Hyperfeintensors für die Kopplung maßgeblich ist. Ebenso wie bei Signal 3 wurden die Gruppen, die diesem Signal zugeordnet wurden, ausschließlich nach den berechneten Kopplungen ausgewählt und die Zuordnung ist in beiden Fällen als vorläufig anzusehen. Das Signal 5 liegt in der Z-Orientierung bei +2,9 MHz und verändert seine Position über +2,4 MHz in der Y-Orientierung zu +2,5 MHz in der X-Orientierung. Eine eindeutige Zuordnung dieses Signals ist schwierig, jedoch kommen die Gruppen 7-CH3, 18-CH3 und 81-CH als Möglichkeiten in Frage. Aufgrund der relativ geringen Anisotropie ist die Zuordnung zu β-Protonen naheliegend. Für die Gruppe 7-CH3 wurde eine Kopplung von +2,2 MHz und für die Gruppe 18-CH3 eine Kopplung von +4,7 MHz berechnet. Die Gruppe 81-CH weist eine berechnete Kopplung von +1,7 MHz auf. Die berechneten Kopplungen der Methylgruppe in Position 7 zeigt die geringste Abweichung mit den experimentellen Werten. Dennoch werden die anderen Gruppen zunächst ebenfalls diesem 73 Ergebnisse und Diskussion Signal zugeordnet, weil das Signal breit und unstrukturiert ist und daher eine Überlagerung verschiedener Signale nicht ausgeschlossen werden kann. Das Signal 6, das in X- und Y-Orientierung auftritt, handelt es sich um ein intensives und stark anisotropes Signal mit einer sehr großen Kopplungskonstante (14,4 MHz), dem keine Gruppe des Ringsystems zugeordnet werden konnte. Diese Kopplung muss also von einer Gruppe verursacht werden, die nicht zum Ringsystem gehört und doch einen starken Kontakt zum angeregten Triplettzustand besitzt. Möglicherweise findet hier eine Wechselwirkung des Elektronenspins mit einem an das zentrale Metall koordinierten Lösemittelmolekül statt. Ist dies der Fall, sollte das Signal bei Verwendung von deuteriertem Lösemittel nicht mehr auftreten, wodurch diese Hypothese bestätigt werden könnte. Bis auf die Gruppen 131-CH2, 132-CH2 und 172-CH2 konnten alle erwarteten Signale unterschiedlicher Genauigkeit zugeordnet werden. Die verbliebenen Gruppen besitzen berechnete Kopplungskonstanten, die im Bereich von 0 bis 0,1 MHz liegen. Diese Kopplungen sind zu klein, sodass sie nicht in einem experimentellen Spektrum zu beobachten sind. Die Zuordnung der Methinprotonen ist eindeutig, da ihre Signale im erwarteten Bereich auftreten. Außerdem kann die Zuordnung der Methylgruppen in Position 12 und 7 als gesichert angesehen werden. Die berechneten Kopplungen der anderen zugeordneten Gruppen zeigen eine unterschiedlich große Abweichung von den experimentell nachgewiesenen Signalen und ihre Zuordnung ist daher als vorläufig anzusehen bis diese Ergebnisse durch weiterführende Messungen bestätigt oder widerlegt werden können. 74 Ergebnisse und Diskussion 3.1.3.1.5. ENDOR-Spektroskopie von ZnPPIX Mb in Puffer/DMSO Der ZnPPIX Apo-Mb Komplex wurde in einem 40/60-Gemisch von 50 mM Phosphatpuffer, pH 7,0, und Glycerin auf gleiche Weise ENDOR-spektroskopisch untersucht. Jedoch kam hier kein Q-Band Spektrometer, sondern ein X-Band Spektrometer zum Einsatz. Dies hat zwar auf die Position der Kopplungssignale keinen Einfluss, allerdings ist durch das geringere Magnetfeld die Separation der einzelnen Kerne mit unterschiedlichen gyromagnetischen Momenten geringer. Deshalb können in den ENDORSpektren des Komplexes (X-Band) auch 15 N-Kopplungen beobachtet werden, in den Spektren des freien Kofaktors (Q-Band) jedoch nicht. Abbildung 36: ENDOR-Spektren (X-Band) von ZnPPIX Apo-Mb bei 10 K. Die Spektren wurden bei drei unterschiedlichen Feldern aufgenommen (ZI – grün, YI – rot, XI – schwarz). Hyperfeinkopplungen mit negativem Vorzeichen erscheinen rechts von der Larmor-Frequenz der ungekoppelten Protonen. Die Verschiebung der einzelnen Signale ist durch gestrichelte Linien angedeutet. Die mit * gekennzeichneten Signale stammen von Stickstoffkopplungen des Rings und von Histidin. Das Spektrum des Proteinkomplexes (Abbildung 36, Tabelle 5, Tabelle 6) weist insgesamt mehr Signale auf, die über einen größeren Bereich verteilt sind. Im ENDOR-Spektrum der Z-Orientierung treten neben der Stickstoffkopplung sechs weitere Signale auf. Die 75 Ergebnisse und Diskussion Kopplung der Ringstickstoffatome und des Histidinstickstoffs, der an das Zink koordiniert ist, treten gemeinsam mit einer Kopplungskonstante von etwa 0,4 MHz auf. Sie werden sowohl bei Z-Orientierung als auch bei Y-Orientierung detektiert, fehlen aber im Spektrum der X-Orientierung. Möglicherweise sind die Signale zu schwach oder das Signal zu Rausch Verhältnis nicht ausreichend, um die Wechselwirkung der 15 N-Kerne in der X-Orientierung zu beobachten. Dominierend im Spektrum der Z-Orientierung ist das Signal der Kopplung der Methinprotonen (Signal 1), das im Proteinkomplex aus zwei, teilweise separierten, Banden bei –10,6 und –9,2 MHz besteht. Die Kopplung ist damit um etwa 1 MHz größer als für den freien Kofaktor. In der Y-Orientierung liegt das Signal der Methinprotonen 5 und 15 bei –9,4 MHz, während die Signale der Protonen 10 und 20 nicht detektiert werden. Die Kopplungen der Protonen 10 und 20 liegen in der X-Orientierung bei –8,4 MHz. Insgesamt hat die Spindichte in diesen Positionen des Moleküls durch die Proteinkomplexierung also geringfügig zugenommen. Die berechneten Werte für die Kopplungen der Methinprotonen am ZnPPIX mit Imidazol als fünftem Liganden sind mit –11,3 MHz bis –12,6 MHz zu groß, während die Ergebnisse für den freien Kofaktor geringfügig zu klein sind. Offensichtlich wird in der Rechnung die Geometrie stärker verzerrt als durch das Protein bzw. der daraus resultierende Effekt wird in der Rechnung überbewertet. Insgesamt zeigen die berechneten Werte der Methinprotonen allerdings eine überraschend hohe Übereinstimmung mit den experimentellen Kopplungskonstanten. Die Kopplung der 82-CH2 Gruppe (Signal 2) liegt im Proteinkomplex nahezu unverändert bei –2,6 MHz (-2,9 MHz ohne Protein), das Signal ist aber vergleichsweise schärfer und in allen drei Spektren sichtbar. In der Y-Orientierung liegt das Signal bei –4,7 MHz und in der Z-Orientierung wieder bei –2,7 MHz. Die Zuordnung dieses Signals zu der 82-CH2 Gruppe erfolgte, weil keine andere Gruppe des Moleküls eine berechnete Kopplung mit einem derart negativen Wert aufweist. In Übereinstimmung mit der Messung am unkomplexierten ZnPPIX und den berechneten Werten (vgl. Abschnitt 5.1.12) wird das Signal 3 bei –0,7 MHz den Methylgruppen in Position 2 und 12 zugeordnet. Auch dieses Signal ist in den Spektren des Proteinkomplexes besser aufgelöst und intensiver. Es liegt in der Y-Orientierung bei –2,0 MHz und in der X-Orientierung bei –1,7 MHz. Die Verlagerung des Signals abhängig von der Richtung des äußeren Magnetfeldes ist qualitativ gut von den Rechnungen 76 Ergebnisse und Diskussion vorhergesagt worden, wie in Tabelle 6 zu sehen ist. Die experimentelle Kopplung in Z-Orientierung ist auf Grund der Komplexierung des Kofaktors durch das Protein praktisch unverändert. Allerdings ist das Signal im Komplex stärker anisotrop. Dies könnte auf die räumlich stark geordnete Struktur des Proteins zurückzuführen sein. Während der unkomplexierte Kofaktor in jeder Richtung von Lösemittelmolekülen umgeben ist, die alle identisch sind und innerhalb der Hydrathülle auch ähnlich ausgerichtet sind, befindet sich der komplexierte Kofaktor im Gegensatz dazu in einer Umgebung, die per se stark anisotrop ist. Tabelle 5: ENDOR-Kopplungen des Komplexes ZnPPIX Mb in Puffer/DMSO. Alle Angaben in MHz mit Vorzeichen relativ zu D. a) Die Kopplungen des Histidinstickstoffs beziehen sich auf die Larmor-Frequenz von 15N. Nummer Position im Molekül Exp. Exp. Exp. Kopplung Kopplung Kopplung Berechnete Kopplung (Aiso) in Z in Y in X 5-H, 10-H, 15-H, -10,6; -9,4 -8,4 20-H -9,2 2 82-CH2 -2,6 -4,7 -2,7 -2,9 3 2-CH3; 12-CH3 -0,7 -2,0 -1,7 -1,0; -0,7 4 32-CH2 +0,5 n.d. n.d. +0,4 5 18-CH3 +2,1 +2,5 +3,2 +2,9 6 7-CH3 +4,3 +4,8 +5,9 +3,3 7 - n.d. +9,0 +9,7 - * 15 |0,4| a |0,4| a n.d. - 1 N -11,8; -12,6; -11,3; -11,5 Dem Signal 4 bei +0,5 MHz wurde die 32-CH2 Gruppe zugeordnet, weil aus der Rechnung hervorgeht, dass die Kopplung dieser Gruppe durch den fünften Liganden das Vorzeichen wechselt. Während im freien Kofaktor diese Gruppe ein Signal bei –0,5 MHz (Signal 3) erzeugte, wird sie in der Rechnung des asymmetrischen Komplexes mit positiver Kopplung aufgeführt. Da jedoch in der Y- und X-Orientierung dieser Gruppe keine 77 Ergebnisse und Diskussion weiteren Kopplungen zugeordnet werden konnten, ist diese Interpretation als unsicher anzusehen. Während die Kopplung der Methylgruppen in Position 7 und 18 für den freien Kofaktor in einem überlagerten Signal auftraten, liegen sie im Komplex getrennt voneinander vor (Signale 5 und 6). Die berechneten Kopplungen liegen sehr eng zusammen (+2,9 MHz bzw. +3,3 MHz) und beide Signale weisen eine relativ geringe Verschiebung bei Variation der Molekülorientierung im Magnetfeld auf. Die Entscheidung Signal 5 zu Position 18 und Signal 6 zu Position 7 zuzuordnen, wurde letztendlich aufgrund der Tatsache gefällt, dass die berechneten Werte für den vollständigen Hyperfeintensor der Gruppe 7-CH3 Werte größer 4 MHz aufweist, während für den vollständigen Hyperfeintensor der Gruppe 18-CH3 ausschließlich Werte kleiner als 4 MHz berechnet wurden. Auch in den Spektren des Proteinkomplexes wurde je ein Signal in der Y- und der X-Orientierung detektiert, das nicht mit dem Kofaktor in Verbindung gebracht werden konnte (Signal 7). Die Kopplung ist mit +9,7 bzw. +9,0 MHz allerdings bedeutend kleiner als in den Spektren des freien Kofaktors (+14,4 MHz). Es könnte sich daher um die γ-CH3 Gruppen der Aminosäure Valin 68 handeln, die in die π-Elektronenwolke des Kofaktors eintauchen (s.a. 3.1.3.1.2 und 3.1.3.2.2). Die Spektren des Proteinkomplexes ließen keine Signalzuordnung der Gruppen 171, 172, 31, 81, 131 und 132 zu. Die Positionen 171, 172, 131 und 132 sind nicht konjugiert und sollten daher nur in sehr geringem Maß Spindichte des Triplettzustandes tragen. Die CH-Gruppen 31 und 81 zeigen wahrscheinlich nur schwache ENDOR Kopplungen, da sie durch das Protein in einer aus der Ringebene herausgedrehten Stellung fixiert sind und dadurch die Konjugation teilweise aufgehoben wird. Die Kopplungen der Methinprotonen und der Methylgruppe in Position 7 sind im Proteinkomplex um etwa 1 MHz größer als für den freien Kofaktor. Dagegen haben die experimentell ermittelten Kopplungen der Gruppen 82-CH2 und 18-CH3 geringfügig abgenommen und die Kopplungen der Methylgruppen 2 und 12 sind praktisch unverändert. Diese Beobachtung kann dahingehend interpretiert werden, dass der Triplettzustand im Proteinkomplex stärker auf die einzelnen Gruppen verteilt ist, also stärker delokalisiert ist, als im Fall des freien Kofaktors. Insbesondere ist eine Zunahme der Kopplungskonstante der Methinprotonen und der Methylgruppe in Position 7 zu beobachten, während die Kopplungskonstanten der anderen Gruppen, die anhand der experimentellen Spektren 78 Ergebnisse und Diskussion identifiziert werden konnten, sich nur geringfügig änderten. Diese Beobachtung wurde durch die quantenchemischen Rechnungen vorhergesagt und ist im wesentlichen auf die geometrische Verzerrung des Kofaktors durch die asymmetrische Ligandierung zurückzuführen. Tabelle 6: Auflistung aller berechneten Hyperfeinkopplungen für den Triplettzustand des ZnPPIX-Imidazol Modellsystems inklusive ihrer vollständigen Hyperfeintensoren. a Komponente des Hyperfeintensors entlang der C-H Bindungsachse (Y-Richtung für Protonen 5 und 15, X-Richtung für Protonen 10 und 20), b„out of plane“- Komponente (Z). Position CH3 Methinprotonen andere ZnPPIX + Imidazol Aiso A1 A2 A3 2 -1,00 -0,24 -1,42 -1,34 12 -0,65 +0,13 -0,99 -1,09 7 +3,31 +4,36 +2,98 +2,60 18 +2,87 +3,88 +2,48 +2,24 5 -11,79 -4,49a -12,73b -18,15 10 -12,56 -4,87a -13,40b -19,40 15 -11,27 -4,27a -12,20b -17,35 20 -11,48 -4,45a -12,27b -17,72 3-1 -0,35 +0,79 -0,86 -0,98 3-2a +0,35 +0,01 +0,16 +0,89 3-2b +0,39 +0,20 +0,21 +0,76 8-1 +0,91 +0,12 -0,14 +2,74 8-2a -2,87 -1,49 -3,31 -3,79 8-2b -2,99 -1,11 -3,25 -4,60 13-1a -0,64 -0,17 -0,33 -1,20 13-1b -0,02 -0,17 -0,33 +0,43 17-1a +1,25 +0,68 +0,91 +2,18 17-1b +0,42 -0,09 -0,39 +1,73 79 Ergebnisse und Diskussion 3.1.3.2. Zink-Phäophorbid a Apo-Myoglobin (ZnPheid a Mb) und ZinkPyrophäophorbid a Apo-Myoglobin (ZnPPheid a Mb) 3.1.3.2.1. UV-Vis-Spektroskopie und Stabilität Erste Versuche Porphyrin-Derivate in Apo-Myoglobin zu binden wurden von Davies und Pearlstein durchgeführt[75]. Der Komplex aus Zink-Pyrophäophorbid a und ApoMyoglobin wurden bereits von Boxer et al. dargestellt und beschrieben[24], während der ZnPheid a Komplex mit Apo-Myoglobin noch nicht beschrieben wurde. Die UV-VisSpektren beider Kofaktoren werden durch die Qy-Bande im Bereich von 660 nm und die Soret-Bande im Bereich von 430 nm dominiert. Ist der Kofaktor ZnPheid a in Ether gelöst, aber durch Pyridin koordiniert, so liegt die Qy-Bande bei 656 nm und die Soret-Bande bei 427 nm. Zusätzlich werden drei kleinere Banden bei 610 nm, 567 nm und 525 nm beobachtet (Abbildung 37). A B Abbildung 37: UV-Vis-Spektren der freien Kofaktoren in Pyridin/Ether (schwarz) und der entsprechenden Myoglobinkomplexe (rot) von Zink-Phäophorbid a (A) und Zink-Pyrophäophorbid a (B). Durch den Einbau in die Proteinumgebung werden alle Banden in den langwelligen Bereich verschoben. Wird dieser Kofaktor im Myoglobin gebunden, so verschieben die Banden zu 661 nm (Qy) und 436 nm (Soret). Zusätzlich werden beide Banden breiter. Dies spricht nicht für die 80 Ergebnisse und Diskussion Koordination in einer geordneten Umgebung. Die beiden Banden bei 525 nm und 567 nm sind stark rot verschoben und überlagern mit der dritten Bande, die jetzt bei 622 nm liegt. Beim Zink-Pyrophäophorbid a tritt eine vergleichbare Verschiebung der Banden auf. Die Qy-Bande ist durch den Einbau in das Apo-Myoglobin von 655 nm zu 661 nm verschoben, die Soret-Bande verschiebt von 427 nm zu 437 nm. Im ungebundenen Zustand werden für diesen Kofaktor drei zusätzliche Banden bei 610 nm, 573 nm und 529 nm gemessen, die, ähnlich wie beim Zink-Phäophorbid a, durch den Einbau in das Apo-Myoglobin zu einer ins Rote verschobenen Bande bei 619 nm verschmelzen. Die Absorptionsbande bei 320 nm im ungebundenen Zustand liegt nach dem Einbau in das Protein bei 336 nm. Das ZnPPheid a weist im Vergleich zu ZnPheid a schmalere und strukturiertere Absorptionsbanden auf. Durch den Einbau in das Protein zeigt sich eine Verbreiterung der Qy-Bande, während die Soret-Bande unverändert bleibt. Für den ZnPPheid a Mb Komplex wird ein Extinktionskoeffizient bei 661 nm von 57 mM-1 cm-1 angegeben[24]. Theoretisch ist ein Bandenverhältnis der Qy-Bande zur Proteinbande bei 280 nm von 3,75 zu erwarten (εMyoglobin (Pferdemuskel) = 15,2 mM-1 cm-1). Experimentell konnte dieses Verhältnis nicht gefunden werden, stattdessen wurde ein Verhältnis von 2 bis 2,3 ermittelt. Da diese Ergebnisse nicht auf die Bildung eines stöchiometrischen Komplexes hindeutet, war es notwendig, die Stöchiometrie experimentell nachzuweisen. Zu diesem Zweck wurde eine bekannte Stoffmenge ApoMyoglobin mit einer Lösung bekannter Konzentration des Kofaktors in Pyridin titriert. Dies erforderte zunächst die Bestimmung des Extinktionskoeffizienten für den Kofaktor in Pyridinlösung, in der ein Einbau in das Apo-Protein erfolgreich war. Zur Bestimmung des Extinktionskoeffizienten wurde ein NMR-Spektrum einer unbekannten Menge Kofaktor in einem bekannten Volumen d5-Pyridin aufgenommen. Zusätzlich befand sich ein bekanntes Volumen undeuteriertes Dimethylformamid (DMF) in der NMR-Probe. Anhand des Spektrums konnte das Verhältnis der Stoffmengen von DMF und Kofaktor bestimmt werden, um anschließend die Konzentration des Kofaktors zu berechnen. Anhand eines quantitativen UV-Vis-Spektrums wurde ein Extinktionskoeffizient von 61,5 mM-1 cm-1 für ZnPheid a in Pyridin bei 661 nm bestimmt. Um das Titrationsexperiment auszuwerten, wird die Absorption der Qy-Bande und der Soret-Bande gegen das Stoffmengenverhältnis von Kofaktor zu Apo-Myoglobin (nKofaktor/nApo-Myoglobin) aufgetragen. Idealerweise nimmt die Absorption beider Banden bis 81 Ergebnisse und Diskussion zu einem Stoffmengenverhältnis von 1 linear zu und bleibt bei einer weiteren Erhöhung des Stoffmengenverhältnisses konstant. Experimentell wird eine lineare Zunahme der Absorption bis zu einem Wert von etwa 1,2 beobachtet. Im weiteren Verlauf der Titration nimmt die Absorption weiterhin linear, aber mit einer deutlich geringeren Steigung, zu (Abbildung 38). A B Abbildung 38: Titration von Apo-Mb mit den Kofaktoren ZnPheid a (A) und ZnPPheid a (B). Es wurde das Stoffmengenverhältnis n(Kofaktor)/n(Apo-Mb) gegen die Absorption der Soret-Bande (■) und der Qy-Bande (●) aufgetragen. Der lineare Anstieg der Absorption nach dem Äquivalenzpunkt ist dadurch zu erklären, dass die Absorptionsbanden des freien Kofaktors und des im Protein gebundenen Kofaktors nahezu an der gleichen Stelle liegen. Daher muss die Absorption bei kontinuierlicher Erhöhung der Konzentration des Kofaktors weiter ansteigen. Die geringere Steigung der Geraden hinter dem Schnittpunkt kommt durch die geringe Löslichkeit der Kofaktoren in Wasser zustande. Ein Teil des Kofaktors aggregiert und fällt aus. Auf diese Weise kommt auch die Abweichung vom linearen Verlauf in der Titrationskurve von ZnPheid a zustande. Die Absorption der Qy-Bande nimmt bei einer hohen Konzentration von gelöstem Kofaktor nicht weiter zu, da sich im UV-Vis-Spektrum eine zweite Bande bei 689 nm ausbildet. Auffällig ist, dass der Schnittpunkt der beiden Geraden nicht wie erwartet bei 1, sondern bei 1,2 liegt. Diese Abweichung hat verschiedene Ursachen. Zum einen liegt ein Teil des ungebundenen Kofaktors frei in Lösung vor und trägt zur Gesamtabsorption der Lösung bei. Zum anderen ist der 82 Ergebnisse und Diskussion experimentell ermittelte Extinktionskoeffizient mit einem gewissen Fehler behaftet. Insbesondere durch das Vorliegen des ungebundenen Kofaktors in der Lösung kann die Abweichung von etwa 20 % begründet werden. Weiterhin wurde durch Anionenaustauscher-Chromatographie gezeigt, dass nach der Reinigung der Probe kein unspezifisch gebundener Kofaktor vorhanden war. Das in der Literatur beschriebene Resultat, dass etwa 20 % des Kofaktors unspezifisch an der Oberfläche des Proteins bindet[24], konnte in dieser Arbeit nicht bestätigt werden. Die Ergebnisse der durchgeführten Titrationen zeigen, dass ein 1:1 Komplex von Kofaktor und Protein gebildet wurde. In den UV-Vis-Spektren wurde nicht das erwartete Verhältnis von Qy-Bande zur Proteinbande gefunden, weil die Kofaktoren auch im Bereich um 280 nm zur Gesamtabsorption der Probe beitragen. Die hier beschriebenen, erfolgreich eingebauten Phäophorbide weisen strukturelle Unterschiede zum nativen Kofaktor auf. In Position 81 liegt eine Ethyl- statt einer Vinylgruppe vor und Ring D ist ungesättigt, wodurch die Methylgruppe in Position 18 aus der Ringebene herausragt. Außerdem besitzen die Phäophorbide einen fünften Ring, der im Fall des Zink-Phäophorbid a eine Methoxycarbonylgruppe besitzt. Dennoch sollte im Vergleich zu Apo-Myoglobin eine signifikante Stabilisierung der Proteinstruktur durch den Einbau der Kofaktoren auftreten. Abbildung 39: Stabilitätsmessung von ZnPPheid a Mb (■), detektiert durch Fluoreszenz, im Vergleich zu Apo-Mb (■) und rekonstituiertem met-Mb (■). Die Stabilität des Proteins liegt bei 19,4 ± 1,4 kJ mol-1 und die Kooperativität liegt bei 15,9 ± 1,1 kJ mol-1 M-1. Damit ist der ZnPPheid a Mb Komplex deutlich stabiler als das Apo-Protein. 83 Ergebnisse und Diskussion Für den ZnPPheid a Mb Komplex konnte eine Stabilisierung des Proteinkomplexes gemessen werden (Abbildung 39). Die freie Entfaltungsenthalpie liegt für diesen Komplex bei 19,4 ± 1,4 kJ mol-1, gegenüber 10 kJ mol –1 für das Apo-Protein. Die Kooperativität nimmt ebenfalls gegenüber dem Apo-Protein (10 kJ mol-1 M-1) zu und liegt für den Komplex bei 15,9 ± 1,1 kJ mol-1 M-1. Dies ist ein Zuwachs um etwa 60 % gegenüber dem Apo-Protein. Für den ZnPheid a Mb Komplex kann dagegen keine signifikante Stabilisierung gemessen werden. Da dieses Resultat nicht durch die UV-Vis-spektroskopischen Messungen erklärt werden kann, soll mit Hilfe der NMR-Spektroskopie zum Verständnis der fehlenden Stabilisierung des ZnPheid a Mb Komplexes beigetragen werden. 3.1.3.2.2. NMR-Spektroskopie Das NMR-Spektrum von ZnPPheid a Mb wurde bereits von Boxer et al. publiziert[24]. Für den Komplex von ZnPheid a Mb sind bisher keine NMR-spektroskopischen Daten in der Literatur bekannt. Das 1 H-NMR-Spektrum (Abbildung 40) von ZnPPheid a Mb zeigt bei einer Messtemperatur von 25°C zwei Peaks bei –0,8 ppm und –2,5 ppm mit gleich großem Integral, die den γ-CH3-Gruppen der Aminosäure Valin 68 zugeordnet werden können. Im Gegensatz zu allen anderen Myoglobinkomplexen, die in dieser Arbeit untersucht wurden, zeigt dieses Spektrum keine Aufspaltung der einzelnen Peaks. Es konnte keine Veränderung des Signals in einem Zeitraum von 13 Monaten beobachtet werden, die auf einen Gleichgewichtsprozess hindeutet, wie er für das ZnPPIX Mb beobachtet wurde (vgl. Abschnitt 3.1.3.1.2). Neben dem Komplex von ZnPPheid a und Apo-Myoglobin (Pferdemuskel) wurde auch ZnPPheid a in rekombinantem Pottwal-Myoglobin (ZnPPheid a SwMb) bei 8°C untersucht. Es werden zwei Peaks bei –1,0 ppm und –2,8 ppm beobachtet. Die Signale liegen bei fast identischen Werten im Vergleich zum ZnPPheid a Mb Komplex (Tabelle 7). Bei Messungen am ZnPPheid a Mb konnte für beide Peaks eine Änderung der Signallage von 0,015 ppm/ °C bestimmt werden. Daher kann die Lage der Signale für ZnPPheid a SwMb bei 25°C berechnet werden. 84 ppm (t1) -0.50 -2.47 -0.78 Ergebnisse und Diskussion -1.00 -1.50 -2.00 -2.50 -3.00 -3.50 Abbildung 40: Ausschnitt aus dem 1H-NMR-Spektrum (400 MHz) von ZnPPheid a Mb bei 25°C. Zwei ppm (t1) -0.50 -2.83 -0.97 einzelne Signale der γ-CH3-Gruppen der Aminosäure Valin 68 deuten auf einen homogenen Komplex hin. -1.00 -1.50 -2.00 -2.50 -3.00 -3.50 Abbildung 41: Ausschnitt aus dem 1H-NMR-Spektrum (400 MHz) von ZnPPheid a SwMb bei 8°C. Ähnlich wie bei dem Komplex mit Myoglobin vom Pferdemuskel werden nur zwei Signale beobachtet. Die ppm (t1) -0.50 -1.00 -1.50 -2.00 -2.50 -2.91 -2.79 -2.67 -1.38 -1.20 Signalverbreiterung ist im wesentlichen auf die geringere Messtemperatur zurückzuführen. -3.00 -3.50 Abbildung 42: Ausschnitt aus dem 1H-NMR-Spektrum (400 MHz) von ZnPheid a Mb bei 8°C. 85 Ergebnisse und Diskussion Durch diese Vorgehensweise ergeben sich Werte von –2,47 ppm für Peak I und –0,78 ppm für Peak II. Die Signalpositionen stimmen mit den bereits publizierten Werten von –2,45 ppm und –0,70 ppm gut überein. Die Abweichung zwischen den Messwerten kann auf das Lösemittel Temperaturabhängigkeit Wasser der zurückgeführt Signalposition werden, das verursacht, eine sodass ausgeprägte bereits kleine Temperaturschwankungen eine sichtbare Auswirkung auf das Spektrum haben. Im Gegensatz dazu zeigt das NMR-Spektrum von ZnPheid a Mb eine deutliche Signalaufspaltung. Während bei Peak II zwei Signale bei –1,20 ppm und –1,38 ppm beobachtet werden, zeigt der Bereich um Peak I drei Signale bei –2,67 ppm, -2,79 ppm und –2,91 ppm. Eine Signalverdopplung kann durch das Auftreten von zwei Einbauisomeren des Kofaktors erklärt werden, jedoch ist das dritte Signal auf diese Weise nicht zu erklären. Offensichtlich ist der Komplex von ZnPheid a Mb inhomogen. Er zeigt nahezu ein Verhältnis von 1:1 der beiden Einbauisomere und zusätzlich ein weiteres Signal, dass nicht näher charakterisiert werden konnte. In früheren Rekonstitutionsexperimenten mit Hämin wurde die Existenz mindestens einer weiteren Einbauorientierung des Kofaktors vorgeschlagen[26]. Diese Komponente konnte in der Literatur jedoch wegen ihrer geringen Menge relativ zu den anderen Isomeren nicht analysiert werden. Tabelle 7: 1H-NMR-Daten (400 MHz) der γ-CH3-Gruppen der Aminosäure Valin 68 in den Komplexen von ZnPPheid a SwMb, ZnPPheid a Mb und ZnPheid a Mb. Peak I ZnPPheid a -2,90 b -1,08 b Mb -2,47 a -0,78 a ZnPPheid a -2,83 b -0,97 b SwMb -2,40 c -0,77 c ZnPheid a Mb -2,91, -2,79, -2,67 b a Peak II Messtemperatur 25°C, b Referenz[23] Peak I Peak II -2,45, -2,59 a -0,70 a -1,38, -1,20 b Messtemperatur 8°C, c Berechnete Werte für 25°C Messtemperatur unter der Annahme eines identischen Temperatureffektes bei ZnPPheid a Mb und ZnPPheid a SwMb Die NMR-Daten der verschiedenen Zn(P)Pheid a Mb Komplexe sind in Tabelle 7 zusammengefasst. Die Komplexe von ZnPPheid a mit Apo-Mb und derjenige von 86 Ergebnisse und Diskussion ZnPPheid a Apo-SwMb zeigen keine Inhomogenität in den NMR-Spektren, während der Komplex von ZnPheid a und Apo-Mb eine deutliche Inhomogenität zeigt. Deshalb wurde dieser Komplex von weiteren Untersuchungen ausgeschlossen. Da die Rekonstitution von ZnPPheid a in Apo-Mb und Apo-SwMb zu identischen Ergebnissen führte, wurden die EPR- und ENDOR-Untersuchungen auf den Komplex des Kofaktors mit Apo-SwMb beschränkt. 3.1.3.2.3. EPR-Spektroskopie Im Rahmen der EPR-Experimente werden die Spektren des freien Kofaktors ZnPPheid a mit denen des ZnPPheid a SwMb verglichen. Beide Spektren (Abbildung 43) zeigen ein Polarisationsmuster aus alternierenden emissiven und absorptiven Linien (eaeaea). Für Chlorophylle und deren Derivate wird angenommen, dass die Nullfeldparameter D > 0 und E < 0 sind[76; 77]. Durch die Simulation der Spektren wurden für den Proteinkomplex Nullfeldparameter von 303·± 4 10-4 cm-1 und –33 ± 4·10-4 cm-1 erhalten, für den freien Kofaktor wurden durch die Simulation nahezu identische Werte von 302 ± 4·10-4 cm-1 und -35 ± 4·10-4 cm-1 ermittelt. In der Simulation musste ein Besetzungsgrad von 70 % der gesamten Spins im Nullfeldsublevel TY angenommen werden, während die restlichen 30 % der Spins das TZ-Sublevel populieren, um eine gute Übereinstimmung den Linienintensitäten zu erzielen. Um die Breite der Banden in guter Näherung zu simulieren, musste ein anisotroper g-Faktor in der Simulation verwendet werden. Im Proteinkomplex wurden die Werte für gx=2,0042, gy=2,0040 und gz=2,0023 eingesetzt, während für die Simulation des freien Kofaktors der Wert für gx auf 2,0038 angepasst werden musste. Die restlichen g-Werte blieben unverändert. Mit diesen Parametern ist es möglich, die Messdaten mit guter Übereinstimmung zu simulieren. Ein Vergleich mit den Nullfeldparametern des MgPPheid a SwMb[78] (297·10-4 cm-1, 37·10-4 cm-1), die durch ODMR (=Optical Detected Magnetic Resonance) detektiert wurden, zeigt eine gute Übereinstimmung. Die Abweichungen liegen innerhalb des Messfehlers. Die Nullfeldparameter von Zink-Chlorophyll a in n-Oktan liegen bei 306·10-4 cm-1 bzw. -42·10-4 cm-1[79], diejenigen für Chlorophyll a in Polymethylmethacrylat 87 Ergebnisse und Diskussion (PMMA) sind mit 306·10-4 cm-1 und -43·10-4 cm-1 angegeben[80]. Damit unterscheiden sich die in früheren Arbeiten berichteten Werte für D kaum von denen in dieser Arbeit. Die Messwerte für E weichen jedoch deutlich von den Angaben für Chlorophyll a ab. Diese Abweichung ist wahrscheinlich auf die fehlende Methylester-Funktion in Position 132 zurückzuführen, die offensichtlich eine unsymmetrischere Verteilung des Elektrons in der x,y-Ebene des Moleküls bewirkt. Möglicherweise könnten die Abweichenden Werte für E aber auch darauf zurückzuführen sein, dass im Gegensatz zu allen früheren Studien keine Wechselwirkung der Chlorophylle untereinander möglich sind. A B Abbildung 43: Transiente EPR Spektren (Q-Band) von ZnPPheid a in Glycerin/Pyridin (A) und ZnPPheid a SwMb in Wasser/Glycerin (B) bei 10 K. Die aufgenommenen Spektren sind in schwarz und die entsprechende Simulation in rot gezeigt. Beide Spektren zeigen alternierend emissive und absorptive Linien (eae aea). In der Simulation der Spektren von 3Chlorophyll a und 3P700[81] im D-Band (130 GHz) wird für Chlorophyll a eine Umkehr der Reihenfolge gx > gy > gz zu gy > gx > gz beobachtet. Eine ähnliche Beobachtung wird für das ZnPPheid a und das ZnPPheid a SwMb gemacht. Der beschriebene Effekt wird durch eine unterschiedliche Lage der Nullfeld-Hauptachsen relativ zu den g-Tensorhauptachsen in beiden Systemen erklärt. Während die Nullfeldparameter nahezu identisch sind, ist die Verteilung der Spindichte auf beiden Molekülen unterschiedlich. Wie klar bereits bei geringen Magnetfeldstärken (QBand, 1,2 T, 35 GHz) der Effekt der g-Faktor Anisotropie hervorsticht, wird an den PulsEPR-Spektren von ZnPPheid a SwMb und ZnPPheid a erkennbar (Abbildung 44). Der 88 Ergebnisse und Diskussion Abstand zwischen den Übergängen Z und X unterscheidet sich um 20 G bzw. 27 G auf beiden Seiten der Spektren. Dieser Effekt ist bei höheren Frequenzen wesentlich stärker zu beobachten, wodurch eine bessere Interpretation ermöglicht wird. Tabelle 8: EPR-Parameter der simulierten Spektren von ZnPPheid a und ZnPPheid a SwMb im Vergleich zu den Parametern von P700 und Chlorophyll a[81] und P680[80]. Probe gx gy gz |D|·10-4 cm-1 |E|·10-4 cm-1 3 ZnPPheid a 2,0038 2,0040 2,0023 302 ± 4 35 ± 4 3 ZnPPheid a SwMb 2,0042 2,0040 2,0023 303 ± 4 33 ± 4 3 P700 2,00369 2,00323 2,00252 280 39 3 Chl a 2,00344 2,00382 2,00265 284 41 3 P680 - - - 287 42 3 ZnChl a (n-Oktan) - - - 306 42 3 Chl a (PMMA) - - - 306 43 B A Abbildung 44: Das Puls-EPR-Spektrum (Q-Band) von 3ZnPPheid a SwMb (A) und ZnPPheid a in Pyridin/Glycerin (B) bei 10 K zeigt deutlich die g-Faktor Anisotropie. Wäre der g-Faktor isotrop, müssten beide eingezeichneten Abstände gleich groß sein. X, Y und Z stehen für die Hauptrichtungen des Nullfeldtensors. Aufgrund der geringeren Feldstärken bei den hier durchgeführten Messungen ist die Präzision der in dieser Arbeit durchgeführten Simulationen geringer als für die publizierten 89 Ergebnisse und Diskussion Daten. Die Unterscheidung der gx- von den gy-Werten ist unter Berücksichtigung der Ungenauigkeit der Simulation nicht eindeutig, wodurch eine Interpretation erschwert wird. Jedoch könnte der Kofaktor ZnPPheid a mit und ohne Protein ein wertvolles System sein, um einen Beitrag zum besseren Verständnis der g-Faktor Anisotropie zu leisten. Zu diesem Zweck sind jedoch weiterführende Studien bei höheren Frequenzen nötig, um die Präzision der Simulationen zu verbessern. 3.1.3.2.4. ENDOR-Spektroskopie von ZnPPheid a in Pyridin/Glycerin Da die EPR-Spektren des freien Kofaktors und des Proteinkomplexes ein alternierendes Polarisationsmuster aufweisen (eaeaea), können in den ENDOR-Spektren Signale mit negativer Amplitude auftreten. Dies impliziert, dass diese Signale nicht aus der untersuchten Orientierung (z. B. XII-Orientierung), sondern aus einer anderen Orientierung (z. B. ZI oder YII) stammen. Ist ein Signal mit positiver Amplitude mit einem Signal mit negativer Amplitude überlagert, kann das Signal teilweise oder vollständig aus dem Spektrum verschwinden, abhängig von der Größe der Signalamplituden. Die ENDOR-Spektren werden wie schon beim ZnPPIX (Mb) anhand von quantenchemischen Rechnungen analysiert, wobei grundsätzlich bei der Z-Orientierung begonnen wird, da die Kopplungen in dieser Orientierung im Idealfall dem Wert für Aiso entsprechen. Das Spektrum des freien Kofaktors zeigt in der ZII-Orientierung fünf deutliche Signale (Abbildung 45). Das Signal 1 bei –10,3 MHz kann den beiden Methinprotonen in Position 10 und 20 zugeordnet werden. Die berechneten Kopplungen für Aiso liegen bei –8,2 MHz und –7,4 MHz und weichen damit von den experimentellen Ergebnissen ab. Allerdings wurde auch in den Berechnungen der Hyperfeinkopplungen der Chlorophyllradikale für die Methinprotonen ausschließlich zu kleine Werte berechnet. Auf der anderen Seite ist dieses Signal ein gutes Beispiel, warum die Methinprotonen im Idealfall als isolierte C-H Fragmente betrachtet werden können. Entspricht die Kopplungskonstante in Z-Orientierung etwa 1·Aiso, so müssten diese Signale in X-Orientierung bei etwa 0,5·Aiso liegen. In dieser Orientierung liegt das Magnetfeld in Richtung der C-H Bindung. In der Tat wird das Signal in der X-Orientierung bei –4,9 MHz gefunden. In der Y-Orientierung ist das 90 Ergebnisse und Diskussion Magnetfeld in der Ringebene, aber senkrecht zur C-H Bindung orientiert. Theoretisch sollte ein Wert von 1,5·Aiso auftreten was sich auch in den experimentellen Spektren nachweisen lässt. Die Signale liegen in Y-Orientierung bei –13,6 MHz. Abbildung 45: ENDOR Spektren (Q-Band) von ZnPPheid a in Pyridin/Glycerin an den Feldpositionen ZII (grün), XII (schwarz) und YII (rot). Alle Spektren sind so dargestellt, dass Kopplungen mit einem positiven Vorzeichen bei Werten größer als Null erscheinen. Kopplungen aus der selektierten Mannigfaltigkeit werden mit positiver Amplitude dargestellt, Kopplungen aus einer anderen Mannigfaltigkeit sind mit negativer Amplitude dargestellt. Die gestrichelten Linien deuten die Verschiebung der Signale abhängig von der Feldposition an. Blaue Linien werden verwendet, wenn ein Signal bei einer Orientierung nicht zugeordnet werden konnte. Blaue Sterne bezeichnen zugeordnete Signale mit negativer Amplitude, blaue Kreise bezeichnen nicht zugeordnete Signale mit negativer Amplitude. Das Signal 2 zeigt eine ähnliche Abhängigkeit von der Magnetfeldorientierung. In der Z-Orientierung liegt es bei –5,6 MHz (1·Aiso) und in der Y-Orientierung liegt es bei –2,4 MHz (0,5·Aiso). In der X-Orientierung wird das Signal bei –6,0 MHz beobachtet, wobei dieser Wert nicht mehr der Erwartung von 1,5·Aiso entspricht. Allerdings wird das Signal des Protons an dieser Position von den Signalen der beiden anderen Methinprotonen überlagert. Trotzdem kann die Kopplung von –5,6 MHz in der Z-Orientierung dem 5-H Methinproton zugeordnet werden, da die experimentellen Kopplungen mit dem erwarteten Trend - mit Ausnahme der X-Orientierung - übereinstimmen. Außerdem wurde für dieses 91 Ergebnisse und Diskussion Proton eine Hyperfeinkopplungskonstante von –6,3 MHz berechnet, die gut mit dem experimentellen Ergebnis übereinstimmt. Das Signal 3 beinhaltet zwei Kopplungen von +0,4 MHz und +0,8 MHz mit scharfen Signalen. Beide Signale treten bei X-orientiertem Magnetfeld mit negativer Amplitude (blaue Sterne) mit einer Kopplungsgröße von +0,5 MHz und +1,4 MHz auf und müssen daher aus einer anderen Mannigfaltigkeit stammen. Die entsprechenden Signale mit positiver Amplitude, die aus der X-Orientierung des Moleküls stammen, sind dagegen nicht identifizierbar. Möglicherweise liegen sie gemeinsam mit anderen Gruppen bei +3,7 MHz. In Y-Orientierung haben die Kopplungen eine Größe von +0,8 MHz und +1,7 MHz und beide Signale treten wieder scharf mit positiver Amplitude aus dem Spektrum hervor. Beide Signale sind isotrop und können den berechneten Kopplungen der Methylgruppen 18-CH3 (+0,4 MHz) und 7-CH3 (+0,6 MHz) zugeordnet werden. Das Signal 4 bei +4,1 MHz wird der 2-CH3 Gruppe zugeordnet. Das Signal liegt in der X-Orientierung bei +6,0 MHz und in der Y-Orientierung bei +4,4 MHz. Obwohl die berechnete Kopplungskonstante (+5,4 MHz) größer ist als die experimentell ermittelte Kopplung in der Z-Orientierung, ist die Zuordnung durch die Messungen am Proteinkomplex sicher (vgl. Abschnitt 3.1.3.2.5). Im Proteinkomplex wird dieses Signal nur schwach und stark verbreitert detektiert. Dies wird durch eine eingeschränkte Drehbarkeit der Gruppe durch die Proteinumgebung und die 3-Vinylgruppe verursacht. Für eine derartige Wechselwirkung kommt kein anderer der Substituenten in Frage. Die Methylgruppe in Position 12 wird dem Signal 5 bei +9,3 MHz zugeordnet. Sie weist als einzige in den Berechnungen eine ausreichend große positive Kopplungskonstante auf (+9,4 MHz). Die Signalposition ändert sich auf +11,0 MHz in der X-Orientierung und +9,2 MHz in der Y-Orientierung. Das Signal ist in den unterschiedlichen Orientierungen nahezu parallel zum Signal der 2-CH3 Gruppe verschoben. Das mit der Ziffer 6 bezeichnete Signal bei +3,7 MHz könnte von der 132-CH2 Gruppe verursacht werden, die nur in Pyrophäophorbiden vorkommt, da in dieser Position die Methoxycarbonlyfunktion entfernt worden ist. Allerdings ist diese Zuordnung nicht sicher, da nur ein Signal in der X-Orientierung detektiert werden konnte. In den beiden anderen Spektren fehlt das entsprechende Signal. 92 Ergebnisse und Diskussion Tabelle 9: Auflistung der zugeordneten ENDOR-Kopplungen für ZnPPheid a in Pyridin/Glycerin in MHz. Orientierung Signal 1 Signal 2 Signal 3 ZII -10,3 -5,6 +0,4; +0,8 +4,1 +9,3 n.d. XII -4,9 -6,0 n.d. +6,0 +11,0 +3,7 YII -13,6 -2,4 +0,8; +1,7 +4,4 +9,2 n.d. Zuordnung 10-H, 20-H 5-H 18-CH3; 7-CH3 2-CH3 -8,2; -7,4 -6,3 +0,4; +0,6 +5,4 berechnete Werte für Aiso Signal 4 Signal 5 Signal 6 12-CH3 132-CH2 +9,4 +3,7 Außer den Methinprotonen und den Methylgruppen konnte keine weitere Gruppe anhand der aufgenommenen Spektren eindeutig identifiziert werden. Neben der Vinylgruppe in Position 3 und der Ethylgruppe in Position 8 konnten keine Signale der Protonen in Position 17 und 18, sowie der CH2-Gruppen der Propionsäure beobachtet werden. Die Zuordnung der 132-CH2 Gruppe ist unsicher, da nur ein Signal in der X-Orientierung beobachtet werden konnte und ein entsprechendes Signal in den anderen Orientierung fehlt. Die Gruppen, denen keine Signale zugeordnet werden konnten, besitzen berechnete Kopplungen zwischen 0 und 1,5 MHz. Die große Anzahl kleiner Kopplungen in diesem Bereich erschwert die Zuordnung der Methylgruppen 7 und 18. Zwar sollten beide Methylgruppen das intensivste Signal in diesem Bereich aufweisen, jedoch können auch andere Gruppen diese Signale verursachen und eine Signalüberlagerung von mehreren Substituenten kann ebenfalls nicht ausgeschlossen werden. Daher ist die Zuordnung dieser Methylgruppen als wahrscheinlich, aber nicht als sicher anzusehen. 93 Ergebnisse und Diskussion 3.1.3.2.5. ENDOR-Spektroskopie von ZnPPheid a SwMb in Puffer/Glycerin Die ENDOR-Messungen des Proteinkomplexes ZnPPheid a SwMb wurden mit einem Gemisch aus 40/60 Vol% aus 50 mM Phosphatpuffer, pH 7,0, und Glycerin durchgeführt. Das Glycerin ist ein Glasbildner, der für eine hohe optische Transparenz der Probe und damit für eine hohe Lichteindringtiefe garantiert. Abbildung 46: ENDOR Spektren (Q-Band) von ZnPPheid a SwMb in H2O/Glycerin an den Feldpositionen ZI (grün), XI (schwarz) und YI (rot). Alle Spektren sind so dargestellt, dass Kopplungen mit einem positiven Vorzeichen bei Werten größer als Null erscheinen. Kopplungen aus der selektierten Mannigfaltigkeit werden mit positiver Amplitude dargestellt, Kopplungen aus einer anderen Mannigfaltigkeit sind mit negativer Amplitude dargestellt. Die gestrichelten Linien deuten die Verschiebung der Signale abhängig von der Feldposition an. Blaue Linien werden verwendet, wenn ein Signal bei einer Orientierung nicht zugeordnet werden konnte. Blaue Sterne bezeichnen zugeordnete Signale mit negativer Amplitude, blaue Punkte bezeichnen nicht zugeordnete Signale mit negativer Amplitude. Die Spektren des ZnPPheid a SwMb Komplexes (Abbildung 46) sind denen des freien Kofaktors sehr ähnlich. Es werden nur geringe Änderungen in den Kopplungskonstanten beobachtet. Entsprechend erfolgt die Zuordnung der Signale in gleicher Weise. Allerdings gibt es einige entscheidende Unterschiede. Während für den freien Kofaktor lediglich ein Signal mit negativer Amplitude beobachtet werden konnte, werden in den Spektren des Proteinkomplexes fünf Signale mit eindeutig negativer Amplitude detektiert. Eine 94 Ergebnisse und Diskussion eindeutige Zuordnung dieser Signale ist nicht möglich. Auffällig ist aber, dass in der X-Orientierung das Signal bei +3,4 MHz im Fall des Proteinkomplexes vollständig negativ ist. Wenn man die Ähnlichkeit der Spektren von freiem Kofaktor und Proteinkomplex berücksichtigt, ist an dieser Position ein starkes Signal mit positiver Amplitude zu erwarten. Offensichtlich wird es von einem Signal mit negativer Amplitude, aber ähnlicher Kopplungskonstante überlagert. Dies könnte ein Hinweis auf die Methylgruppen der Aminosäure Valin 68 sein, die ebenfalls mit dem Elektronenspin wechselwirken. Allerdings kann anhand der zur Verfügung stehenden Daten keine eindeutige Zuordnung durchgeführt werden. Tabelle 10: Auflistung der zugeordneten ENDOR-Kopplungen für ZnPPheid a SwMb in MHz. Orientierung Signal 1 Signal 2 Signal 3 Signal 4 Signal 5 ZI -10,6 -5,9 +0,5; +1,0 +3,9 +9,1 XI -8,2 -9,5 n.d. +6,1 +9,8 YI -15,3 -2,3 +0,8; +1,7 +5,2 +9,5 Zuordnung 10-H, 20-H 5-H 18-CH3; 7-CH3 2-CH3 12-CH3 -8,2; -7,4 -6,3 +0,4; +0,6 +5,4 +9,4 berechnete Werte für Aiso Das Signal 4 ist aufgrund der deutlichen Unterschiede im Vergleich zum freien Kofaktor der Methylgruppe in Position 2 zugeordnet worden. Wie bereits erwähnt liegt im freien Kofaktor an dieser Position ein scharfes Signal vor, dass in den Spektren des Proteinkomplexes nur verbreitert detektiert wurde. Wenn diese Beobachtung auf eine eingeschränkte Drehbarkeit der Methylgruppe aufgrund einer sterischen Wechselwirkung zwischen der Vinylgruppe in Position 3 und der Proteinumgebung zurückzuführen ist, kann dies durch eine temperaturabhängige Studie der ENDOR-Signale des Proteinkomplexes nachgewiesen werden. Mit zunehmender Temperatur sollte die freie Drehbarkeit der Methylgruppe wieder hergestellt werden und das Signal im Proteinkomplex als scharfe Bande auftreten. Allerdings nimmt die Spinrelaxation mit steigender Temperatur schnell zu, wodurch das Temperaturfenster, in dem eine solche Studie mit Puls-ENDOR-Methoden durchführbar ist, eingeschränkt. 95 Ergebnisse und Diskussion Die Unterschiede in den Kopplungskonstanten des Proteinkomplexes im Vergleich zum freien Kofaktor ist sehr gering, während bei dem Kofaktor ZnPPIX ein deutlicher Einfluss der Proteinumgebung festgestellt werden konnte. Eine mögliche Ursache für diese Beobachtung liegt wahrscheinlich in der Wahl des Lösemittels für den freien Kofaktor. In einer Pyridinlösung ist das ZnPPheid a fünffach koordiniert und dementsprechend verzerrt. Diese Geometrie wird durch den Einbau in das Protein, in dem der Kofaktor ebenfalls fünffach koordiniert wird, nicht oder nur in geringem Maß verändert. Wie die quantenchemischen Rechnungen gezeigt haben, hat die Abweichung der Geometrie von einer planaren Struktur zu einer gewölbten Struktur des Kofaktors, eine Änderung der Kopplungskonstanten von bis zu 1 MHz zur Folge. Diese Theorie könnte durch Messung der Kofaktoren ZnPPIX in Pyridinlösung und ZnPPheid a in einem schwach koordinierenden Lösemittel überprüft werden. Ist die Vermutung zutreffend, sollten für den Kofaktor ZnPPIX in einer Pyridinlösung ähnliche ENDOR-Spektren wie für den Proteinkomplex erhalten werden. Umgekehrt müssten die Spektren des Kofaktors ZnPPheid a in einem schwach koordinierenden Lösemittel insgesamt kleinere Kopplungen zeigen. 3.1.3.2.6. Vergleich der ENDOR-Daten von ZnPPheid a SwMb mit nativen Systemen Die aus den ENDOR-Spektren erhaltenen Daten werden mit den Literaturangaben, die für P680, dem „special pair“ des Photosystems II, zur Verfügung stehen, sowie mit dem Mittelwert der isotropen Hyperfeinkopplungskonstanten vom Radikalanion und –kation des Chlorophyll a verglichen. Für den Vergleich beider Radikale mit dem Triplettzustand wird die Näherung herangezogen, dass die Spindichte (ρT) am Kern i des Triplettzustandes gleich dem Mittelwert der Spindichten von Radikalanion (ρAn) und –kation (ρKat) ist (Gleichung 28)[82; 83]. ρ T (C i ) = 1 / 2[ρ Kat (C i ) + ρ An (C i )] (28) 96 Ergebnisse und Diskussion Bisher konnten lediglich in den Radikalen des Chlorophylls sämtliche Methylgruppen, die Methinprotonen und Signale der Vinylgruppe in Position 3 zugeordnet werden. In den nativen Systemen wurden bislang die Signale der Methinprotonen und der Methylgruppe in Position 12 zugeordnet. In dieser Arbeit konnten damit zum ersten Mal die Kopplungskonstanten aller Methylgruppen und der Methinprotonen eines Chlorophyll aDerivates in einer Proteinumgebung beobachtet und zugeordnet werden. Tabelle 11: Vergleich der ENDOR-Daten des Triplettzustandes in Z-Orientierung von P680, dem berechneten Mittelwert der experimentell ermittelten Kopplungskonstanten von Radikalanion und -kation von Chl a, ZnPPheid a SwMb und den für diese Arbeit berechneten Kopplungskonstanten Aiso des ZnPPheid a Imidazol Modellsystems. Position 3 P680[80] AZZ 3 3 P680[71] Exp. in ZI/II Chl a·± [84] ZnPPheid a berechnete Mittelwert: SwMb Werte für 1/2(AC+AA)[80; Exp. in ZI/II Aiso 85] 2-CH3 - - +4,2 +5,4 +5,37 12-CH3 +9,6 +9,5 +8,9 +9,4 +9,44 7-CH3 < +1,0 - +0,8 +1,0 +0,58 18-CH3 - - - +0,5 +0,43 17-H - - +5,0 - +1,03 18-H - - +5,9 - +1,56 132-H - - -1,9 - +3,70 5-H -1,4 -5,5 -2,3 -5,9 -6,33 10-H -5,4 -10,4 -5,8 -10,6 -7,36 20-H -1,4 -8,4 -2,2 -10,6 -8,18 3 -CH < +1,0 - +0,6 - - 32-CH2 -1,4 - -2,4 - - 1 Die Kopplungskonstante der Methylgruppe 12 liegt in allen vier Systemen bei etwa +9,5 MHz (vgl. Tabelle 11). Da es sich bei dieser Methylgruppe um die Kopplung von β-Protonen mit dem Elektronenspin handelt, ist das Signal isotrop und die Abweichung der 97 Ergebnisse und Diskussion isotropen Kopplungskonstante vom Messwert, der ebenfalls anisotrope Komponenten enthält, ist sowohl für P680 als auch für den Mittelwert der Radikale gering. Gleiches ist für die 7-CH3 Gruppe zutreffend, deren Signal bei Werten von ≤ 1 MHz in allen Spektren beobachtet wird. Für beide Gruppen stimmen die berechneten Werte ebenfalls gut mit den Messdaten überein. Die Angaben für die Methinprotonen von P680 weichen in den beiden publizierten Untersuchungen deutlich voneinander ab (Tabelle 11, Spalte 2 und 3). In der vorliegenden Arbeit konnte zwar eine negative Kopplung von –5,5 MHz beobachtet werden, die dem Methinproton in Position 5 zugeordnet wurde, allerdings konnten keine Kopplungen bei –1,4 MHz detektiert werden, weil die ENDOR-Spektren in diesem Bereich eine Überlagerung von Signalen mit negativer Amplitude aufweisen. Im Vergleich mit den experimentellen Daten von 3P680 (Tabelle 11, Spalte 3) tritt eine bemerkenswerte Abweichung von 2 MHz bei dem dritten Methinproton in Position 20 auf. Die Spektren in der vorliegenden Arbeit zeigen ein Überlagerung der breiten Signale der Methinprotonen in Position 10 und 20 zu einer unsymmetrischen Bande. Auf der Seite zu kleineren Kopplungen könnte man durch die Asymmetrie der Bande ein zusätzliches Signal bei etwa –9 MHz vermuten (Abbildung 46), jedoch wurde bei der Interpretation dieser Ergebnisse auf eine Unterscheidung der beiden Signale verzichtet. Insgesamt zeigen die Spektren des hier untersuchten Modellsystems eine gute Übereinstimmung mit den nativen Systemen und bieten auf Grund ihrer Einfachheit eine gute Grundlage, um die Interpretation komplexer nativer Systeme voranzutreiben. 3.1.3.3. Die Komplexe der Zink-Methylpyrophäophorbide mit ApoMyoglobin Um die Zahl verschiedener Kofaktoren in Myoglobin aus Pferdemuskel als Proteinmatrix zu erweitern, wurde der Einbau drei verschiedener Zink-Phäophorbid-Methylester in ApoMyoglobin untersucht. Neben Zink-Methylpyrophäophorbid a (ZnMePPheid a) wurde auch der Einbau von Zink-Methylpyrophäophorbid d (ZnMePPheid d) versucht. Keine der beiden Verbindungen bildet einen stöchiometrischen Komplex mit Apo-Myoglobin. Dies wurde anhand der UV-Vis-Spektren verifiziert. 98 Ergebnisse und Diskussion Abbildung 47: UV-Vis-Spektrum von ZnMePPheid a in Pyridin (schwarz) und in Apo-Mb komplexiert (rot). Das UV-Vis-Spektrum des freien Kofaktors ZnMePPheid a zeigt die typische Bandenstruktur, die bereits für die anderen Zink-Phäophorbide beobachtet wurde (Abbildung 47). Die Qy-Bande bei 654 nm und die Soret-Bande bei 428 nm dominieren das Spektrum. Außerdem werden die drei typischen Banden für den freien Kofaktor in Pyridin bei 610 nm, 571 nm und 527 nm detektiert. Durch den Einbau des Kofaktors in das Protein zeigt sich eine Verschiebung aller Banden zu größeren Wellenlängen. Jedoch ist sie geringer als für den vergleichbaren Kofaktor mit der freien Säure (ZnPPheid a). Die Qy-Bande wird um 4 nm zu 658 nm verschoben und die Soret-Bande um 8 nm zu 436 nm. Die drei Banden zwischen 525 nm und 615 nm verschmelzen durch den Einbau zu einer Bande bei 620 nm. Auch diese Tatsache wurde bereits bei ZnPPheid a beobachtet. Zweifelsfrei wird der Kofaktor im Protein gebunden, aber im Gegensatz zu ZnPPheid a ist das Verhältnis der Qy-Bande zur Proteinbande bei 280 nm nicht größer als 2, sondern deutlich kleiner als 1. Durch die Bildung des Methylesters wird keine Änderung des Extinktionskoeffizienten erwartet, da die Säurefunktion nicht mit dem delokalisierten π-System konjugiert ist. Wie in Abschnitt 3.1.3.2.1 gezeigt wurde, wird der Quotient von E660/E280 von 2,3 als Bildung eines stöchiometrischen Komplexes interpretiert. Weil der entsprechende Quotient E658/E280 im Fall des ZnMePPheid a Mb nur einen Wert von etwa 99 Ergebnisse und Diskussion 0,7 annimmt, lassen diese Ergebnisse den Schluss zu, dass nur etwa 25-30 % der vakanten Bindungsstellen im Myoglobin besetzt sind. Eine erneute Zugabe von freiem Kofaktor führte zu keiner signifikanten Änderung des Bandenverhältnisses. Die geringe Einbauquote kann eindeutig auf die Esterfunktion zurückgeführt werden, da ZnMePPheid a und ZnPPheid a ansonsten identisch sind. Abbildung 48: UV-Vis-Spektrum von ZnMePPheid d in Pyridin (schwarz) und in Apo-Mb komplexiert (rot). Ähnliche Ergebnisse wurden für ZnMePPheid d erhalten (Abbildung 48). Aufgrund der Aldehydfunktion in Position 3 besitzt der Kofaktor ZnMePPheid d ein um etwa 20 nm ins Rote verschobene UV-Vis-Spektrum. Die Soret-Bande liegt bei 443 nm und die Qy-Bande bei 678 nm. Durch den Einbau in das Apo-Myoglobin werden auch bei diesem Kofaktor sämtliche Banden in den langwelligen Bereich verschoben und liegen für den Komplex bei 457 nm (Soret) und 669 nm (Qy). Allerdings war es auch für diesen Kofaktor nicht möglich, einen stöchiometrischen Komplex zu erhalten. Ferner nimmt durch den Einbau in das Protein die Bandenbreite sowohl der Soret- als auch der Qy-Bande deutlich zu. Insbesondere die Basis der Qy-Bande ist stark verbreitert. Anhand des UV-Vis-Spektrums können zwei unterschiedliche Bindungsmöglichkeiten diskutiert werden. Einerseits findet ein Einbau in die Bindungstasche des Proteins statt, woraus die Absorptionsbanden bei 100 Ergebnisse und Diskussion 689 nm und 457 nm resultieren. Auf der anderen Seite lässt die breite Basis der Qy-Bande auf eine unspezifische Bindung, die möglicherweise auf eine Aggregation der Kofaktoren hindeutet, schließen. Diese Interpretation wird durch die Größe der Absorptionsbande bei 438 nm im Verhältnis zur Soret-Bande bei 457 nm unterstützt. Die ähnliche Größe beider Banden kann als Aufspaltung einer Bande interpretiert werden, die durch Aggregation der Kofaktoren hervorgerufen wird. Da die Bindungstasche des Proteins nicht genügend Raum für den Einbau eines Dimers zur Verfügung stellt und das Absorptionsspektrum der Probe nach einer zusätzlichen Aufreinigung durch Gelfiltation unverändert bleibt, könnte eine unspezifische Koordinierung des Kofaktors am Protein vorliegen. Vor dem Hintergrund dieser Ergebnisse wurde auf den Einbau von 3-devinyl-3hydroxymethyl-132-demethoxycarbonylphäophorbid a Methylester verzichtet, da aufgrund der Esterfunktion erwartet wird, dass kein stöchiometrischer Komplex gebildet werden kann und der Proteinkomplex daher nicht für weitere Studien geeignet ist. Zwei wesentliche Faktoren sind bei der Bewertung der Ergebnisse zu berücksichtigen. Zum einen sind die veresterten Kofaktoren nicht in der Lage, durch die Bildung einer Salzbrücke zur Proteinoberfläche an das Protein zu koordinieren. Als Folge davon sinkt die Einbauwahrscheinlichkeit in das Protein. Zum anderen sinkt die Löslichkeit der Kofaktoren durch die Esterbildung noch weiter, da in Form des Esters keine Möglichkeit besteht, stabile Wasserstoffbrücken mit dem Lösemittel Wasser zu bilden. Dadurch wird die Verweilzeit der Kofaktoren in der Lösung kürzer, bevor sie aggregieren und als Niederschlag dem Gleichgewicht entzogen werden. Sind diese beiden Gründe zutreffend, sollte theoretisch durch wiederholte Zugabe des gelösten Kofaktors der Besetzungsgrad der Proteinbindungstasche erhöht werden können. Jedoch gelingt dies in der Praxis nicht bzw. nur in sehr geringem Ausmaß, da die Wahrscheinlichkeit des Zusammentreffens eines gelösten Kofaktors und einer unbesetzten Bindestelle im Protein mit der Anzahl der freien Bindungsstellen im Protein sinkt. Um das Einbauergebnis zu verbessern müsste die Verweilzeit des Kofaktors in Lösung erhöht, also die Aggregation unterdrückt werden. Zu diesem Zweck wäre der Zusatz von organischen Lösemitteln, z. B. Methanol, denkbar. Allerdings ist bei diesen Experimenten zu berücksichtigen, dass bereits geringe Mengen (>5 Vol%) an organischem Lösemittel zur Denaturierung des Proteins führt. Wenn das unterschiedliche Verhalten der veresterten Kofaktoren allein auf die Esterfunktion zurückgeführt werden kann, muss dies auch für die veresterte Form des 101 Ergebnisse und Diskussion nativen Kofaktors zutreffen. In der Literatur ist der Komplex von Apo-Myoglobin und Eisen-Dimethylprotoporphyrin beschrieben[86]. Obwohl alle spektroskopischen Daten identisch sind mit dem Eisen-Protoporphyrin Apo-Mb Komplex, ist das Verhältnis der Soret-Bande zur Proteinbande für den Dimethylester um 25 % kleiner als für den unveresterten Kofaktor. Auch in diesem Fall liegt eine unvollständige Rekonstitution vor. Zusätzlich ist der Proteinkomplex bei pH 8,0 instabil und denaturiert. Diese Beobachtung wird für den nativen Kofaktor nicht gemacht. Diese Ergebnisse zeigen, dass durch eine Esterfunktion am Kofaktor die Bindungsfähigkeit an das Protein signifikant reduziert wird. Die Frage, ob der gebundene Kofaktor tatsächlich stabil gebunden ist und die Dissoziationskonstante des Komplexes durch die Esterfunktion nicht verändert wird, ist noch unbeantwortet. Allerdings könnte eine erhöhte Dissoziationskonstante erklären, warum die Einbaueffizienz nicht durch Zugabe von gelöstem Kofaktor erhöht werden kann. 102 Ergebnisse und Diskussion 3.1.4. Zusammenfassung Das von Boxer et al. entwickelte Rekonstitutionsverfahren, um Porphyrine in ApoMyoglobin einzubauen, konnte erfolgreich etabliert werden. Auf diese Weise wurden die Komplexe von ZnPPIX, ZnPheid a, ZnPPheid a, ZnMePPheid a und ZnMePPheid d mit Apo-Myoglobin erfolgreich dargestellt und charakterisiert. Zusätzlich konnte der Komplex von ZnPPheid a und SwMb dargestellt und charakterisiert werden. Der Kofaktor ZnPPIX konnte stöchiometrisch in Myoglobin eingebaut werden und durch UV-Vis-Spektroskopie charakterisiert werden. Die Proteinstabilität wurde durch den Einbau eines Kofaktors in das Apo-Myoglobin praktisch vollständig wieder hergestellt, wobei für die Kooperativität im Vergleich zu met-Myoglobin ein etwa 25 % geringerer Wert für den ZnPPIX-Komplex erhalten wurde. Mit Hilfe der NMR-Spektroskopie konnten in diesem Komplex zwei Einbauisomere nachgewiesen werden, wie sie bereits für den nativen Kofaktor, das Hämin, nach in vitro Rekonstitutionen beobachtet worden sind. Unter Verwendung der EPR-Spektroskopie konnten im lichtangeregten Triplettzustand mit der Literatur übereinstimmende Werte für die Parameter D und E der Nullfeldaufspaltung erhalten werden. Der Vergleich von freiem und Protein-komplexiertem Kofaktor zeigt diesbezüglich keine signifikanten Unterschiede. Im Gegensatz dazu sind in den ENDORSpektren deutliche Unterschiede der Kopplungskonstanten für den freien Kofaktor und den Proteinkomplex zu beobachten. Insgesamt zeigen die Spektren des Proteinkomplexes größere Kopplungen und eine deutliche Separation der einzelnen Signale. Diese Beobachtung wird als eine stärkere Delokalisation des Elektronenspins auf dem Molekülgerüst des Kofaktors interpretiert. Die Signale der Methinprotonen, sowie der Methylgruppen konnten für beide Systeme unter Verwendung von quantenchemischen Rechnungen zugeordnet werden. Die verwendete Methodik hat bereits bei der Berechnung der Hyperfeinkopplungskonstanten von Chlorophyllradikalen gute Übereinstimmungen mit den experimentellen Daten gezeigt und konnte diese Erwartung auch im Fall der lichtangeregten Triplettzustände erfüllen. Die Proteinkomplexe von ZnPheid a und ZnPPheid a konnten ebenfalls dargestellt werden und zeigten vergleichbare UV-Vis-Spektren. Während der Komplex des ZnPPheid a eine deutliche Stabilisierung der Proteinhülle zeigte, konnte dies für den ZnPheid a Komplex 103 Ergebnisse und Diskussion nicht beobachtet werden. Dies kann auf eine signifikante Inhomogenität dieses Komplexes zurückgeführt werden, die durch NMR-Spektroskopie nachgewiesen werden konnte. Aufgrund dieses Ergebnisses wurde der ZnPheid a Komplex nicht weiter untersucht, da er für weitere Messungen ungeeignet ist. Das ZnPPheid a Mb wurde analog zum ZnPPIX Mb durch EPR- und ENDOR-Spektroskopie untersucht. Auch in diesem Fall konnten gute Übereinstimmungen der Nullfeldparameter mit der Literatur beobachtet werden. Grundsätzlich werden auch in den ENDOR-Spektren dieses Proteinkomplexes, verglichen mit dem freien Kofaktor, größere Kopplungskonstanten gefunden. Allerdings ist die Zunahme der Kopplungsgröße nicht so signifikant im Vergleich mit dem Kofaktor ZnPPIX. Dies kann darauf zurückgeführt werden, dass der freie Kofaktor ohne Protein bereits fünffach koordiniert ist. Die daraus resultierende Geometrieverzerrung verursacht ähnliche Kopplungskonstanten wie sie für den Proteinkomplex beobachtet werden. Die Unterschiede zwischen freiem Kofaktor und dem Proteinkomplex sind auf den zusätzlichen Einfluss der Proteinumgebung, wie z.B. hydrophobe Wechselwirkungen, zurückzuführen. Ein Vergleich der erhaltenen ENDOR-Daten mit früheren Studien am P680, sowie den Chlorophyllradikalen zeigt eine sehr gute Übereinstimmung der Messwerte. Die Komplexe von Zink-Methylpyrophäophorbid a und d konnten zwar dargestellt und durch UV-Vis-Spektroskopie nachgewiesen werden, jedoch sind unter den gewählten Bedingungen keine stöchiometrischen Komplexe gebildet worden, wie die UV-VisSpektren zeigen. Daher wurde auf eine weitergehende EPR- und ENDOR- spektroskopische Charakterisierung verzichtet. In dieser Arbeit konnte damit der Grundstein für eine vollständige Charakterisierung der elektronischen Struktur von monomeren Chlorophyll-Derivaten in einer Myoglobin-Matrix gelegt werden. 104 Ergebnisse und Diskussion 3.1.5. Ausblick Ausgehend von den Ergebnissen dieser Arbeit bietet sich eine Vielzahl an zukünftigen Möglichkeiten. Um vollständige Daten der elektronischen Struktur der untersuchten Kofaktoren zu erhalten, ist die winkelabhängige Messung von Myoglobin-Einkristallen notwendig. Das rekombinante Pottwal-Myoglobin ist gut charakterisiert und die Kristallisationsbedingungen sind beschrieben[87-89], daher sollten Einkristalle mit nicht nativen Kofaktoren dargestellt werden können. Entsprechende Arbeiten wurden bereits begonnen. Außerdem können die bereits dargestellten Komplexe zur Aufklärung der g-Faktor Anisotropie, sowie der asymmetrischen Linien in den EPR-Spektren herangezogen werden, indem sie bei höheren Magnetfeldern untersucht werden. Die Analyse der Kristallstruktur würde neben der vollständigen elektronischen Struktur des nicht-nativen Kofaktors auch seine Positionierung in der Bindungstasche des Proteins aufklären, die bis heute nicht bekannt ist. Möglicherweise können auf diese Weise auch die Rotationsisomere der Kofaktoren, die durch NMR-Spektroskopie nachgewiesen werden konnten, detailliert analysiert werden. Anschließend bietet sich die Möglichkeit, entweder die Proteinumgebung gezielt zu verändern oder weitere Modifikationen an den Kofaktoren durchzuführen. Die Zahl der zu untersuchenden Kofaktoren kann auf alle (Bakterio)Chlorophylle ausgedehnt werden, sofern sie in die Bindungstasche einbauen. Zusätzlich können andere Metalle, z. B. Magnesium, anstatt Zink in den Kofaktoren gebunden werden. Grundlegende Untersuchungen der Bindungseigenschaften unterschiedlicher Porphyrine sind in dieser Arbeit unternommen worden und stehen zusätzlich auch in der Literatur zur Verfügung[19-21; 23; 90]. Die Aminosäurekette des Proteins Myoglobin kann in verschiedenen Positionen molekularbiologisch verändert werden, so dass der Einfluss der Proteinumgebung detailliert studiert werden kann. Die entsprechenden Expressionssysteme für Mutationen im Bereich der Bindungstasche sind bekannt und basieren auf heterologer Expression des Proteins in E. coli, einem Bakterium, das für seine Toleranz gegenüber unterschiedlichen Wachstumsbedingungen und seine hohe Wachstumsgeschwindigkeit bekannt ist. Auf diese Weise können unterschiedliche Myoglobin-Mutanten in wenigen Tagen erhalten werden. 105 Ergebnisse und Diskussion 3.2. Modellpeptide für [4Fe4S]-Zentren 3.2.1. Literaturübersicht Aufgrund der biologischen Relevanz von [4Fe4S]-Zentren sollen hier einige Übersichtsartikel vorgestellt werden. Einen umfassenden Überblick über alle Formen von FeS-Zentren gibt Lovenberg[36]. Die optischen und magnetischen Eigenschaften von [4Fe4S]-Zentren werden von Sweeney und Rabinowitz[91] übersichtlich zusammengefasst. Die EPR- und ENDOR-Daten von [4Fe4S]-Zentren aus verschiedenen Proteinen werden ausführlich von Mousca und Lamotte[92] diskutiert. Den Einfluss der Proteinumgebung auf das Redoxpotential haben Stephens, Jollie und Warshel unter Verwendung von quantenchemischen Methoden untersucht [42] , während speziell die Eigenschaften der FeS-Zentren des Photosystems I von Golbeck et al. zusammengefasst wurden[93]. Aufgrund der biologischen Relevanz von [4Fe4S]-Zentren gab es bereits mehrere Versuche, Modellsysteme für diese Systeme zu entwickeln. Diese synthetischen Peptide wurden in Länge und Sequenz der Aminosäurenkette variiert, um die minimalen Voraussetzungen zu bestimmen, die für die Bindung eines [4Fe4S]-Zentrums nötig sind. Außerdem wurde versucht, das Redoxpotential dieser Kofaktoren gezielt zu beeinflussen. In der Arbeitsgruppe um Dutton et al.[94-96] wurde eine Serie verschiedener Peptide mit einer Länge zwischen vier und 16 Aminosäuren synthetisch hergestellt, um die nötigen Grundvoraussetzungen der Primärsequenz für den Einbau eines [4Fe4S]-Zentrums zu studieren. Die Peptide, die erfolgreich ein [4Fe4S]-Zentrum binden konnten, besaßen das allgemeine Bindungsmotiv von Eisen-Schwefel-Proteinen. Es besteht aus mindestens sieben Aminosäuren, von denen drei Cysteine sind, die durch jeweils zwei Aminosäuren separiert werden (CxxCxxC). Diese Primärsequenz stellt drei der insgesamt vier Thiolatliganden, die die vier Eisenionen des kubischen [4Fe4S]-Zentrums ligandieren, zur Verfügung. Die vierte Position wird, sofern kein vierter Ligand durch das Peptid zur Verfügung gestellt wird, bei in vitro rekonstituierten Peptiden wahrscheinlich durch β-Mercaptoethanol eingenommen. Zusätzlich konnte nachgewiesen werden, dass eine 106 Ergebnisse und Diskussion Aminosäure mit sterisch anspruchsvoller Seitenkette hinter dem ersten Cysteinliganden zu einer deutlichen Verbesserung der Bindungsfähigkeit führt. In natürlichen Proteinen ist diese Position in den meisten Fällen durch Isoleucin oder Valin besetzt. Wahrscheinlich reduziert diese Aminosäure die Zugänglichkeit des Lösemittels Wasser zum Zentrum durch eine hydrophobe Abschirmung, wodurch die Stabilität des Zentrums erhöht wird. Trotz der Variation der Primärsequenz zeigten alle rekonstituierbaren synthetischen Peptide dieser Studie identische EPR Spektren und ein Redoxpotential von –350 mV gegenüber der Normal-Wasserstoffelektrode (NHE). In einem weiteren Ansatz wurde ein Peptid aus 16 Aminosäuren in die „loop“-Region eines vier-Helix-Bündels mit einer Gesamtlänge von 67 Aminosäuren eingebunden[96]. Neben einem Eisen-Schwefel-Zentrum war dieses Modell in der Lage einen zweiten Kofaktor in Form von Hämin zu binden. Damit ist dies das erste Beispiel für die erfolgreiche Kombination zweier unterschiedlicher redox-aktiver Kofaktoren in einem synthetischen Peptid. Auch dieses Modell weist ein Redoxpotential des FeS-Kofaktors von –350 mV auf. Durch Einführung der Bindungssequenz des Zentrums FX aus dem Photosystem I in die „loop“-Region eines bestehenden vier-Helix-Bündels (α4), das in der Arbeitsgruppe von William F. DeGrado entwickelt wurde, konnten Scott et al. [97] ein Peptid-gebundenes Eisen-Schwefel-Zentrum mit dem bis heute negativsten Redoxpotential von –420 mV darstellen. Obwohl die Bindungssequenz des FX-Zentrums aus Photosystem I übernommen wurde, liegt das Redoxpotential näher an dem eines Ferredoxins als an dem des nativen FX-Zentrums (-705 mV)[7]. Die Autoren schließen daraus, dass die Abschirmung des FeS-Zentrums gegenüber dem Lösemittel von größerer Bedeutung für das Redoxpotential ist als die tatsächliche Zusammensetzung der Aminosäuresequenz. 107 Ergebnisse und Diskussion 3.2.2. Design der Peptidsequenzen Alle bisher dargestellten Modellpeptide orientierten sich an der Aminosäuresequenz von Ferredoxinen. Weil die in dieser Arbeit dargestellten synthetischen Peptide Modelle der Eisen-Schwefel-Zentren des Photosystems I (PS I) darstellen sollen, dient die Primärsequenz der Untereinheit PsaC (Abbildung 49) als Vorbild für die Modellpeptide FA und FB. Abbildung 49: NMR-Struktur der Untereinheit PsaC des Photosystems I (30 Einzelstrukturen) in ungebundenem Zustand (PDB 1K0T)[98]. Der N-terminale Bereich und der C-Terminus sind in Lösung unstrukturiert und in rot dargestellt, die Position der Eisenatome ist als rote Kugeln, die der Sulfidionen als gelbe Kugeln dargestellt. Im nativen Vorbild folgen die Aminosäuren, die jeweils das Zentrum FA oder FB ligandieren, nicht direkt aufeinander (Abbildung 50). Stattdessen werden zwei Bindungsmotive für ein [4Fe4S]-Zentrum (CxxCxxCxxxC) gefunden, in denen die ersten drei Cysteine das erste Zentrum binden und das vierte Cystein das andere [4Fe4S]-Zentrum ligandiert. Dieser Umstand musste bei der Entwicklung der Peptidsequenzen berücksichtigt 108 Ergebnisse und Diskussion werden, da die native Sequenz für eine gestreckte Sekundärstruktur optimiert ist, um den Abstand zwischen den beiden Eisen-Schwefel-Zentren zu überbrücken. Im Gegensatz dazu müssen im Peptid alle vier Cysteine ein und dasselbe [4Fe4S]-Zentrum ligandieren. Daher wurde zwischen dem dritten und vierten Cystein ein Mini-„loop“ aus drei Aminosäuren eingeführt. Diese Methode zeigte bereits bei früheren Studien[94; 95], dass alle vier Cysteine dasselbe Zentrum ligandieren. ...-xxxC10xxC13xxC16xxxC20Px-...-xxxC47xxC50xxC53xxxC57Px-... Abbildung 50: Bindungsmotiv der Eisen-Schwefel-Zentren in der Untereinheit PsaC des Photosystems I. Die Liganden des Zentrums FB sind in rot, die Liganden des Zentrums FA sind in blau dargestellt. Jedes Bindungsmotiv wird durch die Aminosäure Prolin terminiert. Für das Peptid FB wurde die Sequenz der Untereinheit PsaC von Tyrosin 7 bis Cystein 16 übernommen, die bereits drei Cysteine als Liganden enthält. Darauf folgt ein „loop“ bestehend aus den Aminosäuren Lysin, Prolin und Glutamat und anschließend das vierte Cystein, gefolgt von einem Prolin. Für das Peptid FA wurden die Aminosäuren Threonin 44 bis Cystein 53 ebenfalls mit dem Loop und dem vierten Cystein versehen. Beide Sequenzen werden durch ein Prolin, gefolgt von einem Tryptophan als spektroskopische Sonde terminiert. Das Modellpeptid FA enthält drei Aminosäuren, die an der Bindung der Untereinheit PsaC an die Untereinheiten PsaA und PsaB beteiligt sind. Dies sind Valin 48, Lysin 51 und Arginin 52. Die Anbindung der Untereinheit erfolgt über ein ausgedehntes Netzwerk von Wasserstoffbrückenbindungen und hydrophoben Wechselwirkungen verschiedener Aminosäuren. Modellpeptid FA: TEDCVGCKRCKPECPW Native Sequenz FA: TEDCVGCKRC Modellpeptid FB: YDTCIGCTQCKPECPW Native Sequenz FB: YDTCIGCTQC Abbildung 51: Sequenzen der beiden Peptidmodelle für die Eisen-Schwefel-Zentren FA und FB, sowie die Teile der nativen Proteinsequenz, die beim Entwurf der Peptide übernommen wurden. 109 Ergebnisse und Diskussion FX1: RFPCDGPGRGGTCQVS FX2: AFPCDGPGRGGTCDIS Abbildung 52: Sequenzen der beiden Peptidmodelle für das Eisen-Schwefel Zentrum FX. Für beide Modelle wurde die Primärsequenz der Loops übernommen, die das Zentrum FX im Photosystem I ligandieren. Das Eisen-Schwefel-Zentrum FX im PS I wird durch zwei flexible Loops aus den Untereinheiten PsaA und PsaB gebunden, die jeweils zwei der insgesamt vier Cysteinliganden zur Verfügung stellen (Abbildung 53). Zwischen den zwei Cysteinresten, die in jedem „loop“ auftreten, liegen acht Aminosäuren, von denen jeweils vier Glycin sind. Diese Anordnung stellt eine große Herausforderung für ein möglichst kurzes synthetisches Peptid dar, da beide Loops ohne die restliche Kette der Untereinheit sehr flexibel sind. Da sich die beiden Loops nicht in einem Peptid von weniger als 20 Aminosäuren vereinen lassen, wurde jeweils ein Fragment, bestehend aus 16 Aminosäuren jedes Loops, synthetisiert. Von der Untereinheit PsaA wurde die Sequenz von Arginin 575 bis Serin 590 gewählt und von Untereinheit PsaB Alanin 562 bis Serin 577. Da jedes Peptid lediglich zwei Cysteinreste enthält, müssen sich für den erfolgreichen Einbau eines [4Fe4S]-Zentrums zwei Peptide zu einem Dimer zusammenlagern. Abbildung 53: Ausschnitt aus der Kristallstruktur von PS I (1JB0), der die Bindung des Eisen-SchwefelZentrums FX zeigt. Die Untereinheiten A und B bringen jeweils zwei der insgesamt vier Cysteinliganden ein. Rund um das FeS-Zentrum besitzt die Polypeptidkette keine ausgedehnten Sekundärstrukturelemente. 110 Ergebnisse und Diskussion 3.2.3. Charakterisierung der Modellpeptide Die unter Verwendung der Festphasenpeptidsynthese (SPPS) dargestellten Peptide wurden durch HPLC-Chromatographie und MALDI-TOF-Massenspektrometrie charakterisiert. Da in der SPPS die Sequenz der Aminosäuren vorgegeben wird, beschränkt sich die Charakterisierung auf Analyse der Homogenität und Identität der Peptide. Die Identität erfolgt durch Vergleich der berechneten mit der experimentell bestimmten Molekülmasse. Das Ziel hierbei ist es, Fehlsequenzen, Nebenreaktionen und verbliebene Schutzgruppen zu identifizieren. Die Homogenität wird durch Verwendung der HPLC-Chromatographie sichergestellt. FA FB FX1 FX2 Abbildung 54: MALDI-TOF-MS Spektren der vier synthetischen Peptide FA, FB, FX1 und FX2. Alle gefundenen Massen entsprechen den berechneten Werten und die Substanzen sind rein (>98 % HPLC). 111 Ergebnisse und Diskussion Die Massenspektren der einzelnen Peptide (Abbildung 54) zeigen jeweils das berechnete Molekülgewicht. Im Spektrum von FA konnte auch das Dimer des Peptides nachgewiesen werden. In Verbindung mit Luftsauerstoff können die Aminosäuren Cystein Disulfidbrücken untereinander ausbilden, die zur Dimerisierung der Peptide führen. Die Disulfidbrücken behindern die Rekonstitution der Peptide mit einem [4Fe4S]-Zentrum jedoch nicht, da sie unter den reduzierenden Rekonstitutionsbedingungen gespalten werden. Anhand der MS-Spektren konnte die Identität aller Peptide bestätigt werden. Die Reinheit betrug basierend auf analytischer HPLC-Chromatographie mindestens 98 %. 3.2.4. UV-Vis-Spektroskopie von FA und FB Beide Peptide zeigen nach Rekonstitution mit [4Fe4S]-Zentren eine breite Absorptionsbande im sichtbaren Bereich des Spektrums. Nach Reduktion mit Dithionit nimmt die Absorption im gesamten Bereich deutlich ab. Der größte Unterschied liegt bei 420 nm, die Abnahme beträgt dort über 50 % der ursprünglichen Absorption. Im oxidierten [4Fe4S]2+-Zustand ist die Absorptionsbande auf eine charge-transfer-Bande zwischen Eisen- und Schwefelkernen zurückzuführen. Die Spektren der oxidierten Verbindung und der reduzierten Spezies sind typisch für ein [4Fe4S]2+/1+-Zentrum[91]. A B Abbildung 55: UV-Vis-Spektren der Modellpeptide FA (A) und FB (B) im oxidierten (schwarz) und Dithionit reduzierten (rot) Zustand in 50 mM TrisHCl-Puffer, pH 8,3. 112 Ergebnisse und Diskussion Die Extinktionskoeffizienten bei 280 nm der Modellpeptide wurden nach Gleichung (29) berechnet. ε 280 (cm −1 M −1 ) = (Trp ) ⋅ 5500 + (Tyr ) ⋅ 1490 + (Cys) ⋅ 125 (29) In dieser Gleichung stehen (Trp), (Tyr) und (Cys) für die Anzahl der Aminosäuren Tryptophan, Tyrosin und Cystein in dem Peptid, dessen Extinktionskoeffizient bei 280 nm berechnet werden soll. Die Anzahl der Aminosäure wird mit ihrem molaren Extinktionskoeffizienten multipliziert und die erhaltenen Werte für die einzelnen Aminosäuren addiert[99; 100]. Auf diese Weise erhält man zuverlässige Ergebnisse für den Extinktionskoeffizienten einer beliebigen Aminosäuresequenz, sofern die mindestens ein Tryptophan enthalten ist. Für die Modellpeptide FA und FB werden auf diese Weise Werte von 6000 cm-1 M-1 (FA) und 7500 cm-1 M-1 (FB) für den Extinktionskoeffizienten bei 280 nm ermittelt. Für [4Fe4S]-Zentren wird ein Extinktionskoeffizient bei 400 nm von ε400 = 15000 cm-1 M-1 angenommen[101; 102]. Mit Hilfe dieser Werte wird der Peptidanteil bestimmt, der erfolgreich ein [4Fe4S]-Zentrum gebunden hat, indem die Konzentration des Modellpeptides und des [4Fe4S]-Zentrums in der Lösung berechnet wird. Das Verhältnis der Konzentrationen liefert den Anteil an Peptid in der Lösung, der ein [4Fe4S]-Zentrum gebunden hat. Für FA liegt der Anteil an Modellpeptid, das erfolgreich ein [4Fe4S]-Zentrum eingebaut hat, bei 10 % und für FB bei 20 %. Die Effizienz der Rekonstitution ist für beide Peptide gering, konnte aber auch durch Modifikation der Rekonstitutionsbedingungen nicht erhöht werden (vgl. Abschnitt 5.5.2). Der Grund für den geringen Einbau von [4Fe4S]-Zentren konnte nicht festgestellt werden, jedoch sind unterschiedliche Möglichkeiten denkbar. Die Konformation der Peptide könnte derart flexibel sein, dass nur ein geringer Teil die korrekte Konformation zur Bindung des Kofaktors annimmt. Auf der anderen Seite kann auch nicht ausgeschlossen werden, dass zwei oder mehr Peptide ein einziges [4Fe4S]-Zentrum ligandiert. Eine Gelfiltration unter anaeroben Bedingungen könnte über die Größe der rekonstituierten Peptide Aufschluss geben und Rückschlüsse auf die Zahl der Peptide pro FeS-Zentrum zulassen. Bisher konnte dieser Versuch nicht durchgeführt werden, da zum Ein- und Ausbau der Gelfiltrationssäule die Glove-Box demontiert werden müsste, und daher keine konstante Nutzung dieses 113 Ergebnisse und Diskussion Gerätes gewährleistet werden konnte. Der Versuch einer Gelelektrophorese unter anaeroben Bedingungen zur Bestimmung der Molekülgröße der rekonstituierten Peptide schlug fehl, da durch die angelegte Spannung das Wasser in der Elektrophoresezelle elektrolytisch zersetzt wird und der gebildete Sauerstoff zur vollständigen Zerstörung der FeS-Zentren führte. 3.2.5. EPR-Spektroskopie der Modellpeptide Um zu überprüfen, ob ein [4Fe4S]-Zentrum von den Peptiden gebunden wurde und in welchem Oxidationszustand es vorliegt, wurde die EPR-Spektroskopie verwendet. Obwohl ein [4Fe4S]-Zentrum im Oxidationszustand +2 aufgrund einer starken Kopplung der Elektronen nicht EPR-aktiv ist, gibt das Spektrum Aufschluss darüber, ob paramagnetische Verunreinigungen vorliegen. Nur in den Oxidationszuständen +1 oder +3 wird für ein [4Fe4S]-Zentrum ein intensives EPR-Signal erwartet. Die EPR-Spektren der Modellpeptide FA und FB zeigen im oxidierten Zustand ein schwaches, isotropes Signal, welches auf eine Verunreinigung durch ein [3Fe4S]-Zentrum zurückgeführt wurde. Die Bildung von geringen Mengen eines [3Fe4S]-Zentrums bei der in vitro Rekonstitution von Eisen-Schwefel-Proteinen ist praktisch unvermeidlich und wird auch bei nativen Proteinen beobachtet. Nach Reduktion der Probe mit Dithionit wird ein rhombisches EPR-Signal mit den g-Werten 2,04, 1,93 und 1,90 für das Peptid FA beobachtet, während für FB die g-Werte 2,05, 1,93 und 1,92 aus dem Spektrum erhalten werden. Identische Spektren werden für native [4Fe4S]-Zentren beobachtet[103]. Die gemessenen EPR-Spektren der Modellpeptide werden mit den Spektren von zwei Mutanten der Untereinheit PsaC des PS I verglichen. Die Mutante C14G C34S der Untereinheit PsaC (vgl. Abschnitt 5.5.3) besitzt ein intaktes FA-Zentrum und ein teilweise zerstörtes FB-Zentrum, dessen Signal nicht im EPR-Spektrum bei g = 2 auftritt. Eine zweite Mutante von PsaC, C51G C34S, ist komplementär dazu und besitzt ein intaktes FB-Zentrum, während der FA-Komplex nicht bei Werten um g = 2 zu detektieren ist. Diese Mutanten bieten die Möglichkeit, die Spektren der Zentren FA und FB getrennt voneinander zu detektieren. 114 Ergebnisse und Diskussion Beide Mutanten zeigen EPR-Spektren, die vollständig identische g-Werte, die genau mit den Werten, die für das Modellpeptid FA erhalten wurden, übereinstimmen. Die g-Werte des Modellpeptides FB zeigen eine geringe Abweichung bezüglich gx und gz. Jedoch ist die Variation gering und die g-Werte liegen in einem für [4Fe4S]-Zentren typischen Bereich[104]. Das Ergebnis der EPR-Spektroskopie zeigt, dass die Peptide ein [4Fe4S]-Zentrum in den Oxidationszuständen +2/+1 binden. Im Vergleich mit den Mutanten der Untereinheit PsaC wird eine gute Übereinstimmung der EPR-spektroskopischen Daten gefunden. A B Abbildung 56: CW-EPR Spektren (X-Band) der Modellpeptide FA (A) und FB (B) im reduzierten Zustand bei 10 K. Die g-Werte sind in den Spektren angegeben (gx > gy > gz) Tabelle 12: Zusammenfassung der erhaltenen g-Werte der Modellpeptide FA und FB, sowie die nativen Vorbilder, die Zentren FA und FB in der Untereinheit PsaC des PS I. Modellpeptid FA Modellpeptid FB FA in C14G C34S PsaC FB in C51G C34S PsaC gx 2,04 2,05 2,04 2,04 gy 1,93 1,93 1,93 1,93 gz 1,90 1,92 1,90 1,90 115 Ergebnisse und Diskussion In das Modellpeptid FX2 konnte ebenfalls erfolgreich ein [4Fe4S]-Zentrum inkorporiert werden (Abbildung 57), allerdings ist bei der Rekonstitution eine weitere EPR-aktive Verbindung gebildet worden, die nicht eindeutig charakterisiert werden konnte. Offensichtlich ist das Peptid nicht ausreichend spezifisch für den Einbau eines [4Fe4S]-Zentrums. Ein Grund dafür ist wahrscheinlich die Flexibilität der Primärsequenz, die im nativen PS I nötig ist, um das Eisen-Schwefel-Zentrum FX zu binden, allerdings für die kurze Peptidsequenz der Modelle negative Auswirkungen hat. Aufgrund dieses Ergebnisses wurden weitere Arbeiten am Modell FX2 zunächst zurückgestellt. Für das Modell FX1 konnte kein Einbau eines Eisen-Schwefel-Zentrums nachgewiesen werden. Hier liegen ähnliche Merkmale wie bei FX2 vor. Da sich die beiden Peptide allerdings nur um drei Aminosäuren unterscheiden, ist die Ursache für das unterschiedliche Verhalten offensichtlich durch diesen Unterschied begründet. Eine detaillierte Analyse konnte jedoch anhand der vorliegenden Daten nicht durchgeführt werden. Abbildung 57: CW-EPR Spektrum (X-Band) des Modellpeptides FX2 im reduzierten Zustand bei 15 K. Das Spektrum eines [4Fe4S]-Zentrums ist mit dem Spektrum einer anderen Spezies überlagert. 116 Ergebnisse und Diskussion 3.2.5.1. Temperaturabhängigkeit des EPR-Signals Im Oxidationszustand +1 sind [4Fe4S]-Zentren in einem Temperaturbereich unter 40 K durch EPR-Spektroskopie beobachtbar. Bei höheren Temperaturen sind die Linien aufgrund von Relaxationsprozessen stark verbreitert und sind nicht oder nur schwer zu detektieren. Um die bisherigen Daten zu bestätigen, wurde die Temperaturabhängigkeit der EPRSignale der beiden Modellpeptide zwischen 5 K und 40 K untersucht und mit den Daten der Mutante C14G C34S PsaC verglichen (Abbildung 58). Beide Modelle zeigen eine ausgeprägte Temperaturabhängigkeit des EPR-Signals im untersuchten Bereich. Bei einer Temperatur von 15 K wird sowohl für FB als auch die Mutante der Untereinheit PsaC die maximale Signalamplitude beobachtet. Das Modellpeptid FA besitzt dagegen eine maximale Signalamplitude bei 12,5 K. Außerdem ist das Signal bei 5 K bereits deutlich größer als für FB und die Mutante von PsaC. Bei höherer Temperatur wird das EPR-Signal durch Relaxationsprozesse verbreitert, die zu einer Abnahme der Signalamplitude führen. Offensichtlich sind die Relaxationsprozesse im Modell FA schneller als die von FB und PsaC bei gleicher Temperatur. Abbildung 58: Temperaturabhängigkeit der EPR-Signale der Modellpeptide FA (■) und FB (●) im Vergleich zu der Mutante C14G C34S PsaC (▲)von PS I bei 10 mW Mikrowellenleistung. 117 Ergebnisse und Diskussion 3.2.5.2. Leistungsabhängigkeit des EPR-Signals Die Abhängigkeit eines EPR-Signals von der eingestrahlten Mikrowellenleistung, insbesondere die Mikrowellenleistung bei halber Sättigung, ist eine charakteristische Größe für jedes System (vgl. Abschnitt 5.1.10). Die [4Fe4S]-Zentren sind im Bereich der konventionellen EPR-Spektroskopie bis zu einer Leistung von 200 mW nicht vollständig zu sättigen. Da die Sättigung mit zunehmender Mikrowellenleistung direkt von der longitudinalen (T1) und der transversalen (T2) Relaxationszeit abhängt, bietet dieses Verfahren Einblicke in die Relaxationsprozesse im Vergleich zu den nativen Vorbildern. Das EPR-Signal der Mutante C51G C34S zeigt im untersuchten Leistungsbereich die geringste Änderung der Signalintensität (Abbildung 59) und ist am schwierigsten zu sättigen. Das Modell FB und die Mutante C14G C34S sind auch in dieser Untersuchung am ähnlichsten und das Modell FA geht bei der geringsten Leistung in die Sättigung über. Da die exakte Leistung innerhalb des verwendeten Resonators nicht bestimmt wurde, wurde zum Vergleich der Modellpeptide mit den Mutanten von PsaC die am Gerät eingestellte Leistung für die Auswertung verwendet. Daher kann aus dieser Abbildung kein absoluter Wert für die Mikrowellenleistung bei halber Sättigung bestimmt werden. Auf den direkten Vergleich der einzelnen Proben hat dies allerdings keine Auswirkung. Abbildung 59: Leistungsabhängigkeit der EPR-Signale von FA (■), FB (■) und den Mutanten C14G C34S PsaC (■) und C51G C34S PsaC (■) bei 15 K. 118 Ergebnisse und Diskussion Ebenso wie die Analyse der Temperaturabhängigkeit der EPR-Signale deutet auch die Abhängigkeit der Signale von der Mikrowellenleistung auf deutlich kleinere Relaxationszeiten des Modellpeptides FA hin. Ein derartiger Unterschied bei im Prinzip identischen Systemen könnte durch Aggregation des Modellpeptides FA erklärt werden. Die gegenseitige Beeinflussung der FeS-Zentren mehrerer Modellpeptide durch ihre fluktuierenden Magnetfelder wäre ein möglicher Grund für die Abnahme der Relaxationszeiten. Die kürzeren Relaxationszeiten des Modells FA wirken sich negativ auf die Signalintensität der Puls-EPR-Spektren aus. Bei vergleichbaren Konzentrationen konnte lediglich etwa 20 % der Signalintensität im Vergleich zu FB in der Puls-EPR-Spektroskopie erhalten werden. Weil die Puls-EPR-Spektroskopie eine Grundvoraussetzung für ENDOR- und ESEEMSpektroskopie darstellt, wurde eine weitere Charakterisierung des Modellpeptides FA dadurch erschwert. 3.2.5.3. ESEEM- und ENDOR-Spektroskopie der Modellpeptide FA und FB Nachdem die Identität der [4Fe4S]-Zentren eindeutig nachgewiesen werden konnte (Abschnitt 3.2.6), wurde ihre Wechselwirkung mit der Peptidumgebung untersucht. Wie bereits erwähnt (vgl. Abschnitt 1.5) besitzen alle Eisen-Schwefel-Zentren Wasserstoffbrückenbindungen zwischen den Sulfidionen innerhalb des Zentrums und mindestens einem Amidproton der Polypeptidkette[38]. Sowohl in ENDOR-Spektren, als auch in ESEEM- (Electron Spin Echo Envelope Modulation) Spektren sollte diese Wechselwirkung sichtbar sein. Außerdem können anhand der ENDOR-Spektren Rückschlüsse auf die Spindichteverteilung innerhalb des Eisen-Schwefel-Zentrums gezogen werden. Die ENDOR-Spektren der Modellpeptide FA und FB in wässriger Lösung zeigen zwei unstrukturierte Banden mit einer Hyperfeinkopplungskonstante von 1,7 MHz (Abbildung 60 und Abbildung 61). Diese Banden können entweder Wasserstoffbrücken eines Amidprotons zu einem Sulfidion des Eisen-Schwefel-Zentrums oder den β-CH2-Protonen der Cysteinliganden zugeordnet werden. Um diese beiden Möglichkeiten eindeutig unterscheiden zu können, wurde das Lösemittel Wasser gegen D2O ausgetauscht. 119 Ergebnisse und Diskussion Alle austauschbaren Protonen, inklusive der Amidprotonen, sind dadurch im 1H-DaviesENDOR nicht mehr sichtbar und lediglich nicht-austauschbare Protonen, wie z. B. CH2-Protonen, verbleiben. Wie in Abbildung 60 gezeigt, hat der Austausch des Lösemittels gegen D2O keinen signifikanten Einfluss auf die Hyperfeinkopplungskonstante. Daher kann dieses Signal den β-CH2-Protonen der ligandierenden Cysteinreste zugeordnet werden. 2 H 1 H 1 H Abbildung 60: ENDOR-Spektren (Q-Band) des Peptides FB bei 4 K. 1H-Davies-ENDOR in H2O (schwarz) bei 12550 G, 1H-Davies ENDOR in D2O (rot) bei 12651 G und 2H-Mims-ENDOR in D2O (grün) bei 12651 G. Die Achse für das letzte Spektrum wurde durch das Verhältnis von gH/gD = 6,514 skaliert, um eine bessere Vergleichbarkeit zu ermöglichen. Das ENDOR-Signal zeigt allerdings keine Struktur, die für eine genauere Analyse herangezogen werden könnte. Die isotropen Kopplungskonstanten der β-CH2-Protonen hängen vom Diederwinkel zwischen den Ebenen Fe-S-C und S-C-H und der Spindichte des ungepaarten Elektrons auf dem Schwefelatom des Cysteins ab. Sie können nach einer erweiterten Heller-McConnel-Gleichung beschrieben werden[92; 105-107] . Auf Grundlage dieser Gleichung könnte eine Struktur des ENDOR-Spektrums im Sinne von Bindungswinkeln und Spindichteverteilung interpretiert werden, wodurch Einsichten in die elektronische und dreidimensionale Struktur gewonnen würden. Durch das Fehlen einer Struktur der ENDOR-Signale ist eine Interpretation in dieser Richtung nicht möglich. 120 Ergebnisse und Diskussion Im Vergleich zum entsprechenden Spektrum von C14G C34S PsaC fällt auf, dass dieses mit 10 MHz deutlich breiter ist als die Spektren der Modellpeptide, die lediglich 4 MHz Breite bei halber Signalhöhe aufweisen (Abbildung 61). Wenn man davon ausgeht, dass die Modellpeptide ähnliche Diederwinkel aufweisen, wie sie in den meisten [4Fe4S]2+/1+-Proteinen auftreten[106; 108] , kann diese Beobachtung mit einer geringeren Spindichte des ungepaarten Elektrons auf den Schwefelatomen der ligandierenden Cysteine erklärt werden. A B Abbildung 61: 1H-Davies-ENDOR-Spektrum des Modellpeptides FA bei 4 K (A) und der Mutante C14G C34S PsaC bei 4 K (B). Neben dem 1H-ENDOR-Spektrum wurde von FB zusätzlich ein 2H-ENDOR-Spektrum nach Austausch des Lösemittels gegen D2O aufgenommen (Abbildung 60). Es zeigt eine signifikante Kopplung eines Deuterium-Kerns mit dem Elektronenspin des EisenSchwefel-Zentrums. Dieses Ergebnis kann als Hinweis auf eine Wasserstoffbrücke interpretiert werden, da die Kopplung durch ein austauschbares Proton verursacht werden muss. Allerdings kann bei der geringen Größe des Peptids nicht ausgeschlossen werden, dass auch das Lösemittel direkt mit dem Elektronenspin wechselwirkt. Eine eindeutige Unterscheidung dieser beiden Möglichkeiten ist mit Hilfe der ESEEM-Spektroskopie an Proben ohne Deuterium Anreicherung möglich. Daher wurden von den Modellpeptiden und der Untereinheit PsaC ESEEM-Spektren (X-Band) bei 4 K aufgenommen (Abbildung 62). Diese Spektren sind für alle drei 121 Ergebnisse und Diskussion untersuchten Verbindungen identisch. Sie zeigen zwei Signale bei 15,4 MHz und 3,5 MHz, die einem Proton und einem Stickstoffatom zugeordnet werden können. Die Detektion des Stickstoffatoms ist aber nur dann möglich, wenn es mit dem Elektronenspin wechselwirkt. Daher handelt es sich bei der beobachteten 2 H-ENDOR-Kopplung mit hoher Wahrscheinlichkeit um eine Wasserstoffbrückenbindung zu einem Amidproton, da sonst keine Stickstoffverbindungen in Frage kommen. Obwohl theoretisch auch eine Kopplung zu den vorhandenen Aminosäuren Lysin in den Modellen FA und FB für diese Wasserstoffbrücke in Frage kommen könnte, ist dies unwahrscheinlich, da bisher in keinem nativen FeS-Protein eine solche Bindung beobachtet werden konnte. A B C Abbildung 62: 3-Puls-ESEEM-Spektren (X-Band) der Modelle FA (A) und FB (B) im Vergleich zu einem 3-Puls-ESEEM-Spektrum von C14G C34S PsaC (C) in H2O bei 4 K. 122 Ergebnisse und Diskussion 3.2.6. Mössbauer-Spektroskopie Die Mössbauer-Spektroskopie bietet die Möglichkeit, ein Eisen-Schwefel-Zentrum sowohl im oxidierten, als auch im reduzierten Zustand zu charakterisieren. Anhand der Quadrupolaufspaltung ∆EQ und der Isomerenverschiebung δ können die verschiedenen Arten von Eisen-Schwefel-Zentren unterschieden werden[109]. Um die dargestellten Mössbauer-Spektren verstehen zu können, ist eine kurze Übersicht über die Redoxzustände und deren Verteilung innerhalb des FeS-Zentrums nötig, allerdings werden nachfolgend nur die relevanten Oxidationszustände [4Fe4S]2+/1+ beschrieben. Im oxidierten [4Fe4S]2+-Zustand liegen formal zwei Fe3+ und zwei Fe2+ im FeS-Zentrum vor. Aufgrund der antiferromagnetischen Kopplung der Ionen untereinander liegt ein System mit dem Gesamtspin S = 0 vor. Im Mössbauer-Spektrum werden jedoch nicht die Signale von zwei unterschiedlichen Paaren (2 x Fe3+, 2 x Fe2+) beobachtet, sondern ein Signal, dass auf zwei identische Paare zurückgeführt werden muss[110-112]. Die Eisenionen liegen daher alle identisch als Fe2,5+ vor. Wird das FeS-Zentrum reduziert und liegt anschließend im [4Fe4S]2+-Zustand vor, wird die formale Betrachtung komplizierter. Im einfachsten Fall liegen jetzt drei Fe2+- und ein Fe3+-Ion im FeS-Zentrum vor. Aus der antiferromagnetischen Kopplung ergibt sich ein Gesamtspin von S = ½. In diesem Zustand liegen ebenfalls zwei Paare vor, die aus 2 x Fe2,5+ und 2 x Fe2+ bestehen. Sind die beiden Paare jeweils lokalisiert, werden im Mössbauer-Spektrum zwei Signale mit Isomerenverschiebungen von δ = 0,49 mm/s (Fe2,5+) und δ = 0,62 mm/s (Fe2+) gefunden. Jedoch müssen diese beiden Paare nicht notwendigerweise lokalisiert vorliegen. Wenn die kubische Struktur des [4Fe4S]-Zentrums unverzerrt vorliegt, können sie ebenfalls über den Kubus delokalisiert sein und verursachen auf diese Weise ein Signal, welches dem Mittelwert der beiden Einzelsignale entspricht, in diesem Fall wäre eine Isomerenverschiebung von etwa δ = 0,55 mm/s zu erwarten. Die Mössbauer-Daten der Modellpeptide FA (Abbildung 63) und FB (Abbildung 64) im oxidierten Zustand zeigen das charakteristische Spektrum für ein kubisches [4Fe4S]-Zentrum mit vier identischen Fe2,5+ Kernen[109]. Im [4Fe4S]2+-Zustand weisen die Modelle für das Zentrum FA und FB eine Isomerenverschiebung von 0,43 mm/s bzw. 0,46 mm/s auf. Die Quadrupolaufspaltung liegt mit einem Wert von 0,99 mm/s für das 123 Ergebnisse und Diskussion Peptid FA und 0,97 mm/s für das Peptid FB ebenfalls bei charakteristischen Wert für [4Fe4S]2+-Zentren. A B Abbildung 63: Mössbauer-Spektren des Modellpeptides FA in oxidiertem (A) und reduziertem (B) Zustand bei 80 K. Die gemessenen Daten sind als schwarze Punkte dargestellt. Die Interpolation der Hauptspezies ist als rote Linie, die Interpolation der Nebenspezies als grüne Linie und die Summe der interpolierten Kurven als schwarze Linie (schwarz) dargestellt. A B Abbildung 64: Mössbauer-Spektren des Modellpeptides FB in oxidiertem (A) und reduziertem (B) Zustand bei 80 K. Die gemessenen Daten sind als schwarze Punkte dargestellt. Die Interpolation der Hauptspezies ist als rote Linie, die Interpolation der Nebenspezies als grüne Linie und die Summe der interpolierten Kurven als schwarze Linie (schwarz) dargestellt. Im oxidierten Zustand wurde keine Verunreinigung durch Fe(II) gefunden. Im reduzierten Zustand wird für die Modelle eine Isomerenverschiebung von 0,51 mm/s (FA) und 0,52 mm/s (FB) beobachtet. Diese Werte können – wie oben beschrieben- als ein delokalisiertes Fe2,5+ Fe2,5+ und ein delokalisiertes Fe2+ Fe2+ Paar interpretiert werden. Die 124 Ergebnisse und Diskussion experimentell ermittelten Werte sind geringfügig kleiner als der Mittelwert der beiden lokalisierten Paare und können durch Delokalisation erklärt werden. Die Quadrupolaufspaltung liegt bei 1,03 mm/s und 0,69 mm/s für FA und FB. Die Abnahme der Quadrupolaufspaltung für FB durch die Reduktion ist ungewöhnlich. Diese Beobachtung kann durch die Überlagerung der einzelnen Feldgradienten der Eisenkerne erklärt werden. Jeder Feldgradient besitzt ein Vorzeichen, das allerdings nicht aus den MössbauerSpektren ermittelt werden kann. Wenn der Feldgradient des durch die Reduktion entstandenen (formalen) Fe2+ innerhalb des Kubus ein entgegengesetztes Vorzeichen zu den anderen Eisenkernen hat, kollabiert die Quadrupolaufspaltung des Spektrums, da sie die Summe aller Feldgradienten darstellt. Tabelle 13: Zusammenfassung der Mössbauer-Parameter der Modellpeptide im oxidierten und reduzierten Zustand im Vergleich zu den Mutanten der Untereinheit PsaC von PS I. Probe δ [mm/s] ∆EQ [mm/s] Linienbreite FA Modellpeptid (ox) 0,43 0,99 0,56 FA Modellpeptid (red) 0,51 1,03 0,63 FB Modellpeptid (ox) 0,46 0,97 0,56 FB Modellpeptid (red) 0,52 0,69 0,61 C14G C34S PsaC (ox), FA 0,44 0,95 0,44 C14G C34S PsaC (red), FA 0,50 0,97 0,55 C51G C34S PsaC (ox), FB 0,44 0,88 0,58 C51G C34S PsaC (red), FB 0,50 0,97 0,55 In den Spektren des Modells FA und der reduzierten Form des Modells FB tritt zusätzlich eine zweite Spezies mit einer Isomerenverschiebung von 1,28 mm/s und einer Quadrupolaufspaltung von 2,99 mm/s auf. Diese Verunreinigung konnte als Fe(II)-Spezies identifiziert werden, die fünf- oder sechsfach durch Oxoliganden gebunden ist. Bei dieser Verbindung handelt es sich um Eisenoxide bzw. –hydroxide, die aufgrund ihrer kolloidalen Struktur über eine Gelfiltration nicht abzutrennen sind. Obwohl diese Verunreinigung bei den Mössbauer-Messungen ohne externes Magnetfeld keine negativen Auswirkungen zeigt, verhindert sie eine Analyse der Proben in Abhängigkeit vom externen Magnetfeld. 125 Ergebnisse und Diskussion Die Eisen(III)kerne sind durch Oxo- und Hydroxobrücken derart magnetisch gekoppelt, dass sie sich wie ein Spin verhalten, der durch seine Relaxation am Kernort der zu untersuchenden Spezies ein fluktuierendes Magnetfeld erzeugt. Durch diesen „superparamagnetische Relaxation“ genannten Prozess tritt im Mössbauer-Spektrum bei externem Magnetfeld eine große Zahl unterschiedlicher Linien auf, die eine Interpretation erschweren oder, wie in diesem Fall, unmöglich machen. Die Peptidmodelle besitzen im Vergleich zu den beiden Mutanten von PsaC sehr ähnliche Mössbauer-Parameter. Unabhängig von der EPR- und der UV-Vis-Spektroskopie konnte durch die Mössbauer-Spektroskopie gezeigt werden, dass die beiden Modellpeptide ein [4Fe4S]2+/1+-Zentrum binden. 3.2.7. Bestimmung des Redoxpotentials der Modellpeptide FA und FB Das Redoxpotential ist der wesentliche Parameter, der die Funktion eines redox-aktiven Kofaktors bestimmt. Daher ist ein Vergleich dieses Wertes mit den nativen FeS-Zentren des PS I ein wesentliches Kriterium für die Qualität der Modellpeptide. Das Redoxpotential der Eisen-Schwefel-Zentren wurde durch eine Redoxtitration unter anaeroben Bedingungen bestimmt und durch UV-Vis-Spektroskopie der Lösung in einer gasdichten Küvette (d = 1cm) kontrolliert. Dabei wurde die Abnahme der Absorption bei 420 nm in Abhängigkeit vom Potential verfolgt und die erhaltenen Daten nach der NernstGleichung interpoliert (Abbildung 65). Die EPR-Proben der Lösung werden in der Glovebox in flüssigem Stickstoff eingefroren und nach Beendigung der Titration bei 15 K und 10 mW Mikrowellenleistung vermessen. Die EPR-Spektren wurden qualitativ ausgewertet, wobei lediglich Proben berücksichtigt wurden, die kein Signal des reduzierten [4Fe4S]-Zentrums aufwiesen bzw. beim negativsten Potential der Titration ein starkes Signal im EPR-Spektrum zeigten. Auf diese Weise wurde der Bereich des Redoxübergangs eingegrenzt. Zusätzlich wurde eine Titration mit den Mediatoren Methylviologen (-680 mV), Phenosafranin (-515 mV) und Indigotetrasulfonat (-280 mV) durchgeführt, um einen optimalen elektrischen Kontakt zwischen Lösung und Elektrode zu garantieren. Die 126 Ergebnisse und Diskussion Konzentration der Mediatoren betrug jeweils 0,5 mM in der Lösung. Diese Titration wurde wie oben beschrieben durch die EPR-Spektroskopie ausgewertet, um den Übergangsbereich des Redoxpotentials eingrenzen zu können. Eine Auswertung dieser Titration durch die UV-Vis-Spektroskopie war nicht möglich, da die Mediatoren deutlich größere Extinktionskoeffizienten aufweisen als die Modellpeptide, die sich zudem für die oxidierte und die reduzierte Form der Mediatoren unterscheiden. Unabhängig von der Verwendung der Mediatoren wurde in allen Messungen ein nahezu identisches Potential für die Modellpeptide ermittelt. A B Abbildung 65: Redoxtitration der Peptidmodelle FA (A) und FB (B) mit Dithionit. Durch UV-VisSpektroskopie bestimmte Messpunkte sind in schwarz dargestellt, EPR-Messpunkte in rot. Bei der Interpolation der Messpunkte wurde die Zahl der Elektronen (n) in der Nernstgleichung als n = 1 definiert, da es sich um eine ein-Elektronen-Reduktion handelt, während das Gleichgewichtspotential angepasst wurde. Aus den Messwerten für FA ließ sich ein Redoxpotential von –491 ± 30 mV ermitteln, während für FB ein kleinerer Wert von –471 ± 30 mV bestimmt wurde. Im Rahmen des Fehlers sind beide Werte identisch. Allerdings können die Datenpunkte für FB deutlich besser interpoliert werden. Bei der Titration von FA werden vollständig oxidierte und vollständig reduzierte Probe bei sehr geringen Potentialunterschieden beobachtet. Dieses Ergebnis lässt den Schluss zu, dass keine vollständige Gleichgewichtseinstellung zwischen oxidierter und reduzierter Form des [4Fe4S]-Zentrums vorliegt, wodurch die Potentialmessung ungenau wird. Außerdem findet die Titration an der unteren Grenze der Reduktionskraft von Dithionit statt, so dass keine 127 Ergebnisse und Diskussion Messdaten bei negativeren Potentialen aufgenommen werden konnten. Die Titration des Modellpeptids FB liefert bessere Ergebnisse, da die Messwerte den interpolierten Verlauf der Nernst-Kurve besser widerspiegeln. Bei dieser Titration können zwei Übergänge identifiziert werden. Die Reduktion der Hauptspezies bei –470 mV und die Reduktion einer zweiten Spezies, die etwa 10 % der Abnahme der UV-Vis-Absorption bei 420 nm verursacht. Das Redoxpotential dieser Verunreinigung liegt bei etwa –200 ± 30 mV und damit im typischen Bereich eines [3Fe4S]-Zentrums[42], das bereits in der EPR-spektroskopischen Charakterisierung als Verunreinigung auftrat. Auf die durch Interpolation der Messpunkte ermittelte Lage des Redoxpotentials der Hauptspezies hat diese Verunreinigung keinen Einfluss. Um die erhaltenen Daten abzusichern, wurde versucht, das Reduktionsmittel Dithionit durch Natriumborhydrid (NaBH4) oder Titan(III)citrat zu ersetzen. Beide Verbindungen weisen ein deutlich negativeres Redoxpotential als Dithionit auf. Allerdings konnte bei der Verwendung dieser Reduktionsmittel kein EPR-Signal eines reduzierten [4Fe4S]-Zentrums beobachtet werden. Aus welchem Grund die Modellpeptide mit Dithionit, aber nicht mit NaBH4 oder Titancitrat reduziert werden können, konnte nicht geklärt werden. Möglicherweise wird das FeS-Zentrum reduktiv zerstört. Die Redoxpotentiale der Eisen-Schwefel-Zentren in der Untereinheit PsaC liegen bei –530 mV für FA und –580 mV für FB[10] bzw. bei –465 mV für FA und –440 mV für FB [9] (vgl. Abschnitt 1.2.2). Die Bestimmung der Redoxpotentiale bei Raumtemperatur, die zu den positiveren Literaturwerten führen, stimmen gut mit den Werten überein, die für die Modellpeptide bei Raumtemperatur erhalten wurden. Für die Modellpeptide sollten diese Werte unabhängig von der Temperatur sein, weil eine Verschiebung des Gleichgewichtes mit der Temperatur, wie sie für die nativen Systeme diskutiert wird, aufgrund der Abwesenheit eines reversiblen Elektronenakzeptors oder –donors, nicht möglich ist. Im nativen PS I kann das Elektron dagegen abhängig von der Temperatur auf den Zentren FA und FB delokalisiert vorliegen, wodurch die Messergebnisse beeinflusst werden könnten. Das negative Redoxpotential wird möglicherweise durch zwei der 16 Aminosäuren entscheidend beeinflusst. Die Modellpeptide FA und FB unterschieden sich in den Positionen 8 und 9 von der Mehrheit der natürlichen Proteine mit [4Fe4S]-Zentren. Bei Ferredoxinen unterschiedlicher Spezies wird in diesen Positionen ein Glycin bzw. ein Alanin gefunden, also unpolare Aminosäuren mit kleinen Seitenketten. Im Modellpeptid 128 Ergebnisse und Diskussion FA sind diese Positionen durch Lysin bzw. Arginin und im Modellpeptid FB durch Threonin bzw. Glutamin besetzt. Der Austausch einer unpolaren Aminosäure gegen eine polare oder geladene Aminosäure sollte einen drastischen Einfluss auf das Redoxpotential des FeS-Zentrums haben. In synthetischen Modellen, die an diesen Positionen die unpolaren Aminosäuren aufwiesen, wurden Redoxpotentiale von –350 mV gefunden[94]. Das Ergebnis der Redoxtitration zeigt, dass die kurze Aminosäuresequenz mit 16 Resten effektiv in der Lage ist, die eingebauten [4Fe4S]-Zentren gegen das Lösemittel abzuschirmen und eine ausreichend hydrophobe Umgebung zu bilden. Trotz ihrer geringen Größe weisen die beiden Peptide FA und FB die negativsten Reduktionspotentiale aller künstlichen [4Fe4S]-Zentren auf. 3.2.8. NMR-Spektroskopie Bei einem Polypeptid von 16 Aminosäuren bietet sich zur Bestimmung der Struktur die NMR-Spektroskopie an, da die Seitenketten der Cysteinreste durch den engen Kontakt zu den paramagnetischen Zentren direkt identifiziert werden können. Durch Verwendung von eindimensionalen und zweidimensionalen NOE-Spektren können die restlichen Aminosäuren identifiziert und eine Struktur in Lösung bestimmt werden, die für die weitere Analyse der EPR- und ENDOR-Daten in Bezug auf die Spindichteverteilung des ungepaarten Elektrons in dem FeS-Zentrum hilfreich wäre. Die 1H-NMR-Spektren der Modellpeptide FA und FB zeigen die Protonen der Aminosäuren im erwarteten Bereich zwischen 0 ppm und 9 ppm. Aufgrund der paramagnetischen Eigenschaften der Probe ist die Auflösung der individuellen Linien jedoch gering. Es konnten weder für das Modell FA noch für das Modell FB paramagnetisch verschobene Signale nachgewiesen werden, wobei ein Messbereich bis 70 ppm untersucht wurde. Das vollständige Fehlen der Signale in der näheren Umgebung der paramagnetischen Zentren führte zu der Annahme, dass die Signale zu stark verbreitert werden und daher nicht mehr detektiert werden können. Die NMR-Spektroskopie wird ebenfalls durch das Auftreten der kolloidalen Eisenpartikel erschwert, die bereits die Mössbauer-Spektroskopie bei angelegtem Magnetfeld verhindert haben und durch die Tatsache, dass die Peptide mit [4Fe4S]-Zentrum lediglich eine Minderheit der gesamten Probe darstellen. Bevor diese 129 Ergebnisse und Diskussion Methode zu dem gewünschten Ergebnis führen kann, muss zunächst der Einbau der FeS-Zentren optimiert und anschließend die unspezifisch gebundenen Eisenionen aus der Probe entfernt werden. 3.2.9. Bindung an das Photosystem I Beide Modellpeptide sollten auch bezüglich ihrer Fähigkeit untersucht werden, an das Photosystem I, nach Entfernung der Untereinheiten C, D und E, zu binden. Zusammen mit den Untereinheiten werden die FeS-Zentren FA und FB des PS I entfernt, während das Zentrum FX, das von den Untereinheiten PsaA und PsaB gebunden wird, im PS I verbleibt. Während das Peptid FA drei Aminosäuren enthält, die direkt an der Bindung der Untereinheit C beteiligt sind, besitzt das Peptid FB keine Aminosäure, die an dieser Wechselwirkung teilnimmt. Daher wird für FA eine höhere Wahrscheinlichkeit der Bindung an das PS I erwartet. 3.2.9.1. Optische Experimente Die Kinetik der Ladungsrekombination nach Anregung durch einen Laserblitz ist für das Photosystem I bereits in früheren Studien untersucht worden[113] (vgl. Abschnitt 5.1.11)[114]. Dabei zeigte sich ein deutlicher Unterschied in der Kinetik der Rückreaktion zwischen nativem PS I und modifiziertem PS I, bei dem die Untereinheiten C, D und E entfernt wurden, weil ohne diese Untereinheiten die terminalen Elektronenakzeptoren fehlen (Abbildung 66). Wenn eines der Modellpeptide in der Nähe des FX-Zentrums bindet und dadurch am Elektronentransfer beteiligt ist, ist eine Änderung der Rückreaktionskinetik zu erwarten. In diesem Fall läge im Anschluss an das Zentrum FX ein weiterer Elektronenakzeptor vor, der in der Kinetik der Ladungsrekombination eine Änderung verursachen würde. Die Experimente zur Bestimmung der Kinetik der Ladungsrekombination wurden zunächst in Gegenwart eines Überschusses je eines der Peptidmodelle durchgeführt. Nachdem eine Änderung der Kinetik beobachtet werden konnte, wurde das überschüssige Peptide durch wiederholte Verdünnung und Konzentration der Probe über einen Mikrokonzentrator 130 Ergebnisse und Diskussion (Membrangröße 100 kDa) entfernt. Bei der anschließenden erneuten Messung wurde eine identische Kinetik beobachtet. Abbildung 66: Arbeitsmodell zum Verständnis der Ladungsrekombinationskinetik im PS I in Gegenwart der Modellpeptide (Mp) basierend auf den Daten des nativen PS I (modifiziert nach Golbeck et al.[93]). Die Wege der Ladungsrekombination im PS I ohne seine stromalen Untereinheiten C, D und E sind in schwarz dargestellt und diejenigen im nativen PS I in rot. Der ideale Fall der Beteiligung der Modellpeptide am Elektronentransfer ist in grün dargestellt. Da vom PS I in Gegenwart der Peptidmodelle keine Kristallstruktur untersucht wurde, wurde das Energieniveau (Mp) an einem logisch sinnvollen Punkt entlang der Abszisse positioniert. Die Kinetik der Rückreaktion im PS I zeigt eine Verlangsamung in Gegenwart eines Überschusses von jeweils einem Modellpeptid im Vergleich zur Referenz (Abbildung 67). Dieses Ergebnis kann dahingehend interpretiert werden, dass die Modelle am Elektronentransfer beteiligt sind. Zusätzlich bleibt der Einfluss der Modellpeptide trotz Aufreinigung durch wiederholtes verdünnen 131 und aufkonzentrieren der Probe Ergebnisse und Diskussion (100 kDa Membrangröße) erhalten. Diese Beobachtung weist auf eine stabile Bindung bzw. einen irreversiblen Einfluss der Modellpeptide hin. Die einzige Position für eine solche Beteiligung am Elektronentransfer ist die Bindungsstelle der Untereinheit PsaC, da die restlichen Kofaktoren der Elektronentransportkette im Inneren des Proteins liegen und daher nicht für die Modellpeptide zugänglich sind. Abbildung 67: Blitzlicht-induzierte transiente Absorptionsänderung von P700 in PS I zur Bestimmung der Kinetik der Ladungsrekombination. Das Photosystem I ohne die Untereinheiten C, D und E (schwarz) zeigt eine schnellere Kinetik im Vergleich zu Messungen in Gegenwart der Modellpeptide FA (blau) und FB (grün). 3.2.9.2. EPR-Experimente Wie die optische Untersuchung der Elektronentransferkinetik gezeigt hat, sind beide Peptide in der Lage, mit dem Photosystem I, dem die Untereinheiten C, D und E fehlen, zu interagieren und dadurch den Elektronentransfer zu beeinflussen Wenn die Modelle am Elektronentransfer beteiligt sind, müsste im EPR-Spektrum bei Belichtung der Probe das Signal des reduzierten [4Fe4S]1+-Zentrums auftreten. Wird das modifizierte Photosystem I in Gegenwart eines der Modellpeptide bei 10 K belichtet, wird kein Signal eines reduzierten FeS-Zentrums beobachtet. Das Kontrollexperiment mit nativem Photosystem I inklusive der Untereinheiten C, D und E zeigt nach der Belichtung bei 10 K das erwartete Interaktionsspektrum der beiden 132 Ergebnisse und Diskussion reduzierten Eisen-Schwefel-Zentren mit g-Werten von 2,056, 1,949 und 1,899 für das Zentrum FA und 2,056, 1,934 und 1,899 für das Zentrum FB[115]. Da die im vorangegangenen Kapitel beschriebenen optischen Messungen bei Raumtemperatur durchgeführt wurden, konnte zu diesem Zeitpunkt nicht ausgeschlossen werden, dass der Elektronentransfer zu den Modellpeptiden durch die tiefe Temperatur beeinflusst wird. Um diese Möglichkeit auszuschließen, wurde das Experiment wiederholt und die Proben unter kontinuierlicher Belichtung eingefroren, so dass der Elektronentransfer bei Raumtemperatur stattfinden konnte, aber eine Ladungsrekombination durch die tiefe Temperatur verhindert wird. Auch in diesem Fall konnte für das vollständige Photosystem I das erwartete Spektrum beobachtet werden, während die Proben, die die Modellpeptide enthielten, keine Signale eines reduzierten [4Fe4S]-Zentrums aufwiesen. Mit Hilfe der EPR-Spektroskopie konnte kein Nachweis für eine Bindung der Peptide an das Photosystem I ohne seine stromalen Untereinheiten C, D und E erbracht werden. Allerdings stellt diese Beobachtung nicht unbedingt einen Widerspruch zu den optischen Messungen dar. Der Einfluss der Ladungsrekombination, der durch die optischen Messungen detektiert wurde, muss nicht unbedingt durch eine direkte Beteiligung des Modellpeptides im Rahmen einer Reduktion stattfinden. Durch eine Bindung an das Photosystem könnte die Geometrie in der Umgebung des FX-Zentrums verändert werden, sodass eine veränderte Kinetik der Ladungsrekombination resultiert. Allerdings gibt es im Augenblick keine Daten, die die Wechselwirkung der Modellpeptide mit dem Photosystem näher beschreiben oder eine fundierte Interpretation zulassen. 133 Ergebnisse und Diskussion 3.2.10. Zusammenfassung In diesem Teil der Arbeit konnten vier unterschiedliche Peptide mit jeweils 16 Aminosäuren Länge unter Verwendung der Fmoc-basierten SPPS dargestellt und charakterisiert werden, mit denen ein [4Fe4S]-Zentrum gebunden werden sollte. Als Vorbild für diese Modelle dienten die drei Eisen-Schwefel-Zentren FA, FB und FX des Photosystems I. Eines der Peptide (FX1) war nicht in der Lage den gewünschten Kofaktor einzubauen, während ein weiteres Peptid (FX2) den gewünschten Kofaktor nicht spezifisch genug eingebaut hat. Hierbei wurde mindestens eine weitere paramagnetische Spezies gebildet, die nicht detailliert charakterisiert werden konnte. Die beiden synthetischen Peptide FA und FB binden den gewünschten Kofaktor. Durch EPR- und MössbauerSpektroskopie konnte die Identität der Kofaktoren in beiden Redoxzuständen [4Fe4S]2+/1+ zweifelsfrei nachgewiesen werden. Die für native FeS-Proteine charakteristische Merkmale, wie Wasserstoffbrückenbindungen und Elektronenspindichte auf den Sulfidionen des FeS-Zentrums, wurden durch ENDOR- und ESEEM-Spektroskopie nachgewiesen. Obwohl eine detaillierte Analyse der ENDOR-Spektren bezüglich der Bindungswinkel der Cystein β-CH2-Gruppen und der Spindichte auf den Sulfidionen des FeS-Zentrums nicht möglich war, zeigen sie bereits deutliche Unterschiede der Modelle zum nativen Vorbild auf. Die Spindichte des ungepaarten Elektrons auf den Sulfidionen ist in den Modellen wahrscheinlich deutlich geringer als in den Zentren des nativen Photosystems I. Die Modelle besitzen Redoxpotentiale von –490 mV (FA) und –470 mV (FB), die im Rahmen des Fehlers identisch sind. Trotz ihrer geringen Größe weisen die beiden Peptide FA und FB damit das negativste Redoxpotential aller bisher dargestellten Peptid-basierten Modelle auf und liegen verhältnismäßig dicht an den Werten ihrer nativen Vorbilder. Im Vergleich zu den Zentren FA und FB in der Untereinheit PsaC des Photosystems I zeigen die experimentell ermittelten Daten des Peptidmodells FB generell eine bessere Übereinstimmung. Sowohl im Redoxpotential, als auch bei den magnetischen Eigenschaften zeigt es eine deutlich bessere Übereinstimmung mit dem nativen Vorbild als das Peptidmodell FA. Letzteres weist deutlich kürzere Relaxationszeiten in der EPRspektroskopischen Untersuchung auf, die möglicherweise auf die Bildung von Aggregaten zurückzuführen ist. 134 Ergebnisse und Diskussion 3.2.11. Ausblick Bei der Fortführung dieses Projektes steht die Analyse, wie das Peptid die Eigenschaften, insbesondere das Redoxpotential, des Kofaktors beeinflusst, klar im Vordergrund. Basierend auf der Analyse von 510 natürlichen Eisen-Schwefel Proteinen[95] kann der Einfluss der einzelnen Aminosäuren auf das Redoxpotential untersucht werden. Dabei sind die Aminosäuren zwischen dem ersten und dritten Cystein der Bindungssequenz von besonderem Interesse. Die in dieser Arbeit ausführlich charakterisierten Peptidmodelle FA und FB unterscheiden sich in den Positionen 8 und 9 ihrer Aminosäuresequenz direkt nach dem zweiten konservierten Cystein deutlich von der Mehrheit der natürlichen Proteine. In den nativen Sequenzen wird in der Position 8 zu 47 % ein Glycin und in der Position 9 zu 18 % ein Alanin gefunden. Während dies die häufigsten Aminosäuren in diesen Positionen sind, werden an diesen Stellen im Modell FA die Aminosäuren Lysin und Arginin bzw. in FB Threonin und Glutamin eingesetzt, weil sie in der nativen Bindungssequenz der jeweiligen Zentren auftreten. Der Einfluss dieser polaren bzw. geladenen (Arg) Aminosäuren auf das Redoxpotential des FeS-Zentrums müsste nachweisbar sein, insbesondere, weil ein recht ähnliches Modell, das FdM-Pa, welches ebenfalls aus 16 Aminosäuren besteht, ein Redoxpotential von –350 mV aufweist[94]. Die Änderung des Redoxpotentials von etwa 100 mV durch den Austausch einer einzelnen polaren gegen eine unpolare Aminosäure würde eindeutig die bisher gängigen Modelle des Einflusses der Peptidkette auf diese Klasse der Kofaktoren unterstützen. In einem zweiten Ansatz kann die Sequenz insgesamt überarbeitet werden, um die geringe Effizienz der Rekonstitution zu verbessern. Da von den Modellpeptiden keine strukturellen Informationen erhalten werden konnten und eine Kristallisation von synthetischen Peptiden häufig schwieriger ist als im Fall von nativen Proteinen, wäre aufgrund der geringen Länge des Peptides ein kombinatorischer Ansatz denkbar. Von den 16 Aminosäuren sind die vier Positionen der Cysteinliganden festgelegt. Außerdem haben frühere Studien ergeben, dass hinter dem ersten Cystein eine Aminosäure mit sterisch aufwändiger Seitenkette positioniert sein sollte, z. B. Isoleucin oder Valin, und das gesamte Bindungsmotiv durch ein Prolin terminiert wird. Die Aminosäure Tryptophan in Position 16 dient als spektroskopische Sonde und ist daher ebenfalls von Veränderungen der Peptidkette ausgenommen. Von den restlichen 9 Positionen könnte zunächst der 135 Ergebnisse und Diskussion künstlich entworfene Mini-„loop“ beibehalten werden, wodurch nur noch sechs variable Positionen bleiben. Als „screening“-Parameter wäre in diesem Fall das durch UV-Vis-Spektroskopie bestimmte Bandenverhältnis der Absorptionen bei 280 nm und 400 nm. Auf diese Weise kann in kurzer Zeit eine optimale Effizienz der Rekonstitution erreicht werden, allerdings würden die in dieser Arbeit beschriebenen Eigenschaften der Modelle bei dieser Vorgehensweise verloren gehen. 136 Zusammenfassung 4. Zusammenfassung Die Charakterisierung der nativen Photosysteme ist aufgrund der Zahl der in ihnen enthaltenen Kofaktoren äußerst schwierig. Jeder einzelne Kofaktor wird durch seine direkte Proteinumgebung derart beeinflusst, dass er seine Aufgabe im Gesamtkonzept optimal ausführen kann. Um ein derart komplexes System untersuchen und verstehen zu können, müssen zunächst die Bausteine, aus denen es besteht, charakterisiert und verstanden werden. Dies ist nur möglich, wenn kleine und einfach strukturierte Modellsysteme zur Verfügung stehen, die vollständig charakterisiert und umfassend variiert werden können. Der lichtangeregte Triplettzustand von Chlorophyllen und Phäophytinen senkt durch seine lange Lebensdauer und seine hohe Reaktivität die Effizienz der photosynthetischen Lichtreaktion und kann zu irreparablen Schäden in den Reaktionszentren führen. Aus diesem Grund muss er in nativen Systemen effizient und schnell gelöscht werden, was durch die Carotinoide in den Photoreaktionszentren erreicht wird. Als spektroskopische Sonde ist er dagegen unverzichtbar, da er die Möglichkeit bietet, direkt die Molekülorbitale des lichtangeregten Singulettzustands (HOMO und LUMO) zu untersuchen. Die wichtigste spektroskopische Methode, um lichtangeregte Triplettzustände zu charakterisieren, ist die EPR-Spektroskopie und die darauf aufbauende ENDOR-Spektroskopie. Ein Ziel dieser Arbeit war die EPR- und ENDOR-spektroskopische Charakterisierung von monomeren Chlorophyll a Derivaten und Zink-Protoporphyrin. Metall-Protoporphyrine sind in nativen Systemen zwar nicht an Photoreaktionen beteiligt, zeichnen sich jedoch durch eine hohe Triplettausbeute nach Lichtanregung aus und dienen in dieser Arbeit stellvertretend für Porphyrine als Testmoleküle. Weil Chlorophylle und deren Derivate in organischen Lösemitteln zur Aggregation neigen und in wässrigen Lösungen nahezu vollständig unlöslich sind, wurde auf eine Proteinbasierte Matrix zur Erzeugung von monomeren Chlorophyllderivaten in Lösung zurückgegriffen. Auf diese Weise konnten die Komplexe von ZnPPIX, ZnPheid a, ZnPPheid a, ZnMePPheid a und ZnMePPheid a mit Apo-Myoglobin dargestellt werden. 137 Zusammenfassung Die Komplexe wurden durch die UV-Vis-Spektroskopie charakterisiert, wobei alle Kofaktoren nach Einbau in die Proteinmatrix eine Verschiebung der Absorptionsbanden in den langwelligen Bereich zeigten. Um die Integrität der Proteinkomplexe zu überprüfen, wurde ihre Stabilität gegenüber der Entfaltung untersucht. Die Tatsache, dass eine Stabilisierung der Proteinstruktur gegenüber der Apo-Form nur bei einigen Kofaktoren gefunden werden konnte, wurde als ein Homogenitäts- bzw. Heterogenitätskriterium verwendet. Diese Interpretation wurde durch die NMR-Spektroskopie unterstützt, mit der die Homogenität der Proteinkomplexe anhand von zwei Signalen untersucht werden konnte, die bei negativen ppm-Werten im Spektrum auftraten und der Aminosäure Valin 68 zugeordnet wurden. Nach der vollständigen Charakterisierung der Proteinkomplexe wurden die Triplettzustände nach Lichtanregung durch die EPR- und ENDORSpektroskopie untersucht. Die Nullfeldparameter, die durch die EPR-spektroskopischen Messungen erhalten wurden, zeigten eine gute Übereinstimmung mit nativen Systemen. Die ENDOR-Spektren konnten durch die Ergebnisse quantenchemischer Rechnungen interpretiert werden. Hierbei konnten erstmals die vier Methinprotonen und alle Methylgruppen des Ringsystems zugeordnet werden. Der Vergleich der freien Kofaktoren und denjenigen, die durch das Protein komplexiert vorlagen, zeigte, dass die Hyperfeinkopplungskonstanten der Methinprotonen und einiger Methylgruppen durch die Komplexierung zunahmen, während die Kopplungskonstanten der restlichen Substituenten nahezu unverändert blieben. Durch die Verwendung unterschiedlicher Kofaktoren konnte ebenfalls festgestellt werden, dass eine unveresterte Propionsäure als Substituent des Kofaktors das Bindungsverhalten verbessert, während ein Methylester negative Auswirkungen hat. Mit den veresterten Kofaktoren konnten keine stöchiometrischen Komplexe dargestellt werden. Die in dieser Arbeit vorgestellten Ergebnisse zeigen das Potential der dargestellten Modellkomplexe, monomere Kofaktoren abhängig von der Proteinumgebung vollständig zu charakterisieren. Durch Variationen in der Proteinumgebung kann der Einfluss des Proteins auf die Eigenschaften des Kofaktors untersucht werden. Insbesondere die Verteilung der Elektronenspindichte nach Lichtanregung auf dem Molekülgerüst kann auf diese Weise detailliert untersucht werden. Ferner besteht die Möglichkeit, die hier durchgeführten Untersuchungen auf eine Vielzahl photoaktiver, biologisch relevanter 138 Zusammenfassung Kofaktoren auszudehnen und eine wichtige Interpretationshilfe für die Prozesse in den photosynthetisch aktiven Proteinen bereitzustellen. Neben den Chlorophyllen und ihren metallfreien Basen, den Phäophytinen, sind [4Fe4S]-Zentren als terminale Elektronenakzeptoren ein wichtiger Bestandteil des Photosystems I. Daher wurden in einem zweiten Teil dieser Arbeit Modellpeptide für diese Zentren entworfen. Die Modellpeptide wurden unter Verwendung der Fmoc-Peptidsynthese mit 16 % Ausbeute dargestellt und durch HPLC-Chromatographie und MALDI-TOF- Massenspektrometrie charakterisiert. In den wichtigsten spektroskopischen Eigenschaften entsprechen die Modelle von FA und FB ihren Vorbildern aus der Untereinheit PsaC des Photosystems I. Nach dem Einbau der FeS-Zentren zeigen die Modellpeptide ein für [4Fe4S]-Zentren charakteristisches UV-Vis-Spektrum mit einer Absorption bei 400 nm, die durch einen charge-transfer von den Sulfid- zu den Eisenionen verursacht wird. Diese Absorptionsbande nimmt durch Reduktion des durch das jeweilige Modellpeptid gebundene FeS-Zentrum um etwa 50 % ab, wie es für ein solches System erwartet wird. Bei der EPR-spektroskopischen Charakterisierung im reduzierten Zustand weisen die Modellpeptide nur geringe Abweichungen von den FeS-Zentren FA und FB des PS I auf. Die g-Werte von 2,04, 1,93, und 1,90 stimmen mit den Daten des nativen Systems überein. Im oxidierten Zustand sind die [4Fe4S]2+-Zentren EPR-inaktiv und können durch diese Methode nicht charakterisiert werden. Diese Lücke konnte durch Verwendung der Mössbauer-Spektroskopie geschlossen werden. Sie bietet die Möglichkeit, verschiedene FeS-Zentren in ihren unterschiedlichen Redoxzuständen zu unterscheiden, wodurch die Peptid-gebundenen [4Fe4S]-Zentren unabhängig von anderen Methoden in beiden Redoxzuständen charakterisiert werden konnten. Durch die ENDOR-Spektroskopie konnte gezeigt werden, dass die Spindichteverteilung innerhalb des FeS-Zentrums von derjenigen der nativen Systeme abweicht, wodurch kleinere Hyperfeinkopplungskonstanten resultierten. Durch die gleiche Methode konnten erste Hinweise auf Wasserstoffbrückenbindungen zwischen dem FeS-Zentrum und Amidprotonen der Polypeptidkette erhalten werden. Diese Ergebnisse konnten durch den Vergleich der ESEEM-Spektren des nativen Systems mit denen der beiden Modellpeptide bestätigt werden. Das Redoxpotential, die wichtigste Eigenschaft für einen Kofaktor des Elektronentransfers, liegt in einem vergleichbaren Bereich, wie er für das native System 139 Zusammenfassung beschrieben ist. Damit wurden in dieser Arbeit Modellpeptide mit dem bisher negativsten Redoxpotential (-470 mV) hergestellt. Trotz ihrer geringen Größe von lediglich 16 Aminosäuren sind die Modellpeptide in der Lage, das FeS-Zentrum gut gegen das umgebende Lösemittel abzuschirmen, so dass ein derart negatives Redoxpotential ermöglicht wird. Da andere Modellsysteme mit ähnlichen Peptidlängen ein Redoxpotential etwa 100 mV positiver aufwiesen, muss dieser Parameter signifikant durch die Polypeptidkette beeinflusst werden. Ein kurzes synthetisches Peptid bietet die beste Möglichkeit, diesen Einfluss zu studieren. Der wesentliche Unterschied der hier dargestellten Modelle zu früheren Ansätzen liegt in den Positionen 8 und 9 der Peptidsequenz. In dieser Arbeit treten polare bzw. geladene Aminosäuren an diesen Positionen in der Sequenz auf, während in früheren Studien unpolare Aminosäuren mit kleinen Seitenketten verwendet wurden. Neben Modellen für die FeS-Zentren FA und FB wurden auch Modellpeptide für das Zentrum FX dargestellt. Sie waren jedoch nicht in der Lage, ausschließlich das gewünschte [4Fe4S]-Zentrum zu binden. Das FX-Zentrum, das durch die Untereinheiten PsaA und PsaB im Photosystem I gebunden wird, ist äußerst schwierig zu modellieren, da zwei flexible „loops“ ohne Sekundärstrukturelemente im PS I die Liganden bereitstellen. Eine solche Struktur kann ohne externe Vorgabe der Konformation wahrscheinlich nicht durch ein kurzes Peptid modelliert werden. Die dargestellten Strukturen sind zu flexibel, wodurch der Entropieverlust des Systems, der durch Bindung eines FeS-Zentrums entsteht, nicht durch die Bindungsenthalpie des Zentrums aufgewogen werden kann. Ein weiteres Ziel der Arbeit war es, zu untersuchen, inwieweit eine Wechselwirkung der Modellpeptide mit dem PS I zu beobachten ist, bzw. ob die Modelle in der Lage sind, die Funktion der natürlichen FeS-Zentren zu übernehmen. Die absorptionsspektroskopische Analyse der Ladungsrekombinationskinetik nach Lichtanregung im PS I zeigt eine Verlangsamung der Kinetik in Gegenwart der Modellpeptide. Überraschenderweise zeigen beide Modellpeptide einen ähnlich starken Einfluss auf die Kinetik, was als Wechselwirkung mit dem PS I interpretiert werden muss. Allerdings weist nur das Modellpeptid FA drei Aminosäuren auf, die an der Bindung der nativen Untereinheit PsaC zum PS I-Kern, bestehend aus den Untereinheiten PsaA und PsaB, beteiligt sind. Eine Wechselwirkung dieses Peptides mit dem PS I-Kern wurde erwartet und experimentell gefunden. Das Modell FB besitzt keine Aminosäure, die im 140 Zusammenfassung natürlichen PS I an der Bindung der Untereinheit PsaC beteiligt ist. Trotzdem zeigt dieses Modell den gleichen Effekt wie das Modell FA. Welche Ursache der Einfluss der Modellpeptide auf die Kinetik der Ladungsrekombination hat, konnte nicht abschließend geklärt werden. Eine Reduktion der Modelle durch Aufnahme eines Elektrons vom FXZentrum aus dem PS I-Kern ist die wahrscheinlichste Art der Interaktion. Diese konnte jedoch durch die EPR-Spektroskopie nicht nachgewiesen werden. Eine rein statische Wechselwirkung, die die Eigenschaften des Zentrums FX ändert, ist zwar ebenfalls möglich, ist aber experimentell äußerst schwer nachzuweisen. Die Untersuchung der Modellpeptide durch die NMR-Spektroskopie, um strukturelle Informationen zu erhalten, erbrachte keine auswertbaren Ergebnisse. In keinem Spektrum konnten paramagnetisch verschobene Signale von Aminosäuren beobachtet werden. Die Untereinheit PsaC dagegen konnte in früheren Arbeiten im isolierten Zustand und gebunden an das PS I vollständig durch die NMR-Spektroskopie charakterisiert werden. Um durch die NMR-Spektroskopie Aufschluss über die Struktur der Modellpeptide in Lösung zu erhalten, muss daher zunächst die Einbaueffizienz der FeS-Zentren in die Modellpeptide erhöht werden. Die in dieser Arbeit dargestellten Modellpeptide bieten das Potential zu einem besseren Verständnis, wie die physikalischen Eigenschaften eines Eisen-Schwefel-Zentrums durch die Proteinumgebung beeinflusst werden, beizutragen. Die Peptidsequenzen können schnell dargestellt und einfach in einer beliebigen Position variiert werden. Nach der Rekonstitution können sie durch verschiedene spektroskopische Methoden bezüglich der Einbaueffizienz und ihrer magnetischen Eigenschaften charakterisiert werden, wodurch Rückschlüsse auf den Einfluss der veränderten Peptidsequenz gezogen werden können. 141 Zusammenfassung 142 Experimenteller Teil 5. Experimenteller Teil 5.1. Geräte und Methoden 5.1.1. NMR-Spektroskopie Die NMR-Spektren wurden nach dem Fast-Fourier-Transform (FFT)-Verfahren mit den Spektrometern DRX-500 und DRX-400 der Firma Bruker aufgenommen. Die chemischen Verschiebungen werden in Bezug auf das verwendete deuterierte Lösemittel angegeben. Bei in Wasser gelösten Proben erfolgte die Lösemittelunterdrückung durch das Verfahren der „Presaturation“, bei dem die Resonanzlinie des Lösemittels Wasser vor jedem Messzyklus gesättigt wird. Die Signalmultiplizitäten werden durch die Abkürzungen s (Singulett), d (Dublett), t (Triplett), q (Quartett) und m (Multiplett) angegeben. Die Kopplungskonstanten J werden in Hertz (Hz) angegeben. Die Zuordnung der Signale erfolgt durch Vergleich mit Literaturdaten. 5.1.2. UV-Vis-Spektroskopie Die UV-Vis-Absorptionsspektren wurden entweder mit einem ATI Unicam UV2-300 Spektrometer oder einem Shimadzu UV-2401PC Spektrometer aufgenommen. Es werden die Wellenlängen der Absorptionsmaxima, sowie deren relative Intensität angegeben. Alle Spektren wurden in Quarzküvetten mit einer Schichtdicke von 1 cm gegen eine Referenzküvette mit Referenzlösemittel gemessen. Banden, die aufgrund von Überlagerungen mit anderen Absorptionsbanden kein ausgeprägtes Maximum aufweisen, werden mit sh (shoulder) gekennzeichnet. 143 Experimenteller Teil 5.1.3. Infrarot-Spektroskopie Die Infrarot-Spektren wurden in einem Bruker IFS 66 FT-IR-Spektrometer oder einem Perkin-Elmer 1600 Spektrometer in KBr als Matrix gemessen. Die Position der Banden wird in Wellenzahlen (cm-1) angegeben. Die Angabe beschränkt sich auf die stärksten Banden der jeweiligen funktionellen Gruppen. 5.1.4. Massenspektrometrie Die Massenspektren der Peptide und Proteine wurden durch Matrix-Assisted Laser Desorption Ionization Time of Flight (MALDI-TOF) Spektroskopie mit einer Voyager DE Pro Workstation in Verbindung mit einem Hochgeschwindigkeits-Digitalisierer der Firma LeCroy gemessen. Die Messgenauigkeit lag bei ±0,1 % des Molekulargewichtes. Für die synthetischen Peptide wurden 2,5-Dihydroxybenzoesäure, 3,5-Dimethoxy-4-hydroxyzimtsäure und α-cyano-4-hydroxyzimtsäure, für Myoglobin wurden zusätzlich 2,6-Dihydroxyacetophenon und 6-Aza-2-thiothymin als Matrices verwendet. Die Aufnahme der Spektren erfolgte bei einer Beschleunigungsspannung von 20000 V und einer Verzögerungszeit von 300 nsec. Die Elektronenionisationsspektren der dargestellten Phäophorbide wurden mit einem MAT 311A oder 8230 Spektrometer der Firma Finnigan mit einer Ionisierungsenergie von 70 eV aufgenommen. Zur Ionisierung kam die Elektrosprayionisation (ESI) zum Einsatz. Als Probenmatrix diente Dimethyloxybenzylalkohol oder m-Nitrobenzylalkohol. 5.1.5. Säulenchromatographie Die präparative Säulenchromatographie wurde in selbstgefüllten Schwerkraftsäulen verschiedener Größe auf Silikagel 60 der Firma Merck (Korngröße 63 – 40 µm) als stationäre Phase durchgeführt. Des weiteren kamen Sepakron-FPGC Säulen der Firma Kronlab mit den Lösemittelpumpen Büchi 688 oder Besta E-100 zum Einsatz. Diese Säulen wurden mit einem Druck von 1 bis 10 bar betrieben. 144 Experimenteller Teil 5.1.6. Hochdruck-Flüssigkeitschromatographie (HPLC) Die HPLC der synthetischen Peptide wurde auf einem HPLC-System der Firma GilsonAbimed durchgeführt unter Verwendung von zwei gekoppelten Gradientenpumpen (Typ 331 und 332) mit einem Detektor vom Typ UV/Vis-156. Dabei wurde eine Säule der Firma Vydac vom Typ 218TP1002 Protein & Peptide C18 (200 x 20 mm, 300 Å) verwendet. Es wurde ein Gradient mit 45 Minuten Länge eines Wasser/AcetonitrilGemisches verwendet, der Anteil an Acetonitril wurde während dieser Zeit von 20 % auf 70 % erhöht. Alle Lösemittel enthielten 0,1 % TFA und wurden entgast. Präparative HPLC Trennungen der Phäophorbide wurden mit einer Pumpe der Firma Gilson-Abimed an einer Nucleosil C-18 Umkehrphasensäule der Firma Macherey & Nagel (250 x 20 mm) als stationärer Phase unter Verwendung eines Detektors der Firma Shimadzu SPD10 AV (VP) durchgeführt. Die mobilen Phasen sind jeweils bei den Verbindungen angegeben. 5.1.7. Fluoreszenzspektroskopie Die Fluoreszenzspektroskopie für die Entfaltungsstudien wurde auf einem Cary Eclipse Fluoreszenzspektrometer der Firma Varian durchgeführt. Typischerweise wurde bei 290 nm die Fluoreszenz der Aminosäure Tryptophan angeregt und diese im Bereich von 300 nm bis 450 nm detektiert. Zur Ermittlung der Wellenlänge des Fluoreszenzmaximums wurden die Messkurven durch ein Polynom 9. Grades interpoliert. Die Probenkonzentration lag zwischen 30µM und 300 µM. 5.1.8. Circular Dichroismus- (CD) Spektroskopie Für die CD-Spektroskopie kam ein Jasco J-715 Spektrometer zum Einsatz. Das Spektrometer wurde vor jeder Messung mit 60 mg (1R)-(-)-Campher-10-sulfonsäure Ammoniumsalz in 100 ml H20 auf einen Wert von θ291=-181,6 mdeg (d=10 mm) kalibriert. 145 Experimenteller Teil Die Spektren wurden im Bereich von 180 nm bis 240 nm in Quarzküvetten mit einer Schichtdicke von 0,1 mm aufgenommen. 5.1.9. Anaerobe Arbeiten und elektrochemische Titration Alle anaeroben Arbeiten wurden in einer Glovebox der Firma Coy Laboratory Products Inc. (Modell A) mit einer Stickstoff- / Wasserstoffatmosphäre 95/5 (Vol-%) durchgeführt. Noch vorhandener Rest-Sauerstoff wurde auf zwei Palladiumkatalysatoren mit Wasserstoff zu Wasser umgesetzt. Der Sauerstoffgehalt wurde kontinuierlich über einen GasanalyseGerät der Firma Coy, Modell 10, kontrolliert und lag unter 1 ppm. Die elektrochemische Titration wurde in einer Quarzküvette mit einer Referenzelektrode (Ag/AgCl, 1 M AgCl) und einer Arbeitselektrode (Glascarbon) durchgeführt. Das Potential der Referenzelektrode wurde nach jeder Anwendung durch ein Zyklovoltammogramm mit Methylviologen (-449 mV gegen NHE) in 50 mM Phosphatpuffer, pH 7,0, kontrolliert. Ein pH-Meter wurde als Anzeigegerät verwendet. Das Potential wurde durch Zugabe einer Dithionitlösung in kleinen Mengen (< 5µL) eingestellt. 5.1.10. Elektron-Paramagnetische Resonanz Spektroskopie (EPR) Die X-Band Triplett-EPR-Spektren der untersuchten Phäophorbide wurden auf einem Bruker Elexsys 580 Super XFT Spektrometer oder einem ESP 380-E FT-EPR Spektrometer mit einem dielektrischen Saphirring-Resonator ER4118X-MD-5W1 (EPR) oder einem EN4118X-MD5-W1 (ENDOR) Resonator aufgenommen. Die Probe wurde durch einen Heliumkryostat Model CF935 mit einer Temperatursteuerung ITC4 der Firma Oxford gekühlt. Für die ENDOR Spektroskopie kam ein RF-Verstärker Typ ENI A 500 zum Einsatz. Die Proben wurden mit einem OPO VisIR 100 Laser von GWU belichtet, der seinerseits von einem Nd:YAG Laser Typ GCR 130 der Firma Spectra Physics betrieben wurde. 146 Experimenteller Teil Die Q-Band Triplett-EPR Spektren wurden auf einem Bruker Elexsys 580 Super QFT Spektrometer mit zylindrischem Eigenbau-Resonator TE011 durchgeführt. Um die Probe belichten zu können war der Resonator mit Schlitzen versehen. Die Probe wurde durch einen Heliumkryostat Model CF935 mit einer Temperatursteuerung ITC4 der Firma Oxford gekühlt. Die Aufzeichnung der transienten EPR-Spektren erfolgte durch das SpecJet-Modul (Oszilloskop) des EPR-Spektrometers. Entlang der Zeitachse wurden 2048 Punkte mit 4 ns Abstand aufgenommen, während entlang der Magnetfeldachse 512 Punkte bei einer Breite des Spektrums von 1200 Gauss aufgezeichnet wurden. Für jeden Wert des Magnetfeldes wurden zehn "Decay"-Kurven addiert. Typische Messbedingungen für die transienten EPR-Spektren: Laserleistung 10 mJ, Wellenlänge 585 nm, Blitzdauer 5 ns, Blitzfrequenz 10 Hz, Temperatur 10 K, Mikrowellenfrequenz 9,436 GHz, Mikrowellenleistung 100 µW bis 1 mW. Die X-Band CW-EPR Spektren der FeS-Peptide wurden auf einem Bruker E 500 Spektrometer mit einem Oxford ESR continuus flow Kryostaten, Model 910, mit einer Temperatursteuerung ITC 503 der Firma Oxford gemessen. Für die Belichtung der Proben des Photosystems I wurde das Licht eines Leica Pradovit Diaprojektors vom Typ P2002 mit einer 250 W Lampe auf die Probe innerhalb des Resonators fokussiert. Um thermische Einflüsse durch die Belichtung zu vermeiden kam ein Wasserfilter mit 3 cm Schichtdicke zum Einsatz. Die Proben wurden bei 15 K insgesamt 10 Minuten belichtet und dabei im Abstand von zwei Minuten um 90 Grad gedreht, um eine optimale Durchdringung der Probe zu erreichen. Typische Bedingungen für X-Band CW-EPR Spektren: Temperatur 10 K, Mikrowellenfrequenz 9,436 GHz, Mikrowellenleistung 10 mW, Modulationsfrequenz 100 kHz, Modulationsamplitude 10,0 G und Zeitkonstante 40 ms. Die Temperaturabhängigkeit der Signale wurde zwischen 5 K und 40 K bei 10 mW Mikrowellenleistung bestimmt. Die Sättigungskurven wurden bei 15 K mit Mikrowellenleistungen zwischen 25 µW und 200 mW aufgenommen. Die Leistungsabhängigkeit der EPR-Signale wurde nach Hales et al.[116] ausgewertet. Dabei kam Gleichung (30) zum Einsatz. 147 Experimenteller Teil b ⎞2 ⎛ P SI 1 ⎛ SI ⎞ und P1 / 2 = 2 = K ⋅ ⎜⎜ 1 / 2 ⎟⎟ mit K = ⎜ ⎟ g T1T2 P ⎝ P ⎠ max ⎝ P + P1 / 2 ⎠ (30) In dieser Gleichung stehen SI für die Amplitude der ersten Ableitung des EPR-Signals, P für die bei diesem Spektrum verwendete Leistung und P1/2 für die Leistung bei halber Sättigung. Der Exponent b nimmt für inhomogen verbreiterte Linien, wie sie in Metalloproteinen beobachtet werden, einen Wert von b = 1 und für homogen verbreiterte Linien einen Wert von b = 3 an. Die Proportionalitätskonstante K wird zur Normierung der Messdaten als Maximalwert des Quotienten der Signalamplitude durch die Wurzel der Mikrowellenleistung definiert. Zur Auswertung wird die normierte Signalintensität (SI·P-1/2·K-1) gegen den Logarithmus der Mikrowellenleistung aufgetragen und ein Wert für b = 1 angenommen. 5.1.11. Optische Spektroskopie am Photosystem I zur Untersuchung der Elektronentransferkinetik Die Messungen der Elektronentransferkinetik wurden in der Arbeitsgruppe von Prof. John H. Golbeck an der Pennsylvania State University durchgeführt. Der Aufbau der Geräte ist in der Literatur detailliert beschrieben[113]. Das Verfahren wird an dieser Stelle kurz beschrieben. Die transienten Absorptionsänderungen des Radikalkations P700+ bei 820 nm wurden im Bereich von Mikrosekunden bis zu einer Minute nach der Anregung aufgenommen. Der Messstrahl von 820 nm wurde durch einen DC 25 F Halbleiter Diodenlaser von Spindler und Hoyer mit einer Ausgangsleistung von 50 mW erzeugt. Um Unregelmäßigkeiten in der Laserleistung zu korrigieren, wurde der Laserstrahl in Proben- und Referenzstrahl geteilt und die Intensität beider Strahlen über Photodiodendetektoren in Echtzeit miteinander verglichen. Die Anregung der Proben erfolgte durch einen Nd-YAG Laser vom Modell DCR-11 der Firma Spectra-Physics bei 532 nm mit einer Leistung von 135 mJ pro Puls und einer Pulslänge von 10 ns. Die Datenaufzeichnung wurde durch eine Photodiode gestartet, die durch das Streulicht des Anregungslasers aktiviert wurde. Die beobachtete 148 Experimenteller Teil Absorptionsänderung jeder Probe wurde durch Bestimmung der Chlorophyllkonzentration jeder Probe nach der Messung normiert. 5.1.12. Durchführung der theoretischen Untersuchungen Die theoretischen Untersuchungen wurden von Dr. Sebastian Sinnecker durchgeführt. Das Verfahren wird im folgenden kurz beschrieben. Die Dichtefunktionaltheorie (DFT) wurde benutzt, um die experimentellen Studien durch quantenchemische Rechnungen zu begleiten[117]. Für die Studien wurden vereinfachte Modellsysteme gewählt, in denen der Phytylester an Position 17 durch die freie Säure ersetzt wurde. Alle Modellsysteme wurden vollständig geometrieoptimiert. Hierzu wurden das BP Dichtefunktional[118-120] und der SV(P) Basissatz eingesetzt[121]. Diese Rechnungen wurden mit Turbomole durchgeführt[122]. Die Hyperfeinkopplungen wurden mit dem Programm ORCA berechnet[123]. Hierbei wurde das B3LYP Hybridfunktional[124; 125] mit der EPR-II Basis kombiniert[126]. Da für Magnesium oder Zink keine EPR-II Basisfunktionen entwickelt wurden, wurde die TZVP Basis für diese Modelle verwendet[127]. Dieses Verfahren lieferte bereits in vergangenen Studien zu Hyperfeinkopplungen von Chlorophyllradikalen zuverlässige Ergebnisse[73; 74]. Dabei handelte es sich jedoch ausschließlich um Systeme mit einem einzelnen ungepaarten Elektron. Daher ergibt sich in dieser Arbeit die interessante Möglichkeit, etwas über die Genauigkeit der eingesetzten Methoden für organische Triplettradikale zu erfahren. 5.2. Synthese und Chemikalien Alle Chemikalien für die Peptidsynthese wurden von Iris Biotech bezogen und besaßen mindestens den Reinheitsgrad „zur Peptidsynthese“. Das PAL-PEG-PS Harz für die Peptidsynthese wurde von Applied Biosystems bezogen. Alle anderen Chemikalien (Roth, Fluka, Merck, Acros, Aldrich) lagen als „p.a.“ Ware vor. Eine Ausnahme ist Natriumdithionit (Aldrich), welches lediglich in der Reinheit „technisch“ bezogen wurde. Alle Lösemittel wurden vor Gebrauch destilliert, sofern sie in der Reinheit technisch 149 Experimenteller Teil geliefert wurden. Wasser wurde ausschließlich aus einer Millipore Wasseraufbereitungsanlage bezogen. Typische Werte für die Leitfähigkeit lagen bei 18,2 MΩ-1 cm-1. 5.2.1. Festphasen-Peptidsynthese (SPPS) Die Synthese der einzelnen Peptide wurde nach der Fmoc/t-Bu Methode mit einem Syntheseautomaten der Firma Advanced Chemtech Model 348Ω durchgeführt. In allen Synthesen wurde ein PAL-PEG-PS Harz als feste Phase verwendet, so dass der C-Terminus jedes Peptides als Carbonsäureamid vorlag. Der N-Terminus der FeS-Peptide war eine freie Aminofunktion, bei allen anderen dargestellten Peptiden wurde die N-terminale Aminogruppe acetyliert. Die Aminosäuren wurden mit folgenden Schutzgruppen eingesetzt. Thr, Asp, Glu, Tyr: tert-butyl; Gln, Cys: Trityl; Trp, Lys: Boc und Arginin wurde durch 2,2,4,6,7-pentamethyl-dihydrobenzofuran-5-sulfonyl (Pbf) geschützt. Die Aminosäuren Gly, Ile, Pro und Val wurden ohne Schutzgruppe eingesetzt. Alle Reagenzien wurden in N-Methyl-2-pyrrolidon (NMP) gelöst, außer Piperidin, das zum Entschützen des N-Terminus verwendet wurde und in DMF gelöst wurde. Die Reagenzien wurden im Verhältnis 1:5:5:10 (Harz:Aminosäure:Kopplungsreagenz:DIPEA) eingesetzt. Als Kopplungsreagenzien kamen sowohl TCTU, TBTU und PyBOP, sowohl einzeln wie auch in Kombination bei Doppelkopplungen, zum Einsatz. Nach jeder Kopplung und jeder Schutzgruppenabspaltung wurde das Harz sechs Mal mit DMF gewaschen. In einer typischen Synthese wurde zunächst das Harz (258 mg, 0,17 mmol/g) für 30 Minuten mit DMF benetzt und anschließend zwei Mal je 15 Minuten mit 25 % Piperidin versetzt, um die N-terminalen Schutzgruppen zu entfernen. In allen Kopplungszyklen wurden Doppelkopplungen mit einer Reaktionszeit von 30 Minuten pro Kopplung durchgeführt. Nach Beendigung der Synthese wurde das Harz vier Mal mit Dichlormethan, zwei Mal mit Methanol und wieder vier Mal mit Dichlormethan gewaschen. Das Harz wurde für 90 Minuten im Argonstrom getrocknet und die synthetisierten Peptide wurden mit einer Mischung aus TFA (36 ml), Thioanisol (2,1 ml) Ethandithiol (1,2 ml) und Anisol (0,9 ml) vom Harz abgespalten und entschützt. Die erhaltene Lösung wurde über Nacht bei 150 Experimenteller Teil –20°C gelagert, um eine vollständige Abspaltung der Schutzgruppen zu garantieren. Anschließend wurde die Abspaltlösung im Vakuum entfernt und die Peptide mit Ether/Pentan im Verhältnis 1:1 gefällt und sedimentiert. Der erhaltene Feststoff wurde viermal in Ether/Pentan resuspendiert und anschließend in Essigsäure/Wasser gelöst, lyophylisiert und mittels HPLC aufgereinigt. 5.2.2. Rekombinantes Pottwal-Myoglobin aus E. coli Das Plasmid für die Überexpression von rekombinantem Pottwal-Myoglobin wurde von Prof. Dr. S. Hirota von der Kyoto Pharmaceutical University, Kyoto, Japan, zur Verfügung gestellt. Die Anzucht der E. coli Zellen, die Isolierung und Aufreinigung des rekombinanten Pottwal-Myoglobins und verschiedene Versuche zur Kristallisation wurden von Herrn Dr. Hideaki Ogata durchgeführt. Der Einbau des Plasmids erfolgte durch die Transformationsmethode unter Verwendung des „BL21(DE3) Competent Cells“ Kits der Firma Merck. Für die Anzucht der Zellen[88; 89; 128; 129] wurde der E. coli-Stamm BL21DE3 mit dem wtMb/pET29b-Vektor verwendet. Das Medium bestand aus Trypton (16 g/L), Hefe Extrakt (10 g/L) und Kochsalz (5 g/L) mit einem pH-Wert von 7,3. Nach der Sterilisation wurden pro Liter Medium 1 mL des Antibiotikums Kanamycinmonosulfat (25mg/mL) zugegeben. Die Zellen wurden zunächst in einer Vorkultur (80 mL) über Nacht bei 37°C kultiviert. Um eine mögliche Kontamination des Mediums auszuschließen, wurde es während der Wachstumsphase der Vorkultur bei 37°C über Nacht gelagert. Es wurde nur Medium verwendet, dass keinerlei Anzeichen einer Kontamination (z.B. Trübung etc.) zeigte. Für die Hauptkultur wurden je 10 mL der Vorkultur auf jeweils 1,6 L Medium in einer Sterilbank überimpft und über Nacht bei 37°C geschüttelt. Der Rest der Vorkultur wurde auf Aga-Platten ausgestrichen. Nach vier bis fünf Stunden erreichte die Hauptkultur einen OD660-Wert zwischen 0,6 und 0,7 relativ zum Medium und die IPTG Induktion mit 320 µL einer 0,5 M IPTGStammlösung pro 1,6 L Medium wurde durchgeführt. Die Zellkultur wurde über Nacht bei 151 Experimenteller Teil 30°C inkubiert und anschließend sedimentiert. Aus acht Liter Medium wurden 30 g Nassgewicht Zellen erhalten. Zum Aufschließen wurden je 10 g Zellen 30 mL eines 20 mM TrisHCl-Puffers, pH 8,0, mit 80 mg Lysozym, 2,65 mg Dithiothreitol, 7,2 mg Protease Inhibitor (Pefabloc SC Plus Roche, ehem. BioMol), 0,25 mg DNase, 0,4 µL einer Stammlösung mit der Konzentration 1 mg/mL RNase A und 14,9 mg EDTA verwendet. Die Zellen wurden für 30 Minuten auf Eis in diesem Puffer resuspendiert, bei –80°C eingefroren und nach dem Auftauen erneut für 30 Minuten gerührt. Anschließend wurde die Suspension bei 40000 g für 20 Minuten in der Zentrifuge sedimentiert. Bei 4°C wurde der klare Überstand bis zu einer Endkonzentration von 55 % mit Ammoniumsulfat versetzt (14 g (NH4)2SO4 in 40 mL) und für 30 Minuten inkubiert. Die denaturierten Proteine wurden bei 40000 g innerhalb von 20 Minuten sedimentiert und der klare, rotbraune Überstand über Nacht gegen vier Liter 20 mM TrisHCl Puffer, pH 8,0, dialysiert. Die Aufreinigung des Myoglobins erfolgte über eine DEAE Sephacel AnionenaustauscherSäule, die mit 20 mM TrisHCl Puffer äquilibriert war. Auf dieser Säule bindet das Myoglobin im Gegensatz zu einigen Verunreinigungen nicht und eluiert als rotbraune Bande. Die erhaltene Fraktion wurde mit einer Spatelspitze Kaliumhexacyanoferrat(III), gelöst in 20 mM TrisHCl Puffer, pH 8,0, oxidiert und mit 10 %iger Essigsäure auf einen pH-Wert von 6,0 eingestellt. Die Lösung wurde auf eine CM Sephadex C25 Säule, die mit 20 mM Phosphatpuffer pH 6,0 äquilibriert war, aufgetragen und durch einen manuellen Stufengradienten mit 50 mM Phosphatpuffer, pH 8,0, eluiert. Abschließen wird die Proteinlösung über Ultrafiltrationsmembranen mit 10 kDa Ausschlussgröße aufkonzentriert. 5.2.3. Darstellung von Apo-Myoglobin Das kommerziell bezogene polykristalline Myoglobin wird in destilliertem Wasser gelöst (10 mg/mL) und unter Eiskühlung mit 1 M Salzsäure (66 µL/mL) auf pH 1-2 eingestellt, wobei eine Farbänderung von rot-braun nach braun zu beobachten war. Diese Lösung wurde drei Mal mit eiskaltem 2-Butanon extrahiert. Die farblose wässrige Phase wird zunächst für vier Stunden gegen NaHCO3 (50 mg/L) und 0,1 mM EDTA und anschließend 152 Experimenteller Teil über Nacht gegen destilliertes Wasser dialysiert. Die Proteinkonzentration wird durch UV/Vis Spektroskopie bestimmt (ε280=15,2 mM-1 cm-1). 5.2.4. Darstellung von Zink-Protoporphyrin (ZnPPIX) Unter Lichtausschluss werden 35 mg Protoporphyrin (PPIX) in 4 ml DMSO gelöst, mit 2 ml einer gesättigten Zinkacetatlösung in DMSO versetzt und über Nacht bei Raumtemperatur gerührt. Der Reaktionsfortschritt kann durch UV-Vis-Spektroskopie verfolgt werden, da das Edukt zwei Banden bei 629 nm und 509 nm aufweist, die bei vollständiger Metallierung nicht mehr auftreten. Nach beendeter Reaktion wird die Reaktionsmischung mit Ether und Wasser ausgeschüttelt. Die organische Phase wird über Natriumsulfat getrocknet und im Vakuum entfernt. Man erhält 35,5 mg (92 %) ZnPPIX als rotbraunen Feststoff. UV/Vis (DMSO): 586 nm (0,19), 547 nm (0,20), 423 nm (1,00) 1 H-NMR (400MHz, DMSO): 10,26, 10,18, 10,16, 10,12 (je s, 1H, 5-, 10-, 15-, 20-H), 8,53, 8,50 (je dd, 1H, J=11,6Hz, J=17,8Hz, X von ABX 31-CH, 81-CH), 6,41, 6,40 (je dd, 1H, J=1,7Hz, J=17,9Hz, A von ABX 31-CH, 81-CH), 6,15, 6,14 (je dd, 1H, J=1,7Hz, J=11,4Hz, B von ABX 31-CH, 81-CH), 3,75, 3,74, 3,63, 3,60 (je s, 3H, 2-, 7-, 12-, 18CH3), 2,7-2,3 (m, 8H, 131-, 132-, 171-, 172-CH2) MS (ESI): berechnet: 667 g/mol, gefunden: 668 [667+H], 690 [667+Na], 706 [667+K] 153 Experimenteller Teil 5.3. Darstellung der (Metall-)Phäophorbide Alle Arbeiten mit Chlorophyllen und deren Derivaten wurden unter Grünlicht durchgeführt. Die Lagerung der Produkte erfolgte ohne Lösemittel bei –80 °C. 5.3.1. Allgemeine Arbeitsvorschriften 5.3.1.1. Standardaufarbeitung Das Reaktionsgemisch wurde mit Ether und Wasser versetzt, bis eine klare Lösung entstand. Die Phasen wurden separiert und die organische Phase dreimal mit Wasser gewaschen. Nach Trocknung über Natriumsulfat wurde das Lösemittel unter Vakuum entfernt. 5.3.1.2. Abspaltung des Phytylesters Das entsprechende Phäophytin wurde mit TFA versetzt (1 ml/ 10 mg Pigment) und 10 bis 15 Minuten bei Raumtemperatur gerührt. Nach Entfernung der TFA im Vakuum konnte der Rückstand ohne Reinigung für die Metallierungsreaktion eingesetzt werden. 5.3.1.3. Metallierung der Phäophorbide Zu dem zu metallierenden Phäophorbid wurde je ein 200facher Überschuss an Zinkacetat und an Natriumacetat gegeben. Bei Pigmenten, die in der 132-Position die Methoxycarbonylgruppe besitzen, wird außerdem eine Spatelspitze Natriumascorbat zugegeben, um die Racemisierung der 132-Postion zu reduzieren. Als Lösemittel wurde Eisessig (10 ml/ 15 mg Pigment) verwendet und die Reaktionsmischung wurde für eine Stunde bei Raumtemperatur gerührt. Der Fortschritt der Reaktion konnte durch UV-VisSpektroskopie verfolgt werden. Nach beendeter Reaktion wurde das Reaktionsgemisch mit der doppelten Menge an Toluol versetzt und das Azeotrop Toluol/Essigsäure am Rotationsverdampfer abgezogen. Der verbliebene Standardaufarbeitung (5.3.1.1) isoliert. 154 Rückstand wird nach der Experimenteller Teil 5.3.2. Gewinnung von Chlorophyll a aus getrockneten Cyanobakterien Es wurden 2 g Bakterien mit einem Gemisch aus Aceton und gesättigter Na2CO3 Lösung (80/20) versetzt und unter Eiskühlung für 15 min mit Ultraschall aufgeschlossen. Die ungelösten Zellrückstände wurden sedimentiert und die organische Phase wurde mit Wasser und anschließend mit gesättigter Kochsalzlösung gewaschen, über Natriumsulfat getrocknet und einrotiert. Der Rückstand wurde durch HPLC-chromatographisch getrennt und man erhält 6,5 mg Chlorophyll a. UV/Vis (Diethylether): 660 nm (0,77), 613 nm (0,12), 576 nm (0,06), 530 nm (0,04), 428 nm (1,00), 409 nm (0,75) HPLC: - stationäre Phase: Nucleosil-5-C18 - mobile Phase: Methanol, 0,8 ml/min - Retentionszeit: 7,14 min Chl a 8,15 min Chl a´ MS (ESI): berechnet: 892 g/mol, gefunden: 915 [892+Na], 931 [892+K] 5.3.3. Umsetzung von Chlorophyll a zu Phäophorbid a Der Phytylester wurde nach 5.3.1.2 hydrolysiert. UV/Vis (Diethylether): 667 nm (0,58), 609 nm (0,08), 561 nm (0,03), 533 nm (0,10), 504 nm (0,12), 466 nm (0,04), 407 nm (1,00) HPLC: - stationäre Phase: Nucleosil-5-C18 - mobile Phase: Methanol / 2 M Ammoniumacetat: 8 / 1 - Retentionszeit: 5,02 min Phäophorbid a 6,38 min Phäophorbid a´ MS (ESI): berechnet: 592 g/mol, gefunden: 593 [592+H], 615 [592+Na] 155 Experimenteller Teil 5.3.4. Darstellung von Pyrophäophytin a aus getrockneten Cyanobakterien Es wurden 500 g Bakterien vier Mal mit je 500 ml Methanol extrahiert. Nach dem Abfiltrieren der unlöslichen Rückstände wurde das Methanol im Vakuum entfernt. Der zähflüssige Rückstand wurde mit 150 ml Collidin (2,4,6-Trimethylpyridin) versetzt und für sechs Stunden unter Argon am Rückfluss gekocht. Anschließend wurde das Collidin im Ölpumpenvakuum abdestilliert (p < 2 mbar, Kp 30 °C) und der verbliebene Rückstand durch MPLC gereinigt. Dazu wurde die Probe in Pentan : Diethylether (3:1) gelöst und zunächst mit Pentan eluiert. Im Verlauf der Trennung wurde das Lösemittel sukzessiv auf reinen Ether umgestellt. Es werden 2,5 g Pyrophäophytin a erhalten. UV/Vis (Diethylether): 667 nm (0,50), 609 nm (0,07), 534 nm (0,09), 505 nm (0,11), 409 nm (1,00) IR (KBr): 1731 cm-1 (CO2Phytyl), 1637, 1618 cm-1 (131-CO) 1 H-NMR (400MHz, CDCl3): 9,49 (s, 1H, 10-H), 9,38 (s, 1H, 5H), 8,57 (s, 1H, 20-H), 8,00 (dd, 1H, J=11,5 Hz, J=17,8 Hz, 31-CH), 6,28, 6,17 (je d, 1H, J=17,8 Hz, 1H, J=11,5 Hz AB von ABX 32-CH2), 5,25, 5,09 (je d, 1H, je J=19,9 Hz, 132-CH2), 4,53 (m, 1H, 18H), 4,29 (m, 1H, 17-H), 3,67 q, 2H, J=7,6 Hz, 81-CH2), 3,65 (s, 3H, 121-CH3), 3,41 (s, 3H, 21-CH3), 3,21 (s, 3H, 71-CH3), 2,76 (m, 1H, 171a-CH2), 2,56 (m, 1H, 172a-CH2), 2,33 (m, 2H, 171b-CH2, 172b-CH2), 1,79 (d, 3H, 181-CH3), 1,67 (t, 3H, J=7,6 Hz, 82-CH3), 0,32, 1,02 (je br s, 1H, NH), 0,7 bis 2,0 (mehrere breite m, Phytylester) MS (ESI): berechnet: 812 g/mol, gefunden: 813 [812+H], 835 [812+Na] 300 mg (0,369 mmol) Pyrophäophytin a wurden nach der Standardvorschrift 5.3.1.2 zu Pyrophäophorbid a mit der entsprechenden Carbonsäuregruppe hydrolysiert. Ohne Analyse oder Aufreinigung wird diese Verbindung direkt metalliert (5.3.1.3). Man erhält 170 mg (0,285 mmol, 77% ) ZnPPheid a. 156 Experimenteller Teil UV/Vis (Diethylether): 653 nm (0,58), 608 nm (0,12), 568 (0,07), 424 nm (1,00) HPLC: 1 - stationäre Phase: Nucleosil-5-C18 - mobile Phase: Methanol / 0,5 M Ammoniumacetat: 8 / 1 - Retentionszeit: 8,14 min ZnPPheid a H-NMR (400MHz, Pyridin): 10,05 (s, 1H, 10-H), 9,82 (s, 1H, 5-H), 8,80 (s, 1 H, 20-H), 8.37 (dd, 1H, J=11,5 Hz, J=18,0 Hz, X von ABX 31-CH), 6,40, 6,16 (je d, 1H, J=17,8 Hz, J=11,5 Hz AB von ABX 32-CH2), 5,58, 5,31 (d, 1H, J=19,5 Hz, d, 1H, J=19,5 Hz 132CH2), 4,68 (dq, 1H, J17,18=2,4 Hz, J18,181=7,5 Hz, 18-H), 4,51 (m, 1H, 17-H), 3,90 (q, 2H, 81-CH2, J=7,7 Hz), 3,88 (s, 3H, 121-CH3), 3,48 (s, 3H, 21-CH3), 3,36 (s, 3H, 71-CH3), 2,98 (m, 1H, 171a-CH2), 2,91 (m, 1H, 172a-CH2), 2,60 (m, 2H, 171b-CH2, 172b-CH2), 1,88 (d, 3H, J8, 181=7,4 Hz,181-CH3), 1.79 (t, 1H, J=7,7 Hz, 82-CH3) MS (ESI): berechnet: 596 g/mol, gefunden: 595 [596-H] 5.3.5. Darstellung von Methylpyrophäophorbid a (132-Decarboxymethylphäophorbid a Methylester) Es wurden 731 mg (0,9 mmol) Pyrophäophytin a in einem Gemisch aus 80 ml Methanol und 20 ml Schwefelsäure gelöst und für 48 Stunden bei Raumtemperatur gerührt. Der Reaktionsansatz wurde mit gesättigter Natriumhydrogencarbonatlösung neutralisiert und mit Diethylether extrahiert. Die vereinten organischen Phasen wurden mit Wasser ausgeschüttelt, über Natriumsulfat getrocknet und am Rotationsverdampfer eingeengt. Der schwarze Rückstand wird in Pentan aufgenommen und über Nacht bei –20 °C gelagert. Die schwarzen Kristalle wurden abfiltriert und man erhält Methylpyrophäophorbid a (336 mg, 74 %). UV/Vis (CH2Cl2): 667 nm (0,45), 609 nm (0,08), 539 nm (0,09), 508 nm (0,10), 413 nm (1,00) -1 IR (KBr): 1743 cm (CO2Me), 1636 cm-1, 1618 cm-1 (131-CO) Schmelzpunkt: 215 – 219°C (Lit.: 217 – 219°C)[130] 157 Experimenteller Teil 1 H-NMR (400MHz, CDCl3): 9,88 (s, 1H, 10-H), 9,72 (s, 1H, 5H), 8,90 (s, 1H, 20-H), 8.03 (dd, 1H, J=11.5Hz, J=17.8Hz 31-CH), 6.30 (dd, 1H, J=17.8Hz, J=11.5Hz, 32-CH2), 5.38, 5.19 (2d, je 1H, J=19.9Hz, 132-CH2), 4,64 (dq, 1H, J18,17=2,0 Hz, J18, 181=7,3 Hz, 18H), 4,42 (m, 1H, 17-H), 3,81 (q, 2H, J=7,7 Hz, 81-CH2), 3,75 (s, 3H , 121-CH3) , 3,66 (s, 3H, OCH3), 3,46 (s, 3H, 21-CH3), 3,33 (s, 3H, 71-CH3), 2,81 (m, 1H, 171a-H) , 2,64 (m, 1H, 172a-H), 2,4 (m, 2H, 171b, 172b-H), 1,90 (d, 3H, J=7,3 Hz, 181-CH3), 1,72 (t, 3H, J=7,6 Hz, 82-CH3), -0,44, -1,73 (je br s, 1H, NH) MS (ESI): berechnet: 548 g/mol, gefunden: 549 [548+H], 571 [548+Na] Die Verbindung wurde nach der Standardmethode 5.3.1.3 zu Zink-MePPheid a metalliert. UV/Vis (CH2Cl2): 654 nm (0,67), 608 nm (0,13), 568 nm (0,07), 426 nm (1,00) IR (KBr) : 1742 cm-1 (CO2Me), 1637 cm-1, 1618 cm-1 (131-CO) HPLC: 1 - stationäre Phase: Nucleosil-5-C18 - mobile Phase: Methanol / 0,5 M Ammoniumacetat: 8 / 1 - Retentionszeit: 13,75 min ZnMePPheid a H-NMR (400MHz, CDCl3): 10,00 (s, 1H, 10-H), 9,76 (s, 1H, 5-H), 8,74 (s, 1H, 20-H), 8,31 (dd, 1H, J=11,5 Hz, J=18 Hz, X von ABX 31-CH), 6,33, 6,10 (je d, 1H, J=11,5 Hz, J=18Hz, AB von ABX 32-CH2), 5,42, 5,20 (je d, 1H, J=20 Hz, 132-CH2), 4,53 (dq, 1H, J17,18=2,4 Hz, J18,181=7,3 Hz, 18-H), 4,35 (m, 1H, 17-H), 3,84 (q, J=7,4 Hz, 2H, 81-CH2), 3,83 3,62, 3,59, 3,43 (je s, 3H, 121-CH3, 71-CH3, OCH3, 21-CH3), 2,8-2,45 (m, 4H, 171CH2, 172-CH2), 1,80 (d, J=7,3 Hz, 3H, 181-CH3), 1,73 (t, J=7,5 Hz, 3H, 82-CH3) MS (ESI): berechnet: 610 g/mol, gefunden: 611 [610+H], 633 [610+Na] 5.3.6. Darstellung von Methylpyrophäophorbid d 380 mg (0,7 mmol) Methylpyrophäophorbid a wurden in 70 ml THF mit 10 mg (0,04 mmol) OsO4 gelöst. In fünf Minuten wurde eine Lösung von 0,8 g (3,7 mmol) NaIO4 und 300 µL Eisessig in 5 ml Wasser zugetropft. Nach 24 Stunden Rühren bei Raumtemperatur wurde die Mischung mit Dichlormethan versetzt und mit Wasser ausgeschüttelt. Die organische Phase wurde über Natriumsulfat getrocknet und einrotiert. Man erhält 182 mg (47 %) Methylpyrophäophorbid d (3-devinyl-3-formyl-132158 Experimenteller Teil demethoxycarbonylphäophorbid a Methylester) als schwarze Kristalle in ausreichender Reinheit zur Metallierung oder weiteren Umsetzung. UV/Vis (CH2Cl2): 689 nm (0,72), 629 nm (0,08), 549 nm (0,14), 517 nm (0,15), 422 nm (1,00) IR (KBr): 1739 (CO2Me), 1690 (131CO), 1673 (31-CHO) 1 H-NMR (500MHz, CDCl3): 11,54 (s, 1H, 31-CHO), 10,44 (s, 1H, 5-H), 9,72 (s, 1H, 10- H), 8,96 (s, 1H, 20-H), 5,38, 5,21 (d, je1H, J=19,7 Hz, 132-CH2), 4,63 (dq, 1H, J18,17=1,5 Hz, J18,181=7,4 Hz, 18-H), 4,42 (m, 1H, 17-H), 3,79 (s, 3H, 121-CH3), 3,73 (s, 3H, OCH3), 3,64 (s, 3H, 21-CH3), 3,33 (s, 3H, 71-CH3), 2,78 (m, 1H, 171a-H), 2,64 (m, 1H, 172a-H), 2,35 (m, 2H, 171b-H, 172b-H), 1,88 (d, 3H, J=7,4 Hz, 181-CH3), 1,71 (t, 3H, J=7,7 Hz, 82CH3) MS (ESI): berechnet: 550 g/mol, gefunden: 551 [550+H], 573 [550+Na] Das Methylpyrophäophorbid d wird analog zu 5.3.1.3 metalliert, jedoch wird als Lösemittel statt Eisessig ein Gemisch aus Dichlormethan-Methanol (50:1) verwendet. UV/Vis (CH2Cl2): 681 nm (0,99), 630 nm (0,13), 574 nm (0,08), 532 nm (0,06), 441 nm (1,00) HPLC: 1 - stationäre Phase: Nucleosil-5-C18 - mobile Phase: Methanol / Wasser: 8 / 1 - Retentionszeit: 9,34 min ZnMePPheid d H-NMR (500MHz, Pyridin): 11,72 (s, 1H, 31-CHO), 10,80 (s, 1H, 10-H), 10,05 (s, 1H, 5-H), 8,94 (s, 1H, 20-H), 5,46, 5,26 (je d, 1H, J= 20 Hz, 132-CH2), 4,60 (m, 1H, 18-H), 4,41 (m, 1H, 17-H), 3,86, (q, 2H, J=7,6 Hz, 81-CH2), 3,83, 3,75, 3,60, 3,36 (je s, 3H, 121-, 21-, 71-CH3, O-CH3), 2,9-2,3 (m, 4H, 171-CH2, 172-CH2), 1,83 (d, 3H, 181-CH3), 1,73 (t, 3H, J=7,6 Hz, 82CH3) MS (ESI): berechnet: 612 g/mol, gefunden: 613 [612+H], 635 [612+Na] 159 Experimenteller Teil 5.4. Rekonstitution von Apo-Myoglobin mit verschiedenen Kofaktoren 5.4.1. Einbau von Zink-Protoporphyrin Zu einer eisgekühlten Apo-Mb Lösung in 50 mM Phosphatpuffer pH 8,0 wurde unter Lichtausschluss eine ZnPPIX Lösung in DMSO zugetropft (1,5 eq). Der Anteil an DMSO am Gesamtvolumen sollte 5 % nicht übersteigen, um eine Denaturierung des Proteins zu vermeiden. Die Probe wurde auf Eis für zwei Stunden gerührt und anschließend filtriert, um den Überschuss an wasserunlöslichem Kofaktor zu entfernen. Die Aufreinigung erfolgte über zwei aufeinanderfolgende Sephadex G 25 Säulen (PD 10), die zuvor mit 50 mM Phosphatpuffer pH 7,0 äquilibriert wurden. Ein vollständiger Einbau kann durch UV/Vis Spektroskopie nachgewiesen werden, wenn das Verhältnis der Soret-Bande und der Proteinbande 10 beträgt (E428/E280 ≈ 10). Alle Proben, die ein Verhältnis E428/E280 > 9 aufwiesen, wurden für weitere Messungen verwendet. UV/Vis (H2O): 596 nm (0,05), 554 nm (0,07), 428 nm (1,00), 280 nm (0,10) 5.4.2. Einbau der Zink-Phäophorbid-Kofaktoren Zu einer Eisgekühlten Apo-Mb Lösung in 20 mM MES Puffer pH 6,2 wurden 2 eq des entsprechenden Zink-Phäophorbids in Pyridin langsam und unter Rühren zugetropft. Nach einer Stunde wird die Lösung zunächst filtriert und anschließend über zwei PD10 Säulen von ungebundenem Kofaktor und Pyridin befreit. UV/Vis (H2O): für ZnPPheid a Mb 660 nm (0,64), 620 nm (0,13), 437 nm (1,00), 415 nm (0,55), 392 nm (0,33), 333 nm (0,28) für ZnPheid a Mb 661 nm (0,76), 622 nm (0,15), 437 nm (1,00), 418 nm (0,64), 388 nm (0,37), 337 nm (0,32) 160 Experimenteller Teil für ZnMePPheid a Mb 658 nm (0,61), 622 nm (0,13), 436 nm (1,00), 417 nm (0,64), sh 390 nm, 335 nm (0,31) für ZnMePPheid d Mb 688 nm (0,85), 458 nm (1,00), 437 nm (0,87), 403 nm (0,61) für ZnPPheid a SwMb 660 nm (0,57), 623 nm (0,13), 437 nm (1,00), 416 nm (0,61), sh 392 nm, 335 nm (0,31) 5.5. Darstellung der [4Fe4S]-Modellpeptide 5.5.1. Die Peptide FA, FB, FX1 und FX2 Alle Peptide wurden wie unter 5.2.1 beschrieben synthetisiert. Die Sequenzen der einzelnen Peptide sind in Tabelle 14 zusammengefasst. Die Ausbeute für FA und FB lag bei 17 % nach der Aufreinigung. Die Reinheit wurde durch analytische HPLC und MALDITOF Massenspektrometrie überprüft. Tabelle 14: Sequenzen der Peptide, die als Modelle für die [4Fe4S]-Zentren im Photosystem I verwendet wurden. Bezeichnung des Peptids Sequenz (von N- zum C-Terminus) MS: FA TEDCVGCKRCKPECPW FB YDTCIGCTQCKPECPW FX1 RFPCDGPGRGGTCQVS FX2 AFPCDGPGRGGTCDIS FA berechnet: 1852,8 gefunden: 1850,5 FB berechnet: 1845,7 gefunden: 1845,6 FX1 berechnet: 1636,9 gefunden: 1636,3 FX2 berechnet: 1552,7 gefunden: 1552,1 161 Experimenteller Teil 5.5.2. Rekonstitution der Peptide Die synthetisierten Peptide wurden anhand von Literaturvorschriften mit [4Fe4S]-Zentren rekonstituiert[131; 132] . Außerdem kam ein abgeändertes Verfahren zum Einsatz. Beide Rekonstitutionsmethoden werden nachfolgend detailliert beschrieben. Sämtliche Arbeiten wurden in einer Glovebox unter Schutzgasatmosphäre durchgeführt. Alle Lösungen und Puffer wurden vor dem Einschleusen in die Glovebox mindestens 30 Minuten mittels Durchleitung von Argon entgast. Zu 50 mL eines 50 mM Tris-HCl Puffers, pH 8,3, wurden 400 µL β-Mercaptoethanol gegeben, gefolgt von 1 mg Peptidlösung in 0,1 mL des gleichen Puffers. Anschließend tropft man nacheinander langsam 150 µL einer 60 mM Eisen(III)chloridlösung in Puffer und 150 µL einer frischen 60 mM Natriumsulfidlösung, ebenfalls in Puffer gelöst, zu. Zwischen der Zugabe der Eisenchloridlösung und der Natriumsulfidlösung lagen 20 Minuten. Die Rekonstitutionsmischung wurde mit einem Septum und Parafilm verschlossen und über Nacht bei +4 °C in einem Kühlschrank außerhalb der Glovebox inkubiert. Die Lösungen von sechs bis 12 dieser Ansätze wurden vereint und durch Ultrafiltration auf ein Volumen von 2 mL aufkonzentriert. Der Überschuss an nicht gebundenem Eisen(III) und Natriumsulfid wurde durch zwei aufeinanderfolgende mit Sephadex G-25 Material gefüllte Säulen (PD 10-Säulen), die mit 50 mM Tris-HCl Puffer, pH 8,3, äquilibriert waren, entfernt. Es wurde eine braune Fraktion erhalten. In einem abgewandelten Verfahren wurden 10 mg Peptid in 1 mL Puffer, 1,5 mL Eisen(III)chloridlösung und 1,5 mL Natriumsulfidlösung in 50 mL Puffer nach dem obigen Schema vereint. Insbesondere bei diesem Verfahren musste die schwarze Lösung nach der Eisen(III)-Zugabe solange inkubiert werden, bis eine klare, beinahe farblose Lösung entstand, um die Bildung von Eisensulfid zu vermeiden. Die Aufreinigung erfolgt wie oben beschrieben. Zur Isotopenmarkierung mit 57 Fe wurde elementares, isotopenreines Eisen in konzentrierter Salzsäure gelöst. Um die Überspannung, die bei der Bildung von Wasserstoff an einer Eisenoberfläche auftritt, zu verhindern, wurde ein Platinblech in den Kolben gegeben. Die entstandene Lösung wurde im Vakuum zur Trockne eingeengt. Der zurückbleibende braun-gelbe Feststoff wurde in Puffer aufgenommen. Zur Herstellung von 162 Experimenteller Teil Proben in D2O wurde die gesamte Prozedur in Deuteriumoxid durchgeführt. Der Isotopeneffekt von Deuterium wurde bei der Angabe der pH-Werte nicht berücksichtigt. Zum Einsatz für EPR- und Mössbauer-Proben wurden die [4Fe4S]-Zentren mit Dithionit oder NaBH4 reduziert. Dazu wurde die Konzentration an Peptid in der Lösung bestimmt und eine stöchiometrische Menge an Dithionit oder NaBH4 in 0,5 M Tris-HCl Puffer, pH 8,3, zu der Lösung gegeben. 5.5.3. Herstellung der PS I Proben Das Photosystem I von Synechocystis 6803 sp. PCC 6803 und die Mutanten C14G C34S und C51G C34S der Untereinheit C wurden freundlicherweise von Donald A. Bryant und John H. Golbeck von der Pennsylvania State University, USA, zur Verfügung gestellt. Um die stromalen Untereinheiten C, D und E zu entfernen, wurde die Lösung der PS I Komplexe mit 6,8 M Harnstoff in 50 mM TrisHCl Puffer, pH 8,3, für 70 Minuten bei Raumtemperatur inkubiert. Anschließend wurde die Lösung über zwei Gelfiltrationssäulen mit Sephadex G-25 als fester Phase gereinigt, die zuvor mit 50 mM TrisHCl Puffer, pH 8,3, mit 0,04 % β-Dodecylmaltosid äquilibriert wurden. Die eluierte grüne Bande wurde durch Zentrifugation in Mikrokonzentratoren (Membrangröße 100 kDa) auf eine Konzentration von 150 µg/mL Chl a aufkonzentriert und mit einer 1 M Stammlösung Natriumascorbat versetzt, so dass eine Endkonzentration von 1 mM Ascorbat vorlag. Zusätzlich wurde DCPIP bis zu einer Endkonzentration von 10 mM zugegeben. Für die EPR-Experimente zum Nachweis der Bindung der Peptide an das PS I wurde ein mindestens 100facher Überschuss an Peptid relativ zum PS I verwendet, wodurch je nach Modellpeptid ein 10-20facher Überschuss an [4Fe4S]-Zentren im Verhältnis zur potentiellen Bindungsstelle vorlag. Für die optische Analyse der Ladungsrekombinationskinetik wurde ein 10facher Überschuss an Peptid verwendet. Nach der Zugabe der Peptide wurde die Probe über Nacht auf Eis inkubiert. Anschließend wurde diese Lösung 10 Minuten unter Eiskühlung im dunkeln inkubiert, um das Photosystem I zu reduzieren und anschließend im EPR-Probengefäß eingefroren. 163 Literatur 6. Literatur 1. Lehninger, A. L., Nelson, D. L., and Cox, M. M.; "Prinzipien der Biochemie", Tschesche, H., 1998, Akademischer Verlag, Heidelberg. 2. Voet, D. and Voet, J. G.; "Biochemie", 1992, VCH-Wiley, Weinheim. 3. Hyde, C. C., Ahmed, S. A., Padlan, E. A., Miles, E. W., Davies, D. R.; "3Dimensional Structure of the Tryptophan Synthase Alpha-2-Beta-2 Multienzyme Complex from Salmonella-Typhimurium", J. Biol. Chem., 1988, 263, 1785717871. 4. Radford, S. E.; "Protein folding: progress made and promises ahead", Trends Biochem. Sci., 2000, 25, 611-618. 5. Cantrell, A., Bryant, D. A.; "Molecular-Cloning and Nucleotide-Sequence of the PsaA and PsaB Genes of the Cyanobacterium Synechococcus SP PCC-7002", Plant Mol. Biol., 1987, 9, 453-468. 6. Brettel, K., Leibl, W.; "Electron transfer in photosystem I", Biochim. Biophys. Acta - Bioenergetics, 2001, 1507, 100-114. 7. Chamorovsky, S. K., Cammack, R.; "Direct Determination of the Midpoint Potential of the Acceptor-X in Chloroplast Photosystem-I by Electrochemical Reduction and Electron-Spin-Resonance Spectroscopy", Photobiochem. Photobiophys., 1982, 4, 195-200. 8. Parrett, K. G., Mehari, T., Warren, P. G., Golbeck, J. H.; "Purification and Properties of the Intact P-700 and FX-Containing Photosystem-I Core Protein", Biochim. Biophys. Acta, 1989, 973, 324-332. 9. Jordan, R., Nessau, U., and Schlodder, E.; "Photosynthesis: Mechanism and Effects", Garab, G., 1998, Kluwer Academic Publishers, Dordrecht. 164 Literatur 10. Evans, M. C. W., Heathcote, P.; "Effects of Glycerol on the Redox Properties of the Electron-Acceptor Complex in Spinach Photosystem-I Particles", Biochim. Biophys. Acta, 1980, 590, 89-96. 11. Nugent, J. H. A., Moller, B. L., Evans, M. C. W.; "Comparison of the ElectronParamagnetic-Res Properties of Photosystem-I Iron-Sulfur Center-A and Center-B in Spinach and Barley", Biochim. Biophys. Acta, 1981, 634, 249-255. 12. Cammack, R., Ryan, M. D., Stewart, A. C.; "EPR-Spectrum of Iron-Sulfur Center B in Photosystem-I of Phormidium-Laminosum", Febs Lett., 1979, 107, 422-426. 13. Capaldi, R. A.; "Structure and Function of Cytochrome-c-Oxidase", Annu. Rev. Biochem., 1990, 59, 569-596. 14. Phillips Jr., G. N.; 2005, "Myoglobin" in Handbook of Metalloproteins, Messerschmidt, A., Huber, R., Poulos, T., and Wieghardt, K., S. 5-15, Wiley, Weinheim. 15. Springer, B. A., Sligar, S. G., Olson, J. S., Phillips, G. N.; "Mechanism of Ligand Recognition in Myoglobin", Chem.Rev., 1994, 94, 699-714. 16. Suzuki, T., Imai, K.; "Evolution of myoglobin", Cell.Mol.Life Sci., 1998, 54, 9791004. 17. Kendrew, J. C., Dickerson, R. E., Strandberg, B. E., Hart, R. G., Davies, D. R.; "Structure of Myoglobin - A- Three-Dimensional Fourier Synthesis at 2 A Resolution", Nature, 1960, 185, 422-427. 18. Olson, J. S., Phillips, G. N.; "Myoglobin discriminates between O-2, NO, and CO by electrostatic interactions with the bound ligand", J.Biol.Inorg.Chem., 1997, 2, 544-552. 19. Yonetani, T., Yamamoto, H., Woodrow, G. V.; "Studies on Cobalt Myoglobins and Hemoglobins, I. Preparation and Optical Properties of Myoglobins and Hemoglobins containing Cobalt Proto-, Meso-, And Deuteroporphyrins and 165 Literatur Thermodynamic Chracterization of their Reversible Oxygenation", J. Biol. Chem., 1974, 249, 682-690. 20. Angiolillo, P. J., Vanderkooi, J. M.; "The Photoexcited Triplet State as a Probe of Chriomophore-Protein Interaction in Myoglobin", Biophys. J., 1998, 75, 14911502. 21. Deeb, R. S., Peyton, D. H.; "Proton NMR-Study of the Interaction of Tin(IV) Protoporphyrin-IX Monomers and Dimers with Apomyoglobin", Biochemistry, 1992, 31, 468-474. 22. Falk, H., Marko, H., Muller, N., Schmitzberger, W., Stumpe, H.; "Reconstitution of Apomyoglobin with Bile-Pigments", Monatsh. Chem., 1990, 121, 893-901. 23. Wright, K. A., Boxer, S. G.; "Solution Properties of Synthetic Chlorophyllide- and Bacteriochlorophyllide-Apomyoglobin Complexes", Biochemistry, 1981, 20, 75467556. 24. Boxer, S. G., Wright, K. A.; "Preparation and Properties of a ChlorophyllApomyoglobin Complex", J. Am. Chem. Soc., 1979, 101, 6791-6794. 25. LaMar, G. N., Pande, U., Hauksson, J. B., Pandey, R. K., Smith, K. M.; "Proton Nuclear Magnetic-Resonance Investigation of the Mechanism of the Reconstitution of Myoglobin that Leads to Metastable Heme Orientational Disorder", J. Am. Chem. Soc., 1989, 111, 485-491. 26. Hauksson, J. B., LaMar, G. N., Pandey, R. K., Rezzano, I. N., Smith, K. M.; "H-1NMR Study of the Role of Individual Heme Propionates in Modulating Structural and Dynamic Properties of the Heme Pocket in Myoglobin", J. Am. Chem. Soc., 1990, 112, 6198-6205. 27. Hauksson, J. B., LaMar, G. N., Pandey, R. K., Rezzano, I. N., Smith, K. M.; "H-1NMR Study of the Role of Individual Heme Propionates in Modulating Structural and Dynamic Properties of the Heme Pocket in Myoglobin", J. Am. Chem. Soc., 1990, 112, 6198-6205. 166 Literatur 28. Hunter, C. L., Lloyd, E., Eltis, L. D., Rafferty, S. P., Lee, H., Smith, M., Mauk, A. G.; "Role of the heme propionates in the interaction of heme with apomyoglobin and apocytochrome b(5)", Biochemistry, 1997, 36, 1010-1017. 29. Cadenas, E.; "Biochemistry of Oxygen-Toxicity", Annu. Rev. Biochem., 1989, 58, 79-110. 30. Gouterman, M., Snyder, L. C., Wagniere, G. H.; "Spectra of Porphyrins .2. 4 Orbital Model", J. Mol. Spectrosc., 1963, 11, 108ff. 31. Hartwich, G.; "Dynamik des Elektronentransfers in Reaktionszentren photosynthetischer Bakterien: Beeinflussung durch gezielten Pigmentaustausch. Dissertation.", 1994, Technische Universität München, München. 32. O.Q.Munro, J.C.Bradley, R.D.Hancock, H.M.Marques, F.Masicano, P.W.Wade.; "Molecular Mechanics Study of the Ruffling of Metalloporphyrins", J. Am. Chem. Soc., 1992, 114, 7218-7230. 33. W.Kaim and B.Schwederski.; "Bioanorganische Chemie", C.Elschenbroich, F.Hensel, and H.Hopf, 2004, Teubner Verlag, Stuttgart. 34. L.Pauling, C.D.Coryell.; "Magnetic properties and structure of hemoglobin, oxyhemoglobin and carbonmonoxyhemoglobin", Proc. Nat. Acad. Sci. USA, 1936, 22, 210. 35. J.J.Weiss.; "Nature of the iron-oxygen bond in oxy-hemoglobin", Nature, 1964, 202, 83. 36. Lovenberg W.; "Iron-Sulfur Proteins", 1977, Academic Press, New York. 37. Beinert, H.; "Iron-sulfur proteins: ancient structures, still full of surprises", J. Biol. Inorg. Chem., 2000, 5, 2-15. 38. Adman, E., Watenpaugh, K. D., Jensen, L. H.; "NH-S Hydrogen-Bonds in Peptococcus-Aerogenes Ferredoxin, Clostridium-Pasteurianum Rubredoxin, and 167 Literatur Chromatium High Potential Iron Protein", Proc. Nat. Acad. Sci. USA, 1975, 72, 4854-4858. 39. Swartz, P. D., Beck, B. W., Ichiye, T.; "Structural origins of redox potentials in FeS proteins: Electrostatic potentials of crystal structures", Biophys. J., 1996, 71, 2958-2969. 40. Jensen, G. M., Warshel, A., Stephens, P. J.; "Calculation of the Redox Potentials of Iron-Sulfur Proteins - The 2-/3- -Couple of [FE4S4-CYS(4)] Clusters in Peptococcus-Aerogenes Ferredoxin, Azotobacter-Vinelandii Ferredoxin-I, and Chromatium-Vinosum High-Potential Iron Protein", Biochemistry, 1994, 33, 10911-10924. 41. Langen, R., Jensen, G. M., Jacob, U., Stephens, P. J., Warshel, A.; "Protein Control of Iron-Sulfur Cluster Redox Potentials", J. Biol. Chem., 1992, 267, 25625-25627. 42. Stephens, P. J., Jollie, D. R., Warshel, A.; "Protein control of redox potentials of iron-sulfur proteins", Chem. Rev., 1996, 96, 2491-2513. 43. Noodleman, L., Case, D. A., Mouesca, J. M., Lamotte, B.; "Valence electron delocalization in polynuclear iron-sulfur clusters", J. Biol. Inorg. Chem., 1996, 1, 177-182. 44. Bominaar, E. L., Borshch, S. A., Girerd, J. J.; "Double-Exchange and Vibronic Coupling in Mixed-Valence Systems - Electronic-Structure of [FE4S4](3+) Clusters in High-Potential Iron Protein and Related Models", J. Am. Chem. Soc., 1994, 116, 5362-5372. 45. Dill, K. A., Shortle, D.; "Denatured States of Proteins", Annu. Rev. Biochem., 1991, 60, 795-825. 46. Shortle, D.; "Probing the Determinants of Protein Folding and Stability with Amino-Acid Substitutions", J. Biol. Chem., 1989, 264, 5315-5318. 168 Literatur 47. Merrifield, R. B.; "Solid Phase Peptide Synthesis .1. Synthesis of a Tetrapeptide", J. Am. Chem. Soc., 1963, 85, 2149ff. 48. Chan, W. C. and White, P. D.; "Fmoc Solid Phase Peptide Synthesis - A Practical Approach", 2000, Oxford University Press Inc., New York. 49. Lloyd-Williams, P., Albericio, F., and Giralt, E.; "Chemical Approaches to the Synthesis of Peptides and Proteins", 1997, CRC Press LLC, Boca Raton, Florida. 50. Knorr, R., Trzeciak, A., Bannwarth, W., Gillessen, D.; "New Coupling Reagents in Peptide Chemistry", Tetrahedron Lett., 1989, 30, 1927-1930. 51. Atherton, N. M.; "Electron Spin Resonance", 1973, Wiley, New York. 52. Gordy, W.; "Theory and Applications of Electron Spin Resonance", 1980, Wiley, New York. 53. Budil, D. E., Thurnauer, M. C.; "The Chlorophyll Triplet-State as a Probe of Structure and Function in Photosynthesis", Biochim. Biophys. Acta, 1991, 1057, 141. 54. Uphaus, R. A., Norris, J. R., Katz, J. J.; "Triplet-States in Photosynthesis", Biochem. Biophys. Res. Commun., 1974, 61, 1057-1063. 55. Angerhofer, A.; 1991, "Chlorophyll Triplets and Radical Pairs" in Chlorophylls, Scheer, H., S. 945-992, CRC Press, Boca Raton. 56. Davies, E. R.; "New Pulse ENDOR Technique", Phys. Lett. A, 1974, A 47, 1-2. 57. Frauenfelder, H.; "The Mössbauer Effect", 1962, Benjamin, New York. 58. Gol´danskii, V. I.; "The Mössbauer Effect and its Applications in Chemistry", 1964, Consultants Bureau, New York. 59. Münck, E.; 2000, "Aspects of 57Fe Mössbauer Spectroscopy" in Physical Methods in Bioinorganic Chemistry, Que Jr., L., S. 287-320, University Science Books, Sausalito, Kalifornien. 169 Literatur 60. Teale, F. W. J.; "Cleavage of the Haem-Protein Link by Acid Methylethylketone", Biochim. Biophys. Acta, 1959, 35, 543. 61. Scheer, H.; "Chlorophylls", Scheer, H., 1991, 157, CRC Press, München. 62. Käss, H., Lubitz, W., Hartwig, G., Scheer, H., Noy, D., Scherz, A.; "ENDOR studies of substituted chlorophyll cation radicals", Spectrochim. Acta Pt. A - Mol. Bio., 1998, 54, 1141-1156. 63. Strell, M., Urumov, T.; "New Methods for Introduction of Metals into Derivatives of Chlorophyll", Justus Liebigs Ann. Chem., 1977, 970-974. 64. Wasielewski, M. R., Svec, W. A.; "Synthesis of Covalently Linked Dimeric Derivatives of Chlorophyll-A, Pyrochlorophyll-A, Chlorophyll-B, and Bacteriochlorophyll-A", J. Org. Chem., 1980, 45, 1969-1974. 65. Smith, K. M., Goff, D. A., Simpson, D. J.; "Meso-Substitution of Chlorophyll Derivatives - Direct Route for Transformation of Bacteriopheophorbides-d into Bacteriopheophorbides-c", J. Am. Chem. Soc., 1985, 107, 4946-4954. 66. Tamiaki, H., Amakawa, M., Shimono, Y., Tanikaga, R., Holzwarth, A. R., Schaffner, K.; "Synthetic zinc and magnesium chlorin aggregates as models for supramolecular antenna complexes in chlorosomes of green photosynthetic bacteria", Photochem. Photobiol., 1996, 63, 92-99. 67. Shosheva, A. Ch., Devedjiev, Y. A. D., Christova, P. K., Atanasov, B. P.; "Preparation, Crystallizatio, and Spectral Properties of Zinc-Subsituted Sperm Whale Myoglobin", Stud. Biophys., 1989 , 129, 77-82. 68. Shulman, R. G., Wüthrich, K., Yamane, T., Patel, D. J., Blumberg, W. E.; "Nuclear Magnetic Resonance Determination of Ligand-Induced Conformational Changes in Myoglobin", J. Mol. Biol., 1970, 53, 143ff. 170 Literatur 69. Wright, K. A., Boxer, S. G.; "Solution Properties of Synthetic Chlorophyllide- and Bacteriochlorophyllide-Apomyoglobin Complexes", Biochemistry, 1981, 20, 75467556. 70. Hoffmann, B. M.; "Triplet-State Electron-Paramagnetic Resonance Studies of Zinc Porphyrins and Zinc-Substituted Hemoglobins and Myoglobins", J. Am. Chem. Soc., 1975, 97, 1688-1694. 71. Lendzian, F., Bittl, R., Telfer, A., Lubitz, W.; "Hyperfine structure of the photoexcited triplet state (3)P680 in plant PSII reaction centres as detennined by pulse ENDOR spectroscopy", Biochim. Biophys. Acta - Bioenergetics, 2003, 1605, 35-46. 72. Krimse, R. and Stach, J.; "ESR-SPektroskopie Anwendungen in der Chemie", Leibnitz, E., 1985, Akademie-Verlag, Berlin. 73. Sinnecker, S., Koch, W., Lubitz, W.; "Chlorophyll a radical ions: A density functional study", J. Phys. Chem. B, 2002, 106, 5281-5288. 74. Sinnecker, S., Koch, W., Lubitz, W.; "Bacteriochlorophyll a radical cation and anion-calculation of isotropic hyperfine coupling constants by density functional methods", Phys. Chem. Chem. Phys., 2000, 2, 4772-4778. 75. Davies, R. C., Pearlstein, R. M.; "Chlorophyllin-Apomyoglobin Complexes", Nature, 1979, 280, 413-415. 76. Thurnauer, M. C., Katz, J. J., Norris, J. R.; "Triplet-State in Bacterial Photosynthesis - Possible Mechanisms of Primary Photo-Act", Proc. Nat. Acad. Sci. USA, 1975, 72, 3270-3274. 77. Thurnauer, M. C., Norris, J. R.; "Ordering of Zero-Field Triplet Spin Sub-Levels in Chlorophylls - Magneto-Photoselection Study", Chem. Phys. Lett., 1977, 47, 100105. 171 Literatur 78. Clarke, R. H., Hanlon, E. B., Boxer, S. G.; "Investigation of the lowest triplet state of the pyrochlorophyllide a - Apomyoglobin complex by zero-field optically detected magnetic resonance spectroscopy", Chem. Phys. Lett., 1982, 89, 41-44. 79. Levanon, H., Norris, J. R.; "Photoexcited Triplet-State and Photosynthesis", Chem. Rev., 1978, 78, 185-198. 80. DiValentin, M., Kay, C. W. M., Giacometti, G., Möbius, K.; "A time-resolved electron nuclear double resonance study of the photoexcited triplet state of P680 in isolated reaction centers of photosystem II", Chem. Phys. Lett., 1996, 248, 434441. 81. Poluektov, O. G., Utschig, L. M., Schlesselman, S. L., Lakshmi, K. V., Brudvig, G. W., Kothe, G., Thurnauer, M. C.; "Electronic structure of the P-700 special pair from high-frequency electron paramagnetic resonance spectroscopy", J. Phys. Chem. B, 2002, 106, 8911-8916. 82. Carrington, A. and McLachlan, A. D.; "Introduction to Magnetic Resonance with Applications to Chemistry and Chemical Physics", 1969, Harper & Row, New York. 83. F.Lendzian, R.Bittl, W.Lubitz.; "Pulsed ENDOR of the photoexcited triplet states of bacteriochlorophyll a and of the primary donor P865 in reaction centers of Rhodobacter sphaeroides R-26", Photosynthesis Research, 1998, 55, 189-197. 84. Huber, M., Lendzian, F., Lubitz, W., Tränkle, E., Möbius, K., Wasielewski, M. R.; "ENDOR and TRIPLE Resonance in Solutions of the Chlorophyll-a and Bis(chlorophyll)cyclophane Radical Cations", Chem. Phys. Lett., 1986, 132, 467473. 85. Hoff, A. J., Lendzian, F., Möbius, K., Lubitz, W.; "Proton and Nitrogen Electron Nuclear Double and Triple Resonance of the Chlorophyll-a Anion in Liquid Solution", Chem. Phys. Lett., 1982, 85, 3-8. 172 Literatur 86. Tamura, M., Asakura, T., Yonetani, T.; "Heme Modification Studies of Myoglobin .1. Purification and Some Optical and EPR Characteristics of Synthesized Myoglobins Containing Unnatural Hemes", Biochim. Biophys. Acta, 1973, 295, 467-479. 87. Liong, E. C., Dou, Y., Scott, E. E., Olson, J. S., Phillips, G. N.; "Waterproofing the heme pocket - Role of proximal amino acid side chains in preventing hemin loss from myoglobin", J. Biol. Chem., 2001, 276, 9093-9100. 88. CARVER, T. E., BRANTLEY, R. E., SINGLETON, E. W., Arduini, R. M., QUILLIN, M. L., Phillips, G. N., Olson, J. S.; "A Novel Site-directed Mutant of Myoglobin with an Unusually High O2 Affinity and Low Autooxidation Rate", J. Biol. Chem., 1992, 267, 14443-14450. 89. Springer, B. A., Sligar, S. G.; "High-Level Expression of Sperm Whale Myoglobin in Escherichia-coli", Proc. Nat. Acad. Sci. USA, 1987, 84, 8961-8965. 90. K.S.Reddy, Angiolillo, P. J., W.W.Wright, M.Laberge, Vanderkooi, J. M.; "Spectral Splitting in the a(Q0,0) Absorption Band of Ferrous Cytochrome c and Other Heme Proteins", Biochemistry, 1996, 35, 12820-12830. 91. Sweeney, W. V., Rabinowitz, J. C.; "Proteins Containing 4FE-4S Clusters - An Overview", Annu. Rev. Biochem., 1980, 49, 139-161. 92. Mouesca, J. M., Lamotte, B.; "Iron-sulfur clusters and their electronic and magnetic properties", Coord. Chem. Rev., 1998, 180, 1573-1614. 93. Vassiliev, I. R., Antonkine, M. L., Golbeck, J. H.; "Iron-sulfur clusters in type I reaction centers", Biochim. Biophys. Acta - Bioenergetics, 2001, 1507, 139-160. 94. Mulholland, S. E., Gibney, B. R., Rabanal, F., Dutton, P. L.; "Characterisation of the Fundamental Protein Ligand Requirements of [4Fe-4S]2+/+ Clusters with Sixteen Amino Acid Marquettes", J. Am. Chem. Soc., 1998, 120, 10296-10302. 173 Literatur 95. Mulholland, S. E., Gibney, B. R., Rabanal, F., Dutton, P. L.; "Determination of Nonligand Amino Acids Critical to [4Fe-4S]2+/+ Assembly in Ferredoxin Maquettes", Biochemistry, 1999, 38, 10442-10448. 96. Gibney, B. R., Mulholland, S. E., Rabanal, F., Dutton, P. L.; "Ferredoxin and ferredoxin-heme maquettes", Proc. Nat. Acad. Sci. USA, 1996, 93, 15041-15046. 97. Scott, M. P., Biggins, J.; "Introduction of a [4Fe-4S (S-cys)4]+1/+2 iron-sulfur center into a four-a helix protein using design parameters from the domain of the FX-cluster in the Photosystem I reaction center", Protein Sci., 1997, 6, 340-346. 98. Antonkine, M. L., Liu, G. H., Bentrop, D., Bryant, D. A., Bertini, I., Luchinat, C., Golbeck, J. H., Stehlik, D.; "Solution structure of the unbound, oxidized photosystem I subunit PsaC, containing [4Fe-4S] clusters F-A and F-B: a conformational change occurs upon binding to photosystem I", J. Biol. Inorg. Chem., 2002, 7, 461-472. 99. Pace, C. N., Vajdos, F., Fee, L., Grimsley, G., Gray, T.; "How to Measure and Predict the Molar Absorption-Coefficient of a Protein", Protein Sci., 1995, 4, 24112423. 100. Edelhoch, H.; "Spectroscopic Determination of Tryptophan and Tyrosine in Proteins", Biochemistry, 1967, 6, 1948ff. 101. Armengaud, J., Gaillard, J., Forest, E., Jouanneau, Y.; "Characterization of a 2[4FE-4S] Ferredoxin Obtained by Chemical Insertion of the FE-S Clusters into the Apoferredoxin-II from Rhodobacter-Capsulatus", Eur. J. Biochem., 1995, 231, 396-404. 102. Kennedy, M. L., Gibney, B. R.; "Proton coupling to [4Fe-4S](2+/+) and [4Fe4Se](2+/+) oxidation and reduction in a designed protein", J. Am. Chem. Soc., 2002, 124, 6826-6827. 103. Moura, J. J. G., Macedo, A. L., Palma, P. N.; "Ferredoxins", Methods Enzymol., 1994, 165-188. 174 Literatur 104. Beinert, H.; "Recent Developments in the Field of Iron-Sulfur Proteins", FASEB J., 1990, 4, 2483-2491. 105. Heller, C., McConnell, H. M.; "Radiation Damage in Organic Crystals .2. Electron Spin Resonance of (CO2H)CH2CH(CO2H) in Beta-Succinic Acid", J. Chem. Phys., 1960, 32, 1535-1539. 106. Doan, P. E., Fan, C., Hoffmann, B. M.; "Pulsed 1,2-H ENDOR and 2H-2H TRIPLE Resonance of H-Bonds and Cysteinyl b -CH2 of the D. gigas Hydrogenase [3Fe4S]+ Cluster", J. Am. Chem. Soc., 1994, 116, 1033-1041. 107. Mouesca, J. M., Rius, G., Lamotte, B.; "Single-Crystal Proton ENDOR Studies of the [FE4S4](3+) Cluster - Determination of the Spin Population-Distribution and Proposal of a Model to Interpret the H-1-NMR Paramagnetic Shifts in HighPotential Ferredoxins", J. Am. Chem. Soc., 1993, 115, 4714-4731. 108. Backes, G., Mino, Y., Loehr, T. M., Meyer, T. E, Cusanovich, M. A., Sweeney, W. V., Adam, E. T., Sandersloehr, J.; "The Environment of FE4S4 Clusters in Ferredoxins and High-Potential Iron Proteins - New Information From X-Ray Crystallography and Resonance Raman-Spectroscopy", J. Am. Chem. Soc., 1991, 113, 2055-2064. 109. Beinert, H., Holm, R. H., Münck, E.; "Iron-Sulfur Clusters: Nature´s Modular, Multipurpose Structures", Science, 1997, 277, 653-659. 110. Dickson, D. P. E., Cammack, R.; "Mössbauer-Effect in Super-Reduced Form of High-Potential Iron-Sulfur Protein From Chromatium", Biochem. J., 1974, 143, 763-765. 111. Thompson, C. L., Johnson, C. E., Dickson, D. P. E., Cammack, R., Hall, D. O., Weser, U., Rao, K. K.; "Mössbauer-Effect in 8-Iron Ferredoxin From ClostridiumPasteurianum - Evidence for State of Iron Atoms", Biochem. J., 1974, 139, 97-103. 175 Literatur 112. Dickson, D. P. E., Johnson, C. E., Cammack, R., Evans, M. C. W., Hall, D. O., Rao, K. K.; "Mössbauer-Effect in High-Potential Iron-Sulsfur Protein From Chromatium - Evidence for State of Iron Atoms", Biochem. J., 1974, 139, 105-108. 113. Vassiliev, I. R., Jung, Y. S., Smart, L. B., Schulz, R., McIntosh, L., Golbeck, J. H.; "A Mixed-Ligand Iron-Sulfur Cluster (C556S(PsaB) or C565S(PsaB)) in the F-XBinding Site Leads to a Decreased Quantum Efficiency of Electron-Transfer in Photosystem", Biophys. J., 1995, 69, 1544-1553. 114. Cohen, R. O., Shen, G. Z., Golbeck, J. H., Xu, W., Chitnis, P. R., Valieva, A. I., van der Est, A., Pushkar, Y., Stehlik, D.; "Evidence for asymmetric electron transfer in cyanobacterial photosystem I: Analysis of a methionine-to-leucine mutation of the ligand to the primary electron acceptor A(0)", Biochemistry, 2004, 43, 4741-4754. 115. Golbeck, J. H., Mehari, T., Parrett, K., Ikegami, I.; "Reconstitution of the Photosystem-I Complex from the P700 and FX-Containing Reaction Center Core Protein and the FA/FB Polypeptide", Febs Lett., 1988, 240, 9-14. 116. Hales, B. J.; "Intinsic and Extrinsic Paramagnets as Probes of Metal-Clusters", Methods Enzymol., 1993, 384-395. 117. Koch, W. and Holthausen, M. C. A.; "A Chemists Guide to Density Functional Theory", 2000, Wiley-VCH, Weinheim. 118. Perdew, J. P.; "Correction", Phys. Rev. B, 1986, 34, 7406. 119. Perdew, J. P.; "Density-Functional Approximation for the Correlation-Energy of the Inhomogenous Electron-Gas", Phys. Rev. B, 1986, 33, 8822-8824. 120. Becke, A. D.; "Density-Functional Exchange-Eenergy Approximation with Correct Asymptotic-Behavior", Phys. Rev. A, 1988, 38, 3098-3100. 121. Schafer, A., Horn, H., Ahlrichs, R.; "Fully Optimized Contracted Gaussian-Basis Sets for Atoms Li to Kr", J. Chem. Phys., 1992, 97, 2571-2577. 176 Literatur 122. Ahlrichs, R., Bär, M., Baron, H.-P., Bauernschmitt, R., Böckinger, S., Ehring, M., Eichkorn, K., Eliiot, S., Furche, F., Haase, F., Häser, M., Horn, H., Hättig, C., Huber, C., Huniar, U., Kattannek, M., Köhn, A., Kölmel, C., Kollwitz, M., May, K., Ochsenfeld, C., Öhm, H., Schäfer, A., Schneider, U., Treutler, O., von Arnim, M., Weigend, F., Weis, P., and Weiss, H.; Turbomole, 5-3-2000, Quantum Chemistry Group, University of Karlsruhe, Karlsruhe, Germany. 123. Neese, F.; ORCA - an ab initio, Density Functional and Semiempirical Program Package, 2004, Max-Planck Institut für Bioanorganische Chemie, Mülheim an der Ruhr, Germany. 124. Becke, A. D.; "Density-Functional Thermochemistry .3. The Role of Exact Exchange", J. Chem. Phys., 1993, 98, 5648-5652. 125. Lee, C. T., Yang, W. T., Parr, R. G.; "Development of the Colle-Salvetti Correlation-Energy Formula into a Functional of the Electron-Density", Phys. Rev. B, 1988, 37, 785-789. 126. Barone, V.; "Recent Advances in Density Functional Methods", Chong, D. P., 1996, 287, Ed. World Scientific Publ. Co., Singapore. 127. Schafer, A., Huber, C., Ahlrichs, R.; "Fully Optimized Contracted Gaussian-Basis Sets of Triple Zeta Valence Quality for Atoms Li to Kr", J. Chem. Phys., 1994, 100, 5829-5835. 128. Hirota, S., Azuma, K., Fukuba, M., Kuroiwa, S., Funasaki, N.; "Heme reduction by intramolecular electron transfer in cysteine mutant myoglobin under carbon monoxide atmosphere", Biochemistry, 2005, 44, 10322-10327. 129. Hirota, S., Mizoguchi, Y., Yamauchi, O., Kitagawa, T.; "Observation of an isotopesensitive low-frequency Raman band specific to metmyoglobin", J. Biol. Inorg. Chem., 2002, 7, 217-221. 177 Literatur 130. Smith, K. M., Goff, D. A., Simpson, D. J.; "Meso-Substitution of Chlorophyll Derivatives - Direct Route for Transformation of Bacteriopheophorbides-d into Bacteriopheophorbides-c", J. Am. Chem. Soc., 1985, 107, 4946-4954. 131. Antonkine, M. L., Bentrop, D., Bertini, I., Luchinat, C., Shen, G. Z., Bryant, D. A., Stehlik, D., Golbeck, J. H.; "Paramagnetic H-1 NMR spectroscopy of the reduced, unbound Photosystem I subunit PsaC: sequence-specific assignment of contactshifted resonances and identification of mixed- and equal-valence Fe-Fe pairs in [4Fe-4S] centers F-A(-) and F-B(-)", J. Biol. Inorg. Chem., 2000, 5, 381-392. 132. Mehari, T., Parrett, K. G., Warren, P. V., Golbeck, J. H.; "Reconstitution of the Iron-Sulfur Clusters in the Isolated FA/FB Protein - EPR Spectral Characterization of Same-Species and Cross-Species Photosystem-I Complexes", Biochim. Biophys. Acta, 1991, 1056, 139-148. 178 Lebenslauf Name: Christoph Breitenstein Geburtsdatum: 27. März 1976 Geburtsort: Oberhausen Ausbildung 1982-1986 Gemeinschaftsgrundschule an der Schlägelstrasse 1986-1995 Otto-Pankok-Gymnasium Mülheim Juni 1995 Abitur 1996-2002 Hochschulstudium Chemie an der Gerhard-Mercator-Universität Gesamthochschule Duisburg Februar 2002 Diplom in Chemie mit dem Thema „Synthese 13C markierter Retinale“ März 2002 Beginn der Promotion am Max-Planck-Institut für Bioanorganische Chemie in Mülheim Wehrdienst 1995-1996 3. U-Flottille, Kiel, Versorger Meersburg